Embed Size (px)
Citation preview

CELLULAR NEUROSCIENCEREVIEW ARTICLE
published: 30 January 2015doi: 10.3389/fncel.2015.00004
Control of cortical neuronal migration byglutamate and GABAHeiko J. Luhmann1*, A. Fukuda2 and W. Kilb1
1 Institute of Physiology, University Medical Center of the Johannes Gutenberg University, Mainz, Germany2 Department of Neurophysiology, Hamamatsu University School of Medicine, Hamamatsu, Shizuoka, Japan
Edited by:Takeshi Kawauchi, Keio UniversitySchool of Medicine/PRESTO, Japan
Reviewed by:Rustem Khazipov, Institut Nationalde la Santé et de la RechercheMédicale, FranceLaura Cancedda, Istituto Italiano diTecnologia, Italy
*Correspondence:Heiko J. Luhmann, Institute ofPhysiology, University MedicalCenter of the Johannes GutenbergUniversity, Duesbergweg 6,D-55128 Mainz, Germanye-mail: [email protected]
Neuronal migration in the cortex is controlled by the paracrine action of the classicalneurotransmitters glutamate and GABA. Glutamate controls radial migration of pyramidalneurons by acting primarily on NMDA receptors and regulates tangential migration ofinhibitory interneurons by activating non-NMDA and NMDA receptors. GABA, acting onionotropic GABAA-rho and GABAA receptors, has a dichotomic action on radially migratingneurons by acting as a GO signal in lower layers and as a STOP signal in upper cortical plate(CP), respectively. Metabotropic GABAB receptors promote radial migration into the CP andtangential migration of interneurons. Besides GABA, the endogenous GABAergic agonisttaurine is a relevant agonist controlling radial migration. To a smaller extent glycine receptoractivation can also influence radial and tangential migration. Activation of glutamate andGABA receptors causes increases in intracellular Ca2+ transients, which promote neuronalmigration by acting on the cytoskeleton. Pharmacological or genetic manipulation ofglutamate or GABA receptors during early corticogenesis induce heterotopic cell clustersin upper layers and loss of cortical lamination, i.e., neuronal migration disorders which canbe associated with neurological or neuropsychiatric diseases. The pivotal role of NMDA andionotropic GABA receptors in cortical neuronal migration is of major clinical relevance, sincea number of drugs acting on these receptors (e.g., anti-epileptics, anesthetics, alcohol) maydisturb the normal migration pattern when present during early corticogenesis.
Keywords: neuronal migration, cerebral cortex, GABA, glutamate, neuronal migration disorders
INTRODUCTIONDuring early brain development, mostly during embryonic phasesand in some species also during early postnatal periods, newlygenerated neurons must migrate from their site of origin totheir final target in a distinct brain area and a certain subregion,e.g., a specific layer at a certain site of the cerebral cortex.The distances that neurons must travel depend on the cell typeand on the species and range from a few hundred micrometersto many millimeters. One exciting question in neuroscienceis how newly generated neurons find their correct way totheir final position. Over the last two decades we learned thatthe neuronal migration process is controlled by a number ofdifferent mechanisms. Transcription factors control the identityand the laminar position of developing neurons (for review, Kwanet al., 2012). Chemical cues, e.g., semaphorines and ephrins,are expressed as gradients in the brain and serve as attractingor repelling signals for migrating cells (Bagnard et al., 2001;Holmberg et al., 2006; Zimmer et al., 2008; Sentürk et al.,2011). In some brain regions the vertical fibers of radial glialcells act as chemico-mechanical guiding structures for migratingneurons (for review, Huang, 2009). Receptors activated byneurotransmitters or certain molecules, e.g., the extracellularmatrix protein reelin, act as GO or STOP signals in neuronalmigration (Huang, 2009).
This review will focus on the role of the two classicalneurotransmitter systems glutamate and GABA in neuronalmigration of cortical neurons. After briefly describing thedifferent modes of neuronal migration and differences inthe migration process between glutamatergic and GABAergicneurons, our current knowledge on the function of glutamateand GABA receptors in neuronal migration will be reviewed.Finally, we will also shortly address the putative role of glycinereceptors in neuronal migration. Since the receptors of bothtransmitters are the target of numerous drugs acting for exampleas anesthetics or anti-epileptics, pathophysiological perturbationsof the migration process by unwanted side effects of these drugsacting on glutamate and GABA receptors during early braindevelopment will be also discussed.
For comprehensive overviews on the molecular and cellularmechanisms of neuronal migration the interested reader isreferred to reviews by Ayala et al. (2007), Valiente and Marín(2010) and the review by David. J. Price on “Neuronal migrationin the cerebral cortex” in this issue. A summary on neocorticallayer formation, the timing of projection and interneuronmigration and a comparison between rats and mice is given in thereviews by Kwan et al. (2012) and Tanaka and Nakajima (2012).An update on the early development of the human cerebral cortexis given by Bystron et al. (2008).
Frontiers in Cellular Neuroscience www.frontiersin.org January 2015 | Volume 9 | Article 4 | 1

Luhmann et al. GABA and glutamate in neuronal migration
MODES OF NEURONAL MIGRATIONDifferent modes of neuronal migration have been described(Figure 1). In 1972, Pasko Rakic published a seminal paper on the“Mode of cell migration to the superficial layers of fetal monkeyneocortex” and described the radial migration of immatureneocortical neurons along the vertical fibers of radial glial cells(Rakic, 1972). Rakic postulated the existence of a “strong surfaceaffinity” between the radial glial fibers and the migrating neuronand suggested that this “developmental mechanism . . . wouldallow for the vertical cell columns of adult neocortex” (Rakic,1972). The radial unit hypothesis of the cerebral cortex was born.The radial glia-dependent locomotion is the dominant migrationmode of newborn pyramidal, glutamatergic neurons in thehippocampus and cerebral cortex and also represents the centralmechanism for the “inside first—outside last” developmentalpattern of the cerebral cortex (neurons marked in red inFigure 1B; Nadarajah et al., 2003). Like building a house, theoldest neurons form the lowest layer 6 and subsequently generatedneurons form layers 5, 4, 3 and finally layer 2. This inside-outlayering also means that radially migrating neurons must passbeyond their predecessors before reaching their final positionin the newly generated cortical layer, which they form (forreview, Cooper, 2008). Recently Le Magueresse et al. (2012)described with time-lapse 2-photon microscopy in acute brainslice preparations of the neonatal mouse a new type of radialmigration of subventricular zone (SVZ)-generated neurons alongastrocytes lining blood vessels, which does not depend on radialglial cells.
A different mode of neuronal migration, which is independentof glial guiding fibers, is the somal translocation (Nadarajahet al., 2003; for review, Cooper, 2008). Somal translocation issmoother and faster than glia-guided radial migration. Here aleading coiled process extends into the marginal zone (MZ) and
is anchored to the basement membrane or to the extracellularmatrix. The soma moves upward in a spring-like manner byrapidly shortening the leading process. It seems likely that glia-independent somal translocation and glia-dependent locomotiondepend on different cytoskeletal machinery and motors andthereby are regulated by different processes.
In contrast to the radial migration of pyramidal cells,neocortical GABAergic interneurons show a tangentialmigration pattern throughout the developing telencephalon(de Carlos et al., 1996; for review, Marín, 2013). Inhibitoryinterneurons migrate tangentially over long distances bygenerating a leading process, which detects chemical cues in theextracellular environment, and subsequent movement of thenucleus towards to the branching point (nucleokinesis). Recentobservations in slice cultures of the mouse embryonic brainindicate that endothelial cells may guide tangential migration(Won et al., 2013) and that tangential migration in the MZ iscontrolled by meningeal vessels (Borrell and Marín, 2006). Themolecular mechanism of this blood vessel-guided migration tothe cortex are not known, but neurotrophic factors such as brain-derived neurotrophic factor (BDNF) and glial cell line-derivedneurotrophic factor (GDNF) may be involved (Le Magueresseet al., 2012). Meninges affect tangential migration in the MZvia secretion of the chemokine CXCL12 which activates CXCR4receptors (Borrell and Marín, 2006). This type of migration maybecome reactivated in the adult brain under pathophysiologicalconditions, e.g., stroke, when SVZ-generated neuroblasts areguided to the peri-infarct zone by blood vessels (Kojima et al.,2010).
Finally, so-called random walk migration has been describedfor medial ganglionic eminence (MGE)-derived corticalinterneurons in the MZ of flat-mount cortices (Tanaka et al.,2009). Interneurons migrated tangentially over periods of up to
FIGURE 1 | Origin and migratory pathways of glutamatergic andGABAergic neurons. (A) Schematic diagram illustrating migration pathway ofthe majority of glutamatergic neurons, originating in the ventricular zone (VZ)of the pallium and radially migrating into the developing cerebral cortex (redarrows). The majority of GABAergic neurons are generated in the medial(MGE) and lateral gangionic eminence (LGE) and reach their final position bytangential migration via deep pathways and superficial cortical layers. (B)Glutamatergic neurons (marked in different shades of red) are generated in
the VZ and migrate radially either by somal translocation or, at later phases, bylocomotion along radial glial cells (light gray). Upon reaching the marginal zone(MZ) they detach and align on top of previously generated neurons of thecortical plate (CP), generating the “inside first—outside last” pattern of thecerebral cortex. The majority of GABAergic neurons (marked in differentshades of blue) reach the cortex via tangential migration in the deep pathwaywithin the subventricular zone (SVZ) or the superficial pathway in the MZ.Some GABAergic interneurons travel also within the subplate (SP).
Frontiers in Cellular Neuroscience www.frontiersin.org January 2015 | Volume 9 | Article 4 | 2

Luhmann et al. GABA and glutamate in neuronal migration
2 days in an unpredictable manner, often changing the rate anddirection of migration. These results suggest that MGE-derivedcortical interneurons, once arriving at the MZ, are released fromregulation by guidance cues and initiate random walk movement(Tanaka et al., 2009).
In summary, radial migration, somal translocation andtangential migration are the dominant forms of neuronalmigration in the developing cerebral cortex. It is not surprisingthat mutations affecting genes, which control these forms ofmigration may cause severe brain malformations, which aregenerally categorized as neuronal migration disorders and whichare often associated with a spectrum of neurological and/orneuropsychiatric diseases (for review, Guerrini et al., 2008;Guerrini and Parrini, 2010).
MIGRATION OF GLUTAMATERGIC NEURONSNeocortical glutamatergic neurons mostly follow a pure radialmigration pattern and for them radial glial cells in the ventricularzone (VZ) fulfill two important and different functions inthe embryonic cortex (Figure 1). On the one hand radialglial cells serve as progenitors and produce by asymmetriccell division neurons and astrocytes, on the other hand radialglial cells serve as migratory guides for the newly generatedglutamatergic neurons. Radial glial cells produces neocorticalpyramidal and layer 4 spiny stellate cells, which migrate tothe cortical plate (CP), thereby forming in the “inside first—outside last” pattern the usually six-layered cerebral cortex. Sisterglutamatergic neurons, which derive from the same mothercell, take up radially aligned positions in the cerebral cortexacross layers and have a higher propensity to form unidirectionalchemical synaptic connections with each other rather thanwith neighboring non-siblings (Yu et al., 2009). These dataindicate that the columnar organization of the cerebral cortexmay be determined to some extent by lineage (Noctor et al.,2001).
During early embryonic development, these glutamatergicneurons initially use somal translocation to migrate radiallyand then follow the vertical track along radial glial fibersfor locomotion (Rakic, 1972). The extracellular matrix proteinreelin, which is secreted from early born Cajal-Retzius neuronslocated in the MZ (for review, Kirischuk et al., 2014), controlsthis radial migration (for review, Valiente and Marín, 2010).Radial migration of single glutamatergic neurons does notoccur continuously following a straightforward route, but rathershows phases of transient migratory arrest and even retrogrademigration (Noctor et al., 2004). Gap junctions play importantroles in the regulation of both proliferation and neuronalmigration. Hemichannels formed by gap junctions mediatethe spread of spontaneous intracellular Ca2+ waves acrossprogenitor cells and provide dynamic adhesive contacts betweenmigrating neurons and radial glial fibers (for review, Eliasand Kriegstein, 2008). For glia-guided neuronal migration theconnexins Cx26 and Cx43 are essential and in the mousetheir deletion disrupts migration to the CP (Elias et al.,2007). For Cx43 it has been demonstrated that deletion of theC-terminal domain modifies neuronal migration (Cina et al.,2009).
Recent immunohistochemical data obtained in embryonicmice demonstrated one population of transient glutamatergicneurons, which is generated early (at embryonic day (E) 12.5) andmigrates tangentially over long distances from their generationsite at the pallial-subpallial boundary to the CP (Teissier et al.,2010). At birth, these early glutamatergic neurons homogeneouslypopulate all neocortical areas, but subsequently die massively byapoptosis. At birth, about 50% of the dying neocortical neuronsbelong to this population of tangential migrating glutamatergicneurons (Teissier et al., 2010).
In summary, glutamatergic neurons use mostly radialmigration along radial glial fibers and somal translocation tomove from their site of generation in the VZ into the developingcerebral cortex.
MIGRATION OF GABAergic NEURONSIn contrast to the large majority of the glutamatergic neurons,cortical GABAergic interneurons are at least in rodents generatedin the subcortical telencephalon; in the lateral, medial, caudaland septal ganglionic eminence (LGE, MGE, CGE, and SGE,respectively; Figure 1A), to a minor extent also in theendopeduncular and preoptic area and also in the corticalSVZ (for review, Gelman and Marin, 2010) see also reviewby Wieland B. Huttner on “Neurogenesis in the developingcerebral cortex” in this issue. A subset of GABAergic neurons,which are 5-HT3 positive, are generated postnatally in theSVZ and migrate into numerous forebrain regions, includingthe cerebral cortex, striatum, and nucleus accumbens (Intaet al., 2008). The origin of GABAergic neocortical interneuronsin higher mammals, including humans, remains controversial,although a recent publication indicate that also in thesespecies a substantial proportion of interneurons originate fromsubcortical telencephalic eminences (Letinic et al., 2002; Ma et al.,2013).
The spatio-temporal expression of various transcriptionfactors control the generation and identity of different typesof cortical GABAergic interneurons at different developmentalperiods (for review, Butt et al., 2007; Jovanovic and Thomson,2011). Dlx1/2 and Mash1 are extensively expressed in theganglionic eminence and determine the GABAergic lineage.Lhx6, which is under the control of Nkx2.1 and Dlx5/6,control the generation of parvalbumin- and somatostatin-immunoreactive interneurons, which are generated first inthe ventral and dorsal area of the MGE, respectively (Wanget al., 2010). The later generation of vasoactive intestinalpolypeptide (VIP) and cholecystokinine (CCK) expressingGABAergic interneurons in the CGE is controlled by thetranscription factors Nkx6.2 and CoupTF1/2. The spatio-temporaldevelopmental profile of cortical GABAergic interneuronspredicts their intrinsic electrophysiological properties andfiring patterns in the mature cortex (Butt et al., 2005).Rapidly adapting firing properties can be observed in matureneuropeptide Y (NPY), reelin, calretinin and/or vasointestinalpeptide expressing cortical interneurons, which are generatedin the CGE. Rapidly adapting NPY-containing interneuronsare also produced in the preoptic area (for review, Marín,2013).
Frontiers in Cellular Neuroscience www.frontiersin.org January 2015 | Volume 9 | Article 4 | 3

Luhmann et al. GABA and glutamate in neuronal migration
From their birth place in the ganglionic eminence forebrainGABAergic interneurons migrate tangentially in the MZ, SVZor intermediate zone (IZ) to the developing cerebral cortex(for review, Marín, 2013). Tangential migration is controlledby the spatio-temporal expression of a number of chemicalcues, acting as attracting or repelling signals. Semaphorines,expressed in the LGE, prevent the entry of migrating interneuronsinto this region and Ephrin EphA5/EphA4 receptors, expressedin the VZ, repel MGE-generated interneurons (for review,Marín, 2013). Tangential migration of cortical GABAergicinterneurons is enhanced by the neurotrophic factors BDNF,NT-4, hepatocyte growth factor, and GDNF. On their wayto the cortex, interneurons use specific routes or migratorystreams (marked in blue in Figure 1B): (i) a superficialroute in the MZ; (ii) a deep route in the IZ/SVZ; and(iii) a route in the subplate (SP). Using an in situ migrationassay, Tanaka et al. (2003) observed that neocortical GABAergicinterneurons initially migrate predominantly in the IZ/SVZand then invade the CP and MZ by departing from themajor migratory stream in the IZ/SVZ. Once arriving in theMZ GABAergic interneurons show random walk migrationand disperse throughout the cortex (Tanaka et al., 2009). Asubpopulation of GABAergic interneurons descend from the MZto be distributed in the CP.
During their tangential migration process, neocorticalGABAergic interneurons progressively acquire responsivenessto GABA. Combining in vitro patch-clamp recordings,neuropharmacological experiments and single-cell PCR inE14.5 mouse acute slices, Carlson and Yeh (2011) characterizedthe functional expression of GABAA receptor subunits intangentially migrating interneurons derived from the MGE.At this age, synapses have not yet formed and responsivenessto GABA reflect the functional expression of synaptic andextrasynaptic GABAA receptors. Early migrating interneuronslocated close to the corticostriate juncture showed a robustexpression of the alpha2 and alpha3 subunits. When enteringthe developing cortex, both subunits were still highly expressedand in addition alpha1 and gamma1-3 subunits were upregulated(Carlson and Yeh, 2011). The functional implications of thesimultaneous activation of multiple GABAA receptor isoformsand the upregulation of receptor isoforms with higher affinity toGABA in the migration process are not known and need to beelucidated.
Some experimental data indicate that migrating interneuronson their way to the cortex may move from one substrate toanother, e.g., following specific axonal projections. Once theyhave reached their final cortical region, cortical GABAergicinterneurons migrate radially to their final layer, which hasbeen already formed by the radial migration of glutamatergicneurons. Thus, GABAergic interneurons invade their targetlayers after glutamatergic projection neurons have reached theirfinal position. The mechanisms underlying this switch fromtangential to radial migration are not completely understood. Itmay be that an intrinsic developmental program or connexinstrigger the tangential-to-radial switch (for review, Marín, 2013).Elias et al. (2010) have demonstrated in embryonic rat brainslices including the MGE that this switch is controlled by
Cx43 and depends on the adhesive properties and the Cterminus of Cx43, but not on the Cx43 channel. These dataindicate that the switch from tangential to radial migrationdepends on a gap junction-mediated interaction betweenmigrating GABAergic interneurons and radial glia cells, similarlyto the glia-dependent migration of glutamatergic neurons.In contrast, whereas reelin signaling is essential for properradial migration of pyramidal neurons, layer acquisition ofneocortical GABAergic interneurons does not depend on reelin,but rather on cues provided by projection neurons (Pla et al.,2006).
In summary, GABAergic interneurons migrate tangentiallyalong specific streams from their site of origin in the subcorticaltelencephalon to their final neocortical site, where they thenmigrate radially to their final cortical layer.
ROLE OF GLUTAMATE IN NEURONAL MIGRATIONThe classical excitatory transmitter glutamate influences neuronalmigration mainly by acting on two ionotropic receptors: (i) theNMDA receptor, a Ca2+-permeable subclass of glutamatereceptor; (ii) the AMPA/kainate receptor, a usually Ca2+-impermeable glutamate receptor. Three (GluR1-3) of thefour known subunits for AMPA receptors are expressed atprenatal stages in the developing cortex, while the GluR4subunit appears only postnatally (Luján et al., 2005). Of thefour subunits assembling kainate receptors, KA-2 and GluR5and GluR6 are already expressed in the embryonic neocortexaround E14 (Bahn et al., 1994). Functional NMDA receptorsare composed from two NR1 and two NR2 subunits. NR1and the highly Ca2+ permeable NR2B subunits are alreadyexpressed at early postnatal stages, while expression of NR2Aemerges at postnatal stages in the neocortex (Luján et al.,2005). Functional NMDA receptors have been found onmigrating glutamatergic and GABAergic interneurons (Beharet al., 1999; Soria and Valdeolmillos, 2002). Metabotropicglutamate receptors, in particular mGlu1 and mGlu5, arealso already expressed in the immature neocortex (López-Bendito et al., 2002a). A direct modulation of neuronalmigration by NMDA receptors has been initially describedby Komuro and Rakic for granule cells of the developing mousecerebellum in vitro. Here, blockade of NMDA receptors byspecific antagonists caused a slow-down of neuronal migration,whereas enhanced activation of NMDA receptors by removal ofmagnesium from the extracellular milieu or by applicationof the cotransmitter glycine accelerated cell movement(Komuro and Rakic, 1993).
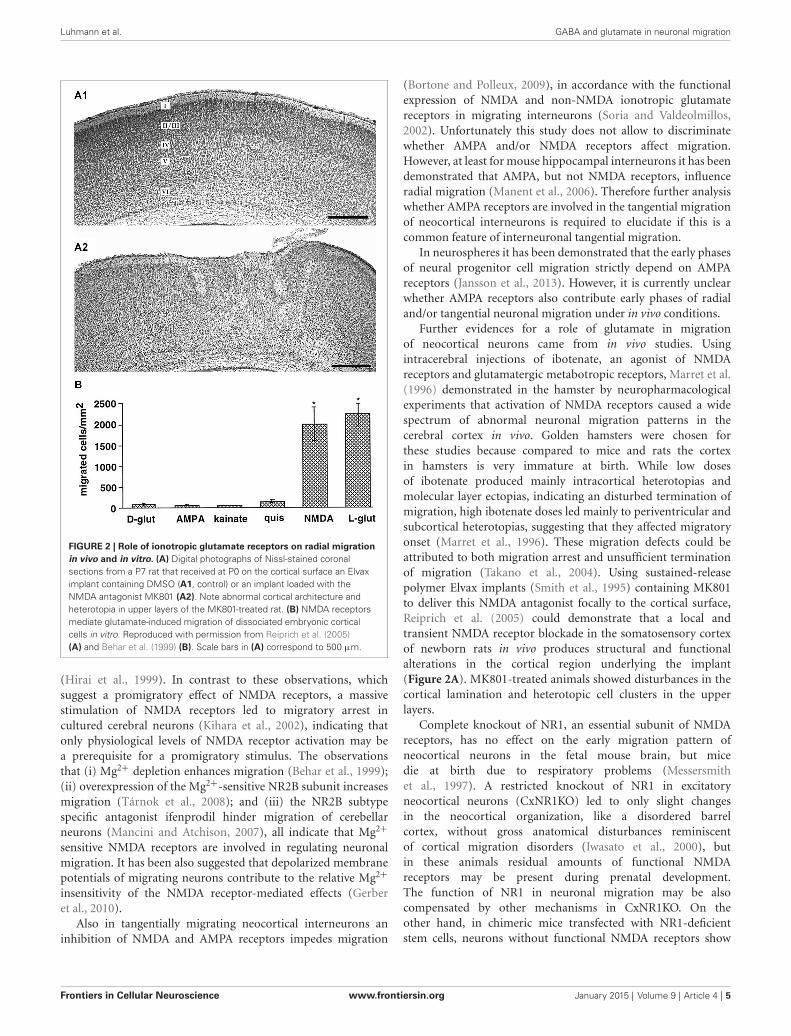
Various in vitro studies using different models of corticalneuronal migration indicate that NMDA receptors also controlradial neuronal migration in the cerebral cortex. In cell dissociatesof murine embryonic cortical cells and cortical slice cultures,Behar et al. (1999) demonstrated that glutamate is a potentchemoattractant. Only activation of NMDA receptors, but notother ionotropic glutamate receptors, stimulated radial migrationof immature neurons out of the cortical VZ/SVZ and applicationof NMDA antagonists blocked migration (Figure 2B; Behar et al.,1999). Inhibition of NMDA receptors using either MK801 orAPV, attenuated radial migration in rat tissue explants in vitro
Frontiers in Cellular Neuroscience www.frontiersin.org January 2015 | Volume 9 | Article 4 | 4
Luhmann et al. GABA and glutamate in neuronal migration
FIGURE 2 | Role of ionotropic glutamate receptors on radial migrationin vivo and in vitro. (A) Digital photographs of Nissl-stained coronalsections from a P7 rat that received at P0 on the cortical surface an Elvaximplant containing DMSO (A1, control) or an implant loaded with theNMDA antagonist MK801 (A2). Note abnormal cortical architecture andheterotopia in upper layers of the MK801-treated rat. (B) NMDA receptorsmediate glutamate-induced migration of dissociated embryonic corticalcells in vitro. Reproduced with permission from Reiprich et al. (2005)(A) and Behar et al. (1999) (B). Scale bars in (A) correspond to 500 µm.
(Hirai et al., 1999). In contrast to these observations, whichsuggest a promigratory effect of NMDA receptors, a massivestimulation of NMDA receptors led to migratory arrest incultured cerebral neurons (Kihara et al., 2002), indicating thatonly physiological levels of NMDA receptor activation may bea prerequisite for a promigratory stimulus. The observationsthat (i) Mg2+ depletion enhances migration (Behar et al., 1999);(ii) overexpression of the Mg2+-sensitive NR2B subunit increasesmigration (Tárnok et al., 2008); and (iii) the NR2B subtypespecific antagonist ifenprodil hinder migration of cerebellarneurons (Mancini and Atchison, 2007), all indicate that Mg2+
sensitive NMDA receptors are involved in regulating neuronalmigration. It has been also suggested that depolarized membranepotentials of migrating neurons contribute to the relative Mg2+
insensitivity of the NMDA receptor-mediated effects (Gerberet al., 2010).
Also in tangentially migrating neocortical interneurons aninhibition of NMDA and AMPA receptors impedes migration
(Bortone and Polleux, 2009), in accordance with the functionalexpression of NMDA and non-NMDA ionotropic glutamatereceptors in migrating interneurons (Soria and Valdeolmillos,2002). Unfortunately this study does not allow to discriminatewhether AMPA and/or NMDA receptors affect migration.However, at least for mouse hippocampal interneurons it has beendemonstrated that AMPA, but not NMDA receptors, influenceradial migration (Manent et al., 2006). Therefore further analysiswhether AMPA receptors are involved in the tangential migrationof neocortical interneurons is required to elucidate if this is acommon feature of interneuronal tangential migration.
In neurospheres it has been demonstrated that the early phasesof neural progenitor cell migration strictly depend on AMPAreceptors (Jansson et al., 2013). However, it is currently unclearwhether AMPA receptors also contribute early phases of radialand/or tangential neuronal migration under in vivo conditions.
Further evidences for a role of glutamate in migrationof neocortical neurons came from in vivo studies. Usingintracerebral injections of ibotenate, an agonist of NMDAreceptors and glutamatergic metabotropic receptors, Marret et al.(1996) demonstrated in the hamster by neuropharmacologicalexperiments that activation of NMDA receptors caused a widespectrum of abnormal neuronal migration patterns in thecerebral cortex in vivo. Golden hamsters were chosen forthese studies because compared to mice and rats the cortexin hamsters is very immature at birth. While low dosesof ibotenate produced mainly intracortical heterotopias andmolecular layer ectopias, indicating an disturbed termination ofmigration, high ibotenate doses led mainly to periventricular andsubcortical heterotopias, suggesting that they affected migratoryonset (Marret et al., 1996). These migration defects could beattributed to both migration arrest and unsufficient terminationof migration (Takano et al., 2004). Using sustained-releasepolymer Elvax implants (Smith et al., 1995) containing MK801to deliver this NMDA antagonist focally to the cortical surface,Reiprich et al. (2005) could demonstrate that a local andtransient NMDA receptor blockade in the somatosensory cortexof newborn rats in vivo produces structural and functionalalterations in the cortical region underlying the implant(Figure 2A). MK801-treated animals showed disturbances in thecortical lamination and heterotopic cell clusters in the upperlayers.
Complete knockout of NR1, an essential subunit of NMDAreceptors, has no effect on the early migration pattern ofneocortical neurons in the fetal mouse brain, but micedie at birth due to respiratory problems (Messersmithet al., 1997). A restricted knockout of NR1 in excitatoryneocortical neurons (CxNR1KO) led to only slight changesin the neocortical organization, like a disordered barrelcortex, without gross anatomical disturbances reminiscentof cortical migration disorders (Iwasato et al., 2000), butin these animals residual amounts of functional NMDAreceptors may be present during prenatal development.The function of NR1 in neuronal migration may be alsocompensated by other mechanisms in CxNR1KO. On theother hand, in chimeric mice transfected with NR1-deficientstem cells, neurons without functional NMDA receptors show
Frontiers in Cellular Neuroscience www.frontiersin.org January 2015 | Volume 9 | Article 4 | 5
Luhmann et al. GABA and glutamate in neuronal migration
a normal distribution within the hippocampus, indicatingthat NMDA receptors on neuronal membranes itself may bedispensable for correct radial migration (Maskos and McKay,2003).
Thus no final conclusion on the role of NMDA receptorsfor migration can currently be given. While pharmacologicalin vivo and in vitro experiments strongly suggest an importantrole of NMDA receptors for radial migration, the observationthat neurons lacking functional NMDA receptors show adequatemigration questions this conclusion. These conflicting results mayeither indicate that the NMDA receptor dependent effects aremediated by non-neuronal target structures like glial cells or thatcompensatory mechanisms may counteract the lack of functionalNMDA receptors.
The source of extracellular glutamate controlling neuronalmigration is not completely known. In vitro studies onhippocampal organotypic slice co-culture assays from munc18-1knockout mice, in which vesicular transmitter release is deleted,indicate that glutamate and also GABA is released in a SNARE-independent manner and both transmitters control neuronalmigration via a paracrine action (Manent et al., 2005). Anothermechanism of extracellular transmitter control are transporters.Glutamate uptake by transporters expressed in astrocytes setextracellular glutamate levels. The expression of glutamatetransporters is relatively low in immature rodent hippocampusand increases during early postnatal development (Thomas et al.,2011), suggesting that extracellular concentrations of glutamatemay be higher during early corticogenesis when neuronalmigration occurs. However, extracellular space is also largerduring early development (for review, Syková, 2004), thereforeoverall extracellular transmitter concentrations in the youngbrain may be not so much higher than in adult. Furthermore,inhibition of glutamate uptake enhances migration (Komuro andRakic, 1993), which indicates that glutamate is sequestered ratherthan released in the vicinity of migration neurons. Related to theglutamatergic system, it has been demonstrated in the cerebellumthat glutamate activates Bergmann glial cells to produce andrelease d-serine, which potentiates glutamate actions on NMDAreceptors and enhances neuronal migration of cerebellar granuleneurons (Kim et al., 2005).
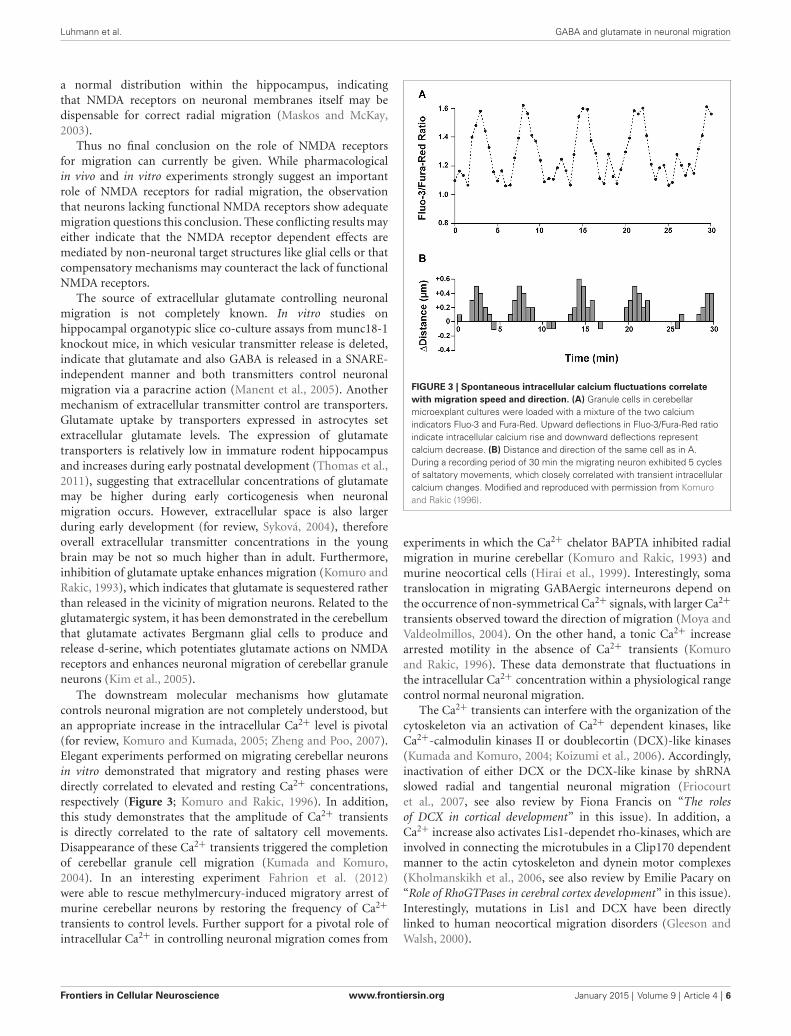
The downstream molecular mechanisms how glutamatecontrols neuronal migration are not completely understood, butan appropriate increase in the intracellular Ca2+ level is pivotal(for review, Komuro and Kumada, 2005; Zheng and Poo, 2007).Elegant experiments performed on migrating cerebellar neuronsin vitro demonstrated that migratory and resting phases weredirectly correlated to elevated and resting Ca2+ concentrations,respectively (Figure 3; Komuro and Rakic, 1996). In addition,this study demonstrates that the amplitude of Ca2+ transientsis directly correlated to the rate of saltatory cell movements.Disappearance of these Ca2+ transients triggered the completionof cerebellar granule cell migration (Kumada and Komuro,2004). In an interesting experiment Fahrion et al. (2012)were able to rescue methylmercury-induced migratory arrest ofmurine cerebellar neurons by restoring the frequency of Ca2+
transients to control levels. Further support for a pivotal role ofintracellular Ca2+ in controlling neuronal migration comes from
FIGURE 3 | Spontaneous intracellular calcium fluctuations correlatewith migration speed and direction. (A) Granule cells in cerebellarmicroexplant cultures were loaded with a mixture of the two calciumindicators Fluo-3 and Fura-Red. Upward deflections in Fluo-3/Fura-Red ratioindicate intracellular calcium rise and downward deflections representcalcium decrease. (B) Distance and direction of the same cell as in A.During a recording period of 30 min the migrating neuron exhibited 5 cyclesof saltatory movements, which closely correlated with transient intracellularcalcium changes. Modified and reproduced with permission from Komuroand Rakic (1996).
experiments in which the Ca2+ chelator BAPTA inhibited radialmigration in murine cerebellar (Komuro and Rakic, 1993) andmurine neocortical cells (Hirai et al., 1999). Interestingly, somatranslocation in migrating GABAergic interneurons depend onthe occurrence of non-symmetrical Ca2+ signals, with larger Ca2+
transients observed toward the direction of migration (Moya andValdeolmillos, 2004). On the other hand, a tonic Ca2+ increasearrested motility in the absence of Ca2+ transients (Komuroand Rakic, 1996). These data demonstrate that fluctuations inthe intracellular Ca2+ concentration within a physiological rangecontrol normal neuronal migration.
The Ca2+ transients can interfere with the organization of thecytoskeleton via an activation of Ca2+ dependent kinases, likeCa2+-calmodulin kinases II or doublecortin (DCX)-like kinases(Kumada and Komuro, 2004; Koizumi et al., 2006). Accordingly,inactivation of either DCX or the DCX-like kinase by shRNAslowed radial and tangential neuronal migration (Friocourtet al., 2007, see also review by Fiona Francis on “The rolesof DCX in cortical development” in this issue). In addition, aCa2+ increase also activates Lis1-dependet rho-kinases, which areinvolved in connecting the microtubules in a Clip170 dependentmanner to the actin cytoskeleton and dynein motor complexes(Kholmanskikh et al., 2006, see also review by Emilie Pacary on“Role of RhoGTPases in cerebral cortex development” in this issue).Interestingly, mutations in Lis1 and DCX have been directlylinked to human neocortical migration disorders (Gleeson andWalsh, 2000).
Frontiers in Cellular Neuroscience www.frontiersin.org January 2015 | Volume 9 | Article 4 | 6

Luhmann et al. GABA and glutamate in neuronal migration
In summary, there is compelling evidence that glutamatecontrols radial migration of glutamatergic neurons, mostprobably by acting on NMDA receptors. The mechanisms ofthe glutamate effect on tangential migration of GABAergicinterneurons is less established and here AMPA receptors are morerelevant.
ROLE OF GABA AND TAURINE IN NEURONAL MIGRATIONThe classical inhibitory neurotransmitter GABA is importantin controlling neuronal migration via ionotropic GABAA andmetabotropic GABAB receptors (Manent and Represa, 2007).GABAA receptors are heteropentamers compiled from in total19 subunits, divided into eight groups, while GABAB receptorsare heterodimers co-assembled from the GABAB2 subunit withone of the two isoforms of the GABAB1 subunit (for a detailedreview, see Farrant and Kaila, 2007; Ulrich and Bettler, 2007).Several GABA receptor subunits are abundantly expressed duringearly cortical development. At E14 the GABAA receptor subunitsα2, α3, α4, β1 and γ1 are expressed, with α3 expressed atparticular high levels during prenatal development (Laurie et al.,1992). Accordingly, GABAA receptor mediated currents areobserved already in proliferative neuroblasts and early postmitoticneurons (LoTurco et al., 1995; Owens et al., 1999). In linewith the paucity of α1 and γ2 expression, immature corticalneurons show GABAA receptor mediated currents with slowkinetics and little desensitization, high GABA affinity and lackof synaptic GABAergic currents before they terminate migrationin the CP (Owens et al., 1999). In addition to this classicalGABAA receptor, ρ subunit containing GABAA-rho receptors,characterized by an exceptionally high GABA affinity and littledesensitization, are found in the SVZ, while they are lacking in CPneurons (Denter et al., 2010). GABAB1 and GABAB2 subunits areexpressed throughout all neocortical lamia during early stages ofcortical development (López-Bendito et al., 2002b). Interestinglytangentially migrating neurons express only GABAB1 subunitsand should thus lack functional GABAB receptors (López-Benditoet al., 2002b). Finally, it is important to consider that immatureneocortical neurons show a high ratio in the expression of NKCC1to KCC2, which renders GABAA mediated responses depolarizing(Yamada et al., 2004).
The implication of GABA receptors in the control of neuronalmigration was first demonstrated by Behar et al. (1996), whocould show by the use of a microchemotaxis chamber thatneuronal migration of dissociated cortical neurons of embryonicrats is stimulated by low concentrations of GABA actingon GABAA/GABAA-rho and GABAB receptors. Femtomolarconcentrations of GABA induced chemotaxis (migration along achemical gradient) and micromolar GABA initiated chemokinesis(increased random movement). In a subsequent study Behar et al.(1998) showed that 1–5 µM GABA stimulated the migration ofGAD-expressing neurons in the CP, whereas 500 fM stimulatedmotility of GAD-expressing neurons in the VZ. In this studythe authors also postulate that GABA can promote migrationvia G-protein activation, mediated by GABAB receptors, andarrest migration via GABAA receptor-mediated depolarization(Behar et al., 1998). Using organotypic neocortical slice culturesBehar et al. demonstrated that specific activation of the different
GABA receptors modified different parts of neural migration tothe CP (Behar et al., 2000): (i) GABAA/GABAA-rho receptoractivation promoted neuronal migration from the VZ/SVZ tothe IZ; (ii) GABAB receptor controlled the migration from theIZ into the CP; and finally (iii) GABAA receptor activationprovided the stop signal to terminate migration in the upper CP(Behar et al., 2000). The two ionotropic GABA receptor subtypes,GABAA and GABAA-rho receptors, play different roles in thecontrol of radial migration (Figure 4A). In organotypic murineneocortical slice cultures application of bicuculline methiodide(BMI) facilitated neuronal migration, while the GABAA-rhospecific antagonist TPMPA attenuated migration (Denter et al.,2010). These data add important information to the model onthe role of GABA in neuronal migration as initially introducedby Behar et al. (2000). GABAA-rho receptors have a highaffinity to GABA and are transiently expressed on migratingneurons in the IZ, where the extracellular GABA concentrationat late embryonic stages is relatively low (Figure 4A). In theIZ, GABA acting primarily on GABAA-rho receptors acts as aGO signal for migrating neurons coming from the VZ/SVZ andpassing through the IZ to the CP. Reaching the CP, migratingneurons no longer express functional GABAA-rho receptors.Due to the intracortical outside directed GABA gradient low-affinity GABAA receptors are now activated and GABA servesas a STOP signal in the upper CP (Denter et al., 2010).Although this model is supported by a number of experimentalstudies, data on the existence of the outside directed GABAgradient are scare. Regional differences in the amplitudes ofGABAA receptor-mediated tonic currents suggest a gradient ofendogenous GABAA receptor agonists in embryonic murinecortex (Furukawa et al., 2014). However, detailed informationabout spatial gradients in extracellular GABA concentrations arecurrently not available (Bolteus et al., 2005). GABA imaging inbrain slices using immobilized enzyme-linked photoanalysis mayprovide a possibility to demonstrate such a gradient (Morishimaet al., 2010).
Blockade of GABAB receptors in organotypic cultures of therat brain led to an accumulation of tangentially migrating neuronsin the VZ/SVZ of the rat neocortex (López-Bendito et al., 2003),suggesting a role of GABAB receptors for final apposition ofGABAergic interneurons. However since no functional GABAB
receptors are expressed on these neurons (López-Bendito et al.,2002b), this effect may be caused by GABAB receptor-dependenteffect on targets upstream from the migration tangential neuronsitself (López-Bendito et al., 2002b).
A direct influence of GABA on the migration of corticalneurons has also be shown in vivo. Furukawa et al. (2014)demonstrated in vivo that continuous blockade of GABAA
receptors with the GABAA antagonist gabazine (SR95531) duringlate embryonic stages accelerated radial migration in the murineneocortex (Figure 4B). In line with this, Elvax implants loadedwith the GABAA antagonist BMI or the agonist muscimol placedon the neocortical surface of newborn rats induced heterotopiccell clusters in upper layers and a loss of neocortical lamination,probably because of an overmigration from the loss of a stopsignal (Figure 4C). Whereas BMI caused this effect by blockingGABAA receptors, long-term application of muscimol induced
Frontiers in Cellular Neuroscience www.frontiersin.org January 2015 | Volume 9 | Article 4 | 7

Luhmann et al. GABA and glutamate in neuronal migration
FIGURE 4 | Role of ionotropic GABA receptors on radial migration.(A) Model of GABAA and GABAA-rho receptor dependent radialmigration in the neonatal cerebral cortex, which shows outsidedirected GABA gradient (gray colored gradient). In the IZ migratingneurons express functional GABAA receptors (blue discs) andGABAA-rho receptors (orange discs), whereas in the CP migratingneurons express only functional GABAA receptors. Due to the outsidedirected GABA gradient the low-affinity GABAA receptors are onlyactivated in the CP, while the lower GABA concentration in the IZ issufficient to activate the high affinity GABAA-rho receptors. Activationof GABAA-rho receptors is necessary to support migration in the IZ(GO sign), while activation of GABAA receptors contributes totermination of migration (STOP sign). (B) Blockade of GABAA
receptors with gabazine facilitates radial migration. Figures illustratered fluorescent protein (RFP)-positive cells in control (left) andgabazine-treated (right) GAD67GFP/GFP fetuses at E17.5, which wereinjected with gabazine at E14.5 immediately after the electroporationof the RFP vectors. (C) Digital photographs of Nissl-stained coronalsections from a P7 rat that received at P0 on the cortical surface anElvax implant containing DMSO (top, control), the GABAA antagonistbicuculline methiodide (middle, BMI) or the GABAA agonist muscimol(bottom). Note upper layer heterotopia due to increased radialmigration in BMI- and muscimol-treated animals. Scale bar in B, Cmiddle and C bottom corresponds to 200 µm, in C top to 500 µm.Modified with permission from Denter et al. (2010) (A), Furukawaet al. (2014) (B), and Heck et al. (2007) (C).
a pronounced receptor desensitization, thereby also reducingGABAA receptor function on migrating rat neurons (Hecket al., 2007). In accordance with the results of in vitro studiesidentifying the role of GABAB receptors (Behar et al., 2001), inutero knockdown of GABAB receptors using RNA interferencetechniques impaired radial migration of the affected pyramidalneuron progenitors in the rat neocortex (Bony et al., 2013).The superficial stream of tangentially migrating GABAergicinterneurons in the MZ of neonatal mice is also impaired afterinhibition of GABAA receptors in vivo (Inada et al., 2011),demonstrating a direct influence of endogenous GABA also ontangential migration.
In summary, these reports demonstrate that ionotropicGABAA and metabotropic GABAB receptors are involved in thecontrol of neuronal migration in the cortex.
One feature of the GABA receptors expressed on migratingneurons is their high GABA affinity, allowing them to sense evenlow ambient GABA concentrations. Microdialysis experimentsin tangential neocortical slices revealed an extracellular GABAconcentration of 25 nmol/l in the MZ of early postnatal rats(Qian et al., 2014), however, this experimental paradigm mayunderestimate the interstitial GABA concentration close to
migrating neurons. Using GABAergic modulation of glutamaterelease Dvorzhak et al. (2010) suggested a juxtasynaptic GABAconcentration of 250 nmol/l during early postnatal stages inthe mouse, with a substantial developmental decrease duringthe first postnatal week. A substantially higher ambient GABAconcentration of ∼0.5 µmol/l was observed in the ganglioniceminence of mice using GABAA receptor expressing sniffer cells asGABA sensors (Cuzon et al., 2006), which may be relevant for thetangential migration of GABAergic interneurons from this region.
It was suggested that tangentially migrating GABAergicneurons are a source for GABA (Manent et al., 2005). In vitroassays indicated that CP neurons itself secrete promigratorysignals acting on GABA receptors and suggested that these signalsmay include GABA and/or taurine (Behar et al., 2001). In linewith this, tangentially migrating neurons in GAD67-GFP knock-in mice had a substantially slower migration rate, which hasbeen attributed to the lower extracellular GABA level in theseanimals (Inada et al., 2011). However, no obvious disorders ingross neocortical organization have been observed after completeblockade of GABA synthesis in GAD65/GAD67 knockout mice(Ji et al., 1999), indicating that other substances can act asGABAergic agonists during prenatal development.
Frontiers in Cellular Neuroscience www.frontiersin.org January 2015 | Volume 9 | Article 4 | 8
Luhmann et al. GABA and glutamate in neuronal migration
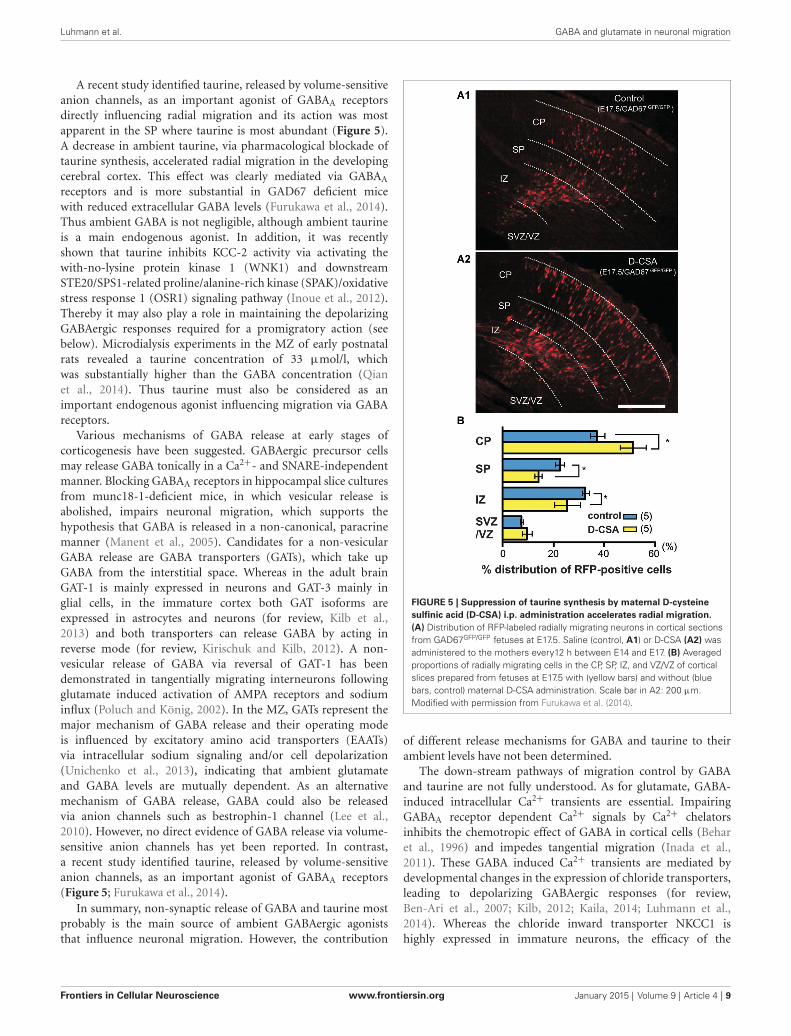
A recent study identified taurine, released by volume-sensitiveanion channels, as an important agonist of GABAA receptorsdirectly influencing radial migration and its action was mostapparent in the SP where taurine is most abundant (Figure 5).A decrease in ambient taurine, via pharmacological blockade oftaurine synthesis, accelerated radial migration in the developingcerebral cortex. This effect was clearly mediated via GABAA
receptors and is more substantial in GAD67 deficient micewith reduced extracellular GABA levels (Furukawa et al., 2014).Thus ambient GABA is not negligible, although ambient taurineis a main endogenous agonist. In addition, it was recentlyshown that taurine inhibits KCC-2 activity via activating thewith-no-lysine protein kinase 1 (WNK1) and downstreamSTE20/SPS1-related proline/alanine-rich kinase (SPAK)/oxidativestress response 1 (OSR1) signaling pathway (Inoue et al., 2012).Thereby it may also play a role in maintaining the depolarizingGABAergic responses required for a promigratory action (seebelow). Microdialysis experiments in the MZ of early postnatalrats revealed a taurine concentration of 33 µmol/l, whichwas substantially higher than the GABA concentration (Qianet al., 2014). Thus taurine must also be considered as animportant endogenous agonist influencing migration via GABAreceptors.
Various mechanisms of GABA release at early stages ofcorticogenesis have been suggested. GABAergic precursor cellsmay release GABA tonically in a Ca2+- and SNARE-independentmanner. Blocking GABAA receptors in hippocampal slice culturesfrom munc18-1-deficient mice, in which vesicular release isabolished, impairs neuronal migration, which supports thehypothesis that GABA is released in a non-canonical, paracrinemanner (Manent et al., 2005). Candidates for a non-vesicularGABA release are GABA transporters (GATs), which take upGABA from the interstitial space. Whereas in the adult brainGAT-1 is mainly expressed in neurons and GAT-3 mainly inglial cells, in the immature cortex both GAT isoforms areexpressed in astrocytes and neurons (for review, Kilb et al.,2013) and both transporters can release GABA by acting inreverse mode (for review, Kirischuk and Kilb, 2012). A non-vesicular release of GABA via reversal of GAT-1 has beendemonstrated in tangentially migrating interneurons followingglutamate induced activation of AMPA receptors and sodiuminflux (Poluch and König, 2002). In the MZ, GATs represent themajor mechanism of GABA release and their operating modeis influenced by excitatory amino acid transporters (EAATs)via intracellular sodium signaling and/or cell depolarization(Unichenko et al., 2013), indicating that ambient glutamateand GABA levels are mutually dependent. As an alternativemechanism of GABA release, GABA could also be releasedvia anion channels such as bestrophin-1 channel (Lee et al.,2010). However, no direct evidence of GABA release via volume-sensitive anion channels has yet been reported. In contrast,a recent study identified taurine, released by volume-sensitiveanion channels, as an important agonist of GABAA receptors(Figure 5; Furukawa et al., 2014).
In summary, non-synaptic release of GABA and taurine mostprobably is the main source of ambient GABAergic agoniststhat influence neuronal migration. However, the contribution
FIGURE 5 | Suppression of taurine synthesis by maternal D-cysteinesulfinic acid (D-CSA) i.p. administration accelerates radial migration.(A) Distribution of RFP-labeled radially migrating neurons in cortical sectionsfrom GAD67GFP/GFP fetuses at E17.5. Saline (control, A1) or D-CSA (A2) wasadministered to the mothers every12 h between E14 and E17. (B) Averagedproportions of radially migrating cells in the CP, SP, IZ, and VZ/VZ of corticalslices prepared from fetuses at E17.5 with (yellow bars) and without (bluebars, control) maternal D-CSA administration. Scale bar in A2: 200 µm.Modified with permission from Furukawa et al. (2014).
of different release mechanisms for GABA and taurine to theirambient levels have not been determined.
The down-stream pathways of migration control by GABAand taurine are not fully understood. As for glutamate, GABA-induced intracellular Ca2+ transients are essential. ImpairingGABAA receptor dependent Ca2+ signals by Ca2+ chelatorsinhibits the chemotropic effect of GABA in cortical cells (Beharet al., 1996) and impedes tangential migration (Inada et al.,2011). These GABA induced Ca2+ transients are mediated bydevelopmental changes in the expression of chloride transporters,leading to depolarizing GABAergic responses (for review,Ben-Ari et al., 2007; Kilb, 2012; Kaila, 2014; Luhmann et al.,2014). Whereas the chloride inward transporter NKCC1 ishighly expressed in immature neurons, the efficacy of the
Frontiers in Cellular Neuroscience www.frontiersin.org January 2015 | Volume 9 | Article 4 | 9

Luhmann et al. GABA and glutamate in neuronal migration
chloride outward transporter KCC2 is low, and this imbalancein chloride transport causes a high intracellular concentrationof chloride ions (Yamada et al., 2004). In accordance withthis hypothesis, a pharmacological blockade of NKCC1 usingbumetanide impairs tangential migration of murine GABAergicinterneurons in vivo (Inada et al., 2011). This hypothesis waschallenged by the observation, that premature expression ofKCC2 by in utero expression at E17/18 causes no obviousmigration deficits of rat neocortical neurons, while causing ahyperpolarizing shift in the chloride reversal potential of GABA-induced currents at early postnatal stages (Cancedda et al., 2007).This result is not too surprising, because ectopically expressedwild type KCC2 is not active in embryonic cerebral corticesand becomes functional only postnatally (Inoue et al., 2012).In addition, the in utero expression was performed at relativelylate stages, so that a substantial part of radial migration tolayer II/III was already accomplished until E21 (Cancedda et al.,2007). Indeed, ectopic expression of constitutive active KCC2mutant at E15 lowered intracellular chloride concentrations,rendered hyperpolarizing GABAA receptor mediated responsesin postmitotic neurons and perturbed their radial migration(Inoue et al., 2012). In migrating murine interneurons thechloride outward transporter KCC2 increases in expressionand becomes functional after they enter the cerebral cortex(Bortone and Polleux, 2009), resulting in a reduced intracellularchloride concentration. The consequent shift in GABAergicaction from excitation to inhibition leads to a decrease inthe frequency of spontaneous intracellular Ca2+ transientsand terminates neuronal migration, thus turning GABA intoa STOP signal for migrating interneurons (Bortone andPolleux, 2009). This scenario is supported by experimentaldata from Inoue et al. as mentioned above (Inoue et al.,2012).
In addition to a direct excitatory effect, depolarizingGABAergic responses are also involved in spontaneous activitypatterns observed in neocortical networks during pre- and earlypostnatal development (for review, Khazipov and Luhmann,2006; Allene and Cossart, 2010; Kilb et al., 2011). In a ratneocortical culture model de Lima et al. (2009) demonstrated arelationship between the expression of spontaneous synchronousnetwork activity and neuronal migration. Although migratinginterneurons did not participate in early cortical network activity,migration was terminated when interneurons became active ina synchronous network. These data indicate that synchronizedGABA and also glutamate release during early network activitycan terminate neuronal migration (de Lima et al., 2009).
In summary, GABA and the endogenous GABAergic agonisttaurine have a strong impact on tangential and radial migration.These neurotransmitters have both, promigratory and migration-terminating actions, depending on the type of GABA receptor andthe intracellular chloride concentration in the migrating neuron.
ROLE OF GLYCINE IN NEURONAL MIGRATIONBeside ionotropic GABA receptors, glycine receptors alsohave an influence on neuronal migration. As GABAA andGABAA-rho receptors, glycine receptors are also transmitter-gated chloride channels, which upon activation by glycine or
taurine mediate a depolarizing or even excitatory action inthe immature cortex (Flint et al., 1998; Kilb et al., 2002,2008). A functional expression of heteromeric glycine receptors,compiled from α2/β subunits, has already been describedin various types of immature neurons, including putativemigratory neurons in the IZ (Flint et al., 1998; Kilb et al.,2002, 2008; Okabe et al., 2004), whereas tangentially migratingneurons express α2 homomeric glycine receptors (Avila et al.,2013). It is therefore not surprising that an activation ofglycine receptors also promoted radial neuronal migration asdemonstrated in organotypic slice cultures from embryonicmouse cerebral cortex (Nimmervoll et al., 2011). However,as pharmacological inhibition of glycine receptors did notinterfere with radial migration, Nimmervoll et al. (2011) suggestthat glycine receptors do not contribute substantially to radialmigration in the neocortex (see also Furukawa et al., 2014).In contrast, tangential migration of cortical interneurons waseffectively attenuated by genetic or pharmacological suppressionof glycine receptor function in organotypic slice cultures frommouse cortex (Avila et al., 2013). In this study, the migrationspeed was not affected by addition of taurine, suggestingthat glycine itself acts as endogenous neurotransmitter. In linewith this suggestion, the estimated extracellular glycine levelsof ∼150 nmol/l (Qian et al., 2014) would allow a partialactivation of α2 subunit containing receptors with their EC50
of ∼0.5 µmol/l (Flint et al., 1998; Okabe et al., 2004), whilethe estimated extracellular taurine concentration of 33 µmol/l(Qian et al., 2014) is most probably ineffective to activateglycine receptors (EC50 for taurine ∼2.5 mM, Okabe et al.,2004).
In summary, these results indicate that glycine receptorscan affect neuronal migration, although these receptors may berelevant only for tangential migration.
ROLE OF GLUTAMATE AND GABA IN NEURONAL MIGRATIONDISORDERSGiven the pivotal role of glutamate and GABA in controllingneuronal migration in the developing cortex, it is not surprisingthat any modulation in the function of these neurotransmittersduring pre- and early postnatal periods may have profoundeffects on the generation of the cortical architecture. Sinceneuronal migration is indirectly also controlled by spontaneousnetwork activity, modulation of these transmitter systems mayalso cause disturbances in early neuronal activity patternssubsequently leading to migration deficits (for review, Kilb et al.,2011).
A direct role of GABA for migration disorders has recentlybeen demonstrated in experimentally induced polymicrogyria.Wang et al. (2014) observed that the accumulation of neurons inthe polymicrogyria forming below a neocortical freeze lesion wasprevented by the administration of GABAA receptor antagonistsin vivo. An altered migration caused by excitatory GABAA
receptors has also been revealed as a cause for the hippocampalgranule cell ectopia observed after febrile seizures (Koyama et al.,2012).
A number of drugs, which are taken by pregnant women forcontrol of psychiatric or neurological disorders (e.g., epilepsy),
Frontiers in Cellular Neuroscience www.frontiersin.org January 2015 | Volume 9 | Article 4 | 10

Luhmann et al. GABA and glutamate in neuronal migration
anesthetics required for surgical operation of pregnant women,or drug abuse (e.g., ethanol consumption during pregnancy)may have profound effects on neuronal migration patternsin the cortex of the unborn child. These drugs often acton glutamate and/or GABA receptors and, when reaching theimmature brain, may change the migration pattern of corticalpyramidal cells and GABA interneurons. Anti-epileptic drugshave a wide range of actions (for review Ikonomidou and Turski,2010): (i) increasing GABAergic action by inhibiting degradationor uptake mechanisms of GABA; (ii) potentiating GABAergicfunction by acting on GABAA receptor subtypes; (iii) reducingpresynaptic glutamate release; (iv) inhibiting glutamate receptorfunction; and (v) modulating neuronal activity by inhibitingvoltage-gated sodium and Ca2+ channels. The most potentanti-epileptic drugs are often unspecific and act via several ofthese mechanisms. In experimental studies, anti-epileptic drugswere administered in clinically relevant doses to pregnant ratsduring the last week of gestation (for review, Manent et al.,2011). Prenatal exposure to the antiepileptic drugs vigabatrinand valproate, which both increase extracellular GABA levels,induced neuronal migration disorders in the hippocampus andcerebral cortex (Manent et al., 2007). In line with this, thebenzodiazepine diazepam, which augments the activation ofGABAA receptors, substantially increased the motility rate ofmigrating GABAergic interneurons (Inada et al., 2011). Incontrast the anticonvulsant carbamazepine, which mainly acts onvoltage-dependent sodium channels and has only minor effectson GABA receptor function, did not cause major disturbancesin neuronal migration (Manent et al., 2007). These data indicatethat changes in the extracellular concentration of GABA have astronger influence on neuronal migration than modifications inspontaneous neuronal activity.
Alcohol exposure during pregnancy is one of the leadingcauses of mental retardation and several neuroanatomicalmalformations, including migration disorders such aslissencephaly and cortical heterotopias (for review, Miller,1992). Ethanol is a drug acting on ionotropic GABA receptors(for review, Lobo and Harris, 2008) and NMDA receptors (forreview, Chandrasekar, 2013), which both are key players inthe control of neuronal migration. Clinically relevant levels ofethanol substantially impaired migration of cerebellar neuronsby attenuating intracellular Ca2+ signals (Kumada et al., 2006).In contrast, exposure to relatively low levels of ethanol inutero elevated ambient GABA level, enhanced the sensitivity ofMGE-derived interneurons to GABA and promoted prematuretangential migration into the cortical anlage (Cuzon et al.,2008). A direct link between neuronal migration disorders andglutamate receptor dysfunction has been found in a mouse modelfor Zellweger disease. Here the migration defect results from amutation in the NMDA receptor mediated Ca2+ mobilization(Gressens et al., 2000).
In summary, exposure of the prenatal human brain to drugsand pharmacological agents acting on glutamate and/or GABAreceptors, may have a profound influence on tangential and radialmigration. The resulting neuronal migration disorders may bedifficult to detect and may result in therapy-resistant neurologicaland neuropsychiatric disorders.
OPEN QUESTIONS AND CONCLUSIONSAlthough the last two decades provided a large amount ofexperimental data on the role of glutamate and GABA onneuronal migration in the cerebral cortex, a number of questionsneed to be addressed in order to understand the function of thesetwo classical neurotransmitters and other transmitters in moredetail.
1. What is the function of different receptor subunits inneuronal migration? It became recently clear that migratingneurons change during the migration process the expressionof their functional receptors, receptor subtypes and subunits.However, the exact role of these developmental changes on themigration process is largely unknown.
2. What are the physiological extracellular concentrations ofglutamate, GABA, taurine and glycine in distinct brainregions (e.g., cortical layers) and at different developmentalperiods? The answer to this question will provide importantinformation to understand tangential and radial migrationpatterns.
3. What are the roles of other transmitter system besideglutamate and GABA? There are a few reports that otherclassical neurotransmitters like dopamine or serotonin are alsoimplicated in the control of neuronal migration (e.g., Crandallet al., 2007; Riccio et al., 2011). However, a detailed analysis ofthe mechanisms of their actions or the identification of otherneurotransmitters affecting migration remains open.
4. Which neuronal migration patterns are physiological andwhich ones are pathophysiological? This question is of majorclinical relevance and may be difficult to answer with in vitroapproaches, where experimental conditions are more or lessartificial.
5. When and how does a neuronal migration disorder result ina clinical manifestation of a neurological or neuropsychiatricdisease? This is another clinically relevant question andaddresses the problem that small structural alterations dueto migration disorders may be difficult to detect withconventional imaging techniques.
6. At which time points are the different migration processesrespond most sensitive to noxious stimuli? Drugs, hypoxia,inflammation and other noxious conditions in the prenatalbrain probably influence the different types and modes ofmigration in a time-dependent manner. E.g., it is unclearin which developmental period of the prenatal human braincertain drugs have strong impact on radial or tangentialmigration.
7. What are the (epi-)genetic causes of neuronal migrationdisorders? Although this field strongly developed over thelast decade (for review, LoTurco and Bai, 2006; Guerrini andParrini, 2010), the influence of genetic and epigenetic factorsin distinct migration processes is often still not known.
8. What are the migratory pathways and mechanisms inprimates? The vast majority of information on neuronalmigration comes from experimental studies on rodents.However, some early events in neocortical development aredifferent between rodents and primates (for review Rakic,2009). A better understanding of neuronal migration and
Frontiers in Cellular Neuroscience www.frontiersin.org January 2015 | Volume 9 | Article 4 | 11

Luhmann et al. GABA and glutamate in neuronal migration
migration disorders in humans, may require further studies inprimates.
In summary, a variety of studies provide substantial evidencethat the classical neurotransmitters GABA and glutamateinfluence neuronal migration and may thus directly contribute tothe pathogenesis of neuronal migration disorders. However, theeffects of these neurotransmitters are not uniform, but depend onthe brain region, identity and maturational state of the migratingneuron and the neurotransmitter receptor subtypes involved.Awareness of the complex interplay between neurotransmitteraction and cellular migration processes may help to preventmigration disorders during fetal development.
ACKNOWLEDGMENTSWe thank our colleagues who contributed to the work reviewedin this paper. This work was supported by the DeutscheForschungsgemeinschaft (SFB 1080 and KI835/2).
REFERENCESAllene, C., and Cossart, R. (2010). Early NMDA receptor-driven waves of activity
in the developing neocortex: physiological or pathological network oscillations?J. Physiol. 588, 83–91. doi: 10.1113/jphysiol.2009.178798
Avila, A., Vidal, P. M., Dear, T. N., Harvey, R. J., Rigo, J. M., and Nguyen, L. (2013).Glycine receptor alpha 2 subunit activation promotes cortical interneuronmigration. Cell Rep. 4, 738–750. doi: 10.1016/j.celrep.2013.07.016
Ayala, R., Shu, T., and Tsai, L. H. (2007). Trekking across the brain: the journey ofneuronal migration. Cell 128, 29–43. doi: 10.1016/j.cell.2006.12.021
Bagnard, D., Vaillant, C., Khuth, S. T., Dufay, N., Lohrum, M., Püschel, A. W.,et al. (2001). Semaphorin 3A-vascular endothelial growth factor-165 balancemediates migration and apoptosis of neural progenitor cells by the recruitmentof shared receptor. J. Neurosci. 21, 3332–3341.
Bahn, S., Volk, B., and Wisden, W. (1994). Kainate receptor gene expression in thedeveloping rat brain. J. Neurosci. 14, 5525–5547.
Behar, T. N., Li, Y. X., Tran, H. T., Ma, W., Dunlap, V., Scott, C., et al. (1996).GABA stimulates chemotaxis and chemokinesis of embryonic cortical neuronsvia calcium-dependent mechanisms. J. Neurosci. 16, 1808–1818.
Behar, T. N., Schaffner, A. E., Scott, C. A., Greene, C. L., and Barker, J. L. (2000).GABA receptor antagonists modulate postmitotic cell migration in slice culturesof embryonic rat cortex. Cereb. Cortex 10, 899–909. doi: 10.1093/cercor/10.9.899
Behar, T. N., Schaffner, A. E., Scott, C. A., O’Connell, C., and Barker, J. L. (1998).Differential response of cortical plate and ventricular zone cells to GABA as amigration stimulus. J. Neurosci. 18, 6378–6387.
Behar, T. N., Scott, C. A., Greene, C. L., Wen, X. L., Smith, S. V., Maric, D., et al.(1999). Glutamate acting at NMDA receptors stimulates embryonic corticalneuronal migration. J. Neurosci. 19, 4449–4461.
Behar, T. N., Smith, S. V., Kennedy, R. T., Mckenzie, J. M., Maric, I., and Barker,A. L. (2001). GABAB receptors mediate motility signals for migrating embryoniccortical cells. Cereb. Cortex 11, 744–753. doi: 10.1093/cercor/11.8.744
Ben-Ari, Y., Gaiarsa, J. L., Tyzio, R., and Khazipov, R. (2007). GABA: a pioneertransmitter that excites immature neurons and generates primitive oscillations.Physiol. Rev. 87, 1215–1284. doi: 10.1152/physrev.00017.2006
Bolteus, A. J., Garganta, C., and Bordey, A. (2005). Assays for measuringextracellular GABA levels and cell migration rate in acute slices. Brain Res. BrainRes. Protoc. 14, 126–134. doi: 10.1016/j.brainresprot.2004.12.005
Bony, G., Szczurkowska, J., Tamagno, I., Shelly, M., Contestabile, A., andCancedda, L. (2013). Non-hyperpolarizing GABA(B) receptor activationregulates neuronal migration and neurite growth and specification bycAMP/LKB1. Nat. Commun. 4:1800. doi: 10.1038/ncomms2820
Borrell, V., and Marín, O. (2006). Meninges control tangential migration of hem-derived Cajal-Retzius cells via CXCL12/CXCR4 signaling. Nat. Neurosci. 9,1284–1293. doi: 10.1038/nn1764
Bortone, D., and Polleux, F. (2009). KCC2 expression promotes the terminationof cortical interneuron migration in a voltage-sensitive calcium-dependentmanner. Neuron 62, 53–71. doi: 10.1016/j.neuron.2009.01.034
Butt, S. J. B., Cobos, I., Golden, J., Kessaris, N., Pachnis, V., and Anderson, S. (2007).Transcriptional regulation of cortical interneuron development. J.Neurosci. 27,11847–11850. doi: 10.1523/jneurosci.3525-07.2007
Butt, S. J., Fuccillo, M., Nery, S., Noctor, S., Kriegstein, A., Corbin, J. G., et al.(2005). The temporal and spatial origins of cortical interneurons predict theirphysiological subtype. Neuron 48, 591–604. doi: 10.1016/j.neuron.2005.09.034
Bystron, I., Blakemore, C., and Rakic, P. (2008). Development of the humancerebral cortex: Boulder committee revisited. Nat. Rev. Neurosci. 9, 110–122.doi: 10.1038/nrn2252
Cancedda, L., Fiumelli, H., Chen, K., and Poo, M. M. (2007). Excitatory GABAaction is essential for morphological maturation of cortical neurons in vivo.J. Neurosci. 27, 5224–5235. doi: 10.1523/jneurosci.5169-06.2007
Carlson, V. C. C., and Yeh, H. H. (2011). GABA(A) receptor subunit profiles oftangentially migrating neurons derived from the medial ganglionic eminence.Cereb. Cortex 21, 1792–1802. doi: 10.1093/cercor/bhq247
Chandrasekar, R. (2013). Alcohol and NMDA receptor: current research and futuredirection. Front. Mol. Neurosci. 6:14. doi: 10.3389/fnmol.2013.00014
Cina, C., Maass, K., Theis, M., Willecke, K., Bechberger, J. F., and Naus, C. C. (2009).Involvement of the cytoplasmic C-terminal domain of connexin43 in neuronalmigration. J. Neurosci. 29, 2009–2021. doi: 10.1523/jneurosci.5025-08.2009
Cooper, J. A. (2008). A mechanism for inside-out lamination in the neocortex.Trends Neurosci. 31, 113–119. doi: 10.1016/j.tins.2007.12.003
Crandall, J. E., McCarthy, D. M., Araki, K. Y., Sims, J. R., Ren, J.-Q., and Bhide, P. G.(2007). Dopamine receptor activation modulates GABA neuron migration fromthe basal forebrain to the cerebral cortex. J. Neurosci. 27, 3813–3822. doi: 10.1523/jneurosci.5124-06.2007
Cuzon, V. C., Yeh, P. W., Cheng, Q., and Yeh, H. H. (2006). Ambient GABApromotes cortical entry of tangentially migrating cells derived from the medialganglionic eminence. Cereb. Cortex 16, 1377–1388. doi: 10.1093/cercor/bhj084
Cuzon, V. C., Yeh, P. W., Yanagawa, Y., Obata, K., and Yeh, H. H. (2008). Ethanolconsumption during early pregnancy alters the disposition of tangentiallymigrating GABAergic interneurons in the fetal cortex. J. Neurosci. 28, 1854–1864. doi: 10.1523/jneurosci.5110-07.2008
de Carlos, J. A., Lopez-Mascaraque, L., and Valverde, F. (1996). Dynamics of cellmigration from the lateral ganglionic eminence in the rat. J. Neurosci. 16, 6146–6156.
de Lima, A. D., Gieseler, A., and Voigt, T. (2009). Relationship between GABAergicinterneurons migration and early neocortical network activity. Dev. Neurobiol.69, 105–123. doi: 10.1002/dneu.20696
Denter, D. G., Heck, N., Riedemann, T., White, R., Kilb, W., and Luhmann, H. J.(2010). GABA(C) receptors are functionally expressed in the intermediate zoneand regulate radial migration in the embryonic mouse neocortex. Neuroscience167, 124–134. doi: 10.1016/j.neuroscience.2010.01.049
Dvorzhak, A., Myakhar, O., Unichenko, P., Kirmse, K., and Kirischuk, S.(2010). Estimation of ambient GABA levels in layer I of the mouse neonatalcortex in brain slices. J. Physiol. 588, 2351–2360. doi: 10.1113/jphysiol.2010.187054
Elias, L. A., and Kriegstein, A. R. (2008). Gap junctions: multifaceted regulators ofembryonic cortical development. Trends Neurosci. 31, 243–250. doi: 10.1016/j.tins.2008.02.007
Elias, L. A., Turmaine, M., Parnavelas, J. G., and Kriegstein, A. R. (2010). Connexin43 mediates the tangential to radial migratory switch in ventrally derived corticalinterneurons. J. Neurosci. 30, 7072–7077. doi: 10.1523/jneurosci.5728-09.2010
Elias, L. A., Wang, D. D., and Kriegstein, A. R. (2007). Gap junction adhesion isnecessary for radial migration in the neocortex. Nature 448, 901–907. doi: 10.1038/nature06063
Fahrion, J. K., Komuro, Y., Li, Y., Ohno, N., Littner, Y., Raoult, E., et al. (2012).Rescue of neuronal migration deficits in a mouse model of fetal Minamatadisease by increasing neuronal Ca2+ spike frequency. Proc. Natl. Acad. Sci. U S A109, 5057–5062. doi: 10.1073/pnas.1120747109
Farrant, M., and Kaila, K. (2007). The cellular, molecular and ionic basis ofGABA(A) receptor signalling. Prog. Brain Res. 160, 59–87. doi: 10.1016/s0079-6123(06)60005-8
Flint, A. C., Liu, X. L., and Kriegstein, A. R. (1998). Nonsynaptic glycine receptoractivation during early neocortical development. Neuron 20, 43–53. doi: 10.1016/s0896-6273(00)80433-x
Friocourt, G., Liu, J. S., Antypa, M., Rakic, S., Walsh, C. A., and Parnavelas, J. G.(2007). Both doublecortin and doublecortin-like kinase play a role in cortical
Frontiers in Cellular Neuroscience www.frontiersin.org January 2015 | Volume 9 | Article 4 | 12

Luhmann et al. GABA and glutamate in neuronal migration
interneuron migration. J. Neurosci. 27, 3875–3883. doi: 10.1523/jneurosci.4530-06.2007
Furukawa, T., Yamada, J., Akita, T., Matsushima, Y., Yanagawa, Y., and Fukuda, A.(2014). Roles of taurine-mediated tonic GABAA receptor activation in the radialmigration of neurons in the fetal mouse cerebral cortex. Front. Cell. Neurosci.8:88. doi: 10.3389/fncel.2014.00088
Gelman, D. M., and Marin, O. (2010). Generation of interneuron diversity in themouse cerebral cortex. Eur. J. Neurosci. 31, 2136–2141. doi: 10.1111/j.1460-9568.2010.07267.x
Gerber, A. M., Beaman-Hall, C. M., Mathur, A., and Vallano, M. L. (2010). Reducedblockade by extracellular Mg2+ is permissive to NMDA receptor activation incerebellar granule neurons that model a migratory phenotype. J. Neurochem.114, 191–202. doi: 10.1111/j.1471-4159.2010.06746.x
Gleeson, J. G., and Walsh, C. A. (2000). Neuronal migration disorders: from geneticdiseases to developmental mechanisms. Trends Neurosci. 23, 352–359. doi: 10.1016/s0166-2236(00)01607-6
Gressens, P., Baes, M., Leroux, P., Lombet, A., Van Veldhoven, P., Janssen, A.,et al. (2000). Neuronal migration disorder in Zellweger mice is secondary toglutamate receptor dysfunction. Ann. Neurol. 48, 336–343. doi: 10.1002/1531-8249(200009)48:3<336::aid-ana8>3.0.co;2-q
Guerrini, R., Dobyns, W. B., and Barkovich, A. J. (2008). Abnormal developmentof the human cerebral cortex: genetics, functional consequences and treatmentoptions. Trends Neurosci. 31, 154–162. doi: 10.1016/j.tins.2007.12.004
Guerrini, R., and Parrini, E. (2010). Neuronal migration disorders. Neurobiol. Dis.38, 154–166. doi: 10.1016/j.nbd.2009.02.008
Heck, N., Kilb, W., Reiprich, P., Kubota, H., Furukawa, T., Fukuda, A., et al. (2007).GABA-A receptors regulate neocortical neuronal migration in vitro and in vivo.Cereb. Cortex 17, 138–148. doi: 10.1093/cercor/bhj135
Hirai, K., Yoshioka, H., Kihara, M., Hasegawa, K., Sakamoto, T., Sawada, T., et al.(1999). Inhibiting neuronal migration by blocking NMDA receptors in theembryonic rat cerebral cortex: a tissue culture study. Brain Res. Dev. Brain Res.114, 63–67. doi: 10.1016/s0165-3806(99)00019-x
Holmberg, J., Genander, M., Halford, M. M., Annerén, C., Sondell, M., Chumley,M. J., et al. (2006). EphB receptors coordinate migration and proliferationin the intestinal stem cell niche. Cell 125, 1151–1163. doi: 10.1016/j.cell.2006.04.030
Huang, Z. (2009). Molecular regulation of neuronal migration during neocorticaldevelopment. Mol. Cell. Neurosci. 42, 11–22. doi: 10.1016/j.mcn.2009.06.003
Ikonomidou, C., and Turski, L. (2010). Antiepileptic drugs and brain development.Epilepsy Res. 88, 11–22. doi: 10.1016/j.eplepsyres.2009.09.019
Inada, H., Watanabe, M., Uchida, T., Ishibashi, H., Wake, H., Nemoto, T., et al.(2011). GABA regulates the multidirectional tangential migration of GABAergicinterneurons in living neonatal mice. Plos One 6:e27048. doi: 10.1371/journal.pone.0027048
Inoue, K., Furukawa, T., Kumada, T., Yamada, J., Wang, T. Y., Inoue, R., et al.(2012). Taurine inhibits K+-Cl- cotransporter KCC2 to regulate embryonicCl- homeostasis via With-no-lysine (WNK) protein kinase signaling pathway.J. Biol. Chem. 287, 20839–20850. doi: 10.1074/jbc.m111.319418
Inta, D., Alfonso, J., von Engelhardt, J., Kreuzberg, M. M., Meyer, A. H., van Hooft,J. A., et al. (2008). Neurogenesis and widespread forebrain migration of distinctGABAergic neurons from the postnatal subventricular zone. Proc. Natl. Acad.Sci. U S A 105, 20994–20999. doi: 10.1073/pnas.0807059105
Iwasato, T., Datwani, A., Wolf, A. M., Nishiyama, H., Taguchi, Y., Tonegawa, S., et al.(2000). Cortex-restricted disruption of NMDAR1 impairs neuronal patterns inthe barrel cortex. Nature 406, 726–731. doi: 10.1038/35021059
Jansson, L. C., Louhivuori, L., Wigren, H. K., Nordström, T., Louhivuori, V.,Castrén, M. L., et al. (2013). Effect of glutamate receptor antagonists onmigrating neural progenitor cells. Eur. J. Neurosci. 37, 1369–1382. doi: 10.1111/ejn.12152
Ji, F. Y., Kanbara, N., and Obata, K. (1999). GABA and histogenesis in fetal andneonatal mouse brain lacking both the isoforms of glutamic acid decarboxylase.Neurosci. Res. 33, 187–194. doi: 10.1016/s0168-0102(99)00011-5
Jovanovic, J. N., and Thomson, A. M. (2011). Development of cortical GABAergicinnervation. Front. Cell. Neurosci. 5:14. doi: 10.3389/fncel.2011.00014
Kaila, K. (2014). Cation-chloride cotransporters in neuronal development,plasticity and disease. Nat. Rev. Neurosci. 15, 637–654. doi: 10.1038/nrn3819
Khazipov, R., and Luhmann, H. J. (2006). Early patterns of electrical activity in thedeveloping cerebral cortex of human and rodents. Trends Neurosci. 29, 414–418.doi: 10.1016/j.tins.2006.05.007
Kholmanskikh, S. S., Koeller, H. B., Wynshaw-Boris, A., Gomez, T., Letourneau,P. C., and Ross, M. E. (2006). Calcium-dependent interaction of lis1 withIQGAP1 and Cdc42 promotes neuronal motility. Nat. Neurosci. 9, 50–57.doi: 10.1038/nn1619
Kihara, M., Yoshioka, H., Hirai, K., Hasegawa, K., Kizaki, Z., and Sawada, T. (2002).Stimulation of N-methyl-D-aspartate (NMDA) receptors inhibits neuronalmigration in embryonic cerebral cortex: a tissue culture study. Brain Res. Dev.Brain Res. 138, 195–198. doi: 10.1016/s0165-3806(02)00490-x
Kilb, W. (2012). Development of the GABAergic system from birth to adolescence.Neuroscientist 18, 613–630. doi: 10.1177/1073858411422114
Kilb, W., Hanganu, I. L., Okabe, A., Sava, B. A., Shimizu-Okabe, C., Fukuda,A., et al. (2008). Glycine receptors mediate excitation of subplate neurons inneonatal rat cerebral cortex. J. Neurophysiol. 100, 698–707. doi: 10.1152/jn.00657.2007
Kilb, W., Ikeda, M., Uchida, K., Okabe, A., Fukuda, A., and Luhmann, H. J. (2002).Depolarizing glycine responses in Cajal-Retzius cells of neonatal rat cerebralcortex. Neuroscience 112, 299–307. doi: 10.1016/s0306-4522(02)00071-4
Kilb, W., Kirischuk, S., and Luhmann, H. J. (2011). Electrical activity patterns andthe functional maturation of the neocortex. Eur. J. Neurosci. 34, 1677–1686.doi: 10.1111/j.1460-9568.2011.07878.x
Kilb, W., Kirischuk, S., and Luhmann, H. J. (2013). Role of tonic GABAergiccurrents during pre- and early postnatal rodent development. Front. NeuralCircuits 7:139. doi: 10.3389/fncir.2013.00139
Kim, P. M., Aizawa, H., Kim, P. S., Huang, A. S., Wickramasinghe, S. R., Kashani,A. H., et al. (2005). Serine racemase: activation by glutamate neurotransmissionvia glutamate receptor interacting protein and mediation of neuronal migration.Proc. Natl. Acad. Sci. U S A 102, 2105–2110. doi: 10.1073/pnas.0409723102
Kirischuk, S., and Kilb, W. (2012). “GAT (GABA transporters),” in Encyclopedia ofSignaling Molecules, ed C. Sangdun (New York: Springer), 756–760.
Kirischuk, S., Luhmann, H. J., and Kilb, W. (2014). Cajal-Retzius cells: update onstructural and functional properties of these mystic neurons that bridged the20th century. Neuroscience 275, 33–46. doi: 10.1016/j.neuroscience.2014.06.009
Koizumi, H., Tanaka, T., and Gleeson, J. G. (2006). Doublecortin-like kinasefunctions with doublecortin to mediate fiber tract decussation and neuronalmigration. Neuron 49, 55–66. doi: 10.1016/j.neuron.2005.10.040
Kojima, T., Hirota, Y., Ema, M., Takahashi, S., Miyoshi, I., Okano, H., et al.(2010). Subventricular zone-derived neural progenitor cells migrate along ablood vessel scaffold toward the post-stroke striatum. Stem Cells 28, 545–554.doi: 10.1002/stem.306
Komuro, H., and Kumada, T. (2005). Ca2+ transients control CNS neuronalmigration. Cell Calcium 37, 387–393. doi: 10.1016/j.ceca.2005.01.006
Komuro, H., and Rakic, P. (1993). Modulation of neuronal migration by NMDAreceptors. Science 260, 95–97. doi: 10.1126/science.8096653
Komuro, H., and Rakic, P. (1996). Intracellular Ca2+ fluctuations modulate therate of neuronal migration. Neuron 17, 275–285. doi: 10.1016/s0896-6273(00)80159-2
Koyama, R., Tao, K., Sasaki, T., Ichikawa, J., Miyamoto, D., Muramatsu, R., et al.(2012). GABAergic excitation after febrile seizures induces ectopic granule cellsand adult epilepsy. Nat. Med. 18, 1271–1278. doi: 10.1038/nm.2850
Kumada, T., and Komuro, H. (2004). Completion of neuronal migration regulatedby loss of Ca(2+) transients. Proc. Natl. Acad. Sci. U S A 101, 8479–8484. doi: 10.1073/pnas.0401000101
Kumada, T., Lakshmana, M. K., and Komuro, H. (2006). Reversal of neuronalmigration in a mouse model of fetal alcohol syndrome by controlling second-messenger signalings. J. Neurosci. 26, 742–756. doi: 10.1523/jneurosci.4478-05.2006
Kwan, K. Y., Sestan, N., and Anton, E. S. (2012). Transcriptional co-regulation ofneuronal migration and laminar identity in the neocortex. Development 139,1535–1546. doi: 10.1242/dev.069963
Laurie, D. J., Wisden, W., and Seeburg, P. H. (1992). The distribution of thirteenGABAA receptor subunit mRNAs in the rat brain. III. Embryonic and postnataldevelopment. J. Neurosci. 12, 4151–4172.
Lee, S., Yoon, B. E., Berglund, K., Oh, S. J., Park, H., Shin, H. S., et al. (2010).Channel-Mediated tonic GABA release from Glia. Science 330, 790–796. doi: 10.1126/science.1184334
Le Magueresse, C., Alfonso, J., Bark, C., Eliava, M., Khrulev, S., and Monyer, H.(2012). Subventricular zone-derived neuroblasts use vasculature as a scaffold tomigrate radially to the cortex in neonatal mice. Cereb. Cortex 22, 2285–2296.doi: 10.1093/cercor/bhr302
Frontiers in Cellular Neuroscience www.frontiersin.org January 2015 | Volume 9 | Article 4 | 13

Luhmann et al. GABA and glutamate in neuronal migration
Letinic, K., Zoncu, R., and Rakic, P. (2002). Origin of GABAergic neurons in thehuman neocortex. Nature 417, 645–649. doi: 10.1038/nature00779
Lobo, I. A., and Harris, R. A. (2008). GABA-A receptors and alcohol. Pharmacol.Biochem. Behav. 90, 90–94. doi: 10.1016/j.pbb.2008.03.006
López-Bendito, G., Luján, R., Shigemoto, R., Ganter, P., Paulsen, O., and Molnár, Z.(2003). Blockade of GABA-B receptors alters the tangential migration of corticalneurons. Cereb. Cortex 13, 932–942. doi: 10.1093/cercor/13.9.932
López-Bendito, G., Shigemoto, R., Fairén, A., and Luján, R. (2002a). Differentialdistribution of group I metabotropic glutamate receptors during rat corticaldevelopment. Cereb. Cortex 12, 625–638. doi: 10.1093/cercor/12.6.625
López-Bendito, G., Shigemoto, R., Kulik, A., Paulsen, O., Fairén, A., andLuján, R. (2002b). Expression and distribution of metabotropic GABAreceptor subtypes GABA-BR1 and GABA-BR2 during rat neocorticaldevelopment. Eur. J. Neurosci. 15, 1766–1778. doi: 10.1046/j.1460-9568.2002.02032.x
LoTurco, J. J., and Bai, J. (2006). The multipolar stage and disruptions inneuronal migration. Trends Neurosci. 29, 407–413. doi: 10.1016/j.tins.2006.05.006
LoTurco, J. J., Owens, D. F., Heath, M. J. S., Davis, M. B. E., and Kriegstein,A. R. (1995). GABA and glutamate depolarize cortical progenitor cells andinhibit DNA synthesis. Neuron 15, 1287–1298. doi: 10.1016/0896-6273(95)90008-x
Luhmann, H. J., Kirischuk, S., Sinning, A., and Kilb, W. (2014). Early GABAergiccircuitry in the cerebral cortex. Curr. Opin. Neurobiol. 26, 72–78. doi: 10.1016/j.conb.2013.12.014
Luján, R., Shigemoto, R., and López-Bendito, G. (2005). Glutamate and GABAreceptor signalling in the developing brain. Neuroscience 130, 567–580. doi: 10.1016/j.neuroscience.2004.09.042
Ma, T., Wang, C. M., Wang, L., Zhou, X., Tian, M., Zhang, Q. Q., et al. (2013).Subcortical origins of human and monkey neocortical interneurons. Nat.Neurosci. 16, 1588–1597. doi: 10.1038/nn.3536
Mancini, J. D., and Atchison, W. D. (2007). The NR2B subunit in NMDA receptorsis functionally important during cerebellar granule cell migration. Neurosci.Lett. 429, 87–90. doi: 10.1016/j.neulet.2007.09.079
Manent, J. B., Beguin, S., Ganay, T., and Represa, A. (2011). Cell-autonomous andcell-to-cell signalling events in normal and altered neuronal migration. Eur. J.Neurosci. 34, 1595–1608. doi: 10.1111/j.1460-9568.2011.07867.x
Manent, J. B., Demarque, M., Jorquera, I., Pellegrino, C., Ben-Ari, Y., Aniksztejn,L., et al. (2005). A noncanonical release of GABA and glutamate modulatesneuronal migration. J. Neurosci. 25, 4755–4765. doi: 10.1523/jneurosci.0553-05.2005
Manent, J. B., Jorquera, I., Ben-Ari, Y., Aniksztejn, L., and Represa, A.(2006). Glutamate acting on AMPA but not NMDA receptors modulates themigration of hippocampal interneurons. J. Neurosci. 26, 5901–5909. doi: 10.1523/jneurosci.1033-06.2006
Manent, J. B., Jorquera, I., Mazzucchelli, I., Depaulis, A., Perucca, E., Ben-Ari,Y., et al. (2007). Fetal exposure to GABA-acting antiepileptic drugs generateshippocampal and cortical dysplasias. Epilepsia 48, 684–693. doi: 10.1111/j.1528-1167.2007.01056.x
Manent, J. B., and Represa, A. (2007). Neurotransmitters and brain maturation:early paracrine actions of GABA and glutamate modulate neuronal migration.Neuroscientist 13, 268–279. doi: 10.1177/1073858406298918
Marín, O. (2013). Cellular and molecular mechanisms controlling the migrationof neocortical interneurons. Eur. J. Neurosci. 38, 2019–2029. doi: 10.1111/ejn.12225
Marret, S., Gressens, P., and Evrard, P. (1996). Arrest of neuronal migration byexcitatory amino acids in hamster developing brain. Proc. Natl. Acad. Sci. U S A93, 15463–15468. doi: 10.1073/pnas.93.26.15463
Maskos, U., and McKay, R. D. G. (2003). Neural cells without functional N-methyl-D-aspartate (NMDA) receptors contribute extensively to normal postnatal braindevelopment in efficiently generated chimaeric NMDA R1-/-<->+/+ mice.Dev. Biol. 262, 119–136. doi: 10.1016/s0012-1606(03)00354-3
Messersmith, E. K., Feller, M. B., Zhang, H., and Shatz, C. J. (1997). Migration ofneocortical neurons in the absence of functional NMDA receptors. Mol. Cell.Neurosci. 9, 347–357. doi: 10.1006/mcne.1997.0646
Miller, M. W. (1992). “Effects of prenatal exposure to ethanol on cell proliferationand neuronal migration,” in Development of the Centarl Nervous System, edM. W. Miller (New York, Chichester, Brisbane, Toronto, Singapore: John Wileyand Sons, Inc.), 47–69.
Morishima, T., Uematsu, M., Furukawa, T., Yanagawa, Y., Fukuda, A., and Yoshida,S. (2010). GABA imaging in brain slices using immobilized enzyme-linkedphotoanalysis. Neurosci. Res. 67, 347–353. doi: 10.1016/j.neures.2010.04.005
Moya, F., and Valdeolmillos, M. (2004). Polarized increase of calcium andnucleokinesis in tangentially migrating neurons. Cereb. Cortex 14, 610–618.doi: 10.1093/cercor/bhh022
Nadarajah, B., Alifragis, P., Wong, R. O. L., and Parnavelas, J. G. (2003). Neuronalmigration in the developing cerebral cortex: observations based on real-timeimaging. Cereb. Cortex 13, 607–611. doi: 10.1093/cercor/13.6.607
Nimmervoll, B., Denter, D. G., Sava, I., Kilb, W., and Luhmann, H. J. (2011).Glycine receptors influence radial migration in the embryonic mouse neocortex.Neuroreport 22, 509–513. doi: 10.1097/wnr.0b013e328348aafe
Noctor, S. C., Flint, A. C., Weissman, T. A., Dammerman, R. S., and Kriegstein,A. R. (2001). Neurons derived from radial glial cells establish radial units inneocortex. Nature 409, 714–720. doi: 10.1038/35055553
Noctor, S. C., Martínez-Cerdeño, V., Ivic, L., and Kriegstein, A. R. (2004). Corticalneurons arise in symmetric and asymmetric division zones and migrate throughspecific phases. Nat. Neurosci. 7, 136–144. doi: 10.1038/nn1172
Okabe, A., Kilb, W., Shimizu-Okabe, C., Hanganu, I. L., Fukuda, A., and Luhmann,H. J. (2004). Homogenous glycine receptor expression in cortical plate neuronsand Cajal-Retzius cells of neonatal rat cerebral cortex. Neuroscience 123, 715–724. doi: 10.1016/j.neuroscience.2003.10.014
Owens, D. F., Liu, X. L., and Kriegstein, A. R. (1999). Changing properties ofGABAA receptor-mediated signaling during early neocortical development.J. Neurophysiol. 82, 570–583.
Pla, R., Borrell, V., Flames, N., and Marin, O. (2006). Layer acquisition by corticalGABAergic interneurons is independent of Reelin signaling. J. Neurosci. 26,6924–6934. doi: 10.1523/jneurosci.0245-06.2006
Poluch, S., and König, N. (2002). AMPA receptor activation induces GABA releasefrom neurons migrating tangentially in the intermediate zone of embryonicrat neocortex. Eur. J. Neurosci. 16, 350–354. doi: 10.1046/j.1460-9568.2002.02068.x
Qian, T. Z., Chen, R. Q., Nakamura, M., Furukawa, T., Kumada, T., Akita, T., et al.(2014). Activity-dependent endogenous taurine release facilitates excitatoryneurotransmission in the neocortical marginal zone of neonatal rats. Front. Cell.Neurosci. 8:33. doi: 10.3389/fncel.2014.00033
Rakic, P. (1972). Mode of cell migration to the superficial layers of fetal monkeyneocortex. J. Comp. Neurol. 145, 61–83. doi: 10.1002/cne.901450105
Rakic, P. (2009). Evolution of the neocortex: a perspective from developmentalbiology. Nat. Rev. Neurosci. 10, 724–735. doi: 10.1038/nrn2719
Reiprich, P., Kilb, W., and Luhmann, H. J. (2005). Neonatal NMDA receptorblockade disturbs neuronal migration in rat somatosensory cortex in vivo.Cereb. Cortex 15, 349–358. doi: 10.1093/cercor/bhh137
Riccio, O., Jacobshagen, M., Golding, B., Vutskits, L., Jabaudon, D., Hornung,J.P., et al. (2011). Excess of serotonin affects neocortical pyramidal neuronmigration. Transl. Psychiat. 1:e47. doi: 10.1038/tp.2011.49
Sentürk, A., Pfennig, S., Weiss, A., Burk, K., and Acker-Palmer, A. (2011). Ephrin Bsare essential components of the Reelin pathway to regulate neuronal migration.Nature 472, 356–360. doi: 10.1038/nature09874
Smith, A. L., Cordery, P. M., and Thompson, I. D. (1995). Manufacture and releasecharacteristics of Elvax polymers containing glutamate receptor antagonists.J. Neurosci. Methods 60, 211–217. doi: 10.1016/0165-0270(95)00014-l
Soria, J. M., and Valdeolmillos, M. (2002). Receptor-activated calcium signalsin tangentially migrating cortical cells. Cereb. Cortex 12, 831–839. doi: 10.1093/cercor/12.8.831
Syková, E. (2004). Extrasynaptic volume transmission and diffusion parameters ofthe extracellular space. Neuroscience 129, 861–876. doi: 10.1016/j.neuroscience.2004.06.077
Takano, T., Sawai, C., and Takeuchi, Y. (2004). Radial and tangentialneuronal migration disorder in ibotenate-induced cortical lesions in hamsters:immunohistochemical study of reelin, vimentin and calretinin. J. Child Neurol.19, 107–115. doi: 10.1177/08830738040190020501
Tanaka, D. H., and Nakajima, K. (2012). Migratory pathways of GABAergicinterneurons when they enter the neocortex. Eur. J. Neurosci. 35, 1655–1660.doi: 10.1111/j.1460-9568.2012.08111.x
Tanaka, D., Nakaya, Y., Yanagawa, Y., Obata, K., and Murakami, F. (2003).Multimodal tangential migration of neocortical GABAergic neuronsindependent of GPI-anchored proteins. Development 130, 5803–5813. doi: 10.1242/dev.00825
Frontiers in Cellular Neuroscience www.frontiersin.org January 2015 | Volume 9 | Article 4 | 14

Luhmann et al. GABA and glutamate in neuronal migration
Tanaka, D. H., Yanagida, M., Zhu, Y., Mikami, S., Nagasawa, T., Miyazaki, J.,et al. (2009). Random walk behavior of migrating cortical interneurons in themarginal zone: time-lapse analysis in flat-mount cortex. J. Neurosci. 29, 1300–1311. doi: 10.1523/jneurosci.5446-08.2009
Tárnok, K., Czöndör, K., Jelitai, M., Czirók, A., and Schlett, K. (2008).NMDA receptor NR2B subunit over-expression increases cerebellar granulecell migratory activity. J. Neurochem. 104, 818–829. doi: 10.1111/j.1471-4159.2007.0501.x
Teissier, A., Griveau, A., Vigier, L., Piolot, T., Borello, U., and Pierani, A. (2010). Anovel transient glutamatergic population migrating from the pallial-subpallialboundary contributes to neocortical development. J. Neurosci. 30, 10563–10574.doi: 10.1523/jneurosci.0776-10.2010
Thomas, C. G., Tian, H., and Diamond, J. S. (2011). The relative roles of diffusionand uptake in clearing synaptically released glutamate change during earlypostnatal development. J. Neurosci. 31, 4743–4754. doi: 10.1523/jneurosci.5953-10.2011
Ulrich, D., and Bettler, B. (2007). GABA(B) receptors: synaptic functions andmechanisms of diversity. Curr. Opin. Neurobiol. 17, 298–303. doi: 10.1016/j.conb.2007.04.001
Unichenko, P., Dvorzhak, A., and Kirischuk, S. (2013). Transporter-mediatedreplacement of extracellular glutamate for GABA in the developing murineneocortex. Eur. J. Neurosci. 38, 3580–3588. doi: 10.1111/ejn.12380
Valiente, M., and Marín, O. (2010). Neuronal migration mechanisms indevelopment and disease. Curr. Opin. Neurobiol. 20, 68–78. doi: 10.1016/j.conb.2009.12.003
Wang, Y., Dye, C. A., Sohal, V., Long, J. E., Estrada, R. C., Roztocil, T., et al. (2010).Dlx5 and Dlx6 regulate the development of parvalbumin-expressing corticalinterneurons. J. Neurosci. 30, 5334–5345. doi: 10.1523/jneurosci.5963-09.2010
Wang, T. Y., Kumada, T., Morishima, T., Iwata, S., Kaneko, T., Yanagawa, Y.,et al. (2014). Accumulation of GABAergic Neurons, causing a focal ambientGABA gradient and downregulation of KCC2 are induced during microgyrusformation in a mouse model of polymicrogyria. Cereb. Cortex 24, 1088–1101.doi: 10.1093/cercor/bhs375
Won, C., Lin, Z. C., Kumar, T. P., Li, S. Y., Ding, L., Elkhal, A., et al.(2013). Autonomous vascular networks synchronize GABA neuron migrationin the embryonic forebrain. Nat. Comm. 4:2149. doi: 10.1038/ncomms3149
Yamada, J., Okabe, A., Toyoda, H., Kilb, W., Luhmann, H. J., and Fukuda, A. (2004).Cl- uptake promoting depolarizing GABA actions in immature rat neocorticalneurones is mediated by NKCC1. J. Physiol. 557, 829–841. doi: 10.1113/jphysiol.2004.062471
Yu, Y. C., Bultje, R. S., Wang, X., and Shi, S. H. (2009). Specific synapses developpreferentially among sister excitatory neurons in the neocortex. Nature 458,501–504. doi: 10.1038/nature07722
Zheng, J. Q., and Poo, M. M. (2007). Calcium signaling in neuronal motility. Annu.Rev. Cell Dev. Biol. 23, 375–404. doi: 10.1146/annurev.cellbio.23.090506.123221
Zimmer, G., Garcez, P., Rudolph, J., Niehage, R., Weth, F., Lent, R., et al. (2008).Ephrin-A5 acts as a repulsive cue for migrating cortical interneurons. Eur. J.Neurosci. 28, 62–73. doi: 10.1111/j.1460-9568.2008.06320.x
Conflict of Interest Statement: The authors declare that the research was conductedin the absence of any commercial or financial relationships that could be construedas a potential conflict of interest.
Received: 13 October 2014; accepted: 06 January 2015; published online: 30 January2015.Citation: Luhmann HJ, Fukuda A and Kilb W (2015) Control of corticalneuronal migration by glutamate and GABA. Front. Cell. Neurosci. 9:4.doi: 10.3389/fncel.2015.00004This article was submitted to the journal Frontiers in Cellular Neuroscience.Copyright © 2015 Luhmann, Fukuda and Kilb. This is an open-access articledistributed under the terms of the Creative Commons Attribution License (CC BY).The use, distribution and reproduction in other forums is permitted, provided theoriginal author(s) or licensor are credited and that the original publication in thisjournal is cited, in accordance with accepted academic practice. No use, distributionor reproduction is permitted which does not comply with these terms.
Frontiers in Cellular Neuroscience www.frontiersin.org January 2015 | Volume 9 | Article 4 | 15





























