Embed Size (px)
Citation preview

Available online at www.sciencedirect.com
Developmental Biology 311 (2007) 679–690www.elsevier.com/developmentalbiology
Genomes & Developmental Control
Control of kidney, eye and limb expression of Bmp7 by an enhancerelement highly conserved between species
Derek Adams a, Michele Karolak a, Elizabeth Robertson b,⁎, Leif Oxburgh a,⁎
a Maine Medical Center Research Institute, 81 Research Drive, Scarborough, Maine 04074, USAb Wellcome Trust Center for Human Genetics, University of Oxford, Roosevelt Drive, Oxford OX3 7BN, UK
Received for publication 12 January 2007; revised 10 August 2007; accepted 20 August 2007Available online 28 August 2007
Abstract
Bmp7 is expressed in numerous tissues throughout development and is required for morphogenesis of the eye, hindlimb and kidney. In thisstudy we show that the majority if not all of the cis-regulatory sequence governing expression at these anatomical sites during development ispresent in approximately 20 kb surrounding exon 1. In eye, limb and kidney, multiple distinct enhancer elements drive Bmp7 expression withineach organ. In the eye, the elements driving expression in the pigmented epithelium and iris are spatially separated. In the kidney, Bmp7expression in collecting ducts and nephron progenitors is driven by separate enhancer elements. Similarly, limb mesenchyme and apicalectodermal ridge expression are governed by separate elements. Although enhancers for pigmented epithelium, nephrogenic mesenchyme andapical ectodermal ridge are distributed across the approximately 20 kb region, an element of approximately 480 base pairs within intron 1 governsexpression within the developing iris, collecting duct system of the kidney and limb mesenchyme. This element is remarkably conserved both insequence and position in the Bmp7 locus between different vertebrates, ranging from Xenopus tropicalis to Homo sapiens, demonstrating thatthere is strong selective pressure for Bmp7 expression at these tissue sites. Furthermore, we show that the frog enhancer functions appropriately intransgenic mice. Interestingly, the intron 1 element cannot be found in the Bmp7 genes of vertebrates such as Danio rerio and Takifugu rubripesindicating that this modification of the Bmp7 gene might have arisen during the adaptation from aquatic to terrestrial life. Mutational analysisdemonstrates that the enhancer activity of the intron 1 element is entirely dependent on the presence of a 10 base pair site within the intron 1enhancer containing a predicted binding site for the FOXD3 transcription factor.© 2007 Elsevier Inc. All rights reserved.
Keywords: Bone morphogenetic protein; BMP; BMP7; OP1; Kidney development; Eye development; Limb development; Ureteric bud
Introduction
The bone morphogenetic protein (BMP) ligands comprise alarge family of growth factors sharing structural homology withthe transforming growth factor β proteins. Their expression isessential for key events in early embryonic patterning anddevelopment of multiple organ systems (Hogan, 1996). BMPsregulate processes as diverse as cell proliferation, apoptosis anddifferentiation in a spectrum of tissues. Despite the variety ofcellular responses elicited by these growth factors, the signalingpathway downstream of BMPs is relatively simple, with receptor
⁎ Corresponding authors.E-mail addresses: [email protected] (E. Robertson),
[email protected] (L. Oxburgh).
0012-1606/$ - see front matter © 2007 Elsevier Inc. All rights reserved.doi:10.1016/j.ydbio.2007.08.036
binding leading to phosphorylation and activation of the Smad1,5 and 8 transcription factors that are translocated to the nucleustogether with Smad4 (Massagué, 1998). Differential transcrip-tional responses to BMPs are largely regulated by association ofthe phosphorylated Smad complex with cell type specificauxiliary transcription factors that activate distinct geneticprograms (Massagué and Wotton, 2000). Our previous workhas shown that the developmental functions of BMP7 can beentirely replaced with the closely related BMP6 and largelyreplaced with the more distantly related BMP4 (Oxburgh et al.,2005). The spatial distribution and overall levels of BMPactivities may thus explain their distinct roles in development.Consistent with this idea, exacerbated phenotypes were revealedby compound mutation of the Bmp5, Bmp6 and Bmp7 genes.These BMP family members are expressed in an overlapping

680 D. Adams et al. / Developmental Biology 311 (2007) 679–690
fashion in the heart, but their individual inactivation fails todisrupt heart development. However, inactivation of Bmp7 andBmp5 (Solloway and Robertson, 1999) or Bmp7 and Bmp6(Kim et al., 2001) leads to retardation in heart development anddefects in valve formation and septation. This strongly indicatesthat at least the 60A subgroup (Bmp5, 6 and 7) is functionallyredundant and that the collective expression of these ligandsdetermines developmental function. In keeping with thisconclusion, individual inactivation of these genes revealsphenotypes that closely correspond with their domains ofunique expression (Dudley et al., 1995; Dudley and Robertson,1997; Kingsley et al., 1992; Solloway et al., 1998). Collectively,these studies strongly suggest that developmental roles of BMPsare determined by the cis-regulatory sequences governing theirexpression rather than via distinct properties of individualligands. Thus, a mechanistic understanding of BMP function indevelopment will require characterization of the basis of tissue-specific expression of these growth factors. Bmp7 is essential fordevelopment of the eye, kidney and hindlimb (Dudley et al.,1995, 1999; Oxburgh et al., 2004). Here we describe for the firsttime the cis-regulatory sequences controlling Bmp7 expressionin these tissues. We have identified an approximately 480 basepair evolutionarily conserved enhancer island within intron 1 ofthe Bmp7 locus governing expression in all three of these tissues.Surprisingly, we find that this enhancer activity is entirelydependent on the presence of a stretch of 10 base pairscontaining a predicted binding site for the transcription factorFOXD3.
Materials and methods
RNA purification, Northern analysis and RACE
RNAwas purified from embryonic day 13.5 (E13.5), E17.5 and adult kidneysusing the Trizol reagent (Invitrogen) according to the manufacturer's instruc-tions. 50 μg total RNA was separated on a 1% formaldehyde denaturing gelalongside an RNA size ladder (ssRNA ladder, New England Biolabs). The gelwas ethidium bromide stained and nucleic acid migration distances calculated.The gel was subsequently blotted to Hybond N nitrocellulose (GE Healthcare)(Sambrook et al., 1989) and hybridized with a 32P-labeled random primed probe(Rediprime, GEHealthcare) representing the entire Bmp7 coding sequence usingstandard Northern blotting procedures (Sambrook et al., 1989). Autoradiographswere measured and compared to the size standards to determine molecularweights of detected bands. Three prime Rapid Amplification of cDNA ends(RACE) was performed on total RNA using the SMART RACE kit (Clontech)according to the manufacturer's instructions. 3′ termini of isolated clones weresequenced and aligned with the genomic sequence of Bmp7 (chromosome 2,172,510,951–172,583,260). Bmp7 cDNA clones isolated from an embryonickidney library (Stratagene) using the Bmp7 coding sequence probe weresequenced, and their 5′ termini compared to the Bmp7 genomic sequence.
Transgenic reporter constructs
A phage clone spanning approximately 20 kilobases (kb) surrounding thefirst exon of Bmp7 was isolated from a 129 SVJ genomic library (Stratagene) byscreening using a 32P-labeled random primed probe representing the first exon ofBmp7 using standard procedures (Sambrook et al., 1989). The genomic clonewas restriction mapped and subcloned into the Hsp68lacZ reporter construct(Sasaki and Hogan, 1996) in five fragments (Fig. 1D). The following restrictionenzymes were used to generate fragments: for 142:1, EcoRI–NsiI, for 216:1NsiI–HindIII, for 216:2 HindIII–NsiI, for 217:1, NsiI–NdeI, for 217:2, NdeI–XbaI. Genomic DNA was digested, separated on 0.8% agarose gels and DNA
was purified from bands of appropriate molecular weights using the GenecleanSpin kit (QBioGene) according to the manufacturer's instructions. PurifiedDNA was polished using T4 DNA polymerase (New England Biolabs)according to the manufacturer's instructions, and enzyme was heat inactivatedfor 20 min at 65 °C before ligation. The 480 base pair (bp) Xenopus tropicalisBmp7 intron 1 element was PCR amplified from genomic DNA using PlatinumHi-Fidelity PCR kit (Invitrogen) with 5′ phosphorylated oligonucleotides 5′-GGCTCGGACG TTCTTGGACG TCTCT-3′ and 5′-AGATCCTTATAATCA-CAACC AGACA-3′. Hsp68lacZ plasmid was linearized with SmaI anddephosphorylated using Calf Intestinal Phosphatase (Both New EnglandBiolabs) according to the manufacturer's instructions. Linearized plasmid wasgel purified using the Geneclean spin kit. Plasmid and insert were ligated usingthe Takara ligation kit (Takara) according to manufacturer's instructions andtransformed into chemically competent DH5α E. coli prepared according tostandard procedures (Sambrook et al., 1989). Ampicillin-resistant colonies werescreened by restriction mapping for the presence of genomic DNA and DNAwas purified from positive clones using the Qiagen Maxiprep kit (Qiagen)according to manufacturer's instructions. Purified plasmid DNA was digestedwith NotI, separated on 0.8% agarose gel, and the anticipated molecular weightcorresponding to the transgene was excised and purified using the Qiagen GelCleanup Kit (Qiagen). Transgene DNA was verified by agarose gel electro-phoresis and suspended to a concentration of 3 ng/μl for pronuclear injection in abuffer containing 10 mM PIPES, 5 mM NaCl and 150 mM KCl.
Subcloning and mutagenesis
Six individual subclones were generated from the 217:1 genomic fragmentby PCR with the Platinum Hi-Fidelity PCR kit (Invitrogen) using the followingoligonucleotide combinations: 5′-GAACATTCTT GCCAAACCAT TCAG-3′and 5′-CGCTATTCTA CGGTGGAAAC AGAC-3′, 5′-TTCCTGTGTTTGGGATGC-3′ and 5′-TTTCTGCTGG TGAGATGAC-3′, 5′-TGTTTCCACCGTAGAATAGC GTC-3′ and 5′-CCATTTTGGT CTCCCAGGTA GTG-3′,5′-TGTGTGTGTG TGTATTACCC ACCG-3′ and 5′-CAACAGTGAC AATG-CTGAGA GACAG-3′, 5′-AGTATTTCTT CCCACCCCTT TCTG-3′ and 5′-AGACACCGCA GGCTGTATGT ATTAC-3′, 5′-CATCAGTAAA GCC-TGGTTGG ATTC-3′ and 5′-TTCCGCAATG TCCCGAAAC-3′. An 5′-TT-GGCGCGCC-3′ sequence containing anAscI site was added at the 5′ end of eachforward amplification oligonucleotide for directional cloning. Similarly, a PCRfragment representing the 480 bp intron 1 enhancer was generated using theoligonucleotide combination 5′-AACATTTGTG TCGGAAGGCA TCGCG-3′and 5′-AGCCCCCAACCCCCCACCCCATAGA-3′. The 217:1 genomic clonewas used as template and PCR products were phosphorylated usingPolynucleotide Kinase and digested with AscI (both New England Biolabs)according to the manufacturer's instructions before ligating into AscI and SmaIlinearized Hsp68lacZ plasmid. Ampicillin resistant clones were verified byrestriction mapping and transgenes were purified.
For deletion mutagenesis, the intron 1 enhancer PCR fragment was polishedusing T4 polymerase and cloned into an EcoRV linearized pBSIIKS plasmid.Orientation of the insert was verified by restriction mapping, and a clonecontaining an insert that could be liberated by AscI and SmaI digestion wasselected. Six individual mutants of this enhancer element were generated byPCR using the following oligonucleotides: I, 5′-AACCCTGGTG TTCGCA-GAGG-3′ and 5′-TCTGGTTCCT GTACCAACAT-3′, II, 5′-CACGTTAAACATGTTGGTAC-3′ and 5′-ATTTCCAAAC CGGAGCCGCT-3′, III, 5′-TTG-GCCGGCC CTTTGAAATA-3′ and 5′-GGATGCCATT GTTAATTTGT-3′, IV,5′-TGGCATCCCA AACACAGGAA-3′ and 5′-TTGTTAATTT GTTCCCAT-GC-3′, V, 5′-TTGGCAGCCC CGGCTCCTGC-3′ and 5′-CAGCCCTCACTCGTGCTCGG-3′, VI, 5′-CCAGAATTAA CTGCAAAGTG-3′ and 5′-TTCCCTGCGA GGAACGGAAG-3′. This strategy results in deletion of: I,40 nt, II, 9 nt, III, 40 nt, IV, 10 nt, V, 36 nt and VI, 28 nt. The 480 nt intron 1enhancer pBSIIKS clone was used as template in each of these reactions. PCRproducts were phosphorylated, ligated, transformed into competent cells andverified by restriction mapping. The inserts of selected clones were sequencedfor confirmation, and AscI/SmaI cloned into the Hsp68lacZ plasmid from whichtransgenes were purified.
For site-directed mutagenesis, the 480 nt intron 1 enhancer pBSIIKS clonewas used as template to replace three base pairs within the putative FOXD3binding sites. Platinum Hi-Fidelity polymerase was used to perform PCR using

681D. Adams et al. / Developmental Biology 311 (2007) 679–690
oligonucleotides 5′-ACCAATTTGT TCCCATGCAG GAG-3′ (mutated bases initalic) and 5′-AATGGCATCC CAAACACAGG AAAGG-3′. PCR productswere DpnI digested to degrade template plasmid DNA according to themanufacturer's instructions (New England Biolabs), and phosphorylated asdescribed above. After gel purification, the PCR product was ligated andtransformed into chemically competent E. coli. Mutagenesis of the putativeFOXD3 binding site and integrity of the sequence were verified by sequencing.Themutagenized fragment was subcloned into the Hsp68lacZ reporter plasmid asdescribed above.
Generation and analysis of transgenic mice
Transgenes were injected into pronuclei of fertilized mouse oocytes of eitherF1CBA/DBAhybrid or SVJmice using standard procedures (Hogan et al., 1994)and injected oocytes were transferred to oviducts of pseudopregnant females ofeither F1 CBA/DBA hybrid or Swiss Webster strains. For transient transgenicanalysis, the day of oviduct transfer was counted as day 0.5 of pregnancy.Transgenic lines were maintained on an ICR background. X-gal staining ofwhole embryos was performed as previously described (Oxburgh et al., 2004).For vibratome sectioning, embryos were prefixed in 1% formaldehyde and 0.2%glutaraldehyde for 60min prior to sectioning. 150 μm sections were cut and fixedfor an additional 30 min before X-gal staining. Stained embryos and vibratomesections were photographed using a stereomicroscope.
For transient transgenic analyses, embryos were harvested for X-galstaining, and DNA was purified from yolk-sacs using an Autogen 850 nucleicacid purification instrument according to the manufacturer's instructions(Autogen Inc). Transgenic embryos were identified by PCR genotyping forthe β-galactosidase cDNA. At least seven positive embryos were analyzed foreach reporter transgene. In most cases, approximately five of these embryosdisplayed X-gal staining. A consensus staining pattern was established bycomparison of embryos, and representative embryos were photographed. In thecase of the reporter transgene containing deletion IV, 15 positive embryos wereanalyzed. For the transgene generated by site-directed mutagenesis, sevenpositive embryos were analyzed.
For establishment of the transgenic strain, staining patterns of embryosderived from four individual founders were compared. Patterns were identical,but strength of staining differed significantly. The strain displaying the strongeststaining was selected for analysis.
Electrophoretic mobility shift assay (EMSA)
Crude nuclear extracts were purified from entire E14.5 kidneys or E16.5limbs by dissociation of tissue in 10mMHEPES, pH 7.9, 1.5mMMgCl2, 10mMKCl and 0.5 mM DTT containing Roche Mini Protease inhibitor (addedaccording to the manufacturer's instructions) in a Dounce homogenizer on ice.After incubation on ice for 20 min, nuclei were sedimented, washed andsolubilized in 10 mM HEPES pH 7.0, 25% glycerol, 400 mM NaCl, 1.5 mMMgCl2, 0.2 mM EDTA, 0.5 mM DTT. 1.5 pmol DNA oligonucleotide duplex ofsequence: 5′-GGATGCCATT GTTAATTTGT TCCCAT-3′ was end-labeledwith 32P by incubation with T4 polynucleotide kinase and γ-32P-ATP accordingto themanufacturer's instructions (NewEnglandBiolabs). After purification overa Sephadex G-25 spin column, labeled oligonucleotide was brought up to 100 μl.2 μl nuclear extract was preincubated for 10 min with either: (i) water, (ii) 1.5pmol unlabeled non-specific scrambled oligonucleotide duplex or (iii) 1.5 pmolunlabeled specific oligonucleotide in Promega binding buffer (Promega Corp) atroom temperature. 1 μl labeled oligonucleotide was added to each reaction andincubated for a further 20min at room temperature. Gel-loading buffer was addedand samples were loaded onto a 4% native polyacrylamide gel and separated at350 V. Gels were subsequently dried and exposed to autoradiography film.
Results
Genomic and transcriptional mapping of the Bmp7 locus
To gain a functional understanding of transcriptionalregulation of the Bmp7 locus (Fig. 1A), and specifically to
ascertain whether transcription might be initiated from an exonupstream of the first annotated exon in the Ensembl database(Birney et al., 2006), we isolated and sequenced cDNA clonesfrom a newborn kidney library. In addition to the previouslypublished initiation site 102 bp upstream of the first AUG, asecond transcriptional initiation site lying a further 270 bpupstream was identified (Fig. 1B). However, no alternate firstexon was identified. In addition to the previously published site255 bp downstream of the stop codon, rapid amplification ofcDNA ends (RACE) identified two alternate transcriptionaltermination sites located 448 bp and 1887 bp downstream of thestop codon, respectively (Fig. 1B). Consistent with this, fourdiscrete mRNA species of 3.8, 3.6, 2.1 and 1.9 kb were detectedby Northern blot analysis (Fig. 1C). All transcripts contain theentire coding sequence, and there is a strong bias in favor oftranscription of isoforms with 1887 bp 3′ untranslated regionsboth in embryonic and adult organs.
Screening for enhancer elements
Comparison of numerous loci indicates that cis-actingelements regulating gene expression are generally concentratedaround sites of transcriptional initiation and termination (Zhanget al., 2007). To identify elements driving developmentalexpression of Bmp7, we therefore analyzed the genomic regionsurrounding the first exon (Figs. 1A, D). Phage clones spanninga 20 kb region approximately 3 kb upstream to 17 kb down-stream of exon 1were isolated from a 129Sv genomic library andsubcloned into the Hsp68lacZpA reporter construct (Sasaki andHogan, 1996). Five reporter constructs spanning the region weregenerated (Fig. 1D). Additionally, the targeting construct used togenerate the Bmp7 reporter mouse by replacement of exon 1with a β-galactosidase cDNA (Godin et al., 1998) was used as atransgene. The genomic construct contains approximately 4 kbof sequence upstream and 3 kb downstream of exon 1. Notably,this reporter (designated T) contains the endogenous Bmp7promoter.
Transient transgenic embryos were harvested at E12.5–E13.5, and X-gal stained. Regulatory sequence surroundingexon 1 of Bmp7 governs developmental expression in neuralcrest (Fig. 1E), kidney (Figs. 1F, G, G'), eye, central nervoussystem, and limb (Figs. 2D, F). Two separate domains of Bmp7expression are seen in the developing kidney, namely in thenephrogenic mesenchyme (Fig. 1F) and collecting ducts (Figs.1G, G'). Cis-regulatory sequence governing nephrogenicmesenchyme expression is located within reporter T but analysisof individual Hsp68lacZpA genomic reporter clones spanningthis entire region failed to identify an element drivingnephrogenic mesenchyme expression, suggesting that expres-sion in this cell population is dependent on cooperation betweentwo distant enhancer elements represented within T but notsmaller subclones. Alternatively, the heterologous Hsp68promoter may be incompatible with cis-regulatory sequencedriving expression in this domain. The 2 kb intron 1 fragmentdesignated 217:1 driving expression in limb, eye and kidneycollecting ducts was chosen for further analysis as it regulatesexpression in the three tissues in which BMP7 signaling is

682 D. Adams et al. / Developmental Biology 311 (2007) 679–690
essential for normal development. This 2 kb region wassubcloned to generate six reporter constructs each containingapproximately 700 to 1200 bp of overlapping sequence for afurther round of transient transgenic analysis (data not shown). A754 bp fragment driving expression in limb, eye and collectingducts of the developing kidney was isolated and designated217XPst.
Developmental expression regulated by the intron 1 enhancer217XPst
Four individual lines of transgenic mice were generatedusing the 217XPst:Hsp68lacZpA (217XPst) reporter transgene.When assayed for β-galactosidase expression at E10 and E12.5,embryos from each of the lines display similar staining patterns,

683D. Adams et al. / Developmental Biology 311 (2007) 679–690
with the exception of variability in signal intensity (supple-mentary data). Representative data are shown. To make accuratecomparisons between expression domains governed by theintron 1 enhancer and those governed by the endogenous Bmp7gene, comparable X-gal stained embryos from the Bmp7lacZreporter mouse were analyzed. Interestingly, the 754 bp intron 1regulatory island appears to determine a significant proportionof endogenous Bmp7 expression. At E8, expression is localizeddiffusely in an area of the tailbud surrounding the Bmp7expressing node (Figs. 2A, B). At E9.5, comparison of 217XPstwith Bmp7 reveals expression in the midbrain hindbrainjunction or isthmus, Wolffian duct and limb mesenchyme(Figs. 2C, D). Weak staining can also be seen in themyocardium of the developing heart (Figs. 2C, D). Althoughstrong expression can also be seen in the forebrain of reportermice, it is localized to the ventral rather than the Bmp7expressing dorsal aspect (Figs. 2C, D). This pattern is seenconsistently in separate lines of 217XPst reporter mice, andindicates that the Bmp7 forebrain enhancer may be onlypartially represented in fragment 217XPst. At E12.5, transgeneexpression can be seen in the collecting duct system (Figs. 3A,B), the iris (Figs. 3C, D) and the limb mesenchyme (Figs. 3E,F). Interestingly, comparison with expression of Bmp7 showsthat the intron 1 enhancer drives expression in discretesubdomains within these developing organs.
Identification of a 480 base pair conserved enhancer elementin intron 1
It is surprising that expression of Bmp7 at so many distinctanatomical sites and phases in development is determined bya single enhancer element. To inquire whether all expressiondriven by this regulatory element might be determined by asingle key transcription factor binding site, or whetherdifferent transcription factor binding sites govern expressionat distinct locations, we next analyzed the primary sequenceof the 217XPst regulatory region. Sequence comparisonbetween 217XPst fragment and Bmp7 loci from chick,human, Xenopus tropicalis, Takifugu rubripes and Daniorerio identified a 480 bp sequence with extensive homologybetween mouse, chick, human and frog (Fig. 4). In contrast,this sequence was absent in fish (data not shown). Sequencehomology searches of entire fish genomes with the Bmp7intronic island also failed to reveal any significant conserva-
Fig. 1. Genomic organization of the murine Bmp7 locus and screening strategy usedexons distributed over approximately 72 kb on mouse chromosome 2. Exon–intron sUTR and encoding mRNA for the prodomain of BMP7 and red boxes corresponding tat top right represents 2.5 kb. (B) Predicted structures of Bmp7 transcripts based on sein addition to the previously published initiation site 102 bp upstream of the first AUGthe AUG (2.1 and 3.8 kb transcripts). Additionally, two 3′ termini were identified: onanother 1887 bp downstream from the UAG (3.6 and 3.8 kb transcripts). Red and bisolated from E13.5, E17.5 and adult kidneys (13.5, 17.5 and ad respectively), and sucoding sequence. Four RNA species were detected; 1.9 kb, 2.1 kb, 3.6 kb and 3.8 kb.distinct RNA transcripts. (D) Strategy used to screen for enhancer elements driving Bmexon 1 (bracketed in panel A) is shown, and lines below denote sizes of genomic subcleach of these subclones was assayed using the Hsp68lacZpA reporter in transgenic216:2 genomic subclone drives expression in neural crest (NC) adjacent to the neuradriven by the T fragment. (G, G') Expression in collecting ducts (CD) of the kidney
tion (data not shown). Thus, this regulatory island appearsfirst in the amphibian Bmp7 homolog and is stronglyconserved through evolution of vertebrates. Predicted tran-scription factor binding sites for OCT-1, AGL-3 and FOXD3are located within this conserved region.
The identification of this conserved 480 bp island within217XPst raises the question of whether it accounts for theβ-galactosidase expression pattern seen using the 217XPstreporter, and if so to what degree this regulatory islanddisplays functional conservation through evolution. To answerthese questions we generated two reporter transgenes contain-ing either the conserved 480 bp region from the mouse or theX. tropicalis Bmp7 locus. The mouse reporter transgenedirected expression in the embryonic eye, limb, Wolffian ductand forebrain comparably with 217XPst, but did not directexpression at the heart (Fig. 4B). Interestingly, the X.tropicalis reporter transgene also directed expression at thesame sites with the exception of the Wolffian duct (Fig. 4C).The exact localization of expression within limb mesenchymediffers between Xenopus and mouse, and we thus concludethat the 480 bp element located in intron 1 is a partiallyconserved enhancer driving Bmp7 expression at key sitessuch as eye and limb mesenchyme.
Homology predictions within the conserved intronic enhancerelement
To begin to ascertain the basis of transcriptional regulationby this enhancer, we identified putative transcription factorbinding sites by sequence comparison. C. elegans has only asingle Bmp (Savage-Dunn, 2001) in contrast to the human,with over 10 closely related ligands (Reddi, 2005). Ourworking hypothesis is that Bmps have undergone duplicationevents, retaining a conserved signaling function but acquiringadditional novel domains of expression. Considering thatother Bmp family members are co-expressed with Bmp7 inthe collecting duct system of the kidney (Bmp3), the eye(Bmp4) and limb mesenchyme (Bmp5), an interestingpossibility is that the Bmp7 intronic enhancer might bepartially conserved from an ancestral Bmp gene. To explorethis idea, we undertook exhaustive sequence alignmentsbetween the Bmp7 intronic enhancer and the entire loci ofall annotated mouse Bmp genes as well as members of thegrowth and differentiation factor (Gdf) family that function
to identify enhancer elements. (A) The Bmp7 gene is comprised of 7 discretetructure is depicted, with black boxes corresponding to exons comprising the 5′o exons comprising the 3′UTR and mRNA for the mature domain of BMP7. Barquencing of newborn kidney cDNA and 3′ RACE of embryonic kidney mRNA:(1.9 and 3.6 kb transcripts), an initiation site was identified 372 bp upstream of
e 448 bp downstream from the UAG stop codon (1.9 and 2.1 kb transcripts), andlack regions correspond to red and black exons in panel A. (C) Total RNAwasbjected to Northern hybridization with a probe corresponding to the entire Bmp7Alternate initiation and termination sites shown in panel B account for these fourp7 expression in the developing kidney. The genomic region surrounding Bmp7ones 142:1, 216:1, 216:2, 217:1 and 217:2. The presence of enhancer elements inmice. Reporter constructs drive expression at diverse anatomical sites. (E) Thel tube (NT). (F) Expression in nephrogenic mesenchyme (NM) of the kidney isis driven by the 217:1 fragment.

Fig. 2. The intron 1 enhancer element drives expression in a significant subset of Bmp7 expression domains: Comparison of Bmp7 expression driven from theendogenous Bmp7 locus with expression driven from the intron 1 enhancer element. Heterozygous Bmp7lacZ reporter mice (Bmp7+/lacZ) and 217XPst (intron 1enhancer) embryos were dissected at E8, E9.5 and E10.5 and whole-mount X-gal stained. (A) At E8, endogenous Bmp7 expression is seen in head-folds (HF), heart(HT) and notochord (NT), with an expanded domain in the caudal notochord corresponding to the position of the node (ND). (B) The intron 1 enhancer element drivesexpression mainly in a diffuse region of the tail bud (TB) surrounding the node and also weakly in the heart. (C) At E9.5, endogenous Bmp7 expression is seen in heart,dorsal forebrain (DFB), isthmus or midbrain–hindbrain junction (IST), optic placode (OPT), otic placode (OTP), Rathke's pouch (RP), limb bud (LB) and Wolffianduct (WD). (D) The intron 1 enhancer element drives expression in a subset of these domains: isthmus, limb bud andWolffian duct. Weak expression can be seen in theheart and ectopic expression is seen in the ventral forebrain (VFB). (E) At E10.5, additional domains of endogenous Bmp7 expression are apparent in mesonephrictubules (MN) and ureteric bud (UB). (F) These domains are represented in the intron 1 enhancer, where an additional ectopic domain of expression can be seen in thedorsal root ganglion (DRG).
684 D. Adams et al. / Developmental Biology 311 (2007) 679–690
via activation of the Smad 1, 5 and 8 effector pathway. Fourregions of 90–100% homology, each spanning approximately25 bp, were identified (Fig. 5A). We also included the threeputative transcription factor binding sites identified byhomology searching using stringent criteria in the Transfacdatabase. Using this rationale, we identified 6 potentialtranscription factor binding sites, designated I–VI.
Analysis of selected deletions within the intron 1 enhancer
To ascertain whether any one of the 6 potential transcriptionfactor binding sites identified in silico is essential for develop-mental expression governed by the 480 bp Bmp7 intron 1enhancer, we generated a series of deletion constructs in whicheach one of these regions was removed in isolation (Fig. 5A). Our

Fig. 3. The intron 1 enhancer element drives expression in specific domains of the eye, limb and kidney. (A) Bmp7 is expressed in both the collecting duct system (CD)and nephrogenic mesenchyme (NM) of the developing kidney. (B) The intron 1 enhancer drives expression only in collecting ducts. (C) In the eye, Bmp7 is expressedin the pigmented epithelium (PE) and nascent iris (IR). (D) The intron 1 enhancer drives expression solely in the nascent iris of the eye. (E) Bmp7 is expressed in theapical ectodermal ridge (AER), limb mesenchyme (LM) and precartilage condensates (PC) of the developing limb. (F) Intron 1 enhancer driven expression is limited tolimb mesenchyme.
685D. Adams et al. / Developmental Biology 311 (2007) 679–690
panel of 6 Hsp68lacZ reporter deletion constructs was then testedin transgenic embryos. Embryos were dissected at E10.5 andstained for β-galactosidase expression. E10.5 was chosen for thisanalysis because the majority of domains of Bmp7 expressionrelevant to this study are evident at this time-point and whole-mount X-gal staining is robust. Since Bmp7 expression in the eyeis only initiated at or around this time-point, we focused ouranalysis on Wolffian duct, ureteric bud, limb and isthmus. Whilesome variability can be seen in amplitude of expression anddegree of ectopic expression between the different constructs,
only the 10 bp deletion spanning the core of the predicted FOXD3binding site abrogates expression (Fig. 5E). Thus, we concludethat the putative FOXD3 binding site within the conserved 480 bpenhancer is absolutely necessary for expression in the limb,isthmus and Wolffian duct.
Site-directed mutagenesis of the putative FOXD3 binding site
To verify that the loss of expression caused by deletion of theputative FOXD3 binding site from the intron 1 enhancer was

Fig. 4. The Bmp7 intron 1 enhancer is strongly conserved from human to frog genomes, but is not represented in the fish. (A) Alignment of Homo sapiens (hsap),Musmusculus (mmus), Gallus gallus (ggal) and Xenopus tropicalis (xtrop) Bmp7 loci identifies a highly homologous 480 bp region located within the murine intron 1enhancer. This region is not represented in either zebrafish or fugu genomes. Three putative transcription factor binding sites conserved across all species werepredicted using the Transfac database: OCT1, AGL3 and FOXD3. (B) The 480 bp region from mouse recapitulates expression in eye (E), limb (L) and Wolffian duct(WD) seen in 217XPst at E10.5. (C) The 480 base pair region from X. tropicalis intron 1 drives Hsp68 reporter gene expression in embryonic eye (E) and limb (L), butnot Wolffian duct (WD) at E11.5.
686 D. Adams et al. / Developmental Biology 311 (2007) 679–690
due to abrogated transcription factor binding at this specific siterather than loss of transcription factor binding secondary toalterations in DNA structure or changes in spacing betweenenhancer elements, we performed site-directed mutagenesis ofthree base pairs within the core of the putative FOXD3 bindingsite (Fig. 6A). Embryos transgenic for a Hsp68lacZ reportercontaining the 480 bp mutated Bmp7 intron 1 sequence did notshow any X-gal staining in limb, isthmus or Wolffian duct (Fig.6B), demonstrating that the putative FOXD3 binding site isindeed required for Bmp7 intron 1 driven expression in thesetissues.
Nuclear factor binding to the predicted FOXD3 binding site
To conclusively determine that nuclear proteins in thedeveloping limb and kidney indeed do associate with the
putative FOXD3 binding site in the conserved enhancer, weperformed an electrophoretic mobility shift assay. A radio-labeled oligonucleotide spanning the predicted FOXD3 bindingsite in intron 1 (Fig. 6C) was used to assay whether crude nuclearextracts of E14.5 kidney and E16.5 limb could retard migrationof this probe in a native polyacrylamide gel. To ensurespecificity of nuclear factor binding, extracts were preincubatedeither with non-specific unlabeled oligonucleotide or unlabeledoligonucleotide identical to the probe. As shown in Fig. 6D, theprobe is retarded by extracts from both embryonic kidney andlimb, and this is competed by addition of unlabeled probeoligonucleotide, demonstrating that this interaction is specific.Thus, nuclear factors expressed in embryonic limb and kidneybind to the predicted FOXD3 binding site required fordevelopmental expression ofBmp7 in these tissues.We concludethat the putative FOXD3 binding site within the conserved intron

Fig. 5. Deletion mutagenesis to localize the transcription factor binding site(s) driving Bmp7 expression in limb mesenchyme (LM), Wolffian duct (WD) and isthmus(IST). (A) Schematic of the Bmp7 intron 1 enhancer element showing the two different strategies used to identify putative transcription factor binding sites: (i) regionsof strong homology with intronic sequence in other TGFβ superfamily ligand genes (red bars), (ii) Transfac-predicted binding sites conserved in all species (blackmarks). Reporter genes I–VI containing deletions spanning single regions were used to generate transgenic animals which were analyzed at E10.5 for expression inisthmus (IST), limb mesenchyme (LM) and Wolffian duct (WD). (B–G) Only deletion IV spanning the predicted FOXD3 binding site significantly affects expressionin the three domains assayed.
687D. Adams et al. / Developmental Biology 311 (2007) 679–690
1 enhancer element of Bmp7 directly binds one or multipletranscription factors that enhance transcription specifically in thelimb and Wolffian duct of the developing embryo.
Discussion
Bmp7 is expressed in numerous tissues throughout develop-ment and is required for morphogenesis of the eye, hindlimb andkidney (Dudley et al., 1995; Godin et al., 1998). In this study, wehave shown that the majority if not all of the cis-regulatorysequence regulating Bmp7 expression at these anatomical sites iscontained within approximately 20 kb of genomic sequence
surrounding exon 1. In each of these organs, multiple distinctenhancer elements drive Bmp7 expression. In the eye, we showthat elements driving expression in the pigmented epitheliumand iris are spatially separated. In the kidney, Bmp7 expressionin collecting ducts and nephron progenitors is driven by separateenhancer elements. Similarly in the limb, mesenchyme andapical ectodermal ridge (AER) expression are governed byseparate elements. Although enhancers for pigmented epithe-lium, nephrogenic mesenchyme and AER are distributed acrossapproximately 20 kb, an element of approximately 480 bpwithinintron 1 governs expression within the developing iris, kidneycollecting duct system and limb mesenchyme.
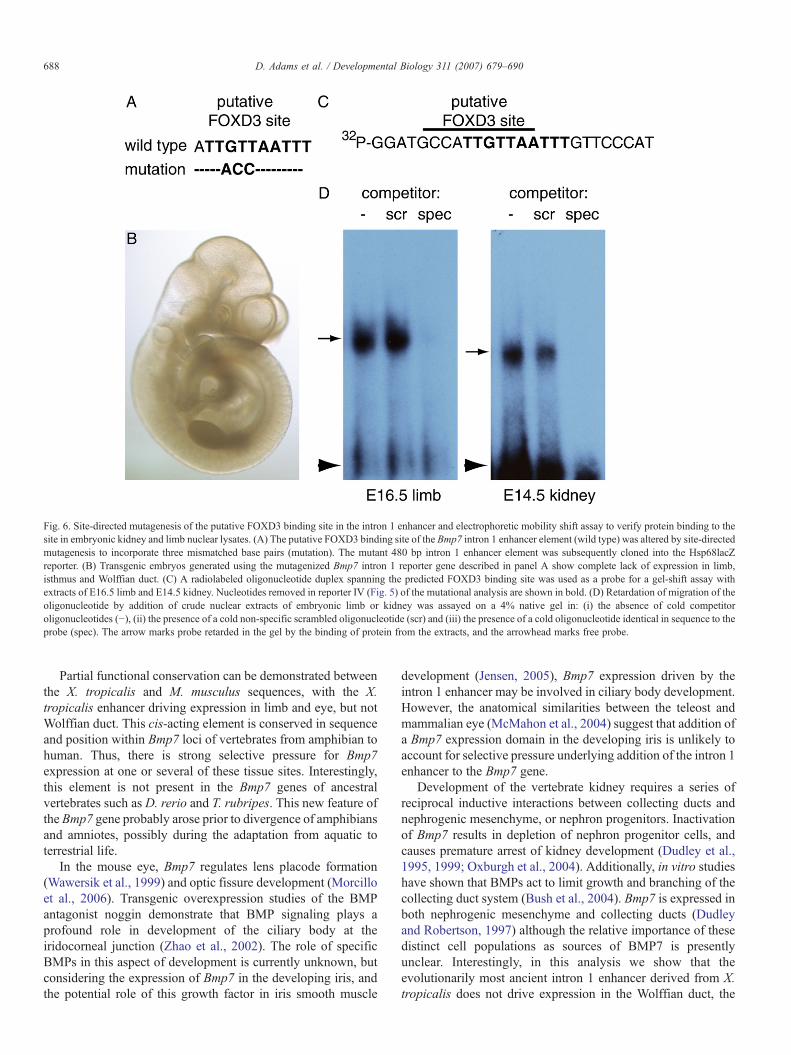
Fig. 6. Site-directed mutagenesis of the putative FOXD3 binding site in the intron 1 enhancer and electrophoretic mobility shift assay to verify protein binding to thesite in embryonic kidney and limb nuclear lysates. (A) The putative FOXD3 binding site of the Bmp7 intron 1 enhancer element (wild type) was altered by site-directedmutagenesis to incorporate three mismatched base pairs (mutation). The mutant 480 bp intron 1 enhancer element was subsequently cloned into the Hsp68lacZreporter. (B) Transgenic embryos generated using the mutagenized Bmp7 intron 1 reporter gene described in panel A show complete lack of expression in limb,isthmus and Wolffian duct. (C) A radiolabeled oligonucleotide duplex spanning the predicted FOXD3 binding site was used as a probe for a gel-shift assay withextracts of E16.5 limb and E14.5 kidney. Nucleotides removed in reporter IV (Fig. 5) of the mutational analysis are shown in bold. (D) Retardation of migration of theoligonucleotide by addition of crude nuclear extracts of embryonic limb or kidney was assayed on a 4% native gel in: (i) the absence of cold competitoroligonucleotides (−), (ii) the presence of a cold non-specific scrambled oligonucleotide (scr) and (iii) the presence of a cold oligonucleotide identical in sequence to theprobe (spec). The arrow marks probe retarded in the gel by the binding of protein from the extracts, and the arrowhead marks free probe.
688 D. Adams et al. / Developmental Biology 311 (2007) 679–690
Partial functional conservation can be demonstrated betweenthe X. tropicalis and M. musculus sequences, with the X.tropicalis enhancer driving expression in limb and eye, but notWolffian duct. This cis-acting element is conserved in sequenceand position within Bmp7 loci of vertebrates from amphibian tohuman. Thus, there is strong selective pressure for Bmp7expression at one or several of these tissue sites. Interestingly,this element is not present in the Bmp7 genes of ancestralvertebrates such as D. rerio and T. rubripes. This new feature ofthe Bmp7 gene probably arose prior to divergence of amphibiansand amniotes, possibly during the adaptation from aquatic toterrestrial life.
In the mouse eye, Bmp7 regulates lens placode formation(Wawersik et al., 1999) and optic fissure development (Morcilloet al., 2006). Transgenic overexpression studies of the BMPantagonist noggin demonstrate that BMP signaling plays aprofound role in development of the ciliary body at theiridocorneal junction (Zhao et al., 2002). The role of specificBMPs in this aspect of development is currently unknown, butconsidering the expression of Bmp7 in the developing iris, andthe potential role of this growth factor in iris smooth muscle
development (Jensen, 2005), Bmp7 expression driven by theintron 1 enhancer may be involved in ciliary body development.However, the anatomical similarities between the teleost andmammalian eye (McMahon et al., 2004) suggest that addition ofa Bmp7 expression domain in the developing iris is unlikely toaccount for selective pressure underlying addition of the intron 1enhancer to the Bmp7 gene.
Development of the vertebrate kidney requires a series ofreciprocal inductive interactions between collecting ducts andnephrogenic mesenchyme, or nephron progenitors. Inactivationof Bmp7 results in depletion of nephron progenitor cells, andcauses premature arrest of kidney development (Dudley et al.,1995, 1999; Oxburgh et al., 2004). Additionally, in vitro studieshave shown that BMPs act to limit growth and branching of thecollecting duct system (Bush et al., 2004). Bmp7 is expressed inboth nephrogenic mesenchyme and collecting ducts (Dudleyand Robertson, 1997) although the relative importance of thesedistinct cell populations as sources of BMP7 is presentlyunclear. Interestingly, in this analysis we show that theevolutionarily most ancient intron 1 enhancer derived from X.tropicalis does not drive expression in the Wolffian duct, the

689D. Adams et al. / Developmental Biology 311 (2007) 679–690
epithelial tubule in the intermediate mesoderm that gives rise tothe permanent kidney. Thus, addition of the intron 1 enhancercannot simply be explained by selective pressures modifyingBmp7 expression domains in the kidney collecting duct.
Polydactyly is seen in approximately 65% of Bmp7 nullembryos (Dudley et al., 1995). Studies of compound mutantsdemonstrate that Bmp7 cooperates with Bmp4 to regulate digitpatterning (Katagiri et al., 1998). Incomplete penetrance of thepolydactyly phenotype is probably due to genetic redundancy.Conditional inactivation of Bmp4 in limb mesenchymesimilarly results in polydactyly due to delayed induction ofthe apical ectodermal ridge (AER) and increased SonicHedgehog (SHH) signaling from the zone of polarizing activity(ZPA) (Selever et al., 2004). BMP signals from limbmesenchyme thus act indirectly to restrict the range of SHHsecreted from the ZPA and limit the formation of anterior digits.We show here that the Bmp7 intron 1 enhancer element drivesexpression strongly in limb mesenchyme, and thus is likely toregulate the digit patterning activity of Bmp7 essential tonormal development. Interestingly, molecular and morphologi-cal studies fail to identify a structure homologous to the tetrapodautopod in the teleost (Ahlberg and Milner, 1994; Sordino et al.,1995). The fossil record shows that digit patterning was afeature of early tetrapods such as Acanthostega and Icthyos-tega, but that the number of digits per autopod was eight(Ahlberg and Milner, 1994). The digit number of the autopodwas subsequently reduced to five in the common ancestor ofamphibians and amniotes. An intriguing possibility is thatadaptation of the autopod from an octa- to pentadactylmorphology more suitable for terrestrial life selected for anovel Bmp7 expression domain in limb mesenchyme driven bythe intron 1 enhancer.
The transcriptional network driving Bmp7 expression ispoorly understood. Through mutational analysis, we haveidentified a FOXD3 transcription factor binding site essentialfor expression in kidney collecting duct and limb mesenchyme.The likelihood that FOXD3 is responsible for activating Bmp7in these developing tissues is small since it is expressedexclusively in the neural crest (Dottori et al., 2001). At presentwe have not identified alternate candidate factors. However,comparison of expression patterns governed by intron 1enhancer elements in different species may yield importantclues in answering this question. For example, in this study wefound that the X. tropicalis enhancer drives expression only ineye and limb whereas the mouse enhancer drives expression inthe Wolffian duct in addition to these locations. Paradoxically,the latter expression patterns appear to be driven by a singleFOXD3 transcription factor binding sites, posing the questionas to how the FOXD3 binding site derived from the X. tropicalisenhancer differs in transcriptional activation from the mouseenhancer when placed into the context of the mouse genome.Sequence comparison in this region shows that these speciesdiffer by two base pairs indicating that transcription factorbinding may differ between them. Close examination of thepattern of limb mesenchyme expression driven by the X.tropicalis enhancer reveals that the expression domain onlypartially overlaps with that of the mouse. Thus the differences in
expression between mouse and Xenopus intron 1 enhancersmay be explained by the fact that multiple FOXD-liketranscription factors with subtly different binding specificitiesassociate with this cis-regulatory sequence in limb mesenchymeand Wolffian duct. Future comparative studies of these Bmp7enhancer elements will address the identities of these factors,expanding our understanding of the transcriptional circuitryregulating developmental expression of Bmp7.
Acknowledgments
Many thanks to the Mouse Transgenic and MRI CoreFacility at Maine Medical Center Research Institute forpronuclear injections and to the Bioinformatics and GenomicsCore for DNA purification. Thanks to Dr. Andrew T. Dudley forhelp in isolating the phage clone used in this study and Drs.Dominic Norris and Elizabeth Bikoff for helpful comments onthe manuscript. The project described was supported by GrantNumber P20 RR18789 to L. O. from the National Center forResearch Resources (NCRR), a component of the NationalInstitutes of Health (NIH) and R01 HD34435 to E. J. R. fromthe NIH. The contents of this paper are solely the responsibilityof the authors and do not necessarily represent the official viewsof the NIH.
Appendix A. Supplementary data
Supplementary data associated with this article can be found,in the online version, at doi:10.1016/j.ydbio.2007.08.036.
References
Ahlberg, P.E., Milner, A.R., 1994. The origin and early diversification oftetrapods. Nature 368, 507–514.
Birney, E., et al., 2006. Ensembl 2006. Nucleic Acids Res. 34, D556–D561.Bush, K.T., et al., 2004. TGF-[beta] superfamily members modulate growth,
branching, shaping, and patterning of the ureteric bud. Dev. Biol. 266,285–298.
Dottori, M., et al., 2001. The winged-helix transcription factor Foxd3 suppressesinterneuron differentiation and promotes neural crest cell fate. Development128, 4127–4138.
Dudley, A.T., Robertson, E.J., 1997. Overlapping expression domains of bonemorphogenetic protein family members potentially account for limited tissuedefects in BMP7 deficient embryos. Dev. Dyn. 208, 349–362.
Dudley, A.T., et al., 1995. A requirement for bone morphogenetic protein-7during development of the mammalian kidney and eye. Genes Dev. 9,2795–2807.
Dudley, A.T., et al., 1999. Interaction between FGF and BMP signalingpathways regulates development of metanephric mesenchyme. Genes Dev.13, 1601–1613.
Godin, R.E., et al., 1998. Regulation of BMP7 expression during kidneydevelopment. Development 125, 3473–3482.
Hogan, B.L.M., 1996. Bone morphogenetic proteins: multifunctional regulatorsof vertebrate development. Genes Dev. 10, 1580–1594.
Hogan, B.L., et al., 1994. Manipulating the Mouse Embryo: A LaboratoryManual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
Jensen, A.M., 2005. Potential roles for BMP and Pax genes in the developmentof iris smooth muscle. Dev. Dyn. 232, 385–392.
Katagiri, T., et al., 1998. Skeletal abnormalities in doubly heterozygous Bmp4and Bmp7 mice. Dev. Genet. 22, 340–348.
Kim, R.Y., et al., 2001. Bmp6 and Bmp7 are required for cushion formation andseptation in the developing mouse heart. Dev. Biol. 235, 449–466.

690 D. Adams et al. / Developmental Biology 311 (2007) 679–690
Kingsley, D.M., et al., 1992. The mouse short ear skeletal morphogenesis locusis associated with defects in a bone morphogenetic member of the TGFβSuperfamily. Cell 71, 399–410.
Massagué, J., 1998. TGF-beta signal transduction. Annu. Rev. Biochem. 67,753–791.
Massagué, J., Wotton, D., 2000. Transcriptional control by the TGF-beta/Smadsignaling system. EMBO J. 19, 1745–1754.
McMahon, C., et al., 2004. Using zebrafish to study the complex geneticsof glaucoma. Comp. Biochem. Physiol., Part C Pharmacol. Toxicol. 138,343–350.
Morcillo, J., et al., 2006. Proper patterning of the optic fissure requires thesequential activity of BMP7 and SHH. Development 133, 3179–3190.
Oxburgh, L., et al., 2004. TGFb superfamily signals are required formorphogenesis of the kidney mesenchyme progenitor population. Develop-ment 131, 4593–4605.
Oxburgh, L., et al., 2005. BMP4 substitutes for loss of BMP7 during kidneydevelopment. Dev. Biol. 286, 637–646.
Reddi, A.H., 2005. BMPs: from bone morphogenetic proteins to bodymorphogenetic proteins. Cytokine Growth Factor Rev. 16, 249–250.
Sambrook, J., et al., 1989. Molecular Cloning: A Laboratory Manual. ColdSpring Harbor Laboratory Press.
Sasaki, H., Hogan, B.L., 1996. Enhancer analysis of the mouse HNF-3 betagene: regulatory elements for node/notochord and floor plate areindependent and consist of multiple sub-elements. Genes Cells 1, 59–72.
Savage-Dunn, C., 2001. Targets of TGF beta-related signaling in Caenorhab-ditis elegans. Cytokine Growth Factor Rev. 12, 305–312.
Selever, J., et al., 2004. Bmp4 in limb bud mesoderm regulates digit pattern bycontrolling AER development. Dev. Biol. 276, 268–279.
Solloway, M.J., Robertson, E.J., 1999. Early embryonic lethality in Bmp5;Bmp7 double mutant mice suggests functional redundancy within the 60Asubgroup. Development 126, 1753–1768.
Solloway, M.J., et al., 1998. Mice lacking Bmp6 function. Dev. Genet. 22,321–339.
Sordino, P., et al., 1995. Hox gene expression in teleost fins and the origin ofvertebrate digits. Nature 375, 678–681.
Wawersik, S., et al., 1999. BMP7 acts in murine lens placode development.Dev. Biol. 207, 176–188.
Zhang, Z.D., et al., 2007. Statistical analysis of the genomic distribution andcorrelation of regulatory elements in the ENCODE regions. Genome Res.17, 787–797.
Zhao, S., et al., 2002. BMP signaling is required for development of the ciliarybody. Development 129, 4435–4442.





























