Embed Size (px)
Citation preview

RESEARCH ARTICLES
Convergent Evolution of Syringyl Lignin Biosynthesis viaDistinct Pathways in the Lycophyte Selaginella andFlowering Plants C W
Jing-Ke Weng,a,1 Takuya Akiyama,b Nicholas D. Bonawitz,a, Xu Li,a John Ralph,b,c and Clint Chapplea,2
a Department of Biochemistry, Purdue University, West Lafayette, Indiana 47907b U.S. Dairy Forage Research Center, U.S. Department of Agriculture–Agricultural Research Service, Madison, Wisconsin 53706c Department of Biochemistry (Enzyme Institute) and Department of Energy Great Lakes Bioenergy Research Center, University
of Wisconsin, Madison, Wisconsin 53726
Phenotypic convergence in unrelated lineages arises when different organisms adapt similarly under comparable selective
pressures. In an apparent example of this process, syringyl lignin, a fundamental building block of plant cell walls, occurs in
two major plant lineages, lycophytes and angiosperms, which diverged from one another more than 400 million years ago.
Here, we show that this convergence resulted from independent recruitment of lignin biosynthetic cytochrome P450-
dependent monooxygenases that route cell wall monomers through related but distinct pathways in the two lineages. In
contrast with angiosperms, in which syringyl lignin biosynthesis requires two phenylpropanoid meta-hydroxylases C39H and
F5H, the lycophyte Selaginella employs one phenylpropanoid dual meta-hydroxylase to bypass several steps of the
canonical lignin biosynthetic pathway. Transgenic expression of the Selaginella hydroxylase in Arabidopsis thaliana
dramatically reroutes its endogenous lignin biosynthetic pathway, yielding a novel lignin composition not previously
identified in nature. Our findings demonstrate a unique case of convergent evolution via distinct biochemical strategies and
suggest a new way to genetically reconstruct lignin biosynthesis in higher plants.
INTRODUCTION
In response to environmental pressures, phylogenetically unre-
lated species sometimes arrive at similar adaptive solutions
through independent mechanisms (Tanaka et al., 2009). Al-
though convergent evolution happens rarely in nature, it repre-
sents an important evolutionary phenomenon, which has been
documented in action at multiple levels of biological processes
(Conant and Wagner, 2003). For example, birds and bats inde-
pendently evolved wings for powered flight (Hedenstrom et al.,
2007), both higher plants and fungi developed the ability to
synthesize growth regulator gibberellins via nonorthologous
pathways (Hedden et al., 2001), and mammals and fungi re-
cruited two distinct families of proteins to methylate Lys residues
on histone tails (Cheng et al., 2005). Elucidation of the molecular
mechanisms underlying phenotypic convergence is key to un-
derstanding how this process contributes to evolution.
Vascular plants arose in the Late Silurian period (;420 million
years ago). They diversified rapidly during the Early Devonian
period (416 to 398 million years ago), when an early split in the
history of land plant evolution gave rise to twomajor lineages: the
lycophytes andeuphyllophytes (Kenrick andCrane, 1997) (Figure
1A). These two lineages are united as a monophyletic group by
the presence of specialized water-conducting tracheary ele-
ments, the cell walls of which are physically reinforced by the
phenolic lignin heteropolymer (Kenrick and Crane, 1997). Lignin
endowsvascular plantswith the rigidity to standupright, prevents
their tracheids and vessel elements from collapsing during long-
distance water transport, and has greatly contributed to the
dominance of these plants in terrestrial environments (Boerjan
et al., 2003). Although lignin appears to be fundamental to the
biochemistry of all vascular plants, its monomer composition
exhibits an intriguing distribution pattern across the major line-
ages. Among the euphyllophytes, ferns and gymnosperms gen-
erally contain p-hydroxyphenyl (H) and guaiacyl (G) lignin units,
derived from p-coumaryl alcohol and coniferyl alcohol, respec-
tively, and angiosperms additionally contain syringyl (S) units
derived from sinapyl alcohol (Boerjan et al., 2003) (Figure 1). This
observation has led to the notion that S lignin is a recent invention
restricted primarily to angiosperms. In fact, S lignin has also been
found in some lycophyte species belonging to the order of
Selaginellales but is absent from species belonging to its extant
sister order Lycopodiales and is thought to have been absent
from its extinct sister order Lepidodendrales, suggesting that
S lignin might have arisen independently in lycophytes and
1Current address: Howard Hughes Medical Institute, Jack H. SkirballCenter for Chemical Biology and Proteomics, The Salk Institute forBiological Studies, La Jolla, CA 92037.2 Address correspondence to [email protected] author responsible for distribution of materials integral to thefindings presented in this article in accordance with the policy describedin the Instructions for Authors (www.plantcell.org) is: Clint Chapple([email protected]).CSome figures in this article are displayed in color online but in blackand white in the print edition.WOnline version contains Web-only data.www.plantcell.org/cgi/doi/10.1105/tpc.109.073528
The Plant Cell, Vol. 22: 1033–1045, April 2010, www.plantcell.org ã 2010 American Society of Plant Biologists

euphyllophytes by evolutionary convergence (Towers and
Gibbs, 1953; White and Towers, 1967; Logan and Thomas,
1985, 1987; Jin et al., 2005; Weng et al., 2008c) (Figure 1A).
Lignin monomer biosynthesis in angiosperms is largely de-
pendent on the expression of three cytochrome P450 monoox-
ygenases (P450s; Figure 1B) (Boerjan et al., 2003). Cinnamate
4-hydroxylase (C4H) catalyzes the phenylpropanoid ring para-
hydroxylation reaction that is required for the formation of all
three types of lignin units (Teutsch et al., 1993), but two distinct
P450s carry out each of the two subsequent hydroxylations
required for S lignin synthesis. p-Coumaroyl shikimate 39-hydroxylase (C39H) catalyzes the first ring meta-hydroxylation
reaction, necessary for the biosynthesis of both G and S lignin
units (Franke et al., 2002a), and ferulate 5-hydroxylase (F5H)
catalyzes the second ringmeta-hydroxylation reaction leading to
the biosynthesis of S lignin units from G-substituted precursors
(Meyer et al., 1998).
We recently isolated from the lycophyte Selaginella moellen-
dorffii a novel P450 (CYP788A1) that, like angiosperm F5Hs, can
divert G-substituted intermediates toward S lignin synthesis
(Weng et al., 2008c). Selaginella F5H (Sm F5H) shares only 37%
amino acid sequence identity with its angiosperm counterparts, a
level of similarity that can be expected from any of two random
plant P450 enzymes from families with unrelated functions,
suggesting that the similar catalytic activities of F5Hs in the two
lineages were derived from convergent evolution (Weng et al.,
2008c). Here, we report the discovery of a novel activity of SmF5H
that reveals an unexpected metabolic distinction between Selag-
inella and angiosperms and suggests that S lignin is elaborated via
distinct biosynthetic routes in these two groups of plants.
Figure 1. S Lignin Biosynthesis via Distinct Biosynthetic Routes in Angiosperms and the Lycophyte Selaginella.
(A) Plant phylogenetic tree showing the distribution of lignin monomer composition across major land plant lineages. Open circle, no lignin; yellow circle,
presence of H and G lignin; red circle, presence of S lignin in addition to H and G lignin. †Lignin composition indicated by the analysis of fossil samples of
extinct Sigillaria species (Logan and Thomas, 1987).
(B) Alternative pathways toward S lignin biosynthesis in Selaginella and angiosperms. The shared routes between Selaginella and angiosperms are
shaded in purple, whereas the divergent branches specific to Selaginella or angiosperms are shaded in pink or light blue, respectively. The two aromatic
ring meta-hydroxylation steps are catalyzed by one bifunctional enzyme, Sm F5H (circled in red) in Selaginella, instead of two enzymes, C39H and F5H
(circled in blue), in angiosperms. At, Arabidopsis thaliana; Sm, Selaginella moellendorffii. PAL, phenylalanine ammonia-lyase; 4CL, 4-hydroxycinnamoyl-
CoA ligase; CCoAOMT, caffeoyl-CoA O-methyl transferase; HCALDH, hydroxycinnamaldehyde dehydrogenase; COMT, caffeic acid O-methyltrans-
ferase; CAD, (hydroxy)cinnamyl alcohol dehydrogenase.
1034 The Plant Cell

RESULTS
Sm F5H Is a Phenylpropanoid DualMeta-Hydroxylase
in Vitro
In our previous study, we demonstrated that like angiosperm
F5Hs, Sm F5H uses coniferaldehyde and coniferyl alcohol as
substrates in preference to ferulic acid (Humphreys et al., 1999;
Weng et al., 2008c). To test whether Sm F5H can use phenyl-
propanoids other than those G-substituted intermediates as
substrates, we performed a series of kinetic assays using Sm
F5H against a wider range of phenylpropanoid pathway inter-
mediates. Parallel comparative assays were conducted using
Arabidopsis thaliana F5H (At F5H) recombinant protein.
Surprisingly, we found that although both At F5H and Sm F5H
can catalyze 5-hydroxylation reactions on G-substituted inter-
mediates equally well, Sm F5H can also efficiently catalyze the
3-hydroxylation of p-coumaraldehyde and p-coumaryl alcohol
(Table 1; see Supplemental Figure 1 online). In contrast with
Sm F5H, At F5H shows little activity toward p-coumaraldehyde
and p-coumaryl alcohol with Km values so high that these
activities are not likely to be relevant in vivo (Table 1; see
Supplemental Figure 1 online). The similarly ring-substituted
acid, p-coumaric acid, is not an optimal substrate for Sm F5H,
consistent with the high Km value observed for ferulic acid.
Neither Sm F5H nor At F5H showed any detectable activity
toward cinnamic acid, cinnamaldehyde, and cinnamyl alco-
hol, indicating the ring para-hydroxyl is required for the meta-
hydroxylase activity. When p-coumaroyl shikimic acid, the
substrate for angiosperm C39H (Schoch et al., 2001), was tested,
no activity was detected in either Sm F5H or At F5H assays,
suggesting that Sm F5H does not display promiscuous activity
toward any given p-coumaroyl derivatives. We also detected no
activity of Sm F5H toward caffeic acid, caffealdehyde, or caffeyl
alcohol, suggesting that the 3-hydroxylO-methylation is required
for the subsequent 5-hydroxylase activity of Sm F5H. Collec-
tively, the kinetic data imply that Selaginellamay have a pathway
for S lignin biosynthesis via the H-substituted aldehyde and
alcohol, a route that is thought to be absent in angiosperms
(Figure 1B).
Sm F5H Partially Rescues the Growth Defects of the
Arabidopsis C39H-Deficient Mutants
In Arabidopsis, mutants defective in each of the two phen-
ylpropanoid meta-hydroxylases, C39H and F5H, have been
isolated and characterized. Whereas various alleles of the
C39H-deficient reduced epidermal fluorescence8 (ref8) mutants
exhibit severe dwarfism, female sterility, greatly reduced soluble
sinapate ester and total lignin content, and a lignin composed of
almost pure H units (Franke et al., 2002a, 2002b; Abdulrazzak
et al., 2006), the F5H-deficient fah1-2 mutant shows a total loss
of sinapate esters and S lignin but normal growth (Chapple et al.,
1992). We have shown that the Sm F5H transgene can rescue
fah1-2mutant biochemical phenotypes (Weng et al., 2008c), but
our revised kinetic analysis of the enzyme’s substrate specificity
suggested that this complementation experiment may have
exploited only a portion of the catalytic repertoire of Sm F5H.
We postulated that if the phenylpropanoid dualmeta-hydroxylase
activity of Sm F5H observed in vitro is relevant in vivo, transgenic
expression of Sm F5Hwould also be able to rescue ref8 because
the p-coumaraldehyde and p-coumaryl alcohol that the mutant
employs for H lignin synthesis would be available as substrates
for the enzyme.
To test this hypothesis, we generated Sm F5H transgenics in
ref8 fah1-2 double mutant backgrounds by crossing plants
carrying one of two ref8 alleles, ref8-1 (a slightly leaky allele
that carries a point mutation as described by Franke et al.
[2002b]) and ref8-2 (a T-DNA insertional null allele), with four
independent transgenic lines previously generated in the fah1-2
background that harbor the Sm F5H transgene under the control
of the Arabidopsis C4H promoter (AtC4H:SmF5H), a promoter
that has been shown to efficiently target expression in vascular
tissue (Weng et al., 2008c). In both cases, visual inspection of the
resulting ref8/fah1-2/AtC4H:SmF5H plants from the F2 genera-
tion indicated a partial but substantial complementation of the
growth phenotype (Figures 2 and 3). Comparedwith ref8 and ref8
Table 1. Kinetic Properties of Sm F5H and At F5H against Phenylpropanoid Intermediates
Substrate
Sm F5H At F5H
Kma (mM) Vmax
a (pkat·mg�1) Vmax/Km Kma (mM) Vmax
a (pkat·mg�1) Vmax/Km
p-Coumaric acid 1.67 3 103 1.67 1.00 3 10�3 N.D.b N.D.b N.D.b
p-Coumaraldehyde 4.50 4.45 0.989 3.71 3 102 0.700 1.89 3 10�3
p-Coumaryl alcohol 5.11 3.58 0.701 5.95 3 102 0.606 1.02 3 10�3
Ferulic acid 1.09 3 103 1.35 1.24 3 10�3 1.85 3 103 2.03 1.10 3 10�3
Coniferaldehyde 2.80 3.40 1.21 3.06 2.18 0.712
Coniferyl alcohol 3.63 2.56 0.705 1.76 1.73 0.983
No activity was detected using either Sm F5H or At F5H against the following substrates at a concentration of 200 mM: cinnamic acid,
cinnamaldehyde, cinnamyl alcohol, p-coumaroyl shikimic acid, caffeic acid, caffealdehyde, and caffeyl alcohol.aExtrapolated using Lineweaver-Burk plots based on triplicate assays. Note that the enzyme assays were performed using yeast microsomes
containing the heterologously expressed catalysts in which the amount of P450 was not quantified. The kinetic constants involving Vmax cannot be
compared between assays conducted using Sm F5H and At F5H.bKinetic constants not determined (N.D.; no detectable activity).
Alternative Route of S Lignin Synthesis 1035

fah1-2 plants, which are severe dwarfs with miniature rosettes,
ref8/fah1-2/AtC4H:SmF5H plants are significantly larger in stat-
ure (Figures 2 and 3). The dark-green/purple color typically
observed in ref8 or ref8 fah1-2 rosette leaves is also greatly
alleviated in the Sm F5H transgenics, indicating a decrease in the
accumulation of anthocyanins. Despite the considerable com-
plementation in growth phenotype, the female sterility pheno-
type of ref8 is not rescued in the Sm F5H transgenics, suggesting
that the alternative phenylpropanoid meta-hydroxylation path-
way mediated by Sm F5H is not sufficient to compensate for the
loss of C39H activity in flower development in ref8 or that the At
C4H promoter does not target Sm F5H expression to the
necessary tissues or cells.
To test whether overexpression of At F5H can rescue ref8,
we generated similar transgenic plants carrying an At C4H
promoter-driven At F5H transgene (AtC4H:AtF5H). None of the
resulting ref8-1/fah1-2/AtC4H:AtF5H transgenic lines showed
any sign of phenotypic complementation (data not shown), which
is consistent with the in vitro observation that At F5H is not
effective as a 3-hydroxylase. The above data suggest that the
complementation of ref8 by Sm F5H is due to its specific
3-hydroxylase activity and not the 5-hydroxylase activity it
shares with At F5H.
Sm F5H Restores S Lignin Biosynthesis in the Arabidopsis
C39H-Deficient Mutants
To examine whether the rescue of the growth phenotype is
associated with a restoration of normal lignin deposition in the
ref8/fah1-2/AtC4H:SmF5H transgenics, we first performed
phloroglucinol-HCl and Maule histochemical staining on stem
cross sections (Figure 4). The phloroglucinol-HCl reagent gives a
red reaction when it reacts with hydroxycinnamaldehyde end
groups in the lignin polymer, whereas the Maule reagent gives a
qualitative indication of lignin monomer composition by staining
G lignin brown and S lignin red. Compared with the wild type and
fah1-2, ref8-1 and the ref8-1 fah1-2 double mutant show much
weaker phloroglucinol-HCl staining and no Maule staining, con-
sistent with decreased total lignin and lack of G and S lignin units
in these mutants. By contrast, the ref8-1/fah1-2/AtC4H:SmF5H
transgenics exhibited strong red Maule staining in both xylem
and interfascicular fiber cells, indicating that S lignin deposition is
restored in these cells. Little phloroglucinol-HCl staining was
detected in sections of these plants, suggesting low quantities of
aldehyde end groups in the transgenic lignin, a character rem-
iniscent of high S lignin as previously reported (Franke et al.,
2000).
We then quantified the total lignin content of the transgenic
plants together with the control plants by Klason analysis (Figure
5). Compared with ref8-1 and the ref8-1 fah1-2 double mutant,
which contain only about one-third of the Klason lignin content
found in the wild type, the ref8-1/fah1-2/AtC4H:SmF5H trans-
genics deposit about three-quarters of the wild-type level of
lignin. Interestingly, the ref8-2/fah1-2/AtC4H:SmF5H trans-
genics also have a Klason total lignin content similar to the
ref8-1/fah1-2/AtC4H:SmF5H transgenics, despite their pheno-
typic difference in growth (Figures 2E and 2H). This observation
is consistent with the previous suggestion that deficiency in
Figure 2. Partial Complementation of the Growth Phenotype of the Arabidopsis ref8 fah1-2 Double Mutant by the Sm F5H Transgene.
(A) Columbia wild type.
(B) fah1-2.
(C) ref8-1.
(D) ref8-1 fah1-2.
(E) A representative line of ref8-1/fah1-2/AtC4H:SmF5H.
(F) ref8-2.
(G) ref8-2 fah1-2.
(H) A representative line of ref8-2/fah1-2/AtC4H:SmF5H.
All plants were photographed at 2 months of age, and all images are shown at an identical scale.
[See online article for color version of this figure.]
1036 The Plant Cell

factors other than lignin (e.g., some unknown growth substance
synthesized via C39H) may contribute to the dwarfism in ref8
mutants (Abdulrazzak et al., 2006).
We further examined the impact of Sm F5H on lignin monomer
composition in the ref8 fah1-2background using the derivatization
followed by reductive cleavage (DFRC) method, a procedure that
specifically releases lignin monomers (as their peracetates) from
b-O-4-linked lignin units (Figure 6, Table 2; see Supplemental
Figure 2 online). Asmentioned previously, compared with the wild
type, which contains both G and S units with traces of H units, S
units in fah1-2 are below detectable limits. The ref8-1 fah1-2 line
resembles ref8-1 in that it deposits essentially only H subunits and
only accumulates low levels of lignin. Both ref8-1/fah1-2/AtC4H:
SmF5H and ref8-2/fah1-2/AtC4H:SmF5H transgenics show a
significant recovery of the total DFRC-releasable lignin monomer
yield. More surprisingly, these plants contain lignin with compa-
rable amounts of releasable H and S units, but very few G units, a
lignin composition not previously identified in nature.
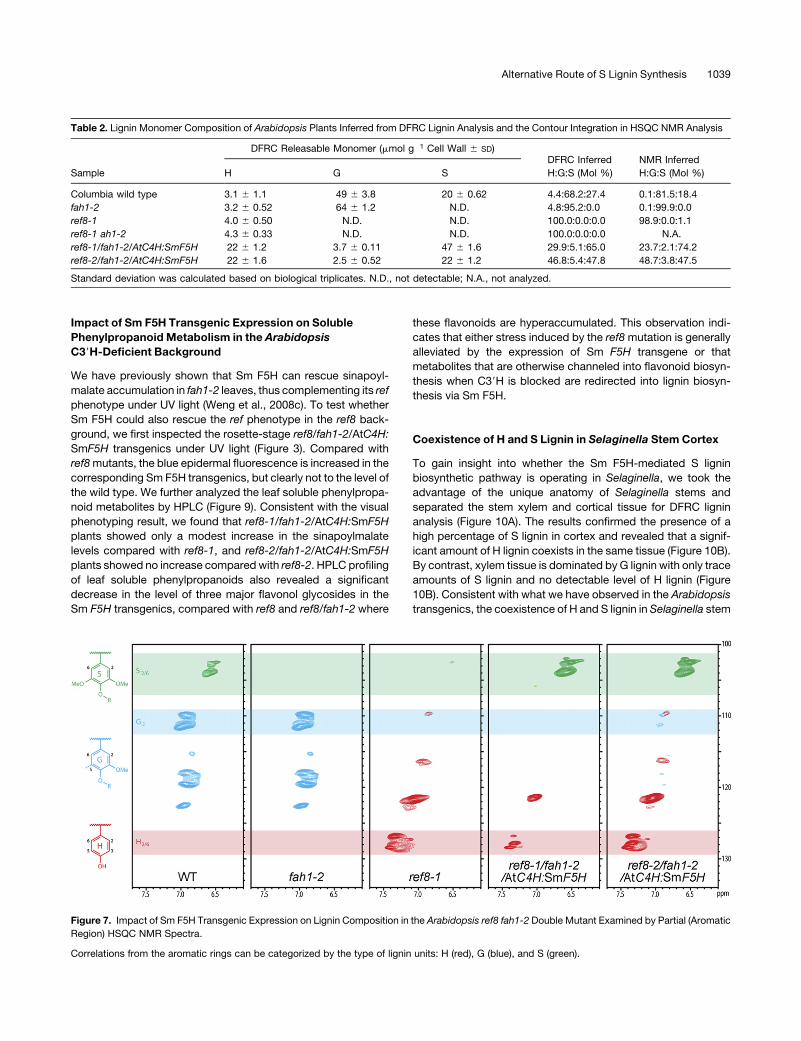
To independently evaluate the novel lignin composition in the
ref8/fah1-2/AtC4H:SmF5H transgenics,we conductedNMRanal-
ysis on whole lignins released from cell walls by treatment with
polysaccharide hydrolases. These data clearly show that the
transgenic expression of Sm F5H can rescue S lignin biosynthesis
inArabidopsis ref8 fah1-2 doublemutants and provide support for
the observation of unique H-S lignins (Figure 7). Contour integra-
tion in two-dimensional 13C–1H-correlated (HSQC) NMR also
allowed better estimates of the H:G:S distribution of the whole
lignin (Ralph et al., 2006; Wagner et al., 2007) (Table 2). The
lignins displayed distinctly different structural attributes result-
ing from the diverse distributions of lignin monomers (see
Figure 3. Photographs of Rosette-Stage Arabidopsis Plants under Visible Light and UV Light.
Blue fluorescence under UV light indicates the presence of sinapoylmalate in the leaf epidermis, whereas red fluorescence indicates its absence and
results from the UV-induced chlorophyll fluorescence. Top, visible light; bottom, UV light.
Figure 4. Impact of Sm F5H Transgenic Expression on Lignin Histochemical Staining in the Arabidopsis ref8 fah1-2 Double Mutant.
Phloroglucinol-HCl (top) and Maule (bottom) histochemical staining of 2-month-old Arabidopsis stem cross sections. The phloroglucinol-HCl reagent
detects aldehyde groups contained in lignin and results in red staining that is generally indicative of the presence of lignin. The Maule reagent is
diagnostic for the presence of S units in lignin, leading to a yellow staining of G lignins and red staining of lignins containing S subunits.
Alternative Route of S Lignin Synthesis 1037

Supplemental Figure 3 online). For example, 5-5–linked struc-
tures (in dibenzodioxocin units) were elevated in the G-rich
fah1-2mutant but greatly reduced in the S-elevated transgenics.
Sm F5H Rescues the Arabidopsis HCT-Deficient Mutant
Taken together, the above results suggest that Sm F5H can
mediate a novel lignin biosynthetic route that bypasses three
enzymes, hydroxycinnamoyl-CoA:shikimate hydroxycinnamoyl
transferase (HCT), C39H, and caffeoyl-CoAO-methyl transferase
(CCoAOMT), which catalyze four consecutive steps in the ca-
nonical pathway in angiosperms (Figure 1B). This hypothesis
predicts that Sm F5H should also be able to rescue the pheno-
types exhibited by an Arabidopsis HCT-deficient mutant, just as
it rescues the ref8 mutant. To test this prediction, we generated
AtC4H:SmF5H transgenics in a genetic background where HCT
is downregulated by RNA interference (RNAi). As has been
reported previously, HCT downregulated plants phenocopy
ref8 in that they show severe dwarfism and a dark-green/purple
coloration of their rosette leaves, although in general they are not
female-sterile like ref8 (Figure 8A) (Hoffmann et al., 2004). While
quantitative RT-PCR shows that HCT transcript levels in
HCTRNAi/AtC4H:SmF5H plants are similar to those of HCTRNAi
plants (Figure 8B), the plants are significantly rescued in growth
and exhibit decreased leaf pigmentation (Figure 8A). DFRC lignin
analysis revealed thatHCTRNAiplants deposit trace amounts of H
lignin, whereas the lignin of HCTRNAi/AtC4H:SmF5H plants con-
tains comparable amount of H and S subunits similar to the lignin
of ref8/fah1-2/AtC4H:SmF5H plants (Figure 8C). These obser-
vations further support the rerouted lignin biosynthetic pathway
mediated by Sm F5H, which bifurcates from the canonical
pathway upstream of HCT.
Figure 5. Restoration of Total Acid-Insoluble Lignin Content by Sm F5H
in the Arabidopsis ref8 fah1-2 Double Mutant Determined by Klason
Analysis.
Error bars represent 1 SD of biological triplicates. N.D., not determined
(no inflorescence stems could be harvested from these plants).
Figure 6. Impact of Sm F5H Transgenic Expression on Lignin Compo-
sition in the Arabidopsis ref8 fah1-2 Double Mutant Examined by DFRC–
Gas Chromatography Analysis.
H/G/S are p-hydroxyphenyl/guaiacyl/syringyl lignin-derived hydroxycin-
namyl alcohol peracetates. c/t, cis/trans; IS, internal standard.
[See online article for color version of this figure.]
1038 The Plant Cell
Impact of Sm F5H Transgenic Expression on Soluble
Phenylpropanoid Metabolism in the Arabidopsis
C39H-Deficient Background
We have previously shown that Sm F5H can rescue sinapoyl-
malate accumulation in fah1-2 leaves, thus complementing its ref
phenotype under UV light (Weng et al., 2008c). To test whether
Sm F5H could also rescue the ref phenotype in the ref8 back-
ground, we first inspected the rosette-stage ref8/fah1-2/AtC4H:
SmF5H transgenics under UV light (Figure 3). Compared with
ref8mutants, the blue epidermal fluorescence is increased in the
corresponding Sm F5H transgenics, but clearly not to the level of
the wild type. We further analyzed the leaf soluble phenylpropa-
noid metabolites by HPLC (Figure 9). Consistent with the visual
phenotyping result, we found that ref8-1/fah1-2/AtC4H:SmF5H
plants showed only a modest increase in the sinapoylmalate
levels compared with ref8-1, and ref8-2/fah1-2/AtC4H:SmF5H
plants showed no increase comparedwith ref8-2. HPLC profiling
of leaf soluble phenylpropanoids also revealed a significant
decrease in the level of three major flavonol glycosides in the
Sm F5H transgenics, compared with ref8 and ref8/fah1-2 where
these flavonoids are hyperaccumulated. This observation indi-
cates that either stress induced by the ref8mutation is generally
alleviated by the expression of Sm F5H transgene or that
metabolites that are otherwise channeled into flavonoid biosyn-
thesis when C39H is blocked are redirected into lignin biosyn-
thesis via Sm F5H.
Coexistence of H and S Lignin in Selaginella Stem Cortex
To gain insight into whether the Sm F5H-mediated S lignin
biosynthetic pathway is operating in Selaginella, we took the
advantage of the unique anatomy of Selaginella stems and
separated the stem xylem and cortical tissue for DFRC lignin
analysis (Figure 10A). The results confirmed the presence of a
high percentage of S lignin in cortex and revealed that a signif-
icant amount of H lignin coexists in the same tissue (Figure 10B).
By contrast, xylem tissue is dominated by G lignin with only trace
amounts of S lignin and no detectable level of H lignin (Figure
10B). Consistent with what we have observed in the Arabidopsis
transgenics, the coexistence of H and S lignin in Selaginella stem
Table 2. Lignin Monomer Composition of Arabidopsis Plants Inferred from DFRC Lignin Analysis and the Contour Integration in HSQC NMR Analysis
Sample
DFRC Releasable Monomer (mmol g�1 Cell Wall 6 SD)DFRC Inferred
H:G:S (Mol %)
NMR Inferred
H:G:S (Mol %)H G S
Columbia wild type 3.1 6 1.1 49 6 3.8 20 6 0.62 4.4:68.2:27.4 0.1:81.5:18.4
fah1-2 3.2 6 0.52 64 6 1.2 N.D. 4.8:95.2:0.0 0.1:99.9:0.0
ref8-1 4.0 6 0.50 N.D. N.D. 100.0:0.0:0.0 98.9:0.0:1.1
ref8-1 ah1-2 4.3 6 0.33 N.D. N.D. 100.0:0.0:0.0 N.A.
ref8-1/fah1-2/AtC4H:SmF5H 22 6 1.2 3.7 6 0.11 47 6 1.6 29.9:5.1:65.0 23.7:2.1:74.2
ref8-2/fah1-2/AtC4H:SmF5H 22 6 1.6 2.5 6 0.52 22 6 1.2 46.8:5.4:47.8 48.7:3.8:47.5
Standard deviation was calculated based on biological triplicates. N.D., not detectable; N.A., not analyzed.
Figure 7. Impact of Sm F5H Transgenic Expression on Lignin Composition in the Arabidopsis ref8 fah1-2 Double Mutant Examined by Partial (Aromatic
Region) HSQC NMR Spectra.
Correlations from the aromatic rings can be categorized by the type of lignin units: H (red), G (blue), and S (green).
Alternative Route of S Lignin Synthesis 1039

cortex suggests that the novel S lignin biosynthetic pathway
derived from H lignin precursors may be active in Selaginella.
DISCUSSION
F5HMediates a C39H-Independent S Lignin Biosynthetic
Pathway in Selaginella
We have provided both in vitro and in vivo evidence to show that
the convergent evolution of S lignin in lycophytes and angio-
sperms is the result of the elaboration of alternative biochemical
pathways. Whereas in angiosperms two phenylpropanoid meta-
hydroxylases, C39HandF5H, are required for S ligninbiosynthesis
from para-hydroxylated intermediates, a unique phenylpropanoid
dualmeta-hydroxylase, Sm F5H, evolved in the lycophyte Selag-
inella where it mediates S lignin biosynthesis via a different
pathway (Figure 1B). Our data suggest that G and S lignin can
be biosynthesized independently of one another through two
distinct sets of enzymes in Selaginella, in comparison to in
angiosperms where S lignin subunits are derived from G lignin
precursors.
We postulate that the C39H-independent S lignin biosynthetic
pathway in Selaginella requires two independent or one bifunc-
tional COMT to catalyze themethylation step after the addition of
each ring meta-hydroxyl by Sm F5H (Figure 1B). Bifunctional
COMTs with activities toward 3- and 5-hydroxylated phenyl-
propanoid aldehydes and alcohols are known in angiosperms
(Parvathi et al., 2001; Do et al., 2007). Such bifunctional COMTs
could be ubiquitous in vascular plants or, like the phenylpropa-
noid 5-hydroxylases, may have emerged independently in Se-
laginella and angiosperms.
It is also important to note that the in vitro kinetics data
showed that SmF5Hdoes not produce 3,4,5-trihydroxycinnamyl
aldehyde/alcohol from p-coumaryl- or caffeyl-substituted
substrates. These data suggest that after the first meta-
hydroxylation step, the dihydroxy-substituted product is
released from the active site of Sm F5H, and subsequent 3-O-
methylation is required for the reentry of the guaiacyl-substituted
substrate into the enzyme active site for the 5-hydroxylation.
These biochemical characteristics resemble those of the re-
cently characterized Arabidopsis CYP98A8, a phenylpropanoid
dual meta-hydroxylase involved in the biosynthesis of N1,N5-
di(hydroxyferuloyl)-N10-sinapoylspermidine (Matsuno et al.,
2009). Interestingly, the phenylpropanoid dualmeta-hydroxylase
activities of Sm F5H and CYP98A8 are distinct from that of the
petunia flavonoid 39,59-hydroxylase, which is capable of cata-
lyzing 39- and 59-hydroxylations of 49-hydroxylated flavonone
substrates without meta-O-methylation after the addition of the
first meta-hydroxyl group (Menting et al., 1994).
The C39H-independent S lignin biosynthetic pathway in
Selaginella also requires a form of hydroxycinnamoyl-CoA
Figure 8. Complementation of the Arabidopsis HCTRNAi Plant by Sm F5H.
(A) Four-week-old Arabidopsis plants showing the partial rescue of growth phenotype of an HCTRNAi plant by the Sm F5H transgene.
(B) Relative HCT transcript levels in plants quantified by quantitative RT-PCR. Error bars represent standard deviations based on assays of biological
duplicates.
(C) DFRC–gas chromatography analysis of lignin monomer composition. H/G/S are p-hydroxyphenyl/guaiacyl/syringyl lignin-derived hydroxycinnamyl
alcohol peracetates. c/t, cis/trans; IS, internal standard.
[See online article for color version of this figure.]
1040 The Plant Cell

reductase (CCR) that can efficiently convert p-coumaroyl-CoA to
p-coumaraldehyde. It is possible that, like Sm F5H, this enzyme
is also specifically expressed in the cortex (Weng et al., 2008c),
where it provides H-substituted precursors for Sm F5H, some of
which escape hydroxylation and lead to the deposition of H lignin
in this tissue.
Unlike Selaginella, angiosperms usually contain only trace
amount of H lignin (Boerjan et al., 2003), an observation that may
be explained by previous studies that have shown that for a
number of angiospermCCR isoforms,p-coumaroyl-CoA is a less
favorable substrate than the G lignin precursor, feruloyl-CoA
(Wengenmayer et al., 1976; Sarni et al., 1984; Baltas et al., 2005).
That carbon flux is not stoichiometrically redirected into H lignin
when G and S lignin is eliminated in the Arabidopsis ref8 mutant
further supports this suggestion (Franke et al., 2002a; Abdulrazzak
et al., 2006). Surprisingly, we found that transgenic expression of
the bifunctional Sm F5H in the ref8 background not only restored
S lignin biosynthesis but also significantly enhanced H lignin
deposition. This result suggests that at least in the lignifying
tissue, Arabidopsis contains CCR isoforms that can support the
biosynthesis of substantial amounts of H-substituted lignin pre-
cursors. Thus, in contrast with what has been thought previously,
it seems that CCR is unlikely to be the factor that limits H lignin
biosynthesis, at least in wild-type Arabidopsis. Although it is
unclear why Sm F5H is more effective in restoring S lignin
synthesis than it is in complementing the sinapoylmalate-
deficient phenotype of the fah1 ref8 double mutant, it is possible
that a CCR that catalyzes the conversion of p-coumaroyl-CoA to
p-coumaraldehyde may be limiting or absent in Arabidopsis leaf
tissue, limiting the availability of substrate for Sm F5H in leaves.
Despite the presence of a C39H-independent S lignin biosyn-
thetic pathway mediated by Sm F5H, the Selaginella genome
contains a C39H ortholog that is >60% identical to angiosperm
C39H at amino acid level (Weng et al., 2008c), which suggests
that Selaginella may also contain a C39H-dependent pathway,
analogous to the one defined in angiosperms (Franke et al.,
2002b). The fact that Selaginella xylem, where Sm F5H is not
highly expressed (Weng et al., 2008c), contains lignin composed
of almost pure G units, is consistent with this notion, although the
exact biochemical function of C39H ortholog in Selaginella is still
to be determined.
SmF5HProvides a Valuable Tool for Genetic Engineering of
Lignin Biosynthesis in Higher Plants
Lignin is a ubiquitous component of the cell wall of vascular
plants and is essential to normal plant development, and plants in
Figure 9. HPLC Quantification of Major Phenylpropanoid Metabolites in
Leaves of 3-Week-Old Arabidopsis.
Sinapoylmalate can be detected in ref8-1 and even the null mutant ref8-
2, which is consistent with a previous report (Abdulrazzak et al., 2006)
and suggests the presence of a C39H independent 3-hydroxylation
pathway in ref8 mutant background. We observed that At F5H pos-
sesses a level of 3-hydroxylase activity toward p-coumaraldehyde and
p-coumaryl alcohol (Table 1). Although the kinetic constants suggest
such reactions cannot take place in vivo, under ref8mutant background,
the levels of relevant substrates might reach local concentrations that
could permit some flux through At F5H. KG1, kaempferol 3-O-b-[b-D-
glucopyranosyl(1/6)- D-glucopyranoside]-7-O-a-L-rhamnopyranoside;
KG2, kaempferol 3-O-b-D-glucopyranoside-7-O-a-L-rhamnopyranoside;
KG3, kaempferol 3-O-a-L-rhamnopyranoside-7-O-a-L-rhamnopyranoside;
SM, sinapoylmalate.
[See online article for color version of this figure.]
Figure 10. Tissue-Specific Lignin Analysis in Selaginella Stems.
(A) A longitudinal section of mature Selaginella stem showing that xylem and cortical tissue can be separated. Bar = 1 mm.
(B) Lignin monomer composition (H, G, and S) in separated stem xylem and cortical tissue quantified by DFRC lignin analysis. N.D., not detectable.
[See online article for color version of this figure.]
Alternative Route of S Lignin Synthesis 1041

which the lignin biosynthetic pathway has been downregulated
often suffer from significantly reduced growth (Weng et al.,
2008b). The discovery of the phenylpropanoid dual meta-
hydroxylase activity of Sm F5H and its role in the unique C39H-independent S lignin biosynthetic pathway suggests it may be a
valuable tool for fundamentally rerouting lignin biosynthesis in
higher plants. In the presence of Sm F5H, several essential lignin
biosynthetic genes in the canonical pathway, including C39H and
HCT as demonstrated in this study, could be downregulated
without causing deleterious effects and resulted in a unique H-S
lignin composition in the transgenic plants, which may not occur
in nature. The mechanism that leads to the unique H-S lignin is
currently unknown but deserves additional investigation in the
future.
S Lignin Could Have Emerged Multiple Times during
Plant Evolution
Some older literature reported that S lignin could be detected in
some fern and gymnosperm species (e.g., cuplet fern, yew plum
pine, sandarac-cypress, and gnetophytes) (reviewed in Weng
et al., 2008a), which suggests that S lignin might have evolved
multiple times during plant evolution, but additional F5H analogs
and/or alternative pathways toward S lignin biosynthesis have
yet to be discovered. The apparently independent occurrence of
S lignin in distantly related plant lineages implies that it may have
an important role in plants’ adaptation to their environment. S
lignin is present in the anatomically analogous fiber cells in stems
of angiosperms and Selaginella, suggesting that S lignin may
function similarly in both lineages for mechanical support and/or
defense against pathogens and herbivores (Li et al., 2001; Weng
et al., 2008c). For example, the evolution of S-lignified fibers in
angiosperms, Selaginella, and some members of the Gnetales
may indicate that the presence of these strengthening cell types
permitted the development of relatively weaker vessel elements
in the vasculature of these plants (Logan and Thomas, 1985;
Carlquist, 1996).
METHODS
Plant Material
Arabidopsis thaliana was grown under a 16-h-light/8-h-dark photoperiod
at 100 mE·m22·s21 at 228C. Columbia-0 was used as the wild type. The
T-DNA insertional ref8-2 null allele was obtained from the ABRCunder the
accession of SALK_036132 (Alonso et al., 2003). Selaginella moellen-
dorffii was obtained from Plant Delights Nursery (Raleigh, NC) and grown
in a local greenhouse under 50% shade cloth.
Transgenic Plants
To generate ref8/fah1-2/AtC4H:SmF5H plants, ref8-1 and ref8-2 hetero-
zygous plants were used as the female parent in a cross with four
independent lines of previously described fah1-2/AtC4H:SmF5H plants
(lines 2, 6, 7, and 8) (Weng et al., 2008c). In the F2 generation, plants with
the genotype REF8/ref8 fah1-2/fah1-2 that were homozygous for the Sm
F5H transgene were selected and allowed to self. All subsequent anal-
yses were performed in the F3 generation on homozygous Sm F5H
transgenics that were genotyped as being ref8 fah1-2 double homozy-
gotes. All the independent transgenic lines showed partial phenotypic
complementation compared with the corresponding ref8 fah1-2 plants.
Transgenics that derived from fah1-2/AtC4H:SmF5H line 6 were used
for further detailed analysis. To generate ref8-1/fah1-2/AtC4H:AtF5H
plants, a similar approach was adopted as previously described using
the fah1-2/AtC4H:AtF5H transgenic plants as the male parent in the
initial cross.
To generate the HCTRNAi binary vector, a 356-bp HCT cDNA fragment
was PCR amplified by the primer pair 59-GGGGACAAGTTTGTACAA-
AAAAGCAGGCTAACATCAGAGATTCCACCA-39 and 59-GGGGACCA-
CTTTGTACAAGAAAGCTGGGTCAACTTCGGGAATAAGC-39. The resulting
PCR product was recombined with the modified version of pDONR221
(Invitrogen) using BP clonase (Invitrogen) to generate the entry clone. The
entry clone was then recombined with a destination binary vector
(modified from pBI121; Clontech) that harbors a cauliflower mosaic virus
35S promoter-driven Gateway RNAi cassette to generate the final con-
struct. The HCTRNAi construct was introduced into Arabidopsis via
Agrobacterium tumefaciens–mediated transformation (Clough and Bent,
1998). To generate HCTRNAi/AtC4H:SmF5H plants, plants with the geno-
type REF8/ref8 fah1-2/fah1-2 that were homozygous for the Sm F5H
transgene were used as the female parent to cross with the HCTRNAi
plants. In the F1 generation, the plants with the genotype REF8/REF8
FAH1/fah1-2 that were hemizygous for both the Sm F5H transgene and
the HCTRNAi transgene were analyzed. In parallel, HCTRNAi plants were
crossed to fah1-2 to generate plants with the genotype REF8/REF8
FAH1/fah1-2 that were hemizygous for the HCTRNAi transgene in the F1
generation, which were used as control plants.
To genotype ref8-1, the primer pair 59-CGAGCTATCATGGAGGAGC-
ATA-39 and 59-CAACAAGAGCATGAGCAGCAG-39 was used in com-
bination with EcoRV digestion, exploiting the cleaved amplified
polymorphic sequencemarker resulting from the point mutation in ref8-1.
Similarly, fah1-2 was genotyped using the primer pair 59-TGGTGTGTA-
CATATATGGATGAAGAA-39 and 59-TAGCAAGAGTGGTGAATATGT-
GAAGT-39 in combination with MseI digestion. To genotype ref8-2, LP
primer 59-TCGTGGTTTCTAATAGCGGTG-39 and RP primer 59-TGTTAA-
GAAAAACAATTAGGGTTTTTG-39, together with the T-DNA left border
BP primer 59-TGGTTCACGTAGTGGGCCATCG-39 were used according
to the previously described method (Alonso et al., 2003). To genotype the
presence of the Sm F5H transgene, the gene-specific primer pair
59-CAAGGTCCTCCACAAGAAGC-39 and 59-CAGTCGAAGCACTGGAT-
GAA-39was used. To genotype the presence of theHCTRNAi transgene, a
primer to the 35S promoter 59-GACCTAACAGAACTCGCCGTAAAGA-39
and an HCT gene-specific primer 59-TAAGGGTAGGAGCAAAATCAC-
CAAA-39 were used.
Yeast Expression of P450s and Enzyme Assays
The construction of Sm F5H and At F5H yeast expression vectors is
described by Weng et al. (2008c) and Humphreys et al. (1999), respec-
tively. Constructs were transformed into the WAT11 yeast strain, and
yeast growth, induction, and preparation of yeast microsomal extracts
were conducted as previously described (Humphreys et al., 1999).
Enzyme kinetic assays were performed essentially as described
(Humphreys et al., 1999). In brief, a reaction system containing 1 mM
NADP+, 10mMglucose-6-phosphate, and 4 units of glucose-6-phosphate
dehydrogenase was incubated for 5 min at 308C in the presence of
substrate to allow the generation of NADPH. The reaction was incubated
for 20 min, at 308C after adding the yeast microsomal extract, and was
terminated by adding glacial acetic acid. All the assays except those
involving hydroxycinnamyl alcohols were extracted with ethyl acetate,
dried in vacuo, resuspended in 50% methanol, and analyzed by HPLC.
The assays testing hydroxycinnamyl alcohols were analyzed by HPLC
directly. For assays testing Sm F5H against p-coumaric acid, a substrate
concentration series of 10, 14, 18, 25, 50, and 200 mM was used. For
1042 The Plant Cell

assays testing SmF5H against p-coumaraldehyde/p-coumaryl alcohol or
Sm F5H/At F5H against coniferaldehyde/coniferyl alcohol, a substrate
concentration series of 0.7, 0.8, 1, 1.3, 2, and 10mMwas used. For assays
testing Sm F5H/At F5H against ferulic acid, a substrate concentration
series of 40, 50, 70, 100, 200, and 1000 mMwas used. The same amount
of microsomal extract from a single prep was used for each enzyme
toward various substrates.
Histochemical Staining
For phloroglucinol-HCl staining, hand sections of 2-month-old Arabidop-
sis stems were stained with 1% phloroglucinol (w/v) in 12% HCl for 5 min
and observed under light microscope. For Maule staining, hand sections
of 2-month-old Arabidopsis stems were fixed in 4% glutaraldehyde,
rinsed in water, and treated for 10 min with 0.5% KMnO4. Sections were
then rinsed with water, treated for 5 min with 10% HCl, rinsed in water,
mounted in concentrated NH4OH, and examined under the light micro-
scope.
Lignin Analysis
Cell wall samples free of soluble metabolites were prepared as previously
described (Meyer et al., 1998). For Klason lignin analysis, 100 mg of cell
wall sample was swelled with 3 mL of 72% H2SO4 for 30 min at 308C and
then diluted with water to 4%H2SO4 and autoclaved at 1208C for 1 h. The
residue was filtered through a preweighed glass filter. The residue was
dried in an 808C oven on the filter overnight before being weighed at room
temperature. The DFRC lignin analysis was performed essentially as
previously reported (Lu and Ralph, 1998). Briefly, cell wall samples were
dissolved in acetyl bromide/acetic acid solution, containing 4,49-
ethylidenebisphenol as an internal standard. The reaction products
were dried down using nitrogen gas, dissolved in dioxane/acetic acid/
water (5/4/1, v/v/v), reacted with Zn dust, purified with C-18 SPE columns
(SUPELCO), and acetylated with pyridine/acetic anhydride (2/3, v/v). The
lignin derivatives were analyzed by gas chromatography/flame ionization
detection using response factors relative to the internal standard of 1.26
for p-coumaryl alcohol peracetate, 1.30 for coniferyl alcohol peracetate,
and 1.44 for sinapyl alcohol peracetate. The same samples were run
through gas chromatography–mass spectrometry in parallel to confirm
the identity of the derived hydroxycinnamyl alcohol peracetates.
Lignin Preparation for NMR Spectroscopy
Dried Arabidopsis stems (0.5 to 1.5 g) were ground in a Retsch MM301
shaker mill for 3 min at 30 s21 and extracted sequentially with water
(sonication, 20 min, four times), 80%methanol (four times), acetone (two
times), chloroform-acetone (1/1, v/v, two times), and acetone (one time)
again. The obtained isolated cell walls (0.3 to 1 g) were ball-milled for 5 h
per 1 g of sample weight (in 20 min on/10 min off cycles) using a Retsch
PM100 ball mill vibrating at 600 rpm with zirconium dioxide vessels (50
mL) containing ZrO2 ball bearings (10 3 10 mm). Ball milled walls were
transferred to centrifuge tubes (50 mL) and digested at 308C with crude
cellulases (Cellulysin; Calbiochem; 30mgg21 of sample, in pH5.0 sodium
acetate buffer, 33 2 d, fresh buffer and enzyme each time), leaving all of
the lignin and residual polysaccharides totaling 0.202 g (20.1% of the
original cell wall after extractions, wild type), 0.232 g (23.1%, fah1-2),
0.0452 g (15.3%, ref8-1), 0.0749 g (11.9%, ref8-1/fah1-2/AtC4H:SmF5H),
and 0.137 g (16.1%, ref8-2/fah1-2/AtC4H:SmF5H). The polysaccharidase-
digested cell wall fractions (70 mg each except for ref8-1, 45 mg) were
subjected to solubilization in DMSO/N-methylimidazole (2/1, v/v). Fol-
lowing acetic anhydride addition (0.5 mL, 1.5 h), the polysaccharidase-
digested cell walls gave acetylated samples for NMR measurement (Lu
and Ralph, 2003).
NMR Spectroscopy
The NMR spectra were acquired on a Bruker Biospin DMX-500 instru-
ment fitted with a sensitive cryogenically cooled 5-mm DCH 1H/13C
gradient probewith inverse geometry (proton coils closest to the sample).
Acetylated lignin preparations (5 to 80 mg) were dissolved in 0.5 mL
CDCl3; the central chloroform solvent peakwas used as internal reference
(dC 77.0, dH 7.26 ppm). HSQC experimental conditions were as described
previously (Wagner et al., 2007). Volume integration of contours in HSQC
plots was accomplished by Bruker’s TopSpin 2.0 software as described
(Ralph et al., 2006).
Quantitative RT-PCR
Total RNA was extracted from 3-week-old rosette leaves of Arabidopsis
plants using the RNeasy plant mini kit (Qiagen). Single-strand cDNAs
were synthesized via reverse transcription using the High Capacity cDNA
reverse transcription kit (Applied Biosystems). The cDNAs was treated
with RNase and used as template for real-time PCR. Quantitative real-
time PCR was performed on the StepOne Real-Time PCR system
(Applied Biosystems) using the DDCT method with default cycling pro-
gram. HCT was amplified using the primer pair 59-GAATTCCATAC-
GAGGGTTTGTCTT-39 and 59-GGGCAATGGCAACGGATA-39, whereas
At1g13320, as an internal standard (Czechowski et al., 2005), was
amplified using the primer pair 59-TAACGTGGCCAAAATGATGC-39 and
59-GTTCTCCACAACCGCTTGGT-39. Both the primer pairs were tested in
a standard curve analysis beforehand, which showed amplification
efficiency higher than 90%.
Leaf Sinapoylmalate and Flavonoid Analysis
Three-week-old Arabidopsis rosette leaves were harvested, ground in
liquid nitrogen, and extracted with 50%methanol (1 mL per 100 mg fresh
weight) for 2 h at 658C, and the extracts were then analyzed by HPLC.
Metaboliteswere separated on aC18 column, using a gradient from1.5%
acetic acid to 35% acetonitrile in 1.5% acetic acid at a flow rate of 1 mL
min21. Sinapoylmalate content was quantified using sinapic acid as
standard. The identification of the threemajor kaempferol glycosides was
according to a previous report (Veit and Pauli, 1999); quantification used
kaempferol as standard.
Accession Numbers
Sequence data from this article can be found in the Arabidopsis Genome
Initiative or GenBank/EMBL databases under the following accession
numbers: Sm F5H, EU032589; At F5H, At4g36220; andHCT, At5g48930.
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure 1. Lineweaver-Burk Representation of the
Kinetic Analysis of Phenylpropanoid Meta-Hydroxylation Reactions
Catalyzed by Yeast-Expressed Sm F5H or At F5H.
Supplemental Figure 2. GC-MS Confirmation of the Identity of
Peracetates of Monolignols Derived from the DFRC Lignin Analysis
Shown in Figure 6.
Supplemental Figure 3. Partial (Sidechain Region) HSQC Spectra
Showing the Structural Changes Resulting from Lignification Using
Altered Monolignol Supplies.
ACKNOWLEDGMENTS
This work is funded by the National Science Foundation (Grant IOB-
0450289). Partial funding to J.R. was via the Department of Energy
Alternative Route of S Lignin Synthesis 1043

(DOE) Office of Science (Grant DE-AI02-06ER64299) and the DOE Great
Lakes Bioenergy Research Center (DOE Office of Science BER DE-
FC02-07ER64494). N.D.B. is supported by a fellowship from the Life
Sciences Research Foundation. We thank J.A. Banks for providing
Selaginella moellendorffii plant materials.
Received December 15, 2009; revised March 8, 2010; accepted March
22, 2010; published April 6, 2010.
REFERENCES
Abdulrazzak, N., et al. (2006). A coumaroyl-ester-3-hydroxylase insertion
mutant reveals the existence of nonredundant meta-hydroxylation
pathways and essential roles for phenolic precursors in cell expan-
sion and plant growth. Plant Physiol. 140: 30–48.
Alonso, J.M., et al. (2003). Genome-wide insertional mutagenesis of
Arabidopsis thaliana. Science 301: 653–657.
Baltas, M., Lapeyre, C., Bedos-Belval, F., Maturano, M., Saint-
Aguet, P., Roussel, L., Duran, H., and Grima-Pettenati, J. (2005).
Kinetic and inhibition studies of cinnamoyl-CoA reductase 1 from
Arabidopsis thaliana. Plant Physiol. Biochem. 43: 746–753.
Boerjan, W., Ralph, J., and Baucher, M. (2003). Lignin biosynthesis.
Annu. Rev. Plant Physiol. Plant Mol. Biol. 54: 519–546.
Carlquist, S. (1996). Wood, bark, and stem anatomy of gnetales: A
summary. Int. J. Plant Sci. 157: S58–S76.
Chapple, C.C., Vogt, T., Ellis, B.E., and Somerville, C.R. (1992). An
Arabidopsis mutant defective in the general phenylpropanoid path-
way. Plant Cell 4: 1413–1424.
Cheng, X., Collins, R.E., and Zhang, X. (2005). Structural and se-
quence motifs of protein (histone) methylation enzymes. Annu. Rev.
Biophys. Biomol. Struct. 34: 267–294.
Clough, S.J., and Bent, A.F. (1998). Floral dip: A simplified method for
Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant
J. 16: 735–743.
Conant, G.C., and Wagner, A. (2003). Convergent evolution of gene
circuits. Nat. Genet. 34: 264–266.
Czechowski, T., Stitt, M., Altmann, T., Udvardi, M.K., and Scheible,
W.R. (2005). Genome-wide identification and testing of superior
reference genes for transcript normalization in Arabidopsis. Plant
Physiol. 139: 5–17.
Do, C.T., Pollet, B., Thevenin, J., Sibout, R., Denoue, D., Barriere, Y.,
Lapierre, C., and Jouanin, L. (2007). Both caffeoyl Coenzyme A 3-O-
methyltransferase 1 and caffeic acid O-methyltransferase 1 are in-
volved in redundant functions for lignin, flavonoids and sinapoyl
malate biosynthesis in Arabidopsis. Planta 226: 1117–1129.
Franke, R., Hemm, M.R., Denault, J.W., Ruegger, M.O., Humphreys,
J.M., and Chapple, C. (2002a). Changes in secondary metabolism
and deposition of an unusual lignin in the ref8 mutant of Arabidopsis.
Plant J. 30: 47–59.
Franke, R., Humphreys, J.M., Hemm, M.R., Denault, J.W., Ruegger,
M.O., Cusumano, J.C., and Chapple, C. (2002b). The Arabidopsis
REF8 gene encodes the 3-hydroxylase of phenylpropanoid metabo-
lism. Plant J. 30: 33–45.
Franke, R., McMichael, C.M., Meyer, K., Shirley, A.M., Cusumano, J.
C., and Chapple, C. (2000). Modified lignin in tobacco and poplar
plants over-expressing the Arabidopsis gene encoding ferulate
5-hydroxylase. Plant J. 22: 223–234.
Hedden, P., Phillips, A.L., Rojas, M.C., Carrera, E., and Tudzynski, B.
(2001). Gibberellin biosynthesis in plants and fungi: A case of con-
vergent evolution? J. Plant Growth Regul. 20: 319–331.
Hedenstrom, A., Johansson, L.C., Wolf, M., von Busse, R., Winter,
Y., and Spedding, G.R. (2007). Bat flight generates complex aero-
dynamic tracks. Science 316: 894–897.
Hoffmann, L., Besseau, S., Geoffroy, P., Ritzenthaler, C., Meyer, D.,
Lapierre, C., Pollet, B., and Legrand, M. (2004). Silencing of
hydroxycinnamoyl-coenzyme a shikimate/quinate hydroxycinnamoyl-
transferase affects phenylpropanoid biosynthesis. Plant Cell 16:
1446–1465.
Humphreys, J.M., Hemm, M.R., and Chapple, C. (1999). New routes
for lignin biosynthesis defined by biochemical characterization of
recombinant ferulate 5-hydroxylase, a multifunctional cytochrome
P450-dependent monooxygenase. Proc. Natl. Acad. Sci. USA 96:
10045–10050.
Jin, Z.F., Matsumoto, Y., Tange, T., Akiyama, T., Higuchi, M., Ishii,
T., and Iiyama, K. (2005). Proof of the presence of guaiacyl-syringyl
lignin in Selaginella tamariscina. J. Wood Sci. 51: 424–426.
Kenrick, P., and Crane, P.R. (1997). The origin and early evolution of
plants on land. Nature 389: 33–39.
Li, L., Cheng, X.F., Leshkevich, J., Umezawa, T., Harding, S.A., and
Chiang, V.L. (2001). The last step of syringyl monolignol biosynthesis
in angiosperms is regulated by a novel gene encoding sinapyl alcohol
dehydrogenase. Plant Cell 13: 1567–1586.
Logan, K.J., and Thomas, B.A. (1985). Distribution of lignin derivatives
in plants. New Phytol. 99: 571–585.
Logan, K.J., and Thomas, B.A. (1987). The distribution of lignin
derivatives in fossil plants. New Phytol. 105: 157–173.
Lu, F., and Ralph, J. (1998). The DFRC method for lignin analysis. 2.
Monomers from isolated lignins. J. Agric. Food Chem. 46: 547–552.
Lu, F., and Ralph, J. (2003). Non-degradative dissolution and acetyla-
tion of ball-milled plant cell walls: high-resolution solution-state NMR.
Plant J. 35: 535–544.
Matsuno, M., et al. (2009). Evolution of a novel phenolic pathway for
pollen development. Science 325: 1688–1692.
Menting, J., Scopes, R.K., and Stevenson, T.W. (1994). Characteri-
zation of flavonoid 3[prime],5[prime]-hydroxylase in microsomal mem-
brane fraction of Petunia hybrida flowers. Plant Physiol. 106: 633–642.
Meyer, K., Shirley, A.M., Cusumano, J.C., Bell-Lelong, D.A., and
Chapple, C. (1998). Lignin monomer composition is determined by
the expression of a cytochrome P450-dependent monooxygenase in
Arabidopsis. Proc. Natl. Acad. Sci. USA 95: 6619–6623.
Parvathi, K., Chen, F., Guo, D., Blount, J.W., and Dixon, R.A. (2001).
Substrate preferences of O-methyltransferases in alfalfa suggest new
pathways for 3-O-methylation of monolignols. Plant J. 25: 193–202.
Ralph, J., Akiyama, T., Kim, H., Lu, F., Schatz, P.F., Marita, J.M.,
Ralph, S.A., Reddy, M.S., Chen, F., and Dixon, R.A. (2006). Effects
of coumarate 3-hydroxylase down-regulation on lignin structure. J.
Biol. Chem. 281: 8843–8853.
Sarni, F., Grand, C., and Boudet, A.M. (1984). Purification and prop-
erties of cinnamoyl-CoA reductase and cinnamyl alcohol dehydro-
genase from poplar stems (Populus X euramericana). Eur. J. Biochem.
139: 259–265.
Schoch, G., Goepfert, S., Morant, M., Hehn, A., Meyer, D., Ullmann,
P., and Werck-Reichhart, D. (2001). CYP98A3 from Arabidopsis
thaliana is a 39-hydroxylase of phenolic esters, a missing link in the
phenylpropanoid pathway. J. Biol. Chem. 276: 36566–36574.
Tanaka, K., Barmina, O., and Kopp, A. (2009). Distinct developmental
mechanisms underlie the evolutionary diversification of Drosophila
sex combs. Proc. Natl. Acad. Sci. USA 106: 4764–4769.
Teutsch, H.G., Hasenfratz, M.P., Lesot, A., Stoltz, C., Garnier, J.M.,
Jeltsch, J.M., Durst, F., and Werck-Reichhart, D. (1993). Isolation
and sequence of a cDNA encoding the Jerusalem artichoke cinna-
mate 4-hydroxylase, a major plant cytochrome P450 involved in the
general phenylpropanoid pathway. Proc. Natl. Acad. Sci. USA 90:
4102–4106.
1044 The Plant Cell

Towers, G.H.N., and Gibbs, R.D. (1953). Lignin chemistry and the
taxonomy of higher plants. Nature 172: 25–26.
Veit, M., and Pauli, G.F. (1999). Major flavonoids from Arabidopsis
thaliana leaves. J. Nat. Prod. 62: 1301–1303.
Wagner, A., Ralph, J., Akiyama, T., Flint, H., Phillips, L., Torr, K.,
Nanayakkara, B., and Te Kiri, L. (2007). Exploring lignification in conifers
by silencing hydroxycinnamoyl-CoA:shikimate hydroxycinnamoyltrans-
ferase in Pinus radiata. Proc. Natl. Acad. Sci. USA 104: 11856–11861.
Weng, J.K., Banks, J.A., and Chapple, C. (2008a). Parallels in lignin
biosynthesis: A study in Selaginella moellendorffii reveals conver-
gence across 400 million years of evolution. Commun. Integr. Biol. 1:
20–22.
Weng, J.K., Li, X., Bonawitz, N.D., and Chapple, C. (2008b). Emerging
strategies of lignin engineering and degradation for cellulosic biofuel
production. Curr. Opin. Biotechnol. 19: 166–172.
Weng, J.K., Li, X., Stout, J., and Chapple, C. (2008c). Independent
origins of syringyl lignin in vascular plants. Proc. Natl. Acad. Sci. USA
105: 7887–7892.
Wengenmayer, H., Ebel, J., and Grisebach, H. (1976). Enzymic synthe-
sis of lignin precursors. Purification and properties of a cinnamoyl-
CoA: NADPH reductase from cell suspension cultures of soybean
(Glycine max). Eur. J. Biochem. 65: 529–536.
White, E., and Towers, G.H.N. (1967). Comparative biochemistry of
lycopods. Phytochemistry 6: 663–667.
Alternative Route of S Lignin Synthesis 1045

DOI 10.1105/tpc.109.073528; originally published online April 6, 2010; 2010;22;1033-1045Plant Cell
Jing-Ke Weng, Takuya Akiyama, Nicholas D. Bonawitz, Xu Li, John Ralph and Clint Chapple and Flowering PlantsSelaginella
Convergent Evolution of Syringyl Lignin Biosynthesis via Distinct Pathways in the Lycophyte
This information is current as of June 22, 2018
Supplemental Data /content/suppl/2010/04/06/tpc.109.073528.DC1.html
References /content/22/4/1033.full.html#ref-list-1
This article cites 42 articles, 17 of which can be accessed free at:
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists





























