Embed Size (px)
Citation preview

CLINICAL MICROBIOLOGY REVIEWS, Jan. 1993, p. 1-210893-8512/93/010001-21$02.00/0Copyright © 1993, American Society for Microbiology
Compounds Active against Cell Walls ofMedically Important Fungi
RICHARD F. HECTORtCutter Biological, Berkeley, California 94710
Vol. 6, No. 1
INTRODUCTION .............................................................................1OVERVIEW OF THE CELL WALL BIOCHEMISTRY OF MEDICALLY IMPORTANT FUNGI ...........1
Yeasts.............................................................................1Filamentous Fungi............................................................................. 3Dimorphic Pathogenic Fungi............................................................................. 4
COMPOUNDS ACTIVE AGAINST FUNGAL CELL WALLS ..........................................................5Inhibitors of Chitin Synthesis.............................................................................. 5
History of polyoxins and nikkomycins.............................................................................. 5Biochemistry of polyoxins and nikkomycins .............................................................................5Use of polyoxins and nikkomycins against medically important fungi ............................................7Nonspecific inhibitors of chitin synthesis............................................................................. 8New compounds............................................................................. 9Inhibitors of chitinases ............................................................................. 9
Inhibitors of Glucan Synthesis ............................................................................. 9History of aculeacins, echinocandins, and papulacandins ............................................................9Biochemistry of beta-glucan inhibitors............................................................................. 10Effects of beta-glucan inhibitors on medically important fungi....................................................12Recent developments with inhibitors of glucan synthesis............................................................13
Benanomicins and Pradimicins .............................................................................. 13Combinations of Cell Wall-Active Substances ............................................................................13
CONCLUSIONS............................................................................. 14REFERENCES
INTRODUCTION
The antifungal agents available to treat systemic mycoticinfections are limited in both numbers and effectiveness.While a number of azole drugs have been introduced or arepresently being evaluated, this newer class of agent has yetto supplant amphotericin B as the drug of choice in life-threatening infections. This situation is all the more unfor-tunate given the dramatic increases in the numbers of fungalinfections in certain populations of immunocompromisedpatients (70, 201, 209).The aforementioned compounds share a target in that they
all interfere with the structural integrity of the fungal cyto-plasmic membrane: amphotericin B by physical disruption ofthe membrane, and azoles by blocking synthesis of theergosterol component (1, 44, 255). Even the allylamines, anew class of agent recently introduced to the market, sharethis target by interfering with squalene epoxidase activity(204).
Prokaryotes have a variety of physiologic and structuralproperties unique to their kingdom that are therefore appro-priate drug targets. In contrast, fungi, as eukaryotic organ-
isms, have limited numbers of identified targets that distin-guish them from mammalian systems. In addition to thecytoplasmic membranes, the cell walls of medically impor-tant fungi (and some protists) have components that are notencountered elsewhere in nature. The carbohydrate poly-mers composed of the alpha- and beta-glucans, chitin, and
t Present address: Shaman Pharmaceuticals, 887 Industrial Road,San Carlos, CA 94070-3312.
mannan all play roles in maintaining the structural integrityof fungal cells in a fashion analogous to the peptidoglycan ofthe bacterial cell wall. It follows that if a compound inter-fered with the synthesis or structure of the fungal cell wall,the results could be catastrophic for the organism, analogousto the effect of beta-lactams on bacteria. A number ofcompounds have been discovered and described over thepast 25 years that have, as either their primary or secondarymechanism of action, the ability to affect the cell walls offungi. This review presents what is known about thesecompounds with respect to their characterizations, mecha-nisms of action, and spectra of activity. Also, commentaryon their potential for use as antifungal drugs either singly orin combination with other compounds will be provided.
OVERVIEW OF THE CELL WALL BIOCHEMISTRY OFMEDICALLY IMPORTANT FUNGI
While a detailed discussion on the structure and compo-sition of fungal cell walls falls outside the scope of thisreview, a concise overview of the subject presented in thecontext of how specific compounds interfering with the cellwall can serve as antifungal agents would be of value.Further information can be found in several excellent re-
views on this subject that cover both saprophytic andmedically important fungi (13, 37, 45, 86, 176, 189).
Yeasts
The majority of information about fungal cell wall compo-sition, synthesis, and genetics has come from studies involv-
1
1-d
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.

CLIN. MICROBIOL. REV.
ing true yeasts, both saprophytic and medically importantspecies. This is not surprising given the ease of handling thisfungal form and the overwhelming numbers of infections dueto Candida spp. (177). Indeed, the vast majority of publishedstudies on the subject of the cell walls of medically importantfungi deal with the single species Candida albicans. How-ever, this review begins with some information on the cellwalls of saprophytic yeasts, since even more is known in thisarea, and then draws parallels with the information availableon C. albicans and other medically important fungi.
Ultrastructural and biochemical studies of Saccharomycescerevisiae have shown that the wall of this yeast is composedof distinct layers differing in composition. The innermostlayer is predominantly composed of microfibrils of beta-(1,3)-glucan with some beta-(1,6) branch points (143, 152,153), while the outer layer is essentially all mannoprotein (9,104). In contrast to the cell walls of most medically impor-tant fungi, the cell wall of S. cerevisiae is relatively poor inchitin and depends on the beta-(1,3)-glucan for maintenanceof its structural integrity (148). However, the bud septumand resulting bud scars are composed primarily of chitin (35,36, 169). Elucidation of the factors governing the regulationof chitin synthesis in S. cerevisiae has been the subject ofintense research efforts over the past two decades, with amolecular approach providing clarification of many of thepreviously unresolved issues. To date, several genes regu-lating different aspects of chitin synthesis in this yeast havebeen described, CHSI, CHS2, CS02, CS03, CS04, andCALI (32, 38, 51, 220, 254). Each has been found to regulatedifferent aspects of chitin synthesis during the growth,budding, and repair of the cell wall of this yeast, withdifferential levels of activity.
In addition to synthesis of components of the cell wall, theability to selectively degrade polymers within the wall duringcell growth is presumed to be of great importance. Moststudies on this subject have centered on the chitinases foundin S. cerevisiae (39, 62, 78, 146, 147). An endochitinase fromthis yeast has been characterized and cloned and was foundto be essential for the separation of mother and daughtercells. The enzyme, which is found in vesicles in the periplas-mic space, may actually be used to "nick" the chitin fibrilsrather than to degrade large portions of the polymer. Thiswould presumably lead to a softening of the chitin mesh incritical areas of the septum, allowing the cells to separate.Not surprisingly, the importance of a balance between chitinsynthesis and chitinase activity during cell growth andseparation has been recognized (18).While mannoprotein represents a major percentage of the
cell wall mass in saphrophytic fungi (37), less is known aboutthe role of this component in the cell wall than about theother polymers. It is known that the outer core of manno-protein, with its branched alpha-(1,2), -(1,3) and -(1,6) link-ages, is the principal immunochemical determinant of thecell wall (8, 226, 239). One report describes speculation thatthe mannoprotein may serve as a protective shield bydetermining the porosity of the cell wall, thereby limitingaccess to the more critical glucan layer by degradativeenzymes produced by competing soil or other environmentalorganisms (269). While some information on endogenousmannosidases from S. cerevisiae is available, experimentswith overproducing strains created by the introduction ofplasmids carrying the relevant genes failed to reveal pheno-typic differences in the whole cells (147).The cell wall of C. albicans shares many similarities with
those of saprophytic yeasts; it has a budding form composedof chitin, beta-glucan, and mannoprotein, with the latter two
components constituting approximately 80% of the wallmass (232). This yeast, however, has a slightly highercomponent of chitin, constituting 0.6 to 1% of the cell wall(50, 232). Although 90% of the chitin is restricted to the budscar site, the remainder can be found distributed throughoutthe cell wall, contributing to its strength (250). Throughspecialized cytochemical techniques, investigators were ableto demonstrate the presence of multiple layers within the cellwall of C. albicans that could be differentiated on the basis ofchemical composition (46, 195). In general, the outer layersare thought to be composed of mannan, mannoprotein, andbeta-(1,6)-glucan, while the inner layers are predominantlybeta-(1,3)-glucan and chitin with some mannoprotein. Theinner layers are responsible for maintaining the structuralintegrity of the cell wall, with an increase in its rigidity dueto a covalent cross-linking of glucan and chitin (235). In cellsentering a stationary phase of growth, however, a moreuniform distribution of these polymers throughout the cellwall has been described (46).
Additional information on the interaction and cross-link-ing of the major components of the cell wall has beenobtained through the study of regenerating protoplasts ofCandida spp. During the earlier stages of regeneration,chitin is synthesized in amounts greater than normally foundin intact yeast or mycelial cells, while glucan and mannansyntheses lag initially (80, 160, 163). After 4 to 5 h ofregeneration, the normal localization of mannan in the outerwall layers and chitin in the inner layers can be observed.By using specific inhibitors of chitin, glucan, and manno-protein synthesis introduced at intervals during the regener-ation process, Elorza et al. were able to document theimportance of the chitin mesh in forming the frameworkupon which glucan and mannoprotein were subsequentlyadded, with the latter components determining the shape ofthe cell (79).As is the case with saprophytic yeasts, the mannoprotein
component of the cell wall is responsible for the antigenicdeterminants in Candida spp. With a backbone of alpha-(1,6)-linked residues and side chains with alpha-(1,2) and-(1,3) linkages, a variety of species-specific antigens havebeen characterized (200, 234, 237). As for S. cerevisiae,degradative enzymes, i.e., chitinase, glucanase, and man-nanase, have been described for C. albicans (12, 14). In-creases in their activities coincide with an increase in syn-thetic activity during log-phase growth, again suggesting theimportance of a balance between synthesis and degradationduring the remodeling of the cell wall.An important distinction between C. albicans and yeasts
such as S. cerevisiae is the ability of the former to grow in atrue hyphal stage (however, a recent report describes theability of S. cerevisiae to grow in a pseudohyphal formduring nitrogen starvation [89]). Besides the obvious mor-phologic differences in the two growth forms, there are someimportant differences in the physical compositions of theircell walls. The mycelial phase is known to contain three tofive times as much chitin as the yeast phase (30, 50, 232),possibly a reflection of the higher levels of chitin synthetaseactivity during germ tube formation (54). While the overallcomposition of glucan, mannan, lipid, and protein is similarin both growth phases (232), the proportions of specificglucans are not. The activity of beta-(1,3)-D-glucan synthaseis significantly higher in the mycelial form, while exo-(1,3)-beta-D-glucan hydrolase activity is greatly reduced (170).Not surprisingly, therefore, the hyphal form was found tocontain a higher percentage of beta-(1,3)-glucan than beta-(1,6)-glucan, a reverse of the situation found in the yeast
2 HECTsOR
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.

COMPOUNDS ACTIVE AGAINST CELL WALLS OF FUNGI 3
form (93). The glucans of the hyphal form also contain ahigher number of branch points, which Gopal et al. specu-lated may contribute to the morphogenic transition betweenyeast and mycelium. In an ultrastructural and cytologicalstudy of germ tube formation, Cassone et al. (48) foundthat the emerging tube originated from the innermostlayers of the blastospore and was rich in chitin. With re-spect to the role of degradative enzymes in germ tubeformation, Molina et al. (170) discovered that glucanaseactivity substantially decreased during the yeast to myce-lium transition, while Sullivan et al. (233) found thatN-acetyl-glucosaminidase secretion was greatly increased in the my-celial form. In the latter report, the authors speculate thatthe secretion was facilitated by a more porous wall in themycelial phase.
Recently, genes encoding for the ChsI and ChsII forms ofchitin synthetase were cloned from C. albicans and demon-strated to have a degree of homology with those found in S.cerevisiae (4, 51). Interestingly, Chen-Wu et al. found dif-ferential expression of the enzymes dependent on whetherthe organism was growing in the yeast or hyphal form, withthe CHS2 form being expressed at much higher levels duringgermination (51). Because of the higher amounts of chitin inthe hyphal form, it is tempting to speculate that control ofmorphogenesis in C. albicans may be partially under thecontrol of one of the CHS genes.
Despite the fact that Cryptococcus neoformans is thecause of significant numbers of infections in mammals, thestructure and composition of this yeast have not beenextensively studied. In particular, little is known about thecell wall because more attention is generally focused on thecapsule surrounding this yeast. Indeed, analytical studies ofthe cell wall are generally hampered by the capsule, whichcontains large amounts of glucuronoxylomannan and smalleramounts of galactoxylomannan (20, 52, 53). An analysis ofthe sugar components of an acapsular mutant of Cryptococ-cus neoformans determined that, overall, glucose consti-tutes 86% of the cell mass, while N-acetylglucosamineaccounts for 7% (129). Various extracts from the walls of thismutant were assayed, and the presence of glucans withalpha-(1,3), beta-(1,6), and beta-(1,3) linkages was deter-mined. The presence of both alpha and beta linkages wasalso demonstrated through the production of spheroplasts byusing polysaccharolytic enzymes capable of degrading thesepolymers (15, 186). Mannose constitutes only 10% of the cellwall and is not found as a neutral sugar, but rather isrestricted to mannoprotein. In contrast to its position in mostother yeasts, the mannoprotein is located primarily in theinner layer of the cell wall and therefore may be involvedonly in nonstructural roles (257). Ultrastructural studieshave shown that the wall of Cryptococcus neoformans hastwo layers, an electron-dense inner layer and a translucentouter layer (49).
Despite the fact that strains of Tnchosporon and Geotri-chum spp. are now more frequently isolated from patients,little information exists in the literature on the structure andcomposition of these yeasts. Weijman (258) has reported therelative concentrations of hexoses in the cell walls of Geot-nichum and Trichosporon spp., but information on the majorclasses of polymers is lacking.
Filamentous Fungi
The dynamics of hyphal growth, usually termed apicalextension, is a subject that has attracted considerable atten-
tion over the years. It is an obvious statement that thefactors controlling apical extension must be fundamentallydifferent from those involved with the spherical growthof yeasts, though the wall components are often the same.An excellent review of these two forms of growth thatdescribes fungi capable of both has been written by Shep-herd (223).
Unfortunately, most of what is known about the compo-sition and growth of the cell walls of filamentous fungi isbased on studies of saprophytic species. However, in thecase of Aspergillus spp., it can be assumed that the medi-cally important species share many features with those notcommonly associated with mycoses. In particular, a numberof papers on the subject of chitin and glucan synthesis inAspergillus nidulans have appeared. The overall composi-tion of the cell wall of this fungus has been characterized bytwo different groups (33, 271), who both found that glucoseaccounted for roughly 26 to 30% of the wall mass, galactosewas 1 to 4%, mannose was only 3 to 5%, and glucosamineand N-acetylglucosamine constituted 14%. Chitin and beta-glucan were the major polymers in the wall, with beta-(1,3)-glucan predominating over the beta-(1,6) form. Alpha-(1,3)-glucan and galactosamine were also detected.
In a study in which factors regulating chitin synthesis wereaddressed, the use of cycloheximide to deregulate chitinsynthesis in growing hyphae resulted in chitin synthesisalong the entire hypha rather than just at the apical tip (134,230). This caused a halt in elongation but an increase inbranching. Using chitin synthetase isolated from apical tipsor from distal portions of untreated hyphae of Aspergillusfumigatus, Archer was able to demonstrate that, while theenzyme from the growing tips was activated, the chitinsynthetase from mature portions was latent (2).
Studies from two different groups employing temperature-sensitive mutants of A. nidulans underscore the importanceof chitin or beta-glucan or both in providing structuralrigidity in the cell wall. Katz and Rosenberger, in a studypublished in 1970 (133), found that their mutant madereduced amounts of chitin but normal amounts of glucose atthe restrictive temperature and grew without lysis only whenthe growth medium was osmotically stabilized. When anintermediate tonicity was used, swollen hyphae were pro-duced. More recently, two reports have described a series oftemperature-sensitive mutants of A. nidulans defective ei-ther in amidotransferase (hence, blocked for chitin synthe-sis) or in the ability to synthesize beta-(1,3)-glucan (22, 23).At the restrictive temperature, germinating conidia of themutants deficient in chitin formed osmotically sensitive cellsthat lysed under hypotonic conditions; the phenotype wasreversed by supplying exogenous N-acetylglucosamine. Themutants with reduced beta-(1,3)-glucan contained normallevels of chitin and alpha-(1,3)-glucan but were similarlyprone to lysis on hypotonic media. Collectively, these datademonstrate the critical roles of both chitin and beta-(1,3)-glucan in this species.Chemical and ultrastructural analyses of a mutant of A.
nidulans lacking the ability to synthesize alpha-(1,3)-glucanrevealed that this polymer was essentially restricted to theouter wall layer (191). The cell wall was also found to containlarger amounts of beta-(1,3)-glucan than the wild type.Interestingly, it has been determined that the alpha-glucanserves as an endogenous carbon source during the stationaryphase of growth (191, 272). While the latter finding impliesthat this polymer is less critical for maintenance of thestructural integrity of the hyphal wall, other evidence sug-gests that alpha-(1,3)-glucan, together with melanin, serves
VOL. 6, 1993
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.

CLIN. MICROBIOL. REV.
to protect hyphae from the action of hydrolytic enzymes,i.e., chitinase and beta-glucanase (191). In the case of theglucan, the latter property would presumably be a conse-quence of the physical barrier formed by this polymer incovering the inner layers of chitin and beta-(1,3)-glucan. Therole of melanin in fungi is reviewed by Wheeler and Bell(263).While little specific evidence is available, it is generally
assumed that newly inserted wall material is not extensivelycross-linked and therefore represents a weak spot in thewall. Support of this hypothesis was provided by Polacheckand Rosenberger (192), who determined that autolytic en-zymes produced byA. nidulans disproportionately degradednew wall material in the hyphal apex in comparison withareas of mature cell wall.Two ultrastructural studies of A. fumigatus found no
evidence for multiple layers in the cell wall per se, thoughflocculent electron-dense material covering the outer wallwas noted (41, 197). One of the reports, however, stated thatthe conidia had two wall layers (197). Treatment of hyphaeofA. fumigatus and Aspergillus niger with the lytic enzymesbeta-glucuronidase and chitinase caused the release of re-ducing and N-acetylated sugars (244). The combination ofthe two enzymes was much more efficient in releasingprotoplasts than use of beta-glucuronidase alone and was notsignificantly enhanced by the addition of pronase. Theseresults confirm the importance of beta-glucan and chitin inmaintaining the rigidity of the cell wall.
In a study characterizing the major polymers in the wall ofA. niger, Stagg and Feather (229) found substantial amountsof chitin and determined that there was a 4:1 ratio ofalpha-glucan to beta-glucan.With regard to the antigenic components of the Aspergil-
lus cell wall, key studies agree that the immunodominantantigens of the cell wall for both A. fumigatus and A. nigerare composed of galactomannan (10, 11, 18) and are sensitiveto the action of pronase (105). The mannan component isprimarily composed of (1,6)-linked residues with (1,2) sidechains, while the beta-D-galactofuranosyl units are (1,6)linked to the mannan core (11).A chemical analysis of the dermatophytic fungi Trichophy-
ton mentagrophytes, Microsporum canis, Microsporumgyp-seum, and Epidermophyton floccosum revealed very similarprofiles of composition among these species, with N-acetyl-hexosamines accounting for 26 to 31% of the wall mass,glucose at 36 to 46%, mannose ranging from 8 to 11%, andprotein at approximately 7% (218). Noguchi et al. performeda carbohydrate analysis on 13 species of dermatophytes andreported findings similar to those mentioned above (175).Specific determination of the wall polymers in T. mentagro-phytes showed similarities to the polymers in Aspergillusspp. in that galactomannans, beta-(1,3)- and beta-(1,6)-glu-cans, and chitin were present. The authors were able todemonstrate the ultrastructural existence of multiple layersin the wall (139), a finding confirmed by others (175). Themicroconidial wall of T. mentagrophytes was found to beremarkably similar to the hyphal wall in chemical composi-tion and ultrastructure (265). Using microconidia, the au-thors determined that the outer wall layer consists of aglycoprotein-lipid complex, the middle layer is made of aproteinaceous rodlet layer, and the inner wall is largelycomposed of a complex of beta-(1,3)-glucan and chitin. Aspecific analysis of the chitin from the cell wall and septaof T. mentagrophytes arthroconidia demonstrated the pres-ence of the typical microfibrillar form of this polymer, butthe authors also found a previously uncharacterized non-
fibrillar chitin that was resistant to bacterial chitinase (194).The latter point would have obvious implications in theability of fungal spores to resist degradative enzymes in thesoil.
Dimorphic Pathogenic Fungi
Blastomyces dermatitidis, Coccidioides immitis, His-toplasma capsulatum, and Paracoccidioides brasiliensis arethe etiologic agents of what are known collectively as theendemic mycoses. These diseases are essentially restrictedto the Western Hemisphere because of ill-defined but abso-lute ecologic requirements of the respective fungal agents.While each of these fungi exists in a filamentous form in itsnative state in the soil, chance encounters with mammalianhosts lead to a conversion to the parasitic phase. While B.dermatitidis, H. capsulatum, and P. brasiliensis grow in ayeast form during infection (or under permissive conditionsin vitro), Coccidioides immitis has a unique endospore-spherule life cycle.The cell wall of P. brasiliensis is composed primarily of
carbohydrates (50 to 80% dry weight) arranged as a singlelayer in the mycelial phase and in two layers in the yeastphase (42). Chitin constitutes 13% of the mycelial form and34% of the yeast phase, being located in the inner wall (43).Beta-(1,3)-glucan accounts for most of the carbohydratefound in the mycelial wall (38%), but there is significantly lessin the yeast wall (6%). Interestingly, while absent in themycelial wall, alpha-(1,3)-glucan is the major polymer in theyeast wall (45%), being found only in the outer layer (43, 132).A fourth polysaccharide, galactomannan, is found in bothgrowth phases and forms about 6% of the total wall. Theremainder of the wall is composed of protein and lipids (5). Aconnection between the alpha-(1,3)-glucan component andthe pathogenic potential of this fungus was discovered when astrain that had been serially passaged over a number of yearslost its virulence in a hamster model as a result of an almostcomplete loss of the alpha-glucan from the cell wall (208).The gross composition of the cell wall from both growth
phases of B. dermatitidis is rather similar to that found withP. brasiliensis. Chitin is found as approximately 14% of themycelial phase but increases to 48% of the yeast phase (132).The overall hexose content is comparable in both growthforms, but the type of glucan differs. While the yeast-phaseglucan is approximately 95% alpha-glucan and 5% beta-glucan, the mycelial phase was found to contain 60% alpha-glucan and 40% beta-glucan (130). Kanetsuna and Carbonell(130) reported that the glucans existed as alpha-(1,3)- andbeta-(1,3)-glucans, respectively.H. capsulatum is unique in that isolates in the yeast phase
can be segregated into two chemotypes by the compositionof the cell wall (73, 190). Chemotype I was found to containapproximately 40% of its wall mass as chitin, while chemo-type II had less than 15% of this polymer. The other majordifference was that, while the glucan in chemotype I is betalinked and has no alpha-(1,3) linkages, chemotype II hasapproximately 60% of its glucan in the alpha-(1,3) configu-ration and the remainder is beta-glucan (73, 131, 198, 199).The yeast phases of both chemotypes have rather smallamounts of mannan and amino acids. The mycelial phase ofchemotype I has smaller amounts of chitin (4%) but a higheramount of amino acids (73). Kanetsuna et al. (131), however,reported that the mycelial phase of a chemotype II isolatehad 26% chitin and 19% glucan, the latter being mostlybeta-glucan. In several of these studies, the authors thought
4 HECTOR
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.

COMPOUNDS ACTIVE AGAINST CELL WALLS OF FUNGI 5
that the beta-glucan and chitin were likely in close associa-tion and perhaps even linked in some fashion.As was determined for C. albicans and S. cerevisiae, at
least two genes encoding chitin synthetases have beendiscovered in B. dermatitidis and H. capsulatum (26). Whenthe sequences of these genes were compared, Bowen et al.found a high degree of homology in the two species, suggest-ing a strong phylogenetic relationship.
In a situation analogous to that previously described for P.brasiliensis, avirulent isogenic strains of H. capsulatumwere analyzed and found to contain up to 1,000-fold lessalpha-(1,3)-glucan than the fully virulent parental strains(137). However, these findings stand in contrast to those forchemotype I strains previously described, which still re-tained virulence (73).With Coccidioides immitis, chemical analyses of the cell
wall have been performed on both the spherule-endosporeand mycelial phases and on arthroconidia. In a series ofpapers, Wheat et al. (260-262) determined that the spherulewall contained 4 to 15% chitin, the mycelia had 5 to 22%,chitin and arthroconidia had 10% of their cell wall mass aschitin. Spherules were found to have a total neutral hexoseconcentration of 66% glucose, 27% mannose, less than 10%3-methylmannose, and a trace of galactose. The mycelialphase had 33% glucose, 46% mannose, 12% galactose, andless than 10% 3-methylmannose. The hexose content ofarthroconidia was similar to that of the mycelial phase.Together, amino acids and N-acetylglucosamine were foundto constitute 40 to 60% of the isolated cell walls. Wheatfound that treatment of the mycelial and arthroconidial wallswith beta-(1,3)-glucanase released 20 and 16%, respectively,of the wall weights as glucose. Importantly, when Collinsand Pappagianis (57) treated endospores with a commer-cially prepared chitinase (which contained beta-glucanase aswell), the enzymes were able to digest 6.5% of the cell mass.However, when mature spherules were used, the chitinaseliberated 24.8% of the cell mass. In an earlier report, thesame authors noted that, when mature spherules were heldin phosphate buffer, the wall appeared to undergo thinningjust prior to release of their endospores, leading to specula-tion that the walls may contain an endogenous chitinase-likeenzyme (58). This was found to be the case several yearslater when synchronously growing cultures of the spherule-endospore phase were found to produce chitinase during theperiod of disruption of the mature spherule, resulting in arelease of the progeny endospores (111).When various growth stages of the spherule-endospore
form of Coccidioides immitis were subjected to enzymaticdegradation followed by transmission electron microscopy,it was discovered that the cell wall of the maturing spheruleundergoes dramatic changes, particularly during the processof endosporulation (108). As is the case for many other fungi,the inner layers of the cell wall were found to contain mainlychitin and beta-(1,3)-glucan. The outer layers were predom-inantly alpha-(1,3)-glucan, and mannoprotein was thought tobe located throughout the cell wall. At the point of endos-porulation, the emerging cleavage planes were determined tobe essentially pure chitin, with beta- and alpha-glucans andmannoprotein added at later points during the maturation ofthe planes into endospore walls. As mentioned previously, atthe point of maturity, the production of chitinase degradesmuch of the chitin in the cell walls, aiding the disruption ofthe spherule (111).
COMPOUNDS ACTIVE AGAINST FUNGAL CELLWALLS
Inhibitors of Chitin Synthesis
History of polyoxins and nikkomycins. Two structurallyrelated groups of fermentation secondary metabolites thatact as specific inhibitors of chitin synthetase (UDP-2-aceta-mido-2-deoxy-D-glucose:chitin 4-beta-acetamidodeoxy-D-glucosyltransferase; EC 2.4.1.16) have been described. Thepolyoxins, first described in 1965 (238), and the nikkomy-cins, described in 1976 (63), are peptide-nucleoside com-pounds produced by Streptomyces cacaoi and Streptomycestendae, respectively. Both groups of compounds were iso-lated and described as a result of programs to discoverfungicides and insecticides for agricultural use (63, 124).The polyoxins were discovered in a screen against Pellic-
ulariafilamentosa f. sp. sasakii, which causes a sheath blightin rice plants. A crude mixture of polyoxins was eventuallymarketed as a fungicide and saw use in a variety of agricul-tural applications. The first producing strain for the nikko-mycins, Streptomyces tendae TU 901, was isolated in 1970from a soil sample collected at the site of the famousfive-story pagoda at the shrine in Nikko, Japan. Although thenikkomycins have been extensively evaluated for commer-cial development, no product has reached the marketplace.Subsequent to the discovery of the nikkomycins, a subspe-cies of S. cacaoi was discovered to produce a number ofsecondary metabolites termed neopolyoxins that were es-sentially identical in structure to the nikkomycins (252, 253).To date, 13 naturally occurring polyoxins, polyoxins A
through M, and 14 naturally derived nikkomycins, nikkomy-cinsB~,B3, C, , Cz, D, E, I, J, M, N, X, Z, pseudo-J, andpseudo-Z, have been described (63, 99-101, 114, 124-127,142, 143).
Biochemistry of polyoxins and nikkomycins. The polyoxinsand nikkomycins collectively include several metabolitecongeners that bear striking structural similarity to UDP-N-acetylglucosamine, the precursor substrate for chitin (Fig.1). The biologically active compounds from each group,composed of unique di- and tripeptides made up of alpha-L-amino acids linked to native or substituted uridine (or toformylimidazoline in the case of nikkomycin X), were foundto act as competitive inhibitors of fungal chitin synthetase(82, 83, 172). These substances are unusual in that otherpreviously described aminoacyl nucleoside antibiotics act byan inhibition of protein synthesis (reviewed in reference 87).Overall, the polyoxins and nikkomycins have rather similarin vitro potency profiles against isolated chitin synthetasesfrom a variety of fungal sources, with reported Ki values of0.6 ,uM for polyoxin A, 0.5 ,uM for nikkomycin X, 2 to 3.5,uM for nikkomycin Z (97, 172), 0.6 to 3.0 ,uM for polyoxin D(14, 92), and 32 ,uM for polyoxin B (97). Given that all ofthese data were generated before the existence of multipleforms of chitin synthetase was discovered, it can only beassumed that the values represent an average from thesedifferent enzymes. The abilities of these compounds toinhibit chitin synthesis is not limited to fungi, however, as itwas determined that chitin synthetase isolated from theinsect Tribolium castaneum was inhibited by nikkomycinand polyoxin D, with calculated Kis of 0.02 and 4 ,uM,respectively (55). Indeed, a mixture of nikkomycin X and Zwas evaluated for potential use as an insecticidal agent (117,270).The polyoxins have been thoroughly studied with respect
to their actual mechanisms of action, with the structure-
VOL. 6, 1993
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.
CLIN. MICROBIOL. REV.
a
b
c
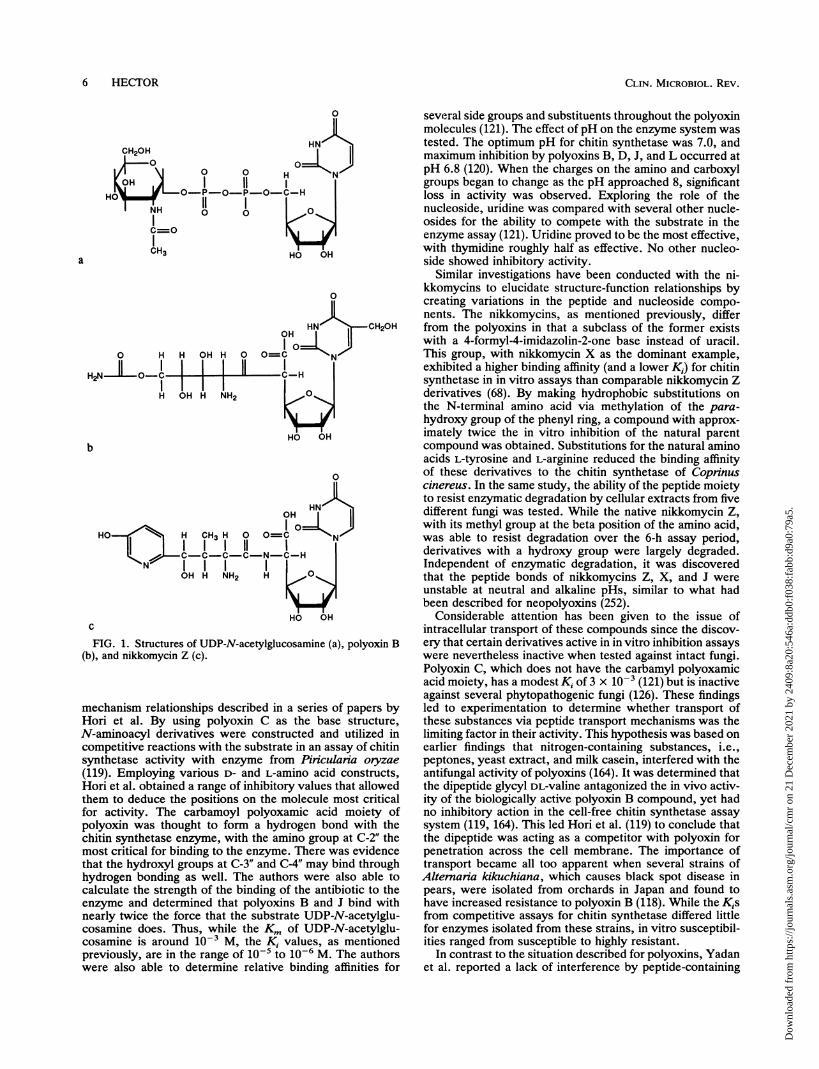
FIG. 1. Structures of UDP-N-acetylglucosamine (a), polyoxin B(b), and nikkomycin Z (c).
mechanism relationships described in a series of papers byHori et al. By using polyoxin C as the base structure,N-aminoacyl derivatives were constructed and utilized incompetitive reactions with the substrate in an assay of chitinsynthetase activity with enzyme from Piniculania oryzae
(119). Employing various D- and L-amino acid constructs,Hori et al. obtained a range of inhibitory values that allowedthem to deduce the positions on the molecule most criticalfor activity. The carbamoyl polyoxamic acid moiety ofpolyoxin was thought to form a hydrogen bond with thechitin synthetase enzyme, with the amino group at C-2" themost critical for binding to the enzyme. There was evidencethat the hydroxyl groups at C-3" and C-4" may bind throughhydrogen bonding as well. The authors were also able tocalculate the strength of the binding of the antibiotic to theenzyme and determined that polyoxins B and J bind withnearly twice the force that the substrate UDP-N-acetylglu-cosamine does. Thus, while the Km of UDP-N-acetylglu-cosamine is around i0-' M, the Ki values, as mentionedpreviously, are in the range of o-5 to 10-6 M. The authorswere also able to determine relative binding affinities for
several side groups and substituents throughout the polyoxinmolecules (121). The effect of pH on the enzyme system wastested. The optimum pH for chitin synthetase was 7.0, andmaximum inhibition by polyoxins B, D, J, and L occurred atpH 6.8 (120). When the charges on the amino and carboxylgroups began to change as the pH approached 8, significantloss in activity was observed. Exploring the role of thenucleoside, uridine was compared with several other nucle-osides for the ability to compete with the substrate in theenzyme assay (121). Uridine proved to be the most effective,with thymidine roughly half as effective. No other nucleo-side showed inhibitory activity.
Similar investigations have been conducted with the ni-kkomycins to elucidate structure-function relationships bycreating variations in the peptide and nucleoside compo-nents. The nikkomycins, as mentioned previously, differfrom the polyoxins in that a subclass of the former existswith a 4-formyl-4-imidazolin-2-one base instead of uracil.This group, with nikkomycin X as the dominant example,exhibited a higher binding affinity (and a lower K1) for chitinsynthetase in in vitro assays than comparable nikkomycin Zderivatives (68). By making hydrophobic substitutions onthe N-terminal amino acid via methylation of the para-hydroxy group of the phenyl ring, a compound with approx-imately twice the in vitro inhibition of the natural parentcompound was obtained. Substitutions for the natural aminoacids L-tyrosine and L-arginine reduced the binding affinityof these derivatives to the chitin synthetase of Coprinuscinereus. In the same study, the ability of the peptide moietyto resist enzymatic degradation by cellular extracts from fivedifferent fungi was tested. While the native nikkomycin Z,with its methyl group at the beta position of the amino acid,was able to resist degradation over the 6-h assay period,derivatives with a hydroxy group were largely degraded.Independent of enzymatic degradation, it was discoveredthat the peptide bonds of nikkomycins Z, X, and J wereunstable at neutral and alkaline pHs, similar to what hadbeen described for neopolyoxins (252).
Considerable attention has been given to the issue ofintracellular transport of these compounds since the discov-ery that certain derivatives active in in vitro inhibition assayswere nevertheless inactive when tested against intact fungi.Polyoxin C, which does not have the carbamyl polyoxamicacid moiety, has a modest Ki of 3 x 10-3 (121) but is inactiveagainst several phytopathogenic fungi (126). These findingsled to experimentation to determine whether transport ofthese substances via peptide transport mechanisms was thelimiting factor in their activity. This hypothesis was based onearlier findings that nitrogen-containing substances, i.e.,peptones, yeast extract, and milk casein, interfered with theantifungal activity of polyoxins (164). It was determined thatthe dipeptide glycyl DL-valine antagonized the in vivo activ-ity of the biologically active polyoxin B compound, yet hadno inhibitory action in the cell-free chitin synthetase assaysystem (119, 164). This led Hori et al. (119) to conclude thatthe dipeptide was acting as a competitor with polyoxin forpenetration across the cell membrane. The importance oftransport became all too apparent when several strains ofAltemaria kikuchiana, which causes black spot disease inpears, were isolated from orchards in Japan and found tohave increased resistance to polyoxin B (118). While the Kisfrom competitive assays for chitin synthetase differed littlefor enzymes isolated from these strains, in vitro susceptibil-ities ranged from susceptible to highly resistant.
In contrast to the situation described for polyoxins, Yadanet al. reported a lack of interference by peptide-containing
6 HECITOR
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.

COMPOUNDS ACTIVE AGAINST CELL WALLS OF FUNGI 7
medium in susceptibility tests using nikkomycin Z against C.albicans (266). It was subsequently discovered by thoseauthors that in this yeast at least two distinct peptidetransport systems facilitate entry of di- and tripeptidesintracellularly and that nikkomycin Z is transported by atleast one of these systems. The mechanism for transport ofpolyoxins, however, is less clear, but data generated withpolyoxin derivatives suggest a peptide transport mechanismas well (173). McCarthy et al. (157) confirmed the existenceof two peptide permeases in C. albicans but reported thatpeptides in the growth medium could interfere with nikko-mycin activity. However, this effect was restricted to certaindipeptides with a stereospecific configuration that werecandidates for active peptide transport by the identifiedpermeases. Interestingly, the converse was also true; nikko-mycins could serve as antagonists to dipeptide uptake. Thepeptide permeases of C. albicans have been further charac-terized in additional reports by McCarthy et al. (155, 156),Shallow et al. (219), Payne et al. (182), and Logan et al.(151).The concomitant findings that peptide transport was in-
volved in the uptake of nikkomycins and polyoxins and C.albicans was not particularly susceptible to the effects ofthese substances led to considerable efforts by severalgroups to obtain active derivatives of these compounds. Therationale was that since the isolated chitin synthetase of thisyeast was susceptible to these compounds in a submilligramrange and the susceptibility of intact yeasts was in the highmilligram range, inadequate uptake of these substancesmight be the limiting factor. In the case of the polyoxins,several derivatives with different amino acid or amino fattyacid compositions were prepared with the hope of improvinguptake and biological activity (81, 138, 144, 221, 222). Withthe nikkomycins, the group led by H. Zahner used a differentapproach in that they first mutagenized the producing strainof Streptomyces tendae and then ran a directed fermentationproviding pyrimidines, purines, imidazoles, or amino acids(24, 25, 66, 67, 69, 101, 114). In this way, new derivativesinvolving substitutions for both the nucleoside and thepeptide moieties were obtained. With both approaches,although several candidates with improved properties intransport, stability, or in vitro inhibition of chitin synthetasewere obtained, none of the derivatives was sufficiently moreactive than the parent compounds against intact fungal cellsto justify their development. In searching for reasons toexplain the lack of improved activity after such herculeanefforts, Khare et al. (138) stated that, among other reasons,inhibition of chitin synthesis may not be sufficient to killfungal cells. Results from susceptibility tests and efficacyexperiments, described below, will demonstrate that this isthe case for certain fungi but not for others.
Complicating the issue of differing susceptibilities is arecent report that suggests that, with S. cerevisiae, the geneproducts of CHSI and CHS2 show differential susceptibili-ties to the inhibitory effects of polyoxins and nikkomycins,with Chsl being more susceptible (34). As it has beenconfirmed that several different medically important fungialso have multiple forms of chitin synthetase (26), it is anobvious statement that a determination of the relative im-portance of each form to the growth of the cell must bemade. Combining this information with data on susceptibil-ities of the different enzymes to polyoxins and nikkomycinsmay explain the broad range in susceptibilities that will bediscussed below.
Lastly, some experimentation to elucidate factors govern-ing genetic control of nikkomycin synthesis in Streptomyces
tendae have been reported, but no specific genes have beenidentified (84, 85). The ability to selectively control theexpression of the more active forms of nikkomycin wouldgreatly improve the economics of this fermentation product.
Use of polyoxins and nikkomycins against medically impor-tant fungi. The polyoxins were first described in 1965 (238),and reports on their effects on a variety of saprophytic fungiappeared shortly thereafter (14, 27, 77, 82, 118, 210). De-scriptions of swollen, protoplast-like structures in cellstreated with these substances suggested an effect not unlikethat seen with bacteria treated with beta-lactam antibiotics.As early as 1977, inhibitors of chitin synthesis were pro-posed as therapeutic agents for fungal diseases (91), but itwas not until 1983 that the first reports on the effects ofpolyoxin D on medically important fungi appeared. Onereason for this delay may have been the perception thatthese substances were, for the most part, inactive againstsaprophytic yeasts, with a difference of 3 orders of magni-tude between the Kis for inhibition of isolated chitin syn-thetase and the concentration necessary to kill the cells (27,36, 125, 135). With the discovery, however, that polyoxin Dwas active in vitro against the parasitic phase of Coccid-ioides immitis in the microgram-per-milliliter range, the useof these agents as antifungal drugs seemed plausible.Hector and Pappagianis (109) found that immature spher-
ules of Coccidioides immitis were highly and rapidly suscep-tible to the effects of polyoxin D, exhibiting morphologiceffects consistent with an osmotically sensitive state. Within9 h after exposure to this substance, these osmoticallysensitive cells invariably continued to swell until they burst.At higher concentrations of polyoxin D, i.e., 200 ,ug/ml, thecompound appeared to cause death directly without the cellsswelling. When cells were treated with approximately 50ug/ml for longer periods, an inhibition of endosporulationensued, in addition to swelling. This inhibition was mani-fested by either a complete lack of cleavage plane formationor the appearance of fewer, irregularly shaped endospores.Interestingly, in contrast to the dramatic effects seen withthe spherule-endospore phase of this fungus, the mycelialphase was able to germinate and grow in the highest con-centrations of polyoxin D tested with no apparent differ-ences distinguishable by light or transmission electron mi-croscopy.
Shortly after the appearance of the report by Hector andPappagianis, results from in vitro studies using polyoxin Dagainst C. albicans and Cryptococcus neofonnans werepublished (16). Four strains of C. albicans were found to besusceptible in the range of 500 to 2,000 ,ug/ml, with treatedcells manifesting effects such as swelling and "chaining" ofcells. Examination of cells treated with the chitin-stainingdye calcofluor white revealed that treated cells had far lessfluorescence than control cells, indicating reduced levels ofchitin incorporated into the cell wall. In particular, theauthors noted the complete lack of fluorescence at the site ofseptum formation, an area normally rich in chitin. Theauthors also describe the chaining phenomenon for S. cere-visiae, which suggests that chitin synthesis in the septum iscritical if the cells are to separate properly. The sequentialaddition of buds to form the chain may be an attempt by thecell to cope with the loss of proper septum formation.Importantly, under conditions of culture that promotedgermination, polyoxin D was able to block germ tube forma-tion with C. albicans, perhaps a result of the higher level ofchitin synthesis needed for the mycelial phase (30, 50). In afollow-up study in which labeled lectin was used to localizechitin in an examination by transmission electron micros-
VOL. 6, 1993
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.

CLIN. MICROBIOL. REV.
copy, treatment of both yeast and mycelial forms of C.albicans with polyoxin D led to the disappearance of chitinin the inner cell wall layers and a lack of septum formation(115). With prolonged incubation, the successive cells in achain grew progressively larger and had thinner walls,suggesting that chitin also plays a role in maintaining thestructural integrity of the entire wall. Last, the authors alsofound Cryptococcus neoformans to be susceptible to theeffects of polyoxin D but concluded that the compound wasfungistatic in nature.By using the polyoxin complex AL (a crude mixture of
polyoxins) on the polymorphic phaeohyphomycosis fungusWangiella dermatitidis, morphologic effects similar to thosedescribed with treated cultures of both Coccidioides immitisand C. albicans were obtained (59). Treatment of wild-typeyeast cells by the drug complex was found to cause irregularbudding and defective septum formation. Exposure of atemperature-sensitive mutant (an isolate used to demon-strate the multicellular form of this fungus) to polyoxin ALinhibited conversion to the multicellular form, an effect notdissimilar to the inhibition of endosporulation of Coccid-ioides immitis. The resulting aberrant form also had areduced ability to bind calcofluor white and was osmoticallysensitive.
Recently, in vitro results with nikkomycins X and Zagainst diverse fungi were reported (113, 185). While thedimorphic fungi Coccidioides immitis and B. dermatitidiswere found to be highly susceptible, C. albicans and Cryp-tococcus neoformans had only moderate susceptibility tothese compounds. Fusarium solani, Fusarium oxysporum,Rhizopus arrhizus, A. fumigatus, and, interestingly, Can-dida tropicalis were found to be resistant to nikkomycins Xand Z at concentrations ranging up to 1 to 8 mg/ml. Whileone could speculate that C. albicans might be less suscepti-ble than Coccidioides immitis because of a lower chitincontent, the reduced susceptibility of Cryptococcus neofor-mans and the mycelial fungi, forms with more chitin than C.albicans, indicates that the issue is more complex. Theextreme resistance of C. tropicalis is also puzzling. What isclear is that for the susceptible fungi chitin is an essentialcomponent of the cell wall and that its inhibition has negativeconsequences. The inhibition of septum formation, as man-ifested in the traditional sense in yeasts or in a modified formas in the process of endosporulation in Coccidioides immitis,directly interferes with the ability of these cells to reproduce.When cell wall synthesis is inhibited, the effects may resultin the rapid disintegration of the cell, as with Coccidioidesimmitis; may lead to aberrant forms such as was seen withyeasts; or may have few discernible effects, as with thefilamentous fungi.More recently, Gottlieb et al. (96) described an intriguing
physiologic consequence of treatment of C. albicans withpolyoxin D. This group discovered that cells treated withthis compound were less able to bind to buccal epithelialcells, with a reduction in binding of as much as 58% incomparison with that of normal cells. These results are allthe more fascinating when one considers that, with thepossible exception of exposed bud scars, all data indicatethat chitin is restricted to the inner layers of the cell wall ofthis yeast and should therefore not be accessible for bindinginteractions. For a review of the factors governing adher-ence of C. albicans, see the report by Calderone and Braun(40).With respect to the activity of the chitin synthetase
inhibitors in animal models of mycoses, the number ofreports is limited. In one study, Becker et al. described
experiments in which polyoxin D and a mixture of nikkomy-cins X and Z were used parenterally in a systemic model ofcandidiasis in mice (17). They found that, while polyoxin Doffered no protection in comparison with controls, a combi-nation of nikkomycins X and Z was able to delay though notprevent deaths. In contrast, Hector et al. found nikkomycinsX and Z to be highly efficacious in mouse models ofcoccidioidomycosis and blastomycosis and moderately effi-cacious against histoplasmosis (112). When given orally, thecompounds were able to completely prevent deaths in miceinfected with a 100% lethal challenge, though nikkomycin Zwas found to be considerably more active than nikkomycinX. The fungicidal nature of nikkomycin Z was proven when,in short-term organ load experiments with models of blasto-mycosis and coccidioidomycosis, the target organs werefound to be essentially sterilized by twice-daily (b.i.d.) dosesof only 50 mg/kg of body weight given for 5 days. Overall,the results with nikkomycin Z were superior to those withgroups given relevant doses of several azoles or amphoteri-cin B. In a later paper, the fungistatic nature of nikkomycinZ against systemic candidiasis was confirmed in a mousemodel in animals dosed with 5 to 50 mg/kg b.i.d. (110).Surprisingly, the authors noted a lack of a dose-responseeffect with the doses employed.
Nonspecific inhibitors of chitin synthesis. A number ofcompounds have been described that have an effect on chitinsynthesis, though it is usually as a consequence of a second-ary mechanism. However, several groups have sought toexploit these effects in the search for more effective fungi-cides and insecticides. One of the first reports on this subjectdescribed several new insecticidal agents belonging to aclass called benzoylphenyl ureas. These agents had noimmediate effect on insects, but treatment resulted in asubsequent disturbance in cuticle deposition and an abortivemoult (259). Although the authors did not recognize that aninhibition of chitin synthesis was responsible for theseeffects, the potential for these agents as pesticides andfungicides was ultimately recognized and described. Whilethe action of the benzoylphenyl ureas on insect cuticles wasclaimed to be identical to that seen with polyoxin D (227),others thought that the specific effects of this class of agentwere a result of an inhibition of serine protease activity thatblocked activation of the chitin synthetase zymogen (150).These and other chymotrypsin inhibitors were demonstratedto prevent chitin synthesis in a cockroach system.To test these agents in a fungal system, examples of
benzoylphenyl ureas, trichloromethylsulfonyls, and organo-phosphorous compounds were compared with nikkomycinin an in vitro chitin synthetase assay that used enzyme fromCoprinus cinereus (31). None of the pesticides had anyinhibitory activity in the fungal system, while nikkomycin Zwas active in the expected micromolar range. Ultimately, itwas confirmed through the use of an insect chitin synthetasesystem that the pesticides did not act as specific inhibitors ofthe enzyme (55). Despite this, a natural product with insec-ticidal activity that the authors claimed acted as a specificinhibitor of chitin synthetase activity from an insect sourcewas described. Plumbagin, which is isolated from the Afri-can medicinal plant Plumbago capensis, had activity com-parable to that of polyoxin D in vitro and caused inhibition ofecdysis in several lepidopterous pests (145). In a subsequentstudy, however, plumbagin was found to inhibit cytochromeP-450-dependent ecdysone 20 monooxygenase activity inadult mosquitoes and larvae of Drosophila melanogaster atsignificantly lower concentrations than had been reported forchitin inhibition, leading Mitchell and Smith to conclude that
8 HECTOR
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.

COMPOUNDS ACTIVE AGAINST CELL WALLS OF FUNGI 9
inhibition of chitin synthesis was a secondary effect of thissubstance (165). Thus, because of the general lack of spec-ificity of these agents, it seems unlikely that the myriad ofinsecticides could serve as a source of antifungal drugs.A number of compounds that affect the synthesis or
structure of components of the cytoplasmic membrane areknown to affect chitin synthesis in fungi. One of the earliestfindings on this subject resulted from attempts to solubilizechitin synthetase from membrane preparations by using thedetergent digitonin. Depending on the concentration em-ployed, this saponin was found to both stimulate and inhibitthe action of fungal chitin synthetase from chitosomes fromMucor rouxii (203). This led to a study of the effects on chitinsynthesis of the traditional antifungal agents amphotericin B,nystatin, pimaricin, and filipin, all known to bind to sterols(196). At concentrations greater than 10,ug/ml, amphotericinB and nystatin acted as noncompetitive inhibitors of chitinsynthetase, while the other compounds were less effective.Given that the MICs of these substances are fully 1 order ofmagnitude lower than the concentrations necessary to causethis effect, any contribution to the antifungal effects by thismechanism would be minor. However, this study was im-portant in that it established that substances affecting themembrane may have secondary effects on chitin synthesis.This has been shown to be true for other membrane-activeantibiotics as well. Studies of the effects of azoles on chitinsynthesis in vivo in C. albicans have demonstrated thatwithin certain ranges of concentrations a net stimulation ofchitin synthesis can ensue (107, 187, 255). In the case of theazoles, the depletion of ergosterol in the cytoplasmic mem-branes of treated cells likely results in a secondary effect onthe activity of the membrane-bound chitin synthetase, per-haps by activation of the zymogenic form of this enzyme(255).
In a recent study, the agricultural fungicide edifenphos,which is known to affect membrane phospholipid composi-tion, was found to cause noncompetitive inhibition of chitinsynthetase from Fusarium graminearum within specific con-centration ranges (21).Two compounds that have an effect on chitin synthesis by
inhibition of other enzyme systems are tetaine and tunicamy-cin. Tetaine, an inhibitor of glucosamine-6-phosphate syn-thetase, affects chitin and mannan but not glucan synthesisin C. albicans, with the effects on the mycelial form morepronounced than those on the yeast form (161). In the case oftunicamycin, while an earlier paper reported that the com-pound was a specific competitive inhibitor of Neurosporacrassa (217), a later study using the substance in an insectsystem found that it acts by blocking the synthesis of alipid-linked saccharide intermediate involved in protein-chitin complexes (264).New compounds. Two recently described secondary me-
tabolites of fermentation that have antifungal activity de-serve mention within the context of chitin synthetase inhib-itors. One, FR-900403, a peptide-nucleoside antibiotic,differs from the polyoxin-nikkomycin complex in that thenucleoside is adenosine and the peptide is linked to thenucleoside by the C-3' residue (128). The compound isreported to be active against C. albicans but not againstfilamentous fungi. The second compound, FR-900848, is anucleoside-fatty acid compound with uridine linked to amonounsaturated fatty acid containing five cyclopropanerings (268). This compound was found to be active againstfilamentous fungi but had no activity against yeasts. Whilethe mechanism(s) of action has not been reported for eitherof these compounds, FR-900848 was reported to cause
swelling and multiple branch points in filamentous fungi,suggesting an effect on the cell wall. Thus, these reportsdemonstrate that active nucleoside-based compounds otherthan nikkomycins and polyoxins can be identified.
Inhibitors of chitinases. In the previous sections on cellwall biochemistry, the importance of a balance betweenchitin synthesis and chitin degradation during growth, repro-duction, and separation was discussed. Just as inhibitors ofchitin synthesis have been shown to possess antifungalproperties, a class of chitinase inhibitors has been describedand found to have affects on whole cells. Allosamidin,derived from the fermentation of a Streptomyces isolate notidentified to species level, was first reported as an inhibitorof insect chitinase (140, 205, 206). The purified material wasreported to inhibit the enzyme from the silkworm Bombyxmon in a competitive fashion in the micromolar range andwas lethal to caterpillars at a dose of 10 ,ug per larva.Subsequently, additional allosamidins, some of which wereconsiderably more active than the original compound, wereidentified and isolated (174, 207). Demethylallosamidin wasfound to inhibit the chitinase of S. cerevisiae at a levelcomparable to that described for the insect system and alsointerfered with the cell division of this yeast (207). Cellstreated with 20 ,ug of this compound per ml grew in clumpsthat did not separate, though the culture continued to growwithout inhibition. The 50% inhibitory concentrations forseveral other congeners against chitinase from C. albicanswere determined to be in the microgram-per-milliliter range.The activity of allosamidin against the chitinase from C.
albicans has been studied, with a 50% inhibitory concentra-tion of 0.3 ,uM calculated (71). The inhibition was deter-mined to be of a competitive type, with a Ki of 0.23 ,uM. Aswas the case with S. cerevisiae, use of this material againstC. albicans had no effect on cell numbers in a broth culture.Thus, while the use of these inhibitors singly as antifungaldrugs seems to be without merit, the possibility of combiningthem with other cell wall-active agents to complement theiractivities is intriguing.
Inhibitors of Glucan Synthesis
History of aculeacins, echinocandins, and papulacandins.To date, three groups of compounds that are specific inhib-itors of fungal glucan synthesis have been described. Theaculeacins, echinocandins, and papulacandins were all dis-covered as fermentation metabolites during screening pro-grams for new antibiotics. The aculeacins, isolated fromAspergillus aculeatus, were described in 1977 as a complexof seven lipoprotein compounds: aculeacins A, B, C, D, E,F, and G (168, 211). Aculeacin A (Fig. 2) was identified asthe major product, accounting for 90% of the production,though aculeacin D was initially characterized as having thestrongest activity. The physical properties have not beenconmpletely described for each component of this complex.The echinocandins, a complex of B, C, and D, are
hexapeptide-lipid antibiotics that are structurally similar tothe aculeacins (Fig. 3). Echinocandin B, first described in1974, is the major product of the complex produced by somespecies of A. nidulans and Aspergillus rugulosus (19, 136,246).The papulacandins, first described in 1977, are a complex
of five compounds, i.e., papulacandins A, B, C, D, and E,isolated from a strain of Papulania sphaerospenna (98, 248).These amphophilic compounds have a unique structure,described as a spirocyclic diglycoside esterified by twolong-chain unsaturated fatty acids (Fig. 4) (247). With the
VOL. 6, 1993
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.

CLIN. MICROBIOL. REV.
HO H0 N-OH H' H H
\O / H '
OH
H OHFIG. 2. Struct
exception of papulacandin D, the compounds differ from oneanother in the degree of unsaturation and in the numbers andpositions of hydroxyl groups on the fatty acid moieties. Withpapulacandin D, a sugar and a fatty acid group are missing.
Biochemistry of beta-glucan inhibitors. Unlike the polyox-ins and nikkomycins, the structures of the beta-glucaninhibitors do not provide clues as to their mechanisms ofaction. All are amphophilic, having either an amino acid ringor sugar residues to which one or more lipid chains areattached. The lipid chains appear to be essential for theiractivity; when the linolenic acid moiety was deacylated fromechinocandin B, the remaining portion was inactive (65).However, the ability to substitute other lipids has led to thecreation of several active compounds of both echinocandin
ture of aculeacin A.
(159, 160) and papulacandin (249, 256). Among the echino-candins, the various congeners have some differences in theamino acid makeup of the cyclic peptide (246). Similarly,some natural derivatives of echinocandin B from othermicrobial sources also have differences in amino acid com-position (7).
Several studies with one or more of these substances haveled to the conclusion that they all act as specific inhibitors ofbeta-(1,3)-glucan synthetase (EC 2.4.1.34; UDP-glucose:1,3-beta-D-glucosyltransferase). The first reports, published in1977, made the general claim that papulacandin B andaculeacin A were able to inhibit synthesis of the structuralglucan component of C. albicans and S. cerevisiae, respec-tively (98, 167). The report on papulacandin B also noted
O H
H'
OH0
OHFIG. 3. Structure of echinocandin B.
10 HECTOR
I
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.

COMPOUNDS ACTIVE AGAINST CELL WALLS OF FUNGI 11
0 HOIll... 8"'
6CH2
HO
FIG. 4. Structure of papulacandin B.
that the substance acted without affecting nucleic acid ormannan synthesis, nor did it cause "leakiness" in thecytoplasmic membrane (98). In a subsequent study, theability of papulacandin B to interfere with glucan synthesiswas confirmed for both C. albicans and S. cerevisiae, thoughthe authors did note that at high concentrations (8 ,ug/ml)papulacandin B could lyse the cells (6). Echinocandin B wassimilarly able to inhibit the synthesis of beta-glucan. Some ofthe first evidence that the mechanisms of these antibioticswere specific for the glucan synthase enzyme came in 1981,when Perez et al., using material from a cell-free pellet ofhomogenized Geotrichum lactis, found that both papulacan-din B and aculeacin A were able to reduce beta-(1,3)-glucansynthesis maximally by 65 to 70% in an in vitro assaysystem, with Kis of 1.2 and 60 ,uM, respectively (183). Theauthors found that the two substances were able to affectboth the Km and the Vm.,. Interestingly, the substancescaused no inhibition when the assay material was derivedfrom stationary-phase cells. It was also determined that,under the conditions of the assay, neither mannan nor chitinsynthesis was affected by the test substances.The specificity of echinocandin B for glucan synthetase
was proven against a mixed-membrane fraction from C.albicans (216). In contrast to the situation reported forpapulacandin B and aculeacin A, this study found that theechinocandin did not affect the Km but did reduce the Vm.,findings characteristic of noncompetitive inhibition. How-ever, the authors obtained variable Kis as well, suggestingthat the kinetics may be more complicated than a simplenoncompetitive inhibition. The authors stated that the use ofa mixed-membrane preparation rather than purified enzymemight be responsible for the lack of conclusive evidence. In
both this study and the study by Perez et al., inhibition bythe test substances was indifferent to the otherwise stimula-tory capacity of added GTP.More recent studies with a derivative of echinocandin B,
cilofungin, have confirmed the specific inhibition of glucansynthesis. Taft et al. determined a K1 of 16 ,uM for thissubstance in a membrane system from N. crassa and a valueof 2.5 ,uM in a system from C. albicans, further concludingthat the inhibition was of a noncompetitive type (240, 241). Itwas determined that isolated chitin synthetase from C.albicans was unaffected by cilofungin, echinocandin B, orpapulacandin B. In the first report utilizing a solubilizedglucan synthetase, Tang and Parr determined the enzymekinetics for the enzyme from C. albicans and the inhibitionconstant for cilofungin (243). The Km of 2.0 mM was similarto that reported earlier for the membrane-bound form (216),and a Ki of 2.5 ,uM was identical to that found by Taft et al.(241). A critical finding was that both the peptide nucleus andthe lipid side chain are required for the competitive inhibi-tion of the enzyme.The beta-glucan inhibitors also have secondary effects on
other components of intact cells. Pfaller et al. (187) reportedthat, in addition to its ability to reduce glucan synthesis in C.albicans, cilofungin caused a reduction in the ergosterol andlanosterol contents of the cytoplasmic membrane and anincrease in the chitin content of the cell wall. The latterfinding is similar to the qualitative effects noted with thisyeast after treatment with papulacandin B (106). With P.brasiliensis, use of papulacandin B, while depressing beta-glucan synthesis, nevertheless caused a substantial increasein the alpha-glucan content of the cell wall of the mycelialform (64). Both papulacandin B and echinocandin B were
3.
VOL. 6, 1993
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.

CLIN. MICROBIOL. REV.
found to depress the activity of some membrane enzymes inC. albicans, i.e., ATPase and 5'-nucleotidase (236).
Effects of beta-glucan inhibitors on medically importantfungi. For the naturally occurring inhibitors of glucan syn-thetase, a very narrow spectrum of activity was determinedat an early stage of their development. Although the papu-lacandins are highly active against Candida spp. at levelsequal or superior to those of azole and polyene antifungalagents, these compounds are inactive against Cryptococcusneofonnans and several filamentous fungi (248). Similarly,aculeacins A to D and F to G have good in vitro activityagainst Candida spp. and S. cerevisiae and inhibitory thoughnot lethal activity against several filamentous fungi (167, 168,211).
Cytological and ultrastructural studies of the effects ofaculeacin A, echinocandin B, and papulacandin B on C.albicans determined that treatment with these antibioticscaused considerable structural changes in the appearance ofthe cells. Similar to the effects noted for the chitin synthetaseinhibitors, treatment of intact yeasts with echinocandin andpapulacandins appeared to cause the formation of osmoti-cally sensitive cells (47, 248). Papulacandin B treatment alsocaused cells to grow in forms resembling pseudohyphae.Yeasts treated with aculeacin A were found to grow inaggregates, the cell wall was often thickened and composedof several layers, and buds failed to separate from mothercells (28). In an important observation, the lysis of aculeacinA-treated S. cerevisiae appeared to be largely restricted tothe growing tips of budding cells, likely a reflection of theincomplete cross-linking of wall components during thedynamic process of bud growth (167). Supporting this con-clusion, the fungicidal activities of papulacandin B andechinocandin B were found to be restricted to activelygrowing yeasts; resting cells were unaffected (47, 248).The papulacandins have been evaluated in a mouse model
of systemic candidiasis and found to have good activity, with50% effective doses of 180 and 80 mg/kg for papulacandin Aand B, respectively (248).The majority of the literature on the effects of glucan
synthetase inhibitors on medically important fungi pertainsto synthetic analogs of the naturally occurring compounds.A report by Debona et al. (65) demonstrated that a variety ofderivatives can be synthesized by substituting different fattyacid or alkyloyl side chains that still retain biological activ-ity. In particular, the 4-n-octyloxybenzoyl analog cilofunginhas generated great interest. In the first reports on thisderivative, Gordee et al. (94, 95) detailed its specific mode ofaction and determined that the compound had impressive invitro activity against a variety of Candida spp. and goodefficacy in mouse models of systemic and superficial candi-diasis. As was determined for the native compounds, cil-ofungin was found to have fungicidal activity, but only foractively growing cells. A number of subsequent reports haveconfirmed the high level of in vitro activity that is essentiallylimited to members of the genus Candida, with highestactivity against C. albicans and C. tropicalis (102, 116, 158,159, 225, 228, 231). Several peculiarities or restrictive con-ditions with in vitro tests with this compound have beendocumented, including demonstration of the Eagle effect(102, 228) and effects of pH (158) and inoculum size (102).An ultrastructural study of the effects of cilofungin on C.
albicans produced findings of aggregate formation, pseudo-mycelium formation, and distortion of the cell wall andcytoplasmic membrane (74) similar to those described pre-viously for aculeacin A and papulacandin B (28). The abilityof cilofungin to inhibit germ tube formation was also noted.
Several reports on the in vivo activity of cilofungin haveappeared subsequent to the initial reports of Gordee, someof which come to conflicting conclusions about the activityof this agent in comparison to amphotericin B. Perfect et al.(184) confirmed the in vivo activity of cilofungin in a pyelo-nephritis model of candidiasis in rabbits, reporting that b.i.d.doses of 50 mg/kg were essentially as effective as 1 mg ofamphotericin B per kg. However, they found cilofungin to beless active than amphotericin B in clearing candidal endo-carditis. In a mouse model of systemic candidiasis, cilofun-gin given in doses of 62.5 mg/kg/day was comparable toamphotericin B given in doses of 0.625 mg/kg/day withrespect to survival data and the ability to clear spleens offungal burden, but amphotericin B was more active ineradicating the yeast from kidneys (103). In contrast, Smithet al., also using a murine model of systemic candidiasis,found that while treatment doses of cilofungin ranging from25 to 100 mg/kg b.i.d. resulted in no significant reduction inmortalities, amphotericin B at 1 mg/kg did confer protectionand promoted reductions in fungal organ loads (224). Possi-ble reasons for the conflicting data emerged from an elegantstudy on the pharmacokinetic behavior of cilofungin inrabbits conducted by Lee et al. (149). In addition to deter-mining such parameters as area under the curve, volume ofdistribution, clearance, and half-life for single doses, theauthors carried out a detailed study of the behavior of thiscompound with repeated interval dosing and assessed thepharmacokinetics during continuous infusion. This groupfound that with single doses cilofungin is rapidly cleared viafirst-order kinetics and had essentially no penetration intothe central nervous system. However, with continuousinfusion of the substance, the profile changed dramaticallysuch that nonlinear saturable kinetics ensued. Under theseconditions, significant penetration into the central nervoussystem and other organs and body spaces was attained. Withthese data in hand, Rouse et al. (202) used a rabbit model ofsystemic candidiasis to compare amphotericin B with cil-ofungin, the latter being infused continuously. In contrast tomany of the previously cited reports, the results indicatedthat cilofungin was significantly more effective than ampho-tericin B in reducing Candida organ counts in kidneys andwas comparable to the latter drug in the clearance in severalother key organs. In a similar approach, Cole et al. foundthat when cilofungin was administered continuously via animplanted osmotic pump in mice with a generalized Candidainfection, improved clearance of several key organs overthat seen with the drug administered intraperitoneally couldbe obtained if the dosage was high enough (56). Therefore, inthe specific case of cilofungin, it seems logical to concludethat the pharmacokinetic behavior of this drug plays acritical role in its in vivo activity. As such, the b.i.d. dosingschedule utilized in the majority of the preclinical experi-ments described herein may not have been optimized toachieve sufficient tissue penetration of this compound.
In contrast to the results of previous studies describing itslimited activity against non-Candida fungi, higher concen-trations of cilofungin had an effect on the mycelial phase ofCoccidioides immitis, in terms of both cell growth andcomposition (88). However, the antibiotic was not able toenhance killing of arthroconidia in a neutrophil phagocytosisassay. Also, the spherule-endospore phase of Coccidioidesimmitis was refractory to the effects of cilofungin in vitro,and no efficacy could be demonstrated in a mouse model ofsystemic coccidioidomycosis. Thus, these results have cer-tain similarities to the previously mentioned findings ofDavila et al. (64), who reported that, while papulacandin B
12 HECTOR
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.

COMPOUNDS ACTIVE AGAINST CELL WALLS OF FUNGI 13
FIG. 5. Structure of benanomicin A.
was able to affect the growth and composition of mycelial-phase Paracoccidioides brasiliensis, it had no effect on theyeast phase by any of the measured parameters.With the positive preclinical findings of efficacy with
cilofungin and the favorable preclinical toxicology test re-sults (95), human studies were begun, and initial results werepromising (60). Unfortunately, the clinical trials with thissubstance were halted shortly thereafter by the findings ofrenal acidosis in certain patients. These effects were pre-sumed to be a consequence of the polyethylene glycolvehicle in which this drug was administered (72).
Recent developments with inhibitors of glucan synthesis.Two recent and important findings for this class of agent are
that (i) water-soluble derivatives that retain bioactivity can
be synthesized and (ii) the spectrum of activity was discov-ered to include Pneumocystis carinii.As is now commonly known, the previously rare pneumo-
nia caused by Pneumocystis carinii marked the first clinicalrecognition of the disease syndrome known as AIDS. Thisorganism, which is commonly designated a parasite but may,in fact, be a fungus (76), contains beta-(1,3)-glucan in the cellwall of the cyst form (154). Subsequently, derivatives ofechinocandin B and papulacandin B were found to be activein a rat model of Pneumocystis pneumonia (7, 256). Thewater-soluble echinocandin B derivatives were also active ina mouse model of candidiasis, offering hope that one of thesenew analogs may serve as an effective and nontoxic alterna-tive to cilofungin.
Benanomicins and Pradimicins
The benanomicins and pradimicins are two structurallysimilar groups of antifungal compounds only recently de-scribed. These compounds, which are benzo[a]naph-thacenequinones with D-alanine substituted at C-15 and withmono- and disaccharide side chains, were discovered by twoseparate groups of investigators (Fig. 5). Benanomicins Aand B were detected in a fermentation of an isolate tenta-tively identified as an Actinomadura sp. (90, 242). Subse-quently, several methylated minor components of the com-plex were identified and described (141). Concurrently,pradimicins A to E were produced from wild-type andmutant strains ofActinomadura hibisca (180, 181, 214, 245).
The pradimicins are nearly identical to the benanomicins; infact, pradimicin C is thought to be identical to benanomicinN, a newer derivative (215). While the mechanism of actionof these compounds has not yet been determined, evidencesuggests that these compounds have a calcium-dependentaffinity for the mannan component of the fungal cell wall andsubsequently cause leakage of the cytoplasmic contents (3,178, 213, 215, 267). Benanomicins A and B have moderateactivity against yeasts and filamentous fungi in vitro as wellas good activity in a mouse model of systemic candidiasiswhen administered subcutaneously. Interestingly, benano-micins A and B also inhibit infection of T cells by humanimmunodeficiency virus (122).
Pradimicins A, B, and C were compared with amphoteri-cin B and ketoconazole in a series of in vitro tests againstdiverse fungi and were reported to have activities interme-diate to those of the latter agents (181). The activities of thepradimicins, when they were given parenterally in a mousemodel of systemic candidiasis, were similar for all threeisomers, with an average 50% protective dose of 9 mg/kg(versus 0.21 mg/kg for amphotericin B and 45 mg/kg forketoconazole). Similar activity for pradimicin A was seen inmouse models of cryptococcosis and aspergillosis.
In an effort to circumvent the drawback of poor oralbioavailability of these compounds, water-soluble pradimicinderivatives that retain the good level of activity of the parentcompounds have been created (178, 179, 213). The fact thatmultiple derivatives of this class of compounds retained goodactivity indicates that they warrant further attention.
In the hope of finding other derivatives, the producingstrain for the pradimicins was mutagenized and grown underconventional conditions or grown under directed fermenta-tion supplemented with D-amino acids (212, 215). Severalnew aglycone derivatives, i.e., pradimicins M, N, 0, and P,were found as a result of the mutagenesis; all were devoid ofantifungal activity. However, two derivatives that incorpo-rated D-serine showed activity comparable to that ofpradimicin A.At this point, it is unclear whether this class of agent will
be pursued.
Combinations of Cell Wall-Active Substances
One approach to increasing the therapeutic index of antiin-fective drugs is to test them in combinations for synergisticinteractions. While there have been a number of reportsconcerning positive interactions of antifungal or antifungaland antibacterial drugs, few combinations have shown sub-stantial improvements over the effects of the single drugs;many have shown antagonism (29, 61, 75, 123, 171, 193, 251).In the case of combinations of the traditional antifungals,azoles, 5-fluorocytosine, and amphotericin B among others,there may not be a relevant physiologic rationale for puttingthese agents together. However, in light of the previousdiscussion of the interrelation of glucans and chitin, thepossibility for synergistic interactions with compounds thataffect components of the cell wall seems plausible.
Indeed, there have been several clues in the literature tosuggest that compounds inhibiting chitin or beta-glucansynthesis have secondary effects on other components of thecell wall. Use of all three classes of beta-glucan inhibitors onselect fungi was found to stimulate the production of chitinor alpha-glucan. Miyata et al. (166) determined that treat-ment of Schizosaccharomyces pombe with aculeacin A didresult in reduced beta-glucan synthesis but greatly enhancedthe production of alpha-glucan. Pfaller et al. (187) reported
VOL. 6, 1993
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.

CLIN. MICROBIOL. REV.
that cilofungin, in addition to its ability to reduce glucansynthesis in C. albicans, caused an increase in the chitincontent of the cell wall. The latter finding is similar to thequalitative effects noted with this yeast after treatment withpapulacandin B (106). With P. brasiliensis, use of papula-candin B, while depressing beta-glucan synthesis, neverthe-less caused a substantial increase in the alpha-glucan contentof the cell wall of the mycelial form (64). These changes mayreflect an effort by the cell to overcome the inhibition of onecritical wall component by an overproduction of a secondpolymer: clearly, the ability to do so would avoid cataclys-mic consequences if these cells were exposed to thesenatural products in the soil. One of the more curious findingsin support of this hypothesis is the fact that, while themycelial phase of C. immitis can grow in concentrations ofpolyoxin D lethal to the parasitic phase, the synthesis ofchitin in the mycelial phase is greatly reduced (109). Apossible interpretation of these findings is that the mycelialphase is able to substitute another component, presumablyglucan, to maintain the structural integrity of the wall. Forundetermined reasons, the parasitic phase does not have thisability.
Circumstantial proof of this concept came with the firstreports on combinations of nikkomycin Z and beta-glucaninhibitors tested against fungi. Whole cells and protoplasts ofC. albicans treated with nikkomycin Z and papulacandin Bwere found to interact synergistically to prevent growth invitro, a result not particularly surprising given that this yeastis susceptible to both agents singly (106). However, the moreinteresting and solid support came when nikkomycin Z andcilofungin were used against mycelial fungi. When isolates ofA. fiumigatus and Cunninghamella bertholletiae were testedin vitro, they were resistant to the single drugs up to thehighest concentrations tested. The surprising result was thatboth fungi were found to be highly susceptible to thecombination of the two agents, with fractional inhibitoryconcentrations as low as 0.04 (<0.5 indicates synergy) (185).Examination of the treated cells after 24 h revealed that theconidia used for inoculum had become osmotically sensitive,swelling to several times their normal sizes. These dramaticresults demonstrate that these filamentous fungi do not havethe ability to resist the simultaneous loss of two importantcell wall components.
If these findings can be extended to other fungi outside thespectrum of activity of these single agents and if the combi-nations can be successfully implemented in therapy trials,then the use of combinations of cell wall-active substancesshould be a welcome addition to the armamentarium avail-able to clinicians.
It is important to point out, however, that if compoundsthat completely inhibit chitin or beta-glucan synthesis couldbe identified, these single drugs would likely be just aseffective as the combinations described above. It is knownthat the papulacandins and nikkomycins depress but do notcompletely halt the synthesis of beta-(1,3)-glucan and chitin,respectively (106, 183). However, in studies described ear-lier in this review (22, 23, 133), temperature-sensitive mu-tants of A. nidulans that were completely blocked for eitherchitin or beta-(1,3)-glucan synthesis showed the same pat-terns of osmotically sensitive cells found when combinationsof cilofungin and nikkomycin Z were used against A. fiumig-atus (185).
Other recent reports describe the synergistic interaction ofcombinations of nikkomycin and azole antifungal agentsagainst C. albicans in vitro and in vivo (110, 162). Thisphenomenon is probably a result of the previously described
secondary effects of azoles on chitin synthesis (107, 188, 255)being compounded by the specific inhibition of this polymerby nikkomycin. The usefulness of this approach has yet to bedetermined because the effect is limited to certain ratios ofthe two compounds, making dosing problematic.
CONCLUSIONSCollectively, the information available on compounds that
interfere with the cell walls of fungi does seem to validate thepremise that these substances offer much the same promisethat penicillin did for antibacterial therapy in the 1940s. Atthe same time, however, the early data have adequatelydocumented the shortcomings of these agents. It becameclear during the preparation of this paper that restricting it toa review of articles on anti-cell wall substances would notimpart an appreciation for some of the more subtle issuesand restrictions germane to each of the drugs. In particular,the problem of a narrow spectrum of activity for many ofthese substances should be assessed in the context of howindividual species of medically important fungi utilize vari-ous wall components. For example, the fact that echinocan-din B and its derivatives are active against species ofCandida but little else becomes understandable when onerealizes that, for this genus, chitin is of importance forseptum formation but beta-glucan plays the dominant role inthe architecture of the yeast wall. Conversely, for thedimorphic pathogenic fungi, the parasitic phase has anabsolute requirement for chitin to maintain the cell wall, butbeta-glucan may be less important. This type of analysisbecomes important, from both a marketing and a therapeuticviewpoint, in the strategies one implements for using thesedrugs.
In contrast to the situation that amphotericin B and someof the azoles enjoy, in which a clinician can assume broad-spectrum coverage for most applications, it is clear that useof single cell wall-active substances will require a reasonableidentification of the fungus involved. The possibility ofcombining agents to expand the spectrum of activity is anattractive but tentative finding. As such, the acceptance anduse of these substances by clinicians will likely depend on asolid demonstration of an improved therapeutic index and/orreduced toxicity compared with what is presently available.It seems clear that the preclinical data generated to date withthe echinocandin derivatives and the nikkomycins providean adequate basis for their continued development. Giventhat the literature is replete with articles describing theincreased numbers of mycoses in patients with some degreeof compromised immune status, a situation that demandsmore potent candidates for chemotherapy, it seems a fore-gone conclusion that the most promising of the compoundsdescribed herein will find use in medical practice. From thevantage point of one who works in the pharmaceuticalindustry, however, "market share" and "narrow spectrum"are mutually exclusive terms. Still, given the success of thenewest entrants into the marketplace, there is reason to beoptimistic that cell wall-active substances will become avail-able in the future.
REFERENCES1. Andreoli, T. E., and M. Monahan. 1968. The interaction of
polyene antibiotics with thin lipid membranes. J. Gen. Physiol.52:300-325.
2. Archer, D. B. 1977. Chitin biosynthesis in protoplasts andsubcellular fractions of Aspergillus fumigatus. Biochem. J.164:653-658.
14 HEC'TOR
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.

COMPOUNDS ACTIVE AGAINST CELL WALLS OF FUNGI 15
3. ASM News. 1991. New antifungal agents being studied. ASMNews 57:6.
4. Au-Young, J., and P. W. Robbins. 1990. Isolation of a chitinsynthase gene (CHS1) from Candida albicans by expression inSaccharomyces cerevisiae. Mol. Microbiol. 4:197-207.
5. Azuma, I., F. Kanetsuna, Y. Tanaka, Y. Yamamura, and L. M.Carbonell. 1974. Chemical and immunological properties ofgalactomannans obtained from Histoplasma duboisii, His-toplasma capsulatum, Paracoccidioides brasiliensis, and Blas-tomyces dermatitidis. Mycopathol. Mycol. Appl. 54:111-125.
6. Baguley, B. C., G. Rommele, J. Gruner, and W. Wehrli. 1979.Papulacandin B: an inhibitor of glucan synthesis in yeastspheroplasts. Eur. J. Biochem. 97:345-351.
7. Balkovec, J. M., R. M. Black, M. L. Hammond, J. V. Heck,R. A. Zambias, G. Abruzzo, K. Bartizal, H. Dropp, C. Trainor,R. E. Schwartz, D. C. McFadden, K. H. Nollstadt, L. A.Pittarelli, M. A. Powles, and D. M. Schmatz. 1992. Synthesis,stability, and biological evaluation of water-soluble prodrugs ofa new echinocandin lipopeptide. Discovery of a potentialclinical agent for the treatment of systemic candidiasis andPneumocystis carinii pneumonia (PCP). J. Med. Chem. 35:194-198.
8. Ballou, C. E. 1970. A study of the immunochemistry of threeyeast mannans. J. Biol. Chem. 245:1197-1203.
9. Ballou, C. E. 1976. Structure and biosynthesis of the mannancomponent of the yeast cell envelope. Adv. Microb. Physiol.14:93-158.
10. Bardalaye, P. C., and J. H. Nordin. 1977. Chemical structure ofthe galactomannan from the cell wall of Aspergillus niger. J.Biol. Chem. 252:2584-2591.
11. Barreto-Bergter, E. M., P. A. J. Gorin, and L. R. Travassos.1981. Cell constituents of mycelia and conidia of Aspergillusfumigatus. Carbohydr. Res. 95:205-218.
12. Barrett-Bee, K. J., and W. Henderson. 1982. Variation in theactivities of enzymes associated with cell wall metabolismduring a growth cycle of Candida albicans. FEMS Microbiol.Lett. 15:275-278.
13. Bartnicki-Garcia, S. 1968. Cell wall chemistry, morphogenesis,and taxonomy of fungi. Annu. Rev. Microbiol. 22:87-108.
14. Bartnick-Garcia, S., and E. Lippman. 1972. Inhibition of Mu-cor rouxii by polyoxin D: effect on chitin synthetase andmorphological development. J. Gen. Microbiol. 71:301-309.
15. Bastide, J. M., E. H. Hadibi, and M. Bastide. 1979. Taxonomicsignificance of yeast sphaeroplast release after enzymic treat-ment of intact cells. J. Gen. Microbiol. 113:147-153.
16. Becker, J. M., N. L. Covert, P. Shenbagamurthi, A. S. Stein-feld, and F. Naider. 1983. Polyoxin D inhibits growth ofzoopathogenic fungi. Antimicrob. Agents Chemother. 23:926-929.
17. Becker, J. M., S. Marcus, J. Tullock, D. Miller, E. Krainer,R. K. Khare, and F. Naider. 1988. Use of the chitin-synthesisinhibitor nikkomycin to treat disseminated candidiasis in mice.J. Infect. Dis. 157:212-214. (Letter.)
18. Bennett, J. E., A. K. Bhattacharjee, and C. P. J. Glaudemans.1985. Galactofuranosyl groups are immunodominant in As-pergillus fumigatus galactomannan. Mol. Immunol. 22:251-254.
19. Benz, F., F. Knuesel, J. Nuesch, H. Treichler, W. Voser, R.Nyfeler, and W. Keller-Schierlein. 1974. Echinocandin B, einneuartiges Polypeptid Antibioticum aus Aspergillus nidulansvar echinulatus: Isolierung and Baudsteine. Helv. Chim. Acta57:2459-2477.
20. Bhattacharjee, A. K., K. J. Kwon-Chung, and C. P. J. Glaude-mans. 1981. Capsular polysaccharide from a parent strain andfrom a possible mutant strain of Cryptococcus neofornansserotype A. Carbohydr. Res. 95:237-248.
21. Binks, P. R., G. D. Robson, M. W. Goosey, A. Humphreys, andA. P. J. Trinci. 1990. Chitin synthesis in Fusarium gramin-earum and its inhibition by edifenphos (Hinosan). J. Gen.Microbiol. 137:615-620.
22. Borgia, P. T. 1992. Roles of the orLA, tsE, and bimG genes ofAspergillus nidulans in chitin synthesis. J. Bacteriol. 174:384-389.
23. Borgia, P. T., and C. L. Dodge. 1992. Characterization ofAspergillus nidulans mutants deficient in cell wall chitin orglucan. J. Bacteriol. 174:377-383.
24. Bormann, C., W. Huhn, H. Zahner, R. Rathmann, H. Hahn,and W. A. Konig. 1985. New nikkomycins produced by mu-tants of Streptomyces tendae. J. Antibiot. 38:9-16.
25. Bormann, C., S. Mattern, H. Schrempf, H.-P. Fiedler, and H.Zahner. 1989. Isolation of Streptomyces tendae mutants withan altered nikkomycin spectrum. J. Antibiot. 42:913-918.
26. Bowen, A. R., J. L. Chen-Wu, M. Momany, R. Young, P. J.Szaniszlo, and P. W. Robbins. 1992. Classification of fungalchitin synthases. Proc. Natl. Acad. Sci. USA 89:519-523.
27. Bowers, B., G. Levin, and E. Cabib. 1974. Effect of polyoxin Don chitin synthesis and septum formation in Saccharomycescerevisiae. J. Bacteriol. 119:564-575.
28. Bozzola, J. J., R. J. Mehta, L. J. Nisbet, and J. R. Valenta.1984. The effect of aculeacin A and papulacandin B on mor-phology and cell wall ultrastructure in Candida albicans. Can.J. Microbiol. 30:857-863.
29. Brajtburg, J., D. Kobayashi, G. Medoff, and G. S. Kobayashi.1982. Antifungal action of amphotericin B in combination withother polyene or imidazole antibiotics. J. Infect. Dis. 146:138-146.
30. Braun, P. C., and R. A. Calderone. 1978. Chitin synthesis inCandida albicans: comparison of yeast and hyphal forms. J.Bacteriol. 135:1472-1477.
31. Brillinger, G. U. 1979. Metabolic products of microorganisms.181. Chitin synthase from fungi, a test model for substanceswith insecticidal properties. Arch. Microbiol. 121:71-74.
32. Bulawa, C. 1992. CSD2, CSD3, and CSD4, genes required forchitin synthesis in Saccharomyces cerevisiae: the CSD2 geneproduct is related to chitin synthases and to developmentallyregulated proteins in Rhizobium species and Xenopus laevis.Mol. Cell. Biol. 12:1764-1776.
33. Bull, A. T. 1970. Chemical composition of wild-type andmutant Aspergillus nidulans cell walls. The nature of polysac-charide and melanin constituents. J. Gen. Microbiol. 63:75-94.
34. Cabib, E. 1991. Differential inhibition of chitin synthetases 1and 2 from Saccharomyces cerevisiae by polyoxin D andnikkomycins. Antimicrob. Agents Chemother. 35:170-173.
35. Cabib, E., and B. Bowers. 1971. Chitin and yeast budding:localization of chitin in yeast bud scars. J. Biol. Chem.246:152-159.
36. Cabib, E., and B. Bowers. 1975. Timing and function of chitinsynthesis in yeast. J. Bacteriol. 124:1586-1593.
37. Cabib, E., R. Roberts, and B. Bowers. 1982. Synthesis of theyeast cell wall and its regulation. Annu. Rev. Biochem. 51:763-793.
38. Cabib, E., A. Sburlati, B. Bowers, and S. J. Silverman. 1989.Chitin synthase I, an auxiliary enzyme for chitin synthesis inSaccharomyces cerevisiae. J. Cell. Biol. 108:1665-1672.
39. Cabib, E., S. J. Silverman, and J. A. Shaw. 1992. Chitinase andchitin synthase 1: counterbalancing activities in cell separationof Saccharomyces cerevisiae. J. Gen. Microbiol. 138:97-102.
40. Calderone, R. A., and P. C. Braun. 1991. Adherence andreceptor relationships of Candida albicans. Microbiol. Rev.55:1-20.
41. Campbell, C. K. 1970. Fine structure of vegetative hyphae ofAspergillus fumigatus. J. Gen. Microbiol. 64:373-376.
42. Carbonell, L. M., and F. Gil. 1982. Ultraestructura del Para-coccidioides brasiliensis, p. 23-34. In G. Del Negro, C. D.Lacaz, and A. M. Fiorello (ed.), Paracoccidioidomicose, blas-tomicose Sul-Americana. Sarvier-Edusp., Sao Paulo, Brazil.
43. Carbonell, L. M., F. Kanetsuna, and F. Gil. 1970. Chemicalmorphology of glucan and chitin in the cell wall of the yeastphase of Paracoccidioides brasiliensis. J. Bacteriol. 101:636-642.
44. Cass, A., A. Finkelstein, and V. Krespi. 1970. The ion perme-ability induced in thin lipid membranes by the polyene antibi-otics nystatin and amphotericin B. J. Gen. Physiol. 56:100-124.
45. Cassone, A. 1989. Cell wall of Candida albicans: its functionsand its impact on the host, p. 248-314. In M. McGinnis and M.Borgers (ed.), Current topics in medical mycology, vol. 3.
VOL. 6, 1993
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.

CLIN. MICROBIOL. REV.
Springer-Verlag, New York.46. Cassone, A., D. Kerridge, and E. F. Gale. 1979. Ultrastructural
changes in the cell wall of Candida albicans following cessa-tion of growth and their possible relationship to the develop-ment of polyene resistance. J. Gen. Microbiol. 110:339-349.
47. Cassone, A., R. E. Mason, and D. Kerridge. 1981. Lysis ofgrowing yeast-form cells of Candida albicans by echinocandin:a cytological study. Sabouraudia 19:97-110.
48. Cassone, A., N. Simonetti, and V. Strippoli. 1973. Ultrastruc-tural changes in the wall during germ-tube formation fromblastospores of Candida albicans. J. Gen. Microbiol. 77:417-426.
49. Cassone, A., N. Simonetti, and V. Strippoli. 1974. Wall struc-ture and bud formation in Cryptococcus neoformans. Arch.Microbiol. 95:205-212.
50. Chattaway, F. W., and M. R. Holmes. 1968. Cell wall compo-sition of the mycelial and blastospore forms of Candida albi-cans. J. Gen. Microbiol. 51:367-376.
51. Chen-Wu, J. L., J. Zwicker, A. R. Bowen, and P. W. Robbins.1992. Expression of chitin synthase genes during yeast andhyphal growth phases of Candida albicans. Mol. Microbiol.6:497-502.
52. Cherniak, A., E. Reiss, M. E. Slodki, R. D. Plattner, and S. 0.Blumer. 1980. Structure and antigenic activity of the capsularpolysaccharide of Cryptococcus neoformans serotype A. Mol.Immunol. 17:1025-1032.
53. Cherniak, A., E. Reiss, and S. H. Turner. 1982. A galactoxy-lomannan of Cryptococcus neoformans serotype A. Carbo-hydr. Res. 103:239-250.
54. Chiew, Y. Y., M. G. Shepherd, and P. A. Sullivan. 1980.Regulation of chitin synthesis during germ-tube formation inCandida albicans. Arch. Microbiol. 125:97-104.
55. Cohen, E., and J. E. Casida. 1980. Inhibition of Tribolium gutchitin synthetase. Pestic. Biochem. Physiol. 13:129-136.
56. Cole, G. T., K. T. Lynn, and K. R. Seshan. 1990. Evaluation ofa murine model of hepatic candidiasis. J. Clin. Microbiol.28:1828-1841.
57. Collins, M., and D. Pappagianis. 1973. Effect of lysozyme andchitinase on the spherules of Coccidioides immitis in vitro.Infect. Immun. 7:817-822.
58. Collins, M. S., D. Pappagianis, and J. Yee. 1977. Enzymaticsolubilization of precipitin and complement fixing antigen fromendospores, spherules, and spherule fraction of Coccidioidesimmitis, p. 429-444. In L. Ajello (ed.), Coccidioidomycosis.Symposia Specialists, Miami.
59. Cooper, C. R., Jr., J. L. Harris, C. W. Jacobs, and P. J.Szaniszlo. 1984. Effects of polyoxin AL on cellular develop-ment in Wangiella dermatitidis. Exp. Mycol. 8:349-363.
60. Copley-Merriman, C. R., H. Gallis, J. R. Graybill, B. N.Doebbeling, and D. L. Hyslop. 1990. Program Abstr. 30thIntersci. Conf. Antimicrob. Agents Chemother., abstr. 582.
61. Corrado, M. L., M. Kramer, M. Cummings, and R. H. Eng.1982. Susceptibility of dematiaceous fungi to amphotericin B,miconazole, ketoconazole, flucytosine and rifampin alone andin combination. Sabouraudia 20:109-113.
62. Correa, J. U., N. Elango, I. Polacheck, and E. Cabib. 1981.Endochitinase, a mannan-associated enzyme from Saccharo-myces cerevisiae. J. Biol. Chem. 257:1392-1397.
63. Dahn, U., H. Hagenmaier, H. Hohne, W. A. Konig, G. Wolf,and H. Zahner. 1976. Stoffwechselprodukte von Mikroorgan-ismen. 154. Mitteilung. Nikkomycin, ein neuer Hemmstoff derChitin-synthese bei Pilzen. Arch. Microbiol. 107:143-160.
64. Davila, T., G. San-Blas, and F. San-Bias. 1986. Effect ofpapulacandin B on glucan synthesis in Paracoccidioides bra-siliensis. J. Med. Vet. Mycol. 24:193-202.
65. Debona, M., B. J. Abbot, D. S. Fukuda, M. Barnhart, K. E.Willard, R. M. Molloy, K. H. Michel, J. R. Turner, T. F.Butler, and A. H. Hunt. 1989. Synthesis of new analogs ofechinocandin B by enzymatic deacylation and chemical reacy-lation of the echinocandin B peptide: synthesis of the antifun-gal agent cilofungin (LY121019). J. Antibiot. 42:389-397.
66. Decker, H., C. Bormann, H.-P. Fiedler, H. Zahner, H. Heitsch,and W. A. Konig. 1989. Isolation of new nikkomycins from
Streptomyces tendae. J. Antibiot. 42:230-235.67. Decker, H., F. Walz, C. Bormann, H. Zahner, H.-P. Fiedler, H.
Heitsch, and W. A. Konig. 1990. Nikkomycins Wz and W., newchitin synthetase inhibitors from Streptomyces tendae. J. An-tibiot. 43:43-48.
68. Decker, H., H. Zahner, H. Heitsch, W. A. Konig, and H.-P.Fiedler. 1991. Structure-activity relationships of the nikkomy-cins. J. Gen. Microbiol. 137:1805-1813.
69. Delzer, J., H.-P. Fiedler, H. Mueller, H. Zahner, R. Rathmann,K. Ernst, and W. A. Konig. 1984. New nikkomycins bymutasynthesis and directed fermentation. J. Antibiot. 37:80-82.
70. Diamond, R. 1991. The growing problem of mycoses in patientsinfected with the immunodeficiency virus. Rev. Infect. Dis.13:480-486.
71. Dickinson, K., V. Keer, C. A. Hitchcock, and D. J. Adams.1989. Chitinase activity from Candida albicans and its inhibi-tion by allosamidin. J. Gen. Microbiol. 135:1417-1421.
72. Doebbeling, B. N., B. D. Fine, Jr., M. A. Pfaller, C. T. Sheetz,J. B. Stokes, and R. P. Wenzel. 1990. Program Abstr. 30thIntersci. Conf. Antimicrob. Agents Chemother., abstr. 583.
73. Domer, J. E. 1971. Monosaccharide and chitin content of cellwall of Histoplasma capsulatum and Blastomyces dermatiti-dis. J. Bacteriol. 107:870-877.
74. Drouhet, E., B. Dupont, L. Improvisi, M. Lesourd, and M. C.Prevost. 1990. Activity of cilofungin (LY121019), a new ii-popeptide antibiotic, on the cell wall and cytoplasmic mem-brane of Candida albicans. Structural modifications in scan-ning and transmission electron microscopy. J. Med. Vet.Mycol. 28:425-436.
75. Dupont, B., and E. Drouhet. 1979. In vitro synergy andantagonism of antifungal agents against yeast-like fungi. Post-grad. Med. J. 55:683-686.
76. Edman, J. C., J. A. Kovacs, H. Masur, D. V. Santi, H. J.Elwood, and M. L. Sogin. 1988. Ribosomal RNA sequenceshows Pneumocystis carinii to be a member of the fungi.Nature (London) 334:519-522.
77. Egucki, J., S. Sasaki, N. Ota, T. Akashiba, T. Tsuchiyama, andS. Suzuki. 1968. Studies on polyoxins, antifungal antibiotics.IX. Mechanism of action on the diseases caused byAlternariaspp. Ann. Phytopathol. Soc. Jpn. 34:280-288.
78. Elango, N., J. U. Correa, and E. Cabib. 1982. Secretorycharacter of yeast chitinase. J. Biol. Chem. 257:1398-1400.
79. Elorza, M. V., A. Murgui, H. Rico, F. Miragall, and R.Sentandreu. 1987. Formation of a new cell wall by protoplastsof Candida albicans: effect of papulacandin B, tunicamycinand nikkomycin. J. Gen. Microbiol. 133:2315-2325.
80. Elorza, M. V., H. Rico, D. Gozalbo, and R. Sentandreu. 1983.Cell wall composition and protoplast regeneration in Candidaalbicans. Antonie van Leeuwenhoek 49:457-469.
81. Emmer, G., N. S. Ryder, and M. A. Grassberger. 1984.Synthesis of new polyoxin derivatives and their activity againstchitin synthase from Candida albicans. J. Med. Chem. 28:278-281.
82. Endo, A., K. Kakiki, and T. Misato. 1970. Mechanism of actionof the antifungal agent polyoxin D. J. Bacteriol. 104:189-196.
83. Endo, A., and T. Misato. 1969. Polyoxin D, a competitiveinhibitor of UDP-N-acetylglucosaminyltransferase in Neuros-pora crassa. Biochem. Biophys. Res. Commun. 37:718-722.
84. Engel, P. 1987. Plasmid transformation of Streptomyces tendaeafter heat attenuation of restriction. Appl. Environ. Microbiol.53:1-3.
85. Engel, P., and A. H. J. Ullah. 1988. Mutation affecting peptidebond formation in nikkomycin biosynthesis. Biochem. Bio-phys. Res. Commun. 156:695-700.
86. Farkas, V. 1979. Biosynthesis of cell walls of fungi. Microbiol.Rev. 43:117-144.
87. Fox, J. J., K. A. Watanabe, and A. Bloch. 1966. Nucleosideantibiotics. Prog. Nucleic Acid Res. Mol. Biol. 5:251-313.
88. Galgiani, J. N., S. H. Sung, K. V. Clemons, and D. A. Stevens.1990. Activity of cilofungin against Coccidioides immitis: dif-ferential in vitro effects on mycelia and spherules correlatedwith in vivo studies. J. Infect. Dis. 162:944-948.
16 HECTOR
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.

COMPOUNDS ACTIVE AGAINST CELL WALLS OF FUNGI 17
89. Gimeno, C. J., P.O. Ljungdahl, C. A. Styples, and G. R. Fink.1992. Unipolar cell divisions in the yeast S. cerevisiae lead tofilamentous growth: regulation by starvation and RAS. Cell68:1077-1090.
90. Gomi, S., M. Sezaki, S. Kondo, T. Hara, H. Naganawa, and T.Takeuchi. 1989. The structures of new antifungal antibiotics,benanomicins A and B. J. Antibiot. 41:1019.
91. Gooday, G. W. 1977. Biosynthesis of the fungal wall-mecha-nisms and implications. J. Gen. Microbiol. 99:1-11.
92. Gooday, G. W., A. deRousset-Hall, and D. Hunsley. 1976.Effect of polyoxin D on chitin synthesis in Coprinus cinereus.Trans. Br. Mycol. Soc. 67:193-200.
93. Gopal, P. K., M. G. Shepherd, and P. A. Sullivan. 1984.Analysis of wall glucans from yeast, hyphal and germ-tubeforming cells of Candida albicans. J. Gen. Microbiol. 130:3295-3301.
94. Gordee, R. S., D. J. Zeckner, L. F. Ellis, A. L. Thakkar, andL. C. Howard. 1984. In vitro and in vivo anti-Candida activityand toxicology of LY121019. J. Antibiot. 37:1054-1065.
95. Gordee, R. S., D. J. Zeckner, L. C. Howard, W. E. Alborn, Jr.,and M. Debono. 1988. Anti-Candida activity and toxicology ofLY121019, a novel polypeptide antifungal antibiotic. Ann.N.Y. Acad. Sci. 544:294-301.
96. Gottlieb, S., Z. Altboum, D. C. Savage, and E. Segal. 1991.Adhesion of Candida albicans to epithelial cells effect ofpolyoxin D. Mycopathologia 115:197-205.
97. Gow, L. A., and C. P. Selitrennikoff. 1984. Chitin synthetase ofNeurospora crassa: inhibition by nikkomycin, polyoxin B, andUDP. Curr. Microbiol. 11:211-216.
98. Gruner, J., and P. Traxler. 1977. Papulacandin, a new antibi-otic, active especially against yeasts. Experientia 33:137.
99. Hagenmaier, H., A. Keckeisen, W. Dehier, H.-P. Fiedler, H.Zahner, and W. A. Konig. 1981. Konstitutionsaufldarung derNikkomycine I, J, M und N. Liebigs Ann. Chem. 1981:1728-1735.
100. Hagenmaier, H., A. Keckeisen, H. Zalner, and W. A. Konig.1979. Aufklarung der Struktur des NucleosidantibiotikumsNikkomycin X. Liebigs Ann. Chem. 1979:1494-1502.
101. Hahn, H., H. Heitsch, R. Rathmann, G. Zimmerman, C.Bormann, H. Zahner, and W. A. Konig. 1987. Partialsyntheseder Nikkomycine B. und K, sowie unnaturlicher Sterioi-somerer und Strukturanaloga. Liebigs Ann. Chem. 1987:803-807.
102. Hall, G. S., C. Myles, K. J. Pratt, and J. A. Washington. 1988.Cilofungin (LY121019), an antifungal agent with specific activ-ity against Candida albicans and Candida tropicalis. Antimi-crob. Agents Chemother. 32:1331-1335.
103. Hanson, L. H., A. M. Perlman, K. V. Clemons, and D. A.Stevens. 1991. Synergy between cilofungin and amphotericin Bin a murine model of candidiasis. Antimicrob. AgentsChemother. 35:1334-1337.
104. Hasenclever, H. F., and W. 0. Mitchell. 1964. A study of yeastsurface antigens by agglutination inhibition. Sabouraudia3:288.
105. Hearn, V. M., and D. W. R. Mackenzie. 1981. Analysis of wallantigens of Aspergillus fiumigatus by two-dimensional immu-noelectrophoresis. J. Med. Microbiol. 14:119-129.
106. Hector, R. F., and P. C. Braun. 1986. Synergistic action ofnikkomycins X and Z with papulacandin B on whole cells andregenerating protoplasts of Candida albicans. Antimicrob.Agents Chemother. 29:389-394.
107. Hector, R. F., and P. C. Braun. 1987. The effect of bifonazoleon chitin synthesis in Candida albicans, p. 369-382. In R. A.Fromtling (ed.), Recent trends in the discovery, developmentand ev41tJation of antifungal agents. J. R. Prous SciencePublishers, Barcelona, Spain.
108. Hector, R. F., and D. Pappagianis. 1982. Enzymatic degrada-tion of the walls of spherules of Coccidioides immitis. Exp.Mycol. 6:136-152.
109. Hector, R. F., and D. Pappagianis. 1983. Inhibition of chitinsynthesis in the cell wall of Coccidioides immitis by polyoxinD. J. BIacteriol. 154:488-498.
110. Hector, R. F., and K. Schaller. 1992. Positive interaction of
nikkomycins and azoles against Candida albicans in vitro andin vivo. Antimicrob. Agents Chemother. 36:1284-1289.
111. Hector, R. F., B. L. Zimmer, and D. Pappagianis. 1985. Therole of endogenous chitinase in the maturation of the spherule-endospore phase of Coccidioides immitis, abstr. F 61, p. 374.Abstr. 85th Annu. Meet. Am. Soc. Microbiol. 1985. AmericanSociety for Microbiology, Washington, D.C.
112. Hector, R. F., B. L. Zimmer, and D. Pappagianis. 1990.Evaluation of nikkomycins X and Z in murine models ofcoccidioidomycosis, histoplasmosis, and blastomycosis. Anti-microb. Agents Chemother. 34:587-593.
113. Hector, R. F., B. L. Zimmer, and D. Pappagianis. 1991.Inhibitors of cell wall synthesis: nikkomycins, p. 341-353. InH. Yamaguchi et al. (ed.), Recent progress in antifungalchemotherapy. Marcel Dekker, New York.
114. Heitsch, H., W. A. Konig, H. Decker, C. Bormann, H.-P.Fiedler, and H. Zahner. 1989. Structure of the new nikkomy-cins pseudo-Z and pseudo-J. J. Antibiot. 42:711-717.
115. Hilenski, L. L., F. Naider, and J. M. Becker. 1986. Polyoxin Dinhibits colloidal gold-wheat germ agglutinin labelling of chitinin dimorphic forms of Candida albicans. J. Gen. Microbiol.132:1441-1451.
116. Hobbs, M., J. Perfect, and D. Duraclk 1988. Evaluation of invitro antifungal activity of LY121019. Eur. J. Clin. Microbiol.Infect. Dis. 7:77-80.
117. Holst, H., D. Malissiovas, and H. Schmutterer. 1978. Wirkun-gen eines chitinsynthesehemmenden Antibiotikums auf Spin-nmilben und Insekten. Mitt. Dtsch. Ges. Allg. Angew. Ento-mol. 1:143-146.
118. Hori, M., J. Eguchi, K. Kakiki, and T. Misato. 1974. Studies onthe mode of action of polyoxins. VI. Effect of polyoxin B onchitin synthesis in polyoxin-sensitive and resistant strains ofAlternaia kikuchiana. J. Antibiot. 27:260-266.
119. Hori, M., K. Kaldki, and T. Misato. 1974. Further study on therelation of polyoxin structure to chitin synthetase inhibition.Agric. Biol. Chem. 38:691-698.
120. Hori, M., K. Kakiki, and T. Misato. 1974. Interaction betweenpolyoxin and active center of chitin synthetase. Agric. Biol.Chem. 38:699-705.
121. Hori, M., K. Kakiki, S. Suzuki, and T. Misato. 1971. Studies onthe mode of action of polyoxins. III. Relation of polyoxinstructure to chitin synthetase inhibition. Agric. Biol. Chem.35:1280-1291.
122. Hoshino, H., J. Seki, and T. Takeuchi. 1989. New antifungalantibiotics, benanomicins A and B inhibit infection of T-cellwith human immunodeficiency virus (HIV) and syncytiumformation by HIV. J. Antibiot. 42:344-346.
123. Hughes, C. E., C. Harris, J. A. Moody, L. R. Peterson, andD. N. Gerding. 1984. In vitro activities of amphotericin B incombination with four antifungal agents and rifampin againstAspergillus spp. Antimicrob. Agents Chemother. 25:560-562.
124. Isono, K., K. Asahi, and S. Suzuki. 1969. Studies on polyoxins,antifungal antibiotics. XIII. The structure of polyoxins. J. Am.Chem. Soc. 91:7490-7505.
125. Isono, K., J. Nagatsu, Y. Kawashima, and S. Suzuki. 1965.Studies on polyoxins, antifungal antibiotics. I. Isolation andcharacterization of polyoxins A and B. Agric. Biol. Chem.29:848-854.
126. Isono, K., J. Nagatsu, K. Kobinata, K. Sasaki, and S. Suzuki.1965. Studies on polyoxins, antifungal antibiotics. V. Isolationand characterization of polyoxins C, D, E, F, G, H, and I.Agric. Biol. Chem. 31:190-199.
127. Isono, K., S. Suzuki, M. Tanaka, T. Nanbata, and K. Shibuya.1970. Bisulfite-catalyzed facile decarboxylation reaction of5-carboxyuracil derivatives and transformation of polyoxins.Tetrahedron 6:425-428.
128. Iwamoto, T., A. Fujie, Y. Tsurumi, K. Nitta, S. Hashimoto, andM. Okuhara. 1990. FR900403, a new antifungal antibioticproduced by a Kernia sp. J. Antibiot. 43:1183-1185.
129. James, P. G., R. Cherniak, R. G. Jones, and C. A. Stortz. 1990.Cell-wall glucans of Cryptococcus neoformans CAP 67. Car-bohydr. Res. 198:23-38.
130. Kanetsuna, F., and L. M. Carbonell. 1971. Cell wall composi-
VOL. 6, 1993
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.

CLIN. MICROBIOL. REV.
tion of the yeastlike and mycelial forms of Blastomyces der-matitidis. J. Bacteriol. 106:946-948.
131. Kanetsuna, F., L. M. Carbonell, F. Gil, and I. Azuma. 1974.Chemical and ultrastructural studies on the cell walls of theyeast-like and mycelial forms of Histoplasma capsulatum.Mycopathol. Mycol. Appl. 54:1-13.
132. Kanetsuna, F., L. M. Carbonell, R. E. Moreno, and J. Ro-driquez. 1969. Cell wall composition of the yeast and mycelialforms of Paracoccidioides brasiliensis. J. Bacteriol. 97:1036-1041.
133. Katz, D., and R. F. Rosenberger. 1970. A mutation inAspergil-lus nidulans producing hyphal walls which lack chitin. Bio-chim. Biophys. Acta 208:452-460.
134. Katz, D., and R. F. Rosenberger. 1971. Hyphal wall synthesisin Aspergillus nidulans: effect of protein synthesis inhibitionand osmotic shock on chitin insertion and morphogenesis. J.Bacteriol. 108:184-190.
135. Keller, F. A., and E. Cabib. 1971. Chitin and yeast budding.Properties of chitin synthetase from Saccharomyces carlsber-gensis. J. Biol. Chem. 246:160-166.
136. Keller-Juslen, C., M. Kuhn, H. R. Loosli, T. J. Petcher, H. P.Weber, and A. Von Wartburg. 1976. Strucktur des Cyclopep-tid-Antibiotikums SL 7810 (=Echinocandin B). TetrahedronLett. 1976:4147-4150.
137. Kempel, K. R., and W. E. Goldman. 1988. Cell walls fromavirulent variants of Histoplasma capsulatum lack alpha-(1,3)-glucan. Infect. Immun. 56:2997-3000.
138. Khare, R. K., J. M. Becker, and F. R. Naider. 1988. Synthesisand anticandidal properties of polyoxin L analogues containingalpha-amino fatty acids. J. Med. Chem. 31:650-656.
139. Kitazima, Y., Y. Banno, T. Noguchi, Y. Nozawa, and Y. Ito.1972. Effects of chemical modification of structural polymerupon the cell wall integrity of Trichophyton. Arch. Biochem.Biophys. 152:811-820.
140. Koga, D., A. Isogai, S. Sakuda, S. Matsumoto, A. Suzuki, S.Kimura, and A. Ide. 1987. Specific inhibition of Bombay morichitinase by allosamidin. Agric. Biol. Chem. 51:471-476.
141. Kondo, S., S. Gomi, K. Uotani, S. Inouye, and T. Takeuchi.1991. Isolation of new minor benanomicins. J. Antibiot. 44:123-129.
142. Konig, W. A., W. Hass, W. Dehler, H.-P. Fiedler, and H.Zahner. 1980. Strukturaufklarung und Partialsynthese des Nu-cleosidantibiotikums Nikkomycin B. Liebigs Ann. Chem.1980:622-628.
143. Kopecka, M., H. J. Phaff, and G. H. Fleet. 1974. Demonstra-tion of a fibrillar component in the cell wall of the yeastSaccharomyces cerevisiae and its chemical nature. J. CellBiol. 62:66-76.
144. Krainer, E., J. M. Becker, and F. Naider. 1991. Synthesis andbiological evaluation of dipeptidyl and tripeptidyl polyoxin andnikkomycin analogues as anticandidal prodrugs. J. Med.Chem. 34:174-180.
145. Kubo, I., M. Uchida, and J. A. Klocke. 1983. An insect ecdysisinhibitor from the African medicinal plant, Plumbago capensis(Plumbaginaceae); a naturally occurring chitin synthetase in-hibitor. Agric. Biol. Chem. 47:911-913.
146. Kuranda, M. J., and P. W. Robbins. 1987. Cloning andheterologous expression of glycosidase genes from Saccharo-myces cerevisiae. Proc. Natl. Acad. Sci. USA 84:2585-2589.
147. Kuranda, M. J., and P. W. Robbins. 1991. Chitinase is requiredfor cell separation during growth of Saccharomyces cerevisiae.J. Biol. Chem. 266:19758-19767.
148. Larriba, G., M. Morales, and J. Ruiz-Herrera. 1981. Biosyn-thesis of beta-glucan microfibrils by cell free extracts fromSaccharomyces cerevisiae. J. Gen. Microbiol. 124:375-383.
149. Lee, J. W., P. Kelley, J. Lecciones, D. Coleman, R. Gordee,P. A. Pizzo, and T. J. Walsh. 1990. Cilofungin (LY121019)shows nonlinear plasma pharmacokinetics and tissue penetra-tion in rabbits. Antimicrob. Agents Chemother. 34:2240-2245.
150. Leighton, T., E. Marks, and F. Leighton. 1981. Pesticides:insecticides and fungicides are chitin synthesis inhibitors.Science 213:905-907.
151. Logan, D. A., J. M. Becker, and F. Naider. 1979. Peptide
transport in Candida albicans. J. Gen. Microbiol. 114:179-186.152. Manners, D. J., A. J. Masson, and J. C. Patterson. 1973. The
structure of a beta-(1,3)-D-glucan from yeast cell walls. Bio-chem. J. 135:19-30.
153. Manners, D. J., A. J. Masson, J. C. Patterson, H. Bjorndal, andB. Lindberg. 1973. The structure of a beta-(1,6)-D-glucan fromyeast cell wall. Biochem. J. 135:31-36.
154. Matsumoto, Y., S. Matsuda, and T. Tegoshi. 1989. Yeast glucanin the cyst wall of Pneumocystis carinii. J. Protozool. 36:21S.
155. McCarthy, P. J., D. J. Newman, L. J. Nisbet, and W. D.Kingsbury. 1985. Relative rates of transport of peptidyl drugsby Candida albicans. Antimicrob. Agents Chemother. 28:494-499.
156. McCarthy, P. J., L. J. Nisbet, J. C. Boehm, and W. D.Kingsbury. 1985. Multiplicity of peptide permeases in Candidaalbicans: evidence from novel chromophoric peptides. J. Bac-teriol. 162:1024-1029.
157. McCarthy, P. J., P. F. Troke, and K. Gull. 1985. Mechanism ofaction of nikkomycin and the peptide transport system ofCandida albicans. J. Gen. Microbiol. 131:775-780.
158. McIntyre, K. A., and J. N. Galgiani. 1989. pH and other effectson the antifungal activity of cilofungin (LY121019). Antimi-crob. Agents Chemother. 33:731-735.
159. Melchinger, W., and J. Muller. 1987. Studies on the in vitrosensitivity of yeast strains isolated from clinical specimens toLY 121019, a new antifungal agent. Mykosen 30:605-608.
160. Mendoza, C. G., and M. N. Ledieu. 1968. Chitin in the new wallof regenerating protoplasts of Candida utilis. Nature (London)220:1035.
161. Milewski, S., H. Chmara, and E. Borowski. 1986. Antibiotictetaine-a selective inhibitor of chitin and mannoprotein bio-synthesis in Candida albicans. Arch. Microbiol. 145:234-240.
162. Milewski, S., F. Mignini, and E. Borowski. 1991. Synergisticaction of nikkomycin X/Z with azole antifungals on Candidaalbicans. J. Gen. Microbiol. 137:2155-2161.
163. Miragall, F., H. Rico, and R. Sentandreu. 1988. Regenerationof the cell wall in protoplasts of Candida albicans: a cytochem-ical study using wheat germ agglutinin and concanavalin A.Arch. Microbiol. 149:286-290.
164. Mitani, M., and Y. Inoue. 1968. Antagonists of antifungalsubstance polyoxin. J. Antibiot. 21:492-496.
165. Mitchell, M. J., and S. L. Smith. 1988. Effect of the chitinsynthetase inhibitor plumbagin and its 2-demethyl derivativejuglone on insect eckysone 20-monooxygenase activity. Expe-rientia 44:990-991.
166. Miyata, M., T. Kanbe, and K. Tanaka. 1985. Morphologicalalterations of the fission yeast Schizosaccharomyces pombe inthe presence of aculeacin A: sphericle wall formation. J. Gen.Microbiol. 131:611-621.
167. Mizoguchi, J., T. Saito, K. Mizuno, and K. Hayano. 1977. Onthe mode of action of a new antifungal antibiotic, aculeacin A:inhibition of cell wall synthesis in Saccharomyces cerevisiae.J. Antibiot. 30:308-313.
168. Mizuno, K., A. Yagi, S. Satoi, M. Takada, M. Hayashi, K.Asano, and T. Matsuda. 1977. Studies on aculeacin. I. Isolationand characterization of aculeacin A. J. Antibiot. 30:297-302.
169. Molano, J., B. Bowers, and E. Cabib. 1980. Distribution ofchitin in the yeast cell wall: an ultrastructural and chemicalstudy. J. Cell. Biol. 85:199-212.
170. Molina, M., R. Cenamor, and C. Nombela. 1987. Exo-(1,3)-beta-glucanase activity in Candida albicans: effect of the yeastto mycelium transition. J. Gen. Microbiol. 133:609-617.
171. Montgomerie, J. Z., J. E. Edwards, Jr., and L. B. Guze. 1975.Synergism of amphotericin B and 5-fluorocytosine for Candidaspecies. J. Infect. Dis. 132:82-86.
172. Muller, H., R. Furter, H. Zahner, and D. M. Rast. 1981.Metabolic products of microorganisms. 203. Inhibition of chi-tosomal chitin synthetase and growth of Mucor rouxii bynikkomycin Z, nikkomycin X and polyoxin A: a comparison.Arch. Microbiol. 130:195-197.
173. Naider, F., P. Shenbagamurthi, A. S. Steinfeld, H. A. Smith, C.Boney, and J. M. Becker. 1983. Synthesis and biologicalactivity of tripeptidyl polyoxins as antifungal agents. Antimi-
18 HECTOR
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.

COMPOUNDS ACTIVE AGAINST CELL WALLS OF FUNGI 19
crob. Agents Chemother. 24:787-796.174. Nishimoto, Y., S. Sakuda, S. Takayama, and Y. Yamada. 1991.
Isolation and characterization of new allosamidins. J. Antibiot.44:716-722.
175. Noguchi, T., Y. Banno, T. Watanabe, Y. Nozawa, and Y. Ito.1975. Carbohydrate composition of the isolated cell walls ofdermatophytes. Mycopathologia 55:71-76.
176. Odds, F. C. 1988. Structure, physiology, and biochemistry ofCandida species, p. 22-59. In F. C. Odds (ed.), Candida andcandidosis: a review and bibliography. 2nd ed. Bailliere Tin-dall, London.
177. Odds, F. C. 1988. Introduction and historical note, p. 3-5. InF. C. Odds (ed.), Candida and candidosis: a review andbibliography, 2nd ed. Bailliere Tindall, London.
178. Oki, T. 1992. A new family of antibiotics: benzo[a]napthacene-quinones. A water-soluble pradimicin derivative, BMY-28864,p. 381-391. In H. Yamaguchi et al. (ed.), Recent progress inantifungal chemotherapy. Marcel Dekker, New York.
179. Old, T., M. Kakushima, M. Nishio, H. Kamel, M. Hirano, Y.Sawada, and M. Konishi. 1990. Water soluble pradimicinderivatives, synthesis and antifungal evaluation of N,N-dime-thyl pradimicins. J. Antibiot. 43:1230-1235.
180. Old, T., K. Saitoh, K. Tomatsu, K. Tomita, M. Konishi, and H.Kawaguchi. 1988. Novel antifungal antibiotic BMY-28567.Ann. N.Y. Acad. Sci. 544:184-187.
181. Oki, T., 0. Tenmyo, M. Hirano, K. Tomatsu, and H. Kamei.1990. Pradimicins A, B, and C: new antifungal antibiotics. II.In vitro and in vivo biological activities. J. Antibiot. 43:763-770.
182. Payne, J. W., K. J. Barrett-Bee, and D. A. Shallow. 1991.Peptide substrates rapidly modulate expression of dipeptideand oligopeptide permeases in Candida albicans. FEMS Mi-crobiol. Lett. 79:15-20.
183. Perez, P., R. Varona, I. Garcia-Acha, and A. Duran. 1981.Effect of papulacandin B and aculeacin A on beta-(1,3)-glucansynthase from Geotrichum lactis. FEBS Lett. 129:249-252.
184. Perfect, J. R., M. M. Hobbs, K. A. Wright, and D. T. Durrack.1989. Treatment of experimental disseminated candidiasis withcilofungin. Antimicrob. Agents Chemother. 33:1811-1812.
185. Perfect, J. R., K. A. Wright, and R. F. Hector. 1991. Syner-gistic interaction of nikkomycin and cilofungin against diversefungi, p. 369-379. In H. Yamaguchi et al. (ed.), Recentprogress in antifungal chemotherapy. Marcel Dekker, NewYork.
186. Peterson, E. M., R. J. Hawley, and R. A. Calderone. 1976. Anultrastructural analysis of protoplast-spheroplast induction inCryptococcus neoformans. Can. J. Microbiol. 22:1518-1521.
187. Pfaller, M., J. Riley, and T. Koerner. 1989. Effects of cilofun-gin (LY121019) on carbohydrate and sterol composition ofCandida albicans. Eur. J. Clin. Microbiol. Infect. Dis. 8:1067-1070.
188. Pfaller, M. A., J. Riley, and T. Koerner. 1990. Effects ofterconazole and other azole antifungal agents on the sterol andcarbohydrate composition of Candida albicans. Diagn. Micro-biol. Infect. Dis. 13:31-35.
189. Phaff, H. J. 1963. Cell wall of yeasts. Annu. Rev. Microbiol.17:15-30.
190. Pine, L., and C. J. Boone. 1968. Cell wall composition andserological reactivity of Histoplasma capsulatum serotypesand related species. J. Bacteriol. 96:789-798.
191. Polacheck, I., and R. F. Rosenberger. 1977. Aspergillus nidu-lans mutant lacking a-(1,3)-glucan, melanin, and cleistothecia.J. Bacteriol. 132:650-656.
192. Polacheck, Y., and R. F. Rosenberger. 1975. Autolytic enzymesin hyphae of Aspergillus nidulans: their action on old andnewly formed walls. J. Bacteriol. 121:332-337.
193. Polak, A., H. J. Scholer, and M. Wall. 1982. Combinationtherapy of experimental candidiasis, cryptococcosis and as-pergillosis in mice. Chemotherapy (Basel) 28:461-479.
194. Pollack, J. H., C. F. Lange, and T. Hashimoto. 1983. "Non-fibrillar" chitin associated with walls and septa of Trichophy-ton mentagrophytes arthrospores. J. Bacteriol. 154:965-975.
195. Poulain, D., G. Tronchin, J. F. Dubremetz, and J. Biguet. 1978.
Ultrastructure of the cell wall of Candida albicans blas-tospores: study of its constitutive layers by the use of acytochemical technique revealing polysaccharides. Ann. Mi-crobiol. (Inst. Pasteur) 129A:141-153.
196. Rast, D. M., and S. Bartnicki-Garcia. 1981. Effects of ampho-tericin B, nystatin, and other polyene antibiotics on chitinsynthase. Proc. Natl. Acad. Sci. USA 78:1233-1236.
197. Reiula, K. E. 1991. Two common fungi associated withfarmer's lung: fine structure of Aspergillus fiumigatus andAspergillus umbrosus. Mycopathologia 113:143-149.
198. Reiss, E. 1977. Serial enzymatic hydrolysis of cell walls of twoserotypes of yeast-form Histoplasma capsulatum with alpha-1,3-glucanase, beta-1,3-glucanase, pronase, and chitinase. In-fect. Immun. 16:181-188.
199. Reiss, E., S. E. Miller, W. Kaplan, and L. Kaufman. 1977.Antigenic, chemical, and structural properties of cell walls ofHistoplasma capsulatum yeast-form chemotypes 1 and 2 afterserial enzymatic hydrolysis. Infect. Immun. 16:690-700.
200. Reiss, E., D. G. Patterson, L. W. Yert, J. S. Holler, and B. K.Ibrahim. 1981. Structural analysis of mannans from Candidaalbicans serotypes A and B and from Torulopsis glabrata bymethylation gas chromatography, mass spectrometry and exo-alpha-mannanase. Biomed. Mass Spectrom. 8:252-255.
201. Richet, H. M., A. Andremont, C. Tancrede, J. L. Pico, andW. R. Jarvis. 1991. Risk factors for candidemia in patients withacute lymphocytic leukemia. Rev. Infect. Dis. 13:487-492.
202. Rouse, M. S., B. M. Tallan, J. M. Steckelberg, N. K. Henry,and W. R. Wilson. 1992. Efficacy of cilofungin therapy admin-istered by continuous intravenous infusion for experimentaldisseminated candidiasis in rabbits. Antimicrob. AgentsChemother. 36:56-58.
203. Ruiz-Herrerra, J., S. Bartnicki-Garcia, and C. E. Bracker.1980. Dissociation of chitosomes by digitonin into 16 S sub-units with chitin synthetase activity. Biochim. Biophys. Acta629:201-216.
204. Ryder, N. S., G. Seidl, and P. F. Troke. 1984. Effect of theantimycotic drug maftifine on growth of and sterol biosynthesisin Candida albicans. Antimicrob. Agents Chemother. 25:483-487.
205. Sakuda, S., A. Isogai, S. Matsumoto, and A. Suzuki. 1986. Thestructure of allosamidin, a novel insect chitinase inhibitor,produced by Streptomyces sp. Tetrahedron Lett. 27:2475-2478.
206. Sakuda, S., A. Isogai, S. Matsumoto, and A. Suzuki. 1987.Search for microbial insect growth regulators. II. Allosamidin,a novel insect chitinase inhibitor. J. Antibiot. 40:296-300.
207. Sakuda, S., Y. Nishimoto, M. Ohi, M. Watanabe, S. Takayama,A. Isogai, and Y. Yamada. 1990. Effects of demthylallosamidin,a potent yeast chitinase inhibitor, on the cell division of yeast.Agric. Biol. Chem. 54:1333-1335.
208. San-Bias, G., F. San-Blas, and L. E. Serrano. 1977. Host-parasite relationships in the yeastlike form of Paracoccidioidesbrasiliensis strain IVIC Pb9. Infect. Immun. 15:343-346.
209. Saral, R. 1991. Candida and Aspergillus infections in immuno-compromised patients: an overview. Rev. Infect. Dis. 13:487-492.
210. Sasaki, S., N. Ota, J. Eguchi, Y. Furukawa, T. Akashiba, T.Tsuchiyama, and S. Suzuki. 1968. Studies on polyoxins, anti-fungal antibiotics. VIII. Mechanism of action on sheath blightof rice plant. Ann. Phytopathol. Soc. Jpn. 34:272-279.
211. Satoi, S. A. Yagi, K. Asano, K. Mizuno, and T. Watanabe.1977. Studies on aculeacin. II. Isolation and characterization ofaculeacins B, C, D, E, F, and G. J. Antibiot. 30:303-307.
212. Sawada, Y., M. Hatori, H. Yamaoto, M. Nishio, T. Miyaki, andT. Oki. 1990. New antifungal antibiotics pradimicins FA-1 andFA-2: D-serine analogs of pradimicins A and C. J. Antibiot.43:1223-1229.
213. Sawada, Y., T. Murakami, T. Ueki, Y. Fukagawa, and T. Oki.1991. Mannan-mediated anticandidal activity of BMY-28864, anew water-soluble pradimicin derivative. J. Antibiot. 44:119-121.
214. Sawada, Y., M. Nishio, H. Yamamoto, M. Hatori, T. Miyaki,M. Konishi, and T. Oki. 1990. New antifungal antibiotics,
VOL. 6, 1993
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.

CLIN. MICROBIOL. REV.
pradimicins D and E: glycine analogs of pradimicins A andC. J. Antibiot. 43:771-777.
215. Sawada, Y., T. Tsuno, H. Yamamoto, M. Nishio, M. Konishi,and T. Oki. 1990. Pradimicins M, N, 0, and P, new dihy-drobenzo[a]naphthacenequinones produced by blocked mu-tants of Actinomadura hibisca P157-2. J. Antibiot. 43:1367-1374.
216. Sawistowska-Schroder, E. T., D. Kerridge, and H. Perry. 1984.Echinocandin inhibition of (1,3)-beta-D-glucan synthase fromCandida albicans. FEBS Lett. 173:134-138.
217. Selitrennikoff, C. P. 1979. Competitive inhibition of Neuros-pora crassa chitin synthetase activity by tunicamycin. Arch.Biochem. Biophys. 195:243-244.
218. Shah, V. K., and S. G. Knight. 1968. Chemical composition ofhyphal walls of dermatophytes. Arch. Biochem. Biophys.127:229-234.
219. Shallow, D. A., K. J. Barrett-Bee, and J. W. Payne. 1991.Evaluation of the dipeptide and oligopeptide permeases ofCandida albicans as uptake routes for synthetic anticandidalagents. FEMS Microbiol. Lett. 79:9-14.
220. Shaw, J. A., P. C. Mol, B. Boewers, S. J. Silverman, M. H.Valdivieso, A. Duran, and E. Cabib. 1991. The function ofchitin synthases 2 and 3 in the Saccharomyces cerevisiae cellcycle. J. Cell Biol. 114:111-123.
221. Shenbagamurthi, P., H. A. Smith, J. M. Becker, and F. Naider.1986. Synthesis and biological properties of chitin synthetaseinhibitors resistant to cellular peptidases. J. Med. Chem.29:802-809.
222. Shenbagamurthi, P., H. A. Smith, J. M. Becker, A. Steinfeld,and F. Naider. 1983. Design of anticandidal agents: synthesisand biological properties of analogues of polyoxin L. J. Med.Chem. 26:1518-1522.
223. Shepherd, M. G. 1988. Morphogenic transformation of fungi,p. 278-304. In M. R. McGinnis (ed.), Current topics in medicalmycology, vol. 2. Springer-Verlag, New York.
224. Smith, K. R., K. M. Lank, C. G. Cobbs, G. A. Cloud, andW. E. Dismukes. 1990. Comparison of cilofungin and ampho-tericin B for therapy of murine candidiasis. Antimicrob.Agents Chemother. 34:1619-1621.
225. Smith, K. R., K. M. Lank, W. E. Dismukes, and C. G. Cobbs.1991. In vitro comparison of cilofungin alone and in combina-tion with other antifungal agents against clinical isolates ofCandida species. Eur. J. Clin. Microbiol. Infect. Dis. 10:588-592.
226. Smith, W. L., T. Nakajima, and C. E. Ballou. 1975. Biosyn-thesis of yeast mannan: isolation of Kluyveromyces lactismannan mutants and a study of the incorporation of N-acetyl-D-glucosamine into the polysaccharide side chains. J. Biol.Chem. 250:3426-3435.
227. Sowa, B. A., and E. P. Marks. 1975. An in vitro system for thequantitative measurement of chitin synthesis in the cockroach:inhibition by TH 6040 and polyoxin D. Insect Biochem.5:855-859.
228. Spitzer, E. D., S. J. Travis, and G. S. Kobayashi. 1988.Comparative in vitro activity of Ly121019 and amphotericin Bagainst clinical isolates of Candida species. Eur. J. Clin.Microbiol. Infect. Dis. 7:80-81.
229. Stagg, C. M., and M. S. Feather. 1973. The characterization ofa chitin-associated D-glucan from the cell walls of Aspergillusniger. Biochim. Biophys. Acta 320:64-72.
230. Sternlight, E., D. Katz, and R. F. Rosenberger. 1973. Sub-apical wall synthesis and wall thickening induced by cyclohex-imide in hyphae ofAspergillus nidulans. J. Bacteriol. 114:819-823.
231. Strippoli, V., E. D. D'Auria, and N. Simonetti. 1988. A study ofthe antifungal activity of LY 121019, a new ehinocandinderivative. Chemioterapia 7:33-37.
232. Sullivan, P. A., Y. Y. Chiew, C. Molloy, M. D. Templeton, andM. G. Shepherd. 1983. An analysis of the metabolism and cellwall composition of Candida albicans during germ-tube forma-tion. Can. J. Biochem. 29:1514-1525.
233. Sullivan, P. A., N. J. McHugh, L. Roman, and M. G. Shepherd.1984. The secretion of N-acetylglucosaminidase during germ-
tube formation in Candida albicans. J. Gen. Microbiol. 130:2213-2218.
234. Sunayama, H., and S. Suzuki. 1970. Studies on the antigenicactivities of yeasts. VI. Analysis of the antigenic determinantsof the mannan of Candida albicans serotype B-792. Jpn. J.Microbiol. 14:371-379.
235. Surarit, R., P. K. Gopal, and M. G. Shepherd. 1988. Evidencefor a glycosidic linkage between chitin and glucan in the cellwall of Candida albicans. J. Gen. Microbiol. 134:1723-1730.
236. Surarit, R., and M. G. Shepherd. 1987. The effects of azole andpolyene antifungals on the plasma membrane enzymes ofCandida albicans. J. Med. Vet. Mycol. 25:403-413.
237. Suzuki, M., and Y. Fukazawa. 1982. Immunochemical charac-terization of Candida albicans cell wall antigens: specificdeterminant of Candida albicans serotype A mannan. Micro-biol. Immunol. 26:387-402.
238. Suzuki, S., K. Isono, J. Nagatsu, T. Mizutani, Y. Kawashima,and T. Mizuno. 1965. A new antibiotic, polyoxin A. J. Antibiot.18:131.
239. Suzuki, S., H. Sunayama, and T. Saito. 1968. Studies on theantigenic activity of yeasts. I. Analysis of the determinantgroups of the mannan of Saccharomyces cerevisiae. Jpn. J.Microbiol. 12:19-24.
240. Taft, C. S., and C. P. Selitrennikoff. 1988. LY121019 inhibitsNeurospora crassa growth and (1-3)-p-D-glucan synthase. J.Antibiot. 41:697-701.
241. Taft, C. S., T. Stark, and C. P. Selitrennikoff. 1988. Cilofungin(LY121019) inhibits Candida albicans (1-3)-r3-D-glucan syn-thase activity. Antimicrob. Agents Chemother. 32:1901-1903.
242. Takeuchi, T., T. Hara, H. Naganawa, M. Okada, M. Hamada,H. Umezawa, S. Gomi, M. Sezaki, and S. Kondo. 1989. Newantifungal antibiotics, benanomicins A and B from an actino-mycete. J. Antibiot. 41:807-811.
243. Tang, J., and T. R. Parr, Jr. 1991. W-1 solubilization andkinetics of inhibition by cilofungin of Candida albicans (1,3)-P-D-glucan synthase. Antimicrob. Agents Chemother. 35:99-103.
244. Thomas, K. R., B. Davis, and J. Mills. 1979. The effect ofbeta-glucuronidase and chitinase on the cell wall ofAspergillusniger and Aspergillus fumigatus. Microbios 25:111-123.
245. Tomita, K., M. Nishio, K. Saitoh, H. Yamamoto, Y. Hoshino,H. Ohkuma, M. Konishi, T. Miyaki, and T. Old. 1990.Pradimicins A, B, and C: new antifungal antibiotics. I. Taxon-omy, production, isolation and physico-chemical properties. J.Antibiot. 43:755.
246. Traber, R., C. Keller-Juslen, H. R. Loosli, M. Huhn, and A.Von Wartburg. 1979. Cyclopeptid-antibiotika aus AspergillusArten. Structur der Echinocandine C und D. Helv. Chim. Acta62:1252-1267.
247. Traxier, P., H. Fritz, H. Fuhrer, and W. J. Richter. 1980.Papulacandins, a new family of antibiotics with antifungalactivity. J. Antibiot. 33:967-978.
248. Traxler, P., J. Gruner, and J. A. L. Auden. 1977. Papulacan-dins, a new family of antibiotics with antifungal activity. I.Isolation, chemical and biological characterization of papula-candins A, B, C, D, and E. J. Antibiot. 30:289-296.
249. Traxler, P., W. Tosch, and 0. Zak. 1987. Papulacandins,synthesis and biological activity of papulacandin B derivatives.J. Antibiot. 40:1146-1164.
250. Tronchin, G., D. Poulain, J. Herbaut, and J. Biguet. 1981.Localization of chitin in the cell wall of Candida albicans bymeans of wheat germ agglutinin. Fluorescence and ultrastruc-tural studies. Eur. J. Cell Biol. 26:121-128.
251. Uno, J., M. L. Shigematsu, and T. Arai. 1983. Novel synergismof two antifungal agents, copiamycin and imidazole. Antimi-crob. Agents Chemother. 24:552-559.
252. Uramoto, M., K. Kobinata, K. Isono, T. Higashiima, T.Miyazawa, E. E. Jenkins, and J. A. McCloskey. 1980. Struc-tures of neopolyoxins A, B, and C. Tetrahedron Lett. 21:3395-3398.
253. Uramoto, M., K. Kobinata, K. Isono, T. Higashiima, T.Miyazawa, E. E. Jenkins, and J. A. McCloskey. 1982. Chemis-try of the neopolyoxins. Pyrimidine and imidazoline nucleoside
20 HECTOR
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.

COMPOUNDS ACTIVE AGAINST CELL WALLS OF FUNGI 21
peptide antibiotics. Tetrahedron 18:1599-1608.254. Valdivieso, M. H., P. C. Mol, J. A. Shaw, E. Cabib, and A.
Duran. 1991. CAL1, a gene required for activity of chitinsynthase 3 in Saccharomyces cerevisiae. J. Cell Biol. 114:101-109.
255. Vanden Bossche, H. 1985. Biochemical targets for antifungalazole derivatives: hypothesis on the mode of action, p. 313-351. In M. R. McGinnis (ed.), Current topics in medicalmycology, vol. 1. Springer-Verlag, New York.
256. VanMiddlesworth, F., M. N. Omstead, D. Schmatz, K. Bartizal,R. Fromtling, G. Bills, K. Nollstadt, S. Honeycutt, M. Zwe-erink, G. Garrity, and K. Wilson. 1991. L-687,781, a newmember of the papulacandin family of B-1,3-D-glucan synthe-sis inhibitors. I. Fermentation, isolation, and biological activ-ity. J. Antibiot. 44:45-51.
257. Vartivarian, S. E., G. H. Reyes, E. S. Jacobson, P. G. James, R.Cherniak, V. R. Mumaw, and M. J. Tingler. 1989. Localizationof mannoprotein in Cryptococcus neofonnans. J. Bacteriol.171:6850-6852.
258. Weiman, A. C. M. 1979. Carbohydrate composition andtaxonomy of Geotrichum, Tnchosporon and allied genera.Antonie van Leeuwenhoek 45:119-127.
259. Wellinga, K., R. Mulder, and J. J. van Daalen. 1973. Synthesisand laboratory evaluation of 1-(2,6-disubstituted benzoyl)-3-phenylureas, a new class of insecticides. I. 1-(2,6-Dichloroben-zoyl)-3-phenylureas. J. Agric. Food Chem. 21:348-354.
260. Wheat, R., and E. Scheer. 1977. Cell walls of Coccidioidesimmitis: neutral sugars of aqueous extract polymers. Infect.Immun. 15:340-341.
261. Wheat, R., T. Terai, A. Kiyomoto, N. F. Conant, E. P. Lowe,and J. Converse. 1967. Studies on the composition and struc-ture of Coccidioides immitis cell walls, p. 237-242. In L. Ajello(ed.), Proceedings of the Second Coccidioidomycosis Sympo-sium. University of Arizona Press, Tucson.
262. Wheat, R. W., C. Tritschler, N. F. Conant, and E. P. Lowe.1977. Comparison of Coccidioides immitis arthrospore, myce-lium, and spherule cell walls, and influence of growth medium
on mycelial cell wall composition. Infect. Immun. 17:91-97.263. Wheeler, M. H., and A. A. Bell. 1988. Melanins and their
importance in pathogenic fungi, p. 338-387. In M. R. McGinnis(ed.), Current topics in medical mycology, vol. 2. Springer-Verlag, New York.
264. Wuesada-Allue, L. A. 1982. The inhibition of insect chitinsynthesis by tunicamycin. Biochem. Biophys. Res. Commun.105:312-319.
265. Wu-Yean, C. D., and T. Hashimoto. 1977. Architecture andchemistry of microconidial walls of Trichophyton mentagro-phytes. J. Bacteriol. 129:1584-1592.
266. Yadan, J.-C., M. Gonneau, P. Sarthou, and F. Le Goffic. 1984.Sensitivity to nikkomycin Z in Candida albicans: role ofpeptide permeases. J. Bacteriol. 160:884-888.
267. Yamaguchi, H., S. Kondo, and T. Takeuchi. 1992. A new familyof antibiotics: benzo[a]naphthacenequinones. A novel antibi-otic, benanomicin A, p. 393-402. In H. Yamaguchi et al. (ed.),Recent progress in antifungal chemotherapy. Marcel Dekker,New York.
268. Yoshida, M., M. Ezaki, M. Hashimoto, M. Yamashita, N.Shigematsu, M. Okuhara, M. Kohsaka, and K. Horikoshi. 1990.A novel antifungal antibiotic, FR-900848. I. Production, isola-tion, physico-chemical and biological properties. J. Antibiot.43:748-754.
269. Zlotnik, H., M. P. Fernandez, B. Bowers, and E. Cabib. 1984.Saccharomyces cerevisiae mannoproteins from an externalcell wall layer that determines wall porosity. J. Bacteriol.159:1018-1026.
270. Zoebelein, G. 1981. Freilandversuche mit AMS 0896 gegenSpinnmilben im Obstbau. Mitt. Dtsch. Ges. Allg. Angew.Entomol. 2:164-171.
271. Zonneveld, B. J. M. 1971. Biochemical analysis of the cell wallofAspegillus nidulans. Biochim. Biophys. Acta 249:506-514.
272. Zonneveld, B. J. M. 1972. Morphogenesis in Aspergillus nidu-lans. The significance of alpha-(1,3) glucan of the cell wall andalpha-(1,3) glucanase for cleistothecium development. Bio-chim. Biophys. Acta 273:174-187.
VOL. 6, 1993
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/c
mr
on 2
1 D
ecem
ber
2021
by
2409
:8a2
0:54
6a:d
db0:
f038
:fab
b:d9
a0:7
9a5.