Embed Size (px)
Citation preview

Marine Biology 106, 267-272 (1990)
Marine . . . . . . . . . . . . . . . Biology
© Springer-Verlag 1990
Corallum morphology and composition of crustacean cryptofauna of the hermatypic coral Madracis mirabilis
J.B. Lewis 1 and P. V. R. Snelgrove 2
1 Department of Biology, McGill University, Montreal, Quebec H3A 1B1, Canada 2 Department of Biology, Woods Hole Oceanographic Institution Woods Hole, Massachusetts 02543, USA
Date of final manuscript acceptance: May 4, 1990. Communicated by R. O'Dor, Halifax
Abstract. The motile crustacean cryptofauna of the her- matypic coral Madracis mirabilis (Duchassaing and Michelotti) was examined from three sites on reefs along the west coast of Barbados in 1985. In addition to site differences in degree of eutrophication, two distinct with- in-corallum habitats were recognized, based on differ- ences in the growth form of the coral host. One form grows in isolated, hemispherical heads, with short, ro- bust, widely separated branches covered by 50 to 80% live tissue. The second form occurs within large continu- ous beds of long, thin, tightly spaced branches tipped by 20 to 30% live tissue. Coral habitats consisting of small isolated heads support a crustacean fauna of high deca- pod and amphipod diversity, with numerous rare and site endemic species and an abundance of relatively large de- capods and amphipods. The continuous beds favour a community of lower decapod and amphipod diversity, fewer rare and site endemic species and with an abun- dance of smaller sized isopods and copepods. It is sug- gested that spatial separation of branches and colonies as well as food availability associated with corallum mor- phology are factors influencing the variation in species abundance and faunal composition of the two habitats.
Introduction
There is a rich and varied invertebrate fauna associated with coral substrata (Grassle 1973, Patton 1973). The motile crustacean cryptofauna that dwells on the surface and in crevices of corals has been implicated in a number of trophic transfer relations within coral-reef ecosystems. The animals are a source of food for fish (Vivien and Peyrot-Clausade 1974) and predatory molluscs (Kohn and Nybakken 1975). They are consumers of epilithic algae (Klumpp et al. 1988) and may utilize mucus pro- duced by corals, or recycle the detritus and organic mate- rial trapped by mucus (Knudsen 1967, Patton 1974, Hutchings 1983). They comprise part of the demersal zooplankton that migrates at night from the reef substra-
turn into the water column (Sale et al. 1978) and may, therefore, be a food scource for neritic organisms and for corals themselves.
Much of the current research on this coral-associated fauna attempts to explain variation in community com- position. Differences in community structure of the fauna between geographical areas, from along depth gradients and from different parts of reefs have been reported by McCloskey (1970), Abele (1976), Coles (1980), Reed etal. (1982), Gotelli and Abele (1983), Gotelli etal. (1985). Variation along a eutrophication gradient has been examined by Snelgrove and Lewis (1989) and differ- ences in community structure resulting from habitat vari- ation within a single location have been reported by Abele and Patton (1976), Coles (1980) and Abele (1984). The effect of trophic relationships on the structure of these communities was studied by Klumpp et al. (1988).
Most of the work on coral cryptofauna has been upon a few species of pocilloporid corals from Pacific reefs. In the tropical eastern Atlantic data exist on the fauna from two species of oculinid corals, neither of which is an abundant reef building coral. In this study we examine the crustacean cryptofauna of a more abundant and widespread Caribbean reef coral, Madracis mirabilis. Other common macrofaunal associates of M. mirabilis were reported previously (Lewis 1960, 1965). Our study is an extension of a survey of the effects of eutrophication on coral reefs in Barbados (Tomascik and Sander 1985, Snelgrove and Lewis 1989) and is focussed on the com- munities from two different growth forms of the corallum of the host coral. We compare the M. mirabilis crustacean fauna with the cryptofauna of other coral hosts and com- ment upon the importance of within corallum habitat variation as a determinant of community composition.
Materials and methods
Madracis mirabilis (Duchassaing and Michelotti) is an erect, branching coral which occurs in two forms in Barbados. One form grows in isolated, hemispherical clumps or heads of approximately

268 J.B. Lewis and P. V. R. Snelgrove: Coral reef cryptofauna
t3°20 '
Bank I Sandy Poyne:
Br/
- - 13° IO'
5¢4o' N 5¢3o'
t Fig. 2. Map of Barbados, West Indies, showing the three sites (Pay- nes Bay, Brighton and Bank Reef) sampled for Madracis mirabilis cryptofauna. Sandy Lane is a reference station established by Tomascik and Sander (1985). Depths are given in parentheses
Fig. 1. Madracis mirabilis. (a) Underwater photograph of isolated clumps at Paynes Bay site. (b) Underwater photograph of a large bed at Bank Reef site. (c) Exposed section of colonies at Bank Reef showing long, tightly spaced branches
10 to 20 cm diameter (Fig. I a). The branches of this form are short, robust, widely separated and covered with 30 to 50 mm (approxi- mately 50 to 80% of branch) live tissue. The second form occurs within large, continuous beds which may be tens of meters in width and up to 0.5 m in vertical thickness (Fig. 1 b, c). The branches of the latter form are long, thin, tightly spaced and bear live tissue only at the tips (approximately 20 to 30% of branch). Both coralla forms occur all along the outer edge of the fringing reefs on the west coast on similar substrate and at the same depth (Lewis 1960).
Two sampling stations were chosen to include a eutrophied and a relatively unpolluted site, from among those established by Snel- grove and Lewis (1989) at the southern end of the fringing reef system (Fig. 2). Brighton is a highly eutrophied site while Bank Reef, which is situated about 1 km offshore, was reported to be less
eutrophied (Tomascik and Sander 1985, Snelgrove and Lewis 1989). Extensive beds of Madracis mirabilis occur at 9 and 15 m at Brighton and Bank Reef, respectively. A third site, Paynes Bay, is conveniently situated midway between the other two sites and 0.5 km south of the Sandy Lane reference station (Fig. 2) of Tomas- cik and Sander (1985). Numerous isolated heads of M. mirabilis occur at a depth of 5 m at Paynes Bay. Because of their close proximity, Paynes Bay and Sandy Lane are assumed to have similar water quality characteristics. Water quality at Sandy Lane was found to be less eutrophied in comparison to Brighton (Tomascik and Sander 1985, 1987). Tomascik and Sander (1985) regarded all three sites as having similar substrate and other physical environ- mental conditions.
The isolated heads at Paynes Bay were sampled by wrapping a polyurethane bag tightly around a clump, prying the head from the substratum and sealing it in the bag. Sampling of the large beds at the Brighton and Bank Reef sites was carried out with a coring device and the preservation, sorting and identification of the crus- tacean fauna followed the methods of Snelgrove and Lewis (1989). Each of the sites was sampled three times between June and Octo- ber, 1985. Five replicate samples were taken each time, for a total of 15 samples per site. No marked temporal changes in water qual- ity variables have been reported on west coast reefs during this season (Lewis et al. 1968, Tomascik and Sander 1985).
Statistical analysis reinterprets the numerical data from Brighton and Bank Reef previously reported by Snelgrove and Lewis (1989) in relation to samples collected from the third site at Paynes Bay. In order to first establish whether there were differ- ences in the sample volumes of coral or the abundance of organisms among the three sites, analyses of variance were performed. Shapiro-Wilk W-tests for normality and F-max tests (Zar 1984) revealed no violations of the assumptions of parametric analysis. Ordination and classification procedures (Field et al. 1982), fol- lowed by determination and comparisons of indices of community structure (Snelgrove and Lewis 1989), were next employed to exam- ine between habitat differences in the crustacean fauna.
The community descriptors species richness (number of species) and eveness (J) were chosen rather than one of the composite in- dices of diversity (eg. H, HI,) because the latter are difficult to interpret without the former two variables (Hurlbert 1971, Pielou 1975). Means of density, richness and evenness were compared in all taxa combined and then separately among adult decapods, am-
J. B. Lewis and P. V. R. Snelgrove: Coral reef cryptofauna 269
E
80
40
o o o o o o o o o o o o o o o o o o o o o e e e • o o o o
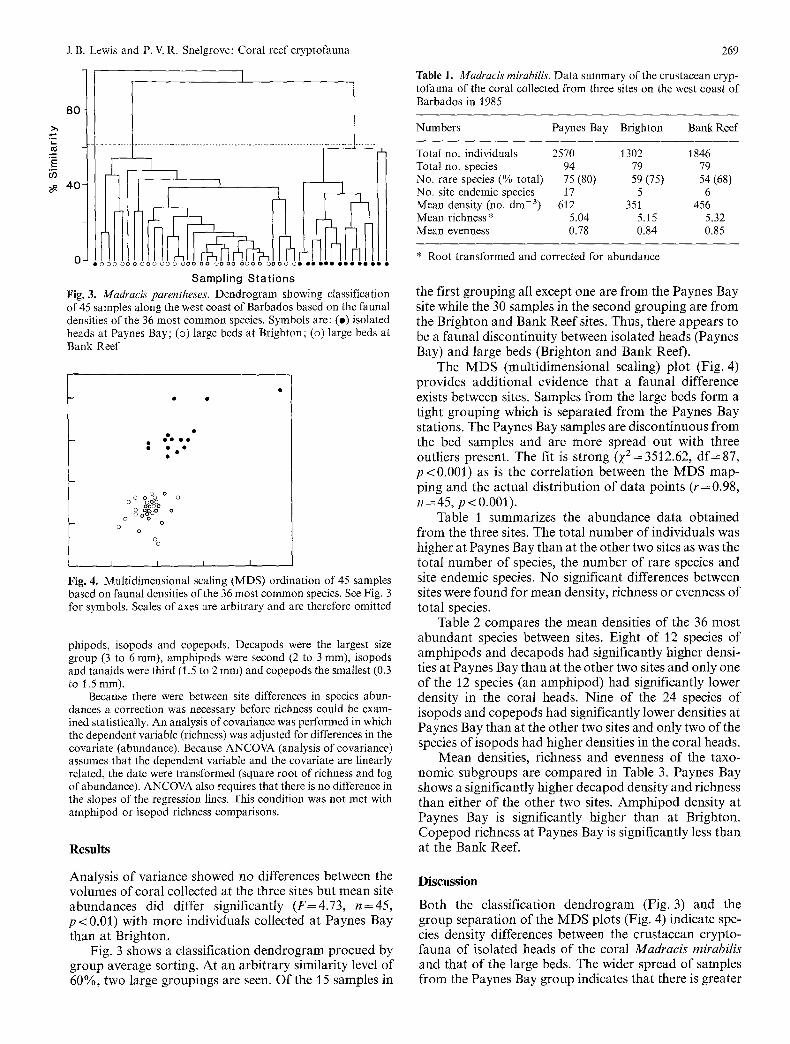
Sampl ing S t a t i o n s Fig. 3. MadracL* parentheses. Dendrogram showing classification of 45 samples along the west coast of Barbados based on the faunal densities of the 36 most common species. Symbols are: (e) isolated heads at Paynes Bay; (o) large beds at Brighton; (o) large beds at Bank Reef
o ° o o o
o o o O o o o
oOg~oO o o o
o o
%
I I I I
Fig. 4. Multidimensional scaling (MDS) ordination of 45 samples based on faunal densities of the 36 most common species. See Fig, 3 for symbols. Scales of axes are arbitrary and are therefore omitted
phipods, isopods and copepods. Decapods were the largest size group (3 to 6 ram), amphipods were second (2 to 3 ram), isopods and tanaids were third (1.5 to 2 ram) and copepods the smallest (0.3 to 1.5 ram).
Because there were between site differences in species abun- dances a correction was necessary before richness could be exam- ined statistically. An analysis of covariance was performed in which the dependent variable (richness) was adjusted for differences in the covariate (abundance). Because ANCOVA (analysis of covariance) assumes that the dependent variable and the covariate are linearly related, the date were transformed (square root of richness and log of abundance). ANCOVA also requires that there is no difference in the slopes of the regression lines. This condition was not met with amphipod or isopod richness comparisons.
Results
Table 1. Madracis mirabilis. Data summary of the crustacean cryp- tofauna of the coral collected from three sites on the west coast of Barbados in 1985
Numbers Paynes Bay Brighton Bank Reef
Total no. individuals 2570 1302 1846 Total no. species 94 79 79 No. rare species (% total) 75 (80) 59 (75) 54 (68) No. site endemic species 17 5 6 Mean density (no. dm -3) 612 351 456 Mean richness * 5.04 5.15 5.32 Mean evenness 0.78 0.84 0.85
* Root transformed and corrected for abundance
the first grouping all except one are f rom the Paynes Bay site while the 30 samples in the second grouping are f rom the Brighton and Bank Reef sites. Thus, there appears to be a faunal discontinuity between isolated heads (Paynes Bay) and large beds (Brighton and Bank Reef).
The M D S (multidimensional scaling) plot (Fig. 4) provides addit ional evidence that a faunal difference exists between sites. Samples f rom the large beds form a tight grouping which is separated f rom the Paynes Bay stations. The Paynes Bay samples are discontinuous f rom the bed samples and are more spread out with three outliers present. The fit is strong ( ~ 2 = 3512.62, d f = 87, p < 0 . 0 0 l ) as is the correlation between the MDS map- ping and the actual distribution of data points ( r=0.98, n = 4 5 , p < 0.001).
Table 1 summarizes the abundance data obtained f rom the three sites. The total number of individuals was higher at Paynes Bay than at the other two sites as was the total number of species, the number of rare species and site endemic species. No significant differences between sites were found for mean density, richness or evenness of total species.
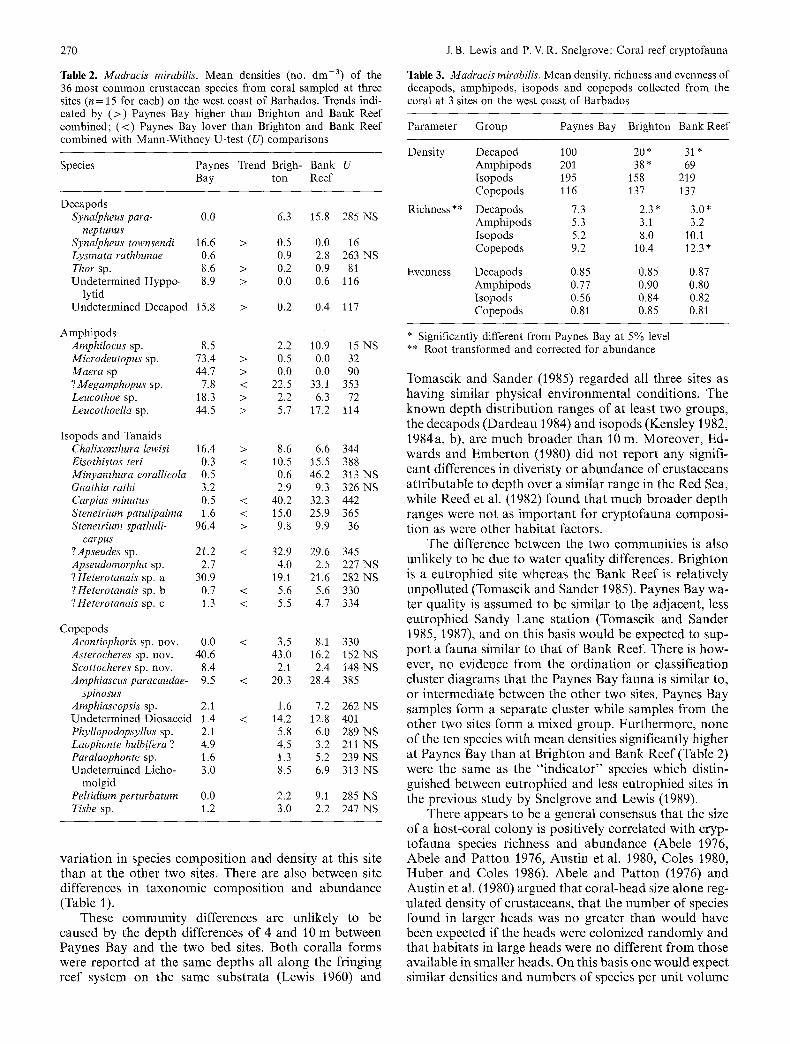
Table 2 compares the mean densities of the 36 most abundant species between sites. Eight of 12 species of amphipods and decapods had significantly higher densi- ties at Paynes Bay than at the other two sites and only one of the 12 species (an amphipod) had significantly lower density in the coral heads. Nine of the 24 species of isopods and copepods had significantly lower densities at Paynes Bay than at the other two sites and only two of the species of isopods had higher densities in the coral heads.
Mean densities, richness and evenness of the taxo- nomic subgroups are compared in Table 3. Paynes Bay shows a significantly higher decapod density and richness than either of the other two sites. Amphipod density at Paynes Bay is significantly higher than at Brighton. Copepod richness at Paynes Bay is significantly less than at the Bank Reef.
Analysis of variance showed no differences between the volumes of coral collected at the three sites but mean site abundances did differ significantly (F=4.73 , n=45 , p<0 .01 ) with more individuals collected at Paynes Bay than at Brighton.
Fig. 3 shows a classification dendrogram procued by group average sorting. At an arbi trary similarity level of 60%, two large groupings are seen. Of t h e / 5 samples in
Discussion
Both the classification dendrogram (Fig. 3) and the group separation of the MDS plots (Fig. 4) indicate spe- cies density differences between the crustacean crypto- fauna of isolated heads of the coral Madracis mirabilis and that of the large beds. The wider spread of samples f rom the Paynes Bay group indicates that there is greater
270
Table 2. Madracis mirabilis. Mean densities (no. dm -3) of the 36 most common crustacean species from coral sampled at three sites (n = 15 for each) on the west coast of Barbados. Trends indi- cated by (>) Paynes Bay higher than Brighton and Bank Reef combined; (<) Paynes Bay lover than Brighton and Bank Reef combined with Mann-Withney U-test (U) comparisons
Species Paynes Trend Brigh- Bank U Bay ton Reef
Decapods Synalpheus para- 0.0
neptunus Synalpheus townsendi 16.6 > Lysrnata rathbunae 0.6 Thor sp. 8.6 > Undetermined Hyppo- 8.9 >
lytid Undetermined Decapod 15.8 >
Amphipods Amphilocus sp. 8.5 2.2 Microdeutopus sp. 73.4 > 0.5 Maera sp 44.7 > 0.0 ?Megamphopus sp. 7.8 < 22.5 Leucothoe sp. 18.3 > 2.2 Leucothoella sp. 44.5 > 5.7
Isopods and Yanaids Chalixanthura lewisi 16.4 > 8.6 Eisothistos teri 0.3 < 10.5 Minyanthura eorallicola 0.5 0.6 Gnathia rathi 3.2 2.9 CaiTias minutus 0.5 < 40.2 Stenetrium patulipalrna 1.6 < 15.0 Stenetrium spathuli- 96.4 > 9.8
carpus ?Apseudes sp. 21.2 < 32.9 Apseudomorpha sp. 2.7 4.0 ?Heterotanais sp. a 30.9 19.1 ?Heterotanais sp. b 0.7 < 5.6 ?Heterotanais sp. c 1.3 < 5,5
Copepods Acontiophoris sp. nov. 0.0 < 3.5 Asterocheres sp. nov. 40.6 43.0 Scottoeheres sp. nov. 8.4 2.1 Amphiascus paracaudae- 9.5 < 20.3
spinosus Amphiaseopsis sp. 2.1 Undetermined Diosaccid 1.4 Phyllopodopsyllus sp. 2.1 Laophonte bulbifera ? 4.9 Paralaophonte sp. 1.6 Undetermined Licho- 3.0
molgid Peltidiurn perturbatum 0.0 Tisbe sp. 1.2
6.3 15.8 285 NS
0,5 0.0 16 0.9 2.8 263 0.2 0.9 81 0.0 0.6 116
0.2 0.4 117
10.9 15 0.0 32 0.0 90
33.1 353 6.3 72
17.2 114
6.6 344 15.5 388 46.2 313 9.3 326
32.3 442 25.9 365 9.9 36
29.6 345 2.5 227
21.6 282 5.6 330 4.7 334
NS
NS
NS NS
NS NS
8.1 330 16.2 152 NS 2.4 148 NS
28.4 385
1.6 7.2 262 NS < 14.2 12.8 401
5.8 6.0 289 NS 4.5 3.2 211 NS 1.3 5.2 239 NS 8.5 6.9 313 NS
2.2 9.1 285 NS 3.0 2.2 247 NS
variat ion in species composi t ion and density at this site than at the other two sites. There are also between site differences in t axonomic composi t ion and abundance (Table 1).
These communi ty differences are unlikely to be caused by the depth differences o f 4 and 10 m between Paynes Bay and the two bed sites. Both coralla forms were reported at the same depths all a long the fringing reef system on the same substrata (Lewis 1960) and
J.B. Lewis and P. V. R. Snelgrove: Coral reef cryptofauna
Table 3. Madracis rnirabilis. Mean density, richness and evenness of decapods, amphipods, isopods and copepods collected from the coral at 3 sites on the west coast of Barbados
Parameter Group Paynes Bay Brighton Bank Reef
Density Decapod 100 20 * 31 * Amphipods 201 38 * 69 1sopods 195 158 219 Copepods 116 137 137
Richness** Decapods 7.3 2.3' 3.0" Amphipods 5.3 3.1 3.2 Isopods 5.2 8.0 10.1 Copepods 9.2 10.4 12.3 *
Evenness Decapods 0.85 0.85 0.87 Amphipods 0.77 0.90 0.80 Isopods 0.56 0.84 0.82 Copepods 0.81 0.85 0.81
* Significantly different from Paynes Bay at 5% level ** Root transformed and corrected for abundance
Tomascik and Sander (1985) regarded all three sites as having similar physical environmenta l conditions. The known depth distr ibution ranges o f at least two groups, the decapods (Dardeau 1984) and isopods (Kensley 1982, 1984a, b), are much broader than 10 m. Moreover , Ed- wards and Ember ton (1980) did no t repor t any signifi- cant differences in diveristy or abundance o f crustaceans at tr ibutable to depth over a similar range in the Red Sea, while Reed et al. (1982) found that much broader depth ranges were no t as impor tan t for c ryp to fauna composi- t ion as were other habi ta t factors.
The difference between the two communit ies is also unlikely to be due to water quali ty differences. Brighton is a eutrophied site whereas the Bank Reef is relatively unpol luted (Tomascik and Sander 1985). Paynes Bay wa- ter quali ty is assumed to be similar to the adjacent, less eutrophied Sandy Lane stat ion (Tomascik and Sander 1985, 1987), and on this basis would be expected to sup- por t a fauna similar to tha t o f Bank Reef. There is how- ever, no evidence f rom the ordina t ion or classification cluster diagrams that the Paynes Bay fauna is similar to, or intermediate between the other two sites. Paynes Bay samples fo rm a separate cluster while samples f rom the other two sites fo rm a mixed group. Fur thermore , none o f the ten species with mean densities significantly higher at Paynes Bay than at Brighton and Bank Reef (Table 2) were the same as the '" indicator" species which distin- guished between eut rophied and less eutrophied sites in the previous study by Snelgrove and Lewis (1989),
There appears to be a general consensus that the size o f a host-coral colony is positively correlated with cryp- to fauna species richness and abundance (Abele 1976, Abele and Pa t ton 1976, Aust in et al. 1980, Coles 1980, Huber and Coles 1986). Abele and Pa t ton (1976) and Aust in et al. (1980) argued that cora l -head size alone reg- ulated density o f crustaceans, tha t the number o f species found in larger heads was no greater than would have been expected if the heads were colonized r andomly and that habitats in large heads were no different f rom those available in smaller heads. On this basis one would expect similar densities and numbers o f species per unit volume

J. B. Lewis and P. V. R. Snelgrove: Coral reef cryptofauna
of Madracis mirabilis colonies. This was not the case however, for the small heads of M. mirabilis contain a larger number of species of decapods and amphipods (but fewer copepods) and higher densities of decapods and amphipods than samples from the beds. The M. mirabilis fauna thus does not conform to the general con- census concerning host size denoted above. Gotelli et al. (•985) also found that coral host size was not a good predictor of cryptofauna species abundance and Ed- wards and Emberton (1980) considered that additional habitat factors might be more important than host size alone.
Paynes Bay colonies bear more living coral surface area at the tips of the branches than do Brighton and Bank Reef colonies. The importance of living tissue on coral branches as a food source for obligate symbiotic crustacea has been reported by a number of authors. Coles (•980) found that large symbiotic decapods were associated with living heads of Pocillopora meandrina whereas small, non-symbiotic forms were characteristic of dead coral. Both the number of species and the number of individuals of symbiotic decapods were positively cor- related with live coral surface area. Abele (•976), Abele and Patton (•976) and Gotelli et al. (•985), also found taxonomic differences between the fauna of dead and living Pocillopora sp. heads and positive correlations be- tween amounts of live coral tissue and both species abun- dances and numbers of individuals. In the Caribbean coral Oculina varicosa Reed and Mikkelsen (•987) found that greater living coral coverage permitted more coral- eating molluscs while algal grazers were more abundant on dead coral. Differences in Madracis mirabilis crypto- fauna may also be partly due to relative abundance of living tissue as a food source. The higher density and species richness of decapods in coral heads is consistent with the suggestion by Coles (•980) that larger crus- taceans tend to be obligate symbionts and hence are more abundant in habitats with a high proportion of live coral tissue. The smaller crustacea (the copepods and isopods) tend to be more abundant in the sites with less live coral- tissue (Table 2) where other food sources are likely to be available. The heterogeneity of dead coral habitats and their greater range of food resources has been noted by Grassle (•973) and Coles (•980).
While the results for the Madracis mirabilis crypto- fauna are similar to those of Coles (•980), Abele (•976) and Abele and Patton (1976) in respect to decapod pref- erence for coral colonies with live tissue, it should be noted that the pocilloporid fauna described by these au- thors consists primarily of obligate commensals. Reed et al. (1982) reported a negative correlation between the number of non-obligate crustaceans and the amount of living Oculina varicosa coral tissue and argued from this, that commensal decapods are less important in the Caribbean than in the Pacific.
In regard to the importance of separation and isola- tion of coral heads, Gotelli and Abele (•983) and Gotelli et al. (1985) noted that small Pocillopora sp. heads with clumped or continuous distribution on reef flats support- ed more individuals than did patchily distributed or iso- lated heads on the reef flanks. However, exposure at low
271
tide on the reef flats and lack of predation by fish might also have caused differences between the fauna of these two habitats. In contrast, in the Caribbean coral Oculina varicosa, Reed et al. (1982) reported lower densities of decapods associated with deep living massive thickets or beds than in the more scattered colonies in shallower water. In this respect the distribution of the fauna of Madracis mirabilis is similar to the decapod fauna of Reed et al. (1982). In addition, the same authors reported that scattered O. varicosa heads supported the highest number of species and more rare species and site endemic species as was the case for M. mirabilis. However, Reed et al. (1982) also noted that their differences could have been due to colony size or periodic seasonal environmen- tal perturbations.
Where there is spatial separation of the isolated heads, movement between the clumps might be expected to be low and dominance of a few, competitively superior species might occur. In large, flat beds however, individ- uals could migrate many metres without leaving the pro- tection of the coral and a much more evenly distributed community would be expected. This may be reflected in the higher evenness of the Brighton and Bank Reef sam- ples and the tighter clumping of the samples from these sites in the MDS plots. However, the small size of the isolated heads sampled in the present study may be as important as the distance between colonies. Coles (1980) suggested that coral heads less than 1000 cm 3 cannot sup- port a predictable symbiotic decapod community. The large number of rare species and of site indigenous spe- cies may be indicative of temporary residents in a com- munity which is too small to be stable.
In recent studies of the motile cryptofauna of corals (Klumpp et al. 1988, Klumpp and Polunin 1989) it is argued that trophic relationships are as important as habitat heterogeneity in determining community compo- sition. However, there is very little information on the feeding habits of the Madracis mirabilis coral fauna to define their trophic relationships. Kensley (1984a) re- ported that coral inhabiting isopods consumed a mixture of algae, detritus and protozoans and regarded them as being widespread and numerous on Caribbean reefs. The most abundant species at Paynes Bay (and most abun- dant overall) was the isopod Stenetrium spathulicarpus. It has previously been recorded on Madracis sp. as well as on other corals and coral rubble by Kensley (1984a). S. patulipalma which was more common at Brighton and Bank Reef, has also been reported on Madracis sp. and coral rubble. Highly modified periopods in both suggest possible feeding specializations in the two species. The most abundant species from both Brighton and Bank Reef was also an isopod, Carpius minutus, followed by two other unidentified isopods, Apseudes sp. and Hetero- tanais sp. Isopod grazers and detritivores are thus com- mon in the large bed habitats. Amphipods were also im- portant algal grazers on the Great Barrier Reef (Klumpp et al. 1988).
In conclusion, the coral habitats composed of small isolated heads with a relatively high proportion of living corm tissue and loose, open branching, favour a crus- tacean fauna of high diversity, with numerous rare and

272
site endemic species and an a b u n d a n c e o f re la t ively large d e c a p o d s and amph ipods . The con t inuous beds, wi th rel- a t ively less t issue per b r a n c h and denser b ranching , fa- vour a c o m m u n i t y o f lower divers i ty , fewer rare and en- demic species and wi th an a b u n d a n c e o f smal ler sized i sopods and copepods . Thus , a c o m b i n a t i o n o f spa t ia l s epa ra t i on o f cora l b ranches and colonies as well as food ava i lab i l i ty and preferences m a y be i m p o r t a n t in influ- encing the fauna l c o m p o s i t i o n o f the two habi ta t s .
Acknowledgements. We thank Dr W Hunte for use of facilities at the Bellairs Research Institute of McGill University in Barbados. Dr J. O. Ramsay of the Department of Psychology, McGill University, kindly offered advice on ordination analysis and Drs J. B. Boers and T. Tomascik provided much useful information in planning the study. B. Conlin assisted in sample collection and K. Silkouskas assisted in sample sorting. The following people kindly assisted in identification of material: E~ Bousfield, Canadian National Muse- um (amphipods); G. Boxshall, British Museum (siphonostoma- toids); M. Dardeau, Dauphin Island Sea Lab (Synalpheid deca- pods); B. Kensley, Smithsonian Institute (isopods); B. M. Marcotte, Bureau of Marine Science, West Boothbay Harbor, Maine (harpacticods); and F. Raft, Canadian National Museum (isopods and tanaids). This work was supported by a Natural Sciences and Engineering Research Council of Canada operating grant to J.B. Lewis and NSERC Postgraduate Fellowship to P.V.R. Snelgrove.
Literature cited
Abele, L. G. (1976). Comparative species richness in fluctuating and constant environments: coral associated decapod crustaceans. Science, N.Y. 192:461-463
Abele, L. G. (1984). Biogeography, colonization and experimental community structure of coral-associated crustaceans. In: Abele, L. G., Thistle, A.B. (eds.) Ecological communities: conceptual issues and evidence. Princeton University Press, Princeton p. 123-137
Abele, L. G., Patton, W. K. (1976). The size of coral heads and the community biology of associated decapod crustaceans. J. Bio- geogr. 3 :35-47
Austin, A. D., Austin, S. A., Sale, P. E (1980). Community struc- ture of the fauna associated with the coral Poeillopora damicor- his (L.) on the Great Barrier Reef. Aust. J. mar. Freshwat. Res. 31:163-174
Coles, S. L. (1980). Species diversity of decapods associated with living and dead reef coral Pocillopora meandrina. Mar. Ecol Prog. Set. 2:281-291
Dardeau, M. (1984). Synalpheus shrimps (Crustacea: Decapoda: Alpheidae). I. The Gambrelloides group with a description of a new species. Mem. Hourglass Cruises, Florida Dept. Natural Resources, Mar. Res. Lab. 7:1-125
Edwards, A, Emberton, H. (1980). Crustacea associated with the scleractinian coral, Stylophora pistillata (Esper.), in the Su- danese Red Sea. J. exp. mar. Biol. Ecol. 42:225-240
Field, J. G., Clarke, K. R., Warwick, R. M. (1982). A practical strategy for analysing multispecies distribution patterns. Mar. Ecol. Prog. Ser. 8:37-52
Gotelli, N. J., Abele, L. G. (1983). Community patterns of coral associated decapods. Mar. Ecol. Prog. Set. 13:131-139
Gotelli, N. J., Gilchrist, S. L., Abele, L. G. (1985). Population biology of Trapezia spp. and other coral-associated decapods. Mar. Ecol. Prog. Set. 89-98
Grassle, J. F. (1973). Variety in coral reef communities. In: Jones, O. A., Endean, R. (eds.) Biology and geology and coral reefs, vol. 2. Biology I. Academic Press, New York, p. 247-270
Huber, M. E., Coles, S. L. (1986). Resource utilization and compe- tition among the five Hawaiian species of Trapezia (Crustacea, Brachyura). Mar. Ecol. Prog. Set. 30:21-31
Hurlbert, S. H. (1971). The nonconcept of species diversity: a cri- tique and alternative parameters. Ecology 52:577-586
Hutchings, P. (1983). Cryptofaunal communities of coral reefs. In:
J.B. Lewis and P. V. R. Snelgrove: Coral reef cryptofauna
Barnes, D. J. (ed.) Perspectives on coral reefs. Brian Closton, Manuka, Australia p. 200-208
Kensley, B. (1982). Anthuridae (Crustacea:Isopoda) of Carrie Bow Cay, Belize. Smithson Contr. mar. Sci. 12:321-353
Kensley, B. (1984a). New marine Isopoda. Smithson. Contr. mar. Sci. 24:1-81
Kensley, B. (1984b). The role of isopod crustaceans in the reef creast community at Carrie Bow Cay, Belize. Pubbl. Staz. zool. Napolie (I: Mar. Ecol.) 5:29-44
Klumpp, D. W., McKinnon, A. D., Mundy, C. N. (1988). Motile cryptofauna of a coral reef: abundance, distribution and trophic potential. Mar. Ecol. Prog. Ser. 45:95-108
Klumpp, D. W, Polunin, N. V. C. (1989). Partitioning among graz- ers of food resources within damselfish territories on a coral reef. J. exp. mar. Biol. Ecol. 125:145-169
Knudsen, J. W. (1967). Trapezia and Tetralia (Decapoda, Brachyura, Yanthidae) as obligate ectoparasites of pocilloporid and acroporid corals. Pacif. Sci. 21:51-57
Kohn, A. J., Nybakken, J. W. (1975). Ecology of Conus on Eastern Indian Ocean fringing reefs: diversity of species and resources utilization. Mar. Biol. 29:211 234
Lewis, J. B, (1960). The coral reefs and coral communities of Bar- bados, W I. Can. J. Zool. 38:1130-1145
Lewis, J. B. (1965). A preliminary description of some marine ben- thic communities from Barbados, West Indies. Can. J. Zool. 43: 1049-1074
Lewis, J. B., Axelsen, F. I., Goodbody, I., Page, C., Chislett, G. R. (1968). Comparative growth rates of some reef corals in the Caribbean. Mar. Sci. Manuscript Rept. McGill University, Montreal 10:1-26
McCloskey, L. R. (1970). The dynamics of the community associ- ated with a marine scleractinian coral. Int. Revue ges. Hydro- biol. 55:13-81
Patton, W K. (1973). Animal associates of living reef corals. In: Jones, O. A., Endean, R. (eds.) Biology and geology of coral reefs, vol. 3. Biology 2. Academic Press, New York, p. 1-36
Patton, W K. (1974). Community structure among the animals inhabiting the coral Poeillopora damieornis at Heron Island, Australia. In: Vernberg, W. B. (ed.) Symbiosis and the sea. University of South Carolina Press, Columbia, p. 219-243
Pielou, E. C. (1975). Ecological diversity. John Wiley & Sons, New York
Reed, J. K., Gore R. H., Scotto, L. E., Wilson, K. A. (1982). Community composition, structure, areal and trophic relation- ships of decapods associated with shallow- and deep-water Oculina varicosa coral reefs: Studies on decapod crustacea from the Indian River region of Florida, XXIV. Bull. mar. Sci. 32: 761-786
Reed, J. K., Mikkelsen, P. M. (1987). The molluscan community associated with the scleractinian coral Oculina varicosa. Bull. mar. Sci. 40:99-131
Sale, P. E, McWilliam, P. S., Anderson, D. T. (1978). Faunal rela- tionships among the near-reef zooplankton at three locations on Heron Reef, Great Barrier Reef, and seasonal changes in this fauna. Mar. Biol. 49:133-145
Snelgrove, P. V. R., Lewis, J. B. (1989). Response of a coral-associ- ated crustacean community to eutrophication, Mar. Biol. 101: 249-257
Tomascik, T., Sander, R (1985). Effects of eutrophication on reef- building corals I. Growth rates of the reef building coral Mon- tastrea annularis. Mar. Biol. 87:143-155
Tomascik, T., Sander, F. (1987). Effects of eutrophication on reef- building corals. II. Structure of scleractinian coral communities on fringing reefs, Barbados, West Indies. Mar. Biol. 94:53-75
Vivien, M. L., Peyrot-Clausade, M. (1974). A comparative study of the feeding behaviour of three coral reef fishes (Holocentridae), with special reference to the Polychaeta of the reef cryptofauna as prey. Proc. 2nd int. Syrup. coral Reefs. 1 :179-192 [Cameron, A. M. et al. (ed.) Great Barrier Reef Committee, Brisbane]
Zar, J. H. (1984). Biostatistical analysis. Prentice Hall Inc., Engel- wood Cliffs





























