Embed Size (px)
Citation preview

AMERICAN ASSOCIATION OF NEUROMUSCULAR & ELECTRODIAGNOSTIC MEDICINE
AMERICAN ASSOCIATION OF NEUROMUSCULAR & ELECTRODIAGNOSTIC MEDICINE
2621 Superior Drive NWRochester, MN 55901
(507) 288-0100
CRANIAL NE
RVE TESTIN
G
AA
NEM
WO
RK
SHO
PJosep
Valls-Solé,
MD, PhD
Mark Hallett,
MD

AMERICAN ASSOCIATION OF NEUROMUSCULAR & ELECTRODIAGNOSTIC MEDICINE
Copyright © September 1997
2621 Superior Drive NWRochester, MN 55901
Workshop handouts are prepared as background didactic material to complement a hands-on workshop
session. This workshop handout was originally prepared in May 1984 and was revised in September
1997. The idea and opinions in this publication are solely those of the author(s) and do not necessarily
represent those of the AANEM.

Cranial Nerve TestingAn AANEM Workshop
Josep Valls-Solé, MD, PhDUnitat d’EMG, Servei de Neurologia
Hospital ClinicBarcelona, Spain
Mark Hallett, MDNational Institute of Neurological Disorders and Stroke
National Institutes of HealthBethesda, MD
ANATOMY OF THE CRANIAL NERVES
The intracranial portions of cranial nerves I (olfactory) and II(optic) lie at the supratentorial level. These nerves are notstudied in routine electrodiagnosis, therefore they are notcovered in this handout. All the remaining cranial nerves havetheir nuclei in the brainstem and run through the posteriorfossa.
Cranial Nerves III, IV, and VI
Cranial nerves III (oculomotor), IV (trochlear), and VI (ab-ducens) are involved with movements of the eyes, and the ocu-lomotor is also involved with autonomic innervation of thepupil. The nuclei of the oculomotor and trochlear nerves arelocated in the mesencephalon, and that of the abducens islocated in the lower pons. They all go through the cavernoussinus and enter the orbit through the superior orbital fissure.The oculomotor nerve innervates the medial, superior, and in-ferior recti, the inferior oblique, and the levator palpebraemuscles, as well as the ciliary and pupilloconstrictor muscles.The trochlear nerve innervates the superior oblique muscle, andthe abducens nerve innervates the lateral rectus. There are noknown sensory fibers carried by cranial nerves III, IV, and VI.Proprioceptive information from extraocular muscles reachesthe brainstem via the trigeminal afferents1 and have an impor-tant contribution to visuo-spatial localization.
Lesions of cranial nerves III, IV, and VI can occur in patientswith metabolic diseases such as diabetes mellitus, and manifestclinically by diplopia. The lesion is usually located in the nerve
trunk, but detailed examination may provide evidence for brain-stem involvement.32 Damage to the parasympathetic fiberscarried by the third cranial nerve cause pupillary mydriasis.Horner’s syndrome, featuring unilateral ptosis, miosis, and lossof sweating, is due to lesions involving the sympathetic system,whose fibers join the third cranial nerve at the cavernous sinus.Horner’s syndrome can be due to preganglionic lesions of thesympathetic tract in the brainstem, as in Wallenberg’s syndrome,or to postganglionic lesions of the sympathetic fibers, as inlesions of the lower cervical roots.
Cranial Nerve V
The trigeminal nerve innervates the muscles involved with mas-tication and receives muscle and cutaneous afferents from allthe face and the skull. The motor nucleus is located in the midpons. The motor axons emerge from the brainstem on thelateral aspect of the pons and run with the mandibular sensorybranch to innervate the masseter, the temporalis, and the ptery-goid muscles. The trigeminal sensory nucleus is a complexstructure that is divided into many subnuclei extending from themesencephalon to the spinal cord. The first-order afferents car-rying proprioceptive sensation have their cell bodies in the mes-encephalic nucleus, which receives input from muscle receptorsof trigeminal, extraocular, and possibly other cranial and facialmuscles. The monosynaptic connection of muscle spindle af-ferents from the masseter muscle with the motor nucleus of thetrigeminal nerve provides the anatomical substrate for the mas-seter reflex. The first-order afferents carrying cutaneous sensa-tion have their cell bodies in the Gasserian ganglion and projectto the pontine and spinal nuclei of the trigeminal nerve. The

pontine nucleus is concerned with tactile sensation, and the bul-bospinal nucleus with temperature and pain sensations. Theophthalmic and maxillary divisions of the trigeminal nerve carrysensation from the upper and middle parts of the face, respec-tively, whereas the mandibular division carries sensory fibersfrom the lower portion of the face, as well as motor fibers.
Lesions of the trigeminal nerve can occur at various levels.Selective lesions of the sensory branches are uncommon, butthe Gasserian ganglion, which lies outside the blood-brainbar-rier, is subject to autoimmune attack. Most common are thesensory syndromes caused by brainstem vascular lesions, suchas Wallenberg’s syndrome.
Cranial Nerve VII
The facial nerve nucleus is located in the pons. The course ofthe facial nerve fibers can be artificially subdivided into four seg-ments: intrapontine, intracranial, interosseous, and extracranial.The most central portion is the intrapontine segment, which isinitially directed posteriorly to hook around the sixth nervenucleus. The intracranial segment is the segment stretchingfrom the emergence of the facial nerve from the brainstem atthe caudal pons, in the area of the cerebellopontine angle, to theinternal auditory meatus. Here the facial nerve begins thelongest and most complex interosseous course of any nerve inthe body. Upon exiting from the skull through the stylomastoidforamen, the extracranial facial nerve branches with some vari-ation into five distal segments: temporalis, zygomaticus, buc-calis, marginalis mandibulae, and colli. The facial nerve does notcarry cutaneous sensory fibers. The so-called sensory root ofthe facial nerve is made up of taste afferents from the anteriortwo thirds of the tongue that run with the nerve intermediariusof Wrisberg to terminate in the nucleus of the tractus solitariustogether with afferents from the ninth and tenth cranial nerves.
The facial nerve may be injured in many ways. Brainstemlesions affecting the nucleus or the pontine segment will causedysfunction similar to that of a nerve lesion. Acoustic neuromasor other tumors may cause a facial nerve lesion at the cerebello-pontine angle. Intrapetrossal nerve swelling is likely the cause ofBell’s palsy. Extracranially, the nerve can be damaged by dis-eases affecting the parotid gland.
Cranial Nerve VIII
Cranial nerve VIII is formed from two nerves, the acoustic andthe vestibular. They run together but have different nuclei anddifferent central connections and mediate different functions.Both nerves are concerned with special somatic functions anddo not contain motor axons. The acoustic nerve carries sensoryinformation from the cochlea to the medial geniculate bodiesand the auditory cortex. The vestibular nerve carries sensory in-
formation from the organs of balance in the ear to the vestibu-lar nuclei in the floor of the fourth ventricle and, from them, toother structures of the brainstem and cerebellum.
The acoustic and vestibular nerves can be damaged in the pos-terior fossa by cerebellopontine angle tumors and be involved ina diversity of disorders.
Cranial Nerves IX and X, and the Cranial Root of XI
The glossopharyngeal and vagus nerves and the cranial root ofthe accessory nerve innervate the laryngeal muscles. The cellsof origin and termination are located in the same nuclei for allthree nerves. The motor nucleus is the nucleus ambiguous, sit-uated within the bulbar reticular formation. The neurons of thenucleus ambiguous innervate striated muscles but are in closecontact with structures of the autonomic nervous system.Autonomic and somatic fibers run along the glossopharyngealand vagus nerves to innervate the muscles of the pharynx,larynx, and most of the visceral organs. The cranial root of theaccessory nerve merges with the vagus nerve after a shortsegment. The sensory nucleus is the nucleus of the tractus soli-tarius, which receives the central projections from the cellslocated in the ganglia of the glossopharyngeal, vagus, and facial(intermedius of Wrisberg) nerves.
Isolated lesions of the glossopharyngeal nerve are very rare. Thevagus nerve is one of the longest nerves in the human body, andcan be damaged in many circumstances. Focal lesions of thenerve branches to the vocal cords can occur with respiratory in-tubation. The distal segments of the vagus nerve are often in-volved in dying-back axonal peripheral neuropathies, and thiscan lead to gastrointestinal and cardiac dysfunction.
The Spinal Root of the XI, and the Cranial Nerve XII
The spinal root of the accessory nerve is made of nerves fromthe first five spinal segments which enter the cranial cavitythrough the foramen magnum. It innervates the sternocleido-mastoid, trapezius, and levator scapula muscles. Although thesemuscles receive additional nerve supply directly from the C2-C4roots, it has been suggested that the spinal accessory nerve pro-vides the sole motor supply, while the innervation from the cer-vical roots is purely proprioceptive. The hypoglossal nervesupplies the muscles of the tongue. Its nucleus is located at thelower part of the medulla, in the floor of the fourth ventricle.The accessory and hypoglossal nerves do not carry cutaneoussensory fibers.
Lesions of the eleventh and twelfth cranial nerves are uncom-mon. The accessory nerve can be damaged in subjects carryingheavy objects on their shoulders. The hypoglossal nerve can bedamaged in brainstem strokes. The accessory and the hy-
2 Cranial Nerve Testing AANEM Workshop

poglossal nerves can both be used for anastomosis with thefacial nerve when it has been damaged.
PHYSIOLOGY
Muscles innervated by cranial nerves are activated during spon-taneous, voluntary, or reflex movements. Excitatory impulsesreach the motoneurons by means of monosynaptic or polysy-naptic connections from the corticonuclear tract, the reticularformation, the trigeminal system, or other sources.37,62,76 Themotoneuron may reach its firing threshold with a single excita-tory input or more often with a combination of inputs from dif-ferent sources. Integration of the sensory afferent inputs intothe commands for voluntary activation is an important aspect ofthe physiology of the motor system.29,31,51 Sensorimotor integra-tion can be examined using the normal brainstem excitatorypolysynaptic reflex known as the blink reflex.
Spontaneous Movements
In normal humans, relevant spontaneous movements involvingcranial muscles occur with activities such as blinking and swal-lowing. The spontaneous blink rate is about 20 blinks perminute in a normal individual in quiet resting, but varies con-siderably according to mental or physical activity. Parkinsonianpatients have a significantly reduced blink rate, while schizo-phrenic patients have an enhanced blink rate. These findingspoint to a direct correlation between dopaminergic systems andblinking frequency.42 The physiology and kinematics of blinkingin animals and humans have been extensively studied byEvinger and his coworkers.18,19
Swallowing is another natural involuntary movement thatoccurs repeatedly in awake humans. Swallowing requires the co-ordinated activity of many muscles innervated by differentcranial nerves. Most of these muscles are not accessible to con-ventional electromyography (EMG) recording techniques, butthey can be studied with combined electrophysiological and me-chanical methods.17 An interesting observation is that the or-bicularis oris and masseter muscles, which are readily accessibleto EMG recordings, have an alternating pattern of activityduring chewing and become synchronous during swallowing.Such change in the functional correlation between trigeminaland facial muscles might be of interest for neurophysiologicalstudies in neurological syndromes involving chewing and swal-lowing disturbances.
Electromyographic Activity at Rest and DuringVoluntary Contraction
Facial and trigeminal muscles, like other muscles, have a certaindegree of background activity in awake human subjects at rest.Such activity can be quantified using reliable methods.38,82 Facialmuscle resting activity is increased in patients with headache39
and is increased on the affected side of patients with aberrant re-generation following recovery from a facial nerve injury.82
Abnormalities in the amount of EMG activity at rest and duringvoluntary contraction have been found in patients with olivo-pontocerebellar atrophy.83
Reflex Responses
The only monosynaptic reflex available to electrophysiologicaltesting in the cranial and facial muscles is the jaw jerk. It is me-diated by the trigeminal nerve. The stretch receptors can bestimulated by tapping on the mandible with a reflex tendonhammer. The afferent branch of the reflex carries the inputs upto the primary afferent terminals in the trigeminal motornucleus, where they synapse with alpha motoneurons to activatethe extrafusal muscle fibers.
Blinking can be reflexly generated by a number of stimuli fromdifferent sources. Elicitation of the blink reflex has been usedin neurophysiological studies for a long time. At present, elec-trical stimulation of the supraorbital nerve is the standardmethod for elicitation of the blink reflex.46,73,74 However, theblink reflex can also be generated by other stimuli such as loudacoustic tones, flashes of light, a tap to the glabella, a puff of airdirected to the cornea, and a small corneal electrical discharge.The study of the blink reflex is one of the most informative neu-rophysiological tests that can be performed in cranial nerves.The relative amounts of levator palpebrae inhibition and orbic-ularis oculi activation, which combine to give rise to the eyelidmovement in a blink, may be different for spontaneous andreflex blinking.
The perioral reflex is another trigemino-facial reflex. This canbe tested clinically by tapping the lips with the finger or with atendon hammer. EMG recordings are obtained with surfaceelectrodes placed on the orbicularis oris.54,78
Jaw opening is a reflex that can be elicited by many kinds ofstimuli; the salient feature being a transient inhibition of sus-tained EMG activity of the masseter muscles.34 Other inhibitoryand excitatory responses are also observed including a burst ofactivity in the digastric muscle53 and the suppression of tonic ac-tivity in the genioglossus muscle.57 The jaw opening reflex mayserve to protect the tongue from an unwanted biting. The mas-seteric inhibitory reflex (MIR) elicited by a mechanical stretchconsists of a single phase,58 whereas that elicited by exterocep-tive stimuli consists of two distinct phases, an MIR1 and anMIR2.23
PRINCIPAL ELECTRODIAGNOSTIC TECHNIQUES
Electrodiagnostic techniques used for the study of cranialnerves are not intrinsically different from those used for othernerves. Needle EMG can be performed in many muscles in-
AANEM Workshop Cranial Nerve Testing 3

nervated by cranial nerves. As in other muscles, the observationof fibrillation potentials and positive sharp waves at rest may in-dicate nerve damage. The electrodiagnostic medicine consul-tant should be aware, however, that the motor unit actionpotentials of the cranial nerve muscles are generally smaller inamplitude and shorter in duration than those of the limbmuscles and, therefore, it may be difficult to distinguish fibrilla-tion potentials from small motor units in muscles that are in-completely relaxed. Standard neurographic techniques for thestudy of motor fibers can be used only for testing the facial andthe spinal accessory nerves. No cranial nerve is available forsensory nerve conduction studies. The study of reflex re-sponses is a very useful tool for the examination of cranial nervefunction.31
Needle Electromyography
Needle EMG of the extraocular muscles is technically difficultand has the potential for ocular damage. It can be performedwith the help of an eyelid securing system. Needle EMG of thel-evator palpebrae is of interest because of the reciprocal behav-ior of this muscle and the orbicularis oculi during eye openingand closing. The normal EMG pattern of the levator palpebraein a subject with eyes open is tonic, interrupted by short silentperiods corresponding to spontaneous blinks. Simultaneousrecording of the levator palpebrae and the orbicularis oculi hasbeen shown to be useful in distinguishingbetween subgroups ofpatients with blepharospasm.2
Trigeminal, facial, spinal accessory, and hypoglossal nerve in-nervated muscles are all accessible to common needle EMGtesting. The vocal cords can also be examined using indirect-transoral or direct transcutaneous methods.10,20,85 For indirectEMG, careful anesthesia of the fauces is needed and precautionwith swallowing should be advised after the examination.Direct EMG is performed by inserting the needle electrodethrough the upper edge of the cricothyroid cartilage, to reachmmmmmmmmmmmmmmmmmmmmmmmmmmmmmthe thyroarytenoid muscle. This technique is also used forinjection of botulinum toxin in patients with spasmodic dys-phonia.85
Direct Stimulation of the Cranial Nerves
Cranial nerves available for direct stimulation and musclerecording are the facial and the spinal accessory nerves (Figures1 and 2).
Facial Nerve
Electrodiagnostic testing of the facial nerve is routinely per-formed in patients with facial palsy. Nerve excitability can betested by applying shocks of increasing intensity and determin-ing the intensity required to produce a muscle twitch. Althoughcomparison of nerve excitability between sides provides useful
clinical information in patients with facial palsy, surface EMGrecording of the compound muscle action potential (CMAP) inthe orbicularis oculi, orbicularis oris, quadratus labii, or nasalisallows more accurate and quantitative assessment. Stimuli areapplied just below the ear and anterior to the mastoid process,over the stylomastoid foramina. Normal latency values in adultsare 3.4 ± 0.8 ms.86 Selective stimulation of a given branch of thefacial nerve can also be performed.
The authors believe that for the assessment of a proximal lesionas in Bell’s palsy, the latency of the direct response is rarely
4 Cranial Nerve Testing AANEM Workshop
Figure 1—Location of the stimulating and recording electrodesfor electrophysiological examination of the facial nerve, modi-fied from DeLisa and MacKenzie16 with permission.
Figure 2—Location of the stimulating and recording electrodes forelectrophysiological examination of the spinal accessory nerve, modifiedfrom DeLisa and MacKenzie16 with permission.

useful. Even with substantial axonal degeneration, the onsetlatency determined by the remaining axons tend to be normal oronly slightly increased. In contrast, the amplitude of the directresponse provides useful information regarding prognosis byelucidating the degree of axonal loss. The amplitude of thedirect response varies substantially from one subject to the next.Thus, a comparison between sides in the same individual ismore meaningful than the absolute value. An amplitude reduc-tion to one half that of the response on the normal side suggestsdistal degeneration. Serial determinations during the first weekafter paralysis may reveal progressive amplitude decrease as anincreasing number of axons degenerate. Segmental evaluationof nerve conduction is possible in the facial nerve by using tran-scranial magnetic stimulation and recording from lower facialmuscles. In this procedure, the magnetic coil applied 8 to 10 cmlateral to the vertex elicits bilateral responses at a latency of 11to 12 ms, but if the coil is moved further laterally, a responseappears only on the ipsilateral side at a latency of about 5 ms.This response is believed to result from direct stimulation of thefacial nerve in the posterior fossa.3
With shocks of very high intensity, the stimulating current mayactivate the masseter muscle, and a volume-conducted potential
may be mistaken as the CMAP of the facial nerve. This woulderroneously suggest a favorable prognosis when in fact the facialnerve has already degenerated. In these instances, visual in-spection of the contracting muscle is essential to verify that therecorded potential originates from the intended muscle. Fromthe authors’ experience, the latency of the volume-conductedresponse from the masseter muscle is usually less than 2 ms, farshorter than that of the facial nerve. Figure 3 shows normalCMAPs of the orbicularis oculi, and a volume conducted re-sponse from the masseter muscle. Surface stimulation of thefacial nerve causes a late bilateral response in the orbicularisoculi, which can be due to activation of cutaneous fibers of thesecond or third branch of the trigeminal nerve or afferent activ-ity in the facial nerve, the so-called "facio-facial reflex."88 Figure4 shows a reflex response elicited by stimulation of the facialnerve, compatible with the "facio-facial reflex."
Spinal Accessory Nerve
In direct stimulation studies of the spinal accessory nerve, theactive electrode is placed over the upper trapezius muscle at theangle of neck and shoulder, and the reference electrode over thetendon near the acromion process. The nerve is stimulated asit descends along the posterior border of the sternocleidomas-toid muscle. In some subjects, the spinal accessory nerve iseasier to stimulate with the patient in the prone position.Normal latency was 1.8 to 3.0 ms in 25 normal subjects aged 10to 60 years.11 Changes in amplitude provide reliable informationwhen compared to the normal side, and an amplitude reductionof 50% or more suggests distal degeneration.
Repetitive stimulation of the spinal accessory nerve can be per-formed in cases with suspected disorders of the neuromusculartransmission. The recommended position for testing the neu-romuscular transmission in the spinal accessory nerve is with thepatient upright in a chair, the arms adducted and extended withthe hand holding onto the bottom of the chair. Exercise is ob-tained by having the patients shrug their shoulders against theirown restraint.
AANEM Workshop Cranial Nerve Testing 5
Figure 3—Responses recorded with surface electrodes placed overthe right (R) and left (L) orbicularis oculi muscles in a patient with leftfacial palsy 10 days from onset of the symptoms. In both parts A andB, the top trace is from stimulation at the right tragus and the bottomtrace is from stimulation at the left tragus. In A, the responses are fromthe orbicularis muscles reflecting activation of the facial nerve. In B, withhigher stimulus intensity, the responses reflect direct activation of themasseter muscles.
Figure 4—Two superimposed single trials recording from the or-bicularis oculi muscle with ipsilateral facial nerve stimulation at the tragus.The first response is the CMAP, and the second response is compati-ble with a "facio-facial reflex."

Hypoglossal Nerve
Hypoglossal nerve conduction can be accomplished by usingsurface electrodes on the top of the tongue and stimulating inthe submandibular region just medial to the angle of themandible.5,63,69 The mean latency was 2.2 ms in one study69 and2.4 ms in another.63
Reflex Responses
Reflex responses are specially useful in electrodiagnostic testingof the cranial nerves. This review will consider trigemino-trigeminal reflexes, trigemino-facial reflexes, and reflexes withafferents from the eighth nerve.
Trigemino-trigeminal Reflexes
Jaw Jerk
The jaw reflex, a myotatic reflex, is activated by sudden stretch-ing of the muscle spindles following a sharp tap to the mandible.This reflex, relayed via the mesencephalic nucleus of the trigem-inal nerve, reflects conduction through the midbrain. Themandibular division of the trigeminal nerve contains the musclespindle afferents as well as the motor axons to the extrafusalmuscle fibers, which constitute the afferent and efferent arcs ofthe masseter reflex, respectively. The cell bodies of the proprio-ceptive spindle afferents lie in the mesencephalic trigeminalnucleus. Branches from these cells make monosynaptic con-nection with the motor neurons of the trigeminal nerve locatedin the pons. The masseter reflex is ordinarily elicited by a me-chanical tap over the mandible. The closure of a microswitchattached to the percussion hammer triggers the oscilloscopesweep. The reflex responses are recorded simultaneously fromthe right and left masseter muscles using two pairs of surfaceelectrodes, the active placed over the muscle belly at the angleof the mandible, and the reference over the mastoid process orthe ear lobe. Since reflex latencies vary with successive trials,comparison of simultaneously recorded right-sided and left-sided responses is more meaningful than absolute values, whichare of the order of 6 to 8 ms in normal subjects. Reflex ampli-tude is also quite variable; the amplitude ratio between simulta-neously recorded right-sided and left-sided responses is,however, relatively constant. Ongerboer de Visser and Goor,66
who use a needle recording electrode, consider the test abnor-mal in patients with consistent unilateral absence of the reflex, aside-to-side latency difference of more than 0.5 ms, or bilateralabsence of the reflex up to the age of 70 years.
Masseteric Inhibitory Reflex
Sustained voluntary activity of the masseter muscles is tran-siently interrupted after stimuli of various kinds such as a tap tothe chin,58 an electrical stimulus to the mental nerve23 or to peri-
oral and intraoral receptors,9 tooth tapping,72 auditory stimula-tion,61 a jet of saline directed to the lower lip,12 or even high-in-tensity stimulation of the limb nerve afferents.27 Factorscontributing to the MIR induced by muscle stretch are the re-fractoriness of motoneurons and the transient arrest in the pro-prioceptive inputs after their synchronized activation in the jawjerk. The MIR is elicited bilaterally following unilateral electricalstimulation of the mental nerve. Electrical stimulation is usuallyapplied to the mental nerve at its exit from the mandibular boneat the chin while subjects are asked to clench their teeth, pro-ducing a sustained contraction of both masseter muscles. Somesubjects with bad dentures are not able to activate their massetermuscles and use the facial muscles instead. In these cases, nosuppression of the EMG activity can be elicited either with a tapto the chin or with an electrical stimulus. This failure of con-traction should not be mistaken for an abnormal MIR. If theMIR is to be used for electrodiagnostic purposes, the subjectshould be able to exert a stable background voluntary contrac-tion and the amount of suppression should be quantified. Thelatter is accomplished by averaging the rectified EMG activity ina sufficient number of trials (Figure 5).
The MIR elicited by a tap to the chin begins at about 10 to 12ms and lasts for 20 to 40 ms, although latency and duration areboth dependent partly on the strength of the tap and the levelof sustained activation of the masseter muscles.84 The MIR fol-lowing electrical stimulation of the mental nerve has two com-ponents, named MIR1 and MIR2. The latency of the MIR1 isof about 10 to 14 ms and the latency of the MIR2 is of about 40
6 Cranial Nerve Testing AANEM Workshop
Figure 5—Masseteric inhibitory reflex to electrical stimulation of themental nerve. Suppression of the ongoing EMG activity is divided intotwo components: MIR1 and MIR2 (A). The same response is recordedafter rectifying and averaging 10 consecutive traces (B). The area of thesuppression can be measured with respect to the background EMG ac-tivity.

to 50 ms. MIR1 is probably organized at the level of the pons,whereas the circuits responsible for MIR2 may extend down tothe pontomedullary junction.68
Trigemino-facial Reflexes
Blink Reflex
The blink reflex is an eyelid closure triggered by a stimulus. Itconsists of activation of the orbicularis oculi muscle and relax-ation of the levator palpebrae muscle. Simple observation ofthe corneal reflex can be done in clinical practice, but EMGrecordings furnish a more complete evaluation of the reflex re-sponses.46,52,73,74 The reflex response consists of two separatecomponents: an early R1 and a later R2 response. Whereas R1is evoked only on the side of stimulation, R2 is recorded bilat-erally with unilateral stimulation. R1 is a pontine reflex, whileR2 is presumably relayed through a more complex route in-cluding the pons and lateral medulla.48,67 R1 is relatively stablewith repeated trials and is therefore better suited for assessingnerve conduction through the trigeminal and facial nerves.Analysis of R2, however, is essential in determining whether alesion involves the afferent or efferent arc of the reflex. With alesion of the trigeminal nerve, R2 is slowed or diminished bilat-erally when the affected side of the face is stimulated (afferentdelay). With a lesion of the facial nerve, R2 is abnormal on theaffected side regardless of the side of stimulation (efferentdelay). Surface electrodes are used for stimulation of the nerveand recording of the evoked muscle action potentials.46 Theactive recording electrode is placed on the lower aspect of theorbicularis oculi muscle and the reference electrode on thelateral surface of the nose; the ground electrode is placed overthe forehead. Electrical stimulation of the supraorbital nerve ismost commonly used, but infraorbital or mental nerve stimula-tion also elicits an R2 in the orbicularis oculi muscle bilaterallyand, less consistently, an R1 on the side of stimulus. Shocks ofthe same intensity are delivered to each side to compare relativeexcitability of the reflexes. If R1 is unstable or not excitable,however, shocks of higher intensities or paired shocks with in-terstimulus intervals of 3 to 5 ms are used to facilitate the re-sponse so that the shortest latency can be determined.
The reflex latencies of R1 and R2 are measured from the stim-ulus artifact to the initial deflection of the evoked potential.If paired shocks are required to elicit R1, its latency is measuredfrom the stimulus artifact of the second shock provided, ofcourse, that the first stimulus elicits no response. With a sub-threshold conditioning shock and a maximal or supramaximaltest shock, the recorded response is elicited only by the secondstimulus so that the latency may be accurately determined fromthe second shock artifact. For each subject, at least eight re-sponses are measured, and the minimal latency is determined.Since R2 is inherently more variable than R1, its absolute latencyis less reliable than the latency difference between the simulta-neously elicited ipsilateral and contralateral responses.
The shock artifact is especially large when the supraorbital nerveis stimulated since the recording electrodes are located only 2 to3 cm away from the cathode of the stimulating electrode.Therefore, R1 tends to occur at the tail of the stimulus artifacteven though its latency is on the order of 10 ms. Careful repo-sitioning of the stimulating electrode may help reduce the stim-ulus artifact. A low-frequency cutoff of 300 Hz may also behelpful. Some investigators use a specially designed amplifierwith a short blocking time (1.0 ms) and low noise (0.5 µV RMSat bandwidth of 2,000 Hz). Stimulus artifact compensation canbe effectively performed by using two stimuli of the same in-tensity but of reversed polarity, separated by an interval equal totheir duration.64
A mechanical tap over the glabella also elicits a blink reflex. Itslatency and other characteristics can be recorded if the tap is de-livered with a sweep triggering hammer.21,52,74 Although the stim-ulus is a gentle tap, this is a cutaneous rather than a stretchreflex,74 probably relayed via the same polysynaptic reflex path-ways as the electrically elicited blink reflex.
Table 1 shows the mean and the standard deviation of the R1and R2 elicited by stimulation of the supraorbital nerve, in com-parison with the latency of the CMAP and the ratio betweenreflex and direct responses (R/D ratio), in 83 healthy subjects, 7to 86 years of age, and in 30 full-term neonates.44,50 An R1 wasrecorded in all but three infants. The R1 latency in neonates
AANEM Workshop Cranial Nerve Testing 7
Table 1 - The Blink Reflex in normal adults and neonates and in patients with various neurological diseases
Number R1 R2 R2c Direct R/D Response Ratio
Normal Adults 166 10.5 (0.8) 30.5 (3.4) 30.5 (4.4) 2.9 (0.4) 3.6 (0.5)
Normal Neonates 60 12.1 (1.0) 35.9 (2.5) often absent 3.3 (0.4) 3.7 (0.4)
Diabetic Polyneuropathy 172 11.4 (1.2) 33.7 (4.6) 34.8 (5.3) 3.4 (0.6) 3.4 (0.5)
Guillain-Barré Syndrome 108 15.1 (6.9) 38.2 (8.9) 38.3 (8.7) 4.5 (3.4) 3.8 (1.0)
Charcot-Marie-Tooth Disease 92 15.7 (4.1) 38.5 (6.7) 38.6 (6.5) 5.7 (3.1) 3.1 (1.0)
Multiple Sclerosis 126 12.3 (2.7) 35.8 (8.4) 35.7 (8.0) 2.9 (0.5) 4.3 (0.9)
was significantly greater than that in adults despite the con-siderably shorter length of the reflex arc in the former. Unlikethe consistent response in adults, an R2 was difficult to elicit ininfants; it was recorded in only 20 of 30 neonates, mostly on theside ipsilateral to the stimulus.50 There were no significant dif-ferences between sides.
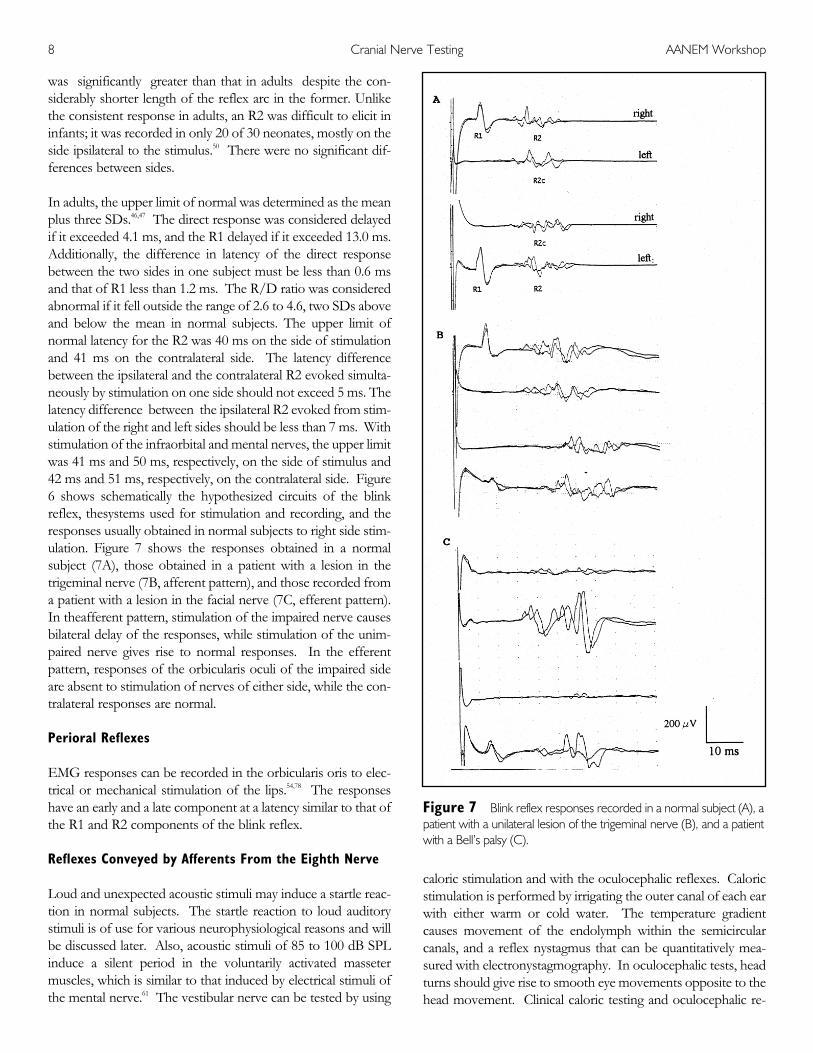
In adults, the upper limit of normal was determined as the meanplus three SDs.46,47 The direct response was considered delayedif it exceeded 4.1 ms, and the R1 delayed if it exceeded 13.0 ms.Additionally, the difference in latency of the direct responsebetween the two sides in one subject must be less than 0.6 msand that of R1 less than 1.2 ms. The R/D ratio was consideredabnormal if it fell outside the range of 2.6 to 4.6, two SDs aboveand below the mean in normal subjects. The upper limit ofnormal latency for the R2 was 40 ms on the side of stimulationand 41 ms on the contralateral side. The latency differencebetween the ipsilateral and the contralateral R2 evoked simulta-neously by stimulation on one side should not exceed 5 ms. Thelatency difference between the ipsilateral R2 evoked from stim-ulation of the right and left sides should be less than 7 ms. Withstimulation of the infraorbital and mental nerves, the upper limitwas 41 ms and 50 ms, respectively, on the side of stimulus and42 ms and 51 ms, respectively, on the contralateral side. Figure6 shows schematically the hypothesized circuits of the blinkreflex, thesystems used for stimulation and recording, and theresponses usually obtained in normal subjects to right side stim-ulation. Figure 7 shows the responses obtained in a normalsubject (7A), those obtained in a patient with a lesion in thetrigeminal nerve (7B, afferent pattern), and those recorded froma patient with a lesion in the facial nerve (7C, efferent pattern).In theafferent pattern, stimulation of the impaired nerve causesbilateral delay of the responses, while stimulation of the unim-paired nerve gives rise to normal responses. In the efferentpattern, responses of the orbicularis oculi of the impaired sideare absent to stimulation of nerves of either side, while the con-tralateral responses are normal.
Perioral Reflexes
EMG responses can be recorded in the orbicularis oris to elec-trical or mechanical stimulation of the lips.54,78 The responseshave an early and a late component at a latency similar to that ofthe R1 and R2 components of the blink reflex.
Reflexes Conveyed by Afferents From the Eighth Nerve
Loud and unexpected acoustic stimuli may induce a startle reac-tion in normal subjects. The startle reaction to loud auditorystimuli is of use for various neurophysiological reasons and willbe discussed later. Also, acoustic stimuli of 85 to 100 dB SPLinduce a silent period in the voluntarily activated massetermuscles, which is similar to that induced by electrical stimuli ofthe mental nerve.61 The vestibular nerve can be tested by using
caloric stimulation and with the oculocephalic reflexes. Caloricstimulation is performed by irrigating the outer canal of each earwith either warm or cold water. The temperature gradientcauses movement of the endolymph within the semicircularcanals, and a reflex nystagmus that can be quantitatively mea-sured with electronystagmography. In oculocephalic tests, headturns should give rise to smooth eye movements opposite to thehead movement. Clinical caloric testing and oculocephalic re-
8 Cranial Nerve Testing AANEM Workshop
Figure 7—Blink reflex responses recorded in a normal subject (A), apatient with a unilateral lesion of the trigeminal nerve (B), and a patientwith a Bell’s palsy (C).

flexes test the integrity of the labyrinth, vestibular nerves, mediallongitudinal fasciculus, and oculomotor system. They are ab-normal in posterior fossa lesions and cranial nerve injuries.
Excitability Recovery Curves
Paired stimulation is a technique commonly used to test the ex-citability of the structures integrated in a reflex system. In theblink reflex, excitability recovery curves are different for theearly and late responses.43 The differences can be attributed tothe larger number of synapses involved in the R2 response.Therefore, an abnormality in the blink reflex recovery curvelimited to the R2 component indicates a disorder of the ex-citability of brainstem interneurons. Using this technique, anexcitability enhancement of brainstem interneurons has beenfound in several neurological disorders.4,43,77,80
The excitability recovery curve has also been examined for themasseteric silent period following paired stimulation of themental nerve. In this case, there was a dissociation betweenMIR1 and MIR2, the latter being more affected than theformer.13 Patients with dystonia have a reduced inhibition of theMIR2.14
Other Techniques
Unexpected loud acoustic stimuli elicit a startle reaction.22,24,89
Usually, the acoustic stimulus is a tone of 90 to 105 dB SPL, de-livered through earphones, although higher intensities havebeen used.7 One problem with testing the startle reaction is thathabituation occurs quite fast with repeated stimuli. For thisreason, the rate of stimulation should be very low, with intervalsof up to 20 minutes.7 The brainstem circuits responsible for thestartle reaction have been studied in the rat. Davis and col-leagues15 showed that the structures implicated in startle arelocated in the medial bulbopontine reticular formation, and in-dicated the nucleus reticularis pontis caudalis as the likely gener-ator of the reaction. The impulses travel with the reticulospinaltract to the alpha motoneurons.33 The startle reaction of normalhuman subjects involves activity in many muscles innervated bycranial nerves as well as the muscles of the limbs and trunk.With acoustic stimuli, the response of the orbicularis oculi hasthe shortest latency and greatest persistence. However, Brownand colleagues7 have suggested that the early activity in the or-bicularis oculi may be separate from the startle reaction. Thiswould leave the sternomastoid muscle as the first muscle to beactivated, in keeping with their hypothesized generation of thestartle reaction in the lower brainstem, closer to the nucleus ofXI than to the nucleus of VII. The issue is not resolved at thepresent time because in fact, the startle reaction can be gener-ated in cats by electrical stimulation of the nucleus reticularispontis caudalis,15 which lies closer to VII than to XI. In anycase, persistence of activity in the sternomastoid muscle after re-
peated stimuli is certainly abnormal in patients with exaggeratedstartle reactions.8,60
If the acoustic stimulus causing a startle is preceded by a weaksignal, such as another weak auditory stimulus, a low-intensityelectrical stimulus or a flash of light, there is inhibition of thestartle response. This is the so-called prepulse inhibition.30,35
Prepulse inhibition has been found to be abnormal in patientswith schizophrenia6 and in patients with Huntington’s disease.75
Prepulse inhibition is probably mediated by brainstem struc-tures.70,71
The blink reflex can also be used to study classical Pavlovianconditioning in humans.25 In this test, repeated presentation ofa pair of time-locked stimuli leads to the generation of a condi-tioned response. Classical conditioning has been largely studiedin rabbits and in normal humans,90 and the role of the cerebel-lum in the appropriate timing of the response has been well es-tablished.87 Classical conditioning is abnormal in patients withcerebellar deficits.79
COMMON NEUROLOGICAL DISORDERS INVOLVINGCRANIAL NERVES
Peripheral Nerve Lesions
Polyneuropathies
Cranial nerves may be involved in toxic or metabolic polyneu-ropathies, but the neurophysiological findings are less promi-nent in cranial nerves than in the nerves of the limbs. Inpatients with Guillain-Barré syndrome, bilateral facial nerveparalysis occurs in about 50% of patients, and involvement ofbulbar cranial nerves in 10% to 30%. In these instances, thestudy of the blink reflex demonstrates a delay in the R1 and R2responses. The trigeminal nerve may be involved in diseases ofconnective tissue,55 in which a trigeminal neuropathy may be theonly neurological manifestation. Trigeminal involvement is rel-atively frequent in patients with Sjögren’s syndrome with a puresensory neuronopathy. In these patients, it is assumed that thedorsal root ganglia and the Gasserian ganglia are damaged bycirculating antibodies because of their lack of protection by theblood-brain barrier. Ganglionic neuronal damage may cause in-volvement of all sensory modalities in the limbs, with wide-spread loss of deep tendon reflexes. Also, there is loss ofcutaneous sensation in the face, with consistent abnormalities intrigemino-facial and trigemino-trigeminal reflexes, but the jawjerk, mediated by neurons of the mesencephalic nucleus,remains normal.81
Cranial nerves may also be involved in hereditary peripheralneuropathies. In the hereditary motor and sensory neuropathy
AANEM Workshop Cranial Nerve Testing 9

type I (HMSN-I), the blink reflex shows a marked delay of R1and R2.46 This is not the case with the HMSN-II, which con-stitutes a clear electrophysiological difference between the twomain forms of Charcot-Marie-Tooth disease.
Trigeminal Nerve Lesions
Despite the fact that the trigeminal nerve is the most importantcranial nerve for carrying sensory inputs from the face to thecentral nervous system, no known technique permits study ofsensory conduction in its peripheral segment. Therefore, elec-trodiagnostic studies rely on the assessment of reflex muscle re-sponses. In peripheral nerve lesions affecting the first branch,there should be an afferent delay of blink reflex responses fol-lowing stimulation of the affected nerve. In lesions affecting thethird branch of the trigeminal nerve, a similar pattern may beobserved with regard to the masseteric silent period followingstimulation of the mental nerve.
Isolated trigeminal nerve lesions are rare. Lesions may beinduced in the Gasserian ganglion following thermocoagulationor compression for treatment of trigeminal neuralgia. An un-common but remarkable clinical syndrome is the numb chinsyndrome, which consists of numbness restricted to the distrib-ution of the mental nerve. This syndrome is usually associatedwith neoplastic processes such as breast cancer or malignantlymphoma, and may result from a metastasis involving the infe-rior alveolar nerve at the mandible, or the mandibular branch ofthe Gasserian ganglion at the base of the skull.56
Facial Nerve Paralysis
The facial nerve is vulnerable to various disorders in several ofits segments. In the idiopathic form of facial paralysis (Bell’spalsy), the nerve injury probably occurs in the external third ofthe petrosal canal, just before the facial nerve exits the cranialbone. Electrodiagnostic abnormalities are observed with directstimulation of the facial nerve, which shows areduced CMAP,and with the blink reflex, which shows absence or delay of R1and R2 on the side of the lesion, with normal responses con-tralaterally, following stimulation to either side. When R1 ispresent, the ratio between the latency of R1 and that of theCMAP is increased, indicating that the site of the lesion is rostralto the point of direct facial nerve stimulation. After a completetransverse section of the facial nerve, excitability of the distalsegment remains normal up to 4 days, but complete loss of ex-citability occurs by the end of the first week, when the nerve un-dergoes wallerian degeneration. Hence, prognosis of a facialnerve paralysis cannot safely be established until the end of thefirst week. Afast and complete recovery can be assumed whenthe amplitude of the CMAP recorded on the affected side ismore than 50% of that of the unaffected side at 1 week afteronset of the paralysis. When the amplitude of the CMAP is lessthan 15% of that of the unaffected side, a permanent motordeficit and an abnormal recovery, with probable aberrant re-
generation, is the most likely. The same rule applies forpatientswith Bell’s palsy; while the commonest lesion is demyelination,there may be secondary axonal degeneration. Table 2 shows aprognostic classification of patients with Bell’s palsy, accordingto results of different tests.26
One of the most common sequelae of a facial palsy with axonaldegeneration is synkinesis due to aberrant regeneration. Duringaberrant axon regeneration, fibers that originally innervated theorbicularis oculi muscle are misdirected to other facialmuscles.49,82 Under such circumstances, blink reflex responsesare present in muscles other than the orbicularis oculi. In 26 of29 patients tested at least 4 months after total facial nerve de-generation, an aberrant blink reflex was recorded in the or-bicularis oris muscle on the affected side.49 For assessing facialsynkinesis after a facial nerve palsy, two pairs of recording elec-trodes are placed on the same side of the face, one pair over theorbicularis oculi and the other over the orbicularis oris orplatysma. When the subject is asked to perform isolated move-ments with the lower or upper facial muscles, simultaneous ac-tivity is observed in muscles not intended to be activated. Theblink reflex spreads to lower facial muscles. Other features ofthe postparalytic facial syndrome include an enhancement of thereflex responses, such as the orbicularis oculi response to supra-orbital nerve stimulation, and the observation of surface EMGmyokymic discharges in the orbicularis oris.82
POSTERIOR FOSSA LESIONS
Extra-axial Lesions
The Cranial Nerve Vascular Compression Syndrome
Hemifacial spasm consists of an involuntary twitching ofmuscles on one side of the face. EMG recordings, usually madefrom the orbicularis oris, orbicularis oculi, or frontalis muscles,show short lasting bursts of activity that may progressivelybuild up to constitute a sustained tonic discharge pattern lastingfor several hundreds of milliseconds. The abnormal burstsoccur simultaneously in many ipsilateral facial muscles, althoughsynkinesis is not always present during periods when there is noabnormal activity. This feature distinguishes hemifacial spasmfrom postparalytic synkinesis in which the aberrant response isalways present. Another anomaly that can be demonstratedneuro- physiologically in patients with hemifacial spasm isephaptic transmission.40,41 In this technique, simultaneousrecordings are made from the orbicularis oculi and the orbicu-laris oris while an electrical stimulus is applied to a branch of thefacial nerve supplying one of these muscles. In normal subjects,a CMAP is obtained only in that muscle, with no response in theother one. By contrast, patients with hemifacial spasm mayhave a response in the muscle whose nerve branch has not beenstimulated, at a latency too short for a transynaptic reflex re-sponse. Such a response is interpreted as originating by ephap-
10 Cranial Nerve Testing AANEM Workshop

tic transmission in a zone poorly myelinated or damaged by vas-cular compression.36 Similar lesions have been suggested tooccur in patients with trigeminal neuralgia. However, vascularcompression of nerves in the posterior fossa is a commonautopsy finding in asymptomatic individuals. It is thereforelikely that additional factors are needed to develop the clinicalsyndrome. One factor may be enhanced neuronal excitability ofthe neurons in the corresponding nuclei. Changes in excitabil-ity of the blink reflex pathway can occur at a premotoneuronallevel in some patients with hemifacial spasm.80 No neurophysi-ological abnormalities have been demonstrated yet in trigeminalneuralgia. The blink reflex is abnormal in paratrigeminal syn-dromes.46,65
Intra-axial Lesions
Facial Myokymia
Undulating, vermicular or wavelike movements of the muscleare known as myokymia. The clinical phenomenon is associ-ated with a characteristic EMG activity, referred to as myokymicdischarges. The phenomenon is due to an enhancement of theaxon membrane neuronal excitability, induced by demyelina-tion, edema, or ischemia.28 The myokymic discharges are seen asrecurrent firing potentials at a variable frequency between 5 and62 Hz. Facial myokymia can be seen in a variety of disorders,including Guillain-Barré syndrome, multiple sclerosis, brainstemtumors, rattlesnake envenomation, and the postparalytic facialsyndrome.
Vascular Lesions of the Brainstem
The blink reflex can be used to evaluate brainstem lesions whichmay or may not be clinically manifest. In patients with
Wallenberg’s syndrome, stimulation of the affected side of theface gives rise to a normal R1, but to a delayed, absent, ormarkedly diminished R2, bilaterally. With stimulation on thenormal side of the face, all responses may be normal, althoughan absent contralateral R2 occurs in a small percentage of pa-tients.48,67 Other trigemino-facial and trigemino-trigeminal re-flexes can also be involved in patients with Wallenberg’ssyndrome. In other brainstem syndromes, the use of variousbrainstem reflexes is recommended because the reflex circuitsare organized at different levels.31,66
Demyelinating Lesions
Conduction through a demyelinated segment in the centralreflex arc can also be measured objectively by the blink reflex.In multiple sclerosis, R1 was slowed on one or both sides in 49of 63 patients who had clinical pontine signs.44
Rostral Lesions
The blink reflex responses can be abnormal not only withlesions affecting the reflex pathways directly, but also withlesions indirectly influencing the excitability of the polysynapticconnections. Thus R2 is absent, or markedly diminished ordelayed, in comatose states regardless of the site of the respon-sible lesion.59 A significant change in the latency of R1 has alsobeen described in acute hemispheric stroke by Fisher and col-leagues21 who used mechanical rather than electrical stimulation.
The blink reflex excitability curve is abnormal in many disordersof the basal ganglia, such as parkinsonism,43 blepharospasm,4 orcervical dystonia.77 The abnormalities found in these patientsare limited to the excitability recovery of the R2, while the re-covery of R1 is normal. This is consistent with an enhanced ex-
AANEM Workshop Cranial Nerve Testing 11
Table 2 - Recovery from a Bell’s Palsy
Characteristics Class I Class II Class III
Nerve Lesion Conduction Block Partial Degeneration Complete DegenerationRecovery Early Delayed Defective
Complete Adequate InadequateClinical AssessmentOnset of Clinical Recovery 10 to 21 days 2 to 6 months More than 6 monthsResidual Asymmetry None Slight Mild to markedResidual Impairment None Minimal Variable
of Motor Function
Neurophysiological Assessment*EMG (denervation signs) Absent Sparse ProfuseEMG (motor units) Reduced Reduced AbsentExcitability of the Retained Reduced Lost
Facial Nerve* Performed at the 14th to 20th day from onset of the facial palsy.

citability of the interneurons in the blink reflex pathway, and notof the facial motoneurons.
REFERENCES
1. Alvarado-Mallart MR, Batini C, Buisseret-Delmas C, Corvisier J:Trigeminal representations of the masticatory and extraocular pro-prioceptors as revealed by horseradish peroxidase retrograde trans-port. Exp Brain Res 1975; 23:167-179.
2. Aramideh M, Ongerboer de Visser BW, Devriese PP, Bour LJ,Speelman JD: Electromyographic features of levator palpebrae su-perioris and orbicularis oculi muscles in blepharospasm. Brain 1994;117:27-38.
3. Benecke R, Meyer BU, Schonle P, Conrad B: Transcranial magneticstimulation of the human brain: Responses in muscles supplied bycranial nerves. Exp Brain Res 1988; 71:623-632.
4. Berardelli A, Rothwell JC, Day BL, Marsden CD: Pathophysiologyof blepharospasm and oromandibular dystonia. Brain 1985;108:593-608.
5. Blank A, Vatine JJ, Ferber I: Electrophysiological assessment of thetongue and the conduction time of the hypoglossal nerve.Electromyogr Clin Neurophysiol 1987; 27(3):131-133.
6. Braff D, Stone C, Callaway E, Geyer M, Bali L: Prestimulus effectson human startle reflex in normals and schizophrenics.Psychophysiology 1978; 15:339-343.
7. Brown P, Rothwell JC, Thompson PD, Britton TC, Day BL,Marsden CD: New observations on the normal auditory startlereflex in man. Brain 1991a; 114:1891-1902.
8. Brown P, Rothwell JC, Thompson PD, Britton TC, Day BL,Marsden CD: The hyperekplexias and their relationship to thenormal startle reflex. Brain 1991b; 114:1903-1928.
9. Cadden SW, Newton JP: A comparison of reflex depressions of ac-tivity in jaw-closing muscles evoked by intra- and peri-oral stimuli inman. Arch Oral Biol 1988; 33:863-869.
10. Cathala HP, Chouard C, Le Guerinel J, Verdy MF: Etude critiquedes différentes techniques d’électromyographie laringée. AnnOtolaryng (Paris) 1968; 85:283-292.
11. Cherington M: Accessory nerve conduction studies. Arch Neurol1968; 18:708-709.
12. Cruccu G, Agostino R, Inghilleri M, Manfredi M, Ongerboer deVisser BW: The masseter inhibitory reflex is evoked by innocuousstimuli and mediated by A beta afferent fibres. Exp Brain Res 1989;77:447-450.
13. Cruccu G, Agostino R, Fornarelli M, Inghilleri M, Manfredi M:Recovery cycle of the masseter inhibitory reflex in man. NeurosciLett 1984; 49:63-68.
14. Cruccu G, Pauletti G, Agostino R, Berardelli A, Manfredi M:Masseter inhibitory reflex in movement disorders, Huntington’schorea, Parkinson’s disease, dystonia, and unilateral masticatoryspasm. Electroencephalogr Clin Neurophysiol 1991; 81:24-30.
15. Davis M, Gendelman DS,Tischler MD, Gendelman PM: A primaryacoustic startle circuit: Lesion and stimulation studies. J Neurosci1982; 2:791-805.
16. DeLisa J, MacKenzie GK: Manual of Nerve Conduction VelocityTechniques. New York, Raven Press, 1982.
17. Ertekin C, Pehlivan M, Aydogdu I, Erta_ M, Uluda_ B, Çelebi G,Çolako_lu Z, Sa_duyu A, Y_ceyar N: An electrophysiological inves-tigation of deglutition in man. Muscle Nerve 1995; 18:1177-1186.
18. Evinger C, Shaw MD, Peck CK, Manning KA, Baker R: Blinkingand associated eye movements in humans, guinea pigs and rabbits. JNeurophysiol 1984; 52:323-339.
19. Evinger C, Manning KA, Sibony PA: Eyelid movements.Mechanisms and normal data. Invest Ophthalmol Vis Sci 1991;32:387-400.
20. Faaborg-Andersen K: Electromyographic investigations of intrinsiclaryngeal muscles in humans. Acta Physiol Scand 1957;41:(suppl):140.
21. Fisher MA, Shahani BT, Young RR: Assessing segmental excitabil-ity after acute rostral lesions: II. The blink reflex. Neurology 1979;29:45-50.
22. Fox JE: Excitatory and inhibitory components of the eyeblink re-sponses to startle evoking stimuli, studied in the human subject.Electroencephalogr Clin Neurophysiol 1978; 44:490-501.
23. Godaux E, Desmedt JE: Exteroceptive suppression and motorcontrol of the masseter and temporalis muscles in normal man.Brain Res 1975b; 85:447-458.
24. Gogan P: The startle and orienting reactions in man. A study oftheir characteristics and habituation. Brain Res 1970; 18:117-135.
25. Gormezano I: Classical conditioning, in Sidowski JB (ed):Experimental Methods and Instrumentation in Psychology. NewYork, McGraw-Hill, 1966, pp 385-420.
26. Granger CV: Prognosis in Bell’s palsy. Arch Phys Med Rehabil1976; 57:33-35.
27. Gurza S, Lowe AA, Sessle BJ: Influences on masseter activity ofstimuli applied to various sites in cats and macaque monkeys. ArchOral Biol 1976; 21:705-707.
28. Gutmann L: Facial and limb myokymia. Muscle Nerve 1991;14:1043-1049.
29. Hallett M, Berardelli A, Delwaide P, Freund HJ, Kimura J, LuckingC, Rothwell JC, Shahani BT, Yanagisawa N: Central EMG and testsof motor control. Report of an IFCN committee.Electroencephalogr Clin Neurophysiol 1994; 90:404-432.
30. Hoffman HS, Ison JR: Reflex modification in the domain of startle:I. Some empirical findings and their implications for how thenervous system processes sensory input. Psychorev 1980; 87:175-189.
31. Hopf HC: Topodiagnostic value of brain stem reflexes. MuscleNerve 1994; 17:475-484.
32. Hopf HC, Gutmann L: Diabetic 3rd nerve palsy: Evidence for a es-encephalic lesion. Neurology 1990; 40:1041-1045.
33. Hori A, Yasuhara A, Naito H, Yasuhara M: Blink reflex elicited byauditory stimulation in the rabbit. J Neurol Sci 1986; 76:49-59.
34. Hufschmidt HJ, Spuler H: Mono- and polysynaptic reflexes of thetrigeminal muscles in human beings. J Neurol Neurosurg Psychiatry1962; 25:332-335.
35. Ison JR, Sanes JN, Foss JA, Pinckney LA: Facilitation and inhibitionof the human startle blink reflexes by stimulus anticipation. BehavNeurosci 1990; 104:418-429.
36. Janetta PJ, Abassy M, Maroon JC, Ramos FM, Albir MS: Etiologyand definitive microsurgical treatment of hemifacial spasm. JNeurosurg 1977; 47:321-328.
37. Jenny AB, Saper CB: Organization of the facial nucleus and corti-cofacial projection in the monkey. A reconsideration of the uppermotor neuron facial palsy. Neurology 1987; 37:930-939.
38. Jensen R, Fuglsang-Frederiksen A, Olesen J: Quantitative surfaceEMG of pericranial muscles. Reproducibility and variability.Electroencephalogr Clin Neurophysiol 1993; 89:1-9.
39. Jensen R, Fuglsang-Frederiksen A, Olesen J: Quantitative surfaceEMG of pericranial muscles in headache. A population study.Electroencephalogr Clin Neurophysiol 1994; 93:335-344.
40. Kamp Nielsen V: Pathophysiology of hemifacial spasm. I. Ephaptictransmission and ectopic excitation. Neurology 1984; 34:418-426.
41. Kamp Nielsen V: Pathophysiology of hemifacial spasm. II. Lateralspread of the supraorbital nerve reflex. Neurology 1984; 34:427-431.
42. Karson CN, Burns RS, Lewitt PA, Foster NL, Newman RP: Blinkrates and disorders of movement. Neurology 1984; 34:677-678.
43. Kimura J: Disorder of interneurons in parkinsonism. The orbicu-laris oculi reflex to paired stimuli. Brain 1973; 96:87-96.
12 Cranial Nerve Testing AANEM Workshop

44. Kimura J: Electrically elicited blink reflex in diagnosis of multiplesclerosis: Review of 260 patients over a seven-year period. Brain1975; 98:413-426.
45. Kimura J: Electrodiagnosis in Diseases of Nerve and Muscle:Principles and Practice, ed 2. Philadelphia, Davis, 1989.
46. Kimura J, Powers JM, Van Allen MW: Reflex response of orbicu-laris oculi muscle to supraorbital nerve stimulation: Study in normalsubjects and in peripheral facial paresis. Arch Neurol 1969; 21:193-199.
47. Kimura J, Rodnitzky RL, Van Allen MW: Electrodiagnostic study oftrigeminal nerve: Orbicularis oculi reflex and masseter reflex intrigeminal neuralgia, paratrigeminal syndrome, and other lesions ofthe trigeminal nerve. Neurology 1970; 20:574-583.
48. Kimura J, Lyon LW: Orbicularis oculi reflex in the Wallenberg syn-drome: Alteration of the late reflex by lesions of the spinal tract andnucleus of the trigeminal nerve. J Neurol Neurosurg Psychiatry1972; 35:228-233.
49. Kimura J, Rodnitzky RL, Okawara S : Electrophysiologic analysis ofaberrant regeneration after facial nerve paralysis. Neurology 1975;25:989-993.
50. Kimura J, Bodensteiner J, Yamada T: Electrically elicited blink reflexin normal neonates. Arch Neurol 1977; 34:246-249.
51. Kimura J, Daube J, Burke D, Hallett M, Cruccu G, Ongerboer deVisser B, Yanagisawa N, Shimamura M, Rothwell JC: Human re-flexes and late responses. Report of an IFCN committee.Electroencephalogr Clin Neurophysiol 1994; 90:393-403.
52. Kugelberg E: Facial reflexes. Brain 1952; 75:385-396.53. Lamarre Y, Lund JP: Load compensation in human masseter
muscles. J Physiol 1975; 253:21-35.54. Larson CR, Folkins JW, McLean MD, Muller EM: Sensitivity of the
human perioral reflex to parameters of mechanical stretch. Brain Res1978; 146:159-164.
55. Lecky BRF, Hughes RAC, Murray NMF: Trigeminal sensory neu-ropathy. A study of 22 cases. Brain 1987; 110:1463-1485.
56. Lossos A, Siegal T: Numb chin syndrome in cancer patients:Etiology, response to treatment, and prognostic significance.Neurology 1992; 42:1181-1184.
57. Lowe AA, Gurza S, Sessle BJ: Excitatory and inhibitory influenceson tongue muscle activity in cat and monkey. Brain Res 1976;113:417-422.
58. Lund JP, Lamarre Y, Lavigne G, Duquet G: Human jaw reflexes.Adv Neurol 1983; 39:739-755.
59. Lyon LW, Kimura J, McCormick WF: Orbicularis oculi reflex incoma: Clinical, electrophysiological, and pathological correlations. JNeurol Neurosurg Psychiatry 1972; 35:582-598.
60. Matsumoto J, Fuhr P, Nigro M, Hallett M: Physiological abnormal-ities in hereditary hyperexplexia. Ann Neurol 1992; 32:41-50.
61. Meier-Ewert K, Gleitsmann K, Reiter F: Acoustic jaw reflex in man:Its relationship to other brainstem and microreflexes.Electroencephalogr Clin Neurophysiol 1974; 36:629-637.
62. Mogoseanu D, Smith AD, Bolam JP: Monosynaptic innervation offacial motoneurones by neurones of the parvicellular reticular for-mation. Exp Brain Res 1994; 101:427-438.
63. Muellbacher W, Mathis J, Hess CW: Electrophysiological assess-ment of central and peripheral motor routes to the lingual muscles.J Neurol Neurosurg Pyschiatry 1994; 57(3):309-315.
64. Nilsson J, Ravits J, Hallett M: Stimulus artifact compensation usingbiphasic stimulation. Muscle Nerve 1988; 11:597-602.
65. Ongerboer de Visser BW, Goor C: Electromyographic and reflexstudy in idiopathic and symptomatic trigeminal neuralgias: Latency ofthe jaw and blink reflexes. J Neurol Neurosurg Psychiatry 1974;37:1225-1230.
66. Ongerboer de Visser BW, Goor C: Jaw reflexes and masseter elec-tromyograms in mesencephalic and pontine lesions: An electrodiag-nostic study. J Neurol Neurosurg Psychiatry 1976; 39:90-92.
67. Ongerboer de Visser BW, Kuypers HGJM: Late blink reflexchanges in lateral meduallary lesions: An electrophysiological andneuroanatomical study of Wallenberg’s syndrome. Brain 1978;101:285-294.
68. Ongerboer de Visser BW, Cruccu G, Manfredi M, KoelmanJHThM: Effects of brainstem lesions on the masseter inhibitoryreflex. Functional mechanisms of reflex pathways. Brain 1989;113:781-792.
69. Redmond MD, Di Benedetto M: Hypoglossal nerve conduction innormal subjects. Muscle Nerve 1988; 11(5):447-452.
70. Rimpel J, Geyer D, Hopf HC: Changes in the blink responses tocombined trigeminal, acoustic and visual repetitive stimulation,studied in the human subjects. Electroencephalogr ClinNeurophysiol 1982; 54:552-560.
71. Saitoh K, Tilson HA, Shaw S, Dyer RS: Possible role of the brain-stem in the mediation of prepulse inhibition in the rat. Neurosci Lett1987; 75:216-222.
72. Sessle BJ, Schmitt A: Effects of controlled tooth stimulation on jawmuscle activity in man. Arch Oral Biol 1972; 17:1597-1607.
73. Shahani BT: The human blink reflex. J Neurol NeurosurgPsychiatry 1970; 33:792-800.
74. Shahani BT, Young RR: Human orbicularis oculi reflexes.Neurology 1972; 22:149-154.
75. Swerdlow NR, Paulsen J, Braff DL, Butters N, Geyer MA, SwensonMR: Impaired prepulse inhibition of acoustic and tactile startle re-sponse in patients with Huntington’s disease. J Neurol NeurosurgPsychiatry 1995; 58:192-200.
76. Tackmann W, Ettlin T, Barth R: Blink reflexes elicited by electrical,acoustic and visual stimuli. I. Normal values and possible anatomi-cal pathways. Eur Neurol 1982; 21:210-216.
77. Tolosa ES, Montserrat L, Bayes A: Blink reflex studies in focal dys-tonias: Enhanced excitability of brainstem interneurons in cranialdystonia and spasmodic torticollis. Mov Dis 1988; 3:61-69.
78. Topka H, Hallett M: Perioral reflexes in orofacial dyskinesia andspasmodic dysphonia. Muscle Nerve 1992; 15:1016-1022.
79. Topka H, Valls-Solé J, Massaquoi SG, Hallett M: Deficit in classicalconditioning in patients with cerebellar degeneration. Brain 1993;116:961-969.
80. Valls-Solé J, Tolosa ES: Blink reflex excitability cycle in hemifacialspasm. Neurology 1989; 39:1061-1066.
81. Valls-Solé J, Graus F, Font J, Pou A, Tolosa ES: Normal proprio-ceptive trigeminal afferents in patients with Sjögren’s syndrome andsensory neuronopathy. Ann Neurol 1990; 28:786-790.
82. Valls-Solé J, Tolosa ES, Pujol M: Myokymic discharges and en-hanced facial nerve reflex responses after recovery from idiopathicfacial palsy. Muscle Nerve 1992; 15:37-42.
83. Valls-Solé J, Lou JS, Hallett M: Brainstem reflexes in patients witholivopontocerebellar atrophy. Muscle Nerve 1994; 17:1439-1448.
84. van Steenberghe D, de Laat A: EMG of Jaw Reflexes in Man.Leuven, Belgium, Leuven University Press, 1989.
85. Watson BC, Schaefer SD, Freeman FJ, Dembowski J, Kondraske G,Roarkk R: Laryngeal electromyographic activity in adductor and ab-ductor spasmodic dysphonia. J Speech Hear Res 1991; 34:473-482.
86. Waylonis GW, Johnson EW: Facial nerve conduction delay. ArchPhys Med Rehabil 1964; 45:539-541.
87. Welsh JP, Harvey JA: Pavlovian conditioning in the rabbit during in-activation of the interpositus nucleus. J Physiol 1991; 444:459-480.
88. Willer JC, Lamour Y: Electrophysiological evidence for a facio-facialreflex in the facial muscles in man. Brain Res 1977; 119:459-464.
89. Wilkins DE, Hallett M, Wess MM: Audiogenic startle reflex of manand its relationship to startle syndrome: A review. Brain 1986;109:561-573.
90. Woodruff-Pak DS: Aging and classical conditioning: Parallel studiesin rabbits and humans. Neurobiol Aging 1988; 9:511-522.
AANEM Workshop Cranial Nerve Testing 13

AMERICAN ASSOCIATION OF NEUROMUSCULAR & ELECTRODIAGNOSTIC MEDICINE
AMERICAN ASSOCIATION OF NEUROMUSCULAR & ELECTRODIAGNOSTIC MEDICINE
2621 Superior Drive NWRochester, MN 55901
(507) 288-0100
CRANIAL NE
RVE TESTIN
G
AA
NEM
WO
RK
SHO
P
Josep Valls
-Solé, MD, PhD
Mark Hallett,
MD





























