Embed Size (px)
Citation preview

Investigative Ophthalmology & Visual Science, Vol. 30, No. 9, September 1989Copyright © Association for Research in Vision and Ophthalmology
Corneal Nerve Regeneration
Correlation between Morphology and Restoration of Sensitivity
A. Margreef de Leeuw and Kwan Y. Chan
Corneal nerve regeneration was determined in albino rabbits after deepithelialization of the corneausing heptanol. Regeneration was monitored up to 10 weeks by measuring corneal tactile sensitivityusing an esthesiometer and by examining stromal and intraepithelial nerve patterns following goldchloride impregnation and acetylcholinesterase staining. Tactile sensitivity was much reduced, possi-bly absent, up to 2 weeks after wounding. From 2.5-4 weeks, sensitivity recovered rapidly to 60% ofprewounding levels and remained unchanged thereafter. In control corneas, a distinct orientation of thebasoepithelial leashes towards the nasal-most limbus was observed in the central two-thirds of thecornea. Three days after wounding, neurites that were oriented radially towards the wound centerextended into the periphery of the wound area from just beyond the wound margin. At 1 week,regenerating axons were present as single neurites and in the form of modified leashes, mainly at theperiphery of the wound area but also more towards the center. At 3 weeks, neurites, regeneratedleashes and networks of terminals with terminal endings were found throughout the regeneratedepithelium. Regional changes in the orientation of the regenerated leashes were observed also. Nofurther change in the intraepithelial nerve pattern was detectable thereafter up to 10 weeks afterwounding. It was concluded that partial restoration of tactile sensitivity following deepithelialization ofthe cornea is a function of the establishment of a near-normal nerve pattern in the regeneratedepithelium and is correlated with the subnormal neural density observed in a previous study. InvestOphthalmol Vis Sci 30:1980-1990,1989
The cornea is innervated mainly by sensory nervesfrom the ophthalmic branch of the trigeminal nerve;in addition, a minor autonomic innervation (espe-cially of the peripheral cornea) has also been recog-nized.1 The afferent sensory fibers originate from uni-polar neurons located in the anteromedial region ofthe trigeminal ganglion and leave the ganglion in thecombined maxillary/ophthalmic division, branchesof which (the long ciliary nerves) continue as sensoryfibers to the eye.2 Upon entering the suprachoroidalspace, the sensory fibers supply the limbal region, thetrabecular meshwork, and the region of the canal ofSchlemm, and then send branches at midstromaldepth into the cornea from its entire circumference.For a certain distance into the cornea, the nerve fibers
From the Department of Ophthalmology, University of Wash-ington, Seattle, Washington.
Supported by NIH Research Grant No. EY-04538, and in partby EY-01730, HD-02274 and an award from Research to PreventBlindness, Inc.
Submitted for publication: September 1, 1988; accepted Febru-ary 27, 1989.
Reprint requests: A. Margreet de Leeuw, PhD, Department ofOphthalmology, RJ-10, University of Washington, Seattle, WA98195.
contain both myelinated and unmyelinated axons,thus resembling peripheral nerves.3
The nociceptive sensitivity of the cornea for me-chanical and thermal stimuli4 is essential for theproper functioning of the cornea.5 Loss of sensitivitydue to partial or total denervation of the cornea fol-lowing viral infections or ocular surgery may result inchronic desiccation and inflammation of the cornea.Studies in experimental animals suggest that damageto corneal innervation affects epithelial regenerationand corneal metabolism.6'7 Regeneration of cornealnerves and restoration of sensory function in humanappear to be limited.8'9
In previous studies regarding corneal nerve regen-eration following various surgical procedures, onlylimited attempts were made to correlate nerve regen-eration patterns with restoration of sensory func-tion.10"14 The time course of sensory reinnervationfollowing various surgical procedures and the type ofregenerated axons supporting sensory function arenot well studied.
In the present study, we investigated an experimen-tal nerve wounding model of the cornea in whichacute damage to corneal nerves was restricted to in-traepithelial axon terminals. Tactile sensitivity of thewounded corneas was determined and correlated
1980
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933149/ on 05/14/2018

No. 9 CORNEAL NERVE REGENERATION / de Leeuw and Chon 1981
with nerve regeneration patterns visualized by twodifferent staining techniques.
Materials and Methods
Animal Model
All procedures in this study adhered to the ARVOResolution on the Use of Animals in Research.
Male New Zealand albino rabbits (2-2.5 kg) wereanesthetized with halothane and one eye was prop-tosed. Corneal epithelium was removed from a circu-lar area using n-heptanol (heptyl alcohol; EastmanKodak Company, Rochester, NY) according to themethod of Cintron et al.15 In short, a 4 mm diameterdisc of No. 50 Whatman filter paper was fixated on awax plug inside the hollow end of a metal cork-borertube, saturated with heptanol and then carefullybrought into contact with the surface of the corneafor 1 min. The wound site was immediately rinsedwith sterile saline and cell debris removed with a sa-line-moistened cotton tip. After staining with 2% so-dium fluorescein, the size of the wound was mea-sured along the x- and y-axis using caliper and ruler(accuracy within 0.25 mm). The wound was placed inthe center of the cornea, unilaterally, on a randomlyselected side. The contralateral eye served as a con-trol. During the first week, the animals were observeddaily for signs of ocular inflammatory reactions. Twoto five rabbits were sacrificed at 3 days and 1,2,3, 4,5, 6 and 10 weeks after wounding.
Tactile Sensitivity
The corneal sensitivity to a mechanical stimuluswas determined twice weekly for 1 week prior towounding and for 3 weeks thereafter, and then onceweekly for up to 10 weeks after wounding using ahand-held esthesiometer (Cochet and Bonnet type;filament diameter: 0.12 mm; Luneau Ophthalmol-ogy, Paris, France). Sensitivities in the center of thewounded area and at the temporal side, 1-1.5 mmoutside the wound margin (see Fig. 1), were com-pared with sensitivities in equivalent sites in the con-trol eye. The nylon filament, while watching it fromthe side, was brought into contact with the cornea at aright angle, until deflection of the filament was noted.For each filament length, the specific site was touchedten times at 1 sec intervals. The frequency of theblink reflex, which included full and partial blinks,was determined for decreasing filament lengths be-tween 5.0 and 1.0 cm with 0.5 cm decrements. Thesefilament lengths corresponded to a range of deflec-tion forces between 13 and 145 mg (calibration bymanufacturer). The frequency of the corneal blink
N
I
Fig. 1. Diagrammatic illustration of the experimental procedureperformed on rabbit cornea. The central wound area (checkered)was an average of 5 mm diameter. Two sites (small circles) werechosen for sensitivity measurement. For nerve staining, the nasal(N) and temporal (T) periphery was trimmed (shaded), the inferior(I) quarter (striped) was for AChE staining after trisection, and theremaining superior (S) portion was for gold chloride impregnation.
reflex was plotted as a function of the logarithm of thedeflection force.16 A straight line determined by lin-ear regression analysis was fitted to the curve, and thedeflection force corresponding to a 50% response wasdetermined. Corneal sensitivity was defined as thereciprocal of the force (in gram) at which a 50% re-sponse was seen. Sensitivities were plotted as a func-tion of time.
Gold Chloride Impregnation andAcetylcholinesterase Staining
The eyes were rapidly enucleated after marking thenasal limbus with a small needle and the corneas re-moved and placed in fixative for subsequent process-ing. The inferior quadrant was cut out and divided inthree strips of equal width for subsequent acetylcho-linesterase staining (Fig. 1). The remaining corneawas trimmed at the nasal and temporal peripheryand processed for impregnation with gold chloride(Fig. 1).
Corneal nerves were stained with gold chloride es-sentially according to Rozsa and Beuerman.3 The fol-lowing steps were carried out at room temperature.Corneas were fixed in 4% paraformaldehyde in 0.1 Msodium cacodylate buffer (pH 5.5) for 4 min. Corneaswere then placed sequentially in fresh lemon juice(briefly centrifuged to remove solid materials) for 10
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933149/ on 05/14/2018
1982 INVESTIGATIVE OPHTHALMOLOGY & VISUAL SCIENCE / September 1989 Vol. 30
10 -i
1.2 1.4 1.6 1.8 2.0 2.2
LOG DEFLECTION FORCE (mg)
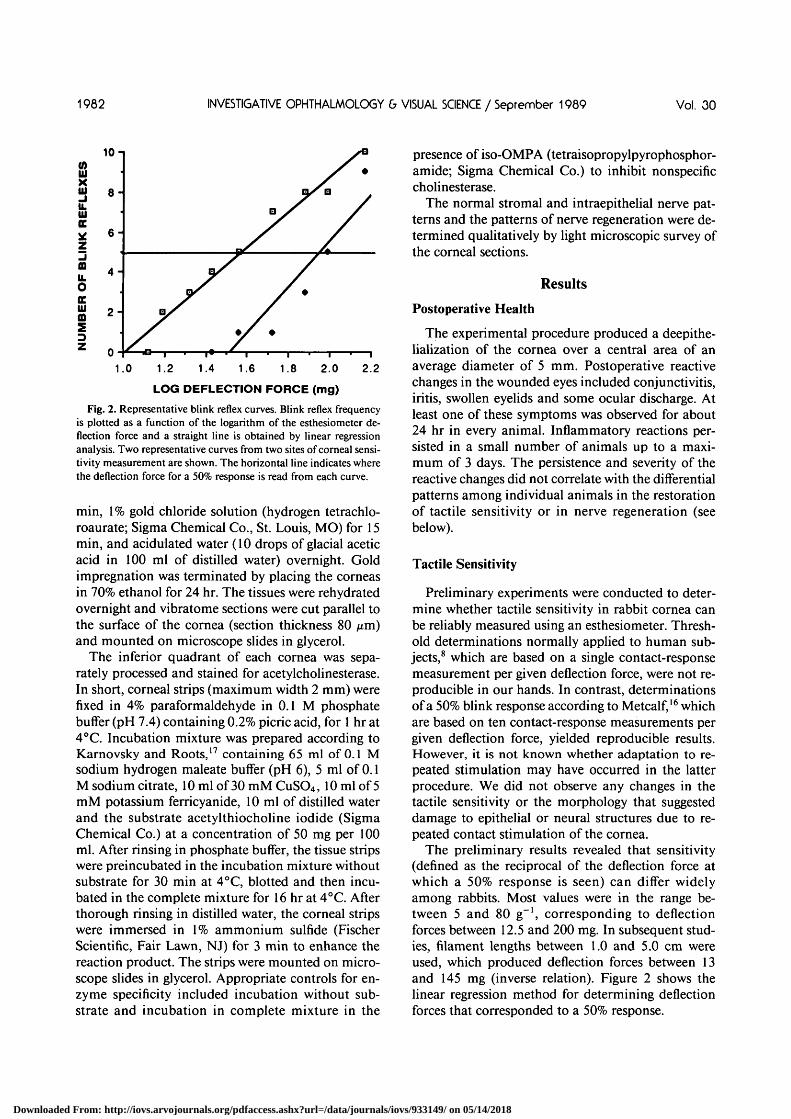
Fig. 2. Representative blink reflex curves. Blink reflex frequencyis plotted as a function of the logarithm of the esthesiometer de-flection force and a straight line is obtained by linear regressionanalysis. Two representative curves from two sites of corneal sensi-tivity measurement are shown. The horizontal line indicates wherethe deflection force for a 50% response is read from each curve.
min, 1% gold chloride solution (hydrogen tetrachlo-roaurate; Sigma Chemical Co., St. Louis, MO) for 15min, and acidulated water (10 drops of glacial aceticacid in 100 ml of distilled water) overnight. Goldimpregnation was terminated by placing the corneasin 70% ethanol for 24 hr. The tissues were rehydratedovernight and vibratome sections were cut parallel tothe surface of the cornea (section thickness 80 nm)and mounted on microscope slides in glycerol.
The inferior quadrant of each cornea was sepa-rately processed and stained for acetylcholinesterase.In short, corneal strips (maximum width 2 mm) werefixed in 4% paraformaldehyde in 0.1 M phosphatebuffer (pH 7.4) containing 0.2% picric acid, for 1 hr at4°C. Incubation mixture was prepared according toKarnovsky and Roots,17 containing 65 ml of 0.1 Msodium hydrogen maleate buffer (pH 6), 5 ml of 0.1M sodium citrate, 10 ml of 30 mM CuSO4, 10 ml of 5mM potassium ferricyanide, 10 ml of distilled waterand the substrate acetylthiocholine iodide (SigmaChemical Co.) at a concentration of 50 mg per 100ml. After rinsing in phosphate buffer, the tissue stripswere preincubated in the incubation mixture withoutsubstrate for 30 min at 4°C, blotted and then incu-bated in the complete mixture for 16 hr at 4°C. Afterthorough rinsing in distilled water, the corneal stripswere immersed in 1% ammonium sulfide (FischerScientific, Fair Lawn, NJ) for 3 min to enhance thereaction product. The strips were mounted on micro-scope slides in glycerol. Appropriate controls for en-zyme specificity included incubation without sub-strate and incubation in complete mixture in the
presence of iso-OMPA (tetraisopropylpyrophosphor-amide; Sigma Chemical Co.) to inhibit nonspecificcholinesterase.
The normal stromal and intraepithelial nerve pat-terns and the patterns of nerve regeneration were de-termined qualitatively by light microscopic survey ofthe corneal sections.
Results
Postoperative Health
The experimental procedure produced a deepithe-lialization of the cornea over a central area of anaverage diameter of 5 mm. Postoperative reactivechanges in the wounded eyes included conjunctivitis,iritis, swollen eyelids and some ocular discharge. Atleast one of these symptoms was observed for about24 hr in every animal. Inflammatory reactions per-sisted in a small number of animals up to a maxi-mum of 3 days. The persistence and severity of thereactive changes did not correlate with the differentialpatterns among individual animals in the restorationof tactile sensitivity or in nerve regeneration (seebelow).
Tactile Sensitivity
Preliminary experiments were conducted to deter-mine whether tactile sensitivity in rabbit cornea canbe reliably measured using an esthesiometer. Thresh-old determinations normally applied to human sub-jects,8 which are based on a single contact-responsemeasurement per given deflection force, were not re-producible in our hands. In contrast, determinationsof a 50% blink response according to Metcalf,16 whichare based on ten contact-response measurements pergiven deflection force, yielded reproducible results.However, it is not known whether adaptation to re-peated stimulation may have occurred in the latterprocedure. We did not observe any changes in thetactile sensitivity or the morphology that suggesteddamage to epithelial or neural structures due to re-peated contact stimulation of the cornea.
The preliminary results revealed that sensitivity(defined as the reciprocal of the deflection force atwhich a 50% response is seen) can differ widelyamong rabbits. Most values were in the range be-tween 5 and 80 g"1, corresponding to deflectionforces between 12.5 and 200 mg. In subsequent stud-ies, filament lengths between 1.0 and 5.0 cm wereused, which produced deflection forces between 13and 145 mg (inverse relation). Figure 2 shows thelinear regression method for determining deflectionforces that corresponded to a 50% response.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933149/ on 05/14/2018
No. 9 CORNEAL NERVE REGENERATION / de Leeuw and Chan 1983
A: CONTROL CENTER B: WOUND CENTER C: CONTROL PERIPHERY 0: WOUND PERIPHERY
• i : • • • •
I i i s • • " I I "j i "
80-
60-
40-
20-
o-
•
• • •
• • ••
• • • •
• • : "A
' • ' i s . 1• •
•
•
• .-* '• •
• I . V - " - .• a a
a
•io A 10 30
DAYS
SO •10 A 10 30
I DAYS
-10 A 10 30 50 -10 A 10 30 SO
I n&va ' DAYS
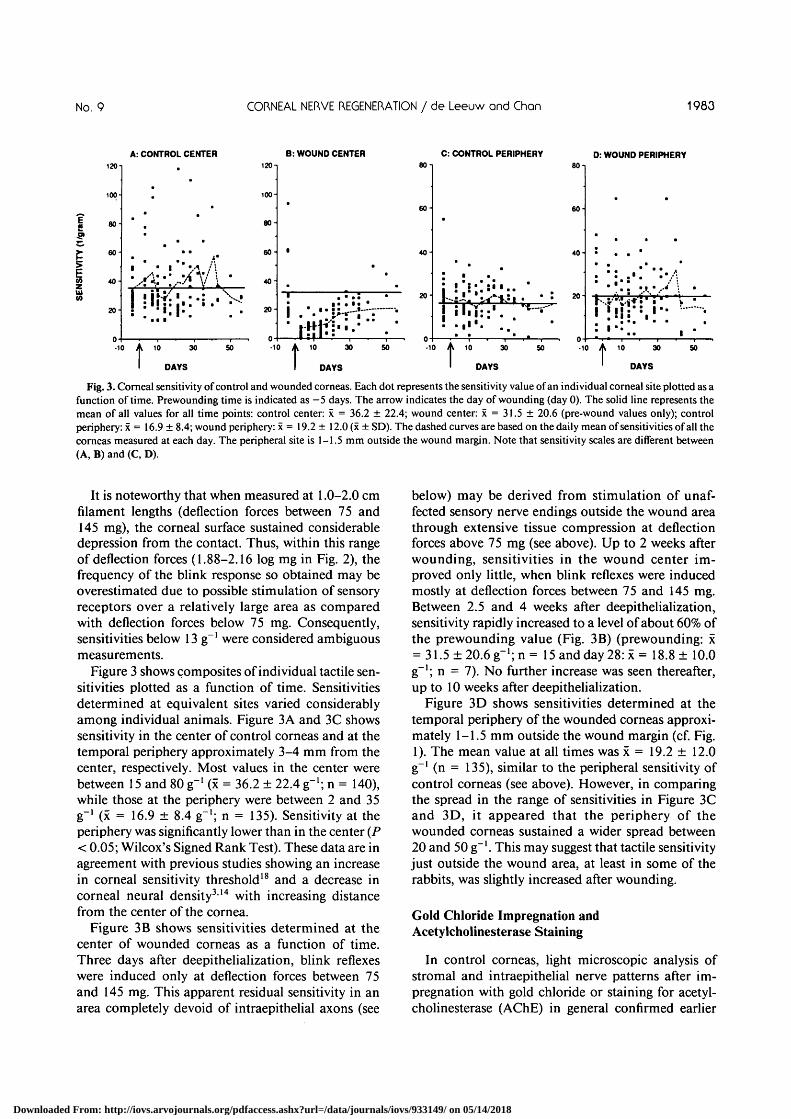
Fig. 3. Corneal sensitivity of control and wounded corneas. Each dot represents the sensitivity value of an individual corneal site plotted as afunction of time. Prewounding time is indicated as —5 days. The arrow indicates the day of wounding (day 0). The solid line represents themean of all values for all time points: control center: x = 36.2 ± 22.4; wound center: x = 31.5 ± 20.6 (pre-wound values only); controlperiphery: x = 16.9 ± 8.4; wound periphery: x = 19.2 ± 12.0 (x ± SD). The dashed curves are based on the daily mean of sensitivities of all thecorneas measured at each day. The peripheral site is 1-1.5 mm outside the wound margin. Note that sensitivity scales are different between(A, B) and (C, D).
It is noteworthy that when measured at 1.0-2.0 cmfilament lengths (deflection forces between 75 and145 mg), the corneal surface sustained considerabledepression from the contact. Thus, within this rangeof deflection forces (1.88-2.16 log mg in Fig. 2), thefrequency of the blink response so obtained may beoverestimated due to possible stimulation of sensoryreceptors over a relatively large area as comparedwith deflection forces below 75 mg. Consequently,sensitivities below 13 g~' were considered ambiguousmeasurements.
Figure 3 shows composites of individual tactile sen-sitivities plotted as a function of time. Sensitivitiesdetermined at equivalent sites varied considerablyamong individual animals. Figure 3A and 3C showssensitivity in the center of control corneas and at thetemporal periphery approximately 3-4 mm from thecenter, respectively. Most values in the center werebetween 15 and 80 g"1 (x = 36.2 ± 22.4 g"1; n = 140),while those at the periphery were between 2 and 35g"1 (x = 16.9 ± 8.4 g"1; n = 135). Sensitivity at theperiphery was significantly lower than in the center (P< 0.05; Wilcox's Signed Rank Test). These data are inagreement with previous studies showing an increasein corneal sensitivity threshold18 and a decrease incorneal neural density314 with increasing distancefrom the center of the cornea.
Figure 3B shows sensitivities determined at thecenter of wounded corneas as a function of time.Three days after deepithelialization, blink reflexeswere induced only at deflection forces between 75and 145 mg. This apparent residual sensitivity in anarea completely devoid of intraepithelial axons (see
below) may be derived from stimulation of unaf-fected sensory nerve endings outside the wound areathrough extensive tissue compression at deflectionforces above 75 mg (see above). Up to 2 weeks afterwounding, sensitivities in the wound center im-proved only little, when blink reflexes were inducedmostly at deflection forces between 75 and 145 mg.Between 2.5 and 4 weeks after deepithelialization,sensitivity rapidly increased to a level of about 60% ofthe prewounding value (Fig. 3B) (prewounding: x= 31.5±20.6g"1;n= 15 and day 28: x = 18.8 ± 10.0g"1; n = 7). No further increase was seen thereafter,up to 10 weeks after deepithelialization.
Figure 3D shows sensitivities determined at thetemporal periphery of the wounded corneas approxi-mately 1-1.5 mm outside the wound margin (cf. Fig.1). The mean value at all times was x = 19.2 ± 12.0g~' (n = 135), similar to the peripheral sensitivity ofcontrol corneas (see above). However, in comparingthe spread in the range of sensitivities in Figure 3Cand 3D, it appeared that the periphery of thewounded corneas sustained a wider spread between20 and 50 g"1. This may suggest that tactile sensitivityjust outside the wound area, at least in some of therabbits, was slightly increased after wounding.
Gold Chloride Impregnation andAcetylcholinesterase Staining
In control corneas, light microscopic analysis ofstromal and intraepithelial nerve patterns after im-pregnation with gold chloride or staining for acetyl-cholinesterase (AChE) in general confirmed earlier
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933149/ on 05/14/2018
1984 INVESTIGATIVE OPHTHALMOLOGY 6 VISUAL SCIENCE / September 1989 Vol. 30
B
i
* • • • %
\
V30 |Jm
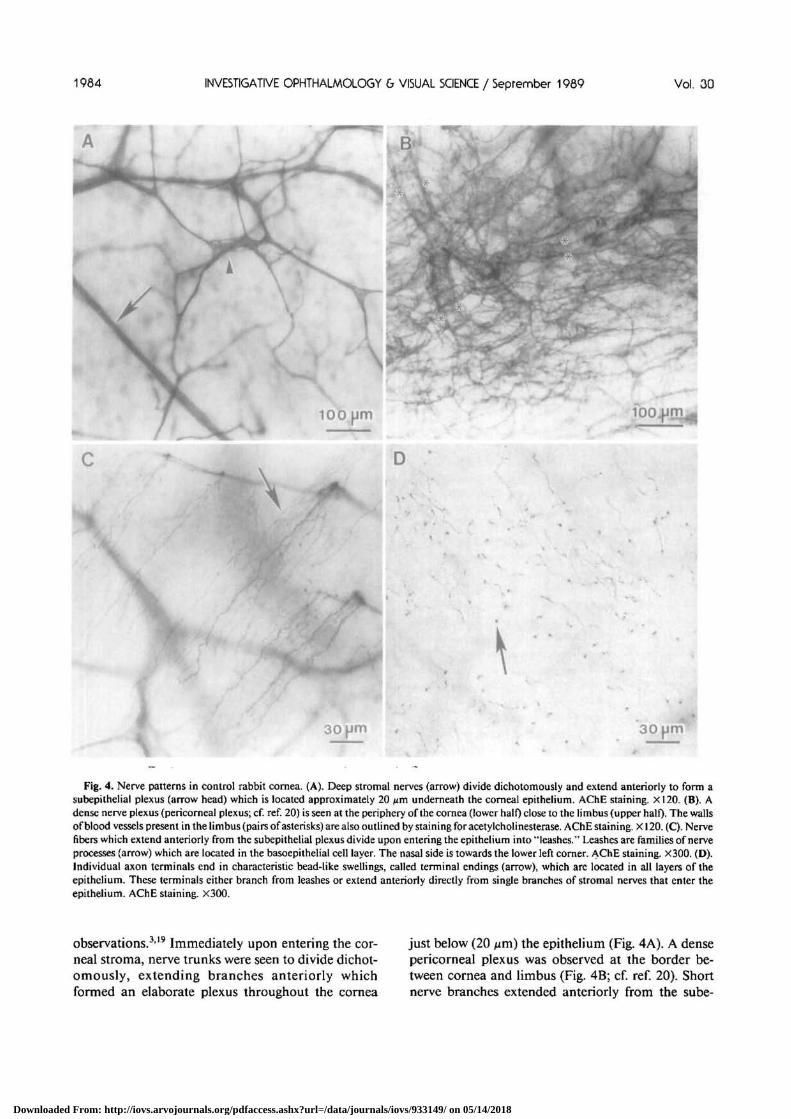
Fig. 4. Nerve patterns in control rabbit cornea. (A). Deep stromal nerves (arrow) divide dichotomously and extend anteriorly to form asubepithelial plexus (arrow head) which is located approximately 20 jzm underneath the corneal epithelium. AChE staining. X120. (B). Adense nerve plexus (pericomeal plexus; cf. ref. 20) is seen at the periphery of the cornea (lower half) close to the limbus (upper half). The wallsof blood vessels present in the limbus (pairs of asterisks) are also outlined by staining for acetylcholinesterase. AChE staining. X120. (C). Nervefibers which extend anteriorly from the subepithelial plexus divide upon entering the epithelium into "leashes." Leashes are families of nerveprocesses (arrow) which are located in the basoepithelial cell layer. The nasal side is towards the lower left corner. AChE staining. X300. (D).Individual axon terminals end in characteristic bead-like swellings, called terminal endings (arrow), which are located in all layers of theepithelium. These terminals either branch from leashes or extend anteriorly directly from single branches of stromal nerves that enter theepithelium. AChE staining. X300.
observations.319 Immediately upon entering the cor-neal stroma, nerve trunks were seen to divide dichot-omously, extending branches anteriorly whichformed an elaborate plexus throughout the cornea
just below (20 nm) the epithelium (Fig. 4A). A densepericomeal plexus was observed at the border be-tween cornea and limbus (Fig. 4B; cf. ref. 20). Shortnerve branches extended anteriorly from the sube-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933149/ on 05/14/2018

No, 9 CORNEAL NERVE REGENERATION / de Leeuw and Chon 1985
N N
Fig. 5. Diagrammatic illustration of the specific orientation ofbasoepithelial leashes. (A). Basoepithelial leashes in the controlcornea have axon processes oriented towards the nasal-mostlimbus. Each symbol for a leash consists of three short axons joinedat an initial point of origin. N, nasal; T, temporal; S, superior; I,inferior. (B). Nerve regeneration following deepithelialization origi-nates from the subepithelial plexus outside the wound area mainlyas distinct single neurites and widely spaced leashes. The orienta-tion of these regenerating axons is perpendicular to the woundmargin. At the nasal side of the wound area, the regeneratinggrowth is opposite to the normal orientation, resulting in redirectedleash growth and neuroma-like nerve patterns. Dotted circle indi-cates wound margin.
pithelial plexus and entered the epithelium (Fig. 4C).Upon entering the epithelium, these nerve branchesdivided into families of fine axons (leashes) whichextended horizontally in the basoepithelial cell layer(Fig. 4C). From these leashes individual axons ex-tended anteriorly and together formed an elaboratenetwork of axon terminals with bead-like terminalendings at all levels of the epithelium (Fig. 4D). Someof the intraepithelial axon terminals were traced backdirectly to stromal nerve fibers, that is, no interme-diate leash formation was found.
The orientation of leashes within each cornea washighly specific and conserved. Within the centraltwo-thirds of the cornea, all leashes were orientedtowards the nasal-most limbus (Fig. 5A). Every cor-nea that was examined revealed the same pattern.Closer to the limbus, leashes were wider and shorterand were oriented towards the periphery, which wasdifferent from the pattern and orientation of theleashes in the center (Fig. 6).
Three days after deepithelialization, regenerated
' • 30pm
Fig. 6. Basoepithelial leashes at the periphery. Leashes at theperiphery of the cornea are wider and shorter than those in thecenter. Their orientation is predominantly towards the periphery(indicated by broken line). AChE staining. X315.
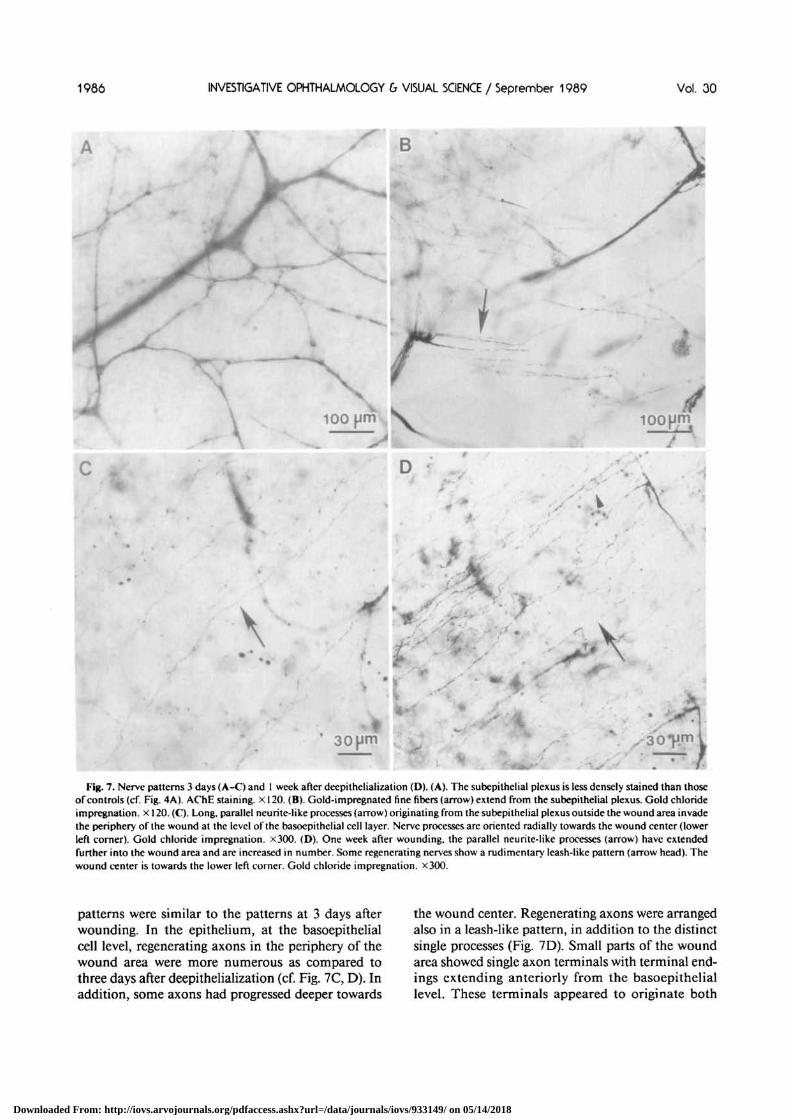
epithelial cells covered the entire wound area. Thisobservation is in agreement with data reported earlierby Chan et al,14 that for a 4-5 mm epithelial woundthe in vivo wound closure time is about 50 hr. Fibersof the subepithelial plexus in the stroma beneath thewound area differed from control stromal nervefibers only by a somewhat decreased staining density,possibly indicating some subtle damage from hep-tanol that diffused into the stroma (cf. Figs. 4A} 7A).Deep stromal nerves were unaffected. With gold chlo-ride, a network of gold-impregnated fine fibers ap-peared to extend from the stromal nerves (Fig. 7B).After staining for AChE such fibers were not seen.Beuerman and Kupke10 also observed these finefibers following mechanical abrasions of the cornealepithelium and suggested this to be a sprouting re-sponse to epithelial denervation at the stromal level.At the epithelial level, the peripheral wound area wasinvaded by mostly parallel nerve processes whichwere of larger diameter than those of leashes and wereoriented perpendicular to the wound margin (Fig.7C). These processes were traced to have branched offfrom stromal nerves located just outside the woundarea (collateral sprouting). However, most of thewounded area except for the periphery was com-pletely devoid of any nerve structures at this stage.
One week after deepithelialization, stromal nerve
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933149/ on 05/14/2018
1986 INVESTIGATIVE OPHTHALMOLOGY 6 VISUAL SCIENCE / September 1989 Vol. 30
B
\
\\
3O#H
Fig. 7. Nerve patterns 3 days (A-C) and 1 week after deepithelialization (D). (A). The subepithelial plexus is less densely stained than thoseof controls (cf. Fig. 4A). AChE staining, x 120. (B). Gold-impregnated fine fibers (arrow) extend from the subepilhelial plexus. Gold chlorideimpregnation, x 120. (C). Long, parallel neurite-like processes (arrow) originating from the subepithelial plexus outside the wound area invadethe periphery of the wound at the level of the basoepithelial cell layer. Nerve processes are oriented radially towards the wound center (lowerleft corner). Gold chloride impregnation. X300. (D). One week after wounding, the parallel neurite-like processes (arrow) have extendedfurther into the wound area and are increased in number. Some regenerating nerves show a rudimentary leash-like pattern (arrow head). Thewound center is towards the lower left corner. Gold chloride impregnation. X300.
patterns were similar to the patterns at 3 days afterwounding. In the epithelium, at the basoepithelialcell level, regenerating axons in the periphery of thewound area were more numerous as compared tothree days after deepithelialization (cf. Fig. 7C, D). Inaddition, some axons had progressed deeper towards
the wound center. Regenerating axons were arrangedalso in a leash-like pattern, in addition to the distinctsingle processes (Fig. 7D). Small parts of the woundarea showed single axon terminals with terminal end-ings extending anteriorly from the basoepitheliallevel. These terminals appeared to originate both
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933149/ on 05/14/2018

No. 9 CORNEAL NERVE REGENERATION / de Leeuw and Chon 1987
from the wound invading leash-like structures andfrom nerve sprouts of the subepithelial plexus under-neath the wound area. The central part of the woundarea was still devoid of any innervation.
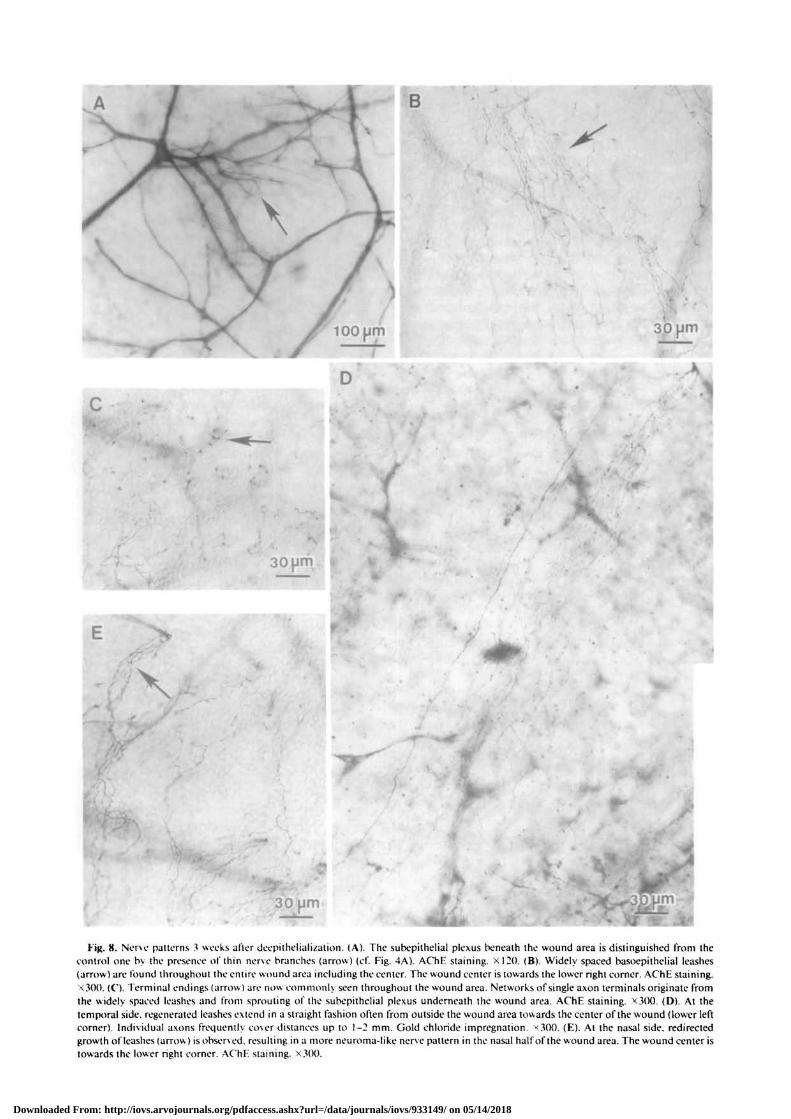
Three weeks after wounding, numerous thin nervefibers were seen to branch from stromal nerves, inboth gold chloride-impregnated and AChE-stainedsections (cf. Figs. 4A, 7B, 8A). These results suggestthat AChE is present in stromal nerve sprouts only1 week or more after wounding. At the epitheliallevel, the entire wound area now showed the presenceof distinct single nerve processes and wide leashes(Fig. 8B) that could be traced back to stromal nervesoutside the wound area. Some leashes, however, wereseen to originate from the subepithelial plexus under-neath the wound area. Networks of single axon termi-nals with terminal endings were found throughoutthe wound area, including the center (Fig. 8C).
The unique orientations of the leashes were alsochanged during nerve regeneration. At the temporalside of the wound area, regenerated leashes werewider and longer than those observed in control cor-neas (cf. Figs. 4C, 8D), with individual axons fre-quently spanning a distance of more than 1 mm.Their nasal-most orientation remained the same. Atthe superior, inferior and especially at the nasal sideof the peripheral wound area, the regenerating leashesoften were curved at the point of origin from thesubepithelial plexus (Figs. 5, 8E). They appeared tohave oriented initially towards the nasal-most limbusfor a short distance and abruptly turned away to aradial orientation. Leashes at the nasal side wereshorter than those at the temporal side, were irregularin form and appeared wavy. Such features resembleneuroma-like intraepithelial nerve patterns describedby others (cf. ref. 11).
Between 3 and 10 weeks after wounding, no furtherqualitative changes were detectable in the regenera-tion pattern of corneal nerves.
Discussion
Various surgical procedures in the eye involve(penetrating) incisions in the cornea or limbus thatinvariably affect corneal innervation. These proce-dures, which include keratotomy, keratectomy, kera-toplasty and perilimbal incision, can be characterizedwith respect to their effect on corneal innervation bythe incision site, incision depth and incision pattern.The time course and pattern of nerve regeneration isexpected to depend in part on where the nerves aresevered, what the extent is of tissue regeneration andwhether corneal tissue is merely incised or replacedby donor tissue.
In the present study, deepithelialization of the cen-tral cornea resulted most probably in complete loss oftactile sensitivity in the center of the wound for aperiod of more than 2 weeks. Partial restoration ofsensitivity (60%) occurred rapidly between 2.5 and 4weeks after wounding, which correlates temporallywith the establishment of a near normal network ofintraepithelial nerves with terminal endings through-out the entire regenerated epithelium. Observationson the pattern of nerve regeneration during the first 2weeks after wounding suggest that intraepithelialreinnervation is gradual and proceeds in a centripetalfashion. Chan et al14 reported in a previous studybased on the same wound model that neural densityin the epithelium at the center of the wound returnsto around 60% of control values 3 to 4 weeks afterdeepithelialization of the cornea. These data onneural density are in agreement temporally and per-centage-wise with the present data on corneal sensi-tivity and suggest that tactile sensitivity is directlyproportional to neural density. This relationship isalso evidenced by the correlation between the cen-tripetal increase of both neural density and tactilesensitivity observed in normal cornea.318
Nerve regeneration following circular keratectomyand keratotomy, which involved anterior stroma, wasstudied morphologically for various periods up to 3months in an experimental rabbit model." Degener-ation of distal nerve fibers within the wound area isfollowed by invasion of the regenerating epitheliumwith two consecutive waves of regenerating neuritesoriginating from the subepithelial plexus surroundingthe wound margin. The first wave of single neuritessubsequently degenerates between 1 and 3 weeksafter surgery. The second wave of neurites reestab-lishes a near-normal intraepithelial nerve pattern be-ginning at 2 weeks after surgery and is thought tosupport restoration of sensory functions.'' In thepresent study, degeneration of the first wave of singleneurites is not a prominent feature. Between 3 and 10weeks after wounding, single neurites are intermixedwith leash-type axons. The nature and function of thesingle neurites remains unknown. It has been sug-gested that collateral sprouting of nerve fibers in reac-tion to damage and regeneration of tissue is inducedby nerve growth factors released from the target tis-sue21 and that these sprouts may serve a function inits regeneration.1112 The time course and pattern offunctional reinnervation of the epithelium observedin the heptanol wound model appear to be similar asin the keratectomy models, which involve either theepithelium only410 or the anterior stroma as well."
In human corneal grafts, nerve regeneration andrestoration of sensitivity is far more slower than in
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933149/ on 05/14/2018
B
30
\
30(Jm
< V
30 pm ira
Fig. 8. Nerve patterns 3 weeks after deepithelialization. (A). The subepithelial plexus beneath the wound area is distinguished from thecontrol one by the presence of thin nerve branches (arrow) (cf. Fig. 4A). AChE staining. XI20. (B). Widely spaced basoepithelial leashes(arrow) are found throughout the entire wound area including the center. The wound center is towards the lower right corner. AChE staining.x.300. (C). 1 erminal endings (arrow) are now commonly seen throughout the wound area. Networks of single axon terminals originate fromthe widely spaced leashes and from sprouting of the subepithelial plexus underneath the wound area. AChE staining. X300. (D). At thetemporal side, regenerated leashes extend in a straight fashion often from outside the wound area towards the center of the wound (lower leftcorner). Individual axons frequently cover distances up to 1-2 mm. Gold chloride impregnation. <300. (E). At the nasal side, redirectedgrowth of leashes (arrow) is observed, resulting in a more neuroma-like nerve pattern in the nasal half of the wound area. The wound center istowards the lower right corner. AChE staining. X300.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933149/ on 05/14/2018

No. 9 CORNEAL NERVE REGENERATION / de Leeuw and Chon 1989
most experimental models.9"'22 The nerve regenera-tion pattern, however, appears to be similar in bothsituations."22 The differences in the regenerationrate may be the result of various conditions inherentin each situation, for example, species differences incorneal nerve pattern323 and regenerative potential,differences in neural density at the site of incision,and different extents of denervation. In human kera-toplasty, incisions are generally close to the limbus inthe periphery of the cornea where neural density islow.9"14 Other factors may also influence the rate ofregeneration, such as preservation of human graftmaterial before transplantation, steroid therapy fol-lowing surgery to prevent immunorejection of thegraft and the ability of the recipient's cornea to reepi-thelialize the donor tissue.24
In penetrating limbal incisions such as performedin iridectomy, trabeculodectomy and cataract sur-gery, corneal nerve trunks are severed proximal totheir branching in the cornea. The entire corneal sec-tor enclosed by the arc of incision is denervated andinsensitive to mechanical stimulation.8"13 Restora-tion of tactile sensitivity is slow both in clinical andexperimental situations.813 In an experimental rabbitmodel, stromal nerve fibers penetrate through thescar tissue between 1 and 3 months after incision,''but the subepithelial plexus and intraepithelial termi-nals are absent or distorted in architecture even after30 months.13 Intraepithelial nerve regeneration fol-lowing limbal incisions does not originate to a greatextent from nerve fibers in the unaffected part of thecornea, in contrast to regeneration following keratec-tomy or keratotomy."'13 These results suggest thatregeneration of functional axons in the cornea is notinduced by tissue denervation per se but rather byclose proximity to regenerating epithelial cells or toother cell types that may react to denervation by se-creting trophic substances. Furthermore, reinnerva-tion of corneal epithelium following limbal incisionsis dependent on prior regeneration of stromal nervesthrough the scar tissue and is, in both clinical andexperimental situations, far slower than that follow-ing experimental keratectomy and keratotomy.8"13
Further studies are necessary to explain the phenom-enon that the more proximally the nerves are cut, themore delayed and incomplete the regeneration pro-cess is.
The innervation pattern of normal rabbit corneavisualized by gold chloride impregnation or AChEstaining is essentially the same as described by Rozsaand Beuerman3 and Tervo and Pakalma,19 respec-tively. The specific orientation of intraepithelialleashes has not been reported before and remains tobe confirmed in other species, including human. It is
not known whether this orientation is an anatomicalcuriosity or has physiological significance. Interest-ingly, during nerve regeneration, two different factorsappear to determine the final orientation of leashes.The tortuous path of regenerating leashes observedespecially at the nasal side of the wound indicates thatat first they are attracted towards the nasal-mostlimbus, as in the normal cornea, and then follow amore centripetal path, possibly under the influence ofa wound-derived factor. It is unlikely that the regen-erating neurites (days 3-7) and leashes (1-3 weeks)are caught up in the wave of migrating epitheliumproceeding towards the wound center (days 1-2) be-cause of temporal differences between the two events(cf. ref. 14). These observations suggest that a zonaldistribution of trophic factors may exist which deter-mine nerve development and regeneration in thecorneal epithelium. This phenomenon may also beinvolved in the establishment of the radial gradient innerve density in normal cornea.314 Concepts of gra-dients and zonal distributions of factors in the corneaneed more attention in the design of future experi-ments.
What is the primary driving force for nerve regen-eration in the cornea? It has been suggested that nervegrowth factor (NGF) may induce collateral sproutingof nociceptive axons in adult animals, probably aspart of the normal regulation of receptive fields.25 Agrowth factor (epithelial neuronotrophic factor,ENF) which is released from corneal epithelium inculture, has been shown to promote neurite out-growth in trigeminal ganglion cells in culture.5 Asurge in the production and secretion of ENF occursduring the period of nerve regeneration in the rabbitcornea.14 It is tempting to speculate that ENF mayhave a function in the regulation of normal innerva-tion of the cornea and may be a primary factor in theinduction of corneal nerve regeneration. However,other factors, including neuropeptides, have beenimplicated in nerve regeneration as well.21 A numberof neuropeptides that are demonstrated in somenerves in the cornea26 await further elucidation oftheir function. Guidance by Schwann cells21 may alsobe essential for regeneration of stromal nerves such asobserved following limbal incisions."13 The relativeimportance of these and other factors in the func-tional sensory reinnervation of the cornea remains tobe established.
Despite a poor understanding of the factors in-volved in corneal nerve regeneration, one can recog-nize one common feature in various regenerationpatterns. Whatever force drives nerve regenerationappears to be insufficient to accomplish a completerestoration of corneal innervation in terms of neural
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933149/ on 05/14/2018

1990 INVESTIGATIVE OPHTHALMOLOGY & VISUAL SCIENCE / Seprember 1989 Vol. 30
pattern, density and sensory function. Possibly, theinductive force in a mature cornea may differ quanti-tatively and qualitatively from that in a developingcornea. Also, the microenvironment in a mature cor-nea may limit full expression of the force inducingnerve regeneration. Comparison of nerve regenera-tion in mature and developing corneas may shedsome light on the phenomenon of subnormal regen-eration in mature corneas. Furthermore, analysis ofthe process of nerve regeneration in an experimentalsituation can be helpful in developing new strategiesfor the enhancement of nerve regeneration in theclinical situation.
Key words: cornea, innervation, sensitivity, wound healing,nerve regeneration
Acknowledgments
The authors are grateful to Dr. Donald Martin for statis-tical analysis, to Heidi Utsugi and Dan Possin for technicalassistance, and to Chuck Stephens for photographic help.Dr. de Leeuw was a postdoctoral fellow at the Departmentof Ophthalmology.
References
1. Ten Tusscher MPM, KJooster J, and Vrensen GFJM: Theinnervation of the rabbit's anterior eye segment: A retrogradetracing study. Exp Eye Res 46:717, 1988.
2. Bossy J: Cranial nerves and special sense organs. In Atlas ofNeuroanatomy and Special Sense Organs, Bossy J, editor. Phil-adelphia, W.B. Saunders Company, 1970, pp. 214-216,240-243.
3. Rozsa AJ and Beuerman RW: Density and organization of freenerve endings in the corneal epithelium of the rabbit. Pain14:105, 1982.
4. Tanelian DL and Beuerman RW: Responses of rabbit cornealnociceptors to mechanical and thermal stimulation. ExpNeurol 84:165, 1984.
5. Chan K.Y: Cornea, trigeminal ganglion, and their interactions.In Vision: Structure and Function, Yew DT, So KF, and TsangDSC, editors. Singapore, World Scientific Publishing, 1988,pp. 47-62.
6. Beuerman RW and Schimmelpfennig B: Sensory denervationof the rabbit cornea affects epithelial properties. Exp Neurol69:169, 1980.
7. Vannas A, Holden BA, and Sweeney DF: Epithelial metabo-lism of the corneal graft is abnormal. Br J Ophthalmol 71:593,1987.
8. Lyne A: Corneal sensitivity after surgery. Trans OphthalmolSocUK 102:302, 1982.
9. Mathers WD, Jester JV, and Lemp MA: Return of humancorneal sensitivity after penetrating keratoplasty. Arch Oph-thalmol 106:210, 1988.
10. Beuerman RW and Kupke K: Neural regeneration followingexperimental wounds of the cornea in the rabbit. In TheStructure of the Eye, Hollyfield JG, editor. Amsterdam, Else-vier North Holland, Inc., 1982, pp. 319-30.
11. Rozsa AJ, Guss RB, and Beuerman RW: Neural remodelingfollowing experimental surgery of the rabbit cornea. InvestOphthalmol Vis Sci 24:1033, 1983.
12. Beuerman RW and Rozsa AJ: Collateral sprouts are replacedby regenerating neurites in the wounded corneal epithelium.Neurosci Lett 44:99, 1984.
13. Chan-Ling T, Tervo K, Tervo T, Vannas A, Holden BA, andEranko L: Long-term neural regeneration in the rabbit follow-ing 180° limbal incision. Invest Ophthalmol Vis Sci 28:2083,1987.
14. Chan KY, Jones RR, Bark DH, Swift J, Parker JA, andHaschke RH: Release of neuronotrophic factor from rabbitcorneal epithelium during wound healing and nerve regenera-tion. Exp Eye Res 45:633, 1987.
15. Cintron C, Hassinger L, Kublin CL, and Friend J: A simplemethod for the removal of rabbit corneal epithelium utilizingn-heptanol. Ophthalmic Res 11:90, 1979.
16. Metcalf JF: Corneal sensitivity and neurohistochemical studiesof experimental herpetic keratitis in the rabbit. Exp Eye Res35:231, 1982.
17. Karnovsky MJ and Roots L: A 'direct-coloring' thiocholinemethod for cholinesterases. J Histochem Cytochem 12:219,1964.
18. Draeger J, Koudelka A, and Lubahn E: Zur Asthesiometry derHornhaut. Klin Mbl Augenheilk 169:407, 1976.
19. Tervo T and Palkama A: Innervation of the rabbit cornea: Ahistochemical and electron-microscopic study. Acta Anat102:164, 1978.
20. Tervo T: Histochemical demonstration of cholinesterase activ-ity in the cornea of the rat and the effect of various denerva-tions on the corneal nerves. Histochemistry 47:133, 1976.
21. Dekker A, Gispen WH, and De Wied D: Minireview: Axonalregeneration, growth factors and neuropeptides. Life Sci41:1667, 1987.
22. Tervo T, Vannas A, Tervo K, and Holden BA: Histochemicalevidence of limited reinnervation of human corneal grafts.Acta Ophthalmol 63:207, 1985.
23. Schimmelpfennig B: Nerve structures in human central cor-neal epithelium. Graefes Arch Clin Exp Ophthalmol 218:14,1982.
24. Reed DB, Hills JF, Mannis MJ, and Johnson CA: Cornealepithelial healing after penetrating keratoplasty using topicalhealon versus balanced salt solution. Ophthalmic Surg 18:525,1987.
25. Diamond J, Coughlin M, Macintyre L, Holmes M, and Vi-sheau B: Evidence that endogenous j3 nerve growth factor isresponsible for the collateral sprouting, but not the regenera-tion, of nociceptive axons in adult rats. Proc Natl Acad SciUSA 84:6596, 1987.
26. Tervo T, Tervo K, Eranko L, Vannas A, Eranko O, and CuelloAC: Substance P immunoreaction and acetylcholinesterase ac-tivity in the cornea and Gasserian ganglion. Ophthalmic Res15:280, 1983.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933149/ on 05/14/2018





























