Embed Size (px)
Citation preview

of June 29, 2018.This information is current as
CellsRole during Rescue of Exhausted CD8 TPlays a Critical CD8-Intrinsic and -Extrinsic
CD40 Ligand Pathway−Cutting Edge: CD40
Rajarshi Bhadra, Jason P. Gigley and Imtiaz A. Khan
ol.1102319http://www.jimmunol.org/content/early/2011/09/25/jimmun
published online 26 September 2011J Immunol
MaterialSupplementary
9.DC1http://www.jimmunol.org/content/suppl/2011/09/27/jimmunol.110231
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2011 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on June 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Cutting Edge: CD40–CD40 Ligand Pathway Plays a CriticalCD8-Intrinsic and -Extrinsic Role during Rescue ofExhausted CD8 T CellsRajarshi Bhadra, Jason P. Gigley, and Imtiaz A. Khan
CD8 exhaustion mediated by an inhibitory pro-grammed death-1–programmed death ligand-1 (PD-L1)pathway occurs in several chronic infections, includingtoxoplasmosis. Although blockade of the programmeddeath-1–PD-L1 pathway revives this response, the roleof costimulatory receptors involved in this rescue hasnot been ascertained in any model of CD8 exhaustion.This report demonstrates that one such costimulatorypathway, CD40–CD40L, plays a critical role duringrescue of exhausted CD8 T cells. Blockade of this path-way abrogates the ameliorative effects of anti–PD-L1treatment on CD8 T cells. Additionally, we demon-strate in an infectious disease model that CD8-intrinsicCD40 signaling is important for optimal CD8 poly-functionality, proliferation, T-bet upregulation, andIL-21 signaling, albeit in the context of CD8 rescue.The critical role of CD40 during the rescue of exhaustedCD8 T cells may provide a rational basis for designingnovel therapeutic vaccination approaches. The Journalof Immunology, 2011, 187: 000–000.
CD8 exhaustion, as reported in chronic viral and tumormodels, involves a hierarchical loss of functions (cy-tokine, proliferation, and cytotoxicity), and, in ex-
treme cases, CD8 T cells can be physically deleted in aprogrammed death-1 (PD-1)–dependent manner (1). A re-cently published study from our laboratory demonstrates thatCD8 T cells from chronically infected mice exhibit pro-gressive functional exhaustion, concomitant with increasedPD-1 expression, in the Toxoplasma model (2). This dysfunc-tion results in reactivation of latent toxoplasmosis leadingto host mortality. In vivo blockade of PD-1 interaction withits receptor PD ligand-1 (PD-L1) preferentially reinvigoratespre-existing PD-1–expressing CD8 T cells, thereby prevent-ing disease recrudescence and host mortality (2). Apart fromTCR signal and cytokine milieu, one of the most importantfactors governing CD8 response is the balance between pos-itive signals from costimulatory receptors and the negative
signals from inhibitory receptors (3). It has been postulatedthat interplay between signals from costimulatory and in-hibitory receptors may play an important role in T cell acti-vation, differentiation, effector function, and exhaustion (4).However, the role of costimulatory receptors during the res-cue of this response has not been explored in any model ofCD8 exhaustion. Therefore, we hypothesized that blockade ofinhibitory PD-1–PD-L1 pathway in chronically infected micewould lead to upregulation of costimulatory receptors, andsuch positive signals would be critical for the rescue of dys-functional CD8 T cells. We identified CD40 as one of themolecules highly upregulated on CD8 T cells in anti–PD-L1–treated, chronically infected mice. A previous report using anHY-TCR transgenic model demonstrated that CD8-intrinsicCD40 signaling is critical for memory generation (5). How-ever, this notion has been challenged by multiple studies ininfectious disease models in which CD40-deficient memoryCD8 T cells has been shown to elicit a normal immune re-sponse in a CD40-sufficient environment (6). Similarly, inthe Toxoplasma model, the CD40–CD40L pathway has beenshown to play a minimal role in T cell response development(7). This discrepancy in CD40 dependence has been attrib-uted to the nature of Ag and the highly inflammatory con-ditions associated with infectious disease (6). However, therole of the CD40–CD40L pathway during rescue of ex-hausted CD8 T cells has not been addressed in any model.The current report demonstrates that the CD40–CD40L
pathway has a minimal effect on CD8 T cell response duringchronic toxoplasmosis. Surprisingly, coadministration of bothanti–PD-L1 and anti-CD40L abrogates anti–PD-L1–medi-ated rescue, suggesting that positive costimulatory signals playa pivotal role during the rescue process. Additionally, wedemonstrate for the first time, to our knowledge, in an in-fectious disease model that CD40 plays a CD8-intrinsic rolenot only in terms of quantum but also quality of the immuneresponse, albeit in the context of reinvigoration of CD8 re-sponse. Moreover, this report demonstrates that CD40 playsa strictly T cell-intrinsic role in modulating the IL-21–IL-21Rpathway, which has been shown to be important for ame-
Department of Microbiology, Immunology and Tropical Medicine, George WashingtonUniversity, Washington, DC 20037
Received for publication August 9, 2011. Accepted for publication September 6, 2011.
This work was supported by National Institutes of Health Grant AI-33325 (to I.A.K.).
Address correspondence and reprint requests to Dr. Imtiaz A. Khan, Department ofMicrobiology, Immunology and Tropical Medicine, Ross Hall, Room 745, GeorgeWashington University, 2300 I Street N.W., Washington, DC 20037. E-mail address:[email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: BM, bone marrow; Gzb, granzyme B; KO, knockout;PD-1, programmed death-1; PD-L1, programmed death ligand-1; p.i., postinfection;TLA, toxoplasma lysate Ag; WT, wild-type.
Copyright� 2011 by TheAmerican Association of Immunologists, Inc. 0022-1767/11/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1102319
Published September 26, 2011, doi:10.4049/jimmunol.1102319 by guest on June 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

liorating CD8 exhaustion in viral models of chronic infection(8).
Materials and MethodsMice, infection, and chimera generation
Age-matched female C57BL/6, CD90.1, CD45.1, CD82/2, and CD402/2
mice were purchased from the National Cancer Institute or The JacksonLaboratory. Mixed bone marrow (BM) chimeras were generated by injectingan qual number of BM cells from CD90.1 and CD402/2 mice into lethallyirradiated CD45.1 recipients. Chimeras were infected at 8 wk postrecon-stitution. Mice were infected with 10 cysts Toxoplasma gondii (ME49) via theoral route and sacrificed at week 7 postinfection (p.i.).
In vivo and in vitro Ab treatment
Mice were injected with anti–PD-L1 (0.2 mg/mouse, MIH5; in-house) and/oranti-CD40L (0.5 mg/mouse, MR1; BioXCell) every third day i.p. starting atweek 5 p.i. for 2 wk. Relevant isotype controls were used throughout. Ago-nistic CD40 (FGK4.5; BioXCell) was used in vitro at a final concentration of10 mg/ml for 6 h at 37˚C.
Tissue preparation and flow cytometry
Single-cell suspension preparation, restimulation, and flow cytometry wereperformed as previously described (2). Briefly, surface staining for 30 min at4˚C was followed by intracellular staining where needed using BD Fix/PermKit (BD Biosciences) as per the manufacturer’s protocol. Live/Dead Aqua(Invitrogen) staining was performed prior to surface staining for Figs. 3 and 4.For cytokine detection in CD8 T cells, 5 3 105 naive splenocytes fromCD82/2 mice was mixed with an equal number of brain or splenic cells frominfected mice, and restimulation was similarly carried out for 16 h in thepresence of toxoplasma lysate Ag (TLA; 30 mg/ml). A total of 0.65 ml/mlmonensin (BD Biosciences) and 0.65 ml/ml brefeldin A (BD Biosciences) wasadded during the final 9 h of stimulation. Cytokine detection in splenic CD4T cells was similarly carried out without CD82/2 splenocyte addition. Afteracquisition using BD FACSAria or BD FACSCalibur or Cytek upgraded BDFACSCalibur (BD Biosciences), data were analyzed using FlowJo (Tree Star).Computation for enumerating polyfunctional cells was performed using thePESTLE and SPICE programs, generously provided to us by Dr. MarioRoederer (National Institutes of Health).
Statistical analysis
Unless otherwise mentioned, statistical analysis was performed using unpairedStudent t tests with p , 0.05 taken as statistically significant. For comparingwild-type (WT) and knockout (KO) T cell data from chimeras within thesame treatment group, the paired t test was used, whereas for comparing WTor KO data between different treatment groups (i.e., isotype versus anti–PD-L1), the unpaired t test was used.
Results and DiscussionAnti–PD-L1 treatment upregulates CD40 but not CD40L on CD8T cells
To identify costimulatory molecules involved in the rescue ofexhausted CD8 T cells, we examined a panel of these receptorson CD8 T cells in chronically infected mice treated with anti–PD-L1 or control Ab (data not shown). CD40 was identifiedas one of the costimulatory molecules showing high levels ofdifferential expression. Minimal CD40 levels were noted onsplenic CD8 T cells from control mice, whereas high ex-pression of this molecule occurred on CD8 T cells from anti–PD-L1–treated animals (Fig. 1A). Furthermore, CD40 waspreferentially expressed on PD-1+ subset (Fig. 1B) in anti–PD-L1 treated mice. Despite high CD40 expression on CD8T cells from anti–PD-L1–treated mice, no differential ex-pression of its receptor, CD40L, was noted between CD8T cells from control and Ab-treated animals (Fig. 1C).Combined, the above data point toward potential involvementof the CD40–CD40L pathway during rescue of exhaustedCD8 T cells.
CD40–CD40L signaling plays a critical role during rescue ofexhausted CD8 T cells
Because CD40 is differentially expressed on reinvigoratedCD8 T cells, we next investigated if CD40–CD40L signalingplayed any role during the rescue process. To address this,chronically infected mice were administered control Ab, anti–PD-L1, anti-CD40L, or both (anti–PD-L1 and anti-CD40L)for 2 wk (weeks 5–7) and then analyzed at week 7 p.i. Inagreement with observations by Reichmann et al. (7), block-ade of the CD40–CD40L pathway alone had no profoundeffect on T cells’ absolute number, and CD40 expression,proliferation, and functionality of CD8 T cells remainedunaltered in both spleen and brain (Fig. 2A–D). Althoughanti–PD-L1 treatment by itself was able to augment theabove parameters (absolute number, CD40 expression, pro-liferation, and effector functions), surprisingly, coadminis-tration of anti-CD40L and anti–PD-L1 abrogated the rescueeffect (Fig. 2A–D) (2). This suggests that the CD40–CD40Lpathway plays a critical role during rescue of dysfunctionalCD8 T cells. T-box family transcription factors T-bet andEomes have been to shown to be important for proliferation,effector functions, and survival of CD8 T cells (9). To furtherinvestigate the role of CD40 signaling on CD8 transcriptionalcontrol, purified splenic CD8 T cells from the above micewere treated in vitro with agonistic CD40 or control Ab. Theexpression of the above transcription factors was then evalu-ated by intracellular staining. In vivo coadministration ofanti–PD-L1 and anti-CD40L abrogated increase in T-betand Eomes expression, and in vitro treatment with agonisticCD40 failed to augment these molecules (Fig. 2E). Surpris-ingly, agonistic CD40 treatment of CD8 T cells from anti–PD-L1–treated mice resulted in further increase in T-bet butnot in Eomes (Fig. 2E). This suggests that although Eomesexpression in CD8 T cells is independent of CD8-intrinsicCD40 signaling, T-bet expression is at least partially de-pendent on CD40 signaling directly on CD8 T cells.
CD8-intrinsic CD40 signaling is important for optimal rescue ofexhausted CD8 T cells
The above data bear the implication that CD40 signalingdirectly on CD8 T cells is capable of modulating T-bet ex-pression. However, it does not address if T cell-intrinsicCD40–CD40L interaction in vivo can elicit a similar effect.Moreover, it does not rule out the possibility of CD40 ex-pression on other cell types via changes in microenvironment
FIGURE 1. High expression of CD40 on CD8 T cells in anti–PD-L1–
treated mice. A, Chronically infected mice were treated with anti–PD-L1 for
2 wk and then sacrificed at week 7 p.i. CD40 expression was analyzed on
splenic CD8 T cells. Dotted line represents background fluorescence
throughout. B represents PD-1 expression on CD40+ and CD402 splenic
CD8 T cells in anti–PD-L1–treated mice. C, Splenic CD8 T cells from
control and anti–PD-L1–treated mice were evaluated for CD40L expression.
The data represent three experiments with at least four mice per group.
2 CUTTING EDGE: ROLE OF CD40 SIGNALING IN EXHAUSTED CD8 T CELL RESCUE
by guest on June 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

such as cytokines or Ag burden, affecting CD8 T cells. Toaddress the CD8-intrinsic role of CD40 during rescue ofexhausted CD8 T cells in vivo, we generated BM chimeras byinjecting similar number of BM cells from CD90.1 (WT)and CD90.2+/+CD402/2 (KO) into lethally irradiated CD45.1WT recipients. After reconstitution, the mixed bone BMchimeras were infected with T. gondii, treated with anti–PD-L1 for 2 wk (weeks 5–7), and then analyzed at week 7 p.i. Incontrol Ab-treated animals, WT and KO CD8 T cells werepresent in ∼ 1:1 ratio, and no difference in PD-1 expressionwas noted (Fig. 3A–C). However, after anti PD-L1 treatment,this ratio was altered to 2:1 in both spleen and brain, sug-gesting that CD40 plays a CD8-intrinsic role in mediatingthe expansion during the rescue process (Fig. 3A, 3B). To fur-ther investigate if differential expansion of WT CD8 T cellsin anti–PD-L1–treated chimeras was due to increased pro-liferation of these cells, we performed intracellular stainingfor Ki-67. As shown in Fig. 3D and 3E, irrespective of CD40expression, anti–PD-L1–treated chimeras showed greater CD8T cell proliferation than controls. However, although CD40deficiency minimally affected proliferation of CD8 T cellsin control chimeras, it had a more pronounced effect in anti–PD-L1–treated chimeras. Next, we measured apoptosis byexamining active caspase-3 expression. Regardless of CD40deficiency, anti–PD-L1 treatment reduced CD8 apoptosis(Fig. 3F, 3G). Nevertheless, WT CD8 T cells were moder-ately more apoptotic than KO cells in both control and
treated chimeras (Fig. 3F, 3G). Combined, this suggests thatproliferation is the major mechanism responsible for pre-ponderance of WT CD8 T cells in anti–PD-L1–treated chi-meras. Next, we examined if CD40 played a CD8-intrinsicrole on effector functions of reinvigorated CD8 T cells. Irre-spective of anti–PD-L1 treatment or CD40 deficiency, min-imal IL-2 production was noted in CD8 T cells (data notshown). WT or KO CD8 T cells produced similar levels ofIFN-g, granzyme B (Gzb), and TNF-a in control chimerasin both spleen and brain (Fig. 3H). Although anti–PD-L1treatment augmented these molecules in both WT and KOCD8 T cells, the latter exhibited far lower levels of Gzb andespecially IFN-g and TNF-a (Fig. 3H). Moreover, the per-centage of both bifunctional and trifunctional CD8 T cellswas highly depressed in KO CD8 T cells in treated animals(Fig. 3I ). Finally we examined the transcriptional profile ofCD8 T cells. Irrespective of CD40 deficiency similar levels of
FIGURE 2. Blockade of CD40–CD40L pathway abrogates anti–PD-L1–
mediated rescue of CD8 T cells. A, Anti–PD-L1, anti-CD40L, or both were
administered to chronically infected mice for 2 wk and then sacrificed at week
7 p.i. Absolute number of CD8 T cells was evaluated in spleen and brain.
CD40 (B) and Ki-67 (C) expression was assessed in these mice by flow
cytometry. D, IFN-g and Gzb production by CD8 T cells was evaluated in
splenocytes or brain cells from infected mice in presence of TLA. E, SplenicCD8 T cells purified from in vivo Ab-treated animals were evaluated for
T-bet (top panel) and Eomes (bottom panel) after 6 h in vitro stimulation with
agonistic CD40 or control Ab. The data represent two experiments with at
least four mice per group. Error bars represent SD throughout.
FIGURE 3. CD40 plays a CD8-intrinsic role during rescue of exhausted
CD8 T cells. Chronically infected mixed BM chimeras were treated with anti–
PD-L1 for 2 wk and analyzed at week 7 p.i. Relative abundance of WT
(CD90.1) and KO (CD90.2) CD8 T cells evaluated in spleen and brain by
flow cytometry is presented as representative flow plots (A) or bar graphs (B).C represents frequency of WT and KO splenic CD8 T cells in control chi-
meras expressing PD-1. D and E, The percentage of proliferating congenic
splenic CD8 T cells was assessed by intracellular staining for Ki-67. Flow
plots in red denote KO CD8 T cells throughout. Data are presented as
histograms (D) or bar graphs (E). Histograms (F) and bar graphs (G) depictactive caspase-expressing CD8 T cells evaluated after incubating splenocytes
from chimeras at 37˚C for 5 h. H, IFN-g, Gzb, and TNF-a production by
congenic CD8 T cells were evaluated in splenocytes or brain cells from
infected mice in presence of TLA. I represents the percentage of bifunctional(top panels) and trifunctional (bottom panels) CD8 T cells in spleen and brain.
Results are representative of two experiments with at least three mice per
group.
The Journal of Immunology 3
by guest on June 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
Eomes were noted in CD8 T cells in anti–PD-L1–treatedchimeras (Supplemental Fig. 1). In contrast, anti–PD-L1 treat-ment augmented T-bet preferentially in WT CD8 T cells(Supplemental Fig. 1). Considering that Eomes has a criticalrole in mediating cytotoxicity, this may explain why KO CD8T cells in anti–PD-L1–treated chimeras have unaffected singleGzb+ CD8 T cells but yet have attenuated IFN-g and TNF-aproduction (9). Together, the data suggest that during rescueof exhausted CD8 T cells, CD40 plays a CD8-intrinsic rolein mediating proliferation and effector functions of CD8T cells. However, unlike anti-CD40L treatment, CD40 de-ficiency on CD8 T cells in a CD40-sufficient environmentdoes not entirely abrogate the ameliorative effects of anti–PD-L1, suggesting that CD40 has an important CD8-extrinsicrole as well.
CD40 plays an important role in mediating IL-21 and IL-21Rexpression
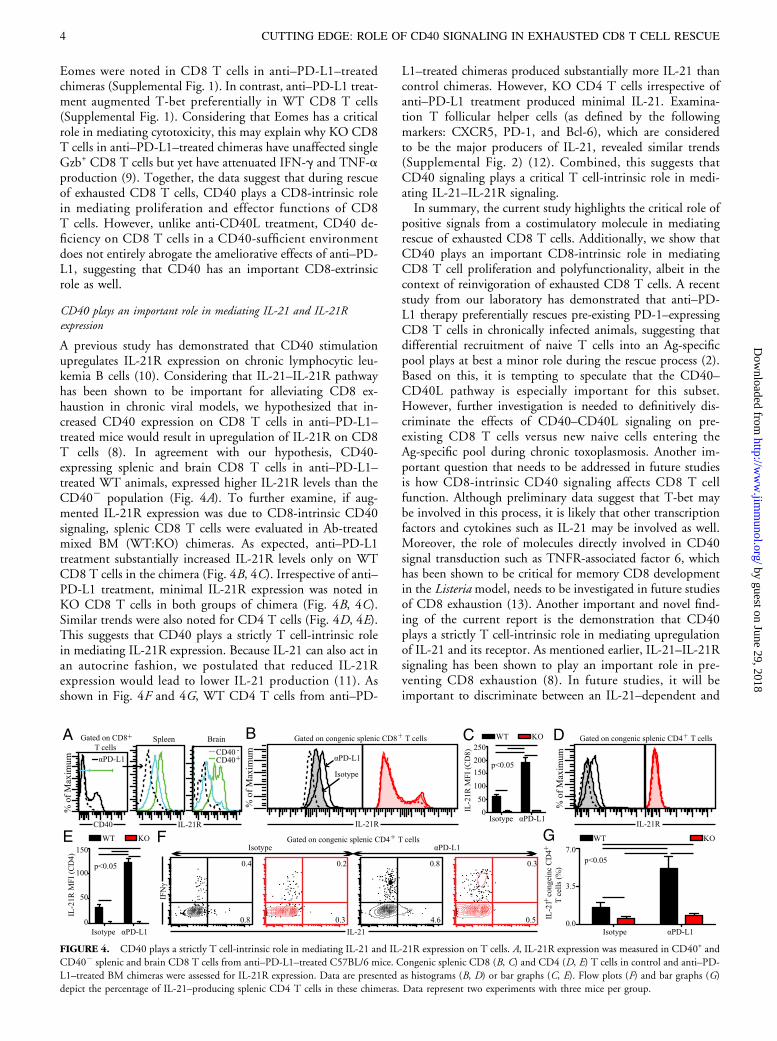
A previous study has demonstrated that CD40 stimulationupregulates IL-21R expression on chronic lymphocytic leu-kemia B cells (10). Considering that IL-21–IL-21R pathwayhas been shown to be important for alleviating CD8 ex-haustion in chronic viral models, we hypothesized that in-creased CD40 expression on CD8 T cells in anti–PD-L1–treated mice would result in upregulation of IL-21R on CD8T cells (8). In agreement with our hypothesis, CD40-expressing splenic and brain CD8 T cells in anti–PD-L1–treated WT animals, expressed higher IL-21R levels than theCD402 population (Fig. 4A). To further examine, if aug-mented IL-21R expression was due to CD8-intrinsic CD40signaling, splenic CD8 T cells were evaluated in Ab-treatedmixed BM (WT:KO) chimeras. As expected, anti–PD-L1treatment substantially increased IL-21R levels only on WTCD8 T cells in the chimera (Fig. 4B, 4C). Irrespective of anti–PD-L1 treatment, minimal IL-21R expression was noted inKO CD8 T cells in both groups of chimera (Fig. 4B, 4C).Similar trends were also noted for CD4 T cells (Fig. 4D, 4E).This suggests that CD40 plays a strictly T cell-intrinsic rolein mediating IL-21R expression. Because IL-21 can also act inan autocrine fashion, we postulated that reduced IL-21Rexpression would lead to lower IL-21 production (11). Asshown in Fig. 4F and 4G, WT CD4 T cells from anti–PD-
L1–treated chimeras produced substantially more IL-21 thancontrol chimeras. However, KO CD4 T cells irrespective ofanti–PD-L1 treatment produced minimal IL-21. Examina-tion T follicular helper cells (as defined by the followingmarkers: CXCR5, PD-1, and Bcl-6), which are consideredto be the major producers of IL-21, revealed similar trends(Supplemental Fig. 2) (12). Combined, this suggests thatCD40 signaling plays a critical T cell-intrinsic role in medi-ating IL-21–IL-21R signaling.In summary, the current study highlights the critical role of
positive signals from a costimulatory molecule in mediatingrescue of exhausted CD8 T cells. Additionally, we show thatCD40 plays an important CD8-intrinsic role in mediatingCD8 T cell proliferation and polyfunctionality, albeit in thecontext of reinvigoration of exhausted CD8 T cells. A recentstudy from our laboratory has demonstrated that anti–PD-L1 therapy preferentially rescues pre-existing PD-1–expressingCD8 T cells in chronically infected animals, suggesting thatdifferential recruitment of naive T cells into an Ag-specificpool plays at best a minor role during the rescue process (2).Based on this, it is tempting to speculate that the CD40–CD40L pathway is especially important for this subset.However, further investigation is needed to definitively dis-criminate the effects of CD40–CD40L signaling on pre-existing CD8 T cells versus new naive cells entering theAg-specific pool during chronic toxoplasmosis. Another im-portant question that needs to be addressed in future studiesis how CD8-intrinsic CD40 signaling affects CD8 T cellfunction. Although preliminary data suggest that T-bet maybe involved in this process, it is likely that other transcriptionfactors and cytokines such as IL-21 may be involved as well.Moreover, the role of molecules directly involved in CD40signal transduction such as TNFR-associated factor 6, whichhas been shown to be critical for memory CD8 developmentin the Listeria model, needs to be investigated in future studiesof CD8 exhaustion (13). Another important and novel find-ing of the current report is the demonstration that CD40plays a strictly T cell-intrinsic role in mediating upregulationof IL-21 and its receptor. As mentioned earlier, IL-21–IL-21Rsignaling has been shown to play an important role in pre-venting CD8 exhaustion (8). In future studies, it will beimportant to discriminate between an IL-21–dependent and
FIGURE 4. CD40 plays a strictly T cell-intrinsic role in mediating IL-21 and IL-21R expression on T cells. A, IL-21R expression was measured in CD40+ and
CD402 splenic and brain CD8 T cells from anti–PD-L1–treated C57BL/6 mice. Congenic splenic CD8 (B, C) and CD4 (D, E) T cells in control and anti–PD-
L1–treated BM chimeras were assessed for IL-21R expression. Data are presented as histograms (B, D) or bar graphs (C, E). Flow plots (F) and bar graphs (G)depict the percentage of IL-21–producing splenic CD4 T cells in these chimeras. Data represent two experiments with three mice per group.
4 CUTTING EDGE: ROLE OF CD40 SIGNALING IN EXHAUSTED CD8 T CELL RESCUE
by guest on June 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

–independent role of CD8-intrinsic CD40 signaling duringthe rescue of exhausted CD8 T cells. The current findingscomplemented with a better understanding of the CD40–IL-21 axis in mediating dysfunctional CD8 T cell rescue willlead to more efficacious therapeutic vaccination strategiesagainst tumors and chronic infections.
DisclosuresThe authors have no financial conflicts of interest.
References1. Freeman, G. J., E. J. Wherry, R. Ahmed, and A. H. Sharpe. 2006. Reinvigorating
exhausted HIV-specific T cells via PD-1-PD-1 ligand blockade. J. Exp. Med. 203:2223–2227.
2. Bhadra, R., J. P. Gigley, L. M. Weiss, and I. A. Khan. 2011. Control of Toxoplasmareactivation by rescue of dysfunctional CD8+ T-cell response via PD-1-PDL-1blockade. Proc. Natl. Acad. Sci. USA 108: 9196–9201.
3. Bhadra, R., J. P. Gigley, and I. A. Khan. 2011. The CD8 T-cell road to immu-notherapy of toxoplasmosis. Immunotherapy 3: 789–801.
4. Crawford, A., and E. J. Wherry. 2009. The diversity of costimulatory and inhibitoryreceptor pathways and the regulation of antiviral T cell responses. Curr. Opin.Immunol. 21: 179–186.
5. Bourgeois, C., B. Rocha, and C. Tanchot. 2002. A role for CD40 expression onCD8+ T cells in the generation of CD8+ T cell memory. Science 297: 2060–2063.
6. Lee, B. O., L. Hartson, and T. D. Randall. 2003. CD40-deficient, influenza-specific
CD8 memory T cells develop and function normally in a CD40-sufficient envi-
ronment. J. Exp. Med. 198: 1759–1764.7. Reichmann, G., W. Walker, E. N. Villegas, L. Craig, G. Cai, J. Alexander, and
C. A. Hunter. 2000. The CD40/CD40 ligand interaction is required for resistance
to toxoplasmic encephalitis. Infect. Immun. 68: 1312–1318.8. Yi, J. S., M. Du, and A. J. Zajac. 2009. A vital role for interleukin-21 in the control
of a chronic viral infection. Science 324: 1572–1576.9. Intlekofer, A. M., N. Takemoto, E. J. Wherry, S. A. Longworth, J. T. Northrup,
V. R. Palanivel, A. C. Mullen, C. R. Gasink, S. M. Kaech, J. D. Miller, et al. 2005.
Effector and memory CD8+ T cell fate coupled by T-bet and eomesodermin. Nat.Immunol. 6: 1236–1244.
10. de Totero, D., R. Meazza, S. Zupo, G. Cutrona, S. Matis, M. Colombo, E. Balleari,
I. Pierri, M. Fabbi, M. Capaia, et al. 2006. Interleukin-21 receptor (IL-21R) is up-
regulated by CD40 triggering and mediates proapoptotic signals in chronic lym-
phocytic leukemia B cells. Blood 107: 3708–3715.11. Nurieva, R., X. O. Yang, G. Martinez, Y. Zhang, A. D. Panopoulos, L. Ma,
K. Schluns, Q. Tian, S. S. Watowich, A. M. Jetten, and C. Dong. 2007. Essential
autocrine regulation by IL-21 in the generation of inflammatory T cells. Nature 448:480–483.
12. Nurieva, R. I., Y. Chung, D. Hwang, X. O. Yang, H. S. Kang, L. Ma, Y. H. Wang,
S. S. Watowich, A. M. Jetten, Q. Tian, and C. Dong. 2008. Generation of T
follicular helper cells is mediated by interleukin-21 but independent of T helper 1,
2, or 17 cell lineages. Immunity 29: 138–149.13. Pearce, E. L., M. C. Walsh, P. J. Cejas, G. M. Harms, H. Shen, L. S. Wang,
R. G. Jones, and Y. Choi. 2009. Enhancing CD8 T-cell memory by modulating
fatty acid metabolism. Nature 460: 103–107.
The Journal of Immunology 5
by guest on June 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from




























