Embed Size (px)
Citation preview

JULIANA PROVASI CARDOSO
Contribuição da hiperexpressão do sistema de efluxo AdeABC e da
hiperprodução de AmpC na resistência aos β-lactâmicos em isolados clínicos de
Acinetobacter baumannii produtores de OXA-23 no Brasil
Tese apresentada à Universidade Federal
de São Paulo - Escola Paulista de Medicina,
para obtenção de título de Mestre em
Ciências.
São Paulo
2013

ii
JULIANA PROVASI CARDOSO
Contribuição da hiperexpressão do sistema de efluxo AdeABC e da
hiperprodução de AmpC na resistência aos β-lactâmicos em isolados clínicos de
Acinetobacter baumannii produtores de OXA-23 no Brasil
Orientadora: Profª. Drª. Ana Cristina Gales
Co-orientador: Dr. Rodrigo Cayô da Silva
Este trabalho foi realizado com o auxílio
financeiro fornecido pelo Conselho Nacional
de Desenvolvimento Científico e
Tecnológico - CNPq.
São Paulo
2013

iii
Cardoso, Juliana Provasi Contribuição da hiperexpressão do sistema de efluxo AdeABC e da hiperprodução de AmpC na resistência aos β-lactâmicos em isolados clínicos de Acinetobacter baumannii produtores de OXA-23 no Brasil Juliana Provasi Cardoso - São Paulo/SP - Brasil, 2013. xxi, 130f. Tese (Mestrado) - Universidade Federal de São Paulo - UNIFESP. Escola Paulista de Medicina. Programa de Pós-graduação em Infectologia. Departamento de Medicina, Disciplina de Infectologia. Título em Inglês: Contribution of the overexpression efflux pump AdeABC and hiperproduction of AmpC in the resistance to β-lactams in clinical isolates of Acinetobacter baumannii producing OXA-23 in Brazil.
1. Acinetobacter baumannii. 2. OXA-23. 3. Sistema de efluxo. 4. Brasil.

iv
UNIVERSIDADE FEDERAL DE SÃO PAULO
ESCOLA PAULISTA DE MEDICINA
DISCIPLINA DE INFECTOLOGIA
Chefe do Departamento:
Professora Drª Maria Tereza Zanella
Coordenador do curso de Pós-graduação:
Professor Dr. Ricardo Sobhie Diaz
Chefe da Disciplina de Infectologia:
Prof. Dr. Celso Franciso Hernandes Granato
São Paulo
2013

v
JULIANA PROVASI CARDOSO
Contribuição da hiperexpressão do sistema de efluxo AdeABC e da
hiperprodução de AmpC na resistência aos β-lactâmicos em isolados clínicos de
Acinetobacter baumannii produtores de OXA-23 no Brasil
BANCA EXAMINADORA:
Presidente:
Profa. Dra. Ana Cristina Gales
Professora Adjunta e Diretora do Laboratório ALERTA da Disciplina de Infectologia do
Departamento de Medicina da Universidade Federal de São Paulo - UNIFESP.
Titulares:
Profa. Dra. Anna Sara Shafferman Levin
Professora Associada do Departamento de Doenças Infecciosas e Parasitárias da
Faculdade de Medicina da Universidade de São Paulo e Presidente da Comissão de
Controle de Infecção Hospitalar do Hospital das Clínicas - HC-FMUSP.
Prof. Dr. José Rodrigues do Carmo Filho
Professor Adjunto da Pontifícia Universidade Católica de Goiás - PUC-GO e Analista
em saúde da Secretaria Municipal de Saúde de Goiânia.
Dra. Paola Cappellano
Médica Infectologista da Comissão de Epidemiologia Hospitalar do Hospital São Paulo
- HSP da Universidade Federal de São Paulo - UNIFESP e Coordenadora do grupo de
Racionalização de Antimicrobianos em Infecções em pacientes com Doenças Onco-
Hematológicas e Transplante de Medula Óssea.
Suplente:
Profa. Dra. Silvia Figueiredo Costa
Professora Associada ao Departamento de Doenças Infecciosas e Parasitárias da
Faculdade de Medicina da Uiversidade de São Paulo - USP e chefe do Laboratório de
Investigação Médica LIM-54 do Hospital das Clínicas da Faculdade de Medicina - HC-
FMUSP.

vi
“Por vezes sentimos que aquilo que fazemos não é senão uma gota de água
no mar. Mas o mar seria menor se lhe faltasse uma gota.”
Madre Tereza de Calcutá

vii
Dedicatória
Dedico este trabalho a todas as pessoas que contribuíram, de alguma forma, para sua
realização e àquelas, que um dia, o poderão usufruir.

viii
AGRADECIMENTOS
Agradeço primeiramente a Deus, por me conceder a vida e a sabedoria, por ser a
minha força nos momentos de fraqueza e por intervir por mim quando tudo parecia ser
impossível;
... à todos da minha família, em especial à minha mãe Valquiria Provasi, ao meu pai
Vitor Eduardo Cardoso, à minha avó Terezinha Bariani Provasi e ao meu avô Ari
Provasi (in memoriam) por não pouparem esforços, me dando a melhor educação, por
me ensinarem princípios, sempre acreditando no meu sucesso e por me apoiarem em
cada decisão difícil da vida;
... agradeço principalmente à minha mãe. Posso imaginar o quanto foi difícil para você
ver sua única filha saindo de casa... mas você sempre me apoiou nas minhas decisões
e sempre me deu muita força para seguir com os meus sonhos, me ajudou a superar
as dificuldades e esteve comigo nos momentos mais importantes. Muito obrigada... por
absolutamente tudo!
... ao meu tio Udo Kehrle, à minha tia Zuleika Provasi por serem meus pais postiços,
sempre me dando bons conselhos, me recebendo com todo coração e por terem me
ajudado tanto nesses anos em que eu mais precisei;
... às minhas primas Aline, Karen e Ellen Kehrle, pelos conselhos acadêmicos,
mesmo quando não entendiam nada sobre o assunto do meu estudo, pela amizade,
pelas muitas risadas, pela companhia, mesmo algumas de vocês estando longe, pela
força e por aguentarem minhas reclamações. Vocês são as irmãs que eu não tive;

ix
... aos meus tios Ricardo José Cardoso, Renato Cardoso Filho, á minha avó Lauria
Carésia Cardoso (in memoriam), e ao meu irmão João Vitor Cardoso, por serem os
primeiros a me receber em São Paulo de braços abertos, pelo carinho com que me
acolheram e por sempre me apoiarem nos meus estudos;
... à minha orientadora Profa. Drª. Ana Cristina Gales, por me acolher em seu
laboratório e acreditar na minha capacidade, por seus ensinamentos, conselhos e
conhecimentos incontestáveis, por ser a grande pesquisadora que é e por permitir a
concretização deste estudo. Não poderia ter realizado meu mestrado num lugar
melhor;
... ao Prof. Dr. Antonio Carlos Campos Pignatari, por todo conhecimento e apoio no
LEMC;
... ao meu co-orientador Dr. Rodrigo Cayô, por todo ensinamento, paciência,
dedicação, amizade, puxões de orelha e também incentivos, pelo apoio fundamental
no final do mestrado e por me guiar nesta jornada dos Acinetos;
... à Dra. Renata Cristina Picão que me recebeu no ALERTA, pela amizade e
carinho, pelos ensinamentos acadêmicos e da vida e por estar sempre à disposição;
... à querida Jéssica Werneck, que me introduziu ao maravilhoso mundo dos
Acinetos;
... à todos do Laboratório ALERTA: Adriana Nicolletti, Adriana Matos, Ana Carolina
Ramos, Bruna Nonato, Cecília Carvalhaes, Dandara Cassu, Danilo Xavier, Eloiza
Campana, Eliete Frigatto, Fernanda Petrolini, Fernanda Rodrigues, Graziela
Braun, Jhonatha Moura, Lorena Fehlberg, Lucas Andrade, Lygia Schandert,

x
Marina Visconde, Rafael Affini, Raquel Girardello, Rodrigo Cayô e Talita Baroni
que contribuíram de alguma forma com a realização deste trabalho, pelos
conhecimentos trocados, pela amizade e pela ótima convivência diária, que fez com
que esses três anos de estudo se tornassem mais divertidos e leves;
... à Raquel Girardello por todo apoio, auxílio e conhecimento que me ofereceu ao
longo do meu mestrado, principalmente na reta final, e por me ensinar a fazer aquelas
contas...;
... em especial à Adriana Matos, Ana Carolina Ramos, Fernanda Rodrigues,
Graziela Braun, Marina Visconde e Talita Baroni que me ajudaram imensamente no
final dos meus experimentos, pelos momentos de estudo, de desabafo e de alegrias
compartilhados. Vocês se tornaram mais que companheiras de laboratório;
... à todos do Laboratório Especial de Microbiologia Clínica - LEMC, pelo apoio e
auxílio para a concretização deste trabalho, em especial a Elke Gump, Talita
Rochetti e Paulo Bispo pelo suporte no Real Time;
... às minhas professoras e também “avó e mãe microbiológicas” da PUC-GO, Cláudia
Duque e Edlaine Rodrigues, por plantarem a sementinha da micro no meu coração;
... ao Prof. Dr. José Rodrigues do Carmo Filho, meu primeiro orientador na Iniciação
Científica, por despertar a pesquisadora que existe em mim;
... aos meus amigos, pela compreensão da minha ausência, por vibrarem a cada
vitória e por entenderem e me apoiarem na decisão de vir para São Paulo... por mais
que a distância nos separe, estarão sempre no meu coração;

xi
... às queridas Vanessa Ferreira e Mônica Simon por me estenderem não só a mão,
mas o braço no momento em que mais precisei... não sei como expressar a minha
gratidão pela ajuda que vocês me ofereceram num momento tão delicado;
... ao meu namorado Rafael Teixeira, que passou tantas noites, feriados e finais de
semana no laboratório comigo, sempre me incentivando a seguir em frente com meu
projeto, compreendendo a minha ausência e me acalmando nos momentos em que eu
parecia explodir, por ser meu companheiro de todas as horas e por ser o ombro amigo
sempre que precisei;
... à banca, que gentilmente aceitou meu convite para participar da minha defesa e
que reservou um tempo para estudar e contribuir com o meu trabalho;
... aos funcionários da UNIFESP, do Laboratório Central, médicos, pessoal da
limpeza e ao porteiro Edilson, pelos serviços e ajudas prestados;
... ao meu gatinho Ozzy, que me fez companhia enquanto eu escrevia minha tese;
... e as minhas “bichinhas” bactérias, em especial aos meus “filhinhos” Acinetos que
aprendi a “amar” e principalmente a respeitar esses seres tão pequenos e complexos,
aos quais tive o prazer de estudar seus mecanismos de resistência mais afundo.

xii
SUMÁRIO
ÍNDICE DE ABREVIATURAS E SIGLAS .................................................................... xiv
ÍNDICE DE TABELAS ............................................................................................... xviii
ÍNDICE DE FIGURAS ................................................................................................. xix
RESUMO .................................................................................................................... xx
ABSTRACT ................................................................................................................ xxi
1. INTRODUÇÃO .......................................................................................................... 1
2. REVISÃO BIBLIOGRÁFICA ...................................................................................... 4
2.1. Agente Etiológico ............................................................................................... 4
2.2. Epidemiologia das Infecções Causadas por A. baumannii ................................. 7
2.3. Opções de Tratamento para Infecções Causadas por A. baumannii ................ 10
2.3.1. Ampicilina/Sulbactam ................................................................................. 10
2.3.2. Minociclina ................................................................................................. 11
2.3.3. Tigeciclina .................................................................................................. 12
2.3.4. Polimixinas ................................................................................................. 14
2.3.5. Carbapenens ............................................................................................. 15
2.4. Mecanismos de Resistência aos Carbapenens em A. baumannii ..................... 18
2.4.1. Produção de β-Lactamases ....................................................................... 18
2.4.1.1. β-Lactamases de Espectro Ampliado (ESβLs).....................................19
2.4.1.2. Metalo-β-Lactamases (MβL).................................................................20
2.4.1.3. Cefalosporinase do Tipo AmpC (ADCs)...............................................21
2.4.1.4. Oxacilinases..........................................................................................22
2.4.1.4.1. OXA-23...................................................................................23
2.4.1.4.2. OXA-24/40..............................................................................29
2.4.1.4.3. OXA-51...................................................................................30
2.4.1.4.4. OXA-58...................................................................................30
2.4.1.4.5. OXA-143, OXA-182 e OXA-235 ............................................30
2.4.2. Impermeabilidade de Membrana Externa................................................... 31
2.4.3. Hiperexpressão de Sistemas de Efluxo ...................................................... 33
2.4.3.1. Sistema AdeABC.......................................................................35
2.4.4. Alteração das Proteínas Ligadoras de Penicilinas (PBPs) ......................... 37
3. OBJETIVOS ............................................................................................................ 39
3.1. OBJETIVO PRINCIPAL .................................................................................... 39
3.2. OBJETIVOS SECUNDÁRIOS .......................................................................... 39
4. MATERIAL E MÉTODOS ........................................................................................ 40

xiii
4.1. Amostras Bacterianas ...................................................................................... 40
4.2. Identificação dos Isolados de A. baumannii ...................................................... 42
4.2.1. “Matrix-Assisted Laser Desorption Ionization Time-Of-Flight Mass
Spectrometry” (MALDI-TOF MS) ......................................................................... 42
4.2.2. Sequenciamento do Gene rpoB ................................................................. 43
4.3. Análise da Similaridade Genética por PFGE .................................................... 45
4.4. “Multilocus Sequence Typing” (MLST) .............................................................. 47
4.5. Testes de Sensibilidade aos Antimicrobianos ................................................... 49
4.5.1. Avaliação Fenotípica da Hiperexpressão de Sistemas de Efluxo ............... 50
4.6. Pesquisa e Identificação de Genes de Resistência .......................................... 51
4.6.1. Avaliação da Hiperprodução de ADC (AmpC) ............................................ 54
4.7. Avaliação das Proteínas de Membrana Externa por SDS-PAGE ...................... 54
4.8. PCR em Tempo Real (qRT-PCR) para Quantificação da Transcrição do Sistema
de Efluxo AdeABC e das Proteínas de Membrana Externa CarO, 33-36 kDa,
OmpHMP e OmpW ................................................................................................. 55
4.8.1. Extração do RNA Total e Síntese de cDNA ............................................... 55
4.8.2. Quantificação Relativa da Transcrição Gênica ........................................... 56
4.8.3. Análise da Quantificação Relativa da Transcrição Gênica ......................... 58
5. RESULTADOS ....................................................................................................... 60
5.1. Identificação dos isolados bacterianos ............................................................. 60
5.2. Análise da similaridade genética (PFGE) e da ancestralidade (MLST) ............. 60
5.3. Perfil de sensibilidade aos antimicrobianos ...................................................... 65
5.4. Avaliação dos genes codificadores de β-lactamases e a relação desses com a
presença de ISAba1 ................................................................................................ 67
5.5. Avaliação fenotípica da hiperexpressão dos sistemas de efluxo ...................... 72
5.6. Avaliação fenotípica da alteração das proteínas de membrana externa pela
técnica de SDS-PAGE ............................................................................................ 76
5.7. Quantificação da expressão dos genes coficadores de porinas e do sistema de
efluxo AdeABC pela técnica de qRT-PCR ............................................................... 79
6. DISCUSSÃO ........................................................................................................... 81
7. CONCLUSÃO ....................................................................................................... 100
8. REFERÊNCIAS BIBLIOGRÁFICAS ...................................................................... 102

xiv
ÍNDICE DE ABREVIATURAS E SIGLAS
ABC - ATP Binding Cassette
ADC - Acinetobacter-derived cephalosporinases
AFLP - Amplified Fragment Lenght Polimorfism
AK - Amicacina
AmpC - Cefalosporinase chromosomal
AMP/SUL - Ampicilina/Sulbactam
ApaI - Acetobacter pasteurianus sub. pasteurianus
ARDRA - Amplified Ribosomal DNA Restriction Analysis
ARI - Acinetobacter Resistant to Imipenem
ATCC - American Type Culture Collection
ATP - Adenosina trifosfato
AZT - Aztreonam
BSA - Albumina de soro bovino
BSAC - British Society of Antimicrobial Chemotherapy
C - Citosina
CAZ - Ceftazidima
CC - Complexo Clonal
CCCP - Carbonyl Cyanide m-Chlorophenyl Hydrazone
cDNA - DNA complementar
CESP - Citrobacter freundii, Enterobacter spp., Serratia marcescens, Providencia
stuartii, Pseudmomonas aeruginosa e Morganella morganii
CHDL - Carbapenem-Hydrolyzing Class D β-Lactamase
CHEF-DR - Contour-Clamped Homogeneous Electric Fields
CIM - Concentração Inibitória Mínima
CIP - Ciprofloxacino
CLSI - Clinical and Laboratory Standards Institute

xv
Ct - Ciclo Threshold
CTX - Cefotaxima
CTX-M - Cefotaximase
Da - Dalton
DHP-1 - Dehidropeptidase-1
DLV - Double Locus Variant
DMT - Drug-Metabolite Transporter
E - Eficiência
EDTA - Ácido Etilenodiamino Tetra-Acético
ERIC-PCR - Enterobacterial Repetitive Intergenic Consensus - Polimerase Chain
Reaction
ESβL - β-lactamase de Espectro Ampliado
EU - Clone Europeu
EUCAST - The European Committee on Antimicrobial Susceptibility Testing
F - Forward
FEP - Cefepima
G - Guanina
GES - Guiana Extended Spectrum
GIM - German Imipenemase
GM - Gentamicina
HMM - High molecular mass
IMP - Imipenemase
IPM - Imipenem
ISAba - Insertion Sequence Acinetobacter baumannii
kDa - Kilodaltons
KPC - Klebsiella pneumoniae carbapenemase
LB - Luria Bertani
LEMC - Laboratório Especial de Microbiologia Clínica

xvi
LEV - Levofloxacino
LMM - Low molecular mass
M - Molar
MALDI-TOF MS - Matrix-Assisted Laser Desorption Ionization Time-Of-Flight Mass
Spectrometry
MC - Minociclina
MATE - Multidrug and Toxic Compound Extrusion
MDR - Multirresistentes
MER - Meropenem
MFS - Major Facilitador Superfamily
MgCl2 - Cloreto de magnésio
MGTs - Transglicosilases monofuncionais
MH - Müeller-Hinton
MLST - Multilocus Sequence Typing
mM - Milimolar
MRSA - Staphylococcus aureus resistente à meticilina
MTPX - Multiplex
MβL - Metalo-β-lactamase
NaCl - Cloreto de sódio
NDM - Nova Deli metalo-β-lactamase
NMP - 1-(1-naphthylmethyl)-piperazine
OMP - Outer Membrane Protein
OmpHMP - Outer membrane protein Heat-Modified Protein
OXA - Oxacilinase
PaβN - Phenylalanine arginyl β-naphtylamide
pb - Pares de base
PBPs - Proteínas ligadoras de penicilina
PCR - Reação em cadeia da polimerase

xvii
PER - Pseudomonas Extended Resistant
PFGE - Pulsed Field Gel Electrophoresis
pH - Potencial Hidrogeniônico
pmol - picomol
PO - Polimixina B
qRT-PCR - Quantificação da transcriptase reversa - reação em cadeia da polimerase
R - Reverse
Rep-PCR - Repetitive Element Palindromic – Polimerase Chain Reaction
RND - Resistance-Nodulation Division
rRNA - RNA ribossômico
SDS-PAGE - Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis
SG - Sequence Group
SHV - Sulfhydryl-Variable β-Lactamase
SIM - Seul Imipenemase
SLV - Single Locus Variant
SMR - Small Multidrug Resistance
SPM - São Paulo metalo-β-lactamase
ST - Sequence Type
TBE - Tris-Boreto-EDTA
TEM - Temoniera β-Lactamase
TGC - Tigeciclina
Tm - Temperatura de melting
TPPCl - Cloreto de tetrafenilfosfónio
TSB - Caldo Tríptico de Soja
VEB - β-Lactamase de Espectro Ampliado Vietnamita
VIM - Verona Imipenemase
VRE - Enterococcus resistente à vancomicina

xviii
ÍNDICE DE TABELAS
Tabela 1 - Variantes de OXA-23 descritas de acordo com a espécie de isolamento, ano e país de origem............................................................................................................................ 07 Tabela 2. Sequência de oligonucleotídeos das zonas internas e flanqueadoras do gene rpoB para identificação de Acinetobacter spp. de acordo com La Scola e colaboradores (2006)......................................................................................................................................... 44 Tabela 3. Oligonucleotídeos empregados para os sete genes utilizados para a caracterização do MLST, de acordo com Nemec e colaboradores (2008)................................ 48
Tabela 4. Sequência de oligonucleotídeos para identificação dos genes codificadores de -lactamases e sequência de inserção......................................................................................... 51 Tabela 5. Sequência de oligonucleotídeos utilizados para a quantificação dos genes codificadores do sistema de efluxo AdeABC, e dos genes das principais proteínas de membrana externa, pela técnica de qRT-PCR.......................................................................... 57 Tabela 6. Temperatura de melting e eficiência dos oligonucleotídeos utilizados na reação de qRT-PCR....................................…………….............................................................................. 58 . Tabela 7. Distribuição dos isolados de A. baumannii produtores de OXA-23 de acordo com a região geográfica e os genótipos determinados pelas técnicas de PFGE e MLST.............. 62 Tabela 8. Perfil de sensibilidade aos antimicrobianos entre os isolados de A. baumannii produtores de OXA-23............................................................................................................... 66 Tabela 9. Caracterização das β-lactamases e perfil de sensibilidade aos antimicrobianos entre os 31 isolados de A. baumannii produtores de OXA-23...................................................
69
Tabela 10. Resultados da CIM para ceftazidima, cefotaxima, imipenem e meropenem dterminadas pela técnica de microdiluição em caldo, realizada na presença e na ausência do inibidor de sistemas de efluxo PAβN.....................................................................................
75
Tabela 11. Perfil das proteínas de membrana externa dos isolados de A. baumannii produtores de OXA-23 baseando nos géis de SDS-PAGE........................................................
77
Tabela 12. Expressão relativa dos genes codificadores das OMPs OmpW, CarO, Omp33-36 e OmpHMP e da bomba de efluxo AdeB dos isolados clínicos comparados a expressão basal dos mesmos genes na ATCC BAA-1709.........................................................................
80

xix
ÍNDICE DE FIGURAS
Figura 01. Estruturas químicas das moléculas dos carbepenens e de seus precursores................................................................................................................................... 16 Figura 2. Distribuição mundial de OXA-23 em isolados clínicos de A. baumannii resistentes aos carbapenens.......................................................................................................................... 25 Figura 3. Distribuição mundial das variantes de OXA-23 descritas em Acinetobacter spp. e em K. pneumoniae....................................................................................................................... 28 Figura 4. Esquema do sistema de efluxo AdeABC.................................................................... 35 Figura 5. Representação do operon do sistema AdeABC............……………………........……. 36 Figura 6: Distribuição dos 31 isolados de A. baumannii produtores de OXA-23 apresentando distintos perfis clonais de acordo com a unidade da federação brasileira................................... 41 . Figura 7. Dendrograma do perfil genotípico determinado pela técnica de PFGE dos isolados de A. baumannii produtores de OXA-23 avaliados neste estudo.…............................................ 61 Figura 8. Árvore filogenética entre os diferentes STs encontrados no presente estudo e os demais STs descritos no mundo até o momento.…………….....................................…............. 64 Figura 9. Géis de SDS-PAGE das proteínas de membrana externa dos diferentes isolados de A. baumannii produtores de OXA-23 no Brasil.…...................................................................…. 78 Figura 10. Modelo de um complexo clonal.......…...................................................................…. 86

xx
RESUMO
A produção de OXA-23 é o principal mecanismo de resistência aos carbapenens em isolados de A. baumannii no Brasil e surtos causados por esses micro-organismos têm sido reportados em todo o mundo. Embora as cefalosporinas de espectro ampliado (ESCph) não sejam hidrolisadas pelas carbapenemases de classe D, isolados brasileiros de A. baumannii produtores de OXA-23 apresentam altos níveis de resistência a esses antimicrobianos. O presente estudo objetivou avaliar a contribuição da hiperexpressão do sistema de efluxo AdeABC e da hiperprodução de AmpC na resistência aos β-lactâmicos em isolados brasileiros de A. baumannii produtores de OXA-23. Para o estudo foram selecionados 31 isolados de A. baumannii produtores de OXA-23 de quatorze hospitais privados localizados em sete estados brasileiros e que apresentavam diferentes padrões de PFGE. A identificação dos isolados de A. baumannii foi realizada pelo MALDI-TOF MS e confirmada pelo sequenciamento do gene rpoB. O perfil clonal foi novamente confirmado pela técnica de PFGE e analisado pelo programa “BioNumerics”. A produção de genes codificadores de β-lactamases e sua associação com a presença de ISAba1 foi confirmada por PCR seguido de sequenciamento. O teste de sensibilidade foi realizado pela metodologia de microdiluição em caldo, segundo o CLSI. Para avaliar fenotipicamente a hiperexpressão de sistemas de efluxo, as CIMs para os β-lactâmicos foram comparadas na presença e na ausência do inibidor de sistemas de efluxo PAβN. A ancestralidade dos diferentes clones foi avaliada pela técnica de MLST. O perfil de proteínas de membrana externa foi avaliado por SDS-PAGE. Os níveis de transcrição dos genes adeB, carO, ompA, ompW e omp33-36 foram avaliados pela técnica de qRT-PCR comparando com os resultados obtidos da ATCC BAA-1709 de A. baumannii multissensível. Todos os isolados foram identificados como A. baumannii. Foi observada a presença de ISAba1 localizada a montante dos genes blaOXA-23, blaOXA-51-like e blaAmpC em 100%, 54,8% e 77,4% dos isolados, respectivamente. Um grande número de variantes da CHDL cromossômica do grupo OXA-51 foi verificada entre os isolados brasileiros de A. baumannii, estando a variante estreitamente relacionada ao respectivo padrão de PFGE/ST. Altos níveis de resistência foram observados para ampicillina/sulbactam, ESCph, aztreonam, carbapenens, fluoroquinolonas, aminoglicosídeos e tigeciclina. Somente a polimixina B e a minociclina apresentaram atividade contra os isolados avaliados (CIM90 de 0,5 µg/mL, ambos). Os 31 perfis de PFGE foram agrupados em 12 STs diferentes, dos quais sete apresentaram sequências alélicas únicas. A maioria dos isolados pertencia aos complexos clonais CC79, CC15 e CC1, sendo o ST79 o mais prevalente entre os isolados avaliados (n=12; 38,7%). Os demais STs encontrados foram: ST15 (n=6; 19,4%), ST1 (n=4; 12,9%) e ST162, ST46, ST188, ST189, ST190, ST191, ST192, ST228 e ST299 (n=1; 3,2%, cada). Os isolados apresentando o gene blaAmpC associado a ISAba1 apresentaram altas taxas de resistência às ESCph (ceftazidima, CIMs de 32 a >1024 µg/mL; cefotaxima, CIMs de 256 a >1024 µg/mL), contrastando com os isolados que não apresentavam tal associação (ceftazidima, CIMs de 8 a 128 µg/mL; cefotaxima, CIMs de 16 a 64 µg/mL). Diminuição da CIMs ≥ 2 diluições para meropenem e ceftazidima foi verificado em 80,6% (n=25) e 54,8% (n=17), respectivamente. A hiperexpressão do gene adeB foi observada em 74,2% (n=23) dos isolados, principalmente naqueles isolados pertencentes aos complexos clonais. Por outro lado, foi observada uma diminuição da expressão do gene carO. Apesar disso, o perfil de SDS-PAGE demonstrou uma grande reorganização das proteínas de membrana externa entre os isolados avaliados, destacando-se a ausência de uma porina de 21 kDa, compatível com a OmpW. A resistência às ESCph nos isolados brasileiros de A. baumannii produtores de OXA-23 está relacionada a hiperprodução de AmpC, além disso, a hiperexpressão de sistemas de efluxo contribui na elevação das CIMs para meropenem e ceftazidima. A maioria dos clones de A. baumannii produtores de OXA-23 circulantes no Brasil, pertencem aos principais complexos clonais descritos no mundo e apresentam um fenótipo MDR. A resistência aos β-lactâmicos entre os isolados de A. baumannii produtores de OXA-23 no Brasil é multifatorial, o que dificulta o seu completo entendimento.

xxi
ABSTRACT
The production of OXA-23 is the main mechanism of carbapenem-resistance among isolates of A. baumannii in Brazil, and outbreaks caused by these microrganisms have been reported worldwide. Although extended-spectrum cephalosporins (ESCph) are not hydrolyzed by the carbapenemases of class D, OXA-23-producing A. baumannii isolated in Brazil show high resistant rates to these antimicrobials agents. The present study aimed to evaluate the contribution of overexpression of efflux pumps AdeABC and overproduction of AmpC to the β-lactams resistance among A. baumannii Brazilian isolates producers of OXA-23. For this study, we selected 31 un-related isolates of A. baumannii producing OXA-23 from fourteen private hospitals located in seven Brazilian states. The identification of the isolates as A. baumannii was performed by MALDI-TOF MS and confirmed by sequencing of the rpoB gene. The genetic relatedness was evaluated by PFGE technique and analyzed by the BioNumerics software. The production of β-lactamase encoding genes and its association with the presence of ISAba1 was confirmed by PCR followed by sequencing. The susceptibility testing was performed by broth microdilution method according to CLSI. To assess the phenotype overexpression of efflux pumps, the MICs for the β-lactams were compared in the presence and absence of the inhibitor efflux pumps, PAβN. The filogenetic relationship among different clones was assessed by MLST. The outer membrane protein profiles were evaluated by SDS-PAGE. Transcription levels of the genes adeB, carO, ompA, ompW and omp33-36 were assessed by qRT-PCR technique compared to those obtained for the ATCC BAA-1709, multi-drug susceptible A. baumannii strain. All isolates were identified as A. baumannii. It was observed the presence of ISAba1 located upstream of the gene blaOXA-23, blaOXA-51-like and blaAmpC in 100%, 54.8% and 77.4% of the isolates, respectively. A large number of variants of the OXA-51 group was found among the A. baumannii Brazilian isolates, which were closely related to the corresponding PFGE pattern/ST. High resistant rates were observed for ampicillin/sulbactam, ESCph, aztreonam, carbapenems, fluoroquinolones, aminoglycosides and tigecycline. Only polymyxin B and minocycline showed activity against these isolates (MIC90, 0.5 μg/mL, for both drugs). The 31 PFGE profiles were grouped into 12 different STs, of which seven had unique allelic sequences. Most isolates belonged to clonal complexes CC79, CC15 and CC1, being ST79 the most prevalent among the isolates (n=12, 38.7%) evaluated. The remaining STs founded were: ST15 (n=6, 19.4%), ST1 (n=4, 12.9%) and ST162, ST46, ST188, ST189, ST190, ST191, ST192, ST228 and ST299 (n=1, 3.2%; each). The isolates presenting the blaAmpC gene associated with ISAba1 showed high resistant rates to ESCph (ceftazidime MICs from 32 to >1024 μg/ml; cefotaxime MICs from 256 to >1024 μg/mL), in contrast to those isolates that did not carry ISAba1 upstream blaAmpC and showed lower ceftazidime (8 to 128 μg/mL) and cefotaxime MICs (16 to 64 μg/mL). A ≥ 2 dilution decrease in the MICs of meropenem and ceftazidime was observed for 80.6% (n=25) and 54.8% (n = 17) of OXA-23-producing Acinetobacter baumannii, respectively. Overexpression of the gene adeB was observed in 74.2% (n=23) of isolates, particularly those isolates belonging to clonal complexes. Moreover, we observed a decrease of carO gene expression in the majority of the isolates. Nevertheless, the profile of SDS-PAGE showed a major reorganization of the outer membrane proteins among isolates, highlighting the absence of a 21 kDa porin, consistent with the OmpW. Resistance to ESCph in A. baumannii Brazilian isolates producers of OXA-23 is related to overproduction of AmpC coupled with the overexpression of efflux pumps, which contribute to the increased MICs for ceftazidime and meropenem. Most of OXA-23-producing A. baumannii clones circulating in Brazil belong to the major clonal complexes described in the world Resistance to β-lactams among isolates of A. baumannii producing OXA-23 in Brazil is multifactorial, which hinders its complete understanding.

INTRODUÇÃO
1
1. INTRODUÇÃO
Acinetobacter spp. são micro-organismos amplamente distribuídos na natureza,
possuindo a característica de se adaptarem a diversos locais e condições que seriam
desfavoráveis para muitos outros micro-organismos (Higgins et al., 2010). Essas
características estão relacionadas ao fato de que isolados de Acinetobacter spp.
necessitam de poucos nutrientes para se manterem metabolicamente ativos, o que os
tornam facilmente adaptados ao ambiente nosocomial (Higgins et al., 2010). Além de
serem intrinsicamente resistentes a diversos antimicrobianos, possuem uma grande
capacidade em adquirir novos genes de resistência, sendo assim, são considerados
patógenos de grande importância clínica, principalmente, no ambiente hospitalar.
Cepas de Acinetobacter spp. multirresistentes (MDR) tem se tornado uma
preocupação mundial. Uma vez instaladas em uma determinada unidade ou, até
mesmo, em todo o hospital, esses micro-organismos podem causar surtos de difícil
controle e erradicação (Villegas & Hartstein, 2003). O fenótipo MDR limita,
consideravelmente, as opções terapêuticas para o tratamento das infecções causadas
por Acinetobacter spp., dentre as quais, se destacam os carbapenens, a tigeciclina, a
ampicilina/sulbactam e as polimixinas (Karageorgopoulos & Falagas, 2008).
Entretanto, na última década, as taxas de resistência aos carbapenens vem
aumentando consideravelmente em todo o mundo.
De acordo com os dados do Programa SENTRY (Gales et al., 2012), a taxa de
resistência aos carbapenens entre as amostras de Acinetobacter spp. isoladas em
centros brasileiros, aumentou 60% em uma década, passando de 12,6%, no período
de 1997 a 1999, para 71,4% no período de 2008 a 2010. A causa desse aumento
pode ser justificada, em parte, pela disseminação de clones produtores da
carbapenemase de classe D OXA-23, não somente no Brasil, mas em todo o mundo
(Perez et al., 2007; Werneck et al., 2011a). Apesar das altas taxas de resistência aos
carbapenens verificadas nos isolados brasileiros de A. baumannii, esses

INTRODUÇÃO
2
antimicrobianos ainda constituem a primeira opção para o tratamento empírico de
infecções causadas por esses micro-organismos, pois se difundem amplamente pelos
tecidos, são bem tolerados e apresentam reações adversas mínimas (Nicolau, 2008).
No Brasil, a produção de OXA-23 é o mecanismo de resistência aos
carbapenens mais frequentemente relatado em isolados clínicos de A. baumannii;
porém, sua produção isolada não afeta, de modo significativo, a atividade dos
carbapenens, fazendo-se necessária a presença da sequência de inserção ISAba1
para aumentar a expressão da mesma e, dessa forma, elevar o grau de resistência a
esses antimicrobianos (Poirel & Nordmann, 2006). Embora a produção de OXA-23
associada à ISAba1 justifique a resistência aos carbapenens nos isolados brasileiros,
o mesmo não ocorre com as altas Concentrações Inibitórias Mínimas (CIMs) para às
cefalosporinas de amplo espectro verificadas nesses isolados (Werneck et al., 2011a;
Gales et al., 2012).
A OXA-23 é uma CHDL (Carbapenem-Hydrolyzing Class D β-Lactamase)
mundialmente disseminada e, portanto, foco de muitos estudos, o que acarreta no
desinteresse em estudar outros mecanismos de resistência aos β-lactâmicos pouco
conhecidos, porém não menos importantes. Vários sistemas de efluxo foram descritos
em A. baumannii e, quando hiperexpressos, conseguem ejetar diferentes classes de
antimicrobianos, contribuindo para o fenótipo MDR. Entretanto, seriam os carbapenens
e as cefalosporinas de amplo espectro ejetados eficientemente para o meio
extracelular? O sistema de efluxo AdeABC é o mais importante e estudado dentre os
sistemas descritos em A. baumannii e estudos anteriores demonstraram, ainda que
indiretamente, que a hiperexpressão desse sistema pode contribuir com a resistência
aos carbapenens, principalmente quando associado à produção de CHDLs (Héritier et
al., 2005a, Héritier et al., 2005b).
O estudo de Peleg e colaboradores (2007a) analisou a ação do inibidor de
sistema de efluxo PAβN (Phenyl-Arginine-β-Naphthylamide) em isolados clínicos de A.
baumannii com elevadas CIMs para tigeciclina, cefepima e ceftazidima. Os autores

INTRODUÇÃO
3
verificaram uma diminuição expressiva das CIMs para esses antimicrobianos na
presença do inibidor, bem como para os carbapenens. No mesmo ano, foi
demonstrado ser o sistema de efluxo AdeABC, o responsável pela resistência a
tigeciclina nesses isolados (Peleg et al., 2007b).
Acredita-se que a associação da hiperexpressão de sistemas de efluxo com a
produção de OXA-23 em A. baumannii poderia aumentar significativamente as CIMs
dos β-lactâmicos, podendo levar à falência terapêutica quando estes forem utilizados
clinicamente. As amostras de Acinetobacter spp. isoladas em hospitais brasileiros
geralmente são resistentes a todos os β-lactâmicos, incluindo as cefalosporinas de
amplo espectro, que não são substratos das CHDLs. Sendo assim, outros
mecanismos presentes concomitantemente, poderiam estar contribuindo com esse
fenótipo, como o aumento da produção de AmpC associado a ISAba1, ou o aumento
da expressão de sistemas de efluxo (Bergogne-Bérézin & Towner, 1996; Bou &
Martínez-Beltrán, 2000; Corvec et al., 2003; Segal et al., 2004).
A real contribuição dos sistemas de efluxo e/ou da hiperprodução de AmpC na
elevação da CIMs para os carbapenens e para as cefalosporinas de amplo espectro
em isolados brasileiros de A. baumannii produtores de OXA-23 ainda é desconhecida.
Diante do provavável papel dos mecanismos apresentados e da interação dos
mesmos, o presente trabalho teve por objetivo estudar a contribuição do sistema de
efluxo AdeABC e da hiperprodução de AmpC na resistência aos β-lactâmicos em
isolados clínicos de A. baumannii produtores de OXA-23 no Brasil.

REVISÃO BIBLIOGRÁFICA
4
2. REVISÃO BIBLIOGRÁFICA
2.1. Agente Etiológico
A história do gênero Acinetobacter começou na Holanda em 1911, onde o
pesquisador Martinus Willem Beijerinck descreveu em uma amostra de solo, o primeiro
micro-organismo originalmente denominado como Micrococcus calco-aceticus
(Beijerinck, 1911). Durante muitos anos, espécies como Micrococcus calco-aceticus,
Alcaligenes hemolysans, Mima polymorpha, Moraxella lwoffii, Herellea vaginicola e
Bacterium anitratum foram classificadas como sendo espécies diferentes. Somente em
1954, Brisou & Prevot (1954) propuseram o gênero Acinetobacter - que em grego
significa bacilo não móvel - para separar as espécies sem motilidade das demais
espécies móveis dentro do gênero Achromobacter a qual pertenciam.
Em 1968, Baumann e colaboradores (1968) fizeram um amplo estudo e
reclassificaram essas espécies, agrupando-as no gênero Acinetobacter, proposto por
Brisou e Prevot, 14 anos antes (Howard et al., 2012). Essas bactérias continham
diversas características bioquímicas em comum, sendo elas: (i) diferentes formas de
apresentação na coloração de Gram, ou seja, bacilos Gram negativos em sua fase
exponencial e morfologia de cocobacilos Gram positivos ou lábeis em sua fase
estacionária, devido a sua resistência à descoloração pelo álcool; (ii) produção de
cápsula por algumas cepas; (iii) assimilação de nitrato; (iv) crescimento somente na
presença de oxigênio; (v) não fastidiosas; (vi) não fermentadoras da glicose; (vii) não
produtoras de oxidase; (viii) produtoras de catalase; (ix) imóveis; (x) não formadoras
de esporos; (xi) não produtoras de pigmentos e (xii) relação G+C em torno de 39 a
47% (Baumann et al., 1968).
Somente em 1971, através do admirável trabalho de Baumann e colaboradores
realizado em 1968, o Subcomitê de Taxonomia de Moraxella e Bactérias Relacionadas
reconheceu o gênero Acinetobacter (Howard et al., 2012). Em 1986, Bouvet & Grimont
propuseram mais quatro novas espécies: A. baumannii, A. junii, A. haemolyticus e A.

REVISÃO BIBLIOGRÁFICA
5
johnsonii e, desde então, novas espécies tem sido descritas. Em 1991, Gerner-Smidt e
colaboradores propuseram um grupo incluindo as espécies de Acinetobacter de maior
importância clínica e que possuíam características fenotípicas semelhantes,
denominando-o “Complexo Acinetobacter calcoaceticus-baumannii”. Esse complexo
inclui as espécies A. calcoaceticus, A. baumannii, A. pittii e A. nosocomialis. Como
possuem características bioquímicas muito semelhantes, a distinção dessas espécies
constitui um grande desafio para os laboratórios clínicos de microbiologia. Entretanto,
com os avanços da biologia molecular, essas quatro espécies conseguem ser bem
diferenciadas. A exceção do A. calcoaceticus, que é comumente encontrado no meio
ambiente, as outras três especies pertencentes ao complexo A. calcoaceticus-
baumannii são frequentemente isoladas no ambiente hospitalar e, portanto, possuem
grande relevância clínica (Espinal et al., 2012).
Atualmente, isolados do gênero Acinetobacter spp. pertencem ao Reino
Bacteria, Filo Proteobacteria, Classe Gammaproteobacteria, Ordem Pseudomonadales
e Familia Moraxellaceae (Rossau et al., 1991; Vaneechoutte et al., 2011). Até o
momento, foram descritas 27 espécies reconhecidas e validadas e mais seis espécies
propostas, mas ainda não reconhecidas pela comunidade internacional (Towner, 2009;
Cayô, 2012). Nos laboratórios de pesquisa é comum identificar essas espécies por
metodologias moleculares, de modo a se obter resultados confiáveis e
discriminatórios. Porém, a implementação dessas técnicas nem sempre são custo-
efetivas na rotina laboratorial.
Em muitos laboratórios de microbiologia, as técnicas bioquímicas manuais para
identificação bacteriana ainda são empregadas. Entretanto, tais técnicas são
demoradas e para algumas bactérias, como no caso do gênero Acinetobacter, não são
discriminatórias o suficiente devido à semelhança catabólica entre suas espécies,
principalmente dentro complexo A. calcoaceticus-baumannii. Tais problemas também
comprometem o uso dos aparelhos automatizados como o Phoenix® (BD, New Jersey,
USA), o MicroScan-WalkAway® (Siemens, Munich, Germany) e o Vitek® (Biomerieux,

REVISÃO BIBLIOGRÁFICA
6
Marcy l'Etoile, France) na diferenciação das espécies de Acinetobacter spp. (Espinal et
al., 2012).
Recentemente, a técnica “Matrix Assisted Laser Desorption/Ionization – Time
Of Flight Mass Spectrometry” (MALDI-TOF MS) foi desenvolvida para a identifcação
rápida e acurada de micro-organismos (Seng et al., 2009). O MALDI-TOF MS é uma
técnica de proteômica que ioniza proteínas e as identifica pela sua relação
massa/carga e, nos últimos anos, vem sendo empregada na rotina laboratorial por
apresentar boa acurácia na identificação de muitas espécies bacterianas, com uma
ótima relação custo/benefício e, principalmente, pela rapidez na liberação dos
resultados (Seng et al., 2009, Espinal et al., 2012). O MALDI-TOF MS constitui uma
boa metodologia para a identificação do complexo A. calcoaceticus-baumannii desde
que se inclua o espectro de A. nosocomialis em seu banco de dados (Espinal et al.,
2012).
Além do MALDI-TOF MS, uma vasta gama de técnicas moleculares são
utilizadas na identificação e diferenciação das espécies de Acinetobacter spp., dentre
as quais podemos citar: i) a hibridização DNA-DNA que é utilizada na avaliação da
taxonomia para novas espécies descritas (Dijkshoorn & Nemec, 2008); ii) o
sequenciamento da porção 16S do rRNA (Ibrahim et al., 1997); iii) o ARDRA
“Amplified Ribosomal DNA Restriction Analysis” (Vaneechoutte et al., 1995); iv) o
AFLP “Amplified Fragment Lenght Polimorfism” (Janssen et al., 1996); v) a ribotipagem
(Gerner-Smidt et al., 1991), vi) o sequenciamento das regiões espaçadoras
intergênicas da porção 16S-23S do rRNA (Chang et al., 2005) e vii) a análise de
restrição das regiões espaçadoras intergênicas da porção 16S-23S ribossomal
(Dolzani et al., 1995). Além destas, outras técnicas como a análise dos perfis das
proteínas do envelope celular (Dijkshoorn et al., 1987; Dijkshoorn et al., 1990) e a
tipagem do antígeno O por anticorpos monoclonais, constituem outras técnicas
utilizadas para identificação de amostras de Acinetobacter spp. (Pantophlet et al.,
2002).

REVISÃO BIBLIOGRÁFICA
7
Dentre tantas técnicas descritas, o sequenciamento do gene rpoB, codificador
da porção β da RNA polimerase, é considerado como o mais fidedigno, pois diferencia
não somente as espécies do complexo A. calcoaceticus-baumannii como também as
demais espécies do gênero (Gundi et al., 2009). Esse gene de apenas uma cópia é
altamente variável, apresentando quatro regiões que são altamente discriminatórias,
sendo duas regiões internas (zona 1, 350 pb e zona 2, 450 pb) ao gene rpoB e duas
regiões flanqueadoras variáveis, sendo a primeira de 301 pb a 310 pb e a segunda de
86 pb a 177 pb (La Scola et al., 2006).
2.2. Epidemiologia das Infecções Causadas por A. baumannii
A. baumannii, ao contrário de outras espécies deste gênero, raramente é
encontrado no meio ambiente (Peleg et al., 2008), sendo isolado principalmente no
ambiente nosocomial, como, equipamentos médicos, fômites e colonizando
funcionários e pacientes do hospital (McConnell et al., 2013). O habitat natural de A.
baumannii ainda não é conhecido e, apesar de muito especulado, nenhum estudo
concluiu o seu nicho ecológico até o momento (Peleg et al., 2008). Entretanto, a
sazonalidade parece estar correlacionada à frequência de isolamento de A. baumannii
em amostras clínicas, sendo mais prevalente em climas tropicais, como mostrado no
estudo de Chu e colaboradores (1999) em Hong Kong. Esse estudo relatou que
durante o verão, 53% dos estudantes de medicina e enfermeiras avaliados tiveram sua
pele colonizada por A. baumannii, enquanto no inverno, essa porcentagem reduziu
para 32%. Fora do ambiente hospitalar, um dado interessante foi publicado no estudo
conduzido por La Scola & Raoult (2004), o qual encontrou A. baumannii no corpo de
21% dos Pediculus humanus estudados provenientes de moradores de rua. Eles
concluíram que a alta prevalência desse patógeno em piolhos humanos
eventualmente poderia causar bacteremia nesses indivíduos. Já Berlau e
colaboradores (1999b) estudaram a prevalência de Acinetobacter em vegetais e
constataram que de 177 amostras, 27% continham A. baumannii. Esse dado contradiz

REVISÃO BIBLIOGRÁFICA
8
com a informação de que A. baumannii raramente é encontrado fora do ambiente
hospitalar. Por outro lado, A. baumannii raramente é encontrado colonizando o corpo
humano, sendo isolado de 0,5% a 3% na pele (Seifert et al., 1997, Berlau et al.,
1999a) e em 0,8% do trato gastrointestinal de humanos saudáveis (Dijkshoorn et al.,
2005).
Embora sua fonte natural ainda permaneça uma incógnita, sabe-se que este é
o principal micro-organismo relacionado a surtos hospitalares, capaz de causar
infecção em diversos sítios corpóreos de pacientes hospitalizados (Peleg et al., 2008).
As infecções mais comumente causadas por A. baumannii serão descritas a seguir.
A pneumonia associada à ventilação mecânica é uma das infecções mais
comuns causada por A. baumannii no ambiente hospitalar e é precedida pela
colonização das vias aéreas (Dijkshoorn et al., 2007), cuja taxa de mortalidade nestes
casos fica entre 40% a 70% (Fagon et al., 1996; Garnacho et al., 2003). A pneumonia
adquirida na comunidade causada por este patógeno é rara e tem sido associada a
fatores predisponentes como alcoolismo ou doença pulmonar obstrutiva crônica.
Nesses casos, a taxa de mortalidade também é alta, variando entre 40% e 60%
(Anstey et al., 1992; Chen et al., 2001; Anstey et al.,2002; Leung et al., 2006).
Embora as queimaduras também sejam um importante sítio de infecção por A.
baumannii, é difícil a diferenciação entre colonização e infecção nesses casos. Além
disso, o tratamento possui uma dificuldade a mais, pois há uma menor penetração do
antimicrobiano nesses locais (McConnell et al., 2013). Durante a guerra do
Afeganistão e Iraque, por exemplo, 22% das infecções causadas por A. baumannii, em
soldados americanos feridos em campo de batalha, tiveram como sítio infeccioso a
pele queimada, sendo que 53% desses isolados eram MDR (Keen et al., 2010). Outro
problema comum em combatentes de guerra é a infecção de pele e partes moles
(Murray et al., 2006; Johnson et al., 2007; Scott et al., 2007; Sebeny et al., 2008). Scott
e colaboradores (2007) coletaram amostras de pele de soldados feridos, do solo e das
áreas de tratamento nos campos de batalha do Iraque, para verificar a presença de A.

REVISÃO BIBLIOGRÁFICA
9
baumannii. Os autores isolaram A. baumannii somente em 0,6% da pele dos pacientes
e em 2% das amostras de solo. Entretanto, esse micro-organismo foi isolado em todas
as áreas de tratamento coletadas, sugerindo que essa era a fonte de contaminação.
Fora do contexto militar, A. baumannii também foi encontrado em feridas de
sobreviventes do tsunami que ocorreu no final de 2004 na Ásia (Garzoni et al., 2005;
Maegele et al., 2005). Osteomielite causada por A. baumannii também é outra
infecção frequente em combatentes de guerra (Davis et al., 2005; Murray et al., 2006;
Johnson et al., 2007; Petersen et al., 2007; Schafer & Mangino, 2008).
Outro quadro infeccioso causado por A. baumannii são as meningites. Nos
últimos anos tem-se observado um aumento dos casos de meningite causada por
esse patógeno, principalmente associada a pacientes submetidos a neurocirurgias
(Siegman-Igra et al., 1993; Katragkou et al., 2006; Cascio et al., 2010). Rodríguez
Guardado e colaboradores (2008) identificaram 51 casos de meningite pós-
neurocirúrgica causadas por A.baumannii em dois hospitais universitários, entre 1990
a 2004, que apresentaram uma elavada taxa de mortalidade (33%). Esses casos
representaram 10,9% de todos os casos de meningites identificados nos dois hospitais
durante o período de estudo. Em um estudo similar, Metan e colaboradores (2007)
avaliaram 28 casos de meningite causada por A. baumannii e observaram uma
mortalidade de 71%. Embora pouco frequentes, casos de meningite por A. baumannii
adquirida na comunidade também já foram relatados (Chang et al., 2000; Taziarova et
al., 2007; Lowman et al., 2008; Ozaki et al., 2009). Outros quadros infecciosos
causados por A. baumannii, ainda que raros, como endocardite associada à prótese
valvar (Olut & Erkek, 2005) e ceratite em paciente submetido à cirurgia ocular (Kau et
al., 2002) também foram relatados.
A habilidade de sobreviver em ambientes desfavoráveis, observada em
isolados clínicos de A. baumannii constitui sua principal característica em relação a
outros Gram negativos e deve-se, em parte, a sua capacidade em formar biofilmes,
resistindo à dissecação (Donlan & Costerton, 2002; Gaddy & Actis, 2009). A

REVISÃO BIBLIOGRÁFICA
10
dificuldade em erradicar A. baumannii do ambiente hospitalar, torna os surtos
causados por esse patógeno de difícil controle (Wilks et al., 2006).
2.3. Opções de Tratamento para Infecções Causadas por A. baumannii
Diante das características apresentadas, A. baumannii é uma bactéria de difícil
tratamento já que poucos são os antimicrobianos disponíveis clinicamente que
apresentam atividade contra esses micro-organismos na atualidade. Infelizmente,
algumas cepas apresentam resistência a todos os antimicrobianos que serão aqui
abordados.
2.3.1. Ampicilina/Sulbactam
Sulbactam é um inibidor de β-lactamase que possui atividade contra
Acinetobacter spp. (Brauers et al., 2005), pois se liga à proteína ligadora de penicilina
2 (PBP2), uma das enzimas responsáveis pele síntese de peptideoglicano da parede
celular bacteriana (Cayô et al., 2011b). No Brasil, sulbactam é comercializado
juntamente com a ampicilina, porém, apenas o sulbactam apresenta atividade contra
isolados de A. baumannii (Corbella et al., 1998; Brauers et al., 2005).
O tratamento com ampicilina/sulbactam vem sendo descrito como uma
alternativa eficaz em infecções causadas por Acinetobacter spp. (Jiménez-Mejías et
al., 1997, Tuon et al., 2010). Mesmo nos casos de infecções causadas por
Acinetobacter spp. resistentes aos carbapenens, o uso da ampicilina/sulbactam tem
sido preferido ao das polimixinas (Oliveira et al., 2008). Embora a monoterapia não
seja recomendada para os casos de infecções graves (Maragakis & Perl, 2008), a
associação de ampicilina/sulbactam com outros antimicrobianos, como amicacina,
rifampicina ou colistina tem apresentado resultados promissores (Appleman et al.,
2000; Savov et al., 2002; Ko et al., 2004, Kiffer et al., 2005; Tong et al., 2006; Song et
al., 2007; Peleg, 2007c; Shrivastava et al., 2009). O uso de ampicilina/sulbactam é
indicado e eficaz no tratamento de meningites, pneumonia associada à ventilação

REVISÃO BIBLIOGRÁFICA
11
mecânica e bacteremia associada ao uso de cateter (Jiménez-Mejías et al., 1997;
Corbella et al., 1998; Wood et al., 2002, Oliveira et al., 2008).
As principais vantagens desse antimicrobiano são o custo reduzido e a sua boa
tolerabilidade (Jellison et al., 2001; Oliveira et al., 2008). Apesar disso, alguns estudos
tem mostrado um constante aumento na resistência a esse antimicrobiano, como
apresentado no trabalho de Yang e colaboradores (2010), em Taiwan, onde a taxa de
resistência observada foi em torno de 70%. Já Wieczorek e colaboradores (2012)
apresentaram um estudo de resistência in vitro, onde a CIM para sulbactam em
isolados de A. baumannii passou de 0,5 µg/mL para 4 µg/mL, quando os isolados
foram induzidos a 0,9x o valor da CIM inicial. Mesmo não sendo mais expostos ao
sulbactam, a CIM dos isolados de A. baumannii não retornaram ao seu valor original,
sugerindo que o uso do sulbactam poderia favorecer o surgimento de cepas
resistentes, para as quais o tratamento com esse antimicrobiano, mesmo em altas
doses, poderia não ser efetivo.
2.3.2. Minociclina
A minociclina é um derivado da tetraciclina que foi introduzida na década de 60
(Neonakis et al., 2011) e que voltou a ser utilizada pela redução de opções
terapêuticas eficazes e pelo surgimento de cepas de A. baumannii MDR. Seu
mecanismo de ação consiste em completar os ribossomos, impedindo a ligação do
tRNA e, consequentemente, inibir a transcrição da síntese proteica (Chopra et al.,
1992). A minociclina apresenta concentrações sanguíneas e teciduais ideais, boa
penetração no sistema nervoso central e uma menor taxa de resistência comparada à
tetraciclina. Ela apresenta atividade contra micro-organismos Gram positivos e Gram
negativos (Bishburg & Bishburg, 2009), incluindo isolados de A. baumannii (Pei et al.,
2012; Fernández-Cuenca et al., 2013).
Embora poucos estudos descrevam o uso da minociclina, os resultados
demonstram que esse antimicrobiano pode ser uma boa opção terapêutica no

REVISÃO BIBLIOGRÁFICA
12
tratamento de isolados de A. baumannii MDR. Griffith e colaboradores (2008)
relataram o sucesso do tratamento com minociclina em sete de oito casos de ferida
traumática, ocasionada por isolados MDR pertencentes ao complexo A. calcoaceticus-
baumannii. Já Wood e colaboradores (2003), descreveram o tratamento efetivo em
pacientes com pneumonia. Entretanto, algumas cepas de A. baumannii apresentam
resistência a esse antimicrobiano que, aparentemente, possuem dois mecanismos de
resistência: o primeiro seria decorrente dos genes moduladores tetB e otr, específicos
de sistemas de efluxo, responsáveis por fazer a extrusão da tetraciclina e da
minociclina, mas não da tigeciclina (Chopra et al., 1992; Huys et al., 2005a); e o
segundo é a síntese da proteína Tet(M) e Tet(O), que se ligam ao ribossomo,
modificando sua conformação e impedindo a ligação da minociclina, da tetraciclina e
da doxiciclina (Chopra & Roberts, 2001; Ribera et al., 2003).
2.3.3. Tigeciclina
A tigeciclina é um antimicrobiano semissintético da família das glicilciclinas, que
foi sintetizado a partir de modificação na molécula de minociclina. Seu mecanismo de
ação consiste em inibir a síntese proteica da bactéria, ligando-se a subunidade 30S do
ribossomo. Possui atividade contra micro-organismos Gram negativos, incluindo os
isolados produtores de carbapenemases, Gram positivos [incluindo isolados de
Sthaphylococcus aureus resistentes à oxacilina (ORSA) e Enterococcus spp.
resistentes à vancomicina (VRE)] e anaeróbios (Bergeron et al., 1996).
Para este antimicrobiano, não existem critérios de sensibilidade estabelecidos
pelo “Clinical and Laboratory Standards Institute” (CLSI, 2012), “British Society for
Antimicrobial Chemotherapy” (BSAC, 2010) e “The European Committee on
Antimicrobial Susceptibility Testing” (EUCAST, 2012), para categorizar os isolados de
Acinetobacter spp. (Neonakis et al., 2011). Entretanto, pelo seu valor de CIM baixo, a
tigeciclina atinge baixas concentrações séricas e, as poucas opções de tratamento

REVISÃO BIBLIOGRÁFICA
13
observadas para isolados de A. baumannii MDR, fazem da tigeciclina um
antimicrobiano oportuno (Peleg et al., 2007a; Neonakis et al., 2011).
Vários estudos foram realizados como intuito de avaliar a eficácia clínica da
tigeciclina em infecções causadas por A. baumannii, já que esse antimicrobiano possui
boa atividade in vitro contra esses isolados (Neonakis et al., 2011). Poulakou e
colaboradores (2009) relataram que, de 15 pacientes infectados com A. baumannii
MDR, 11 obtiveram melhora ou cura com a monoterapia por tigeciclina. Esse resultado
se contrapõe ao estudo de Gallagher & Rouse (2008), no qual os autores relataram
que, dos 28 pacientes com infecção causada por A. baumannii MDR avaliados,
apenas oito obtiveram melhoria ou cura com a monoterapia com tigeciclina. Da mesma
forma, Schafer e colaboradores (2007) retrataram um caso de pneumonia e
bacteremia causado por um isolado de A. baumannii MDR, que veio a desenvolver
resistência a tigeciclina durante o tratamento. Resultado semelhante foi observado por
Reid e colaboradores (2007), que verificaram o aumento da CIM de tigeciclina em um
isolado de A. baumannii ao longo do tratamento de infecção urinária pós-transplante
de rim e fígado. Por essas razões, os autores concluíram que pacientes acometidos
por infecções causadas por A. baumannii MDR, que fazem uso de tigeciclina, devem
ser monitorados quanto ao desenvolvimento de resistência durante o tratamento (Reid
et al., 2007).
A terapia combinada pode constituir uma alternativa para desfavorecer o
desenvolvimento de resistência à tigeciclina. Estudos tem relatado sinergismo das
combinações de tigeciclina com carbapenens, levofloxacino, amicacina ou rifampicina
(Entenza et al., 2009; Lim et al., 2009; Principe et al., 2009). Resultados controversos
da combinação da tigeciclina com a colistina tem sido reportados (Petersen et al.,
2006; Arroyo et al., 2009; Dizbay et al., 2010). Estudos prospectivos ainda são
necessários, incluindo um maior número de pacientes, para melhor definir a real
eficácia da tigeciclina nas infecções causadas por A. baumannii (Neonakis et al.,
2011).

REVISÃO BIBLIOGRÁFICA
14
2.3.4. Polimixinas
Polimixina B e polimixina E (também denominada colistina) são polipeptídeos
provenientes da bactéria Paenibacillus polymyxa que foram descobertos em 1947
(Girardello, 2012). Seu mecanismo de ação consiste em permeabilizar a parede
celular bacteriana, deslocando as moléculas de cálcio e magnésio, ligadas ao
lipopolissacarídeo bacteriano, o que desestabilizaria os componentes da membrana
celular externa e interna (Hancock, 1997).
Durante muito tempo, a polimixina teve o seu uso restrito devido à sua
toxicidade (Wolinsky & Hines, 1962; Koch-Weser et al., 1970; Gales et al., 2001;
Falagas et al., 2007). Entrentanto, estudos recentes relatam que as polimixinas não
são tão tóxicas como se imaginava (Conway et al., 1997; Kasiakou et al., 2005; Reina
et al., 2005; Falagas et al., 2007) e, devido as restritas opções terapêuticas,
atualmente, esses antimicrobianos são empregados com uma frequência cada vez
maior no tratamento das infecções causadas por bactérias Gram negativas MDR.
As polimixinas apresentam diversas formas de administração, tópica, inalatória
ou parenteral (Karageorgopoulos & Falagas, 2008; Landman et al., 2008). Elas são
utilizadas para tratar infecções de corrente sanguínea, feridas, infecção do trato
urinário (Gounden et al., 2009), pneumonias e meningites causadas por micro-
organismos Gram negativos MDR (Karageorgopoulos & Falagas, 2008). Entretanto, o
relato do fenômeno de heterorresistência que ocorre quando subpopulações
geneticamente idênticas são mais resistentes que o clone original é preocupante
(Perez et al., 2007), já tendo sido reportado em isolados de A. baumannii (Li et al.,
2006). No estudo de Li e colaboradores (2006) foi observado que de 16 isolados, 15
apresentavam heterorresistência, indicando que esse fenômeno pode ser frequente.
Felizmente ainda são raros os relatos de resistência às polimixinas (Gales et
al., 2001; Gales et al., 2006) e muitos estudos tem demonstrado sucesso no
tratamento de infecções causadas por A. baumannii utilizando antimicrobianos dessa
classe (Levin et al., 1999; Michalopoulos et al., 2005b; Holloway et al., 2006; Kallel et

REVISÃO BIBLIOGRÁFICA
15
al., 2007; Falagas et al., 2010). Nos casos de isolados resistentes aos carbapenens, o
tratamento de primeira escolha é aquele que emprega as polimixinas (Garnacho-
Montero et al., 2003; Michalopoulos et al., 2005a; Matthaiou et al., 2008).
2.3.5. Carbapenens
Em 1967, com o surgimento de isolados produtores de β-lactamases e a
ineficiência da penicilina contra esses agentes, a indústria Beecham Pharmaceuticals
(London, UK) começou um estudo para encontrar um inibidor de β-lactamase
proveniente de fonte natural (Basker, 1982). Apenas em 1976, foi isolado em várias
culturas de Streptomyces olivaceus, o ácido olivânico, que não foi considerado apenas
um inibidor de β-lactamase, mas também um antimicrobiano com um espectro mais
amplo do que outros β-lactâmicos disponíveis naquela época. Devido a sua pouca
penetração na célula bacteriana e a sua instabilidade química, esse composto teve o
seu desenvolvimento interrompido (Brown et al, 1976; Butterworth et al., 1979; Hood et
al., 1979). Em paralelo, pesquisadores da Merck, Sharp and Dohme Research
Laboratories (Harlow, UK), se voltavam para a descoberta de antimicrobianos que
bloqueavam a síntese de peptideoglicano e analisavam um novo antimicrobiano obtido
de culturas de Streptomyces cattleya que foi, posteriormente, nomeado como
tienamicina. Coincidentemente, esse composto possuía o mesmo núcleo
carbapenêmico do ácido olivânico, mas se diferenciava quanto à estrutura química
(Albers-Schonberg et al., 1978; Kahan et al., 1979). Anos mais tarde, foi desenvolvido
o imipenem, cuja molécula precursora foi a tienamicina, que teve o início da sua
utilização clínica em 1985 (Miyadera et al., 1983; Papp-Wallace et al., 2011). Desde
então, novas moléculas foram sendo desenvolvidas e aprimoradas e, atualmente,
existem seis carbapenens para uso clínico que se diferem pela sua estrutura
molecular, como apresentado na Figura 1. No Brasil, somente o ertapenem, o
imipenem e o meropenem estão disponíveis para uso clínico.
REVISÃO BIBLIOGRÁFICA
16
1 2
3 4 5
6 7 8
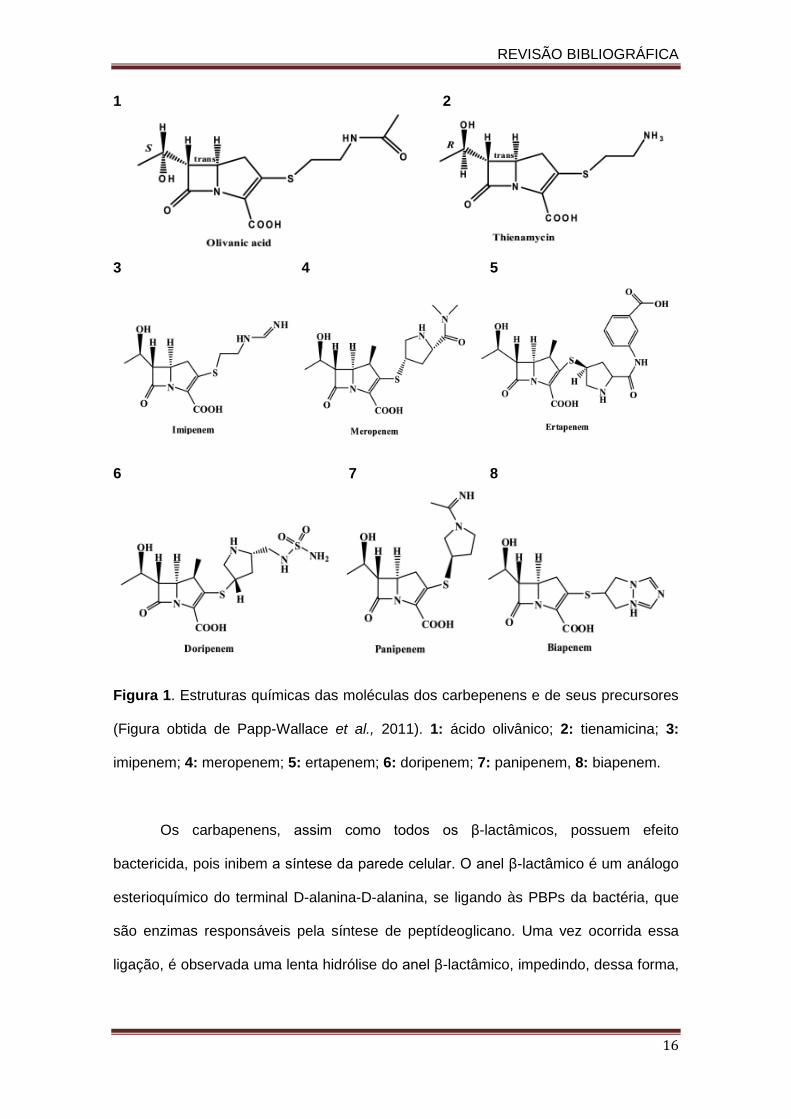
Figura 1. Estruturas químicas das moléculas dos carbepenens e de seus precursores
(Figura obtida de Papp-Wallace et al., 2011). 1: ácido olivânico; 2: tienamicina; 3:
imipenem; 4: meropenem; 5: ertapenem; 6: doripenem; 7: panipenem, 8: biapenem.
Os carbapenens, assim como todos os β-lactâmicos, possuem efeito
bactericida, pois inibem a síntese da parede celular. O anel β-lactâmico é um análogo
esterioquímico do terminal D-alanina-D-alanina, se ligando às PBPs da bactéria, que
são enzimas responsáveis pela síntese de peptídeoglicano. Uma vez ocorrida essa
ligação, é observada uma lenta hidrólise do anel β-lactâmico, impedindo, dessa forma,

REVISÃO BIBLIOGRÁFICA
17
que as PBPs sintetizem o peptideoglicano, principal composto da parede celular
(Zapun et al., 2008; Drawz & Bomono, 2010).
Os carbapenens são os antimicrobianos utilizados como primeira opção
terapêutica nas infecções causadas por A. baumannii, por possuirem um amplo
espectro de ação, que engloba tanto bactérias Gram negativas como Gram positivas e
por apresentarem boa tolerabilidade e eficácia (Papp-Wallace et al., 2011). Os
carbapenens mais utilizados no Brasil são o imipenem, meropenem e ertapenem e
algumas observações a respeitos desses compostos devem ser consideradas: (i) o
imipenem, primeiro carbapenem descrito, é degradado pela enzima dehidropeptidase-
1 (DHP-1) e, por isso, é administrado juntamente com a cilastatina, um inidor dessa
enzima (Zhanel et al., 2007); (ii) o meropenem não é tão potente quanto o imipenem
na ação contra A. baumannii, mas não é uma regra geral, pois varia de acordo com a
cepa (Oliver et al., 2004); e (iii) o ertapenem tem um espectro restrito e não possui
atividade contra isolados de P. aeruginosa e Acinetobacter spp. (Oliver et al., 2004).
Altas taxas de resistência aos carbapenens verificadas entre os isolados de A.
baumannii tem sido descritas em todo o mundo (Poirel & Nordmann, 2006; Peleg et
al., 2008). Os principais mecanismos de resistência aos carbapenens descritos nesse
micro-organismo são: (i) a produção de carbapenemases do tipo OXA, principalmente
a OXA-23 associada ao elemento de inserção ISAba1, OXA-58 associada aos
elementos de inserção ISAba2, ISAba3 e ISAba18 e OXA-24/40 (Poirel & Nordmann,
2006; Turton et al., 2006); (ii) perda ou redução da porina CarO (29kDa) (Mussi et al.,
2005); (iii) redução da expressão da PBP2 (Fernández-Cuenca et al., 2003a) e (iv) a
hiperexpressão do sistema de efluxo AdeABC (Marqué et al., 2005). Todos esses
mecanismos podem estar presentes no mesmo isolado, uma característica muito
comum em cepas MDR de A. baumannii, que será discutida posteriormente.
Na tentativa de combater infecções mais graves, a terapia combinada surge
como uma importante alternativa. Entretanto, o surgimento de micro-organismos
resistentes é um risco que deve ser levado em consideração. Para tratar isolados de

REVISÃO BIBLIOGRÁFICA
18
A. baumannii MDR, várias opções de terapia combinada tem sido propostas
(Rodríguez-Hernández et al., 2000; Ermertcan et al., 2001; Fernández-Cuenca et al.,
2003b; Wareham et al., 2006; Kattan et al., 2008; Song et al., 2009), mas nenhuma
mostrou-se ser totalmente eficaz.
2.4. Mecanismos de Resistência aos Carbapenens em A. baumannii
Como descrito anteriormente, A. baumannii é um micro-organismo peculiar por
apresentar características importantes de sobrevivência em ambientes hostis e por
apresentar grande capacidade em adquirir diversos mecanismos de resistência,
muitas vezes associados. Serão apresentados a seguir os quatro mecanismos
descritos, até o momento, que conferem resistência aos β-lactâmicos em isolados
clínicos de A. baumannii.
2.4.1. Produção de β-Lactamases
As β-lactameses são enzimas cromossomais ou plasmidiais que hidrolisam o
anel β-lactâmico (Bush et al., 1995). Essas enzimas são produzidas por bactérias
Gram positivas que as secretam para o meio extracelular e por bactérias Gram
negativas, que as armazenam em seu espaço periplasmático sendo, portanto,
estrategicamente mais eficientes (Livermore, 1993). A capacidade dessas enzimas de
conferirem mais resistência aos β-lactâmicos ou não, depende de sua localização, da
sua cinética, das condições físico-químicas (Livermore, 1995), da quantidade de
enzima produzida, da habilidade em hidrolisar o anel β-lactâmico e da velocidade com
que o antimicrobiano penetra na célula bacteriana (Bush et al.,1995).
Devido a grande diversidade bioquímica e molecular das β-lactamases
encontradas, em 1980, Ambler propôs classificá-las em quatro classes de acordo com
suas sequências de aminoácidos, sendo elas: classe A - serino β-lactamases,
incluindo as β-Lactamases de Espectro Ampliado (ESβL), penicilinases e
carbenicilinases; classe B - Metalo-β-Lactamases (MβL); classe C - cefalosporinases

REVISÃO BIBLIOGRÁFICA
19
cromossomais e plasmidiais do tipo AmpC; e classe D - oxacilinases (Ambler, 1980).
Posteriormente, Bush e colaboradores (1995) atualizaram a classificação de Bush
(1989a,b,c) que por sua vez, foi uma atualização de Sykes (1982). A classe molecular
de Ambler foi incorporada a essa nova classificação que é baseada nas características
da enzima, substratos específicos e perfil de inibição por inibidores da β-lactamase. A
classificação ocorrida no ano de 1995 foi revisada para a inclusão de novas β-
lactamases não descritas anteriormente (Bush & Jacoby, 2010). Cada uma das
classes será detalhada a seguir.
2.4.1.1. β-Lactamases de Espectro Ampliado (ESβLs)
As ESβLs pertencem à classe A de Ambler (Ambler, 1980) e ao grupo 2be de
Bush (Bush & Jacoby, 2010). Essas enzimas possuem em seu sítio ativo, um éster de
serina que hidrolisa penicilinas, cefalosporinas de amplo espectro e aztreonam e são
inibidas pelos inibidores de β-lactamases, como o ácido clavulânico, o sulbactam e o
tazobactam (Bush & Jacoby, 2010). Entretanto, como os genes que codificam as
ESβLs geralmente estão localizados em plasmídeos, muitas vezes, os isolados
produtores dessas enzimas apresentam resistência cruzada à outras classes de
antimicrobianos, como os aminoglicosídeos, a tetraciclina, o cloranfenicol e a
sulfametoxazol/trimetoprim. Isso acontece, porque os genes que conferem resistência
a esses compostos também são carreados pelos mesmos plasmídeos que carreiam os
genes codificadores de ESβLs (Winokur et al., 2001; Turner, 2005).
Embora as ESβLs sejam encontradas em abundância nas enterobactérias,
apenas CTX-M-2, CTX-M-15, CTX-M-43, PER-1, PER-2, PER-7, TEM-92, TEM-150,
SHV-5, e VEB-1, já foram descritas em A. baumannii (Vahaboglu et al., 1997; Poirel et
al., 2003; Nagano et al., 2004; Celenza et al., 2006; Pasterán et al., 2006; Endimiani et
al., 2007; Naas et al., 2007; Shakil et al., 2010; Bonnin et al., 2011a, Bonnin et al.,
2011b).

REVISÃO BIBLIOGRÁFICA
20
Outro subgrupo da classe A é o 2f, onde estão inseridas as β-lactamases que
hidrolisam os carbapenens. Essas enzimas possuem resíduos de serina em seu sítio
ativo e são sensíveis aos inibidores das serino-β-lactamases, como o ácido clavulânico
e o tazobactam (Bush & Jacoby, 2010). As enzimas do tipo GES e KPC são as
principais representantes desse grupo. As carbapenemases do tipo GES raramente
são descrita em Acinetobacter spp., e até o momento, GES-11, GES-12 e GES-14
foram relatadas nesse micro-organismo em áreas geográficas específicas (Moubareck
et al., 2009; Bogaerts et al., 2010; Bonnin et al., 2011a). Já as carbapenemases do
tipo KPC, comumente encontrada em isolados de K. pneumoniae, são mundialmente
disseminadas e causadoras de inúmeros surtos, inclusive no Brasil (Hirsch & Tam,
2010). Entretanto, os representantes desse subgrupo foram raramente descritos em
cepas de A. baumannii, sendo relatados KPC-2, KPC-3, KPC-4 e KPC-10 (Robledo et
al., 2010).
2.4.1.2. Metalo-β-Lactamases (MβL)
Na classe B de Ambler estão inseridas as MβL que utilizam íons Zn2++ como
cofator e, por isso, são inibidas por agentes quelantes como o EDTA (Walsh et al.,
2005). As MβL hidrolisam todos os β-lactâmicos, exceto o aztreonam (Walsh et al.,
2005). De acordo com a classificação de Bush (Bush & Jacoby, 2010), existem três
subgrupos, sendo o subgrupo B1 o de maior importância clínica, composto por
enzimas adquiridas que já foram descritas em isolados de A. baumannii (Queenan &
Bush, 2007; Cornaglia et al., 2011). Essas enzimas estão, em sua grande maioria,
localizadas em elementos genéticos móveis denominados integrons de classe 1, onde
estão inseridos os genes cassetes que as codificam. Por sua vez, os integrons podem
estar inseridos tanto no plasmídeo como no cromossomo bacteriano (Walsh et al.,
2005).
Em seu artigo de revisão, Cornaglia e colaboradores (2011) relataram que já
foram descritas, em cepas de A. baumannii isoladas em diferentes países, as

REVISÃO BIBLIOGRÁFICA
21
seguintes MβLs: IMP-1, IMP-2, IMP-4, IMP-5, IMP-8, IMP-10, VIM-1, VIM-2, VIM-3,
VIM-4, VIM-11, SIM-1 e NDM-1. Recentemente, foram descritas a NDM-2 nos
Emirádos Árabes (Ghazawi et al., 2012) e a GIM-1 na Coréia (Hong et al., 2012).
2.4.1.3. Cefalosporinase do Tipo AmpC (ADCs)
As enzimas do tipo AmpC pertencem a classe C de Ambler (Ambler, 1980) e ao
grupo 1 de Bush (Bush & Jacoby, 2010). Essas enzimas hidrolisam com eficiência as
penicilinas e as cefamicinas e, com menor eficiência, o aztreonam e as cefalosporinas
de amplo espectro. Entretanto, os carbapenens e as cefalosporinas de quarta geração
não são, geralmente, hidrolisados por essa enzima (Ni et al., 2005; Jacoby, 2009). O
gene que codifica a β-lactamase do tipo AmpC pode estar localizado tanto no
cromossomo quanto no plasmídeo (Philippon et al., 2002; Jacoby, 2009). As enzimas
AmpCs cromossomais estão presentes em diversas bactérias, mas, principalmente,
naquelas incluídas no grupo CESP (Citrobacter freundii, Enterobacter spp., Serratia
marcescens, Providencia stuartii, P. aeruginosa e Morganella morganii), onde tem a
sua produção induzida na presença de β-lactâmicos (Bush et al., 1995).
Para induzir a expressão da AmpC é necessário que os três genes reguladores
(ampD, ampG e ampR) estejam presentes no genoma bacteriano (Kuga et al., 2000).
Antimicrobianos como o imipenem e a cefoxitina são considerados fortes indutores da
produção dessa enzima (Jones, 1998). Em A. baumannii, essa enzima também é
cromossomal e intrínseca, porém, a indução não ocorre de modo efetivo devido à
ausência do gene ampR. Dessa foram, a AmpC é somente expressa em níveis basais,
não hidrolisando as cefalosporinas de amplo espectro em A. baumannii (Bou &
Martínez-Beltrán, 2000). Entretanto, quando o elemento de inserção ISAba1 está
inserido a montante do gene blaAmpC, ocorre a hiperexpressão dessa enzima
conferindo resistência às cefalosporinas de amplo espectro (Boo et al., 2009; Lin et al.,
2010).

REVISÃO BIBLIOGRÁFICA
22
Rodríguez-Martínez e colaboradores (2010) denominaram uma nomenclatura
específica para a AmpC em Acinetobacter, chamada de ADC (Acinetobacter-Derived
Cephalosporinases), pois apresentam um único ancestral comum e se distanciam
muito das AmpCs descritas em outros micro-organismos (Hujer et al., 2005). A
primeira ADC descrita em A. baumannii foi a ADC-7, que hidrolisa fortemente
ceftazidima e cefotaxima, mas não hidrolisa carbapenens (Hujer et al., 2005), sendo a
última descrita, a ADC-68 (KC866352.1). A hiperprodução de ADC associada à perda
de porina e/ou à hiperexpressão de sistemas de efluxo pode levar a diminuição da
sensibilidade aos carbapenens (Bradford et al., 1997; Jacoby, 2009).
2.4.1.4. Oxacilinases
As oxacilinases (OXAs) pertencem a classe D de Ambler (Ambler, 1980) e ao
grupo 2 de Bush (Bush & Jacoby, 2010). São chamadas assim por terem a capacidade
de hidrolisar a oxacilina e a cloxacilina e por hidrolisar, consideravelmente, as
aminopenicilinas e as carboxipenicilinas. Essas enzimas são fortemente inibidas, in
vitro, pelo cloreto de sódio (Poirel et al., 2010) e apresentam resíduo de serina em seu
sítio ativo como as enzimas das classes A e C de Ambler (Poirel et al., 2010).
Na subdivisão de Bush (Bush & Jacoby, 2010), as oxacilinases são
classificadas em três subgrupos, sendo eles: 2d - OXAs de espectro limitado, capazes
de hidrolisar somente a oxacilina e a cloxacilina, comumente encontradas em
enterobactérias (Bush & Jacoby, 2010); 2de - OXAs de espectro ampliado, capazes de
hidrolisar oxiaminocefalosporinas, cefepima e cefotaxima, comumente encontradas em
P. aeruginosa (Aubert et al., 2001; Bush & Jacoby, 2010); e 2df - carbapenemases do
tipo OXA ou CHDLs, frequentemente encontradas em A. baumannii. As CHDLs
hidrolisam fracamente os carbapenens e os genes que as codificam podem estar
localizados tanto no cromossomo como em plasmídeos (Bush et al., 1995; Walther-
Rasmussen & Høiby, 2006; Bush & Jacoby, 2010). Por apresentarem a capacidade de
hidrolisar os carbapenens, constituem o grupo de oxacilinases de maior importância.

REVISÃO BIBLIOGRÁFICA
23
Já foram descritos sete subtipos de CHDLs em todo o mundo, sendo elas:
OXA-23, OXA-24/40, OXA-51, OXA-58, OXA-143, OXA-182 e OXA-235 (Brown &
Amyes, 2006; Walther-Rasmussen & Høiby, 2006; Higgins et al., 2009; Kim et al.,
2010; Poirel et al., 2010, Higgins et al., 2013).
2.4.1.4.1. OXA-23
A OXA-23 foi inicialmente descrita em 1985 na cidade de Edimburgo - Escócia,
em um isolado de A. baumannii resistente ao imipenem, proveniente de hemocultura
(Paton et al., 1993). Inicialmente, foi denomindada ARI-1 “Acinetobacter Resistant to
Imipenem” e, posteriormente, por possuir similaridade menor que 40% com as outras
enzimas pertencentes à classe D de Ambler (Ambler, 1980), passou a ser denominada
OXA-23 (Donald et al., 2000), sendo o primeiro grupo de CHDL descrita.
Em um estudo conduzido por Poirel e colaboradores (2008) foi identificado o
gene blaOXA-23 no cromossomo de isolados sensíveis aos carbapenens de
Acinetobacter radioresistans, considerada uma espécie comensal presente na pele de
pacientes internados. Os autores concluíram que A. radioresistens constitui o
reservatório natural do gene que codifica a OXA-23, o qual foi, posteriormente,
transferido para A. baumannii. O gene codificador dessa enzima foi disseminado
através da sequência de inserção ISAba1 mediado por plasmídeo proveniente de um
A. baumannii. O plasmídeo teria se inserido no cromossomo de A. radioresistans, se
transposto, mobilizando as regiões a montante e a jusante do gene blaOXA-23 no
cromossomo de A. radioresistens. A estrutura gerada formou assim uma estrutura
semelhante a um transposon e, ao mesmo tempo, conferia expressão aumentada ou
hiperexpressão da OXA-23. O plasmídeo carreando o gene blaOXA-23 foi, então,
transferido para um isolado de A. baumannii e, dessa forma, disseminado para outros
isolados.
A OXA-23 é uma carbapenemase mundialmente disseminada, presente em
mais de 50 países. Por essa razão é a oxacilinase mais estudada em Acinetobacter

REVISÃO BIBLIOGRÁFICA
24
spp., principalmente em A. baumannii, salvo a exceção de um caso de OXA-23
descrito em Proteus mirabilis, proveniente de um surto ocorrido na França (Bonnet et
al., 2002). A Figura 2 mostra a distribuição de OXA-23 em isolados de A. baumannii
no mundo.
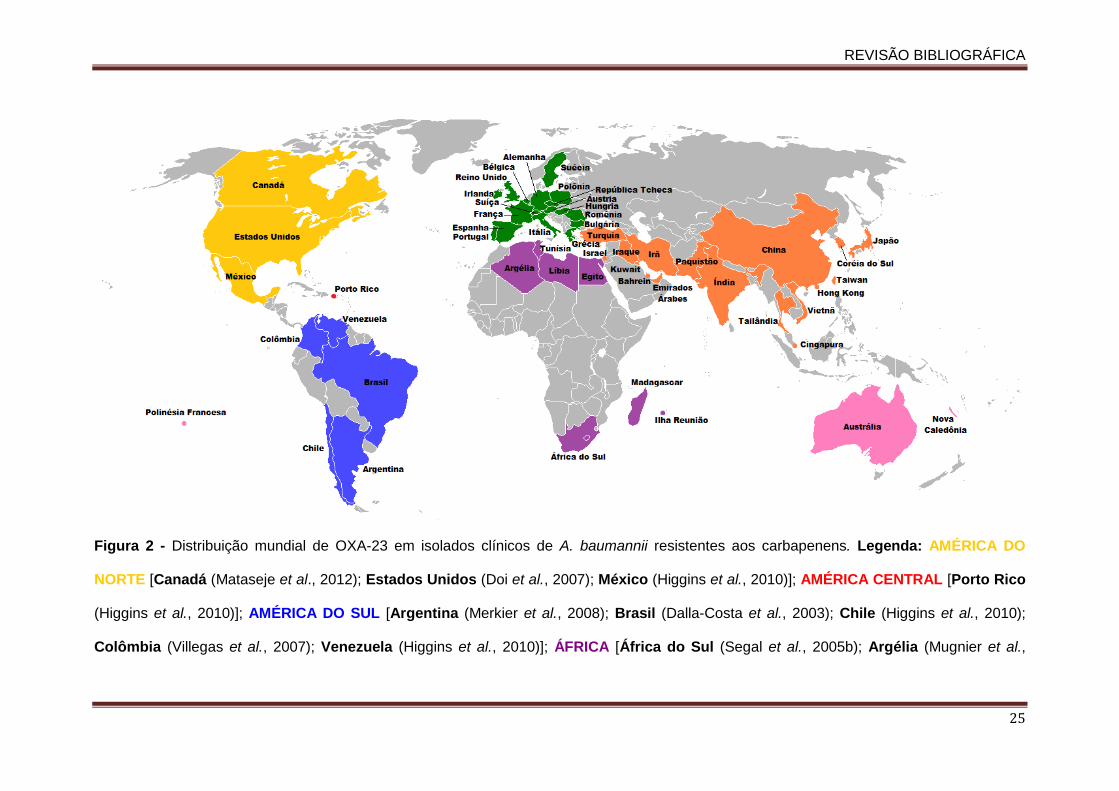
REVISÃO BIBLIOGRÁFICA
25
Figura 2 - Distribuição mundial de OXA-23 em isolados clínicos de A. baumannii resistentes aos carbapenens. Legenda: AMÉRICA DO
NORTE [Canadá (Mataseje et al., 2012); Estados Unidos (Doi et al., 2007); México (Higgins et al., 2010)]; AMÉRICA CENTRAL [Porto Rico
(Higgins et al., 2010)]; AMÉRICA DO SUL [Argentina (Merkier et al., 2008); Brasil (Dalla-Costa et al., 2003); Chile (Higgins et al., 2010);
Colômbia (Villegas et al., 2007); Venezuela (Higgins et al., 2010)]; ÁFRICA [África do Sul (Segal et al., 2005b); Argélia (Mugnier et al.,

REVISÃO BIBLIOGRÁFICA
26
2010); Egito (Mugnier et al., 2010); Ilha de Reunião (Mugnier et al., 2010); Líbia (Mugnier et al., 2010); Madagascar (Andriamanantena et al.,
2010); Tunísia (Hammami et al,. 2011)]; EUROPA [Alemanha (Kohlenberg et al., 2009); Áustria (Higgins et al., 2010); Bélgica (Bogaerts et
al., 2008); Bulgária (Stoeva et al., 2008); Escócia (Paton et al., 1993); Espanha (Higgins et al., 2010); França (Héritier et al., 2005a); Grécia
(Higgins et al., 2010); Hungria (Higgins et al., 2010); Inglaterra (Turton et al., 2005); Irlanda (Higgins et al., 2010); Itália (Mendes et al.,
2009b); Polônia (Higgins et al., 2010); Portugal (Higgins et al., 2010); Rep. Tcheca (Krizova et al., 2012); Romênia (Bonnin et al., 2011c);
Suécia (Karah et al., 2011); Suíça (Higgins et al., 2010)]; ÁSIA [Bahrein (Mugnier et al., 2010); China (Wang et al., 2003); Coréia (Jeon et al.,
2005); Emirados Árabes Unidos (Mugnier et al., 2008); Hong Kong (Mendes et al., 2009a); Índia (Mendes et al., 2009a); Irã (Feizabadi et al.,
2008); Iraque (Kusradze et al., 2011); Israel (Higgins et al., 2010); Japão (Kouyama et al., 2012); Kuwait (Al-Sweih et al., 2012); Paquistão
(Higgins et al., 2010); Cingapura (Afzal-Shah et al., 2001); Tailândia (Mendes et al., 2009a); Taiwan (Lu et al., 2009); Turquia (Meric et al.,
2008); Vietnã (Mugnier et al., 2010)]; OCEANIA [Austrália (Higgins et al., 2010); Nova Caledônia (Le Hello et al., 2008); Polinésia Francesa
(Naas et al., 2005)].

REVISÃO BIBLIOGRÁFICA
27
Vários estudos têm relatado variantes da OXA-23, incluindo uma variante
descrita em K. pneumoniae na Austrália. Até o momento, foram relatadas 13 variantes
de OXA-23 (Tabela 1), concentradas principalmente no sudeste da Ásia e, dessas,
mais de 50% foram descritas na Tailândia, como pode ser observado na Figura 3.
Tabela 1 - Variantes de OXA-23 descritas de acordo com a espécie de isolamento,
ano e país de origem.
Variante da OXA-23 Espécie Ano País Referência
OXA-27 A. baumannii 2001 Cingapura Afzal-Shah et al., 2001
OXA-49 A. baumannii 2003 China AY288523
OXA-73 K. pneumoniae 2006 Austrália AY762325
OXA-133 A. radioresistans 2009 Índia Mendes et al., 2009a
OXA-146 A. baumannii 2009 China FJ194494
OXA-165 A. baumannii 2010 Tailândia HM488986
OXA-166 A. baumannii 2010 Tailândia HM488987
OXA-167 A. baumannii 2010 Tailândia HM488988
OXA-168 A. baumannii 2010 Tailândia HM488989
OXA-169 A. baumannii 2010 Tailândia HM488990
OXA-170 A. baumannii 2010 Tailândia HM488991
OXA-171 A. baumannii 2010 Tailândia HM488992
OXA-225 A. baumannii 2012 Alemanha JN638887

REVISÃO BIBLIOGRÁFICA
28
Figura 3. Distribuição mundial das variantes de OXA-23 descritas em Acinetobacter spp. e em K. pneumoniae.

REVISÃO BIBLIOGRÁFICA
29
A OXA-23, assim como a maioria das CHDLs, hidrolisam fracamente os
carbapenens e, para que o gene que a codifica seja hiperexpresso, é necessária a
presença de um elemento de inserção a montante do gene, geralmente a ISAba1
(Poirel & Nordmann, 2006). No estudo de Héritier e colaboradores (2005a) foi
analisada a eficiência dos três principais grupos de CHDLs, OXA-23, OXA-24/40 e
OXA-58 frente aos carbapenens. Os autores observaram que quando a CHDL possuia
um elemento de inserção a montante do gene (ISAba1 para OXA-23 e ISAba3 para
OXA-58), o valor da CIM para os carbapenens aumentava significativamente. Cepas
transformantes que possuiam o plasmídeo pFER ou pMAD, naturalmente produtores
de OXA-23 ou OXA-58, foram mais resistentes aos carbapenens que os
transformantes com plasmídeos recombinates pOXA-23 e pOXA-58.
2.4.1.4.2. OXA-24/40
A segunda CHDL descrita em A. baumannii foi a OXA-24, identificada por Bou
e colaboradores (2000a) na Espanha, em 1997. Sua sequência é idêntica à OXA-40
(Lopez-Otsoa et al., 2002) e, por muitos autores, é denominada OXA-24/40. É a
terceira CHDL mais disseminada no mundo e, atualmente, possui seis variantes (OXA-
25, OXA-26, OXA-72, OXA-139, OXA-160 e OXA-207), sendo a OXA-72 a única
variante descrita no Brasil (Werneck et al., 2011a). Uma das principais características
dessa enzima é que não está associada à sequência de inserção para aumentar a
expressão de seu gene, como verificado para as CHDLs OXA-23, OXA-51 e OXA-58.
Sendo assim, as carbapenemases do grupo OXA-24/40 são as CHDLs mais potentes
encontradas em A. baumannii (Poirel et al., 2010). O processo de mobilização dessas
enzimas ocorre por um processo único entre as CHDLs, por meio de recombinação
homóloga (Poirel et al., 2010). Além disso, a OXA-24/40 já foi descrita em P.
aeruginosa na Espanha (Sevillano et al., 2009).

REVISÃO BIBLIOGRÁFICA
30
2.4.1.4.3. OXA-51
A terceira CHDL descrita em A. baumannii foi identificada pela primeira vez em
dois isolados clínicos de A. baumannii em 1994 na Argentina (Brown et al., 2005). É
uma CHDL que hidrolisa fracamente os carbapenens e necessita de uma sequência
de inserção, geralmente ISAba1 ou ISAba9, a montante do seu gene para aumentar
sua eficiência na resistência aos carbapenens (Figueiredo et al., 2009). A OXA-51 é
naturalmente encontrada em isolados de A. baumannii (Héritier et al., 2005b) e, por
muito tempo, a presença desse gene em cepas de Acinetobacter spp. foi utilizada
como teste presuntivo para a identificação de A. baumannii. Entretanto, a presença do
gene blaOXA-51-like em um isolado de Acinetobacter spp. não é mais preditivo de A.
baumannii, uma vez que esse gene foi encontrado em um plasmídeo carreado por um
isolado clínico de A. nosocomialis (Lee et al., 2009). Atualmente, existem mais de 65
variantes de OXA-51distribuídas pelo mundo (http://www.lahey.org/Studies/).
2.4.1.4.4. OXA-58
A OXA-58 foi a quarta CHDL descrita em A. baumannii e é, também, a segunda
mais disseminada no mundo. Foi identificada pela primeira vez em 2003 na França
(Poirel et al., 2005) e possui atualmente três variantes, sendo elas a OXA-96, a OXA-
97 e a OXA-164 (http://www.lahey.org/Studies/). Assim como as variantes de OXA-51 e
de OXA-23, as CHDLs do grupo da OXA-58 necessitam de uma sequência de
inserção a montante do gene que a codifica para aumentar a sua expressão e assim,
conferir resistência aos carbapenens. A OXA-58 pode estar associada a diferentes
sequências de inserção como a ISAba1, ISAba2, ISAba3 e ISAba18 (Poirel &
Nordmann, 2006).
2.4.1.4.5. OXA-143, OXA-182 e OXA-235
Recentemente, três novos grupos de CHDLs foram descritas em A. baumannii.
A OXA-143, descrita em isolados de A. baumannii no Brasil em 2004, possui uma

REVISÃO BIBLIOGRÁFICA
31
identidade de 88% de sequência de aminoácidos com a OXA-24/40. Essa enzima
apresenta apenas uma variante descrita, a OXA-231, na cidade de Londrina, Paraná,
em 2012 (Gionco et al., 2012). Assim como a OXA-24/40, também não necessita de
uma sequência de inserção para aumentar a expressão de seu gene, embora o
contexto genético do gene blaOXA-143-like não seja o mesmo do gene blaOXA-24/40-like
(Higgins et al., 2009; Gionco et al., 2012). A OXA-182 foi identificada em 2007 na
Coréia do Sul e não apresenta nenhuma variante até o momento. Possui uma
identidade maior que 80% de sequências de aminoácidos com a OXA-24/40 e, assim
como essa, também não necessita de uma sequência de inserção para aumentar a
expressão de seu gene (Kim et al., 2010). A OXA-235 foi identificada nos Estados
Unidos e no México entre 2005 a 2009. Possui uma identidade de 85% de sequência
de aminoácidos com a OXA-134 (uma CHDL descrita em A. lwoffii) e apresenta duas
variantes, a OXA-236 e a OXA-237, ambas descritas nos Estados Unidos. Esse
mesmo estudo mostrou que o gene blaOXA-235 está alocado entre duas ISAba1, sendo
esse elemento de inserção necessário tanto para sua mobilização como para o
aumento da sua expressão (Higgins et al., 2013).
2.4.2. Impermeabilidade de Membrana Externa
Porinas são proteínas de membrana externa ou (“Outer Membrane Proteins” -
OMPs), que formam canais, permitindo o transporte de moléculas hidrofílicas de baixo
peso molecular para dentro e para fora da célula bacteriana (Vila et al., 2007).
Variações estruturais ou diminuição da expressão das porinas, em resposta a
presença de antimicrobianos, são estratégias de sobrevivência que muitas bactérias
desenvolveram. As porinas podem significar um importante fator no desenvolvimento
da resistência contra os antimicrobianos (Vila et al., 2007). Como descrito no trabalho
de Sato e Nakae (1991), a permeabilidade da membrana externa de A. baumannii é
duas a sete vezes menor que em P. aeruginosa. O pequeno número e tamanho das
porinas em A. baumannii poderia explicar a diminuição da permeabilidade da

REVISÃO BIBLIOGRÁFICA
32
membrana externa (menos que 5%), quando comparada a outros bacilos Gram
negativos. Esse fato poderia auxiliar na resistência instrínseca a diversos
antimicrobianos verificado neste micro-organismo (Obara & Nakae, 1991). Vários
estudos avaliando amostras de A. baumannii resistentes aos carbapenens,
observaram modificação na estrutura ou na perda de porinas (Limansky et al., 2002;
Dupont et al., 2005; Mussi et al., 2005; Tomás et at., 2005).
Através da análise da sequência de aminoácidos, foi possível detectar que as
estruturas das porinas em Gram negativos são semelhantes, apresentando ausência
de regiões de resíduos hidrofóbicos, carga total ligeiramente negativa, baixo índice de
instabilidade, conteúdo elevado de glicina e ausência de resíduos de cisteína (Vila et
al., 2007). As proteínas OmpHMP, OmpW, CarO, 33-36 kDa e OprD-like são OMPs
descritas em isolados de A. baumannii e que teriam um papel importante na
resistência aos carbapenens nesse micro-organismo. A OmpHMP (“Heat-Modifiable
Protein”) é uma proteína monomérica de 35,6 KDa e uma das OMPs mais importantes.
Possui homologia com outras proteínas monoméricas, como a OmpA em
Enterobactérias e a OprF em Pseudomonas spp. (Nitzan et al., 1999; Gribun et al.,
2003). Essa OMP também pode ser denominada OmpAAb em Acinetobacter spp.
(Sugawara & Nikaido, 2012).
A OmpW é uma proteína monomérica pequena de 22 kDa homóloga à OprG
da família Pseudomonadaceae (Siroy et al., 2006). Pode estar relacionada à
resistência a colistina em mutantes de A. baumannii (Vila et al., 2007) e à ceftriaxona
em amostras de Salmonella typhimurium (Hong et al., 2006). No intuito de se conhecer
sobre a contribuição da OmpW na resistência aos carbapenens, Tiwari e
colaboradores (2012) avaliaram a proteômica da OmpW em cepas resistentes de A.
baumannii em comparação a um controle sensível (ATCC 19606). Uma diminuição da
expressão da OmpW nas amostras resistentes aos carbapenens em relação a ATCC
foi detectada.

REVISÃO BIBLIOGRÁFICA
33
A CarO é uma porina de 29 kDa que, quando ausente em isolados de A.
baumannii não produtores de carbapenemase, pode estar associada a resistência ao
imipenem (Limansky et al., 2002). Por outro lado, o estudo publicado por Mussi e
colaboradores (2005) demonstrou que a inativação do gene carO por um elemento de
inserção, levava a perda dessa porina e, consequentemente, à resistência tanto ao
imipenem como ao meropenem. Foi verificado ainda que isoformas modificadas dessa
porina não transportavam eficientemente os carbapenens, em comparação a isoforma
selvagem (Vashist et al., 2010).
Outras duas porinas estão relacionadas à resistência aos carbapenens em
isolados de A. baumannii: a Omp33-36 kDa que na verdade possui 31 kDa (Tomás et
al., 2005) e uma porina de 43 kDa, homóloga à OprD (D2) de P. aeruginosa, chamada
de OprD-like (Dupont et al., 2005). Embora não tenha sido observado nenhum sítio de
ligação específico para os carbapenens na CarO (Siroy et al., 2005), a OprD-like seria
um canal específico para esses compostos (Vila et al., 2007).
2.4.3. Hiperexpressão de Sistemas de Efluxo
Os sistemas de efluxo estão presentes em todos os tipos de células,
protegendo-as dos efeitos tóxicos de substâncias químicas. A membrana externa das
bactérias Gram negativas age como uma barreira, limitando a entrada de agentes
nocivos à célula bacteriana. Os sistemas de efluxo expulsam tais agentes do interior
da célula (Vila et al., 2007). Até o momento, seis famílias de efluxo foram descritas por
Poole (2002), tanto em bactérias Gram positivas quanto em bactérias Gram negativas:
ABC “ATP Binding Cassette”, MFS “Major Facilitador Superfamily”, SMR “Small
Multidrug Resistance”, MATE “Multidrug and Toxic Compound Extrusion”, DMT “Drug-
Metabolite Transporter” e RND “Resistance-Nodulation Division”.
Os sitemas de efluxo do tipo ABC são transportadores de multidrogas
dependentes de ATP como fonte de energia para ejetar os antimicrobianos do interior

REVISÃO BIBLIOGRÁFICA
34
das células. Os membros dessa família raramente estão envolvidos com fenótipo de
resistência aos antimicrobianos em bactérias Gram negativas (Poole, 2002).
A família MFS transporta grupos específicos de antimicrobianos, como a
tetraciclina (tetA e tetB), a minocilina e o cloranfenicol (tetB e cmlA) (Martí et al., 2006;
Vila et al., 2007; Coyne et al., 2011). A família SMR contém o sistema de efluxo AbeS.
A inativação do gene regulador abeS confere um maiore nível de resistência ao
cloranfenicol, às fluoroquinolonas, à eritromicina e à novobiocina. Além disso, esse
sistema também confere resistência aos corantes e detergentes (Coyne et al., 2011).
A família MATE, contém o sistema de efluxo AbeM que utiliza dois mecanismos
de geração de energia: a força próton motriz e o gradiente de íons de sódio (Piddock,
2006). No entanto, esse sistema utiliza com maior frequência a força próton motriz
para ejetar os antimicrobianos da célula bacteriana (Su et al., 2005). A hiperexpressão
desse sistema confere um aumento, maior que quatro vezes, na CIM para
norfloxacino, ofloxacino, ciprofloxacino, gentamicina, triclosan e brometo de etídio.
Além disso, o mesmo sistema aumenta em duas vezes a CIM para canamicina,
eritromicina, cloranfenicol, cloreto de tetrafenilfosfónio (TPPCl) e trimetoprim (Su et al.,
2005). A família DMT ainda é pouco estudada, mas parece estar mais associada à
expulsão de metabólitos do interior da célula bacteriana (Zakataeva et al., 2006).
A família RND é, provavelmente, a mais estudada por estar relacionada à
resistência aos β-lactâmicos, uma das classes de antimicrobianos mais utilizadas.
Essa família utiliza a força próton motriz para expulsar os antimicrobianos da célula.
Ela contém o sistema de efluxo AdeABC, o mais importante dentre os demais
membros dessa família em A. baumannii e que será melhor abordado no item 2.4.3.1.
Além do sistema AdeABC, outros representantes dessa família foram descritos. O
sistema AdeFGH, que é regulado pelo gene adeG, confere resistência às
fluoroquinolonas, ao cloranfenicol, ao trimetoprim, à clindamicina e confere uma
diminuição da sensibilidade às tetraciclinas, à tigeciclina e ao sulfametoxazol (Coyne
et al., 2010a; Magnet et al., 2001; Marchand et al., 2004; Coyne et al., 2011). O

REVISÃO BIBLIOGRÁFICA
35
sistema AdeIJK leva à resistência intrínseca às cefalosporinas, ao aztreonam, às
fluoroquinolonas, à tetraciclinas, às lincosamidas, à rifampicina, ao cloranfenicol, ao
trimetoprim, à novobiocina e ao ácido fusídico (Damier-Piolle et al., 2008; Coyne et al.,
2010b) e, recentemente, seu gene regulador, o adeN, foi descoberto e codifica o
regulador transcricional TetR (Rosenfeld et al., 2012).
2.4.3.1. Sistema AdeABC
O sistema de efluxo AdeABC é o mais importante em A. baumannii. É
composto pelo sistema de três proteínas (AdeA, proteína de fusão, AdeB, componente
trans-membrana, ou seja, a bomba propriamente dita e o AdeC, a proteína de
membrana externa), como mostrado na Figura 4.
Figura 4. Esquema do sistema de efluxo AdeABC. Fonte:
http://www.zsyy.com.cn/alfb_Show.asp?id=2485&Nclass=28&N=N.
A hiperexpressão desse sistema confere resistência, principalmente, aos
aminoglicosídeos, aos β-lactâmicos, às fluoroquinolonas, às tetraciclinas, à tigeciclina,
aos macrolídeos, ao cloranfenicol e ao trimetoprim (Magnet et al., 2001; Héritier et al.,
2005a). Cefepima, cefpiroma e cefotaxima são os β-lactâmicos mais afetados e sofrem
pouco impacto pelos outros sistemas dessa família. O sistema AdeABC é regulado por
dois genes, adeR (resposta regulatória) e adeS (sensor quinase) (Magnet et al., 2001).
Mutações nos genes adeRS são responsáveis pela hiperexpressão do sistema

REVISÃO BIBLIOGRÁFICA
36
AdeABC, conferindo resistência às classes de antimicrobianos descritas
anteriormente. Quando um dos dois genes é inativado, a sensibilidade aos
antimicrobianos é observada, pois a proteína AdeR funciona como um ativador
transcricional (Magnet et al., 2001; Marchand et al., 2004; Coyne et al., 2010b; Coyne
et al., 2011). Transposição de uma cópia de ISAba1 dentro do gene adeS, também
pode levar a hiperexpressão desse sistema, como observado em amostras clínicas no
trabalho de Ruzin e colaboradores (2007). O operon do sistema AdeABC está
representado na Figura 5.
Figura 5. Representação do operon do sistema AdeABC.
Um estudo in vitro com inibidores de sistemas de efluxo como o “carbonyl
cyanide m-chlorophenylhydrazone” (CCCP), “1-(1-naphthylmethyl)-piperazine” (NMP)
e “phenyl-arginine-beta-naphthylamide” (PAβN), mostraram resultados controversos e
diferenças nas atividades dos carbapenens (Bou et al., 2000b; Quale et al., 2003;
Pournaras et al., 2006). A hiperexpressão do sistema AdeABC, provavelmente,
contribui para a resistência aos carbapenens, mas outros mecanismos podem estar
envolvidos, como por exemplo, o estresse osmótico (Coyne et al., 2011). O sistema
AdeABC está presente em muitos isolados de A. baumannii (Bratu et al., 2008; Chen
et al., 2009; Hujer et al., 2006; Huys et al., 2005a; Nemec et al., 2007; Srinivasan et al.,
2009) e parece estar presente apenas em isolados clínicos (Huys et al., 2005b). A
hiperexpressão desse sistema já foi descrita in vivo, como sendo responsável pela
resistência à tigeciclina (Peleg et al., 2007a) e ao ciprofloxacino (Higgins et al., 2004).
adeS adeR adeA adeB adeC

REVISÃO BIBLIOGRÁFICA
37
2.4.4. Alteração das Proteínas Ligadoras de Penicilinas (PBPs)
As PBPs são enzimas que possuem uma origem evolutiva comum e catalisam
a síntese de peptídeoglicano, principal componente da parede celular bacteriana.
Essas proteínas estão associadas à morfogênese e à divisão celular (Sauvage et al.,
2008). São classificadas em dois grupos, de acordo com o seu peso molecular
aparente pela técnica de SDS-PAGE, pelas suas sequências de aminoácidos e suas
funções enzimáticas e celulares: (i) HMM - PBPs de alto peso molecular, subdividida
em duas classes: classe A, PBPs com atividade de transpeptidase e glicosiltranferase
(Bertsche et al., 2005; Born et al., 2006); classe B, PBPs com atividade somente de
transpeptidase (Den Blaauwen et al., 2003; Piette et al.,2004); (ii) LMM - PBPs de
baixo peso molecular ou de classe C que são D,D-carboxipeptidases e/ou
endopeptidases, envolvidas na divisão celular, na maturação do peptideoglicano e
reciclagem da parede celular (Ghosh et al., 2008). As transglicosilases monofuncionais
ou MGTs estão presentes em algumas bactérias. Essas proteínas tem um único
domínio de glicosiltranferase similar á classe A das PBPs, porém, sua função ainda
não é conhecida (Spratt et al.,1996; Cayô et al., 2011b).
Os β-lactâmicos atuam mimetizando o terminal D-Ala-D-Ala, atuando como
inibidores suicidas ao se ligar covalentemente às PBPs (Zapun et al., 2008). Tal
ligação leva à desestabilização da parede da célula bacteriana, levando à lise celular
(Wilke et al., 2005). Dessa forma, mutações nas PBPs levam à resistência aos β-
lactâmicos (Zapun et al., 2008). Em um estudo conduzido por Obara & Nakae (1991),
avaliaram as PBPs de A. calcoaceticus sensíveis a imipenem detectando seis tipos
diferentes de PBPs no gel de SDS-PAGE que foram deletadas. Amostras que eram
resistentes à cefoxitina, à ceftazidima ou à cefoperazona, apresentaram alteração na
expressão das PBPs e na redução da expressão das porinas.
No estudo conduzido por Cayô e colaboradores (2011b), sete genes de PBPs
(PBP1a, PBP1b, PBP2, PBP3, PBP5/6, PBP6b, PBP7/8) e um gene de MGT (MgtA)
foram descritos em isolados de A. baumannii, mostrando a diversidade e

REVISÃO BIBLIOGRÁFICA
38
complexidade dessas enzimas. Na presença de alternativas eficientes, tais como a
produção de CHDLs e/ou a perda de porinas, a pressão seletiva dos β-lactâmicos
sobre as PBPs em A. baumannii, pode ser insuficiente para selecionar mutações que
estariam diretamente envolvidas na resistência sobre esses antimicrobianos. Por outro
lado, é possível que outros mecanismos complexos associados à regulação de genes
codificadores de PBPs, bem como eventos pós-transcricionais, poderiam causar
resistência aos β-lactâmicos nesse patógeno (Cayô et al., 2011b).
Em 2011, um estudo realizado por Yun e colaboradores (2011) avaliou a
regulação proteômica de isolados de A. baumannii resistentes ao imipenem sob
condições de estresse causado por diferentes antimicrobianos. Os autores
observaram que, na presença de imipenem, ocorre um aumento dos níveis de
transcrição dos genes codificadores da PBP1a, PBP3, PBP5/6, dos sistemas de efluxo
AdeABC e AdeJIK, e da AmpC. Além disso, foi observada uma redução da expressão
das OMPs OmpHMP e OmpW, demonstrando a complexa associação de mecanismos
de resistência em isolados de A. baumannii resistentes aos carbapenens.

OBJETIVOS
39
3. OBJETIVOS
3.1. OBJETIVO PRINCIPAL
Avaliar a contribuição da hiperexpressão do sistema de efluxo AdeABC e da
hiperprodução de AmpC na resistência aos β-lactâmicos em isolados clínicos
de A. baumannii produtores de OXA-23 no Brasil.
3.2. OBJETIVOS SECUNDÁRIOS
Verificar a ancestralidade das amostras de A. baumannii produtoras de OXA-23
no Brasil;
Correlacionar à ancestralidade e os padrões genéticos de isolados de A.
baumannii descritos no mundo, com os diferentes clones e subclones de A.
baumannii descritos no Brasil;
Detectar fenotipicamente a hiperexpressão de sistemas de efluxo utilizando o
inibidor PAβN;
Associar o nível de expressão do gene adeB aos seus respectivos grupos
clonais e perfis de sensibilidade correspondentes;
Correlacionar a hiperprodução de AmpC associado à presença de ISAba1 com
a resistência às cefalosporinas de amplo espectro;
Avaliar o perfil das proteínas de membrana externa, correlacionando o perfil de
SDS-PAGE com os resultados de expressão da qRT-PCR.

MATERIAL E MÉTODOS
40
4. MATERIAL E MÉTODOS
4.1. Amostras Bacterianas
Em um estudo prévio publicado por Gales e colaboradores (2011) foi avaliada a
atividade do doripenem frente a uma coleção de 806 isolados clínicos Gram negativos,
provenientes de infecção do trato respiratório inferior, infecção de corrente sanguínea
e de infecção intra-abdominal. Nesse estudo, um total de 103/806 isolados (12,78%)
foi identificado como sendo Acinetobacter spp. Os isolados foram obtidos em 18
hospitais privados com mais de 200 leitos localizados em oito estados brasileiros de
quatro regiões geográficas distintas. Inicialmente, no estudo conduzido por Werneck e
colaboradores (2011a) foi realizada uma avaliação da produção de CHDLs nesses
isolados, no qual foi verificado que 76/103 (73,78%) eram produtores de OXA-23.
Além disso, no mesmo estudo foi observada uma alta diversidade genética entre os
isolados produtores de OXA-23, sendo descritos 16 clones e 15 subclones (Werneck
et al., 2011a).
Para a avaliação do impacto do sistema de efluxo AdeABC e da hiperprodução
de AmpC na resistência aos β-lactâmicos em isolados clínicos brasileiros de A.
baumannii produtores de OXA-23, foi selecionado um representante de cada grupo
clonal descrito por Werneck e colaboradores (2011a), incluindo os subclones,
totalizando assim 31 isolados de A. baumannii. A Figura 6 representa a distribuição
dos 31 isolados de A. baumannii selecionados para o presente estudo, de acordo com
a unidade da federação.
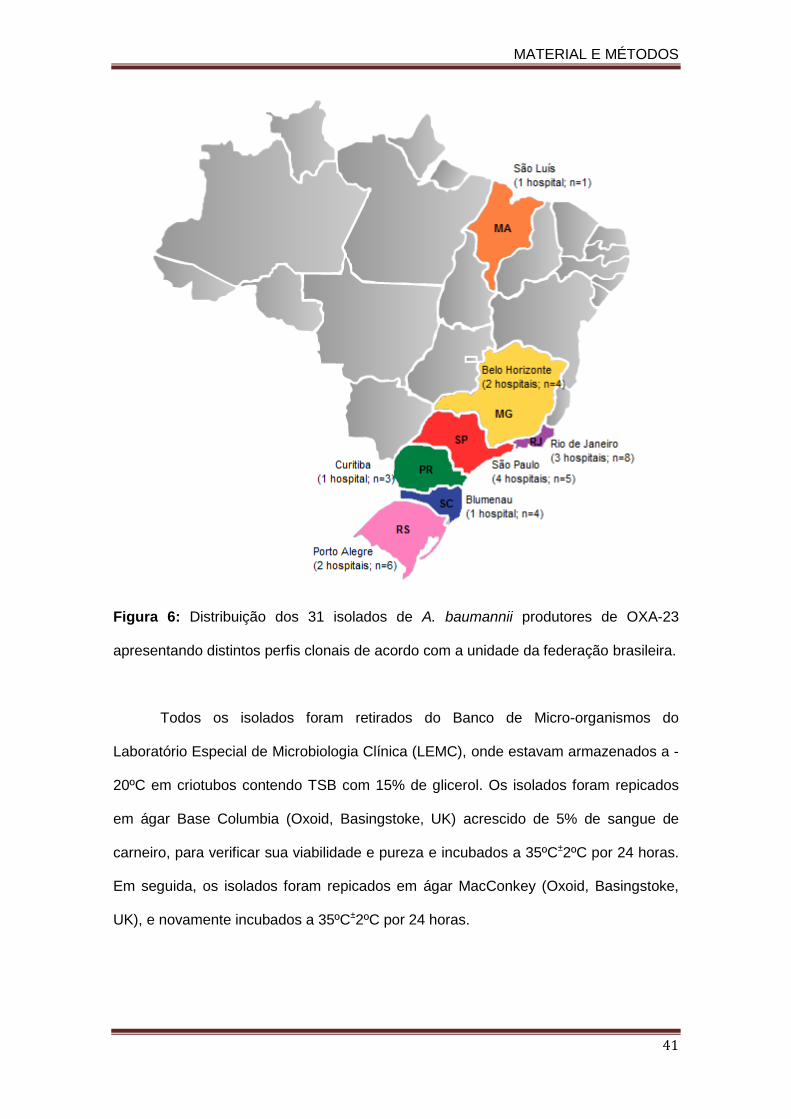
MATERIAL E MÉTODOS
41
Figura 6: Distribuição dos 31 isolados de A. baumannii produtores de OXA-23
apresentando distintos perfis clonais de acordo com a unidade da federação brasileira.
Todos os isolados foram retirados do Banco de Micro-organismos do
Laboratório Especial de Microbiologia Clínica (LEMC), onde estavam armazenados a -
20ºC em criotubos contendo TSB com 15% de glicerol. Os isolados foram repicados
em ágar Base Columbia (Oxoid, Basingstoke, UK) acrescido de 5% de sangue de
carneiro, para verificar sua viabilidade e pureza e incubados a 35ºC±2ºC por 24 horas.
Em seguida, os isolados foram repicados em ágar MacConkey (Oxoid, Basingstoke,
UK), e novamente incubados a 35ºC±2ºC por 24 horas.

MATERIAL E MÉTODOS
42
4.2. Identificação dos Isolados de A. baumannii
4.2.1. “Matrix-Assisted Laser Desorption Ionization Time-Of-Flight Mass
Spectrometry” (MALDI-TOF MS)
Após o subcultivo dos 31 isolados de A. baumannii que estavam armazenados
no banco de micro-organismos, todos os isolados tiveram sua identificação
inicialmente realizada pela metodologia MALDI-TOF MS, utilizando o espectrômetro de
massa Microflex LT e o software MALDI Biotyper 2.0 (Bruker Daltonics,
Massachusetts, USA). A extração das proteínas totais de cada isolado foi realizada de
acordo com as normas do fabricante (Bruker Daltonics, Massachusetts, USA), como
se segue: em um tubo de 1,5 mL, uma alça de 1 µL contendo colônias bacterianas, foi
diluída em 300 µL de água destilada estéril. Em seguida, foi adicionado 900 µL de
etanol absoluto (Carlo Erba, Arese, IT). Esta mistura foi cuidadosamente agitada e
centrifugada a 13.000 rpm durante 2 minutos. O sobrenadante foi descartado e o
precipitado ressuspenso vigorosamente com 50 µL de ácido fórmico a 70% (Sigma-
Aldrich, St. Louis, USA) e misturado com 50 µL de acetonitrila pura (Sigma-Aldrich, St.
Louis, USA). Esta solução foi centrifugada a 13.000 rpm durante 2 minutos e 30 µL do
sobrenadante foi armazenado a -20°C, para posterior análise pelo aparelho.
Para a identificação das amostras clínicas, 1 µL do extrato proteico foi pipetado
em triplicata, numa placa de aço inoxidável, reutilizável, contendo 96 regiões
demarcadas para inoculação das amostras, fornecida pelo fabricante. O inóculo foi
deixado à temperatura ambiente para secagem e posteriormente adicionado 1 µL da
matriz [500 mg/mL de ácido alfa-4-ciano-hidroxicinamico (Bruker Daltonics,
Massachusetts, USA)] diluída em uma solução de ácido trifluoroacético a 2,5%
(Acetonitrila 50%, água 50% e ácido trifluoroacético 2,5%), e novamente deixado à
temperatura ambiente para secagem. Como calibrante, foi utilizado o extrato proteico
da cepa Escherichia coli DH5α. Após o processamento das amostras e do calibrante, a
placa foi inserida no equipamento para realização da identificação bacteriana.

MATERIAL E MÉTODOS
43
A identificação pela técnica do MALDI-TOF MS é baseada de acordo com o
valor do “score” liberado pelo aparelho. Um valor ≥ 2,3 indica que a identificação do
gênero e da espécie é confiável; entre 2,0 a 2,29 indica que a identificação do gênero
é confiável e da espécie é provável; e entre 1,7 a 1,9 indica que a identificação do
gênero é provável. Quando os valores de “score” são inferiores a 1,69 o aparelho
reporta como micro-organismo não identificado, pois não reconhece o perfil proteico
armazenado em seu banco de dados, devendo a amostra ser novamente testada.
Somente foi considerada a identificação com score ≥ 2,3 para o presente estudo.
4.2.2. Sequenciamento do Gene rpoB
Após a análise pelo MALDI-TOF MS, as amostras tiveram sua identificação
confirmada pelo sequenciamento do gene rpoB, codificador da subunidade β da RNA
polimerase, como descrito por La Scola e colaboradores (2006) e validado
posteriormente por Gundi e colaboradores (2009). Quatro regiões, sendo duas
internas e duas que flanqueavam o gene rpoB, foram padronizadas para a
identificação das diferentes espécies do gênero Acinetobacter (La Scola et al., 2006;
Gundi et al., 2009). Os oligonucleotídeos utilizados neste estudo estão descritos na
Tabela 2. Para esse estudo, foram amplificadas as duas regiões internas ao gene
rpoB, utilizando os primers Ac969F e Ac1598R, produzindo um amplicon de 800 pb.

MATERIAL E MÉTODOS
44
Tabela 2. Sequência de oligonucleotídeos das zonas internas e flanqueadoras do
gene rpoB para identificação de Acinetobacter spp. de acordo com La Scola e
colaboradores (2006).
Região Oligonucleotídeos Sequência 5´-3’ Tamanho do fragmento
(pb)
Tm (°C)
Zona 1
Ac969 F TAYCGYAAAGAYTTGAAAGAAG
350
49,7
Ac1093 R
CMACACCYTTGTTMCCRTGA
55,2
Zona 2
Ac1055 F
GTGATAARATGGCBGGTCGT
450
55,5
Ac1598 R
CGBGCRTGCATYTTGTCRT
57,1
rplL
AcintLB F
GAAGARCTTAAGAMDAARCTTG
301-310
60
AcintLB R
CGTTTCTTTTCGGTATATGAGT 60
rpoC
AcintBC F
GTTCTTTAGGTATCAACATTGAA
86-177
60
AcintBC R
GACGCAAGACCAATACGRAT 59
Em um tubo de microcentrífuga de 1,5 mL foi adicionado 350 μL de água Milli-
Q® estéril (Billerica, MA, EUA) e em seguida, diluídas de 3 a 5 colônias bacterianas
previamente isoladas (item 4.1). As reações em cadeia da polimerase (PCR) foram
preparadas em fluxo laminar, contendo master-mix (“GoTaq Green Master Mix”,
Promega, Madison, EUA), água estéril (Water, Molecular Biology Grade, Eppendorf
AG, Hamburg, Alemanha) e oligonucleotídeos (Invitrogen, Carlsbad, EUA). A PCR foi
realizada em Termociclador Mastercycler Gradient (Eppendorf, Hamburg, Germany)
com desnaturação inicial a 94°C por 10 minutos, seguida por 35 ciclos de
desnaturação a 94°C por 30 segundos, anelamento dos oligonucleotídeos a 48°C por
30 segundos e extensão das fitas a 72°C por 1 minuto e uma extensão final a 72°C
por 10 minutos.
Os fragmentos gerados pela PCR foram purificados com o “Kit PCR CleanUp”
(Qiagen, Hilden, Germany) e submetidos ao sequenciamento utilizando 2 µL de

MATERIAL E MÉTODOS
45
“BigDye Terminator v 3.1” (Applied Biosystems, Warrington, UK), diluído em 6 µL de
“Buffer Big Dye Terminator 5x” (Applied Biosystems, Warrington, UK), adicionado a 1
µL do oligonucleotídeo diluído a 3,2 pmol e 5 µL de DNA purificado a 75-100 ng. A
reação foi completada com água ultrapura para a obtenção de um volume final de 20
µL. As condições de ciclagem foram: desnaturação do DNA a 95°C por 1 minuto;
anelamento dos oligonucleotídeos a 50°C por 30 segundos e extensão das fitas a
60°C por 3 minutos. O sequenciamento foi realizado no aparelho 3500 Genetic
Analyzer (Applied Biosystems, Warrington, UK). As sequências de DNA obtidas e as
sequências proteicas derivadas foram analisadas utilizando o programa “Lasergene
Software Package” (DNAStar, Madison, EUA) e então, comparadas com as
sequências depositadas no “GenBank” (http://www.ncbi.nlm.nih.gov/blast/).
Como descrito por Gundi e colaboradores (2009), um isolado foi considerado
como pertencente a uma determinada espécie do gênero Acinetobacter quando a
homologia de sequência foi superior a 98%. Caso algum isolado apresentasse alta
homologia para mais de uma espécie, as duas regiões flanqueantes ao gene rpoB
(Tabela 2) (La Scola et al., 2006) seriam utilizadas com o objetivo de se chegar à
identificação final.
4.3. Análise da Similaridade Genética por PFGE
A relação genética entre os isolados de A. baumannii pertencentes aos 16
clones e 15 subclones selecionados para o estudo, foi confirmada pela técnica de
PFGE. As amostras foram incubadas por 18 horas em 3 mL de caldo TSB (Oxoid,
Basingstoke, UK) e, então, centrifugadas por 15 minutos a 3.000 rpm. O sobrenadante
foi desprezado e o precipitado, contendo as células bacterianas, diluído em 1 mL de
solução salina, homogeneizado e transferido para tubo de microcentrífuga
previamente pesado. Os tubos foram centrifugados por 1 minuto a 12.000 rpm e o
sobrenadante removido. O sedimento bacteriano foi diluído em solução salina, na
proporção 1:1 entre o volume de diluente e o peso do material obtido pela

MATERIAL E MÉTODOS
46
centrifugação. Esse último valor foi calculado pela subtração do peso do tubo vazio
pelo peso do tubo contendo o precipitado de células. Após minuciosa
homogeneização, 7 L da suspensão bacteriana foi transferida para outro tubo
contendo 300 L de tampão TEN (Tris 1 M pH 7,0; Tris base 1 M pH 8,0, EDTA 0,5 M
pH 7,0, EDTA 0,5 M pH 8,0, NaCl 4 M e água destilada) e, então, 340 L de agarose
“low melting” 2% (Invitrogen, Eragny, France) foi adicionado e homogeneizado a esta
mistura, sendo colocada em moldes (Bio-Rad Laboratories, Califórnia, USA) para a
formação de blocos de agarose.
Depois de solidificados, os blocos de agarose foram incubados a 37°C, por no
mínimo 5 horas em solução EC [Tris base 1M pH 7,0, Tris base 1 M pH 8,0, EDTA 0,5
M pH 7,0, EDTA 0,5 M pH 8,0, NaCl, N-lauril sarcosil, Brij 58 (Sigma-Aldrich, St. Louis,
USA), deoxicolato de sódio e água destilada] juntamente com 200 L de lisozima
(Sigma-Aldrich, St. Louis, USA) por amostra. Após a incubação dos blocos de
agarose, os reagentes foram aspirados e os blocos lavados duas vezes com 2 mL de
CHEF-TE (Tris 1 M pH 7,0; Tris base 1 M pH 8,0, EDTA 0,5 M pH 7,0, EDTA 0,5 M pH
8,0 e água destilada) por 30 minutos à temperatura ambiente. O tampão CHEF-TE foi
removido e substituído por 2 mL de solução ES (EDTA 625 mM pH 9,3 e N-lauril
sarcosil 5%) e 100 L de proteinase K (Sigma-Aldrich, St. Louis, USA) a 20 mg/mL. As
amostras foram incubadas por 12 horas a 50°C. Após o tratamento com proteinase K,
foram realizadas quatro lavagens com CHEF-TE por 60 minutos cada e, então,
armazenadas a 5°C.
Para a digestão do DNA bacteriano, os blocos de agarose foram incubados
inicialmente por quatro vezes de 60 minutos cada, com 200 L de DNS (Tris base 1 M
pH 8,0, MgCl2 1 M e água destilada) à temperatura ambiente. O DNS foi então
substituído por 50 L do “tampão de enzima de restrição” (Tampão NEB 10x, BSA e
água) (Uniscience, São Paulo, Brazil) e, em seguida, os blocos de agarose foram
incubados por 60 minutos a 5°C. Após a incubação, este tampão foi substituído

MATERIAL E MÉTODOS
47
novamente por 50 L do “tampão de enzima de restrição” contendo 3 L da enzima
ApaI e incubado por 60 minutos a 5°C. Após este período, os blocos de agarose foram
incubados a 25°C por 12 a 18 horas para a clivagem do DNA.
Após a digestão enzimática do DNA, a eletroforese de campo pulsado foi
realizada em gel de agarose 1% (Invitrogen, Eragny, France) com TBE 0,5X (Tris base
ácido bórico EDTA e água destilada), no sistema CHEF-DR II (Bio-Rad Laboratories,
California, USA) à temperatura de 14oC e utilizando corrente elétrica de 200 Volts
(6V/cm) com switch time inicial de 5 e final de 35 segundos durante 19 horas. O gel foi
corado com brometo de etídio 10 mg/mL e fotografado sob iluminação ultravioleta em
Transiluminador (Bio-Rad Laboratories, California, USA). O gel foi analisado por
inspeção visual, utilizando os critérios de Tenover e colaboradores (1995) e pelo
programa Bionumerics, para avaliar a similaridade genética entre os isolados.
4.4. “Multilocus Sequence Typing” (MLST)
Para a determinação do “Sequence Typing” (ST) dos 31 isolados clínicos de A.
baumannii produtores de OXA-23, foi realizada a técnica de MLST do Institut Pasteur,
publicada por Nemec e colaboradores (2008). Cada amostra foi submetida à
amplificação e ao sequenciamento parcial dos sete genes padronizados para o gênero
Acinetobacter spp. (cpn60, fusA, gltA, pyrG, recA, rplB e rpoB), utilizando os
oligonucleotídeos descritos previamente (Nemec et al., 2008), como mostrado na
Tabela 3.

MATERIAL E MÉTODOS
48
Tabela 3. Oligonucleotídeos empregados para os sete genes utilizados para a
caracterização do MLST, de acordo com Nemec e colaboradores (2008).
Oligonucleotídeo Sequência 5’ – 3’ Tamanho do
fragmento (pb) Condição de
corrida
cpn60 F
ACTGTACTTGCTCAAGC
405
94° 2 min
94° 30 seg 50° 30seg 72° 30 seg
72° 5 min
35x
cpn60 R
TTCAGCGATGATAAGAAGTGG
fusA F
ATCGGTATTTCTGCKCACATYGAT
633
fusA R
CCAACATACKYTGWACACCTTTGTT
gltA F
AATTTACAGTGGCACATTAGGTCCC
483
gltA R
GCAGAGATACCAGCAGAGATACACG
pyrG F
GGTGTTGTTTCATCACTAGGWAAAGG
297
pyrG R
ATAAATGGTAAAGAYTCGATRTCACCMA
recA F
CCTGAATCTTCYGGTAAAAC
372
recA R
GTTTCTGGGCTGCCAAACATTAC
rplB F
GTAGAGCGTATTGAATACGATCCTAACC
330
rplB R
CACCACCACCRTGYGGGTGATC
rpoB F
GGCGAAATGGC(AGT)GA(AG)AACCA
456 rpoB R
GA(AG)TC(CT)TCGAAGTTGTAACC
As sequências de DNA obtidas, foram analisadas utilizando o programa
“Lasergene Software Package” (DNAStar, Madison, EUA) e, então, as sequências
para cada um dos sete genes avaliados, foram depositadas em conjunto por isolado
avaliado, diretamente na página do MLST do Institut Pasteur, França
(http://www.pasteur.fr/cgi-bin/genopole/PF8/mLstdbnet.pl?page=allseq&file=acin_profiles.xmL),
a fim de se obter os respectivos alelos e, consequentemente, o número do ST (Nemec
et al., 2008; Diancourt et al., 2010). Para as sequências que não tiveram seus alelos
determinados pelo site, os eletroferogramas dos respectivos alelos foram enviados ao

MATERIAL E MÉTODOS
49
curador do site, juntamente com os dados epidemiológicos exigidos para a obtenção
do número do novo alelo e do respectivo ST.
4.5. Testes de Sensibilidade aos Antimicrobianos
Todos os 31 isolados de A. baumannii tiveram sua concentração inibitória
mínima (CIM) obtida para os 14 antimicrobianos selecionados para o estudo, pela
técnica de microdiluição em caldo, seguindo as recomendações do CLSI M7-A9 (CLSI,
2012). Foram testados os seguintes antimicrobianos: ampicilina/sulbactam,
cefotaxima, ceftazidima, cefepima, aztreonam, imipenem, meropenem, ciprofloxacina,
levofloxacina, amicacina, gentamicina, minociclina, tigeciclina e polimixina B. Todos os
sais de antimicrobianos foram obtidos da Sigma-Aldrich (Sigma-Aldrich, St. Louis,
USA).
As soluções estoque dos antimicrobianos testados foram preparadas, em
concentrações 20 vezes superiores às concentrações finais, obtidas ou alcançadas
nas placas de microdiluição em caldo, e armazenadas a -70ºC até o seu
processamento. A partir da solução estoque, foram realizadas duas diluições de 1:20,
até a obtenção da maior diluição a ser testada. A partir desta, foram realizadas
diluições seriadas, das quais, 100 μL foi dispensado na placa de microdiluição. As
placas foram armazenadas em freezer -70°C até a realização dos testes.
Após o crescimento bacteriano em placas de ágar MacConkey (Oxoid,
Basingstoke, UK) por 24 horas, com auxílio de uma alça de semeadura, foi preparado
um inóculo na escala 0,5 de McFarland em 5 mL de caldo Müeller-Hinton (MH) (Oxoid,
Basingstoke, UK), para obtenção de uma concentração bacteriana em torno de 1,5 x
108 unidades formadoras de colônias (UFC)/mL. Uma alíquota 132 μL da suspensão a
0,5 de McFarland foi transferida para um tubo contendo 19.868 μL caldo MH (Oxoid,
Basingstoke, UK) para a obtenção de uma concentração bacteriana final em torno de 5
x 105 UFC/mL. Após a homogeneização, 100 μL dessa suspensão bacteriana foi
inoculada em cada poço da placa de microdiluição em caldo, implicando assim, em

MATERIAL E MÉTODOS
50
uma diluição de 1:2 do inóculo bacteriano e das concentrações de antimicrobianos
testadas. A concentração final de bactérias em cada poço da placa de microdiluição
em caldo foi de aproximadamente 2,5 x 105 UFC/mL.
A seguir, as placas foram incubadas a 35°C±2ºC, em aerobiose por 20 a 24
horas. A leitura das placas de microdiluição em caldo foi realizada visualmente. A CIM
foi considerada como a menor concentração do antimicrobiano capaz de inibir
completamente o crescimento bacteriano. Os resultados foram interpretados de
acordo com os pontos de corte estabelecidos pelo CLSI (2012), exceto para a
tigeciclina, onde foram utilizados os pontos de corte sugeridos por Jones e
colaboradores (2007). Para o controle de qualidade dos testes de sensibilidade, foram
utilizadas as cepas “American Type Culture Collection” (ATCC®) E. coli ATCC® 25922,
P. aeruginosa ATCC® 27853, E. coli ATCC® 35218 e S. aureus ATCC® 29213.
4.5.1. Avaliação Fenotípica da Hiperexpressão de Sistemas de Efluxo
Inicialmente, para verificar se a concentração a ser utilizada do inibidor nos
testes de sensibilidade não seria capaz de inibir o crescimento bacteriano, o que
poderia influenciar nos resultados, os 31 isolados foram inoculados em placas de ágar
Müeller-Hinton (Oxoid, Basingstoke, UK) a uma suspensão de 0,5 de McFarland,
contendo discos com as concentrações de 10 μg/mL, 15 μg/mL, 25 μg/mL, 50 μg/mL e
100 μg/mL do inibidor e, então, incubados a 35°C±2ºC por 24 horas, para posterior
avaliação do crescimento bacteriano. Optou-se por utilizar a concentração de 15
μg/mL, pois não existe um valor padronizado da concentração do inibidor para este
teste.
Para a avaliação fenotípica da hiperexpressão dos sistemas de efluxo, as CIMs
foram determinadas para os antimicrobianos β-lactâmicos ceftazidima, cefotaxima,
imipenem e meropenem, como descrito no item 4.5, na ausência e na presença do
inibidor de sistemas de efluxo, fenilalamida arginina β-nafitilamida - PAβN (Sigma-
Aldrich, St Louis, EUA), em concentrações fixas de 15 μg/mL. Uma diminuição da CIM

MATERIAL E MÉTODOS
51
≥ 2 diluições na presença do PAβN comparada a CIM do antimicrobiano sozinho foi
considerada indicativa da hiperexpressão de sistemas de efluxo.
4.6. Pesquisa e Identificação de Genes de Resistência
Todos os 31 isolados de A. baumannii tiveram a produção de OXA-23,
confirmada pela técnica de PCR multiplex descrita por Woodford e colaboradores
(2006). A presença de OXA-143 foi confirmada por PCR convencional, usando os
oligonucleotídeos descritos por Higgins e colaboradores (2009), como descritos na
Tabela 4. Os demais genes codificadores de β-lactamases, também foram avaliados
neste estudo, e os respectivos oligonucleotídeos utilizados para a amplificação destes
genes estão mostrados na Tabela 4.
Tabela 4. Sequência de oligonucleotídeos para identificação dos genes codificadores
de -lactamases e sequência de inserção.
Oligonucleotídeo Sequência 5’-3’ Tamanho do fragmento
(pb)
Tm (°C)
Referência
Pré-TEM F
GTATCCGCTCATGAGACAATA
955
51,7
Bermudes et al., 1999
Pré-TEM R
TCTAAAGTATATATGAGTAAACTTGGTCTG
52,6
MTPX SHV F
CTTGACCGCTGGGAAACGG
200
59,0
Laboratório Alerta
MTPX SHV R
AGCACGGAGCGGATCAACGG
62,6
MTPX CTX-M 1,2 F
ATGTGCAGYACCAGTAA
512
49,3
Laboratório Alerta
MTPX CTX-M 1,2 R
CGCTGCCGGTTTTATCSCCC
61,6
MTPX CTX-M-8 F
AACRCRCAGACGCTCTAC
333
55,0
Laboratório Alerta MTPX CTX-M-8 R
TCGAGCCGGAASGTGTYAT
57,6
MTPX CTX-M-14 F
GGTGACAAAGAGARTGCAACGGAT
876
58,6
Laboratório Alerta MTPX CTX-M-14 R
TTACAGCCCTTCGGCGATGA
58,9
MTPX GES F
AGCAGCTCAGATCGGTGTTG
750 57,5

MATERIAL E MÉTODOS
52
MTPX GES R
CCGTGCTCAGGATGAGTTG
55,8 Laboratório Alerta
MTPX SPM F
CTAAATCGAGAGCCCTGCTTG
798
55,6
Mendes et al., 2007b
MTPX SPM R
CCTTTTCCGCGACCTTGATC
56,3
MTPX IMP F
GAATAG(A/G)(A/G)TGGCTTAA(C/T)TCTC
188
49,2
Mendes et al., 2007b
MTPX IMP R
CCAAAC(C/T)ACTA(G/C)GTTATC
45,5
MTPX VIM F
GTTTGGTCGCATATCGCAAC
382
54,8
Mendes et al., 2007b
MTPX VIM R
AATGCGCAGCACCAGGATAG
58,0
MTPX GIM F
TCAATTAGCTCTTGGGCTGA
72
54,9
Mendes et al., 2007b MTPX GIM R
CGGAACGACCATTTGAATGG
54,3
MTPX SIM F
GTACAAGGGATTCGGCATCG
569
55,9
Mendes et al., 2007b
MTPX SIM R
TGGCCTGTTCCCATGTGAG
57,5
Pré-NDM F
GGCGTTAGATTGGCTTACACC
1146
56,1
Laboratório Alerta
Pré-NDM R
CTGGGTCGAGGTCAGGATAG
56,4
KPC F
TCGCTAAACTCGAACAGG
785
51,9
Bratu et al., 2005
KPC R
TTACTGCCCGTTGACGCCCAATCC
63,7
MTPX OXA-1 F TATCTACAGCAGCGCCAGTG
601
60,0
Laboratório Alerta
MTPX OXA-1 R TGCACC AGTTTTCCC ATACA 60,0
MTPX OXA-2 F CGATAGTTGTGGCAGACGAA
504
60,0
Laboratório Alerta MTPX OXA-2 R TCTTTGCACGCAGTATCCAG 60,0
MTPX OXA-10 F GTCTTTCGAGTACGGCATTA
720
52,1
Laboratório Alerta MTPX OXA-10 R
ATTTTCTTAGCGGCAACTTAC
51,2
MTPX OXA-18 F ATGCAACGGAGCCTGTCC
109
66,8
Laboratório Alerta MTPX OXA-18 R GGCAGGGTGTTGAGGAACT 57,8
MTPX OXA-23 F
GATCGGATTGGAGAACCAGA
501
54,0
Woodford et al., 2006
MTPX OXA-23 R
ATTTCTGACCGCATTTCCAT
52,5

MATERIAL E MÉTODOS
53
MTPX OXA-24 F
GGTTAGTTGGCCCCCTTAAA
246
54,9
Woodford et al., 2006
MTPX OXA-24 R
AGTTGAGCGAAAAGGGGATT
54,5
MTPX OXA-51 F
TAATGCTTTGATCGGCCTTG
353
53,3
Woodford et al., 2006
MTPX OXA-51 R
TGGATTGCACTTCATCTTGG
53,0
MTPX OXA-58 F
AAGTATTGGGGCTTGTGCTG
599
55,6
Woodford et al., 2006
MTPX OXA-58 R
CCCCTCTGCGCTCTACATAC
57,2
Pré-OXA-143 F
AGTTAACTTTCAATAATTG
825
40,1
Higgins et al., 2009
Pré-OXA-143 R
TTGGAAAATTATATAATCCC
42,3
OXA-69 F CTAATAATTGATCTACTCAAG
975
52,0
Héritier et al., 2005b
OXA-69 R
CCAGTGGATGGATGGATAGATTATC
52,0
Pré-AmpC F
GCGCCGTGAATTCTTAAGTG
2740
60,7
Este estudo
Pré-AmpC R
AATAATGCCATTCGCTTTAATTG 59,4
ISAba 1 F
GTCAGTTGCACTTGGTCG
1091 53,9 Este estudo
Em fluxo laminar, uma solução mãe foi preparada contendo “GoTaq® Green
Master Mix” (Promega, Madison, USA), água estéril e oligonucleotídeos a 20 mM
(Invitrogen, Eragny, France). Após leve agitação, 19 μL da mistura foi transferido para
cada tubo de reação, que continha 1 μL de uma suspensão da bactéria a ser testada
(item 4.2.2). As condições de termociclagem para cada um dos oligonucleotídeos
foram realizadas conforme publicado previamente pelos respectivos autores (Tabela
4). Após a amplificação do DNA, a revelação do produto foi feita em eletroforese em
gel de agarose (Invitrogen, Eragny, France) a 2%, contendo brometo de etídio a 2,5
mg/mL, imerso em TBE à 0,5X. A visualização foi feita sob luz ultravioleta em
transiluminador (Bio-Rad Laboratories, California, USA). Os produtos de PCR foram
purificados e sequenciados, como descritos anteriormente no item 4.2.2.

MATERIAL E MÉTODOS
54
4.6.1. Avaliação da Hiperprodução de AmpC (ADC)
Para a avaliação da hiperprodução de AmpC entre os isolados de A. baumannii
produtores de OXA-23, foi pesquisada a presença da sequência de inserção ISAba1,
responsável pela hiperexpressão da β-lactamase cromossômica, a montante do gene
blaADC-like por PCR. Para isso, foram utilizados os primers ISAba1.F (5’-
GTCAGTTGCACTTGGTCG-3’) e ADC.R (5’-TGCAAGGCAAGGTTACCACT-3’). A
visualização de um fragmento de 780 pb indicou a presença da ISAba1 a montante do
gene blaADC.
4.7. Avaliação das Proteínas de Membrana Externa por SDS-PAGE
Para verificar possíveis alterações no perfil das proteínas de membrana
externa dos isolados clínicos de A. baumannii produtores de OXA-23, estes foram
submetidos à técnica de SDS-PAGE. Os isolados foram cultivados em 15 mL de caldo
LB (Oxoid, Basingstoke, UK) a 37°C, sob agitação, por 24 horas. Os inóculos foram
centrifugados a 4.000 rpm, por 15 minutos e a 4°C. O sedimento bacteriano foi então
ressuspenso em 1 mL de tampão Tris-Mg (Tris 10 mM; MgCl2 5 mM), pH 7,3 e
sonicado por cinco ciclos de 30 segundos. Em cada ciclo, a sonicação é interrompida
por um segundo a cada cinco segundos. Após essa etapa, foi realizada outra
centrifugação a 5.000 rpm, por 5 minutos e a 4°C. O sobrenadante foi transferido para
outro tubo e centrifugado novamente por 30 minutos, a 17.000 rpm e a 4°C. O
sobrenadante foi descartado e o sedimento foi ressuspenso em 800 μL de sarcosil a
2% e incubado por 30 minutos em temperatura ambiente. Transcorrido esse período,
foi realizada uma nova centrifugação de 30 minutos, a 17.000 rpm e a 4°C. O
sedimento foi então, lavado com tampão Tris-Mg (Tris 10 mM; MgCl2 5 mM), pH 7,3 e
centrifugado por 30 minutos, a 17.000 rpm e a 4ºC. O sobrenadante foi descartado e o
sedimento, correspondente à porção da membrana externa, foi ressuspenso em 40 μL

MATERIAL E MÉTODOS
55
do tampão Tris-Mg (Tris 10 mM; MgCl2 5 mM), pH 7,3. O extrato proteico foi
armazenado a -20°C até sua utilização.
Para a calibração da curva e, subsequente quantificação dos extratos proteicos
foi utilizado o reagente de Bradford (Bio-Rad Laboratories, California, USA), acrescido
de albumina de soro bovino nas concentrações 100, 50, 25, 12,5 6,25 e 3,125 µg/mL
para obtenção da curva de calibração, utilizando o espectrofotômetro digital “NanoVue
Plus Spectrophotometer” (GE Healthcare, Canadá). Após a quantificação das
proteínas pelo método de Bradford, as concentrações dos extratos proteicos foram
igualadas, levando-se em consideração a menor concentração identificada, em
tampão de amostra “Laemmi Sample Buffer” (Bio-Rad Laboratories, California, USA) e
aquecida a 100ºC por 5 minutos. Um total de 20 µL do extrato proteico foi adicionado
ao gel de poliacrilamida a 17%. A eletroforese foi realizada a uma voltagem de 180 V
por 50 minutos. O “Precision Plus Protein™ Dual Color Standards” (Bio-Rad
Laboratories, California, USA), foi utilizado como marcador de peso molecular.
4.8. PCR em Tempo Real (qRT-PCR) para Quantificação da Transcrição do
Sistema de Efluxo AdeABC e das Proteínas de Membrana Externa CarO,
33-36 kDa, OmpHMP e OmpW
A técnica de qRT-PCR foi utilizada para quantificar a transcrição relativa do
gene adeB, que compõe o sistema de efluxo AdeABC, e dos genes codificadores das
proteínas de membrana externa 33-36kDa, CarO, OmpHMP e OmpW que estão
envolvidas no aumento da resistência aos carbapenens.
4.8.1. Extração do RNA Total e Síntese de cDNA
Os isolados foram cultivados em 20 mL de Caldo LB (Oxoid, Basingstoke, UK)
a 37°C, sob agitação, até atingirem a fase logarítmica de crescimento, correspondente
à absorbância de 0,5 a 0,6 a um comprimento de onda de 600 nm em
espectrofotômetro BioMate 5, ThermoSpectronic (Thermo Fisher Scientific, Delaware,

MATERIAL E MÉTODOS
56
USA). Um mililitro da cultura bacteriana foi adicionado a 1 mL de RNA Protect Bacteria
Reagent (Qiagen, Hilden, Germany), homogeneizado por completo em vórtex e
incubado por 5 minutos em temperatura ambiente. Transcorrido esse período, foi
realizada uma centrifugação por 10 minutos a 12.000 rpm em temperatura ambiente.
O RNA foi então, extraído a partir do sedimento bacteriano resultante, utilizando o
“RNeasy Mini Kit” (Qiagen, Hilden, Germany), de acordo com as recomendações do
fabricante. Para retirada de resíduos de DNA bacteriano foi realizado um tratamento
com “DNAse Free-RNAse Set” (Qiagen, Hilden, Germany), durante a extração do
RNA. O RNA total foi quantificado no espectrofotômetro digital “NanoVue Plus
Spectrophotometer” (GE Healthcare, Canadá).
A reação de transcriptase reversa foi realizada utilizando 30 ng de RNA de
cada amostra, com “High Capacity cDNA Reverse Transcription Kit” (Applied
Biosystems, Warrington, UK), de acordo com as recomendações do fabricante. A
reação foi incubada por 10 minutos a 25°C, seguido por 120 minutos a 37ºC, em
Termociclador Mastercycler Gradient (Eppendorf, Hamburg, Germany). O cDNA foi
armazenado em freezer -80°C até sua utilização.
4.8.2. Quantificação Relativa da Transcrição Gênica
O cDNA, preparado a partir da extração de RNA das amostras clínicas de A.
baumannii (item 4.8.1), foi submetido à reação de qRT-PCR, utilizando sequências de
oligonucleotídeos (Invitrogen, Eragny, France) específicos para cada gene que
compõe o sistema de efluxo e proteínas de membrana externa, mostrados na Tabela
5.

MATERIAL E MÉTODOS
57
Tabela 5. Sequência de oligonucleotídeos utilizados para a quantificação dos genes
codificadores do sistema de efluxo AdeABC, e dos genes das principais proteínas de
membrana externa, pela técnica de qRT-PCR.
Oligonucleotídeo Sequência 5’ - 3’ Tamanho do
fragmento (pb) Referências
33-36 kDa F GAAGCTGGTGCAATGTTG
196 Pereira, 2009
33-36 kDa R TAGTGCCATCGATGTTGC
CarO F
GCAATGGCAGATGAAGC
111 Pereira, 2009
CarO R TAAAGCACCACCGTAACC
OmpHMP F
CTCTTGCTGGCTTAAACG
254 Pereira, 2009
OmpHMP R TGTGTGACCTTCGATACG
OmpW F
TTAGCATCAGCAGGTTGG
120 Pereira, 2009
OmpW R TATTGGTATCGGGGCAAC
AdeB F
GGATTATGGCGACAGAAGGA
106 Pereira, 2009
AdeB R AATACTGCCGCCAATACCAG
16S F
CAGCTCGTGTCGTGAGATGT
150 Pereira, 2009
16S R CGTAAGGGCCATGATGACTT
As reações de qRT-PCR foram realizadas em triplicata, utilizando 12,5 μL de
“Platinum SyBR Green qPCR” (Invitrogen, Eragny, France), adicionado de 0,5 μL de
cada oligonucleotídeo a uma concentração de 10 µM (Invitrogen, Eragny, France). À
mistura, foi adicionado 3 μL de cDNA e 8,5 μL de água ultrapura, para obtenção de um
volume final de 25 μL. A reação de PCR foi realizada no equipamento 7500 Real Time
PCR System (Applied Biosystems, Warrington, UK). As condições de termociclagem
foram: 2 minutos a 50°C, seguidos de 10 minutos a 95°C e 45 ciclos de 15 segundos a
95°C e 1 minuto a 60°C.

MATERIAL E MÉTODOS
58
4.8.3. Análise da Quantificação Relativa da Transcrição Gênica
Para a determinação da eficiência de cada oligonucleotídeo testado, foi
realizada uma curva de desnaturação “melting curve,” utilizando um extrato de DNA a
partir de um inóculo bacteriano do isolado ATCC 19606, na escala 0,5 de McFarland.
Desse inóculo inicial, foi realizada uma diluição seriada de 1:10 até a diluição de 10-8,
como mostrado na Tabela 6.
Tabela 6. Temperatura de melting e eficiência dos oligonucleotídeos utilizados na
reação de qRT-PCR.
Oligonucleotídeos Temperatura de
Melting °C Eficiência (%) Slope
Concentração do inóculo
OmpW
78,3 124,2 -2,851 10
-1-10
-8
OmpHMP
81,1 117,45 -2,964 10
-1-10
-8
CarO
79,3 119,4 -2,93 10
-1-10
-8
AdeB
77,9 125,7 -2,828 10
-1-10
-8
16s
84,3 112,1 -3,063 10
-1-10
-8
Todas as reações de qRT-PCR foram realizadas em triplicata e o gene
codificador da porção 16S ribossomal foi utilizado como gene de referência para a
normalização da transcrição dos genes alvo. A eficiência das reações de amplificação
dos genes estudados foi determinada de acordo com a fórmula E = (10-1/slope -1) x
100, onde “slope” representa a inclinação da reta no gráfico de regressão linear das
médias dos valores de Ct, observadas nas três replicatas das reações de qRT-PCR
para as diluições seriadas do extrato do isolado ATCC BAA-1709.
Ct “Ciclo Threshold” é considerado o ciclo em que foi detectada fluorescência
durante a amplificação da reação de qRT-PCR, acima do limite basal de fluorescência
observado no início da reação. Como parâmetro para identificação de hiperexpressão

MATERIAL E MÉTODOS
59
de efluxo, foi considerado 40 vezes acima do limite basal da cepa controle ATCC BAA-
1709 A. baumannii.

RESULTADOS
60
5. RESULTADOS
5.1. Identificação dos isolados bacterianos
Todos os 31 isolados selecionados para o presente estudo foram identificados
como sendo A. baumannii pela técnica de MALDI-TOF MS, com score superior a 2,3.
Esse resultado foi posteriormente confirmado pelo sequenciamento da região interna
do gene rpoB, a qual apresentou homologia de 99% com as sequências padrão de A.
baumannii depositadas no GenBank.
5.2. Análise da similaridade genética (PFGE) e da ancestralidade (MLST)
Após a confirmação da espécie, todos os isolados de A. baumannii tiveram o
seu perfil genético confirmado pela técnica de PFGE, sendo observada uma
variabilidade genética entre os isolados brasileiros de A. baumannii produtores de
OXA-23. A análise dos géis de PFGE, a partir do dendrograma obtido utilizando o
programa BioNumerics (Figura 7) e Tenover e colaboradores (1995), demonstrou a
presença de 16 padrões e 15 padrões relacionados distribuídos nos 14 hospitais
privados localizados em sete estados diferentes, como mostrado na Tabela 7.
Nenhum padrão clonal específico pode ser relacionado a uma determinada instituição
ou região geográfica, exceto pelo padrão D, que foi encontrado apenas no Rio de
Janeiro. Dentro de um mesmo hospital, vários padrões de A. baumannii produtores de
OXA-23 podem estar circulando durante um mesmo período de tempo (Hospital 2 no
Rio de Janeiro e Hospital 12 em Porto Alegre). Por outro lado, isolados com padrões
relacionados foram encontrados em diferentes hospitais localizados em áreas
geográficas distintas (padrões I e J).

RESULTADOS
61
Figura 7. Dendrograma do perfil genotípico determinado pela técnica de PFGE dos
isolados de A. baumannii produtores de OXA-23 avaliados neste estudo.
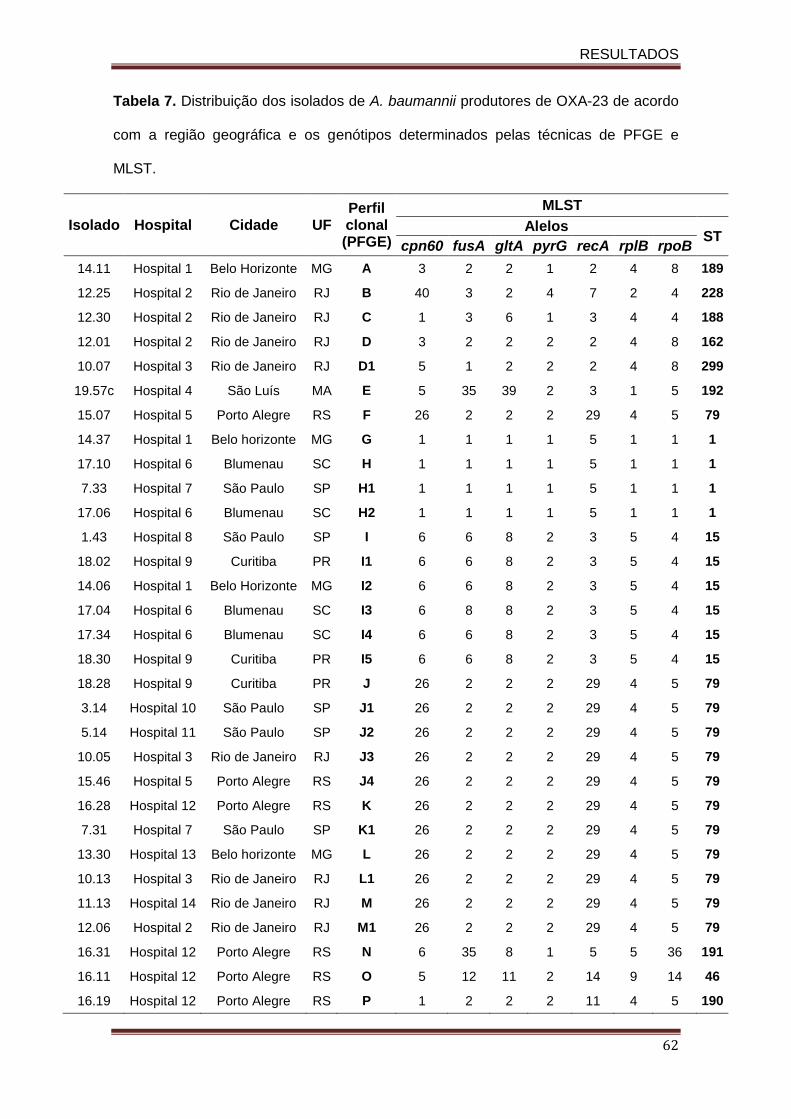
RESULTADOS
62
Tabela 7. Distribuição dos isolados de A. baumannii produtores de OXA-23 de acordo
com a região geográfica e os genótipos determinados pelas técnicas de PFGE e
MLST.
Isolado Hospital Cidade UF Perfil clonal (PFGE)
MLST
Alelos ST
cpn60 fusA gltA pyrG recA rplB rpoB
14.11 Hospital 1 Belo Horizonte MG A 3 2 2 1 2 4 8 189
12.25 Hospital 2 Rio de Janeiro RJ B 40 3 2 4 7 2 4 228
12.30 Hospital 2 Rio de Janeiro RJ C 1 3 6 1 3 4 4 188
12.01 Hospital 2 Rio de Janeiro RJ D 3 2 2 2 2 4 8 162
10.07 Hospital 3 Rio de Janeiro RJ D1 5 1 2 2 2 4 8 299
19.57c Hospital 4 São Luís MA E 5 35 39 2 3 1 5 192
15.07 Hospital 5 Porto Alegre RS F 26 2 2 2 29 4 5 79
14.37 Hospital 1 Belo horizonte MG G 1 1 1 1 5 1 1 1
17.10 Hospital 6 Blumenau SC H 1 1 1 1 5 1 1 1
7.33 Hospital 7 São Paulo SP H1 1 1 1 1 5 1 1 1
17.06 Hospital 6 Blumenau SC H2 1 1 1 1 5 1 1 1
1.43 Hospital 8 São Paulo SP I 6 6 8 2 3 5 4 15
18.02 Hospital 9 Curitiba PR I1 6 6 8 2 3 5 4 15
14.06 Hospital 1 Belo Horizonte MG I2 6 6 8 2 3 5 4 15
17.04 Hospital 6 Blumenau SC I3 6 8 8 2 3 5 4 15
17.34 Hospital 6 Blumenau SC I4 6 6 8 2 3 5 4 15
18.30 Hospital 9 Curitiba PR I5 6 6 8 2 3 5 4 15
18.28 Hospital 9 Curitiba PR J 26 2 2 2 29 4 5 79
3.14 Hospital 10 São Paulo SP J1 26 2 2 2 29 4 5 79
5.14 Hospital 11 São Paulo SP J2 26 2 2 2 29 4 5 79
10.05 Hospital 3 Rio de Janeiro RJ J3 26 2 2 2 29 4 5 79
15.46 Hospital 5 Porto Alegre RS J4 26 2 2 2 29 4 5 79
16.28 Hospital 12 Porto Alegre RS K 26 2 2 2 29 4 5 79
7.31 Hospital 7 São Paulo SP K1 26 2 2 2 29 4 5 79
13.30 Hospital 13 Belo horizonte MG L 26 2 2 2 29 4 5 79
10.13 Hospital 3 Rio de Janeiro RJ L1 26 2 2 2 29 4 5 79
11.13 Hospital 14 Rio de Janeiro RJ M 26 2 2 2 29 4 5 79
12.06 Hospital 2 Rio de Janeiro RJ M1 26 2 2 2 29 4 5 79
16.31 Hospital 12 Porto Alegre RS N 6 35 8 1 5 5 36 191
16.11 Hospital 12 Porto Alegre RS O 5 12 11 2 14 9 14 46
16.19 Hospital 12 Porto Alegre RS P 1 2 2 2 11 4 5 190

RESULTADOS
63
A relação filogenética entre os diferentes padrões e padrões relacionados de A.
baumannii produtores de OXA-23 no Brasil também foi avaliada através da técnica de
MLST (Tabela 7). Os resultados obtidos demonstram 12 STs diferentes entre os 31
isolados avaliados. O ST mais prevalente foi o ST79, que foi encontrado em 12
isolados (38,7%) presentes em dez hospitais brasileiros, localizados em Porto Alegre,
Curitiba, São Paulo, Rio de Janeiro e Minas Gerais e que apresentavam cinco perfis
de PFGE distintos (F, J, K, L e M). O ST15 foi o segundo mais frequente, encontrado
em seis isolados, todos pertencentes ao padrão I, presentes em quatro hospitais
localizados em Belo Horizonte, São Paulo, Blumenau e em Curitiba. O ST1 foi o
terceiro mais prevalente, sendo encontrado em quatro isolados que apresentavam dois
perfis de PFGE diferentes, localizados em hospitais de Belo Horizonte, Blumenau e
São Paulo. Dois isolados encontrados nas cidades de Porto Alegre e do Rio de
Janeiro, apresentaram dois STs (ST46 e ST162, respectivamente) já depositados no
banco de MSLT do “Institut Pasteur”, mas pouco frequentes em nosso meio. Por outro
lado, sete isolados incluídos em sete perfis de PFGE distintos, apresentaram
sequências alélicas únicas e, portanto, foram classificados como novos STs (ST188 ao
ST192, ST228 e ST299). Esses novos STs foram encontrados nas cidades do Rio de
Janeiro, Porto Alegre, Belo Horizonte e em São Luís. Dentre esses, dois isolados do
Rio de Janeiro, relacionados clonalmente (padrões D e D1), apresentaram dois STs
diferentes devido a mutações nos alelos dos genes cpn60 e fusA.

RESULTADOS
64
Figura 8. Árvore filogenética entre os diferentes STs encontrados no presente estudo e os demais STs descritos no mundo até o momento. Legenda:
números em rosa, STs identificados no estudo; pontos em azul, complexos clonais; pontos em verde, STs descritos até o momento; pontos isolados,
“singletons”; dois ou mais pontos verdes interligados, amostras geneticamente correlacionada (eBURST realizado dia 31/05/2013).

RESULTADOS
65
O ST79 está inserido dentro do CC79 e no SG3, como mostrado na Figura 8.
Outros três STs encontrados no presente estudo, também fazem parte do CC79, que
incluem o ST162, ST189 (um SLV [“Single Locus Variant”] do ST162) e o ST190 (a
DLV [“Double Locus Variant”] do ST79). Sendo assim, quase metade (48.39%,
n=15/31) dos isolados brasileiros de A. baumannii produtores de OXA-23 avaliados
neste estudo pertecem ao CC79. Embora o ST299 tenha sido classificado como
“singleton”, ele é um DLV do ST162 e, portanto, está relacionado ao CC79. Entretanto,
como não existe um ST apresentando um SLV entre o ST299 e o ST162, ele não pode
ser incluído no CC79. O ST15 (n=6) e o ST1 (n=4) pertencem, respectivamente, ao
CC15/SG7 e ao CC1/SG2, dois importantes grupos clonais epidêmicos e
multirresistentes disseminados pelo mundo. O ST46, ST188 e o ST228 estão
correlacionados aos ST149, ST250 e ST25, respectivamente. A inclusão desses três
alelos no banco de dados do MLST permitiu a criação de novos grupos clonais
chamados SG20 (ST188 e ST250), SG21 (ST228 e ST25) e SG28 (ST46 e ST149).
Os novos STs, ST191 e ST192 não apresentaram relação genética com nenhum ST
descrito e, portanto, foram classificados como “singletons”.
5.3. Perfil de sensibilidade aos antimicrobianos
O perfil de sensibilidade para os 14 antimicrobianos testados pela técnica de
microdiluição em caldo demonstrou que, os diferentes padrões de A. baumannii
produtores de OXA-23 circulantes no país apresentam fenótipo MDR, como mostrado
na Tabela 8. Embora as cefalosporinas de amplo espectro não sejam hidrolisadas
pelas CHDLs, os isolados brasileiros produtores de OXA-23 apresentam altas taxas de
resistência a esses antimicrobianos, com CIM50 variando de 128 µg/mL para cefepima
a 1024 µg/mL para ceftazidima. Entretanto, apesar da ceftazidima ser a cefalosporina
menos potente contra isolados brasileiros de A. baumannii foi a que apresentou as
menores taxas de resistência (48,4%), ainda que o número de isolados com
resistência intermediária tenha sido alto (38,7%). Como era esperado, as taxas de

RESULTADOS
66
resistência aos carbapenens foram altas (98,8% para imipenem e meropenem). Altas
taxas de resistência foram também observadas para as fluoroquinolonas, embora o
levofloxacino tenha sido oito vezes mais potente in vitro que o ciprofloxacino (CIM50, 16
µg/mL e CIM50, 128 µg/mL, respectivamente). Altos percentuais de resistência foram
detectados para ambos compostos (83,9% e 96,8%, respectivamente). Entre os
aminoglicosídeos, a gentamicina foi 16 vezes mais potente que a amicacina (CIM50, 8
µg/mL e CIM50, 128 µg/mL, respectivamente) e apresentou menores taxas de
resistência (41,9% e 74,2% respectivamente).
Tabela 8. Perfil de sensibilidade aos antimicrobianos entre os isolados de A.
baumannii produtores de OXA-23.
Antimicrobianos CIM (µg/mL) % por categoria
b
CIM50 CIM90 Intervalo S I R
β-lactâmicos
Ampicilina/Sulbactam 128/64 128/64 32/16 - 128/64 0 0 100
Ceftazidima 1024 >1024 8 - >1024 3,2 38,7 48,4
Cefotaxima 512 1024 16 - >1024 0 16,1 83,9
Cefepima 128 256 64 - 1024 0 0 100
Aztreonama 256 >256 16 - >256 - - -
Imipenem 32 64 8 - 64 0 3,2 96,8
Meropenem 32 64 8 - 64 0 3,2 96,8
Fluoroquinolonas
Ciprofloxacina 128 >256 0,5 - >256 3,2 0 96,8
Levofloxacina 16 32 ≤0,25 - 128 3,2 12,9 83,9
Aminoglicosídeos
Gentamicina 8 >1024 ≤0,5 - >1024 45,2 12,9 41,9
Amicacina 128 512 2 - >512 22,6 3,2 74,2
Tetraciclinas
Minociclina 0,125 0,5 0,06 - 4 100 0 0
Glicilciclinas
Tigeciclina 4 16 0,5 - 16 12,9 38,7 48,4
Lipopeptídeos
Polimixina B ≤0,25 0,5 ≤0,25 - 1 100 0 0
a. Não existem pontos de corte para aztreonam para isolados de A. baumannii (CLSI, 2012); b.
Legenda: S, sensível; I, intermediário e R, resistente.

RESULTADOS
67
Ampicilina/sulbactam, uma das poucas opções terapêuticas utilizadas no
tratamento de infecções por isolados de A. baumannii MDR, mostrou-se não ser ativa
frente aos isolados brasileiros produtores de OXA-23 (100% de resistência). O mesmo
foi observado para tigeciclina, onde somente 12,9% dos isolados de A. baumannii
avaliados neste estudo apresentaram sensibilidade a esse antimicrobiano (CIM50, 4
µg/mL). Dentre os 14 antimicrobianos testados, somente minociclina (CIM50, 0,125
µg/mL) e polimixina B (CIM50, ≤0,25 µg/mL) demonstraram ter excelente atividade
contra os isolados de A. baumannii produtores de OXA-23, que foram 100% sensíveis
a estes antimicrobianos.
5.4. Avaliação dos genes codificadores de β-lactamases e a relação
desses com a presença de ISAba1
A pesquisa dos principais genes codificadores de β-lactamases foi realizada
para todos os isolados de A. baumannii e os resultados apresentados na Tabela 9.
Não foram detectados genes codifidores de ESβLs nos isolados de A. baumannii
produtores de OXA-23 avaliados neste estudo. Somente foi encontrado o gene blaTEM-1
codificador de uma β-lactamase de espectro restrito em 14 isolados (45,2%),
principalmente nos isolados pertencentes ao ST79 (padrões de PFGE F, J, K, L e M) e
em apenas um isolado do ST228 (padrão B). O sequenciamento total do gene blaOXA-
23-like para todos os 31 isolados avaliados no presente estudo, demonstrou que a
variante OXA-23 continua sendo a CHDL mais prevalente nesta coleção, não tendo
sofrido mutações ao longo dos anos. Além disso, todos os padrões produtores de
OXA-23 avaliados neste estudo, apresentam a sequência de inserção ISAba1
associada ao gene blaOXA-23, aumentando, consequentemente, a sua expressão.
Apesar da grande maioria dos isolados de A. baumannii produtores de OXA-23
apresentarem CIMs altas para imipenem e meropenem (CIMs ≥ 16 µg/mL), o isolado
17.10 (padrão H) apresentou CIM de 8 µg/mL para esses antimicrobianos. Quando foi
comparado o conteúdo genético de -lactamases entre amostras que eram

RESULTADOS
68
geneticamente relacionadas, mas que apresentavam CIMs diferentes para os
carbapenens, nenhuma diferença foi encontrada.
Como esperado, o gene blaAmpC foi encontrado em todos os isolados avaliados
e a sua associação com a ISAba1 foi verificada em 80,6% dos casos (n=25/31). A
maioria dos isolados apresentando ISAba1 a montante do gene blaAmpC (23/25, 92%)
são os isolados incluídos nos principais complexos clonais disseminados em nosso
país, CC1, CC15 e CC79. Os seis isolados que não apresentaram ISAba1 associado a
essa cefalosporinase cromossômica eram representantes de padrões de PFGE
minoritários (padrão O/ST46, padrão C/ST188, padrão A/ST189, padrão P/ST190,
padrão N/ST191 e padrão E/ST192), encontrados principalmente na cidade de Porto
Alegre (Tabela 7). Embora o ST190 seja relacionado ao CC79, o isolado incluído
nesse ST não apresentou a ISAba1 associada ao gene blaAmpC. Entretanto, o padrão
D/ST162, incluído no CC79 e o seu padrão relacionado D1/ST299, um “singleton”,
apresentaram ISAba1 inserida a montante da AmpC, além do padrão B/ST228,
representante do SG21. Todos os isolados que apresentaram ISAba1 a montante do
gene blaAmpC apresentaram altos níveis de resistência às cefalosporinas de amplo
espectro (ceftazidima, CIMs de 32 a >1024 µg/mL; cefotaxima, CIMs de 256 a >1024
µg/mL). Ao contrário, os isolados cujo gene blaAmpC não estava associado a este
elemento de inserção, apresentaram CIMs menores (ceftazidima, CIMs de 8 a 16
µg/mL; cefotaxima, CIMs de 16 a 32 µg/mL), com exceção do isolado 16.31 (padrão
N/ST191) que apresentou CIMs elevadas para ambas às cefalosporinas (ceftazidima,
CIM 128 µg/mL; cefotaxima, CIM 64 µg/mL). Embora tenha sido observada uma
pequena diferença das CIMs para aztreonam, quando comparado os dois grupos para
as amostras que possuíam e não possuíam a ISAba1 a montante da AmpC, (CIMs de
64 a > 256 µg/mL e CIMs de 16 a 64 µg/mL), respectivamente, essa diferença foi mais
discreta quando comparadas com CIMs de ceftazidima e cefotaxima. As CIMs para
cefepima, a princípio, parecem não ter sido influenciadas pela presença ou ausência
de ISAba1.
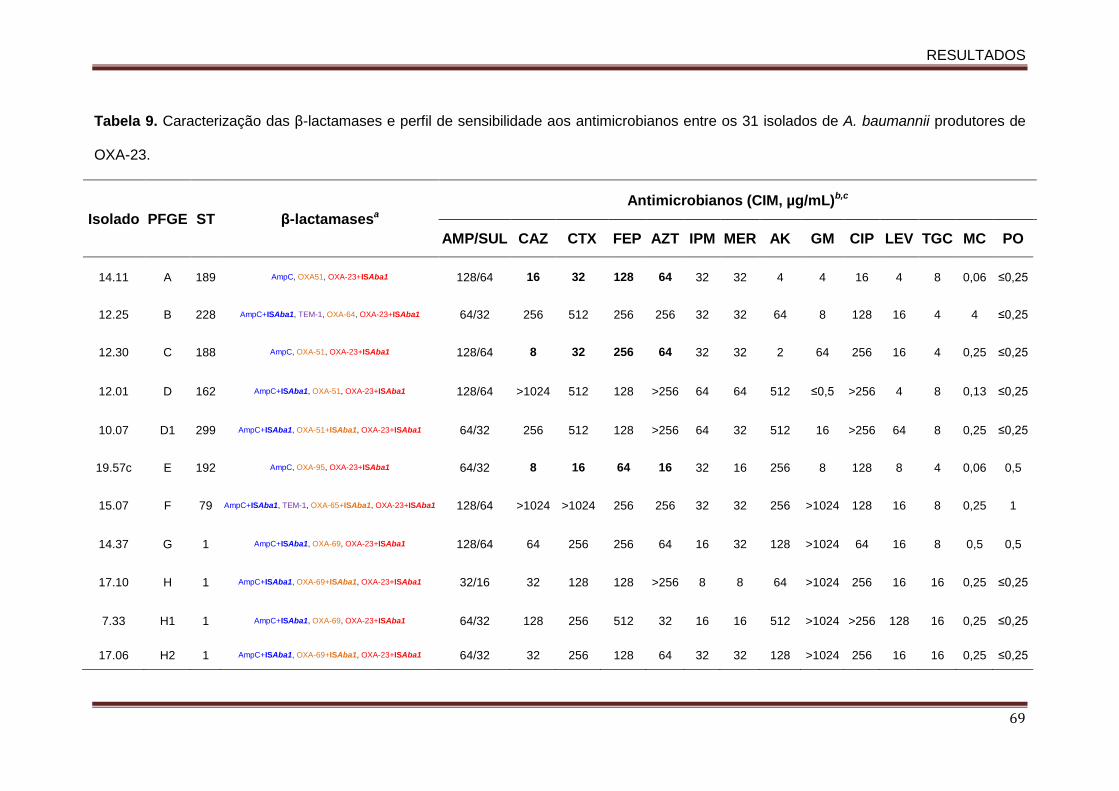
RESULTADOS
69
Tabela 9. Caracterização das β-lactamases e perfil de sensibilidade aos antimicrobianos entre os 31 isolados de A. baumannii produtores de
OXA-23.
Isolado PFGE ST β-lactamasesa
Antimicrobianos (CIM, µg/mL)b,c
AMP/SUL CAZ CTX FEP AZT IPM MER AK GM CIP LEV TGC MC PO
14.11 A 189 AmpC, OXA51, OXA-23+ISAba1 128/64 16 32 128 64 32 32 4 4 16 4 8 0,06 ≤0,25
12.25 B 228 AmpC+ISAba1, TEM-1, OXA-64, OXA-23+ISAba1 64/32 256 512 256 256 32 32 64 8 128 16 4 4 ≤0,25
12.30 C 188 AmpC, OXA-51, OXA-23+ISAba1 128/64 8 32 256 64 32 32 2 64 256 16 4 0,25 ≤0,25
12.01 D 162 AmpC+ISAba1, OXA-51, OXA-23+ISAba1 128/64 >1024 512 128 >256 64 64 512 ≤0,5 >256 4 8 0,13 ≤0,25
10.07 D1 299 AmpC+ISAba1, OXA-51+ISAba1, OXA-23+ISAba1 64/32 256 512 128 >256 64 32 512 16 >256 64 8 0,25 ≤0,25
19.57c E 192 AmpC, OXA-95, OXA-23+ISAba1 64/32 8 16 64 16 32 16 256 8 128 8 4 0,06 0,5
15.07 F 79 AmpC+ISAba1, TEM-1, OXA-65+ISAba1, OXA-23+ISAba1 128/64 >1024 >1024 256 256 32 32 256 >1024 128 16 8 0,25 1
14.37 G 1 AmpC+ISAba1, OXA-69, OXA-23+ISAba1 128/64 64 256 256 64 16 32 128 >1024 64 16 8 0,5 0,5
17.10 H 1 AmpC+ISAba1, OXA-69+ISAba1, OXA-23+ISAba1 32/16 32 128 128 >256 8 8 64 >1024 256 16 16 0,25 ≤0,25
7.33 H1 1 AmpC+ISAba1, OXA-69, OXA-23+ISAba1 64/32 128 256 512 32 16 16 512 >1024 >256 128 16 0,25 ≤0,25
17.06 H2 1 AmpC+ISAba1, OXA-69+ISAba1, OXA-23+ISAba1 64/32 32 256 128 64 32 32 128 >1024 256 16 16 0,25 ≤0,25
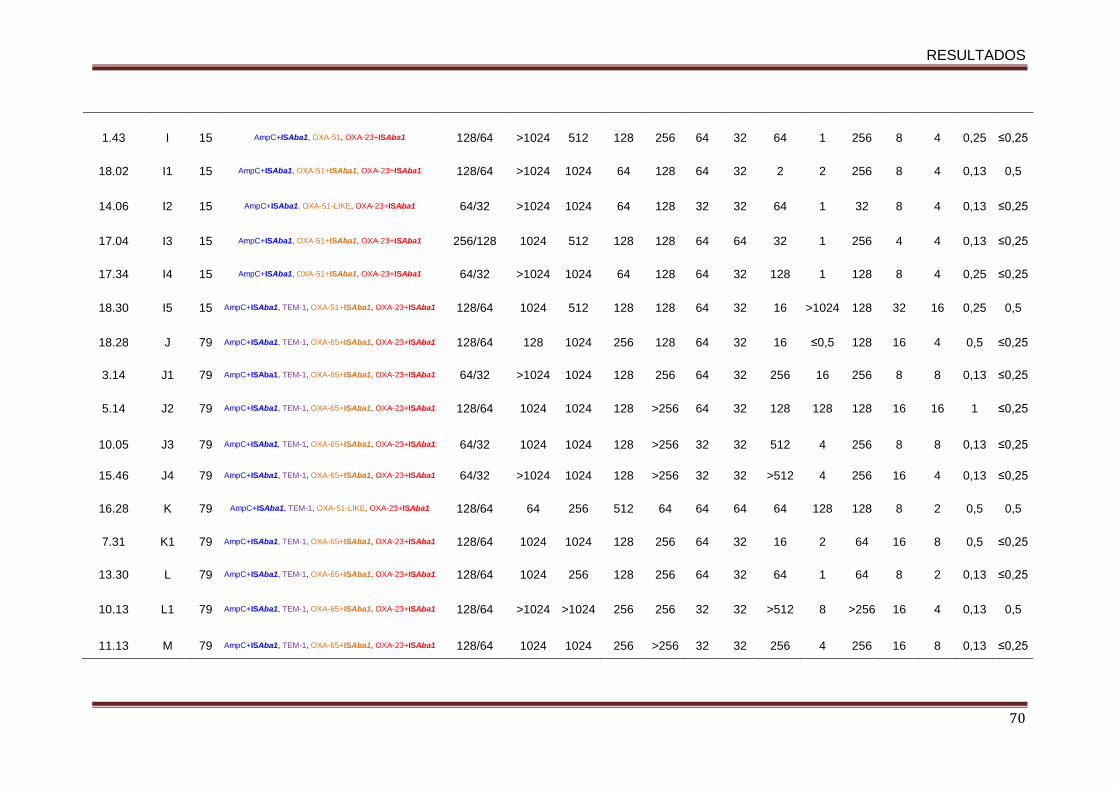
RESULTADOS
70
1.43 I 15 AmpC+ISAba1, OXA-51, OXA-23+ISAba1 128/64 >1024 512 128 256 64 32 64 1 256 8 4 0,25 ≤0,25
18.02 I1 15 AmpC+ISAba1, OXA-51+ISAba1, OXA-23+ISAba1 128/64 >1024 1024 64 128 64 32 2 2 256 8 4 0,13 0,5
14.06 I2 15 AmpC+ISAba1, OXA-51-LIKE, OXA-23+ISAba1 64/32 >1024 1024 64 128 32 32 64 1 32 8 4 0,13 ≤0,25
17.04 I3 15 AmpC+ISAba1, OXA-51+ISAba1, OXA-23+ISAba1 256/128 1024 512 128 128 64 64 32 1 256 4 4 0,13 ≤0,25
17.34 I4 15 AmpC+ISAba1, OXA-51+ISAba1, OXA-23+ISAba1 64/32 >1024 1024 64 128 64 32 128 1 128 8 4 0,25 ≤0,25
18.30 I5 15 AmpC+ISAba1, TEM-1, OXA-51+ISAba1, OXA-23+ISAba1 128/64 1024 512 128 128 64 32 16 >1024 128 32 16 0,25 0,5
18.28 J 79 AmpC+ISAba1, TEM-1, OXA-65+ISAba1, OXA-23+ISAba1 128/64 128 1024 256 128 64 32 16 ≤0,5 128 16 4 0,5 ≤0,25
3.14 J1 79 AmpC+ISAba1, TEM-1, OXA-65+ISAba1, OXA-23+ISAba1 64/32 >1024 1024 128 256 64 32 256 16 256 8 8 0,13 ≤0,25
5.14 J2 79 AmpC+ISAba1, TEM-1, OXA-65+ISAba1, OXA-23+ISAba1 128/64 1024 1024 128 >256 64 32 128 128 128 16 16 1 ≤0,25
10.05 J3 79 AmpC+ISAba1, TEM-1, OXA-65+ISAba1, OXA-23+ISAba1 64/32 1024 1024 128 >256 32 32 512 4 256 8 8 0,13 ≤0,25
15.46 J4 79 AmpC+ISAba1, TEM-1, OXA-65+ISAba1, OXA-23+ISAba1 64/32 >1024 1024 128 >256 32 32 >512 4 256 16 4 0,13 ≤0,25
16.28 K 79 AmpC+ISAba1, TEM-1, OXA-51-LIKE, OXA-23+ISAba1 128/64 64 256 512 64 64 64 64 128 128 8 2 0,5 0,5
7.31 K1 79 AmpC+ISAba1, TEM-1, OXA-65+ISAba1, OXA-23+ISAba1 128/64 1024 1024 128 256 64 32 16 2 64 16 8 0,5 ≤0,25
13.30 L 79 AmpC+ISAba1, TEM-1, OXA-65+ISAba1, OXA-23+ISAba1 128/64 1024 256 128 256 64 32 64 1 64 8 2 0,13 ≤0,25
10.13 L1 79 AmpC+ISAba1, TEM-1, OXA-65+ISAba1, OXA-23+ISAba1 128/64 >1024 >1024 256 256 32 32 >512 8 >256 16 4 0,13 0,5
11.13 M 79 AmpC+ISAba1, TEM-1, OXA-65+ISAba1, OXA-23+ISAba1 128/64 1024 1024 256 >256 32 32 256 4 256 16 8 0,13 ≤0,25

RESULTADOS
71
12.06 M1 79 AmpC+ISAba1, TEM-1, OXA-65+ISAba1, OXA-23+ISAba1 256/128 >1024 >1024 1024 >256 32 32 256 8 >256 8 8 0,13 ≤0,25
16.31 N 191 AmpC, OXA-51, OXA-23+ISAba1 128/64 128 64 128 256 64 32 256 ≤0,5 128 8 4 0,13 0,5
16.11 O 46 AmpC, OXA-318, OXA-23+ISAba1 64/32 8 32 64 32 64 64 128 32 0,5 ≤0,25 1 0,13 ≤0,25
16.19 P 190 AmpC, OXA-242, OXA-23+ISAba1 64/32 8 32 64 32 32 32 4 16 16 4 0,5 0,06 ≤0,25
a. Em azul estão incluídas as cefalosporinases cromossômicas do tipo AmpC (ADC) intrínseca de A. baumannii, em roxo estão incluídas as β-lactamases de
espectro restrito do tipo TEM-1, em laranja estão incluídas as CHDLs cromossômicas e intrínsecas de A. baumannii pertencentes ao grupo das OXA-51, em
vermelho estão incluídas as CHDLs adquiridas de A. baumannii pertencentes ao grupo das OXA-23, a presença de ISAba1 adiante das β-lactamases do tipo
AmpC, OXA-51-like e OXA-23 estão assinaladas em negrito; b. Legenda: AMP/SUL, ampicilina/Sulbactam; CAZ, ceftazidima; CTX, ceftriaxona; FEP,
cefepima; AZT, aztreonam; IPM, imipenem; MER, meropenem; AK, amicacina; GM, gentamicina; CIP, ciprofloxacina; LEV, levofloxacina; TGC, tigeciclina;
MC, minociclina; PO, polimixina B; c. As CIMs para as cefalosporinas de amplo espectro em negrito representam os isolados que não possuem ISAba1 a
montante do gene blaAmpC.

RESULTADOS
72
Um total de sete variantes diferentes de oxacilinases pertencentes ao grupo
das CHDLs cromossômicas do tipo OXA-51 e intrínsecas de A. baumannii foram
verificadas nos 31 isolados avaliados no estudo, sendo elas: OXA-51, OXA-64, OXA-
65, OXA-69, OXA-95, OXA-242 e OXA-318. Duas outras variantes do grupo OXA-51,
encontradas nos isolados 14.06 (padrão I2/ST15) e 16.28 (padrão K/ST79) ainda não
tiveram a sua nomenclatura definitiva determinada e, portanto, foram referidas como
OXA-51-like. Novos sequenciamentos estão sendo realizados no momento, para
confirmar a caracterização de uma nova variante presente nesses isolados. De um
modo geral, isolados de A. baumannii pertencentes a um mesmo ST, tenderam a
apresentar a mesma variante de OXA-51, como a OXA-51 no ST15, a OXA-65 no
ST79 e a OXA-69 no ST1. O mesmo não ocorreu entre amostras que apresentavam
STs incluídos em um mesmo complexo clonal, como é o caso do ST162 e do ST189,
incluídos no CC79, e cujos isolados apresentaram a variante OXA-51, se diferindo do
ST79 que apresenta a variante OXA-65. Da mesma forma, o isolado que apresentava
o ST190, também incluído no CC79, carreia uma nova variante nomeada OXA-242.
Para os demais isolados que apresentaram STs diferentes formando SGs ou não
(“singletons”), apresentaram, de um modo geral, variantes diferentes daquelas
verificadas nos CCs, como a OXA-64 (padrão B/ST228/SG21), a OXA-95 (padrão
E/ST192/“singleton”) e a nova variante OXA-318 (padrão O/ST46/SG28). A presença
de ISAba1 a montante das CHDLs do grupo da OXA-51 foi mais relacionada aos
complexos clonais, principalmente a OXA-65 nos isolados apresentando o ST79. Por
outro lado, não foi identificada ISAba1 a montante do gene blaOXA-51-like nas amostras
de A. baumannii incluídas em STs minoritários.
5.5. Avaliação fenotípica da hiperexpressão dos sistemas de efluxo
A avaliação fenotípica do impacto dos sistemas de efluxo na resistência às
cefalosporinas de amplo espectro e aos carbapenens foi realizada através da
avaliação da redução das CIMs para esses antimicrobianos, na presença do inibidor

RESULTADOS
73
de sistemas de efluxo PAβN. Os resultados obtidos estão incluídos na Tabela 10. No
presente estudo, não foi considerada significativa a diminuição de apenas uma diluição
na CIM para qualquer um dos quatro β-lactâmicos testados na presença do inibidor, já
que tal diferença entre dois testes dilucionais poderia ser consequente à variabilidade
inter testes intrínseca à técnica laboratorial. Em 17/31 (54,8%) isolados de A.
baumannii apresentaram diminuição da CIM para ceftazidima ≥ 2 diluições na
presença do PAβN, principalmente naqueles isolados apresentando altos níveis de
resistência (CIM ≥ 1024 µg/mL). Por outro lado, os isolados sensíveis ou com
resistência intermediária à ceftazidima (CIM ≤ 16 µg/mL), não apresentaram
diminuição significativa na CIM na presença do PAβN. O isolado 16.31 que
apresentava altos CIMs para ceftazidima e cefotaxima no teste com o inibidor,
apresentou diminuição da CIM para ceftazidima de 128 µg/mL para 16 µg/mL. De
modo contrário, os sistemas de efluxo parecem não ter impacto na ejeção da
cefotaxima, já que somente quatro amostras apresentaram diminuição significativa da
CIM, podendo outros mecanismos de resistência, em conjunto, estarem associados ao
aumento da CIM, ou até mesmo mecanismos desconhecidos.
Resultados semelhantes também foram observados quando se avaliou o
impacto dos sistemas de efluxo na elevação das CIMs para imipenem e meropenem.
Para imipenem, uma diminuição significativa foi observada somente em 9/31 isolados
(29%), demonstrando um papel secundário na resistência a esse antimicrobiano em
isolados de A. baumannii produtores de OXA-23. Por outro lado, os sistemas de efluxo
demonstram ser importantes na elevação da CIMs para meropenem em isolados de A.
baumannii com esse fenótipo, já que foi observada uma dimuição significativa na CIMs
em 80,6% (25/31) dos isolados testados. Desses, 18 isolados deixaram de ser
resistentes e passaram a apresentar resistência intermediária (CIM, 8 µg/mL) e quatro
se tornaram sensíveis ao meropenem (CIM, 4 µg/mL). Dentre os nove isolados que
apresentaram diminuição significativa na CIM para imipenem, apenas dois mudaram

RESULTADOS
74
de categoria de sensibilidade, apresentando resistência intermediária a esse
antimicrobiano (CIM, 8 µg/mL).
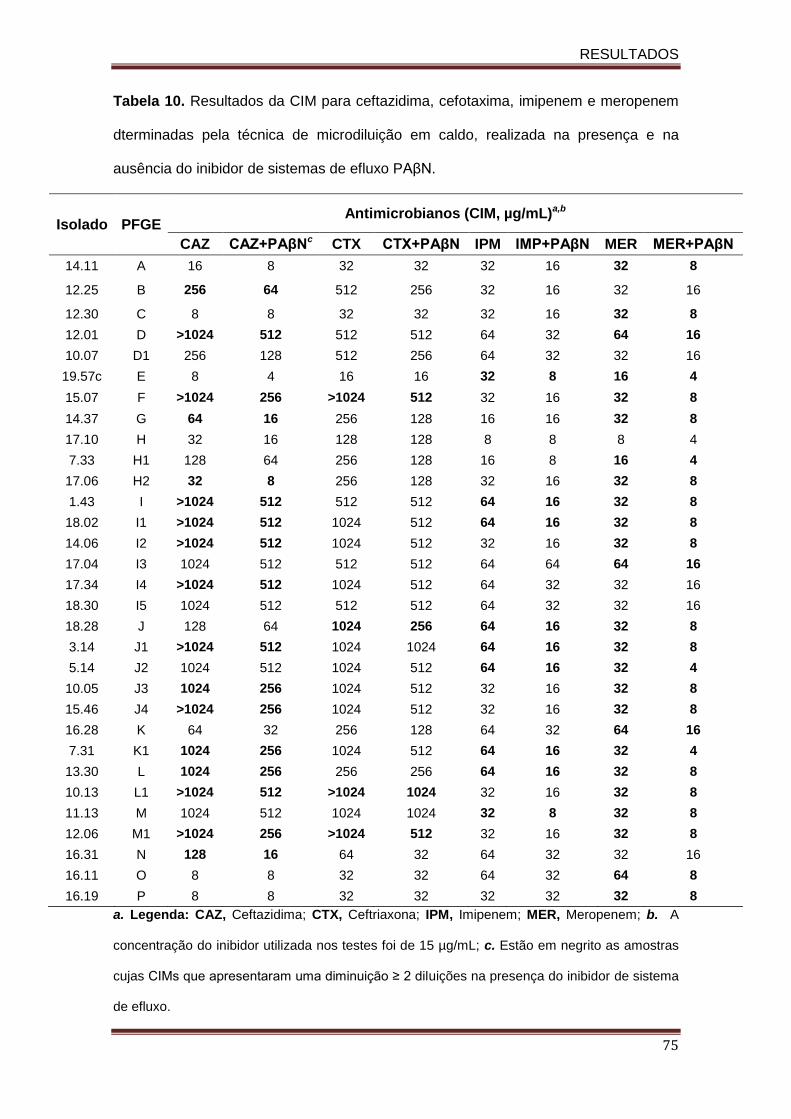
RESULTADOS
75
Tabela 10. Resultados da CIM para ceftazidima, cefotaxima, imipenem e meropenem
dterminadas pela técnica de microdiluição em caldo, realizada na presença e na
ausência do inibidor de sistemas de efluxo PAβN.
Isolado PFGE Antimicrobianos (CIM, µg/mL)a,b
CAZ CAZ+PAβNc CTX CTX+PAβN IPM IMP+PAβN MER MER+PAβN
14.11 A 16 8 32 32 32 16 32 8
12.25 B 256 64 512 256 32 16 32 16
12.30 C 8 8 32 32 32 16 32 8
12.01 D >1024 512 512 512 64 32 64 16
10.07 D1 256 128 512 256 64 32 32 16
19.57c E 8 4 16 16 32 8 16 4
15.07 F >1024 256 >1024 512 32 16 32 8
14.37 G 64 16 256 128 16 16 32 8
17.10 H 32 16 128 128 8 8 8 4
7.33 H1 128 64 256 128 16 8 16 4
17.06 H2 32 8 256 128 32 16 32 8
1.43 I >1024 512 512 512 64 16 32 8
18.02 I1 >1024 512 1024 512 64 16 32 8
14.06 I2 >1024 512 1024 512 32 16 32 8
17.04 I3 1024 512 512 512 64 64 64 16
17.34 I4 >1024 512 1024 512 64 32 32 16
18.30 I5 1024 512 512 512 64 32 32 16
18.28 J 128 64 1024 256 64 16 32 8
3.14 J1 >1024 512 1024 1024 64 16 32 8
5.14 J2 1024 512 1024 512 64 16 32 4
10.05 J3 1024 256 1024 512 32 16 32 8
15.46 J4 >1024 256 1024 512 32 16 32 8
16.28 K 64 32 256 128 64 32 64 16
7.31 K1 1024 256 1024 512 64 16 32 4
13.30 L 1024 256 256 256 64 16 32 8
10.13 L1 >1024 512 >1024 1024 32 16 32 8
11.13 M 1024 512 1024 1024 32 8 32 8
12.06 M1 >1024 256 >1024 512 32 16 32 8
16.31 N 128 16 64 32 64 32 32 16
16.11 O 8 8 32 32 64 32 64 8
16.19 P 8 8 32 32 32 32 32 8
a. Legenda: CAZ, Ceftazidima; CTX, Ceftriaxona; IPM, Imipenem; MER, Meropenem; b. A
concentração do inibidor utilizada nos testes foi de 15 µg/mL; c. Estão em negrito as amostras
cujas CIMs que apresentaram uma diminuição ≥ 2 diluições na presença do inibidor de sistema
de efluxo.

RESULTADOS
76
5.6. Avaliação fenotípica da alteração das proteínas de membrana externa
pela técnica de SDS-PAGE
O perfil de proteínas de membrana externa para os 31 isolados de A.
baumannii produtores de OXA-23 avaliados nesse estudo, estão demonstrados na
Tabela 11, baseando-se na análise visual dos géis de SDS-PAGE incluídos na Figura
9. Os resultados obtidos demonstraram uma alta variabilidade no perfil de proteínas de
membrana externa entre os isolados clínicos de A. baumannii produtores de OXA-23
no Brasil. Ainda que essa variação tenha sido independente do padrão genético
apresentado pelo isolado, foi possível observar que a maioria dos isolados (28/31) não
apresentaram uma banda de 21 kDa compatível com a OmpW. As proteínas de 29
kDa e 31 kDa, compatíveis com as porinas CarO e Omp33-36 relacionadas à
resistência aos carbapenens, apresentaram perfis variáveis. O isolado 17.10 (marcado
em cinza na tabela) foi o único isolado a apresentar todas as bandas com tamanhos
de 21 kDa, 29 kDa, 31 kDa, 35,6 kDa e 42 kDa, compatíveis com as OMPs OmpW,
CarO, Omp33-36, OmpHMP e OprD, respectivamente. Coincidentemente, esse
isolado apresentava resistência intermediária a imipenem e meropenem (CIMs de 8
µg/mL).

RESULTADOS
77
Tabela 11. Perfil das proteínas de membrana externa dos isolados de A. baumannii
produtores de OXA-23 baseando nos géis de SDS-PAGE.
Isolado PFGE
CIM (µg/mL)
Proteínas de Membrana Externaa
- OmpWc Omp25 CarO 33-36 OmpHMP OprD
IPM MER 18 kDab 21 kDa 25 kDa 29 kDa 31 kDa 35,6 kDa 42 kDa
14.11 A 32 32 (-) (+) (+) (-) (-) (+) (-)
12.25 B 32 32 (+) (+) (+) (-) (+) (+) (+)
12.30 C 32 32 (+) (-) (+) (-) (+) (+) (+)
12.01 D 64 64 (+) (-) (+) (-) (+) (+) (+)
10.07 D1 64 32 (+) (-) (-) (+) (+) (+) (-)
19.57c E 32 16 (+) (-) (-) (+) (+) (+) (-)
15.07 F 32 32 (+) (-) (-) (+) (+) (+) (+)
14.37 G 16 32 (+) (-) (-) (+) (+) (+) (+)
17.10 H 8 8 (+) (+) (+) (+) (+) (+) (+)
7.33 H1 16 16 (+) (-) (+) (-) (+) (+) (+)
17.06 H2 32 32 (+) (-) (+) (+) (+) (+) (+)
1.43 I 64 32 (+) (-) (+) (+) (+) (+) (+)
18.02 I1 64 32 (+) (-) (+) (+) (+) (+) (+)
14.06 I2 32 32 (+) (-) (+) (+) (+) (+) (+)
17.04 I3 64 64 (-) (-) (+) (-) (-) (+) (-)
17.34 I4 64 32 (-) (-) (+) (-) (-) (+) (-)
18.30 I5 64 32 (-) (-) (+) (-) (-) (+) (-)
18.28 J 64 32 (+) (-) (-) (+) (+) (+) (+)
3.14 J1 64 32 (+) (-) (-) (+) (+) (+) (+)
5.14 J2 64 32 (+) (-) (-) (+) (+) (+) (+)
10.05 J3 32 32 (+) (-) (-) (+) (+) (+) (+)
15.46 J4 32 32 (+) (-) (+) (-) (+) (+) (+)
16.28 K 64 64 (+) (-) (-) (+) (+) (+) (+)
7.31 K1 64 32 (+) (-) (+) (+) (+) (+) (+)
13.30 L 64 32 (+) (-) (-) (+) (+) (+) (+)
10.13 L1 32 32 (+) (-) (-) (+) (+) (+) (+)
11.13 M 32 32 (+) (-) (+) (+) (+) (+) (+)
12.06 M1 32 32 (+) (-) (+) (+) (+) (+) (+)
16.31 N 64 32 (+) (-) (+) (+) (+) (+) (+)
16.11 O 64 64 (+) (-) (+) (+) (+) (+) (+)
16.19 P 32 32 (+) (-) (-) (+) (+) (+) (-)
ATCC 19606 - - - (+) (+) (+) (+) (+) (+) (+)
a. Legenda: (-) - ausência de banda no gel, (+) - presença de banda no gel com expressão
semelhante à ATCC, (+), - presença de banda no gel com expressão inferior à ATCC, (+)-
presença de banda no gel com tamanho inferior a ATCC. b. Os pesos em kDa para cada OMP
foram estimados baseados no padrão de peso molecular aplicada em cada gel separadamente.
c. prováveis OMPs correspondentes ao tamanho da proteína encontrada no gel de SDS-PAGE.
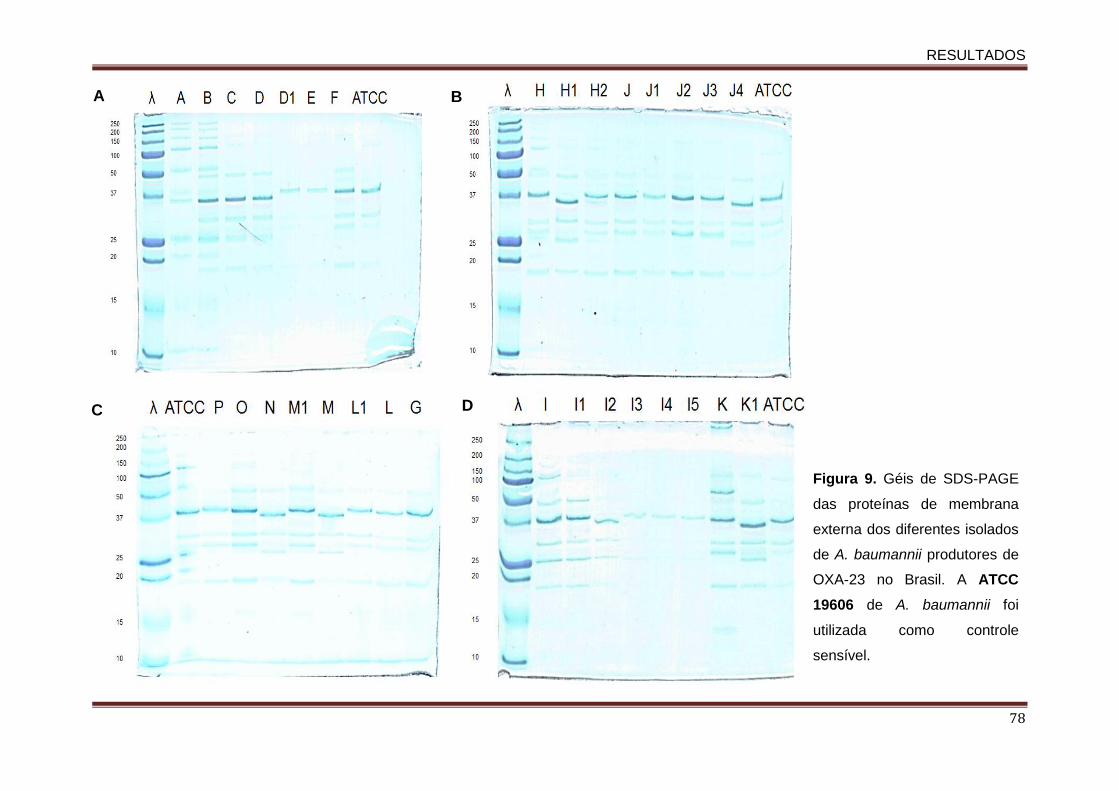
RESULTADOS
78
A B
C D
Figura 9. Géis de SDS-PAGE
das proteínas de membrana
externa dos diferentes isolados
de A. baumannii produtores de
OXA-23 no Brasil. A ATCC
19606 de A. baumannii foi
utilizada como controle
sensível.

RESULTADOS
79
Outro resultado importante foi a alteração no tamanho das bandas das
proteínas de 25 kDa e 35,6 kDa, compatíveis com as OMPs Omp25 e OmpHMP,
proteínas que não estão relacionadas à resistência aos carbapenens, em dois e dez
isolados de A. baumannii, respectivamente.
5.7. Quantificação da expressão dos genes codificadores de porinas e do
sistema de efluxo AdeABC pela técnica de qRT-PCR
As amostras de A. baumannii foram submetidas à metodologia de qRT-PCR
para a quantificação da expressão dos genes ompW, carO, omp33-36, ompHMP e
adeB. Os valores dos níveis de expressão foram obtidos comparando com a cepa
ATCC BAA-1709 de A. baumannii, que apresenta sensibilidade a todos os β-
lactâmicos, e estão incluídos na Tabela 12. De acordo com os resultados obtidos, a
grande maioria do isolados (23/31, 74,2%) de A. baumannii produtores de OXA-23 no
Brasil, hiperexpressam o sistema de efluxo AdeABC, quando comparados à cepa
ATCC BAA-1709. Embora os níveis de expressão tenham variado de acordo com o
padrão de PFGE, foi possível observar que os padrões que estão incluídos nos
complexos clonais CC79, CC15 e CC1 tendem a hiperexpressar mais esse o sistema,
principalmente os isolados pertencentes ao ST1. Apesar disso, não foi possível
correlacionar o aumento da expressão desse sistema de efluxo a um fenótipo de
resistência. Por outro lado, os resultados do qRT-PCR demonstraram que quase todos
os isolados clínicos (30/31, 96,8%) expressavam menos a porina CarO, quando
comparados a cepa ATCC. Resultado semelhante foi observado para a OmpHMP e a
Omp33-36 em 14 e 15 dos 31 (45,2%) isolados clínicos avaliados respectivamente.
Entretanto, a diminuição da OmpW somente foi observada em quatro isolados
(12,9%).
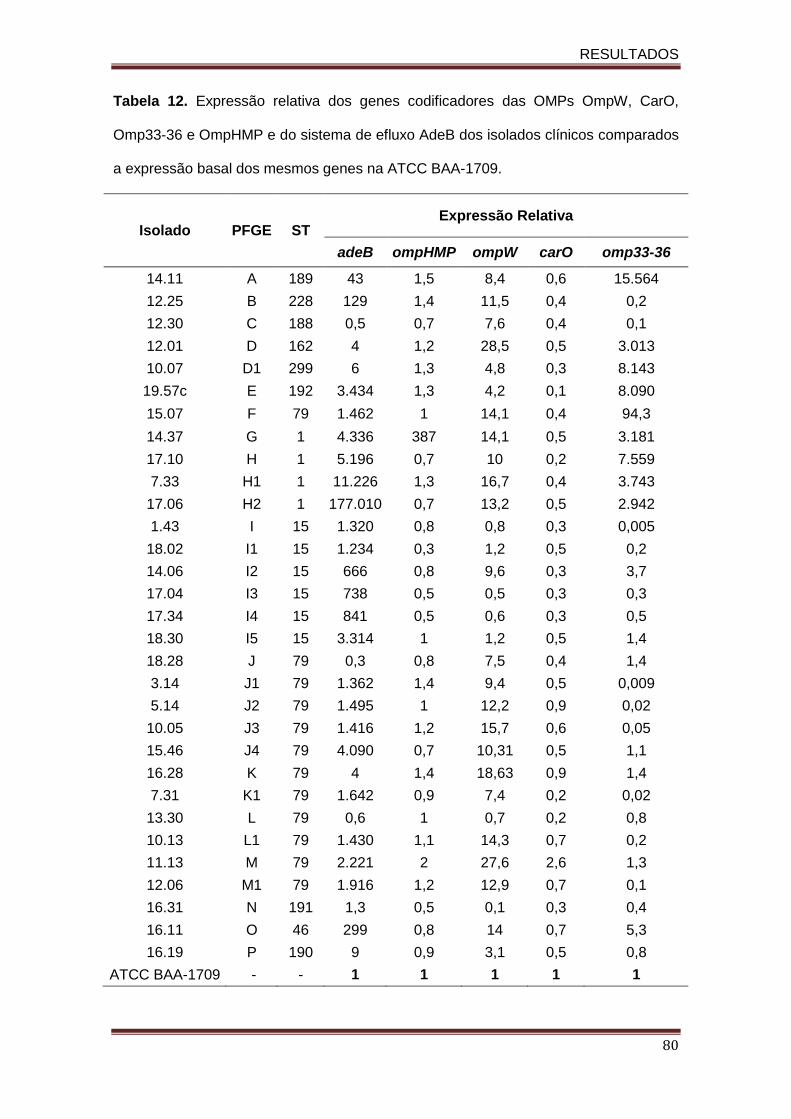
RESULTADOS
80
Tabela 12. Expressão relativa dos genes codificadores das OMPs OmpW, CarO,
Omp33-36 e OmpHMP e do sistema de efluxo AdeB dos isolados clínicos comparados
a expressão basal dos mesmos genes na ATCC BAA-1709.
Isolado PFGE ST Expressão Relativa
adeB ompHMP ompW carO omp33-36
14.11 A 189 43 1,5 8,4 0,6 15.564
12.25 B 228 129 1,4 11,5 0,4 0,2
12.30 C 188 0,5 0,7 7,6 0,4 0,1
12.01 D 162 4 1,2 28,5 0,5 3.013
10.07 D1 299 6 1,3 4,8 0,3 8.143
19.57c E 192 3.434 1,3 4,2 0,1 8.090
15.07 F 79 1.462 1 14,1 0,4 94,3
14.37 G 1 4.336 387 14,1 0,5 3.181
17.10 H 1 5.196 0,7 10 0,2 7.559
7.33 H1 1 11.226 1,3 16,7 0,4 3.743
17.06 H2 1 177.010 0,7 13,2 0,5 2.942
1.43 I 15 1.320 0,8 0,8 0,3 0,005
18.02 I1 15 1.234 0,3 1,2 0,5 0,2
14.06 I2 15 666 0,8 9,6 0,3 3,7
17.04 I3 15 738 0,5 0,5 0,3 0,3
17.34 I4 15 841 0,5 0,6 0,3 0,5
18.30 I5 15 3.314 1 1,2 0,5 1,4
18.28 J 79 0,3 0,8 7,5 0,4 1,4
3.14 J1 79 1.362 1,4 9,4 0,5 0,009
5.14 J2 79 1.495 1 12,2 0,9 0,02
10.05 J3 79 1.416 1,2 15,7 0,6 0,05
15.46 J4 79 4.090 0,7 10,31 0,5 1,1
16.28 K 79 4 1,4 18,63 0,9 1,4
7.31 K1 79 1.642 0,9 7,4 0,2 0,02
13.30 L 79 0,6 1 0,7 0,2 0,8
10.13 L1 79 1.430 1,1 14,3 0,7 0,2
11.13 M 79 2.221 2 27,6 2,6 1,3
12.06 M1 79 1.916 1,2 12,9 0,7 0,1
16.31 N 191 1,3 0,5 0,1 0,3 0,4
16.11 O 46 299 0,8 14 0,7 5,3
16.19 P 190 9 0,9 3,1 0,5 0,8
ATCC BAA-1709 - - 1 1 1 1 1

DISCUSSÃO
81
6. DISCUSSÃO
Desde a sua primeira descrição na Escócia em 1985 (Scaife et al., 1995), a
CHDL OXA-23 tornou-se um dos principais mecanismos de resistência aos β-
lactâmicos em isolados clínicos de A. baumannii (Mugnier et al., 2010), inclusive surtos
causados por esses micro-organismos têm sido reportados em várias partes do mundo
(Zhou et al., 2007; Calhoun et al., 2008; Stoeva et al., 2008; Carvalho et al., 2009). Um
estudo conduzido por Mugnier e colaboradores (2010) que avaliou cepas de A.
baumannii produtoras de OXA-23 isoladas em diferentes países, demonstrou que o
gene blaOXA-23 podia estar inserido em diferentes contextos genéticos, que incluem os
transposons Tn2007, Tn2008 e, principalmente, o Tn2006, além da ISAba1. Eles
também observaram que o gene blaOXA-23 associado ao Tn2006, pode estar localizado
tanto no cromossomo como em plasmídeos. Os autores concluíram que a alta
variabilidade genética observada entre os isolados de A. baumannii produtores de
OXA-23 pode dificultar o controle da sua disseminação inter e intra-hospitalar.
No Brasil, a OXA-23 é a principal carbapenemase encontrada em isolados de
A. baumannii e a sua disseminação pelos hospitais brasileiros alcançou um nível
epidêmico. A primeira descrição dessa CHDL no Brasil ocorreu, em 2003, na cidade
de Curitiba (Dalla-Costa et al., 2003). Nesse estudo os autores avaliaram oito
amostras clínicas de A. baumannii resistentes aos carbapenens isoladas em dois
hospitais da capital paranaense, em 1999. Após a realização do PFGE, verificou-se a
presença de um mesmo clone e a presença de OXA-23, que até aquele momento não
havia sido detectada em nosso país. Desde então, este clone vem se disseminando e
se diversificando pelo Brasil (Carvalho et al., 2009; Martins et al., 2009; Corrêa et al.,
2012).
Em 2009, Carvalho e colaboradores realizaram um estudo no qual avaliaram
96 amostras de A. baumannii resistentes ao imipenem coletadas em oito hospitais do
Rio de Janeiro, no período de 2006 a 2007. Todos os isolados eram produtores de

DISCUSSÃO
82
OXA-23 e diferente do estudo de Dalla-Costa e colaboradores (2003), foram
observados a presença de cinco clones. No mesmo ano, Martins e colaboradores
(2009) avaliaram um surto ocorrido em 2007, na cidade de Porto Alegre causado por
isolados de A. baumannii produtores de OXA-23. Os autores do estudo também
constataram uma maior variabilidade genética entre as 53 amostras avaliadas (cinco
padrões de PFGE). Esses dois estudos são importantes, porque demonstraram que a
OXA-23 era a carbapenemase mais frequente e disseminada entre isolados de A.
baumannii no Brasil. Embora em estudo anterior tivesse descrito a presença da MβL
IMP-1 em isolados de A. baumannii em um hospital da cidade São Paulo (Gales et al.,
2003), posteriormente ficou demonstrado que a presença dessa MβL estava restrita
aos hospitais da grande São Paulo (Sader et al., 2005; Tognim et al., 2006; Mendes et
al., 2007a).
Em 2010, Schimith e colaboradores (2010) avaliaram a evolução temporal de
isolados de A. baumannii produtores de OXA-23 na cidade de Curitiba. Este estudo
demonstrou que o clone inicialmente descrito, em 1999, por Dalla-Costa e
colaboradores (2003), persistiu no hospital de Curitiba até 2004, quando então, foi
substituído por outros dois clones que prevaleceram nos anos seguintes. No mesmo
estudo, foi verificada uma alta mortalidade (45,2%) entre os pacientes que
apresentaram infecções por isolados de A. baumannii produtores de OXA-23,
principalmente nos casos de pneumonia relacionada à ventilação mecânica (Schimith
et al., 2010). Resultados similares foram obtidos por Carneiro e colaboradores (2010)
que avaliaram 172 amostras de A. baumannii isolados em um hospital de Londrina. Os
autores também observaram uma alta frequência de pneumonia relacionada à
ventilação mecânica associada a altas taxas de mortalidade (52,9%). No mesmo
estudo, ficou demonstrado o papel do ambiente nosocomial na disseminação de cepas
MDR de A. baumannii, uma vez que foi comprovado pela análise do PFGE que o
padrão genético das cepas isoladas dos leitos da UTI era idêntico às cepas que
estavam causando infecção nos pacientes internados na mesma unidade hospitalar.

DISCUSSÃO
83
Os dados obtidos com os estudos de 2009 e 2010 demonstraram uma mudança
drástica no perfil clonal dos isolados brasileiros de A. baumannii produtores de OXA-
23 (Carvalho et al., 2009; Martins et al., 2009; Carneiro et al., 2010; Schimith et al.,
2010), o que contribuiria para o sucesso desse patógeno nos hospitais do nosso país.
Em 2011, Furtado e colaboradores também relataram altas taxas de
mortalidade (61,9%) entre os casos de infecção causada por isolados de A. baumannii
produtores de OXA-23 em uma UTI de um hospital da cidade de São Paulo, inclusive
superiores àquelas descritas por Schimith e colaboradores (2010). No mesmo ano,
Ferreira e colaboradores (2011) relataram a presença de OXA-23 em isolados de A.
baumannii recuperados de águas residuais de três hospitais localizados em Porto
Alegre, demonstrando o papel do esgoto hospitalar na perpetuação de patógenos e
genes de resistência no meio ambiente, os quais poderiam se estabelecer como
reservatórios naturais. No mesmo ano, dois estudos demonstraram que a dinâmica da
resistência aos carbapenens no Brasil era mais complexa do que se imaginava
(Antonio et al., 2011; Werneck et al., 2011b). No estudo de Antonio e colaboradores
(2011), foi relatada a alta prevalência de OXA-143 em isolados de A. baumannii no
estado de São Paulo, demonstrando que a OXA-23 não era a única CHDL prevalente
em nosso país. Até esse momento, pouco se sabia sobre a frequência de OXA-143 no
Brasil, mesmo essa CHDL tendo sido descrita dois anos antes em isolados de
Acinetobacter spp. coletados de uma UTI brasileira, em 2004 (Higgins et al., 2009). Já
no estudo conduzido por Werneck e colaboradores (2011b), foi questionado se a
prevalência de OXA-143 poderia ser maior em hospitais públicos da cidade de São
Paulo do que em hospitais privados, como verificado por Antonio e colaboradores
(2011). Os autores também demonstram que em hospitais privados, a OXA-23 era a
carbapenemase mais frequente, e encontrava-se disseminada em isolados brasileiros
de A. baumannii. Estes autores também reportaram que o perfil genético destes
isolados era altamente diversificado, superior inclusive ao verificado nos estudos
publicados anteriormente.

DISCUSSÃO
84
Embora pouco se saiba sobre a prevalência de OXA-143 em muitos estados
brasileiros, estudos recentes demonstram que essa CHDL está restrita à região
sudeste do país, principalmente nos estados de São Paulo e Minas Gerais (Mostachio
et al., 2012). Em um estudo retrospectivo, em que foram avaliadas amostras de
Acinetobacter spp. coletadas por um período de 18 anos em um hospital da cidade de
São Paulo, demonstrou que a OXA-143 era mais antiga do que se pensava, tendo sido
encontrada em um isolado de 1995 (Cayô et al., 2011a). O mais interessante é que o
primeiro isolado produtor de OXA-23 descrito no estudo somente foi encontrado em
2003, quase uma década após o aparecimento da OXA-143 na instituição. Apesar
disso, os isolados produtores de OXA-23 se disseminaram rapidamente nos anos que
se seguiram, tornando-se a principal carbapenemase responsável pela resistência aos
carbapenens naquela instituição (Cayô et al., 2011a). Esses dados mostraram a
grande adaptabilidade dos isolados produtores de OXA-23 ao ambiente nosocomial, o
que em parte, justifica seu sucesso em manterem-se por longos períodos neste
ambiente e causando surtos de difícil controle. Estudos mais recentes comprovam a
alta prevalência de OXA-23 em isolados de A. baumannii resistentes aos carbapenens
em diferentes estados no Brasil (Corrêa et al., 2012; Gales et al., 2012; Martins AF et
al., 2012; Martins N et al., 2012).
No presente estudo, foram avaliados 31 isolados de A. baumannii produtores
de OXA-23 provenientes de 14 hospitais privados localizados em sete estados
brasileiros e representativos dos principais padrões genéticos e padrões relacionados
descritos no estudo de Werneck e colaboradores (2011b). Os 16 padrões de PFGE e
15 padrões relacionados demonstraram o alto grau de diversificação que os isolados
brasileiros de A. baumannii produtores de OXA-23 alcançaram, bem diferente daquele
perfil monoclonal descrito 10 anos antes por Dalla-Costa e colaboradores (2003). A
análise do padrão genético desses isolados, nos mostra que em um mesmo hospital,
vários clones podem estar circulando ao mesmo tempo. Além disso, isolados
relacionados clonalmente foram encontrados em diferentes hospitais, o que demonstra

DISCUSSÃO
85
uma transmissão intra-hospitalar e uma evolução subsequente desses clones, de
forma distinta, em cada instituição. Essa grande variabilidade clonal entre os isolados
de A. baumannii produtores de OXA-23 tem sido relatada por diferentes estudos em
vários hospitais brasileiros nos últimos quatro anos (Carvalho et al., 2009; Martins et
al., 2009; Carneiro et al., 2010; Schimith et al., 2010; Ferreira et al., 2011; Furtado et
al., 2011).
Atualmente, duas metodologias de MLST são utilizadas para a avaliação da
ancestralidade de cepas de A. baumannii, se diferenciando basicamente em alguns
genes utilizados (Bartual et al., 2005; Nemec et al., 2008). A grande dificuldade na
interpretação dos resultados do MLST para A. baumannii está no fato de que cada
website curador responsável pelos respectivos bancos de dados nomeiam os STs de
forma distinta. Apesar disso, os STs obtidos podem ser correlacionados, e a melhor
maneira de conseguir essa correlação, é por meio do sequenciamento dos dois
esquemas de genes para ambas as metodologias. O estudo de Stietz e colaboradores
(2013) correlaciona os principais complexos clonais descritos pelas duas metodologias
e disseminados no mundo, demonstrando que o CC92, CC113 e CC103 de acordo
com Bartual e colaboradores (2005), correspondem aos complexos clonais CC2, CC79
e CC15 de Nemec e colaboradores (2008), respectivamente.
Para auxiliar na compreensão do mapa de filogenia, apresentado na Figura 8,
alguns conceitos importantes serão definidos a seguir. Cada padrão genético e seu
padrão relacionado obtidos pela técnica de PFGE correspondem, geralmente, a um
mesmo ST, que podem estar inseridos ou não em um Complexo Clonal (CC) e a um
“Sequence Group” (SG). Determina-se complexo clonal, o número do ST que está no
centro de um grupo, pois foi o primeiro ST encontrado (A exemplo da Figura 10, o ST4
está inserido no CC1, pois o ST1 foi o primeiro a ser reportado).

DISCUSSÃO
86
Figura 10: Modelo de um complexo clonal.
Fonte: http://eburst.mlst.net/v3/instructions/3.asp.
Quando um isolado bacteriano apresenta uma mutação em um alelo em
comparação ao ST anteriormente descrito, ele é denominado “Single Locus Variant
(SLV)”. Já se ele apresenta duas mutações em relação ao primeiro ST descrito, ele é
denominado “Double Locus Variant (DLV)”. Um “Singleton” é um ST que não está
inserido em um complexo clonal e, portanto, não possui “parentesco” com nenhum ST
anteriormente descrito, como exemplificado na Figura 10 pelo ST12. O SG é definido
pelo “eBURST”, quando a análise de todas as amostras é feita, separando-se em
grupos de ordem decrescente de quantidade de STs correlacionadas e não possui,
necessariamente, o mesmo número do complexo clonal. O SG será sempre variável e
um complexo clonal pode não pertencer sempre a um mesmo SG, pois conforme
novos STs vão sendo incorporados aos complexos clonais, o número de STs dentro
desses complexos aumenta, podendo um complexo clonal antes chamado de SG2,
passar a ser chamado de SG1, pois o SG1 apresenta agora, uma maior quantidade de
STs que o SG2. Amostras correlacionadas com dois ou mais STs, mas que não
formam um CC, também formam um SG.
As sequências alélicas obtidas para os setes genes dos 31 isolados de A.
baumannii produtores de OXA-23 avaliados no presente estudo, foram comparadas
com as sequências já existentes no banco de dados do MLST do “Institut Pasteur”
(http://www.pasteur.fr/cgi-bin/genopole/PF8/mlstdbnet.pl?page=download_profiles&file=acin_profiles.xml). A partir
Singleton . ST12

DISCUSSÃO
87
dessa comparação foi possível a obtenção da árvore filogenética (Figura 8), feita
através do site http://eburst.mlst.net, a qual permite verificar a relação de
ancestralidade entre os diferentes clones/STs e seus respectivos CCs em todo o
mundo.
Vários estudos demonstram que os clones europeus EU-I, EU-II e EU-III são os
mais disseminados em todo o mundo, principalmente na Europa e Ásia (Turton et al.,
2007; Towner et al., 2008; Mugnier et al., 2010; Diancourt et al., 2010; Fu et al., 2010).
No Brasil, há um estudo relatando que 70% das amostras de A. baumannii produtores
de OXA-23 isoladas em hospitais do Rio de Janeiro foram semelhantes ao clone EU-II
(Grosso et al., 2011). Estes clones europeus correspondem aos complexos clonais
CC1, CC2 e CC3, respectivamente pela metodologia, de Nemec (Krizova et al., 2011;
Mezzatesta et al., 2012). No Brasil, os STs mais disseminados são o ST1/CC1,
ST15/CC15 e ST79/CC79 (Mugnier et al., 2010, Grosso et al., 2011; Martins et al.,
2013; Coelho-Souza et al., 2013). Os dados obtidos no presente estudo corroboram
com a literatura, uma vez que a maioria dos isolados de A. baumannii produtores de
OXA-23 avaliados pertencem ao ST79, seguido do ST15 e do ST1. O ST79,
juntamente dos ST162, ST189 e ST190 fazem parte do CC79, um dos principais
complexos clonais descritos, sendo reportado também na Argentina (Stietz et al.,
2013). Esses dados demonstram que os principais clones brasileiros de A. baumannii
circulantes fazem parte de clones mundialmente disseminados. Além disso, foi
possível observar a inclusão de novos STs, demonstrando a provável transmissão do
gene blaOXA-23 para amostras com genótipos distintos. Embora nenhum padrão clonal
específico possa ser relacionado a uma determinada instituição no presente estudo, foi
possível observar que, dentro de um mesmo hospital, vários padrões de PFGE de A.
baumannii produtores de OXA-23 podem estar circulando ao mesmo tempo. Além
disso, isolados com padrões relacionados foram encontrados em diferentes hospitais
localizados em áreas geográficas distintas, sugerindo que possa ter ocorrido a

DISCUSSÃO
88
transmissão inter-hospitalar destes clones, os quais posteriormente se diferenciaram
geneticamente de maneira gradual.
Outro dado importante é que os principais padrões de A. baumannii
disseminados mundialmente apresentam um fenótipo MDR, o que torna o panorama
ainda mais preocupante (Mugnier et al., 2010; Grosso et al., 2011; Martins et al., 2013;
Coelho-Souza et al., 2013; Stietz et al., 2013). Quando avaliamos os dados de
sensibilidade aos antimicrobianos entre os 31 isolados avaliados no estudo,
verificamos altas taxas de resistência a quase todos os antimicrobianos testados no
isolados brasileiros. Somente a minociclina e a polimixina B, apresentaram atividade
frente aos isolados de A. baumannii produtores de OXA-23. Embora o uso da
minociclina tenha caído em desuso devido à introdução de agentes antimicrobianos
mais potentes (Neonakis et al., 2011), a sua concentração sérica/tecidual ideal e sua
notável penetração no sistema nervoso central, aliado ao aumento de infecções por
isolados de A. baumannii MDR, fez com que esse antimicrobiano voltasse a ganhar
destaque (Bishburg & Bishburg, 2009; Neonakis et al., 2011). Os relatos na literatura,
ainda que escassos, têm demonstrado resultados animadores para o tratamento de
infecções de pele (Griffith et al., 2008) e pneumonia (Wood et al., 2003; Griffith et al.,
2008) causadas por A. baumannii. Infelizmente, a minociclina na sua formulação
endovenosa, não é comercializada para uso clínico no Brasil.
As polimixinas (B e E/Colistina) apresentam uma atividade bactericida contra
isolados de A. baumannii, e as taxas de resistência tem se mantido relativamente
baixas, variando, em média, de 1,7% a 3% entre isolados de Acinetobacter spp.
(Gales et al., 2006; Souli et al., 2006; Gales et al., 2011). A excelente atividade das
polimixinas frente a isolados MDR de A. baumannii tem sido comprovada por
diferentes estudos, que comprovam que esse antimicrobiano, em muitos casos, é uma
das únicas opções disponíveis para o tratamento de infecções causadas por esse
micro-organismo (Levin et al., 1999; Kallel et al., 2007; Griffith et al., 2008; Bishburg &
Bishburg, 2009; Falagas et al., 2010, Rossi, 2011). Embora casos de resistência a

DISCUSSÃO
89
esses compostos sejam raros em isolados de A. baumannii, Ko e colaboradores
(2007) relataram uma taxa de 27,9% de resistência à polimixinas nos 214 isolados de
A. baumannii avaliados em dois hospitais sul-coreanos (Ko et al., 2007). Além da
monoterapia, estudos têm demonstrado que o uso das polimixinas em associação com
carbapenens, quinolonas e aminoglicosídeos no tratamento de infecções causadas por
isolados de A. baumannii MDR, apresentaram resultados promissores (Yoon et al.,
2004; Tan et al., 2007; Pankuch et al., 2008).
Todos os isolados de A. baumannii produtores de OXA-23 avaliados no
presente estudo apresentaram resistência à ampicilina/sulbactam. Apesar de ser
comercializado no Brasil juntamente com a ampicilina, o sulbactam, que é um inibidor
de β-lactamase com uma estrutura semelhante aos β-lactâmicos, é o que possui
atividade contra isolados de A. baumannii, porque se liga à PBP2 (Zavascki et al.,
2010). Altas taxas de resistência ao sulbactam também foram observadas em algumas
regiões geográficas, ainda que inferiores às observadas no presente estudo, variando
de 46,7% na Espanha (Fernández-Cuenca et al., 2004) a 70% em Tawain (Yang et al.,
2010). Apesar dos resultados desanimadores apresentados por esses estudos, alguns
autores obtiveram excelentes resultados utilizando esse antimicrobiano como opção
terapêutica para o tratamento de infecções causadas por A. baumannii (Levin et al.,
2002; Levin et al., 2003; Levin, 2003; Levin et al., 2008; Oliveira et al., 2008). Outro
fator que pode ter influenciado na resistência a esse antimicrobiano, são os pontos de
cortes inadequados para essas amostras, que poderiam ser revisados.
Levin e colaboradores (2003) observaram uma taxa de cura de 67,5% ao
tratarem 40 pacientes com infecções por A. baumannii MDR, principalmente infecção
de corrente sanguínea e pneumonia, com ampicilina/sulbactam. Em outro estudo, o
mesmo grupo de autores avaliou retrospectivamente, o desfecho clínico de pacientes
com infecções por Acinetobacter spp. resistentes aos carbapenens que haviam
recebido terapia com polimixina B ou ampicilina/sulbactam (Oliveira et al., 2008). Os
autores observaram que a resposta clínica à ampicilina/sulbactam foi superior àquela

DISCUSSÃO
90
apresentada pela polimixina no tratamento de A. baumannii MDR. Neste estudo, foi
também evidenciado que o uso de polimixina B era um fator independente de
mortalidade, apresentando alto valor de “Acute Physiological and Chronic Health
Evaluation II” - APACHE II (Oliveira et al., 2008). Devido aos resultados encorajadores,
recomenda-se que para infecções causadas por isolados de A. baumannii sensíveis à
ampicilina/sulbactam, deve-se priorizar esse antimicrobiano como opção terapêutica
(Neonakis et al., 2011). Além disso, alguns estudos sugerem que, para cepas MDR, o
ideal é utilizar a ampicilina/sulbactam combinada a outras drogas como à amicacina, à
rifampicina e/ou à colistina (Appleman et al., 2000; Savov et al., 2002; Ko et al., 2004,
Kiffer et al., 2005; Tong et al., 2006; Song et al., 2007; Peleg, 2007c; Maragakis & Perl,
2008, Shrivastava et al., 2009). Foi também demonstrado que o custo do tratamento
com ampicilina/sulbactam é significativamente menor quando comparado com
imipenem/cilastatina (1.500 dólares - imipenem-cilastatina vs 500 dólares -
ampicilina/sulbactam, p=0.004) (Jellison et al., 2001). A desvantagem no uso do
sulbactam seria o fato de que em muitos países, como o Brasil, somente existe a
formulação de sulbactam associada à ampicilina (proporção de 2:1
ampicilina/sulbactam).
Os dados de sensibilidade para tigeciclina entre os isolados de A. baumannii
produtores de OXA-23 foram desencorajadores, o que compromete o uso desse
antimicrobiano. Embora o uso da tigeciclina para o tratamento de infecções causadas
por A. baumannii seja “off-label”, as CIMs relativamente baixas (CIM50 0,5-1 mg/L e
CIM90 1-2 mg/L) obtidas em diferentes estudos, tornaram o uso desse antimicrobiano
atraente contra A. baumannii (Reinert et al., 2007; Garrison et al., 2009; Schimith et al.,
2010, Fernandéz-Canigia & Dowzicky, 2012, Zarrilli et al., 2012, Ergin et al., 2013,
Froment et al., 2013). Entretanto, apesar da boa atividade in vitro contra isolados de A.
baumannii MDR, a eficácia clínica da tigeciclina continua a ser controversa
(Karageorgopoulos et al., 2008; Gallagher & Rouse, 2008). Além disso, a resistência a
tigeciclina em A. baumannii ocorre pela hiperexpressão de sistemas de efluxo,

DISCUSSÃO
91
principalmente o sistema AdeABC, presente no cromossomo desse micro-organismo
(Peleg et al., 2007b), o que demonstra uma predisposição em adquirir naturalmente
resistência a esse antimicrobiano. Talvez as altas CIMs observadas para tigeciclina no
presente estudo possam ser justificadas pela hiperexpressão do sistema de efluxo
AdeABC verificado para quase todos os isolados nos testes de qRT-PCR.
Ainda que a maioria dos isolados de A. baumannii produtores de OXA-23
apresentaram resistência à amicacina, 45,2% dos isolados ainda mantém
sensibilidade à gentamicina. Vários estudos relatam a baixa atividade dos
aminoglicosídeos contra A. baumannii, geralmente associada à presença de metilases
em seu genoma (Doi & Arakawa, 2007; Zhou et al., 2010, Aghazadeh et al., 2013).
Assim como mostrado por estudos prévios (Perez et al., 2007; Gales et al., 2012), os
isolados de A. baumannii apresentaram altos níveis de resistência às fluoroquinolonas.
Embora a produção de OXA-23 justifique a resistência aos carbapenens
observada nos isolados de A. baumannii avaliados no estudo, o mesmo não se aplica
às cefalosporinas de espectro ampliado que não são hidrolisadas por essas β-
lactamases (Bush & Jacoby, 2010; Poirel et al., 2010). Nenhum gene codificador de
ESβL, dentre aqueles pesquisados, foi encontrado nos isolados brasileiros de A.
baumannii produtores de OXA-23 que pudesse justificar a resistência às
cefalosporinas de espectro ampliado. Embora as ESLs sejam muito prevalentes em
enterobactérias, raramente são descritas em isolados de A. baumannii (Vahaboglu et
al., 1997; Poirel et al., 2003; Nagano et al., 2004; Celenza et al., 2006; Pasterán et al.,
2006; Endimiani et al., 2007; Naas et al., 2007; Shakil et al., 2010; Bonnin et al.,
2011a, Bonnin et al., 2011b), o que corrobora com os nossos resultados. Somente a β-
lactamase de espectro restrito TEM-1 foi identificada nos isolados de A. baumannii
produtores de OXA-23. A presença dessa enzima já foi descrita em estudos prévios
com frequências que variaram de 18% a 71% (Ben et al., 2011; Manageiro et al.,
2012). No estudo conduzido por Ben e colaboradores (2011), os autores aplicaram um
teste de regressão multivariada e concluíram que a coprodução de OXA-23 (p=0,004 e

DISCUSSÃO
92
OR=10,52) e TEM-1 (p=0,005 e OR=6,14) é um bom preditor de resistência a
imipenem em isolados de A. baumannii.
Em isolados de A. baumannii, a expressão da AmpC cromossômica, que é
intrínseca dessa espécie, não é induzida pelo fato de que neste micro-organismo o
gene ampR está ausente, sendo, portanto, expressa em níveis basais, e, dessa forma,
não confere resistência às oximinocefalosporinas (Bou & Martínez-Beltrán, 2000).
Entretanto, a resistência às oximinocefalosporinas em A. baumannii está geralmente
relacionada à hiperprodução de AmpC, quando a sequência de inserção ISAba1 está
inserida a montante deste gene, conferindo um promotor forte (Boo et al., 2009; Lin et
al., 2010). Como era de se esperar, todos os isolados avaliados no presente estudo
apresentavam o gene blaAmpC e 25 dos 31 isolados apresentavam a ISAba1 a
montante do gene. Esses isolados possuíam altos níveis de resistência às
cefalosporinas de amplo espectro. Por outro lado, os seis isolados que não
apresentavam essa associação mostravam CIMs mais baixos para às cefalosporinas
de amplo espectro, demonstrando o papel da hiperprodução de AmpC na resistência a
esses antimicrobianos em isolados de A. baumannii produtores de OXA-23. A
importância da ISAba1 foi ressaltada em estudos prévios, demonstrando, inclusive, a
presença dessa sequência de inserção em determinados STs, principalmente,
naqueles mais prevalentes (Lin et al., 2010; Ben et al., 2011).
A presença de sequências de inserção, principalmente da ISAba1, permitiu ao
A. baumannii controlar a expressão gênica de maneira única. A grande vantagem está
no grande número de cópias dessas sequências ao longo do genoma de A. baumannii
(Segal et al., 2005a), aumentando a expressão ou inativando vários genes
codificadores de β-lactamases como a AmpC (Ben et al., 2011), a OXA-51 (Lopes et
al., 2012a), a OXA-23 (Lee et al., 2011) e a OXA-58 (Lopes et al., 2012b), e da porina
CarO (Mussi et al., 2005). Outro ponto importante do presente estudo foi a alta
frequência de ISAba1 a montante dos genes blaAmpC, blaOXA-51-like e blaOXA-23
concomitantemente nos isolados pertencentes aos CC79, CC15 e CC1. A

DISCUSSÃO
93
hiperprodução de todos esses genes contribui para os altos valores de CIMs para os
β-lactâmicos verificados nesses clones.
As CHDLs hidrolisam fracamente os carbapenens, sendo necessária a
presença de uma sequência de inserção para aumentar a expressão da maioria
dessas enzimas (Bush et al., 1995; Poirel & Nordmann, 2006; Walther-Rasmussen &
Høiby, 2006; Figueiredo et al., 2009, Bush & Jacoby, 2010). Embora cinco isolados
apresentassem ISAba1 a montante do gene blaOXA-51-like, não foi possível predizer o
impacto dessa associação na resistência aos carbapenens, já que todos os isolados
apresentaram também a ISAba1 a montante do gene blaOXA-23, o que logo justificaria a
resistência aos carbapenens. Apesar disso, vários estudos que avaliaram isolados de
A. baumannii resistentes aos carbapenens, que não carreavam genes codificadores de
nenhuma carbapenemase adquirida, concluíram que o gene blaOXA-51-like associado a
ISAba1 também está relacionado ao fenótipo de resistência aos carbapenens nesses
isolados (Héritier et al., 2006; Turton et al., 2006; Segal et al., 2007, Evans et al.,
2013).
Sistemas de efluxo são transportadores expressos em qualquer célula viva,
protegendo-as dos efeitos tóxicos de substâncias químicas orgânicas (Vila et al.,
2007). Esses sistemas podem ser específicos para um determinado substrato ou
podem transportar uma gama de compostos estruturalmente diferentes (Piddock,
2006; Coyne et al., 2011). Dessa forma, o fenótipo MDR está frequentemente
associado à hiperexpressão desses sistemas, já que reduzem, ao mesmo tempo, o
acúmulo de muitos antimicrobianos no interior da bactéria (Piddock, 2006; Vila et al.,
2007; Coyne et al., 2011). Embora esses sistemas possam conferir diminuição da
sensibilidade a muitos antimicrobianos, nem sempre essa diminuição resulta em
resistência (Piddock, 2006).
No presente estudo não foi considerado significativa a diminuição de apenas
uma diluição na CIM para qualquer um dos quatro β-lactâmicos testados na presença
do inibidor, já que tal diferença entre dois testes dilucionais poderia ser consequente à

DISCUSSÃO
94
variabilidade atribuída à própria técnica de sensibilidade utilizada. Sendo assim, neste
estudo, as CIMs para ceftazidima e, principalmente, para meropenem diminuíram (≥ 2
diluições) na presença do inibidor de sistemas de efluxo PAβN. Além disso, o isolado
que apresentava CIMs elevadas para ceftazidima e cefotaxima, mesmo sem
apresentar ISAba1 associada a AmpC, apresentou uma diminuição da CIM para
ceftazidima de 128 µg/mL para 16 µg/mL, mas não para cefotaxima. Esses dados
demonstram um possível impacto da hiperexpressão dos sistemas de efluxo na
resistência à ceftazidima em isolados que não hiperproduzem a AmpC e no aumento
dos níveis de resistência ao meropenem em isolados que produzem OXA-23. Dados
similares foram obtidos por Peleg e colaboradores (2007b) que avaliaram a influência
do inibidor PAβN nas CIMs para diferentes antimicrobianos. Embora eles tenham
enfatizado a diminuição das CIMs para tigeciclina na presença do inibidor, a CIMs para
meropenem, imipenem e ceftazidima decresceram consideravelmente, o mesmo não
ocorrendo para cefepima. Apesar disso, naquele estudo, os isolados avaliados eram
sensíveis aos carbapenens, com CIMs que variavam de 0,06 a 4 µg/mL,
demonstrando que os sistemas de efluxo contribuem na elevação da CIMs, ainda que
não sejam capazes de tornar os isolados resistentes aos carbapenens. Ainda que de
forma mais discreta, as CIMs para imipenem também diminuíram de duas a três
diluições. Ao contrário, no estudo conduzido por Hou e colaboradores (2012) que
relataram uma diminuição das CIMs para imipenem de 4 a 32 vezes em isolados de A.
baumannii resistentes a esse composto, o mesmo não foi verificado nos isolados
sensíveis a imipenem. Eles observaram também um aumento de 4, 3 e 11 vezes na
expressão das bombas adeB, AdeJ e AdeM.
Os dados de qRT-PCR demonstraram uma hiperexpressão do sistema de
efluxo AdeABC entre os isolados brasileiros de A. baumannii produtores de OXA-23,
principalmente naqueles isolados que estão incluídos nos três principais complexos
clonais disseminados em nosso país (CC79, CC15 e CC1). Esse aumento da
expressão pode ser um dos responsáveis pelo fenótipo MDR verificado entre os

DISCUSSÃO
95
isolados brasileiros, contribuindo, principalmente, na elevação das CIMs verificadas
para tigeciclina nesse grupo amostral. Além disso, é importante lembrar que outros
sistemas de efluxo da Família RND já foram descritos em isolados de A. baumannii, e
que poderiam estar contribuindo na ejeção de ceftazidima e meropenem, como
verificado no teste fenotípico com o PAβN (Coyne et al., 2011).
Os resultados obtidos até o momento na literatura sugerem que o sistema
AdeABC esteja relacionado à resistência ao meropenem, mas não ao imipenem em
isolados de A. baumannii produtores de OXA-23 que tiveram o gene adeB deletado
(Wong et al., 2009). Provavelmente, somente a hiperexpressão do sistema AdeABC
não seja suficiente para causar altos níveis de resistência aos carbapenens,
necessitando da associação de outros mecanismos (Coyne et al., 2011). No estudo
conduzido por Yoon e colaboradores (2013) verificou-se uma alta frequência de
hiperexpressão do sistema de efluxo AdeABC em isolados MDR de A. baumannii. Os
autores associaram essa hiperexpressão a mutações nos genes regulatórios adeR e
adeS.
Uma grande variedade de oxacilinases do grupo das OXA-51 foi descrita entre
os isolados de A. baumannii produtores de OXA-23. De um modo geral, a variante de
OXA-51 produzida está relacionada ao ST. Apesar disso, isolados apresentando perfil
de PFGE relacionados podem sofrer mutações no gene blaOXA-51-like, originado novas
variantes, como observado no presente estudo. Uma alta variabilidade de oxacilinases
intrínsecas em A. baumannii já foi relatada previamente, demonstrando que tais
enzimas variam conforme o padrão de PFGE (Héritier et al., 2005a). Essa estreita
correlação entre a variante de OXA-51 e seu padrão de PFGE, permitiu a Zander e
colaboradores (2012) correlacionar a variante ao perfil clonal obtido pelo DiversiLab,
de modo que os autores concluíram que a tipagem baseada no sequenciamento dos
genes blaOXA-51-like, demonstra ser uma promissora ferramenta na caracterização
molecular de isolados de A. baumannii e identificar as principais linhagens clonais
disseminadas no mundo. Como as variantes de OXA-51 variam conforme o padrão

DISCUSSÃO
96
clonal, o estudo de Ikonomidis e colaboradores (2007) realizado na Grécia,
demonstrou uma baixa variabilidade de oxacilinases pertencentes a esse grupo,
provavelmente em consequência a uma menor variabilidade de padrões observada
entre os isolados de A. baumannii naquele país.
Embora a presença do gene blaOXA-51-like, intrínseco do cromossomo de A.
baumannii, tenha sido detectado em todos os isolados avaliados no presente estudo, a
disseminação desse gene mediada por plasmídeos para outras espécies de
Acinetobacter não-baumannii em Taiwan (Lee et al., 2012), comprometeu a utilização
desse gene como prova presuntiva de identificação para A. baumannii. Apesar disso,
os nossos resultados indicam que todos os isolados produtores de OXA-51 foram
identificados como sendo A. baumannii pelo sequenciamento do gene rpoB, indicando
que em nosso meio, o uso do gene blaOXA-51-like ainda é um bom teste para diferenciar
os isolados como sendo baumannii de não-baumannii. Outro dado importante, foi a
excelente correlação entre os resultados obtidos pelo MALDI-TOF e o sequenciamento
do gene rpoB.
Acredita-se que o tamanho pequeno e o limitado número de porinas em
Acinetobacter spp. seja o responsável pela resistência intrínseca aos antimicrobianos
observada neste micro-organismo (Vila et al., 2007). O conceito que se tinha sobre a
permeabilidade de A. baumannii foi em parte confirmado por um interessante estudo
publicado recentemente (Sugawara & Nikaido, 2012). Nesse estudo, foi observado que
a permeabilidade da membrana externa de A. baumannii para cefalotina e cefaloridina
era 100 vezes menor do que a permeabilidade observada para a E. coli K12,
demonstrando que naturalmente A. baumannii apresenta uma baixa permeabilidade
de membrana externa (Sugawara & Nikaido, 2012). Além disso, foi observado que
embora a OmpA, também conhecida como OmpHMP (36,5 kDa), apresente uma
capacidade mínima de formar poros, houve uma diminuição de duas a três vezes na
permeabilidade para cefalotina e cefaloridina quando o gene ompA foi deletado
(Sugawara & Nikaido, 2012). Os autores concluem que a baixa permeabilidade dessa

DISCUSSÃO
97
porina aliada à hiperexpressão de β-lactamases intrínsecas, como as CHDLs do
cluster OXA-51 e as cefalosporinases do tipo ADC/AmpC, e de sistemas de efluxo,
poderiam ser essenciais para os altos níveis de resistência intrínseca observado para
muitos antimicrobianos em isolados clínicos de A. baumannii (Sugawara & Nikaido,
2012).
Ainda que sejam poucos os estudos que avaliaram o papel das porinas na
resistência aos carbapenens em A. baumannii, os dados obtidos mostraram que essas
proteínas desempenham um papel importante na resistência aos carbapenens
(Limansky et al., 2002; Dupont et al., 2005; Mussi et al., 2005; Tomás et al., 2005).
Sendo assim, Limansky e colaboradores (2002) demonstraram que a resistência ao
imipenem em um isolado de Acinetobacter spp. não produtor de carbapenemase,
estava associada à perda de uma proteína de 29 kDa, designada CarO. Resultado
semelhante foi observado por um estudo conduzido por Mussi e colaboradores (2005),
no qual foi demonstrado que a resistência ao imipenem e ao meropenem em A.
baumannii também estava associada a perda dessa porina, pela inativação do gene
carO por uma IS. Recentemente, outros dois estudos, comparando a proteômica de
isolados sensíveis e resistentes de A. baumannii, observaram alterações estruturais na
CarO, principalmente nas estruturas primárias e quartenárias dessa porina (Siroy et
al., 2006; Vashist et al., 2010). Foi também observado que as isoformas alteradas não
transportavam eficientemente os carbapenens como a porina “selvagem” (Vashist et
al., 2010). Dessa forma, isolados de A. baumannii resistentes, provavelmente,
alterariam a CarO de modo a diminuir a entrada do carbapenem na célula bacteriana.
Da mesma forma, outras duas porinas foram descritas relacionadas à resistência aos
carbapenens em A. baumannii: uma proteína chamada 33-36 kDa (Tomás et al.,
2005), com um tamanho estimado de 31 kDa, e outra de 43 kDa, que demonstra
homologia peptídica com a OprD (D2) de P. aeruginosa (Dupont et al., 2005). Como
nenhum sítio de ligação ao imipenem pode ser detectado na CarO, acredita-se que
esta porina seja um canal inespecífico de entrada para os carbapenens (Siroy et al.,

DISCUSSÃO
98
2005), enquanto a proteína de 43 kDa (OprD-like) seria o canal específico para estes
antimicrobianos (Vila et al., 2007).
Outra porina descrita em A. baumannii é a OmpW (21 kDa), que demonstra
uma alta homologia com a OmpW de E. coli e P. aeruginosa. Embora sua função em
A. baumannii ainda não esteja bem esclarecida, Vila e colaboradores (2007)
observaram uma diminuição in vitro da expressão dessa porina em mutantes de A.
baumannii resistentes à colistina (Vila et al., 2007). Entretanto, Tiwari e colaboradores
(2012) avaliando a proteômica de isolados de A. baumannii resistentes aos
carbapenens, observaram uma diminuição da expressão de OmpW, comparada com a
cepa ATCC 19606 de A. baumannii sensível aos carbapenens. Ainda que em estudo
similar, Siroy e colaboradores (2005) não tenham observado alteração na expressão
da OmpW, eles relataram a presença de pelo menos quatro isoformas diferentes
dessa OMP no gel de SDS-PAGE (Siroy et al., 2006). Vashist e colaboradores (2010)
também observaram a presença de isoformas de OmpW e, em decorrência desse
achado, os autores inferiram que era possível que a OmpW, sendo uma porina
pequena, fosse importante para realizar a função de transporte na ausência ou
alteração das porinas principais em isolados resistentes de A. baumannii (Vashist et
al., 2010).
É importante ressaltar que a análise de géis de proteínas de membrana externa
pela técnica de SDS-PAGE é sempre relativa, por apresentar diferentes fatores
durante a realização da técnica que podem alterar o perfil das bandas proteicas.
Apesar do protocolo utilizado no presente estudo objetivar diminuir a influência desses
fatores, a presença de formas aberrantes da mesma proteína em isolados clínicos de
A. baumannii, faz com que elas apresentem tamanhos diferentes no gel de SDS-
PAGE (Siroy et al., 2006; Vashist et al., 2010), tornando a análise do gel para esse
micro-organismo uma tarefa complexa. Ainda assim, o SDS-PAGE continua sendo
uma técnica útil para avaliar o perfil de OMPs em bacilos Gram negativos resistentes
aos carbapenens (Limansky et al., 2002; Dupont et al., 2005; Mussi et al., 2005;

DISCUSSÃO
99
Tomás et al., 2005). Sendo assim, a análise dos géis de SDS-PAGE demonstrou um
perfil de OMPs variável entre os isolados de A. baumannii produtores de OXA-23,
apresentando proteínas com tamanhos alterados compatíveis com a OmpHMP e a
Omp25, duas proteínas, cuja importância na resistência aos carbapenens ainda
permanece desconhecida. Talvez essas alterações sejam uma ação compensatória
nessas proteínas de modo a contrabalancear a pressão seletiva dos β-lactâmicos
sobre as proteínas utilizadas por esses compostos para atravessar a membrana
externa bacteriana, como a CarO, Omp33-36 e OprD (Vashist et al., 2010). Além
disso, para a maioria dos isolados analisados, não foi verificado uma OMP de
aproximadamente 21 kDa compatível com a OmpW. Entretanto, comparando os dados
de expressão obtidos pela técnica de qRT-PCR, verificou-se que a porina OmpW
continuava a ser expressa nos isolados clínicos. O peso molecular muito similar entre
as proteínas importantes e relacionadas à resistência aos carbapenens pode ter
dificultado a análise visual do gel, já que as OMPs OmpW (21 kDa), Omp25 (25 kDa),
CarO (29 kDa) e Omp33-36 (31 kDa) estão incluídas em uma faixa de peso molecular
de apenas 10 kDa.
Os resultados obtidos com o presente estudo demonstraram a complexidade
envolvida na resistência aos β-lactâmicos em isolados brasileiros de A. baumannii
produtores de OXA-23. Vários são os mecanismos que podem estar expressos ou
reprimidos ao mesmo tempo elevando as CIMs. Talvez uma das grandes vantagens
evolutivas que permitiram a esses micro-organismos o sucesso alcançado no
ambiente hospitalar seja a presença de diferentes sequências de inserção no seu
genoma, promovendo um controle da expressão gênica com o menor gasto energético
possível. Essa característica é única entre os micro-organismos patogênicos e,
embora alarmante, torna essa bactéria ainda mais fascinante.

CONCLUSÃO
100
7. CONCLUSÃO
A hiperexpressão do sistema de efluxo AdeABC contribuiu com o aumento nos
níveis de resistência principalmente ao meropenem e à ceftazidima;
A presença de ISAba1 a montante do gene blaAmpC poderia justificar os altos
níveis de resistência às cefalosporinas de amplo espectro encontradas neste
estudo;
Uma alta diversidade de STs foi verificada nesses isolados, demonstrando uma
diversificação ao longo dos anos;
A maioria dos isolados brasileiros de A. baumannii incluídos no CC79, CC15 e
CC1 apresentam ISAba1 a montante dos genes blaAmpC, blaOXA-51-like e blaOXA-23
em contraposição a maioria dos isolados não inseridos em um Complexo
Clonal;
A maioria dos isolados brasileiros está relacionada aos ST79, ST15 e ST1
incluídos nos CC79, CC15 e CC1, respectivamente, sendo estes os mais
disseminados pelo Brasil;
O ST1, inserido no CC1, está relacionado ao EU-I, um dos mais disseminados
no mundo;
Um grande número de variantes pertencentes ao grupo das CHDLs
cromossômicas e intrínsecas de A. baumannii foi observado nos isolados,
variando de acordo com o padrão de PFGE;

CONCLUSÃO
101
Altos níveis de expressão do sistema de efluxo AdeABC foram verificados na
maioria dos isolados de A. baumannii produtores de OXA-23 quando
comparados a cepa ATCC BAA-1907 multisensível;
Os altos níveis de expressão do gene adeB se correlacionam, em sua maioria,
ao ST1, ST15 e ST79, sendo o ST, onde estão incluídos os padrões G e H, os
que mais hiperexpressam;
Não foi possível correlacionar o alto nível de expressão do sistema de efluxo
aos altos níveis de resistência aos antimicrobianos;
Somente a minociclina e a polimixina B apresentaram excelente atividade
frente aos isolados de A. baumannii produtores de OXA-23;
A presença de ISAba1 à montante da AmpC está relacionada aos altos níveis
de resistência às cefalosporinas de terceira geração, no entanto, isolados que
não possuem a ISAba1 à montante da AmpC, apresentaram menor taxa de
resistência aos antimicrobianos de uma forma geral,
Uma complexa organização do perfil de proteínas de membrana externa foi
observada entre os isolados avaliados no estudo, demonstrando que a
impermabilidade de membrana é um fator considerável.

REFERÊNCIAS BIBLIOGRÁFICAS
102
8. REFERÊNCIAS BIBLIOGRÁFICAS Afzal-Shah M, Woodford N, Livermore DM. Characterization of OXA-25, OXA-26, and OXA-27, molecular class D β-lactamases associated with carbapenem resistance in clinical isolates of Acinetobacter baumannii. Antimicrob Agents Chemother. 2001; 45(2):583-8. Aghazadeh M, Rezaee MA, Nahaei MR, Mahdian R, Pajand O, Saffari F, Hassan M, Hojabri Z. Dissemination of Aminoglycoside-Modifying Enzymes and 16S rRNA Methylases Among Acinetobacter baumannii and Pseudomonas aeruginosa Isolates. Microb Drug Resist. 2013 Apr 11. [Epub ahead of print] Albers-Schonberg G, Arison BH, Hensens OD, Hirshfield J, Hoogsteen K, Kaczke EA, Rhodes RE, Kahan JS, Kahan FM, Ratcliffe W, Walton E, Ruswinkle LJ, Morin RB, Christensen BG. Structure and absolute configuration of thienamycin. Journal of the American Chemical Society. 1978;100(20):6491-9. Al-Sweih NA, Al-Hubail M, Rotimi VO. Three distinct clones of carbapenem-resistant Acinetobacter baumannii with high diversity of carbapenemases isolated from patients in two hospitals in Kuwait. J Infect Public Health. 2012;5(1):102-8. Ambler RP. The structure of β-lactamases. Philos. Trans. R. Soc. Lond B Biol. Sci. 1980; 289: 321-31. Andriamanantena TS, Ratsima E, Rakotonirina HC, Randrianirina F, Ramparany L, Carod JF, Richard V, Talarmin A. Dissemination of multidrug resistant Acinetobacter baumannii in various hospitals of Antananarivo Madagascar. Ann Clin Microbiol Antimicrob. 2010;30(9):17. Anstey NM, Currie BJ, Withnall KM. Community-acquired Acinetobacter pneumonia in the Northern Territory of Australia. Clin Infect Dis. 1992;14(1):83-91. Anstey NM, Currie BJ, Hassell M, Palmer D, Dwyer B, Seifert H. Community-acquired bacteremic Acinetobacter pneumonia in tropical Australia is caused by diverse strains of Acinetobacter baumannii, with carriage in the throat in at-risk groups. J Clin Microbiol. 2002;40(2):685-6. Antonio CS, Neves PR, Medeiros M, Mamizuka EM, Elmor de Araújo MR, Lincopan N. High prevalence of carbapenem-resistant Acinetobacter baumannii carrying the blaOXA-
143 gene in Brazilian hospitals. Antimicrob Agents Chemother. 2011;55(3):1322-3. Appleman MD, Belzberg H, Citron DM, Heseltine PN, Yellin AE, Murray J, Berne TV. In vitro activities of nontraditional antimicrobials against multiresistant Acinetobacter baumannii strains isolated in an intensive care unit outbreak. Antimicrob Agents Chemother. 2000;44(4):1035-40. Arroyo LA, Mateos I, Gonzalez V, Aznar J.In vitro activities of tigecycline, minocycline, and colistin–tigecycline combination against multi- and pandrug-resistant clinical isolates of Acinetobacter baumannii group. Antimicrob Agents Chemother. 2009;53(3):1295–6. Aubert D, Poirel L, Chevalier J, Leotard S, Pages JM, Nordmann P. Oxacillinase-mediated resistance to cefepime and susceptibility to ceftazidime in Pseudomonas aeruginosa. Antimicrob Agents Chemother. 2001;45(6):1615-20.

REFERÊNCIAS BIBLIOGRÁFICAS
103
Bartual SG, Seifert H, Hippler C, Luzon MA, Wisplinghoff H, Rodríguez-Valera F. Development of a multilocus sequence typing scheme for characterization of clinical isolates of Acinetobacter baumannii. J Clin Microbiol. 2005;43(9):4382-90. Basker MJ. The carbapenem family. J Antimicrob Chemother. 1982;10(1):4-7. Baumann P, Doudoroff M, Stanier RY. A study of the Moraxella group. II. Oxidative-negative species (genus Acinetobacter). J Bacteriol. 1968;95(5):1520-41. Beijerinck MW. Über pigmentbildungbei essigbakterien. Centr. Bakteriol. Parasitenk. 1911;2(29):169-76. Ben RJ, Yang MC, Hsueh JC, Shiang JC, Chien ST. Molecular characterisation of multiple drug-resistant Acinetobacter baumannii isolates in southern Taiwan. Int J Antimicrob Agents. 2011;38(5):403-8. Bergeron J, Ammirati M, Danley D, James L, Norcia M, Retsema J, Strick CA, Su WG, Sutcliffe J, Wondrack L. Glycylcyclines bind to the high-affinity tetracycline ribosomal binding site and evade Tet(M) and Tet(O) mediated ribosomal protection. Antimicrob Agents Chemother. 1996;40(9):2226-8 Bergogne-Bérézin E, Towner KJ. Acinetobacter spp. as nosocomial pathogens: microbiological, clinical, and epidemiological features. Clin Microbiol Rev. 1996; 9(2):148-65. Berlau J, Aucken H, Malnick H, Pitt T. Distribution of Acinetobacter species on skin of healthy humans. Eur J Clin Microbiol Infect Dis. 1999a;18(3):179-83. Berlau J, Aucken HM, Houang E, Pitt TL. Isolation of Acinetobacter spp. including A. baumannii from vegetables: implications for hospital-acquired infections. J Hosp Infect. 1999b;42(3):201-4. Bermudes H, Jude F, Chaibi EB, Arpin C, Bebear C, Labia R, Quentin C. Molecular characterization of TEM-59 (IRT-17), a novel inhibitor-resistant TEM-derived β-lactamase in a clinical isolate of Klebsiella oxytoca. Antimicrob Agents Chemother. 1999;43(7):1657-61. Bertsche U, Breukink E, Kast T, Vollmer W. In vitro murein peptidoglycan synthesis by dimers of the bifunctional transglycosylase-transpeptidase PBP1B from Escherichia coli. J Biol Chem. 2005;280(45):38096-101. Bishburg E, Bishburg K. Minocycline--an old drug for a new century: emphasis on methicillin-resistant Staphylococcus aureus (MRSA) and Acinetobacter baumannii. Int J Antimicrob Agents. 2009;34(5):395-401. Bogaerts P, Cuzon G, Naas T, Bauraing C, Deplano A, Lissoir B, Nordmann P, Glupczynski Y. Carbapenem-resistant Acinetobacter baumannii isolates expressing the blaOXA-23 gene associated with ISAba4 in Belgium. Antimicrob Agents Chemother. 2008;52(11):4205-6. Bogaerts P, Naas T, El Garch F, Cuzon G, Deplano A, Delaire T, Huang TD, Lissoir B, Nordmann P, Glupczynski Y. GES extended-spectrum β-lactamases in Acinetobacter baumannii isolates in Belgium. Antimicrob Agents Chemother. 2010;54(11):4872-8.

REFERÊNCIAS BIBLIOGRÁFICAS
104
Bonnet R, Marchandin H, Chanal C, Sirot D, Labia R, De Champs C, Jumas-Bilak E, Sirot J. Chromosome-encoded class D β-lactamase OXA-23 in Proteus mirabilis. Antimicrob Agents Chemother. 2002;46(6):2004-6. Bonnin RA, Nordmann P, Potron A, Lecuyer H, Zahar JR, Poirel L. Carbapenem-hydrolyzing GES-type extended-spectrum β-lactamase in Acinetobacter baumannii. Antimicrob Agents Chemother. 2011a;55(1):349-54. Bonnin RA, Potron A, Poirel L, Lecuyer H, Neri R, Nordmann P. PER-7, an extended-spectrum beta-lactamase with increased activity toward broad-spectrum cephalosporins in Acinetobacter baumannii. Antimicrob Agents Chemother. 2011b;55(5):2424-7. Bonnin RA, Poirel L, Licker M, Nordmann P. Genetic diversity of carbapenem-hydrolysing β-lactamases in Acinetobacter baumannii from Romanian hospitals. Clin Microbiol Infect. 2011c;17(10):1524-8. Boo TW, Walsh F, Crowley B. Molecular characterization of carbapenem-resistant Acinetobacter species in an Irish university hospital: predominance of Acinetobacter genomic species 3. J Med Microbiol. 2009;58(2):209-16. Born P, Breukink E, Vollmer W. In vitro synthesis of cross-linked murein and its attachment to sacculi by PBP1A from Escherichia coli. J Biol Chem. 2006;281(37):26985-93 Bou G, Martínez-Beltrán J. Cloning, nucleotide sequencing, and analysis of the gene encoding an AmpC β-lactamase in Acinetobacter baumannii. Antimicrob Agents Chemother. 2000;44(2):428-32. Bou G, Oliver A, Martínez-Beltrán J. OXA-24, a novel class D beta-lactamase with carbapenemase activity in an Acinetobacter baumannii clinical strain. Antimicrob Agents Chemother. 2000a;44(6):1556-61. Bou G, Cervero G, Dominguez M A, Quereda C, Martinez-Beltran J. Characterization of a nosocomial outbreak caused by a multiresistant Acinetobacter baumannii strain with a carbapenem-hydrolyzing enzyme: high-level carbapenem resistance in A. baumannii is not due solely to the presence of β-lactamases. J. Clin. Microbiol. 2000b;38(9):3299–3305. Bouvet PMJ, Grimont PAD. Taxonomy of the genus Acinetobacter with the recognition of Acinetobacter baumannii sp. nov. Acinetobacter haemolyticus sp. nov. Acinetobacter johnsonii sp. nov. and Acinetobacter junii sp. nov. and emended descriptions of Acinetobacter calcoaceticus and Acinetobacter lwofii. International Journal of Systematic Bacteriology. 1986;36(2):228-40. Bradford PA, Urban C, Mariano N, Projan SJ, Rahal JJ, Bush K. Imipenem resistance in Klebsiella pneumoniae is associated with the combination of ACT-1, a plasmid-mediated AmpC beta-lactamase, and the foss of an outer membrane protein. Antimicrob Agents Chemother. 1997;41(3):563-9. Bratu S, Landman D, Alam M, Tolentino E, Quale J. Detection of KPC carbapenem-hydrolyzing enzymes in Enterobacter spp. from Brooklyn, New York. Antimicrob Agents Chemother. 2005;49(2):776-8.

REFERÊNCIAS BIBLIOGRÁFICAS
105
Bratu S, Landman D, Martin DA, Georgescu C, Quale J. Correlation of antimicrobial resistance with β-lactamases, the OmpA-like porin, and efflux pumps in clinical isolates of Acinetobacter baumannii endemic to New York City. Antimicrob Agents Chemother. 2008;52(9):2999-3005. Brauers J, Frank U, Kresken M, Rodloff AC, Seifert H. Activities of various β-lactams and β-lactam/β-lactamase inhibitor combinations against Acinetobacter baumannii and Acinetobacter DNA group 3 strains. Clin Microbiol Infect. 2005;11(1):24-30. Brisou J, Prevot AR. Studies on bacterial taxonomy X The revision of species under Acromobacter group. Ann Inst Pasteur (Paris). 1954;86(6):722-8. BSAC. British Society ofor Antimicrobial Chemotherapy - Methods for Antimicrobial Susceptibility Testing.Version 9.1 March 2010. Birmingham, UK. Brown AG, Butterworth D, Cole M, Hanscomb G, Hood JD, Reading C, Rolinson GN. Naturally-occurring β-lactamase inhibitors with antibacterial activity. J Antibiot (Tokyo). 1976;29(6):668-9. Brown S, Young HK, Amyes SG. Characterisation of OXA-51, a novel class D carbapenemase found in genetically unrelated clinical strains of Acinetobacter baumannii from Argentina. Clin Microbiol Infect. 2005;11(1):15-23. Brown S, Amyes S. OXA (β)-lactamases in Acinetobacter: the story so far. J Antimicrob Chemother. 2006;57(1):1-3. Bush K. Characterization of beta-lactamases. Antimicrob Agents Chemother. 1989a;33(3):259-63. Bush K. Classification of beta-lactamases: groups 1, 2a, 2b, and 2b'. Antimicrob Agents Chemother. 1989b;33(3):264-70. Bush K. Classification of beta-lactamases: groups 2c, 2d, 2e, 3, and 4. Antimicrob Agents Chemother. 1989c;33(3):271-6. Bush K, Jacoby GA, Medeiros AA. A functional classification scheme for β-lactamases and its correlation with molecular structure. Antimicrob. Agents Chemother. 1995;39(6):1211-33. Bush K, Jacoby GA. Updated functional classification of β-lactamases. Antimicrob Agents Chemother. 2010;54(3):969-976. Butterworth D, Cole M, Hanscomb G, Rolinson GN. Olivanic acids, a family of β-lactam antibiotics with β-lactamase inhibitory properties produced by Streptomyces species. I. Detection, properties and fermentation studies. J Antibiot (Tokyo). 1979;32(4):287-94. Calhoun JH, Murray CK, Manring MM. Multidrug-resistant organisms in military wounds from Iraq and Afghanistan. Clin Orthop Relat Res. 2008;466(6):1356-62. Carneiro M, Barbosa PI, Vespero EC, Tanita MT, Carrilho CM, Perugini M, Saridakis HO, Possuelo LG, Renner JD, Valim AR, Tognin MC, Martins AF, Barth AL. Carbapenem-resistant OXA-23-producing Acinetobacter baumannii isolates causing ventilator-associated pneumonia. Am J Infect Control. 2010;38(8):667-9.

REFERÊNCIAS BIBLIOGRÁFICAS
106
Carvalho KR, Carvalho-Assef AP, Peirano G, Santos LC, Pereira MJ, Asensi MD. Dissemination of multidrug-resistant Acinetobacter baumannii genotypes carrying blaOXA-23 collected from hospitals in Rio de Janeiro, Brazil. Int J Antimicrob Agents. 2009;34(1):25-8. Cascio A, Conti A, Sinardi L, Iaria C, Angileri FF, Stassi G, David T, Versaci A, Iaria M, David A. Post-neurosurgical multidrug-resistant Acinetobacter baumannii meningitis successfully treated with intrathecal colistin. A new case and a systematic review of the literature. Int J Infect Dis. 2010;14(7):572-9. Cayô R, Petrolini FVB, Carvalhaes CG, Cardoso JP, Machado AMO, Gales AC. Temporal Dynamic of Carbapenemase-producing Acinetobacter baumannii (Acb) in a Brazilian Teaching Hospital Through an 18-Year Period: Earlier Dissemination of blaOXA-143 in Brazil. In: 51st ICAAC Interscience Conference on Antimicrobial Agents and Chemotherapy, 2011a, Chicago, IL. 51st ICAAC Interscience Conference on Antimicrobial Agents and Chemotherapy, 2011a. Cayô R, Rodríguez MC, Espinal P, Fernández-Cuenca F, Ocampo-Sosa AA, Pascual A, Ayala JA, Vila J, Martínez-Martínez L. Analysis of genes encoding penicillin-binding proteins in clinical isolates of Acinetobacter baumannii. Antimicrob Agents Chemother. 2011b;55(12):5907-13. Cayô R – Tese de Doutorado - Dinâmica da resistência aos β-lactâmicos em amostras clínicas de Acinetobacter spp. isoladas no Brasil e na Espanha. 2012. pp 7. Celenza G, Pellegrini C, Caccamo M, Segatore B, Amicosante G, Perilli M. Spread of blaCTX-M-type and blaPER-2 β-lactamase genes in clinical isolates from Bolivian hospitals. J Antimicrob Chemother. 2006;57(5):975-8. Chang WN, Lu CH, Huang CR, Chuang YC. Community-acquired Acinetobacter meningitis in adults. Infection. 2000;28(6):395-7. Chang HC, Wei YF, Dijkshoorn L, Vaneechoutte M, Tang CT, Chang TC. Species-level identification of isolates of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex by sequence analysis of the 16S-23S rRNA gene spacer region. J Clin Microbiol. 2005; 43(4):1632-9. Chen MZ, Hsueh PR, Lee LN, Yu CJ, Yang PC, Luh KT. Severe community-acquired pneumonia due to Acinetobacter baumannii. Chest. 2001;120(4):1072-7. Chen CH, Young TG, Huang CC. Predictive biomarkers for drug-resistant Acinetobacter baumannii isolates with blaTEM-1, AmpC-type bla and integrase 1 genotypes. J Microbiol Immunol Infect. 2006;39(5):372-9. Chen Y, Pi B, Zhou H, Yu Y, Li L. Triclosan resistance in clinical isolates of Acinetobacter baumannii. J Med Microbiol. 2009;58(Pt 8):1086-91. Chen TL, Lee YT, Kuo SC, Hsueh PR, Chang FY, Siu LK, Ko WC, Fung CP. Emergence and distribution of plasmids bearing the blaOXA-51-like gene with an upstream ISAba1 in carbapenem-resistant Acinetobacter baumannii isolates in Taiwan. Antimicrob Agents Chemother. 2010;54(11):4575-81. Chopra I, Hawkey PM, Hinton M. Tetracyclines, molecular and clinical aspects. J Antimicrob Chemother. 1992;29(3):245-77.

REFERÊNCIAS BIBLIOGRÁFICAS
107
Chopra I, Roberts M. Tetracycline antibiotics: mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol Mol Biol. 2001;65(2):232-60. Chu YW, Leung CM, Houang ET, Ng KC, Leung CB, Leung HY, Cheng AF. Skin carriage of Acinetobacter in Hong Kong. J Clin Microbiol. 1999;37(9):2962-7. CLSI. Clinical Laboratory Standard Institute. Methods For Dilution Antimicrobial Susceptibility Tests For Bacteria That Grow Aerobically. Document M7-A9, Wayne, Pa, USA, 2012. CLSI. Clinical Laboratory Standard Institute. Performance standards for antimicrobial susceptibility testing; Twenty-Second Informational Supplement. Document M100-S22, Wayne, Pa, USA. 2012. Coelho-Souza T, Reis JN, Martins N, Martins IS, Menezes AO, Reis MG, Silva NO, Dias RC, Riley LW, Moreira BM. Longitudinal surveillance for meningitis by Acinetobacter in a large urban setting in Brazil. Clin Microbiol Infect. 2013;19(5):E241-4. Conway SP, Pond MN, Watson A, Etherington C, Robey HL, Goldman MH. Intravenous colistin sulphomethate in acute respiratory exacerbations in adult patients with cystic fibrosis. Thorax. 1997;52(11):987-93. Corbella X, Ariza J, Ardanuy C et al. Efficacy of sulbactamalone and in combination with ampicillin in nosocomial infections caused by multiresistant Acinetobacter baumannii. J Antimicrob Chemother. 1998;42(6):793–802. Cornaglia G, Giamarellou H, Rossolini GM. Metallo-β-lactamases: a last frontier for β-lactams? Lancet Infect Dis. 2011;11(5):381-93. Corrêa LL, Botelho LA, Barbosa LC, Mattos CS, Carballido JM, de Castro CL, Mondino PJ, de Paula GR, de Mondino SS, de Mendonça-Souza CR. Detection of bla(OXA-23) in Acinetobacter spp. isolated from patients of a university hospital. Braz J Infect Dis. 2012;16(6):521-6. Corvec S, Caroff N, Espaze E, Giraudeau C, Drugeon H, Reynaud A. AmpC cephalosporinase hyperproduction in Acinetobacter baumannii clinical strains. J Antimicrob Chemother. 2003;52(4):629-35. Coyne S, Guigon G, Courvalin P, Périchon B. Screening and quantification of the expression of antibiotic resistance genes in Acinetobacter baumannii with a microarray. Antimicrob Agents Chemother. 2010a;54(1):333-40. Coyne S, Rosenfeld N, Lambert T, Courvalin P, Périchon B. Overexpression of resistance-nodulation-cell division pump AdeFGH confers multidrug resistance in Acinetobacter baumannii. Antimicrob Agents Chemother. 2010b;54(10):4389-93. Coyne S, Courvalin P, Périchon B. Efflux-mediated antibiotic resistance in Acinetobacter spp. Antimicrob Agents Chemother. 2011;55(3):947-53. Dalla-Costa LM, Coelho JM, Souza HA, Castro ME, Stier CJ, Bragagnolo KL, Rea-Neto A, Penteado-Filho SR, Livermore DM, Woodford N. Outbreak of carbapenem-resistant Acinetobacter baumannii producing the OXA-23 enzyme in Curitiba, Brazil. J Clin Microbiol. 2003;41(7):3403-6.

REFERÊNCIAS BIBLIOGRÁFICAS
108
Damier-Piolle L, Magnet S, Brémont S, Lambert T, Courvalin P. AdeIJK, a resistance-nodulation-cell division pump effluxing multiple antibiotics in Acinetobacter baumannii. Antimicrob Agents Chemother. 2008;52(2):557-62. Davis KA, Moran KA, McAllister CK, Gray PJ. Multidrug-resistant Acinetobacter extremity infections in soldiers. Emerg Infect Dis. 2005;11(8):1218-24. Den Blaauwen T, Aarsman ME, Vischer NO, Nanninga N. Penicillin-binding protein PBP2 of Escherichia coli localizes preferentially in the lateral wall and at mid-cell in comparison with the old cell pole. Mol Microbiol. 2003;47(2):539-47. Diancourt L, Passet V, Nemec A, Dijkshoorn L, Brisse S. The population structure of Acinetobacter baumannii: expanding multiresistant clones from an ancestral susceptible genetic pool. PLoS One. 2010;5(4):e10034. Dijkshoorn L, Michel MF, Degener JE. Cell envelope protein profiles of Acinetobacter calcoaceticus strains isolated in hospitals. J Med Microbiol. 1987;23(4):313-9. Dijkshoorn L,Tjernberg I, Pot B, Michel MF,Ursing J,Kersters K.Numerical analysis of cell envelope protein profiles of Acinetobacter strains classified by DNA-DNA hybridization. Systematic and Appllied Microbiology. 1990;13(4):338-44. Dijkshoorn L, van Aken E, Shunburne L, van der Reijden TJ, Bernards AT, Nemec A, Towner KJ. Prevalence of Acinetobacter baumannii and other Acinetobacter spp. in faecal samples from non-hospitalised individuals. Clin Microbiol Infect. 2005;11(4):329-32. Dijkshoorn L, Nemec A, Seifert H. An increasing threat in hospitals: multidrug-resistant Acinetobacter baumannii. Nat Rev Microbiol. 2007;5(12):939-51. Dijkshoorn L, Nemec A. The diversity of the genus Acinetobacter. In: Acinetobacter Molecular Microbiology by Ulrike Gerische. 1 ed. Caister Academic Press, Norfolk, UK, 2008;1-35 Dizbay M, Tozlu DK, Cirak MY, Isik Y, Ozdemir K, Arman D.In vitro synergistic activity of tigecycline and colistin against XDR-Acinetobacter baumannii.J Antibiot (Tokyo). 2010;63(2):51–3. Doi Y, Adams JM, Yamane K, Paterson DL. Identification of 16S rRNA methylase-producing Acinetobacter baumannii clinical strains in North America. Antimicrob Agents Chemother. 2007;51(11):4209-10. Doi Y, Arakawa Y. 16S ribosomal RNA methylation: emerging resistance mechanism against aminoglycosides. Clin Infect Dis. 2007;45(1):88-94. Dolzani L, Tonin E, Lagatolla C, Prandin L, Monti-Bragadin C. Identification of Acinetobacter isolates in the A. calcoaceticus-A. baumannii complex by restriction analysis of the 16S-23S rRNA intergenic-spacer sequences. J Clin Microbiol. 1995;33(5):1108-13. Donald HM, Scaife W, Amyes SG, Young HK. Sequence analysis of ARI-1, a novel OXA β-lactamase, responsible for imipenem resistance in Acinetobacter baumannii 6B92. Antimicrob Agents Chemother. 2000;44(1):196-9

REFERÊNCIAS BIBLIOGRÁFICAS
109
Donlan RM, Costerton JW. Biofilms: survival mechanisms of clinically relevant microorganisms. Clin Microbiol Rev. 2002;15(2):167-93. Drawz SM, Bonomo RA. Three decades of β-lactamase inhibitors. Clin Microbiol Rev. 2010;23(1):160-201. Dupont M, Pagès JM, Lafitte D, Siroy A, Bollet C. Identification of an OprD homologue in Acinetobacter baumannii. J Proteome Res. 2005;4(6):2386-90. Endimiani A, Luzzaro F, Migliavacca R, Mantengoli E, Hujer AM, Hujer KM, Pagani L, Bonomo RA, Rossolini GM, Toniolo A. Spread in an Italian hospital of a clonal Acinetobacter baumannii strain producing the TEM-92 extended-spectrum β-lactamase. Antimicrob Agents Chemother. 2007;51(6):2211-4 Entenza JM, Moreillon P. Tigecycline in combination with other antimicrobials: a review of in vitro, animal and case report studies. Int J Antimicrob Agents. 2009;34(8):1–9. Ergin A, Hascelik G, Eser OK. Molecular characterization of oxacillinases and genotyping of invasive Acinetobacter baumannii isolates using repetitive extragenic palindromic sequence-based polymerase chain reaction in Ankara between 2004 and 2010. Scand J Infect Dis. 2013;45(1):26-31. Ermertcan S, Hoşgör M, Tünger O, Coşar G. Investigation of synergism of meropenem and ciprofloxacin against Pseudomonas aeruginosa and Acinetobacter strains isolated from intensive care unit infections. Scand J Infect Dis. 2001;33(11):818-21 Espinal P, Seifert H, Dijkshoorn L, Vila J, Roca I. Rapid and accurate identification of genomic species from the Acinetobacter baumannii (Ab) group by MALDI-TOF MS. Clin Microbiol Infect. 2012;18(11):1097-103. EUCAST. European Committee on Antimicrobial Susceptibility Testing. Breakpoint tables for interpretation of MICs and zone diameters. Version 2.0. Växjö, Sweden. 2012 Fagon JY, Chastre J, Domart Y, Trouillet JL, Gibert C. Mortality due to ventilator-associated pneumonia or colonization with Pseudomonas or Acinetobacter species: assessment by quantitative culture of samples obtained by a protected specimen brush. Clin Infect Dis. 1996;23(3):538-42. Evans BA, Hamouda A, Amyes SG. The rise of carbapenem-resistant Acinetobacter baumannii. Curr Pharm Des. 2013;19(2):223-38. Falagas ME, Bliziotis IA, Tam VH. Intraventricular or intrathecal use of polymyxins in patients with Gram-negative meningitis: a systematic review of the available evidence. Int J Antimicrob Agents. 2007;29(1):9-25. Falagas ME, Rafailidis PI, Ioannidou E, Alexiou VG, Matthaiou DK, Karageorgopoulos DE, Kapaskelis A, Nikita D, Michalopoulos A. Colistin therapy for microbiologically documented multidrug-resistant Gram-negative bacterial infections: a retrospective cohort study of 258 patients. Int J Antimicrob Agents. 2010;35(2):194-9. Feizabadi MM, Fathollahzadeh B, Taherikalani M, Rasoolinejad M, Sadeghifard N, Aligholi M, Soroush S, Mohammadi-Yegane S. Antimicrobial susceptibility patterns and distribution of blaOXA genes among Acinetobacter spp. isolated from patients at Tehran hospitals. J Infect Dis. 2008;61(4):274-8.

REFERÊNCIAS BIBLIOGRÁFICAS
110
Fernández-Canigia L, Dowzicky MJ. Susceptibility of important Gram-negative pathogens to tigecycline and other antibiotics in Latin America between 2004 and 2010. Ann Clin Microbiol Antimicrob. 2012;22;11:29. Fernández-Cuenca F, Martínez-Martínez L, Conejo MC, Ayala JA, Perea EJ, Pascual A. Relationship between β-lactamase production, outer membrane protein and penicillin-binding protein profiles on the activity of carbapenems against clinical isolates of Acinetobacter baumannii. J Antimicrob Chemother. 2003a;51(3):565-74. Fernández-Cuenca F, Martínez-Martínez L, Pascual A, Perea EJ. In vitro activity of azithromycin in combination with amikacin, ceftazidime, ciprofloxacin or imipenem against clinical isolates of Acinobacter baumannii. Chemotherapy. 2003b;49(1-2):24-6. Fernández-Cuenca F, Pascual A, Ribera A, Vila J, Bou G, Cisneros JM, Rodríguez-Baño J, Pachón J, Martínez-Martínez L; Grupo de Estudio de Infección Hospitalaria. [Clonal diversity and antimicrobial susceptibility of Acinetobacter baumannii isolated in Spain. A nationwide multicenter study: GEIH-Ab project (2000)]. Enfermedades Infecciosas y Microbiología Clínica. 2004;22(5): 267-271. Fernández-Cuenca F, Tomás-Carmona M, Caballero-Moyano F, Bou G, Martínez- Martínez L, Vila J, Pachón J, Cisneros JM, Rodríguez-Baño J, Pascual A; grupo del proyecto GEIH-REIPI-Ab 2010. In vitro activity of 18 antimicrobial agents against clinical isolates of Acinetobacter spp.: multicenter nationalstudy GEIH-REIPI-Ab 2010. Enferm Infecc Microbiol Clin. 2013;31(1):4-9. Ferreira AE, Marchetti DP, De Oliveira LM, Gusatti CS, Fuentefria DB, Corção G. Presence of OXA-23-producing isolates of Acinetobacter baumannii in wastewater from hospitals in southern Brazil. Microb Drug Resist. 2011;17(2):221-7. Figueiredo S, Poirel L, Papa A, Koulourida V, Nordmann P. Overexpression of the naturally occurring blaOXA-51 gene in Acinetobacter baumannii mediated by novel insertion sequence ISAba9. Antimicrob Agents Chemother. 2009;53(9):4045-7 Froment Gomis P, Jean-Pierre H, Rousseau-Didelot MN, Compan B, Michon AL, Godreuil S. Tigecycline: CMI 50/90 towards 1766 Gram-negative bacilli (3rd generation cephalosporins resistant enterobacteriaceae), Acinetobacter baumannii and Bacteroides fragilis group, University Hospital - Montpellier, 2008-2011. Pathol Biol (Paris). 2013 Mar 7. doi:pii: S0369-8114(13)00041-2. Fu Y, Zhou J, Zhou H, Yang Q, Wei Z, Yu Y, Li L. Wide dissemination of OXA-23-producing carbapenem-resistant Acinetobacter baumannii clonal complex 22 in multiple cities of China. J Antimicrob Chemother. 2010;65(4):644-50. Furtado GH, Cavalcante AC, Medeiros EA, Gales AC, Oliveira VG, Girardello R. Bloodstream infections with OXA-23-producing Acinetobacter baumannii isolates in a university-affiliated hospital in Brazil: epidemiology and clinical outcomes. Am J Infect Control. 2011;39(8):706-8. Gaddy JA, Actis LA. Regulation of Acinetobacter baumannii biofilm formation. Future Microbiol. 2009;4(3):273-8. Gales AC, Reis AO, Jones RN. Contemporary assessment of antimicrobial susceptibility testing methods for polymyxin B and colistin: review of available interpretative criteria and quality control guidelines. J Clin Microbiol. 2001;39(1):183-90.

REFERÊNCIAS BIBLIOGRÁFICAS
111
Gales AC, Tognim MC, Reis AO, Jones RN, Sader HS. Emergence of an IMP-like metallo-enzyme in an Acinetobacter baumannii clinical strain from a Brazilian teaching hospital. Diagn Microbiol Infect Dis. 2003;45(1):77-9. Gales AC, Jones RN, Sader HS. Global assessment of the antimicrobial activity of polymyxin B against 54 731 clinical isolates of Gram-negative bacilli: report from the SENTRY antimicrobial surveillance programme (2001-2004). Clin Microbiol Infect. 2006;12(4):315-321. Gales AC, Azevedo HD, Cereda RF, Girardello R, Xavier DE; INVITA-A-DORI Brazilian Study Group. Antimicrobial activity of doripenem against Gram-negative pathogens: results from INVITA-A-DORI Brazilian study. Braz J Infect Dis. 2011;15(6):513-20. Gales AC, Castanheira M, Jones RN, Sader HS. Antimicrobial resistance among Gram-negative bacilli isolated from Latin America: results from SENTRY Antimicrobial Sueveillance Program (Latin America, 2008-2010). Diagnostic Microbiology and Infectious Diseases. 2012;73(4):354-60. Gallagher JC, Rouse HM. Tigecycline for the treatment of Acinetobacter infections: a case series. Ann Pharmacother. 2008;42(9):1188–94. Garnacho J, Sole-Violan J, Sa-Borges M, Diaz E, Rello J. Clinical impact of pneumonia caused by Acinetobacter baumannii in intubated patients: a matched cohort study. Crit Care Med. 2003;31(10):2478-82 Garnacho-Montero J, Ortiz-Leyba C, Jiménez-Jiménez FJ, Barrero-Almodóvar AE, García-Garmendia JL, Bernabeu-WittelI M, Gallego-Lara SL, Madrazo-Osuna J. Treatment of multidrug-resistant Acinetobacter baumannii ventilator-associated pneumonia (VAP) with intravenous colistin: a comparison with imipenem-susceptible VAP. Clin Infect Dis. 2003;36(9):1111-8. Garrison MW, Mutters R, Dowzicky MJ. In vitro activity of tigecycline and comparator agents against a global collection of Gram-negative and Gram-positive organisms: tigecycline Evaluation and Surveillance Trial 2004 to 2007. Diagnostic Microbiology and Infectious Disease. 2009;65(3): 288-299. Garzoni C, Emonet S, Legout L, Benedict R, Hoffmeyer P, Bernard L, Garbino J. Atypical infections in tsunami survivors. Emerg Infect Dis. 2005;11(10):1591-3. Gerner-Smidt P, Tjernberg I, Ursing J. Reliability of phenotypic tests for identification of Acinetobacter species.J Clin Microbiol. 1991;29(2):277-82. Ghazawi A, Sonnevend A, Bonnin RA, Poirel L, Nordmann P, Hashmey R, Rizvi TA, B Hamadeh M, Pál T. NDM-2 carbapenemase-producing Acinetobacter baumannii in the United Arab Emirates. Clin Microbiol Infect. 2012;18(2):34-6. Ghosh AS, Chowdhury C, Nelson DE. Physiological functions of D-alanine carboxypeptidases in Escherichia coli. Trends Microbiol. 2008;16(7):309-17. Gionco B, Pelayo JS, Venancio EJ, Cayô R, Gales AC, Carrara-Marroni FE. Detection of OXA-231, a new variant of blaOXA-143, in Acinetobacter baumannii from Brazil: a case report. J Antimicrob Chemother. 2012;67(10):2531-2. Girardello R – Tese de doutorado - Caracterização dos mecanismos de resistência à polimixina B em isolados clínicos de Acinetobacter baumannii. 2012. 14 pp.

REFERÊNCIAS BIBLIOGRÁFICAS
112
Gounden R, Bamford C, van Zyl-Smit R, Cohen K, Maartens G. Safety and effectiveness of colistin compared with tobramycin for multi-drug resistant Acinetobacter baumannii infections. BMC Infect Dis. 2009;9:26. Gribun A, Nitzan Y, Pechatnikov I, Hershkovits G, Katcoff DJ. Molecular and structural characterization of the HMP-AB gene encoding a pore-forming protein from a clinical isolate of Acinetobacter baumannii. Curr Microbiol. 2003;47(5):434-43. Griffith EM, Yun CH, Horvath LL, Murray KC. Minocycline therapy for traumatic wound infections caused by the multidrug-resistant Acinetobacter baumannii-Acinetobacter calcoaceticus complex.Inf Dis Clin Pract. 2008;16(1):16-9. Grosso F, Carvalho KR, Quinteira S, Ramos A, Carvalho-Assef AP, Asensi MD, Peixe L. OXA-23-producing Acinetobacter baumannii: a new hotspot of diversity in Rio de Janeiro? J Antimicrob Chemother. 2011; 66(1):62-5. Gundi VA, Dijkshoorn L, Burignat S, Raoult D, La Scola B. Validation of partial rpoB gene sequence analysis for the identification of clinically important and emerging species. Microbiology. 2009;155(7):2333-41. Hammami S, Ghozzi R, Saidani M, Ben Redjeb S. Carbapenem-resistant Acinetobacter baumannii producing the carbapenemase OXA-23 in Tunisia. Tunis Med. 2011;89(7):638-43. Hancock RE. Peptide antibiotics. Lancet.1997;349(9049):418-422. Héritier C, Poirel L, Lambert T, Nordmann P. Contribution of acquired carbapenem-hydrolyzing oxacillinases to carbapenem resistance in Acinetobacter baumannii. Antimicrob Agents Chemother. 2005a;49(8):3198-202. Héritier C, Poirel L, Fournier PE, Claverie JM, Raoult D, Nordmann P. Characterization of the naturally occurring oxacillinase of Acinetobacter baumannii. Antimicrob Agents Chemother. 2005b;49(10):4174-9. Héritier C, Poirel L, Nordmann P. Cephalosporinase over-expression resulting from insertion of ISAba1 in Acinetobacter baumannii. Clin Microbiol Infect. 2006;12(2):123-30. Higgins PG, Wisplinghoff H, Stefanik D, Seifert H. Selection of topoisomerase mutations and overexpression of adeB mRNA transcripts during an outbreak of Acinetobacter baumannii. J Antimicrob Chemother. 2004;54(4):821-3. Higgins PG, Poirel L, Lehmann M, Nordmann P, Seifert H. OXA-143, a novel carbapenem-hydrolyzing class D β-lactamase in Acinetobacter baumannii. Antimicrob Agents Chemother. 2009;53(12):5035-8. Higgins PG, Dammhayn C, Hackel M, Seifert H. Global spread of carbapenem-resistant Acinetobacter baumannii. J Antimicrob Chemother. 2010;65(2):233-8. Higgins PG, Pérez-Llarena FJ, Zander E, Fernández A, Bou G, Seifert H. OXA-235, a novel Class D β-Lactamase involved in resistance to carbapenems in Acinetobacter baumannii. Antimicrob Agents Chemother. 2013;57(5):2121-6.

REFERÊNCIAS BIBLIOGRÁFICAS
113
Hirsch EB, Tam VH. Detection and treatment options for Klebsiella pneumoniae carbapenemases (KPCs): an emerging cause of multidrug-resistant infection. J Antimicrob Chemother. 2010;65(6):1119-25. Holloway KP, Rouphael NG, Wells JB, King MD, Blumberg HM. Polymyxin B and doxycycline use in patients with multidrug-resistant Acinetobacter baumannii infections in the intensive care unit. Ann Pharmacother. 2006;40(11):1939-45. Hong H, Patel DR, Tamm LK, van den Berg B. The outer membrane protein OmpW forms an eight-stranded β-barrel with a hydrophobic channel. J Biol Chem. 2006; 281(11):7568-77 Hong MK, Lee JH, Kwon DB, Kim JK, Tran TH, Nguyen DD, Jeong BC, Lee SH, Kang LW. Crystallization and preliminary diffraction studies of GIM-1, a class B carbapenem-hydrolyzing β-lactamase. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2012; 68(10):1226-8. Hood JD, Box SJ, Verrall MS. Olivanic acids, a family of β-lactam antibiotics with β-lactamase inhibitory properties produced by Streptomyces species. II. Isolation and characterisation of the olivanic acids MM 4550, MM 13902 and MM 17880 from Streptomyces olivaceus. J Antibiot (Tokyo). 1979;32(4):295-304 Hou PF, Chen XY, Yan GF, Wang YP, Ying CM. Study of the correlation of imipenem resistance with efflux pumps AdeABC, AdeIJK, AdeDE and AbeM in clinical isolates of Acinetobacter baumannii. Chemotherapy. 2012;58(2):152-8. Howard A, O'Donoghue M, Feeney A, Sleator RD. Acinetobacter baumannii: An emerging opportunistic pathogen. Virulence. 2012;3(3):243-50. Hujer KM, Hamza NS, Hujer AM, Perez F, Helfand MS, Bethel CR, Thomson JM, Anderson VE, Barlow M, Rice LB, Tenover FC, Bonomo RA. Identification of a new allelic variant of the Acinetobacter baumannii cephalosporinase, ADC-7 β-lactamase: defining a unique family of class C enzymes. Antimicrob Agents Chemother. 2005; 49(7):2941-8. Hujer KM, Hujer AM, Hulten EA, Bajaksouzian S, Adams JM, Donskey CJ, Ecker DJ, Massire C, Eshoo MW, Sampath R, Thomson JM, Rather PN, Craft DW, Fishbain JT, Ewell AJ, Jacobs MR, Paterson DL, Bonomo RA. Analysis of antibiotic resistance genes in multidrug-resistant Acinetobacter sp. isolates from military and civilian patients treated at the Walter Reed Army Medical Center. Antimicrob Agents Chemother. 2006;50(12):4114-23. Huys G, Cnockaert M, Vaneechoutte M, Woodford N, Nemec A, Dijkshoorn L, Swings J. Distribution of tetracycline resistance genes in genotypically related and unrelated multiresistant Acinetobacter baumannii strains from different European hospitals. Res Microbiol. 2005a;156(3):348-55. Huys G, Cnockaert M, Nemec A, Swings J. Sequence-based typing of adeB as a potential tool to identify intraspecific groups among clinical strains of multidrug-resistant Acinetobacter baumannii. J Clin Microbiol. 2005b;43(10):5327-31. Ibrahim A, Gerner-Smidt P, Liesack W. Phylogenetic relationship of the twenty-one DNA groups of the genus Acinetobacter as revealed by 16S ribosomal DNA sequence analysis. Int J Syst Bacteriol. 1997;47(3):837-41.

REFERÊNCIAS BIBLIOGRÁFICAS
114
Ikonomidis A, Pournaras S, Markogiannakis A, Maniatis AN, Manolis EN, Tsakris A. Low genetic diversity of the intrinsic OXA-51-like class D carbapenemases among Acinetobacter baumannii clinical isolates in Greece. Epidemiol Infect. 2007;135(5):877-81. Jacoby GA. AmpC β-lactamases. Clin.Microbiol. Rev. 2009;22(1):161-82. Janssen P, Coopman R, Huys G, Swings J, Bleeker M, Vos P, Zabeau M, Kersters K. Evaluation of the DNA fingerprinting method AFLP as an new tool in bacterial taxonomy. Microbiology. 1996;142 (7):1881-93. Jellison TK, Mckinnon PS, Rybak MJ. Epidemiology, resistance, and outcomes of Acinetobacter baumannii bacteremia treated with imipenem-cilastatin or ampicillin-sulbactam. Pharmacotherapy. 2001;21(2):142-8. Jeon BC, Jeong SH, Bae IK, Kwon SB, Lee K, Young D, Lee JH, Song JS, Lee SH. Investigation of a nosocomial outbreak of imipenem-resistant Acinetobacter baumannii producing the OXA-23 β-lactamase in Korea. J Clin Microbiol. 2005;43(5):2241-5. Jiménez-Mejías ME, Pachón J, Becerril B, Palomino-Nicás J, Rodríguez-Cobacho A, Revuelta M. Treatment of multidrug-resistant Acinetobacter baumannii meningitis with ampicillin/sulbactam. Clin Infect Dis. 1997;24(5):932-5. Johnson EN, Burns TC, Hayda RA, Hospenthal DR, Murray CK. Infectious complications of open type III tibial fractures among combat casualties. Clin Infect Dis. 2007;45(4):409-15. Jones RN. Important and emerging β-lactamase-mediated resistances in hospital-based pathogens: the AmpC enzymes. Diagn Microbiol Infect Dis.1998; 31(3):461-6. Jones RN, Ferraro MJ, Reller LB, Schreckenberger PC, Swenson JM, Sader HS. Multicenter studies of tigecycline disk diffusion susceptibility results for Acinetobacter spp. J Clin Microbiol. 2007;45(1):227-30. Kahan JS, Kahan FM, Goegelman R, Currie SA, Jackson M, Stapley EO, Miller TW, Miller AK, Hendlin D, Mochales S, Hernandez S, Woodruff HB, Birnbaum J. Thienamycin, a new β-lactam antibiotic. I. Discovery, taxonomy, isolation and physical properties. J Antibiot (Tokyo). 1979;32(1):1-12. Kallel H, Hergafi L, Bahloul M, Hakim A, Dammak H, Chelly H, Hamida CB, Chaari A, Rekik N, Bouaziz M. Safety and efficacy of colistin compared with imipenem in the treatment of ventilator-associated pneumonia: a matched case-control study. Intensive Care Med. 2007;33(7):1162-7. Karageorgopoulos DE, Falagas ME. Current control and treatment of multidrug-resistant Acinetobacter baumannii infections. Lancet Infect Dis. 2008;8(12):751-62. Karah N, Giske CG, Sundsfjord A, Samuelsen Ø. A diversity of OXA-carbapenemases and class 1 integrons among carbapenem-resistant Acinetobacter baumannii clinical isolates from Sweden belonging to different international clonal lineages. Microb Drug Resist. 2011;17(4):545-9. Kasiakou SK, Michalopoulos A, Soteriades ES, Samonis G, Sermaides GJ, Falagas ME. Combination therapy with intravenous colistin for management of infections due to

REFERÊNCIAS BIBLIOGRÁFICAS
115
multidrug-resistant Gram-negative bacteria in patients without cystic fibrosis. Antimicrob Agents Chemother. 2005;49(8):3136-46. Katragkou A, Kotsiou M, Antachopoulos C, Benos A, Sofianou D, Tamiolaki M, Roilides E. Acquisition of imipenem-resistant Acinetobacter baumannii in a pediatric intensive care unit: A case-control study. Intensive Care Med. 2006;32(9):1384-91. Kattan JN, Villegas MV, Quinn JP. New developments in carbapenems. Clin Microbiol Infect. 2008;14(12):1102-11. Kau HC, Tsai CC, Kao SC, Hsu WM, Liu JH. Corneal ulcer of the side port after phaco emulsification induced by Acinetobacter baumannii. J Cataract Refract Surg. 2002;28(5):895-7. Keen EF 3rd, Robinson BJ, Hospenthal DR, Aldous WK, Wolf SE, Chung KK, Murray CK. Prevalence of multidrug-resistant organisms recovered at a military burn center. Burns. 2010;36(6):819-25. Kiffer CR, Sampaio JL, Sinto S, Oplustil CP, Koga PC, Arruda AC, Turner PJ, Mendes C. In vitro synergy test of meropenem and sulbactam against clinical isolates of Acinetobacter baumannii. Diagn Microbiol Infect Dis. 2005;52(4):317-22. Kim CK, Lee Y, Lee H, Woo GJ, Song W, Kim MN, Lee WG, Jeong SH, Lee K, Chong Y. Prevalence and diversity of carbapenemases among imipenem-nonsusceptible Acinetobacter isolates in Korea: emergence of a novel OXA-182. Diagn Microbiol Infect Dis. 2010;68(4):432-8. Ko WC, Lee HC, Chiang SR, Yan JJ, Wu JJ, Lu CL, Chuang YC. In vitro and in vivo activity of meropenem and sulbactam against a multidrug-resistant Acinetobacter baumannii strain. J Antimicrob Chemother. 2004;53(2):393-5. Ko KS, Suh JY, Kwon KT, Jung SI, Park KH, Kang CI, Chung DR, Peck KR, Song JH. High rates of resistance to colistin and polymyxin B in subgroups of Acinetobacter baumannii isolates from Korea. Journal of Antimicrobial Chemotherapy. 2007;60(5): 1163-1167. Koch-Weser J, Sidel VW, Federman EB, Kanarek P, Finer DC, Eaton AE. Adverse effects of sodium colistimethate. Manifestations and specific reaction rates during 317 courses of therapy. Ann Intern Med. 1970;72(6):857-68. Kohlenberg A, Brümmer S, Higgins PG, Sohr D, Piening BC, de Grahl C, Halle E, Rüden H, Seifert H. Outbreak of carbapenem-resistant Acinetobacter baumannii carrying the carbapenemase OXA-23 in a German university medical centre. J Med Microbiol. 2009;58(11):1499-507. Kouyama Y, Harada S, Ishii Y, Saga T, Yoshizumi A, Tateda K, Yamaguchi K. Molecular characterization of carbapenem-non-susceptible Acinetobacter spp. In Japan: predominance of multidrug-resistant Acinetobacter baumannii clonal complex 92 and IMP-type metallo-β-lactamase-producing non-baumannii Acinetobacter species. J Infect Chemother. 2012;18(4):522-8. Krizova L, Dijkshoorn L, Nemec A. Diversity and evolution of AbaR genomic resistance islands in Acinetobacter baumannii strains of European clone I. Antimicrob Agents Chemother. 2011;55(7):3201-6.

REFERÊNCIAS BIBLIOGRÁFICAS
116
Krizova L, Bonnin RA, Nordmann P, Nemec A, Poirel L. Characterization of a multidrug-resistant Acinetobacter baumannii strain carrying the blaNDM-1 and blaOXA-23 carbapenemase genes from the Czech Republic. J Antimicrob Chemother. 2012;67(6):1550-2. Kuga A, Okamoto R, Inoue M. ampR gene mutations that greatly increase class C β-lactamase activity in Enterobacter cloacae. Antimicrob Agents Chemother. 2000;44(3):561-7. Kusradze Ia, Diene SM, Goderdzishvili M, Rolain JM. Molecular detection of OXA carbapenemase genes in multidrug-resistant Acinetobacter baumannii isolates from Iraq and Georgia. Int J Antimicrob Agents. 2011;38(2):164-8. La Scola B, Raoult D. Acinetobacter baumannii in human body louse. Emerg Infect Dis. 2004;10(9):1671-3. La Scola B, Gundi VA, Khamis A, Raoult D. Sequencing of the rpoB gene and flanking spacers for molecular identification of Acinetobacter species. J Clin Microbiol. 2006;44(3):827-32. Landman D, Georgescu C, Martin DA, Quale J. Polymyxins revisited. Clin Microbiol Rev. 2008;21(3):449-65. Le Hello S, Falcot V, Lacassin F, Baumann F, Nordmann P, Naas T. Molecular epidemiology of carbapenem-resistant Acinetobacter baumannii in New Caledonia. Clin Microbiol Infect. 2008;14(10):977-81. Lee K, Kim MN, Choi TY, Cho SE, Lee S, Whang DH, Yong D, Chong Y, Woodford N, Livermore DM; KONSAR Group. Wide dissemination of OXA-type carbapenemases in clinical Acinetobacter spp. isolates from South Korea. Int J Antimicrob Agents. 2009;33(6):520-4. Lee Y, Kim CK, Lee H, Jeong SH, Yong D, Lee K. A novel insertion sequence, ISAba10, inserted into ISAba1 adjacent to the bla(OXA-23) gene and disrupting the outer membrane protein gene carO in Acinetobacter baumannii. Antimicrob Agents Chemother. 2011;55(1):361-3. Lee YT, Kuo SC, Chiang MC, Yang SP, Chen CP, Chen TL, Fung CP. Emergence of carbapenem-resistant non-baumannii species of Acinetobacter harboring a blaOXA-51-like
gene that is intrinsic to A. baumannii. Antimicrob Agents Chemother. 2012;56(2):1124-7. Leung WS, Chu CM, Tsang KY, Lo FH, Lo KF, Ho PL. Fulminant community-acquired Acinetobacter baumannii pneumonia as a distinct clinical syndrome. Chest. 2006;129(1):102-9. Levin AS, Barone AA, Penço J, Santos MV, Marinho IS, Arruda EA, Manrique EI, Costa SF. Intravenous colistin as therapy for nosocomial infections caused by multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii. Clin Infect Dis. 1999;28(5):1008-11. Levin AS. Multiresistant Acinetobacter infections: a role for sulbactam combinations in overcoming an emerging worldwide problem. Clin Microbiol Infect. 2002;8(3):144-53. Review.

REFERÊNCIAS BIBLIOGRÁFICAS
117
Levin AS, Levy CE, Manrique AE, Medeiros EA, Costa SF. Severe nosocomial infections with imipenem-resistant Acinetobacter baumannii treated with ampicillin/sulbactam. Int J Antimicrob Agents. 2003;21(1):58-62. Levin AS. Treatment of Acinetobacter spp infections. Expert Opin Pharmacother. 2003;4(8):1289-96. Review. Levin AS, Oliveira MS. The challenge of multidrug resistance: the treatment of gram-negative rod infections. Shock. 2008;30 Suppl 1:30-3. Li J, Rayner CR, Nation RL, Owen RJ, Spelman D, Tan KE, Liolios L. Heteroresistance to colistin in multidrug-resistant Acinetobacter baumannii. Antimicrob Agents Chemother. 2006;50(9):2946-50. Lim TP, Tan TY, Lee W, Sasikala S, Tan TT, Hsu LY, Kwa AL. In vitro activity of various combinations of antimicrobials against carbapenem-resistant Acinetobacter species in Singapore. J Antibiot (Tokyo). 2009; 62(12):675-9. Limansky AS, Mussi MA, Viale AM. Loss of a 29-kilodalton outer membrane protein in Acinetobacter baumannii is associated with imipenem resistance. J Clin Microbiol. 2002;40(12):4776-8 Lin YC, Hsia KC, Chen YC, Sheng WH, Chang SC, Liao MH, Li SY. Genetic basis of multidrug resistance in Acinetobacter clinical isolates in Taiwan. Antimicrob Agents Chemother. 2010;54(5):2078-84. Livermore DM. Determinants of the activity of β-lactamase inhibitor combinations. J Antimicrob Chemother. 1993;31(Suppl A):9-21.
Livermore DM. -Lactamases in laboratory and clinical resistance. Clin. Microbiol. 1995;8(4):557-84. Lopes BS, Evans BA, Amyes SG. Disruption of the blaOXA-51-like gene by ISAba16 and activation of the blaOXA-58 gene leading to carbapenem resistance in Acinetobacter baumannii Ab244. J Antimicrob Chemother. 2012a;67(1):59-63. Lopes BS, Al-Hassan L, Amyes SG. ISAba825 controls the expression of the chromosomal bla(OXA-51-like) and the plasmid borne bla(OXA-58) gene in clinical isolates of Acinetobacter baumannii isolated from the USA. Clin Microbiol Infect. 2012b;18(11):E446-51. Lopez-Otsoa F, Gallego L, Towner KJ, Tysall L, Woodford N, Livermore DM. Endemic carbapenem resistance associated with OXA-40 carbapenemase among Acinetobacter baumannii isolates from a hospital in northern Spain. J Clin Microbiol. 2002;40(12):4741-3. Lowman W, Kalk T, Menezes CN, John MA, Grobusch MP. A case of community-acquired Acinetobacter baumannii meningitis - has the threat moved beyond the hospital? J Med Microbiol. 2008;57(5):676-8. Lu PL, Doumith M, Livermore DM, Chen TP, Woodford N. Diversity of carbapenem resistance mechanisms in Acinetobacter baumannii from a Taiwan hospital: spread of plasmid-borne OXA-72 carbapenemase. J Antimicrob Chemother. 2009;63(4):641-7.

REFERÊNCIAS BIBLIOGRÁFICAS
118
Maegele M, Gregor S, Steinhausen E, Bouillon B, Heiss MM, Perbix W, Wappler F, Rixen D, Geisen J, Berger-Schreck B, Schwarz R. The long-distance tertiary air transfer and care of tsunami victims: injury pattern and microbiological and psychological aspects. Crit Care Med. 2005;33(5):1136-40. Magnet S, Courvalin P, Lambert T. Resistance-nodulation-cell division-type efflux pump involved in aminoglycoside resistance in Acinetobacter baumannii strain BM4454. Antimicrob Agents Chemother. 2001;45(12):3375-80. Manageiro V, Jones-Dias D, Ferreira E, Louro D; Antimicrobial Resistance Surveillance Program in Portugal, Caniça M. Genetic diversity and clonal evolution of carbapenem-resistant Acinetobacter baumannii isolates from Portugal and the dissemination of ST118. Int J Antimicrob Agents. 2012;40(5):398-403. Maragakis LL, Perl TM. Acinetobacter baumannii: epidemiology, antimicrobial resistance, and treatment options. Clin Infect Dis. 2008;46(8):1254-63. Marchand I, Damier-Piolle L, Courvalin P, Lambert T. Expression of the RND-type efflux pump AdeABC in Acinetobacter baumannii is regulated by the AdeRS two-component system. Antimicrob Agents Chemother. 2004;48(9):3298-304. Marqué S, Poirel L, Héritier C, Brisse S, Blasco MD, Filip R, Coman G, Naas T, Nordmann P. Regional occurrence of plasmid-mediated carbapenem-hydrolyzing oxacillinase OXA-58 in Acinetobacter spp. in Europe. J Clin Microbiol. 2005;43(9):4885-8. Martí S, Fernández-Cuenca F, Pascual A, Ribera A, Rodríguez-Baño J, Bou G, Miguel Cisneros J, Pachón J, Martínez-Martínez L, Vila J; Grupo de Estudio de Infección Hospitalaria (GEIH). Prevalence of the tetA and tetB genes as mechanisms of resistance to tetracycline and minocycline in Acinetobacter baumannii clinical isolates. Enferm Infecc Microbiol Clin. 2006;24(2):77-80. Martins AF, Kuchenbecker R, Sukiennik T, Boff R, Reiter KC, Lutz L, Machado AB, Barth AL. Carbapenem-resistant Acinetobacter baumannii producing the OXA-23 enzyme: dissemination in Southern Brazil. Infection. 2009;37(5):474-6. Martins AF, Kuchenbecker RS, Pilger KO, Pagano M, Barth AL; CMCIES-PMPA/SMS Task Force. High endemic levels of multidrug-resistant Acinetobacter baumannii among hospitals in southern Brazil. Am J Infect Control. 2012;40(2):108-12. Martins N, Martins IS, de Freitas WV, de Matos JA, Magalhães AC, Girão VB, Dias RC, de Souza TC, Pellegrino FL, Costa LD, Boasquevisque CH, Nouér SA, Riley LW, Santoro-Lopes G, Moreira BM. Severe infection in a lung transplant recipient caused by donor-transmitted carbapenem-resistant Acinetobacter baumannii. Transpl Infect Dis. 2012;14(3):316-20. Martins N, Martins IS, de Freitas WV, de Matos JA, Girão VB, Coelho-Souza T, Maralhães AC, Cacci LC, de Figueiredo MP, Dias RC, Costa-Lourenço AP, Ferreira AL, Dalla-Costa L, Nouér SA, Santoro-Lopes G, Riley LW, Moreira BM. Imported and Intensive Care Unit-Born Acinetobacter baumannii Clonal Complexes: One-Year Prospective Cohort Study in Intensive Care Patients. Microb Drug Resist. 2013;19(3):216-23. Mataseje LF, Bryce E, Roscoe D, Boyd DA, Embree J, Gravel D, Katz K, Kibsey P, Kuhn M, Mounchili A, Simor A, Taylor G, Thomas E, Turgeon N, Mulvey MR.

REFERÊNCIAS BIBLIOGRÁFICAS
119
Carbapenem-resistant Gram-negative bacilli in Canada 2009-10: results from the Canadian Nosocomial Infection Surveillance Program (CNISP); Canadian Nosocomial Infection Surveillance Program. J Antimicrob Chemother. 2012;67(6):1359-67. Matthaiou DK, Michalopoulos A, Rafailidis PI, Karageorgopoulos DE, Papaioannou V, Ntani G, Samonis G, Falagas ME. Risk factors associated with the isolation of colistin-resistant gram-negative bacteria: a matched case-control study. Crit Care Med. 2008;36(3):807-11. Erratum in: Crit Care Med. 2008;36(7):2224. McConnell MJ, Actis L, Pachón J. Acinetobacter baumannii: human infections, factors contributing to pathogenesis and animal models. FEMS Microbiol Rev. 2013;37(2):130-55. Mendes RE, Castanheira M, Toleman MA, Sader HS, Jones RN, Walsh TR. Characterization of an integron carrying blaIMP-1 and a new aminoglycoside resistance gene, aac(6')-31, and its dissemination among genetically unrelated clinical isolates in a Brazilian hospital. Antimicrob Agents Chemother. 2007a;51(7):2611-4. Mendes RE, Kiyota KA, Monteiro J, Castanheira M, Andrade SS, Gales AC, Pignatari AC, Tufik S. Rapid detection and identification of metallo-β-lactamase-encoding genes by multiplex real-time PCR assay and melt curve analysis. J Clin Microbiol. 2007b;45(2):544-7. Mendes RE, Bell JM, Turnidge JD, Castanheira M, Jones RN. Emergence and widespread dissemination of OXA-23, -24/40 and -58 carbapenemases among Acinetobacter spp. in Asia-Pacific nations: report from the SENTRY Surveillance Program. J Antimicrob Chemother. 2009a;63(1):55-9. Mendes RE, Spanu T, Deshpande L, Castanheira M, Jones RN, Fadda G. Clonal dissemination of two clusters of Acinetobacter baumannii producing OXA-23 or OXA-58 in Rome, Italy. Clin Microbiol Infect. 2009b;15(6):588-92. Meric M, Kasap M, Gacar G, Budak F, Dundar D, Kolayli F, Eroglu C, Vahaboglu H. Emergence and spread of carbapenem-resistant Acinetobacter baumannii in a tertiary care hospital in Turkey. FEMS Microbiol Lett. 2008; 282(2):214-8. Merkier AK, Catalano M, Ramírez MS, Quiroga C, Orman B, Ratier L, Famiglietti A, Vay C, Di Martino A, Kaufman S, Centrón D. Polyclonal spread of blaOXA-23 and blaOXA-58
in Acinetobacter baumannii isolates from Argentina. J Infect Dev Ctries. 2008;2(3):235-40. Metan G, Alp E, Aygen B, Sumerkan B. Acinetobacter baumannii meningitis in post-neurosurgical patients: clinical outcome and impact of carbapenem resistance. J Antimicrob Chemother. 2007;60(1):197-9. Mezzatesta ML, D'Andrea MM, Migliavacca R, Giani T, Gona F, Nucleo E, Fugazza G, Pagani L, Rossolini GM, Stefani S. Epidemiological characterization and distribution of carbapenem-resistant Acinetobacter baumannii clinical isolates in Italy. Clin Microbiol Infect. 2012;18(2):160-6. Michalopoulos A, Kasiakou SK, Rosmarakis ES, Falagas ME. Cure of multidrug-resistant Acinetobacter baumannii bacteremia with continuous intravenous infusion of colistin. Scand J Infect Dis. 2005a;37(2):142-5.

REFERÊNCIAS BIBLIOGRÁFICAS
120
Michalopoulos AS, Tsiodras S, Rellos K, Mentzelopoulos S, Falagas ME. Colistin treatment in patients with ICU-acquired infections caused by multiresistant Gram-negative bacteria: the renaissance of an old antibiotic. Clin Microbiol Infect. 2005b;11(2):115-21. Miyadera T, Sugimura Y, Hashimoto T, Tanaka T, Iino K, Shibata T, Sugawara S. Synthesis and in vitro activity of a new carbapenem, RS-533. The Journal of Antibiotics. 1983;36(8):1034-9. Mostachio AK, Levin AS, Rizek C, Rossi F, Zerbini J, Costa SF. High prevalence of OXA-143 and alteration of outer membrane proteins in carbapenem-resistant Acinetobacter spp. isolates in Brazil. Int J Antimicrob Agents. 2012;39(5):396-401. Moubareck C, Brémont S, Conroy MC, Courvalin P, Lambert T. GES-11, a novel integron-associated GES variant in Acinetobacter baumannii. Antimicrob Agents Chemother. 2009;53(8):3579-81. Mugnier P, Poirel L, Pitout M, Nordmann P. Carbapenem-resistant and OXA-23-producing Acinetobacter baumannii isolates in the United Arab Emirates. Clin Microbiol Infect. 2008;14(9):879-82. Mugnier PD, Poirel L, Naas T, Nordmann P. Worldwide dissemination of the blaOXA-23 carbapenemase gene of Acinetobacter baumannii. Emerg Infect Dis. 2010;16(1):35-40. Murray CK, Roop SA, Hospenthal DR, Dooley DP, Wenner K, Hammock J, Taufen N, Gourdine E. Bacteriology of war wounds at the time of injury. Mil Med. 2006;171(9):826-9. Mussi MA, Limansky AS, Viale AM. Acquisition of resistance to carbapenems in multidrug-resistant clinical strains of Acinetobacter baumannii: natural insertional inactivation of a gene encoding a member of a novel family of β-barrel outer membrane proteins. Antimicrob Agents Chemother. 2005;49(4):1432-40. Naas T, Levy M, Hirschauer C, Marchandin H, Nordmann P. Outbreak of carbapenem-resistant Acinetobacter baumannii producing the carbapenemase OXA-23 in a tertiary care hospital of Papeete, French Polynesia. J Clin Microbiol. 2005;43(9):4826-9. Naas T, Namdari F, Réglier-Poupet H, Poyart C, Nordmann P. Panresistant extended-spectrum β-lactamase SHV-5-producing Acinetobacter baumannii from New York City. J Antimicrob Chemother. 2007;60(5):1174-6. Nagano N, Nagano Y, Cordevant C, Shibata N, Arakawa Y. Nosocomial transmission of CTX-M-2 β-lactamase-producing Acinetobacter baumannii in a neurosurgery ward. J Clin Microbiol. 2004;42(9):3978-84. Nemec A, Maixnerová M, van der Reijden TJ, van den Broek PJ, Dijkshoorn L. Relationship between the AdeABC efflux system gene content, netilmicin susceptibility and multidrug resistance in a genotypically diverse collection of Acinetobacter baumannii strains. J Antimicrob Chemother. 2007;60(3):483-9. Nemec A, Krízová L, Maixnerová M, Diancourt L, van der Reijden TJ, Brisse S, van den Broek P, Dijkshoorn L. Emergence of carbapenem resistance in Acinetobacter baumannii in the Czech Republic is associated with the spread of multidrug-resistant strains of European clone II. J Antimicrob Chemother. 2008;62(3):484-9.

REFERÊNCIAS BIBLIOGRÁFICAS
121
Neonakis IK, Spandidos DA, Petinaki E. Confronting multidrug-resistant Acinetobacter baumannii: a review. Int J Antimicrob Agents. 2011;37(2):102-9. Ni M, Zhang D, Qi J. Analysis of AmpC β-lactamase gene in Pseudomonas aeruginosa. J. Huazhong. Univ Sci. Technolog. Med. Sci. 2005;25(1):17-23. Nicolau DP. Carbapenems: a potente class of antibiotics. Expert Opinion on Pharmacotherapy. 2008;9(1):23-37. Nitzan Y, Pechatnikov I, Bar-El D, Wexler H. Isolation and characterization of heat-modifiable proteins from the outer membrane of Porphyromonas asaccharolytica and Acinetobacter baumannii. Anaerobe. 1999;5(1):43-50. Obara M, Nakae T. Mechanisms of resistance to β-lactam antibiotics in Acinetobacter calcoaceticus. J Antimicrob Chemother. 1991;28(6):791-800 Oliveira MS, Prado GV, Costa SF, Grinbaum RS, Levin AS. Ampicillin/sulbactam compared with polymyxins for the treatment of infections caused by carbapenem-resistant Acinetobacter spp. J Antimicrob Chemother. 2008;61(6):1369-75. Oliver A, Levin BR, Juan C, Baquero F, Blázquez J. Hypermutation and the preexistence of antibiotic-resistant Pseudomonas aeruginosa mutants: implications for susceptibility testing and treatment of chronic infections. Antimicrob Agents Chemother. 2004;48(11):4226-33. Olut AI, Erkek E. Early prosthetic valve endocarditis due to Acinetobacter baumannii: a case report and brief review of the literature. Scand J Infect Dis. 2005;37(11-12):919-21. Ozaki T, Nishimura N, Arakawa Y, Suzuki M, Narita A, Yamamoto Y, Koyama N, Nakane K, Yasuda N, Funahashi K. Community-acquired Acinetobacter baumannii meningitis in a previously healthy 14-month-old boy. J Infect Chemother. 2009;15(5):322-4. Pankuch GA, Lin G, Seifert H, Appelbaum PC. Activity of meropenem with and without ciprofloxacin and colistin against Pseudomonas aeruginosa and Acinetobacter baumannii. Antimicrobial Agents and Chemotherapy. 2008;52(1): 333-336. Pantophlet R, Severin JA, Nemec A, Brade L, Dijkshoorn L, Brade H. Identification of Acinetobacter isolates from species belonging to the Acinetobacter calcoaceticus-Acinetobacter baumannii complex with monoclonal antibodies specific for O Antigens of their lipopolysaccharides. ClinDiagn Lab Immunol. 2002;9(1):60-5. Papp-Wallace KM, Endimiani A, Taracila MA, Bonomo RA. Carbapenems: past, present, and future. Antimicrob Agents Chemother. 2011;55(11):4943-60. Pasterán F, Rapoport M, Petroni A, Faccone D, Corso A, Galas M, Vázquez M, Procopio A, Tokumoto M, Cagnoni V. Emergence of PER-2 and VEB-1a in Acinetobacter baumannii strains in the Americas. Antimicrob Agents Chemother. 2006;50(9):3222-4. Paton R, Miles RS, Hood J, Amyes SG, Miles RS, Amyes SG. ARI 1: β-lactamase-mediated imipenem resistance in Acinetobacter baumannii. Int J Antimicrob Agents. 1993;2(2):81-7.

REFERÊNCIAS BIBLIOGRÁFICAS
122
Pei G, Mao Y, Sun Y. In vitro activity of minocycline alone and in combination with cefoperazone-sulbactam against carbapenem-resistant Acinetobacter baumannii. Microb Drug Resist. 2012;18(6):574-7. Peleg AY, Potoski BA, Rea R, Adams J, Sethi J, Capitano B, Husain S, Kwak EJ, Bhat SV, Paterson DL. Acinetobacter baumannii bloodstream infection while receiving tigecycline: a cautionary report. J Antimicrob Chemother.2007a;59(1):128-31. Peleg AY, Adams J, Paterson DL. Tigecycline Efflux as a Mechanism for Non susceptibility in Acinetobacter baumannii. Antimicrob Agents Chemother. 2007b;51(6):2065-9. Peleg AY. Optimizing therapy for Acinetobacter baumannii. Semin Respir Crit Care Med. 2007c;28(6):662-71. Peleg AY, Seifert H, Paterson DL. Acinetobacter baumannii: emergence of a successful pathogen. Clin Microbiol Rev. 2008;21(3):538-82. Pereira AS. Determinação da Concentração de Antimicrobiano Capaz de Prevenir o Aparecimento de Mutantes Resistentes em Amostras Clínicas de Acinetobacter spp. Tese de Doutorado. Universidade Federal de São Paulo. São Paulo, 2009. 171 pp. Perez F, Hujer AM, Hujer KM, Decker BK, Rather PN, Bonomo RA. Global challenge of multidrug-resistant Acinetobacter baumannii. Antimicrob Agents Chemother. 2007;51(10):3471-84. Petersen PJ, Labthavikul P, Jones CH, Bradford PA. In vitro antibacterial activities of tigecycline in combination with other antimicrobial agents determined by chequer board and time-kill kinetic analysis.J Antimicrob Chemother. 2006;57(3):573-6. Petersen K, Riddle MS, Danko JR, Blazes DL, Hayden R, Tasker SA, Dunne JR. Trauma-related infections in battlefield casualties from Iraq. Ann Surg. 2007;245(5):803-11. Philippon A, Arlet G, Jacoby GA. Plasmid-determined AmpC-type β-lactamases. Antimicrob Agents Chemother. 2002;46(1):1-11. Piddock LJ. Clinically relevant chromosomally encoded multidrug resistance efflux pumps in bacteria. Clin Microbiol Rev. 2006;19(2):382-402. Piette A, Fraipont C, Den Blaauwen T, Aarsman ME, Pastoret S, Nguyen-Distèche M. Structural determinants required to target penicillin-binding protein 3 to the septum of Escherichia coli. J Bacteriol. 2004;186(18):6110-7. Poirel L, Menuteau O, Agoli N, Cattoen C, Nordmann P. Outbreak of extended-spectrum beta-lactamase VEB-1-producing isolates of Acinetobacter baumannii in a French hospital. J Clin Microbiol. 2003;41(8):3542-7. Poirel L, Marqué S, Héritier C, Segonds C, Chabanon G, Nordmann P. OXA-58, a novel class D β-lactamase involved in resistance to carbapenems in Acinetobacter baumannii. Antimicrob Agents Chemother. 2005;49(1):202-8. Poirel L, Nordmann P. Carbapenem resistance in Acinetobacter baumannii: mechanisms and epidemiology. Clin Microbiol Infect. 2006;12(9):826-36.

REFERÊNCIAS BIBLIOGRÁFICAS
123
Poirel L, Figueiredo S, Cattoir V, Carattoli A, Nordmann P. Acinetobacter radioresistens as a silent source of carbapenem resistance for Acinetobacter spp. Antimicrob Agents Chemother. 2008;52(4):1252-6. Poirel L, Naas T, Nordmann P. Diversity, epidemiology and genetics of class D β-lactamases. Antimicrob Agents and Chemoter. 2010;54(1):24-38. Poole K. Outer membranes and efflux: the path to multidrug resistance in Gram-negative bacteria. Curr Pharm Biotechnol. 2002;3(2):77-98. Poulakou G, Kontopidou FV, Paramythiotou E, Kompoti M, Katsiari M, Mainas E, Nicolaou C, Yphantis D, Antoniadou A, Trikka-Graphakos E, Roussou Z, Clouva P, Maguina N, Kanellakopoulou K, Armaganidis A, Giamarellou H. Tigecycline in the treatment of infections from multi-drug resistant gram-negative pathogens. J Infect. 2009;58(4):273-84. Pournaras S, Markogiannakis A, Ikonomidis A, Kondyli L, Bethimouti K, Maniatis A N, Legakis N J, Tsakris A. Outbreak of multiple clones of imipenem-resistant Acinetobacter baumannii isolates expressing OXA-58 carbapenemase in an intensive care unit. J Antimicrob Chemother. 2006;57(3):557–561. Principe L, D’Arezzo S, Capone A, Petrosillo N, Visca P. In vitro activity of tigecycline in combination with various antimicrobials against multidrug resistant Acinetobacter baumannii. Ann Clin Microbiol Antimicrob. 2009;8:18. Quale J, Bratu S, Landman D, Heddurshetti R. Molecular epidemiology and mechanisms of carbapenem resistance in Acinetobacter baumannii endemic in New York City. Clin Infect Dis. 2003;37(2):214-20. Queenan AM, Bush K. Carbapenemases: the versatile β-lactamases. Clin Microbiol Rev. 2007;20(3):440-58 Reid GE, Grim SA, Aldeza CA, Janda WM, Clark NM. Rapid development of Acinetobacter baumannii resistance to tigecycline. Pharmacotherapy. 2007;27(8):1198-1201. Reina R, Estenssoro E, Sáenz G, Canales HS, Gonzalvo R, Vidal G, Martins G, Das Neves A, Santander O, Ramos C. Safety and efficacy of colistin in Acinetobacter and Pseudomonas infections: a prospective cohort study. Intensive Care Med. 2005;31(8):1058-65. Reinert RR, Low DE, Rossi F, Zhang X, Wattal C, Dowzicky MJ. Antimicrobial susceptibility among organisms from the Asia/Pacific Rim, Europe and Latin and North America collected as part of TEST and the in vitro activity of tigecycline. Journal of Antimicrobial and Chemotherapy. 2007;60(5): 1018-1029. Ribera A, Ruiz J, Vila J. Presence of the Tet M determinant in a clinical isolate of Acinetobacter baumannii. Antimicrob Agents Chemother. 2003;47(7):2310-2. Robledo IE, Aquino EE, Santé MI, Santana JL, Otero DM, León CF, Vázquez GJ. Detection of KPC in Acinetobacter spp. in Puerto Rico. Antimicrob Agents Chemother. 2010;54(3):1354-7. Rodríguez Guardado A, Blanco A, Asensi V, Pérez F, Rial JC, Pintado V, Bustillo E, Lantero M, Tenza E, Alvarez M, Maradona JA, Cartón JA. Multidrug-resistant

REFERÊNCIAS BIBLIOGRÁFICAS
124
Acinetobacter meningitis in neurosurgical patients with intraventricular catheters: assessment of different treatments. J Antimicrob Chemother. 2008;61(4):908-13. Rodríguez-Hernández MJ, Pachón J, Pichardo C, Cuberos L, Ibáñez-Martínez J, García-Curiel A, Caballero FJ, Moreno I, Jiménez-Mejías ME. Imipenem, doxycycline and amikacin in monotherapy and in combination in Acinetobacter baumannii experimental pneumonia. J Antimicrob Chemother. 2000;45(4):493-501. Rodríguez-Martínez JM, Nordmann P, Ronco E, Poirel L. Extended-spectrum cephalosporinase in Acinetobacter baumannii. Antimicrob Agents Chemother. 2010;54(8):3484-8. Rosenfeld N, Bouchier C, Courvalin P, Périchon B. Expression of the resistance-nodulation-cell division pump AdeIJK in Acinetobacter baumannii is regulated by AdeN, a TetR-type regulator. Antimicrob Agents Chemother. 2012;56(5):2504-10. Rossau R, Van Landschoot A, Gillis M, De Ley J. Taxonomy of Moraxellaceae fam. nov., a new bacterial family to accommodate the genera Moraxella, Acinetobacter, and Psychrobacterand related organisms. International Journal of Systematic Bacteriology. 1991;41(2):310-9. Rossi F. The challenges of antimicrobial resistance in Brazil. Clinical Infectious Diseases. 2011;52(9):1138-1143. Ruzin A, Keeney D, Bradford PA. AdeABC multidrug efflux pump is associated with decreased susceptibility to tigecycline in Acinetobacter calcoaceticus-Acinetobacter baumannii complex. J Antimicrob Chemother. 2007;59(5):1001-4. Sader HS, Castanheira M, Mendes RE, Toleman M, Walsh TR, Jones RN. Dissemination and diversity of metallo-beta-lactamases in Latin America: report from the SENTRY Antimicrobial Surveillance Program. International Journal of Antimicrobial Agents. 2005;25(1): 57-61. Sato K, Nakae T. Outer membrane permeability of Acinetobacter calcoaceticus and its implication in antibiotic resistance. J Antimicrob Chemother. 1991;28(1):35-45. Sauvage E, Kerff F, Terrak M, Ayala JA, Charlier P. The penicillin-binding proteins: structure and role in peptidoglycan biosynthesis. FEMS Microbiol Rev. 2008;32(2):234-58. Savov E, Chankova D, Vatcheva R, Dinev N. In vitro investigation of the susceptibility of Acinetobacter baumannii strains isolated from clinical specimens to ampicillin/sulbactam alone and in combination with amikacin. Int J Antimicrob Agents. 2002;20(5):390-2. Scaife W, Young HK, Paton RH, Amyes SG. Transferable imipenem-resistance in Acinetobacter species from a clinical source. J Antimicrob Chemother. 1995;36(3):585-6. Schafer JJ, Goff DA, Stevenson KB, Mangino JE. Early experience with tigecycline for ventilator-associated pneumonia and bacteremia caused by multidrug-resistant Acinetobacter baumannii. Pharmacotherapy. 2007;27(7):980–7. Schafer JJ, Mangino JE. Multidrug-resistant Acinetobacter baumannii osteomyelitis from Iraq. Emerg Infect Dis. 2008;14(3):512-4.

REFERÊNCIAS BIBLIOGRÁFICAS
125
Schimith Bier KE, Luiz SO, Scheffer MC, Gales AC, Paganini MC, Nascimento AJ, Carignano E, Dalla Costa LM. Temporal evolution of carbapenem-resistant Acinetobacter baumannii in Curitiba, southern Brazil. Am J Infect Control. 2010;38(4):308-14. Scott P, Deye G, Srinivasan A, Murray C, Moran K, Hulten E, Fishbain J, Craft D, Riddell S, Lindler L, Mancuso J, Milstrey E, Bautista CT, Patel J, Ewell A, Hamilton T, Gaddy C, Tenney M, Christopher G, Petersen K, Endy T, Petruccelli B. An outbreak of multidrug-resistant Acinetobacter baumannii-calcoaceticus complex infection in the US military health care system associated with military operations in Iraq. Clin Infect Dis. 2007;44(12):1577-84. Sebeny PJ, Riddle MS, Petersen K. Acinetobacter baumannii skin and soft-tissue infection associated with war trauma. Clin Infect Dis. 2008;47(4):444-9. Segal H, Nelson EC, Elisha BG. Genetic environment and transcription of AmpC in an Acinetobacter baumannii clinical isolate. Antimicrob Agents Chemother. 2004;48(2):612-4. Segal H, Garny S, Elisha BG. Is IS(ABA-1) customized for Acinetobacter? FEMS Microbiol Lett. 2005a;243(2):425-9. Segal H, Elisha BG. Use of Etest MBL strips for the detection of carbapenemases in Acinetobacter baumannii. J Antimicrob Chemother. 2005b;56(3):598. Segal H, Jacobson RK, Garny S, Bamford CM, Elisha BG. Extended -10 promoter in ISAba1 upstream of blaOXA-23 from Acinetobacter baumannii. Antimicrob Agents Chemother. 2007;51(8):3040-1. Seifert H, Dijkshoorn L, Gerner-Smidt P, Pelzer N, Tjernberg I, Vaneechoutte M. Distribution of Acinetobacter species on human skin: comparison of phenotypic and genotypic identification methods. J Clin Microbiol. 1997;35(11):2819-25. Seng P, Drancourt M, Gouriet F, La Scola B, Fournier PE, Rolain JM, Raoult D. Ongoing revolution in bacteriology: routine identification of bacteria by matrix-assisted laser desorption ionization time-of-flight mass spectrometry. Clin Infect Dis. 2009;49(4):543-51. Sevillano E, Gallego L, García-Lobo JM. First detection of the OXA-40 carbapenemase in P. aeruginosa isolates, located on a plasmid also found in A. baumannii. Pathologie-Biologie. 2009;57(6):493-495. Shakil S, Khan AU. Detection of CTX-M-15-producing and carbapenem resistant Acinetobacter baumannii strains from urine from an Indian hospital. J Chemother. 2010;22(5):324-7. Shrivastava SM, Shukla SK, Chaudhary M. Comparison of antimicrobial efficacy of a fixed dose combination of ceftazidime+sulbactam with ceftazidime and sulbactam alone against five bacteria. Folia Microbiol (Praha). 2009;54(5):391-4. Siegman-Igra Y, Bar-Yosef S, Gorea A, Avram J. Nosocomial Acinetobacter meningitis secondary to invasive procedures: report of 25 cases and review. Clin Infect Dis. 1993;17(5):843-9.

REFERÊNCIAS BIBLIOGRÁFICAS
126
Siroy A, Molle V, Lemaître-Guillier C, Vallenet D, Pestel-Caron M, Cozzone AJ, Jouenne T, Dé E. Channel formation by CarO, the carbapenem resistance-associated outer membrane protein of Acinetobacter baumannii. Antimicrob Agents Chemother. 2005;49(12):4876-83. Siroy A, Cosette P, Seyer D, Lemaître-Guillier C, Vallenet D, Van Dorsselaer A, Boyer-Mariotte S, Jouenne T, Dé E. Global comparison of the membrane subproteomes between a multidrug-resistant Acinetobacter baumannii strain and a reference strain. J Proteome Res. 2006;5(12):3385-98. Song JY, Kee SY, Hwang IS, Seo YB, Jeong HW, Kim WJ, Cheong HJ. In vitro activities of carbapenem/sulbactam combination, colistin, colistin/rifampicin combination and tigecycline against carbapenem-resistant Acinetobacter baumannii. J Antimicrob Chemother. 2007;60(2):317-22. Song JY, Cheong HJ, Lee J, Sung AK, Kim WJ. Efficacy of monotherapy and combined antibiotic therapy for carbapenem-resistant Acinetobacter baumannii pneumonia in an immunosuppressed mouse model. Int J Antimicrob Agents. 2009;33(1):33-9. Souli M, Kontopidou FV, Koratzanis E, Antoniadou A, Giannitsioti E, Evangelopoulou P, Kannavaki S, Giamarellou H. In vitro activity of tigecycline against multiple-drug-resistant, including pan-resistant, gram-negative and gram-positive clinical isolates from Greek hospitals. Antimicrobial Agents and Chemotherapy. 2006;50(9): 3166-3169. Spratt BG, Zhou J, Taylor M, Merrick MJ. Monofunctional biosynthetic peptidoglycan transglycosylases. Mol Microbiol. 1996;19(3):639-40. Srinivasan VB, Rajamohan G, Pancholi P, Stevenson K, Tadesse D, Patchanee P, Marcon M, Gebreyes WA. Genetic relatedness and molecular characterization of multidrug resistant Acinetobacter baumannii isolated in central Ohio, USA. Ann Clin Microbiol Antimicrob. 2009;17(8):21. Stietz MS, Ramírez MS, Vilacoba E, Merkier AK, Limansky AS, Centrón D, Catalano M. Acinetobacter baumannii extensively drug resistant lineages in Buenos Aires hospitals differ from the international clones I-III. Infect Genet Evol. 2013;14:294-301. Stoeva T, Higgins PG, Bojkova K, Seifert H. Clonal spread of carbapenem-resistant OXA-23-positive Acinetobacter baumannii in a Bulgarian university hospital. Clin Microbiol Infect. 2008;14(7):723-7. Su XZ, Chen J, Mizushima T, Kuroda T, Tsuchiya T. AbeM, an H+ coupled Acinetobacter baumannii multidrug efflux pump belonging to the MATE family of transporters. Antimicrob Agents Chemother. 2005;49(10):4362-4 Sugawara E, Nikaido H. OmpA is the principal nonspecific slow porin of Acinetobacter baumannii. J Bacteriol. 2012;194(15):4089-96. Sykes RB. The classification and terminology of enzymes that hydrolyze beta-lactam antibiotics. J Infect Dis. 1982;145(5):762-5. Tan TY, Ng LS, Tan E, Huang G. In vitro effect of minocycline and colistin combinations on imipenem-resistant Acinetobacter baumannii clinical isolates. Journal of Antimicrobial Chemotherapy. 2007;60(2): 421-423.

REFERÊNCIAS BIBLIOGRÁFICAS
127
Taziarova M, Holeckova K, Lesnakova A, Sladeckova V, Bartkovjak M, Seckova S, Bukovinova P, Hvizdak F, Svabova V, Findova L, Kisac P, Beno P, Bauer F, Bauer M, Karvaj M, Rudinsky B, Sabo I, Bielova M, Luzica R, Wiczmandyova O, Huttova M, Ondrusova A. Gram-negative bacillary community acquired meningitis is not a rare entity in last two decades. Neuro Endocrinol Lett. 2007;2(3):18-9. Tenover FC, Arbeit RD, Goering RV, Mickelsen PA, Murray BE, Persing DH, Swaminathan B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol. 1995;33(9):2233-9. Tian GB, Adams-Haduch JM, Taracila M, Bonomo RA, Wang HN, Doi Y. Extended-spectrum AmpC cephalosporinase in Acinetobacter baumannii: ADC-56 confers resistance to cefepime. Antimicrob Agents Chemother. 2011;55(10):4922-5. Tiwari V, Vashistt J, Kapil A, Moganty RR. Comparative proteomics of inner membrane fraction from carbapenem-resistant Acinetobacter baumannii with a reference strain. PLoS One. 2012;7(6):e39451. Tognim MC, Gales AC, Penteado AP, Silbert S, Sader HS. Dissemination of IMP-1
metallo--lactamase-producing Acinetobacter species in a Brazilian teaching hospital. Infect Control Hosp Epidemiol. 2006;27(7):742-7. Tomás MM, Beceiro A, Pérez A, Velasco D, Mpoure R, Villanueva R, Martínez-Beltrán J, Bou G. Cloning and functional analysis of the gene enconding the 33 to 36 kilodalton outer membrane protein associated with carbapenem resistance in Acinetobacter baumannii. Antimicrob Agents Chemother. 2005;49(12):5172-5. Tong W, Wang R, Chai D, Li Z, Pei F. In vitro activity of cefepime combined with sulbactam against clinical isolates of carbapenem-resistant Acinetobacter spp. Int J Antimicrob Agents. 2006;28(5):454-6. Towner KJ, Levi K, Vlassiadi M; ARPAC Steering Group. Genetic diversity of carbapenem-resistant isolates of Acinetobacter baumannii in Europe. Clin Microbiol Infect. 2008;14(2):161-7. Towner KJ. Acinetobacter: an old friend, but a new enemy. J Hosp Infect. 2009;73(4):355-63. Tuon FF, Penteado-Filho SR, Amarante D, Andrade MA, Borba LA. Mortality rate in patients with nosocomial Acinetobacter meningitis from a Brazilian hospital. Braz J Infect Dis. 2010;14(5):437-40. Turner PJ. Extended-spectrum β-lactamases. Clin Infec Dis. 2005;41(4):273-5. Turton JF, Kaufmann ME, Glover J, Coelho JM, Warner M, Pike R, Pitt TL. Detection and typing of integrons in epidemic strains of Acinetobacter baumannii found in the United Kingdom. J Clin Microbiol. 2005;43(7):3074-82. Turton JF, Ward ME, Woodford N, Kaufmann ME, Pike R, Livermore DM, Pitt TL. The role of ISAba1 in expression of OXA carbapenemase genes in Acinetobacter baumannii. FEMS Microbiol Lett. 2006;258(1):72-7.

REFERÊNCIAS BIBLIOGRÁFICAS
128
Turton JF, Gabriel SN, Valderrey C, Kaufmann ME, Pitt TL. Use of sequence-based typing and multiplex PCR to identify clonal lineages of outbreak strains of Acinetobacter baumannii. Clin Microbiol Infect. 2007;13(8):807-15. Vaneechoutte M, Dijkshoorn L, Tjernberg I, Elaichouni A, de Vos P, Claeys G, Verschraegen G. Identification of Acinetobacter genomic species by amplified ribosomal DNA restriction analysis. J Clin Microbiol. 1995; 33(1):11-5. Vaneechoutte M, Dijkshoorn L, Nemec A, Kämpfer P, Wauters G. Acinetobacter, Cryseobacterium, Moraxella and other nonfermentative Gram-negative rods. In: Manual of Clinical Microbiology By Versalovic J, Carroll KC, Funke G, Jorgensen JH, Landry ML, Warnock DW. 10 ed. American Society for Microbiology - ASM, Washington, DC, 2011;1:714-738. Vahaboglu H, Oztürk R, Aygün G, Coşkunkan F, Yaman A, Kaygusuz A, Leblebicioglu H, Balik I, Aydin K, Otkun M. Widespread detection of PER-1-type extended-spectrum beta-lactamases among nosocomial Acinetobacter and Pseudomonas aeruginosa isolates in Turkey: a nationwide multicenter study. Antimicrob Agents Chemother. 1997;41(10):2265-9. Vashist J, Tiwari V, Kapil A, Rajeswari MR. Quantitative profiling and identification of outer membrane proteins of β-lactam resistant strain of Acinetobacter baumannii. J Proteome Res. 2010;9(2):1121-8. Vila J, Martí S, Sánchez-Céspedes J. Porins, efflux pumps and multidrug resistance in Acinetobacter baumannii. J Antimicrob Chemother. 2007;59(6):1210-5. Villegas MV, Hartstein AI. Acinetobacter outbreaks, 1977-2000. Infect Control Hosp Epidemiol. 2003;24(4):284-95. Villegas MV, Kattan JN, Correa A, Lolans K, Guzman AM, Woodford N, Livermore D, Quinn JP. Dissemination of Acinetobacter baumannii clones with OXA-23 Carbapenemase in Colombian hospitals. Antimicrob Agents Chemother. 2007;51(6):2001-4. Walsh TR, Toleman MA, Poirel L, Nordmann P. Metallo-β-lactamases: the quiet before the storm? Clin Microbiol Rev. 2005;18(2):306-25. Walther-Rasmussen J, Høiby N. OXA-type carbapenemases. J Antimicrob Chemother. 2006;57(3):373-83. Wang H, Liu YM, Chen MJ, Sun HL, Xie XL, Xu YC. Mechanism of carbapenems resistance in Acinetobacter baumannii. Zhongguo Yi Xue Ke Xue Yuan Xue Bao. 2003; 25(5):567-72. Wareham DW, Bean DC. In vitro activity of polymyxin B in combination with imipenem, rifampicin and azithromycin versus multidrug resistant strains of Acinetobacter baumannii producing OXA-23 carbapenemases. Ann Clin Microbiol Antimicrob. 2006;21;5:10. Werneck JS, Picão RC, Carvalhaes CG, Cardoso JP, Gales AC. OXA-72-producing Acinetobacter baumannii in Brazil: a case report. J Antimicrob Chemother. 2011a;66(2):452-4.

REFERÊNCIAS BIBLIOGRÁFICAS
129
Werneck JS, Picão RC, Girardello R, Cayô R, Marguti V, Dalla-Costa L, Gales AC, Antonio CS, Neves PR, Medeiros M, Mamizuka EM, Elmor de Araújo MR, Lincopan N. Low prevalence of blaOXA-143 in private hospitals in Brazil. Antimicrob Agents Chemother. 2011b;55(9):4494-5; author reply 4495. Wieczorek P, Sacha P, Ojdana D, Milewski R, Jurczak A, Kaczyńska K,Tryniszewska E. In vitro resistance development in Acinetobacter baumannii to sulbactam and cefoperazone. Med Dosw Mikrobiol. 2012;64(1):55-62.
Wilke MS, Lovering AL, Strynadka NC. -lactam antibiotic resistance: a current structural perspective. Curr Opin Microbiol. 2005;8(5):525-33. Wilks M, Wilson A, Warwick S, Price E, Kennedy D, Ely A, Millar MR. Control of an outbreak of multidrug-resistant Acinetobacter baumannii-calcoaceticus colonization and infection in an intensive care unit (ICU) without closing the ICU or placing patients in isolation. Infect Control Hosp Epidemiol. 2006;27(7):654-8. Winokur PL, Canton R, Casellas JM, Legakis N. Variations in the prevalence of strains expressing an extended-spectrum β-lactamase phenotype and characterization of isolates from Europe, the Americas, and the Western Pacific region. Clin Infect Dis. 2001;32(2):94-103. Wolinsky E, Hines JD. Neurotoxic and nephrotoxic effects of colistina patients with renal disease. N Engl J Med. 1962;266:759-62. Wong EW, Yusof MY, Mansor MB, Anbazhagan D, Ong SY, Sekaran SD. Disruption of adeB gene has a greater effect on resistance to meropenems than adeA gene in Acinetobacter spp. isolated from University Malaya Medical Centre. Singapore Medical Journal. 2009;50(8):822-826. Wood GC, Hanes SD, Croce MA, Fabian TC, Boucher BA. Comparison of ampicillin-sulbactam and imipenem-cilastatin for the treatment of Acinetobacter ventilator-associated pneumonia. Clin Infect Dis. 2002;34(11):1425-30. Wood GC, Hanes SD, Boucher BA, Croce MA, Fabian TC. Tetracyclines for treating multidrug-resistant Acinetobacter baumannii ventilator-associated pneumonia. Intensive Care Med. 2003;29(11):2072-6. Woodford N, Ellington MJ, Coelho JM, Turton JF, Ward ME, Brown S, Amyes SG, Livermore DM. Multiplex PCR for genes encoding prevalent OXA carbapenemases in Acinetobacter spp. Int J Antimicrob Agents. 2006;27(4):351-3. Yang SC, Chang WJ, Chang YH, Tsai YS, Yang TP, Juan CW, Shiau MY. Prevalence of antibiotics resistance and OXA carbapenemases genes in multidrug-resistant Acinetobacter baumannii isolates in central Taiwan. Eur J ClinMicrobiol Infect Dis. 2010;29(5):601-4. Yoon J, Urban C, Terzian C, Mariano N, Rahal JJ. In vitro double and triple synergistic activities of Polymyxin B, imipenem, and rifampin against multidrug-resistant Acinetobacter baumannii. Antimicrobial Agents and Chemotherapy. 2004;48(3):753-757. Yoon EJ, Courvalin P, Grillot-Courvalin C. RND-Type Efflux Pumps in Multidrug-Resistant Clinical Isolates of Acinetobacter baumannii: Major Role for AdeABC

REFERÊNCIAS BIBLIOGRÁFICAS
130
Overexpression and AdeRS Mutations. Antimicrob Agents Chemother. 2013;57(7):2989-95. Yun SH, Choi CW, Kwon SO, Park GW, Cho K, Kwon KH, Kim JY, Yoo JS, Lee JC, Choi JS, Kim S, and Kim SI. Quantitative proteomic analysis of cell wall and plasma membrane fractions from Multidrug-Resistant Acinetobacter baumannii. Journal of Proteome Research. 2011;10(2):459-469. Zakataeva NP, Kutukova EA, Gronskiĭ SV, Troshin PV, Livshits VA, Aleshin VV. Export of metabolites by the proteins of the DMT and RhtB families and its possible role in intercellular communication]. Mikrobiologiia. 2006;75(4):509-20. Zander E, Nemec A, Seifert H, Higgins PG. Association between β-lactamase-encoding bla(OXA-51) variants and DiversiLab rep-PCR-based typing of Acinetobacter baumannii isolates. J Clin Microbiol. 2012;50(6):1900-4. Zapun A, Contreras-Martel C, Vernet T. Penicillin-binding proteins and β-lactam resistance. FEMS Microbiology Reviews. 2008;32(2):361-385. Zarrilli R, Pournaras S, Giannouli M, Tsakris A. Global evolution of multidrug-resistant Acinetobacter baumannii clonal lineages. Int J Antimicrob Agents. 2013;41(1):11-9. Zavascki AP, Carvalhaes CG, Picão RC, Gales AC. Multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii: resistance mechanisms and implications for therapy. Expert Review of Anti-infective Therapy. 2010;8(1):71-93. Zhanel GG, Wiebe R, Dilay L, Thomson K, Rubinstein E, Hoban DJ, Noreddin AM, Karlowsky JA. Comparative review of the carbapenems. Drugs. 2007;67(7):1027-52. Zhou H, Pi BR, Yang Q, Yu YS, Chen YG, Li LJ, Zheng SS. Dissemination of imipenem-resistant Acinetobacter baumannii strains carrying the ISAba1 blaOXA-23
genes in a Chinese hospital. J Med Microbiol. 2007;56(8):1076-80. Zhou Y, Yu H, Guo Q, Xu X, Ye X, Wu S, Guo Y, Wang M. Distribution of 16S rRNA methylases among different species of Gram-negative bacilli with high-level resistance to aminoglycosides. Eur J Clin Microbiol Infect Dis. 2010;29(11):1349-53.





























