Embed Size (px)
Citation preview

1
Corso di Immunologia
A.A. 2008-09
Genetica immunoglobuline
Generation of diversity (GOD-I)

2
The organization and expression of the immunoglobulin gene families.
The origins of antibody diversity.
KEYWORDS:
V gene, C gene, J region, D region, Leader, Enhancer, Promoter, Antibody
diversity, Germ line theory, Somatic mutation theory, N region insertions,
Junctional diversity, Combinatorial association, Multispecificity, Clonal
selection
Cosa dovremmo conoscere alla fine di questa lezione
3
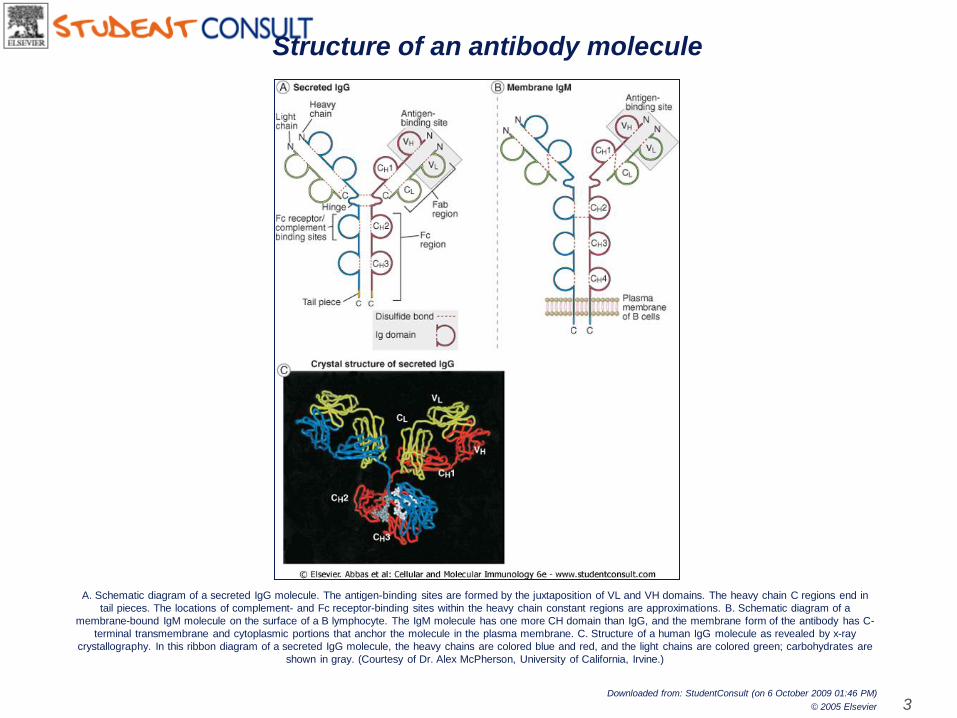
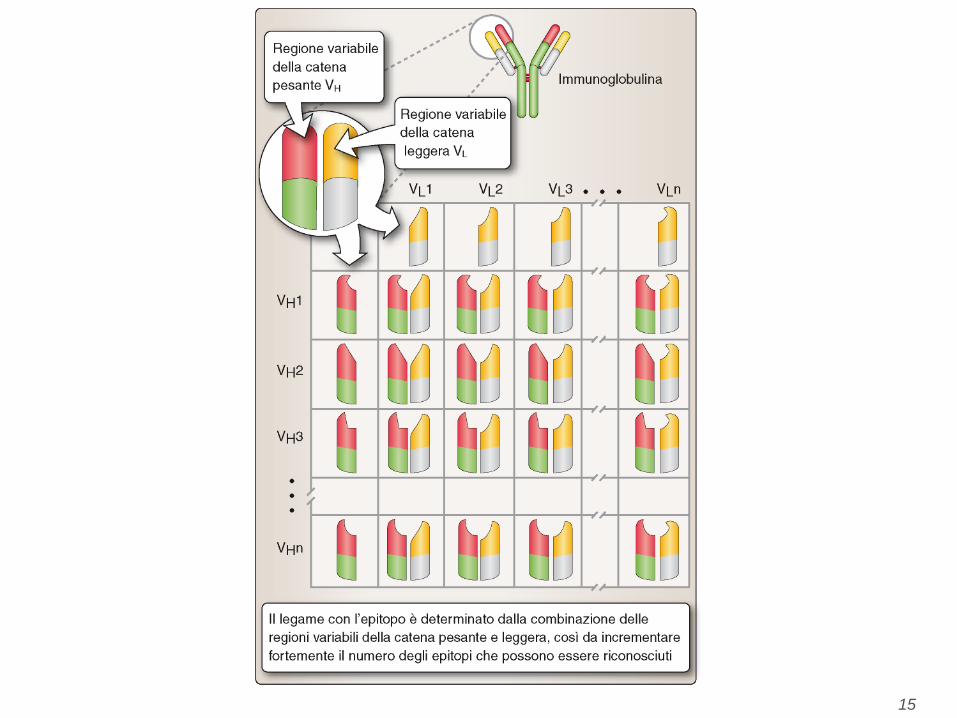
A. Schematic diagram of a secreted IgG molecule. The antigen-binding sites are formed by the juxtaposition of VL and VH domains. The heavy chain C regions end in
tail pieces. The locations of complement- and Fc receptor-binding sites within the heavy chain constant regions are approximations. B. Schematic diagram of a
membrane-bound IgM molecule on the surface of a B lymphocyte. The IgM molecule has one more CH domain than IgG, and the membrane form of the antibody has C-
terminal transmembrane and cytoplasmic portions that anchor the molecule in the plasma membrane. C. Structure of a human IgG molecule as revealed by x-ray
crystallography. In this ribbon diagram of a secreted IgG molecule, the heavy chains are colored blue and red, and the light chains are colored green; carbohydrates are
shown in gray. (Courtesy of Dr. Alex McPherson, University of California, Irvine.)
Downloaded from: StudentConsult (on 6 October 2009 01:46 PM)
© 2005 Elsevier
Structure of an antibody molecule

4
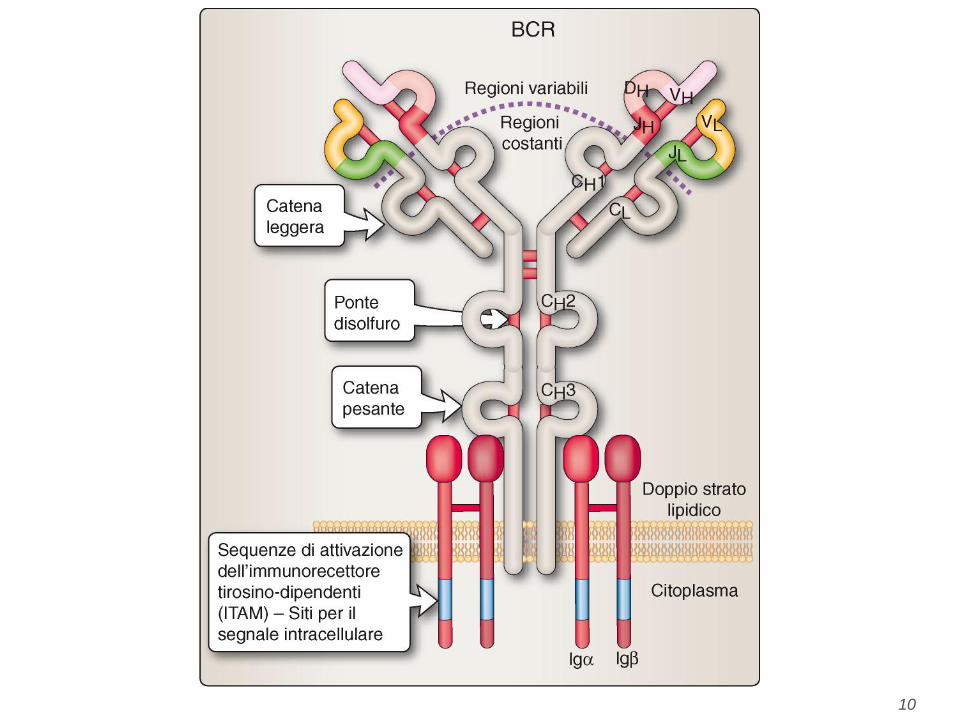
Figure 4-24 Terminologia necessaria
5
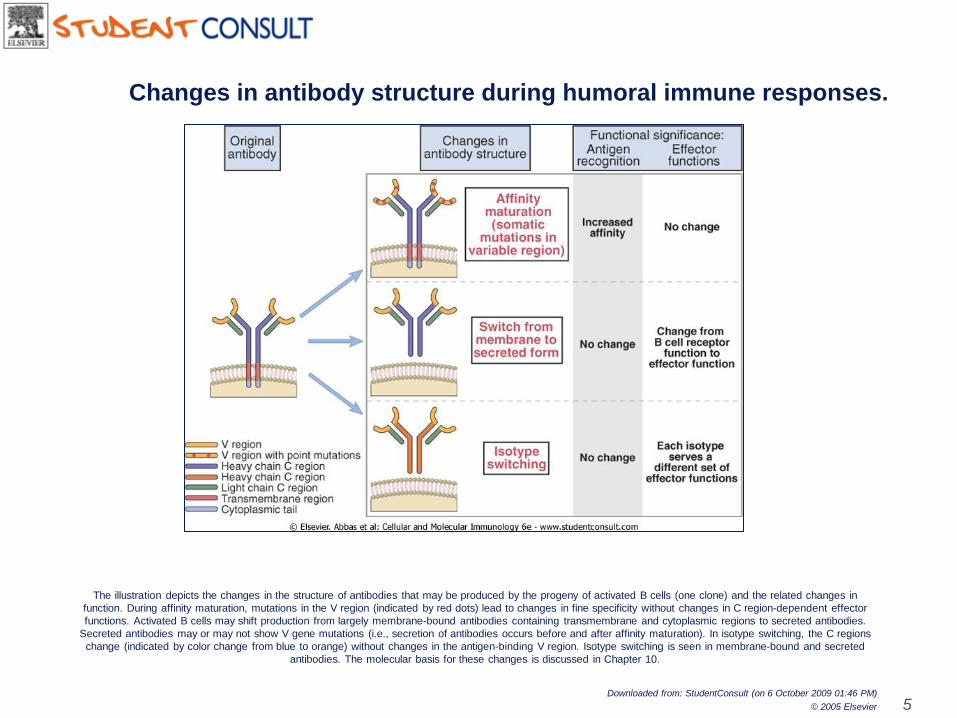
The illustration depicts the changes in the structure of antibodies that may be produced by the progeny of activated B cells (one clone) and the related changes in
function. During affinity maturation, mutations in the V region (indicated by red dots) lead to changes in fine specificity without changes in C region-dependent effector
functions. Activated B cells may shift production from largely membrane-bound antibodies containing transmembrane and cytoplasmic regions to secreted antibodies.
Secreted antibodies may or may not show V gene mutations (i.e., secretion of antibodies occurs before and after affinity maturation). In isotype switching, the C regions
change (indicated by color change from blue to orange) without changes in the antigen-binding V region. Isotype switching is seen in membrane-bound and secreted
antibodies. The molecular basis for these changes is discussed in Chapter 10.
Downloaded from: StudentConsult (on 6 October 2009 01:46 PM)
© 2005 Elsevier
Changes in antibody structure during humoral immune responses.

6
Background Due geni per una proteina.
Nel 1965 Dreyer e Bennet ipotizzarono che le regioni V e C delle
immunoglobuline fossero codificate da 2 geni distinti e tale
ipotesi é stata confermata sperimentalmente da Tonegawa.
Lo studio della variabilità dei geni delle immunoglobuline
rappresenta un bellissimo modello di ricombinazione
somatica.

7
History
Amino acid sequencing data revealed that a single C region
could be associated with many different V regions. Also, it was
shown that a single idiotype could be associated with different
C regions (eg. IgM and IgG). To explain these data it was
suggested that perhaps the two regions of the Ig molecule
were coded by separate genes and that the V and C region
genes were somehow joined before an Ig molecule was made
(i.e. there were two genes for one polypeptide).

8
History
This was a revolutionary concept but with the advent of
recombinant DNA technology, it has been shown to be the
correct. The Ig heavy and light chains are coded by three
separate gene families each one on a separate chromosome -
one for the heavy chain and one for each of the light chain
types. Each of these gene families has several V region genes
and one or more C region genes. The V and C regions genes
are not however immediately adjacent to each other.

9
Immunoglobulin Genetics History
• Same C region could associate with many V regions
– IgG Ab with different specificities
• Same V region could associate with many C regions
– Same idiotype on IgG and IgM Ab
• V and C regions coded for by separate genes
10

11
12

13
14
15

16
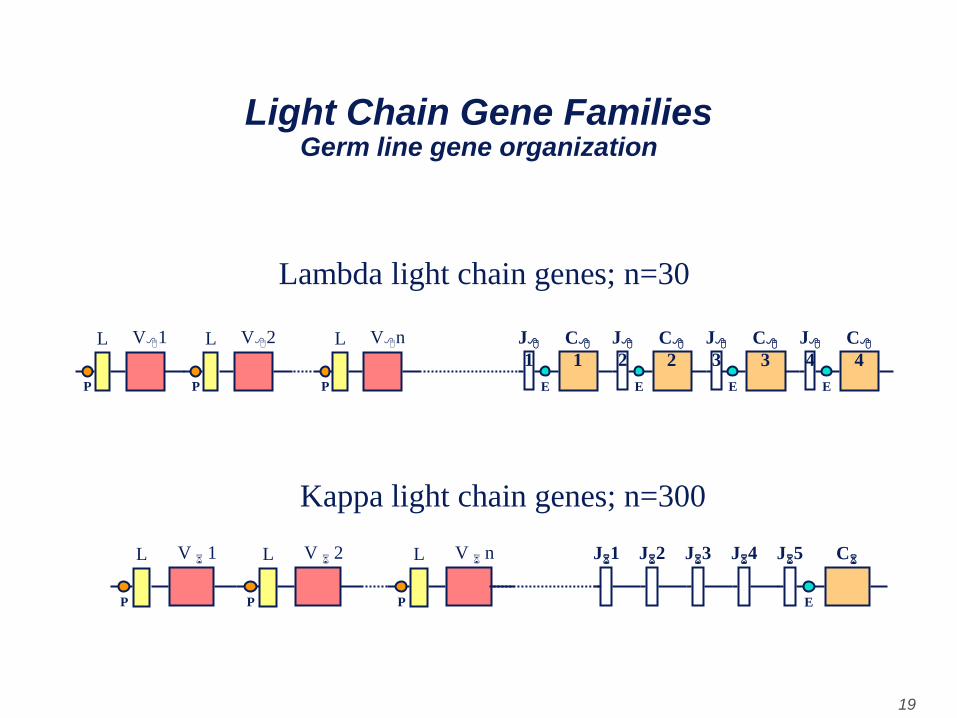
Immunoglobulin Genetics Germ line gene organization
• The organization of the kappa and lambda light
chain genes in the germ line or undifferentiated
cells is depicted in the next Figure.

17
Immunoglobulin Genetics Germ line gene organization
Lambda light chains
• The lambda gene family is composed of 4 C
region genes, one for each subtype of 8 chain,
and approximately 30 V region genes. Each of the
V region genes is composed of two exons, one (L)
that codes for a leader region and the other (V)
that codes for most of the variable region.
Upstream of each of the C genes there is and
additional exon called J (joining). The L, V, J and
C exons are separated by introns (intervening
non-coding sequences).

18
Immunoglobulin Genetics Germ line gene organization
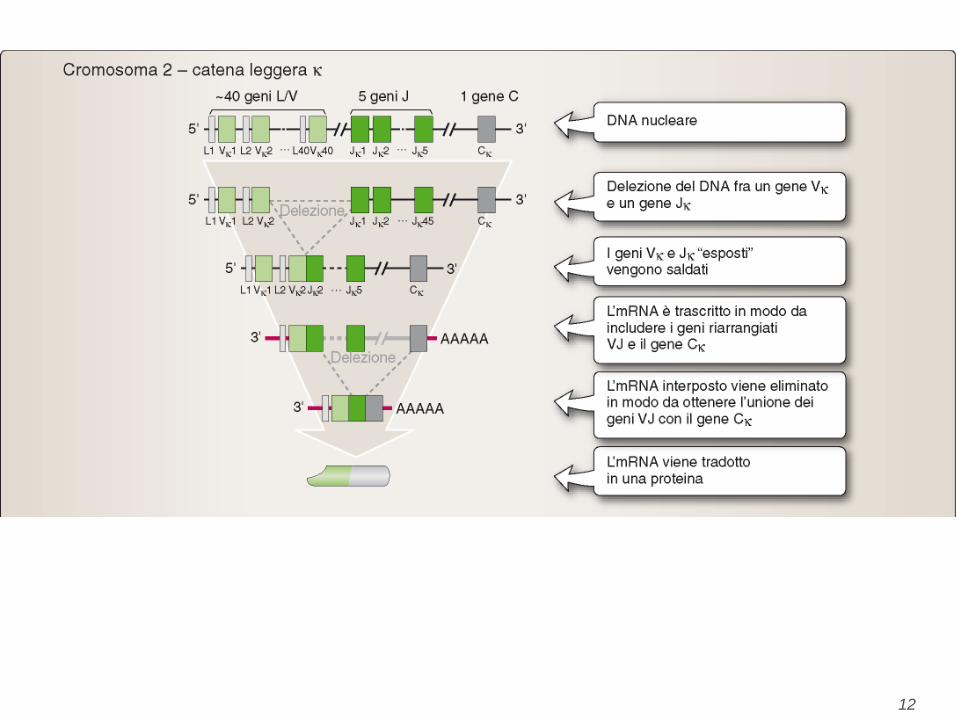
Kappa light chains
• The kappa light chain gene family contains only
one C region gene, since there is only one type of
kappa light chain. There are many V region genes
(approximately 250) each of which has a leader
exon and a V exon. In the 6 gene family there are
several J exons located between the V and C
genes. All of the exons are separated by introns.
19
Light Chain Gene Families Germ line gene organization
J
1
C
1
E
J
2
C
2
E
J
3
C
3
E
J
4
C
4
E P
L V
1
P
L V
n
P
L V
2
Lambda light chain genes; n=30
P
L V 1
P
L V n
P
L V 2 J
2 J
3 J
5 C
E
J1 J
4
Kappa light chain genes; n=300

20
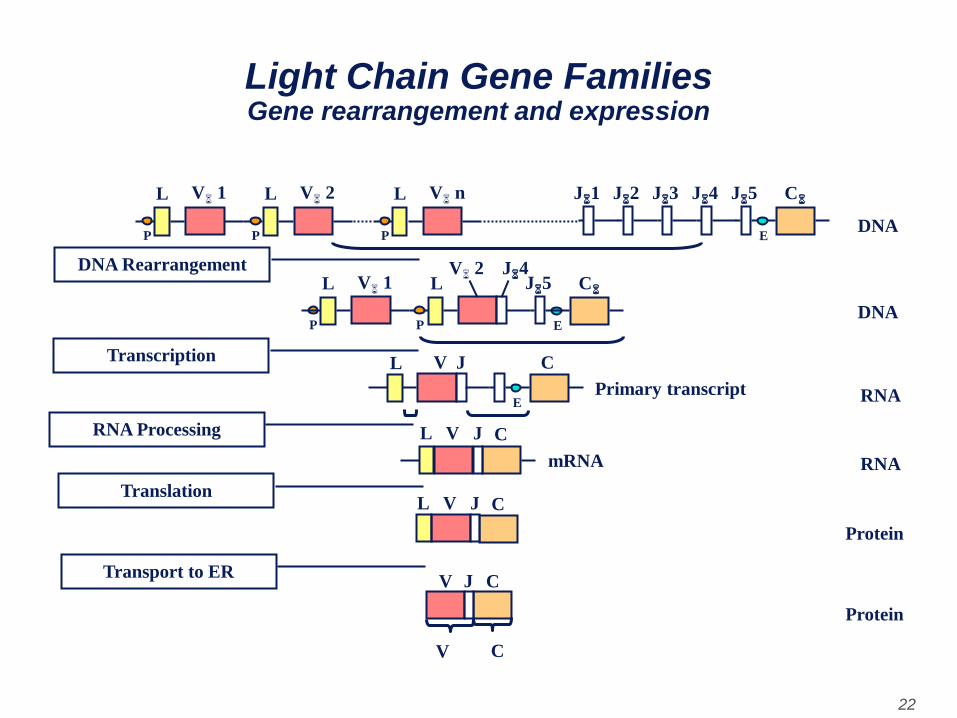
• As a cell differentiates into a mature B cell that will
make a light chain, there is a rearrangement of
the various genes (exons) and the gene begins to
be expressed. As a cell commits to become a B
cell making a light chain, there is a rearrangement
of the genes at the DNA level such that one of the
V genes is brought next to one of the J regions.
Light Chain Gene Families Gene rearrangement and expression

21
• This occurs by a recombination event which
removes the intron between the V and J regions.
The selection of which V gene is used is not
totally random; there is some preference for the
use of V genes nearest to the J regions. However,
with time all V genes can be used so that all
combinations of V genes and J regions can be
generated.
Light Chain Gene Families Gene rearrangement and expression
22
Light Chain Gene Families Gene rearrangement and expression
L V 1 L V
n L V
2 J
2 J
3 J
5 C
J
1 J
4
DNA P P P E
P P E
L V 1 L J
5 C
V 2 J
4
DNA
DNA Rearrangement
V C J
V C
Protein
Transport to ER
Protein
Translation C L V J
E
L V C
J
RNA
Transcription
Primary transcript
C L V J RNA Processing
RNA mRNA

23
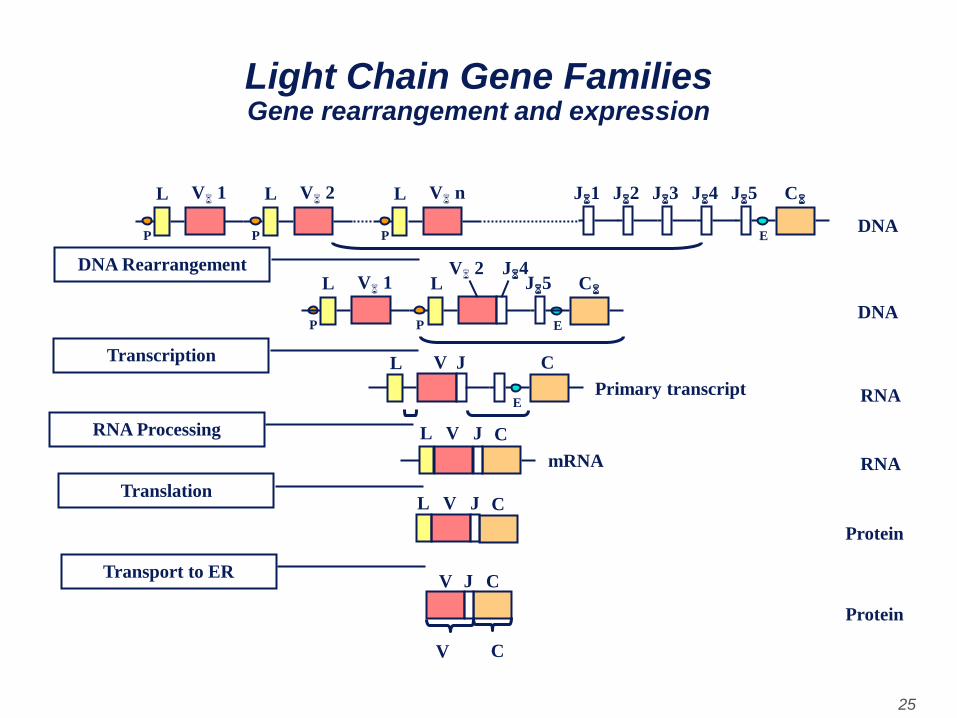
• A consequence of this DNA rearrangement is that
the gene becomes transcriptionally active
because a promoter (P), which is associated with
the V gene, is brought close to an enhancer (E),
which is located in the intron between the J and C
regions.
Light Chain Gene Families Gene rearrangement and expression

24
• As transcription initiates from the promoter a pre-
mRNA is made which contains sequences from
the L, V J and C regions as well as sequences for
the introns between L and V and between J and
C. This pre-mRNA is processed (spliced) in the
nucleus and the remaining introns are removed.
The resulting mRNA has the L, V J and C exons
contiguous.
Light Chain Gene Families Gene rearrangement and expression
25
Light Chain Gene Families Gene rearrangement and expression
L V 1 L V
n L V
2 J
2 J
3 J
5 C
J
1 J
4
DNA P P P E
P P E
L V 1 L J
5 C
V 2 J
4
DNA
DNA Rearrangement
V C J
V C
Protein
Transport to ER
Protein
Translation C L V J
E
L V C
J
RNA
Transcription
Primary transcript
C L V J RNA Processing
RNA mRNA
26
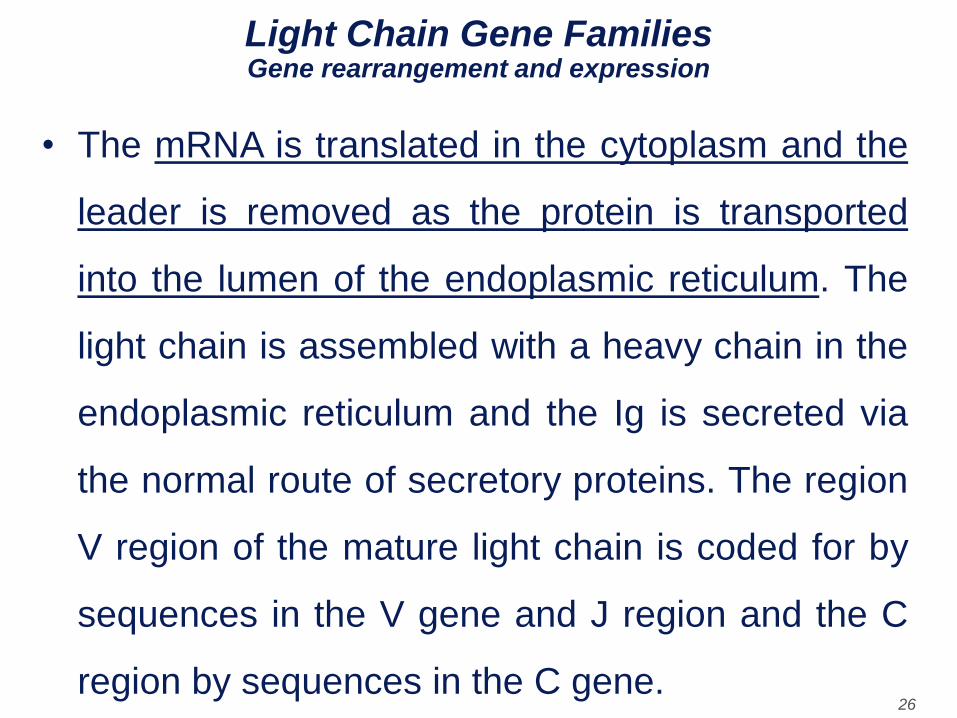
• The mRNA is translated in the cytoplasm and the
leader is removed as the protein is transported
into the lumen of the endoplasmic reticulum. The
light chain is assembled with a heavy chain in the
endoplasmic reticulum and the Ig is secreted via
the normal route of secretory proteins. The region
V region of the mature light chain is coded for by
sequences in the V gene and J region and the C
region by sequences in the C gene.
Light Chain Gene Families Gene rearrangement and expression
27
Light Chain Gene Families Gene rearrangement and expression
L V 1 L V
n L V
2 J
2 J
3 J
5 C
J
1 J
4
DNA P P P E
P P E
L V 1 L J
5 C
V 2 J
4
DNA
DNA Rearrangement
V C J
V C
Protein
Transport to ER
Protein
Translation C L V J
E
L V C
J
RNA
Transcription
Primary transcript
C L V J RNA Processing
RNA mRNA

28
Heavy Chain Gene Family Germ line gene organization
Introns separate exons coding for H chain domains
Heavy chain genes; Vn=1000, Dn=15
P
L V1
P
L Vn
P
L V2 D2 D3 D1 Dn
C
C
C
3
C
C
2
C
1
C
4
C
1
C
2
J2 J3 J5 J1 J4
E
CH1 H CH2 CH3 CH4

29
• In the heavy chain gene family there are many C
genes, one for each class and subclass of Ig.
Each of the C genes is actually composed of
several exons, one for each domain and another
for the hinge region. In the heavy chain gene
family there are many V region genes, each
composed of a leader and V exon.
Heavy Chain Gene Family Germ line gene organization

30
• In addition to several J exons, the heavy chain
gene family also contains several additional exons
called the D (diversity) exons. All of the exons are
separated by introns.
Heavy Chain Gene Family Germ line gene organization

31
Heavy Chain Gene Family Germ line gene organization
Introns separate exons coding for H chain domains
Heavy chain genes; Vn=1000, Dn=15
P
L V1
P
L Vn
P
L V2 D2 D3 D1 Dn
C
C
C
3
C
C
2
C
1
C
4
C
1
C
2
J2 J3 J5 J1 J4
E
CH1 H CH2 CH3 CH4
32
Heavy Chain Gene Family Gene rearrangement and expression
P
L V1
P
L Vn
P
L V2 D2 D3 D1 Dn C
C
J2 J3 J5 J1 J4
E
DJ rearrangement
P
L Vn
P
L V1
P
L V2 D2
D1 C
C
J5 J4
E DNA
VDJ rearrangement
D2 C
C
J5 J4
E P
L V1
P
L V2
DNA
Transcription
D2 C
C
J5 J4
E
L V2
RNA Primary transcript

33
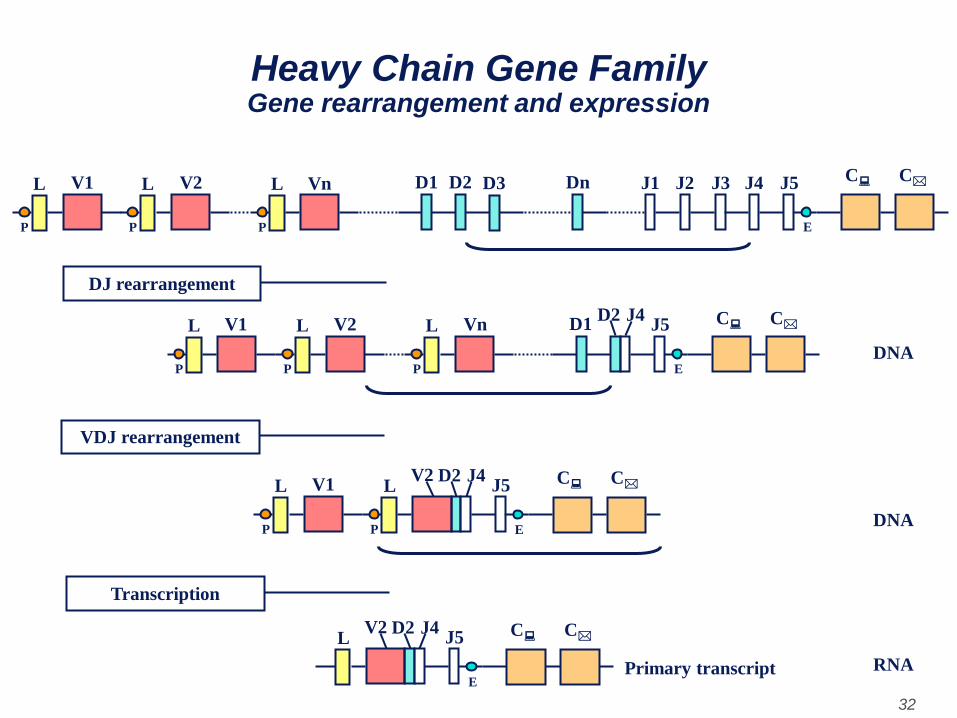
• As a cell differentiates into a mature B cell that will
make a heavy chain, there is a rearrangement of
the various genes segments (exons) and the gene
begins to be expressed.
Heavy Chain Gene Family Germ line gene organization

34
• As a cell commits to become a B cell making a heavy
chain, there are two rearrangements at the DNA level.
First, one of the D regions is brought next to one of the J
regions and then one of the V genes is brought next to
the rearranged DJ region. This occurs by two
recombination events which remove the introns between
the V, D and J regions. As with the light chains the
selection of the heavy chain V gene is not totally random
but eventually all of the V genes can be used.
Heavy Chain Gene Family Germ line gene organization

35
Heavy Chain Gene Family Gene rearrangement and expression
P
L V1
P
L Vn
P
L V2 D2 D3 D1 Dn C
C
J2 J3 J5 J1 J4
E
DJ rearrangement
P
L Vn
P
L V1
P
L V2 D2
D1 C
C
J5 J4
E DNA
VDJ rearrangement
D2 C
C
J5 J4
E P
L V1
P
L V2
DNA
Transcription
D2 C
C
J5 J4
E
L V2
RNA Primary transcript

36
• A consequence of these DNA rearrangements is that the
gene becomes transcriptionally active because a
promoter (P), which is associated with the V gene, is
brought close to an enhancer (E), which is located in the
intron between the J and C: regions. As transcription
initiates from the promoter a pre-mRNA is made which
contains sequences from the L, V, D, J C: and C*
regions as well as sequences for the introns between L
and V, between J and C:, and between C: and C*.
Heavy Chain Gene Family Germ line gene organization
37
Heavy Chain Gene Family Gene rearrangement and expression
P
L V1
P
L Vn
P
L V2 D2 D3 D1 Dn C
C
J2 J3 J5 J1 J4
E
DJ rearrangement
P
L Vn
P
L V1
P
L V2 D2
D1 C
C
J5 J4
E DNA
VDJ rearrangement
D2 C
C
J5 J4
E P
L V1
P
L V2
DNA
Transcription
D2 C
C
J5 J4
E
L V2
RNA Primary transcript

38
• The pre-mRNA is processed (spliced) in the nucleus and
the remaining introns, including those between the
exons in the C genes, are removed. The pre-mRNA can
be processed in two ways, one to bring the VDJ next to
the C gene and the other to bring the VDJ next to the C*
gene. The resulting mRNAs have the L, V, D, J and C: or
C* exons contiguous and will code for a mu and a
gamma chain, respectively.
Heavy Chain Gene Family Germ line gene organization

39
Heavy Chain Gene Family Gene rearrangement and expression
E Primary transcript
E
J4 D2 V2 C
C
J5 L C
C
J5 L D2 J4 V2
RNA Processing
An
mRNA for C
C
L D J V C
L D J V
An
mRNA for C
Translation
C
heavy chain
C
L D J V C
L D J V
C
heavy
chain
Transport to ER
C
heavy chain
V C
C
D J V C
D J V
C
heavy
chain
V C

40
• The mRNAs are translated in the cytoplasm and the
leader is removed as the protein is transported into the
lumen of the endoplasmic reticulum. The heavy chain is
assembled with a light chain in the endoplasmic
reticulum and the Ig is secreted via the normal route of
secretory proteins. The region V region of the mature
heavy chain is coded for by sequences in the V gene, D
region and J region and the C region by sequences in
the C gene.
Heavy Chain Gene Family Germ line gene organization

41
Heavy Chain Gene Family Gene rearrangement and expression
E Primary transcript
E
J4 D2 V2 C
C
J5 L C
C
J5 L D2 J4 V2
RNA Processing
An
mRNA for C
C
L D J V C
L D J V
An
mRNA for C
Translation
C
heavy chain
C
L D J V C
L D J V
C
heavy
chain
Transport to ER
C
heavy chain
V C
C
D J V C
D J V
C
heavy
chain
V C

42

43
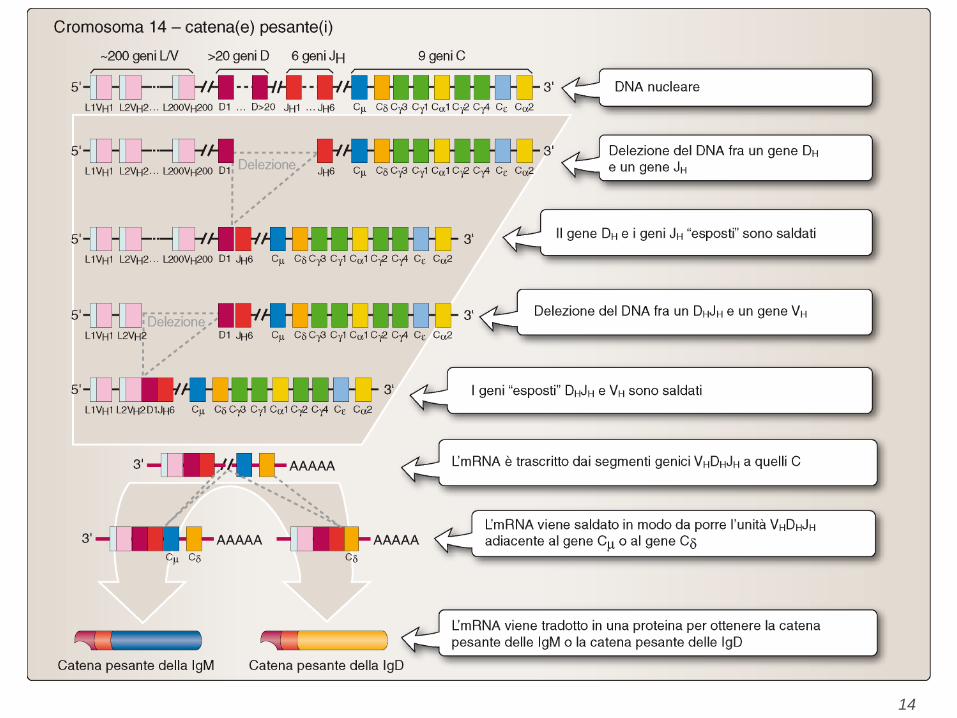
Generation of diversity (1)
• I geni per le catene H, kappa e lambda (L) sono su cromosomi diversi (12,
16, 6 nel topo e 14, 2, 22 nell'uomo) e un primo livello di variabilita`
consiste nella scelta di unire una particolare catena H con una L (livello a).
• I geni V sono composti di 2 subunita` per L (chiamate V e J) e di 3 per H
(chiamate V, D, J, situate in quest'ordine lungo il cromosoma). La subunita`
J e` posta a valle di V ed a monte di C: la sua funzione e` di congiungere
(joining) V a C. La subunita` D prende il nome da "Diversity". Recenti
approssimazioni suggeriscono che vi sono circa 300 geni VH, 300 Vk, 2 Vl
(questo spiega in parte perche` la maggior parte delle Ig sono composte
dalla catena L kappa e non L lambda), 20 geni DH, 5 geni JH, Jk e Jl. Quindi
tramite ricombinazione casuale possiamo ottenere milioni di catene (livello
b) con diverse regioni V.

44
Generation of diversity (2) Nelle cellule non differenziate i geni che codificano per V, J e C (catene L) e i
geni che codificano per V, J, D e C (catene H) sono distanti ed il
differenziamento comporta una delezione di gran parte del DNA frapposto
con finale riarrangiamento dei geni V, J, D e C che verranno a trovarsi
vicino ma non attigui per la presenza di introni. Tale riarrangiamento rende
il gene trascrizionale attivo. Il complesso V-J-(D)-C viene trascritto in un
lungo pre-mRNA che a livello nucleare viene modificato mediante taglio
degli introni e ricongiunzione (splicing) (livello c). Il meccanismo con cui
V-J e V-D-J si uniscono e` dovuto alla presenza, immediatamente dopo e
prima di ciascun segmento genico, di sequenze omologhe di 7 (eptamero)
e di 9 (nonamero) basi che risultando complementari consentiranno
l'unione V-J, V-D-J. Dodici basi dopo l'eptamero e prima del nonamero e 23
basi presenti dopo il nonamero e prima dell'eptamero rappresentano i
corrispottevi segnali di giunzione.

45
Generation of diversity (3)
• La diversita` anticorpale e` inoltre aumentata considerevolmente dal fatto
che si verificano slittamenti di alcuni nucleotidi nelle giunzioni V-D-J e V-J,
con conseguente creazione di diverse sequenze amminoacidiche. Questi
slittamenti modificano solo di 1-2 aminoacidi la regione variabile, ma
questa modificazione e` sufficiente per alterare la conformazione del
dominio V e quindi il legame Ag-Ig (livello d ).
• Inoltre si e` osservato che i geni V vanno incontro a mutazione somatica
nel corso della proliferazione cellulare indotta dall' interazione con l'
antigene molto piu` frequentemente dei geni C. Le mutazioni somatiche
(livello e) avvengono principalmente per le IgG ed IgA e sono responsabili
della modificazione dell'affinita` per l' antigene delle Ig. Questi livelli di
diversita` portano a produrre almeno 1-100 miliardi di molecole Ig diverse
partendo da un limitatissimo numero di geni.

46

47
• Flanking the V, J and D exons there are unique
sequences referred to as recombination signal
sequences (RSS), which function in recombination.
Each RSS consists of a conserved nonamer and a
conserved heptamer that are separated by either 12 or
23 base pairs. The 12bp and 23 bp spaces correspond
to one or two turns of the DNA helix.
Mechanism of DNA Rearrangements
48
Mechanism of DNA Rearrangements
• Recombination
signal sequences
(RSS)
– Nonamer
– Heptamer
– 1 or 2 turn signals
• Rag-1 and Rag-2
evoluzione
L V
2
J C
1
Lambda light chains
L V
1
J C
2
Kappa light chains
Heavy chains
L V
2
J C
2 1 1
D
Heptamer
CACAGTG
GTGTCAC
Nonamer
ACAAAAACC
TGTTTTTGG
23 bp
23 bp
Two turn
RSS 2
Nonamer
GGTTTTTGT
CCAAAAACA
Heptamer
CACTGTG
GTGACAC
12 bp
12 bp
One turn
RSS 1

49
• Recombination only occurs between a 1 turn and a 2
turn signal. In the case of the 8 light chains there is a 1
turn signal upstream of the J exon and a 2 turn signal
downstream of V8. In the case of the 6 light chains there
is a 1 turn signal downstream of the V6 gene and a 2
turn signal upstream of the J exon. In the case of the
heavy chains there are 1 turn signals on each side of the
D exon and a 2 turn signal downstream of the V gene
and a 2 turn signal upstream of the J exon. Thus, this
ensures that the correct recombination events will occur.
Mechanism of DNA Rearrangements

50
Mechanism of DNA Rearrangements
• Recombination
signal sequences
(RSS)
– Nonamer
– Heptamer
– 1 or 2 turn signals
• Rag-1 and Rag-2
L V
2
J C
1
Lambda light chains
L V
1
J C
2
Kappa light chains
Heavy chains
L V
2
J C
2 1 1
D
Heptamer
CACAGTG
GTGTCAC
Nonamer
ACAAAAACC
TGTTTTTGG
23 bp
23 bp
Two turn
RSS 2
Nonamer
GGTTTTTGT
CCAAAAACA
Heptamer
CACTGTG
GTGACAC
12 bp
12 bp
One turn
RSS 1

51
• The recombination event results in the removal of the
introns between V and J in the case of the light chains or
between the V, D, and J in the case of the heavy chains.
The recombination event is catalyzed by two proteins,
Rag-1 and Rag-2. Mutations in the genes for these
proteins results in a severe combined immunodeficiency
disease (both T and B cells are deficient), since these
proteins and the RSS are involved in generating both
the B and T cell receptors for antigen.
Mechanism of DNA Rearrangements

52
Mechanism of DNA Rearrangements
• Recombination
signal sequences
(RSS)
– Nonamer
– Heptamer
– 1 or 2 turn signals
• Rag-1 and Rag-2
L V
2
J C
1
Lambda light chains
L V
1
J C
2
Kappa light chains
Heavy chains
L V
2
J C
2 1 1
D
Heptamer
CACAGTG
GTGTCAC
Nonamer
ACAAAAACC
TGTTTTTGG
23 bp
23 bp
Two turn
RSS 2
Nonamer
GGTTTTTGT
CCAAAAACA
Heptamer
CACTGTG
GTGACAC
12 bp
12 bp
One turn
RSS 1

53
• An individual B cell only produces one type of light chain
and one class of heavy chain. (N.B. The one exception is
that a mature B cell can produce both mu and gamma
heavy chains but the antibody specificity is the same since
the same VDJ region is found on the mu and gamma
chains. Since any B cell has both maternal and paternal
chromosomes which code for the Ig genes there must be
some orderly way in which a cell expresses its Ig genes so
as to ensure that only one type of light chain and one class
of heavy chain is produced.
Order of Ig Gene Expression
54
Order of Ig Gene Expression
+
-
Heavy chain gene
VDJ recombination
Productive rearrangement ?
Yes
No
Transcribe/translate
1st
2nd
or
Apoptosis
Progenitor B cell
Pre B cell
() Heavy chain
Paternal Maternal
55
Order of Ig Gene Expression
+
-
Light chain gene
VJ recombination
Productive rearrangement ?
Yes
No
Transcribe/translate
Light chain
4 th
Apoptosis
1st -
2nd -
3rd -
4th -
or
Pre B cell
Mature B cell
+

56
• When a cell successfully rearranges a heavy chain gene, it
then begins to rearrange one of its kappa light chain genes. It
is a random event whether the maternal or paternal kappa
light chain genes are selected. If the rearrangement is
unsuccessful (i.e. it does not produce a functional kappa light
chain), then it attempts to rearrange the kappa genes on the
other chromosome. If a cell successfully rearranges a kappa
light chain gene, it will be a B cell that makes an Ig with a
kappa light chain.
Order of Ig Gene Expression Kappa light chains

57
• If a cell is unsuccessful in rearranging both of its kappa light
chain genes, it then attempts to make a lambda light chain. It
is a random event whether the maternal or paternal lambda
light chain genes are selected. If the rearrangement is
unsuccessful (i.e. it does not produce a functional lambda light
chain), then it attempts to rearrange the lambda genes on the
other chromosome. If a cell successfully rearranges a lambda
light chain gene, it will be a B cell that makes an Ig with a
lambda light chain.
Order of Ig Gene Expression Lambda light chains

58
Order of Ig Gene Expression Explains:
• Why an individual B cell only produces one
kind of Ig with one kind of heavy and one
kind of light chain
• Why an individual B cell makes only makes
antibodies of one specificity
• Why there is allelic exclusion

59
Origin of Antibody Diversity Bakground
• Antibody diversity refers to the sum total of all
the possible Ab specificities that an organism can
make. It is estimated that we can make 107 – 108
different Ab molecules. One of the major
questions in immunology has been how can we
make so many different antibody molecules.
Theories which have attempted to explain the
origin of antibody diversity fall into two major
categories.

60
Origin of Antibody Diversity History
• Definition – Diversity (repertoire) is the total of
all the Ab specificities that an organism is
capable of expressing
• Theories
– Germ line theory - This theory states that we have a
different V region gene for each possible antibody we
can make.
– Somatic mutation theory - This theory state that we
have only one or a few V region genes and the
diversity is generated by somatic mutations which
occur in these genes.

61
Origin of Antibody Diversity Current concepts
• Our current thinking is that both the germ line
and somatic mutation theories have some merit.
It is thought that antibody diversity is generated
by the following mechanisms.
1 - Large number of V genes
a) 30 lambda V genes
b) 300 kappa V genes
c) 1000 heavy chain V genes

62
Origin of Antibody Diversity Current concepts
2 - V-J and V-D-J joining
The region where the light chain V gene and J region or
the heavy chain V gene and D and J regions come together
is in the 3rd hypervariable region. Since it is random
which V and which J or D regions come together, there
is a lot of diversity that can be generated by V-J and V-
D-J joining.

63
Origin of Antibody Diversity Current concepts
3. Junctional diversity
Inaccuracies in V-J and V-D and D-J recombination.
Recombination between V-J and V-D-J is not always
perfect and additional diversity can arise by errors that
occur in the recombination event that brings the V
region next to the J or D regions or the D region next to
the J region. It is estimated that these inaccuracies can
triple the diversity generated by V-J and V-D-J joining.
The diversity generated by this mechanisms is occurring
in the 3rd hypervariable region and thus, is directly
affecting the combining site of the Ab.

64
Generation of diversity (4) Schema dei livelli che portano alla Diversita` anticorpale.
a) combinazione H-L
b) numero dei geni VH, DH, JH, VL, JL
c) meccanismi ricombinatoriali (VH x DH x JH;
VL x JL)
d) slittamenti giunzionali (VH-DH; DH-JH; VL-
JL)
e) mutazione somatica

65
Origin of Antibody Diversity Current concepts
• Multiple V genes
• V-J and V-D-J joining
• Junctional diversity
• N region insertions
– Amino acid
sequences not
encoded in the germ
line
D region
T G G C C C
Pro Trp
C C C C C C
V gene
T G G
D region
C C C C C C
V gene
T G G
C G G C C C
Pro Arg
C C C C C C
V gene
T G G
D region
C G C C C C
Pro Pro

66
-GTCAATG
-GTCAATG- +
Break DNA
TdT adds nucleotides
Ligate
N region insertion Intron
J D
J
J
J
D
D
D
Generation of N region insertions

67
Origin of Antibody Diversity Current concepts
4. N region insertion
At the junction between D and J segments there is often an
insertion of a series of nucleotides which is catalyzed by
the enzyme terminal transferase. (Terminal transferase
catalyzes the radon polymerization of nucleotides into
DNA without the need for a template. This leads to
further diversity in the 3rd hypervariable region.

68
Origin of Antibody Diversity Current concepts
5. Somatic mutation
There is evidence that somatic mutations are occurring in
the V gene, particularly in the place that codes for the
2nd hypervariable region. Thus, somatic mutation
probably contributes to Ab diversity to some extent.

69
Origin of Antibody Diversity Current concepts
6. Combinatorial association
Any individual B cell has the potential to make any one of
the possible heavy chains and any one of the possible
light chains. Thus, different combinations of heavy and
light chains within an individual B cell adds further
diversity.

70
Origin of Antibody Diversity Current concepts
7. Combinatorial association
Due to cross reactions between antigenic determinants of
similar structure an antibody can often react with more
than one antigenic determinant. This is termed
multispecificity. Multispecificity also contributes to Ab
diversity.

71

72
Generation of Diversity
B cell receptor (Immunoglobulin)
Heavy Light
V gene segments 1000 300
D gene segments 15 -
J gene segments 4 4
N region insertion ++ -
Junctional diversity +++ +
Somatic mutation + +
V x D x J
1000 x 15 x 4 V x J
300 x 4
Total 6 x 104 1.2 x 10
3
Combinatorial association 7.2 x 107
• Ag independent process • Clonal selection
These calculations do not take into consideration the contributions of lambda light
chains, somatic mutation junctional diversity, N region insertions or multispecificity.

73
Generation of Diversity
The process of gene rearrangement of the heavy and
light chains and the combinatorial association of
these chains occurs during B cell development and
is independent of antigen. Clones of B cells
expressing all of the possible antibody specificities
are produced during development and antigen
simply selects those clones which have the
appropriate receptor. The selected clones are then
activated, proliferate and differentiate into antibody
secreting plasma cells.

74
Commutazione di classe (1)
• Per commutazione di classe si intende la capacità che hanno
i linfociti B che esprimono sulla membrana IgM ed IgD, di
esprimere Ig con diversa regione C ma con la stessa regione V
(medesimo allotipo ed idiotipi ma diverso isotipo). Alla base
della commutazione di classe ci sono una serie di
riarrangiamenti che consistono nel trasferimento delle sequenze
geniche V-D-J da C ad altre sequenze C che codificano per gli
altri isotipi di catene pesanti. Tali fenomeni coinvolgono la
ricombinazione di speciali segmenti, chiamati regioni " Switch"
(regioni S), che precedono di circa 2-3 kbp ogni gene CH
eccetto C . Ciascun gene CH é costituito da tanti esoni quanti
sono i singoli domini che costituiscono le porzioni costanti delle
catene pesanti (3 per IgG, IgA, IgD e 4 per IgM ed IgE. Tra gli
esoni CH1 e CH2 dei geni C sono presenti delle porzioni geniche
H (hinge) che codificano per la regione cerniera.

75
Commutazione di classe (2)
Sono stati proposti due meccanismi molecolari per spiegare la
commutazione di classe e quindi la ricombinazione S-S. Tale
evento infatti può avvenire o in seguito a delezione di
materiale genico e quindi essere un evento unidirezionale ed
irreversibile o con scambio tra cromatidi fratelli e quindi
introduzione di più copie di geni C senza eliminazione di
materiale genetico. Inoltre, le interazioni specifiche tra
differenti regioni S possono essere attivate da proteine che
riconoscono sequenze specifiche delle regioni S.
Ogni singolo clone linfocitario può commutare più volte fino
all'espressione di IgA, il cui gene é ultimo nella regione C
(organizzazione genica del topo).
Il fenomeno della commutazione di classe è regolato da fattori
solubili prodotti dai linfociti T (citochine).

76

77
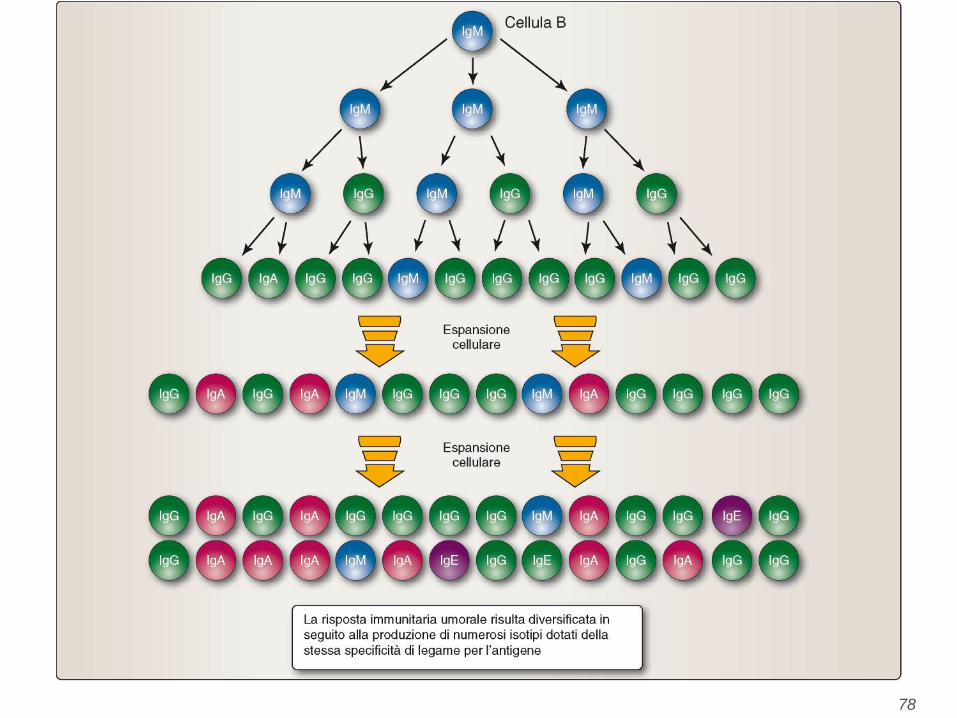
78

79
Commutazione Ig-membrana - Ig-secretoria (Igm-Igs).
Le Ig di membrana sono ancorate alla cellula grazie alla
presenza nella porzione carbossiterminale di 41 aa idrofobici.
Tale porzione è codificata da due piccoli esoni (M1 e M2) che
giacciono al di là degli esoni che codificano per la porzione CH.
Quando un linfocita B in seguito alla interazione con l'antigene
inizia a secernere Ig, avviene un fenomeno di "switch" che
coinvolge la processazione del RNA trascritto. Infatti la
trascrizione dell'RNA termina prima degli esoni M1 e M2 ed
una piccola porzione (S) dell' ultimo esone CH regola la
porzione carbossiterminale. Quindi se la trascrizione si arresta
in corrispondenza di S, si ottiene una Ig secretoria, mentre se
la trascrizione procede includendo gli esoni M1 e M2, la
saldatura rimuove S e si avrà una Ig di membrana. Nella
plasmacellula esiste un meccanismo in grado di far bloccare la
trascrizione prima di M1 e M2 e così tutte le Ig saranno di
tipo secretorio.

80
I linfociti risultano dotati di doppi cromosomi (alleli), uno di
origine paterna ed uno di origine materna. Quindi nel corso
del differenziamento, ogni linfocita dovra` scegliere non solo
quale catena kappa o lambda (esclusione isotipica) ma
anche quale allele utilizzare (esclusione allelica). Questi
due fenomeni di esclusione comportano che ogni linfocita B
debba scegliere uno dei 4 geni L ed uno dei 2 H. Durante lo
sviluppo prima viene attivato un gene H (quindi si troveranno
solo catene H intracitoplasmatiche, linfocita pre-B) .
Esclusione allelica ed esclusione isotipica (1)

81
Avvenuta l'espressione del gene H, avviene il riarrangiamento
dei geni L, in particolare prima riarrangiano i geni k, ed
appena il riarrangiamento é funzionale, cessa ogni altro tipo
di riarrangiamento a carico dei geni L. Le catene M e k si
assemblano e muovono verso la membrana dove si
ancorano. Se invece i due loci falliscono, i geni
riarrangiano e si avranno Ig di membrana
Esclusione allelica ed esclusione isotipica (2)

82
• A cell first attempts to rearrange one of its heavy chain genes;
in some cells the maternal chromosome is selected and in
others the paternal chromosome is selected. If the
rearrangement is successful so that a heavy chain is made,
then no further rearrangements occur in the heavy chain
genes. If, on the other hand, the first attempt to rearrange the
heavy chain genes is unsuccessful (i.e. no heavy chain is
made), then the cell attempts to rearrange the heavy chain
genes on its other chromosome. If the cell is unsuccessful in
rearranging the heavy chain genes the second time, it is
destined to be eliminated.
Order of Ig Gene Expression Heavy chains





























