Embed Size (px)
Citation preview

ELSEVIER Mutation Research 324 (1994) 13-17 Mutation Research Letters
Cytogenetic adaptive response with multiple small X-ray doses in mouse germ cells and its biological influence on the offspring
of adapted males
Lu Cai *, Ping Wang, Xiang-Gen Piao Institute of Radiation Medicine, Norman Bethune University of Medical Sciences, Changchun 130021, People's Republic of China
(Received 17 November 1993; revision received 18 January 1994; accepted 26 January 1994)
Abstract
Cytogenetic adaptive response of mouse germ cells was studied by exposing male mice to a sequence of 4 conditioning doses of 0.05 Gy each (D1) administered at 10-day intervals and subsequently to a single challenging dose of 1.5 Gy (D2). In concurrent experiments, male mice after treatment with D1 doses alone were mated to unirradiated females and the F1 males were given the D2 dose. Chromosomal aberrations in both spermatocytes and bone-marrow cells and UV-induced UDS in splenocytes of these mice were studied. Adapted mice (i.e., D1 + D2 exposures) responded with a significantly lower frequency of chromosomal aberrations than the non-adapted (D2 exposure only) controls. The relative reduction in frequencies was, however, similar to that observed in earlier work with a single conditioning dose of 0.05 Gy. The frequencies of chromosomal aberrations in spermatocytes and bone-marrow cells as well as the levels of UV-induced UDS in splenocytes of the F1 males in the group D1 to fathers + D2 to F1 males were the same as those in F1 males which received only the D2 exposure.
Key words: Adaptive response; Germ cells; Chromosome aberration; DNA synthesis, unscheduled; Low-dose radiation; Multiple exposure
I. Introduction
Since the first publication of Olivieri et al. (1984), a considerable body of evidence has accu- mulated which demonstrates that a low-dose ion- izing radiation can render cells less susceptible to
* Corresponding author, present address: Department of Pathology, The University of Western Ontario, London, Ont. N6A 5C1, Canada; Tel. 5194381084; Fax 5196613370.
a second high dose radiation. This phenomenon, termed adaptive response, has now been exten- sively documented in human lymphocytes (Wiencke et al., 1986; Shadley and Wolff, 1987; Shadley et al., 1987; Shadley and Wiencke, 1989; Wolff et al., 1988; Sankaranarayanan et al., 1989; Vijayalaxmi and Burkart, 1989; Cai and Liu, 1990; Wang et al., 1991; Shadley and Dai, 1992; Bai and Chert, 1993), rabbit lymphocytes and mouse bone-marrow cells (Cai and Liu, 1990, 1992; Liu et al., 1992), mouse splenocytes (Wojcik and
0165-7992/94/$07.00 © 1994 Elsevier Science B.V. All rights reserved SSDI 0165-7992(94)00009-A

14 L. Cai et al. /Mutat ion Research 324 (1994) 13-17
Tuschl, 1990), Chinese hamster V79 cell (Iku- shima, 1987, 1989; Osamak and Horvat, 1992), Drosophila germ cells (Fritz-Niggli and Shaeppi- Buechi, 1991), fish cells (Kurihara et al., 1993) and plant cells (Cortes et al., 1990; Rieger et al., 1992).
In some studies DNA-repair capacity was used as a measure of the adaptive response. Tuschl et al. (1983) observed an enhanced unscheduled DNA synthesis (UDS) following UV-irradiation in lymphocytes of persons occupationally exposed to low doses of ionizing radiation. Enhanced UDS levels and elevated rates of DNA-strand break repair have also been found in mouse cells ex- posed to low-dose radiation (Liu et al., 1990; Wojcik and Tuschl, 1990; Gaziev et al., 1991; James et al., 1991).
In an extension of these studies to mouse germ cells, we (Cai and Liu, 1990, 1992; Cai et al., 1993) showed that when male mice received a single conditioning dose of 0.05 Gy of X-rays followed by a large challenging dose of 1.5 Gy, (i) the frequencies of chromosomal aberrations in their germ cells (scored in their spermatocytes) were lower than those in the "non-adapted" ones and (ii) the sensitivity of F1 mice from "adapted" males to radiation-induced dominant lethality was also lower than that from non-adapted ones.
The studies reported in this paper were de- signed (i) to assess the magnitude of cytogenetic adaptive response in mouse spermatocytes ex- posed to 4 small conditioning X-ray doses (0.05 Gy each, at intervals of 10 days; D1 dose) fol- lowed by a large challenging dose (1.5 Gy; D2 dose) 40 days later and (ii) to examine whether the above conditioning dose delivered to parental males affects the DNA-repair capacity, as mea- sured by UV-induced UDS in splenocytes of F1 males irradiated with 1 Gy, or their sensitivity to the X-ray induction of chromosomal aberrations in bone-marrow cells and germ cells.
2. Materials and methods
Irradiation A Phillips therapeutic X-ray machine operated
at 200 kVp and 10 mA in the presence of 1 mm
Al and 0.5 mm Cu filter plates was used for X-irradiation. The dose rates used were 50 mGy/min for the first low-dose radiation (D1) and 440 mGy/min for the second large-dose ra- diation (D2). The D1 irradiation was given in 4 separate exposures of 0.05 Gy each with 10-day interval between exposures. The D2 dose was 1.5 Gy for the cytogenetic analysis in germ cells and bone-marrow cells and was 1.0 Gy for the analysis of splenocyte UDS.
Animals In all experiments, adult male mice (6-8-week-
old) of the Kunming strain were used. One group of mice received D1 irradiation and 40 days later, the D2 irradiation as D1 + D2 group. Another group of mice, as D2 alone group, only received the D2 irradiation at corresponding time with D1 + D2 group. Subsequently, after a colchicine (4/~g/g body weight) pre-treatment for 4 h, they were killed by decapitation and the testes were processed for cytogenetic studies. During the 40- day interval of D1 and D2 irradiation, the male mice who had just received D1 irradiation alone were mated individually to 3 virgin females. When their progeny became mature and weighed about 22-25 g, they were divided into two groups: one group received 1.5 Gy of X-rays and was used for analysis of chromosomal aberrations in their testes and bone marrow; the other group received 1 Gy of X-rays and was used in UV-induced UDS studies in their splenocytes.
Chromosome preparation Details of the methods used for cytogenetic
analysis were presented in our earlier publication (Cai and Liu, 1990). Briefly, bone-marrow cells and germ ceils were exposed to hypotonic 0.5% KC1 for 15 min and 1% sodium citrate for 30 min, respectively. All of them were fixed in an acetic acid:methanol solution (1:3). The fixed cells were then dropped onto wet slides and stained with Giemsa for 20 min. When the cytogenetic analysis was carried out, the double-blind method was used. Gaps and chromosome lesions less than the width of chromosome were not included in scoring. The Chi-square test was used for the analysis of the results.

L. Cai et al. / Mutation Research 324 (1994) 13-17 15
Analysis of UDS UDS analysis was carried out according to
methods described in Liu et al. (1990). Mouse splenocyte suspensions were made in Iscove's Modified Dulbecco Medium, e.g. IMDM, the red blood cells were lysed with Tris-buffered isotonic ammonium chloride. After washing with IMDM, the cells were finally suspended in IMDM con- taining 5% FCS and 5 x 10 -2 M hydroxyurea and the concentration of ceils was adjusted to 5 × 106 cells per ml. The cell suspensions were preincubated in a CO 2 incubator at 37°C for 30 min. The splenocyte suspension from each mouse was divided into two groups, one of these was dispensed into a small petri dish (depth of culture medium equal to or less than 1 mm) and irradi- ated with ultraviolet rays (30-W germicidal lamp, 75 cm distance, exposure time 20 sec, h = 254 nm); the other one was used as a control. After 0.5/zCi 3H-TdR in 20/zl was added to each dish, the samples were incubated under CO 2 in dark- ness at 37°C for 4 h. At the end of incubation, all samples were placed in the refrigerator for 10 min to terminate the reaction. The cells were collected on glass filters which were counted in the liquid scintillator after drying. UV-induced UDS level was calculated by subtracting the cpm values in the controls from the respective values in the UV-exposed groups.
3. Results and discussion
Table 1 presents the results on the induction of adaptive response in mouse germ cells. As can
be seen, (i) the D1 irradiation alone causes an increase in the frequency of chromosomal aberra- tions in mice and (ii) in the "adapted" mice (i.e., those which received D1 and D2), these frequen- cies are significantly lower than what would be expected if the effects of the adapting and chal- lenging doses are additive. These results thus confirmed our earlier results with a single condi- tioning dose of 0.05 Gy followed by a challenging of 1.5 Gy (Cai and Liu, 1990, 1992; Cai et al., 1993). When the relative reductions in the fre- quencies of chromosomal aberrations in the pres- ent study (with 4 x 0.05 Gy, D1 dose) are com- pared with those obtained earlier (Cai and Liu, 1990: a single D1 of 0.05 Gy), they are nearly the same namely about 29% versus 33%, respectively. This comparison suggests that fractionation of the conditioning dose of 0.2 Gy into equal doses, does not result in any higher adaptation.
Our results are thus qualitatively similar to those with human lymphocytes reported by Fan et al. (1990) who found that the frequencies of chromosomal aberrations in lymphocytes pre-ex- posed to a single 0.01 Gy versus 2 x 0.01 Gy (challenge dose: 1.5 Gy) were nearly the same. Bai and Chen (1993) however, found that when lymphocytes received 0.005 Gy twice in the same cell cycle (a single 0.005 Gy did not induce any adaptive response) the adaptive response was similar to or in excess of that observed after a single adaptive dose of 0.01 Gy. All these find- ings, including our own, point to the possible existence of a saturation mechanism for optimal adaptation.
Worth noting here are the differences in the
Table 1 Induction of the cytogenetic adaptive response by multiple low-dose X-rays in germ cells of adapted mice
Groups Numbe r of Chromatid aberrations Total Cells with
cells Breaks Exchanges chromosome chromosome scored aberrations aberrations
Control 500 0.40 + 0.84 0.00 0.40 + 0.84 0.4 + 0.84 D1 500 2.00 + 0.58 0.00 2.00 + 0.58 2.0 + 0.58 D2 500 33,40 + 1.51 1.80 _+ 1.80 34.80 + 3.49 33.20 + 3.71 D1 + D2 500 23,60 + 2.25 b,c 1.40 + 0.80 25.00 __+_ 2.16 b,c 25.00 5:1.94 b,c
(35,00) a (36.40) (34.80)
a The numbers given in parentheses are expected values, which are equal to the sum of the effects of D1 alone and D2 alone minus the control effect. b,c p < 0.01 vs. expected (b) and D2 alone (c), respectively.
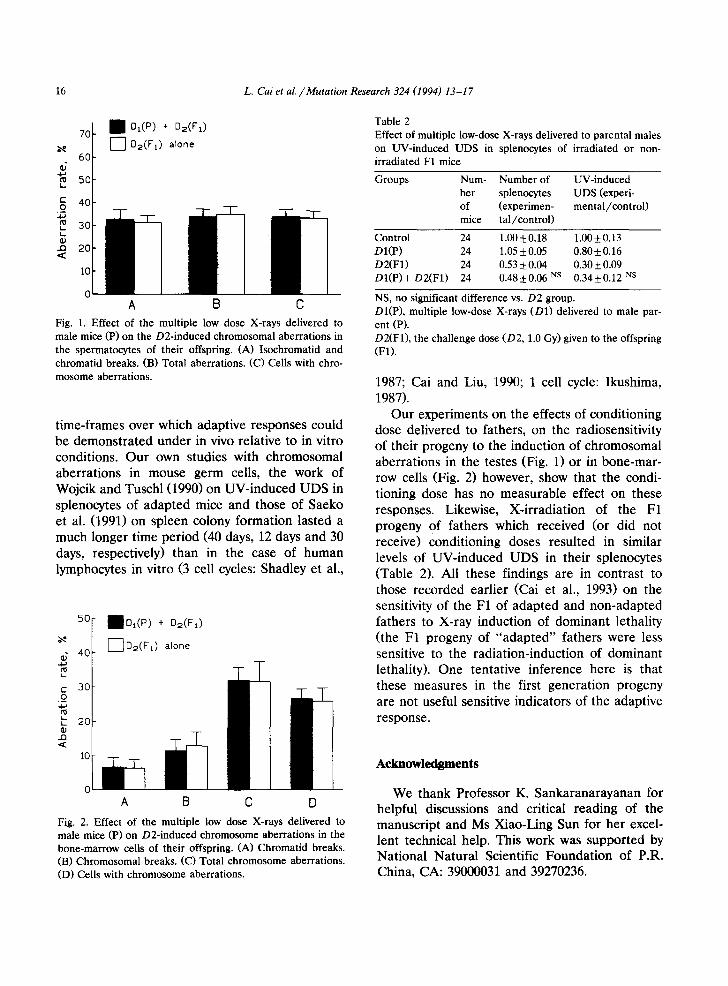
16 L. Cai et al. /Mutation Research 324 (1994) 13-17
.° f 6O
~ 5o
C 4 0 0 4J ~ 3o
.a 20
10
• Ol(P) + D2(F1)
[ ] D2(F1) alone
0 A B C
Fig. 1. Effect of the multiple low dose X-rays delivered to male mice (P) on the D2-induced chromosomal aberrations in the spermatocytes of their offspring. (A) Isochromatid and chromatid breaks. (B) Total aberrations. (C) Cells with chro- mosome aberrations.
time-frames over which adaptive responses could be demonstrated under in vivo relative to in vitro conditions. Our own studies with chromosomal aberrations in mouse germ cells, the work of Wojcik and Tuschl (1990) on UV-induced UDS in splenocytes of adapted mice and those of Saeko et al. (1991) on spleen colony formation lasted a much longer time period (40 days, 12 days and 30 days, respectively) than in the case of human lymphocytes in vitro (3 cell cycles: Shadley et al.,
- 40
C 30 o
~. 2(? .13 .<
10
1 D l ( P ) + D2(F1)
[ ~ D2(F1) alone
Table 2 Effect of multiple low-dose X-rays delivered to parental males on UV-induced UDS in splenocytes of irradiated or non- irradiated F1 mice
Groups Num- Number of UV-induced ber splenocytes UDS (experi- of (experimen- mental/control) mice tal/control)
Control 24 1.005:0.18 1.00_+0.13 DI(P) 24 1.055:0.05 0.80+0.16 D2(F1) 24 0.53 5:0.04 0.30 _+ 0.09 DI(P)+ D2(F1) 24 0.485:0.06 NS 0.34+0.12 NS
NS, no significant difference vs. D2 group. DI(P), multiple low-dose X-rays (D1) delivered to male par- ent (P). D2(F1), the challenge dose (D2, 1.0 Gy) given to the offspring (F1).
1987; Cai and Liu, 1990; 1 cell cycle: Ikushima, 1987).
Our experiments on the effects of conditioning dose delivered to fathers, on the radiosensitivity of their progeny to the induction of chromosomal aberrations in the testes (Fig. 1) or in bone-mar- row cells (Fig. 2) however, show that the condi- tioning dose has no measurable effect on these responses. Likewise, X-irradiation of the F1 progeny of fathers which received (or did not receive) conditioning doses resulted in similar levels of UV-induced UDS in their splenocytes (Table 2). All these findings are in contrast to those recorded earlier (Cai et al., 1993) on the sensitivity of the F1 of adapted and non-adapted fathers to X-ray induction of dominant lethality (the F1 progeny of "adapted" fathers were less sensitive to the radiation-induction of dominant lethality). One tentative inference here is that these measures in the first generation progeny are not useful sensitive indicators of the adaptive response.
Acknowledgments
0
A B C D
Fig. 2. Effect of the multiple low dose X-rays delivered to male mice (P) on D2-induced chromosome aberrations in the bone-marrow cells of their offspring. (A) Chromatid breaks. (B) Chromosomal breaks. (C) Total chromosome aberrations. (D) Cells with chromosome aberrations.
We thank Professor K. Sankaranarayanan for helpful discussions and critical reading of the manuscript and Ms Xiao-Ling Sun for her excel- lent technical help. This work was supported by National Natural Scientific Foundation of P.R. China, CA: 39000031 and 39270236.

L. Cai et al. /Mutation Research 324 (1994) 13-17 17
References
Bai, Y.L., and D.Q. Chen (1993) Accumulative effect of two low doses of irradiation in inducing an adaptive response in human lymphocytes, Mutation Res., 302, 191-196.
Cai, L., and S.Z. Liu (1990) Induction of cytogenetic adaptive response of somatic and germ cells in vivo and in vitro by low-dose X-irradiation, Int. J. Radiat. Biol., 58, 187-194.
Cai, L., and S.Z. Liu (1992) Induction of cytogenetic adaptive response in somatic and germ cells by low-level exposure in vitro and in vivo to X- or y-irradiation, in: T. Sugahara, L.A. Sagan and T. Aoyama (Eds.), Low Dose Irradiation and Biological Defense Mechanisms, Elsevier, Amster- dam, pp. 287-290.
Cai, L., J. Jiang, X.-L. Wang and B. Wang (1993) Induction of an adaptive response to dominant lethality and to chromo- some damage of mouse germ cells by low dose radiation, Mutation Res., 303, 157-161.
Cortes, F., I. Dominguez, S. Mateos, J. Pinero and J. Mateos (1990) Evidence for an adaptive response to radiation damage in plant cells conditioned with X-rays or incorpo- rated tritium, Int. J. Radiat. Biol., 56, 537-541.
Fan, S., Vijayalaxmi, G. Mindek and W. Burkart (1990) Adap- tive response to 2 low doses of X-rays in human blood lymphocytes, Mutation Res., 243, 53-56.
Fritz-Niggli, H., and C. Shaeppi-Buechi (1991) Adaptive re- sponse to dominant lethality of mature (Class A) and immature (class B) oocyte of D. melanogaster to low doses of ionizing radiation: Effects in repair-proficient (yw) and repair-deficient strains (mei 41 °5 and mus 30261), Int. J. Radiat. Biol., 59, 175-184.
Gaziev, A.I., L.A. Formenko, Ja.K. Kozhanovskaya and N.P. Sirota (1991) Adaptive repair response in the tissue of mice exposed to chronic gamma irradiation, in: J.D. Chapma, W.C. Dewey and G.F, Whitmore (Eds.), Radia- tion Research, A Twentieth-Century Perspective, Vol. I, Academic Press, San Diego, [Abstract].
Ikushima, T. (1987) Chromosomal response to ionizing radia- tion reminiscent of an adaptive response in cultured Chi- nese hamster cells, Mutation Res., 180, 215-221.
Ikushima, T. (1989) Radio-adaptive response: Characteriza- tion of a cytogenetic repair induced by low-level ionizing radiation in cultured Chinese hamster cells, Mutation Res., 227, 241-246.
James, S.J., S.M. Enger and T. Makinodan (1991) DNA strand breaks and DNA repair response in lymphocytes after chronic in vivo exposure to very low doses of ionizing radiation in mice, Mutation Res., 249, 255-263.
Kurihara, Y., M. Rienkjkarn and H. Etoh (1993) Cytogenetic adaptive response to low-dose of X-rays in cultured fish cells, J. Radiat. Res., in press.
Liu, S.Z., L. Cai and J.B. Sun (1990) Effect of low-dose radiation on repair of DNA and chromosome damage, Acta Biol. Hung., 41, 149-157.
Liu, S.Z., L. Cai and S.Q. Sun (1992) Induction of a cytoge- netic adaptive response by exposure of rabbits to very low dose-rate gamma radiation, Int. J. Radiat. Biol., 62, 187- 190.
Olivieri, G., J. Bodycote and S. Wolff (1984) Adaptive re- sponse of human lymphocytes to low concentrations of radioactive thymidine, Science, 223, 594-597.
Osamak, M., and D. Horvat (1992) Chromosomal analysis of Chinese hamster V79 ceils exposed to multiple y-rays fraction: induction of adaptive response to mitomycin C, Mutation Res., 282, 259-263.
Rieger, R., A. Michaelis and S. Takehisa (1992) Low tempera- ture between conditioning and challenge treatment pre- vents the "adaptive response" of Vicia faba root tip meristem cells, Mutation Res., 282, 69-72.
Saenko, A.S., T.N. Semenets and O.V. Semina (1991) The enhanced radioresistance (adaptive response) in vivo of splenic colony-forming units (CFU-S) following the expo- sure of mice to 6°Co gamma rays at low doses, Radiobi- ologiia, 31, 716-717 (in Russian).
Sankaranarayanan, K., A.-V. Duyn, M.J. Loos and A.T. Natarajan (1989) Adaptive response of human lympho- cytes to low-level radiation from radioisotopes or X-rays, Mutation Res., 211, 7-12.
Shadley, J.D., and G.Q. Dai (1992) Cytogenetic and survival adaptive response in G1 phase human lymphocytes, Muta- tion Res., 265, 273-281.
Shadley, J.D., and J.K. Wiencke (1989) Induction of the adaptive response by X-rays is dependent on radiation intensity, Int. J. Radiat. Biol., 56, 107-118.
Shadley, J.D., and S. Wolff (1987) Very low doses of X-rays can cause human lymphocytes to become less susceptible to ionizing radiation, Mutagenesis, 2, 95-96.
Shadley, J.D., V. Afzal and S. Wolff (1987) Characterization of the adaptive response to ionizing radiation induced by low dose of X-rays to human lymphocytes, Radiat. Res., 111,511-517.
Tuschl, H., R. Kovac and H. Altmann (1983) UDS and SCE in lymphocytes of persons occupationally exposed to low lev- els of ionizing radiation, Health Physics, 45, 1-7.
Vijayalaxmi and W. Barkart (1989) Resistance and cross-resis- tance to chromosome damage in human blood lympho- cytes adapted to bleomycin, Mutation Res., 211, 1-5.
Wang, Z.-Q., S. Saigusa and M.S. Sasaki (1991) Adaptive response to chromosome damage in cultured human lym- phocytes primed with low doses of X-rays, Mutation Res., 246, 179-186,
Wiencke, J.K., V. Afzal, G. Olivieri and S. Wolff (1986) Evidence that the [3H]thymidine-induced adaptive re- sponse of human lymphocytes to subsequent doses of X-rays involves the induction of a chromosomal repair mechanism, Mutagenesis, 1, 375-380.
Wojcik, A., and H. Tuschl (1990) Indications of an adaptive response in C57BL mice pre-exposed in vivo to low doses of ionizing radiation, Mutation Res., 243, 67-73.
Wolff, S., V. Afzal, J.K. Wiencke, G. Olivieri and A. Michael (1988) Human lymphocytes exposed to low dose ionizing radiations became refractory to high doses of radiation as well as to chemical mutagens that induce double-strand breaks in DNA, Int. J. Radiat. Biol., 53, 49-55.





























