Embed Size (px)
Citation preview

q 2004 The Paleontological Society. All rights reserved. 0094-8373/04/3001-0000/$1.00
Paleobiology, 30(1), 2004, pp. 82–107
Dark and disturbed: a new image of early angiosperm ecology
Taylor S. Feild, Nan Crystal Arens, James A. Doyle, Todd E. Dawson, andMichael J. Donoghue
Abstract.—Better understanding of the functional biology of early angiosperms may clarify eco-logical factors surrounding their origin and early radiation. Phylogenetic studies identify Ambor-ella, Nymphaeales (water lilies), Austrobaileyales, and Chloranthaceae as extant lineages thatbranched before the radiation of core angiosperms. Among living plants, these lineages may rep-resent the best models for the ecology and physiology of early angiosperms. Here we combinephylogenetic reconstruction with new data on the morphology and ecophysiology of these plantsto infer early angiosperm function. With few exceptions, Amborella, Austrobaileyales, and Chlor-anthaceae share ecophysiological traits associated with shady, disturbed, and wet habitats. Thesefeatures include low and easily light-saturated photosynthetic rates, leaf anatomy related to thecapture of understory light, small seed size, and clonal reproduction. Some Chloranthaceae, how-ever, possess higher photosynthetic capacities and seedlings that recruit in canopy gaps and othersunny, disturbed habitats, which may have allowed Cretaceous Chloranthaceae to expand intomore diverse environments. In contrast, water lilies possess ecophysiological features linked toaquatic, sunny habitats, such as absence of a vascular cambium, ventilating stems and roots, andfloating leaves tuned for high photosynthetic rates in full sun. Nymphaeales may represent an earlyradiation into such aquatic environments. We hypothesize that the earliest angiosperms werewoody plants that grew in dimly lit, disturbed forest understory habitats and/or shady streamsidesettings. This ecology may have restricted the diversity of pre-Aptian angiosperms and living basallineages. The vegetative flexibility that evolved in the understory, however, may have been a keyfactor in their diversification in other habitats. Our inferences based on living plants are consistentwith many aspects of the Early Cretaceous fossil record and can be tested with further study ofthe anatomy, chemistry, and sedimentological context of Early Cretaceous angiosperm fossils.
Taylor S. Feild. Department of Botany, University of Toronto, Toronto, Ontario M5S 3B2, Canada. E-mail:[email protected]
Nan Crystal Arens. Department of Geoscience, Hobart and William Smith Colleges, Geneva, New York14456
James A. Doyle. Section of Evolution and Ecology, University of California, Davis, California 95616Todd E. Dawson. Department of Integrative Biology, University of California, Berkeley, California 94720-
3140Michael J. Donoghue. Department of Ecology and Evolution, Yale University, New Haven, Connecticut
06520
Accepted: 21 May 2003
Introduction
Flowering plants—angiosperms—today in-clude more than 257,000 species and have dom-inated most terrestrial ecosystems since the LateCretaceous (Crane 1987; Crane and Lidgard1989; Crane et al. 1995; Wing and Boucher 1998;Lupia et al. 1999). Much discussion has focusedon the ecology of the earliest angiosperms.What sorts of habitats did they occupy? Howdid they function? Answers remain elusive, ow-ing in part to disparate views on the ecologicalstarting point for flowering plant evolution(Doyle and Donoghue 1993; Sanderson andDonoghue 1994).
Previous Interpretations of Early AngiospermEcology. The traditional representation of early
angiosperms is that of woody magnoliids, withlarge bisexual flowers consisting of numerousfree perianth parts, stamens, and carpels, as inextant Magnoliales and Winteraceae (e.g., Arberand Parkin 1907; Takhtajan 1969; Thorne 1976;Cronquist 1988; Gottsberger 1988). By analogywith these living taxa, the first flowering plantswould be slowly growing and maturing trees orshrubs with large leaves that photosynthesizedat low rates. These plants would have estab-lished in wet, low-light environments below theforest canopy (Table 1) (Bews 1927; Axelrod1952; Takhtajan 1969; Thorne 1976; Cronquist1988).
In contrast, others suggested that the first an-giosperms were weedy, drought-tolerant shrubs

83EARLY ANGIOSPERM ECOPHYSIOLOGY
TA
BL
E1.
Sum
mar
yof
hy
pot
hes
esfo
rth
ean
cest
ral
ecop
hy
siol
ogy
ofan
gio
sper
ms.
Sou
rces
for
ecol
ogic
ald
etai
lsof
each
hy
pot
hes
isar
en
oted
;‘‘
?’’
den
otes
that
no
exp
lici
tst
atem
ents
wer
em
ade
byau
thor
sre
gar
din
ga
par
ticu
lar
char
acte
rist
ic,
but
trai
tsw
ere
pre
dic
ted
from
des
crip
tion
sof
ance
stra
lh
abit
ats.
‘‘P
aleo
her
b’’
clad
esin
clu
de
Ari
stol
och
iace
ae,L
acto
ris,
Pip
eral
es,m
onoc
ots,
Cer
atop
hyllu
m,N
ym
ph
aeal
es.
Woo
dy
mag
nol
iid
(Tak
hta
jan
1969
;T
hor
ne
1976
)
Wee
dy
xeri
csh
rub
tori
par
ian
wee
d(S
tebb
ins
1974
)
Ru
der
al/
her
b(T
aylo
ran
dH
icke
y19
92,
1996
)A
quat
icor
igin
(Su
net
al.2
002)
‘‘D
ark
and
dis
turb
ed’’
(th
isst
ud
y)
Ch
arac
ters
Mod
ern
anal
ogs
Mag
nol
iale
s,W
inte
ra-
ceae
Shru
bby
Eri
cace
ae,D
il-
lien
acea
e,an
dTa
s-m
ann
ia(W
inte
race
ae)
‘‘P
aleo
her
b’’
clad
esp
lus
Ch
lora
nth
acea
e?
Ny
mp
hae
ales
,Cer
ato-
phyl
lum
Am
bore
lla,
Ch
lora
nth
a-ce
ae,A
ust
rob
aile
yale
s
Gro
wth
hab
itL
arg
eto
smal
ltr
ees
Shru
bor
sub
-sh
rub
,h
igh
lybr
anch
edR
hiz
omat
ous
her
bs
Aqu
atic
her
bSm
all
tree
/sh
rub
Seed
lin
ges
tabl
ish
men
t?
Wet
,sh
ady,
and
stab
lesi
tes
Seas
onal
lyd
ry,s
un
ny
Var
iabl
yw
et,s
un
ny,
and
dis
turb
edh
abi-
tats
?W
et,
shad
y,an
dst
able
Wet
,sh
ady,
and
dis
-tu
rbed
hab
itat
s
Dis
turb
ance
Low
Hig
hH
igh
,lar
ge-
scal
e(e
.g.,
clea
rin
gs,
floo
dp
lain
),st
ream
sid
es
?L
owH
igh
,sm
all-
scal
e(e
.g.,
stre
amsi
des
,ste
epsl
opes
)Se
edsi
ze?
Lar
ge
?Sm
all
Smal
l?
Smal
lV
eget
ativ
ere
pro
du
ctio
n?
Low
Hig
hH
igh
?H
igh
Ph
otos
yn
thes
is?
Low
?H
igh
orlo
w,d
epen
d-
ing
up
ond
rou
ght
se-
veri
ty
Hig
h?
Hig
hto
mod
erat
eL
ow
Res
ourc
esL
igh
tL
owd
uri
ng
esta
blis
h-
men
t,h
igh
late
ras
can
opy
tree
s
Hig
hH
igh
Hig
hL
ow
Wat
erH
igh
Low
Var
iabl
e,fl
ood
ing
wit
hp
erio
dic
dry
ing
Hig
h,a
quat
icH
igh

84 TAYLOR S. FEILD ET AL.
FIGURE 1. Phylogenetic systematics of basal angio-sperm lineages included in this study, as identified byrecent molecular phylogenetic analyses (see Doyle andEndress 2000). The number of species in each clade isdenoted in parentheses (data from Smith 1976; Verd-court 1986; Todzia 1988; Schneider and Williamson1993; Williamson and Schneider 1993; Saunders 1998,2000; Kong et al. 2002).
that lived in open, disturbed habitats of semi-arid tropical to subtropical regions (Table 1)(Stebbins 1965, 1974; Axelrod 1970). From there,other authors suggested that angiospermsmoved into disturbed streamside habitats inmesic environments (Doyle and Hickey 1976;Hickey and Doyle 1977). These suggestions builton arguments that variable conditions mighthave favored the evolution of the reproductiveand vegetative hallmarks of angiosperms, suchas short generation time, the closed carpel, re-duction of ovules and gametophytes, double fer-tilization and endosperm, and flexible seedlinggrowth (Stebbins 1965, 1974), and on paleoeco-logical inferences from the diversity of EarlyCretaceous angiosperm pollen in the NorthernGondwana tropics, associated with indicators ofaridity, and the morphology and sedimentaryassociations of Early Cretaceous angiospermleaves (Brenner 1976; Doyle and Hickey 1976;Hickey and Doyle 1977). Subsequent studies,however, showed that angiosperm pollen wasequally diverse in parts of Northern Gondwanathat show evidence for wetter climates (Doyle etal. 1982; Brenner 1996).
A related view reconstructed the first angio-sperms as fast-growing, rhizomatous, and semi-herbaceous plants of sunny, unstable stream-sides (Table 1). This interpretation arose fromearly phylogenetic analyses that rooted the an-giosperms among the ‘‘paleoherbs,’’ variouslyincluding Nymphaeales, Piperaceae, Saurura-ceae, Lactoris, Aristolochiaceae, and Chlorantha-ceae (Donoghue and Doyle 1989; Taylor andHickey 1992). The paleoherb hypothesis was ar-gued to be consistent with small angiospermseed sizes and the paucity of angiosperm woodin Early Cretaceous floras (Taylor and Hickey1990, 1992, 1996; Doyle et al. 1994; Nixon et al.1994; Wing and Boucher 1998; Eriksson et al.2000). In this scenario, early angiosperms tol-erated disturbance and had high leaf photosyn-thetic capacity and short generation times (Tay-lor and Hickey 1992, 1996; Wing and Boucher1998).
Finally, the possibility that the first angio-sperms were aquatic has enjoyed renewed atten-tion with the discovery of Archaefructus, a re-markably complete fossil with finely dissectedleaves from Barremian–Aptian lake deposits ofChina. Archaefructus was interpreted as an
aquatic herb (Sun et al. 1998, 2002), and a cla-distic analysis by Sun et al. (2002) placed it be-low the common ancestor of all living angio-sperms.
A New Perspective. Discussions of the ances-tral ecology and habitat of angiosperms havesuffered from two main difficulties. First, pre-vious hypotheses were not well constrained byrobust phylogenies. However, independent mo-lecular analyses have recently converged onsimilar rootings of the angiosperm tree. Thesestudies placed Amborella, Nymphaeales (Ca-bombaceae and Nymphaeaceae, the water lil-ies), and Austrobaileyales (consisting of Austro-baileya, Trimenia, the star anise Illicium, Kadsura,and Schisandra) at the base of the angiospermlineage (Fig. 1) (Soltis et al. 1997, 2000; Mathewsand Donoghue 1999; Parkinson et al. 1999; Qiuet al. 1999; Barkman et al. 2000; Graham andOlmstead 2000; Zanis et al. 2002). Chlorantha-ceae, which occupied various near-basal posi-tions in the molecular phylogenies just cited,branched immediately above this basal gradewhen molecular and morphological data werecombined (Doyle and Endress 2000). In moststudies the root was between Amborella and all

85EARLY ANGIOSPERM ECOPHYSIOLOGY
other angiosperms, although trees in which Am-borella plus Nymphaeales form a clade sister tothe rest cannot be rejected (Barkman et al. 2000;Zanis et al. 2002). However, rooting the angio-sperms near Magnoliales, Chloranthaceae, orCeratophyllum can now be ruled out. Significant-ly, several morphological characteristics of Am-borella, Nymphaeales, Austrobaileyales, andChloranthaceae are known in Early Cretaceousfossil flowers, pollen, seeds, and leaves (Up-church 1984b, 1995; Mohr and Friis 2000; Friiset al. 1997, 1999, 2000, 2001; Doyle 2001), thussupporting the view that these taxa may rep-resent appropriate ecological models for earlyangiosperms.
Here Amborella, Austrobaileyales, Chloran-thaceae, and Nymphaeales are referred to as‘‘basal’’ lineages, as contrasted with the muchlarger clade nested among them that includes allremaining angiosperms (called ‘‘core angio-sperms’’). The term ‘‘basal’’ is often criticizedbecause it is sometimes misused as a synonymfor ‘‘primitive.’’ In fact, basal groups can behighly autapomorphic. Also, only nodes are ac-tually basal. While recognizing these points, webelieve ‘‘basal’’ is a useful shorthand for ‘‘aclade whose stem lineage is attached to a basalor near-basal node.’’ More importantly, whenseveral lineages branch off successively below amajor clade, as is the case in angiosperms, anystates that they share can be inferred to be an-cestral, even if each of these lineages is autapo-morphic in other characters.
A second difficulty is lack of detailed eco-physiological data for putative basal lineageswith which to interpret the paleo-ecophysiologyof Early Cretaceous angiosperms. Previous in-terpretations of early angiosperm ecology reliedon structure-function relations taken from de-rived groups, in which the characters may haveevolved convergently (Doyle and Hickey 1976;Doyle 1977, 1978; Hickey and Doyle 1977; Taylorand Hickey 1996; Wing and Boucher 1998; Er-iksson et al. 2000). Although this approach maybe useful, direct physiological evidence dem-onstrates that inferences of physiological func-tion based on distantly related proxies can bemisleading (Feild et al. 1998).
In this study we take a fresh look at the prob-lem of early angiosperm ecology by combiningan improved knowledge of phylogeny with eco-
physiological observations on Amborella, Austro-baileyales, Chloranthaceae, and Nymphaeales,aimed at recognition of common ecophysiolog-ical patterns in these groups. We focus on traitsrelated to growth under varying light and dis-turbance regimes, including comparative leafanatomy, leaf photosynthetic performance,growth habit, seed size, and seedling establish-ment. The result is a new image of how the firstflowering plants may have functioned and theenvironment they first occupied. Finally, weconsider how these results relate to the EarlyCretaceous fossil record.
Methods
Study Species and Field Sites
Seventy-two species of extant basal angio-sperms were studied, representing Amborella, allwater lily genera (Barclaya, Brasenia, Cabomba,Euryale, Nuphar, Nymphaea, Ondinea, and Victo-ria), all genera in Austrobaileyales (Austrobaileya,Illicium, Kadsura, Schisandra, Trimenia), andChloranthaceae (Ascarina, Chloranthus, Hedyos-mum, Sarcandra). We examined individuals innatural populations (field sites in Australia, Chi-na, Costa Rica, Fiji, French Polynesia, New Ca-ledonia, New Zealand, Papua New Guinea, andU.S.A.; Appendix 1), a common garden, and bo-tanical gardens. Where taxonomic samplingcould not be exhaustive, species were selected toencompass the habitat range of each lineage.Fieldwork was conducted from August 1999 toOctober 2002.
Plants grown in the common garden (Au-burn, Alabama, 328N,858W) were obtained fromlocal nurseries or raised from cuttings takenfrom natural populations. Plants were grownfor 18 months under three light environments—100%, 30%, and 5% full sunlight—before mea-surements were taken.
Comparative Data
Six ecological and physiological characterswere obtained from the field, herbarium vouch-ers, and published sources. Data sources forspecies character-state values are given in Ap-pendix 2. The complete character matrix for thespecies sampled is provided in Appendix 3.
Photosynthetic Performance: Maximum Leaf Pho-tosynthetic Rate and Light-Saturation Point. Chlo-

86 TAYLOR S. FEILD ET AL.
rophyll (Chl) fluorescence emission was used toquantify leaf photosynthetic performance bydetermining maximum photosynthetic rate andthe light intensity at which the photosyntheticsystem saturates (Bilger et al. 1995; Brodribband Hill 1997; Rascher et al. 2000). We took Chlmeasurements of living plants in natural or gar-den settings with a portable pulse-amplitudemodulated fluorometer (Mini-PAM, H. Walz,Effeltrich, Germany) operated using currentlyaccepted protocols (Bilger et al. 1995; Brodribband Hill 1997; Rascher et al. 2000). From light-response curves of Chl fluorescence emission,we calculated maximum ETR (ETRmax) and thelight intensity (PPFDsat) at which this flux oc-curred, and we used these calculations for in-terspecific comparisons of leaf photosyntheticperformance (Appendix 4). Appendix 4 alsoprovides a comprehensive discussion of proce-dures and a verification of sampling methods.
Seed Size. Seed sizes were expressed involume to facilitate comparison with pub-lished data on Cretaceous angiosperm seeds(Tiffney 1984; Wing and Boucher 1998; Eriks-son et al. 2000). Most seeds from extant plantswere collected in the field and measured di-rectly. Some seed volumes were determinedby nondestructive measurement of herbariumvoucher specimens (specific sources from theUniversity of California and Jepson herbariaare available from T. S. Feild) or from pub-lished studies (Appendix 2). For each species,the length (L) and breadth (B) of five seedswere measured with a micrometer (0.1 mmprecision) and averaged. We used the approx-imation of Tiffney (1984), where seed thick-ness (T) 5 0.66 B. To calculate seed volume(V), seeds were approximated as ellipsoidswith V 5 4/3 (ab2), where a 5 L/2 and b 5(B 1 T)/4 (Eriksson et al. 2000).
Seedling Habitats. To evaluate how seed-lings and small saplings (,40 cm tall, ,0.5 cmstem diameter) of extant basal angiospermswere associated with habitat, eight plots(approx. 10 m 3 10 m) were laid out and sur-veyed for seedling abundance. Plots were ran-domly established in a variety of habitats, in-cluding large (100 m2) to small (less than 20m2) canopy gaps, understory, understorystreamside zones, and exposed forest edges,ridgecrests, pastures, and stream banks. Spe-
cies were easily identified by leaf characters.Seedling and sapling abundance was low fornearly all species studied, ranging from twoto 45 plants/plot. These descriptions are lim-ited because they rely on single-time obser-vations and may include plants of differentage.
Disturbance regime was categorized on thebasis of physical attributes of the site (see be-low) and whether seedling establishment oc-curred in soil litter (undisturbed) or mineralsoil (disturbed). This method of site charac-terization parallels that of other workers andproves useful in designating relative catego-ries of disturbance frequency (Sakai et al.1995; Arens 2001; Dai et al. 2002). To quantifylight environment, eight measurements of dif-fuse PPFD were made with a hand-held lightmeter (Li-190; Li-COR, Lincoln, NB) aboveeach seedling and averaged. Light availabilitywas expressed as a percentage of average fulldiffuse sunlight (495 mmol photons m22 s21;SD 55; n 5 39 sites) to avoid considerable het-erogeneity introduced by sunflecks (Chazdon1988). Measurements of full diffuse light weremade at midday on cloudy days.
By combining light intensity data with sitecharacteristics, three seedling habitat stateswere defined. Shady, stable, and aquatic (0) hab-itats have low light (,2% full diffuse sunlight)and included submerged aquatic zones withlow current or wave action (e.g., muddy pondand stream bottoms rich in organic matter).Shady, disturbed (1) habitats have low light (,2% full diffuse sunlight) and were recently dis-turbed. Examples included exposed mineralsoils on steep understory slopes, margins ofunderstory watercourses, understory trailsand irrigation canals, and sites in the shade ofregenerating vegetation near roadsides. Sun-ny, disturbed (2) habitats have higher light (5–100% full diffuse sunlight) and experiencedrecent disturbance. These sites includedbrighter subcanopy to open disturbed areas,such as open stream margins, cleared slopesand ravines, small canopy gaps, and exposedlandslip margins.
Leaf Anatomy. Leaf anatomy was observedin vibratome cross-sections (50 mm thick)from leaves preserved in 20% ethanol, 15%glacial acetic acid, 15% formaldehyde, and

87EARLY ANGIOSPERM ECOPHYSIOLOGY
50% water. Three leaves were taken from thesame branches used for Chl a fluorescencemeasurements. Leaf sections were cleared forone hour in a solution of 50% bleach, rinsedwith water, and stained with toluidine blue.Three leaf mesophyll anatomy states were de-fined (terminology following Esau 1977):spongy parenchyma only (0), consisting entirelyof spherically shaped mesophyll cells; single,weakly differentiated layer of palisade parenchyma(1), with one tier of columnar cells organizedinto a distinct layer at the upper leaf surface;and multiple layers of palisade parenchyma (2),with two to three stacked layers of columnarcells near the upper leaf surface.
Growth Forms. Four growth form catego-ries were recognized. Tree or shrub (0) included(following Hickman 1993) woody perennialsproducing secondary xylem with a distincttrunk-crown architecture (trees) and woodyperennials producing several trunks from thebase and lacking a distinct trunk-crown sys-tem (shrubs). Twining woody vine (1) includedwoody perennials completely or partly de-pendent upon other plants for mechanicalsupport and climbing with circumnutatedgrowth. Herbs with some secondary growth (2)had above-ground shoots generally lackingsecondary xylem, but rhizomes and bases ofannual shoots developing limited secondaryxylem. Finally, herbs lacking secondary growth(3) developed only primary xylem.
Phylogeny and Character Reconstruction
Ancestral states and patterns of characterevolution were reconstructed by assembling acomposite phylogenetic tree of extant basalangiosperms, including Amborella, Nymphae-ales, Austrobaileyales, Chloranthaceae, and aplaceholder for the remaining ‘‘core angio-sperm’’ clade (including monocots, eudicots,Piperales, Winterales, Laurales, and Magno-liales). As the backbone, we used the singlemost parsimonious tree found in Doyle andEndress’s (2000) analysis of three genes (rbcL,atpB, 18S rDNA) plus morphology, whichidentified Chloranthaceae as sister to the coreangiosperms. The Chloranthaceae were ex-panded by using a parsimony analysis of mor-phological data (Eklund et al. 2004), but withrelationships within Chloranthus based on a
molecular study (ITS and trnL-F [Kong et al.2002]). Nymphaeales were expanded by usingthe single most parsimonious tree from ananalysis by Les et al. (1999) of three genes(rbcL, matK, 18S rDNA) plus morphology. ForIllicium, the molecular tree of Hao et al. (2000),based on ITS sequences, was used. Interspe-cific relations of Schisandra and Kadsura wererepresented by one of eight most parsimoni-ous trees from an analysis of ITS and mor-phology (Hao et al. 2001). Relationships in theTrimenia clade have not been analyzed. Forpurposes of inferring discrete character evo-lution, we took a conservative approach andleft them as an unresolved polytomy.
MacClade 4.03 (Maddison and Maddison2001) was used to reconstruct states of dis-crete and continuous characters at ancestralnodes and branches. All characters were treat-ed as unpolarized and unordered, meaning alltransitions among states were equally proba-ble. The sensitivity of ancestral state recon-structions for discrete characters to differentcoding schemes (binary versus multistate)and types (unordered versus ordered) wasalso examined. Because neither alternativecoding schemes nor ordered character statesaffected our conclusions, we took the conser-vative approach of scoring characters as un-ordered. Coding characters for the core angio-sperm clade posed problems because of lackof data; core angiosperms could only bescored for mesophyll and growth habit (fol-lowing Doyle and Endress 2000).
Ancestral states for continuously varyingtraits (ETRmax, PPFDsat, seed size) at the root ofthe tree were determined with two widelyused methods, squared-change parsimonyand linear parsimony (Maddison 1991), usingMacClade. Squared-change parsimony as-signs a single ancestral value to each internalnode to minimize the sum of squares changeover the tree. Linear parsimony reconstructsancestral node values by minimizing totalchanges. Linear parsimony reconstructionswere explored by using the maximum andminimum state options in MacClade. To re-construct node values for continuous charac-ters, we used MacClade to randomly resolvethe Trimenia polytomy. Different random res-olutions in Trimenia altered state values for the
88 TAYLOR S. FEILD ET AL.
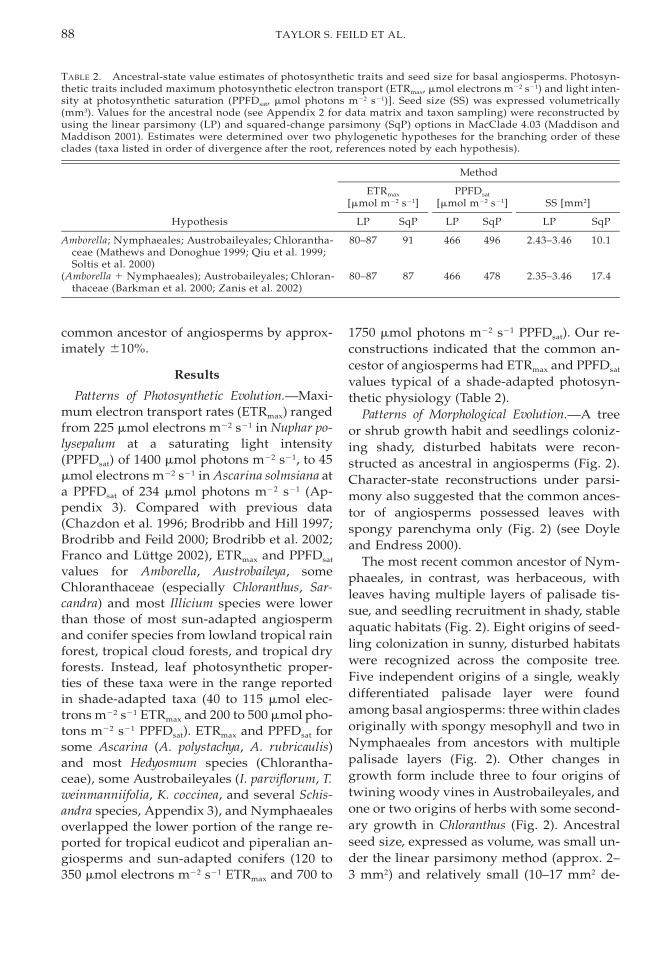
TABLE 2. Ancestral-state value estimates of photosynthetic traits and seed size for basal angiosperms. Photosyn-thetic traits included maximum photosynthetic electron transport (ETRmax, mmol electrons m22 s21) and light inten-sity at photosynthetic saturation (PPFDsat, mmol photons m22 s21)]. Seed size (SS) was expressed volumetrically(mm3). Values for the ancestral node (see Appendix 2 for data matrix and taxon sampling) were reconstructed byusing the linear parsimony (LP) and squared-change parsimony (SqP) options in MacClade 4.03 (Maddison andMaddison 2001). Estimates were determined over two phylogenetic hypotheses for the branching order of theseclades (taxa listed in order of divergence after the root, references noted by each hypothesis).
Hypothesis
Method
ETRmax
[mmol m22 s21]
LP SqP
PPFDsat
[mmol m22 s21]
LP SqP
SS [mm2]
LP SqP
Amborella; Nymphaeales; Austrobaileyales; Chlorantha-ceae (Mathews and Donoghue 1999; Qiu et al. 1999;Soltis et al. 2000)
80–87 91 466 496 2.43–3.46 10.1
(Amborella 1 Nymphaeales); Austrobaileyales; Chloran-thaceae (Barkman et al. 2000; Zanis et al. 2002)
80–87 87 466 478 2.35–3.46 17.4
common ancestor of angiosperms by approx-imately 610%.
Results
Patterns of Photosynthetic Evolution. Maxi-mum electron transport rates (ETRmax) rangedfrom 225 mmol electrons m22 s21 in Nuphar po-lysepalum at a saturating light intensity(PPFDsat) of 1400 mmol photons m22 s21, to 45mmol electrons m22 s21 in Ascarina solmsiana ata PPFDsat of 234 mmol photons m22 s21 (Ap-pendix 3). Compared with previous data(Chazdon et al. 1996; Brodribb and Hill 1997;Brodribb and Feild 2000; Brodribb et al. 2002;Franco and Luttge 2002), ETRmax and PPFDsat
values for Amborella, Austrobaileya, someChloranthaceae (especially Chloranthus, Sar-candra) and most Illicium species were lowerthan those of most sun-adapted angiospermand conifer species from lowland tropical rainforest, tropical cloud forests, and tropical dryforests. Instead, leaf photosynthetic proper-ties of these taxa were in the range reportedin shade-adapted taxa (40 to 115 mmol elec-trons m22 s21 ETRmax and 200 to 500 mmol pho-tons m22 s21 PPFDsat). ETRmax and PPFDsat forsome Ascarina (A. polystachya, A. rubricaulis)and most Hedyosmum species (Chlorantha-ceae), some Austrobaileyales (I. parviflorum, T.weinmanniifolia, K. coccinea, and several Schis-andra species, Appendix 3), and Nymphaealesoverlapped the lower portion of the range re-ported for tropical eudicot and piperalian an-giosperms and sun-adapted conifers (120 to350 mmol electrons m22 s21 ETRmax and 700 to
1750 mmol photons m22 s21 PPFDsat). Our re-constructions indicated that the common an-cestor of angiosperms had ETRmax and PPFDsat
values typical of a shade-adapted photosyn-thetic physiology (Table 2).
Patterns of Morphological Evolution. A treeor shrub growth habit and seedlings coloniz-ing shady, disturbed habitats were recon-structed as ancestral in angiosperms (Fig. 2).Character-state reconstructions under parsi-mony also suggested that the common ances-tor of angiosperms possessed leaves withspongy parenchyma only (Fig. 2) (see Doyleand Endress 2000).
The most recent common ancestor of Nym-phaeales, in contrast, was herbaceous, withleaves having multiple layers of palisade tis-sue, and seedling recruitment in shady, stableaquatic habitats (Fig. 2). Eight origins of seed-ling colonization in sunny, disturbed habitatswere recognized across the composite tree.Five independent origins of a single, weaklydifferentiated palisade layer were foundamong basal angiosperms: three within cladesoriginally with spongy mesophyll and two inNymphaeales from ancestors with multiplepalisade layers (Fig. 2). Other changes ingrowth form include three to four origins oftwining woody vines in Austrobaileyales, andone or two origins of herbs with some second-ary growth in Chloranthus (Fig. 2). Ancestralseed size, expressed as volume, was small un-der the linear parsimony method (approx. 2–3 mm2) and relatively small (10–17 mm2 de-

89EARLY ANGIOSPERM ECOPHYSIOLOGY
FIGURE 2. Phylogenetic mapping of growth habit, leaf mesophyll anatomy, and habitat of seedling establishmentin extant basal angiosperms. The composite phylogenetic tree was assembled as described in ‘‘Materials and Meth-ods.’’
pending on the phylogenetic hypothesis; Ta-ble 2) under squared-change parsimony.
Discussion
The Dark and Disturbed Pattern
Mapping ecophysiological characters ontothe phylogeny of extant basal angiospermssuggests that their common ancestor was atree or shrub that established in disturbedhabitats under low light (Fig. 2). In contrast,the herbaceous habit, higher leaf photosyn-thetic rates and light-saturation points, pali-sade mesophyll tissue, and seedling recruit-ment in more brightly lit, disturbed habitats
were derived in separate lineages. This inter-pretation of the ancestral ecology is novel incombining two habitat characteristics thathave not been previously associated: under-story shade and disturbance. Earlier hypoth-eses reconstructed the earliest angiosperms asshade-tolerant but slow-growing residents ofstable understory habitats, or weedy, shade-intolerant colonizers of open disturbed habi-tats (Table 1). Blanc (1986: p. 110) suggestedthat angiosperms were originally understoryshrubs, like some Chloranthaceae, but he didnot emphasize the role of disturbance. Belowwe consider how inferred ancestral ecophysi-

90 TAYLOR S. FEILD ET AL.
ological characters relate to shady and dis-turbed habitats. We also discuss preliminaryresults on basal angiosperm water-use physi-ology that indicate early angiosperms wereadapted to wet habitats.
Ancestral Characters Related to Shade. A leafcross-section dominated by spongy parenchy-ma was ancestral in angiosperms (Fig. 2). Thespheroid cellular shapes making up spongyparenchyma create numerous light-reflectingair-water interfaces that produce more light-scattering than palisade tissue. Increased scat-tering consequently lengthens the path oflight through the leaf, increasing the likeli-hood that photons will be harvested for car-bon gain—an important trait in light-limitedforest understories (Vogelmann et al. 1996;Smith et al. 1997). The reconstructed ancestralphotosynthetic physiology (maximum photo-synthetic electron transport rate and light-sat-uration point) was typical of shade-adaptedtropical understory plants (Table 1, Appendix3) (Bilger et al. 1995; Chazdon et al. 1996;Brodribb and Hill 1997; Brodribb and Feild2000; Brodribb et al. 2002; Franco and Luttge2002). Low leaf photosynthetic capacity offersan advantage in shady understory habitats be-cause such leaves are less costly to build andmaintain (Givnish 1979, 1988; Chazdon et al.1996). Low photosynthetic capacity, however,can constrain a plant’s ability to exploit high-light open habitats because of greater risk ofphotoinhibition (Demmig-Adams et al. 1995;Smith et al. 1997).
Ancestral Characters Related to Disturbance.Our data suggest that early angiosperms re-cruited in disturbed habitats. Generally, thedisturbed sites colonized by seedlings of Am-borella, most Austrobaileyales, and Chloran-thaceae were small (2–20 m2). Examples in-cluded limbfall gaps, understory streambanks, and steep mineral soil patches exposedby erosion or the activity of large animals (Ap-pendix 3). Excepting water lilies, basal angio-sperm seedlings established on unstable sub-strates prone to shifting, such as brittle clay,stony sand and shaly soils, and sites betweenor in moss mat crevices on rotting logs. Traitsthat permit recruitment on disturbed sites andallow plants to survive root and shoot dam-
age are essential for survival in these habitats(Greig 1993; Bond and Midgely 2001, 2003).
One such trait is small seed size. The in-ferred common ancestor of angiosperms pro-duced small seeds (Tables 1, 2) and probablysmall embryos (Doyle and Endress 2000; For-bis et al. 2002). Although large seeds areviewed as advantageous in shady environ-ments because abundant carbon reserves areneeded to establish in low light (Salisbury1942; Harper et al. 1970; Grime 1979; Leish-man et al. 2000; but see Kelly 1995), recentwork demonstrates that small seeds (volumesless than ca. 10 mm3) can also be viable inshady, disturbed habitats, such as steep un-derstory slopes, exposed mineral soil wash-outs, and stream banks—exactly the habitatswhere most extant basal angiosperms occur,as opposed to undisturbed forest floor mi-crosites choked with litter (Lusk 1995; Grubb1998; Lusk and Kelly 2003). Despite limitedcarbon reserves, small seeds establish in thesehabitats because they are more readily lodgedin small litter-free microsites than largerseeds (Lusk and Kelly 2003).
The diversity of vegetative regenerationmodes found in basal angiosperms also in-creases tolerance of disturbance (Table 3). Am-borella, Austrobaileyales, and Chloranthaceaesprout abundantly from the base of adult(flowering) and juvenile (nonflowering)plants. Their seedlings become highly rami-fied, passing through a creeping establish-ment phase consisting of a decumbent ligno-tuber with several basal sprouts (Table 3)(Blanc 1986). Austrobaileyales and Chloran-thaceae, in addition, display layering of de-bris-pinned shoots, rhizomes, and ability ofsnapped shoots to reroot and establish in thehumid forest understory or near creeks (Table3). Extensive clonal growth, resulting in shootredundancy and high meristem dispersionacross the forest floor, makes basal angio-sperms diffuse and resilient targets for limb-falls, trampling, and herbivore damage. Thesekinds of disturbance are those most typical ofthe forest understory (Greig 1993; Sakai et al.1995; Canham et al. 1999; Gavin and Peart1999; Bond and Midgely 2001, 2003). Our re-sults support the conjecture of Wells (1969)

91EARLY ANGIOSPERM ECOPHYSIOLOGY
TABLE 3. Modes of clonal growth and regeneration among basal angiosperms. Definitions for vegetative modeswere modified from Greig 1993. Basal sprouting refers to the production of stem suckers, whereby new trunks arisefrom the base of the original plant from basal buds in plants that have flowered (adults) or not (juveniles). Creepingseedlings refers to a pattern of rhizomatous seedling development that is monocot-like, characterized by a decum-bent (‘‘creeping’’) lignotuber, from which several ascending shoots emerged. Layering was defined as the produc-tion of adventitious roots when trunks or branches become pinned to the ground and then new stems grow upward.Shoot fragmentation was defined as the ability of snapped-off pieces of stems to re-root and eventually form a newplant. Rhizomes and stolons are underground or above-ground stems that give rise to closely spaced, intercon-nected shoot systems. Observations are based on observations of field-grown plants or from literature sources (Blanc1986; Schneider and Williamson 1993; Williamson and Schneider 1993; Barrat-Segretain 1996). n is the total numberof species observed; see Appendix 1 for the species studied. ‘‘Common’’ means that all or more than 75% of specieswere observed regenerating in a particular way, whereas ‘‘infrequent’’ refers to between 30% and 74% of species,and ‘‘rare’’ to fewer than 29% of species. The presence (‘‘yes’’) and absence (‘‘no’’) for particular mode was usedfor lineages containing one or two species.
Taxon nBasal sproutingAdult/Juvenile
Creepingseedlings Layering
Recruitmentby fragments
Rhizomesor stolons
AmborellaNymphaeales
17
yescommon
yesno
noinfrequent
nocommon
noinfrequent
AustrobaileyalesAustrobaileya 1 yes yes yes no yesIlliciumKadsura and SchisandraTrimenia
1415
4
commoncommoncommon
commoncommoncommon
commoncommoninfrequent
rarerareinfrequent
rarecommoninfrequent
ChloranthaceaeAscarinaChloranthusHedyosmumSarcandra
8662
commoncommoncommonyes/yes
commoncommoncommonyes
infrequentinfrequentinfrequentyes
commonnocommonno
rarecommonrareyes
that sprouting is an ancestral trait of angio-sperms.
Ancestral Characters Related to Moist Condi-tions. Preliminary data on basal angio-sperms suggest that early angiosperms wereadapted to high water availability. Amborella,Austrobaileyales, and Chloranthaceae arelargely restricted to tropical and subtropicalhabitats, predominantly nonseasonal mon-tane cloud forests, with high rainfall (3000 to10,000 mm yr21) and mist (Todzia 1988; Feildet al. 2000, 2003a; Feild unpublished data2002). Species from drier temperate regions(.1700–2200 mm) occur in wet microsites,such as perennial understory water coursesand springs (Stone 1968; Kwit et al. 1998; Luoand Li 1999; Feild unpublished data 2002).Consistent with a preference for wet habitats,the physiologies of most basal angiospermsare tuned to low evaporative demand. Terres-trial basal angiosperm leaves possess low sto-matal densities (45–115, x 5 65 stomata mm22,SD 5 4, n 5 75 taxa) as well as low instanta-neous (Feild et al. 2003a) and integrated leafwater-use efficiencies (the ratio of carbon gain
to transpirational water loss), as inferred fromleaf stable carbon isotopic composition (Far-quhar et al. 1982; Feild unpublished data2002). Amborella and Austrobaileya stems pos-sess lower ability to transport water comparedwith most tropical eudicots (Feild et al. 2000,2001, 2003a; Feild unpublished data 2002).Consistent with xylem hydraulic measure-ments, vascular features characteristic ofmoisture-loving plants pervade woods of Am-borella, Austrobaileyales, and Chloranthaceae,including vesselless xylem with scalariformpitting (Amborella) and wood with short, an-gular vessels (4–120 mm long in stems 2–5mm in diameter) dominated by scalariformpits with incompletely hydrolyzed pit mem-branes (Carlquist 1975, 1984; Carlquist andSchneider 2002; Feild unpublished data 2002).More detailed exploration of these lines of ev-idence would be desirable.
Exceptions to the Dark and DisturbedPattern
Water Lilies. The aquatic habit of Nym-phaeales contrasts with that reconstructed for

92 TAYLOR S. FEILD ET AL.
the common ancestor of extant angiosperms(Fig. 2). Inferred specializations for freshwateraquatic zones include herbaceousness (Fig. 2)and evolution of aerenchymatous gas-ex-change canals, running from leaf to root,which allow downward oxygen transport intoanaerobic muddy soils and upward flow oftoxic methane (Sculthorpe 1967; Dacey 1980).Some water lily leaves also show modifica-tions for underwater photosynthesis, such asfinely dissected shape (Cabomba) and lack ofcuticle, stomata, and/or intercellular air-spac-es (Sculthorpe 1967). Consequently, only float-ing ‘‘lily pad’’ and emergent leaves of Nym-phaeales are potentially comparable to leavesof terrestrial basal angiosperms. Even in theseleaves, large functional differences, related tothe occupation of brightly lit open-water hab-itats, were found. For example, water liliesevolved considerably higher leaf photosyn-thetic rates and light-saturation points (exceptCabomba; Appendix 3), as well as multiple-lay-ered palisade mesophyll tissue (Fig. 2) andleaves with dense stomata (up to 500 mm22
[Schneider and Williamson 1993]). Accordingto the most parsimonious distribution of an-cestral states, water lily ecophysiology bearsweakly on that of the first angiosperms be-cause aquatic habitats appear to be derived.
If Nymphaeales are so specialized, one mayask if the ecology of the other basal linesmight be equally unlike that of the first angio-sperms. However, in light of current phylog-enies, the alternative that the features relatedto dark and disturbed habitats arose indepen-dently in Amborella, Austrobaileyales, andChloranthaceae is less parsimonious than theview that they were retained from the first an-giosperms. It is interesting to note, however,that one extant water lily, Barclaya rotundifolia,may have reinvaded wet, shady, disturbed for-est understory habitats (Fig. 2) (Schneider andCarlquist 1995).
Interestingly, there is fossil evidence for ear-ly angiosperm aquatics. Friis et al. (2001) com-pared a Barremian–Aptian flower from Por-tugal with Nymphaeales, but in the absence ofvegetative parts, there is no direct evidence itwas aquatic. More clearly aquatic plants, re-sembling Nymphaeales in having palmatelyveined leaves attached to a short axis, are
known from the Aptian–Albian of Brazil(Mohr and Friis 2000). Another early aquaticis Archaefructus (Sun et al. 1998, 2002), fromthe Barremian–Aptian of China, with a her-baceous habit and finely dissected, featheryleaves. A cladistic analysis placed Archaefruc-tus below all extant angiosperms, raising thepossibility that the aquatic habit is ancestral(Sun et al. 2002). Examination of the data set,however, indicated that it is equally parsimo-nious to place Archaefructus within extant an-giosperms, as an extinct member of Nym-phaeales, and it might also be a less reducedrelative of Ceratophyllum or an extinct basal eu-dicot lineage (Friis et al. 2003). Even if Ar-chaefructus is basal, the ancestral habit wouldstill be equivocal, because the woody plantAmborella is at the next node. Furthermore, thehypothesis that aquatic herbs were ancestralmust deal with the observation that Amborellaand other basal angiosperms (except Nym-phaeales) develop normal secondary vasculartissue, as in other seed plants, which wouldhave to reoriginate in essentially the ancestralform, lacking any of the anomalies usually as-sociated with secondary woodiness.
Chloranthaceae. Our analyses also suggestthat some Chloranthaceae may have beenamong the first terrestrial lineages to ‘‘breakout’’ of the dark and disturbed environment.Unlike Amborella and most Austrobaileyales,Ascarina and Hedyosmum independentlyevolved seedling recruitment in wet, dis-turbed, and higher-light habitats, such ascloud forest land slips, large forest light-gaps,and roadsides (Fig. 2) (Todzia 1988; Martinand Ogden 2002). The ecophysiological traitsunderlying expansion into these zones requirefurther study. However, greater ecologicalflexibility in Chloranthaceae is not linked toherbaceousness. Herbs with limited second-ary xylem appear to be derived (once or twice)within Chloranthus, and these herbaceousgroups occur in shady, disturbed habitats(Fig. 2) (Luo and Li 1999). Hedyosmum orientale,which occurs in sunny, disturbed habitats andhas been described as herbaceous (Verdcourt1986; Todzia 1988), is a possible exception. Re-cent results, however, showed that H. orienta-le’s growth habit and stem vasculature are dif-ferent from those of Chloranthus herbs; plants

93EARLY ANGIOSPERM ECOPHYSIOLOGY
reach 3.5 m in height and produce woody,canelike shoots with up to 3 cm of secondaryxylem (Feild unpublished data 2002).
Consistent with greater ecological ampli-tude, the fossil record indicates that Chloran-thaceae were the first conspicuous and wide-spread extant angiosperm lineage (althoughtheir abundance may be overestimated byhigh pollen production for wind pollination[Endress 1987]). Fossil pollen types compara-ble to Ascarina and Hedyosmum, named ‘‘Cla-vatipollenites’’ and Asteropollis respectively,were abundant in Early Cretaceous lowlandsediments worldwide (Archangelsky andGamerro 1967; Playford 1971; Volkheimer andSalas 1975; Brenner 1996; Walker and Walker1984; Burger 1990, 1993; Archangelsky andTaylor 1993; Dettmann 1994; Hughes 1994; Ek-lund et al. 2004). ‘‘Clavatipollenites’’ is not nec-essarily related to Ascarina, as it represents theancestral pollen type for the family, but Aster-opollis has been associated with Hedyosmum-like fruits (Friis et al. 1997, 2000; Eklund et al.2004). Many Early Cretaceous leaf fossils, alsofound worldwide, show chloranthaceous fea-tures, including chloranthoid teeth, cuticularstriations, and stomata with variable subsidi-ary cell arrangement (Upchurch 1984b, 1995;Pons 1984; Romero and Archangelsky 1986;Upchurch and Dilcher 1990; Cantrill andNichols 1996; Kong 2001). These foliar fea-tures, however, also occur in Amborella andAustrobaileyales and may be plesiomorphic inangiosperms (Upchurch 1984b; Doyle and En-dress 2000; Doyle 2001; Feild et al. 2003b).
Fossil Evidence Bearing on the Hypothesis
Although reconstructing ancestral traits onthe basis of extant phylogeny is a powerful tool,it is desirable to test hypotheses thus generatedwith evidence from the fossil record. In the fol-lowing section, we critically examine existingfossil data bearing on each component of thedark and disturbed hypothesis, and suggestways in which morphological and sedimento-logical observations might be used, in combi-nation, to diagnose ephemeral, shady, and wethabitats (Wing and DiMichele 1992; Arens 1997;Davies-Vollum and Wing 1998; Behrensmeyer etal. 2000). Many modern basal angiospermsgrow on erosional substrates that are unlikely to
be preserved in the sedimentological record(Behrensmeyer et al. 2000). However, streammargins and channels—two disturbed habi-tats—are common terrestrial depositional set-tings (Wing and DiMichele 1992).
Much discussion of early angiosperm ecologywas based on the Potomac Group (Aptian–Al-bian) of eastern North America (e.g., Fontaine1989; Berry 1911; Brenner 1963; Doyle 1969;Wolfe et al. 1975; Doyle and Hickey 1976; Hickeyand Doyle 1977; Upchurch 1984a, b; Crane 1987;Crane et al. 1993, 1994). Doyle and Hickey (1976;Hickey and Doyle 1977) argued that the mor-phology and sedimentary facies associations ofPotomac leaves indirectly supported Stebbins’s(1965, 1974) view that the first angiospermswere disturbance-tolerant species of semiaridtropical environments, as opposed to woodyplants of stable forests. Later, Taylor and Hickey(1990, 1996) suggested that the record in the Po-tomac Group and elsewhere supported a her-baceous origin. However, most aspects of thePotomac record are equally or more consistentwith the dark and disturbed hypothesis.
Fossil Evidence for Light Habitat. Light envi-ronment can be discerned from a variety of leafmorphological features. Low-light environ-ments are believed to favor larger leaves (Giv-nish 1988), although correlations across speciesfor light habitat and leaf size are inconsistent(Niinemets and Kalevi 1994; Ackerly and Don-oghue 1998). At the anatomical scale, spongymesophyll is more strongly correlated with low-light conditions (Smith et al. 1997; Kott and Brit-ton 1985; Arens 1997). Some cuticle features,such as stomata only on the leaf undersurface,large stomata, and low stomatal density, alsoappear linked to shady habitats (Givnish 1988;Smith et al. 1997).
Doyle and Hickey (1976; Hickey and Doyle1977) argued that many lower Potomac angio-sperms were low shrubs of sunny stream-mar-gin habitats, judging from their small leaf size,poor petiole differentiation, and disorganizedleaf venation (plus sedimentary evidence dis-cussed below). Because some of these havechloranthoid features (Upchurch 1984a), itwould not be surprising if they grew in openhabitats. However, there are indications that oth-er angiosperms were in the forest understory. Attwo lower Potomac localities, Fredericksburg

94 TAYLOR S. FEILD ET AL.
and Fish Hut above Dutch Gap, both no longerextant, large angiosperm leaves called Ficophyl-lum occur at low abundance in rich assemblagesof ferns, conifers, and cycadophytes (Fontaine1889; Berry 1911; Hickey and Doyle 1977; Doylepersonal observations on material at the Smith-sonian Institution, 1971). On the basis of this as-sociation, Doyle and Hickey (1976; Hickey andDoyle 1977) concluded that these plants lived inthe shady forest understory. They suggestedthat the understory was one of the first new hab-itats occupied by early angiosperms, but itcould equally well represent the ancestral hab-itat. There is stronger evidence for occupation ofopen habitats in the upper Potomac (Albian),where platanoid leaves are often dominant instream-margin sediments (Hickey and Doyle1977; Crane et al. 1993).
Because leaf size correlates inconsistentlywith light habitat, future work will need to fo-cus on anatomical features of Early Cretaceousangiosperm leaves. Internal anatomy, however,is only rarely preserved in fossil leaves, al-though it is known from permineralized and fu-sainized (charcoalified) material (e.g., Alvin1974; Schabilion and Reihman 1985) and fromcompressions in fine-grained, clay-rich sedi-ments (Chandrasekharam 1972; Gandolfo et al.2001). Leaf cuticle is more commonly preservedbut must be interpreted with care. For example,stomatal density and size may also respond toatmospheric CO2 concentrations (Van de Wateret al. 1994; McElwain and Chaloner 1996; Beer-ling and Royer 2002). To control for this effect,these characters should be considered relative toa broad range of other taxa from the same strati-graphic horizon. No studies of Potomac Groupleaves have yet specifically focused on ecomor-phological correlates with light environment.Upchurch (1984b), however, noted that the sto-mata of many lower Potomac angiosperms werearound 30 mm wide, which is large comparedwith most extant, tropical sun-adapted angio-sperms (Bongers and Popma 1990).
Fossil Evidence for Disturbance. Some mor-phological features can point toward toleranceof disturbance, the most easily quantified beingsmall seed size (discussed above; Harper et al.1970; Grubb 1998; Lusk and Kelly 2003). In ad-dition, evidence for vigorous vegetative repro-duction (Bond and Midgely 2001, 2003), such as
rhizomes with abundant sprouting, could indi-cate adaptation to disturbance. Although it isunusual to find plant fossils complete enough toreveal growth form, such extraordinary fossilsare known (e.g., Mohr and Friis 2000; Sun et al.2002).
Existing seed size data from Barremian–Ap-tian angiosperm floras (Wing and Boucher 1998;Eriksson et al. 2000) are comparable to those inliving basal angiosperms (Tables 1, 2). Conse-quently, the extant data imply that the small sizeof these fossils does not rule out a woody habitor low-light, closed forest habitats, as some haveargued (Taylor and Hickey 1996; Eriksson et al.2000). Instead, small seed size may reflect a gen-eralized adaptation to disturbance (Grubb 1998;Lusk and Kelly 2003).
Previous arguments that early angiospermswere adapted to disturbance focused on the as-sociation of lower Potomac leaves with coarse-grained sediments deposited in or near activechannels (Doyle and Hickey 1976; Hickey andDoyle 1977; Crane 1987). At the Fredericksburglocality, plant fossils occurred in a lens of finesand in a sand and gravel sequence (Fontaine1889), suggesting rapid filling of an abandonedchannel in a braided stream system. The matrixfrom Fish Hut above Dutch Gap is darker, or-ganic-rich clay containing significant sand andmica (Doyle personal observations, 1971), sug-gesting a crevasse splay, where sand was de-posited on the floodplain after a levee break.Similar deposits occur at Dutch Gap Canal, ca.1.5 km downstream (Upchurch and Doyle1981). At Drewrys Bluff, angiosperm leaves oc-cur in sands (Doyle and Hickey 1976) and a thinclay lens, apparently representing a channel fill(Upchurch 1984a; Crane and Upchurch 1987).
Crevasse-splay deposits such as those atDutch Gap potentially offer ‘‘snapshots’’ of un-derstory microsites on the vegetated floodplain.If autochthonous preservation can be demon-strated (e.g., Calder et al. 1996), such depositsmay allow cryptic disturbed understory mi-crosites to be identified in the fossil record. Fu-ture work should focus on more detailed sedi-mentological analyses of early angiosperm flo-ras, combining bed form observations withgrain size distributions and mineralogy. Thismay allow for more confident distinction ofephemeral substrates from more stable habitats

95EARLY ANGIOSPERM ECOPHYSIOLOGY
(Wing and DiMichele 1992; Davies-Vollum andWing 1998; Davies-Vollum 1999; Behrensmeyeret al. 2000; Royer et al. 2003).
Fossil Evidence for Wet Conditions. Leaf andcuticle characters can often discriminate be-tween wet and dry habitats (Wing and Di-Michele 1992; Wolfe 1993; Wilf et al. 1998). Forinstance, leaf size correlates well with moistureregime (Givnish 1979; Wilf et al. 1998), althoughinfluences of whole-plant branching pattern aswell as flower and fruit placement can occasion-ally override this relation (Bond and Midgely1988; Ackerly and Donoghue 1998). Thick cuti-cles, sunken stomata, and stomata protected byvestibules, papillae, or trichomes are generallyindicative of dry conditions (Wing and Di-Michele 1992; Upchurch 1995). In some cases,however, these features may reflect high herbiv-ory, low nutrient conditions, and extremely highrainfall (e.g., cloud forests [Feild et al. 1998,2003a; see also Wing and DiMichele 1992]).
Doyle and Hickey (1976; Hickey and Doyle1977) argued that the Potomac record support-ed a semiarid origin of the angiosperms (Steb-bins 1974). However, this was based not on ev-idence of aridity in the Potomac area, but ratheron the supposition that the disturbed streammargins where the plants were preserved werethe habitats most likely to be occupied by‘‘weedy’’ immigrants from dry areas. Both leaffloras and paleoclimatic models imply that thePotomac Group climate was moist and subtrop-ical (Upchurch and Wolfe 1987; Beerling andWoodward 2001). The angiosperms range fromsmall leaves with chloranthoid teeth to largerleaves of the Ficophyllum type. Most of the taxastudied anatomically have thin cuticles and sto-mata in the plane of the leaf surface, except Eu-calyptophyllum, which has a thicker cuticle andsunken stomata (Upchurch 1984a,b), traits likelyrelated to broadly mesic conditions (Wing andDiMichele 1992; Feild et al. 1998). Similaritiesbetween these cuticles and those of modern bas-al angiosperms (Upchurch 1984a,b) also sug-gest that they were ecologically comparable.
Several authors took the diversity of early an-giosperm pollen in Northern Gondwana, cou-pled with dominance of xeromorphic Cheirole-pidiaceae and Gnetales, rarity of fern spores,and salt deposits, as support for a semiarid or-igin (Brenner 1976; Doyle et al. 1977; Hickey and
Doyle 1977). However, angiosperms were as ormore diverse in northern South America andthe Middle East, where there are fewer Cheiro-lepidiaceae, more ferns, and coal deposits, in-dicating a wetter climate (Doyle et al. 1982;Schrank 1983, 1990; Brenner 1996). Climatemodels also predict higher rainfall in these areas(Parrish 1987; Beerling and Woodward 2001).Barremian-Aptian angiosperm leaves from Co-lombia (Pons 1984) are smaller than Ficophyllum,but they overlap lower Potomac angiospermleaves in size and other features (unfortunately,no cuticle was preserved). The Colombian florais dominated by matoniaceous ferns (Weichselia),with some cycadophytes and broad-leaved co-nifers (Pons 1984); this flora represents the wetextreme in Northern Gondwana. Together theseobservations indicate that early angiosperms oc-curred in both wet and dry tropical areas, andit would be premature to assume they originat-ed in one environment rather than the other.
Interpretations of whole-plant water balancederived from cuticular anatomy of ancientplants would be strengthened by independentestimates of leaf gas-exchange performance. Sta-ble carbon isotopic composition (d13C, the ratioof 13CO2 to 12CO2) is the most promising proxyfor leaf water-use efficiency (Farquhar et al.1982; Condon et al. 1993). Using well-under-stood relationships from modern angiosperms,it should be possible to apply d13C measure-ments of fossil cuticles to infer relative leaf wa-ter-use efficiency of early angiosperms within afossil assemblage. Although the absolute d13Cvalue of an individual fossil can be influencedby a variety of factors, including ecology, theisotopic composition of atmospheric CO2, andtaphonomy (Arens et al. 2000; Arens and Jahren2000, 2002; Beerling and Royer 2002), samplinga range of taxa within an assemblage may allowseriation of plants along a gradient of water-useefficiency. For example, Nguyen Tu et al. (1999)used d13C of fossil cuticles to infer leaf water-useefficiency of angiosperms, conifers, cycads, andginkgoes and ordinate them along a salinitygradient.
Implications for the Early Evolution ofAngiosperms
The dark and disturbed hypothesis providesa new perspective on the selective environment

96 TAYLOR S. FEILD ET AL.
in which early angiosperms evolved. Stebbins(1965, 1974) hypothesized that the distinctivefeatures of angiosperms arose under seasonallydry climates and unstable habitats, where rapidreproduction and high vegetative flexibilitywere favored. This view has been variouslymodified by other authors (Axelrod 1970; Doyleand Hickey 1976; Hickey and Doyle 1977; Doyle1978; Doyle and Donoghue 1986, 1993; Crane etal. 1995; Taylor and Hickey 1996). Our analysisraises another possibility: Key angiosperm traits(e.g., vessels, reticulate leaf venation, and thecarpel) may be linked to low light and highmoisture availability, coupled with frequent dis-turbance.
Current phylogenies imply that vesselswere absent in the first angiosperms (Ambor-ella is vesselless, and Nymphaeales are ves-selless or have cells intermediate between tra-cheids and vessel elements [Schneider et al.1995; Feild et al. 2000]). Vessels, however,probably appeared early in angiosperm evo-lution, as Austrobaileyales and Chlorantha-ceae are vessel-bearing (Doyle and Endress2000; Carlquist and Schneider 2002). Previousinterpretations of vessel origin in angio-sperms emphasized increased hydraulic effi-ciency and rapid transpiration in open, trop-ical environments (Doyle et al. 1982; Doyleand Donoghue 1986). In contrast, we proposethat vessels confer disturbance tolerance inshady understory habitats, where evaporativedemand is low. By providing a more openpath for water moving through the stem, ves-sels allow a given amount of leaf area to besupported at lower shoot construction cost, asfound especially in basal angiosperm under-story lianas (Brodribb and Feild 2000; Feild etal. 2000, 2003a; Sperry 2003). This interpreta-tion is bolstered by vessel structure in basaltaxa such as Austrobaileyales and some Chlor-anthaceae, which lack features maximizinghydraulic conductivity, such as simple perfo-ration plates (some Schisandra species are anexception [Carlquist 1999]) and completelyopen pit membranes (Carlquist and Schneider2002). Thus, the initial selection pressures forvessels may have been increased vegetativegrowth efficiency under carbon-limited (sha-dy), humid conditions, rather than water
stress in dry climates or under high transpi-ration load in full sun.
Reticulate leaf venation may also providephysiological advantages in forest understories(Givnish 1979; Roth-Nebelsick et al. 2001). Byincreasing within-leaf hydraulic redundancyand structural support, reticulate venation mayhave facilitated the evolution of larger photo-synthetic surfaces capable of more efficient useof understory sunflecks.
Envisioning early angiosperms in a humidclimate also has implications for the origin of thecarpel. Carpels in Amborella, Nymphaeales,Austrobaileyales, and Chloranthaceae are urn-like (ascidiate) and not sealed by postgenital(cellular) fusion (Endress 2001). Instead, theyare ‘‘plugged’’ to varying degrees by mucilage-like pectin and arabinogalactan secretions(Bernhardt et al. 2003; T. L. Sage unpublisheddata 2003 for Amborella and Illicium). Althoughthese extracellular polysaccharides are stronglyhydrophilic, water stored in them is easily lib-erated at low xylem tensions (Morse 1990).Thus, this mode of carpel closure may offer littleprotection from drought. Furthermore, suchcarpels would have promoted outbreeding oracted as effective shields against fungal invasionand/or insect damage (Stebbins 1974; Doyleand Donoghue 1986) only under conditions oflocally high humidity (e.g., forest understories,cloud forests, or aquatic habitats). Interestingly,many of the oldest known fossil angiospermflowers appear to have ascidiate carpels (Friis etal. 1999, 2000; Doyle 2001).
Selective Pressures and Diversification. The lowspecies richness and ecophysiological variationof extant basal lineages (Fig. 1), combined withthe rarity of angiosperm fossils before the Bar-remian–Aptian, suggest that the traits promot-ing angiosperm diversification arose after theorigin of the crown group (i.e., the common an-cestor of all extant taxa; Doyle and Donoghue1993; Sanderson and Donoghue 1994; Magallonand Sanderson 2001). Instead of being the keyto diversification, the dark and disturbed life-style may have been a way for angiosperms togain a root-hold in well-established Mesozoicplant communities.
Greater architectural flexibility and disper-sion of meristems may have offered early angio-sperms a crucial advantage in disturbed under-

97EARLY ANGIOSPERM ECOPHYSIOLOGY
story habitats. For instance, rhizomatous growthand lianoid habits, with heavy reliance on veg-etative propagation (Table 3), increase the dis-covery and exploitation of ephemeral, resource-rich patches in the forest understory (Cook1983). Multistemmed growth, characterized bylarge ‘‘bud banks’’ for shoot iteration followingdamage, also allows long-term persistence indisturbed habitats (Greig 1993; Bond and Mid-gely 2001, 2003). With evolution of vessels, car-bon costs for shoot and root reiteration mayhave decreased (see above). In contrast, thesquat, unbranched, sparsely leaved forms ofmany Bennettitales (e.g., Cycadeoidea; Crane1985) and cycads were probably less able to usepatchy understory resources and may have beenat greater risk of fatal damage from limbfall dis-turbance. Extant understory cycads (e.g., Zamia)recover slowly from limbfall-induced shootdamage because they have a limited capacity toresprout and add new leaves slowly (Clark andClark 1988; Clark et al. 1992). In their relianceon vegetative growth, early angiosperms mayhave been more like ferns than other seedplants. Flexible growth and varied regenerationmodes (vegetative as well as sexual) may havebeen particularly advantageous in understoriesdisturbed by dinosaurs, a significant agent inMesozoic forests (Bakker 1978; Wing and Tiff-ney 1987; Barrett and Willis 2001).
If the dark and disturbed ecology did notpromote diversity, later innovations may havebeen responsible for spurring angiosperm ra-diation into a wide range of new habitats. Asangiosperms broke out of the understory, wesuggest that vessels and reticulate leaf vena-tion were co-opted for new adaptive roles.Modifications of vessels (e.g., longer function-al length and simple perforation plates joiningvessel elements) increased hydraulic conduc-tivity. This vascular system, coupled with in-creasing vein density, could support the high-er transpiration rates required to dissipateheat in sunny environments and enable theevolution of higher photosynthetic rate. Thus,the exaptation of vessels for sunnier and/ordrier landscapes may be more closely linkedto the explosive diversification of angio-sperms in the Albian and Late Cretaceous. Inturn, these new habitats would have exposedangiosperms to many new pressures for
changes in pollination and dispersal mode (cf.Doyle et al. 2003).
Pre-Cretaceous Angiosperm History. Our hy-pothesis also bears on one of the most vexingquestions in paleobotany: Where was the angio-sperm line before the Cretaceous? Axelrod(1952, 1970) argued that crown-group angio-sperms originated and diversified crypticallybefore the Cretaceous. This scenario, however, isunlikely given the primitive character suites ofEarly Cretaceous angiosperms and their mor-phological diversification through the period(Doyle 1969, 1977, 1978, 2001; Doyle and Hickey1976; Doyle and Donoghue 1993; Crane et al.1995). If crown-group angiosperms existed ear-lier, they must have been rare and plesiomorph-ic. In this case, features essential for angiospermdiversification arose after the origin of thecrown group, unless some extrinsic factor sup-pressed diversification of the clade (Doyle andDonoghue 1993). The dark and disturbed hy-pothesis provides a plausible scenario: Low-di-versity basal groups were initially restricted todark, wet, and disturbed forest understorieswhere population sizes were low. Later, Chlor-anthaceae and core angiosperms evolved great-er sun-tolerance and diversified both phyloge-netically and in habitat preference.
Significantly, everwet tropical climates wererare in the Triassic and Jurassic. Both sedi-mentological evidence and climate modelssuggest that atmospheric circulation in tropi-cal latitudes was strongly monsoonal (Parrishet al. 1982; Ziegler et al. 1987; Barron et al.1994; Rees et al. 2000; Beerling and Woodward2001). In the Early Cretaceous tropics, paleo-botanical and sedimentary evidence and cli-mate models indicate that mesic climates wererestricted to northern South America, theMiddle East, and Southeast Asia (Parrish1987; Doyle et al. 1982; Barron et al. 1994;Doyle 1999; Rees et al. 2000; Beerling andWoodward 2001). If wet tropical habitats wererare, it becomes more plausible that pre-Cre-taceous angiosperms, or their immediate pre-cursors, escaped detection. However, unlikethe upland theory of Axelrod (1952, 1970) orthe semiarid theory of Stebbins (1965, 1974),the dark and disturbed hypothesis is testableby study of known and yet-to-be-discoveredfossil floras of appropriate age.

98 TAYLOR S. FEILD ET AL.
Conclusion
Cogent arguments on the forces driving an-giosperm origin and diversification dependon an accurate picture of early angiospermecology. To develop such a picture, wemapped ecophysiological data from extantbasal angiosperms onto their phylogeny to re-construct the ecology of their common ances-tors. This analysis yielded an ecological hy-pothesis of the first angiosperms that is subtly,but importantly, different from previousideas: Angiosperms may have first occupiedshaded, disturbed, and possibly wet under-story habitats.
Acknowledgments
We are grateful for the generous help andsupport from a large number of people whohave made this global research project possible.In Fiji, we thank A. Jokan and M. Tuiwawa (Uni-versity of South Pacific, Suva) for help in re-search logistics and establishing local contacts.We thank of J. Turaga (Bouma village) for aid-ing sevusevu rites to conduct fieldwork on Tav-euni. We also appreciate help from T. Jaffre(IRD, Noumea, New Caledonia), P. J. Franks(James Cook University, Cairns, Australia), J.-Y.Meyer (French Polynesia), M. Turnbull and M.McGlone (University of Canterbury, New Zea-land), and R. M. K. Saunders (University ofHong) in coordinating fieldwork and helpfuldiscussions. H.-Z. Kong and Y.-B. Luo at the In-stitute for Botany in Beijing arranged permis-sion to conduct fieldwork in China, and wethank L.-B. Luo and family for their help in lo-cating plants and hospitality in Xinning Co. InPapua New Guinea, we acknowledge S. Saulei(University of Papua New Guinea), L. Balun(Forestry College, Bulolo), and R. Kiprianis (LaeHerbarium) for aiding fieldwork on Trimenia.We also appreciate experimental advice andfield assistance in Costa Rica and New Caledon-ia from T. Brodribb (Harvard University). Wealso thank several people for discussions thathave led to the ideas presented here: D. D. Ack-erly, B. Baldwin, D. Baum, K. Boyce, S. Burgess,N. M. Holbrook, S. Mathews, B. Mishler, T. Sage,S. Patek, S. Poe, and C. Webb. We acknowledgeP. S. Herendeen, S. Wing, L. J. Hickey, D. W. Tay-lor, and especially G. R. Upchurch for additional
helpful discussions on the Early Cretaceous an-giosperm fossil record. P. S. Herendeen and S.Wing made helpful suggestions for improve-ment of the manuscript. Special thanks go to D.S. Miller for permission to collect plants of Schis-andra glabra in Florida, L. Lowman (RidgecrestNursery), G. Williams-Lineara as well as E. V. J.Tanner for providing some additional ecologicalobservations on Hedyosmum. Finally, we thankH. S. Feild and C. T. Feild for constructing andmaintaining the experimental Illicium and Schis-andra common garden in Auburn, Alabama, aswell as E. Feild for field assistance in Costa Rica,watering plants in Berkeley, and all around sup-port in the many long months of fieldwork forthis project. This research was funded by a dis-covery grant from the Natural Sciences and En-gineering Research Council of Canada and a re-search fellowship to T. S. Feild from the MillerInstitute for Basic Research in Science at the Uni-versity of California, Berkeley. Research wasalso facilitated by support from the National Sci-ence Foundation Deep Time Research Coordi-nation Network (RCN0090283).
Literature Cited
Ackerly, D. D., and M. J. Donoghue. 1998. Leaf size, sapling al-lometry, and Corner’s rules: phylogeny and correlated evo-lution in maples (Acer). American Naturalist 152:767–791.
Alvin, K. L. 1974. Leaf anatomy of Weichselia based on fusaini-zed material. Palaeontology 17:587–598.
Arber, E. A. N., and J. Parkin. 1907. On the origin of angio-sperms. Journal of the Linnean Society (Botany) 38:29–80.
Archangelsky, S., and J. C. Gamerro. 1967. Spore and pollentypes of the Lower Cretaceous in Patagonia (Argentina). Re-view of Palaeobotany and Palynology 1:211–217.
Archangelsky, S., and T. N. Taylor. 1993. The ultrastructure of insitu Clavatipollenites pollen from the Early Cretaceous of Pa-tagonia. American Journal of Botany 80:879–885.
Arens, N. C. 1997. Responses of leaf anatomy to light environ-ment in the tree fern Cyathea caracasana (Cyatheaceae) and itsapplication to some ancient seed ferns. Palaios 12:84–94.
———. 2001. Variation in performance of the tree fern Cyatheacaracasana (Cyatheaceae) across the successional mosaic of anAndean cloud forest. American Journal of Botany 88:545–551.
Arens, N. C., and A. H. Jahren. 2000. Carbon isotope excursionin atmospheric CO2 at the Cretaceous-Tertiary boundary: ev-idence from terrestrial sediments. Palaios 15:314–322.
———. 2002. Chemostratigraphic correlation of four fossil-bear-ing sections in southwestern North Dakota. Geological Soci-ety of America Special Paper 361:75–93.
Arens, N. C., A. H. Jahren, and R. Amundson. 2000. Can C3
plants faithfully record the carbon isotopic composition of at-mospheric carbon dioxide? Paleobiology 26:137–164.
Axelrod, D. I. 1952. A theory of angiosperm evolution. Evolution6:29–60.
———. 1970. Mesozoic paleogeography and early angiospermhistory. Botanical Review 36:277–319.

99EARLY ANGIOSPERM ECOPHYSIOLOGY
Bakker, R. T. 1978. Dinosaur feeding behaviour and the originof flowering plants. Nature 274:661–663.
Barkman, T. J., G. Chenery, J. R. McNeal, J. Lyons-Weiler, W. J.Ellisens, G. Moore, A. D. Wolfe, and C. W. de Pamphilis. 2000.Independent and combined analyses of sequences from allthree genomic compartments converge on the root of flow-ering plant phylogeny. Proceedings of the National Academyof Sciences USA 97:13166–13171.
Barrat-Segretain, M.-H. 1996. Germination and colonization dy-namics of Nuphar lutea (L.) Sm. in a former river channel.Aquatic Botany 55:31–38.
Barrett, P. M, and K. J. Willis. 2001. Did dinosaurs invent flow-ers? Dinosaur-angiosperm coevolution revisited. BiologicalReviews 76:411–447.
Barron, E. J., P. J. Fawcett, D. Pollard, and S. L. Thompson. 1994.Model simulations of Cretaceous climates: the role of geog-raphy and carbon dioxide. Pp. 99–108 in J. R. L. Allen, B. J.Hoskins, B. W. Sellwood, R. A. Spicer, and P. J. Valdes, eds.Palaeoclimates and their modelling: with special reference tothe Mesozoic Era. Chapman and Hall, London.
Beerling, D. J., and D. L. Royer. 2002. Fossil plants as indicatorsof the Phanerozoic global carbon cycle. Annual Review ofEarth and Planetary Sciences 30:527–556.
Beerling, D. J., and F. I. Woodward. 2001. Vegetation and the ter-restrial carbon cycle: modelling the first 400 million years.Cambridge University Press, Cambridge.
Behrensmeyer, A. K., S. M. Kidwell, and R. A. Gastaldo. 2000.Taphonomy and paleobiology. Paleobiology 26:103–147.
Bernhardt, P., T. L. Sage, P. Weston, H. Azuma, M. Lam, L. B.Thien, and J. Bruhl. 2003. The pollination of Trimenia moorei(Trimeniaceae): floral volatiles, insect/wind pollen vectors,and stigmatic self-incompatibility in a basal angiosperm. An-nals of Botany 92:445–458.
Berry, E. W. 1911. Systematic paleontology, Lower Cretaceous,Pteridophyta-Dicotyledonae. Pp. 214–508 in W. B. Clark, ed.Maryland Geological Survey, Lower Cretaceous. Johns Hop-kins University Press, Baltimore.
Bews, J. W. 1927. Studies in the ecological evolution of angio-sperms. New Phytologist 26:1–21.
Bilger, W., U. Schreiber, and M. Bock. 1995. Determinants of thequantum efficiency of photosystem II and nonphotochemicalquenching of chlorophyll fluorescence in the field. Oecologia102:425–432.
Bjorkman, O., and B. Demmig. 1987. Photon yield of O2 evolu-tion and chlorophyll fluorescence characteristics at 77 Kamong vascular plants of diverse origins. Planta 170:489–504.
Blanc, P. 1986. Edification d’arbres par croissanced’etablissement de type monocotyledonien: l’exemple deChloranthaceae. Pp. 101–123 in Colloque international surl’arbre 1986. Naturalia Monspeliensia, numero hors serie,Montpellier.
Bond, W. J., and J. J. Midgely. 1988. Allometry and sexual dif-ferences in leaf size. American Naturalist 131:901–910.
———. 2001. Ecology of sprouting in woody plants: the persis-tence niche. Trends in Ecology and Evolution 16:45–51.
———. 2003. The evolutionary ecology of sprouting. Interna-tional Journal of Plant Sciences 164(Suppl.):S103–S114.
Bongers, F., and J. Popma. 1990. Leaf characteristics of the trop-ical rain forest flora of Los Tuxtlas, Mexico. Botanical Gazette151:354–365.
Brenner, G. J. 1963. The spores and pollen of the Potomac Groupof Maryland. Maryland Department of Geology, Mines andWater Resources Bulletin 27:1–215.
———. 1976. Middle Cretaceous floral provinces and early mi-grations of angiosperms. Pp. 23–47 in C. B. Beck, ed. Origin
and early evolution of angiosperms. Columbia UniversityPress, New York.
———. 1996. Evidence for the earliest stage of angiosperm pol-len evolution: a paleoequatorial section from Israel. Pp. 91–115 in D. W. Taylor and L. J. Hickey, eds. Flowering plant or-igin, evolution and phylogeny. Chapman and Hall, New York.
Brodribb, T. J., and T. S. Feild. 2000. Stem hydraulic supply islinked to leaf photosynthetic capacity: evidence from NewCaledonian and Tasmanian rainforests. Plant, Cell and En-vironment 23:1381–1388.
Brodribb, T., and R. S. Hill. 1997. Light response characteristicsof a morphologically diverse group of southern hemisphereconifers as measured by chlorophyll fluorescence. Oecologia110:10–17.
Brodribb, T. J., N. M. Holbrook, and M. V. Gutierrez. 2002. Hy-draulic and photosynthetic coordination in seasonally drytropical forest. Plant, Cell and Environment 25:1435–1444.
Burger, D. 1990. Early Cretaceous angiosperms from Queens-land, Australia. Review of Palaeobotany and Palynology 65:153–163.
———. 1993. Early and middle Cretaceous angiosperm pollengrains from Australia. Review of Palaeobotany and Palynol-ogy 78:183–234.
Calder, J. H., M. R. Gibling, C. F. Eble, A. C. Scott, and D. J. Mac-Neil. 1996. The Westphalian D fossil lepidodendrid forest atTable Head, Sydney Basin, Nova Scotia: sedimentology, pa-leoecology and floral response to changing edaphic condi-tions. International Journal of Coal Geology 31:277–313.
Canham, C. D., R. K. Kobe, E. F. Latty, and R. L. Chazdon. 1999.Interspecific and intraspecific variation in tree seedling sur-vival: effects of allocation to roots versus carbohydrate re-serves. Oecologia 121:1–11.
Cantrill, D. J., and G. J. Nichols. 1996. Taxonomy and paleo-ecology of Early Cretaceous (Late Albian) angiosperm leavesfrom Alexander Island, Antarctica. Review of Palaeobotanyand Palynology 92:1–28.
Carlquist, S. 1975. Ecological strategies of xylem evolution. Uni-versity of California Press, Berkeley.
———. 1984. Wood anatomy of Trimeniaceae. Plant Systematicsand Evolution 144:103–118.
———. 1987. Presence of vessels in wood of Sarcandra (Chlor-anthaceae): comments on vessel origin in angiosperms.American Journal of Botany 74:1765–1771.
———. 1992a. Wood anatomy and stem of Chloranthus; summaryof wood anatomy of Chloranthaceae, with comments on re-lationships, vessellessness, and the origin of monocotyledons.International Association of Wood Anatomists Bulletin 13:3–16.
———. 1992b. Wood anatomy of Hedyosmum (Chloranthaceae)and the tracheid-vessel element transition. Aliso 13:447–462.
———. 1999. Wood and bark anatomy of Schisandraceae: im-plications for phylogeny, habit, and vessel evolution. Aliso 18:45–55.
Carlquist, S., and E. L. Schneider. 2002. The tracheid-vessel el-ement transition in angiosperms involves multiple indepen-dent features: cladistic consequences. American Journal ofBotany 89:185–195.
Chandrasekharam, A. 1972. Spongy mesophyll remains in fossilleaf compressions. Science 177:354–356.
Chazdon, R. L. 1988. Sunflecks and their importance to forestunderstory plants. Advances in Ecological Research 18:1–63.
Chazdon, R. L., R. W. Pearcy, D. W. Lee, and N. Fetcher. 1996.Photosynthetic responses of tropical forest plants to contrast-ing light environments. Pp. 5–55 in S. S. Mulkey, R. L. Chaz-don, and A. P. Smith, eds. Tropical forest plant ecophysiology.Chapman and Hall, London.

100 TAYLOR S. FEILD ET AL.
Clark, D. B., and D. A. Clark. 1988. Leaf production and the costof reproduction in the neotropical rain forest cycad, Zamiaskinneri. Journal of Ecology 76:1153–1163.
Clark, D. B., D. A. Clark, and M. H. Grayum. 1992. Leaf demog-raphy of a neotropical rain forest cycad, Zamia skinneri (Za-miaceae). American Journal of Botany 79:28–33.
Condon, A. G., R. A. Richards, and G. D. Farquhar. 1993. Rela-tionships between carbon isotopic discrimination, water useefficiency, and transpiration efficiency for dryland wheat.Australian Journal of Agricultural Research 44:1693–1711.
Cook, R. E. 1983. Clonal plant populations. American Scientist71:244–253.
Crane, P. R. 1985. Phylogenetic analysis of seed plants and theorigin of angiosperms. Annals of the Missouri Botanical Gar-den 72:716–793.
———. 1987. Vegetational consequences of the angiosperm di-versification. Pp. 107–144 in E. M. Friis, W. G. Chaloner, andP. R. Crane, eds. The origins of angiosperms and their biolog-ical consequences. Cambridge University Press, Cambridge.
Crane, P. R., and S. Lidgard. 1989. Angiosperm diversificationand paleolatitudinal gradients in Cretaceous floristic diver-sity. Science 246:675–678.
Crane, P. R., and G. R. Upchurch. 1987. Drewria potomacensis gen.et sp. nov., an Early Cretaceous member of Gnetales from thePotomac Group of Virginia. American Journal of Botany 74:1722–1736.
Crane, P. R., K. R. Pedersen, E. M. Friis, and A. N. Drinnan. 1993.Early Cretaceous (early to middle Albian) platanoid inflores-cences associated with Sapindopsis leaves from the PotomacGroup of eastern North America. Systematic Botany 18:328–344.
Crane, P. R., E. M. Friis, and K. R. Pedersen. 1994. Paleobotanicalevidence on the early radiation of magnoliid angiosperms.Plant Systematics and Evolution 8(Suppl.):51–72.
———. 1995. The origin and early diversification of angio-sperms. Nature 374:27–33.
Cronquist, A. 1988. The evolution and classification of floweringplants, 2d ed. New York Botanical Garden, New York.
Dacey, J. W. H. 1980. Internal winds in water lilies: an adaptationfor life in anaerobic sediments. Science 210:1017–1019.
Dai, N., S. Kenji, and A. Sakai. 2002. Seedling establishment ofdeciduous trees in various topographic positions. Journal ofVegetation Science 13:35–44.
Davies-Vollum, K. S. 1999. The formation of beds underlyingcarbonaceous shales in aquic paleosols: examples from theBighorn Basin of Wyoming. International Journal of Coal Ge-ology 41:239–255.
Davies-Vollum, K. S., and S. L. Wing. 1998. Sedimentological,taphonomic, and climatic aspects of Eocene swamp deposits(Willwood Formation, Bighorn Basin, Wyoming). Palaios 13:28–40.
Demmig-Adams, B., W. W. Adams, B. Logan, and A. Verhoeven.1995. Xanthophyll cycle dependent energy dissipation andflexible photosystem II efficiency in plants acclimated to lightstress. Australian Journal of Plant Physiology 22:249–260.
Dettmann, M. E. 1994. Cretaceous vegetation: the microfossil re-cord. Pp. 143–188 in R. S. Hill, ed. History of the Australianvegetation. Cambridge University Press, Cambridge.
Donoghue, M. J., and J. A. Doyle. 1989. Phylogenetic analysis ofangiosperms and the relationships of Hamamelidae. Pp. 14–45 in P. R. Crane and S. Blackmore, eds. Evolution, system-atics, and fossil history of the Hamamelidae, Vol. 1. Claren-don, Oxford.
Doyle, J. A. 1969. Cretaceous angiosperm pollen of the AtlanticCoastal Plain and its evolutionary significance. Journal of theArnold Arboretum 50:1–35.
———. 1977. Patterns of evolution in early angiosperms. Pp.501–546 in A. Hallam, ed. Patterns of evolution. Elsevier, Am-sterdam.
———. 1978. Origin of angiosperms. Annual Review of Ecologyand Systematics 9:365–392.
———. 1999. The rise of angiosperms as seen in the African Cre-taceous pollen record. In K. Heine, ed. Palaeoecology of Af-rica 26:3–29.
———. 2001. Significance of molecular phylogenetic analysesfor paleobotanical investigations on the origin of angio-sperms. Palaeobotanist 50:167–188.
Doyle, J. A., and M. J. Donoghue. 1986. Seed plant phylogenyand the origin of angiosperms: an experimental cladistic ap-proach. Botanical Review 52:321–431.
———. 1993. Phylogenies and angiosperm diversification. Pa-leobiology 19:141–167.
Doyle, J. A. and P. K. Endress. 2000. Morphological phylogeneticanalysis of basal angiosperms: comparison and combinationwith molecular data. International Journal of Plant Sciences161(Suppl.):S121–S153.
Doyle, J. A., and L. J. Hickey. 1976. Pollen and leaves from themid-Cretaceous Potomac Group and their bearing on earlyangiosperm evolution. Pp. 139–206 in C. B. Beck, ed. Originand early evolution of angiosperms. Columbia UniversityPress, New York.
Doyle, J. A., P. Biens, A. Doerenkamp, and S. Jardine. 1977. An-giosperm pollen from the pre-Albian Cretaceous of Equato-rial Africa. Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine 1:451–473.
Doyle, J. A., S. Jardine, and A. Doerenkamp. 1982. Afropollis, anew genus of early angiosperm pollen, with notes on the Cre-taceous palynostratigraphy and paleoenvironments of North-ern Gondwana. Bulletin des Centres de Recherches Explora-tion-Production Elf-Aquitaine 6:39–117.
Doyle, J. A., M. J. Donoghue, and E. A. Zimmer. 1994. Integra-tion of morphological and ribosomal RNA data on the originof angiosperms. Annals of the Missouri Botanical Garden 81:419–450.
Doyle, J. A., H. Eklund, and P. S. Herendeen. 2003. Floral evo-lution in Chloranthaceae: implications of a phylogenetic anal-ysis of morphological characters. International Journal ofPlant Sciences 164(Suppl.):S365–S382.
Edwards, G. E., and N. R. Baker. 1993. Can assimilation in maizeleaves be predicted accurately from chlorophyll fluorescenceanalysis? Photosynthesis Research 37:89–102.
Eklund, H., J. A. Doyle, and P. S. Herendeen. 2004. Morpholog-ical phylogenetic analysis of living and fossil Chloranthaceae.International Journal of Plant Sciences (in press).
Endress, P. K. 1987. The Chloranthaceae: reproductive struc-tures and phylogenetic position. Botanische Jahrbucher furSystematik 109:153–226.
———. 2001. The flowers in extant basal angiosperms and in-ferences on ancestral flowers. International Journal of PlantSciences 162:1111–1140.
Eriksson, O., E. M. Friis, and P. R. Crane. 2000. Seed size, fruitsize, and dispersal systems in angiosperms from the EarlyCretaceous to the Late Tertiary. American Naturalist 156:47–58.
Esau, K. 1977. Anatomy of seed plants. Wiley, New York.Farquhar, G. D., M. H. O’Leary, and J. A. Berry. 1982. On the
relationship between carbon isotopic discrimination and theintercellular carbon dioxide concentration in leaves. Austra-lian Journal of Plant Physiology 9:121–137.
Feild, T. S., M. A. Zwieniecki, M. J. Donoghue, and N. M. Hol-brook. 1998. Stomatal plugs of Drimys winteri (Winteraceae)

101EARLY ANGIOSPERM ECOPHYSIOLOGY
protect leaves from mist but not drought. Proceedings of theNational Academy of Sciences USA 95:14256–14259.
Feild, T. S., T. Brodribb, T. Jaffre, and N. M. Holbrook. 2001. Re-sponses of leaf anatomy, photosynthetic light-use, and xylemhydraulics to light in the basal angiosperm Amborella tricho-poda (Amborellaceae). International Journal of Plant Sciences162:999–1008.
Feild, T. S., M. A. Zwieniecki, T. Brodribb, T. Jaffre, M. J. Don-oghue, and N. M. Holbrook. 2000. Structure and function oftracheary elements in Amborella trichopoda. International Jour-nal of Plant Sciences 161:705–712.
Feild, T. S., P. J. Franks, and T. L. Sage. 2003a. The shade-adaptedecophysiology of Austrobaileya scandens. International Journalof Plant Sciences 164:313–324.
Feild, T. S., N. C. Arens, and T. E. Dawson. 2003b. The ancestralecology of flowering plants: emerging perspectives from ex-tant basal lineages. International Journal of Plant Sciences164(Suppl.):S129–S142.
Fontaine, W. M. 1889. The Potomac or Younger Mesozoic flora.U.S. Geological Survey Monograph 15.
Forbis, T. A., S. K. Floyd, and A. de Queiroz. 2002. The evolutionof embryo size in angiosperms and other seed plants: impli-cations for the evolution of seed dormancy. Evolution 56:2112–2125.
Franco, A. C., and U. Luttge. 2002. Midday depression in savan-na trees: coordinated adjustments in photochemical efficien-cy, photorespiration, CO2 assimilation, and water-use efficien-cy. Oecologia 131:356–365.
Friis, E. M., P. R. Crane, and K. R. Pedersen. 1997. Fossil historyof magnoliid angiosperms. Pp. 121–156 in K. Iwatsuki and P.R. Raven, eds. Evolution and diversification of land plants.Springer, Tokyo.
Friis, E. M., K. R. Pedersen, and P. R. Crane. 1999. Early angio-sperm diversification: the diversity of pollen associated withangiosperm reproductive structures in Early Cretaceous flo-ras from Portugal. Annals of the Missouri Botanical Garden86:259–296.
———. 2000. Reproductive structure and organization of basalangiosperms from the Early Cretaceous (Barremian or Ap-tian) of western Portugal. International Journal of Plant Sci-ences 161(Suppl.):S169–S182.
———. 2001. Fossil evidence of water lilies (Nymphaeales) in theEarly Cretaceous. Nature 410:357–360.
Friis, E. M., J. A. Doyle, P. K. Endress, and Q. Leng. 2003. Ar-chaefructus: angiosperm precursor or specialized early angio-sperm? Trends in Plant Science 8:369–373.
Gandolfo, M. A., K. C. Nixon, and W. L. Crepet. 2001. TuronianPinaceae of the Raritan Formation, New Jersey. Plant System-atics and Evolution 226:187–203.
Gavin, D. G., and D. R. Peart. 1999. Vegetative life history of adominant rain forest canopy tree. Biotropica 31:288–294.
Genty, B., J.-M. Briantais, and N. R. Baker. 1989. The relationshipbetween the quantum yield of photosynthetic electron trans-port rate and quenching of chlorophyll fluorescence. Bio-chimica Biophysica Acta 990:87–92.
Givnish, T. J. 1979. On the adaptive significance of leaf form. Pp.375–407 in O. T. Solbrig, S. Jain, G. B. Johnson, and P. H. Raven,eds. Topics in plant population biology. Columbia UniversityPress, New York.
———. 1988. Adaptation to sun and shade: a whole-plant per-spective. Australian Journal of Plant Physiology 15:63–92.
Gottsberger, G. 1988. The reproductive biology of primitive an-giosperms. Taxon 37:630–643.
Graham, S. W., and R. G. Olmstead. 2000. Utility of 17 chloro-plast genes for inferring the phylogeny of the basal angio-sperms. American Journal of Botany 87:1712–1730.
Greig, N. 1993. Regeneration mode in neotropical Piper: habitatand species comparisons. Ecology 74:2125–2135.
Grime, J. P. 1979. Plant strategies and vegetation processes. Wi-ley, Chichester, U.K.
Grubb, P. J. 1998. Seeds and fruits of tropical rainforest plants:interpretation of the range in seed size, degree of defense, andflesh/seed quotients. Pp. 1–24 in D. M. Newbery, H. H. T.Prins, and N. D. Brown, eds. Dynamics of tropical commu-nities. Blackwell Scientific, Oxford.
Hao, G., R. M. K. Saunders, and M.-L. Chye. 2000. A phyloge-netic analysis of the Illiciaceae based on sequences of internaltranscribed spacers (ITS) of nuclear ribosomal DNA. PlantSystematics and Evolution 223:81–90.
Hao, G., M.-L. Chye, and R. M. K. Saunders. 2001. A phyloge-netic analysis of the Schisandraceae based on morphologyand nuclear ribosomal ITS sequences. Botanical Journal of theLinnean Society 135:401–411.
Harper, J. L., P. H. Lovell, and K. G. Moore. 1970. The shapesand sizes of seeds. Annual Review of Ecology and System-atics 1:327–356.
Hickey, L. J., and J. A. Doyle. 1977. Early Cretaceous fossil evi-dence for angiosperm evolution. Botanical Review 43:1–104.
Hickman, J. C. 1993. The Jepson manual of higher plants of Cal-ifornia. University of California Press, Berkeley.
Hughes, N. F. 1994. The enigma of angiosperm origins. Cam-bridge University Press, Cambridge.
Kelly, C. K. 1995. Seed size in tropical trees: a comparative studyof factors affecting seed size in Peruvian angiosperms. Oec-ologia 102:377–388.
Kong, H.-Z. 2001. Comparative morphology of leaf epidermis inthe Chloranthaceae. Botanical Journal of the Linnean Society136:281–296.
Kong, H.-Z., Z.-D. Chen, and A.-M. Lu. 2002. Phylogeny ofChloranthus (Chloranthaceae) based on nuclear ribosomal ITSand plastid trnL-F sequence data. American Journal of Botany89:940–946.
Kott, L. S., and D. M. Britton. 1985. Role of morphological char-acteristics of leaves and the sporangial region in the taxonomyof Isoetes in northeastern North America. American Fern Jour-nal 75:44–55.
Krause, G. H., and E. Weis. 1991. Chlorophyll fluorescence andphotosynthesis: the basics. Annual Review of Plant Physiol-ogy and Plant Molecular Biology 42:313–349.
Kwit, C., M. W. Swartz, W. J. Platt, and J. P. Geaghen. 1998. Thedistribution of tree species in steepheads of the ApalachicolaRiver Bluffs, Florida. Journal of the Torrey Botanical Society125:309–318.
Leishman, M. R., I. Wright, A. T. Moles, and M. Westoby. 2000.The evolutionary ecology of seed size. Pp. 31–57 in M. Fenner,ed. Seeds: the ecology of regeneration in plant communities.CABI Publishing, Wallingford, U.K.
Les, D. H., E. L. Schneider, D. J. Padgett, P. S. Solis, D. E. Soltis,and M. Zanis. 1999. Phylogeny, classification, and floral evo-lution of water lilies (Nymphaeaceae: Nymphaeales): a syn-thesis of non-molecular, rbcL, matK, and 18S rDNA data. Sys-tematic Botany 24:28–46.
Luo, Y.-B., and Z.-Y. Li. 1999. Pollination ecology of Chloranthusserratus (Thunb.) Roem. et Schult. and Ch. fortunei (A. Gray)Solms-Laub. (Chloranthaceae). Annals of Botany 83:489–499.
Lupia, R., S. Lidgard, and P. R. Crane. 1999. Comparing paly-nological abundance and diversity: implications for biotic re-placement during the Cretaceous angiosperm radiation. Pa-leobiology 25:305–340.
Lusk, C. K. 1995. Seed size, establishment sites, and species co-existence in a Chilean rainforest. Journal of Vegetation Science6:249–256.

102 TAYLOR S. FEILD ET AL.
Lusk, C. K., and C. K. Kelly. 2003. Interspecific variation in seedsize and safe sites in a temperate rain forest. New Phytologist158:535–541.
Maddison, W. P. 1991. Squared-change parsimony reconstruc-tions of ancestral states for continuous-valued characters ona phylogenetic tree. Systematic Zoology 40:305–314.
Maddison, W. P., and D. R. Maddison. 2001. MacClade 4: anal-ysis of phylogeny and character evolution, Version 4.03. Sin-auer, Sunderland, Mass.
Magallon, S., and M. J. Sanderson. 2001. Absolute diversificationrates in angiosperm clades. Evolution 55:1762–1780.
Martin, T. J., and J. Ogden. 2002. The seed ecology of Ascarinalucida: a rare New Zealand tree adapted to disturbance. NewZealand Journal of Botany 40:397–404.
Mathews, S., and M. J. Donoghue. 1999. The root of angiospermphylogeny inferred from duplicate phytochrome genes. Sci-ence 286:947–950.
McElwain, J. C., and W. G. Chaloner. 1996. The fossil cuticle asa skeletal record of environment change. Palaios 11:376–388.
Metcalfe, C. R. 1987. Anatomy of the dicotyledons, 2d ed., Vol.III. Magnoliales, Illiciales, and Laurales (sensu Armen Takh-tajan). Clarendon, Oxford.
Mohr, B. A. R., and E. M. Friis. 2000. Early angiosperms fromthe Lower Cretaceous Crato Formation (Brazil), a preliminaryreport. International Journal of Plant Sciences 161(Suppl.):S155–S167.
Morse, S. R. 1990. Water balance in Hemizonia luzulifolia: the roleof extracellular polysaccharides. Plant, Cell and Environment13:39–48.
Nguyen Tu, T. T., H. Bocherens, A. Mariotti, F. Baudin, D. Pons,J. Broutin, S. Derenne, and C. Largeau. 1999. Ecological dis-tribution of Cenomanian terrestrial plants based on 13C/12Cratios. Palaeogeography, Palaeoclimatology, Palaeoecology145:79–93.
Niinemets, U., and K. Kalevi. 1994. Leaf weight per area and leafsize of 85 Estonian woody species in relation to shade toler-ance and light availability. Forest Ecology and Management70:1–10.
Niklas, K. J. 1997. The evolutionary biology of plants. Universityof Chicago Press, Chicago.
Nixon, K. C., W. L. Crepet, D. Stevenson, and E. M. Friis. 1994.A reevaluation of seed plant phylogeny. Annals of the Mis-souri Botanical Garden 81:484–533.
Parkinson, C. L., K. L. Adams, and J. D. Palmer. 1999. Multigeneanalyses identify the three earliest lineages of extant flower-ing plants. Current Biology 9:1485–1488.
Parrish, J. T. 1987. Global palaeogeography and palaeoclimateof the Late Cretaceous and Early Tertiary. Pp. 51–73 in E. M.Friis, W. G. Chaloner, and P. R. Crane, eds. The origins of an-giosperms and their biological consequences. CambridgeUniversity Press, Cambridge.
Parrish, J. T., A. M. Ziegler, and C. R. Scotese. 1982. Rainfall pat-terns and the distribution of coals and evaporites in the Me-sozoic and Cenozoic. Palaeogeography Palaeoclimatology Pa-laeoecology 40:67–101.
Pearcy, R. W. 1990. Sunflecks and photosynthesis in plant can-opies. Annual Review of Plant Physiology and Plant Molec-ular Biology 41:421–453.
Philipson, W. R. 1986. Trimeniaceae. Flora Malesiana Series I 10:327–333.
Playford, G. 1971. Palynology of Lower Cretaceous (Swan River)strata of Saskatchewan and Manitoba. Palaeontology 14:533–565.
Pons, D. 1984. Le Mesozoıque de Colombie: macroflores et mi-croflores. Editions du CNRS, Paris.
Qiu, Y.-L., J. Lee, F. Bernasconi-Quadroni, D. E. Soltis, P. S. Soltis,
M. Zanis, E. A. Zimmer, Z. Chen, V. Savolainen, and M. W.Chase. 1999. The earliest angiosperms: evidence from mito-chondrial, plastid and nuclear genomes. Nature 402:404–407.
Rascher, U., M. Liebig, and U. Luttge. 2000. Evaluation of instantlight-response curves of chlorophyll fluorescence parametersobtained with a portable chlorophyll fluorometer on site inthe field. Plant, Cell and Environment 23:1397–1405.
Rees, P. M., A. M. Ziegler, and P. J. Valdes. 2000. Jurassic phytoge-ography and climates: new data and model comparisons. Pp.297–318 in B. T. Huber, K. G. MacLeod, and S. L. Wing, eds. Warmclimates in earth history. Cambridge University Press, Cam-bridge.
Roberts, M. L., and R. R. Haynes. 1983. Ballistic seed dispersal inIllicium (Illiciaceae). Plant Systematics and Evolution 143:227–232.
Rodenburg, W. F. 1971. A revision of the genus Trimenia (Trimeni-aceae). Blumea 19:3–15.
Romero, E. J., and S. Archangelsky. 1986. Early Cretaceous angio-sperm leaves from southern South America. Science 234:1580–1582.
Roth-Nebelsick, A., D. Uhl, V. Mosbrugger, and H. Kerp. 2001. Evo-lution and function of leaf venation architecture: a review. Annalsof Botany 87:553–566.
Royer, D. L., L. J. Hickey, and S. L. Wing. 2003. Ecological conser-vatism in the ‘‘living fossil’’ Ginkgo. Paleobiology 29:84–104.
Sakai, A., T. Ohsawa, and M. Ohsawa. 1995. Adaptive significanceof sprouting of Euptelea polyandra, a deciduous tree growing onsteep slopes with shallow soil. Journal of Plant Research 108:377–386.
Salisbury, E. J. 1942. The reproductive capacity of plants: studies inquantitative biology. G. Bell, London.
Sanderson, M. J., and M. J. Donoghue. 1994. Shifts in diversificationrate with the origin of angiosperms. Science 264:1590–1593.
Saunders, R. M. K. 1998. Monograph of Kadsura (Schisandraceae).Systematic Botany Monographs 54:1–106.
———. 2000. Monograph of Schisandra (Schisandraceae). SystematicBotany Monographs 58:1–146.
Schabilion, J. T., and M. A. Reihman. 1985. Anatomy of petrifiedNeuropteris scheuchzeri pinnules from the Middle Pennsylvanianof Iowa: a paleoecological interpretation. Neuvieme Congres In-ternational de Stratigraphie et de Geologie du Carbonifere,Compte Rendu, 5:3–12.
Schneider, E. L., and S. Carlquist. 1995. Vessels in the roots of Bar-claya rotundifolia (Nymphaeaceae). American Journal of Botany82:1343–1349.
Schneider, E. L., and P. S. Williamson. 1993. Nymphaeaceae. Pp.486–493 in K. Kubitzki, ed. The families and genera of vascularplants. Springer, New York.
Schneider, E. L., S. Carlquist, K. Beamer, and A. Kohn. 1995. Ves-sels in Nymphaeaceae: Nuphar, Nymphaea, and Ondinea. In-ternational Journal of Plant Sciences 156:857–862.
Schrank, E. 1983. Scanning electron and light microscopic in-vestigations of angiosperm pollen from the Lower Cretaceousof Egypt. Pollen et Spores 25:213–242.
———. 1990. Palynology of the clastic Cretaceous sediments be-tween Dongola and Wadi Muqaddam, northern Sudan. Ber-liner Geowissenschaftliche Abhandlungen, Reihe A 120:149–167.
Sculthorpe, C. D. 1967. The biology of aquatic vascular plants.Arnold, London.
Smith, A. C. 1947. The families Illiciaceae and Schisandraceae.Sargentia 7:1–224.
———. 1976. Studies of Pacific Island plants, XXXIII. The genusAscarina (Chloranthaceae) in the southern Pacific. Journal ofthe Arnold Arboretum 57:405–425.
Smith, W. K., T. C. Vogelmann, E. H. DeLucia, D. T. Bell, and K.

103EARLY ANGIOSPERM ECOPHYSIOLOGY
A. Shepherd. 1997. Leaf form and photosynthesis. Bioscience47:785–793.
Smits, A. J. M., R. van Ruremonde, and G. van der Velde. 1990.Germination requirements and seed banks of some nym-phaeid macrophytes: Nymphaea alba L., Nuphar lutea (L.) Sm.,and Nymphoides peltata (Gmel.) O. Kuntze. Freshwater Biology24:315–326.
Soltis, D. E., P. S. Soltis, D. L. Nickrent, L. A. Johnson, W. J. Hahn,S. B. Hoot, J. A. Sweere, R. K. Kuzoff, K. A. Kron, M. W. Chase,S. M. Swensen, E. A. Zimmer, S.-M. Chaw, L. J. Gillespie, W. J.Kress, and K. J. Sytsma. 1997. Angiosperm phylogeny inferredfrom 18S ribosomal DNA sequences. Annals of the Missouri Bo-tanical Garden 84:1–49.
Soltis, D. E., P. S. Soltis, M. W. Chase, M. E. Mort, D. C. Albach, M.Zanis, V. Savolainen, W. H. Hahn, S. B. Hoot, M. F. Fay, M. Axtell,S. M. Swensen, K. C. Nixon, and J. S. Farris. 2000. Angiospermphylogeny inferred from 18S rDNA, rbcL, and atpB sequences.Botanical Journal of the Linnean Society 133:381–461.
Sperry, J. S. 2003. Evolution of water transport and xylem structure.International Journal of Plant Sciences 164(Suppl.):S115–S127.
Stebbins, G. L. 1965. The probable growth habit of the earliest flow-ering plants. Annals of the Missouri Botanical Garden 52:457–468.
———. 1974. Flowering plants: evolution above the species level.Harvard University Press, Cambridge.
Stone, D. E. 1968. Cytological and morphological notes on thesoutheastern endemic Schisandra glabra (Schisandraceae). Journalof the Elisha Mitchell Scientific Society 84:351–356.
Sun, G., D. L. Dilcher, S. Zheng, and Z. Zhou. 1998. In search of thefirst flower: a Jurassic angiosperm, Archaefructus, from northeastChina. Science 182:1692–1695.
Sun, G., Q. Ji, D. L. Dilcher, S. Zheng, K. C. Nixon, and X. Wang.2002. Archaefructaceae, a new basal angiosperm family. Science296:899–904.
Swamy, B. G. L., and I. W. Bailey. 1950. Sarcandra, a vesselless genusof the Chloranthaceae. Journal of the Arnold Arboretum 31:117–129.
Takhtajan, A. L. 1969. Flowering plants: origin and dispersal.Smithsonian Institution Press, Washington, D.C.
Taylor, D. W., and L. J. Hickey. 1990. An Aptian plant with attachedleaves and flowers: implications for angiosperm origin. Science247:702–704.
———. 1992. Phylogenetic evidence for the herbaceous origin of an-giosperms. Plant Systematics and Evolution 180:137–156.
———. 1996. Evidence for and implications of an herbaceous originfor angiosperms. Pp. 232–266 in D. W. Taylor and L. J. Hickey, eds.Flowering plant origin, evolution, and phylogeny. Chapman andHall, New York.
Thien, L. B., D. A. White, and L. Y. Yatsu. 1983. The reproductivebiology of a relict: Illicium floridanum Ellis. American Journal ofBotany 70:719–727.
Thorne, R. F. 1976. A phylogenetic classification of the Angiosper-mae. Evolutionary Biology 9:35–106.
Tiffney, B. H. 1984. Seed size, dispersal syndromes, and the rise ofthe angiosperms: evidence and hypothesis. Annals of the Mis-souri Botanical Garden 71:551–576.
Todzia, C. A. 1988. Chloranthaceae: Hedyosmum. Flora NeotropicaMonograph 48:1–139.
Upchurch, G. R. 1984a. Cuticular anatomy of angiosperm leavesfrom the Lower Cretaceous Potomac Group. I. Zone I leaves.American Journal of Botany 71:192–202.
———. 1984b. Cuticle evolution in Early Cretaceous angio-sperms from the Potomac Group of Virginia and Maryland.Annals of the Missouri Botanical Garden 71:522–550.
———. 1995. Dispersed angiosperm cuticles: their history, prep-
aration, and application to the rise of angiosperms in Creta-ceous and Paleocene coals, southern western interior of NorthAmerica. International Journal of Coal Geology 28:161–227.
Upchurch, G. R., and D. L. Dilcher. 1990. Cenomanian angio-sperm leaf megafossils, Dakota Formation, Rose Creek local-ity, Jefferson County, southeastern Nebraska. U.S. GeologicalSurvey Bulletin 1915:1–55.
Upchurch, G. R., and J. A. Doyle. 1981. Paleoecology of the co-nifers Frenelopsis and Pseudofrenelopsis (Cheirolepidiaceae)from the Cretaceous Potomac Group of Maryland and Virgin-ia. Pp. 167–202 in R. C. Romans, ed. Geobotany II. Plenum,New York.
Upchurch, G. R., and J. A. Wolfe. 1987. Mid-Cretaceous to EarlyTertiary vegetation and climate: evidence from fossil leavesand woods. Pp. 75–105 in E. M. Friis, W. G. Chaloner, and P.R. Crane, eds. The origins of angiosperms and their biologicalconsequences. Cambridge University Press, Cambridge.
Van de Water, P. K., S. W. Leavitt, and J. L. Betancourt. 1994.Trends in stomatal density and 13C/12C ratios of Pinus flexilisneedles during last glacial-interglacial cycle. Science 264:239–243.
Verdcourt, B. 1986. Chloranthaceae. Flora Malesiana Series I 10:123–144.
Vogelmann, T. C., J. N. Nishio, and W. K. Smith. 1996. Leaves andlight capture: light propagation and gradients of carbon fixationwithin leaves. Trends in Plant Science 1:65–70.
Volkheimer, W., and A. Salas. 1975. Die alteste Angiospermen-Pa-lynoflora Argentiniens von der Typuslokalitat der unterkreta-zischen Huitrın-Folge des Neuquen-Beckens. MikrofloristischeAssoziation und biostratigraphische Bedeutung. Neues Jahrbuchfur Geologie und Palaontologie Monatshefte 1975:424–436.
Walker, J. W., and A. G. Walker. 1984. Ultrastructure of Lower Cre-taceous angiosperm pollen and the origin and early evolution offlowering plants. Annals of the Missouri Botanical Garden 71:464–521.
Wells, P. V. 1969. The relation between mode of reproduction andextent of species in woody genera of the California chaparral.Evolution 23:264–267.
Wilf, P., S. L. Wing, D. R. Greenwood, and C. L. Greenwood. 1998.Using fossil leaves as paleoprecipitation indicators: an Eocene ex-ample. Geology 26:203–206.
Williamson, P. S., and E. L. Schneider. 1993. Cabombaceae. Pp. 157–161 in K. Kubitzki, ed. The families and genera of vascular plants.Springer, New York.
Williamson, P. S., E. Schneider, and L. Malins. 1989. Tuber and leafstructure of Ondinea purpurea Den Hartog (Nymphaeaceae).Western Australian Naturalist 18:52–61.
Wing, S. L., and L. D. Boucher. 1998. Ecological aspects of the Cre-taceous flowering plant radiation. Annual Review of Earth andPlanetary Sciences 26:379–421.
Wing, S. L., and W. A. DiMichele. 1992. Ecological characterizationof plants. Pp. 140–180 in A. K. Behrensmeyer, J. D. Damuth, W.A. DiMichele, R. Potts, H.-D. Sues, and S. L. Wing, eds. Terrestrialecosystems through time: evolutionary paleoecology of terrestri-al plants and animals. University of Chicago Press, Chicago.
Wing, S. L., and B. H. Tiffney. 1987. The reciprocal interaction ofangiosperm evolution and tetrapod herbivory. Review of Palaeo-botany and Palynology 50:179–210.
Wolfe, J. A. 1993. A method of obtaining climatic parametersfrom leaf assemblages. U.S. Geological Survey Bulletin 2040:1–71.
Wolfe, J. A., J. A. Doyle, and V. M. Page. 1975. The bases of an-giosperm phylogeny: paleobotany. Annals of the MissouriBotanical Garden 62:801–824.
Zanis, M. J., D. E. Soltis, P. S. Solits, S. Mathews, and M. J. Don-

104 TAYLOR S. FEILD ET AL.
oghue. 2002. The root of the angiosperms revisited. Proceed-ings of the National Academy of Sciences USA 99:6848–6853.
Ziegler, A. M., A. L. Raymond, T. C. Gierlowski, M. A. Howell,D. B. Rowley, and A. L. Lottes. 1987. Coal, climate, and ter-restrial productivity: the present and Early Cretaceous com-pared. Pp. 25–49 in A. C. Scott, ed. Coal and coal-bearing stra-ta: recent advances. Geological Society, London.
Appendix 1
Study Sites and Species Included in the Study
Amborellales1. Amborella, A. trichopoda Baillon., just below the summit of
Massif Aoupinie, 800–1000 m, Province Nord, New Caledonia,second population on Plateau de Dogny, 650–750 m, ProvinceSud, New Caledonia.Austrobaileyales
1. Austrobaileya, A. scandens C. T. White, at the base of Mt.Bartle Frere, 650 m, Queensland, Australia.
2. Kadsura, K. coccinea (Lemaire) A. C. Smith, west of SokKwu Wan, Lamma Island, Hong Kong, second population atthe home of Mr. Liu Shu-Zhian, riverside, Xinning Co., Hunan,China; K. heteroclita (Roxburgh) Craib, 1100 m, Luohangdong,Ziyunshan, Forestry Park, Xinning Co. Hunan, China; K. ja-ponica (L.) Dunal, collected in South Korea, grown in a com-mon garden, Berkeley, California, U.S.A.; K. longipenduculataFinet & Gagnepain, 1100 m, Luohangdong, Ziyunshan, For-estry Park, Xinning Co. Hunan, China; K. oblongifolia Merrill,1000 m, Mt. Dialoushan, Hainan Dao, China.
3. Illicium, I. angustisepalum A. C. Smith, Sunset Peak, 460 m,Lantau Island, Hong Kong; I. anisatum L., collected from Ja-pan, grown in a common garden, Lee Co., Auburn, Alabama,U.S.A.; I. dunnianum Tutch. Wu Kau Tang, New Territories,Hong Kong, I. floridanum Ellis, Highway 169, toward PhenixCity, Lee Co., Alabama, U.S.A., second population locatedalong Apalachicola River, near Bristol, Liberty Co., Florida,U.S.A.; I. henryi Diels, collected from China, grown in a com-mon garden, Lee Co., Auburn, Alabama, U.S.A.; I. lanceolatumA. C. Smith, Fanshuijian, Shuhuanshan, Xinning Co., Hunan,China, a second population at 750 m, Bajiaozhai, Nangshan,Xinning Co., Hunan, China; I. majus Hook.f. & Thoms., 850 m,Luohandong, Ziyushan, Forestry Park, Xinning Co., Hunan,China; I. mexicanum A. C. Smith, collected from Veracruz,Mexico, grown in a common garden, Lee Co., Auburn, Ala-bama, U.S.A.; I. micranthum Dunn, Ma On Shan 450 m, HongKong; Lizhishan Village, Xinning Co., Hunan, China; two kmdown from home of Mr. Liu Shu-Zhian, riverside, Xinning Co.,Hunan, China; I. oligodandrum Merril & Chun (1100 m, ridgetop forest, Mt. Dialoushan, Hainan Dao, China; I. parviflorumMichx. Ex Vent., Alexander Springs, Lake Co., near Ocala,Florida, U.S.A.; I. simonsii Maxim, collected from Sichuan,China, grown in the UC Botanic Garden, Berkeley, California,U.S.A.; I. ternstroemioides, A. C. Smith, 1100 m, ridge-top for-est, Mt. Dialoushan, Hainan Dao, China.
4. Schisandra, S. arisanensis subspecies viridis (A. C. Smith)R. M. K. Saunders, Lizhishan Village, Xinning Co., Hunan,China; S. bicolor W.-C. Cheng 1200 m, Luohangdong, Ziyun-shan, Forestry Park, Xinning Co. Hunan, China; S. chinensis(Turcz.) Baill. collected from South Korea, grown in a commongarden, Berkeley, California, U.S.A.; S. glabra (Brickell) RehderCrowley’s Ridge, Three Rivers State Park, Whynne, Arkansas;property of Mr. Dan S. Miller, Tallahassee, Florida; plants ofboth populations collected and grown in a common garden,
Lee Co. Auburn, Alabama, U.S.A.; S. grandiflora (Wallich)Hooker f. & Thomson collected in China, grown in a commongarden, Berkeley, California, U.S.A.; S. henryi Clarke Luohang-dong, Ziyunshan, Forestry Park, Xinning Co. Hunan, China; S.lancifolia (Rehder & E. H. Wilson) A. C. Smith collected in Chi-na, grown in a common garden, Berkeley, California, U.S.A.;S. propinqua (Wallich) Baillon grown in a common garden,Berkeley, California, U.S.A.; S. repanda (Siebold & Zuccarini)Radlkofer collected in Japan, grown in a common garden,Berkeley, California, U.S.A.; S. sphenanthera Rehder & E. H.Wilson 800 m, Sazijiang, Ziyunshan Forestry Farm, XinningCo., Hunan, China.
5. Trimenia , T. moorei (Oliv. in Bth.) Philipson StockyardCreek, near Armidale, New South Wales, Australia; T. neoca-ledonica Baker below the summit of Massif Aoupinie, near vil-lage of Goapin, Province Sud, New Caledonia; T. papuana , 2300m, Mt. Gumi, Morobe Province, Papua New Guinea; T. wein-manniifolia Seeman summit of Des Vouex Peak, 990 m, Boumavillage, Taveuni, Fiji.Chloranthaceae
1. Ascarina, A. lucida Hook.f., Saltwater Forest, 25 m, nearHokitika, New Zealand; A. phillipensis, 2200 m, Kuper Range,Morobe Province, Papua New Guinea; A. polystachya J. R. &Forester, Mt. Marau, 780 m, Tahiti, French Polynesia; A. rub-ricaulis Solms-Laub., Mt. Dzumac, 990 m, Province Sud, NewCaledonia; A. solmsiana Schlechter, below the summit of Mt.Dzumac, 980 m, Province Sud, New Caledonia; A. swamyanaA. C. Smith, below Des Vouex Peak, 1000 m, Bouma Village,Tavenui, Fiji.
2. Chloranthus, C. fortunei 350 m, Sankoudu, Shuichaoyuan,Jinshizhen, Xinning Co., Hunan, China; C. henryi 1100 m, Luo-handong, Ziyushan, Forestry Park, Xinning Co., Hunan, Chi-na; C. japonicus Siebold collected in China, raised in a commongarden at UC Berkeley, California, U.S.A.; C. serratus Roem &Schlt. 90 m, New Territories, Hong Kong; 450 m, Sankoudu,Shuichaoyuan, Jinshizhen, Xinning Co., China; C. spicatus Peinative to China, greenhouse cultivated, UC Berkeley, Califor-nia, U.S.A.; C. sessilifolius Pei 700–800 m, Fanshuijianm Shun-hungshan, Forestry Park, Xinning Co., Hunan, China.
3. Hedyosmum, H. bonplandianum Humboldt, Bonpland &Kunth 1450 m, Estacion Cacao, Volcan Cacao, Guancaste, Cos-ta Rica; H. costaricense Burger 1300 m, Rıo Grande de Orosı,Cartago, Costa Rica; H. goudotianum var. goudotianum Solms-Laubach 2500 m, 7 mi. W of Ojo del Agua, Cerro de la Muerte,Cordillera de Talamanca, Cartago, Costa Rica; H. orientaleMerrill & Chun 1150 m, Mt. Dialoushan, Hainan Dao, China;H. mexicanum Cordemoy 2500 m, along the Pan AmericanHighway, Cordillera de Talamanca, Cartago, Costa Rica.
4. Sarcandra, S. glabra (Thunb.) Nakai Victoria Peak 350 m,Hong Kong; 450 m, Ma On Shan, Hong Kong; Sunset Peak, 640m, Lantau Island, Hong Kong; plants collected from Japan,raised in a common-garden at UC Berkeley, California, U.S.A.;S. hainanensis (Pei) Swamy & Bailey 1000 m, Mt. Dialoushan,Hainan Dao, China.Nymphaeales
1. Brasenia, Brasenia schreberi J. Gmelin Lake Audrain, Do-rado Co., California, U.S.A.
2. Cabomba, C. caroliniana Gray, collected from Texas, grownin an experimental aquarium, Berkeley, California, U.S.A.
3. Nuphar, Nuphar lutea (L.) Sibth. & Sm. subsp. polysepala(Engelm.) E. Beal Lily Pad Lake, Marin Co., California, U.S.A.
4. Nymphaea, N. odorata Aiton Tom Pond, Harvard Forest, Pe-tersham, Massachusetts, U.S.A.

105EARLY ANGIOSPERM ECOPHYSIOLOGY
Appendix 3Characters and character states (discrete characters all unordered) used in phylogenetic character reconstructionanalyses: 1. Maximum leaf photosynthetic rate (ETRmax, mmol electrons m22 s21); 2. Light-saturation point (PPFDsat, mmolm22 s21); 3. Seed size (SV, mm3); 4. Leaf mesophyll (0) spongy parenchyma only, (1) single-layered palisade parenchyma,(2) multiple-layered palisade parenchyma; 5. Seedling establishment habitat (0) shady, stable, and aquatic, (1) shady,disturbed, (2) sunny, disturbed; 6. Growth habit (0) tree or shrub, (1) twining woody vine, (2) herbs with some sec-ondary growth, (3) herbs lacking secondary growth. * denotes that the phylogenetic position is uncertain.
Species
Character
1 ETRmax 2 PPFDsat 3 SV 4 Mesophyll5 Seeding
habitat6 Growth
habit
Amborella trichopodaAscarina coursiiAscarina diffusaAscarina lucidaAscarina rubricaulisAscarina philippinensis
87 6 11??
92 6 23133 6 24
?
466 6 48??
598 6 86670 6 57
?
3.46??
0.330.260.99
000000
122221
000000
Ascarina polystachya*Ascarina solmsianaAscarina swamyanaAustrobaileya scandens
144 6 1345 6 477 6 848 6 8
535 6 29234 6 9320 6 15290 6 5
0.240.650.95
73.20
0000
2111
0001
Barclaya longifoliaBarclaya rotundifoliaBrasenia schreberiCabomba carolinianaChloranthus angustifolius
??
135 6 850 6 5
?
??
585 6 13400 6 10
?
??
2.511.03?
12210
01001
33332
Chloranthus erectusChloranthus fortuneiChloranthus henryiChloranthus japonicusChloranthus nervosus
?72 6 1346 6 166 6 12
?
?389 6 14347 6 2374 6 5
?
2.000.881.221.52?
00000
11111
02222
Chloranthus oldhamiiChloranthus serratusChloranthus spicatusChloranthus sessilifolius‘‘Core Angiosperms’’
?54 6 748 6 849 6 5
?
?309 6 59395 6 34284 6 10
?
?0.762.011.94?
00001
1111?
2202?
Euryale feroxHedyosmum arborescensHedyosmum bonplandianumHedyosmum brasiliense
??
150 6 22?
??
600 6 12?
63.14?
1.681.02
2000
0222
3000
Appendix 2
Data Sources Used for Phylogenetic Character-StateReconstruction
Character SourcesSeeds Hedyosmum Todzia (1988); Illicium Rob-
erts and Haynes (1983); Kadsura Saunders(1998); Schisandra Saunders (2000)
Seedling habitats Chloranthus Luo and Li (1999); OndineaWilliamson et al. 1989; Nuphar Smits et al.(1990); Barrat-Segretain (1996); TrimeniaPhilipson (1986)
Leaf anatomy Amborella Metcalfe (1987); Ascarina Met-calfe (1987), Todzia (1988); AustrobaileyaFeild et al. (2003a); Barclaya E. L. Schneiderpersonal communication 2002; Braseniaand Cabomba Williamson and Schneider(1993); Chloranthus and Hedyosmum Todzia(1988); Euryale, Nuphar, Nymphaea, andOndinea Sculthorpe (1967), Williamson etal. (1989), Schneider and Williamson(1993); Sarcandra Swamy and Bailey(1950); Trimenia Rodenburg (1971)
Growth forms Amborella Feild et al. (2001); AscarinaSmith (1976), Blanc (1986), Verdcourt(1986); Austrobaileya Feild et al. (2003a);Barclaya Schneider and Carlquist(1995); Brasenia and Cabomba William-son and Schneider (1993); ChloranthusVerdcourt (1986), Carlquist (1992a),Luo and Li (1999), Kong et al. (2002);Hedyosmum Todzia (1988), Carlquist(1992b); Illicium Smith (1947), Robertsand Haynes (1983), Thien et al. (1983);Kadsura Smith (1947), Saunders (1998);Euryale, Nuphar, Nymphaea, Ondinea,and Victoria Williamson et al. (1989),Schneider and Williamson (1993), Wil-liamson and Schneider (1993), Les et al.(1999); Sarcandra Swamy and Bailey(1950), Verdcourt (1986), Carlquist(1987); Schisandra Smith (1947), Stone(1968), Saunders (2000); Trimenia Ro-denburg (1971), Philipson (1986)
106 TAYLOR S. FEILD ET AL.
Appendix 3. Continued.
Hedyosmum costaricense*Hedyosmum goudotianum*Hedyosmum mexicanum*
102 6 16141 6 11134 6 9
500 6 14570 6 15550 6 14
??
0.84
000
222
000
Hedyosmum nutansHedyosmum orientaleIllicium angustisepalumIllicium anisatumIllicium dunnianum
?80 6 972 6 779 6 9
100 6 10
?490 6 12405 6 21337 6 25495 6 10
0.56??
12.19?
00111
22111
00000
Illicium floridanumIllicium henryiIllicium lanceolatumIllicium majusIllicium mexicanum*
101 6 6115 6 16
99 6 1668 6 15
100 6 10
501 6 19526 6 21490 6 12358 6 20495 6 12
5.944.01???
11111
11111
00000
Illicium verumKadsura coccineaKadsura heteroclitaKadsura japonica
110 6 8121 6 9
59 6 177 6 11
488 6 13608 6 53303 6 37400 6 35
7.8864.58
8.914.24
1011
121?
0111
Illicium micranthumIllicium oligodandrum*Illicium parviflorumIllicium simonsii*Illicium ternstroemioides*
67 6 1077 6 9
125 6 1187 6 11
100 6 9
409 6 17393 6 15525 6 13460 6 15450 6 28
??
7.84??
11111
11111
00000
Kadsura longipedunculataKadsura oblongifoliaNuphar lutea ssp. polysepala
53 6 2745 6 8
225 6 15
389 6 44290 6 48
1400 6 20
2.822.842.02
102
210
113
Nymphaea odorataOndinea purpureaSarcandra glabraSarcandra hainanensisSchisandra arisanensis
78 6 5?
40 6 546 6 8
111 6 14
425 6 34?
200 6 4282 6 21575 6 53
5.610.711.111.125.75
2200?
00112
33001
Schisandra bicolor*Schisandra chinensisSchisandra glabraSchisandra grandifloraSchisandra henryi
61 6 2115 6 13
69 6 890 6 11
106 6 13
325 6 5525 6 28325 6 33477 6 8536 6 61
6.426.636.574.412.47
?1001
02122
11111
Schisandra lancifolia*Schisandra propinquaSchisandra repanda*Schisandra sphenantheraTrimenia macurura*
105 6 1189 6 1156 6 11
110 6 26?
520 6 22480 6 8320 6 8560 6 103
?
18.654.50
12.064.92?
?0?10
21221
11111
Trimenia moorei*Trimenia neocaledonica*Trimenia papuana*Trimenia weinmanniifolia*Victoria amazonica
87 6 1178 6 12
?113 6 11
?
500 6 48470 6 25
?510 6 48
?
1.525.474.131.42?
00002
21220
10003
Appendix 4
Field Analysis of Photosynthetic Performance UsingChlorophyll Fluorescence Emission
We chose this measure of photosynthesis because many fieldsites were exceedingly wet and remote, making standard leafgas-exchange flux measurements impossible. Maximum leafphotosynthetic rate and the point at which photosynthesisreaches light saturation were calculated from chlorophyll (Chl)a fluorescence data. These calculations exploit the positive re-lationship between PS II efficiency and increasing photosyn-thetic photon flux density (PPFD, mmol photons m22 s21), termeda ‘‘rapid light response curve’’ (Bilger et al. 1995; Rascher et al.2000). Chl a fluorescence provides information on the rate atwhich light energy is transferred to photosystem (PS) II, a por-tion of the biochemical mechanism by which plants generate thereducing power (NADPH) needed to reduce carbon dioxide inthe photosynthetic dark reactions (Krause and Weis 1991). PS II
efficiency (fPSII) expresses the number of active PS II reactioncenters and the efficiency with which these centers transducelight to chemical energy (Genty et al. 1989). fPSII was calculatedas:
f 5 (F 9 2 F)/F 9PSII m m (1)
F is the steady-state fluorescence emission of the leaf while pho-tosynthesizing, and Fm9 is the maximum light-adapted fluores-cence of the leaf when a saturating pulse (duration 5 0.8 s, in-tensity 5 4500 mmol m22 s21) is superimposed on the prevailinglight to momentarily close all active PS II reaction centers (Gen-ty et al. 1989).
fPSII was determined at eight light intensities, from 2% to100% full sunlight (i.e., 20–2000 mmol m22 s21), taking measure-ments from lowest to highest light intensity. The leaf was illu-minated at each intensity until F values were stable at least 15s, which required 30–90 s, and a saturating pulse was then ap-plied. Although fPSII light response measurements under short

107EARLY ANGIOSPERM ECOPHYSIOLOGY
illumination times are likely to be quasi–steady state, Rascher etal. (2000) demonstrated when leaves are fully photosyntheti-cally induced, rapid light response curves are identical thoseproduced by longer illumination times. This result was con-firmed for greenhouse-raised plants of Austrobaileya scandens,Chloranthus spicatus, Illicium floridanum, Illicium henryi, Illiciumparviflorum, Kadsura japonica, Sarcandra glabra, and Schisandralancifolia (Feild unpublished data 2001). Consequently, care wastaken that leaves in the field were exposed to morning light forat least one to two hours to allow full induction of photosyn-thesis and stomatal opening (Pearcy 1990). Measurements weremade between 1030 and 1230 h to avoid time-dependent effectson photosynthetic performance (Rascher et al. 2000). Mean leaftemperatures averaged 218C (6 28C).
Estimates of fPSII were used for calculating the apparent rateof photosynthetic electron transport (ETR). ETR expresses therelative rate at which electrons move through PS II, and is gen-erally close to the rate at which NADP1 is reduced and madeavailable for carbon fixation (Edwards and Baker 1993; Bilger etal. 1995; Brodribb and Hill 1997). ETR (mmol electrons m22 s21)was calculated as:
ETR 5 (f ) 3 (PPFD) 3 (0.5) 3 aPSII (2)
fPSII is calculated in equation (1). Photosynthetic photon fluxdensity (PPFD) is the incident light intensity (mmol m22 s21) anda is percent leaf light absorbance. The constant 0.5 accounts forthe fact that absorbed light is evenly distributed between PSIIand PSI. Measurements of PPFD (in the waveband from 380 to710 nm) were made near the leaf surface by the microquantumsensor of the Mini-PAM, calibrated against a Li-190 light sensor(Li-COR, Lincoln, Nebraska). PPFD values were corrected forthe 2 mm difference between the distance of the photodiode andleaf plane as described by Rascher et al. (2000). The internal hal-ogen lamp of the Mini-PAM provided light, which was directed
to the leaf via a fiber optic bundle. The average value of a 5 0.84for green leaves (Bjorkman and Demmig 1987) was used here.This value is a good approximation because all leaves measuredwere healthy and of similar color. Note, however, that ETR val-ues will not be precise because leaf absorbance differences be-tween species were not measured. Polynomial regression wasused to fit the light response data from each species, and satu-rating PPFD (PPFDsat) and maximum ETR (at PPFDsat) were readfrom these curves as described by Brodribb and Hill (1997).PPFDsat was taken as the light intensity at the peak of the ETRversus PPFD response curve.
Some sampled species occurred in a variety of light environ-ments. For example, Hedyosmum bonplandianum (Chlorantha-ceae) occurred in open habitats on cloud forest land slips (about30–50% full sun, defined as ;2000 mmol photons m22 s21 [Chaz-don et al. 1996]) under persistent cloud cover, semi-shaded for-est light-gaps (10–25% full sun), and in forest understory sites(,2% full sun). For such species, five replicate light curves weretaken from five individuals from each of the three light envi-ronments. Curves were averaged within each environment. Theaverage curve yielding the greatest mean values of ETRmax andPPFDsat was then used for interspecific comparisons. For waterlilies, photosynthetic measurements were restricted to specieswith floating leaves (e.g., Brasenia, Cabomba, Nuphar, Nymphaea)because leaf photosynthetic performance in submersed and ae-rial environments is not commensurate (Niklas 1997). Light en-vironment for each species was characterized with local mea-surements of PPFD taken with a photodiode-based meter (Li-190, Li-COR) held horizontally above leaves of the same branch-es sampled for anatomical and physiological studies.Measurements were taken from 1000 and 1230 h on represen-tative clear periods and cloudy days. Leaves from branches withmaximum photosynthetic capacity were used for assessment ofanatomical/morphological leaf characters.