Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 264, No. 18, Issue of June 25, pp. 10388-10395.1989 Printed in U.S.A.
Cytochrome P-450 hPCN3, a Novel Cytochrome P-450 IIIA Gene Product That Is Differentially Expressed in Adult Human Liver cDNA AND DEDUCED AMINO ACID SEQUENCE AND DISTINCT SPECIFICITIES OF cDNA-EXPRESSED hPCN1 AND hPCN3 FOR THE METABOLISM OF STEROID HORMONES AND CYCLOSPORINE*
(Received for publication, January 13, 1989)
Toshifumi AoyamaS, Shigeru YamanoSQ, David J. Waxmanll, David P. Lapensonll, Urs A. Meyer 11 , Volker Fischer**, Rachel TyndaleSS, Tadanobu InabaSS, Werner KalowSS, Harry V. GelboinS, and Frank J. GonzalezS From the $Laboratory of Molecular Carcinogenesis, National Cancer Institute, National Institutes of Health, Bethesda, Maryland 20892, the TDepartment of Biological Chemistry and Molecular Pharmacology and Dana Farber Cancer Institute, Harvard Medical School, Boston, Massachusetts 021 15, the 11 Department of Pharmacology Biocenter, University of Basel and the **Sandoz BioDharmaceutical Deoartment. Sandoz Ltd.. Basel, Switzerland, and the $$Department of Pharmacology, University ~ ~ ~~
of Toronto, Tbronto, Ontario M j S IA8, Canada
Immunoblotting analysis of human liver microsome preparations revealed that human cytochrome P-450 PCNl (hPCN1, M, -52,000) was expressed in each of 40 individual specimens examined. In about 10-20% of the livers, an immunologically related protein hav- ing a lower electrophoretic mobility (M. -52,500) was also detected. A single liver was found that expressed only the lower mobility protein, designated hPCN3, and RNA isolated from this liver was used to construct a Xgtll library. The library was screened with an hPCNl cDNA probe resulting in the isolation of a unique full-length cDNA that was sequenced and shown to encode hPCN3. The deduced amino acid se- quence of this cDNA contained 502 residues, a calcu- lated molecular mass of 57,115 daltons, and displayed 84% similarity with hPCN1. The deduced amino-ter- minal sequence of hPCN3 was identical to that of HFLa, a major cytochrome P-450 expressed in human fetal liver that is immunologically cross-reactive with several family I11 cytochrome P-450s. hPCNl and hPCN3 cDNAs were expressed in Hep G2 cells using a vaccinia virus expression system and shown to encode active enzymes, both characterized by reduced CO- binding spectra with X,,, at 450 nm. Enzymatic analy- sis revealed that both cytochrome P-450s were simi- larly active in catalyzing oxidation of the calcium channel blocking drug nifedipine. Both enzymes also catalyzed 6@-hydroxylation of the steroid hormones testosterone, progesterone, and androstenedione, al- though hPCNl exhibited several-fold higher expressed activity than hPCN3. Several minor oxidation prod- ucts of these steroids (e.g. 15&hydroxytestosterone), comprising up to -20% of the total metabolites, were formed by hPCNl but not hPCN3, indicating that hPCN3 is a more highly regiospecific monooxygenase catalyst with steroid substrates. Clear differences were also detected in their catalytic activities toward the
* This work was supported in part by Grant DK 33765 from the National Institutes of Health (to D. J. W.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The nucleotide sequence(s) reported in this paper has been submitted
504813. to the GenBankTM/EMBL Data Bank with accession number(s)
§ Supported by a Japanese Overseas Cancer Fellowship of the Foundation for Promotion of Cancer Research.
immunosuppressive drug cyclosporine, with two hy- droxylated metabolites (M1 and M17) and one demeth- ylated metabolite (M21) formed by hPCNl but only one metabolite (Ml) formed by hPCN3. These studies establish that hPCN3 is a newly described cytochrome P-450 that is differentially expressed in the adult hu- man population and that has overlapping substrate specificity compared to hPCNl for metabolism of ste- roid and drug substrates.
The P-450 cytochromes are the terminal components of the microsomal multisubstrate monooxygenase system. Within the P-4501*’ gene superfamily are three families that constitute the major xenobiotic metabolizing enzymes found in liver and other tissues (Gonzalez, 1988). These are of paramount im- portance in the metabolism of many dietary substances, en- vironmental pollutants, and drugs. Metabolism by P-450s is usually the first step toward the eventual elimination of these chemicals from the body. P-450s are also involved in the metabolic activation of carcinogens to electrophilic metabo- lites that can mutate DNA and transform cells.
The human P-450 I11 gene family codes for enzymes that appear to be involved in the metabolism of endogenous steroid substrates (Waxman et al., 1988), as well as numerous clini- cally important drugs including nifedipine (Guengerich et al., 1986), 17a-ethynylestradiol (Guengerich, 1988), and cyclo- sporine (Kronbach et al., 1988). These and other conclusions regarding the substrate specificity of human family I11 P-450s are, however, primarily based on immunoinhibition and other indirect experiments carried out in isolated liver microsomes. Enzymatic analyses of the purified cytochromes have gener- ally yielded very low catalytic activities. These experiments also do not distinguish between closely related P-450 family I11 forms, which cross-react extensively with the available antibody probes. In the rat, at least two closely related gene family I11 P-450s are expressed in liver tissue, where they are
The abbreviations used are: P-450, cytochrome P-450; TLC, thin layer chromatography; SDS, sodium dodecyl sulfate.
It is now clear that P-450 HLp (Molowa et al., 1986) is a distinct gene product from hPCNl (Gonzalez et al., 1988; Nebert et al., 1989), and we have arbitrarily designated it hPCN2 for the purpose of discussion in this report. The hPCN1, hPCNZ, and hPCN3 genes have been named CYP3A4, CYP3A3, and CYP3A5, respectively (Nebert et al., 1989).
10388

Structure and Regulation of Human Cytochrome P-450 PCN3 10389
subject to markedly different regulation (Gonzalez et al., 1986; Graves et al., 1987). In humans, two P-450s have also been identified in this gene family. These are designated P-450 HLp2 (Molowa et al., 1986) (hPCN2) and P - 4 5 0 ~ ~ (Beaune et al., 1986) or hPCNl (Gonzalez et al., 1988) (P-450 IIIA4) and are 98% identical in their cDNA-deduced amino acid se- quences. Multiple base substitutions, insertions, and deletions in the 3"untranslated regions of the mRNAs coding for P- 450NF/hPCN1 and HLp/hPCN2 strongly suggest that they are products of different genes (Gonzalez et al., 1988). In an earlier study of the interindividual differences in expression of hPCN1, several liver samples were observed to express an immunochemically related protein that migrated slightly slower on denaturing gels than the hPCNl protein and was apparently different from hPCN2 (Waxman et al., 1988). In an effort to characterize that protein, a larger number of livers were examined in the current study, and a single liver was found to express primarily the lower mobility protein. In the present report, we describe the cDNA cloning and expression of this protein, designated P-450 hPCN3, and provide evi- dence that it is related to the major form of P-450 expressed in fetal liver and is also, for yet unknown reasons, expressed in only about 10-20% of adult liver samples. Although both hPCN1 and hPCN3 were found to catalyze nifedipine oxida- tion, they exhibited distinct specificities for the hydroxylation of steroid hormones and for cyclosporine metabolism.
EXPERIMENTAL PROCEDURES
Materials-A monoclonal antibody (13-7-10) that reacts with hPCNl and hPCN3 was obtained from P. Beaune, Hospital Necker, Paris and P. Kremers, Universite de Liege, Belgium (Beaune et al., 1985). Human liver samples K10, K12, K14, K15, K16, K18, K19, K20, and K21 were obtained from the University of Toronto liver bank. KDL 12, KDL 24, and fetal liver samples were from the liver bank at the Biocenter, University of Basel, and livers M1 and M2 were from the University of Miami kidney transplant unit. Phospha- tase-conjugated goat anti-mouse IgG antibody and other reagents used for Western blotting were purchased from KPL Laboratories. Kits for cDNA synthesis were from Bethesda Research Laboratories. Phosphorylated EcoRI linkers, Sepharose CL-4B, and cesium triflu- oroacetic acid were obtained from Pharmacia LKB Biotechnology Inc. All other enzymes were obtained from New England Biolabs or Bethesda Research Laboratories. Phage packaging extracts were pur- chased from Stratagene Cloning Systems. Sequenase was obtained from United States Biochemical Cop. Vaccinia virus strain WR, pSC11, and human TK- 143 cells were obtained from Dr. Bernard Moss at the National Institutes of Health. Hep G2 (ATCC HB 8065) and CV-1 (ATCC CCL70) cells were from the American Type Culture Collection.
Preparation of Liuer Microsomes-Human livers obtained from kidney donors were frozen immediately after resection. Microsomes were prepared from human liver by polytron disruption of the tissue in 0.25 M sucrose at 0-4 "C followed by centrifugation at 10,000 X g for 10 min. The supernatant was then centrifuged at 100,000 X g for 50 min, and the microsomal pellet was washed in 0.15 M KCl. Microsomes were suspended in 100 mM potassium phosphate buffer, pH 7.0, containing 20% w/v glycerol and protein determinations were carried out using the BCA protein determination kit (Pierce Chemical Co.). Western blotting was performed by the method of Towbin et al. (1979), using 10% acrylamide, 0.3% bisacrylamide-containing SDS- polyacrylamide gels (Laemmli, 1970).
Nucleic Acids Methods-RNA was isolated from frozen liver spec- imens using the guanidinium thiocyanate-CsC1 procedure (Chirgwin et al., 19791, except that CsCl was replaced with cesium trifluoroacetic acid. A Xgtll library was constructed using a method similar to that described by Watson and Jackson (1985). A double-stranded cDNA synthesis kit, obtained from Bethesda Research Laboratories, was used for the first step. The cDNA was made blunt ended with T4 DNA polymerase methylated with EcoRI methylase and ligated to EcoRI linkers. The cDNA was then digested with EcoRI and size- selected by using a Sepharose CL-4B column. cDNA greater than 1 kilobase pair was ligated to phosphatase-treated Xgtll (Young and Davis, 1983). The library was screened using a full-length hPCNl
probe (Gonzalez et a!., 1988), and the phage with the largest inserts were identified and processed further. The inserts were subcloned into pUC9 and then sequenced using the M13 shotgun cloning pro- cedure (Deininger, 1983) and the dideoxy chain termination method (Sanger et al., 1977) except that modified T7 DNA polymerase, Sequence@, was substituted for Klenow polymerase (Tabor and Rich- ardson, 1987). Sequence data were analyzed using the Beckman Microgenie@ program.
Vaccinia Virus Expression of P-450 cDNA-The vaccinia virus system was used to express hPCNl and hPCN3 cDNAs. The cDNA inserts were first inserted into the recombination plasmid pSCll (Chakrabarti et al., 1985). The plasmids containing the hPCNl and hPCN3 inserts were then allowed to integrate into the vaccinia virus using the methods described by Mackett et al. (1984) and Chakrabarti et al. (1985). Briefly, CV-1 cells were preinfected with vaccinia virus and then transfected with a mixture of vaccinia virus DNA and pSCll containing the cDNA insert. Virus particles were harvested after several days and used to infect human TK- cells in the presence of 5-bromodeoxyuridine. Recombinant virus particles were selected based on their ability to grow in the presence of this agent and on their ability to produce native @-galactosidase (Chakrabarti et al., 1985). The basis of this selection is that thymidine kinase (TK) sequences in pSCl1 integrate into the active TK gene of the vaccinia virus producing an inactive TK gene (due to the disruptive affect of the integrated plasmid). Only viruses that do not have an active TK gene are capable of growing in the presence of 5-bromodeoxyuridine. Recombinant viruses that contain the pSCll integrated DNA are selected for the presence of the Lac-Z (@-galactosidase) gene which is encoded by pSC11. Final identification of vaccinia virus producing hPCNl and hPCN3 was carried out by immunoblotting of cell lysate protein using monoclonal antibody 13-7-10 (Beaune et al., 1985).
Enzymatic Analysis of Expressed P-450"For production of ex- pressed P-450 protein, recombinant vaccinia virus hPCNl and hPCN3, designated vPCNl and vPCN3, respectively, were used to infect Hep G2 cells. Cells were harvested 48 h after infection and then sonicated briefly to disrupt cells. Sonicates, or in some cases isolated microsomes, were assayed for nifedipine oxidase activity (Guengerich et al., 1986) and cyclosporine metabolism (Kronbach et al., 1988) as described previously. Hydroxylations of testosterone, progesterone, and androstenedione were assayed as described (Wax- man et al., 1988). Hydroxysteroid metabolites were identified by the general methods described previously (Waxman et al., 1983, 1988) with specific details provided in Table IV. Compositions of TLC solvents A-M (Table IV) are provided in those earlier references. Composition of other TLC solvents used in the present study are as follows: solvent N = ethyl acetate/n-hexane/acetic acid (16/8/1), solvent 0 = benzene/ethyl acetate (3/1), solvent P = methylene chloride/100% ethanol (97/3), solvent Q = ethyl acetate/chloroform (l/l), solvent R = benzene/ethyl acetate/acetone (10/1/1) and solvent X = chloroform/ethyl acetate/100% ethanol (4/1/0.2). Ethoxycou- marin 0-dealkylase assays were performed as described by Greenlee and Poland (1978). Benzo(a)pyrene hydroxylase assays were carried out using the method of Nebert and Gelboin (1968).
RESULTS
Immunoblot Analysis of Human Liver Microsomal Protein- Immunoblot analysis of isolated human Iiver microsomes using a monoclonal antibody reactive with hPCN1 revealed a band of 52,000 daltons as the major reactive polypeptide (Fig. 1). This band was shown to correspond to hPCNl (see below). In three of the liver samples shown in this experiment (K20, KDL 12, and KDL 24), however, a band of 52,500 daltons was also detected with the monoclonal antibody. Moreover, in liver K15, the 52,500-dalton band was the major immuno- reactive protein, with little or no hPCNl detectable. There was no apparent correlation between the level of the 52,500- dalton protein and the age, sex, or medication status of the donor^.^ In order to further characterize this 52,500-dalton protein, liver K15 was used to isolate cDNAs encoding this protein.
Isolation. and Sequence of the hPCN3 cDNA-A cDNA
A table describing the characteristics of these liver donors is available from the authors upon request.

10390 St ruc ture and Regulation of Human Cytochrome P-450 PCN3
PCN3 I P C N l
FIG. 1. Immunoblot analysis of human liver microsomes using monoclonal antibody 13-7-10 against hPCN1. Human liver microsomal protein (20 pg) was electrophoresed on a 10% SDS- polyacrylamide gel and transferred to nitrocellulose. The paper was then developed as described under “Experimental Procedures” to visualize the hPCNl and hPCN3 proteins.
library derived from K15 liver mRNA was prepared in the vector X g t l l . Screening of this library with a probe for hPCN1 resulted in the isolation of a cDNA, designated hPCN3, that was closely related to but distinct from hPCN1. The complete nucleotide and deduced amino acid sequence of this cDNA is shown in Fig. 2 and compared with the cDNA and deduced amino acid sequence of hPCNl (Gonzalez et al., 1988). hPCN3 and hPCN1 cDNAs displayed 89% nucleotide and 84% de- duced amino acid sequence similarity. In addition, hPCN3 displayed 82% cDNA-deduced amino acid sequence similarity with hPCN2’ (Table I). hPCN2 is identical to hPCNl except for 10 amino acid differences (Fig. 2). However, multiple sequence differences were found in the nontranslated regions between the hPCNl and hPCN2 cDNAs indicating that both are derived from different genes (Gonzalez et al., 1988). hPCN3 also displayed 72 and 71% amino acid similarities with rat PCNl and PCN2, respectively, and 73% similarity with rabbit P-450 3c (Table I). hPCNl also shows no preferred sequence similarity with rat PCN1, rat PCN2, or rabbit P- 450 3c.
Alignment of the hPCN1 and hPCN3 sequences revealed that regions of high nucleotide and amino acid similarities are interspersed with regions of low similarity. This is most apparent in the segment from amino acid residues 206-269 in which 64 amino acid residues contain 24 mismatches (62% similarity). In contrast, between residues 80 and 205, a high (92%) similarity exists along 126 amino acids. Another region of low (69%) similarity is seen between residues 28 and 79 (Fig. 2). This pattern of interspersed high and low sequence similarities is indicative of gene conversion events in which one gene has donated a region of its sequence to another, often adjacent, gene.
Comparison of the cDNA-deduced amino acid sequenceof hPCN3 with published P-450 protein sequences revealed that the partial amino terminal sequence of human fetal liver P- 450 form HFLa (Kitada et al., 1987), which is immunologically related to several family IIIA P-450s, displayed sequence identity with hPCN3. Seven amino acids reported for HFLa, corresponding to residues 4-10 of hPCN3, were identical to those of hPCN3. Others have purified a human fetal liver P- 450, designated P-450 HLp2, that presumably is identical to
HFLa (Wrighton and Vandenbranden, 1989). The sequence of 29 of the 30 amino-terminal residues of HLp2 are the same as those deduced for hPCN3 including residues 2, 6, 10, 28, and 29 that are distinct from hPCNl (residue 20 is Ile in HLp2 and Val in hPCN3). These data suggest that hPCN3 is closely related or perhaps identical to the fetal P-450 HFLa/ HLp2 proteins.
Expression of hPCNI and hPCN3 Using Vaccinia Virus- In order to characterize P-450 forms hPCNl and hPCN3 with respect to their electrophoretic, spectral, and catalytic prop- erties, their respective cDNAs were inserted into vaccinia virus as outlined under “Experimental Procedures.” Hep G2 cells were infected with the reultant recombinant virus par- ticles, vPCNl and vPCN3, and cell lysates were then analyzed by immunoblotting with a monoclonal antibody reactive with hPCN1. vPCN1 and vPCN3 were thus shown to direct the expression of proteins that were reactive with the monoclonal antibody and that exhibited mobilities on SDS-polyacryl- amide gels indistinguishable from the 52,000- and 52,500- dalton proteins of the KDL 24 liver (Fig. 3A). This was verified by mixing experiments, which demonstrated that the expressed hPCN1 had the same mobility as the 52,000-dalton protein found in liver M2 and other livers, whereas the expressed hPCN3 migrated with the 52,500-dalton protein of liver K15 (Fig. 3B).
Spectral and Enzymatic Analysis of hPCN3-M’ lcrosomes were isolated from cells infected with vPCN1, vPCN2, and wild type vaccinia (vWT) and analyzed for P-450 content by formation of the reduced carbon monoxide-bound P-450 ab- sorption spectrum (Omura and Sato, 1964). Microsomes from vWT did not display detectable absorption at 450 nm when reduced in the presence of carbon monoxide, indicating the absence of significant endogenous P-450 in these cells (Fig. 4). In contrast, hPCNl and hPCN3 each produced absorption spectra with peaks a t 450 nm. These results, quantitated in Table 11, indicate that the specific P-450 content of cells expressing hPCNl is somewhat more than those that ex- pressed hPCN3. Due to the lack of purified human hPCNl and hPCN3 proteins to use as standards we were unable to quantify the specific heme content of these P-450s produced in vaccinia virus. However, studies of other P-450 proteins that we have expressed in this system, for which purified P- 450 standards are available, indicate that virtually all of the protein produced is spectrally active.
The expressed enzymes were assayed for activities against some common P-450 substrates. Both hPCNl and hPCN3 were able to catalyze 0-deethylation of 7-ethoxycoumarin (Table 111), although hPCNl was considerably more active than hPCN3 with this substrate. hPCNl was also more active than hPCN3 in the oxidation of benzo(a)pyrene. These com- pounds were relatively poor substrates for both enzymes, however.
Recent studies established that cDNA-expressed hPCNl can metabolize nifedipine (Gonzalez et al., 1988), which con- firmed earlier work using reconstitution and immunoinhibi- tion experiments (Guengerich et al., 1986). Analysis of cells infected with vPCNl and vPCN3 revealed that both enzymes can actively catalyze nifedipine oxidation (Table 111).
Previous studies have established that human liver micro- somes oxidize steroid hormones primarily at the 6P position and that this activity is catalyzed by one or more members of P-450 gene subfamily IIIA (Waxman et al., 1988; Kawano et al., 1987). The substrate specificities of hPCNl and hPCN3 were therefore characterized using the steroids testosterone, progesterone, and androstenedione (Table IV). Both P-450 forms catalyzed 6P-hydroxylation of all three steroids. hPCNl

Structure and Regulation of Human Cytochrome P-450 PCN3 10391 hPCNl GCA A T C T G A A G G G G G G A A G T A T CT G C CA hPCN3 GGGAAGCTCCAGGCAAACAGCCCAGCAAACAGCAGCACTCAGCTAAAAGGAAGACTCACAGAACACAGTTGAAGAAGGAAAGTGGCGATGGACCTCATCCCAAATTTGGCGGTGGAA 117 hPCN3 hPCN1
MetAspLeulleProAsnLeuAlaVatG~u 10 Ala Asp Met
ACCTGGCTTCTCCTGGCTGTCAGCCTGGTGCTCCTCTATCTATATGGGACCCGTACACATGGACTTTTTMGAGACTGGGMTTCCAGGGCCCACACCTCTGCCTTTGTTGGGAAATGTT 237 ThrTrpLeuLeuLeuAlaValSerLeuValLeuLeuTyrLeuTyrGlyThrArgThrHisGlyLeuPheLysArgLeuGlylleProGlyProThrProLeuProLeuLeuGlyASnVal 50
A A T AG T T A
28 H i sSer LYS Phe I l e -
C A A C T T T T G T G A T C G CTTT T AG T T A TTGTCCTATCGTCAGGGTCTCTGGAAATTTGACACAGAGTGCTATAAAAAGTATGGAAAAATGTGGGGAACGTATGAAGGTCAACTCCCTGTGCTGGCCATCACAGATCCCGACGTGATC 357 LeuSerTyrArgGlnGlyLeuTrpLysPheAspThrGluCysTyrLysLysTyrGlyLysMetTrpGlyThrTyrGluGlyGlnLeuProValLeuAlalleThrAspProAspValIle 90
HisLys PheCysMet Met His Va 1 Phe Asp Gln Met
( C T ) (CGA) 79 80 "
AGAACAGTGCTAGTGAAAGAATGTTATTCTGTCTTCACAAATCGAAGGTCTTTAGGCCCAGTGGGATTTATGAAAAGTGCCATCTCTTTAGCTGAGGATGAAGAATGGAAGAGAATACGG 477 ArgThrValLeuValLysGluCysTyrSerValPheThrAsnArgArgSerLeuGlyProValGlyPheMetLysSerAla~leSerLeuAlaGluAspGluGluTrpLysArglleArg 130
A C G C T T A T A
LYS (Leu)
ProPhe I l e (Glu)
Leu
T G T G T C C (CG)
TCATTGCTGTCTCCAACCTTCACCAGCGGAAAACTCAAGGAGATGTTCCCCATCATTGCCCAGTATGGAGATGTATTGGTGAGAAACTTGAGGCGGGAAGCAGAGAMGGCAAGCCTGTC 597 SerLeuLeuSerProThrPheThrSerGlyLysLeuLysGluMetPheProIlelleAlaGlnTyrGlyAspValLeuValArgAsnLeuArgArgGluAlaGluLysGlyLysProVal 170
Va 1 Thr
( T I G
. . . . C A
. .
ACCTTGAAAGACATCTTTGGGGCCTACAGCATGGATGTGATTACTGGCACATCATTTGGAGTGAACATCGACTCTCTCAACAATCCACAAGACCCCTTTGTGGAGAGCACTAAG~GTTC 717 ThrLeuLysAspIlePheGlyAlaTyrSerMetAspValIleThrGlyThrSerPheGl~alAsnIleAspSerLeuAsnAsnProGlnAspProPheValGluSerThrLysLysPhe 210
A A C C T
Va l Ser Asn Leu (Ser) (Val) (Leu) 205206
T G A T G C C G C T A C T A G G G AGTT C A
LeuLysPheGlyPheLeuAs~roLeuPheLeuSerIleIleLeuPheProPheLeuThrProValPheGluAlaLeuAsnValSerLe~heProLysAspThr~leAsnPheLeuSer 250 CTAAAATTTGGTTTCTTAGATCCATTATTTCTCTCAATAATACTCTTTCCATTCCTTACCCCAGTTTTTGAAGCATTAAATGTCTCTCTGTTTCC~GATACCATAAATTTTTTAAGT a37
Arg Asp Phe ThrVal I l e I l eLeu Va l I LeCysVal ArgGluValThr Arg
(GI A G AG G A T C G G TP A
AAATCTGTAAACAGAATGAAGAAAAGTCGCCTCAACGACAAACAAAAGCACCGACTAGATTTCCTTCAGCTGATGATTGACTCC GAATTCGAAAGAAACTGAGTCCCACAAAGCTCTG 957 LysSerValAsnArgMetLysLysSerArgLeuAsnAspLysGlnLysHisArgLeuAspPheLeuGlnLeuMetIleAspSer nAsnSerLysGluThrGluSerHisLysAIaLeu 290
LYS GlU Glu Thr (Val)
C
Va 1 269 y\ HisLys)
TCTGATCTGGAGCTCGCAGCCCAGTCAATAATCTTCATTTTTGCTGGCTATGAAACCACCAGCAGTGTTCTTTCCTTCACTTTATATGAACTGGCCACTCACCCTGATGTCCAGCAGAAA 1077 SerAspLeuGluLeuAlaAlaGlnSerIleIlePheIlePheAlaGlyTyrGluThrThrSerSerValLeuSerPheThrLeuTyrGluLe~laThrHisProAspValGtnGlnLys 330
T G A T T G C T A G
Va 1 I leMet
GG A CTGCAAAAGGAGATTGATGCAGTTTTGCCCAATAAGGCACCACCTACCTATGATGCCGTGGTACAGATGGAGTACCTTGACATGGTGGTGAATGAAACACTCAGATTATTCCCAGTTGCT 1197 LeuGlnLysGlulleAspAlaValLeuProAsnLysAlaProProThrTyrAs~laValValGlnMetGluTyrLeuAs~etValValAsnGluThrLeuArgLeuPheProValAla 370
A C A T C T G A
Glu Thr Leu 1 l e
G GTC A G A G GTGG A GC G (TG)
ATTAGACTTGAGAGGACTTGCAAGAAAGATGTTGAAATCAATGGGGTATTCATTCCCAAAGGGTCAATGGTGGTGATTCCAACTTATGCTCTTCACCATGACCCAAAGTACTGGACAGAG 1317 IleArgLeuGluArgThrCysLysLysAspValGluIleAsnGlyValPheIleProLysGlySerMetValValIleProThrTyrAlaLeuHisHisAspProLysTyrTrpThrGlu 410 Met Va l Met ValVal Met Ser Arg
(Trp)
A T A C AAC A G
Lys Leu Asn Asn Ser
CCTGAGGACTTCCGCCCTGMAAGGTTCAGTAAGAAG---MGGACAGCATAGATCCTTACATATACACACCCTTTGGAACTGGACCCAGAMCTGCATTGGCATGAGGTTTGCTCTCATG 1434 ProGluGluPheArgProGluArgPheSerLysLys LYS~SPSertteAS~roTyrl~eTyrThrProPheGlyThrGlyProArgAsn~lleGlyHetArgPheAlaL~let 449
T G G C G CTAAAGGTGGATTCAAGAGATGGAACCCTMGTGGAGAATGAGTTATTCTAAGGACTTCTACTTTGGTCTTCAAGAAAGCTGTGCCCCAGAACACCAGAGATTTCAACTTAGTC----AA 1670
CC A T C G C T TG CC A C T T G T G
LeuLysValAspSerArgAspClyThrLeuSerGlyGlu Glu Va 1 Ala
502
G TC G TAAAACCTTGA-GATGGGCTTAATCTAATGTA
C C GACTGCATAAATAACCGGGGATTCTGTACATGCATTGAGCTCTCTCA~TGTCTGTGTAGAGTGTTATACTTGGGAATATAAAGGA 1708
GGTGACCAAATCAGTGTGAGGAGGTAGATTTGGCTCCTTTGCTTCTCACGGGACTATTTCCACCACCCCCAGTTAGCACCATTAACTCCTCCTGAGCTCTGATAAGAGAATCAACATTTC
TCAATAATTTCCTCCACAAATTATTAATGAAAATAAGAATTATTTTGATGGCTCTAACAATGACATTTATATCACATGTTTTCTCTGGAGTATTCTATAAGTTTTATGTTAAATC~
AGACCACTTTACAAAGTA - FIG. 2. Complete sequence of hPCN3 cDNA. The cDNA sequence of hPCN3 was determined and its amino
acids was deduced. The nucleotide and amino acid sequence of hPCNl is shown above and below those of hPCN3 with only the residues of hPCNl displayed that do not match hPCN3. hPCN2 was aligned with hPCNl only across the amino acid coding region. hPCN2 is identical to hPCNl across this region except for several nucleotide substitutions resulting in 10 amino acid differences (in parentheses). The ATA triplet between residues 922 and 923 is found in hPCN2 but not in hPCNl or hPCN3. The 3'-noncoding region of hPCN2 was not aligned. This alignment can be found elsewhere (Gonzalez et al., 1988). The underlined numbers refer to those amino acids and segments of hPCN3 discussed in the text. The cysteine residue at position 441 that binds the heme iron and the polyadenylation signals are underlined.

10392 Struc ture and Regulation of Human Cytochrome P-450 PCN3 TABLE I
Comparison of cDNA-deduced amino acid sequence of hPCN3 with sequences of other members of P-450 family III
cDNA-deduced amino acid similarities were calculated using the Mirogenie alignment program. The rat and rabbit sequence data were taken from Gonzalez et al. (1986) and Dalet et al. (1988), respectively.
hPCN3 hPCNl Rabbit3c RatPCNl % similarity
hPCN3 100 hPCNl 84 100 hPCN2 82 98 72 Rabbit 3c 73 74 100 Rat PCNl 72 73 69 100 Rat PCN2 71 72 67 89
A
PCN3 'PCNl
FIG. 3. Comparison of vaccinia expressed hPCNl and hPCN3 proteins with the 52,000- and 52,500-dalton proteins present in human liver microsomes. Hep G2 cell lysate protein (20 pg) derived from cells infected with vPCN1, vPCN3, and vWT were subjected to Western blotting analysis and compared to the KDL 24 microsomal protein ( A ) . Mixing experiments were also performed using a preparation of human microsomes that contained only hPCN3 (K15) and a preparation that contained only hPCNl (M2). Microsomal protein (10 pg) and vPCNl or vPCN3 infected lysate protein (10 pg) were mixed and analyzed ( B ) .
I I 450 500 550
WAVELENGTH (nm)
FIG. 4. Spectral quantitation of vacinia expressed hPCNl and hPCN3. Microsomes were isolated from cells infected with vPCNl and vPCN3. Samples were bubbled with carbon monoxide and then reduced with dithionite and compared by difference spec- troscopy to samples that had only been reduced. Spectra were re- corded with an Aminco DW 2000 spectrophotometer. -, vPCN1; . . . ., v P C N ~ ; -- -, vWT.
TABLE I1 Carbon monoxide difference spectrum and specific P-450 content of
vaccinia virus-expressed human hPCNl and hPCN3 Microsomes were prepared from Hep G2 cells infected with recom-
binant vaccinia virus and suspended with 100 mM potassium phos- phate, pH 7.25, containing 20% w/v glycerol and 0.2% Emulgen 913. The microsomal fractions (14 mg/ml protein for hPCN3 and hPCN1, respectively) were then analyzed for P-450 and P-420 content as shown in Fig. 4.
Expressed P-450 450 nm 420 nm' Total Specific contentb
PM pmollml: hPCNl 0.96 0.09 1.05 75 hPCN3 0.83 0.04 0.87 62
Represents denatured form of the cytochrome. *Picomoles spectral (P-450 + P-420)/mg Hep G2 microsomal
protein.
TABLE I11 Activities of vaccinia virus-expressed hPCNl and hPCN3
Hep G2 cells were infected with recombinant vaccinia virus ex- pressing human hPCNl (vPCN) and hPCN3 (vPCN3) or with wild type vaccinia virus (vWT). Cells were harvested 60 h after infection, sonicated, and assayed for activities as described under "Experimental Procedures" either directly or after preparation of microsomes. Cells infected with vWR produced no detectable activities. Values are the average of duplicate determinations. ND, not detectable.
Activities hPCNl hPCN3 pmollminlmg protein
7-Ethoxycoumarin 0-deethylase 17 (122)" 1.6 (12)" Benzo(a)pyrene hydroxylase 6.2 3.0 Nifedipine oxidase 290 260 Cyclosporine
M17 formation 180 110 M1 formation 170 ND M21 formation 160 ND
Activities determined using isolated microsome preparations.
was apparently more active than hPCN3 in the vaccinia- infected Hep G2 cell microsomes (4-7-fold) when compared on a per mg Hep G2 microsomal protein basis. However, both cytochromes exhibited catalytic rates that were high (corre- sponding to 60-hydroxylation turnover numbers of 3 min" (hPCN3) or 15 min" (hPCN1) with testosterone as substrate) and are comparable to those exhibited by many purified P- 450s. Several minor metabolites were also formed by the expressed P-450s, at rates totalling up to -20% of the 60- hydroxylase rates. These include the testosterone metabolite 15~-hydroxytestosterone, the progesterone metabolite 16a- hydroxyprogesterone, several unidentified steroid metabolites (Table IV), and A'-testosterone4 (Nagata et al., 1986). Quan- titation of these metabolites (Table IV) indicated that hPCNl and hPCN3 exhibit similar metabolite profiles with andro- stenedione as substrate. With testosterone and progesterone, however, hPCNl but not hPCN3 catalyzed formation of the minor metabolites in characteristic ratios relative to the 6p- hydroxylated compounds (Table IV). Interestingly, hPCNl exhibits the same activity profile with these steroid substrates as reported previously for human liver microsomes (testoster- one > progesterone > androstenedione; Waxman et al., 1988). Moreover, the testosterone metabolite profile of hPCNl (60/ 150 product ratio = 100/4) is the same as that exhibited by human liver microsomes (60/150 = 100/3-4) (Waxman et dl., 1988). These observations strengthen the earlier conclusion that hPCNl is the major catalyst of steroid 60-hydroxylation in human liver microsomes.
IT. Aoyama, S. Yamano, D. J. Waxman, D. P. Lapenson, U. A. Meyer, V. Fischer, R. Tyndale, T. Inaba, W. Kalow, H. V. Gelboin, and F. J. Gonzalez, unpublished observations.
Structure and Regulation of Human Cytochrome P-450 PCN3 10393
TABLE IV Steroid hydroxylase activities of vaccinia
virus-expressed hPCNI and hPCN3 Hep G2 cells were infected with vPCNl and vPCN3. Cells were
harvested 60 h after infection, sonicated, and microsomal fractions isolated by ultracentrifugation. Steroid hydroxylase activities were determined at 50 WM steroid substrate as described under “Experi- mental Procedures” after subtraction of low level activities exhibited by mock-infected cells analyzed in parallel (typically 52-3 pmol/ min/mg). Highly similar metabolite profiles were obtained upon analysis of Hep G2 cell sonicates, which exhibited -4-fold lower hydroxylase activities/mg protein than the isolated microsomes.
hPCNl hPCN3 hPCNl/hPCN3 pmol/min/mg
Testosterone metabolites” 60-Hydroxytestosterone 1126 170 6.6 15P-Hydroxytestosterone 45 c 2 >22 Unknown lb 32 C2 >16
6P-Hydroxyprogesterone 873 129 6.8 16a-Hydroxypr~gesterone~ 117 <2 >58
6P-Hydroxyandrostene- 406 103 3.9
Unknown 3’ 34 12 2.8 Unknown 4g 30 3 10
Progesterone metabolites‘
Androstenedione metabolites‘
dione
a Metabolites resolved on silica gel TLC plates developed sequen- tially in solvents AB (see methods for TLC solvent compositions). Identifications are based on co-chromatography of TLC-purified me- tabolites with authentic standards in solvents AA and BB. Each metabolite was also distinguished from 16 other available monohy- droxytestosterones using these methods. Possible formation of 20- OH-testosterone (which is formed by immunoreactive hPCNl in human liver microsomes) (Waxman et al., 1988) could not be assessed owing to interference from other testosterone metabolites formed by cells infected with wild type virus.
This metabolite comigrated with llp-OH-testosterone and 19- OH-testosterone in solvents AA, but was distinguished from both these compounds (and all other available standards) in solvents BB.
Metabolites resolved on TLC using solvent NRR. Co-chromatog- raphy of TLC-purified metabolites with authentic standards was carried out in three independent TLC solvent systems, designated OL, M, and X. Both metabolites were also distinguished from 14 other available monohydroxyprogesterones, as well as the 200-hy- droxy and BOP-hydroxy derivatives of progesterone. A minor metab- olite that migrated in the vicinity of Pa-OH-progesterone was formed by hPCNl but not hPCN3, but could not be reliably quantitated owing to the interference by progesterone metabolites (perhaps re- duction products) also formed by the uninfected cells.
Tentative identification. e Metabolites resolved on TLC using solvents PQ. Co-chromatog-
raphy of TLC-purified metabolites with authentic standards was carried out in TLC solvent systems PQ, HH, and AA. Each metabolite was also distinguished from 11 other available monohydroxyandro- stenediones. ’ Metabolite migrates close to [“C] 16P-hydroxyandrostenedione formed enzymatically by purified rat liver P-450 PB-4 (IIB1).
Metabolite comigrates with 15a-hydroxyandrostenedione and 601- hydroxyandrostenedione in solvent HH and AA, but was distin- guished from both these compounds (and all other available stand- ards) in solvents PQ.
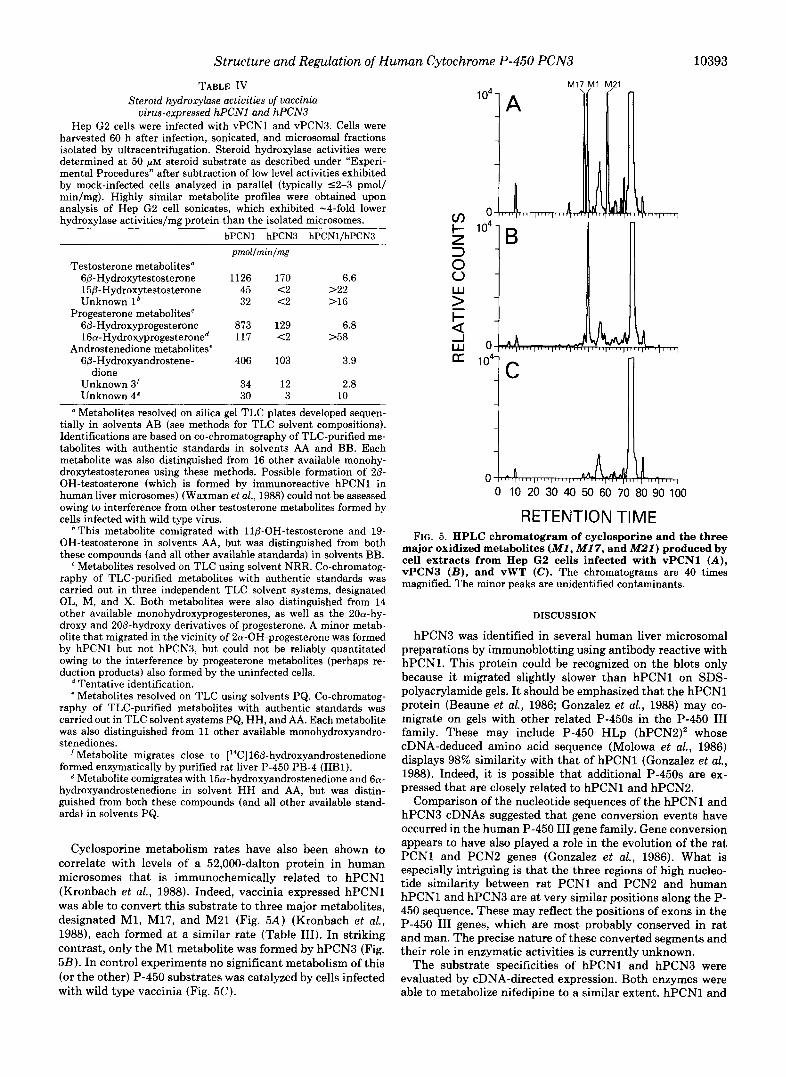
Cyclosporine metabolism rates have also been shown to correlate with levels of a 52,000-dalton protein in human microsomes that is immunochemically related to hPCN1 (Kronbach et al., 1988). Indeed, vaccinia expressed hPCN1 was able to convert this substrate to three major metabolites, designated M1, M17, and M21 (Fig. 5A) (Kronbach et al., 1988), each formed at a similar rate (Table 111). In striking contrast, only the M1 metabolite was formed by hPCN3 (Fig. 5B). In control experiments no significant metabolism of this (or the other) P-450 substrates was catalyzed by cells infected with wild type vaccinia (Fig. 5C).
A M1
L
!1
ul r
C I
1 0 10 20 30 40 50 60 70 80 90 100
RETENTION TIME FIG. 5. HPLC chromatogram of cyclosporine and the three
major oxidized metabolites ( M I , MI 7, and M22) produced by cell extracts from Hep G2 cells infected with vPCNl ( A ) , vPCN3 ( B ) , and vWT (C). The chromatograms are 40 times magnified. The minor peaks are unidentified contaminants.
DISCUSSION
hPCN3 was identified in several human liver microsomal preparations by immunoblotting using antibody reactive with hPCN1. This protein could be recognized on the blots only because it migrated slightly slower than hPCNl on SDS- polyacrylamide gels. I t should be emphasized that the hPCNl protein (Beaune et al., 1986; Gonzalez et al., 1988) may co- migrate on gels with other related P-450s in the P-450 I11 family. These may include P-450 HLp (hPCN2)’ whose cDNA-deduced amino acid sequence (Molowa et al., 1986) displays 98% similarity with that of hPCNl (Gonzalez et al., 1988). Indeed, it is possible that additional P-450s are ex- pressed that are closely related to hPCNl and hPCN2.
Comparison of the nucleotide sequences of the hPCNl and hPCN3 cDNAs suggested that gene conversion events have occurred in the human P-450 I11 gene family. Gene conversion appears to have also played a role in the evolution of the rat PCNl and PCN2 genes (Gonzalez et al., 1986). What is especially intriguing is that the three regions of hlgh nucleo- tide similarity between rat PCNl and PCN2 and human hPCNl and hPCN3 are at very similar positions along the P- 450 sequence. These may reflect the positions of exons in the P-450 I11 genes, which are most probably conserved in rat and man. The precise nature of these converted segments and their role in enzymatic activities is currently unknown.
The substrate specificities of hPCNl and hPCN3 were evaluated by cDNA-directed expression. Both enzymes were able to metabolize nifedipine to a similar extent. hPCNl and

10394 Structure and Regulation of Human Cytochrome P-450 PCN3
hPCN3 also catalyzed steroid hormone 66-hydroxylation; however, hPCNl was several-fold more active and somewhat less regiospecific towards the steroid substrates than hPCN3. In addition, the two P-450s exhibited strikingly different activities toward cyclosporine. hPCNl catalyzed the forma- tion of three oxidized metabolites of cyclosporine, MI, M17 and M21. M1 and M17 are formed by hydroxylations on amino acids 9 and 1, respectively, and M21 by a demethylation on amino acid 4 (Kronbach et al., 1988). In contrast, hPCN3 produced only the M1 metabolite. These data indicate that hPCNl is a less regiospecific catalyst than hPCN3 with this drug substrate, suggesting that hPCNl may be able to bind cyclosporine at more than one site or with multiple orienta- tions at the active site heme pocket.
The present demonstration that two closely related human gene family I11 P-450s exhibit similar (albeit distinguishable) substrate specificities raises the question of whether one form or the other serves as the primary catalyst of microsomal reactions previously associated with human family I11 P-450s. In the case of nifedipine oxidation, hPCNl and hPCN3 ex- hibited similar catalytic activities when expressed in Hep G2 cells, suggesting that both enzymes contribute significantly to human liver microsomal metabolism of this substrate, at least in individuals that express hPCN3 at reasonable levels. It seems, therefore, that the variability in expression of both hPCN15 and hPCN3 in individual human livers contributes to the wide range of nifedipine oxidation rates observed in clinical studies (Kleinbloesem et al., 1984). In the case of steroid 66-hydroxylation, however, hPCNl appears to be sev- eral-fold more active than hPCN3, suggesting that hPCNl may be the major catalyst of these reactions in human liver microsomes.6 The observation that levels of hPCN13 (Wax- man et al., 1988) but not hPCN3 (data not shown) measured in human liver microsomes correlate well with microsomal steroid 6/3-hydroxylation rates ( r = 0.83 for n = 25 livers) is consistent with this conclusion.
On the basis of immunoblotting analysis, hPCN3 was found t o be expressed in only about 10-20%7 of adult livers analyzed. In contrast, hPCNl was expressed in all the livers examined (albeit at a variable level) with the exception of K15, where only hPCN3 was found. Although K15 apparently lacks sig- nificant expression of hPCN1, this liver was normal for other P-450-mediated oxidations including the metabolism of diaze- pam and ethoxyresorufin (Inaba et al., 1988), mephenytoin' and sparteine (Tyndale et al., 1988). The differential expres- sion of hPCN3 did not correlate with sex, age, or known drug treatment. It remains possible that the K15 liver may also contain hPCNl at a much lower content than hPCN3. How- ever, only hPCN3-type clones were obtained after screening of a Xgtll library produced from K15 RNA.
The differential expression of hPCN3 in adult livers de- scribed in the present study could have several possible expla- nations. For instance (i) the hPCN3 gene may be partially defective in man with only a small number of alleles still active in the population. (ii) Expression of the hPCN3 gene may be inducible by some unknown dietary, drug, or hormonal factors such as apparently occurs with hPCNl and/or hPCN2 (Molowa et al., 1986) as well as the corresponding rat and
Since the available antibody probes are unlikely to distinguish between hPCNl and hPCN2 in Western blot analysis, it must be assumed that the previous analyses provide the total hPCNl + hPCN2 content.
A possible added complexity, which cannot be assessed at this time, is the role of P-450 HLp (hPCN2) in these reactions. ' These values are ranges based on the different liver banks that
were included in these analyses (total number of samples = 40). T. Inaba, unpublished data.
rabbit proteins in this family. This possibility cannot be evaluated at this time due to the limited information available regarding the medical histories of the individuals from which the livers were derived. Finally, (iii) genes involved in the regulation of hPCN3, such as those encoding receptors or other trans-acting factors, may be differentially expressed in the human population. In any case, it would be of great interest to devise methods to phenotype individuals for expression hPCN3 in uiuo. Two approaches are possible. First, restriction fragment length polymorphisms would be useful if one could be found that is linked either to the expressed hPCN3 gene or to its lack of expression. Another possibility is to find a substrate that is specific to hPCN3 than can be used in a urine phenotyping test. Indeed RFLP linkage to mutant P-450 genes (Skoda et al., 1988) and urine metabolite analysis (Idle and Smith, 1979) have been used to determine defective human debrisoquine 4-hydroxylase genes.
On the basis of gel mobility analysis (data not shown) and the identity with partial amino-terminal protein sequence (Kitata et al., 1987; Wrighton and Vandenbranden, 1989), it appears that hPCN3 may be the same as HFLa/HLp2, a major P-450 expressed in human fetal liver. This finding would suggest that the differential expression of hPCN3 in adult liver reflects a regulatory difference according to either point ii or point iii above. It is not known whether rat PCNl or PCN2 are expressed in fetal hepatocytes. Rat PCN2, which is expressed in adult male rats and is suppressed in females in the post-suckling period, is not seen until just after birth (Waxman et al., 1985; Gonzalez et al., 1986). These data indicate a marked difference in the regulation of P-450 I11 genes between rats and man. However, it cannot be ruled out that hPCN3/HFLa expression in the fetus is due to transpla- cental induction as suggested earlier (Kitata et al., 1987). The function of hPCN3/HFLa in the human fetus is currently unknown; however, it is possible that this enzyme plays a role in fetal steroid metabolism. Expression of hPCN3 in vaccinia virus and screening of large numbers of substrates may pro- vide a better understanding of the functions of this P-450 enzyme.
Acknowledgments-We wish to thank Dr. Raimund Peter, Univer- sity of Erlanger, BRD, for performing the nifedipine oxidase assays, and Dr. Elzbieta Holsztynska, Dana Farber Cancer Institute, for assistance with amino acid sequence analysis.
Note Added in Proof-While this manuscript was in the press, the complete sequence of fetal HFLa cDNA-deduced protein was pub- lished (Komori et al., 1989). This P-450 is distinct from hPCN3 and displays 88,86, and 82% amino acid similarities with hPCN1, hPCN2, and hPCN3, respectively.
REFERENCES
Beaune, P., Kremers, P., Letawe-Goujon, F., and Gielen, J. E. (1985) Biochem. Phnrmacol. 34, 3547-3552
Beaune, P., Umbenhauer, D. W., Bork, R. W., Lloyd, R. S., and
8068 Guengerich, F. P. (1986) Proc. Natl. Acad. Sci. U. S. A. 83, 8064-
Chakrabarti, S., Brechling, K., and Moss, B. (1985) Mol. Cell. Biol.
Chirgwin, J. M., Przyhyla, A. E., MacDonald, R. J., and Rutter, W.
Dalet, C., Clair, P., Daujat, M., Fort, P., Blanchard, J. M., and Maurel,
Deininger, P. L. (1983) Anal. Biochern. 129, 216-223 Gonzalez, F. J. (1988) Pharmacol. Reu. 40, 243-288 Gonzalez, F. J., Song, B. J., and Hardwick, J. P. (1986) Mol. Cell.
Biol. 6,2969-2976 Gonzalez, F. J., Schmid, B. J., Umeno, M., McBride, 0. W., Hardwick,
J. P., Meyer, U. A., Gelboin, H. V., and Idle, J. R. (1988) DNA 7,
6,3403-3409
J. (1979) Biochemistry 18,5294-5299
P. (1988) DNA 7,39-46
79-86

Structure and Regulation of Human Cytochrome P-450 PCN3 10395
Graves, P. E., Kaminsky, L. S., and Halpert, J . (1987) Biochemistry
Greenlee, W. F., and Poland, A. (1978) J. Pharmacol. Exp. Ther.
Guengerich, F. P. (1988) Molec. Pharmacol. 33,500-508 Guengerich, F. P., Martin, M. V., Beaune, P. H., Kremers, P., Wolff,
T., and Waxman, D. J. (1986) J. Biol. Chem. 261,5051-5060 Idle, J . R., and Smith, R. L. (1979) Drug Metab. Rev. 9 , 301-317 Inaba, T., Tait, A,, Nakuno, M., Mahon, W. A., and Kalow, W. (1988)
Drug Metab. Dispos. 16 , 605-608 Kawano, S., Kamataki, T., Yasumori, T., Yamazoe, Y., and Kato, R.
(1987) J. Biochem. 102,493-501 Kitada, M., Kamataki, T., Itahashi, K., Rikihisa, T., and Kanakubo,
Y. (1987) J. Biol. Chem. 2 6 2 , 13534-13537 Kleinbloesem, C. H., van Brummelen, P., Faber, H., Danhof, M.,
Vermeulen, N. P. E., and Breimer, D. D. (1984) Biochem. Phar- macol. 33,3721-3724
Komori, M., Nishio, K., Ohi, H., Kitada, M., and Kamataki, T. (1989) J. Biochem. (Tokyo) 105,161-163
Kronbach, T., Fischer, V., and Meyer, U. A. (1988) Clin. Pharmacol. Ther. 43,630-635
Laemmli, U. K. (1970) Nature 227,680-685 Mackett, M., Smith, G. L., and Moss, B. (1984) J. Virol. 49,857-864 Molowa, D. T., Schuetz, E. G., Wrighton, S. A., Watkins, P. B.,
Kremers, P., Menolez-Picon, G., Parker, G. A,, and Guzelian, P. S. (1986) Proc. Natl. Acad. Sci. U. S. A. 8 3 , 5311-5315
Natata, K., Liberato, D. J., Gillete, J. R., and Sasame, H. S. (1986) Drug Metab. Dispos. 14 , 559-565
26,3887-3894
205,596-608
Nebert, D. W., and Gelboin, H. V. (1968) J. Biol. Chem. 2 4 3 , 6242- 6249
Nebert, D. W., and Gonzalez, F. J . (1987) Annu. Reu. Biochem. 5 6 , 945-993
Nebert, D. W., Nelson, D. R., Adesnik, M., Coon, M. J., Estabrook, R. W., Gonzalez, F. J., Guengerich, F. P., Gunsalus, I. C., Johnson, E. F., Kemper, B., Leven, W., Phillips, L. R., Sato, R., and Water- man, M. R. (1989) DNA 8,1-14
Omura, T., and Sato, R. (1964) J. Biol. Chem. 239 , 2379-2385 Saneer. F.. Nicklen. S.. and Coulsen. A. R. (1977) Proc. Natl. Acad.
Sii. U. S . A. 74, 5463-5467 ~I
~~
Skoda, R. C.. Gonzalez. F. J., Demierre. A.. and Mever. U. A. (1988) Proc. Natl.' Acad. Sci.' U. S.' A. 8 5 , 5240-5243
" I , ,
Tabor, S., and Richardson, C. C. (1987) Proc. Natl. Acad. Sci.
Towbin, H., Staehlin, T., and Gordon, J. (1979) Proc. Natl. Acad. Sci.
Tyndale, R., Inaba, T., and Kalow, W. (1989) Drug Metab. Dispos.,
U. S. A. 84,4767-4771
U. S. A. 76,4350-4354
in press Watson, C. J., and Jackson, F. J. (1985) DNA Cloning (Glover, D.
Waxman, D. J.. KO. A.. and Walsh. C. (1983) J. Biol. Chem. 2 5 8 . M., ed) Vol. 1, IRL, Washington, D. C.
11937-i1947 , . . , . ,
Waxman, D. J., Dannan, G. A., and Guengerich, F. P. (1985) Bio-
Waxman, D. J., Attisano, C., Guengerich, F. P., and Lapenson, D. P.
Wrighton, S. A,, and Vandenbranden, M. (1989) Arch. Biochem.
Young, R. A., and Davis, R. W. (1983) Proc. Natl. Acad. Sci. U. S. A.
chemistry 24,4409-4417
(1988) Arch. Biochem. Biophys. 263,424-436
Biophys. 268 , 144-151
80,1194-1198





























