Embed Size (px)
Citation preview

University of the Pacific University of the Pacific
Scholarly Commons Scholarly Commons
All Dugoni School of Dentistry Faculty Articles All Faculty Scholarship
6-1-1991
Cytolytic and ion channel forming properties of the N-terminus of Cytolytic and ion channel forming properties of the N-terminus of
lymphocyte perforin lymphocyte perforin
David M. Ojcius University of the Pacific, [email protected]
Pedro M. Persechini Universidade Federal do Rio de Janeiro, [email protected]
Li-Mou Zheng The Rockefeller University
Paulo C. Notaroberto Universidade Federal do Rio de Janeiro
Sandro C. Adeodato Universidade Federal do Rio de Janeiro
See next page for additional authors
Follow this and additional works at: https://scholarlycommons.pacific.edu/dugoni-facarticles
Part of the Biochemistry Commons, Immunity Commons, Immunology of Infectious Disease
Commons, and the Medical Immunology Commons
Recommended Citation Recommended Citation Ojcius, D. M., Persechini, P. M., Zheng, L., Notaroberto, P. C., Adeodato, S. C., & Young, J. D. (1991). Cytolytic and ion channel forming properties of the N-terminus of lymphocyte perforin. Proceedings of the National Academy of Sciences of the United States of America, 88(11), 4621–4625. https://scholarlycommons.pacific.edu/dugoni-facarticles/133
This Article is brought to you for free and open access by the All Faculty Scholarship at Scholarly Commons. It has been accepted for inclusion in All Dugoni School of Dentistry Faculty Articles by an authorized administrator of Scholarly Commons. For more information, please contact [email protected].

Authors Authors David M. Ojcius, Pedro M. Persechini, Li-Mou Zheng, Paulo C. Notaroberto, Sandro C. Adeodato, and John Ding-E. Young
This article is available at Scholarly Commons: https://scholarlycommons.pacific.edu/dugoni-facarticles/133

Proc. Nail. Acad. Sci. USAVol. 88, pp. 4621-4625, June 1991Immunology
Cytolytic and ion channel-forming properties of the N terminus oflymphocyte perforinDAVID M. OJCIUS*, PEDRO M. PERSECHINIt, Li MOU ZHENG*, PAULO C. NOTAROBERTOt,SANDRO C. ADEODATOt, AND JOHN DING-E YOUNG**Laboratory of Cellular Physiology and Immunology, The Rockefeller University, New York, NY 10021; and tInstituto Biofisica, Universidade Federal do Riode Janeiro, 21941 Rio de Janeiro, Brazil
Communicated by Zanvil A. Cohn, March 4, 1991
ABSTRACT Perforin lyses cells by binding to the targetcell membrane, where it polymerizes into large nonspecificpores. It is shown here that the first 34 amino acids of theN-terminal region of either human or murine perforin aresoluble in aqueous medium and spontaneously insert intomembranes. The N-terminal peptides lyse liposomes and nu-cleated cells, and they form ion channels in planar bilayers,some ofwhich are comparable to those previously described forperforin. The lytic activity of the N-terminal domains does notrequire calcium, is independent of the lipid headgroup com-position, and can be inhibited by heparin. Tumor cells incu-bated with the N-terminal peptides undergo the same morpho-logical changes as those induced by native perforin. None of thepeptides corresponding to the putative membrane-spanningdomains from the central region of perforin is cytolytic. Takentogether, these results suggest that the N-terminal region is animportant part of the pore-forming domain of perforin.
Cytotoxic T lymphocytes and natural killer cells endow thebody with an effective barrier against cancerous and virallyinfected cells. These effector lymphocytes have been shownto produce a 70-kDa pore-forming protein, called perforin orcytolysin, that binds to target cell (TC) membranes, where itpolymerizes into barrel-stave pores with internal diameters ofup to 20 nm (1-5). Purified perforin has potent lytic activity,and it can lyse a wide variety of targets, including erythro-cytes, nucleated cells, and artificial lipid vesicles. In lipidbilayers, perforin forms voltage-insensitive and ion-non-selective channels that remain preferentially in the openstate. The channels are heterogeneous in size, averaging 400pS/unit in 0.1 M NaCl (6).The central one-third of the perforin amino acid sequence
displays extensive homology with the amino acid sequencesof C6, C7, C8, and C9, the terminal components of themembrane attack complex (MAC) of complement, except atthe C and N terminii, which are specific to perforin (7).Because of its homology with the putative membrane-spanning domain of the MAC components, the central regionof perforin was thought to contain the perforin lytic domain.To identify the perforin lytic domain, synthetic peptides weretherefore made based on the sequence of four regions fromthe central region predicted to have amphiphilic structures.However, unlike monomeric C9, which binds to a preassem-bled C5b-C6-C7-C8 complex on the TC membrane, mono-meric perforin binds to membrane lipids in the absence ofanyprotein receptor. Thus, the perforin-specific peripheries werealso investigated for a domain that might be directly lytic tomembranes. In this work, a highly lytic domain was in factlocated in the N terminus of perforin.
EXPERIMENTAL PROCEDURESCells, Antibodies, Perforin Purification, and Synthesis of
Peptides. The murine cytolytic cell lines CTLL-R8 andCTLL-2 and the murine tumor cell lines P815, Yac-1, andEL-4 were maintained as described (8). Perforin was purifiedfrom CTLL-R8 cells as described (8). One hemolytic unit(HU) is defined as the amount of perforin required to lyse 107erythrocytes in 140 gl (8).
Peptides were synthesized at the Sloan-Kettering Micro-chemistry Core Facility on an Applied Biosystems peptidesynthesizer following standard procedures. The peptideswere purified to homogeneity by HPLC, after which theywere lyophilized and analyzed for amino acid content.
Preparation of Lipid Vesicles and Measurement of Carboxy-fluorescein (CF) Release. Phosphatidylcholine (PC) large unil-amellar vesicles (LUVs) were made with 50% PC (type IIS;Sigma) and 50o cholesterol (Calbiochem-Behring) by thereverse evaporation method (9) in the presence of 100 mMCF, 50 mM NaCl, and 10 mM Hepes (pH 7.4). External CFwas removed on a Sephadex G-50 column using 150 mMNaCl and 10 mM Hepes (pH 7.4). CF release from 1-2%(wt/vol) LUVs was measured in an SLM Aminco (Urbana,IL) 500C spectrofluorometer at an excitation wavelength of490 nm and an emission wavelength of525 nm (21-240C). Forcalibration, 100%o lysis was determined by diluting a 1%Triton X-100 solution 50-fold into the cuvette. Phosphatidyl-serine (PS) LUVs were made as described above, except thatPC was replaced by PS (Avanti Polar Lipids).
Cell Lysis Assays. Perforin- or peptide-mediated lysis ofnucleated cells was detected by the 5"Cr-release assay (8).Spontaneous release was considered as the 51Cr release fromcells incubated with medium alone or, in the case of compe-tition assays with liposomes, from cells incubated in mediumwith the liposomes but without peptide. The radioactivityfrom cells lysed with 1% SDS was used as the total release,and 51Cr release was calculated as % specific release =(experimental release - spontaneous release/total release -spontaneous release) x 100.The competition assay was used for comparing the ability
of liposomes to absorb peptide from a reaction mediumcontaining peptide and 51Cr-labeled nucleated cells. Indi-cated concentrations ofPC or PS LUVs were resuspended intriplicate in 100 ,ul ofphosphate-buffered saline (PBS, pH 7.4)containing 14 ,uM human N-terminal peptide (HuNP) andmixed in microtiter wells with 100 p.1 of 51Cr-labeled cells (5x 105 cells per ml). Percent specific inhibition was calculatedas (1 - % specific release in experiment/% specific releasein control) x 100. For the control, PBS without liposomeswas mixed with peptide and 51Cr-labeled cells.
Abbreviations: LUV, large unilamellar vesicle; HU, hemolyticunit(s); CF, carboxyfluorescein; MAC, membrane attack complex;PS, phosphatidylserine; PC, phosphatidylcholine; TC, target cell;HuNP, human N-terminal peptide; MuNP, murine N-terminal pep-tide; CP, C-terminal peptide.
4621
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Proc. Nati. Acad. Sci. USA 88 (1991)
Perforin-Mediated Hemolysis. Purified perforin in PBS wasadded, 1304ul per well, in triplicates to 96-well plates; 10 /ulof erythrocyte suspension (109 cells per ml) and 10 /4 of 20mM CaCl2 were then added sequentially. For experimentswhere the effects of inhibitors were assayed, 120 /4 of PBSper well contained the indicated final concentration of hep-arin or antibodies. Hemolysis was determined spectropho-tometrically as described (10). The effects of heparin orantibodies on the membrane binding and insertion activitiesof perforin were dissociated following a previously publishedprotocol (10, 11). Polyclonal antisera (IgG fraction) againstperforin (12) and against a peptide consisting of amino acids1-19 of the human perforin sequence (13) were alreadydescribed. Nonspecific IgG was used as a negative control.
Electron Microscopy. After treatment with murine N-ter-minal peptide (MuNP) or HuNP, cells were fixed by mixingwith the same volume of 4% glutaraldehyde, 1% paraform-aldehyde, and 0.1 M phosphate (pH 7.2) at 4TC for 2 hr. Afterrinsing, the cell pellets were postfixed with 2% OS04 in thesame buffer for 30 min. The pellets were dehydrated withethanol and propylene oxide and embedded in Epon 812. Forcomparison, P815 cells were incubated for 1 hr with 33 HUof perforin per ml.
Planar Bilayers and Current Measurements. Lipid bilayermembranes were formed by the painting technique (14) usingeither a mixture of 30% phosphatidylethanolamine/PC/PS(10:3:3) in decane or 15% asolecithin (type II; Sigma) indecane. The experimental chamber was divided into two2.5-ml compartments as described (15). The cis side con-tained 150 mM NaCl, 1 mM MgCl2, 1 mM CaCl2, and 10 mMHepes (pH 7.4). The trans side was held at virtual ground andcontained 5 mM NaCl, 150 mM KCl, 1 mM MgCl2, 1 mMEGTA, and 10 mM Hepes (pH 7.4). Peptides were added tothe cis side.
RESULTSSelection of Peptides. Seven synthetic peptides were made
based on the published sequence of murine and humanperforin (16-19). As a Kyte-Doolittle hydropathic plot pre-dicts that four perforin regions could form amphiphilic do-mains (Fig. 1), peptides were made corresponding to murineamino acids 189-218 (a,), 237-266 (13i), 402-422 (a2), and104-132 (132). a, would be amphiphilic if in an a-helicalconfiguration and was initially assumed to be a good candi-date for membrane spanning/pore-forming domain; compa-rable amphipathic stretches are present in all of the MACcomponents. Although only partially conserved in C8 and C9and not at all in C7, t3i would be very amphiphilic if in the,8-sheet configuration and contains at the C end a cluster of
+ ++ + - +a1 HkHRIUSSY kTHflTVDL GGRISYIT&L
+ --+++ ++
Pi JNVEMQVSIG Q,&SVSSEXK ACEEKKKQHK
+- + + +
P2 INNDHRYPLD YNrR.EEAR &SVAGSHSK
+
02 RAEHLWGDT TATDAYULKVF F
+- + ++MUNP T=TRSEf KQKHKFVPGV WMAGEgT TJRR
+ +- +++- ++HuNP £QHTAARSE9 KRSHKEVPGA LAGEGVDVT SLRR
CP YAjQGLLDj PGNRSGAVW
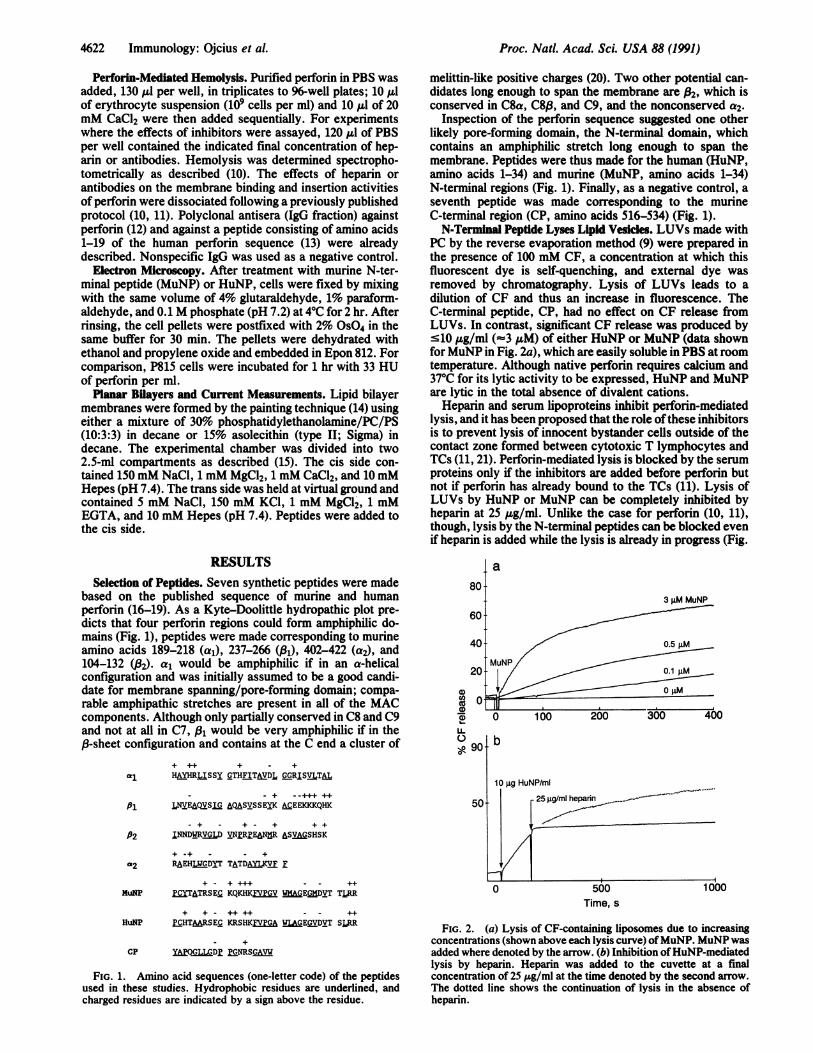
FIG. 1. Amino acid sequences (one-letter code) of the peptidesused in these studies. Hydrophobic residues are underlined, andcharged residues are indicated by a sign above the residue.
melittin-like positive charges (20). Two other potential can-didates long enough to span the membrane are 2, which isconserved in C8a, C8.8, and C9, and the nonconserved a2.
Inspection of the perforin sequence suggested one otherlikely pore-forming domain, the N-terminal domain, whichcontains an amphiphilic stretch long enough to span themembrane. Peptides were thus made for the human (HuNP,amino acids 1-34) and murine (MuNP, amino acids 1-34)N-terminal regions (Fig. 1). Finally, as a negative control, aseventh peptide was made corresponding to the murineC-terminal region (CP, amino acids 516-534) (Fig. 1).N-Terminal Peptide Lyses Lipid Vesides. LUVs made with
PC by the reverse evaporation method (9) were prepared inthe presence of 100 mM CF, a concentration at which thisfluorescent dye is self-quenching, and external dye wasremoved by chromatography. Lysis of LUVs leads to adilution of CF and thus an increase in fluorescence. TheC-terminal peptide, CP, had no effect on CF release fromLUVs. In contrast, significant CF release was produced by'10 pg/ml (%3 pAM) of either HuNP or MuNP (data shownforMuNP in Fig. 2a), which are easily soluble in PBS at roomtemperature. Although native perforin requires calcium and37TC for its lytic activity to be expressed, HuNP and MuNPare lytic in the total absence of divalent cations.Heparin and serum lipoproteins inhibit perforin-mediated
lysis, and it has been proposed that the role ofthese inhibitorsis to prevent lysis of innocent bystander cells outside of thecontact zone formed between cytotoxic T lymphocytes andTCs (11, 21). Perforin-mediated lysis is blocked by the serumproteins only if the inhibitors are added before perforin butnot if perforin has already bound to the TCs (11). Lysis ofLUVs by HuNP or MuNP can be completely inhibited byheparin at 25 pug/ml. Unlike the case for perforin (10, 11),though, lysis by the N-terminal peptides can be blocked evenif heparin is added while the lysis is already in progress (Fig.
a80 .
61
41
21
a)co
0
3 ILM MuNP
o0 0.5 IiM.MuNP
!0 | /0.1 PMif. ~~~~~~0LM
v I 1110 100 200 300 400
b
10 9g HuNP/ml
25 ±g/mI hepannS
/~~~~~~~~~~~. . ...
=IFl0
Time, s
FIG. 2. (a) Lysis of CF-containing liposomes due to increasingconcentrations (shown above each lysis curve) ofMuNP. MuNP wasadded where denoted by the arrow. (b) Inhibition ofHuNP-mediatedlysis by heparin. Heparin was added to the cuvette at a finalconcentration of 25 ,ug/ml at the time denoted by the second arrow.The dotted line shows the continuation of lysis in the absence ofheparin.
4622 Immunology: Ojcius et al.
50i
500 1 000
Proc. Natl. Acad. Sci. USA 88 (1991) 4623
2b). Heparin has no effect on lysis of liposomes due to thelytic bee venom toxin melittin at 10 Ag/ml (data not shown).As a first step toward defining the minimum length for lytic
activity of the N-terminal domain, a peptide was madecorresponding to the first 19 amino acids of HuNP. The lyticactivity of this peptide was comparable to that ofHuNP (datanot shown), implying that the remainder of the sequence ofHuNP or MuNP may not be required for lysis of LUVs.The peptides corresponding to the perforin regions dis-
playing homologies with the MAC components were alsoinvestigated. a1, f1, a2, and P2 were dissolved at 5 mg/ml inan 8 M urea buffer, and their ability to lyse LUVs was tested;the urea control (80 mM urea) was not lytic. Neither a2 nor
/32 (50 Ag/ml) had any noticeable effect on CF release fromLUVs. On the other hand, relatively high concentrations(250 Ag/ml) of a1 or Pf gave rise to small but reproducible CFrelease from LUVs, suggesting that, though only slightly lyticto liposomes and inoffensive to nucleated cells (see below),these perforin domains may participate with the N-terminaldomain in subsequent perforin insertion and polymerization.N-Terminal Peptide Lyses Tumor Cells and Cytolytic Lym-
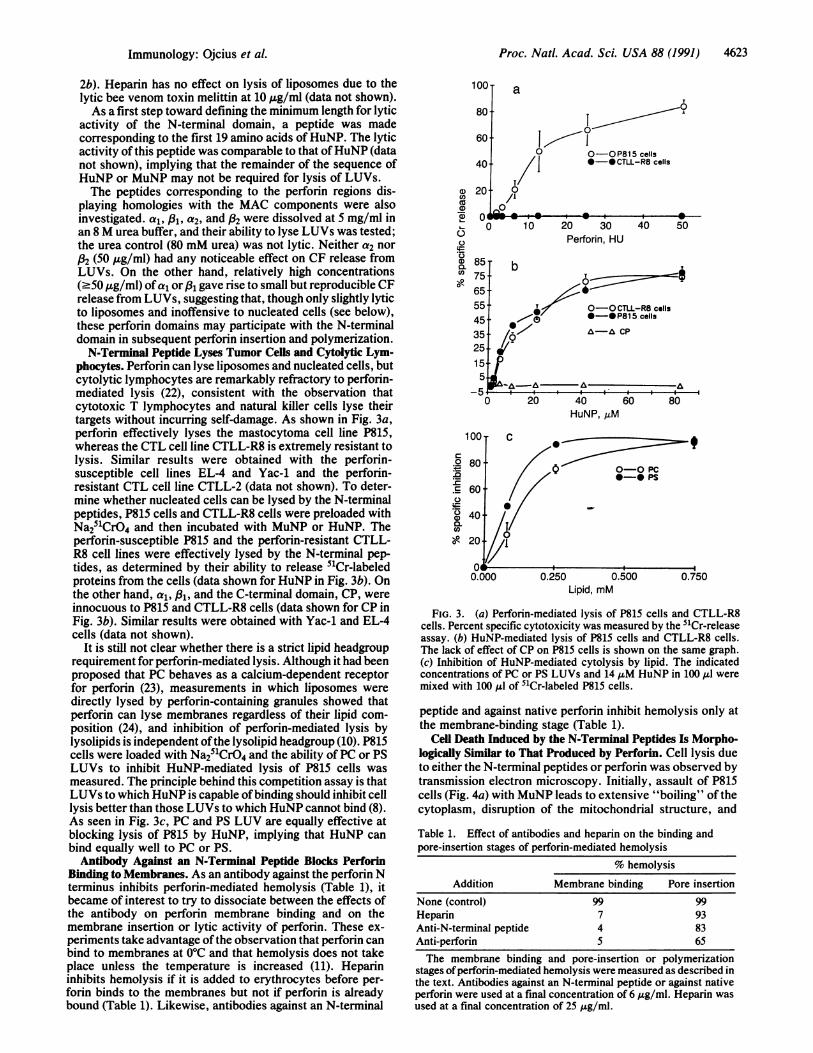
phocytes. Perforin can lyse liposomes and nucleated cells, butcytolytic lymphocytes are remarkably refractory to perforin-mediated lysis (22), consistent with the observation thatcytotoxic T lymphocytes and natural killer cells lyse theirtargets without incurring self-damage. As shown in Fig. 3a,perforin effectively lyses the mastocytoma cell line P815,whereas the CTL cell line CTLL-R8 is extremely resistant tolysis. Similar results were obtained with the perforin-susceptible cell lines EL-4 and Yac-1 and the perforin-resistant CTL cell line CTLL-2 (data not shown). To deter-mine whether nucleated cells can be lysed by the N-terminalpeptides, P815 cells and CTLL-R8 cells were preloaded withNa251CrO4 and then incubated with MuNP or HuNP. Theperforin-susceptible P815 and the perforin-resistant CTLL-R8 cell lines were effectively lysed by the N-terminal pep-tides, as determined by their ability to release 51Cr-labeledproteins from the cells (data shown for HuNP in Fig. 3b). Onthe other hand, a1, 831, and the C-terminal domain, CP, wereinnocuous to P815 and CTLL-R8 cells (data shown for CP inFig. 3b). Similar results were obtained with Yac-1 and EL-4cells (data not shown).
It is still not clear whether there is a strict lipid headgrouprequirement for perforin-mediated lysis. Although it had beenproposed that PC behaves as a calcium-dependent receptorfor perforin (23), measurements in which liposomes weredirectly lysed by perforin-containing granules showed thatperforin can lyse membranes regardless of their lipid com-position (24), and inhibition of perforin-mediated lysis bylysolipids is independent ofthe lysolipid headgroup (10). P815cells were loaded with Na251CrO4 and the ability of PC or PSLUVs to inhibit HuNP-mediated lysis of P815 cells wasmeasured. The principle behind this competition assay is thatLUVs to which HuNP is capable ofbinding should inhibit celllysis better than those LUVs to which HuNP cannot bind (8).As seen in Fig. 3c, PC and PS LUV are equally effective atblocking lysis of P815 by HuNP, implying that HuNP canbind equally well to PC or PS.Antibody Against an N-Terminal Peptide Blocks Perforin
Binding to Membranes. As an antibody against the perforin Nterminus inhibits perforin-mediated hemolysis (Table 1), itbecame of interest to try to dissociate between the effects ofthe antibody on perforin membrane binding and on themembrane insertion or lytic activity of perforin. These ex-periments take advantage ofthe observation that perforin canbind to membranes at 0°C and that hemolysis does not takeplace unless the temperature is increased (11). Heparininhibits hemolysis if it is added to erythrocytes before per-forin binds to the membranes but not if perforin is alreadybound (Table 1). Likewise, antibodies against an N-terminal
01)CO)
a)0)
C.)
CO)OR
1oo0- aT
80
60 ul° O-OP815 cells
40 **-@CTLL-R8 cells
20 °0/I
00 1 p--100 10 20 30 40 50
Perforin, HU
85-
75.
65-
55.
45.
8575
155
T{ o-OCTLL-R8 cells0 0-0*-P815 cells
//0"."A-A CID
-&/& a A
0 20 40 60 80HuNP, MM
0
:5
.r-C)C.)E0)COIOR
0-0 Pc*0- PS
0.000 0.250 0.750
Lipid, mM
FIG. 3. (a) Perforin-mediated lysis of P815 cells and CTLL-R8cells. Percent specific cytotoxicity was measured by the 51Cr-releaseassay. (b) HuNP-mediated lysis of P815 cells and CTLL-R8 cells.The lack of effect of CP on P815 cells is shown on the same graph.(c) Inhibition of HuNP-mediated cytolysis by lipid. The indicatedconcentrations of PC or PS LUVs and 14 MAM HuNP in 100 ,ul weremixed with 100 MAl of 51Cr-labeled P815 cells.
peptide and against native perforin inhibit hemolysis only atthe membrane-binding stage (Table 1).
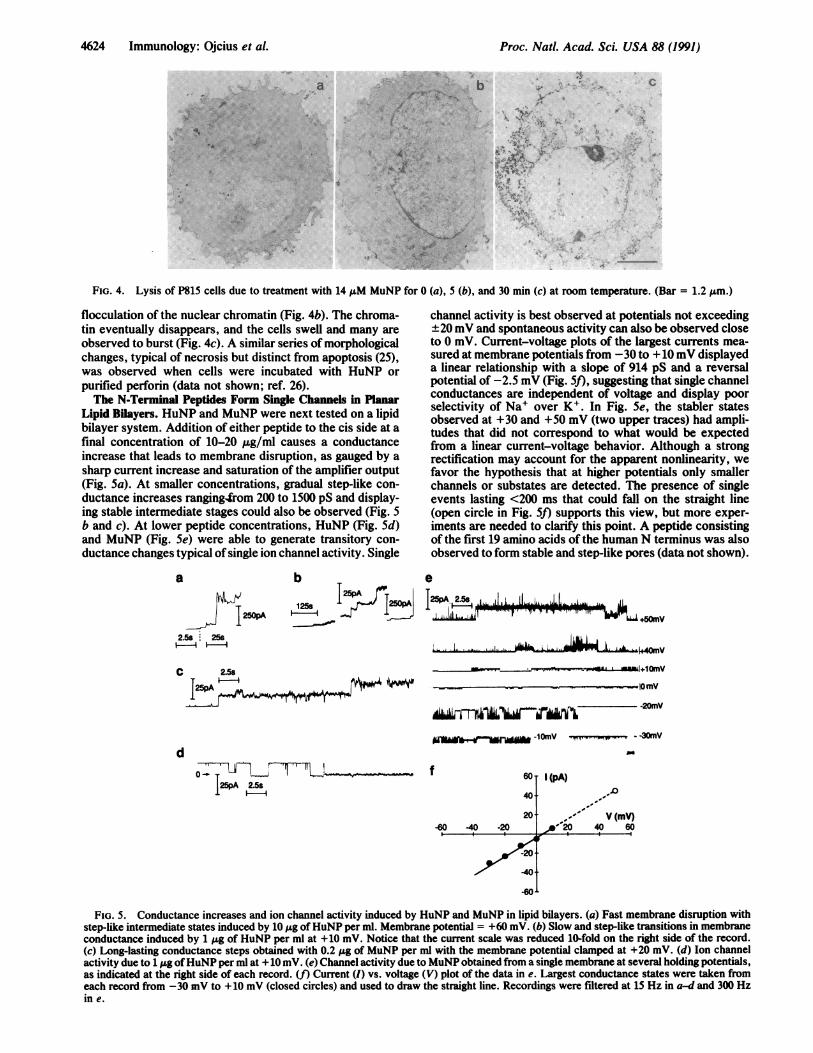
Cell Death Induced by the N-Terminal Peptides Is Morpho-logically Similar to That Produced by Perforin. Cell lysis dueto either the N-terminal peptides or perforin was observed bytransmission electron microscopy. Initially, assault of P815cells (Fig. 4a) with MuNP leads to extensive "boiling" of thecytoplasm, disruption of the mitochondrial structure, and
Table 1. Effect of antibodies and heparin on the binding andpore-insertion stages of perforin-mediated hemolysis
% hemolysisAddition Membrane binding Pore insertion
None (control) 99 99Heparin 7 93Anti-N-terminal peptide 4 83Anti-perforin 5 65
The membrane binding and pore-insertion or polymerizationstages ofperforin-mediated hemolysis were measured as described inthe text. Antibodies against an N-terminal peptide or against nativeperforin were used at a final concentration of 6 ,g/ml. Heparin wasused at a final concentration of 25 ug/ml.
Immunology: Ojcius et al.
Proc. Natl. Acad. Sci. USA 88 (1991)
ae - > , x ' .w . ^ 2? ,.; . ss A... ... . t . e sty }
4s}<x ->8,, .;:r.i : . u,, .X: ::::: : ......... : : wi s<^w". : :' . .. ,.' s e_alr UE :, ..
^ X i;' .,,^S , i,+.? ,. .. ..
........ 'a _.,.;., ..
.- X '.e ^ 6-AS. . ,.,
* *A* ..
:4!'.' i f2 a
Fl
:U A. I.'.4
s A, . +
A. -,:
:'..::..wo.
.:r Si.>
to <, F. W:.
I
C. . is S
... b.- ..i .t s o--t>S S
DSe . ..s . s
a .C /.1, .... . He dL ,-Boss
he .'. i
ant., an:
t $>;te i:. a:x t #
.e ;. < ..A.. .. -:. s ' ...> Ok * .*AbLS't'
a> A.,
\Off *
.8 2 '' $ Al'. I:'
,;, 0, ..
FIG. 4. Lysis of P815 cells due to treatment with 14NM MuNP for 0 (a), 5 (b), and 30 min (c) at room temperature. (Bar = 1.2 Am.)
flocculation of the nuclear chromatin (Fig. 4b). The chroma-tin eventually disappears, and the cells swell and many areobserved to burst (Fig. 4c). A similar series of morphologicalchanges, typical of necrosis but distinct from apoptosis (25),was observed when cells were incubated with HuNP orpurified perforin (data not shown; ref. 26).The N-Terminal Peptides Form Single Channels in Planar
Lipid Bilayers. HuNP and MuNP were next tested on a lipidbilayer system. Addition of either peptide to the cis side at afinal concentration of 10-20 jig/ml causes a conductanceincrease that leads to membrane disruption, as gauged by asharp current increase and saturation of the amplifier output(Fig. 5a). At smaller concentrations, gradual step-like con-ductance increases ranging4rom 200 to 1500 pS and display-ing stable intermediate stages could also be observed (Fig. 5b and c). At lower peptide concentrations, HuNP (Fig. Sd)and MuNP (Fig. 5e) were able to generate transitory con-ductance changes typical of single ion channel activity. Single
a
I 250PA2.5 25s
i
b
125s-
channel activity is best observed at potentials not exceeding±20mV and spontaneous activity can also be observed closeto 0 mV. Current-voltage plots of the largest currents mea-sured at membrane potentials from -30 to +10 mV displayeda linear relationship with a slope of 914 pS and a reversalpotential of -2.5 mV (Fig. 5f), suggesting that single channelconductances are independent of voltage and display poorselectivity of Na' over K+. In Fig. 5e, the stabler statesobserved at +30 and +50 mV (two upper traces) had ampli-tudes that did not correspond to what would be expectedfrom a linear current-voltage behavior. Although a strongrectification may account for the apparent nonlinearity, wefavor the hypothesis that at higher potentials only smallerchannels or substates are detected. The presence of singleevents lasting <200 ms that could fall on the straight line(open circle in Fig. 5J) supports this view, but more exper-iments are needed to clarify this point. A peptide consistingof the first 19 amino acids of the human N terminus was alsoobserved to form stable and step-like pores (data not shown).
0
12W.AiL5I _filL-.1 -," I
25pA 15P
C 2.5s
i 4w_
d
-20mV
lmrwd -1OmV . - -3OmV
60T I(pA)
40f201
-60 -40 -20
.20
40
V (mV)A-20 40 60
.60J
FIG. 5. Conductance increases and ion channel activity induced by HuNP and MuNP in lipid bilayers. (a) Fast membrane disruption withstep-like intermediate states induced by 10 jig ofHuNP per ml. Membrane potential = +60 mV. (b) Slow and step-like transitions in membraneconductance induced by 1 Ag of HuNP per ml at +10 mV. Notice that the current scale was reduced 10-fold on the right side of the record.(c) Long-lasting conductance steps obtained with 0.2 jig of MuNP per ml with the membrane potential clamped at +20 mV. (d) Ion channelactivity due to 1 jig ofHuNP per ml at + 10 mV. (e) Channel activity due to MuNP obtained from a single membrane at several holding potentials,as indicated at the right side of each record. (f) Current (I) vs. voltage (V) plot of the data in e. Largest conductance states were taken fromeach record from -30 mV to +10 mV (closed circles) and used to draw the straight line. Recordings were filtered at 15 Hz in a-d and 300 Hzin e.
J25pA 2t5s IIIf
Ul -J --I' _WL 11 L-11 W"'L+50rnV
4624 Immunology: Oicius et al.
ir
IT.
p IMAIL
4
ILI. " --..L'A- -1- '-A- "k4,OMV1-1 "It LI"- 1+10mv
lfimv

Proc. NatL. Acad. Sci. USA 88 (1991) 4625
DISCUSSION
The lipid bilayer results (Fig. 5) showed that HuNP andMuNP can form large ion channels with conductances com-parable to those already described for perforin (6). Othercharacteristics ofHuNP and MuNP, such as the formation ofchannels displaying multiple sizes, the induction of succes-sive and long-lasting high conductance states in a stair-likepattern, and the abrupt conductance increase at higher doses,also resemble those of perforin. These data, together with thefact that HuNP and MuNP are lytic to various cell types andcan induce liposome damage, strongly suggest that the N-ter-minal region is an important part of the pore-forming domainof perforin. The existence of several channel sizes and/orsubstates observed in several records is consistent with theidea that the N-terminal peptides, like perforin, form high-conductance channels or pores by polymerization. Additionof one or more peptides to aggregates of different sizes couldlead to various conductance changes.As opposed to perforin, which requires Ca2l for its lytic
activity to be expressed (1-4), MuNP and HuNP are lytic inthe absence of any divalent cations. It is possible that theN-terminal domain may be buried within the perforin protein,in which case Ca2l would be required to induce a confor-mational change in perforin that might uncover the lyticdomain. Once the lytic domain is exposed and bound to theTC membrane, it could facilitate the insertion of the rest ofthe perforin molecule into the membrane, at which stageseveral membrane-spanning domains could participate simul-taneously during perforin polymerization into the 20-nmpores. Heparin and other blood serum proteins would pre-vent lysis of innocent bystander cells by binding to theN-terminal domain only if perforin has not yet bound to theTC membrane; the lytic domain, presumably inserted be-tween the membrane and the bulk of the protein, would beinaccessible to the serum proteins once perforin has bound tothe membrane. Consistent with this model, antibodies againstthe N-terminal domain are able to intercept perforin-mediated lysis only if perforin has not yet bound to themembrane. On the other hand, whatever mechanism protectscytolytic lymphocytes against self-lysis would have to rec-ognize some perforin region other than the N-terminal do-main.
In preliminary experiments, most of the lytic activity of theN-terminal domain has been located in the first 19 aminoacids. Although many small peptides are known to formbarrel-stave channels, probably the best characterized is a
family of toxins related to alamethicin and produced by thefungus Trichoderma viride (27). Alamethicin itself has 20amino acids, and the other members of the family arebetween 15 and 24 amino acids long.
Spectroscopic studies have been undertaken to determinethe secondary structure of the perforin N-terminal domain.Unlike alamethicin, the perforinN terminus does not containan amphipathic a-helical stretch long enough to span themembrane. An alternative mode of pore formation is sug-gested if the N terminus is divided into three domains,consisting of amino acids 1-5 (domain A), 6-16 (domain B),and 17-19 (domain C). Domain A is mildly hydrophobic anddomain C is strongly hydrophobic, whereas domain B ispredominantly hydrophilic. However, the hydrophobic andcharged residues in the murine domain B are distributed insuch a way that an amphipathic /B-strand could be formed.The human domain B has almost the same structure, exceptthat an arginine at position 12 disrupts the amphipathiccharacter. However, as previously pointed out (28), interac-tions between proteins and membranes are less specific thanthose between proteins and proteins, so that substitution ofa single residue in a particular location by one of an appar-
ently opposite hydrophobicity does not represent an intoler-able deviation from the peptide's amphipathicity. An attrac-tive feature of the amphipathic /-sheet model for domain Bis that amphipathic /3-sheets have a tendency to self-associate(29) and can traverse the membrane as long as they containat least 10 amino acids. If it turns out that experimentalstudies confirm this model, then the perforn N terminuswould join the growing category of transmembrane peptidesand proteins that lack hydrophobic sequences long enough tospan the membrane. The best known examples are porins,which create pores across the outer membrane ofEscherichiacoli. The bulk of this largely hydrophilic protein exists withinthe lipid bilayer, and its secondary structure appears toconsist mostly of antiparallel /3-pleated sheets (30).
We are grateful to Zanvil A. Cohn, Antonio C. Carvalho, Ralph M.Steinman, and Masako 0. Massuda for support and helpful discus-sions. This work was financed by the Juvenile Diabetes FoundationInternational, the American Diabetes Association, the AmericanCancer Society, and Conselho Nacional de Desenvolvimento Cien-tifico e Tecnologico and Financiadora de Estudos e Projetos/FundoNacional de Desenvolvimento Cientifico e Technologico.
1. Henkart, P. A. (1985) Annu. Rev. Immunol. 3, 31-58.2. Podack, E. R. (1985) Immunol. Today 6, 21-27.3. Tschopp, J. & Jongeneel, C. V. (1988) Biochemistry 27, 2641-2646.4. Young, J. D.-E (1989) Physiol. Rev. 69, 250-314.5. Ojcius, D. M. & Young, J. D.-E (1990) Cancer Cells 2, 138-145.6. Young, J. D.-E, Hengartner, H., Podack, E. R. & Cohn, Z. A.
(1986) Cell 44, 849-859.7. Stanley, K. K. & Luzio, P. (1988) Nature (London) 334, 475-476.8. Jiang, S., Ojcius, D. M., Persechini, P. M. & Young, J. D.-E (1990)
J. Immunol. 144, 998-1003.9. Szoka, F. & Papahadjopoulos, D. (1980) Annu. Rev. Biophys.
Bioeng. 9, 467-508.10. Ojcius, D. M. & Young, J. D.-E (1990) Mol. Immunol. 27, 257-261.11. Young, J. D.-E, Damiano, A., DiNome, M. A., Leong, L. G. &
Cohn, Z. A. (1987) J. Exp. Med. 165, 1371-1382.12. Young, J. D.-E, Cohn, Z. A. & Podack, E. R. (1986) Science 233,
184-190.13. Young, L. H. Y., Joag, S. V., Zheng, L. M., Lee, C.-P., Lee, Y.-S.
& Young, J. D.-E (1990) Lancet 336, 1019-1021.14. Mueller, P., Rudin, D. O., Tien, H. T. & Wescott, W. C. (1963) J.
Phys. Chem. 67, 534-535.15. Alvarez, 0. (1986) in Ion Channel Reconstitution, ed. Miller, C.
(Plenum, New York), pp. 115-130.16. Shinkai, Y., Takio, K. & Okumura, K. (1988) Nature (London) 334,
525-527.17. Lichtenheld, M. G., Olsen, K. J., Lu, P., Lowrey, D. M., Hameed,
A., Hengartner, H. & Podack, E. R. (1988) Nature (London) 335,448-451.
18. Lowrey, D. M., Aebischer, T., Olsen, K., Lichtenheld, M., Rupp,F., Hengartner, H. & Podack, E. R. (1989) Proc. Natl. Acad. Sci.USA 86, 247-251.
19. Kwon, B. S., Wakulchik, M., Liu, C.-C., Persechini, P. M., Tra-pani, J. A., Haq, A. K., Kim, Y. & Young, J. D.-E (1989) Biochem.Biophys. Res. Commun. 158, 1-10.
20. Terwilliger, T. C., Weissman, L. & Eisenberg, D. (1982) Biophys.J. 37, 353-361.
21. Tschopp, J., Masson, D. & Schafer, S. (1986) J. Immunol. 137,1950-1953.
22. Young, J. D.-E & Liu, C.-C. (1988) Immunol. Today 9, 14-15.23. Tschopp, J., Schafer, S., Masson, D., Peitsch, M. & Heusser, C.
(1989) Nature (London) 337, 272-274.24. Blumenthal, R., Millar, P. J., Henkart, M. P., Reynolds, C. W. &
Henkart, P. A. (1984) Proc. Natl. Acad. Sci. USA 81, 5551-5555.25. Wyllie, A. H., Kerr, J. F. R. & Currie, A. R. (1980) Int. Rev. Cytol.
68, 251-306.26. Zheng, L. M., Zychlinsky, A., Liu, C.-C., Ojcius, D. M. & Young,
J. D.-E (1991) J. Cell Biol. 112, 279-288.27. Mathew, M. K. & Balaram, P. (1983) Mol. Cell. Biochem. 50,
47-64.28. Taylor, J. W. & Kaiser, E. T. (1987) Methods Enzymol. 154,
473-498.29. Osterman, D. G. & Kaiser, E. T. (1985)J. Cell. Biochem. 29,57-72.30. Kleffel, B., Garavito, R. M., Baumeister, W. & Rosenbusch, J. P.
(1985) EMBO J. 4, 1589-1592.
Immunology: Ojcius et al.





























