Embed Size (px)
Citation preview

Rev. Écol. (Terre Vie), vol. 65, 2010.
– 139 –
COMPOSITION ET VARIATIONS DU PEUPLEMENT ICHTYOLOGIQUE DE L’HERBIER SUPERFICIEL À POSIDONIA OCEANICA (L.) DELILE,
DANS LA BAIE D’ANNABA (ALGÉRIE)
F. Derbal1,*2& M. H. Kara1
SUMMARY. — Composition and variations of the fish assemblage of Posidonia oceanica (L.) Delile, seagrass meadows in Annaba bay (Algeria). — During an annual cycle (July 2004 - June 2005), two coastal sites, one far away from anthropic action (site A : Aïn Achir) and the other subjected to immediate discharge of waste water (site B : la Caroube), were sampled during the day (8 h -18 h) and by night (19 h - 7 h), with a net (50 m long, 2 m high) located at a depth of -5 m. According to the frequency of fish species captured, their ecological status was determined. The Shannon-Weaver diversity index (H’) was calculated to better understand the structure and organization of fish assemblages in both sites. In site A, 19 species belonging to 8 families were fished during the day. At night, the fish assemblage was represented by 15 species and 9 families. The family Sparidae was the most diversified, followed by Labridae and Serranidae. Labridae were the most numerous family by day and by night. Numbers of species and individuals collected in site B during the day (16) as well as by night (11) were significantly lower than those of site A. During daytime, diversity (H’) and equitability (R) indices were respectively 2.51 and 0.33 in site A and of 2.87 and 0.43 in site B. At night, diversity index was lower (site A : 0.64 and site B : 2.55) and equitability was close to 0.41 in the two sites.
rÉSUMÉ. — Au cours d’un cycle annuel (juillet 2004 - juin 2005), deux sites côtiers, l’un éloigné de l’action anthropique (site A : Aïn-Achir) et l’autre recevant les rejets directs des eaux usées de la ville d’An-naba (site B : la Caroube), ont été échantillonnés de jour et de nuit à l’aide d’un filet trémail. En fonction de sa fréquence, le statut écologique de chaque espèce a été déterminé. L’indice de diversité biologique de Shannon-Weaver (H’) a été calculé pour mieux comprendre la structure et l’organisation du peuplement de poissons dans les deux sites. Dans le site A, 19 espèces appartenant à 8 familles ont été pêchées de jour. La nuit, le peuplement ichtyologique est représenté par 15 espèces et 9 familles. La famille des Sparidés est la plus diversifiée, suivie de celle des Labridés et des Serranidés. Les Labridés sont les mieux représentés numériquement, aussi bien de jour que de nuit. Dans le site B, les nombres d’espèces et d’individus capturés, aussi bien de jour (16 espèces) que de nuit (11 espèces), sont nettement inférieurs à ceux obtenus dans le site A. De jour, les indices de diversité (H’) et d’équitabilité (R) sont respectivement de 2,51 et de 0,33 dans le site A et de 2,87 et de 0,43 dans le site B. La nuit, l’indice de diversité est plus faible (site A : 0,64 ; site B : 2,55) et l’équitabilité est proche de 0,41 dans les deux sites.
Les herbiers à Magnoliophytes marines constituent l’un des écosystèmes les plus remar-quables des zones littorales, tant en milieu tropical qu’en milieu tempéré. Ainsi, leur protection et leur conservation sont devenues une priorité internationale, comme c’est le cas pour Posi-donia oceanica en Méditerranée (Anonyme, 1999 ; Boudouresque et al., 2006). Les herbiers à P. oceanica ont un rôle écologique de tout premier ordre et constituent de ce fait un pôle de biodiversité unique (Boudouresque, 2004). Leur grande production de biomasse est à la base
1 Laboratoire Bioressources Marines, Université Badji-Mokhtar, Annaba, Algérie.* Auteur correspondant. E-mail : [email protected]

– 140 –
de nombreux réseaux trophiques (Pergent-Martini et al., 1994 ; Havelange et al., 1997) et leurs formations constituent des sites de fraie, de recrutement, d’abri et de nurserie pour de nom-breuses espèces macrozoobenthiques (Nakaoka, 2005).
En Méditerranée nord-occidentale, les travaux sur les peuplements ichtyologiques asso-ciés aux petits fonds mixtes (rochers-herbiers) ou à herbiers de phanérogames sont assez nom-breux, aussi bien en zone ouverte non protégée (Bell & Harmelin-Vivien, 1982 ; Guidetti & Bussoti, 2000 ; Valle et al., 2001 ; Frau et al., 2003 ; Letourneur et al., 2003 ; Moranta et al., 2006 ; Deudero et al., 2008) qu’au sein des réserves marines (Harmelin-Vivien, 1982, 1984 ; Francour, 1989, 2000 ; MacPherson et al., 2002). Divers aspects ont été étudiés : la structure des peuplements ichtyologiques (Bell & Harmelin-Vivien, 1982 ; Harmelin-Vivien, 1984 ; Valle et al., 2001) et ses variations spatiales et temporelles (Harmelin-Vivien, 1982 ; Moranta et al., 2006 ; Deudero et al., 2008), les niches écologiques et les habitats préférentiels spéci-fiques (Frau et al., 2003 ; Clynick, 2006), l’effet de la pêche (Sanchez-Jerez & Ramos Espla, 1996 ; Harmelin-Vivien, 2000) et de la dégradation physique de l’herbier sur les peuplements ichtyologiques (Fernandez et al., 2005).
Sur les côtes algériennes, les travaux dans ce domaine ont concerné la composition et la structure du peuplement macrozoobenthique invertébré inféodé à la posidonie (Semroud, 1993 ; Mezali et al., 2006). Dans la région d’Annaba, P. oceanica occupe de vastes étendues, dont certaines sont sérieusement menacées de dégradation (pollution domestique et industrielle). Cependant, les données sur ces herbiers et sur la faune qui leur est associée sont ponctuelles et sommaires (Pergent et al., 1993 ; Derbal & Kara, 2005 ; Abdelli-Menail, 2007), en particulier celles concernant les peuplements ichtyologiques (Derbal & Kara, 2001 ; Derbal et al., 2005).
Le travail présenté ici a pour objectif la caractérisation du peuplement ichyologique de l’herbier superficiel à P. oceanica de la baie d’Annaba en fonction des influences anthropiques. En effet, l’extension des activités touristiques et le développement urbain le long du littoral pourraient constituer, à moyen et à long termes, une menace sur les habitats et les biocénoses marines. Dans ce contexte, la connaissance et le suivi de l’évolution spatio-temporelle du peuplement ichtyologique côtier, pourraient constituer un élément révélateur de son niveau de résilience vis-à-vis des perturbations environnementales littorales (Francour, 2000).
MATÉRIEL ET MÉTHODES
ZONE D’ÉTUDE ET LOCALISATION DES SITES D’ÉCHANTILLONNAGE
L’étude du peuplement ichtyologique de l’herbier à P. oceanica a été réalisée à l’ouest de la baie d’Annaba, dans une zone protégée des vents dominants du nord-ouest. Deux sites côtiers ont été échantillonnés ; le premier est éloigné de l’action anthropique (site A : Aïn-Achir) et le second se trouve à proximité de rejets domestiques temporaires en période hivernale (site B : la Caroube) (Fig. 1). Les deux sites sont soumis à la pêche artisanale et récréative.
L’herbier de « Aïn-Achir » est clairsemé et repose, à -5 m, sur un fond rocheux entrecoupé de zones de sable grossier jusqu’à -10 m de profondeur. Ce secteur, de mode légèrement agité à calme, est éloigné des apports terrigènes et des rejets urbains. La visibilité des eaux superficielles est moyenne (3 - 5 m) en période hivernale et automnale, et bonne ( > 6 m) en période printanière et estivale. Les courants locaux sont de moyenne amplitude (courant moyen). Les températures extrêmes mesurées en surface varient entre 13° C (février 2005) et 27° C (août 2004).
L’herbier de la plage « la Caroube » est également clairsemé et longe presque la totalité de la plage. Il repose, à moins de 3 m, sur un fond meuble constitué essentiellement de sable fin à moyen, mélangé à de la vase terrigène. Quelques zones rocheuses, recouvertes d’algues Codium tomentosum, se trouvent dans l’herbier.
Contrairement au site A, le site B est une plage qui reçoit des rejets domestiques (détergents, eaux ménagères). Il est totalement abrité des vents dominants et est soumis à une action hydrodynamique de très faible amplitude (0,5 nœud). La visibilité y est généralement faible à médiocre en automne et en hiver (≤ 2 m) et s’améliore progressivement au printemps et en été ( > 3 m). Les variations de la température sont similaires à celles de la zone A.
ÉCHANTILLONNAGE
Entre juillet 2004 et juin 2005, nous avons noté la température, la visibilité (1 ou bonne : ≥ 6 m ; 2 ou moyenne : 3 - 5 m ; 3 ou médiocre : ≤ 2 m), la force du courant (1 : feuilles de posidonie immobiles = absence de courant ; 2 : inclinaison des feuilles de 45° = courant de faible amplitude ; 3 : inclinaison des feuilles de 90° = courant fort) et l’état de la mer (1 : mer calme ; 2 : mer ridée ; 3 : mer légèrement agitée ; 4 : mer houleuse ; 5 : mer très agitée). Les données physiques ont été recueillies à l’occasion de chaque pêche.

– 141 –
Figure 1. — Zone d’étude et localisation des sites d’échantillonnage (*).
L’échantillonnage a été effectué, de jour comme de nuit, en utilisant un filet trémail (longueur = 50 m, hauteur = 2 m, maille = 40 mm). Cet engin de pêche est le moins sélectif des filets maillants puisqu’il peut capturer aussi bien des petits poissons que des individus de grande taille.
En raison des fluctuations de la richesse spécifique et de l’abondance des individus en fonction du temps, du nycthémère, des facteurs environnementaux et des saisons (Harmelin-Vivien, 1982, 1984), nous avons procédé à un échantillonnage fréquent, à raison d’une pêche par semaine, en alternant les sites.
ANALYSE DES CAPTURES
Afin de comprendre la structure du peuplement ichtyologique en place et son degré d’organisation, les captures ont été analysées dans chaque site et différents indices écologiques ont été déterminés :
— la richesse spécifique (S) : nombre d’espèces pêchées dans chaque site ;— l’abondance spécifique (A) : nombre d’individus pêchés pour une espèce i ;— la fréquence d’une espèce : rapport exprimé en pourcentage du nombre de prélèvements dans lesquels se trouve
l’espèce i sur le nombre total des prélèvements. Cet indice permet de déterminer le statut écologique ou la classe de la biocénose par rapport à son biotope. Trois classes ont été définies en respectant la méthode de Bouchon-Navaro et al. (1992) (classe résidente : F ≥ 75 % ; classe temporaire : 25 % < F < 75 % ; classe occasionnelle : F ≤ 25 %) ;
— l’indice de diversité de Shannon (H’) (Shannon & Weaver, 1949), utilisé en écologie comme mesure de la diversité spécifique :
avec : S : nombre total d’espèces présentes ; ni : effectif de l’espèce i dans l’échantillon ; N : effectif total. H’ varie entre 0, dans le cas où le peuplement n’est constitué que d’une seule espèce et log2 S dans le cas où toutes les espèces présentes le sont avec une abondance équivalente (H’ = 4,5 ou 5 pour les peuplements les plus diversifiés) ;
— l’indice d’équitabilité ou de régularité R (Pielou, 1966) permet de mesurer l’équirépartition des espèces du peuplement par rapport à une répartition théorique égale pour l’ensemble des espèces (Barbault, 1992) ;
avec : Hmax = log2S.La valeur de R varie entre 0 (une seule espèce domine) et 1 (toutes les espèces ont la même abondance).
– 142 –
RÉSULTATS
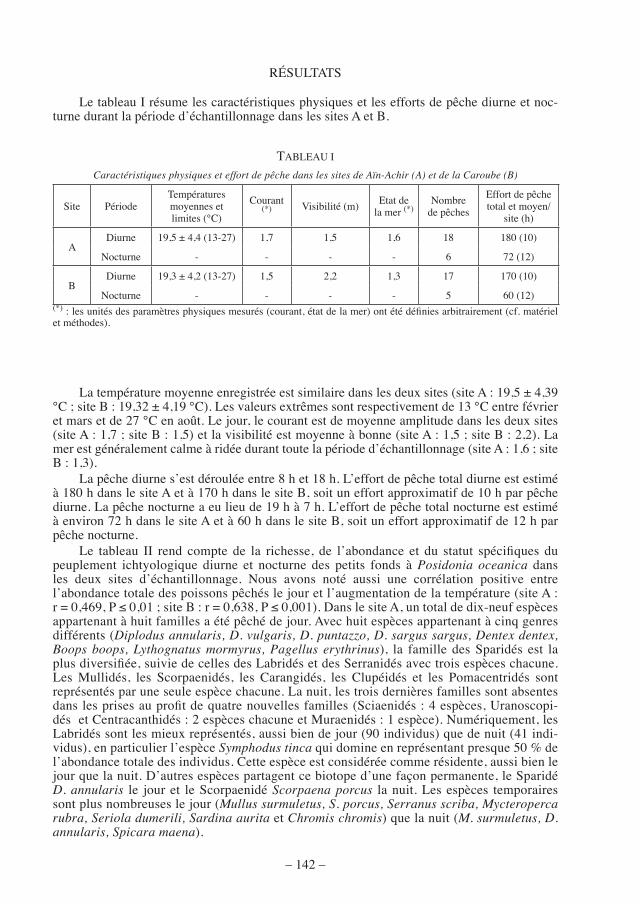
Le tableau I résume les caractéristiques physiques et les efforts de pêche diurne et noc-turne durant la période d’échantillonnage dans les sites A et B.
TABLEAU I
Caractéristiques physiques et effort de pêche dans les sites de Aïn-Achir (A) et de la Caroube (B)
Site PériodeTempératures moyennes et limites (°C)
Courant (*) Visibilité (m) Etat de
la mer (*)Nombre
de pêches
Effort de pêche total et moyen/
site (h)
ADiurne 19,5 ± 4,4 (13-27) 1,7 1,5 1,6 18 180 (10)
Nocturne - - - - 6 72 (12)
BDiurne 19,3 ± 4,2 (13-27) 1,5 2,2 1,3 17 170 (10)
Nocturne - - - - 5 60 (12)(*) : les unités des paramètres physiques mesurés (courant, état de la mer) ont été définies arbitrairement (cf. matériel et méthodes).
La température moyenne enregistrée est similaire dans les deux sites (site A : 19,5 ± 4,39 °C ; site B : 19,32 ± 4,19 °C). Les valeurs extrêmes sont respectivement de 13 °C entre février et mars et de 27 °C en août. Le jour, le courant est de moyenne amplitude dans les deux sites (site A : 1,7 ; site B : 1,5) et la visibilité est moyenne à bonne (site A : 1,5 ; site B : 2,2). La mer est généralement calme à ridée durant toute la période d’échantillonnage (site A : 1,6 ; site B : 1,3).
La pêche diurne s’est déroulée entre 8 h et 18 h. L’effort de pêche total diurne est estimé à 180 h dans le site A et à 170 h dans le site B, soit un effort approximatif de 10 h par pêche diurne. La pêche nocturne a eu lieu de 19 h à 7 h. L’effort de pêche total nocturne est estimé à environ 72 h dans le site A et à 60 h dans le site B, soit un effort approximatif de 12 h par pêche nocturne.
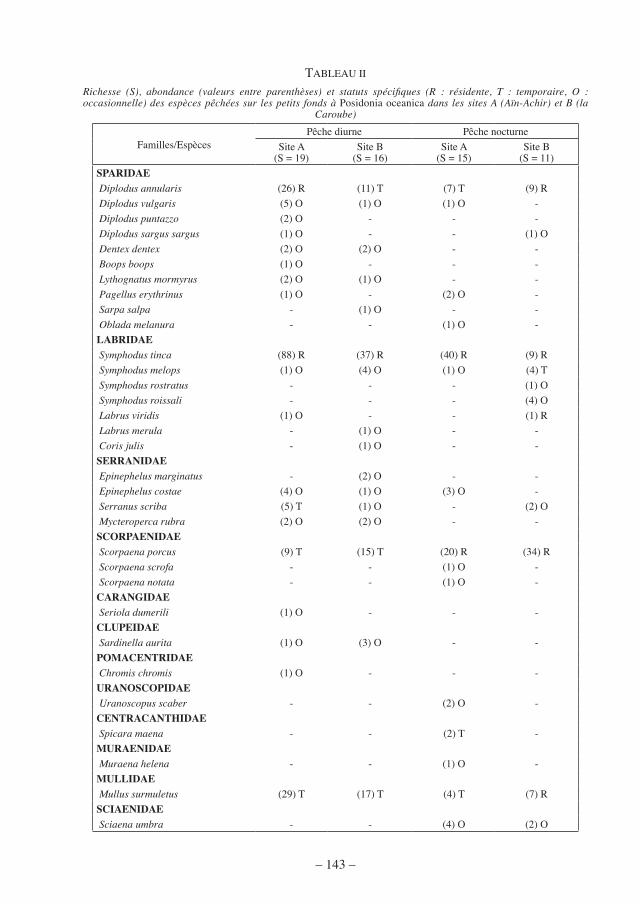
Le tableau II rend compte de la richesse, de l’abondance et du statut spécifiques du peuplement ichtyologique diurne et nocturne des petits fonds à Posidonia oceanica dans les deux sites d’échantillonnage. Nous avons noté aussi une corrélation positive entre l’abondance totale des poissons pêchés le jour et l’augmentation de la température (site A : r = 0,469, P ≤ 0,01 ; site B : r = 0,638, P ≤ 0,001). Dans le site A, un total de dix-neuf espèces appartenant à huit familles a été pêché de jour. Avec huit espèces appartenant à cinq genres différents (Diplodus annularis, D. vulgaris, D. puntazzo, D. sargus sargus, Dentex dentex, Boops boops, Lythognatus mormyrus, Pagellus erythrinus), la famille des Sparidés est la plus diversifiée, suivie de celles des Labridés et des Serranidés avec trois espèces chacune. Les Mullidés, les Scorpaenidés, les Carangidés, les Clupéidés et les Pomacentridés sont représentés par une seule espèce chacune. La nuit, les trois dernières familles sont absentes dans les prises au profit de quatre nouvelles familles (Sciaenidés : 4 espèces, Uranoscopi-dés et Centracanthidés : 2 espèces chacune et Muraenidés : 1 espèce). Numériquement, les Labridés sont les mieux représentés, aussi bien de jour (90 individus) que de nuit (41 indi-vidus), en particulier l’espèce Symphodus tinca qui domine en représentant presque 50 % de l’abondance totale des individus. Cette espèce est considérée comme résidente, aussi bien le jour que la nuit. D’autres espèces partagent ce biotope d’une façon permanente, le Sparidé D. annularis le jour et le Scorpaenidé Scorpaena porcus la nuit. Les espèces temporaires sont plus nombreuses le jour (Mullus surmuletus, S. porcus, Serranus scriba, Mycteroperca rubra, Seriola dumerili, Sardina aurita et Chromis chromis) que la nuit (M. surmuletus, D. annularis, Spicara maena).
– 143 –
TABLEAU II
Richesse (S), abondance (valeurs entre parenthèses) et statuts spécifiques (R : résidente, T : temporaire, O : occasionnelle) des espèces pêchées sur les petits fonds à Posidonia oceanica dans les sites A (Aïn-Achir) et B (la
Caroube)
Familles/EspècesPêche diurne Pêche nocturne
Site A(S = 19)
Site B(S = 16)
Site A(S = 15)
Site B(S = 11)
SPARIDAE Diplodus annularis (26) R (11) T (7) T (9) R Diplodus vulgaris (5) O (1) O (1) O - Diplodus puntazzo (2) O - - - Diplodus sargus sargus (1) O - - (1) O Dentex dentex (2) O (2) O - - Boops boops (1) O - - - Lythognatus mormyrus (2) O (1) O - - Pagellus erythrinus (1) O - (2) O - Sarpa salpa - (1) O - - Oblada melanura - - (1) O -LABRIDAE Symphodus tinca (88) R (37) R (40) R (9) R Symphodus melops (1) O (4) O (1) O (4) T Symphodus rostratus - - - (1) O Symphodus roissali - - - (4) O Labrus viridis (1) O - - (1) R Labrus merula - (1) O - - Coris julis - (1) O - -SERRANIDAE Epinephelus marginatus - (2) O - - Epinephelus costae (4) O (1) O (3) O - Serranus scriba (5) T (1) O - (2) O Mycteroperca rubra (2) O (2) O - -SCORPAENIDAE Scorpaena porcus (9) T (15) T (20) R (34) R Scorpaena scrofa - - (1) O - Scorpaena notata - - (1) O -CARANGIDAE Seriola dumerili (1) O - - -CLUPEIDAE Sardinella aurita (1) O (3) O - -POMACENTRIDAE Chromis chromis (1) O - - -URANOSCOPIDAE Uranoscopus scaber - - (2) O -CENTRACANTHIDAE Spicara maena - - (2) T -MURAENIDAE Muraena helena - - (1) O -MULLIDAE Mullus surmuletus (29) T (17) T (4) T (7) RSCIAENIDAE Sciaena umbra - - (4) O (2) O

– 144 –
Dans le site B, les nombres d’espèces et d’individus capturés le jour (16 espèces, 100 indi-vidus) et la nuit (11 espèces, 74 individus) sont inférieurs à ceux obtenus dans le site A. Une seule espèce (S. tinca) est considérée comme résidente le jour contre quatre espèces (S. tinca, M. surmuletus, D. annularis, S. porcus) la nuit. La famille des Sparidés est la plus diversifiée le jour avec cinq espèces (D. annularis, D. vulgaris, D. dentex, Sarpa salpa, L. mormyrus), suivie des Labridés (S. tinca, S. melops, Labrus merula et Coris julis) et des Serranidés (S. scriba, Epinephelus marginatus, E. costae et M. rubra) avec quatre espèces chacune. Les Serranidés E. marginatus et E. costae et le Sciaenidé Sciaena umbra sont pêchés occasionnellement. La nuit, la diversité spécifique et le nombre d’individus capturés sont en faveur des Labridés, en particulier des espèces S. tinca et Labrus viridis qui sont résidentes dans l’herbier. Par contre, les Sparidés ne sont représentés que par deux espèces, D. annularis qui est résidente et D. sar-gus sargus qui est occasionnelle. Les familles des Mullidés et des Scorpaenidés ne sont repré-sentées chacune que par une espèce résidente, M. surmuletus et S. porcus, respectivement.
Le tableau III présente les indices de diversité (H’) et d’équitabilité (R) pour les pêches diurnes et nocturnes dans les deux sites échantillonnés. De jour, les indices de diversité et d’équitabilité sont respectivement de 2,51 et de 0,33 dans le site A et de 2,87 et de 0,43 dans le site B. La nuit, l’indice de diversité est plus faible (site A : 0,64 et site B : 2,55) et l’équitabilité est proche de 0,40 dans les deux secteurs.
TABLEAU III
Indices de diversité de Shannon-Weaver (H’) et d’équitabilité (R) des pêches diurnes et nocturnes dans les sites de Aïn Achir et de la Caroube
Période Aïn Achir La Caroube
Nombre de pêches (Np)Diurne 18 17Nocturne 6 5
Richesse spécifique (S)Diurne 19 16Nocturne 15 11
Abondance (A)Diurne 182 100Nocturne 90 74
Indice de diversité (H’)Diurne 2,51 2,87Nocturne 2,64 2,55
Equitabilité (R)Diurne 0,33 0,43Nocturne 0,41 0,41
DISCUSSION
Le peuplement ichtyologique des petits fonds à Posidonia oceanica de la baie d’An-naba est composé essentiellement de Sparidés, Labridés et Scorpaenidés. Sur les côtes de l’Est algérien, ces trois familles sont représentées respectivement par 20, 13 et 5 espèces (Derbal & Kara, 2001). Celles-ci constituent la base du peuplement ichtyologique diurne de l’herbier à posidonie dans de nombreuses régions de la Méditerranée (Bell & Harmelin-Vivien, 1982 ; Guidetti & Bussoti, 2000 ; Valle et al., 2001 ; Frau et al., 2003 ; Moranta et al., 2006 ; Deudero & Frau, 2004 ; Deudero et al., 2008).
Dans la baie d’Annaba, la richesse spécifique (26 espèces au total à Aïn-Achir et 21 espè-ces à la Caroube) est nettement inférieure à celle obtenue par Zei (1962) avec un petit chalut dans la région de Split en Adriatique (38 espèces), par Massuti (1965) dans la baie de Palma de Mallorca aux Baléares (29 espèces), par Bell & Harmelin-Vivien (1982) sur les côtes mar-seillaises (49 espèces), par Harmelin-Vivien (1982, 1984) dans le Parc National de Port-Cros (53 espèces) et dans le parc naturel de Galeria en Corse (41 espèces). Avec un chalut à perche de petite dimension, Reñones et al. (1995), récoltent plus en profondeur (25 - 30 m), dans la

– 145 –
baie de Palma (îles Baléares), un total de 41 espèces appartenant à 20 familles. Ce même nom-bre d’espèces a été échantillonné avec un engin similaire sur l’herbier à P. oceanica dans trois sites différents de la Méditerranée occidentale (Mallorca, Formentera et Alacant, en Espagne) (Moranta et al., 2006). Au sein de la réserve marine de l’île Mèdes et dans trois localités non protégées au nord-ouest de la Méditerranée, 57 espèces ont été inventoriées (MacPherson et al., 2002). Dans la province d’Alicante, Valle et al. (2001) recensent à l’aide d’un gangui, sur un fond de 20-22 m, 38 espèces le jour contre 53 espèces la nuit, appartenant à 21 familles. Dans l’herbier du Parc National de Port-Cros, Francour & Harmelin (1988) ont inventorié, par relevé visuel, un total de 19 espèces appartenant à 6 familles, alors que Guidetti & Bussotti (2000) recensent dans le golfe d’Olbia (Sardaigne nord-orientale, Italie) 23 espèces apparte-nant à 8 familles.
Qualitativement, le peuplement ichtyologique de la baie d’Annaba est dominé dans le site A par la famille des Sparidés (9 espèces) et dans le site B par la famille des Labridés (7 espèces), soit 33,3 % et 36,8 % du nombre total des espèces pêchées de jour et de nuit. La présence permanente du crénilabre Symphodus tinca (espèce résidente) dans les 2 sites, aussi bien de jour que de nuit, est une caractéristique des Labridés qui fréquentent les écosystèmes littoraux (Michel et al., 1987), notamment l’herbier à posidonie. Dans la péninsule ibérique, les Labridés sont représentés essentiellement par S. rostratus (Valle et al., 2001). L’espèce Symphodus melops, considérée comme accidentelle sur les côtes tunisiennes (Bradaï et al., 2004) et dans le bassin Levantin (Golani, 1996), a été pêchée occasionnellement dans les deux secteurs (5 individus/site), aussi bien de jour que de nuit. Selon Quignard & Pras (1986), sa répartition se limite au littoral européen jusqu’en Grèce. D’autres espèces du genre Thalas-soma (T. pavo) et Symphodus (S. cinereus, S. doderleini, S. mediterraneus, S. melanocercus, S. ocellatus), rencontrées régulièrement en Méditerranée nord-occidentale (Harmelin-Vivien, 1982, 1984 ; Francour, 2000 ; Guidetti & Bussotti, 2000 ; Valle et al., 2001 ; Frau et al., 2003 ; Moranta et al., 2006), sont absentes de nos captures durant toute la période d’échantillonnage. Cette absence peut être due en partie aux différences dans les stratégies d’échantillonnage adoptées par les différents auteurs (relevés visuels, filets maillants fixes, chaluts), aux exigen-ces écologiques des labres, mais aussi aux caractéristiques de leurs habitats (habitat mixte, herbier superficiel, profond, clairsemé ou dense). En effet, Guidetti & Bussoti (2000) mettent en évidence dans une pelouse mixte à Cymodocea nodosa et Zostera noltii, une corrélation significative entre la densité de plusieurs espèces de Labridés (Labrus viridis, S. tinca, S. cine-reus, Coris julis) et la densité des faisceaux de ces phanérogames. Par contre, Frau et al. (2003) ont montré l’existence de différences spécifiques des habitats préférentiels au sein du même genre, comme c’est le cas de S. mediterraneus, S. rostratus, S. roissali et S. melanocercus qui ont une affinité pour les habitats mixtes (indice d’affinité Ia > 0,7), alors que S. ocellatus et S. doderleini ont une affinité presque similaire (Ia ≈ 0,5) pour les habitats rocheux et mixtes. Dans notre cas, l’habitat échantillonné est considéré comme un herbier à P. oceanica de type clairsemé (Bouhayene, 2002).
En fonction du nycthémère, les espèces résidentes varient selon le site échantillonné. Ainsi, le sparaillon D. annularis est considéré comme une espèce résidente le jour à l’inverse de la rascasse S. porcus qui apparaît plus fréquemment dans les prises nocturnes au niveau du site A. Ceci est dû au fait que le tramail, qui est un engin passif, ne capture les poissons que lors de leurs déplacements horizontaux. D. annularis est une espèce à activité diurne, donc susceptible d’être capturée surtout le jour, tandis que S. porcus présente une activité essentiel-lement nocturne et est donc capturée surtout la nuit. Dans le site B, les espèces qui résident dans l’herbier la nuit sont M. surmuletus, D. annularis et S. porcus. Dans les eaux du Parc National de Port-Cros, D. annularis et S. porcus sont considérées comme des espèces résiden-tes de l’herbier à posidonie (Harmelin-Vivien, 1982). Dans la province d’Alicante, les prises diurnes et nocturnes sont constituées essentiellement de D. annularis, S. scriba et S. rostratus (Valle et al. 2001).
Les espèces qualifiées de résidentes peuvent être soit permanentes et passer la totalité de leur cycle ontogénétique au sein de l’herbier, soit saisonnières et ne passer dans ce biotope qu’une partie de leur cycle. Dans le site A, les espèces résidentes ne représentent que 10,5 % du nombre des espèces pêchées, alors que 36,8 % sont des espèces temporaires et 52,6 % sont

– 146 –
des espèces occasionnelles. Les espèces résidentes sont encore moins fréquentes dans le site B (plage la Caroube) (6,3 %) de même que les espèces temporaires (18,8 %), tandis que la plupart des espèces (75 %) du peuplement ichtyologique de cette zone sont considérées comme occa-sionnelles. Dans le golfe de Galeria (Harmelin-Vivien, 1984), les espèces résidentes, qu’elles soient permanentes ou saisonnières, représentent environ 56 % du nombre d’espèces pêchées, et les visiteurs temporaires et les migrants occasionnels forment environ 22 % chacun. Ces proportions sont nettement différentes de celles obtenues dans notre région, mais sont voisines de celles trouvées pour le peuplement ichtyologique des herbiers des côtes marseillaises, res-pectivement 61 %, 16 % et 22 % (Bell & Harmelin-Vivien, 1982) ou pour celui des herbiers du Parc National de Port-Cros, avec respectivement 60 %, 19 % et 22 % (Harmelin-Vivien, 1982). À Port-Cros, les espèces résidentes, permanentes et saisonnières, appartiennent aux Syngnathidés, Gobiesocidés, Serranidés, Centracanthidés, Labridés, Scorpaenidés et Gobiidés (Harmelin-Vivien, 1982).
En raison des différences dans les méthodes d’inventaires (méthodes destructives : filets, chaluts ; méthodes non destructives : comptage visuel, vidéo sous-marine), de l’effort d’échan-tillonnage et des spécificités environnementales locales entre les rives nord et sud du bassin méditerranéen (différences biogéographiques), il est difficile de comparer la richesse spéci-fique ou familiale ou les abondances et de statuer ainsi sur l’espèce. L’action, parfois conju-guée, de ces facteurs peut biaiser fortement la diversité du peuplement ichtyologique en place ; d’ailleurs, cette question a été soulevée par de nombreux auteurs (Harmelin-Vivien et al. 1985 ; Harmelin-Vivien & Francour, 1992 ; Francour, 1999 ; De Girolamo & Mazzoldi, 2001 ; Stobart et al., 2007). À titre d’exemple, en utilisant le chalut à perche de petites dimensions (1,5 m x 0,5 m d’ouverture), Harmelin-Vivien (1982) dénombre 53 espèces dans l’herbier du Parc National de Port-Cros et 41 espèces dans la région de Galeria en Corse. Ce même nombre d’espèces est récolté avec un engin similaire dans la baie de Palma, autour des îles Baléares (Reñones et al., 1995). En échantillonnant entre 18 et 38 m avec un chalut de dimensions pres-que similaires (l : 1,8 m, h : 0,8 m, L : 8 m), Deudero et al. (2008), ont inventorié 51 espèces sur les côtes sud-ouest de l’île de Mallorca (Baléares).
Le calcul de l’indice de Shannon (H’) montre une certaine similarité de la diversité spéci-fique et une forte stabilité de la structure du peuplement ichtyologique en place dans les deux sites A et B, et ce malgré la présence temporaire de quelques effluents domestiques le long de la plage la Caroube. À l’inverse, Guidetti et al. (2002) soulignent que certaines espèces côtières peuvent répondre négativement (réduction de la population, fuite vers les eaux du large) à l’im-pact provoqué par la décharge d’eaux usées. Harmelin-Vivien (1982, 1984) considère que le peuplement ichtyologique présente de fortes variations en fonction du rythme nycthéméral et des fluctuations de moindre amplitude en fonction de la saison et même du site d’échantillon-nage. Selon ce même auteur, la richesse spécifique est plus élevée la nuit (49 espèces) que le jour (38 espèces), quels que soient le lieu et la saison. L’indice de diversité H’ augmente aussi la nuit. Dans la baie de Palma, Reñones et al. (1995) observent aussi des différences nycthé-mérales quantitatives (en nombre et en biomasse) de la famille des Centracanthidés dans les échantillons nocturnes, et des familles des Labridés et des Scorpaenidés dans les échantillons diurnes. Le tableau IV compare l’indice de Shannon H’ dans différentes localités de Méditer-ranée et d’Atlantique.
Le nombre d’espèces pêchées dans l’herbier de posidonie de la baie d’Annaba est net-tement inférieur à celui trouvé au sein d’autres herbiers de phanérogames marines en zone tempérée. Ceci est très probablement dû aux dissemblances biogéographiques du peuplement ichtyologique côtier le long des côtes méditerranéennes mais également à la technique de pêche, sélective, que représentent les filets tramails. Il est vraisemblable qu’une récolte réalisée à l’aide d’un engin actif, comme c’est le cas du petit chalut à perche, aurait fourni un nombre plus élevé d’espèces. Si la richesse spécifique diffère d’un secteur géographique à l’autre, ce sont bien souvent les mêmes familles qui constituent la base du peuplement ichtyologique de l’herbier à P. oceanica (Labridés, Sparidés, Scorpaenidés). La présence, quoique occasion-nelle, de certaines espèces rares telles que E. marginatus, E. costae, S. umbra et M. helena peut être considérée comme un indicateur de stabilité de la zone d’étude. Toutefois, l’extension de l’urbanisation pourrait avoir des effets négatifs au plan écologique d’une manière générale et

– 147 –
sur l’ichtyodiversité locale en particulier. Une étude à long terme de l’importance relative de chacune des espèces qui fréquentent l’herbier, notamment au niveau des secteurs perturbés par l’action anthropique, permettrait d’estimer le degré de variabilité spatio-temporelle des peuplements de poissons. De ce fait, elle pourrait constituer un bon indicateur de stabilité des peuplements ichtyologiques littoraux. En effet, les analyses pluriannuelles permettent non seu-lement de mieux expliquer le niveau de résilience des peuplements ichtyologiques des herbiers vis-à-vis des perturbations environnementales, mais aussi de dégager des conclusions pertinen-tes dans l’étude des peuplements ichtyologiques littoraux (Francour, 2000).
TABLEAU IV
Variations de l’indice de diversité de Shannon-Weaver (H’) dans différentes régions de Méditerranée et d’Atlantique (a : pêche diurne, b : pêche nocturne)
Zones d’étude H’ RéférencesBaie d’Annnaba (Algérie) Présente étude- Aïn Achir 2,51a - 2,63b
- La Caroube 2,86a - 2,55b
Ile de Corse (France) Harmelin-Vivien (1984)- Stollo 3,26a - 4,15- Galeria 3,20a - 3,11b
Ria de Arosa (NO, Espagne) 0,81 - 1,23 Chesney & Iglesias (1979)Ouest de l’Écosse 1,23 - 1,95 Nash & Gibson (1982)Ria de Muros-Noya (NO, Espagne) 1,72 - 2,21 Iglesias (1983)Ilia de Pontevedra (NO, Espagne) 1,64 - 1,78 Iglesias & Gonzalez-Gurriarfin (1984)Baie de Málaga (Espagne) Reina-Hervás & Serrano (1987)- secteur occidental 2,14- secteur est 2,12
RÉFÉRENCES
abdelli-Menail, S. (2007). — Contribution à l’écobiologie de la grande nacre Pinna nobilis dans le golfe d’Annaba. Mémoire de Magister en Sciences de la Mer, Université d’Annaba - Badji Mokhtar, Algérie.
anonyMe, (1999). — Action plan for the conservation of marine vegetation in the Mediterranean Sea. United Nations Environment Programme, Mediterranean Action Plan Publ., Tunisia.
barbault, r. (1992). — Écologie des peuplements, structure, dynamique et évolution. Masson, Paris.bell, J.d. & HarMelin-ViVien, M.l. (1982). — Fish fauna of French Mediterranean Posidonia oceanica seagrass
meadows. 1. Community structure. Téthys, 10 : 337-347.boucHon-naVaro, y., boucHon, c. & louiS, M. (1992). — L’ichtyofaune des herbiers de phanérogames marines de
la baie de Fort-de-France (Martinique, Antilles Françaises). Cybium, 16 : 307-330.boudoureSque, C.F. (2004). — Marine biodiversity in the Mediterranean : status of species, populations and com-Marine biodiversity in the Mediterranean : status of species, populations and com-
munities. Sci. Rep. Port-Cros natl Park, 20 : 97-146.boudoureSque, c.F., bernard, G., bonHoMne, P., cHarbonnel, e., diViacco, G., MeineSz, a., PerGent, G.,
PerGent-Martini, c., ruitton, S. & tuneSi, l. (2006). — Préservation et conservation des herbiers à Posidonia oceanica. Ramoge Pub. : 1-202.
bouHayene, S. (2002). — Contribution à la connaissance des herbiers à Posidonia oceanica dans la baie d’Annaba (Est algérien) : Phénologie, biomasse, lépidochronologie et production primaire. Mémoire de Magistère en Sciences de la Mer, Université Badji-Mokhtar, Algérie.
bradaï, n.M., quiGnard, J.P., bouaïn, a., Jarbaoui, o., ouanneS-GHorbel, a., ben abdallaH, l., zaouali, J. S. & ben SaleM, S. (2004). — Ichtyofaune autochtone et exotique des côtes tunisiennes : recensement et biogéographie. Cybium, 28 : 315-328.
cHeSney, e.J.Jr. & iGleSiaS, J. (1979). — Seasonal distribution, abundance and diversity of demersal fishes in the Inner Ria de Arosa, Northwest Spain. Estuar. cstl mar. Sci., 8 : 227-239.

– 148 –
clynick, B.G. (2006). — Assemblages of fish associated with coastal marinas in north-western Italy. J. Mar. Biol. Ass. UK, 86 : 847-852.
de GirolaMo, M. & Mazzoldi, c. (2001). — The application of visual census on Mediterranean rocky habitats. Mar. Env. Res., 51 : 1-16.
derbal, F., Francour, P., tHibault, t., kara, M.H. & Faure, e. (2005). — Écologie de quelques espèces cibles (Diplodus cervinus cervinus, Diplodus puntazzo, Epinephelus costae, Sciaena umbra) par relevés visuels sur le littoral est de l’Algérie. Conférence Internationale des Limnologues et Océanographes, Lyon-Vaulx-en-Velin, 4-7 juillet 2005. France.
derbal, F. & kara, M.H. (2001). — Inventaire des poissons des côtes de l’Est algérien. Rapp. Comm. Int. Mer Médit., 36 : 258.
derbal, F. & kara, M.H. (2005). — Contribution à l’inventaire de la faune invertébrée des côtes de l’Est algérien. Pp 1-687 in : A. Chouikhi & M Menioui. (eds), International Workshop Proceedings. Inter-Islamic Science and a Technology Network on Oceanography, Marine and a Coastal Protected Areas.
deudero, S. & Frau, a. (2004). — Role of mixed and fragmentated habitats in fish habitat preference. Rapp. Comm. Int. Mer. Médit., 37 : 513.
deudero, S., Morey, G., Frau, a., Moranta, J. & Moreno, i. (2008). — Temporal trends of littoral fishes at deep Posidonia oceanica seagrass meadows in a temperate costal zone. J. Mar. Syst., 70 : 182-195.
Fernandez, t.V., Milazzo, M., badalaMenti, F. & d’anna, G. (2005). — Comparison of the fish assemblages asso-ciated with Posidonia oceanica after the partial loss and consequent fragmentation of the meadow. Estuar. Cstl mar. Sci., 65 : 645-653.
Francour, P. (1989). — Les peuplements ichtyologiques de la réserve de Scandola : influence de la réserve intégrale. Trav. sci. Parc nat. Région. Rés. Nat. Corse, 21 : 33-93.
Francour, P. (1999). — A critical review of adult and juvenile fish sampling techniques in Posidonia oceanica sea-grass beds. Naturalista sicil., 23 : 33-57.
Francour, P. (2000). — Évolution spatio-temporelle à long terme des peuplements de poissons des herbiers à Posi-donia oceanica de la réserve naturelle de Scandola (Corse, Méditerranée Nord-Occidentale). Cybium, 24(3) suppl. : 85-95.
Francour, P. & HarMelin, J.G. (1988). — Inventaire de la faune ichtyologique marine de Port-Cros. Trav. Sci. Parc Natl. Port-Cros., 14 : 65-79.
Frau, a., deudero, S., cerdeno, S. & alou, l. (2003). — No habit preference in mixed meadows and rocky bottoms for Mediterranean Labridae and Sparidae fish species. Bol. Inst. Esp. Oceanogr., 19 : 483-491.
Golani, d. (1996). — The marine ichtyofauna of the eastern Levant. History, inventory and characterization. Isr. J. Zool., 42 : 15-55.
Guidetti, P. & buSSoti, S. (2000). — Fish fauna of a mixed meadow composed by the seagrasses Cymodocea nodosa and Zostera noltii in the Western Mediterranean. Oceanologica Acta, 23 : 759-770.
Guidetti, P., Fanelli, G., FraScHetti, S., terlizzi, a. & boero, F. (2002). — Coastal fish indicate human-induced changes in the Mediterranean littoral. Mar. Envir. Res., 53 : 77-92.
HarMelin-ViVien, M.l. (1982). — Ichtyofaune des herbiers de posidonies du Parc National de Port-Cros. I. Compo-ort-Cros. I. Compo-sition et variations spatio-temporelles. Trav. Sci. Parc Natl. Port-Cros., 8 : 69-92.
HarMelin-ViVien, M.l. (1984). — Ichtyofaune des herbiers de posidonies du Parc Naturel Régional de Corse. Pp 291-301 in : C. F. Boudouresque, A. Jeudy De Grissac & J. Olivier (eds). First Int. Workshop Posidonia oceanica. GIS Posidonie, Marseille.
HarMelin-ViVien, M.l. (2000). — Influence of fishing on the trophic structure of fish assemblages in the Mediter-ranean sea grass beds. Pp 39-41 in : F. Briand (ed.). Fishing down the Mediterranean food webs. CIESM Workshop Series 12. Monaco.
HarMelin-ViVien, M.l. & Francour, P. (1992). — Trawling or visual censuses ? Methodological bias in the assess-Methodological bias in the assess-ment of fish populations in seagrass beds. P.S.Z.N.I. Mar. Ecol., 13 : 41-51.
HarMelin-ViVien, M., HarMelin, J.G., cHauVet, c., duVal, c., Galzin, r., leJeune, P., barnabé, G., blanc, F., cHeValier, r., duclerc, J. & laSSerre, G. (1985). — Évaluation visuelle des peuplements et populations de poissons. Problèmes et méthodes. Rev. Ecol. (Terre Vie), 40 : 467-539.
HaVelanGe, S., lePoint, G., dauby, P. & bouqueGneau, J.M. (1997). — Feeding of the sparid fish Sarpa salpa in a seagrass ecosystem : Diet and carbon flux. P.S.Z.N.I. Mar. Ecol., 18 : 289-297.
iGleSiaS, J. (1983). — Cambios estacionales de la megafauna bentonica de la Ria de Muros Noya : I. Peces demersales. Bol. Inst. Esp. Oceanogr., 1 : 131-144.
iGleSiaS, J. & Gonzalez-GurriarFin, e. (1984). — Primeros datos sobre la megafauna bentonica de la Ria de Pon-tevedra : peces demersales y crustaceos decapodos (Brachyura). Cuad. Cien. Mar., Sem. Est., Galegos, 1 : 303-319.
letourneur, y., ruitton, S. & Sartoretto, S. (2003). — Environmental and benthic habitat factors structuring the spatial distribution of a summer infralittoral fish assemblage in the north-western Mediterranean Sea. J. Mar. Biol. Ass. UK, 83 : 193-204.
MacPHerSon, e., Gordoa, a. & Garcia-rubieS, a. (2002). — Biomass size spectra in littoral fishes in protected and unprotected areas in the NW Mediterranean. Estuar. cstl mar. Sci., 55 : 777-788.
MaSSuti, M. (1965). — Estudio de los fondos de pesca de Baleares. I : Ciclo anual de los peces de las praderas de Caulerpa y Posidonia oceanica capturados por un pequeño arte de arrastre en la bahia de Palma de Mallorca. Bol. Inst. Esp. Oceanogr., 119 : 1-57.

– 149 –
Mezali, k., zuPo, o. & Francour, P. (2006). — Population dynamics of Holothuria tubulosa and Holothuria polii of an Algerian Posidonia oceanica meadow. Biol. Mar. Med., 13 : 158-161.
MicHel, c., leJeune, P. & VoSS, J. (1987). — Biologie et comportement des Labridae européens. Rev. Fr. Aquariol. Herpétol., 48 : 1-80.
Moranta, J., PalMer, M., Morey, G., ruiz, a. & MoraleS-nin, b. (2006). — Multi-scale spatial variability in fish assemblages associated with Posidonia oceanica meadows in the Western Mediterranean Sea. Estuar. cstl mar. Sci., 68 : 579-592.
nakaoka, M. (2005). — Plant-animal interactions in seagrass beds : ongoing and future challenges for understanding population and community dynamics. Pop. Ecol., 47 : 167-177.
naSH, r.d.M. & GibSon, r.n. (1982). — Seasonal fluctuations and compositions of two populations of small demersal fishes on the west coast of Scotland. Estuar. cstl mar. Sci., 15 : 485-495.
PerGent, G., SeMroud, r., baba aHMed, r., derbal, F., kHatal, y., reMili, a., robert, P. & BoudoureSque, C.F. (1993). — Données préliminaires sur la répartition et l’état de l’herbier à Posidonia oceanica de la région d’El-Kala. Sci. Rep. Port-Cros natl. Park, Fr., 15 : 253-563.
PerGent-Martini, c., rico-raiMondino, V. & PerGent, G. (1994). — Primary production of Posidonia oceanica in the Mediterranean basin. Mar. Biol., 120 : 9-15.
Pielou, E.C. (1966). — The measurement of diversity in different types of biological collections. J. Theor. Biol., 13 : 131-144.
quiGnard, J.P. & PraS, a. (1986). — Labridae. Pp 919-942, in : P.J.P. Withehead, M.L. Bauchot, J.C. Hureau, J. Niel-son & E. Tortonese (eds). Fishes of the Northeastern Atlantic and the Mediterranean. UNESCO, Paris.
reina-HerVáS, J.a. & Serrano, P. (1987). — Structural and seasonal variations of inshore fish populations in Málaga Bay, Southeastern Spain. Mar. Biol., 95 : 501-508.
reñoneS, o., MaSSuti, e., Moranta, J., coll, J. & Moreno, i. (1995). — Fish fauna of Posidonia oceanica seagrass meadows in Palma Bay (Balearic Islands). Cybium, 19 : 201-206.
SancHez-Jerez, P. & raMoS eSPla, a.a. (1996). — Detection of environmental impacts by bottom trawling on Posi-donia oceanica (L.) Delile meadows : sensitivity of fish and macroinvertebrate communities. J. Aqu. Ecos. Health., 5 : 239-253.
SeMroud, r. (1993). — Contribution à la connaissance de l’écosystème à Posidonia oceanica (L.) Delille, dans la région d’Alger (Algérie) : étude de quelques compartiments. Thèse de Doctorat d’Etat, Université des Scien-ces et Techniques Houari Boumediene, Algérie.
SHannon, c.e. & WeaVer, W. (1949). — The mathematical theory of communication. University of Illinois Press., Urbana.
Stobart, b., Garcia-cHarton, J.a., eSPeJo, c., rocHel, e., Goñi, r., reñoneS, o., Herrero, a., crec’Hriou, r., Polti, S., MarcoS, c., PlaneS, S. & Perez-ruzaFa, a. (2007). — A baited underwater video technique to assess shallow-water Mediterranean fish assemblages : Methodological evaluation. J. Exp. Mar. Biol. Ecol., 345 : 158-174.
Valle, c., bayle SeMPere, J.t. & raMoS eSPla, A.A. (2001). — Estudio multiescalar de la ictiofauna asociada a praderas de Posidonia oceanica (L.) Delile, 1813 en Alicante (sudeste iberico). Bol. Esp. Oceanogr., 17 : 49-60.
zei, M. (1962). — Preliminary observations on the life in Posidonia oceanica beds. Pubbl. Staz. Zool. Napoli (Suppl.), 32 : 1-5.