Embed Size (px)
Citation preview

Constitutive and Companion Cell-SpecificOverexpression of AVP1, Encoding a Proton-PumpingPyrophosphatase, Enhances Biomass Accumulation,Phloem Loading, and Long-Distance Transport1[OPEN]
Aswad S. Khadilkar2,3, Umesh P. Yadav2, Carolina Salazar4, Vladimir Shulaev, Julio Paez-Valencia5,Gaston A. Pizzio, Roberto A. Gaxiola, and Brian G. Ayre*
Department of Biological Sciences, University of North Texas, Denton, Texas 76203 (A.S.K., U.P.Y., C.S., V.S.,B.G.A.); and School of Life Sciences, Arizona State University, Tempe, Arizona 85287 (J.P.-V., G.A.P., R.A.G.)
ORCID IDs: 0000-0002-0689-985X (A.S.K.); 0000-0001-9325-6002 (U.P.Y.); 0000-0003-4798-3959 (C.S.); 0000-0002-4336-6470 (G.A.P.);0000-0001-9348-9850 (R.A.G.); 0000-0002-7399-200X (B.G.A.).
Plant productivity is determined in large part by the partitioning of assimilates between the sites of production and the sites ofutilization. Proton-pumping pyrophosphatases (H+-PPases) are shown to participate in many energetic plant processes,including general growth and biomass accumulation, CO2 fixation, nutrient acquisition, and stress responses. H+-PPases havea well-documented role in hydrolyzing pyrophosphate (PPi) and capturing the released energy to pump H+ across the tonoplastand endomembranes to create proton motive force (pmf). Recently, an additional role for H+-PPases in phloem loading andbiomass partitioning was proposed. In companion cells (CCs) of the phloem, H+-PPases localize to the plasma membrane ratherthan endomembranes, and rather than hydrolyzing PPi to create pmf, pmf is utilized to synthesize PPi. Additional PPi in theCCs promotes sucrose oxidation and ATP synthesis, which the plasma membrane P-type ATPase in turn uses to create more pmffor phloem loading of sucrose via sucrose-H+ symporters. To test this model, transgenic Arabidopsis (Arabidopsis thaliana) plantswere generated with constitutive and CC-specific overexpression of AVP1, encoding type 1 ARABIDOPSIS VACUOLARPYROPHOSPHATASE1. Plants with both constitutive and CC-specific overexpression accumulated more biomass in shootand root systems. 14C-labeling experiments showed enhanced photosynthesis, phloem loading, phloem transport, anddelivery to sink organs. The results obtained with constitutive and CC-specific promoters were very similar, such that thegrowth enhancement mediated by AVP1 overexpression can be attributed to its role in phloem CCs. This supports the modelfor H+-PPases functioning as PPi synthases in the phloem by arguing that the increases in biomass observed with AVP1overexpression stem from improved phloem loading and transport.
Plant productivity is largely determined by the ac-quisition and partitioning of organic carbon and othernutrients among the various organs of the plant body.
Suc is the principal form of organic carbon transportedlong distance via the phloem vascular system. In theinitial steps of carbon partitioning, Suc enters the phloemcompanion cell (CC)-sieve element (SE) complex togenerate a hydrostatic pressure gradient that pusheswater and dissolved compounds from source to recipi-ent sink organs. Phloem loading from the apoplasm isthe best described method for accumulating solute in tothe CC-SE complex. Suc generated in mesophyll cellsmoves cell to cell, presumably via plasmodesmata, toenter the vascular bundle (Giaquinta, 1983). Efflux tothe apoplasm occurs from phloem parenchyma cellsthrough the SWEET transporters AtSWEET11 andAtSWEET12 (Chen et al., 2012). From the apoplasmicspace, Suc is then actively accumulated into the CC-SEcomplex by Suc transporter/carrier proteins (SUTs orSUCs) energized by the proton motive force (pmf;Lalonde et al., 2004; Ayre, 2011).
Although phloem loading from the apoplasm ismechanistically well understood, little is known abouthow the process is regulated. SUT genes that participatein phloem loading are transcriptionally regulated by Succoncentrations (Vaughn et al., 2002), leaf maturation
1 This workwas supported by theNational Science Foundation (grantno. IOS 1121819 to B.G.A. and V.S. and grant no. 1122148 to R.A.G.).
2 These authors contributed equally to the article.3 Present address: Department of Biochemistry, University of
Texas Southwestern Medical Center, Dallas, TX 75390–9038.4 Present address: GEVO, Englewood, CO 80112.5 Present address: Department of Botany, University of Wisconsin,
Madison, Wisconsin 53706–1313.* Address correspondence to [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policy de-scribed in the Instructions for Authors (www.plantphysiol.org) is:Brian G. Ayre ([email protected]).
R.A.G. and B.G.A. conceived the research plans and supervised theexperiments; A.S.K. and U.P.Y. conducted the majority of the researchwith additions from J.P.-V., G.A.P., C.S., and V.S.; A.S.K., U.P.Y., V.S.,R.A.G., and B.G.A. analyzed the data; A.S.K., U.P.Y., R.A.G., andB.G.A. wrote the article with contributions from all authors.
[OPEN] Articles can be viewed without a subscription.www.plantphysiol.org/cgi/doi/10.1104/pp.15.01409
Plant Physiology�, January 2016, Vol. 170, pp. 401–414, www.plantphysiol.org � 2016 American Society of Plant Biologists. All Rights Reserved. 401 www.plantphysiol.orgon March 23, 2018 - Published by Downloaded from
Copyright © 2016 American Society of Plant Biologists. All rights reserved.

(Riesmeier et al., 1993; Truernit and Sauer, 1995), diurnalcycles (Kühn et al., 1997), and possibly pressure (Smithand Milburn, 1980; Aloni et al., 1986). Posttranslationalcontrol may be mediated by protein sorting and protein-protein interactions (Krügel et al., 2008; Liesche et al.,2010). Overexpression of SUT genes specifically in CCs toenhance loading and carbon partitioning to sink organshas been proposed as a mechanism to improve plantproductivity (Ainsworth and Bush, 2011; Braun et al.,2014). In a test of this hypothesis, evidence for enhancedloading and partitioning was obtained in Arabidopsis(Arabidopsis thaliana), but plantswere stunted, apparentlybecause improved Suc transport disrupted nutrient ho-meostasis in the sink organs (Dasgupta et al., 2014).
Further regulation of phloem loading and partition-ing may come from Suc availability in the apoplasm orfrom the strength of the pmf required for Suc uptake,which is generated by plasma membrane (PM)-localized,P-type ATPases. The CCs of species that load fromthe apoplasm have high levels of ATPase gene expres-sion (DeWitt and Sussman, 1995) and greater pmf thanthe surrounding phloem parenchyma (Hafke et al.,2005). To generate the ATP required by the ATPases, aportion of the loaded Suc must be oxidized. A mor-phological hallmark of CCs is that they are cytoplas-mically dense with many mitochondria, which may becell-specific adaptations to accommodate this metabo-lism (van Bel and Knoblauch, 2000). In principle, com-plete oxidation of a single molecule of Suc can generateapproximately 60 ATPs and thus provide pmf to loadapproximately 60 molecules of Suc.
During the initial steps of Suc oxidation in CCs, su-crose synthase (SUS) splits Suc into Fru and UDP-Glc,and then UDP-Glc pyrophosphorylase (UGPase) cata-lyzes UDP-Glc + pyrophosphate (PPi)↔Glc-1-P + UTP(Yang and Russell, 1990; Nolte and Koch, 1993). Geneticevidence for this pathway derives in part from strongSUS expression in CCs. Biochemical support derivesfrom CC-specific expression of an Escherichia coli py-rophosphatase, ppaI, which hydrolyzes PPi to preventthe production of Glc-1-P and UTP and results inthe stunted growth characteristic of deficient phloemtransport. Importantly, phloem-specific expression of ayeast invertase (rolCpro:SUC2) restored wild-type pheno-types by allowing Suc to enter glycolysis through a SUS-and PPi-independent pathway (Lerchl et al., 1995).
In addition to P-type ATPases, plants possessmembrane-boundH+-pumping pyrophosphatases (H+-PPases) that have a well-established role in capturingthe energy of PPi hydrolysis by pumping protonsacross the tonoplast or other endomembrane to con-tribute to pmf (Zhen et al., 1997; Baltscheffsky et al.,1999; Maeshima, 2000). PPi is a by-product in the syn-thesis of many biopolymers, including proteins, nucleicacids, starch, and cell wall carbohydrates, and is thus aconvenient source of energy in metabolically activecells. Prior work established that type I H+-PPaseis broadly expressed, with highest levels in rapidlygrowing and vascular tissues (Li et al., 2005; Yang et al.,2007; Pizzio et al., 2015).
While the activity of an E. coli soluble PPase in thecytoplasm results in debilitated plants, overexpressionof H+-PPases results in plants with greater salt anddrought tolerance, improved nutrient uptake, andgreater biomass accumulation (Li et al., 2005, 2008, 2010;Park et al., 2005; Yang et al., 2007; Bao et al., 2008; Lvet al., 2008, 2009; Schilling et al., 2014). In Arabidopsis,ARABIDOPSIS VACUOLAR PYROPHOSPHATASE1(AVP1) overexpression resulted in 40% to 60% greaterleaf area and up to 8.4-fold increase in root growth,relative to the wild type (Li et al., 2005; Gonzalezet al., 2010). Similar increases in biomass were ob-served with AVP1 and orthologs from different speciesoverexpressed in a diversity of plants, including rice(Oryza sativa), cotton (Gossypium hirsutum), maize (Zeamays), alfalfa (Medicago sativa), wheat (Triticum aestivum),barley (Hordeum vulgare), and creeping bentgrass (Agro-stis stolonifera; Brini et al., 2007; Bao et al., 2008; Li et al.,2008, 2010; Lv et al., 2008, 2009; Pasapula et al., 2011;Schilling et al., 2014). Overexpressing H+-PPase genesalso enhances nutrient uptake by affecting the abundanceand activity of the PMH+-ATPase in amanner consistentwith apoplasmic pH alterations and rhizosphere acidifi-cation (Li et al., 2005; Yang et al., 2007). Consequently,plants overexpressing AVP1 outperform controls whengrown under phosphate and nitrate limitations and ac-cumulate more potassium (Yang et al., 2007; Undurragaet al., 2012; Paez-Valencia et al., 2013). In addition, themetabolite profile inArabidopsis overexpressingAVP1 isshifted toward nitrogen metabolism and changes in car-bon signaling (Gonzalez et al., 2010).
It is proposed that H+-PPase overexpression improvesthis diversity of physiological attributes because it local-izes to different membranes and has different functionsdepending on the cells in which it is being expressed(Paez-Valencia et al., 2011; Gaxiola et al., 2012; Pizzioet al., 2015). Although first characterized as a vacuolarH+-PPase (Rea et al., 1992; Maeshima, 2000), studies withgold-conjugated H+-PPase-specific antibodies and pro-teomic approaches show a dual localization at the tono-plast and the PM (Long et al., 1995; Robinson et al., 1996;Langhans et al., 2001; Alexandersson et al., 2004; Paez-Valencia et al., 2011; Regmi et al., 2015). Although duallocalization is well supported, a mechanistic model forthis alternative targeting has not been put forward.
It was suggested that PM-localized H+-PPase couldcontribute to phloem pmf to help energize phloemloading and enhance long-distance transport. Althoughthis is an attractive model, H+-PPases on the CCPM cannot thermodynamically operate hydrolyticallyto pump H+ into the apoplasm (Davies, 1997), basedon estimations of the free energy of the H+-PPasepump action for in vivo conditions. Instead, it is arguedthat the reverse reaction, in which the PM pmf isused to synthesize PPi, is thermodynamically feasible(Davies, 1997). Therefore, it is highly noteworthy thatin vivo data obtained with the H+-PPase from thegram-negative proteobacterium Rhodospirillum rubrumare consistent with the capacity of this enzyme toplay two distinct roles depending on location and
402 Plant Physiol. Vol. 170, 2016
Khadilkar et al.
www.plantphysiol.orgon March 23, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
conditions: it can act as an intracellular H+ pump in theacidocalcisomes (Seufferheld et al., 2004) or as a PPisynthase in the chromatophore membranes during il-lumination (Baltscheffsky et al., 1966, 1999). Further-more, Rocha Facanha and de Meis (1998) presented invitro evidence with tonoplast fractions of maize coleop-tiles and seeds consistent with the reverse function of theH+-PPase and suggested that the H+-PPase could operatein the maintenance of cytosolic PPi levels. Supported bythese studies, it was proposed thatH+-PPases localized tothe PM of CCs could use pmf to maintain the cytosolicPPi levels required for Suc respiration via SUS, UGPase,and PPi-dependent phosphofructokinase (see figure 1 inGaxiola et al., 2012). A recent report showed evidenceconsistent with a PPi synthase function for PM-localizedAVP1 in Arabidopsis CCs, with direct implications inphloem function (Pizzio et al., 2015).To directly test for an AVP1 function in phloem
loading and transport, transgenic Arabidopsis plantswere generated with constitutive and CC-specificoverexpression of AVP1. Plants with both expression
patterns accumulated more biomass in shoot and rootsystems. 14C-labeling experiments showed enhancedphotosynthesis, phloem loading, phloem transport,and delivery to sink organs. Changes in the levels of keymetabolites that report the carbon status of the plantswere sought but not identified, suggesting that steady-state levels are not altered. Since constitutive and CC-specific expression patterns gave very similar results,this work supports the model for a noncanonical H+-PPase function in the phloem by arguing that the in-creases in biomass observed withAVP1 overexpressionstem from improved phloem loading and transport.
RESULTS
Overexpression of AVP1 Enhances Plant Growth
To dissect the roles of AVP1 in enhancing biomassaccumulation and carbohydrate distribution, transgenicArabidopsis plants were generated with constitutiveAVP1 complementary DNA (cDNA) overexpression
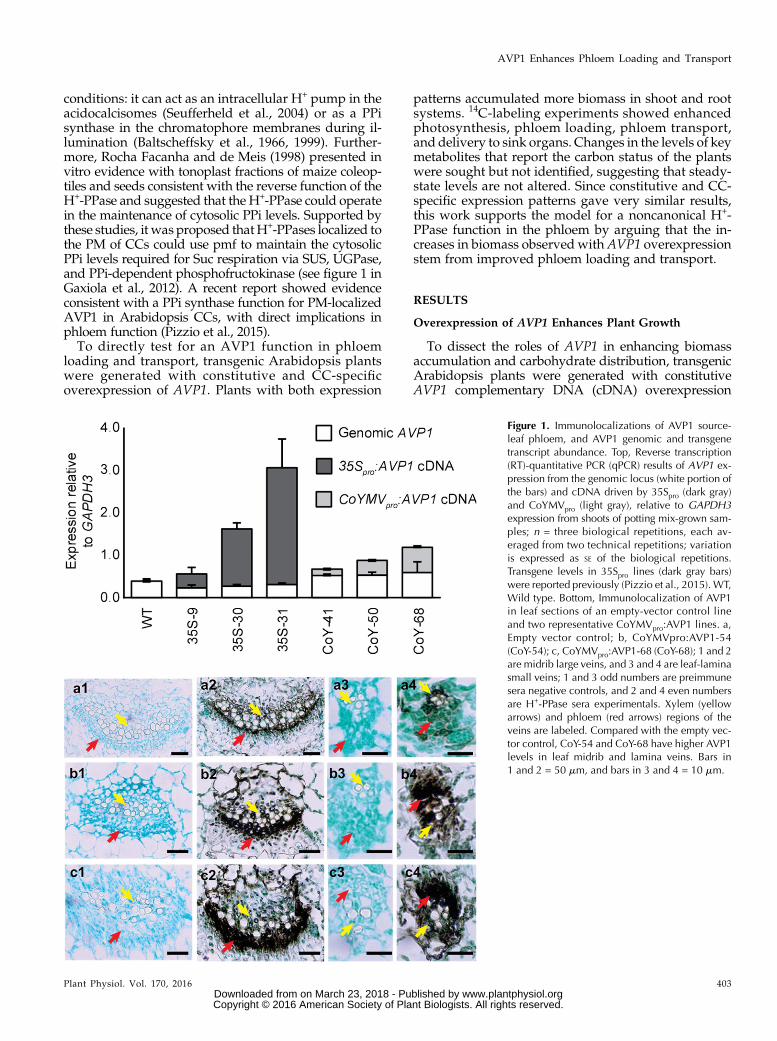
Figure 1. Immunolocalizations of AVP1 source-leaf phloem, and AVP1 genomic and transgenetranscript abundance. Top, Reverse transcription(RT)-quantitative PCR (qPCR) results of AVP1 ex-pression from the genomic locus (white portion ofthe bars) and cDNA driven by 35Spro (dark gray)and CoYMVpro (light gray), relative to GAPDH3expression from shoots of potting mix-grown sam-ples; n = three biological repetitions, each av-eraged from two technical repetitions; variationis expressed as SE of the biological repetitions.Transgene levels in 35Spro lines (dark gray bars)were reported previously (Pizzio et al., 2015). WT,Wild type. Bottom, Immunolocalization of AVP1in leaf sections of an empty-vector control lineand two representative CoYMVpro:AVP1 lines. a,Empty vector control; b, CoYMVpro:AVP1-54(CoY-54); c, CoYMVpro:AVP1-68 (CoY-68); 1 and 2are midrib large veins, and 3 and 4 are leaf-laminasmall veins; 1 and 3 odd numbers are preimmunesera negative controls, and 2 and 4 even numbersare H+-PPase sera experimentals. Xylem (yellowarrows) and phloem (red arrows) regions of theveins are labeled. Compared with the empty vec-tor control, CoY-54 and CoY-68 have higher AVP1levels in leaf midrib and lamina veins. Bars in1 and 2 = 50 mm, and bars in 3 and 4 = 10 mm.
Plant Physiol. Vol. 170, 2016 403
AVP1 Enhances Phloem Loading and Transport
www.plantphysiol.orgon March 23, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
from the cauliflower mosaic virus 35S promoter (35Spro)and phloem CC-specific expression from a promoterelement derived from Commelina yellow mottle virus(CoYMVpro). CoYMVpro has been shown to have strongCC-specific expression (Medberry et al., 1992; Matsudaet al., 2002). For each construct, 25 to 30 independentlytransformed lines were characterized for vegetative ro-sette growth at 16dpost germination. Three lines for eachconstruct that showed increases in rosette area, relative tothewild type, were selected for further study. From thoseconstitutively expressing AVP1 cDNA from 35Spro,these were 35S-9, 35S-30, and 35S-31; and among the lineswith CC-specific overexpression from CoYMVpro, these
were CoY-41, CoY-50, and CoY-68. These lines werebriefly described previously (Pizzio et al., 2015), and theirenhanced growth on potting mix is elaborated inSupplemental Figure S1 (for shoot and root growthin hydroponic conditions, see Fig. 6, A and B, below). Inaddition, AVP1-1, which is a well-described 35Spro:AVP1-overexpressing line (Li et al., 2005; Yang et al.,2007; Pizzio et al., 2015), was included in all analysesas an additional control (Fig. 1). Growth rate anal-ysis showed that the transgenic lines grew faster(Supplemental Fig. S1C) than the wild type.
To establish theAVP1 transcript levels, RT-qPCRwasconducted on total RNA isolated from the wild type
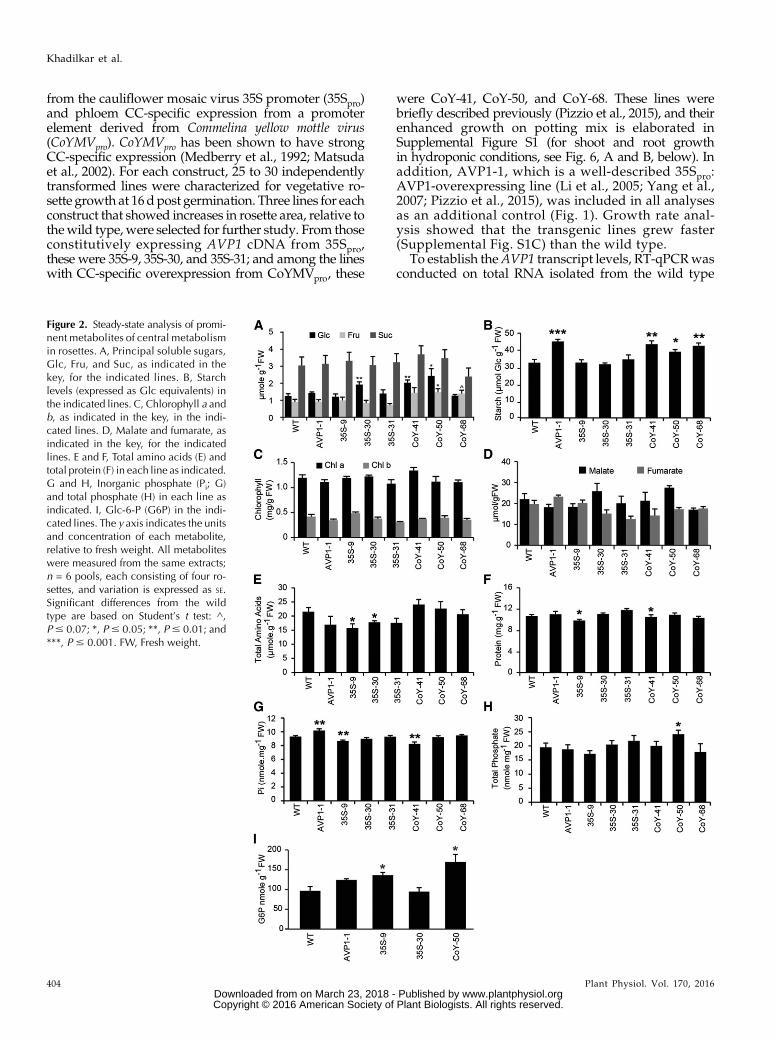
Figure 2. Steady-state analysis of promi-nent metabolites of central metabolismin rosettes. A, Principal soluble sugars,Glc, Fru, and Suc, as indicated in thekey, for the indicated lines. B, Starchlevels (expressed as Glc equivalents) inthe indicated lines. C, Chlorophyll a andb, as indicated in the key, in the indi-cated lines. D, Malate and fumarate, asindicated in the key, for the indicatedlines. E and F, Total amino acids (E) andtotal protein (F) in each line as indicated.G and H, Inorganic phosphate (Pi; G)and total phosphate (H) in each line asindicated. I, Glc-6-P (G6P) in the indi-cated lines. The y axis indicates the unitsand concentration of each metabolite,relative to fresh weight. All metaboliteswere measured from the same extracts;n = 6 pools, each consisting of four ro-settes, and variation is expressed as SE.Significant differences from the wildtype are based on Student’s t test: ^,P# 0.07; *, P# 0.05; **, P# 0.01; and***, P # 0.001. FW, Fresh weight.
404 Plant Physiol. Vol. 170, 2016
Khadilkar et al.
www.plantphysiol.orgon March 23, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
and each transgenic line. Oligonucleotides were designedfor specific quantification of total AVP1 transcriptand transgene-derived transcript and were standard-ized relative to the housekeeping gene GAPDH3(Fig. 1). In the representative lines, expression levelsvaried as expected, but in all cases they exceeded wild-type levels. Relative to AVP1 transcript levels in wild-type plants, endogenous AVP1 transcript declinedapproximately 50% in the 35Spro:AVP1 lines and increasedmoderately in the CoYMVpro:AVP1 lines. Immunohis-tochemistry was conducted to confirm phloem-specific overexpression in two of the CoYMVpro:AVP1lines (Fig. 1). AVP1 protein was detected in the phloemand other cells of the vascular bundle of wild-typecontrols as expected (Paez-Valencia et al., 2011) but wasenhanced substantially in the phloem of transgenic linesin both the midrib and the smaller veins of the leaf.
Overexpression of AVP1 Influences Carbon Partitioning
Wehypothesize thatAVP1 overexpression influencescarbon flux through primary metabolism and parti-tioning through the plant. To assess the effect of AVP1overexpression on Suc synthesis, the major nonstruc-tural carbohydrates (Glc, Fru, Suc, and starch) werequantified in rosettes of the wild type and the repre-sentative transgenic lines at 21 d post germination (Fig. 2).All plantswere harvested into liquid nitrogen 6 to 8 h intothe 12-h illuminated period. Relative to the wild type, therepresentative 35Spro lines did not show consistent alter-ations in nonstructural carbohydrates, but line AVP1-1did accumulate more starch, consistent with priorreports (Pizzio et al., 2015). In contrast, the CoYMVpro:AVP1 lines consistently hadmore hexose andmore starch(Fig. 2, A and B). Soluble carbohydrates were measuredenzymatically and by high-performance anion-exchangechromatography (data not shown), and the results wereconsistent with the two techniques.In addition, a broader collection of prominent carbon-
containing compounds were measured in rosettesto assess the overall carbon status of the plants. Levelsof chlorophyll A and B were measured as metrics foroverall plant health and photosynthetic capacity, andno alterations were observed (Fig. 2C). No significant
and consistent changes were observed in the majororganic acids malate and fumarate or in protein andtotal amino acid levels (Fig. 2, D–F). It was shown previ-ously that AVP1 overexpression can enhance nutrientuptake, including the uptake of phosphate (Yang et al.,2007, 2014; Gaxiola et al., 2012; Pizzio et al., 2015). Thus,inorganic phosphate and total phosphateweremeasured.Some lines had significantlymore or less phosphate, but aconsistent pattern was not evident (Fig. 2, G and H).Phosphorylated intermediates, represented by Glc-6-P,showed moderate increases in two representative 35Spro:AVP1 lines but not the third; levels were also increasedin the CoYMVpro:AVP1 line that was tested (Fig. 2I).
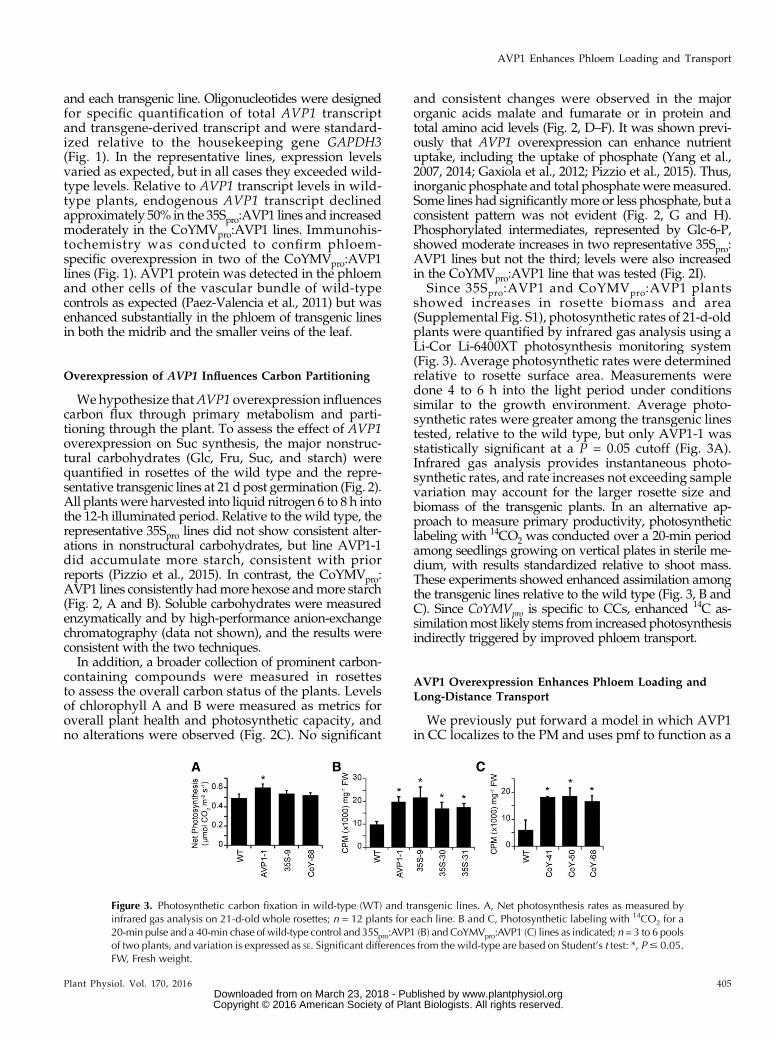
Since 35Spro:AVP1 and CoYMVpro:AVP1 plantsshowed increases in rosette biomass and area(Supplemental Fig. S1), photosynthetic rates of 21-d-oldplants were quantified by infrared gas analysis using aLi-Cor Li-6400XT photosynthesis monitoring system(Fig. 3). Average photosynthetic rates were determinedrelative to rosette surface area. Measurements weredone 4 to 6 h into the light period under conditionssimilar to the growth environment. Average photo-synthetic rates were greater among the transgenic linestested, relative to the wild type, but only AVP1-1 wasstatistically significant at a P = 0.05 cutoff (Fig. 3A).Infrared gas analysis provides instantaneous photo-synthetic rates, and rate increases not exceeding samplevariation may account for the larger rosette size andbiomass of the transgenic plants. In an alternative ap-proach to measure primary productivity, photosyntheticlabeling with 14CO2 was conducted over a 20-min periodamong seedlings growing on vertical plates in sterile me-dium, with results standardized relative to shoot mass.These experiments showed enhanced assimilation amongthe transgenic lines relative to the wild type (Fig. 3, B andC). Since CoYMVpro is specific to CCs, enhanced 14C as-similationmost likely stems from increasedphotosynthesisindirectly triggered by improved phloem transport.
AVP1 Overexpression Enhances Phloem Loading andLong-Distance Transport
We previously put forward a model in which AVP1in CC localizes to the PM and uses pmf to function as a
Figure 3. Photosynthetic carbon fixation in wild-type (WT) and transgenic lines. A, Net photosynthesis rates as measured byinfrared gas analysis on 21-d-old whole rosettes; n = 12 plants for each line. B and C, Photosynthetic labeling with 14CO2 for a20-min pulse and a 40-min chase of wild-type control and 35Spro:AVP1 (B) and CoYMVpro:AVP1 (C) lines as indicated; n = 3 to 6 poolsof two plants, and variation is expressed as SE. Significant differences from the wild-type are based on Student’s t test: *, P# 0.05.FW, Fresh weight.
Plant Physiol. Vol. 170, 2016 405
AVP1 Enhances Phloem Loading and Transport
www.plantphysiol.orgon March 23, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
synthase to increase pyrophosphate levels (Fuglsanget al., 2011; Paez-Valencia et al., 2011; Gaxiola et al.,2012; Pizzio et al., 2015). In this model, increased py-rophosphate levels promote Suc oxidation for ATPsynthesis, which in turn enhances pmf across the PM toenergize phloem loading. Complete oxidation of onemolecule of Suc may yield up to approximately60 ATPs, and a small increase in Suc oxidation candramatically increase CC membrane potential forimproved loading. Earlier work showed that AVP1overexpression enhances pmf, since 35Spro:AVP1 linesexude more H+ to the rhizosphere than the wild type(Pizzio et al., 2015). Earlier work also showed that theinitial velocity and steady state of the pmf H+ gradientsgenerated by the P-type H+-ATPase (i.e. ATP depen-dent and sensitive to 0.1 mM Na3VO4) exhibited valuessignificantly greater (approximately 50%–70%; P =0.05) in 35Spro:AVP1-overexpressing lines comparedwith the wild type (Undurraga et al., 2012).
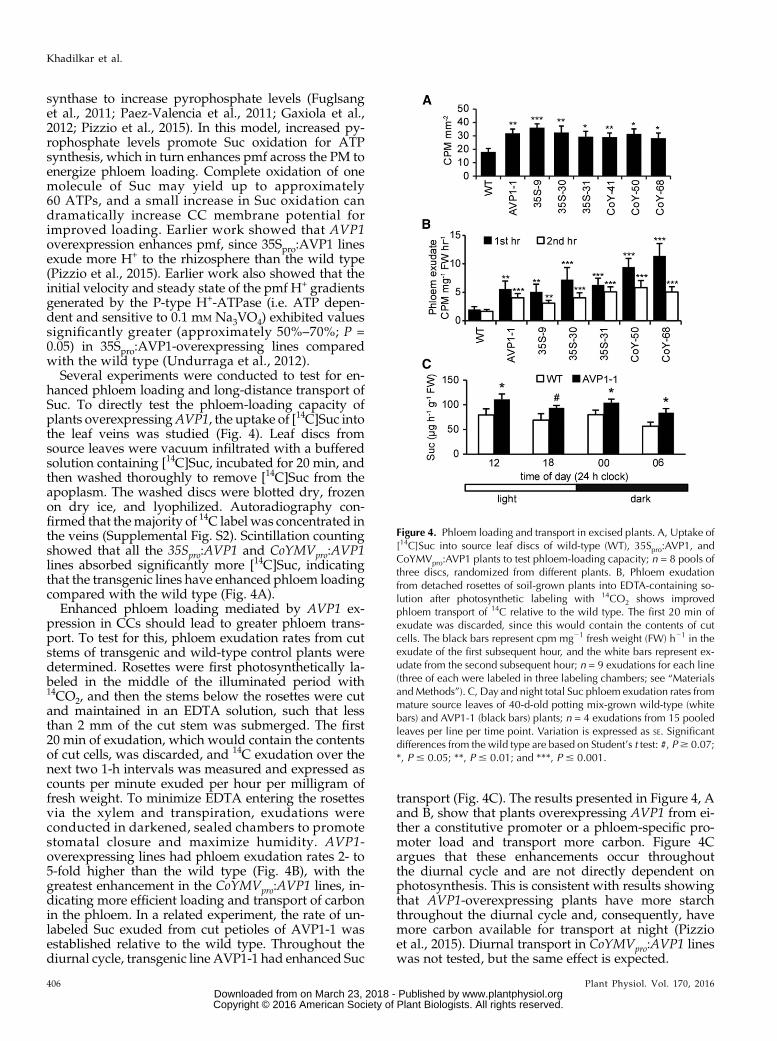
Several experiments were conducted to test for en-hanced phloem loading and long-distance transport ofSuc. To directly test the phloem-loading capacity ofplants overexpressingAVP1, the uptake of [14C]Suc intothe leaf veins was studied (Fig. 4). Leaf discs fromsource leaves were vacuum infiltrated with a bufferedsolution containing [14C]Suc, incubated for 20 min, andthen washed thoroughly to remove [14C]Suc from theapoplasm. The washed discs were blotted dry, frozenon dry ice, and lyophilized. Autoradiography con-firmed that themajority of 14C label was concentrated inthe veins (Supplemental Fig. S2). Scintillation countingshowed that all the 35Spro:AVP1 and CoYMVpro:AVP1lines absorbed significantly more [14C]Suc, indicatingthat the transgenic lines have enhanced phloem loadingcompared with the wild type (Fig. 4A).
Enhanced phloem loading mediated by AVP1 ex-pression in CCs should lead to greater phloem trans-port. To test for this, phloem exudation rates from cutstems of transgenic and wild-type control plants weredetermined. Rosettes were first photosynthetically la-beled in the middle of the illuminated period with14CO2, and then the stems below the rosettes were cutand maintained in an EDTA solution, such that lessthan 2 mm of the cut stem was submerged. The first20 min of exudation, which would contain the contentsof cut cells, was discarded, and 14C exudation over thenext two 1-h intervals was measured and expressed ascounts per minute exuded per hour per milligram offresh weight. To minimize EDTA entering the rosettesvia the xylem and transpiration, exudations wereconducted in darkened, sealed chambers to promotestomatal closure and maximize humidity. AVP1-overexpressing lines had phloem exudation rates 2- to5-fold higher than the wild type (Fig. 4B), with thegreatest enhancement in the CoYMVpro:AVP1 lines, in-dicating more efficient loading and transport of carbonin the phloem. In a related experiment, the rate of un-labeled Suc exuded from cut petioles of AVP1-1 wasestablished relative to the wild type. Throughout thediurnal cycle, transgenic line AVP1-1 had enhanced Suc
transport (Fig. 4C). The results presented in Figure 4, Aand B, show that plants overexpressing AVP1 from ei-ther a constitutive promoter or a phloem-specific pro-moter load and transport more carbon. Figure 4Cargues that these enhancements occur throughoutthe diurnal cycle and are not directly dependent onphotosynthesis. This is consistent with results showingthat AVP1-overexpressing plants have more starchthroughout the diurnal cycle and, consequently, havemore carbon available for transport at night (Pizzioet al., 2015). Diurnal transport in CoYMVpro:AVP1 lineswas not tested, but the same effect is expected.
Figure 4. Phloem loading and transport in excised plants. A, Uptake of[14C]Suc into source leaf discs of wild-type (WT), 35Spro:AVP1, andCoYMVpro:AVP1 plants to test phloem-loading capacity; n = 8 pools ofthree discs, randomized from different plants. B, Phloem exudationfrom detached rosettes of soil-grown plants into EDTA-containing so-lution after photosynthetic labeling with 14CO2 shows improvedphloem transport of 14C relative to the wild type. The first 20 min ofexudate was discarded, since this would contain the contents of cutcells. The black bars represent cpm mg21 fresh weight (FW) h21 in theexudate of the first subsequent hour, and the white bars represent ex-udate from the second subsequent hour; n = 9 exudations for each line(three of each were labeled in three labeling chambers; see “MaterialsandMethods”). C, Day and night total Suc phloem exudation rates frommature source leaves of 40-d-old potting mix-grown wild-type (whitebars) and AVP1-1 (black bars) plants; n = 4 exudations from 15 pooledleaves per line per time point. Variation is expressed as SE. Significantdifferences from the wild type are based on Student’s t test: #, P$ 0.07;*, P # 0.05; **, P # 0.01; and ***, P # 0.001.
406 Plant Physiol. Vol. 170, 2016
Khadilkar et al.
www.plantphysiol.orgon March 23, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
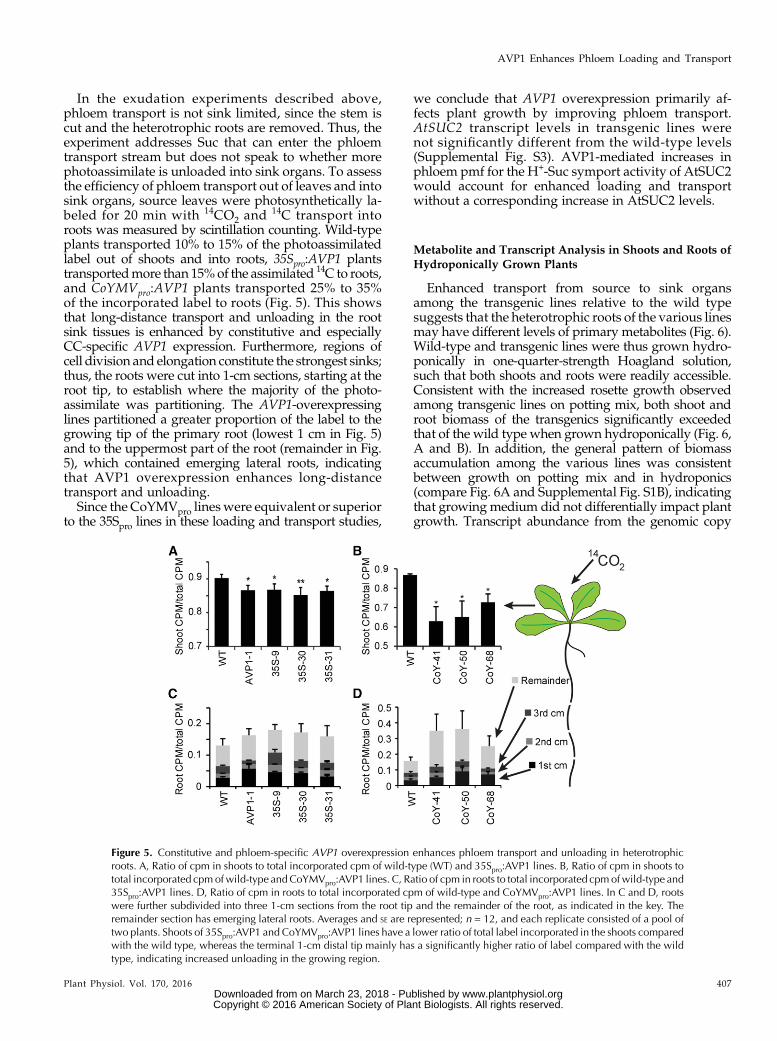
In the exudation experiments described above,phloem transport is not sink limited, since the stem iscut and the heterotrophic roots are removed. Thus, theexperiment addresses Suc that can enter the phloemtransport stream but does not speak to whether morephotoassimilate is unloaded into sink organs. To assessthe efficiency of phloem transport out of leaves and intosink organs, source leaves were photosynthetically la-beled for 20 min with 14CO2 and
14C transport intoroots was measured by scintillation counting. Wild-typeplants transported 10% to 15% of the photoassimilatedlabel out of shoots and into roots, 35Spro:AVP1 plantstransportedmore than 15%of the assimilated 14C to roots,and CoYMVpro:AVP1 plants transported 25% to 35%of the incorporated label to roots (Fig. 5). This showsthat long-distance transport and unloading in the rootsink tissues is enhanced by constitutive and especiallyCC-specific AVP1 expression. Furthermore, regions ofcell division and elongation constitute the strongest sinks;thus, the roots were cut into 1-cm sections, starting at theroot tip, to establish where the majority of the photo-assimilate was partitioning. The AVP1-overexpressinglines partitioned a greater proportion of the label to thegrowing tip of the primary root (lowest 1 cm in Fig. 5)and to the uppermost part of the root (remainder in Fig.5), which contained emerging lateral roots, indicatingthat AVP1 overexpression enhances long-distancetransport and unloading.Since the CoYMVpro lineswere equivalent or superior
to the 35Spro lines in these loading and transport studies,
we conclude that AVP1 overexpression primarily af-fects plant growth by improving phloem transport.AtSUC2 transcript levels in transgenic lines werenot significantly different from the wild-type levels(Supplemental Fig. S3). AVP1-mediated increases inphloem pmf for the H+-Suc symport activity of AtSUC2would account for enhanced loading and transportwithout a corresponding increase in AtSUC2 levels.
Metabolite and Transcript Analysis in Shoots and Roots ofHydroponically Grown Plants
Enhanced transport from source to sink organsamong the transgenic lines relative to the wild typesuggests that the heterotrophic roots of the various linesmay have different levels of primary metabolites (Fig. 6).Wild-type and transgenic lines were thus grown hydro-ponically in one-quarter-strength Hoagland solution,such that both shoots and roots were readily accessible.Consistent with the increased rosette growth observedamong transgenic lines on potting mix, both shoot androot biomass of the transgenics significantly exceededthat of the wild type when grown hydroponically (Fig. 6,A and B). In addition, the general pattern of biomassaccumulation among the various lines was consistentbetween growth on potting mix and in hydroponics(compare Fig. 6A and Supplemental Fig. S1B), indicatingthat growing medium did not differentially impact plantgrowth. Transcript abundance from the genomic copy
Figure 5. Constitutive and phloem-specific AVP1 overexpression enhances phloem transport and unloading in heterotrophicroots. A, Ratio of cpm in shoots to total incorporated cpm of wild-type (WT) and 35Spro:AVP1 lines. B, Ratio of cpm in shoots tototal incorporated cpmofwild-type andCoYMVpro:AVP1 lines. C, Ratio of cpm in roots to total incorporated cpmofwild-type and35Spro:AVP1 lines. D, Ratio of cpm in roots to total incorporated cpm of wild-type and CoYMVpro:AVP1 lines. In C and D, rootswere further subdivided into three 1-cm sections from the root tip and the remainder of the root, as indicated in the key. Theremainder section has emerging lateral roots. Averages and SE are represented; n = 12, and each replicate consisted of a pool oftwo plants. Shoots of 35Spro:AVP1 and CoYMVpro:AVP1 lines have a lower ratio of total label incorporated in the shoots comparedwith the wild type, whereas the terminal 1-cm distal tip mainly has a significantly higher ratio of label compared with the wildtype, indicating increased unloading in the growing region.
Plant Physiol. Vol. 170, 2016 407
AVP1 Enhances Phloem Loading and Transport
www.plantphysiol.orgon March 23, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
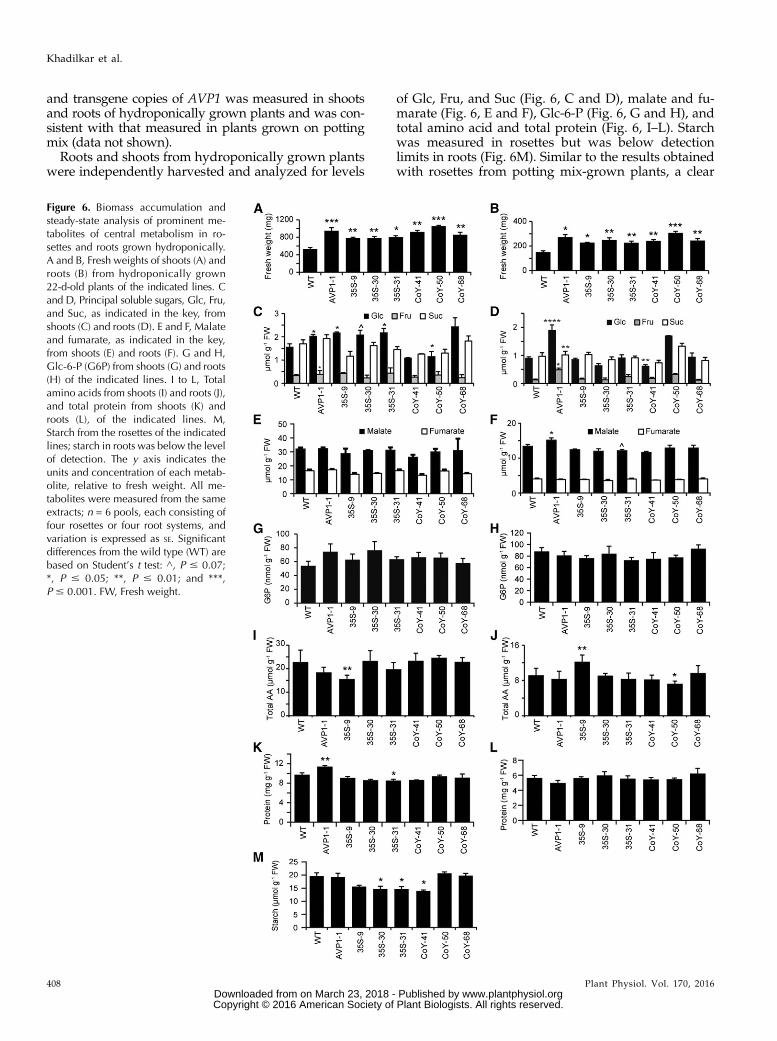
and transgene copies of AVP1 was measured in shootsand roots of hydroponically grown plants and was con-sistent with that measured in plants grown on pottingmix (data not shown).
Roots and shoots from hydroponically grown plantswere independently harvested and analyzed for levels
of Glc, Fru, and Suc (Fig. 6, C and D), malate and fu-marate (Fig. 6, E and F), Glc-6-P (Fig. 6, G and H), andtotal amino acid and total protein (Fig. 6, I–L). Starchwas measured in rosettes but was below detectionlimits in roots (Fig. 6M). Similar to the results obtainedwith rosettes from potting mix-grown plants, a clear
Figure 6. Biomass accumulation andsteady-state analysis of prominent me-tabolites of central metabolism in ro-settes and roots grown hydroponically.A and B, Fresh weights of shoots (A) androots (B) from hydroponically grown22-d-old plants of the indicated lines. Cand D, Principal soluble sugars, Glc, Fru,and Suc, as indicated in the key, fromshoots (C) and roots (D). E and F, Malateand fumarate, as indicated in the key,from shoots (E) and roots (F). G and H,Glc-6-P (G6P) from shoots (G) and roots(H) of the indicated lines. I to L, Totalamino acids from shoots (I) and roots (J),and total protein from shoots (K) androots (L), of the indicated lines. M,Starch from the rosettes of the indicatedlines; starch in roots was below the levelof detection. The y axis indicates theunits and concentration of each metab-olite, relative to fresh weight. All me-tabolites were measured from the sameextracts; n = 6 pools, each consisting offour rosettes or four root systems, andvariation is expressed as SE. Significantdifferences from the wild type (WT) arebased on Student’s t test: ^, P # 0.07;*, P # 0.05; **, P # 0.01; and ***,P # 0.001. FW, Fresh weight.
408 Plant Physiol. Vol. 170, 2016
Khadilkar et al.
www.plantphysiol.orgon March 23, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

pattern of statistically significant differences in metab-olite levels did not emerge among the wild type andthe lines overexpressing AVP1 either constitutively orspecifically in the phloem. Considering the evidence formore reduced carbon reaching the roots, this metaboliteanalysis argues that steady-state metabolite levels aremaintained among all the lines but that the AVP1-overexpressing lines may have more carbon fluxthrough central metabolism.RT-qPCR was used to measure the transcript abun-
dance of genes that may be influenced by constitutiveor CC-specific AVP1 overexpression in shoots androots of representative hydroponically grown plants(Supplemental Figs. S4 and S5). These included genesencoding SUSs (SUS1, AT5G20830; SUS2, AT5G49190;SUS3, AT4G02280; SUS4, AT3G43190; SUS5, AT5G37180;and SUS6, AT1G73370), UDP-Glc pyrophosphatases(UGP1, AT3G03250; and UGP2, AT5G17310), PMH+-ATPases (AHA1, AT2G18960; AHA2, AT4G30190;AHA3, AT5G57350; and AHA4, AT3G47950), cytosolicalkaline invertases (CIN1, At1g35580; and CIN2,AT4G09510); phosphofructokinase a-subunits (PFKa1,AT1G20950; andPFKa2,AT1G20950), phosphofructokinaseb-subunits (PFKb1,AT1G12000; andAtPFKb2,AT4G04040),and SWEETs involved in phloem loading (SWEET11,AT3G48740; and SWEET12,AT5G23660). Strong changesin gene expression were not observed. AtSUS1 wasmoderately repressed, while AtSUS2 was moderatelyenhanced, in the shoot of the representative CoYMVpro:AVP1 line. AtUGP2 transcript was moderately reducedin the roots of 35Spro:AVP1 lines.
DISCUSSION
Enhancing phloem loading and long-distance par-titioning is a potential means to improve plant pro-ductivity through two principal mechanisms: (1)decreasing feedback inhibition on photosynthesis insource leaves, and (2) mobilizingmore resources to sinkorgans for growth, storage, general metabolism, andnutrient uptake (Ainsworth and Bush, 2011; Braunet al., 2014; Yadav et al., 2015). Overexpressing the H+-PPase AVP1 and orthologs in numerous plant speciesincreased biomass transfer to sink organs and overallwhole-plant biomass accumulation (Li et al., 2005; Lvet al., 2008; Gonzalez et al., 2010). It also increased salttolerance and drought resistance (Gaxiola et al., 2001;Gao et al., 2006; Brini et al., 2007; Bao et al., 2008; Liet al., 2008, 2010; Lv et al., 2009; Pasapula et al., 2011;Schilling et al., 2014) and improved growth in nutrient-limited conditions (Yang et al., 2007; Pei et al., 2012;Paez-Valencia et al., 2013; Li et al., 2014). Consistentwith improved nutrient use and uptake, plants over-expressing AVP1 acidified the rhizosphere and showedgreater expression of transporters involved in nitrogen,potassium, and phosphate uptake than wild-type con-trols (Yang et al., 2007; Undurraga et al., 2012; Pizzioet al., 2015). Enhanced growth and energization of therhizosphere are outcomes expected for plants with
increased phloem loading providing more Suc to sinkorgans.
A model was developed previously that proposesthat H+-PPases contribute to phloem loading by func-tioning as PPi synthase at the CC PM to promote theoxidation of a portion of the Suc loaded into the CC(Fig. 7; Gaxiola et al., 2012). Additional PPi functioningas substrate instead of product favors Suc oxidation atsteps mediated by UDP-Glc pyrophosphorylase andPPi-dependent phosphofructokinase (Fig. 7B). Thisoxidation results in additional ATP, which is hydro-lyzed by P-type PM H+-ATPases to create more pmf tofurther energize the phloem system. AVP1 is stronglyexpressed in source-leaf phloem (Pizzio et al., 2015),and PM localization of H+-PPases in the SE-CC complexis well documented (Long et al., 1995; Langhans et al.,2001; Paez-Valencia et al., 2011); the ability of H+-PPasesto function as PPi synthases energized by pmf is alsowell documented (Rocha Facanha and de Meis, 1998;Seufferheld et al., 2004).
In this study, we used constitutive and CC-specificAVP1 overexpression to directly test for improvedphloem loading and long-distance partitioning. Bothconstitutive overexpression with 35Spro:AVP1 andCC-specific overexpression with CoYMVpro:AVP1 resultedin plants with larger rosettes and root systems (Figs.1 and 6). Both constructs also resulted in improvedphotosynthesis (Fig. 3), consistent with expectations forenhanced phloem transport. A series of complementaryexperiments with 14C labeling directly demonstratedimproved phloem loading and long-distance transportto sink organs (Figs. 4 and 5). Loading of [14C]Suc intothe veins of mature source leaves was greater than thatin wild-type controls. After photosynthetic labelingwith 14CO2, up to five times more 14C label was trans-ported through the phloem and out of cut stems. Inintact plants, significantly more 14C label was trans-ported out of shoots and into roots systems, withCoYMVpro:AVP1 plants consistently showing thegreatest transport levels. Measuring biochemical andtransport processes directly in the phloem CCs ofsource and sink tissues in living plants is notoriouslydifficult (Turgeon and Wolf, 2009). However, by usingCoYMVpro to overexpress AVP1, we have geneticallyisolated the biochemical and physiological impacts toCCs. Since CoYMVpro:AVP1 plants perform equiva-lently, and by some metrics better than 35Spro:AVP1plants (Fig. 5), we conclude that most, if not all, of thebeneficial impact of AVP1 overexpression arises fromits activity in phloem.
In an effort to identify how AVP1 overexpressioninfluences primary metabolism, steady-state levels ofnumerous prominent compounds of central metabo-lism were measured. With constitutive AVP1 over-expression, line AVP1-1, which was used here as apreviously characterized control, showed increases inmidday rosette starch of plants grown on potting mix,consistent with earlier results (Pizzio et al., 2015), butthe other 35Spro:AVP1 lines did not (Fig. 2). The lineswith AVP1 overexpression specifically in CCs had
Plant Physiol. Vol. 170, 2016 409
AVP1 Enhances Phloem Loading and Transport
www.plantphysiol.orgon March 23, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

elevated Glc and starch in plants grown on potting mix.These results suggest that, in addition to loading andtransporting more carbon (Figs. 4 and 5), the higherphotosynthesis in these lines (Fig. 3) results in greaterstorage reserves, with independent transgenic linesfluctuating moderately in this capacity.
It is not reasonable to compare metabolite levels be-tween rosettes from plants grown in potting mix andthose grown hydroponically because of the differentconditions, but hydroponic growth allows ready ac-cess to the heterotrophic roots and permits compari-sons between source and sink organs. Rosettes of the35Spro lines had more hexose, particularly Glc (Fig. 6C).Gonzalez et al. (2010) also noted higher levels of hexoseat midday in AVP1-1 plants grown in vitro with one-half-strength Murashige and Skoog medium. The otherrosette metabolites we measured did not differ consis-tently and significantly from the wild type when grownhydroponically (Fig. 6), and this too is consistent withprior results (Gonzalez et al., 2010). In roots, AVP1-1showed increases from the wild type in Glc, Fru,Suc, and, to a lesser extent, malate, indicating a generaltrend to having more reduced carbon in roots. Line35S-9 showed slightly lower amino acid levels in rosettesand slightly higher levels in roots, suggesting improvedamino acid transport, but this was not consistentlyobserved in the other transgenic lines.
Although prominent changes in steady-state levels ofmeasured metabolites in most lines were not observed,the higher levels of carbon fixation, transport, andgrowth make it clear that carbon flux through the cen-tral metabolism is increased in both source and sinkorgans. Flux analysis with isotopic 13C or 14C labelingwill be necessary to better determine the impact ofAVP1 overexpression on Suc oxidation and/or syn-thesis (i.e. PPi as a substrate during Suc oxidation andas a product during Suc synthesis; Fig. 7; Heise et al.,2014; Ma et al., 2014).
There is substantial interest in metabolic engineeringof source/sink relationships to understand and in-crease plant productivity (Yadav et al., 2015). Over-expressing Suc-H+ symporters specifically in CCsresulted in more loading and transport but, unexpect-edly, resulted in stunted growth rather than more ro-bust growth. This effect was linked to a potentialdisruption in homeostasis between transported carbonand nutrient availability in sink organs, particularlyphosphate (Dasgupta et al., 2014). In the work pre-sented here, AVP1 overexpression constitutively andspecifically in CCs similarly increased phloem loadingand carbon transport (Fig. 4), but unlike SUT over-expression, the desired effect of enhanced biomass wasachieved. The reason why one method to enhancephloem transport resulted in stunted growth and theother resulted in enhanced growth may relate to thenutrient balances required in the growing sink organs.When phloem loading and transport were increased bySUT overexpression, Suc was presumably the onlymetabolite for which transport was increased. Theproposal that this caused a nutrient imbalance in sinksis supported by SUT overexpression lines showing in-creased expression of phosphate transporters, purpleacid phosphatases, and increased acidification of therhizosphere, all of which indicate that the plants areperceiving a phosphate limitation. In addition, thestunted phenotype of the SUT overexpression lines wasalleviated by supplementing the growth medium withadditional phosphate (Dasgupta et al., 2014).
With AVP1 overexpression in the phloem, increasedSuc loading and transport were achieved indirectly bypromoting Suc oxidation in CCs, leading ultimately toincreased ATP production and stronger pmf via PMP-type H+-ATPases. AVP1-mediated stimulation of PMATPase activity is documented (Yang et al., 2007;Undurraga et al., 2012). Interestingly, although Suctransport was enhanced, increased expression of
Figure 7. A model for different membrane locali-zations and functions for AVP1 in mesophyll cellsand phloem companion cells. A, AVP1 H+-PPaselocalized to the tonoplast of mesophyll cells pro-motes the hydrolysis of PPi for enhanced Sucsynthesis and to energize the vacuole by pumpingprotons into the lumen. B, Suc/H+ symporters onthe PM of CCs load Suc from the apoplasm; most isdestined for long-distance transport through sieveelements, but a portion is oxidized to energize thephloem network (top of schematic). AVP1 H+-PPase localized to the PM of CCs utilizes the pmfto synthesize PPi.More PPi substrate promotes Sucoxidation to create more ATP (middle of sche-matic). More ATP is hydrolyzed by P-type ATPaseson CC PMs to create more pmf (bottom of sche-matic). Thus, increased Suc oxidization promotedby AVP1-mediated PPi synthesis results in anoverall more energized phloem network.
410 Plant Physiol. Vol. 170, 2016
Khadilkar et al.
www.plantphysiol.orgon March 23, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

AtSUC2, the Suc transporter responsible for phloemloading in Arabidopsis, was not observed in an ear-lier study (Gonzalez et al., 2010) or in our own anal-ysis. Similarly, the expression of AtSWEET11 andAtSWEET12, which are involved in Suc release to theapoplasm, did not increase, nor did transcript levelsof several steps in Suc metabolism (SupplementalFigs. S3–S5). This implies that enhanced Suc trans-port resulted from more flux through the pathway andstronger pmf rather than more metabolic enzymes andmembrane-bound transporters. In addition to increas-ing Suc loading, this stronger pmf around the phloemCCs likely also increased the phloem-mediated transportof other nutrients to maintain balance in the phloem sap.Supporting this, and in contrast to the situation withAtSUC2, transcriptome analysis of AVP1-1 rosettesshowed more than 2-fold up-regulation of several aminoacid transporter genes (Gonzalez et al., 2010), and ouranalysis showed up-regulation of transporters for nitrate,potassium, and phosphate (Paez-Valencia et al., 2013;Pizzio et al., 2015). Enhanced transport of a balancednutrient supply to sink organsmay account for the largerand more energized root systems with higher waterand nutrient uptake efficiencies observed among AVP-overexpressing plants (Gaxiola et al., 2012). In turn, theseenhanced root systems likely account for the overallmorerobust phenotypes and enhanced productivity of plantsoverexpressing AVP1.
CONCLUSION
Amodel put forward previously (Gaxiola et al., 2012)proposes that H+-PPase AVP1, and its orthologs, hasopposite functions depending on cell type and intra-cellular localization (Fig. 7). In nonphloem cells, themodel proposes localization on endomembranes andcanonical function as proton-pumping pyrophospha-tase to energize the endomembrane compartments(principally, but not exclusively, the tonoplast; Fig. 7A).In phloem CCs, however, localization to the PM is welldocumented. Here, the H+-PPase runs backward as asynthase to synthesize PPi at the expense of the pmf.Increased levels of PPi in CCs promote ATP synthesisby Suc oxidation, which in turn is used by the PMP-type ATPase to create more pmf around the phloem,resulting ultimately in more phloem loading andtransport of Suc and presumably other nutrients loadedinto the phloem by symport with protons (Fig. 7B).Here, we show directly by physiological and 14C-labeling studies that overexpression of AVP1 resultsin enhanced phloem loading and transport of Suc tosink organs. We show that a CC-specific promoter,CoYMVpro, for overexpression has the same impact asthe constitutive promoter, 35Spro. This indicates thatmost, if not all, growth enhancements and increases intolerance to water and nutrient deficiencies observed inboth model and crop plant systems may be attributableto a previously unrecognized function for H+-PPases incontrolling phloem loading and transport. By comparing
these results with other efforts to manipulate phloemtransport, our findings indicate that a more holistic ap-proach of increasing the pmf around the phloem may bemore effective than efforts to engineer transport systemsfor individual metabolites. The simultaneous over-expression of AVP1 and genes encoding transporters forspecific nutrients may be additive.
MATERIALS AND METHODS
Plasmid Construction and Plant Transformation
AVP1 cDNA was amplified by PCR from pRT103-AVP1 (Gaxiola et al.,2001) using forward oligonucleotide AVP1-F (59-CACCATGGTGGCG-CCTGCTTTGTTAC-39) and reverse oligonucleotide AVP1-R (59-GAAG-TACTTGAAAAGGATACCACC-39). Phusion Hot start polymerase (NewEngland Biolabs) was used according to the manufacturer’s instructions. ThePCR product was cloned with a pENTR/D-TOPO Cloning Kit (Invitrogen),resulting in pENTR:AVP1. pENTR:AVP1 was recombined with pMDC32(Curtis and Grossniklaus, 2003) using the Gateway LR Clonase II enzyme mix(Invitrogen) to generate pMDC32:AVP1. pMDC32 contains a dual 35Senhancer. pENTR:AVP1 was recombined with pGPTV:CoYMVpro:cmr:ccdB(Dasgupta et al., 2014) to get pGPTV:CoYMVpro:AVP1. These vectors wereelectroporated into Agrobacterium tumefaciens strain GV3101mp90.
Wild-type Columbia-0 (Arabidopsis Biological Resource Center stock no.CS-70000) was transformed with the above constructs by the floral dip method(Clough and Bent, 1998). T1 generation seedlings with pMDC32:AVP1 wereselected on sterile one-half-strength Murashige and Skoog medium (Phyto-technology Laboratories) containing 40 mg L21 hygromycin. Seeds were ger-minated in the dark and grown for 5 d; resistant seedlings were selected basedon their longer hypocotyls (Harrison et al., 2006) and transferred to Fafard 3Bpotting mix (Sun Gro Horticulture). For selecting plants transformed withpGPTV:CoYMVpro:AVP1, T1 seeds were germinated on Fafard 3B potting mixand sprayed with 20 mg L21 glufosinate ammonia (Finale; Farnam) post ger-mination. Twenty-five or more independent lines were identified for bothCoYMVpro:AVP1 and 35Spro:AVP1. Lack of segregation of the resistance markerin the T3 generation was used to identify homozygous lines. From these, threerepresentative lines for each construct were selected for further experimentsbased on cDNA expression and growth morphology.
Plant Material and Growth Conditions
For all plant growth, Arabidopsis (Arabidopsis thaliana) seeds were stratifiedfor 72 h at 4°C, and plants were grown in a controlled environment chamber(Percival AR95L; Percival Scientific) with 12 h of 180mmol photonsm22 s21 lightat 22°C and 12 h of dark at 18°C. All experiments were conducted with T3 or T4generation seeds. For the determination of fresh weight, metabolite analysis,and analysis of transcript abundance, seeds were sown on 9-cm square pots,and seedlings were thinned to four well-spaced plants per pot. At 28 d postgermination, plants were harvested randomly to create six pools with fourplants in each pool. Each pool was frozen in liquid nitrogen and cryogenicground in 15-mL polycarbonate vials (catalog no. PCRV 15-100-23; OPS Diag-nostics) with 8-mm (5/16th inch) stainless steel balls (approximately 2 g each) ina GenoGrinder 2010 (SPEX Sample Prep). The vials, steel balls, and aluminumcryo block for supporting the vials in the GenoGrinder were all prechilled inliquid nitrogen; samples were maintained in liquid nitrogen during processingand otherwise stored at 280°C. For growth rate analysis (Fig. 1C) and photo-synthesis measurements, seeds were sown in 65-mm round pots, and seedlingswere thinned to one plant per pot. Plants were photographed 16, 20, 22, and24 d after germination. Rosette area was measured with ImageJ version 1.42f(Rasband, 2014). Growth on vertically oriented square culture dishes for pho-tosynthetic labeling with 14CO2 was described previously (Dasgupta et al.,2014). Growth on potting mixture for photosynthetic labeling with 14CO2 wasdescribed previously (Dasgupta et al., 2014). Plants were grown hydroponicallyas described (Guo et al., 2008) except that one-quarter-strength Hoagland so-lution (Phytotechnology Laboratories) was used and replaced every 7 d. Me-dium trays and black expanded polyvinylcarbonate sheets to cover the traysand support the shoots were obtained from US Plastics (catalog nos. 49275 and42500, respectively). Holes were drilled in the polyvinylcarbonate sheets for12 plants to be grown in each tray. For each line, six pools of four plants,
Plant Physiol. Vol. 170, 2016 411
AVP1 Enhances Phloem Loading and Transport
www.plantphysiol.orgon March 23, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

randomized from different trays, were harvested 22 d after germination: shoots,cut at the hypocotyl, were weighed and frozen directly, and the roots werequickly blotted dry by gently pressing between paper towels prior to weighingand freezing. The frozen samples were cryo-ground and stored as describedabove. All tissues were harvested 5 to 6 h into the light period.
AVP1 Immunolocalization
Arabidopsis source leaves from plants grown in potting mix were fixed in3.7% (v/v) formaldehyde, 50% (v/v) ethanol, and5% (v/v) acetic acid overnightat 4°C. Fixed material was dehydrated in a graded ethanol series (50%, 60%,70%, 80%, 90%, and 100%, all v/v), and absolute ethanol was replaced byhistological clearing agent (xylene; Sigma-Aldrich). Tissues were embedded inParaplast at 60°C. The embedded tissue was sliced into 10- to 15-mm sectionsand placed onto poly-Lys-coated slides. Sections were deparaffinized in xylene,hydrated in graded alcohol (100%, 90%, 80%, 70%, 60%, 50%, and 30%, withdeionized water), and pretreated with Antigen Retrieval Solution (BioGenex)under a steamer at 65°C for 20 min. Endogenous peroxidase activity wasquenched with 3% hydrogen peroxide, and the slides were treated with PowerUniversal Reagent (BioGenex) to reduce nonspecific binding and incubatedwith polyclonal anti H+-PPase antibody (Park et al., 2005). Controls were donewith preimmune serum at a 1:1,000 dilution in phosphate-buffered saline for allsamples. Then, three rinses with phosphate-buffered saline-Tween (0.1%, v/v)were followed by incubation with peroxidase-labeled polymer conjugatedto goat anti-rabbit immunoglobulins as provided by the EnVision system(DAKO), following the manufacturer’s instructions.
Transcript Analysis
TotalRNAwas isolated from50mgof cryo-ground rosette tissues (see above)using Trizol (Life Technologies). Total RNA was further treated with TurboRNase-free DNase-I (Life Technologies) or with the RTS DNase Kit (Mo BioLaboratories). One milligram of total RNA was reverse transcribed with 50 mM
oligo(dT) (Life Technologies) and either SuperScript III reverse transcriptase(Life Technologies) or GoScript Reverse Transcriptase (Promega) according tothe manufacturer’s instructions. The gene-specific primers used for qPCR aredescribed in Supplemental Table S1. Real-time qPCR was carried out on anApplied Biosystems ViiA 7 device (Life Technologies) with GO Taq qPCR Mix(Promega) using the following protocol: 2 min of denaturation at 95°C followedby 40 cycles of 95°C for 10 s, 65°C for 30 s, and 70°C for 30 s; or with JumpStartTaq Ready Mix (Sigma-Aldrich) using the following thermal profile: 2 min ofdenaturation at 94°C followed by 40 cycles of 94°C for 20 s, 60°C for 30 s, and65°C for 60 s. A final melting curve confirmed a single PCR product. Threebiological and two technical replicates were used for analysis, and transcriptlevels were normalized against GAPDH3 transcript abundance.
Extraction and Measurement of Metabolites
Aliquots of 20mg of cryo-ground tissue (see above) were extractedwith 80%ethanol or TCA (Gibon et al., 2002). The major transient soluble carbohydratesand starchwere extractedwith ethanol and quantified enzymatically (Stitt et al.,1989) using a Synergy H Reader (Bio Tek) or by high-performance anion-exchange chromatography with pulsed amperometric detection (Dasguptaet al., 2014). Starch was measured enzymatically from the insoluble materialafter ethanolic extraction of soluble carbohydrates (Hendriks et al., 2003) or by acommercially available starch assay kit (Megazyme). Total protein was mea-sured using the Bradford assay (Bradford, 1976). Glc-6-P was determinedaccording to Gibon et al. (2004). Malate and fumarate were determinedaccording to Cross et al. (2006). Inorganic phosphate and total phosphate weremeasured using an ammonium molybdate assay (Chiou et al., 2006). Totalamino acids were measured using a fluorescamine method (Gibon et al., 2009).
Photosynthesis Measurement
Photosynthesisperunitof rosette surfaceareawasmeasuredwithaLi-CorLi-6400XT infrared gas analyzer equipped with a whole-plant Arabidopsischamber and red, green, and blue light source according to the equipmentmanufacturer’s instructions. Plants were grown for 21 d in 65-mm-diameterround pots that fit in the whole-plant Arabidopsis chamber. The surface ofpottingmixwas sealedwith pottery clay to avoid gas exchangewith the pottingmedium. Gas-exchangemeasurements were performedwith 400 mmol s21 CO2,a gas flow rate of 500 mmol s21, and photosynthetically active radiation at
225 mmol m22 s21 to reasonably mimic environmental conditions used duringgrowth. Chamber block temperature and relative humidity were established byambient air conditions and were 23°C and approximately 60% respectively.Plants were allowed to acclimate for 120 s in the chamber before taking mea-surements. The acclimatization time was determined empirically. Two mea-surements were taken per plant in 30-s intervals. Plants were digitallyphotographed, and rosette area was determined with ImageJ software(Rasband, 2014) to standardize photosynthetic rates per unit area. Saturatinglight levels were not used for infrared gas analysis, since this would not haverepresented the physiological state during growth.
14C Labeling to Measure Phloem Loading and Transport
Phloem loading of [14C]Suc into source leaf discswas conducted as described(Dasgupta et al., 2014), except that 5-mm leaf discs were excised from sourceleaves rather than using whole leaves. Six replicates, each consisting of six leafdiscs randomized from six plants, were used for each control and experimentalline. Photosynthetic labeling of plants grown on potting mix for phloem exu-dation and the EDTA exudation method was described previously (Dasguptaet al., 2014). Photosynthetic labeling of seedlings grown in vertically orientedsquare culture dishes was described previously (Cao et al., 2013; Dasguptaet al., 2014). Shoots and roots were collected separately, and roots were cut at1-cm intervals from the tip of the root. 14C levels (cpm) in shoots and root sectionswere determined by scintillation counting as described (Cao et al., 2013; Dasguptaet al., 2014). Two plants were pooled for analysis. Label in shoots wasstandardized relative to shoot mass, and label in roots was standardized rela-tive to segment length. All experiments in this study were conducted with 12 hof light and 12 h of dark rather than 14 h of light and 10 h of dark.
Tomeasure unlabeled Suc exuded from cut petioles at different times duringthe diurnal cycle (Fig. 4C), 14 to 16 mature rosette source leaves were harvestedfrom 40-d-old adult plants for each time point. Phloem exudation was done asreported by Tetyuk et al. (2013). In brief, leaves were cut and immediatelyplaced in 20 mM K2-EDTA. Then, all leaves were stacked on top of each other,petioles were recut under 20mMK2-EDTA, and cut endswere placed in reactiontubes with 1 mL of fresh 20mMK2-EDTA solution for 1 h at 21°C. Suc content inexudates was determined by using the Suc/D-Glc/D-Fru assay kit (Megazyme).
Sequence data for the genes studied in this article are listed in SupplementalTable 1.
Supplemental Data
The following supplemental materials are available.
Supplemental Figure S1. Growth characteristics of AVP1 lines.
Supplemental Figure S2. Autoradiograms of representative leaf disks.
Supplemental Figure S3. AtSUC2 expression in wild-type and AVP1 lines.
Supplemental Figure S4. Expression of genes relating to carbon fluxthrough central metabolism in shoots.
Supplemental Figure S5. Expression of genes relating to carbon fluxthrough central metabolism in roots.
Supplemental Table S1. List of gene sequences and primers used inRT-qPCR analysis.
ACKNOWLEDGMENTS
We thank Drs. Stevens Brumbley and Richard Dixon, University of NorthTexas, for the use of equipment.
Received September 9, 2015; accepted October 30, 2015; published November 3,2015.
LITERATURE CITED
Ainsworth EA, Bush DR (2011) Carbohydrate export from the leaf: a highlyregulated process and target to enhance photosynthesis and produc-tivity. Plant Physiol 155: 64–69
412 Plant Physiol. Vol. 170, 2016
Khadilkar et al.
www.plantphysiol.orgon March 23, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Alexandersson E, Saalbach G, Larsson C, Kjellbom P (2004) Arabidopsisplasma membrane proteomics identifies components of transport, signaltransduction and membrane trafficking. Plant Cell Physiol 45: 1543–1556
Aloni B, Wyse RE, Griffith S (1986) Sucrose transport and phloem un-loading in stem of Vicia faba: possible involvement of a sucrose carrierand osmotic regulation. Plant Physiol 81: 482–486
Ayre BG (2011) Membrane-transport systems for sucrose in relation towhole-plant carbon partitioning. Mol Plant 4: 377–394
Baltscheffsky H, Von Stedingk LV, Heldt HW, Klingenberg M (1966)Inorganic pyrophosphate: formation in bacterial photophosphorylation.Science 153: 1120–1122
Baltscheffsky M, Schultz A, Baltscheffsky H (1999) H+-proton-pumpinginorganic pyrophosphatase: a tightly membrane-bound family. FEBSLett 452: 121–127
Bao AK, Wang SM, Wu GQ, Xi JJ, Zhang JL, Wang CM (2008) Over-expression of the Arabidopsis H+-PPase enhanced resistance to salt anddrought stress in transgenic alfalfa (Medicago sativa L.). Plant Sci 176:232–240
Bradford MM (1976) A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding. Anal Biochem 72: 248–254
Braun DM, Wang L, Ruan YL (2014) Understanding and manipulatingsucrose phloem loading, unloading, metabolism, and signalling to en-hance crop yield and food security. J Exp Bot 65: 1713–1735
Brini F, Hanin M, Mezghani I, Berkowitz GA, Masmoudi K (2007) Over-expression of wheat Na+/H+ antiporter TNHX1 and H+-pyrophosphataseTVP1 improve salt- and drought-stress tolerance in Arabidopsis thalianaplants. J Exp Bot 58: 301–308
Cao T, Lahiri I, Singh V, Louis J, Shah J, Ayre BG (2013) Metabolic en-gineering of raffinose-family oligosaccharides in the phloem reveals al-terations in carbon partitioning and enhances resistance to green peachaphid. Front Plant Sci 4: 263
Chen LQ, Qu XQ, Hou BH, Sosso D, Osorio S, Fernie AR, Frommer WB(2012) Sucrose efflux mediated by SWEET proteins as a key step forphloem transport. Science 335: 207–211
Chiou TJ, Aung K, Lin SI, Wu CC, Chiang SF, Su CL (2006) Regulation ofphosphate homeostasis by microRNA in Arabidopsis. Plant Cell 18: 412–421
Clough SJ, Bent AF (1998) Floral dip: a simplified method forAgrobacterium-mediated transformation of Arabidopsis thaliana. PlantJ 16: 735–743
Cross JM, von Korff M, Altmann T, Bartzetko L, Sulpice R, Gibon Y,Palacios N, Stitt M (2006) Variation of enzyme activities and metabolitelevels in 24 Arabidopsis accessions growing in carbon-limited condi-tions. Plant Physiol 142: 1574–1588
Curtis MD, Grossniklaus U (2003) A Gateway cloning vector set for high-throughput functional analysis of genes in planta. Plant Physiol 133:462–469
Dasgupta K, Khadilkar AS, Sulpice R, Pant B, Scheible WR, Fisahn J,Stitt M, Ayre BG (2014) Expression of sucrose transporter cDNAs spe-cifically in companion cells enhances phloem loading and long-distancetransport of sucrose but leads to an inhibition of growth and the per-ception of a phosphate limitation. Plant Physiol 165: 715–731
Davies JM (1997) The bioenergetics of vacuolar H+ pumps. In RA Leigh, DSanders, eds, The Plant Vacuole, Vol 25. Academic Press, San Diego, pp340–363
DeWitt ND, Sussman MR (1995) Immunocytological localization of anepitope-tagged plasma membrane proton pump (H+-ATPase) in phloemcompanion cells. Plant Cell 7: 2053–2067
Fuglsang AT, Paez-Valencia J, Gaxiola RA (2011) Plant proton pumps:regulatory circuits involving H+-ATPase and H+-PPase. In M Geisler, KVenema, eds, Transporters and Pumps in Plant Signaling. Springer-Verlag, Heidelberg, Germany, pp 39–64
Gao F, Gao Q, Duan X, Yue G, Yang A, Zhang J (2006) Cloning of an H+-PPase gene from Thellungiella halophila and its heterologous expressionto improve tobacco salt tolerance. J Exp Bot 57: 3259–3270
Gaxiola RA, Li J, Undurraga S, Dang LM, Allen GJ, Alper SL, Fink GR(2001) Drought- and salt-tolerant plants result from overexpression ofthe AVP1 H+-pump. Proc Natl Acad Sci USA 98: 11444–11449
Gaxiola RA, Sanchez CA, Paez-Valencia J, Ayre BG, Elser JJ (2012) Ge-netic manipulation of a “vacuolar” H+-PPase: from salt tolerance to yieldenhancement under phosphorus-deficient soils. Plant Physiol 159: 3–11
Giaquinta RT (1983) Phloem loading of sucrose. Annu Rev Plant PhysiolPlant Mol Biol 34: 347–387
Gibon Y, Blaesing OE, Hannemann J, Carillo P, Höhne M, Hendriks JH,Palacios N, Cross J, Selbig J, Stitt M (2004) A Robot-based platform tomeasure multiple enzyme activities in Arabidopsis using a set of cyclingassays: comparison of changes of enzyme activities and transcript levelsduring diurnal cycles and in prolonged darkness. Plant Cell 16: 3304–3325
Gibon Y, Pyl ET, Sulpice R, Lunn JE, Höhne M, Günther M, Stitt M (2009)Adjustment of growth, starch turnover, protein content and centralmetabolism to a decrease of the carbon supply when Arabidopsis isgrown in very short photoperiods. Plant Cell Environ 32: 859–874
Gibon Y, Vigeolas H, Tiessen A, Geigenberger P, Stitt M (2002) Sensitiveand high throughput metabolite assays for inorganic pyrophosphate,ADPGlc, nucleotide phosphates, and glycolytic intermediates based on anovel enzymic cycling system. Plant J 30: 221–235
Gonzalez N, De Bodt S, Sulpice R, Jikumaru Y, Chae E, Dhondt S, VanDaele T, De Milde L, Weigel D, Kamiya Y, et al (2010) Increased leafsize: different means to an end. Plant Physiol 153: 1261–1279
Guo B, Jin Y, Wussler C, Blancaflor EB, Motes CM, Versaw WK (2008)Functional analysis of the Arabidopsis PHT4 family of intracellularphosphate transporters. New Phytol 177: 889–898
Hafke JB, van Amerongen JK, Kelling F, Furch ACU, Gaupels F, van BelAJE (2005) Thermodynamic battle for photosynthate acquisition be-tween sieve tubes and adjoining parenchyma in transport phloem. PlantPhysiol 138: 1527–1537
Harrison SJ, Mott EK, Parsley K, Aspinall S, Gray JC, Cottage A (2006) Arapid and robust method of identifying transformed Arabidopsis thali-ana seedlings following floral dip transformation. Plant Methods 2: 19
Heise R, Arrivault S, Szecowka M, Tohge T, Nunes-Nesi A, Stitt M,Nikoloski Z, Fernie AR (2014) Flux profiling of photosynthetic carbonmetabolism in intact plants. Nat Protoc 9: 1803–1824
Hendriks JH, Kolbe A, Gibon Y, Stitt M, Geigenberger P (2003) ADP-glucosepyrophosphorylase is activated by posttranslational redox-modification inresponse to light and to sugars in leaves of Arabidopsis and other plantspecies. Plant Physiol 133: 838–849
Krügel U, Veenhoff LM, Langbein J, Wiederhold E, Liesche J, FriedrichT, Grimm B, Martinoia E, Poolman B, Kühn C (2008) Transport andsorting of the Solanum tuberosum sucrose transporter SUT1 is affected byposttranslational modification. Plant Cell 20: 2497–2513; erratum KrügelU, Veenhoff LM, Langbein J, Wiederhold E, Liesche J, Friedrich T,Grimm B, Martinoia E, Poolman B, Kühn C (2009) Plant Cell 21: 4059–4060
Kühn C, Franceschi VR, Schulz A, Lemoine R, Frommer WB (1997)Macromolecular trafficking indicated by localization and turnover ofsucrose transporters in enucleate sieve elements. Science 275: 1298–1300
Lalonde S, Wipf D, Frommer WB (2004) Transport mechanisms for organicforms of carbon and nitrogen between source and sink. Annu Rev PlantBiol 55: 341–372
Langhans M, Ratajczak R, Lützelschwab M, Michalke W, Wächter R,Fischer-Schliebs E, Ullrich CI (2001) Immunolocalization of plasma-membrane H+-ATPase and tonoplast-type pyrophosphatase in theplasma membrane of the sieve element-companion cell complex in thestem of Ricinus communis L. Planta 213: 11–19
Lerchl J, Geigenberger P, Stitt M, Sonnewald U (1995) Impaired photoassimilatepartitioning caused by phloem-specific removal of pyrophosphate can becomplemented by a phloem-specific cytosolic yeast-derived invertase intransgenic plants. Plant Cell 7: 259–270
Li B, Wei A, Song C, Li N, Zhang J (2008) Heterologous expression of theTsVP gene improves the drought resistance of maize. Plant Biotechnol J6: 146–159
Li J, Yang H, Peer WA, Richter G, Blakeslee J, Bandyopadhyay A,Titapiwantakun B, Undurraga S, Khodakovskaya M, Richards EL, et al(2005) Arabidopsis H+-PPase AVP1 regulates auxin-mediated organdevelopment. Science 310: 121–125
Li X, Guo C, Gu J, Duan W, Zhao M, Ma C, Du X, Lu W, Xiao K (2014)Overexpression of VP, a vacuolar H+-pyrophosphatase gene in wheat(Triticum aestivum L.), improves tobacco plant growth under Pi and Ndeprivation, high salinity, and drought. J Exp Bot 65: 683–696
Li Z, Baldwin CM, Hu Q, Liu H, Luo H (2010) Heterologous expression ofArabidopsis H+-pyrophosphatase enhances salt tolerance in transgeniccreeping bentgrass (Agrostis stolonifera L.). Plant Cell Environ 33: 272–289
Liesche J, He HX, Grimm B, Schulz A, Kühn C (2010) Recycling of Sola-num sucrose transporters expressed in yeast, tobacco, and in maturephloem sieve elements. Mol Plant 3: 1064–1074
Plant Physiol. Vol. 170, 2016 413
AVP1 Enhances Phloem Loading and Transport
www.plantphysiol.orgon March 23, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Long AR, Williams LE, Nelson SJ, Hall JL (1995) Localization of mem-brane pyrophosphatase activity in Ricinus communis seedlings. J PlantPhysiol 146: 629–638
Lv S, Zhang K, Gao Q, Lian L, Song Y, Zhang J (2008) Overexpression ofan H+-PPase gene from Thellungiella halophila in cotton enhances salttolerance and improves growth and photosynthetic performance. PlantCell Physiol 49: 1150–1164
Lv SL, Lian LJ, Tao PL, Li ZX, Zhang KW, Zhang JR (2009) Overexpressionof Thellungiella halophila H+-PPase (TsVP) in cotton enhances droughtstress resistance of plants. Planta 229: 899–910
Ma F, Jazmin LJ, Young JD, Allen DK (2014) Isotopically nonstationary13C flux analysis of changes in Arabidopsis thaliana leaf metabolism dueto high light acclimation. Proc Natl Acad Sci USA 111: 16967–16972
Maeshima M (2000) Vacuolar H+-pyrophosphatase. Biochim Biophys Acta1465: 37–51
Matsuda Y, Liang GQ, Zhu YL, Ma FS, Nelson RS, Ding B (2002) TheCommelina Yellow Mottle Virus promoter drives companion-cell-specific gene expression in multiple organs of transgenic tobacco.Protoplasma 220: 51–58
Medberry SL, Lockhart BEL, Olszewski NE (1992) The commelina yellowmottle virus promoter is a strong promoter in vascular and reproductivetissues. Plant Cell 4: 185–192
Nolte KD, Koch KE (1993) Companion-cell specific localization of sucrosesynthase in zones of phloem loading and unloading. Plant Physiol 101:899–905
Paez-Valencia J, Patron-Soberano A, Rodriguez-Leviz A, Sanchez-Lares J,Sanchez-Gomez C, Valencia-Mayoral P, Diaz-Rosas G, Gaxiola R(2011) Plasma membrane localization of the type I H+-PPase AVP1 insieve element-companion cell complexes from Arabidopsis thaliana.Plant Sci 181: 23–30
Paez-Valencia J, Sanchez-Lares J, Marsh E, Dorneles LT, Santos MP,Sanchez D, Winter A, Murphy S, Cox J, Trzaska M, et al (2013) En-hanced proton translocating pyrophosphatase activity improves nitro-gen use efficiency in Romaine lettuce. Plant Physiol 161: 1557–1569
Park S, Li J, Pittman JK, Berkowitz GA, Yang H, Undurraga S, Morris J,Hirschi KD, Gaxiola RA (2005) Up-regulation of a H+-pyrophosphatase(H+-PPase) as a strategy to engineer drought-resistant crop plants. ProcNatl Acad Sci USA 102: 18830–18835
Pasapula V, Shen G, Kuppu S, Paez-Valencia J, Mendoza M, Hou P, ChenJ, Qui X, Zhu L, Zhang X, et al (2011) Expression of an Arabidopsisvacuolar H+-pyrophosphatase gene (AVP1) in cotton improves droughtand salt tolerance and increases fiber yield in field conditions. PlantBiotechnol J 9: 88–99
Pei L, Wang J, Li K, Li Y, Li B, Gao F, Yang A (2012) Overexpression ofThellungiella halophila H+-pyrophosphatase gene improves low phos-phate tolerance in maize. PLoS ONE 7: e43501
Pizzio GA, Paez-Valencia J, Khadilkar AS, Regmi KC, Patron-SoberanoA, Zhang S, Sanchez-Lares J, Furstenau T, Li J, Sanchez-Gomez C, et al(2015) Arabidopsis type I proton-pumping pyrophosphatase expressesstrongly in phloem, where it is required for pyrophosphate metabolismand photosynthate partitioning. Plant Physiol 167: 1541–1553
Rasband WS (2014) ImageJ. http://rsb.info.nih.gov/ijRea PA, Kim Y, Sarafian V, Poole RJ, Davies JM, Sanders D (1992) Vac-
uolar H+-translocating pyrophosphatases: a new category of ion trans-locase. Trends Biochem Sci 17: 348–353
Regmi KC, Zhang S, Gaxiola RA (2015) Apoplasmic loading in the ricephloem suggested by the presence of sucrose synthase and plasmamembrane localized proton pyrophosphatase. Ann Bot (Lond) (in press)
Riesmeier JW, Hirner B, Frommer WB (1993) Potato sucrose transporterexpression in minor veins indicates a role in phloem loading. Plant Cell5: 1591–1598
Robinson DG, Haschke HP, Hinz G, Hoh G, Maeshima M, Marty F (1996)Immunological detection of tonoplast polypeptide in the plasma mem-brane of pea cotyledon. Planta 198: 95–103
Rocha Facanha A, de Meis L (1998) Reversibility of H+-ATPase and H+-pyrophosphatase in tonoplast vesicles from maize coleoptiles and seeds.Plant Physiol 116: 1487–1495
Schilling RK, Marschner P, Shavrukov Y, Berger B, Tester M, Roy SJ,Plett DC (2014) Expression of the Arabidopsis vacuolar H⁺-pyrophosphatasegene (AVP1) improves the shoot biomass of transgenic barley and increasesgrain yield in a saline field. Plant Biotechnol J 12: 378–386
Seufferheld M, Lea CR, Vieira M, Oldfield E, Docampo R (2004) TheH+-pyrophosphatase of Rhodospirillum rubrum is predominantly locatedin polyphosphate-rich acidocalcisomes. J Biol Chem 279: 51193–51202
Smith JAC, Milburn JA (1980) Phloem turgor and the regulation of sucroseloading in Ricinus communis L. Planta 148: 42–48
Stitt M, Lilley RM, Gerhardt R, Heldt HW (1989) Metabolite levels inspecific cells and subcellular compartments of plant leaves. In MethodsEnzymol, Vol 174. Academic Press, pp 518–552
Tetyuk O, Benning UF, Hoffmann-Benning S (2013) Collection andanalysis of Arabidopsis phloem exudates using the EDTA-facilitatedmethod. J Vis Exp e51111
Truernit E, Sauer N (1995) The promoter of the Arabidopsis thaliana SUC2sucrose-H+ symporter gene directs expression of beta-glucuronidase tothe phloem: evidence for phloem loading and unloading by SUC2.Planta 196: 564–570
Turgeon R, Wolf S (2009) Phloem transport: cellular pathways and mo-lecular trafficking. Annu Rev Plant Biol 60: 207–221
Undurraga SF, Santos MP, Paez-Valencia J, Yang H, Hepler PK, FacanhaAR, Hirschi KD, Gaxiola RA (2012) Arabidopsis sodium dependent andindependent phenotypes triggered by H+-PPase up-regulation are SOS1dependent. Plant Sci 183: 96–105
van Bel AJE, Knoblauch M (2000) Sieve element and companion cell: thestory of the comatose patient and the hyperactive nurse. Aust J PlantPhysiol 27: 477–487
Vaughn MW, Harrington GN, Bush DR (2002) Sucrose-mediated tran-scriptional regulation of sucrose symporter activity in the phloem. ProcNatl Acad Sci USA 99: 10876–10880
Yadav UP, Ayre BG, Bush DR (2015) Transgenic approaches to alteringcarbon and nitrogen partitioning in whole plants: assessing the potentialto improve crop yields and nutritional quality. Front Plant Sci 6: 275
Yang H, Knapp J, Koirala P, Rajagopal D, Peer WA, Silbart LK, MurphyA, Gaxiola RA (2007) Enhanced phosphorus nutrition in monocots anddicots over-expressing a phosphorus-responsive type I H+-pyrophosphatase.Plant Biotechnol J 5: 735–745
Yang H, Zhang X, Gaxiola RA, Xu G, Peer WA, Murphy AS (2014) Over-expression of the Arabidopsis proton-pyrophosphatase AVP1 enhancestransplant survival, root mass, and fruit development under limitingphosphorus conditions. J Exp Bot 65: 3045–3053
Yang NS, Russell D (1990) Maize sucrose synthase-1 promoter directsphloem cell-specific expression of Gus gene in transgenic tobacco plants.Proc Natl Acad Sci USA 87: 4144–4148
Zhen RG, Kim EJ, Rea PA (1997) The molecular and biochemical basis ofpyrophosphate-energized proton translocation at the vacuolar mem-brane. In RA Leigh, D Sanders, eds, The Plant Vacuole, Vol 25. Aca-demic Press, pp 298–337
414 Plant Physiol. Vol. 170, 2016
Khadilkar et al.
www.plantphysiol.orgon March 23, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.





























