Embed Size (px)
Citation preview

NEUROBIOLOGY OF LEARNING AND MEMORY 70, 197–211 (1998)ARTICLE NO. NL983848
Consolidation of Visual Associative Long-TermMemory in the Temporal Cortex of Primates
Yasushi Miyashita,*,† Masashi Kameyama,* Isao Hasegawa,* andTetsuya Fukushima*
*Department of Physiology, The University of Tokyo School of Medicine, 7-3-1,Hongo, Bunkyo-ku, Tokyo 113, Japan; and †Mind Articulation Project, ICORP,
Japan Science and Technology Corporation, Yushima, Tokyo 113, Japan
Neuropsychological theories have proposed a critical role for the interaction betweenthe medial temporal lobe and the neocortex in the formation of long-term memory forfacts and events, which has often been tested by learning of a series of paired wordsor figures in humans. We have examined neural mechanisms underlying the memory‘‘consolidation’’ process by single-unit recording and molecular biological methods inan animal model of a visual pair-association task in monkeys. In our previous studies,we found that long-term associative representations of visual objects are acquiredthrough learning in the neural network of the anterior inferior temporal (IT) cortex.In this article, we propose the hypothesis that limbic neurons undergo rapid modifica-tion of synaptic connectivity and provide backward signals that guide the reorganiza-tion of neocortical neural circuits. Two experiments tested this hypothesis: (1) we exam-ined the role of the backward connections from the medial temporal lobe to the ITcortex by injecting ibotenic acid into the entorhinal and perirhinal cortices, whichprovided massive backward projections ipsilaterally to the IT cortex. We found thatthe limbic lesion disrupted the associative code of the IT neurons between the pairedassociates, without impairing the visual response to each stimulus. (2) We then testedthe first half of this hypothesis by detecting the expression of immediate-early genesin the monkey temporal cortex. We found specific expression of zif268 during the learn-ing of a new set of paired associates in the pair-association task, most intensively inarea 36 of the perirhinal cortex. All these results with the visual pair-association tasksupport our hypothesis and demonstrate that the consolidation process, which was firstproposed on the basis of clinico-psychological evidence, can now be examined in pri-mates using neurophysiolocical and molecular biological approaches. q 1998 Academic Press
Key Words: memory consolidation; inferior temporal cortex; perirhinal cortex; pairassociation; ibotenic acid; immediate-early genes; transcription factor; zif268.
The cerebral cortex is organized so that hierarchically different subareascommunicate by forward and backward neuronal projections (Van Essen etal., 1992). Neuropsychological studies have proposed that the backward con-nections from the medial temporal lobe to the neocortices play a role in theformation of long-term memory for facts and events (declarative memory) (Mish-kin, 1982; Squire, 1987). Behaviorally, the declarative memory has often beentested by learning of a series of paired words or figures in humans (Squire,
Address correspondence and reprint requests to Professor Yasushi Miyashita, Department ofPhysiology, The University of Tokyo School of Medicine, 7-3-1, Hongo, Bunkyo-ku, Tokyo 113,Japan. Fax: /81-3-5800-6847. E-mail: [email protected].
197 1074-7427/98 $25.00Copyright q 1998 by Academic Press
All rights of reproduction in any form reserved.
AID NLM 3848 / 6v16$$$$81 09-10-98 09:02:12 nlmoa AP: NLM

198 MIYASHITA ET AL.
1987; Wechsler, 1987). We have been examining the neural mechanisms under-lying declarative memory by single-unit recording with an animal model of avisual pair-association task in monkeys.
In our previous studies, we provided several lines of evidence indicating thatlong-term representations of visual objects are acquired through learning inthe neural network of the anterior inferior temporal (IT) cortex in primates(Miyashita, 1993; Miyashita, 1995; Higuchi & Miyashita, 1996; Naya, Sakai, &Miyashita, 1996). We identified two mechanisms: one is tuning and the otheris association. Specifically, in the IT cortex of monkeys performing the visualpair-association task, we found a group of neurons that manifested selectiveresponses to both of the paired associates (pair-coding neurons). It providesstrong evidence that single IT neurons acquire stimulus selectivity throughassociative learning (Miyashita, 1993; Sakai & Miyashita, 1991).
In this article, we propose the hypothesis that limbic–neocortical interac-tions are critical in the consolidation process since limbic neurons undergorapid modification of synaptic connectivity and provide backward signals thatguide the reorganization of neocortical neural circuits. Two experiments werecarried out to test the hypothesis. First, we tested the role of the backwardsignals from the medial temporal lobe to the IT cortex. Ibotenic acid wasinjected unilaterally into the entorhinal and perirhinal cortices, which pro-vided massive backward projections ipsilateral to the IT cortex. We found that(i) in spite of the lesion, the sampled neurons responded strongly and selec-tively to the visual patterns and (ii) the paired associates failed to elicit signifi-cantly correlated responses in the cells tested with the lesion. The results showthat the limbic lesion disrupted the associative code of the IT neurons betweenthe paired associates, without impairing the visual response to each stimulus.
Second, we ask why limbic–neocortical interactions are so important. Wehypothesize that limbic neurons undergo rapid modification of synaptic connec-tivity and provide backward signals that guide the reorganization of neocorti-cal neural circuits. We then investigated the molecular basis of such rapidsynaptic modifiability by detecting the expression of immediate-early genes inthe monkey temporal cortex. We found strong expression of zif268 during thelearning of a new set of paired associates in the pair-association task. Thisexpression was quite selective, since we found no zif268 expression in a controlcondition, i.e., during the learning of a new stimulus set in a visual discrimina-tion task. The results suggest that the expression of zif268 plays a role in therapid synaptic modification in the inferior temporal gyrus.
EXPERIMENT 1
There is strong evidence that the IT cortex receives massive backward projec-tions from the medial temporal lobe, especially from the perirhinal and entorhi-nal cortices (Van Hoesen, 1982; Webster et al., 1991). The hypothesis for therole of the backward connections in the consolidation process specifically pre-dicts that the lesion of the perirhinal and entorhinal cortex would impair theformation of the associative code for pictures in the IT cells (Fig. 1). We nowtest this prediction.
We combined single-unit recording techniques with a surgically manipulatedchronic monkey preparation so that individual IT neurons become devoid ofthe backward signals but can receive forward visual signals when visual stim-
AID NLM 3848 / 6v16$$$$81 09-10-98 09:02:12 nlmoa AP: NLM

199CONSOLIDATION OF VISUAL MEMORY IN THE TEMPORAL CORTEX
FIG. 1. Backward connections from the limbic system to the inferior temporal cortex. The inferiortemporal cortex receives the forward visual signal from the prestriate cortices as well as the backwardsignal from the limbic system. What is the role of this backward signal in memory formation?
uli are presented. We prepared a specific experimental design to run thisproject (Fig. 2). Backward neuronal connections from the limbic system to theIT cortex are interrupted by the lesion of the perirhinal/entorhinal cortices.However, bilateral lesion would impair the monkeys’ behavior in the pair-association task (Murray et al., 1993). Thus the perirhinal/entorhinal corticesshould be lesioned unilaterally. With the unilateral lesion, however, the ITcortex of the lesioned side could receive information from the contralateral ITcortex since receptive fields of IT neurons cover bilateral hemifields before
FIG. 2. Experimental design. Backward connections from the limbic system to the inferior tempo-ral cortex are interrupted by lesion of the perirhinal/entorhinal cortices. However, bilateral lesionwould impair the monkeys’ behavior in the pair-association task. Thus the perirhinal/entorhinalcortices should be lesioned unilaterally. With the unilateral lesion, however, the inferior temporalcortex of the lesioned side could receive information from the contralateral inferior temporal cortexsince receptive fields of the inferior temporal neurons cover bilateral hemifields before the lesion. Inorder to remove the interhemispheric signal from the healthy contralateral inferior temporal cortex,we surgically cut the anterior commissure at the beginning of the experiment.
AID NLM 3848 / 6v16$$$$82 09-10-98 09:02:12 nlmoa AP: NLM

200 MIYASHITA ET AL.
the lesion. In order to remove the interhemispheric signal from the healthycontralateral IT cortex, we surgically cut the anterior commissure at the begin-ning of the experiment. With this surgically manipulated chronic monkey prep-aration, we found selective loss of the ability of IT neurons to represent associa-tions between picture pairs, while the ability of the neurons to respond toparticular visual stimuli was left intact.
Methods of Experiment 1
Animals and surgical procedures. Two adult monkeys (Macaca fuscata)were used in accordance with the University of Tokyo Animal Care Committee.Surgery of the anterior commissure (AC) was carried out under aseptic condi-tions and anesthesia with sodium pentobarbital (approximately 25–30 mg/kg). The right hemisphere was retracted from the falx with a brain spoon. Anaspirator was used to make a sagittal incision no more than 5 mm in lengthin the corpus callosum, entering the lateral ventricle at the level of the inter-ventricular foramen. The AC was sectioned at the floor of the lateral ventriclelateral to the fornix.
The lesions of the entorhinal and perirhinal cortices were made by injectinga small volume of the neurotoxin, ibotenic acid. Ibotenic acid has been reportedto kill cell bodies selectively while leaving fibers of passage in the underlyingwhite matter unharmed (Olney, 1983). A Hamilton syringe needle was insertedthrough a guide tube under the control of microdrives. A grid of 1-ml injectionsof ibotenic acid (15 mg/ml, 0.03 ml/min) was made with approximately 2-mmspacing. One monkey (M) received injections at 22 tracks, and the other mon-key (N) received injections at 19 tracks.
Behavioral task. The pair-association task was described previously (Sa-kai & Miyashita, 1991). Colored fractal patterns were used as the visual stim-uli. We generated the patterns according to the algorithm described previously(Miyashita et al., 1991) and arranged them into 12 pairs. Sorting into pairswas basically random, avoiding apparent geometrical resemblances such asrotational symmetry.
In the task, each trial begins with the monkey continually pressing a leverin front of him. Following a fixation stimulus, a cue stimulus was presentedin the center of the video monitor for 0.5 s. The cue stimulus was selectedrandomly from the 24 pictures in the set. After a delay period of 4 s, twostimuli for choice, the paired associate of the cue and one from a different pair,were shown randomly in two of four possible positions (arranged in two rowsof two columns). The monkey obtained fruit juice as a reward for touching thecorrect paired associate within 1.2 s. If the monkey released the lever beforethe choice, that trial was aborted. Eye position was monitored with a magneticsearch coil. The criterion for acquisition of prelesion learning (the set A stimuli)was 2 consecutive days of 26 correct responses in 30 trials. The two monkeystook 326 { 57 trials per picture to reach this criterion. The criterion after theibotenic acid lesion was set to be slightly easier, to 2 consecutive days of 23correct responses in 30 trials. In relearning of the set A paired associates,10 and 50 trials per picture were necessary for monkey M and monkey N,respectively. In learning of the new set (set B) after the lesion, one monkey(M) took 101 trials and the other (N) took 93 trials per picture to reach thecriterion.
AID NLM 3848 / 6v16$$$$82 09-10-98 09:02:12 nlmoa AP: NLM

201CONSOLIDATION OF VISUAL MEMORY IN THE TEMPORAL CORTEX
Electrophysiology and data collection. Recording and analysis of extracellu-lar spike discharges of single neurons were described in a previous publication(Miyashita et al., 1989). After the monkey has learned the task, a head-holdingdevice, scleral eye coil, and a recording chamber for microelectrode were im-planted under aseptic conditions while the animal was anesthetized with so-dium pentobarbital. The activity of single neurons was recorded with a glass-insulated tungsten microelectrode. The electrode was inserted through theintact dura along a stainless steel guide tube by means of a hydraulic micro-drive manipulator (MO-95, Narishige). The action potentials of single cellswere amplified, passed through band-pass filter circuits (30–10 kHz), andconverted into digital pulses by a time-window discriminator (EN-611J, Nihon-koden).
At the beginning of unit recording, locations of responsive cells were mappedin the anterior IT cortex, and an area (about 2 1 3 mm, hatched box in Fig.2) with a high density of responsive cells was identified; then all neuronalrecordings were done from this cortical area both before and after the lesionwas made. Each location of the electrode track was measured by X-ray imagingand verified histologically at the end of the experiments with reference toelectrolytic lesion marks made by applying DC currents of 15–20 mA (tipnegative) for 60–120 s.
Data analysis. Evaluation of cue responses was done by collecting spikenumbers over 80–480 ms at the beginning of the cue interval for each pictureand over 400 ms immediately preceding the appearance of the fixation stimu-lus. The neuron was regarded as responsive if the two distributions weresignificantly different (paired t test, p õ .05).
The pair index, PI, was used to analyze pair-coding responses of these cells.The PI was defined in a previous study (Sahai & Miyashita, 1991). The re-sponse variability of the cells was evaluated before and after the lesion by aresponse variability index. The index was defined in each cell as the ratio ofthe standard deviation of firing rates among the trials for the optimum stimu-lus over the mean firing rate for the optimum stimulus. The larger values ofthis index indicate more trial-to-trial fluctuations of the responses.
Histology. At the conclusion of the experiment, the monkey was deeplyanesthetized with an overdose of sodium pentobarbital (45 mg/kg, im) andthen perfused through the aorta with 0.9% saline followed by 10% formalin inphosphate buffer (pH 7.4). The brain was removed from the skull and sectionedcoronally at 40 mm on a freezing microtome. Serial sections were stained withcresyl violet as well as with the modified Gallyas method. We did not observesigns of gliosis, cell loss, or demyelination in the cortex of the recording sitesexcept for some electrode tracks.
Results of Experiment 1
Two adult monkeys (M. fuscata) were trained in a pair-association task.Before learning of the task, the anterior commissure (AC) of the monkey wastransected. The septal nuclei and fornix (Fx), as well as the underlying dience-phalic structures, were spared. This surgery disconnected the anterior tempo-ral cortex of each hemisphere from the other (Fig. 2), since most commissuralfibers of this area traverse the AC rather than the corpus callosum (Pandya
AID NLM 3848 / 6v16$$$$82 09-10-98 09:02:12 nlmoa AP: NLM

202 MIYASHITA ET AL.
et al., 1973; Demeter et al., 1990). After recovery, the monkey was trainedwith a set (set A) of the paired associates to the criterion performance level,and then extracellular spike discharges of single neurons were recorded as aprelesion control in the anterior IT cortex, as reported in previous studies(Miyashita, 1988; Miyashita & Chang, 1988; Sakai & Miyashita, 1991).
We deprived IT neurons of backward neural information by unilateral ibo-tenic acid lesions (see Methods) of the entorhinal and perirhinal cortices thatprovide massive backward projections ipsilateral to the IT cortex (Van Hoesen,1982; Webster et al., 1991). The lesion covered both the medial and the lateralbanks of the rhinal sulcus completely and most of the entorhinal and perirhinalcortex. The lesioned cortex suffered atrophy, but fibers in the underlying whitematter were left unharmed.
After the ibotenic acid injection, the monkeys were trained with the preoper-atively learned set (set A) and a new set of paired associates (set B) to thecriterion of 2 consecutive days of 23 correct responses in 30 trials. Then extra-cellular spike discharges of single neurons were recorded from the same areaas that in the prelesion control. Average performance for set A stimuli duringneural recording was 81% (before lesion) and 80% (after lesion), and that forset B stimuli was 72% after lesion.
Prior to the ibotenic acid lesion, we examined the responses of 92 cells to theset A stimuli. The distribution of the PI in the 92 cells showed that the pairedassociates elicited significantly correlated responses in the control (p õ .01; Wil-coxon’s signed-rank test, n Å 92), which confirmed the conclusion of our previousreport (Sakai & Miyashita, 1991). After the lesion, we examined the responsesof 72 cells to the set A stimuli and those of 75 cells to the set B stimuli. Thedistribution of the PI of these cells showed that the paired associates did notelicit significantly correlated responses for either the set A or the set B stimuli.
Figure 3 compares the responsiveness of the cells in the three groups (two-way ANOVA and t test). The results of the two monkeys were not significantlydifferent (p ú .25) in a two-way ANOVA for any of the following indices, F(1,233) Å 1.11 for PI, F(1, 233) Å 0.69 for SPON, F(1, 233) Å 0.00 for MAX, F(1,233) Å 1.05 for VAR, and have been pooled. After the lesion of the entorhinaland perirhinal cortices, the cells responded to both the set A pictures (MAX,35 { 2 spikes/s; mean { SEM, n Å 72) and the set B pictures (38 { 2; n Å75), even more strongly than the control (31 { 1; n Å 92). However, the PIwere reduced (p õ .05) after the lesion (0.04 { 0.35 for set A, 00.19 { 0.27for set B) compared to that of control (1.00 { 0.33). The spontaneous dischargerates (SPON) were not significantly different among the three groups (p õ.2). We also tested with a response variability index (VAR) whether the cellsexhibited any sign of pathological firings after the lesion. The larger values ofthis index indicate more trial-to-trial fluctuations of the responses (Methods).The index did not differ significantly between the control group (0.25 { 0.01)and the lesioned groups either with set A (0.28 { 0.02; p ú .2) or with set Bstimuli (0.26 { 0.02; p ú .6). We conclude that lesion of the entorhinal andperirhinal cortices disrupted the associative code of the IT neurons betweenthe paired associates, without impairing the visual response to each stimulus.
EXPERIMENT 2
Immediate-early genes (IEGs), a class of genes that show rapid and transientbut protein synthesis-independent increases in transcription, have been pro-
AID NLM 3848 / 6v16$$$$82 09-10-98 09:02:12 nlmoa AP: NLM

203CONSOLIDATION OF VISUAL MEMORY IN THE TEMPORAL CORTEX
FIG. 3. Effects of the lesion of the entorhinal and perirhinal cortices upon neuronal respon-siveness in the pair-association task. (A) Pair index (PI). (B) Spontaneous discharge rate (SPON).(C) Maximum discharge rate with spontaneous discharge subtracted (MAX). (D) Response variabil-ity (VAR). These indices were compared between two groups: (1) neurons that were recorded priorto the lesion using the set A stimuli (stippled bar, n Å 92) and (2) those recorded after the lesionusing the set B stimuli (hatched with thin line, nÅ 75). Data are shown as means{ SEM (modifiedfrom H. Okuno & Y. Miyashita, 1996, Expression of the transcription factor Zif268 in the temporalcortex of monkeys during visual paired associate learning, European Journal of Neuroscience, 8,2118–2128, Blackwell Science Ltd.).
posed to be involved in genomic responses in neurons related to learning andmemory (Morgan & Curran, 1991). A number of IEGs encode transcriptionfactors such as Fos, Jun, and Zif268, and these IEGs, especially zif268, havebeen hypothesized to play crucial roles in the transduction of neuronal electricsignal into more permanent synaptic organization. For example, the inductionof zif268 in the dentate gyrus is closely correlated with the induction of long-term potentiation (LTP) (Abraham et al., 1993; Worley et al., 1993). In song-birds, zif268 is strongly induced in specific regions of the auditory telencepha-lon when the birds hear the song of their own species, but induced only slightlywhen the birds hear songs of other species (Mello et al., 1992). However, therehave been no reports that seek to determine the roles of IEGs in the consolida-tion process of cognitive memories in primates.
AID NLM 3848 / 6v16$$$$82 09-10-98 09:02:12 nlmoa AP: NLM

204 MIYASHITA ET AL.
In the present study, we attempted to investigate brain areas in which IEGswere expressed during visual long-term memory formation in the primate. Wetrained monkeys to learn two different cognitive memory tasks, a visual pair-association task and a visual discrimination task. The visual pair-associationtask required the monkeys to memorize visual stimulus–stimulus associations(Sakai & Miyashita, 1991; Murray et al., 1993). The other task, the visualdiscrimination task, required them to memorize stimulus–reward associations(Iwai & Mishkin, 1969). The types of learning required in these tasks aredifferent since they are sensitive to lesions in different brain areas and thevisual pair-association task was more sensitive to the medial temporal lesion(Iwai & Mishkin, 1969; Murray et al., 1993; Suzuki et al., 1993). This factprompted us to test the roles of IEGs in the consolidation process of cognitivememories in primates. We found selective expression of zif268 only during thelearning of the pair-association task.
Methods of Experiment 2
Animals and visual memory tasks. The subjects were six male macaquemonkeys (M. fuscata), all weighing between 7.0 and 9.6 kg at the time ofperfusion. Three monkeys were trained to learn a visual pair-association taskand the others were trained to learn a visual discrimination task. A set of 12pairs of the Fourier descriptors was used as a visual stimulus set in both pair-association and visual discrimination tasks. The procedure of the visual pair-association task was essentially the same as described under Experiment 1except that monkeys sequentially learned the training set and the test set ofpaired associates in this study. In the other task, the visual discriminationtask, monkeys were required to touch the rewarded stimulus to obtain fruitjuice. The monkeys also sequentially learned the training and test sets ofvisual stimuli in this task.
The monkeys were first trained to perform the tasks with a set of stimuluspictures (training set). After the monkeys had learned the task rule with thetraining set, a new set of stimuli (test set) was introduced. The monkeysexecuted approximately 600 trials in a training session each day in both thepair-association task and the discrimination task after the introduction of thetest set. On the 8th to 10th day with the test set, the monkeys learning thepair-association task were anesthetized and immediately perfused followingthat day’s training session as described below. The monkeys learning the dis-crimination task were perfused after the training session on the 5th or 6thday. The monkeys in the both groups were exposed to an equivalent numberof visual stimuli (pair-association, 684 { 102 stimuli per hour; discrimination,664 { 72 stimuli per hour) during the 2 h before the perfusion.
Tissue preparation, immunohistochemistry, and antibodies. After thetraining session on the perfusion day, the monkeys were immediately givenan overdose of pentobarbital (ú40 mg/kg) and then perfused transcardially.The brain blocks were frozen in dry-ice powder, and sections (32 mm) were cutusing a cryostat and stained immunohistochemically as described previously(Okuno et al., 1995). In brief, after being washed with PBS, the sections wereincubated with blocking solution containing PBS plus 10% normal goat serumand 0.3% Triton X-100 for 60 min at room temperature, followed by incubationwith blocking buffer containing the primary antibody (see below) at 47C for 24
AID NLM 3848 / 6v16$$$$82 09-10-98 09:02:12 nlmoa AP: NLM

205CONSOLIDATION OF VISUAL MEMORY IN THE TEMPORAL CORTEX
h and then by incubation with blocking buffer containing horseradish peroxi-dase-conjugated goat anti-rabbit IgG antibody (Jackson ImmunoResearch;Catalog No. 111-035-003; diluted 1:400) at room temperature for 2 h. Afterbeing washed, the sections were reacted with coloring solution (0.2 mg/ml 3,3 *-diaminobenzidine tetrahydrochloride, 0.05% H2O2, 0.08% nickel chloride in 50mM Tris–Cl, pH 7.4) for 10 min at room temperature.
All primary antibodies used in this study were rabbit polyclonal antibodies.An anti-Zif268 antibody was raised against a synthetic oligopeptide (19 aminoacid residues) corresponding to the carboxy terminus of Zif268. The antigenicsequence is evolutionally conserved among various species, including rodentsand humans. The specificity and reactivity of the anti-Zif268 antibody to themonkey zif268 gene product were determined previously (Okuno et al., 1995);the anti-Zif268 antibody specifically recognized an 86-kDa protein in the nu-clear extracts from monkey cerebral cortex in immunoprecipitation experi-ments. An anti-c-Fos antibody was purchased from Oncogene Science Inc.(Catalog No. PC05).
Image analysis. To visualize the spatial distribution of Zif268, an image(resolution 4096 1 4096 pixels for 1.57 1 102 mm2) was obtained from each ofthe serial Zif268-immunostained sections at 0.5-mm intervals (20 sections foreach subject) by a CCD camera attached to a microscope, and optical density(OD) was measured in each pixel (pixel OD) using a computer-aided imageanalyzer (IBAS-V2.0, Zeiss).
The cortical area (layers II to VI) in each section was segmented into 0.5-mm-wide strips radially from the white matter to the cortical layer I (see Fig.4A, left). In each segment, the pixel ODs could be classified into two groups,pixel ODs originating from Zif268-immunostaining reaction products in thenuclei and those originating from background staining. The pixel ODs originat-ing from background staining were estimated by fitting a Gaussian distributionto the lowest peak of the distribution of pixel OD using the maximum-likelihoodmethod. The density of Zif268-immunostaining reaction products in each seg-ment was calculated by summing all pixel ODs after subtracting ODs originat-ing from background staining and by normalizing to the area of the segment.The segments were reconstructed into a straight-line, unfolded, two-dimen-sional map (see Fig. 4A, right), essentially according to the proceduresdescribed by Van Essen and Maunsell (1980). The density of the immunostain-ing reaction products is indicated on the unfolded map in pseudocolor represen-tation.
Results of Experiment 2
To investigate the formation of associative memory of the visual stimuli butnot skill-based or habit-like memory incidental to the task paradigm, we firsttrained the monkeys to learn a rule or strategy of the tasks, which is consideredto be related to the latter memory classes, using a set of 24 pictures (trainingset; see Methods). After the monkey’s performance reached a plateau levelwith the training set, a new stimulus set (test set) was introduced to accessformation of the new associative memory of the new visual stimuli. The mon-key’s performance was at a chance level (i.e., 50% correct responses) in thefirst session with the test set and then improved in subsequent test sessions.
AID NLM 3848 / 6v16$$$$82 09-10-98 09:02:12 nlmoa AP: NLM
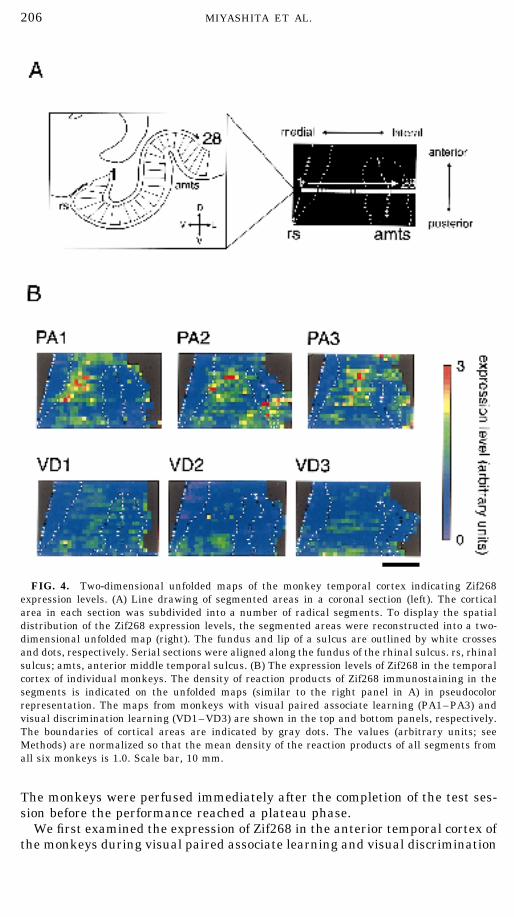
206 MIYASHITA ET AL.
FIG. 4. Two-dimensional unfolded maps of the monkey temporal cortex indicating Zif268expression levels. (A) Line drawing of segmented areas in a coronal section (left). The corticalarea in each section was subdivided into a number of radical segments. To display the spatialdistribution of the Zif268 expression levels, the segmented areas were reconstructed into a two-dimensional unfolded map (right). The fundus and lip of a sulcus are outlined by white crossesand dots, respectively. Serial sections were aligned along the fundus of the rhinal sulcus. rs, rhinalsulcus; amts, anterior middle temporal sulcus. (B) The expression levels of Zif268 in the temporalcortex of individual monkeys. The density of reaction products of Zif268 immunostaining in thesegments is indicated on the unfolded maps (similar to the right panel in A) in pseudocolorrepresentation. The maps from monkeys with visual paired associate learning (PA1–PA3) andvisual discrimination learning (VD1–VD3) are shown in the top and bottom panels, respectively.The boundaries of cortical areas are indicated by gray dots. The values (arbitrary units; seeMethods) are normalized so that the mean density of the reaction products of all segments fromall six monkeys is 1.0. Scale bar, 10 mm.
The monkeys were perfused immediately after the completion of the test ses-sion before the performance reached a plateau phase.
We first examined the expression of Zif268 in the anterior temporal cortex ofthe monkeys during visual paired associate learning and visual discrimination
AID NLM 3848 / 6v16$$$$82 09-10-98 09:02:12 nlmoa AP: NLM

207CONSOLIDATION OF VISUAL MEMORY IN THE TEMPORAL CORTEX
learning. During visual paired associate learning, intensely Zif268-immuno-positive neurons were observed in the inferior temporal gyrus, the area thatlies between the rhinal sulcus and the anterior middle temporal sulcus; theintensely Zif268-immunopositive neurons accumulated in patches in the ven-tral surface of the inferior temporal gyrus rather than in the banks of the rhinalsulcus or the anterior middle temporal sulcus. The patches were centered inlayer IV and spread into both superficial (II/III) and deep (V and VI) layers.The patchy pattern was found in several consecutive sections and was specificto Zif268 since other IEG products (cFos and JuD) did not show such patternsin the adjacent sections.
The distribution of Zif268 expression in the monkey temporal cortex wasvisualized by image analysis and displayed on a two-dimensional unfoldedmap (Fig. 4). In each monkey with visual paired associate learning, Zif268 wasexpressed at high levels in a strip parallel to the rhinal sulcus in an anterior–posterior axis, particularly at several spots in this strip (PA1–PA3). Zif268was expressed at relatively low levels and was distributed more homogeneouslyin the monkeys with visual discrimination learning (VD1–VD3). The inferiortemporal gyrus is composed of three cytoarchitectonically and connectionallydistinct areas (area 35, area 36, and the ventral part of area TE) (Suzuki andAmaral, 1994). The expression of Zif268 in the monkeys during visual pairedassociate learning was prominent in area 36. By contrast, in area 35, whichis medially adjacent to area 36, Zif268 expression levels were low during bothtypes of visual learning. A two-way ANOVA for the mean expression level (twolearning tasks and three brain areas) revealed significant effects of task [F(1,4) Å 12.05, p Å .025] and of area [F(2, 8) Å 17.73, p Å .001]. The expressionin area 36 was significantly higher during visual paired associate learningthan during visual discrimination learning [1.34 { 0.07 and 0.96 { 0.04({SEM, arbitrary units), respectively; p Å .01]. In area 35, the expressionlevels in the two monkey groups were similar and were not significantly differ-ent (pair-association, 0.65 { 0.09; discrimination, 0.58 { 0.05; p ú .5). Inventral TE, the difference did not reach statistical significance (p Å .22), butthere might be a tendency for higher expression of Zif268 in the monkeys withvisual paired associate learning (pair-association, 1.11 { 0.15; discrimination,0.85 { 0.08).
GENERAL DISCUSSION
In this article, we examined the neural mechanisms underlying the memoryconsolidation process by single-unit recording and molecular biological meth-ods during a visual pair-association task in monkeys. In Experiment 1, wetested the role of the backward signal from the medial temporal lobe to theIT cortex. We made lesions in the entorhinal and perirhinal cortices and re-corded unit activity in the IT cortex. In Experiment 2, we investigated themolecular basis of circuit reorganization by detecting the expression of immedi-ate-early genes in monkey temporal cortex. The results of Experiment 1 andExperiment 2 supported the hypothesis that limbic–neocortical interactionsare critical in the consolidation process since limbic neurons undergo rapidmodification of synaptic connectivity and provide backward signals that guidethe reorganization of neocortical neural circuits.
In Experiment 1, the results provided new evidence about the role of the
AID NLM 3848 / 6v16$$$$83 09-10-98 09:02:12 nlmoa AP: NLM

208 MIYASHITA ET AL.
backward signal. The pair-association learning task has been used to testdeclarative memory in humans, and the role of the medial temporal lobe indeclarative memory formation was proposed on the basis of neuropsychologicaland behavioral evidence. The present experiment evaluated single neuron re-sponses for visual pair-association and supported the view that the IT neuronshave the ability to represent the long-term mnemonic code between picturepairs and that the ability is critically dependent on backward signal from theentorhinal and perirhinal cortices to the IT neurons.
In this experiment, we combined single-unit recording techniques with surgi-cal manipulations in chronic monkey preparations; this chronic monkey prepa-ration was designed so that a single IT neuron becomes devoid of limbic back-ward signal but can receive forward visual signal in a normal behavioral con-text (Fig. 2). It was demonstrated by a lesion study that monkeys with bilateralremoval of the entorhinal and perirhinal cortices neither relearn visual pair-association that was learned prelesionally nor learn new associations amongnew visual stimuli (Murray et al., 1993). In the present experiment monkeyswith a unilateral lesion learned both the old and the new associations easily.The milder effects of the unilateral lesion at the behavioral level should bedue to the intact temporal lobe in the contralateral hemisphere (Fig. 2). Atthe level of neuronal response, the lesion wiped out the ability for associationof the IT neurons that had been isolated from the intact temporal lobe in thecontralateral hemisphere by the anterior commissurectomy.
In the primate inferior temporal cortex and part of the superior temporalcortex, neurons selectively respond to complex objects such as faces, hands,Fourier descriptors, and fractal patterns (Miyashita, 1993). Several lines ofexperimental evidence showed that some, if not all, of these responses wereacquired through learning in adulthood (Miyashita, 1993; Higuchi & Miya-shita, 1996). The present results show that the backward connection from thelimbic cortex is indispensable for some kinds of these acquired responses, suchas pair-association coding, but may not be indispensable for others. Indeed,the fractal patterns introduced after the lesion (set B) could activate IT neuronshighly selectively, which suggests preservation of the tuning mechanism thatmolds and sharpens neuronal response selectivity to an object through repeti-tive stimulus presentation (Sakai, Naya, & Miyashita, 1994). These resultsgive concrete evidence for the decomposition of the primate long-term memorysystem into several different neural mechanisms.
In Experiment 2, we detected the altered expression of Zif268 in the monkeyinferior temporal gyrus, especially in area 36, during visual paired associatelearning compared with that during visual discrimination learning. Duringvisual paired associate learning, Zif268-immunopositive neurons accumulatedin patches that spread into the superficial and deep layers in the inferiortemporal gyrus. High-level expression of Zif268 was localized in a strip alongthe rhinal sulcus (Fig. 4B). In contrast to the strong patchy expression ofZif268, the expression of other IEG products (Fos and JunD) did not showsuch patterns. The results indicating that Zif268 expression was most evidentin area 36 during paired associate learning are consistent with the results ofExperiment 1.
In Experiment 2, we used visual discrimination learning as a control for visualpaired associate learning because, under the no task condition, it was difficultto regulate mental activity related to memory formation or even to regulate
AID NLM 3848 / 6v16$$$$83 09-10-98 09:02:12 nlmoa AP: NLM

209CONSOLIDATION OF VISUAL MEMORY IN THE TEMPORAL CORTEX
motor activity in the monkeys. The difference in Zif268 expression level be-tween visual paired associate learning and visual discrimination learning mostlikely originated from differences in neuronal activities related to memoryrequirements for the two tasks, for several reasons. First, the experimentalconditions were almost identical for the two tasks; i.e., the same task apparatuswith the same touch sensor, as well as the same visual stimuli and the samereward, were used, suggesting that the differential expression of Zif268 wasnot related to general motor or sensory activity. Second, the difference wasnot associated with the amount of sensory input; i.e., the number of trials inthe session immediately before the perfusion was not related to the Zif268expression level in any of the three brain areas in the inferior temporal gyrus.Third, Zif268 expression levels in area 35 or in the hippocampus were almostthe same for the two monkey groups, confirming the reproducibility of immuno-staining among different sections and in different monkeys.
Although it is not yet well characterized in the mammalian brain, studiesin both invertebrates and vertebrates have revealed that formation of long-term memory requires new protein and mRNA synthesis whereas formationof short-term memory does not (Bailey & Kandel, 1993). The requirement ofde novo protein and mRNA synthesis for long-term memory suggests thatneuronal activities induced by learning initiate a cascade of gene expression.The first step of the gene cascade is thought to be the expression of IEGs.Transcription factors encoded by IEGs play a role in the establishment of long-term changes in the properties of synapses (Morgan & Curran, 1991; Abrahamet al., 1993). The induced transcription factors regulate expression of late-response genes that probably contribute to synaptic plasticity, especially mor-phological changes. For example, synapsin-I and neurofilament-light genes areactivated by Zif268 (Thiel et al., 1994). Moreover, many IEGs including zif268have cyclic AMP-responsive element (CRE) sequences in their promoter re-gions and their expression can be controlled by CRE-binding proteins (CREBs)(Vaccarino et al., 1993). Recent studies have shown CREB involvement in theformation and consolidation of various classes of long-term memory in severalspecies (Bourtchuladze et al., 1994; Yin et al., 1994). Taken together, theexpression of Zif268 in the temporal cortex observed in this study suggeststhat Zif268 may participate in the gene cascade related to formation of visualassociative memory in primates.
REFERENCES
Abraham, W. C., Mason, S. E., Demmer, J., Williams, J. M., Richardson, C. L., Tate, W. P., Lawlor,P. A., & Dragunow, M. (1993). Correlations between immediate early gene induction and thepersistence of long-term potentiation. Neuroscience, 56, 717–727.
Bailey, C. H., & Kandel, E. R. (1993). Structural changes accompanying memory storage. AnnualReview of Physiology, 55, 397–426.
Bourtchuladze, R., Frenguelli, B., Blendy, J., Cioffi, D., Schultz, G., & Silva, A. J. (1994). Deficientlong-term memory in mice with a targeted mutation of the cAMP-responsive element-bindingprotein. Cell, 79, 59–68.
Demeter, S., Rosene, D. L., & Van Hoesen, G. W. (1990). Fields and origin and pathways of theinterhemisheric commissures in the temporal lobe of the macaques. Journal of ComparativeNeurology, 302, 29–53.
Higuchi, S., & Miyashita, Y. (1996). Formation of mnemonic neuronal responses to visual pairedassociates in inferotemporal cortex is impaired by perirhinal and entorhinal lesions. Proceed-ings of the National Academy of Sciences of the USA, 93, 739–743.
AID NLM 3848 / 6v16$$$$83 09-10-98 09:02:12 nlmoa AP: NLM

210 MIYASHITA ET AL.
Iwai, E., & Mishkin, M. (1969). Further evidence on the locus of the visual area in the temporallobe of the monkey. Experimental Neurology, 25, 585–594.
Mello, C. V., Vicario, D. S., & Clayton, D. F. (1992). Song presentation induces gene expression inthe song bird forebrain. Proceedings of the National Academy of Sciences of the USA, 89,6818–6822.
Mishkin, M. (1982). A memory system in the monkey. Philosophical Transactions of the RoyalSociety of London (Biology), 298, 85–95.
Miyashita, Y. (1988). Neuronal correlate of visual associative long-term memory in the primatetemporal cortex. Nature, 335, 817–820.
Miyashita, Y. (1993). Inferior temporal cortex: Where visual perception meets memory. AnnualReview of Neuroscience, 16, 245–263.
Miyashita, Y. (1995). How the brain creates imagery: Projection to primary visual cortex. Science,268, 1719–1720.
Miyashita, Y., & Chang, H. S. (1988). Neuronal correlate of pictorial short-term memory in theprimate temporal cortex. Nature, 331, 68–70.
Miyashita, Y., Date, A., & Okuno, H. (1993). Configuration encoding of complex visual forms bysingle neurons of monkey temporal cortex. Neuropsychologia, 31, 1119–1131.
Miyashita, Y., Higuchi, S., Sakai, K., & Masui, N. (1991). Generation of fractal patterns for probingthe visual memory. Neuroscience Research, 12, 307–311.
Miyashita, Y., Rolls, T., Cahusac, P. M. B., Niki, H., & Feigenbaum, J. D. (1989). Activity ofhippocampal formation neurons in the monkey related to a stimulus–response associationtask. Journal of Neurophysiology, 61, 669–678.
Morgan, J. I., & Curran, T. (1991). Stimulus–transcription coupling in the nervous system:Involvement of the inducible proto-oncogenes fos and jun. Annual Review of Neuroscience,14, 421–451.
Murray, E. A., Gaffan, D., & Mishkin, M. (1993). Neural substrates of visual stimulus–stimulusassociation in rhesus monkeys. Journal of Neuroscience, 13, 4549–4541.
Naya, Y., Sakai, K., & Miyashita, Y. (1996). Activity of primate inferotemporal neurons relatedto a sought target in pair-association task. Proceedings of the National Academy of Sciencesof the USA, 93, 2664–2669.
Okuno, H., & Miyashita, Y. (1996). Expression of the transcription factor ZIF268 in the temporalcortex of monkeys during visual paired associate learning. European Journal of Neuroscience,8, 2118–2128.
Okuno, H., Saffen, D. W., & Miyashita, Y. (1995). Subdivision-specific expression of Zif268 in thehippocampal formation of the macaque monkey. Neuroscience, 66, 829–845.
Olney, J. W. (1983). Excitotoxins (pp. 82–95). London: Macmillan.
Pandya, D. N., Karol, E. A., & Lele, P. P. (1973). The distribution of the anterior commissure inthe squirrel monkey. Brain Research, 49, 177–180.
Sakai, K., & Miyashita, Y. (1991). Neural organization for the long-term memory of paired associ-ates. Nature, 354, 152–155.
Squire, L. R. (1987). Memory and brain. New York: Oxford Univ. Press.
Squire, L. R., & Zola-Morgan, S. (1991). The medial temporal lobe memory system. Science, 253,1380–1386.
Suzuki, W. A., & Amaral, D. G. (1994). Topographic organization of the reciprocal connectionsbetween the monkey entorhinal cortex and the perirhinal and parahippocampal cortices.Journal of Neuroscience, 14, 1856–1877.
Thiel, G., Schoch, S., & Petersohn, D. (1994). Regulation of synapsin I gene expression by thezinc finger transcription factor zif268/egr-1. Journal of Biological Chemistry, 269, 15294–15301.
Vaccarino, F. M., Hayward, M. D., Le, H. N., Hartigan, D. J., Duman, R. S., & Nestler, E. J. (1993).Induction of immediate early genes by cyclic AMP in primary cultures of neurons from ratcerebral cortex. Molecular Brain Research, 19, 76–82.
Van Essen, D. C., & Maunsell, J. H. R. (1980). Two-dimensional maps of the cerebral cortex.Journal of Comparative Neurology, 191, 225–281.
AID NLM 3848 / 6v16$$$$83 09-10-98 09:02:12 nlmoa AP: NLM

211CONSOLIDATION OF VISUAL MEMORY IN THE TEMPORAL CORTEX
Van Essen, D. C., Anderson, C. H., & Felleman, D. J. (1992). Information processing in the primatevisual system: An integrated systems perspective. Science, 255, 419–423.
Van Hoesen, G. W. (1982). The parahippocampal gyrus: New observations regarding its corticalconnections in the monkey. Trends in the Neurosciences, 5, 345–353.
Webster, M. J., Ungerleider, L. G., & Bachevalier, J. (1991). Connections of inferior temporal areasTE and TEO with medial temporal-lobe structures in infant and adult monkeys. Journal ofNeuroscience, 11, 1095–1116.
Wechsler, D. (1987). Wechsler memory scale—Revised. San Antonio, TX: The Psychological Corp./Harcourt Brace Jovanovich.
Worley, P. F., Bhat, R. V., Baraban, J. M., Erickson, C. A., McNaughton, B. L., & Barnes, C. A.(1993). Thresholds for synaptic activation of transcription factors in hippocampus: Correlationwith long-term enhancement. Journal of Neuroscience, 13, 4776–4786.
Yin, J. C. P., Wallach, J. S., Del Vecchio, M., Wilder, E. L., Zhou, H., Quinn, W. G., & Tully, T.(1994). Induction of a dominant negative CREB transgene specifically blocks long-term mem-ory in Drosophila. Cell, 79, 49–58.
AID NLM 3848 / 6v16$$$$83 09-10-98 09:02:12 nlmoa AP: NLM








![Dense Associative Memory for Pattern Recognitionpapers.nips.cc/...memory-for-pattern-recognition.pdf · Pattern recognition and models of associative memory [1] are closely related](https://img.pdfslide.net/doc/110x75/5fb0165218a43607cd66c0bd/dense-associative-memory-for-pattern-pattern-recognition-and-models-of-associative.jpg)




















