Embed Size (px)
Citation preview

Convergent Signaling Pathways Regulate ParathyroidHormone and Fibroblast Growth Factor-23 Action onNPT2A-mediated Phosphate Transport*
Received for publication, June 18, 2016 Published, JBC Papers in Press, July 18, 2016, DOI 10.1074/jbc.M116.744052
W. Bruce Sneddon‡, Giovanni W. Ruiz§, Luciana I. Gallo§, Kunhong Xiao‡, Qiangmin Zhang‡, Youssef Rbaibi§,Ora A. Weisz§¶, Gerard L. Apodaca§¶, and Peter A. Friedman‡�1
From the ‡Laboratory for GPCR Biology, Department of Pharmacology and Chemical Biology and the Departments of �StructuralBiology, ¶Cell Biology, and §Medicine, University of Pittsburgh School of Medicine, Pittsburgh, Pennsylvania 15261
Parathyroid hormone (PTH) and FGF23 are the primary hor-mones regulating acute phosphate homeostasis. Human renalproximal tubule cells (RPTECs) were used to characterize themechanism and signaling pathways of PTH and FGF23 on phos-phate transport and the role of the PDZ protein NHERF1 in medi-ating PTH and FGF23 effects. RPTECs express the NPT2A phos-phate transporter, �Klotho, FGFR1, FGFR3, FGFR4, and the PTHreceptor. FGFR1 isoforms are formed from alternate splicing ofexon 3 and of exon 8 or 9 in Ir-like loop 3. Exon 3 was absent, butmRNA containing both exons 8 and 9 is present in cytoplasm.Using an FGFR1c-specific antibody together with mass spectrom-etry analysis, we show that RPTECs express FGFR-�1C. The dataare consistent with regulated FGFR1 splicing involving a novelcytoplasmic mechanism. PTH and FGF23 inhibited phosphatetransport in a concentration-dependent manner. At maximallyeffective concentrations, PTH and FGF23 equivalently decreasedphosphate uptake and were not additive, suggesting a sharedmechanism of action. Protein kinase A or C blockade preventedPTH but not FGF23 actions. Conversely, inhibiting SGK1, blockingFGFR dimerization, or knocking down Klotho expression dis-rupted FGF23 actions but did not interfere with PTH effects. C-ter-minal FGF23(180–251) competitively and selectively blockedFGF23 action without disrupting PTH effects. However, both PTHand FGF23-sensitive phosphate transport were abolished byNHERF1 shRNA knockdown. Extended treatment with PTH orFGF23 down-regulated NPT2A without affecting NHERF1. Weconclude that FGFR1c and PTHR signaling pathways converge onNHERF1 to inhibit PTH- and FGF23-sensitive phosphate trans-port and down-regulate NPT2A.
Parathyroid hormone (PTH)2 and FGF23 display tworemarkable features: 1) PTH and FGF23 exhibit parallel inhibi-
tion of renal phosphate transport mediated by NPT2A(sodium-dependent phosphate transporter-2a) but opposingactions on 1,25(OH)2-vitamin D; 2) despite being structurallyand functionally distinct classes of membrane-delimited recep-tors, PTH and FGF receptors activate kinases that obligatorilyphosphorylate NHERF1 at conserved sites required for theirphosphaturic action. Phosphorus is essential for growth andmaintenance of the skeleton and for generating high energyphosphate compounds. Evolutionary adaptation in humansand other terrestrial vertebrates to phosphorus-rich dietsinvolves cell and molecular mechanisms ensuring the efficienturinary elimination of excess inorganic phosphate. The renalproximal tubule is the primary site of phosphate homeostasisand hormone-dependent phosphate transport. The NPT2Asodium-dependent phosphate cotransporter (SLC34A1) inproximal tubules is regulated by PTH and FGF23 (1, 2). PTHand FGF23 reduce phosphate uptake by sequestering anddown-regulating NPT2A, thereby enhancing urinary phos-phate excretion (3, 4). PTH actions are mediated by its cognateG protein-coupled PTH receptor (PTHR) (5, 6). Both PKA andPKC have been implicated in PTH-dependent inhibition ofNPT2A (7–12). Using a signaling-selective form of the PTHRthat activates PKA or signaling-biased PTH analogs, Juppnerand co-workers (13, 14) determined that adenylyl cyclase andPKA regulate acute PTH effects on Npt2a and phosphate trans-port, whereas persistent reductions of phosphate transportrequire PKC.
FGF23 actions on NPT2A are facilitated by receptor tyrosinekinases FGFR1 and FGFR4 with the obligate participation of thecoreceptor �Klotho (15, 16). FGFR1 signaling by ERK1/2 andserum and glucocorticoid-activated kinase (SGK1) leads toinhibition of phosphate transport (17).
NPT2A harbors a canonical C-terminal PDZ recognitionmotif (-TRL) that binds to the PDZ scaffolding protein,Na�/H� exchanger regulatory factor 1 (NHERF1) (18, 19).NHERF1 assembles a ternary complex with NPT2A and ezrin,thereby linking the transporter to the actin cytoskeleton.NHERF1 is essential for the inhibitory action of PTH andFGF23 on phosphate transport (12, 17, 20). NHERF1-null miceand humans harboring mutations in SLC9A3R1 exhibit pro-nounced phosphate wasting, nephrolithiasis, and skeletal dis-
* This work was supported by National Institutes of Health Grants DK069998and DK105811 (to P. A. F.), DK101484 and DK100357 (to O. A. W.), and P30DK079307 (to G. A. and O. A. W.) and the Imaging Core of the PittsburghCenter for Kidney Research. The authors declare that they have no conflictsof interest with the contents of this article. The content is solely the respon-sibility of the authors and does not necessarily represent the official viewsof the National Institutes of Health.
1 To whom correspondence should be addressed: University of PittsburghSchool of Medicine, Department of Pharmacology and Chemical Biology,E1356 Thomas E. Starzl Biomedical Science Tower, 200 Lothrop St., Pitts-burgh, PA 15261. E-mail: [email protected].
2 The abbreviations used are: PTH, parathyroid hormone; PTHR, PTH receptor;Bis-I, bis-indolylmaleimide I; FGFR, fibroblast growth factor receptor;
RPTEC, telomerase-immortalized human renal proximal tubule cell; Nle,norleucine; Dsi-RNA, Dicer substrate siRNA.
crossmarkTHE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 291, NO. 36, pp. 18632–18642, September 2, 2016
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A.
18632 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 291 • NUMBER 36 • SEPTEMBER 2, 2016
by guest on Decem
ber 26, 2019http://w
ww
.jbc.org/D
ownloaded from

orders (21–23). Both PTH and FGF23 phosphorylate NHERF1(17, 24 –26). These findings imply that activation of distincttypes of cell surface receptors results in phosphorylation ofNHERF1, resulting in disassembly of the NPT2A-NHERF1-ezrin complex (27), internalization and down-regulation ofNPT2A, and cessation of phosphate transport. Although thecomponents of these pathways have been described, the lack ofa suitable cell model has hindered a comprehensive analysis ofPTH and FGF23 actions on NPT2A and cellular phosphatetransport. We report here that PTH and FGF23 inhibit NPT2A-dependent phosphate transport in immortalized human renalproximal epithelial cells (RPTECs). PTH actions mediated bythe PTHR involve PKA and PKC, whereas FGF23 effects pro-ceed through FGFR1c and SGK1 and require �Klotho. Inter-ruption of PTHR signaling inhibited PTH but not FGF23actions on NPT2A. Conversely, disrupting FGFR1-associatedproteins eradicated FGF23 but not PTH effects on phosphatetransport. NHERF1 knockdown prevented both PTH andFGF23 action, indicating that PTHR and FGFR1c signalingpathways converge at the level of NHERF1.
Results and Discussion
RPTECs retain many features of proximal tubule epithelialcells, including the formation of tight junctions, PTH-stimu-lated cAMP accumulation, and sodium-dependent phosphateuptake (28). To extend these observations, we characterized theexpression of NPT2A, NHERF1, PTHR, and FGFR and ana-lyzed the regulation of phosphate transport by PTH and FGF23.
NPT2A concentrated in a tightly organized cluster at theapical pole of the cells, reminiscent of apical recycling endo-somes described in other kidney cell types (Fig. 1A) (29, 30).NHERF1 was conspicuously expressed at the cell surface anddisplayed extensive colocalization with NPT2A (Fig. 1A,
merge). NPT2A also localized to the primary cilium; however,NHERF1 was absent from this structure (see arrows in Fig. 1Aand magnification in Fig. 1B). A similar distribution wasobserved using three different anti-NPT2A antibodies (data notshown). This localization contrasts with observations by Wadeet al. (31), who found NPT2A and NHERF1 in microvilli ofmouse proximal tubule cells. We also established that PTHRwas prominently expressed at both apical and basolateral cellmembranes (Fig. 1C) as described for native proximal tubules(10, 32, 33). FGFR1 was primarily expressed at the apical mem-brane (Fig. 1D).
RPTECs express mRNA transcripts for FGFR1, FGFR3, andFGFR4 mRNA (Fig. 2A). These results are compatible with recentfindings that FGFR1, FGFR3, and FGFR4 are expressed in murineproximal tubules (34, 35). This finding is consistent with studiesusing recombinant extracellular Klotho domain and FGFR-Fcfusion constructs of FGFR1b and FGFR1c that showed that FGF23binds only to FGFR1c (36). Based on these and other findings (37,38) FGFR1, with its cofactor �Klotho, is considered the cognateFGF23 receptor regulating phosphate transport.
FGFR1, along with FGFR2 and FGFR4, exhibit isotype varia-tion, ligand specificity, and regulation that are determined byextensive alternative splicing (39, 40). The primary sites ofFGFR1 splicing include Ig-like loops I and III (41, 42). The pat-tern of FGFR1 gene splicing and corresponding protein struc-tures are shown in Fig. 2B. The � or � isoforms of FGFR aregenerated by inclusion or exclusion of exon 3, which encodesthe first Ig-like loop (43). RT-PCR experiments using exon-spanning primers (Table 1) disclosed the absence of exon 3 (Fig.2C), hence indicating that FGFR1 is the � form, FGFR-�1, andlacks the Ig-I loop.
Further FGFR variation arises from alternative splicing in Ig-likeloop III. The N-terminal portion of loop III is encoded by exon 7.The C-terminal half of loop III is the extracellular ligand-bindingdomain of the receptor (44) and is generated by alternate inclusionof either exon 8, which then forms FGFR1b, or exon 9 to formFGFR1c. Regulatory RNA-binding proteins determine the spliceevents for FGFR1b and FGFR1c isoforms (42, 45). FGFR1b is con-sidered the epithelial form of the receptor, whereas FGFR1c isaccepted as expressed by mesenchymal cells (41). Thus, it wouldbe reasonable to suppose that epithelial proximal tubule cellswould express FGFR1b despite FGFR1c being accepted as thecanonical FGF23 receptor. We designed exon-spanning primers(Table 1) to delineate the FGFR1 isoform in RPTECs. Using for-ward primers spanning the exon 7/8 boundary (FGFR1b) or theexon 7/9 boundary (FGFR1c) and reverse primers that span theexon 10/8 boundary (FGFR1b) or the exon 10/9 boundary(FGFR1c), we determined that RPTECs use exon 9 and thusexpress FGFR1c (Fig. 2D).
Strikingly, using the FGFR1b forward primer with theFGFR1c reverse primer, mRNA containing both exons 8 and 9was robustly displayed in the cytoplasm (Fig. 2D). As notedabove, exon 3, however, was undetectable (Fig. 2B). This sug-gests that splicing of exon 3 for the FGFR1�/� receptor isoformoccurs in the nucleus by a canonical process, whereas exon splicingleading to tissue-specific expression of FGFR1b/c proceeds inthe cytoplasm. Such noncanonical post-transcriptional mRNA
FIGURE 1. NPT2A, NHERF1, PTHR, and FGFR1 expression in RPTECs. A,distribution of NPT2A (green), NHERF1 (red), and nuclei (blue) in filter-grownRPTECs. Single optical sections from the apical region of the cell are shown.Arrows, NPT2A association with cilia. B, magnified views of a single cell. C, x-zplane depicting PTHR (green) and nuclear (blue) staining of RPTECs grown oncoverslips. D, FGFR1 (green) and F-actin (red) staining of RPTECs grown onfilters. The x-z plane is shown.
PTH and FGF23 Signaling and Function
SEPTEMBER 2, 2016 • VOLUME 291 • NUMBER 36 JOURNAL OF BIOLOGICAL CHEMISTRY 18633
by guest on Decem
ber 26, 2019http://w
ww
.jbc.org/D
ownloaded from

processing is emerging as a cell-specific regulatory step thatdetermines the translated splice variant (46).
Using a polyclonal FGFR1 antibody that does not distinguishbetween alternatively spliced FGFR1 isoforms, RPTECs expressa single protein band of 72 kDa (Fig. 2E). A band of similar sizewas detected using an FGFR1c-specific antibody (47), confirm-ing that FGFR1c is the only species of FGFR1 protein expressedby RPTECs. We validated this finding by mass spectrometry,where we detected a unique tryptic peptide consistent with
FGFR1c (Fig. 2F). Protein containing both exons 8 and 9 wasnot found. Given that exons 8 and 9 each encode the mutuallyexclusive C-terminal portion of loop IIIc, it is unlikely that aprotein harboring both domains exists or would fold correctly.Thus, RPTECs express FGFR-�1c.
Taken together, these results establish that RPTECs expressNPT2A, NHERF1, PTHR, and FGFR-�1c, the proteins involvedin hormone-regulated phosphate transport. Based on theseconsiderations, we proceeded to characterize the actions of
FIGURE 2. FGFR mRNA and protein expression in RPTECs. A, representative 3% agarose gel of FGFR expression in RPTECs determined by RT-PCR (Table 2).200- and 100-bp markers are shown in the left lane. B, schematic representations of the FGFR1 gene and protein structure. The FGFR-�1c splice variant harborstwo extracellular immunoglobulin-like domains (II and III) with alternatively spliced exon 9 in loop III, a transmembrane domain (TM), and two intracellulartyrosine kinase domains (TK). C, FGFR1-� splice variant in a characteristic 2% agarose gel with ladder in the left lane. The 500-bp PCR product is consistent withthe �-splice variant lacking exon 3. D, FGFR1b/c splice variants. Lane 1, FGFR1b; lane 2, FGFR1c; lane 3, mRNA containing both exons 8 and 9. Lane 3 PCR wasperformed using the forward primer for FGFR1b and the reverse primer for FGFR1c (Table 1). An illustrative 2% agarose gel is shown with a 100-bp ladder in theleft lane. The PCR product in lane 2 is consistent with the exon 9 FGFR1c splice variant. Lane 3 shows an mRNA species containing both exons 8 and 9. E, FGFR1cis the primary species of FGFR1 detected by immunoblotting. FGFR1 and FGFR1c in RPTECs were detected by non-selective and isotype-selective antisera,respectively, as described under “Experimental Procedures.” The results are illustrative of n � 3 independent experiments. F, MS/MS spectrum for the identifiedspecific peptide 355SDFHSQMAVHKLAK368 from FGFR1c. The peak heights show the relative abundances of the corresponding fragmentation ions, with theannotation of the identified matched N terminus-containing b ions in blue and the C terminus-containing y ions in red. Charge state: �3, observed m/z:538.93969, theoretical m/z: 538.94008, precursor mass error: �0.72, Xcorr: 0.719.
PTH and FGF23 Signaling and Function
18634 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 291 • NUMBER 36 • SEPTEMBER 2, 2016
by guest on Decem
ber 26, 2019http://w
ww
.jbc.org/D
ownloaded from

PTH and FGF23 on phosphate transport. PTH and FGF23inhibited phosphate uptake in a concentration-dependentmanner with an EC50 of 30 and 51 pM, respectively (p � 0.02)(Fig. 3A). At maximally effective concentrations of 100 nM,FGF23 and PTH similarly inhibited phosphate uptake by 40 � 9and 43 � 7%, respectively (Fig. 3B). The combined addition ofFGF23 and PTH modestly augmented the inhibition of phos-phate uptake by an additional 20% (Fig. 3B). The non-additiveresponse of PTH and FGF23 probably reflects a common finalmechanism of action.
C-terminal FGF23(180 –251) virtually abolished the actionsof FGF23(28 –251) on phosphate transport but had no effect onits own; it did not interfere with PTH-sensitive phosphatetransport (Fig. 3C). FGF23(180 –251) competes with full-lengthFGF23 for binding to the FGFR1-�Klotho complex and reducesrenal phosphate excretion (48). Additional controls showedthat neither FGF2 nor PTH(7–34) affected phosphate transport(data not shown), further demonstrating the specificity ofFGF23 and PTH action on phosphate transport.
PTHR and FGFR1c mediate the phosphaturic actions of PTHand FGF23, respectively (5, 49, 50). Both receptors areexpressed on RPTECs (Fig. 2). The PTHR signals primarilythrough Gs and Gq, leading to downstream activation of PKAand PKC, which are stimulated in parallel following occupancyof proximal tubule PTHR (51). FGFR1 dimerizes upon activa-
tion, thereby stimulating downstream kinases associated withRas/ERK and SGK1 (17, 52). To define the mechanism of PTHand FGF23 action on phosphate transport in RPTECs, we firstcharacterized the signaling pathways responsible for theireffects by employing a panel of pharmacological protein kinaseinhibitors. The PKA inhibitor H89 and PKC inhibitor bis-in-dolylmaleimide I (Bis-I) blocked the actions of PTH (Fig. 4A)but had no effect on FGF23-sensitive phosphate uptake (Fig.4B). In contrast, the SGK1 inhibitor GSK650394 blocked theactions of FGF23 (Fig. 4B) but not PTH (Fig. 4A). These resultsare consistent with observations that acute NPT2A down-reg-ulation by PTH involves cAMP and PKA (14). The findings arealso compatible with a recent study showing that acute PTHaction involves PKA (14), whereas FGF23 effects are mediatedby SGK1 (17). The ERK1/2 inhibitor PD-98059 abolished theaction of FGF23 and inhibited PTH-sensitive uptake by 50%(Fig. 4, A and B). PTH activation of ERK occurs downstream ofPKC activation (53, 54). This may explain the partial inhibitionof PTH-sensitive phosphate transport by PD98059.
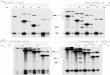
The finding that RPTECs express FGFR1c (Fig. 2, E and F)implies that knockdown of this isoform with an siRNA targetedto exon 9 should inhibit FGF23-sensitive phosphate transport.siRNA specific to either exon 8 (si8, FGFR1b) or exon 9 (si9,FGFR1c) was used to knock down FGFR1 expression. A sampleimmunoblot showing FGFR1c abundance after transfection ofRPTECs with siRNA is shown in Fig. 5A. si8 and si9 reducedFGFR1c protein expression by 82 and 89%, respectively (Fig.5B). Detection with a polyclonal anti-FGFR1 antibody yieldedsimilar results (data not shown). Unexpectedly, knocking downexon 8 (si8) was almost as effective as knocking down exon 9(si9) at reducing FGFR1c protein expression. si8 reducedFGF23-dependent phosphate transport by 40%, and si9 elimi-nated FGF23 modulation (Fig. 5C). These data are consistentwith FGFR1c being the primary mediator of the phosphaturiceffects of FGF23. Furthermore, the ability of si8 to interferewith FGFR1c expression supports the supposition that themRNA emerging from the nucleus contains both exons 8 and 9,
FIGURE 3. PTH and FGF23 inhibit phosphate transport. A, RPTECs were treated for 2 h with 100 nM PTH(1–34) or FGF23. Phosphate uptake was measured for10 min, as detailed under “Experimental Procedures.” Data represent the mean � S.E. (error bars) of n � 6 independent experiments performed in triplicate.Data were normalized for each experiment, where phosphate uptake under control, untreated conditions, was defined as 0% inhibition. Data were fit to asigmoidal relation, and Kd values were calculated with Prism. B, RPTECs on 12-well plates were treated for 2 h with the indicated concentrations of PTH or FGF23.Phosphate uptake was measured for 10 min, as outlined under “Experimental Procedures.” Data represent the mean � S.E. of n � 6 independent experimentsperformed in triplicate. *, p � 0.05 versus control; **, p � 0.01 versus control. C, C-terminal FGF23(180 –251) fragment blocks FGF23 but not PTH inhibitablephosphate uptake. Data are the mean � S.E. of n � 4 experiments. **, p � 0.01 versus FGF23.
TABLE 1FGFR1 exon splicing primers
PTH and FGF23 Signaling and Function
SEPTEMBER 2, 2016 • VOLUME 291 • NUMBER 36 JOURNAL OF BIOLOGICAL CHEMISTRY 18635
by guest on Decem
ber 26, 2019http://w
ww
.jbc.org/D
ownloaded from

as shown in Fig. 2C, and that regulated splicing in the cytoplasmdetermines the translation and expressed form of FGFR1.
FGF23 effects mediated by FGFR1 require the presence ofthe coreceptor �Klotho (41). We confirmed that RPTECs
express �Klotho (Fig. 6A). We then assessed the involvement of�Klotho as an FGFR1 cofactor to support FGF23 effects onphosphate transport. We envisioned that disrupting �Klothowould interfere with FGF23 actions but not those of PTH. Sta-ble formation of the FGF23-FGFR1-�Klotho complex andFGFR dimerization is regulated by sulfated glycosaminoglycans(4). Sodium chlorate inhibits proteoglycan sulfation by remov-ing cell surface glycosaminoglycans without affecting proteinsynthesis or other posttranslational modifications (55). Over-night treatment of RPTECs with 50 mM NaClO3 suppressedFGF23 but not PTH effects on phosphate transport in RPTECs(Fig. 6B). These data are consistent with the constitutive andsufficient expression of �Klotho by RPTECs and its require-ment for FGF23-mediated signaling and NPT2A-dependentphosphate transport and with previous work showing thatNaClO3 reversibly interferes with FGF23 action (4).
Recent work shows that unliganded FGFR1 is able to formhomodimers or heterodimerize with �Klotho. These receptorcomplexes undergo conformational changes upon ligand occu-pancy (56). To establish directly its cofactor role in FGF23-sensitive phosphate transport, we knocked down �Klotho inRPTECs and determined its effect on FGF23 and PTH action onphosphate uptake. Three different siRNAs were screened.siRNA3 decreased �Klotho expression by 80% (Fig. 7A), andthis was associated with the most pronounced disruption withthe inhibition of phosphate uptake by FGF23 (Fig. 7B). Theconverse was also true insofar as siRNA1, which failed to reduce�Klotho expression, had no discernable inhibitory action onFGF23-sensitive phosphate transport. Thus, the extent of�Klotho knockdown corresponded to the degree of suppres-sion of FGF23-sensitive phosphate transport. As predicted,�Klotho knockdown did not interfere with PTH inhibition ofphosphate uptake (Fig. 7B), underscoring the distinct signalingand cofactor requirements for FGF23 and PTH actions. Theresults also imply that �Klotho acts specifically and autono-mously on FGF23.
Contrary to these findings, a recent study suggested thatexogenous recombinant �Klotho binds PTH, thereby interfer-ing with PTH binding and signaling (57). Although we can not
FIGURE 4. Signaling pathways mediating PTH and FGF23 effects on phosphate transport. RPTECs were treated for 2 h with 100 nM PTH(1–34) (A) or FGF23(B) in the presence or absence of the specified inhibitors: BisI (PKC), GSK-650394 (SGK-1), H89 (PKA), and PD (PD-98059, ERK1/2). Inhibitors were used at 1 �M
except for GSK, where 10 �M was employed. Phosphate uptake was measured as before. Data represent the mean � S.E. (error bars) of n � 6 experiments. *, p �0.05; **, p � 0.01 versus PTH or FGF23.
FIGURE 5. Effect of FGFR1 exon 9 and exon 8 knockdown on FGF23-regu-lated phosphate transport. A, FGFR1 knockdown by siRNA for exon 8 (si8),exon 9 (si9), or a scrambled control (scr) was assessed by immunoblottingusing antibody that specifically recognizes FGFR1c. A representative result ispresented. B, quantification of si8 and si9 knockdown of FGFR1c. Data repre-sent the mean � S.E. (error bars) of n � 4 experiments. C, siRNA effects onFGF23-sensitive phosphate transport. Data represent the mean � S.E. of n �4 experiments. *, p � 0.05; **, p � 0.01.
PTH and FGF23 Signaling and Function
18636 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 291 • NUMBER 36 • SEPTEMBER 2, 2016
by guest on Decem
ber 26, 2019http://w
ww
.jbc.org/D
ownloaded from

speak to the effects of exogenous Klotho on PTH action onphosphate transport, blocking FGFR dimerization or down-regulating �Klotho selectively impaired FGF23 actions withoutinterfering with the inhibitory effect of PTH on phosphatetransport. An alternative explanation for the apparent inhibi-tory effect of Klotho on PTH action may be ascribed to signalingcross-talk between GPCRs and receptor tyrosine kinases thatarises from stimulation of ERK1/2 and endocytosis of unligan-ded receptor (58).
PTH and FGF23 acting through their respective GPCR andreceptor tyrosine kinase, stimulate distinct signaling pathways,but both require NHERF1 to inhibit phosphate transport (12,17). This raised the hypothesis that the signaling events initi-ated at PTHR and FGFR1 converge on NHERF1 to facilitateendocytosis and inhibit NPT2A-dependent phosphate trans-port. According to this view, NHERF1 knockdown should dis-rupt the actions of both FGF23 and PTH. Consistent with thisprediction, shNHERF1 (shN1) reduced NHERF1 expression by80% (Fig. 8A) and concomitantly virtually abolished both PTHand FGF23-inhibitable phosphate transport (Fig. 8B). Thesefindings support the idea that although the actions of FGF23and PTH stem from different classes of membrane receptors,their effects intersect at the level of NHERF1. This is compatiblewith the requirement for NHERF1 to mediate the inhibitoryactions of both PTH and FGF23 on NPT2A (12, 17, 20). Phos-phorylation of NHERF1 results in disassembly of the NPT2A-NHERF1-ezrin complex (27), internalization and down-regula-tion of NPT2A, and cessation of phosphate transport.
NHERF1 harbors 38 Ser and Thr residues. Identifying theparticular residues phosphorylated following activation ofPTHR and FGFR1 in a native cell model will be essential tounderstand the structural determinants and sites of PTH- andFGF23-dependent NHERF1 post-translational modifications.
Treatment with PTH or FGF23 for 2 h down-regulatesNPT2A in RPTECs (Fig. 9A). After PTH or FGF23 exposure,NPT2A was less concentrated at the apical pole of the cell, with
FIGURE 6. �Klotho expression and function. A, �Klotho is expressed byRPTECs. Shown is an immunoblot of 20 �g of protein lysate duplicate wellsprobed with rabbit polyclonal anti-Klotho antibody. Molecular mass markersrepresenting 50 and 75 kDa are shown. B, chlorate treatment of RPTECs inter-fered with FGF23 but not PTH inhibition of phosphate transport. RPTECs weretreated overnight with 50 mM NaClO4, followed by a 2-h treatment with 100nM PTH(1–34) or FGF23, as indicated. Phosphate uptake was measured asdescribed under “Experimental Procedures.” Data represent the mean � S.E.(error bars) of n � 4 independent experiments. Data were normalized for eachexperiment, where phosphate uptake under control, untreated conditionswas defined as 0% inhibition. **, p � 0.01 versus FGF23 alone.
FIGURE 7. siRNA �Klotho knockdown blocks FGF23- but not PTH-inhibit-able phosphate transport. �Klotho expression was knocked down in RPT-ECs using siRNA as described under “Experimental Procedures.” A, knock-down of �Klotho was assessed by immunoblotting. A representativeexperiment is depicted. The data for three siRNA duplexes plus a scrambledcontrol (scr) are shown. B, the effects of �Klotho knockdown on PTH- andFGF23-dependent phosphate transport were measured. Data represent themean � S.E. (error bars) of n � 5 experiments. Data were normalized for eachexperiment, where phosphate uptake under control, untreated conditionswas defined as 0% inhibition. **, p � 0.01; ***, p � 0.001 versus scrambled.
FIGURE 8. shRNA NHERF1 knockdown inhibits FGF23- and PTH-regulatedphosphate transport. NHERF1 expression was knocked down in RPTECs byshRNA transfection as described under “Experimental Procedures.” A, knock-down of NHERF1 was assessed by immunoblotting. A representative experi-ment is depicted. The data for NHERF1 shRNA and a scrambled control (scr)are shown. B, effects of NHERF1 knockdown on PTH- and FGF23-dependentphosphate transport were assessed. Data represent the mean � S.E. (errorbars) of n � 4 experiments. Data were normalized for each experiment, wherephosphate uptake under control, untreated conditions was defined as 0%inhibition. **, p � 0.01 versus FGF23 or PTH.
PTH and FGF23 Signaling and Function
SEPTEMBER 2, 2016 • VOLUME 291 • NUMBER 36 JOURNAL OF BIOLOGICAL CHEMISTRY 18637
by guest on Decem
ber 26, 2019http://w
ww
.jbc.org/D
ownloaded from

a significant amount now present in the cytoplasm (Fig. 9B). Incontrast, NHERF1 remained associated with the apical mem-branes and showed diminished colocalization with NPT2A. Aphenomenologically similar process has been described forPTH action on PTHR trafficking, where PTHR is endocytosedwhile NHERF1 remains at the cell surface (59).
Studies of FGF23 action on phosphate transport using opossumkidney cells have proven controversial due to variable results (4, 50,60, 61). The discrepancies among these reports and the work heremay originate from the multiple available opossum kidney cellsubtypes (62) as well as the uncertainty regarding FGFR1 and itsproteoglycan requirement (63, 64). The present findings under-score the robust fidelity of RPTECs in recapitulating the hormonesensitivity and phosphate transport phenotype of native proximaltubules. RPTECs constitutively express all of the known compo-nents that regulate the renal phosphate transport mechanism,including PTHR and FGFR1c, along with its regulatory respectivecofactor, and trans-acting elements that respond to PTH andFGF23. Thus, RPTECs should prove valuable for further anddetailed studies of the mechanism of FGF23 and PTH action incells from a eutherian mammal.
In summary, the present findings establish that PTH andFGF23 inhibit phosphate uptake through distinct mechanismsthat impinge on a final common effector. Ligand binding of thePTHR stimulates PKA, PKC, and ERK1/2. FGF23, workingthrough FGFR1c and �Klotho, stimulates SGK1 and ERK. Thetwo signaling cascades converge at NHERF1. The present stud-ies demonstrate that �Klotho is expressed in RPTECs, acting ina cis manner that is required for FGF23 but not PTH action.Also, we clarify that FGFR-�1c is the splice variant expressed by
proximal tubule cells that, together with �Klotho, serves as theFGF23 receptor. We also present data consistent with the viewthat the final splicing decision for FGFR1c expression involves anovel cytoplasmic mechanism that may be a model for othergrowth factor receptors.
Experimental Procedures
Peptides—Human [Nle8,18,Tyr34]PTH(1–34) was purchasedfrom Bachem (H9110). Recombinant human R179Q-FGF23(25–251) (referred to henceforth as FGF23), which isresistant to furin cleavage and inactivation, was obtained fromR&D Systems (2604-FG-025). The C-terminal FGF23(180 –251) fragment was a generous gift from Dr. Moosa Mohammadi(New York University School of Medicine).
Inhibitors—The PKA inhibitor H-89 (B1427), SGK1 inhibitorGSK-650394 (SML0773), PKC inhibitor Bis-I (B6292), and theERK1/2 inhibitor PD98059 (P215) were purchased from Sigma.Sodium chlorate (catalog no. 403016) and other routine re-agents were from Sigma.
Antibodies—Rabbit polyclonal anti-NHERF1 (ab3452), mono-clonal anti-NHERF1 (Abcam ab31111), rabbit polyclonal anti-FGFR1 (ab10646), and rabbit polyclonal anti-Klotho (ab98111)antisera were purchased from Abcam. Monoclonal anti-FGFR1c(47) was acquired under license from MedImmune.
Cell Lines and Cell Culture—Human renal proximal tubuleepithelial cells immortalized with hTERT (RPTECs) (11) wereobtained from ATCC under license from Geron Corp. They werecultured in defined medium (DMEM/F-12 (Mediatech, 10-090-CV) supplemented with 5 pM triiodo-L-thyronine, 10 ng/mlrecombinant human epidermal growth factor, 25 ng/ml prosta-glandin E1, 3.5 �g/ml ascorbic acid, 1 mg/ml insulin, 0.55 mg/mltransferrin, 0.5 �g/ml sodium selenite, 25 ng/ml hydrocortisone)plus 1% penicillin and streptomycin and 0.1 mg/ml G418.
Phosphate Uptake—RPTECs were seeded on 12-well plates.When the cells reached confluence (2–3 days after passaging),they were treated with 100 nM PTH(1–34) or FGF23 incell culture medium. After 2 h, the hormone-supplementedmedium was aspirated, and the wells were washed three times with1 ml of sodium-replete wash buffer (140 mM NaCl, 4.8 mM KCl, 1.2mM MgSO4, 0.1 mM KH2PO4, 10 mM HEPES, pH 7.4). The cellswere incubated with 1 �Ci of [32P]orthophosphate (PerkinElmerLife Sciences, NEX053) in 1 ml of sodium-replete wash buffer for10 min. Phosphate uptake was terminated by placing the plate onice and rinsing the cells three times with sodium-free wash buffer(140 mM N-methyl-D-glucamine, 4.8 mM KCl, 1.2 mM MgSO4, 0.1mM KH2PO4, 10 mM HEPES, pH 7.4). The cells in each well wereextracted overnight at 4 °C using 500 �l of 1% Triton X-100(Sigma). A 250-�l aliquot was counted in a Beckmann CoulterLS6500 scintillation counter. Data were normalized to phosphateuptake under control conditions defined as 100%.
Immunoblotting—Immunoblotting was performed as de-scribed (12). RPTECs were lysed with 1% Nonidet P-40 (50 mM
Tris, 150 mM NaCl, 5 mM EDTA, 1% Nonidet P-40) supple-mented with protease inhibitor mixture I (Calbiochem). Lysiswas performed for 15 min on ice. Solubilized materials wereresolved on 10% SDS-polyacrylamide gels and transferred toImmobilon-P membranes (Millipore) using the semidrymethod (Bio-Rad). Membranes were blocked overnight at 4 °C
FIGURE 9. PTH and FGF23 down-regulation of NPT2A. A, NPT2A proteinlevels in RPTECs after a 2-h treatment with PTH or FGF23. A representativeexperiment is presented. B, immunofluorescence experiments depicting thechange in localization of NPT2A in response to PTH or FGF23 treatment. RPT-ECs were left untreated (control) or treated for 2 h with 100 nM PTH or FGF23.Green, NPT2A; red, NHERF1; blue, nuclei. The x-z plane is depicted.
PTH and FGF23 Signaling and Function
18638 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 291 • NUMBER 36 • SEPTEMBER 2, 2016
by guest on Decem
ber 26, 2019http://w
ww
.jbc.org/D
ownloaded from

with 5% nonfat dried milk in Tris-buffered saline plus Tween 20(TBST) and incubated with the indicated antibodies (poly-clonal anti-NHERF1 at 1:3000, polyclonal anti-Klotho at1:3000, polyclonal anti-FGFR1 at 1:3000, monoclonal anti-FGFR1c at 1:500) overnight at 4 °C. The membranes werewashed four times for 10 min in TBST and then incubated withgoat anti-rabbit IgG (for anti-NHERF, anti-Klotho, and anti-FGFR1 primary antibodies) or goat anti-human IgG (for anti-FGFR1c primary antibody) conjugated to horseradish peroxi-dase at a 1:5000 dilution for 1 h at room temperature.Membranes were washed four times for 10 min in TBST. Pro-tein bands were detected by Luminol-based enhanced chemi-luminescence (EMD Millipore WBKLS0500).
Immunolocalization—RPTECs were grown to confluence ina 10-cm dish, trypsinized, washed with sterile PBS, and centri-fuged at 600 � g, and the cell pellet was resuspended in 16 ml ofmedium. Polycarbonate Transwell filters (12-mm diameter;Corning Costar 3401) were coated with a 0.2-ml solution oftype IV collagen (5 mg of Sigma type VI collagen dissolved in anaqueous solution of 0.2% (v/v) glacial acetic acid), diluted 1:10in 10 mM sodium carbonate, pH 9.0, buffer. Following a 60-minincubation at room temperature, the collagen solution wasaspirated, and the Transwell filters were rinsed with PBS andallowed to dry in a cell culture hood. A 0.5-ml aliquot of the cellsuspension was placed in the upper compartment of the coatedTranswell filters, whereas 1.2 ml was added to the lower cham-ber. The cells were incubated at 37 °C in a cell culture incubator,equilibrated with 95% O2, 5% CO2 for 7–10 days. The culturemedium was changed every 3 days by aspirating the two cham-bers and adding 0.5 ml of medium to the upper chambermedium and 1.2 ml of medium to the lower chamber. Cellscultured on Transwell filters were treated with PTH or FGF23on both basolateral and apical surfaces, as indicated, and themedium was aspirated and replaced with 100 mM sodium caco-dylate buffer, pH 7.4, containing 4% (v/v) paraformaldehyde for30 min at room temperature. The fixative was aspirated andreplaced with quench solution (20 mM glycine, pH 8.0, and 75mM NH4Cl dissolved in PBS). The filters were subsequentlyplaced in block solution (0.1% (v/v) saponin, 1% (v/v) teleostfish skin gelatin, dissolved in PBS) containing 5% (v/v) horseserum. The cells were incubated overnight at 4 °C with mousemonoclonal anti-NHERF1 antibody (diluted 1:30 in block solu-tion) and one of three primary anti-Npt2a antibodies (diluted1:200 in block solution): rabbit anti-Npt2a antibody (a gift fromDr. Mark Knepper, National Institutes of Health) or goat anti-Npt2a (Santa Cruz, sc-33928). After three washes with blockingsolution, the cells were incubated for 1 h at room temperaturewith a mixture of ToPro3 (diluted 1:1000; Life Technologies)
minimal cross-reacting donkey anti-rabbit (or goat) IgG sec-ondary antibody conjugated with Alexa488 (diluted 1:200) andwith donkey anti-mouse IgG secondary antibody conjugatedwith CY3 (diluted 1:200). Following three washes with blockingsolution and then PBS, the filters were postfixed with 4% (v/v)paraformaldehyde (in cacodylate buffer) for 10 min, rinsed withPBS, removed from the holders, and mounted on glass slidesusing Slowfade mounting medium (ThermoFisher). Confocalimages of NPT2A localization were obtained using a Leica CW-STED confocal microscope (in normal confocal mode; LeicaMicrosystems Inc., Buffalo Grove, IL), outfitted with a �63glycerol objective (numerical aperture � 1.3) and low noisehybrid detectors. The captured images were contrast-correctedusing Volocity (PerkinElmer Life Sciences), exported as TIFFfiles, and assembled in Adobe Illustrator.
PTHR localization was performed in an analogous fashion onRPTECs grown on coverslips as described (27) using an anti-body raised against the C terminus of the human PTHR (65).
siRNA Knockdown—siRNA for �Klotho knockdown waspurchased from Integrated DNA Technologies (Coralville, IA).HuSH shRNA for NHERF1 knockdown was obtained from Ori-gene (TR309271). Dicer substrate short interfering RNAs (Dsi-RNAs) for knockdown of FGFR1b (exon 8) and FGFR1c (exon9) were designed using the IDTDsi-RNA tool. The Dsi-RNAsfor FGFR1b were rGrGrArGrUrUrArArUrArCrCrArCr-CrGrArCrArArArGrAGA (sense) and rUrCrUrCrUrUrUr-GrUrCrGrGrUrGrGrUrArUrUrArArCrUrCrCrArG (antisense).The Dsi-RNAs for FGFR1c were rGrUrGrGrUrArCrCrArAr-GrArArGrArGrUrGrArCrUrUrCCA (sense) and rUrGrGrArAr-GrUrCrArCrUrCrUrUrCrUrUrGrGrUrArCrCrArCrUrC (anti-sense). siRNA, Dsi-RNA, and shRNA were transfected intoRPTECs using Lipojet (100468, SignaGen Laboratories, Gaithers-burg, MD). Cells were transfected on 60-mm dishes. 48 h aftertransfection, the cells were trypsinized and passaged onto 12-wellplates for phosphate uptake assays. Protein lysates were extractedfrom a 100-�l aliquot from these cells to assess by immunoblottingthe extent of knockdown.
RT-PCR—Total RNA was isolated using TRIzol (Invitrogen).First strand cDNA synthesis was completed using the Accu-script high fidelity first strand cDNA synthesis kit (Agilent,200820). Amplification was accomplished using Phusion HFPolymerase (New England Biolabs, M0350). Exon-spanningprimers (Table 1) were designed to fingerprint the specificFGFR1 subtype (�/�) and alternatively spliced isoforms presentin RPTECs. For FGFR1b, we used a forward primer that spansthe exon 7/8 boundary and a reverse primer that spans the exon10/8 boundary. For FGFR1c, we employed a forward primerthat spans the exon 7/9 boundary and a reverse primer that
TABLE 2FGFR Primers
Gene Orientation Primer pairs Accession number Product size
bpFGFR1 5�3 3� CTCGGGACAGACTGGTCTTAGG NM_023110.2 150
3�3 5� CGTCCGACTTCAACATCTTCACFGFR3 5�3 3� GACCGAGGACAACGTGATGA NM_000142.4 160
3�3 5� TGAGTGTAGACTCGGTCAAACAAGFGFR4 5�3 3� CCCTCGAATAGGCACAGTTACC NM_002011.4 120
3�3 5� AGCGGAACTTGACGGTGTTC
PTH and FGF23 Signaling and Function
SEPTEMBER 2, 2016 • VOLUME 291 • NUMBER 36 JOURNAL OF BIOLOGICAL CHEMISTRY 18639
by guest on Decem
ber 26, 2019http://w
ww
.jbc.org/D
ownloaded from

spans the exon 10/9 boundary. Primers were purchased fromIntegrated DNA Technologies. A second series of previouslypublished primer sequences (Table 2) (66) was used to identifyFGFR isoforms expressed in RPTECs. PCRs were performed ona PCR Sprint thermal cycler (Hybaid) with a protocol consistingof 95 °C for 5 min, followed by 40 cycles at 95 °C for 10 s; 61 °Cfor 10 s, 72 °C for 30 s; 72 °C for 10 min and hold at 4 °C. ThePCR was analyzed on a 2% agarose gel, and the relevant bandswere excised, purified, and sequenced.
Mass Spectrometry—RPTEs were grown to confluence onthree 15-cm dishes. On ice, the cells were washed three timeswith 5 ml of cold PBS. The cells were then scraped in 5 ml of PBSand transferred to a 50-ml centrifuge tube. The cells were pel-leted at 200 � g at 4 °C for 4 min in a Beckman CoulterAllegraX-12R centrifuge. The supernatant was discarded, andthe pellet was resuspended in a low salt buffer containing 10 mM
HEPES, pH 7.5, 1.5 mM MgCl2, 10 mM KCl, and protease inhib-itors. The sample was sonicated in a Fisher FB120 sonicator for1 min (8 s on, 16 s off, 30% power) and centrifuged for 30 min at13,000 rpm at 4 °C in an Eppendorf 5415R centrifuge. Theresulting pellet was resuspended in 5 ml of high salt buffer towash the cell membrane (4.25 ml of low salt buffer supple-mented with 0.75 ml of 5 M NaCl) and centrifuged again for 30min at 13,000 rpm at 4 °C. The pellet was resuspended in 1 ml oflow salt buffer and sonicated on ice for 8 s at 30% power. Thesample was then centrifuged at 2000 rpm for 10 min at 4 °C toremove unbroken cells and free nuclei. After another highspeed centrifugation, the sample was divided into pellet andsupernatant fractions. The pellet was resuspended in 100 �l ofLaemmli buffer and resolved on an 8% SDS-polyacrylamide gel.In parallel, a small amount of pellet and supernatant fractionswas examined by Western blotting to ensure that the protein ofinterest was in the pellet sample. The protein in cut gel pieceswas reduced by 10 mM DTT and alkylated by 55 mM iodoacet-amide, and in-gel digestion by trypsin was performed at 37 °Covernight. The digested peptides were resuspended in 0.1% for-mic acid and subjected to LC-MS/MS analysis using the LTQOrbitrap Velos mass spectrometer (Thermo Scientific).
Statistics—Statistical analysis was performed using GraphPadPrism version 6. Single comparisons between groups were ana-lyzed by analysis of variance with post hoc testing using the Bon-feronni multiple comparisons test, whereas repeated measureswere analyzed using a paired t test. Differences between three ormore groups were analyzed by two-way analysis of variance andpost hoc testing using Tukey’s multiple comparisons test. Differ-ences greater than p � 0.05 were assumed to be significant.
Author Contributions—P. A. F., W. B. S., O. A. W., and G. L. A.designed the study, analyzed results, and wrote the paper. W. B. S.,K. X., Q. Z., G. W. R., L. I. G., and Y. R. performed the describedexperiments.
Acknowledgments—We are grateful to Dr. Moosa Mohammadi (NewYork University School of Medicine) for generously sharing reagentsand to Dr. Iris Lindberg (University of Maryland) for vital guidanceon preparing FGF23. MedImmune supplied FGFR1c antiserum, andwe are grateful for their interest and support.
References1. Bergwitz, C., and Juppner, H. (2010) Regulation of phosphate homeostasis
by PTH, vitamin D, and FGF23. Annu. Rev. Med. 61, 91–1042. Gattineni, J. (2014) Inherited disorders of calcium and phosphate metab-
olism. Curr. Opin. Pediatr. 26, 215–2223. Biber, J., Malmstrom, K., Reshkin, S., and Murer, H. (1990) Phosphate
transport in established renal epithelial cell lines. Methods Enzymol. 191,494 –505
4. Yu, X., Ibrahimi, O. A., Goetz, R., Zhang, F., Davis, S. I., Garringer, H. J.,Linhardt, R. J., Ornitz, D. M., Mohammadi, M., and White, K. E. (2005)Analysis of the biochemical mechanisms for the endocrine actions of fi-broblast growth factor-23. Endocrinology 146, 4647– 4656
5. Weinman, E. J., and Lederer, E. D. (2012) PTH-mediated inhibition of therenal transport of phosphate. Exp. Cell Res. 318, 1027–1032
6. Murer, H., Forster, I., Hernando, N., and Biber, J. (2008) Proximal tubularhandling of phosphate. in Seldin and Giebisch’s The Kidney Physiology andPathophysiology (Alpern, R. J., and Hebert, S. C., eds) pp. 1979 –1987,Elsevier, Burlington, MA
7. Cole, J. A., Eber, S. L., Poelling, R. E., Thorne, P. K., and Forte, L. R. (1987)A dual mechanism for regulation of kidney phosphate transport by para-thyroid hormone. Am. J. Physiol. 253, E221–E227
8. Lederer, E. D., Sohi, S. S., Mathiesen, J. M., and Klein, J. B. (1998) Regula-tion of expression of type II sodium-phosphate cotransporters by proteinkinases A and C. Am. J. Physiol. 275, F270 –F277
9. Pfister, M. F., Forgo, J., Ziegler, U., Biber, J., and Murer, H. (1999) cAMP-dependent and -independent downregulation of type II Na-Pi cotrans-porters by PTH. Am. J. Physiol. 276, F720 –F725
10. Traebert, M., Volkl, H., Biber, J., Murer, H., and Kaissling, B. (2000) Lu-minal and contraluminal action of 1–34 and 3–34 PTH peptides on renaltype IIa Na-Pi cotransporter. Am. J. Physiol. Renal Physiol. 278,F792–F798
11. Capuano, P., Bacic, D., Roos, M., Gisler, S. M., Stange, G., Biber, J.,Kaissling, B., Weinman, E. J., Shenolikar, S., Wagner, C. A., and Murer,H. (2007) Defective coupling of apical PTH receptors to phospholipaseC prevents internalization of the Na�-phosphate cotransporter NaPi-IIa in Nherf1-deficient mice. Am. J. Physiol. Cell Physiol. 292,C927–C934
12. Cunningham, R., Xiaofei, E., Steplock, D., Shenolikar, S., and Weinman,E. J. (2005) Defective PTH regulation of sodium-dependent phosphatetransport in NHERF-1�/� renal proximal tubule cells and wild-type cellsadapted to low phosphate media. Am. J. Physiol. Renal Physiol. 289,F933–F938
13. Guo, J., Song, L., Liu, M., Segawa, H., Miyamoto, K., Bringhurst, F. R.,Kronenberg, H. M., and Juppner, H. (2013) Activation of a non-cAMP/PKA signaling pathway downstream of the PTH/PTHrP receptor is essen-tial for a sustained hypophosphatemic response to PTH infusion in malemice. Endocrinology 154, 1680 –1689
14. Nagai, S., Okazaki, M., Segawa, H., Bergwitz, C., Dean, T., Potts, J. T., Jr.,Mahon, M. J., Gardella, T. J., and Juppner, H. (2011) Acute down-regula-tion of sodium-dependent phosphate transporter NPT2a involves pre-dominantly the cAMP/PKA pathway as revealed by signaling-selectiveparathyroid hormone analogs. J. Biol. Chem. 286, 1618 –1626
15. Hu, M. C., Shi, M., Zhang, J., Pastor, J., Nakatani, T., Lanske, B., Razzaque,M. S., Rosenblatt, K. P., Baum, M. G., Kuro-o, M., and Moe, O. W. (2010)Klotho: a novel phosphaturic substance acting as an autocrine enzyme inthe renal proximal tubule. FASEB J. 24, 3438 –3450
16. Gattineni, J., Bates, C., Twombley, K., Dwarakanath, V., Robinson, M. L.,Goetz, R., Mohammadi, M., and Baum, M. (2009) FGF23 decreases renalNaPi-2a and NaPi-2c expression and induces hypophosphatemia in vivopredominantly via FGF receptor 1. Am. J. Physiol. Renal Physiol. 297,F282–F291
17. Andrukhova, O., Zeitz, U., Goetz, R., Mohammadi, M., Lanske, B., andErben, R. G. (2012) FGF23 acts directly on renal proximal tubules to in-duce phosphaturia through activation of the ERK1/2-SGK1 signalingpathway. Bone 51, 621– 628
18. Hernando, N., Deliot, N., Gisler, S. M., Lederer, E., Weinman, E. J., Biber,J., and Murer, H. (2002) PDZ-domain interactions and apical expression of
PTH and FGF23 Signaling and Function
18640 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 291 • NUMBER 36 • SEPTEMBER 2, 2016
by guest on Decem
ber 26, 2019http://w
ww
.jbc.org/D
ownloaded from

type IIa Na/Pi cotransporters. Proc. Natl. Acad. Sci. U.S.A. 99,11957–11962
19. Mamonova, T., Kurnikova, M., and Friedman, P. A. (2012) Structural basisfor NHERF1 PDZ domain binding affinity. Biochemistry 51, 3110 –3120
20. Weinman, E. J., Steplock, D., Shenolikar, S., and Biswas, R. (2011) Fibro-blast growth factor-23-mediated inhibition of renal phosphate transportin mice requires sodium-hydrogen exchanger regulatory factor-1 (NH-ERF-1) and synergizes with parathyroid hormone. J. Biol. Chem. 286,37216 –37221
21. Shenolikar, S., Voltz, J. W., Minkoff, C. M., Wade, J. B., and Weinman, E. J.(2002) Targeted disruption of the mouse NHERF-1 gene promotes inter-nalization of proximal tubule sodium-phosphate cotransporter type IIaand renal phosphate wasting. Proc. Natl. Acad. Sci. U.S.A. 99,11470 –11475
22. Morales, F. C., Takahashi, Y., Kreimann, E. L., and Georgescu, M. M.(2004) Ezrin-radixin-moesin (ERM)-binding phosphoprotein 50 orga-nizes ERM proteins at the apical membrane of polarized epithelia. Proc.Natl. Acad. Sci. U.S.A. 101, 17705–17710
23. Karim, Z., Gerard, B., Bakouh, N., Alili, R., Leroy, C., Beck, L., Silve, C.,Planelles, G., Urena-Torres, P., Grandchamp, B., Friedlander, G., and Prie,D. (2008) NHERF1 mutations and responsiveness of renal parathyroidhormone. N. Engl. J. Med. 359, 1128 –1135
24. Weinman, E. J., and Lederer, E. D. (2012) NHERF-1 and the regulation ofrenal phosphate reabsoption: a tale of three hormones. Am. J. Physiol.Renal Physiol. 303, F321–F327
25. Weinman, E. J., Steplock, D., Zhang, Y., Biswas, R., Bloch, R. J., and She-nolikar, S. (2010) Cooperativity between the phosphorylation of Thr95 andSer77 of NHERF-1 in the hormonal regulation of renal phosphate trans-port. J. Biol. Chem. 285, 25134 –25138
26. Potter, B. A., Ihrke, G., Bruns, J. R., Weixel, K. M., and Weisz, O. A. (2004)Specific N-glycans direct apical delivery of transmembrane, but not solu-ble or glycosylphosphatidylinositol-anchored forms of endolyn in Madin-Darby canine kidney cells. Mol. Biol. Cell 15, 1407–1416
27. Wang, B., Means, C. K., Yang, Y., Mamonova, T., Bisello, A., Altschuler,D. L., Scott, J. D., and Friedman, P. A. (2012) Ezrin-anchored PKA coor-dinates phosphorylation-dependent disassembly of a NHERF1 ternarycomplex to regulate hormone-sensitive phosphate transport. J. Biol.Chem. 287, 24148 –24163
28. Wieser, M., Stadler, G., Jennings, P., Streubel, B., Pfaller, W., Ambros, P.,Riedl, C., Katinger, H., Grillari, J., and Grillari-Voglauer, R. (2008) hTERTalone immortalizes epithelial cells of renal proximal tubules withoutchanging their functional characteristics. Am. J. Physiol. Renal Physiol.295, F1365–F1375
29. Wang, E., Brown, P. S., Aroeti, B., Chapin, S. J., Mostov, K. E., and Dunn,K. W. (2000) Apical and basolateral endocytic pathways of MDCK cellsmeet in acidic common endosomes distinct from a nearly-neutral apicalrecycling endosome. Traffic 1, 480 – 493
30. Apodaca, G., Katz, L. A., and Mostov, K. E. (1994) Receptor-mediatedtranscytosis of IgA in MDCK cells is via apical recycling endosomes. J. CellBiol. 125, 67– 86
31. Wade, J. B., Liu, J., Coleman, R. A., Cunningham, R., Steplock, D. A.,Lee-Kwon, W., Pallone, T. L., Shenolikar, S., and Weinman, E. J. (2003)Localization and interaction of NHERF isoforms in the renal proximaltubule of the mouse. Am. J. Physiol. Cell Physiol. 285, C1494 –C1503
32. Amizuka, N., Lee, H. S., Kwan, M. Y., Arazani, A., Warshawsky, H., Hendy,G. N., Ozawa, H., White, J. H., and Goltzman, D. (1997) Cell-specificexpression of the parathyroid hormone (PTH)/PTH-related peptide re-ceptor gene in kidney from kidney-specific and ubiquitous promoters.Endocrinology 138, 469 – 481
33. Ba, J., Brown, D., and Friedman, P. A. (2003) CaSR regulation of PTH-inhibitable proximal tubule phosphate transport. Am. J. Physiol. RenalPhysiol. 285, F1233–F1243
34. Gattineni, J., Alphonse, P., Zhang, Q., Mathews, N., Bates, C. M., andBaum, M. (2014) Regulation of renal phosphate transport by FGF23 ismediated by FGFR1 and FGFR4. Am. J. Physiol. Renal Physiol. 306,F351–F358
35. Lee, J. W., Chou, C. L., and Knepper, M. A. (2015) Deep sequencing in
microdissected renal tubules identifies nephron segment-specific tran-scriptomes. J. Am. Soc. Nephrol. 26, 2669 –2677
36. Urakawa, I., Yamazaki, Y., Shimada, T., Iijima, K., Hasegawa, H., Okawa,K., Fujita, T., Fukumoto, S., and Yamashita, T. (2006) Klotho convertscanonical FGF receptor into a specific receptor for FGF23. Nature 444,770 –774
37. Han, X., Yang, J., Li, L., Huang, J., King, G., and Quarles, L. D. (2016)Conditional deletion of fgfr1 in the proximal and distal tubule identifiesdistinct roles in phosphate and calcium transport. PLoS One 11, e0147845
38. Liu, S., Vierthaler, L., Tang, W., Zhou, J., and Quarles, L. D. (2008) FGFR3and FGFR4 do not mediate renal effects of FGF23. J. Am. Soc. Nephrol. 19,2342–2350
39. Mistry, N., Harrington, W., Lasda, E., Wagner, E. J., and Garcia-Blanco,M. A. (2003) Of urchins and men: evolution of an alternative splicing unitin fibroblast growth factor receptor genes. RNA 9, 209 –217
40. Ornitz, D. M., and Itoh, N. (2015) The fibroblast growth factor signalingpathway. Wiley Interdiscip. Rev. Dev. Biol. 4, 215–266
41. Belov, A. A., and Mohammadi, M. (2013) Molecular mechanisms of fibro-blast growth factor signaling in physiology and pathology. Cold SpringHarb. Perspect. Biol. 10.1101/cshperspect.a015958
42. Gong, S. G. (2014) Isoforms of receptors of fibroblast growth factors.J. Cell. Physiol. 229, 1887–1895
43. Johnson, D. E., Lu, J., Chen, H., Werner, S., and Williams, L. T. (1991) Thehuman fibroblast growth factor receptor genes: a common structural ar-rangement underlies the mechanisms for generating receptor forms thatdiffer in their third immunoglobulin domain. Mol. Cell. Biol. 11,4627– 4634
44. Johnson, D. E., and Williams, L. T. (1993) Structural and functional diver-sity in the FGF receptor multigene family. Adv. Cancer Res. 60, 1– 41
45. Hovhannisyan, R. H., and Carstens, R. P. (2007) Heterogeneous ribonu-cleoprotein m is a splicing regulatory protein that can enhance or silencesplicing of alternatively spliced exons. J. Biol. Chem. 282, 36265–36274
46. Buckley, P. T., Khaladkar, M., Kim, J., and Eberwine, J. (2014) Cytoplasmicintron retention, function, splicing, and the sentinel RNA hypothesis. Wi-ley Interdiscip. Rev. RNA 5, 223–230
47. Lelliott, C. J., Ahnmark, A., Admyre, T., Ahlstedt, I., Irving, L., Keyes, F.,Patterson, L., Mumphrey, M. B., Bjursell, M., Gorman, T., Bohlooly-Y, M.,Buchanan, A., Harrison, P., Vaughan, T., Berthoud, H. R., and Linden, D.(2014) Monoclonal antibody targeting of fibroblast growth factor receptor1c ameliorates obesity and glucose intolerance via central mechanisms.PLoS One 9, e112109
48. Goetz, R., Nakada, Y., Hu, M. C., Kurosu, H., Wang, L., Nakatani, T., Shi,M., Eliseenkova, A. V., Razzaque, M. S., Moe, O. W., Kuro-o, M., andMohammadi, M. (2010) Isolated C-terminal tail of FGF23 alleviates hy-pophosphatemia by inhibiting FGF23-FGFR-Klotho complex formation.Proc. Natl. Acad. Sci. U.S.A. 107, 407– 412
49. Murer, H., and Biber, J. (2010) Phosphate transport in the kidney. J. Neph-rol. 23, S145–S151
50. Yamashita, T., Konishi, M., Miyake, A., Inui, K., and Itoh, N. (2002) Fibro-blast growth factor (FGF)-23 inhibits renal phosphate reabsorption byactivation of the mitogen-activated protein kinase pathway. J. Biol. Chem.277, 28265–28270
51. Friedman, P. A., Coutermarsh, B. A., Kennedy, S. M., and Gesek, F. A.(1996) Parathyroid hormone stimulation of calcium transport is mediatedby dual signaling mechanisms involving PKA and PKC. Endocrinology137, 13–20
52. Kouhara, H., Hadari, Y. R., Spivak-Kroizman, T., Schilling, J., Bar-Sagi, D.,Lax, I., and Schlessinger, J. (1997) A lipid-anchored Grb2-binding proteinthat links FGF-receptor activation to the Ras/MAPK signaling pathway.Cell 89, 693–702
53. Sneddon, W. B., Liu, F., Gesek, F. A., and Friedman, P. A. (2000) ObligateMAP kinase activation in parathyroid hormone stimulation of calciumtransport but not calcium signaling. Endocrinology 141, 4185– 4193
54. Sneddon, W. B., Yang, Y., Ba, J., Harinstein, L. M., and Friedman, P. A.(2007) Extracellular signal-regulated kinase activation by parathyroid hor-mone in distal tubule cells. Am. J. Physiol. Renal Physiol 292,F1028 –F1034
55. Safaiyan, F., Kolset, S. O., Prydz, K., Gottfridsson, E., Lindahl, U., and
PTH and FGF23 Signaling and Function
SEPTEMBER 2, 2016 • VOLUME 291 • NUMBER 36 JOURNAL OF BIOLOGICAL CHEMISTRY 18641
by guest on Decem
ber 26, 2019http://w
ww
.jbc.org/D
ownloaded from

Salmivirta, M. (1999) Selective effects of sodium chlorate treatment on thesulfation of heparan sulfate. J. Biol. Chem. 274, 36267–36273
56. Comps-Agrar, L., Dunshee, D. R., Eaton, D. L., and Sonoda, J. (2015) Un-liganded fibroblast growth factor receptor 1 forms density-independentdimers. J. Biol. Chem. 290, 24166 –24177
57. Takenaka, T., Inoue, T., Miyazaki, T., Hayashi, M., and Suzuki, H. (2016)Xeno-klotho inhibits parathyroid hormone signaling. J. Bone Miner. Res.31, 455– 462
58. Paradis, J. S., Ly, S., Blondel-Tepaz, E., Galan, J. A., Beautrait, A., Scott,M. G., Enslen, H., Marullo, S., Roux, P. P., and Bouvier, M. (2015) Receptorsequestration in response to �-arrestin-2 phosphorylation by ERK1/2governs steady-state levels of GPCR cell-surface expression. Proc. Natl.Acad. Sci. U.S.A. 112, E5160 –E5168
59. Ardura, J. A., Wang, B., Watkins, S. C., Vilardaga, J. P., and Friedman, P. A.(2011) Dynamic Na�-H� exchanger regulatory factor-1 association anddissociation regulate PTH receptor trafficking at membrane microdo-mains. J. Biol. Chem. 286, 35020 –35029
60. Bowe, A. E., Finnegan, R., Jan de Beur, S. M., Cho, J., Levine, M. A., Kumar,R., and Schiavi, S. C. (2001) FGF-23 inhibits renal tubular phosphate trans-port and is a PHEX substrate. Biochem. Biophys. Res. Commun. 284,977–981
61. Shimada, T., Mizutani, S., Muto, T., Yoneya, T., Hino, R., Takeda, S.,
Takeuchi, Y., Fujita, T., Fukumoto, S., and Yamashita, T. (2001) Cloningand characterization of FGF23 as a causative factor of tumor-inducedosteomalacia. Proc. Natl. Acad. Sci. U.S.A. 98, 6500 – 6505
62. Cole, J. A., Forte, L. R., Krause, W. J., and Thorne, P. K. (1989) Clonalsublines that are morphologically and functionally distinct from parentalOK cells. Am. J. Physiol. 256, F672–F679
63. Ibrahimi, O. A., Zhang, F., Hrstka, S. C., Mohammadi, M., and Linhardt,R. J. (2004) Kinetic model for FGF, FGFR, and proteoglycan signal trans-duction complex assembly. Biochemistry 43, 4724 – 4730
64. Goetz, R., Beenken, A., Ibrahimi, O. A., Kalinina, J., Olsen, S. K., Eliseen-kova, A. V., Xu, C., Neubert, T. A., Zhang, F., Linhardt, R. J., Yu, X., White,K. E., Inagaki, T., Kliewer, S. A., Yamamoto, M., et al. (2007) Molecularinsights into the klotho-dependent, endocrine mode of action of fibroblastgrowth factor 19 subfamily members. Mol. Cell. Biol. 27, 3417–3428
65. Lupp, A., Klenk, C., Rocken, C., Evert, M., Mawrin, C., and Schulz, S.(2010) Immunohistochemical identification of the PTHR1 parathyroidhormone receptor in normal and neoplastic human tissues. Eur. J. Endo-crinol. 162, 979 –986
66. Roidl, A., Foo, P., Wong, W., Mann, C., Bechtold, S., Berger, H. J., Streit, S.,Ruhe, J. E., Hart, S., Ullrich, A., and Ho, H. K. (2010) The FGFR4 Y367Cmutant is a dominant oncogene in MDA-MB453 breast cancer cells. On-cogene 29, 1543–1552
PTH and FGF23 Signaling and Function
18642 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 291 • NUMBER 36 • SEPTEMBER 2, 2016
by guest on Decem
ber 26, 2019http://w
ww
.jbc.org/D
ownloaded from

Zhang, Youssef Rbaibi, Ora A. Weisz, Gerard L. Apodaca and Peter A. FriedmanW. Bruce Sneddon, Giovanni W. Ruiz, Luciana I. Gallo, Kunhong Xiao, Qiangmin
Growth Factor-23 Action on NPT2A-mediated Phosphate TransportConvergent Signaling Pathways Regulate Parathyroid Hormone and Fibroblast
doi: 10.1074/jbc.M116.744052 originally published online July 18, 20162016, 291:18632-18642.J. Biol. Chem.
10.1074/jbc.M116.744052Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/291/36/18632.full.html#ref-list-1
This article cites 65 references, 24 of which can be accessed free at
by guest on Decem
ber 26, 2019http://w
ww
.jbc.org/D
ownloaded from