Embed Size (px)
Citation preview

Comparative analysis of the forestomachmucosa in red deer during prenatal deve-lopment
A.J. MASOT*2, A.J. FRANCO1 AND E. REDONDO2
1 Department of Veterinary Anatomy, Faculty of Veterinary Medicine, University of Extremadura, Cáceres, SPAIN2 Department of Veterinary Histology, Faculty of Veterinary Medicine, University of Extremadura, Cáceres, SPAIN
* Corresponding author: E-mail: [email protected]
SUMMARY
Red deer is an excellent topic of study given its economic importance inrelation to hunting, and for being a bearer of a genetic heritage of incalcula-ble wealth. For this reason, the comparative histological evaluation of the on-togenesis of forestomach mucosa in deer is the object of our study.Histomorphometric and immunohistochemical analyses were carried out on50 embryos and fetuses of deer from the initial stages of prenatal life untilbirth, divided into five experimental groups. At 60 days (25% of gestation) anoutline of the ruminal and reticular compartments appears. At 67 days theomasum wall was differentiated. The mucosa of the non-glandular stomachcompartments, was composed of three layers: epithelial layer, pluripotentialblastemic tissue and serosa. The stratification of the epithelial layer of fores-tomach mucosa was accompanied by changes in its structure with the ap-pearance of the ruminal pillars and papillae in the rumen (at 25% ofgestation); of the primitive reticular cellule, and primary ribs (40% of gesta-tion) in the reticulum; and of the primary laminae (25% of gestation) in theomasum. In comparison with domestic ruminants, red deer were shown asbeing less precocious with regard to development of the non-glandular sto-mach mucosa, in their capacity to secrete neutral mucopolysaccharides; andin their neuroendocrine nature, as determined by the detection of positiveneuroendocrine and/or glial cells.
Keywords : Red deer, mucosa, forestomach, prenatal de-velopment.
RÉSUMÉ
Étude comparative de la muqueuse des estomacs non glandulaires ducerf rouge pendant le développement prénatal.
Le cerf rouge est un excellent sujet d'étude vu son importance économiquedans la relation à la chasse, et pour véhiculer un héritage génétique de valeurincalculable. Pour cette raison, l'étude histologique comparative du dévelop-pement de la muqueuse des estomacs non glandulaires, a été l'objet de notreétude. Cinquante embryons et fœtus de cerfs (répartis dans cinq groupes ex-périmentaux) depuis les étapes initiales de la vie prénatale jusqu'à la nais-sance, ont été étudiés par immunohistochimique et morphométrie. Ladifférenciation du rumen et bonnet s'est produite à 60 jours (25% de gesta-tion) ; celle de l'omasum à 67 jours. La muqueuse des compartiments non-glandulaires de l'estomac, était composée de trois couches : épithélium, tissublastémique pluripotentiel et séreux. La stratification de la lame épithéliale dela muqueuse était accompagnée de changements dans sa structure, avec l'ap-parition des piliers et des papilles du rumen (à 25% de gestation) ; des celluleset les crêtes primaires du bonnet (à 40% de gestation) ; et des lamelles pri-maires de l'omasum (à 25% de gestation). En comparaison avec les rumi-nants domestiques, la muqueuse des estomacs non glandulaires du cerf rougea été moins précoce dans sa capacité à sécréter des mucopolysaccharides neu-tres ; et dans sa fonction neuroendocrine, évaluée par la détection des cel-lules neuroendocriniennes et/ou gliales.
Mots-clés : Cerf rouge, muqueuse, estomacs non glandu-laires, développement prénatal.
IntroductionRed deer in the Iberian Peninsula constitute a heritage to be
conserved for two reasons: as a hunting species and as an eco-nomic activity. As a species for hunting, it is a bearer of a ge-netic wealth whose conservation is vital, due to the fact that itsautochthonous nature is necessary for the maintenance of theecosystem. On the other hand, the economic activity that is de-rived from this species not only depends on game hunting butfrom its increasingly intense iruption in the food industry, giventhat the meat of deer offers significant culinary potential.
The hunting of red deer constitutes one of the main econo-mic uses of Mediterranean forests and “dehesas” (open oakforests) in the south west of the Iberian Peninsula. The mana-gement of its population has undergone a notable evolutionin past years [1]. The traditional system of intensive farmingmethods is gradually being replaced by a mixed system of in-tensive-semi-intensive methods, where new management me-thods require intervention in many areas, food being the areathat most concerns us here.
Food supplementation, on the one hand, give rise to ecolo-gical and behavioural changes in the species, since the addi-tion of food in the most unfavourable season can result in thenumber of heads that an area can support rising by a signifi-cantly increase. Moreover the period of food shortage in themost southerly areas coincides with the rutting season [1]. Onthe other hand, added to the undesirable ecological effect fromthe spatial concentration of food are the changes that comeabout from such supplements on the histophysiology of thestomach of deer.
The high capacity of the ruminants to convert fibrous foodsinto great nutritious value nutrients has impelled, to a greatextent, the study of the structure and function of the digestivetract in these animals. This degradative process of ingestedfood, conditions the morphology of the mucosa of the fores-tomach of the ruminants, especially that of the rumino-reti-cular complex; the omasal mucosa is also affected albeit to alesser extent due to the smaller role of the omasum in the de-gradation processes.
Revue Méd. Vét., 2007, 158, 7, 397-409
Many investigations have been made on the prenatal deve-
lopment of the gastric compartments of the ruminants [3-11],
nevertheless few investigations have been done on the prena-
tal development of the mucosa of the nonglandular gastric
compartments in red deer. Even fewer studies of a compara-
tive nature have been done [12-14].
The object of this work has been to do a comparative histo-
logical study of the mucosa of nonglandular gastric compart-
ments of red deer throughout its development. The study has
been carried out in an extensive deer farming system, without
food supplements, with the objective of establishing a mor-
phologic database of prenatal development of the reticulo-
rumen and omasum without the possible influences that come
from these supplements. The study is also intended to serve as
a reference base for future studies that look at these afore-
mentioned influences.
Materials and MethodsANIMALS
Embryos and fetuses of red deer (Cervus elaphus hispani-cus) (n=25) from the initial prenatal stages until birth werestudied. The specimens were arranged in five groups, five ani-mals each, with reference to the most relevant histomorpho-genic characteristics (Tables I and II). To obtain embryos andfetuses at various stages of development, a total of 125 lapa-rotomies on the same number of females were performed. Thefemales were taken from ten hunting grounds from extensiveand non enclosed-type estates from the Sierra of San Pedro(Northeast of the Province of Cáceres, Spain).
SAMPLING AND PROCESSING
Once the non-glandular stomach was separated, it was ana-lysed by visual and stereomicroscopic inspection. The colou-ring and the consistency of the mucosa were determined.
Square specimens measuring 1.5 x 0.5 cm were taken fromthe medial region of each compartment (rumen, reticulum andomasum) of each red deer. Tissue for histological study werefixed in 4% buffered formaldehyde, processed by conventio-nal paraffin-embendding methods, and sections 5 µm thickwere cut in transversly and stained with haematoxylin andeosin (H-E); Periodic Acid-Schiff (pH 7.2) and PAS-alcianblue (pH 7.2) for specific differentiation of neutral and acidmucopolysaccharides; Van Giesson (VG); Masson’s trichrome(MT) and Reticuline of Gomori (RG).
MORPHOMETRIC ANALYSIS
Specimens for morphometric analysis were embedded inparaffin, stained with H-E, and viewed through a microscope(Optiphot, Nikon Inc, Tokyo, Japan) equipped with a video
camera. The image was reflected onto the screen of a semi-au-tomatic image analyser (Vid IV, Rego and Cía, Madrid,Spain). Variables studied were height of epithelium and totalwall thickness. Eight specimens (sections) were selected foreach group, and 50 measurements were made for each speci-men.
The results are shown as the average ± standard error. Thedata were analysed using analysis of the variance. In the caseswhere the ANOVA was significant, a post hock (Tuckey) ana-lysis was carried out in order to study the significant diffe-rences among the distinct groups. A value of P 0.05 wasconsidered significant.
Tissue growth models were created, using a personal com-puter and statistics program (Statgraphics V 2.1 (1986)). InFigs 25-27 the graphs represent the averages of the real
398 MASOT (A.J.) AND COLLABORATORS
Revue Méd. Vét., 2007, 158, 7, 397-409
GROUP I(CRL cm)(1.4 to 3.6 )30-60 days
GROUP IICRL (cm)(4.5 to 7.2)67-90 days
GROUP IIICRL (cm)(8 to 19 )
97-135 days
GROUP IVCRL (cm)(21 to 33 )
142-191 days
GROUP VCRL (cm)(36 to 40 )
205-235 days
RUMEN
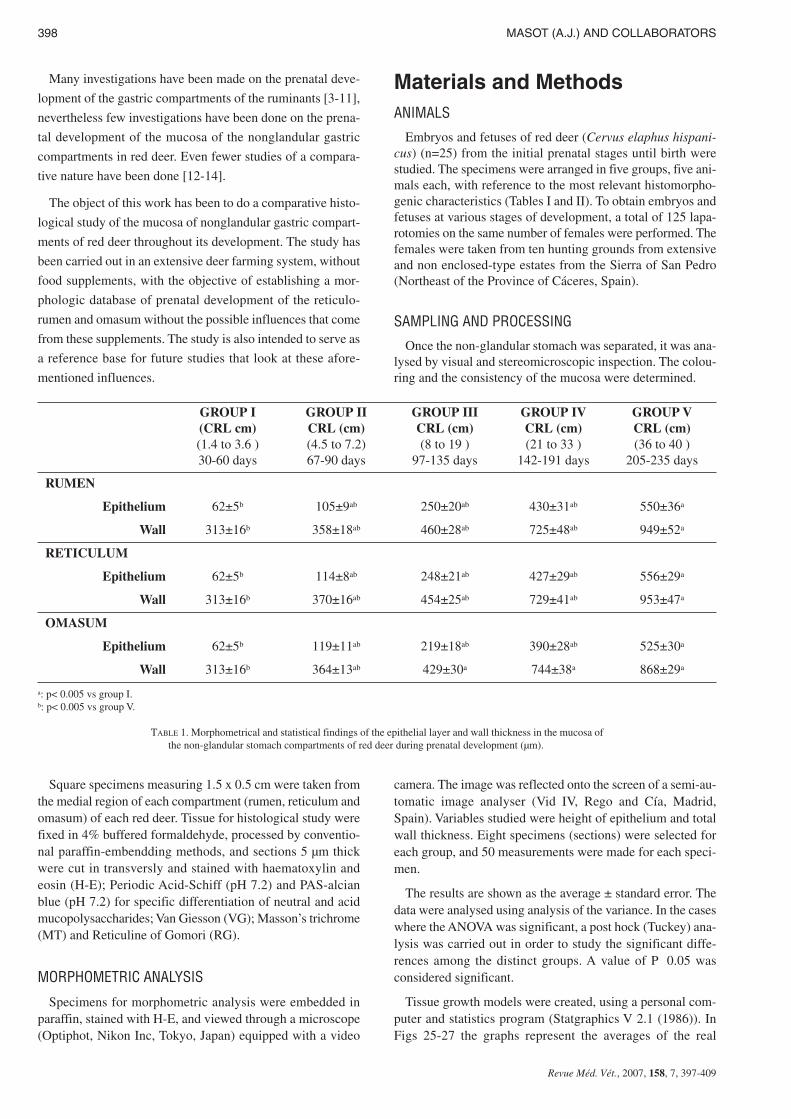
Epithelium 62±5b 105±9ab 250±20ab 430±31ab 550±36a
Wall 313±16b 358±18ab 460±28ab 725±48ab 949±52a
RETICULUM
Epithelium 62±5b 114±8ab 248±21ab 427±29ab 556±29a
Wall 313±16b 370±16ab 454±25ab 729±41ab 953±47a
OMASUM
Epithelium 62±5b 119±11ab 219±18ab 390±28ab 525±30a
Wall 313±16b 364±13ab 429±30a 744±38a 868±29a
a: p< 0.005 vs group I.b: p< 0.005 vs group V.
TABLE 1. Morphometrical and statistical findings of the epithelial layer and wall thickness in the mucosa ofthe non-glandular stomach compartments of red deer during prenatal development (µm).

growth values next to the adjusted line of regression. The suc-cess of fit of this adjustment was measured using the rate ofdetermination r2. In all cases, embryo body length (crown torump; CRL(cm)) was used as the independent variable; thethickness of each tissue stratum served as the dependent va-riable.
IMMUNOCYTOCHEMICAL ANALYSIS
ExtrAvidin Peroxidase Staining (EAS) was performed on
deparaffinized sections from the rumen reticulum and oma-
sum to detect the neuroendocrine cells markers [non-neuron
enolase (NNE)] and glial cells markers [glial fibrillary acidic
PRENATAL DEVELOPMENT OF FORESTOMACH 399
Revue Méd. Vét., 2007, 158, 7, 397-409
RUMEN
GROUP I30-60 Days
GROUP II67-90 Days
GROUP III97-135 Days
GROUP IV142-191 Days
GROUP V205-235 Days
E LP-S TM S E LP-S TM S E LP-S TM S E LP-S TM S E LP-S TM S
NNE
- - - - - - - - + + + - + ++ ++ - + ++ ++ -
GFAP
- - - - - - - - - - - - - ++ ++ ++ - ++ ++ ++
VI
M
- - - - - + + + - + + + - ++ ++ ++ - ++ ++ ++
RETICULUM
GROUP I30-60 Days
GROUP II67-90 Days
GROUP III97-135 Days
GROUP IV142-191 Days
GROUP V205-235 Days
E LP-S TM S E LP-S TM S E LP-S TM S E LP-S TM S E LP-S TM S
NNE
- - - - - - - - + + + - + ++ ++ - + ++ ++ -
GFAP
- - - - - - - - - - - - - ++ ++ ++ - ++ ++ ++
VI
M
- - - - - + + + - ++ ++ ++ - ++ ++ ++ - +++ +++ +++
OMASUM
GROUP I30-60 Days
GROUP II67-90 Days
GROUP III97-135 Days
GROUP IV142-191 Days
GROUP V205-235 Days
E LP-S TM S E LP-S TM S E LP-S TM S E LP-S TM S E LP-S TM S
NNE
- - - - + + + - + ++ ++ - + +++ +++ - + +++ +++ -
GFAP
- - - - - + + + - + + + - ++ ++ + - +++ +++ ++
VI
M
- - - - - + + + - + + + - ++ ++ + - +++ +++ ++
- Non immunoreactivity+ Low IR++ Moderate IR+++ High IRE = Epithelium - Lp-S = Lamina propria-submucosa - T = Tunica muscularis - S = Serosa
TABLE 2. Neuropeptides present in the mucosa of the non-glandular stomach compartments of red deer during prenatal development.

protein (GFAP), vimentin (VIM) and non-neuron enolase(NNE)].
Tissue was deparaffinized, hydrated and treated sequentiallywith 0.5% hydrogen peroxide in methanol for 30 min in orderto block endogenous peroxidase activity. Non-specific tissuebinding sites were blocked by incubation in 1% normal goatserum for 30 min. Samples were incubated with the followingdilution of primary antisera in PBS: 1: 200 monoclonal anti-human NNE (Sigma/Aldrich Química, Madrid, Spain, no.S5768); 1: 400 monoclonal anti-human GFAP (Sigma/AldrichQuímica, Madrid, Spain, no. G-3893); 1:20 monoclonal anti-human VIM (Sigma/Aldrich Química, Madrid, Spain, no. V-5255 for 3 hours at 20ºC. Biotinylated goat anti-mouse IgG (1:200 dilution) (Sigma/Aldrich Química, Madrid, Spain no.B7151) was then added to the sections for 30 min. Sectionswere finally incubated with diluted (1:50) ExtrAvidin-Horse-radish Peroxidase (Sigma/Aldrich Química, Madrid, Spain no.E2886) for 1 h. After diaminobenzidine reaction, nuclearcounterstaining with Mayer hematoxylin was applied. Finallythe section were mounted with Entellan (Merck 7961).
The specificity of the staining reaction was determined incontrol experiments. These comprised, substitution of the pri-mary antibody by PBS or normal mouse serum 1:100; oromission of both primary and secondary antibodies; and priorabsorption of the primary antibody (overnight preincubationof the primary antisera with the respective peptide 50-100µM). The antibody/peptide mixture was applied to sections inthe identical manner and concentration of the primary anti-body.
ResultsHISTOMORPHOGENESIS OF EPITHELIAL LAYER
Group I (1.4 to 3.6 cm C-R, 30-60 days, 1-25% of gestation)
Primitive gastric tube
At 30-60 days of prenatal development, the primitive out-line of the gastric tube showed a thin cavity surrounded by athick wall (313 ± 16 µm). The wall was composed of twolayers: one internal epithelial layer (62 ± 5 µm), adjacent tothe tube and the other external, of pluripotential blastemic tis-sue. The epithelium was pseudostratified and was not cilia-ted.
At 60 days of gestation the outline of the ruminal (Figure1a) and reticular (Figure 1b) compartments appear. The diffe-rentiation of the omasal individualised compartment from theprimitive gastric tube had still not taken place.
Group II (4.5 to 7.2 cm C-R, 67-90 days, 25-35% of gesta-tion)
Ruminal epithelial layer
The wall of the rumen displayed a thickness of 358±18 µm.We witnessed an evident thickening of the stratified epithe-lial layer (105± 9 µm) that covers the ruminal lumen, attribu-table to an increase in the number of its layers (Figure 1c). Inthe basal zone of the epithelium we witnessed nuclear conden-sation with a visible layer of 2 or 3 cells that formed the stra-tum basale. The presence of projections in the epithelial layer
toward the ruminal lumen form the outline of the pillars thatwill later divide the rumen into its different sacs.
Reticular epithelial layer
The reticular wall displayed a thickness of 370±16 µm. Theepithelial layer (114 ± 8 µm) of the differentiated reticulumwas stratified and two layered with a dark-staining basal zonerich in germinativum cells and another more external apicalzone of globular cells of clear cytoplasm (Figure 1d).
Omasal epithelial layer
At 67 days of embryonic life, omasal differentiation tookplace. The wall of the omasum (364 ± 13µm), smooth untilnow, displayed an undulating surface. The epithelial layer dis-played a thickness of 119 ± 11µm. It was stratified, composedof 5-7 layers, and within it two zones could be distinguished:basal or germinal, with 4-5 layers of dark-staining cells and alarge central nucleus, and another smaller apical zone made upof poorly defined globular cells and a small nucleus (Figure1e). At 67 days of intrauterine life, the epithelium exhibitedundulations of similar height, of 5-6 in number that made upthe primary primitive laminae (Figure 1e). At 90 days of ges-tation (35%) the outline of a new generation of omasal lami-nae, smaller that the first - the secondary laminae - emergedfrom the epithelial enlargements of the interlaminar spaces(Figure 1f).
Group III (8 to 19 cm C-R, 97-135 days, 35-50% of gesta-tion)
Ruminal epithelial layer
The ruminal wall displayed a thickness 460 ± 28 µm Thestratified epithelial layer of rumen continued its growth (250± 20 µm). It displayed two clearly defined zones : a basal zoneor stratum germinativum, formed by 3 to 5 layers of morpho-logically oval cells, with a large central nucleus taking upmost of the cell and a basophilic cytoplasm; and another ex-ternal zone of 10-12 layers of polyhedral cells, with small nu-cleus and light-staining cytoplasm that we will call the stratumgranulosum. The ruminal pillars showed a thin epithelium ger-minativum (Figure 1g).
Reticular epithelial layer
The reticular wall displayed a thickness of 454 ± 25 µm.The epithelial layer (248 ± 21 µm) was made up of a basalzone, or stratum germinativum, formed by 3 or 4 layers ofoval cells, and of basophilic cytoplasm; and an apical layerconsisting of polyhedral cells of clear cytoplasm arranged ina mosaic pattern (Figure 1h).
At 97 days prenatal development an undulation of the epi-thelial germinativum stratum took place, as the first differen-tiation of the future cells of the reticulum: the first primitivereticular ribs (Figure 1h). As intrauterine development pro-gressed, the reticulum showed the ribs of the cells as evagi-nations, becoming more and more prominent, from the basalstratum towards the light zone of the epithelium, at more orless regular intervals (Figure 2a). In their growth, these pri-mary ribs pulled with them the lamina propria, without affec-ting the submucosa.
400 MASOT (A.J.) AND COLLABORATORS
Revue Méd. Vét., 2007, 158, 7, 397-409
PRENATAL DEVELOPMENT OF FORESTOMACH 401
Revue Méd. Vét., 2007, 158, 7, 397-409
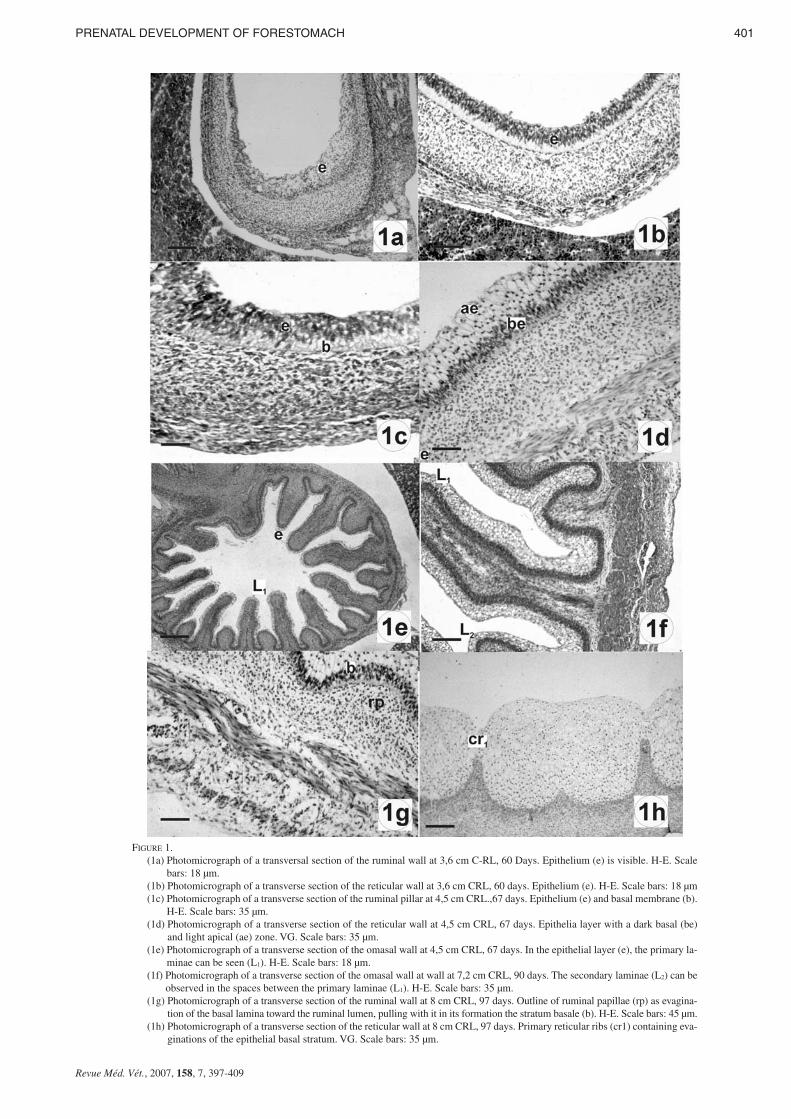
FIGURE 1. (1a) Photomicrograph of a transversal section of the ruminal wall at 3,6 cm C-RL, 60 Days. Epithelium (e) is visible. H-E. Scale
bars: 18 µm. (1b) Photomicrograph of a transverse section of the reticular wall at 3,6 cm CRL, 60 days. Epithelium (e). H-E. Scale bars: 18 µm (1c) Photomicrograph of a transverse section of the ruminal pillar at 4,5 cm CRL.,67 days. Epithelium (e) and basal membrane (b).
H-E. Scale bars: 35 µm.(1d) Photomicrograph of a transverse section of the reticular wall at 4,5 cm CRL, 67 days. Epithelia layer with a dark basal (be)
and light apical (ae) zone. VG. Scale bars: 35 µm. (1e) Photomicrograph of a transverse section of the omasal wall at 4,5 cm CRL, 67 days. In the epithelial layer (e), the primary la-
minae can be seen (L1). H-E. Scale bars: 18 µm.(1f) Photomicrograph of a transverse section of the omasal wall at wall at 7,2 cm CRL, 90 days. The secondary laminae (L2) can be
observed in the spaces between the primary laminae (L1). H-E. Scale bars: 35 µm.(1g) Photomicrograph of a transverse section of the ruminal wall at 8 cm CRL, 97 days. Outline of ruminal papillae (rp) as evagina-
tion of the basal lamina toward the ruminal lumen, pulling with it in its formation the stratum basale (b). H-E. Scale bars: 45 µm.(1h) Photomicrograph of a transverse section of the reticular wall at 8 cm CRL, 97 days. Primary reticular ribs (cr1) containing eva-
ginations of the epithelial basal stratum. VG. Scale bars: 35 µm.

Omasal epithelial layer
The omasal wall displayed a thickness of 429 ± 30µm µmThe epithelial layer of the omasal mucosa (219 ± 18µm) wasstratified with: a basal area of 3-4 layers of germinal cells(stratum germinativum), of a dark cytoplasm and a nucleuswith pyknosis; another apical zone of 6-10 layers of globularcells and a clear cytoplasm that formed the stratum granulo-sum; and externally, the stratum corneum, of anuclear and flatcells. At 97 days of gestation, the outline of a third laminargeneration - the tertiary laminae - appeared between the pri-mary and secondary laminae. Also during this stage, at 135days of intrauterine life, the development of the quaternarylaminae took place, appearing as small elevations of the stra-tum basale of the epithelium among the other laminae, al-though never between the primary and secondary laminae(Figure 2b).
Around mid-gestation, some lateral evaginations of connec-tive tissue towards the epithelial surface were formed fromthe stratum basale of the primary laminae. These were the pri-mitive corneum papillae (Figure 2b).
Group IV (21 to 33 cm C-R, 142-191 days, 45-70% of ges-tation)
Ruminal epithelial layer
The thickness of the ruminal wall was 725 ± 48 µm. In thisstage of development the ruminal epithelium increased consi-derably in thickness with respect to previous stages (430 ± 31µm) and showed greater stratification (Figure 2c). The stra-tum germinativum, in the basal zone, was formed by extre-mely dark stained cells. The stratum granulosum, in the apicalzone, was formed by polyhedral vesiculiform cells of clearcytoplasm and nuclei were polarised toward one of the mem-branes. The appearance of intercellular bridges, directly abovethe stratum basale represented the morphologic expression ofthe lucidum-spinosum stratum. The stratum corneum also ap-peared, with morphologically flat elongated cells in contactwith the lumen. The outline of the ruminal papillae began toappear as evaginations of the basal zone toward the ruminallumen, pulling with it in its formation the stratum basale, thelamina propria and the submucosa (Figure 2d).
Reticular epithelial layer
The thickness of the reticular wall was 729 ± 41 µm. The re-ticular epithelial layer (427 ± 29 µm) (Figure 2e), was finallyformed by the following strata:
- Basal or germinativum formed by a unique layer of dark-staining cells.
- Granulosum, wider and formed by numerous layers oflarge polyhedral cells of a clear cytoplasm.
- Corneum, the most superficial, formed by a unique layerof flat anuclear cells.
The primary reticular ribs continued to move deeper to-wards the epithelial surface from the basal membrane (Figure2e). From the ribs, the corneum papillae emerge as lateralgrowths of a papillary type, coming from the growth and pro-liferation of the basal cells of the stratum germinativum of theepithelial layer (Figure 2e). During this stage, we witnessed
the appearance of secondary reticular ribs, from the basalmembrane, moving towards the epithelial surface: they werealways smaller in size than the primary ones.
Omasal epithelial layer
The thickness of the omasal wall was 744 ± 38µm. The epi-thelial layer of omasum (390 ± 28µm) displayed a greaternumber of layers in all strata (Figure 2f). The globular cells ofthe stratum corneum appeared more elongated. At 191 days ofprenatal development, between the granulosum and corneumstrata, a transition zone of cells of a clear cytoplasm to othersof a lightly eosinophylic colouring could be seen, correspon-ding to the poorly defined lucidum-spinosum stratum. Thecorneum papillae were abundant in the primary as well as inthe secondary laminae (Figure 2g).
Group V (36 to 40 cm C-R, 205-235 days, 75-100% of ges-tation)
Ruminal epithelial layer
The thickness of the walls was 949 ± 52 µm. The thickestepithelial layer of the rumen (550 ± 36 µm) was composed(Figure 2h) of a barely stratified epithelium formed by: stra-tum basale or germinativum of small cells arranged in a row;stratum granulosum; lucidum-spinosum stratum, where thepresence of intercellular bridges and stratum corneum incontact with the ruminal lumen. The growth of the ruminalpapillae was considerable; at 205 days they reached the hi-ghest third of the epithelium, to be positioned close to the epi-thelial surface at 235 days (Figure 2h). Some papillae, onbeing transversely cut, showed a cylindrical form. The papil-lary body appeared delimited by the stratum basale and dis-played a highly cellular connective tissue with a smallquantity of collagen and reticulin that formed the frameworkof the papilla.
Reticular epithelial layer
The thickness of the walls was 953 ± 47 µm. The epithe-lium of the reticulum was keratinized squamous stratified, cor-responding to a tegumentary-type mucosa. It displayed athickness of 556 ± 29 µm and in contrast to the previous stageit showed the differentiation of the lucidum-spinosum stra-tum, made up of condensed cells with dark cytoplasm. Du-ring this stage, the epithelium was lost in the existent spacesamong the primary ribs, resulting in the appearance of inter-nally delimited reticular cells (Figure 3a).
Omasal epithelial layer
The thickness of the walls was 868 ± 29µm. The wall of theomasum, from 205 days of gestation displayed a mucosa withbarely stratified epithelium (525 ± 30µm), where a greaternumber of cells in all of the formative strata could be seen.What stood out was: 1.- the greater differentiation displayedby the lucidum-spinosum stratum; and 2.- the presence of 2-3 layers of flat cells in the corneum stratum. In fetuses at term,we saw numerous longitudinal laminae in the omasal mucosa,distributed in 4 distinct sizes of laminae. The corneum papil-lae increased in number and size, and was seen in all 4 sizesof laminae (Figure 3b).
402 MASOT (A.J.) AND COLLABORATORS
Revue Méd. Vét., 2007, 158, 7, 397-409
PRENATAL DEVELOPMENT OF FORESTOMACH 403
Revue Méd. Vét., 2007, 158, 7, 397-409
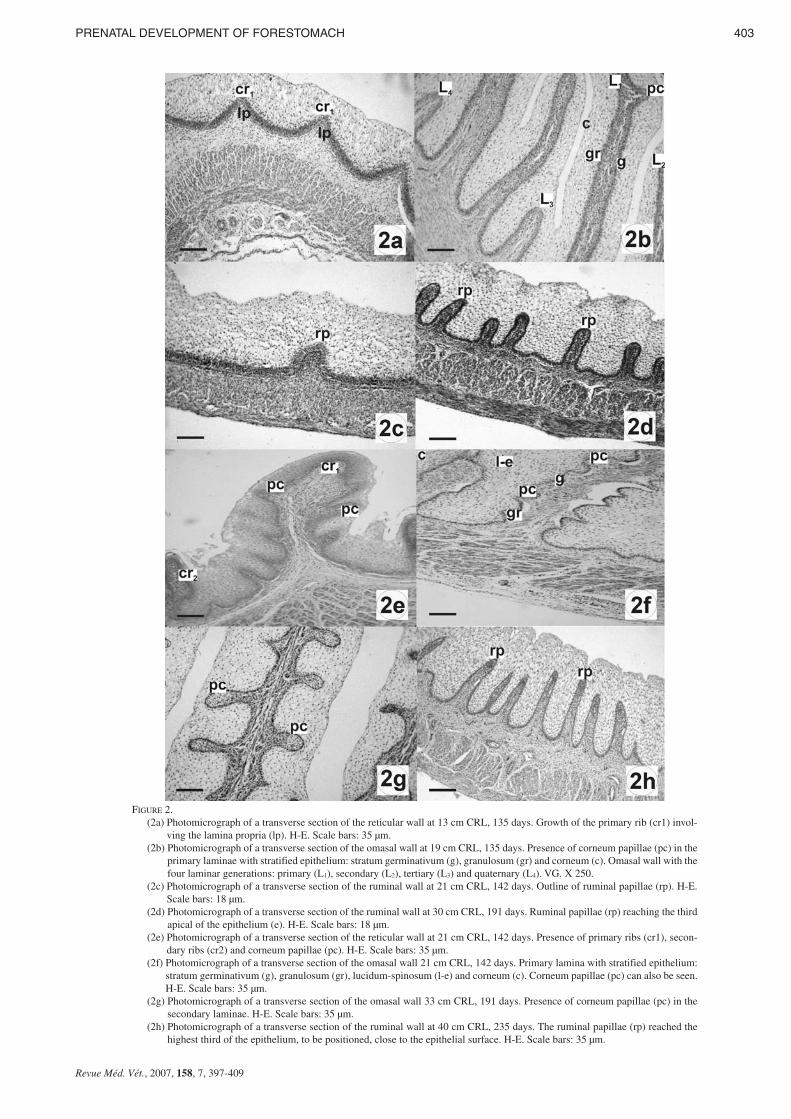
FIGURE 2. (2a) Photomicrograph of a transverse section of the reticular wall at 13 cm CRL, 135 days. Growth of the primary rib (cr1) invol-
ving the lamina propria (lp). H-E. Scale bars: 35 µm.(2b) Photomicrograph of a transverse section of the omasal wall at 19 cm CRL, 135 days. Presence of corneum papillae (pc) in the
primary laminae with stratified epithelium: stratum germinativum (g), granulosum (gr) and corneum (c). Omasal wall with thefour laminar generations: primary (L1), secondary (L2), tertiary (L3) and quaternary (L4). VG. X 250.
(2c) Photomicrograph of a transverse section of the ruminal wall at 21 cm CRL, 142 days. Outline of ruminal papillae (rp). H-E.Scale bars: 18 µm.
(2d) Photomicrograph of a transverse section of the ruminal wall at 30 cm CRL, 191 days. Ruminal papillae (rp) reaching the thirdapical of the epithelium (e). H-E. Scale bars: 18 µm.
(2e) Photomicrograph of a transverse section of the reticular wall at 21 cm CRL, 142 days. Presence of primary ribs (cr1), secon-dary ribs (cr2) and corneum papillae (pc). H-E. Scale bars: 35 µm.
(2f) Photomicrograph of a transverse section of the omasal wall 21 cm CRL, 142 days. Primary lamina with stratified epithelium:stratum germinativum (g), granulosum (gr), lucidum-spinosum (l-e) and corneum (c). Corneum papillae (pc) can also be seen.H-E. Scale bars: 35 µm.
(2g) Photomicrograph of a transverse section of the omasal wall 33 cm CRL, 191 days. Presence of corneum papillae (pc) in thesecondary laminae. H-E. Scale bars: 35 µm.
(2h) Photomicrograph of a transverse section of the ruminal wall at 40 cm CRL, 235 days. The ruminal papillae (rp) reached thehighest third of the epithelium, to be positioned, close to the epithelial surface. H-E. Scale bars: 35 µm.

HISTOCHEMICAL BEHAVIOUR OF THE EPITHELIUM
Ruminal Epithelial layer
Neutral mucopolysaccharides appeared at 67 days and gra-dually decreased throughout the period of prenatal develop-ment until birth. They were found around the tegumentarymucosa in the deeper layers of the epithelium, without affec-ting the stratum corneum. Acid mucopolysaccharides, mucins,and mucoid compounds were not found during development.
Reticular Epithelial layer
The histochemical reaction towards the neutral and acid mu-copolysaccharides mucins and mucoid compounds began at67 days of intrauterine life, and was similar in localization andintensity to that described for the rumen.
Omasal Epithelial layer
The neutral mucopolysaccharides were seen, from 97 daysof gestation, in the germinativum, granulosum and lucidum-spinosum strata, of the tegumentary epithelium of the omasallaminae. The histochemical staining was of medium intensity,decreasing throughout prenatal development to become exi-guous in perinatal stages.
IMMUNOHISTOCHEMICAL OBSERVATION
Ruminal Epithelial layer
Immunohistochemical findings in the epithelium of rumenare summarized in table II and figures 3c,d.
The immunodetection of NNE was positive from 97 daysof gestation in the epithelial layer (Figure 3c), lamina propriaand the submucosa. This rise was progressive until 142 daysof prenatal development, remaining until birth.
The positive immunoreaction for GFAP was observed at142 days of prenatal life in the lamina propria, tunica muscu-laris and serosa and was prolonged until birth. The VIM anti-gen (Figure 3d) was detected in an identical location as that ofGFAP in earlier stages of prenatal development (67 days ges-tation).
Reticular Epithelial layer
Immunohistochemical findings in the epithelium of reticu-lum are summarized in table II and figures 3e,f.
The immunodetection of NNE was positive from 97 daysof gestation in the epithelium (Figure 3e), lamina propria andtunica muscularis and was prolonged until birth.
A positive immunoreaction for GFAP appeared from 142days of foetal development in the lamina propria, tunica mus-cularis and serosa. With regard to VIM, was noted over 67days of gestation in the lamina propria (Figure 3f), tunicamuscularis and serosa; immunopositivity was increased du-ring prenatal development.
Omasal Epithelial layer
Immunohistochemical findings in the epithelium of oma-sum are summarized in table II and figures 3g,h.
The presence of NNE positive cells was observed from 67days of gestation, located in the epithelium, lamina propia-
submucosa (Figure 3g) and in the tunica muscularis. The im-munoreactivity toward these cells increased throughout oma-sal histogenesis, being very high in perinatal stages.
The immunodetection of positive glial cells (GFAP) andVIM, began at 67 days of intrauterine life in the lamina pro-pia-submucosa (Figure 3h), tunica muscularis and serosa. Instages around birth, the immunoreactivity was very intense.
HISTOMORPHOMETRIC OBSERVATIONS
Ruminal Epithelial layer
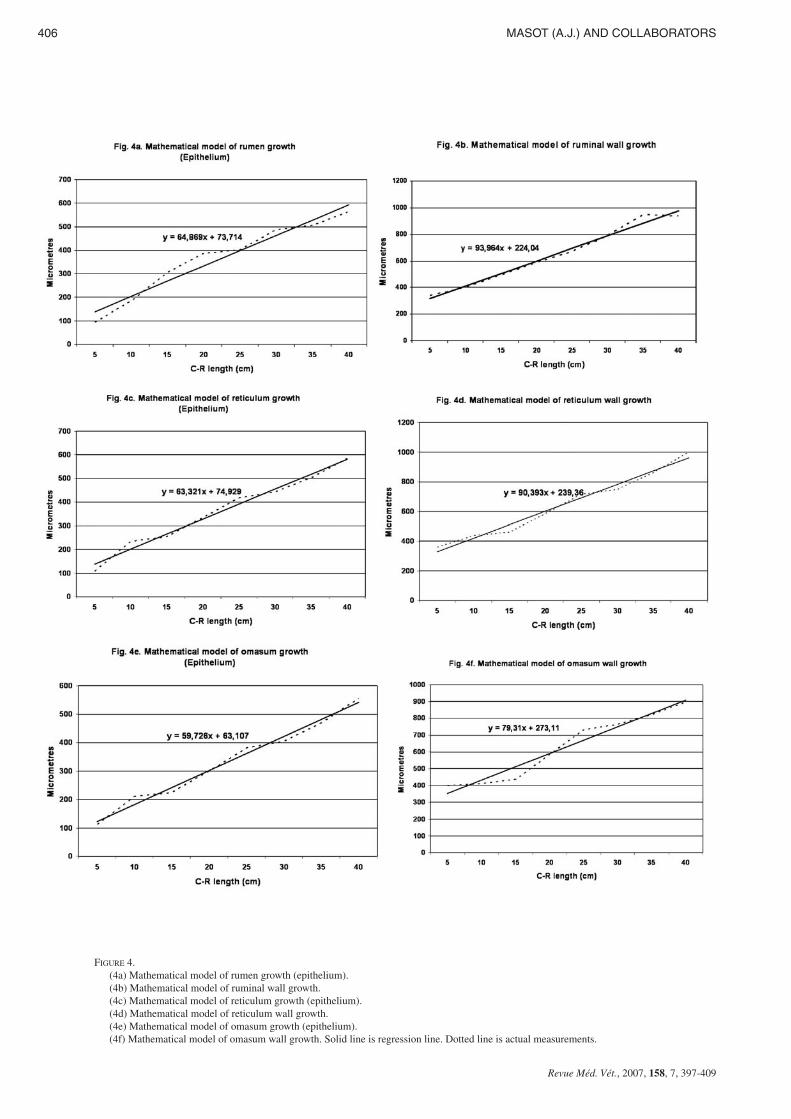
Table I and figures 4a,b shows wall and epithelial layerthickness in the rumen of red deer during prenatal develop-ment.
A factorial ANOVA indicated that the mean value of groupI ruminal epithelium was significantly less than in groups II toV (F=14.68; Tukey test: p ≤ 0.003). On the other way, themean value of wall of group I was significantly greater than ingroups II to V (F=9.50 and Tukey test p ≤ 0.001).
The ruminal epithelial layer experienced rapid growth upuntil 174 days of prenatal development; from 174 until 200days of gestation there is a phase of stabilisation. Subsequentlythe growth of the epithelial layer becomes rapid once againuntil birth. The integral ruminal wall demonstrated gradual andprogressive growth with age and perinatal stabilisation.
Reticular Epithelial layer
In the reticulum (table I, Figures 4c,d), the mean value ofgroup I epithelium was significantly less than in groups II toV (F=16.48; Tukey test: p ≤ 0.002). On the other hand, themean value of wall of group I was also significantly higherthan in groups II to V (F=8.40 and Tukey test p ≤ 0.001).
The morphometric evolution of the reticular epithelial layerand the integral reticular wall (table I, Figures 4c,d), displayedidentical growth phases, stabilisation and regression to thosedescribed in rumen.
Omasal Epithelial layer
The omasal epithelial layer increased progressively in thick-ness throughout histogenesis (table I, Figures 4e,f), in such away that the thickness of the epithelium of Group I was si-gnificantly less than that of groups II, III, IV and V (F=18.54;Tukey test: p ≤ 0.0003).
The thickness of the omasal wall increased gradually throu-ghout prenatal development, until it reached its maximum inperinatal stages (Table I, Figures 4e,f). The thickness of thewall of Group I was significantly less than that of Groups IIto V (F=7.80 and Tukey test p ≤ 0.003).
DiscussionThe appearance of the rumen and the reticulum of the pri-
mitive gastric tube was observed at approximately 3,6 cmCRL, 60 days of prenatal development, 25% of gestation[12,13]. In comparison with other ruminants, prenatal deve-lopment of the deer rumen and reticulum was later than thatin sheep, where it was placed at around 22% of the gestation
404 MASOT (A.J.) AND COLLABORATORS
Revue Méd. Vét., 2007, 158, 7, 397-409
PRENATAL DEVELOPMENT OF FORESTOMACH 405
Revue Méd. Vét., 2007, 158, 7, 397-409
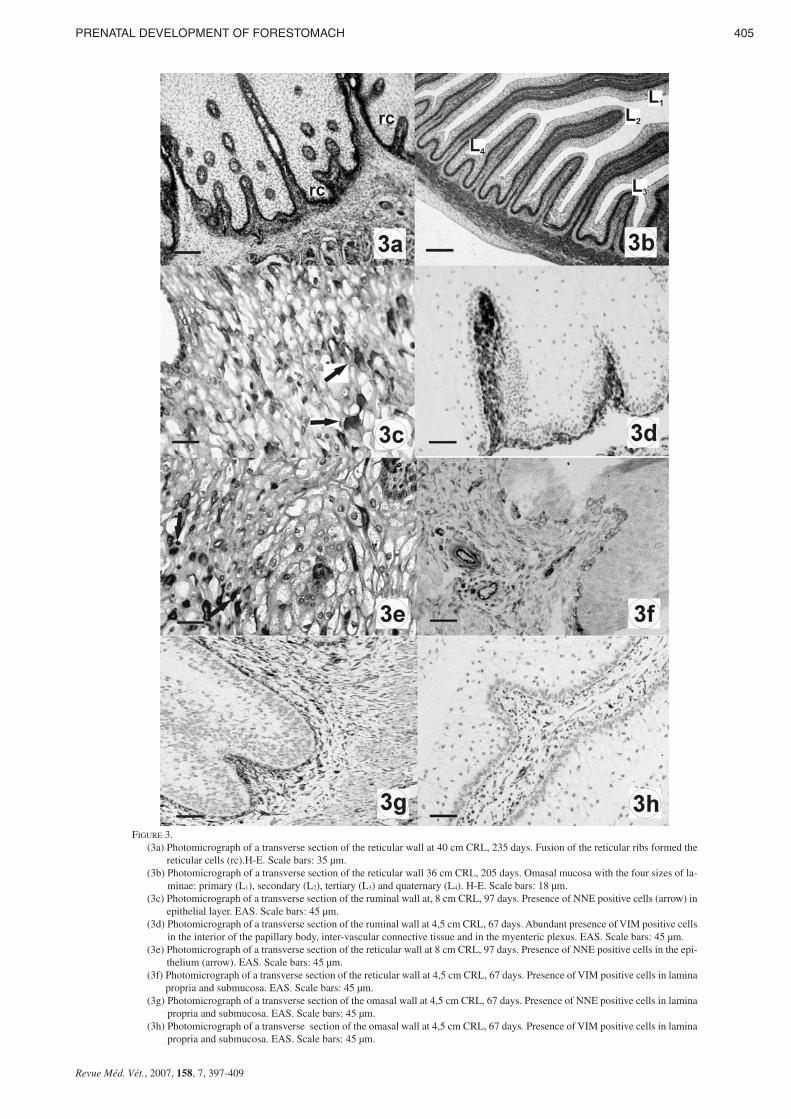
FIGURE 3.(3a) Photomicrograph of a transverse section of the reticular wall at 40 cm CRL, 235 days. Fusion of the reticular ribs formed the
reticular cells (rc).H-E. Scale bars: 35 µm.(3b) Photomicrograph of a transverse section of the reticular wall 36 cm CRL, 205 days. Omasal mucosa with the four sizes of la-
minae: primary (L1), secondary (L2), tertiary (L3) and quaternary (L4). H-E. Scale bars: 18 µm.(3c) Photomicrograph of a transverse section of the ruminal wall at, 8 cm CRL, 97 days. Presence of NNE positive cells (arrow) in
epithelial layer. EAS. Scale bars: 45 µm.(3d) Photomicrograph of a transverse section of the ruminal wall at 4,5 cm CRL, 67 days. Abundant presence of VIM positive cells
in the interior of the papillary body, inter-vascular connective tissue and in the myenteric plexus. EAS. Scale bars: 45 µm.(3e) Photomicrograph of a transverse section of the reticular wall at 8 cm CRL, 97 days. Presence of NNE positive cells in the epi-
thelium (arrow). EAS. Scale bars: 45 µm.(3f) Photomicrograph of a transverse section of the reticular wall at 4,5 cm CRL, 67 days. Presence of VIM positive cells in lamina
propria and submucosa. EAS. Scale bars: 45 µm.(3g) Photomicrograph of a transverse section of the omasal wall at 4,5 cm CRL, 67 days. Presence of NNE positive cells in lamina
propria and submucosa. EAS. Scale bars: 45 µm.(3h) Photomicrograph of a transverse section of the omasal wall at 4,5 cm CRL, 67 days. Presence of VIM positive cells in lamina
propria and submucosa. EAS. Scale bars: 45 µm.
406 MASOT (A.J.) AND COLLABORATORS
Revue Méd. Vét., 2007, 158, 7, 397-409
FIGURE 4.(4a) Mathematical model of rumen growth (epithelium).(4b) Mathematical model of ruminal wall growth.(4c) Mathematical model of reticulum growth (epithelium).(4d) Mathematical model of reticulum wall growth.(4e) Mathematical model of omasum growth (epithelium).(4f) Mathematical model of omasum wall growth. Solid line is regression line. Dotted line is actual measurements.

period [3,4,6,15]; and in goat [10], at around 24% of gesta-tion; although authors [16] established a period of develop-ment later still, and placed it at about 97 days (65% ofgestation). In the cow it’s reported it very early at 1.7 cm CRL,equivalent to a 30 days or 11% gestation period [2].
The differentiation of the omasum, took place, later than inthe rumen and reticulum [12,13], at 4,5 cm CRL, 67 days(25% of gestation). In sheep the omasal differentiation wascalculated at 33 days of embryonic life (22% of gestation). Inthe organogenesis of goat [17] the primordial omasum appearsat 28 days (19% of gestation); and in cow [2,18] at 30 days(11% of gestation).
At 4,5 cm CRL, 67 days (25% of gestation) the rumen, re-ticulum and omasum, were formed by 3 layers: an internal ormucosal, a middle or muscular and an external or serosallayer. In terms of the mucosa, the transition took place froman epithelium of an “embryonic type” [7] toward a stratifiedepithelium with two zones: a basal zone or stratum germina-tivum, and another smaller apical zone or stratum granulo-sum. This bizonal stratification was previously reported inprenatal development in the rumen of buffalo [9], in therumen, reticulum and omasum of sheep [4-6] and in therumen, reticulum and omasum of red deer [12-14].
The stratum corneum of the epithelial layer of the rumenand reticulum of red deer, appeared at about 142 days gesta-tion (50%). There is no general agreement on this matter withregard to previous research. Thus, although we agree withFRANCO et al. [4-6] who placed it in sheep at 83 days (55%of gestastion), in this same species, it’s described at birth [19]and at 57 days (38% of gestation, [15]); in the goat [10] at 104days (70% of gestation); in buffalo [8,20] at 102 days (32% ofgestation) and in the cow [2,7,21] it is described in the deve-lopmental stages close to birth. At 97 days of prenatal deve-lopment (35% of gestation) the corneum stratum appeared inthe omasum. There was agreement that the corneum stratumappeared earlier in the omasal epithelial layer of red deer thanit did in the rumen and reticulum [12,13]. Our observationspresented us with a stratified omasal mucosa from 97 days ofgestation until birth, a fact that has also been observed in pe-rinatal sheep [22] and in goat [10]. A light keratinization wasalso evident, clearly in agreement with that described in theomasal mucosa of goat [23].
In red deer, the progressive stratification of the omasal epi-thelial layer throughout ontogenesis was accompanied by anincrease in the thickness of this layer, until it reached its maxi-mum in the perinatal stages. This finding was similar to des-cribed in cattle [24]. However, in the goat [25] its thicknessremained unchanging throughout prenatal development. Insheep [26], showed a reduction in the perinatal stages.
The stratification of the epithelial layer of the non-glandu-lar stomach compartments, was completed with the appea-rance of the lucidum-spinosum stratum at 142 days (50% ofgestation) in the rumen and omasum of red deer [12,14], andat 205 days gestation (75%) in the reticulum [13]. The ap-pearance of the lucidum-spinosum stratum has only been des-cribed in the forestomach of the buffalo in advanced stages ofdevelopment [27-29]; and in sheep during perinatal stages[22].
The stratification of the epithelial layer of the forestomachmucosa was accompanied by modifications in its structures,with the appearance of the ruminal pillars and papillae in therumen; of the primitive reticular cells (and primary ribs) inthe reticulum; and of the primary laminae in the omasum.
In the rumen the presence of papilliform projections towardthe lumen that formed the outline of the pillars was timed ataround 67 days (25% of gestation). Similar results was des-cribed in sheep [4,15] and cow [2], at 39 (26% of gestation),42 (28% of gestation) and 44 days (16% of gestation), res-pectively. All parietal layers took part in its formation, al-though the serosa to a lesser extent. The outline of the ruminalpapillae started to appear at 142 days, as evaginations of thebasal zone toward the ruminal lumen, pulling with it the stra-tum basale, the lamina propria and the submucosa [12]. Opi-nions differ with regard to the timing of its appearance; thusin sheep [4,15,16] put it at 64 (43% of gestation), 103 (69%of gestation) and 61 days (41% of gestation). In the goat [10]at around 136 days (91% of gestation); in the cow [2,30,31] instages close to birth and in buffalo [27] at 74 days of prenataldevelopment (23% of gestation).
The final structure of the reticular epithelial layer resultedin the formation of the primitive reticular cells. We observedthem in their first differentiation as small evaginations of thebasal zone toward the apical zone of the epithelium (primaryribs) at 117 days of prenatal life (40% gestation). In sheep[6,15,16] it’s described at 95 days (63% gestation), 72 (48%gestation) and 69 (46% gestation); In goat [10], between 69and 102 days (46-68% gestation) and in buffalo [9,28] at 70and 200 days (22% and 62% gestation).
At 142 days (50% gestation) some lateral growths that for-med the corneum papillae appeared from the primary reticu-lar ribs. The differentiation of these structures in sheep wasreported at around 55% gestation [6], at 63% gestation [16]and in perinatal stages [10].
The secondary reticular ribs appeared at 142 days, at ap-proximately 50% gestation, as a growth of the primary ribs, al-though they never reached the same size [3,6,13]. Just as withthe primary ribs, there was no unanimity in terms of their his-todifferentiation. Thus, they were situated at approximately40% gestation in buffalo [9]; in the range 55-70% in sheep[6,15,16] and at 69% in the goat [10].
The characteristic cells of the reticulum require two types ofrib growth for their formation: longitudinal, from the basalmembrane and toward the epithelial surface (responsible forthe growth in length of the ribs), and transversal or an expan-sion of the ribs toward the adjacent ribs, joining with them toform the cells. The uneven height of the both primary and se-condary reticular ribs could explain the formation of cells ofvarying size [6,13].
The appearance of primary laminae in the omasum of reddeer (67 days, 25% of gestation) was later than that describedfor the omasum of sheep, where it was placed at around 30-35 days, 20-22% of gestation [15,26].
The temporal order of appearance of the 4 sizes of laminaeof the omasum of red deer was: primary at 67 days (25% ofgestation), secondary at 90 days (30%), tertiary at 97 days
PRENATAL DEVELOPMENT OF FORESTOMACH 407
Revue Méd. Vét., 2007, 158, 7, 397-409

(35%) and quaternary lamina at 135 (50% of gestation). Insheep, there was a significant discrepancy in terms of the ti-ming of the appearance of the laminae. Thus, FRANCO et al.[5] described the following sequence: at 21%, at 26%, at 33%and at 40% of gestation respectively. DEL RIO ORTEGA [15]place it at 24%, at 28%, at 32% and at 33% of gestation res-pectively. LUBIS & O’SHEA [26] at 21%, at 25%, at 30%and at 40%, and FAT EL-BAB et al. [16] at 21%, at 26%, at33% and at 52%. In cattle [2,18] and in buffalo [28] the pro-cess was earlier, the presence of the 4 sizes of laminae beingdescribed at around 20% of gestation.
The majority of the authors described the laminae of fourdifferent sizes in the omasal mucosa: in sheep at around 33%[15], 40% [5,26] or 52% of gestation [16]; in the goat [25] at33%; in cattle [24] at about 33% and in red deer [14] at 50%of gestation. However, there have been exceptions such as thatnoted in sheep [19], in goat [25] and in buffalo [28], with theappearance of a fifth laminar size in perinatal stages. Therewas unanimity, however, that the appearance of the laminaroutlines was a sequential process, since the very growth of alaminar generation involved the birth of the next generationuntil the appearance of the 4 sizes of laminae. In this process,all architectural components were involved: mucosa, laminapropia-submucosa and tunica muscularis. The exception wasthe serosa, which had a surrounding function, a fact previouslynoted in the prenatal development of sheep [5] and red deer[14].
At around mid-gestation (135 days), lateral evaginations ofthe connective tissue were formed from the basal stratum ofthe primary and secondary laminae towards the epithelial sur-face. These were the primitive corneum papillae. From 205days of prenatal life (75% of gestation) the corneum papillaecould be observed in the 4 different sizes of laminae. In theprenatal development of sheep [5], these corneum papillaewere described in the stratum basale of the primary laminae at69 days (45% of gestation). At 79 days (53% of gestation)they started to become apparent in the second laminar gene-ration, but it was not until 75% of gestation (133 days) thatthey appeared in the two remaining generations. A significantdiscrepancy prevailed in fixing the timing of the appearanceof the corneum papillae, given that in this species: at 64 days,42% of gestation [15]; at 69 days, 46% [26] and at 123 days,82% [16]. In the goat [25] the development of the corneumpapillae was referenced at 67 days (45% of gestation) and incattle [24] at 90 days (32% of gestation). The arrangement ofthe corneum papillae is in agreement with the histophysio-logy. It is controlled in such a way that the movement of thefolds act on the alimentation and introduce it towards the in-terlaminar spaces [5,14].
Results indicate that the tegumentary mucosa of the non-glandular stomach compartments of red deer lacked secretorycapacity during initial embryonic stages; from 67, and 97days,neutral mucopolysaccharides were detected in therumen-reticulum, and omasum, respectively. In sheep [4] weredescribed at 89 days (60% of gestation). Mechanical protec-tion of the first products of embryonic metabolism against ag-gressions is a function located in the stratum corneum of thetegumentary mucosa. On the other hand, in the same way asFRANCO et al. [4] in sheep pointed out, in deer there existed
a gradual relation between neutral mucopolysaccharidecontent in the deeper epithelial layers and the gradual adapta-tion of the mucosa of the non-glandular stomach compart-ments, in its function as chemical protector in postnatal life,in acting as a buffer system in the neutralisation of the acidcompounds produced during ruminal fermentation [12] andfrom the acid compounds incorporated in the amniotic fluid,given that the ingestion of this is a normal process during ges-tation [13,14].
The presence of NNE positive cells in the mucosa of thenon-glandular stomach compartments of deer was not detec-ted until 67 days of prenatal development in the omasum, and97 days in the rumen and reticulum. These cells were locatedin the epithelial layer, lamina propria and submucosa, and tu-nica muscularis. There are not references about the presenceof NNE positive cells in these structures. Similar opinionswere reported in the non-glandular stomach compartments ofred deer [12-14]. Several authors described the presence ofNSE positive cells in rumen, reticulum and omasun of cattle[32,33,34]; in the forestomach of the sheep [35].
The glial cells were detected at 142 days for GFAP, in therumen and reticulum; and at 67 days in the omasum. The VIMwere detected at 67 days in rumen, reticulum and omasum,demonstrating that VIM is a marker of more primitive glialcells [12,13,14]. The localization of the glial cells in the fo-restomach mucosa of deer in development involved the laminapropria, submucosa, tunica muscularis and serosa. In perina-tal sheep [36] and in bovine reticulum [37] immunoreactivityfor glial cells described in the reticular papillae, sugges thatsome of the functions of the reticular mucosa could be intrin-sically regulated by the submucosal plexus.
From the observations of our study, in terms of the prenatalstructure of the forestomach mucosa, we can deduce that thered deer is less precocious than small and large domestic ru-minants. Thus, its secretory capacity, detected by the presenceof neutral mucopolysaccharides were evident in more advan-ced stages of prenatal development than that detected insheep, goat and cow.
AcknowledgmentsThis study was supported by a research grant from Conse-
jería de Infraestructura y Desarrollo Tecnológico of Junta deExtremadura (Project No. 3PR05B028). The authors wouldlike to thank Mrs. Mª Mercedes Carrasco Toral and Mrs. JuanaOllero Granados (Hospital San Pedro de Alcántara, ServicioExtremeño de Salud) and Mr. Juan L. Rodríguez Cruz (Fa-culty of Veterinary Medicine, University of Extremadura) fortheir invaluable technical assistance and contributions.
References1. - CARRANZA J: Aplicaciones de la etología al manejo de las pobla-
ciones de ciervo en el suroeste de la Península Ibérica: producción yconservación. Etología, 1999, 7, 5-18.
2. - VIVO J.M., ROBINA A., REGODÓN S., GUILLEN M.T., FRANCOA., MAYORAL A.I: Histogenetic evolution of bovine gastric com-partments during prenatal period. Histol. Histopathol., 1990, 5, 461-476.
408 MASOT (A.J.) AND COLLABORATORS
Revue Méd. Vét., 2007, 158, 7, 397-409

3. - FRANCO A., VIVO J.M., GUILLEN M.T., REGODÓN S., ROBINAA: Evolución parietal del retículo ovino de raza merina desde los 68días de gestación hasta el nacimiento. Histol. Med., 1989, 5, 57-58.
4. - FRANCO A., REGODÓN S., ROBINA A., REDONDO E.: Histo-morphometric analysis of the rumen of the sheep during development.Am. J. Vet. Res., 1992, 53, 1209-1217.
5. - FRANCO A., (1993b) ROBINA A., REGODÓN S., VIVO J.M.,MASOT A.J., Redondo E: Histomorphometric analysis of the oma-sum of sheep during development. Am. J. Vet. Res., 1993a, 54, 1221-1229.
6. - FRANCO A., ROBINA A., REGODÓN S., VIVO J.M., MASOT A.J.,REDONDO E: Histomorphometric analysis of the reticulum of thesheep during development. Histol. Histopathol., 1993c, 8, 547-556.
7. - WARNER E.D: The organogenesis and early histogenesis of the bo-vine stomach. Am. J. Anat., 1958, 102, 33-63.
8. - PANCHAMUKHI B.G., SRIVASTAVA H.C: Histogenesis of therumen of the buffalo (Bubalus bubalis) stomach. Anatomia HistologiaEmbryologia, 1979, 8, 97-105.
9. - PANCHAMUKHI B.G., SRIVASTAVA H.C: Histogenesis of the re-ticulum of the buffalo (Bubalus bubalis) stomach. Indian J. AnimalSci., 1980, 50, 1064-1070.
10. - MOLINARI E., JORQUERA B: Intrauterine development stages ofthe gastric compartments of the goat (Capra hircus). Anat. Histol.Embryol., 1988, 17, 121-137.
11. - FRANCO A., ROBINA A., GUILLEN M.T., MAYORAL A.I., RE-DONDO E: Histomorphometric analysis of the abomasum of sheepduring development. Anat. Anz., 1993b, 175, 119-125.
12. - FRANCO A., MASOT A.J., GÓMEZ L., REDONDO E: Morpho-metric and immunohistochemical study of the rumen of red deer du-ring prenatal development. J. Anat., 2004a, 204, 501-513.
13. - FRANCO A., REDONDO E., MASOT A.J: Morphometric and im-munohistochemical study of the reticulum of red deer during prena-tal development. J. Anat., 2004b, 205, 277-289
14. - REDONDO E., FRANCO A., MASOT A.J: Morphometric and im-munohistochemical study of the omasum of red deer during prenataldevelopment. J. Anat., 2005, 206, 543-555.
15. - DEL RIO ORTEGA S: Desarrollo prenatal del estómago de la oveja.Doctoral Thesis. Facultad de Veterinaria. Zaragoza. 1973
16. - FAT-EL BAB M.R., SCHWART R., ALI A: Micromorphological stu-dies on the stomach of sheep during prenatal life. Anat. Histol. Em-bryol., 1983, 12, 139-153.
17. - MUTOH K., WAKURI H: Early organogenesis of the caprine sto-mach. Nippon Juigaku Zasshi, 1989, 51, 474-84.
18. - VIVO J.M., ROBINA A: The development of the bovine stomach:morphologic and morphometric analysis. II. Observations of the mor-phogenesis associated with the omasum and abomasum. Anat HistolEmbryol., 1991, 20, 10-17.
19. - WARDROP J.D: Some preliminary observations on the histologicaldevelopment on the forestomach of the lamb. I. Histological changesdue to age in the period from 46 days of foetal life to 77 days of pos-natal life. J. Agric. Sci., 1961, 57, 335-341.
20. - TIWARI G.P., JAMDAR N.M: Studies of the gross and histologicalstructure and development on the forestomach of indian water buffalocalf in early postnatal life with reference to normal feeding. 2. Reti-culum. Indian J. Anim. Sci., 1970, 57, 335-340.
21. - ASARI M., FUKAYA K., KANO Y., EGUICHI Y: Development ofthe fetus and neonatal bovine stomach . Anat. Histol. Embryol., 1981,10, 264-274.
22. - GROENEWALD H.B: Ultrastructure of the epithelium of the rumen,reticulum and omasum of grey, white and black karakul lambs. On-derstepoort J. Vet. Res., 1993, 60, 197-204.
23. - YAMAMOTO Y., ATOJI Y., AGUNGPRIYONO S., SUZUKI Y:Morphological study of the forestomach of the japanese serow (Ca-pricornis crispus). Anat. Histol. Embryol., 1998, 27, 73-81.
24. - TOTZAUER I., SINOWATZ F: Fetal development of the omasum ofcattle (Bos taurus). Tierarztl. Prax., 1990, 18, 577-83.
25. - RAMKRISHNA V., TIWARI G.P: Histological and histochemicalobservations on the forestomach of goat during pre-natal life. ActaAnat (Basel), 1979, 103, 292-300.
26. - LUBIS D., O'SHEA J.D: Development of the omasum in sheep. ActaAnat., 1978, 100, 400-410.
27. - OSMAN A.H.R., BERG R: Studies on the histogenesis of the tunicamucosa of the stomach of the egyptian water buffalo (Bos. bubbalis)1. Histogenesis of the ruminal mucosa. Anatomical Anzeiger, 1981a,149, 232-240.
28. - OSMAN A.H.R., BERG R: Studies on the histogenesis of the tunicamucosa of the stomach of the egyptian water buffalo (Bos. bubbalis)2. Histogenesis of the reticular mucosa. Anat. Anzeiger, 1981b, 150,516-520.
29. - OSMAN A.H.R., BERG R: Studies on the histogenesis of the tunicamucosa of the stomach of the egyptian water buffalo (Bos. bubbalis)4. Histogenesis of the omasal mucosa. Anat. Anz., 1982, 151, 467-471.
30. - ARIAS J.l., CABRERA R., VALENCIA A: Observations on the his-tological development of the rumen papillae of cow. Morphologicalchanges due to age. Anat. Histol. Embryol. 7, 140-151.
31. - AMASAKI H., DAIGO M: Prenatal development of subepitelial vas-culature related to appearance of ruminal papillae in the cow rumen.Anat. Anzeiger, 1987, 164, 139-147.
32. - KITAMURA N., YAMADA J., YAMASHITA T: Immunohistoche-mical study on the distribution of neuron-specific enolase - and pep-tide-containing nerves in the reticulorumen and the reticular grooveof cattle. J. Comp. Neurol., 1986, 248, 223-234.
33. - KITAMURA N., YAMADA J., YAMASHITA T: Immunohistoche-mical study on the distribution of neuron-specific enolase - and pep-tide-containing nerves in the omasum of cattle. J. Comp. Neurol.,1987, 256, 590-599.
34. - KITAMURA N., YAMADA J., YAMAMOTO Y., YAMASHITA T:Subsatance P-immunoreactive neurons of the bovine forestomachmucosa: their presumptive role in a sensory mechanism. Arch. Histol.Cytol., 1993, 56, 399-410.
35. - GROENEWALD H.B: Neuropeptides in the myenteric ganglia andnerve fibres of the forestomach and abomasum of grey, white andblack karakul lambs. Onderstepoort J. Vet. Res., 1994, 61, 207-213.
36. - YAMAMOTO Y., ATOJI Y., SUZUKI Y: Morphological study of thesubmucosal and mucosal plexuses of the sheep forestomach. Anat.Anz., 1995, 177, 405-412.
37. - TEIXEIRA A.F., WEDEL T., KRAMER H.J., KUHNEL W: Struc-tural differences of the enteric nervous system in the cattle foresto-mach revealed by whole mount immunohistochemistry. Anat. Anz.,1998, 180, 393-400.
PRENATAL DEVELOPMENT OF FORESTOMACH 409
Revue Méd. Vét., 2007, 158, 7, 397-409





























