Embed Size (px)
Citation preview

Pflügers Archiv, Bd. 267, S. 313--330 (1958)
Aus dem Pharmako]ogischen Institut der Universität Heidelberg und dem Physiologischen Institut der Universität l~reiburg i. Br.
Das Verhalten der energiereiehen Phosphat-Verbindungen des Myokards im ~ormalzustand
und bei experimenteller Insuffizienz durch Phenylbutazon {Versuche am Herz-Lungen-Präparat des Meerschweinchens) ~
Von H. HOCHREIN und H. J. DÖRLNG
unter Mitarbeit von 1~. DROH und A. ~J~NOPP
Mit 7 Textabbildungen
(Eingegangen am 6. März 1958)
Die Frage, ob die tterzinsnffizienz von einer Veränderung des Stoff- wechsels energiereicher Phosphor-Verbindungen begleitet ist, wurde bisher recht verschieden beantwortet. Während WOLL]~~~]~I~¢~R (1947) ebenso wie F•w•z u. HAw~ (1953) bei der Barbitnrat-Insuffizienz am tterz-Lungel~-Präparat (HLP) des Hundes eine Zunahme von Kreatin- phosphat beobachteten, wurde in anderen Versuchen von W~ICK~R (1935) und MÜGGE (1933) eine Abnahme des Kreatinphosphat-Gehaltes bei Beeinträehtigung der Myokardleistung besehrieben. BOGATZKI U. STAU~ (1956) fanden am Chinin-gesehädigten Rattenherzen ebenfalls ein Absinken der energiereichen Phosphate.
Eine weitere Möglichkeit, gut steuerbare Herzinsuffizienzen zu er- zeugen, besteht nach vorausgegangenen Untersuchungen von I-Ioc~~~,rz¢, WILXE u. STRA]?AZAKIS (1958) in der Infusion von Phenylbutazon. Da diese Form experimenteller tterzinsuffizienz bisher keiner biochemisehen Analyse unterzogen wurde, sollten die folgenden :Untersuchungen klären : ~,
a) ob die Phenylbutazon-Insuffizienz mit einem Anstieg oder miß einer Abnahme der energiereichen Phosphate des Herzmuskels einher- geht,
b) o b überhaupt bei der PhenylbutazomInsuffizienz eine Beziehung zwischen de r mechaniSchen Leistung des Myokards und dem Verhalten der: energiereichen Phosphor-,Verbindungen festzustellen ist.
* Ein vorläufiger Bericht über Tei]ergebnisse wurde bereits auf der 23. T~gg. der Deutsch. Pharmakol. Ges.. in Freiburg/Br. (13.--15. Juni t957) w»rgelegt (vgl. N~unyn-Sehmiedeberg's Arch. exp. Path. Pharm~k. 282, 290--292 (1957).
Pflügers Arch. ges. Physiol., ttd, 267 22

314 I'i. I'IOC]tI~,EI~ und H. J. Dö~I~¢:
Experimenteller Teil
Methodik
Tierexperimentelle Versuchsanordnung S~mtiiehe Ergebnisse wurden am Herz-Lungen-Präparat des Meerschwein-
chens (Tiergewicht 150B20 g) gewonnen. Die spezielle Pr~paration erfolgte nach der von BnGovIc und STE~~ (1953) an Ratten ausgearbeiteten Methode. Der künstliche Kreislauf wurde mit 15 ml ]~attenblut, die mit 4 ml 0,9~oigcr NaC1- Lösung verdürmt waren, aufgefüllt. Zur Gerinnungsverhütung wurde außerdem jeweils 1 ml einer 5~oigen Thrombocid-Lösung 1 zugesetzt. Das Minutenvolumen wurde mit Hilfe des Bubble-flow-meters nach B~Av~, ALT~OFF U. T~VGNE~ (1954) registriert. Zur Messung der Herzfrequenz diente eine Marey'sche Kapsel. Der ar- terielle Druck wurde mittels eines Quecksilbermanometers in mm Hg, der Druck im rechten Vorhof mittels eines Wassermanometers in mm H~O geschrieben. Die ~arkose der Tiere erfolgte nach einer von GIA«~ (1940) erstmalig angegebenen Me- thode durch kombinierte Unterkühlung.Erstiekung. Die Reetal-Tempcratur sinkt dabei bis auf 24 °C ab; die bei der Herstellung des Herz-Lungen-Pri~parates not- wendigen Manipulationen werden in diesem Zustand der Hypothermie weit besser als bei normaler Körpcrtemperatur vertragen. Künstliche Beatmung wurde erst nach Eröffnung des Thorax eingeleitet. Die Wicderaufw~rmung des Herz-Lungen- Präparates wurde nach Herstellung des künstlichen Kreislaufs durch Einleitung des auf 37 °C temperierten Blutgemisches aus dem Vorratsgef~ß zustande gebracht. Der Spiegel des Vorratsgef~ßes befand sieh auf der Grundstufe 200 mm über dem Herzen und wurde bei den Belastungsstufen jeweils um 50 mm erhöht.
Als Maß für die Lcistungsfi~higkeit des Herzens wurde der ,Competence index" (C. I.) nach KRAYE~ (1931) und WOLLn~B~RGE~ (194~7) errechnet:
C. I. ~ Erhöhung des V0rratsgefaßes - - Anstieg des re. Vorhofdruckes Erhöhung des Vorratsgefi~I~es
Der C.I. ist beim vollkompensicrten Herzen gleich 1,0, wenn bei Erhöhung des venösen Angebotes keine Drucksteigerung im rechten Vorhof erfolgt. Dagegen erreicht der C. I. an der Grenze der Leistungsfähigkeit (Su/fizienzgrenze) den Wert Null; hierbei entspricht die Drucksteigerung im rechten Vorhof (in Millimeter HH20 ) genau der Erhöhung des Vorratsgefa]~es (in Millimeter).
Experimentelle Schädigungen des Herzens wurden durch langsame Infusion von Phenylbutazon ~ - - und in einigen Fällen durch Fluoressigsi~ure 3 __ in den künstlichen Kreislauf hervorgerufen. Zu dem Zeitpunkt, ~n dem die gewünschten Insuffizienzzeiehen eingetreten waren, wurde das völlig freiliegende Herz durch Scherenschlag unterhalb der Herzbasis abgetrennt und augenblicklich in flüssigem Stickstoff eingefroren. Zu Vergleiehszwecken wurden auch normale Herzen künst- lich beatmeter - - mit Evipan (20 mg/100 g intraperitoneal) r~rkotisierter - - Meer- schweinchen nach Thoraxeröffnung gewonncB und in identischer Weise verarbeitet.
Chemische Versuchsanordnung 1. Gewinnung des tIerzmuskelextraktes und Bestimmung von Kreatin-
phosphat und säurelösUchem Gesamt-Phosphor. Das eingefrorene Myokardgewebe (linker Ventrikel, Ventrikelseptum und Herzspitze) wurde von anhaltendem Blut
1 Thromboeid wurde freundlicherweise von der Fa. Dr. W. Benend, München- Solln, zur Verfügung gestellt.
Phenylbutazon wurde als 20~oige w~ßrige Lösung verwendet, für deren Über- lassung wir der F~. Dr. K. Thomae, Biberaeh a. d. I~iss, danken.
8 F]uoressigs~ure, krist~llln, der Fu. Fluka, St. Gallen, Schweiz.

Phosphat-Verbindungen des Myokards bei Insuffizienz 315
befreit und in einem vorgekühlten Porzellanmörser unter öfterem Naehgießen von flüssigem Stickstoff fein pulverisiert. Das Muskelpulver wurde dannin 2 In] eisgekühlte 5%ige Trichloressigs~ure eingebracht und sofort zurückgewogen. Die Einwaage be- trug gewöhnlich etwa 0,3 g. Bei gleichzeitiger Kühlung ( - - 1 bis 0°C) wurde diese Gewebsmenge 3 min extrahiert und dann durch anschließendes zweimaliges Zen- trifugieren (3 min bei 4000U/min, 2min bei 3000U/min) von dem übers tehenden-- die säurelösliehen P-Verbindungen enthaltenden - - Ext rakt abgetrennt. Zur Krea- tinphosphat-Bestimmung nach EGGLETON u. EGGLETON (1929) wurden sofort 0,5 ml Extrakt abgezweigt. Zur Bestimmung des säurelöslichen Gesamtphosphors wurden außerdem mittels einer ,Agla',-Mikrometerspritze 0,02 ml Extrakt entnommen, mit 0,7 ml Veraschungsreagens nach I-I~NES u. ISltERWOOD (]949), (3 Teile 98%ige H2S04 -t- 2 Teile 60%ige HC]O~) versetzt und 20 min feucht verascht. Alle Phos- phatanalysen erfolgten nach der kolorimetrischen Methode von BERE~BLm~ U. CH~Zg (1938) unter Verwendung eines Elektrophotometers (Elko II , Zeiss-Opton, Filter S 75).
B. Papierehromatographisehe Arbeitswe|se. Der I~est des Extraktes wurde in der Tiefkühltruhe bei - - 1 8 ° C aufbewahrt und kam nach sp/~testens 30 min zur papierchromatographischen Aufarbeitung entsprechend dem von Fr.ECKE~STEI~" U. JAN~:E (1953), FL~CKEZqSTEIN U. Gv.RLAC~ (1953), bzw. FLECKENSTEIN (1955) angegebenen Prinzip. Eine Modifikation gegenüber dem ursprünglichen Verfahren bestand in dem Zusatz von Butanol zum I. Lösungsmittel sowie in dem Austausch des ursprünglichen II . Lösungsmittels gegen die neuen Lösungsmittel H a und I Ib , deren Zusammensetzung von GERLAC~, WEBE~ U. DöRING (1955) ausgearbeitet wurde.
Im einzelnen wurde wie folgt verfahren: Nach Auftropfen der Extrakte mit Ag]a-Mikrometerspritzen (20 Tropfen zu je 0,005 tal) auf Whatman-Chromato- graphiepapiere der Größe 20 × 50 cm (vorbehandelt mit äthylendiamintetraessig- saurere Natrium) wurden alle Chromatogramme zunachst 19--20 Std mit Lösungs- mittel I absteigend chromatographiert.
Zusammensetzung von Lösungsmittel I: Diisopropy]äther . . . . . . 40 ml n-Butanol . . . . . . . . . 30 ml Ameisensäure 80~oig . . . . 25 ml
Dabei trennen sich Orthophosphat -k Kreatinphosphat 1 und eineun bekannte, phosphorhaltige Komponente (,Verbindung X") von den übrigen organischen Phos- phorverbindungen ab. Auf einem besonderen Chromatogramm wurden nur die beiden genannten Fraktionen quantitativ bestimmt. Auf einem zweiten, gleicher- maßen behandelten Papier wurden die Areale, die das gesamte Orthophosphat ein- schließlich Kreatinphosphat enthalten, abgeschnitten und das restliche Papier in entgegengesetzter l%ichtung mit Lösungsmittel I I a und I I b absteigend ehromato- graphiert.
Zusammensetzung von Lösungsmittel Yla und IIb: 8-Oxychinolin . . . . . . . . 18 mg Äthanol . . . . . . . . . . . 5 ml Äthanol . . . . . . . . . . 5 ml n-Propanol . . . . . . . . . 25 ml n-Propanol . . . . . . . . . 25 ml n-Butanol . . . . . . . . . . 30 ml n-Butanol . . . . . . : . . 30 ml Ammoniak 25~oig . . . . . . 30 ml Ammoniak 25~oig . . . . . . 30 ml Wasser . . . . . . . . . . : 10 ml Wasser . . . . . . . . . . . 10 ml
1 Kreatinphosphat wird in Lösungsmittel I bei 18°C hydrolytisch gespalten; der Phosphatantefl wandert dann zusammen mit dem - - schon urspri~hglich in den Extrakten enthaltenen - - Orthophosphat als eine Fraktion.
22*

316 H. I-Iocmz~rs und H. J. Dö~~~G:
Die Anwendungszeß von Lösungsmittel I I a betrug 2 × 24 Std bei 26°C mit einer einstündigen Zwischentrocknung nach den ersten 24 Std. Lösungsmittel I I b wurde im Anschluß an die Behandlung mit Lösungsmittel I I a unter sonst gleichen Bedingungen 1 × 24 oder 2 X 24 Std zur Einwirkung gebracht. Dabei trennten sich ATP, ADP, AMP und 3 nicht näher identifizierte I~ucleotide, von denen zwei wahrscheinlich GTP und GDP sind, voneinander ab. Glucose-6-phosphat wird bei diesem Vorgehen meist nicht vollständig von der ADP-l~raktion getrennt, so d~l] unter Umständen eine nochmalige Anwendung des Lösungsmittels I notwendig wird (Laufriehtung wie bei Lösungsmittel I I a und I Ib , Laufzeit 20 Std). Die Po- sition der Nucleotide veränderte sich hierbei praktisch nicht, während Glucose-6- lohosphat Yon Lösungsmittel I weiter fortbewegt wird.
In 4 Versuchen wurden die Trichloressigsäure-Extrakte zu Kontrollzwecken auch 2-dimensional chromatogralohiert. Hierzu wurden auf einem besonderen Bogen der üblichen Größe nur an einer Stelle der Startlinie 20 × 0,005 ml Extrakt aufgetragen. Die Behandlung erfolgte mit Lösungsmittel I, I I a und I Ib wie üblich. l~aeh der letzten Anwendung von Lösungsmittel I I b wurde senkrecht zur ersten Laufriehtung aufsteigend mit Lösungsmittel I I I [Zusammensetzung: 15 ml Tri- ehloressigsäure (30% ig), 25 ml Ame£sensäure (80%ig), 40 ml n-Butanol, 20 ml n-Pro- panol, 25 ml Aceton; vgl. GERLAeH, W~BEa, DöRrs¢ (1955)] 18 Std chromato- graphiert. Die Analysenwerte für ATP und ADP stimmten mit den eindimensional gewonnenen genau überein. Das 2-dimensionale Vorgehen hat den Vorteil, daß außer ATP und ADP auch A~P, GTP, GDP, ein unbekanntes l~ucleotid, sowie einige phosphorylierte Triosen und Hexoscn erfaßt werden können. In den Chromato- grammen grenzte sich außerdem ein P-haltiges Areal (bis 7% P) von den anderen Verbindungen ab, dessen sichere Identifizierung noch aussteht. Auch Glucose-6- phosphat, Fruetose-l,6-diphosphat und Triosephosphat können nach dem Vor- gehen von ~LEeKE~ISTEI~, JAI'IXE U. GERLAet{ (1957) papierchromatographiseh iso- liert werden [vergl. FL~eXv,~sT~rs (1957)]. In den folgenden Versuchen wurde jedoch auf die Bestimmung dieser kleinen Fraktionen verzichtet.
Ergebnisse I. Die säurelöslichen Phosphorverbindungen
des Meerschweinchen-Herzens in situ
1. Normale ~Ieerschweinchenherzen in E v i p a n - N a r k o s e . Die Her- s te l lung eines H e r z - L u n g e n - P r ~ p a r a t e s i s t zweifellos eine einschneL dende Opera t ion , die bekann t l i ch schon nach einigen S tunde n zu e inem Versagen des ~¢fyokards führ t . Ob dabe i auch S tö rungen des in ter - med iä r en Phosphats tof fwechse ls - - e twa als u n m i t t e l b a r e Folge des operativen Eingriffs -- stärker in den Vordergrund treten, war aus der
Literatur nicht klar ersichtnch. Die erste Aufgabe mußte daher darin bestehen, a m ungeschäd ig ten 1Vieersehweinchen-lViyokard in s i tu , I~or- m a l w e r t e " für die versch iedenen säurelösl iehen P -Verb indungen zu er- mi t te ln , die dan~ als Vergleichsbasis ffir die Ana lysenwer te be im Herz- L u n g e n - P r ä p a r a t d ienen konnten . I n Tab. 1 s ind die bei 6 ungesehäd ig ten Meerschweinchenherzen in s i tu e rmi t t e l t en Ergebnisse zusammen- gestel l t . Die mi t t l e r en K o n z e n t r a t i o n e n der einzelnen P -Verb indungen s ind dabe i jeweils abso lu t in #mo l /g M y o k a r d (bzw. in # m o l P/g Myokard ) sowie r e l a t i v in P rozen t des säurelösl ichen Gesamt -Phosphor s wieder-
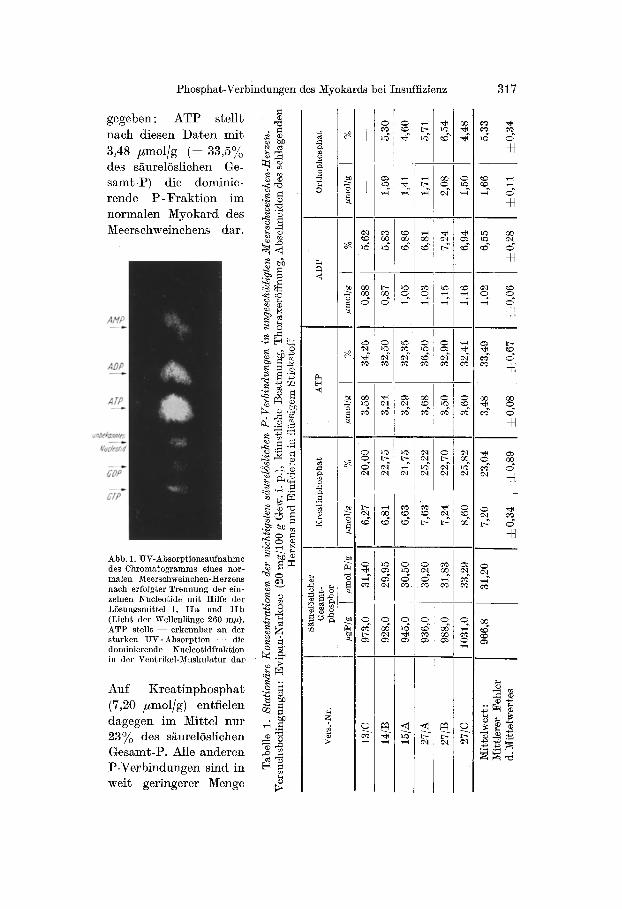
Phosphat-Verbindungen des ~yokards bei Insuffizienz 317
g e g e b e n : A T P s t e l l t
n a c h d i e sen D a t e n m i t
3,48 # m o l / g ( = 33 ,5%
des s äu re lö s l i chen Ge-
s amt - l ) ) d ie d o m i n i e -
r e n d e l) - F r a k t i o n i m
n o r m a l e n M y o k a r d des
M e e r s c h w e i n c h e n s da r .
ünb6
g~
Abb. 1. UV-Absorptionsaufnahme des Chromatogramms eiaes nor- malen Meerschweinchen- t/erzens nach erfolgter Trennung der ein- zelnen Nucleotide mit Hilfe der Lösungsmittel I, IIa und IIb (Licht der Wellenlänge 260 tal0. ATP stell~ - - erkennbar an der starken UV-Absorption --- die dominierende l~ucleotid fraktion in der Ventrikel-Muskulatur dar
A u f K r e a t i n p h o s p h a t
(7,20 # m o l / g ) en t f i e l en
d a g e g e n i m Mi t t e l n u r
23~o des s äu re lö s l i chen
G e s a m t - t ) . Alle a n d e r e n
l ) - V e r b i n d u n g e n s ind in
we i t g e r i n g e r e r Menge
~) va
:©
¢¢
.~ ~0
0~
I I
r :
«q
° 4
-H
oo
¢»
-H
o0
oo
-H
-H
318 H. ttoe~~~iN und H. J. Dö~I~O:
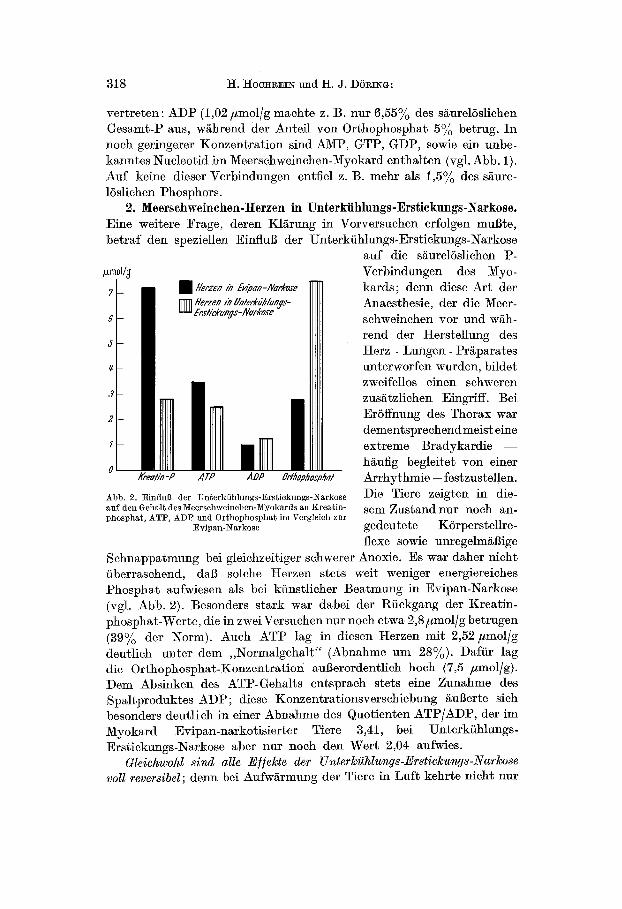
vertreten : ADP (1,02 #mol/g machte z. B. nur 6,55% des säurelöslichen Gesamt-t ) aus, während der Anteil von Orthophosphat 5% betrug. In noch geringerer Konzentrat ion sind AMI ), GTI), GDI), sowie ein unbe- kanntes Nucleotid im Meerschweinchen-Myokard enthalten (vgl. Abb. 1). Auf keine dieser Verbindungen entfiel z. B. mehr als 1,5% des säure- löslichen Phosphors.
2. Meerschweinchen-Herzen in Unterkühlungs-Erstiekungs-Narkose. Eine weitere Frage, deren Klärung in Vorversuchen erfolgen mußte, betraf den speziellen Einfluß der Unterkühlungs-Erstiekungs-Narkose
auf die säurelöslichen l)- pmol/j
7
&
5
«
3 ~H 7
6 Krea/in-P
• gerzen «~ Ev/p«~-Harkose
Ersl/ck«ng.¢-IVarkos«
I~ ,o I ArP ADP OHbopho«p»o!
Abb. 2. Einfluß der Unterkühlungs-Erstickungs-I7arkose auf den Gehalt des lVieerschweinchen-~yokards an Kreabin- phosphat, AT]?, AD~P und Ol'thophospha~ im Vergleich zur
Evipan-~arkose
Verbindungen des Myo- kards; denn diese Art der Anaesthesie, der die Meer- schweinchen vor und wäh- rend der Herstellung des Herz - Lungen- l)räparates unterworfen wurden, bildet zweifellos einen schweren zusätzlichen Eingriff. Bei Eröffnung des Thorax war dementsprechend meist eine extreme Bradykardie - - häufig begleitet von einer Arrhythmie -- festzustellen. Die Tiere zeigten in die- sem Zustand nur noch an- gedeutete Körperstellre- flexe sowie unregelmäßige
Schnappatmung bei gleichzeitiger schwerer Anoxie. Es war daher nicht überraschend, daß solche Herzen stets weit weniger energiereiehes l)hosphat aufwiesen als bei künstlicher Beatmung in Evipan-Narkose (vgl. Abb. 2). Besonders stark war dabei der Rückgang der Kreatin- phosphat-Werte, die in zwei Versuchen nur noch etwa 2,8 #mol/g betrugen (39% der Norm). Auch ATP lag in diesen Herzen mit 2,52 #mol/g deutlich unter dem ,Normalgehal t" (Abnahme um 28%). Dafür lag die Orthophosphat-Konzentration außerordentlich hoch (7,5 #mol/g). Dem Absinken des ATl)-Gehalts entspraeh stets eine Zunahme des Spaltproduktes ADl); diese Konzentrationsversehiebung äußerte sich besonders deutlich in einer Abnahme des Quotienten ATP/ADP, der im Myokard Evipan-narkotisierter Tiere 3,41, bei Unterkühlungs- Erstiekungs-Narkose aber nur noch den Wert 2,04 anfwies.
Gleichwohl sind alle E/]ekte der Unterkühlungs-Erstickungs-Æarkose voll reversibel; denn bei Aufwärmung der Tiere in Luft kehrte nicht nur

Phospha~-Verbindungen des Myokards bei Insuffizienz 319
die Myokard-Funktion, sondern auch der Gehalt an ATP und Kreatin- phosphat zur Norm zurück. Nachträgliche Schäden waren selbst wäh- rend einer Beobachtungsperiode von mehreren Wochen nicht festzu- stehen. Es ist hieraus zu schließen, daß die Unterkühlungs-Erstickungs- Narkose als Anaesthesie-Methode brauchbar ist. Der Vorzug dieses Nar- kose-Verfahrens liegt unter anderem in dem Verzieht auf chemische Narkosemittel, die - - wie z. B. die Barbiturate - - den Eintr i t t der Insuffizienz eines Herz-Lungen-Präparates in bekannter Weise begünstigen können.
H. Die säurelöslichen Phosphor-Verbindungen des Meerschweinchen-Herzens unter den
Bedingungen des Herz-Lungen-Präparates
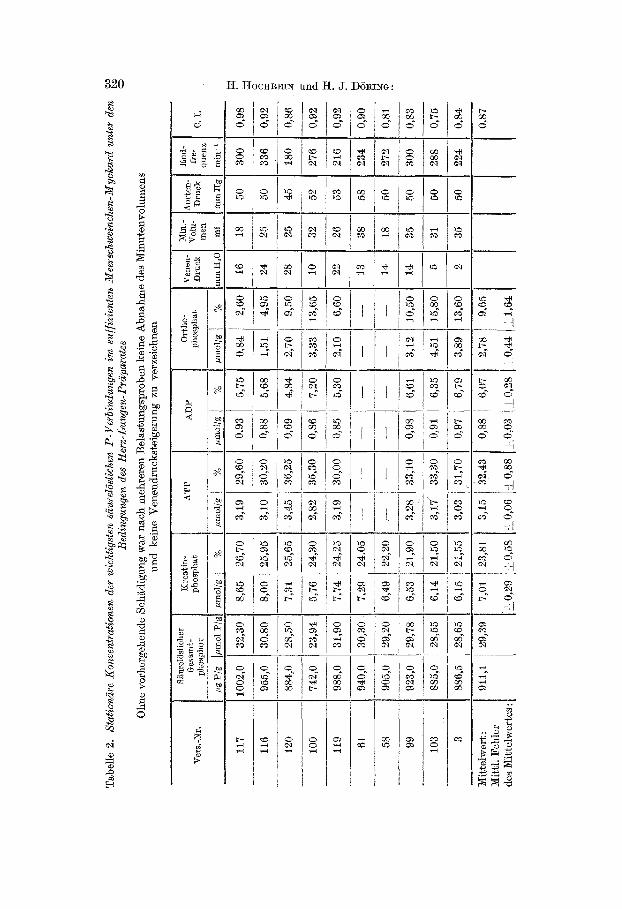
1. Suffiziente Herzen. Als ,suffiziente Herzen" wurden diejenigen bezeichnet, die nach mehreren Belastungen kein Absinken des Minutenvolumens und keine Steigerung des rechten Vorhofdruckes aufwiesen bzw. bei 50 mm hydrostatischer Volumen-Belastung mit einem Druckanstieg von höchstens l0 m m H20 reagierten (C. I. 1,0--0,80) (vgl. Abb. 3). Die Herzen kamen dann auf der Grundstufe der Belastung bei einem mittleren Minutenvolnmen von 28 ml zur Untersuchung. Tab. 2 enthält die an 10 suffizienten tIerz- Lungen-Präparaten gewonnenen Daten für Kreatinphosphat, ATP, ADP und Ortho- phosphat. Von besonderem Interesse ist dabei ein Vergleich mit den in Tab. 1 wiedergegebe- nen ,Normal-Werten" . Tatsächlich ergibt sich aus dieser Gegènüberstellung sehr deutlich, daß das su]]iziente Meerschweinchen-M yol~ard unter den Bedingungen des Herz-Lungen-Präparates praktisch noch den gleichen Gehalt an A T P , ADP und Kreatinphosphat au]weist wie das ungeschädigte Myolcard in situ. Nur die Ortho- phosphat-Frakt ion zeigte bei den Herz-
Abb. 3. Reaktiousweise eines un- geschädigten Herz-Lungen-Prä- parates vom Meerschweinchen im Zustand der Suffizienz. Regis~rie- rung des arteriellen Drucks in m m Hg (oben), des Minutenvolu- mens (5[itte) und des Drucks im reehtenVorhofin m m H20 (unten). Nach 4 maliger Belastung des Her- zens, wobei der Spiegel des Vor- ratsgefäßes jeweils von der Grund- stufe (20 cm über dem Herzen) auf 25 cm und 30 cm erhöht wurde, kehrte der Druck im rechten Vor- hofjeweils zur Ausgangshöhe zu- rück. Das Herz kann deshalb als suffizient bezeichnet werden, weil nach Niveau-Erhöhung des Vor- ratsgefäßes um 50 m m nur eine Erhöhung des Vorhofdruekes mn 5 m m H20 e ia t ra t (Competenee index = 0,90). Dafür stieg das Minutenvolumen von 26 ml auf
36 ml an
Lungen-Präparaten eine gewisse Tendenz zur Erhöhung (mittlerer An- stieg von 5,3% auf 9,7% des säurelöslichen Myokard-Phosphors). Bei geeigneter operativer Technik treten demnach im Herz-Lungen-Präpa- ra t des Meerschweinchens keine nachhaltigen Störungen des myokar- dialcn Phosphat-Stoffwechsels aufi Auch die durch Unterkühlungs-
3 2 0 H. HOCttl~EIN und H. J. DÖRINO:
~ ~ ~ o
O
cD
° ~
bi0
¢9
L~ b~
c9 z.O
õ cD
o
o0
o0 ~o
a'q
o0
¢'0
oo ~q
¢~ o0
l ~ ¢q
¢¢ ¢q
oo
~0
oo oo a'~ a'q
a'q g~ a'q
z~
~0
~z o0
~o LO
o0
o0
~z
¢o
LO ¢.o
oo ",~
~p oo
÷1
oo o'~
c~ ~ oo
I
Phosphat-Verbindungen des Myokards bei Insuffizienz 321
Erstiekungs-Narkose bedingten Veränderungen (Abnahme der ATP- und Kreatinphosphat-Bestände) sind offensichtlich sehr flüchtig. Dieses cha- rakteristische Verhalten der energiereichen P-Verbindungen läßt sieh qualitativ schon aus den in Abb. 4 wiedergegebenen UV-Absorptions- aufnahmen der Adenin-Nueleotide ablesen: I m ungeschädigten Meer- sehweinehen-Myoeard in situ (a) stellt ATP - - erkennbar als s tark UV- absorbierendes Areal - - die dominierende Nueleotid-Fraktion dar. Wäh- rend der Unterkühlungs-Erstik- kungs-Narkose (b) sinkt jedoch die ATP-Frakt ion unter An-
r e i ehe rung von ADP ab; ATP und ADP zeigen nunmehr beinahe gleiehstarke UV-Ab- sorptionen. Nach Fertigstellung des Herz-Lungen-Präparates (e) ist von dieser ADP-Anreiche- ~ rung nichts mehr zu sehen: I m suffizienten Myokard imponiert ...... ATP wieder - - ebenso wie beim ungesehädigten Herzen in situ - - als mengenmäßig stärkstes Adenin-Nueleotid.
2. Herzen bei Schädigung bis zur Suffizienz- Grenze. Durch Infusion von Phenylbutazon a b c ( I0--16 mg/20 ml Blut) oder Abb.~. UV-Absorptionsaufnahmen der Adenin-
Nueleotide des Meersehweinchen-Myokards beim durch extreme Volumbelastnng Herzen in sitn (a), während Unterkählungs-Erstik- kann ein der Snffizienzgrenze kungs-Narkose(b)undimsuffizienten}Ierz-Lnngen-
P räpa ra t (e). Während der Unterkühlungs-Erstik- nahekommendes Dekompensa- kungs-Narkose sinkt die ATP-Frakt ion un~er An- t ionsstadium erreicht werden, reieherung von ADP ab; ATP und ADP weisen
daher im ]gild (b) beinahe gleichstarke UV-Absorp- Der Zeitpunkt des beginnenden tion auf. Im fer t igenHerz-Lungen-Präparat (c) en~-
spricht das Verhältnis derAdenin-Nucleotide wieder Myokard-Versagens ist dabei demungesehädigten:~£yokardinsi tu;ATPimponiert mit Hilfe regelmäßig wieder- wieder als mengenmäßig stärkstes Nncleotid
holtet Volumbelastungen wäh- rend und nach der Phenylbutazon-Infusion ziemlich genau zu er- fassen. Die Versuchsanordnnng im einzelnen ergibt sieh aus Abb. 5 (Experiment Nr. 43). Hier wurde zunächst bei 4maliger Belastung ein an- nähernd normales Verhalten des Minutenvolumens und des rechten Vor- hofdruckes (Anstieg auf 25 mm H~O) registriert. Bei der 5. Belastung t ra ten dann kurz nach der Beendigung der Phenylbutazon-Infusion die ersten Zeichen der beginnenden Insuffizienz zu Tage: Das Minutenvolu- men stieg nicht mehr auf die - - bei den vorausgegangenen Belastungen erreichten - - Werte an; dafür nahm der rechte Vorhofdruck merklich

3 2 2 I t . H o e m ~ ~ i ~ u n d I-I. J . D ö ~ r ~ G :
zu. Noch ausgepr/~gtere Symptome der fortschreitenden Dekompen- sation wurden bei der 6. und letzten Belastung beobachtet. Hier schnellte der Druck im rechten Vorhof auf über 60 mm HeO in die Höhe, ohne daß das Minutenvolumen entsprechend reagierte. Der C. I. betrug in diesem Zustand 0,46. Das Herz wurde daraufhin abgeschnitten und für
die chemische Aufarbei- tung in flüssigem Stick- stoff eingefroren.
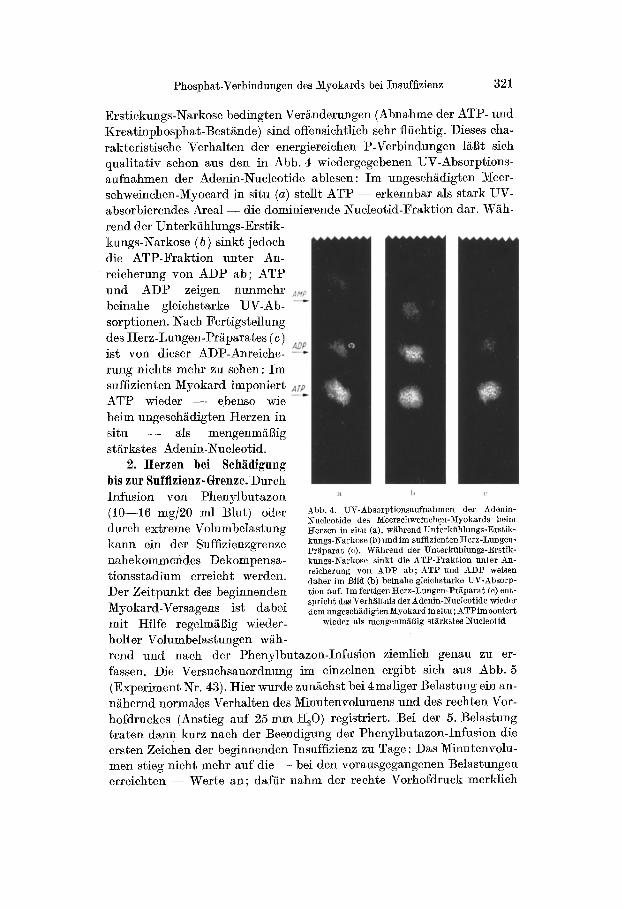
In Tab. 3 sind 8 Ex- perimente dieser Art zu- sammengestellt; der C. I. lag dabei als Ausdruck der Myokard-Schädigung zum Zeit]?nnkt der Herz- Entnahme zwischen 0,30 und 0,70. Obwohl der Zu- stand der einzelnen Her- zen sicherlich nicht gleich- artig war, kann doch aus den Mittelwerten für ATP, ADP, Kreatin]?hosphat und Orthophosphat eine sehr charakteristis cheVer - schiebung dieser Phos- ]?hat-Fraktionen erkannt werden; denn der Gehalt
AbD. 5. l%eaktionsweise des Herz-Lungen-Präparates bei a n Kreatin]?hos]?hat und Phenylbutazon-Sehädigung bis zur Suffizienzgrenze (Ex- ATP war in allen geschg- periment :Nr. 43). Nach l~henylbutazon-Iniusion (Dauer der Infusion von )]larke I bis )£arke I I ) stellen sieh die Zeichen digten Herzen erniedrigt; einer fortschreitenden Dekompensation ein: Bei Niveau- Erhöhung des Vorratsgefäites von 20 cm iiber dem Herzen die Ortho]?hosphat-Werte (Grnndstufe) auf 25 cm und 30 cm, steigt jetzt auch der zeigten dagegen eine rechte Vorhof4ruck beträchtlich an, z. B. beim letzten
Belastungstest auf 60 m m t130 (näheres s. Text) betraehtllche ErhÖhung. Wi~hrend z. B. im unge-
schädigten Myokard 23% des säurelöslichen Phosphors auf Kreatin- phosphat und nur 5,3 ~o auf Orthophos]?hat entfielen, enthielten die Herzen an der Grenze der Suffizienz gleichviel oder sogar mehr Ortho]?hosphat als Kreatin]?hos]?hat. Die Abnahme von Kreatinphos]?hat betrug im Ver- gleich zum normalen Myokard etwa 1/3 des Gesamtbestandes, die ATP- Werte waren auf etwa 85% des ,l~ormal-Wertes" reduziert.
3. Insuffiziente tterzen. Eine über die Suffizienzgrenze hinausgehende Dekompensation (Insuffizienz) wurde in unseren Versuchen durch li~n- gere Phenylbutazon-Einwirkung oder durch Fluoressigsäure-Infusion
'@
o
¢o
¢4
cD
Phosphat-Verbindungen des Myokards bei Insuffizienz
Y-
-H
o~ o o
• o ~
323
324 H. HOCttREISr und H. J. ])öI~ING:
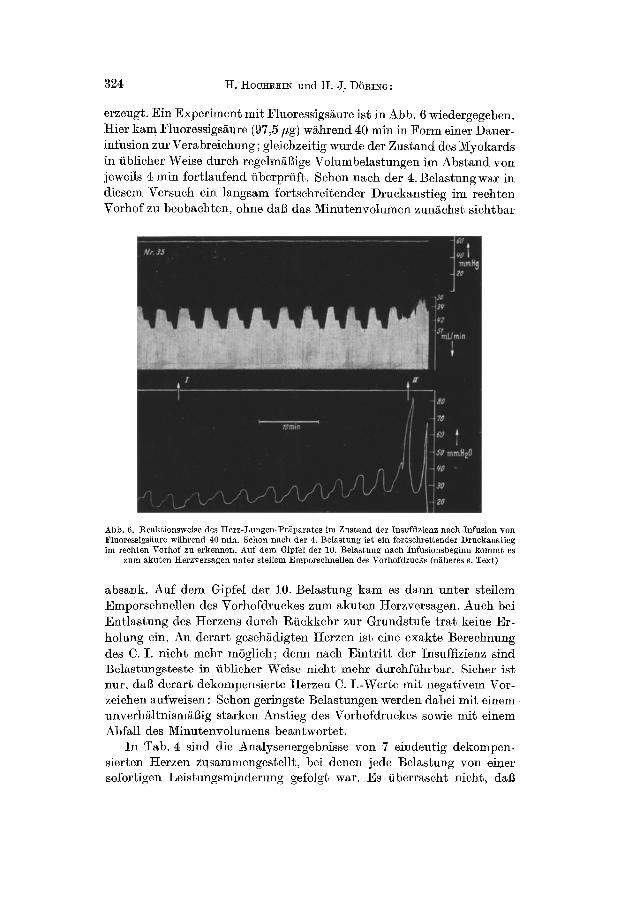
erzeugt. Ein Experiment mit Fluoressigsäure ist in Abb. 6 wiedergegeben. Hier kam Fluoressigsäure (97,5/hg) während 40 min in Form einer Dauer- infusion zur Verabreiehung ; gleichzeitig wurde der Zustand des Myokards in üblicher Weise durch regelmäßige Volumbelastungen im Abstand von jeweils 4 min fortlaufend überprüft. Schon nach der 4. Belastung war in diesem Versuch ein langsam fortschreitender Druckanstieg im rechten Vorhof zu beobachten, ohne daß das Minutenvolumen zunächst sichtbar
Abb. 6. Reaktionsweise des Herz-Lungen-Präparates im Zustand der Insuffizienz nach Infusion von Fluoressigsäm'e während 40 min. Schon nach der 4. Belastung ist ein fortschreitender Druckanstieg im rechten Vorhof zu erkennen. Auf dem Gipfel der 10. Belastung nach Infusionsbeginn kommt es
zum akuten I{erzversagen unter steilem Emporschnellen des Vorhofdrucks (näheres s. Text)
absank. Auf dem Gipfel der 10. Belastung kam es dann unter steilem Emporschnellen des Vorhofdruekes zum akuten Herzversagen. Auch bei Entlastung des Herzens durch Rückkehr zur Grundstufe t ra t keine Er- holung ein. An derart geschädigten Herzen ist eine exakte Berechnung des C. I. nicht mehr möglich; denn nach Eintr i t t der Insuffizienz sind Belastungsteste in üblicher Weise nicht mehr durchführbar. Sicher ist nur, daß derart dekoml0ensierte Herzen C. I .-Werte mit negativem Vor- zeichen aufweisen : Schon geringste Belastungen werden dabei mit einem unverhältnismäßig starken Anstieg des VorhofdI~lekes sowie mit einem Abfall des Minutenvolumens beantwortet .
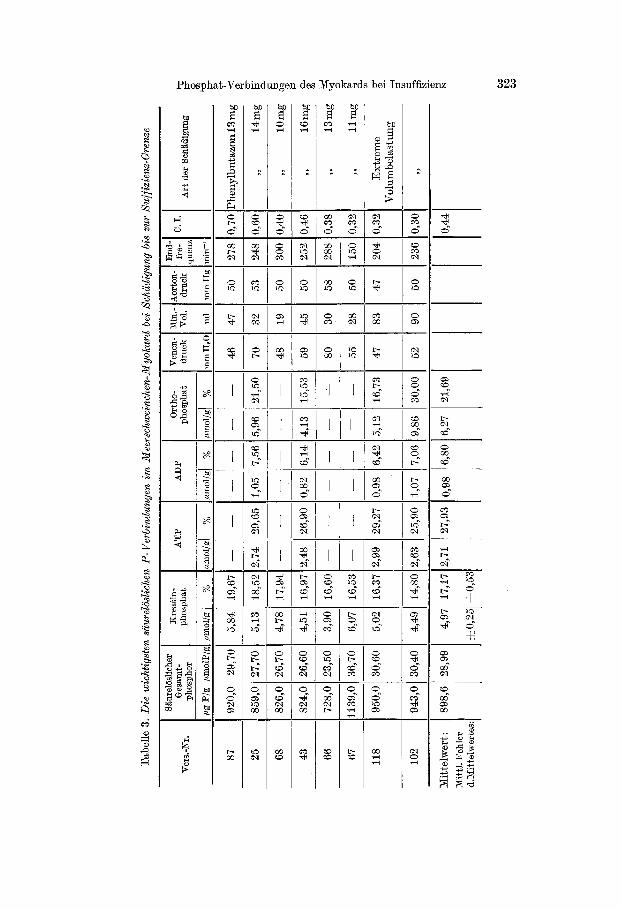
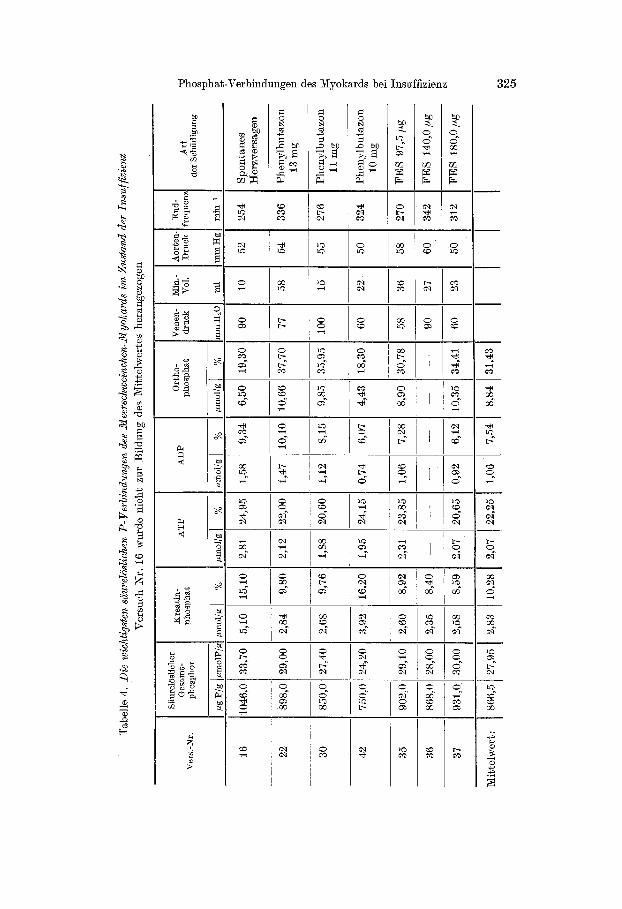
In Tab. 4 sind die Analysenergebnisse von 7 eindeutig dekompen- sierten Herzen zusammengestellt, bei denen jede Belastung von einer sofortigen Leistungsminderung gefolgt war. Es überrascht nicht, daß
,o~ &
b
~'~
~4
o~
4 ©
1)hosphat-Verbindungen des Myokards bei Insuffizienz
r~
o b~
ù~ blO
o k~ © b~
, ~ bi)
©
ù-,1
ù.-t
e~
t ~
~ E
e~
~2
o6
e~
I ®
~ ~ M ~ M d d
æ L-- 4 æ o6 ~
325
cD oo
326 H. Hocm~~i~ und I-I. J. DÖR~G:
die Verschiebungen der intracellulären P-Frakt ionen in diesem Zustand der vollen Insuffizienz noch ausgeprägter sind als an der Grenze der Suffizienz. Dementsprechend war in allen Fällen mehr Orthophosphat als Kreat inphosphat in den insuffizienten Herzen zu finden: Während
Tabelle 5. Relativer Gehalt des Meerschweinchen-Myokards an Kreatinphosphat, A T P, AD P und Orthophosphat bei verschiedenen ~' un~tionszuständen
(Mittelwerte aus Tab. 1--4)
:Normalwer~e bei ~yokard in situ
%
Suffizientes lVlyokard bei t tLP
%
:Hyokard an Suffizienzgrenze
bei HLP %
Insuffizientes ~yokard bei HLP
%
Kreatin-P
ATP
ADP
Ortho- phosphat
23,0
33,5
6,6
5,3
23,8
32,4
6,1
9,7
17,2 10,3
27,9 22,3
6,8 7,5
21,7 31,4
z. B. im Mittel aus 6 Versuchen nur noch 10,3% des säurelöslichen Phos- phors als Kreat inphosphat vorlagen, erreichte die 0r thophosphat-Frak- tion mit 31,4% einen besonders hohen ABteil. Auch die ATP-Werte zeigten eine Senkung auf etwa 2/3 des Normalbestandes. Die beste Über- sicht über diese charakteristischen Beziehungen zwischen dem Leistungs- zustand des Myokards und dem relativen Gehalt an ATP, ADP, Kreat in- phosphat und Orthophosphat ergibt sich aus Tab. 5.
~o
o , « - - -
i g,2
0
/o o foo
i o
J
;'2 lq 16" /8 20 22 2q 26" 222 #,"~,a/l»p/l~pha/ [/z % vom sJufel~2l Æex. -P)
Abb. 7. Beziehung zwischen Competence iudex und KreatinphosDhat-Gehalt des ~yokards (Werte aus Tab. 2 von ungesch~digten Herz-Lungen-Präparaten und aus Tab. 3 von gesch~dig~en Herzen mit
eingesehränkter Leistungsreserve). Es ergibt sich, dai] miß abnehmender Leistungsfähigkeit des ttcrzmuskels (sinkender C. I.) ein niedrigerer Kreatinphosphat-Gehalt gefunden wird
Tatsächlich scheint der Ab/all an energiereichem Phosphat bzw. der An-
stieg der Orthophosphat-Fra/ction ~eweils um so grSfier zu sein, ein je stär- keres Ausmaß die - - zum mechanischen Versagen des Herzens/ührende - - "

Phosphat-Verbindungen des Myok~rds bei Insuffizienz . 327
Myokardschädigung besitzt. Der gleiche Schluß ergibt sich auch aus Abb. 7; hier ist der Kreatinphosphatgehalt der Meerschweinehenherzen mit den gleichzeitig gemessenen C. Z.-Werten in Beziehung gesetzt.
Besprechung Die dargelegten Untersuchungen zeigen zunächst, daß der unge-
schädigte Herzmuskel des Meerschweinchens unter den Bedingungen des Herz-Lungen-Präparates i~hnlich hohe ATP- und Kreatinphosphat- Bestände aufweist wie das intakte Meerschweinchenherz in situ. Der operative Eingriff bei der Herstellung des Herz-Lungen-Präparates hat also bei geeigneter Technik keine nachhaltige Beeinträchtigung des intermediären Phosphatstoffwechsels zur Folge. Das Herz-Lungen-Prä- parat des Meerschweinchens bietet dementsprechend gute Ausgangs- bedingungen, um das Verhalten der energiereichen Phosphate bei ver- schiedenen Funktionszuständen des Herzmuskels zu prüfen. Auch das Herz-Lungen-Präparat des Hundes weist nach Ergebnissen von F~wAz u. ttAWA (1953) bei guter Beatmung ähnlich hohe ATP- und Kreatin- phosphat-Bestände auf wie das Hunde-Myokard in situ. Davon abwei- chende Ergebnisse anderer Autoren [Bl;a~s u. C~~rlCXSHA~K (1947); POLLACK, FLOCK, ESSEX u. BOLLMANN (1934); WOLL~,~]~]~~GER (1947)] deuten allerdings darauf hin, daß sich beim Nerz-Lungen-Präparat des Hundes häufig auch größere Differenzen gegenüber dem Herzen in situ einstellen können. Das Meerschweinchen scheint daher unter Umständen für derartige Studien besser geeignet zu sein als der Hund. [Hocm~EI~, WLLKE U. ST~PAZXKXS (1958)].
Das Herz-Lungen-Präparat stellt wohl die beste experimentelle Me- thode dar, um in einfacher und übersiehtlieher Weise den Funktions- zustand des Herzmuske]s durch Messung des arteriellen Widerstandes, Minutenvolumens und rechten Vorhofdrueks zu erfassen, Insbesondere ist es auf diese Weise möglich, mittels wiederholter Belastungstests den sog. ,Competence index" zu erreehnen, der jeden Leistungszustand des Herzens exakt wiedergibt. In den vorliegenden Untersuchungen konn- ten so 3 Gruppen von Herzen abgegrenzt werden:
a) Su/]iziente Herzen mit einem Competence index von 1,0--0,80;
b) Herzen mit eingeschränkter Leistungsreserve infolge Phenylbutazon- Verabreiehung oder Überlastung (Competence index 0,70--0,30) ;
cJ Insu/]iziente Herzen infolge von Spontanversagen, Phenylbutazon- oder Fluoressigsäure -Vergiftung.
Diesen Veränderungen des Funktionszustandes gehen nach unseren Er- gebnissen charakteristische Veränderungen im Gehalt des Myokards an ener- giereichen Phosphaten parallel:Als besonders au/]älliges Kriterium der sich ausbildenden Insu]/izienz muß dabei der ]ortschreitende Zusammenbruch

328 H. HocII~EIN und H. J. Dö~ING:
der Kreatinphosphat-Bestände - - unter steilem Anstieg der Orthophosphat- Fraktion - - gelten, während die A TP-Wer te weniger reduziert werden. Die Myokardinsuffizienzen unter dem Einfluß von Phcnylbutazon, Fluoressig- säure und Chinin [BOGATZKI U. STARB (1956)] lassen in dieser Hinsicht deutliche Gemeinsamkeiten erkennen. Ähnliche Verhältnisse dürften auch beim Eintr i t t einer Insuffizienz infolge extremer Volumbelastung oder bei spontaner Insuffizienz des isolierten Herzens vorliegen [vgl. MÜGGE (1933); W~IC~:~~ (1935)]. Die eingangs erwähnte Zunahme von ATP und Kreatinphosphat beim Herzvcrsagen nach Barbiturat-Ver- abreichung dürfte also nur ein Spezialfall sein, der nicht der Regel ent- spricht. Von Interesse ist, daß auch der Zusammenbruch der bioclektri- schen und mechanischen Tätigkeit des Myokards bei Anoxie, akuter Co- ronar-Unterbindung oder Vergiftung des Herzens mit 2,4-Dinitrophenol stets von einem tiefen Absturz der Kreatinphosphat-Wcrte begleitet ist [vgl. FL]~CXE~STEIN (1957)]. Wir glauben, daß alle diese Befunde nicht nur für die spezielle tterzphysiologie Bedeutung haben, sondern Ausdruck einer allgemeinen Gesetzmäßig]ceit sind, die auch für andere erregbare Gewebe (Nerv, Gehirn, quergestreifter Muskel, elektrisches Organ von Fischen) Gültigkeit besitzt: In allen diesen Gewebcn kommt es nämlich zu einer Erschöpfung der bioelektrischen bzw. mechanischen Tätigkeit, sobald die Kreatinphosphat-Bestände auf niedrige Werte sinken [vgl. hierzu LII~G u. G~I~AI~D (1949) ; G]~I~LAC~I, Döl~ING u. FL~CK~EN- STEI~ (1958); FLECKEI~STEIb! (1955); NACl~MANSOI~N, Cox, COAT~S U. M_~CH)~DO (1943)]. Über die Natur dieser speziellen Verknüpfung zwi- schen Krcatinphosphat-Gehalt und Funktionszustand herrscht heute noch keine definitive Klarheit. Dies gilt sicherlich auch für das Myokard; tiefere Einblicke in die Zusammenhänge zwischen tlerzlcistung und Krea- t inphosphat dürften daher erst möglich sein, wenn sich die zugrunde liegende allgemeinere Gesetzmäßigkeit dem Verständnis erschließt.
Summary ATP is the most prominent phosphorus compound in the guinea pig's
heart muscle since more than 32~o of the acid soluble phosphorus are eovered by ATP whereas creatine phosphate (24%) and ADP (6o/0) de Iiot reach the high ATP level. No major difference could be found bet- ween these values obtained on the heart in s~tu and the amounfs of or- ganophosphatcs analysed in the myocardium under the special condi- tions of the heart-hlng preparation. Thus in our experiments all changes of the cardiac mctabolism necessarily connected with this operation as weh as additional disturbances due to a special way of anesthesia (coo- ling of the animals in an achte state of anoxia) were quickly reversible. Obviously the concentrations of the high-energy phosphate compounds remain fairly constant as long as the heart- lung preparations will be~ in

Phosphat-Verbindungen des Myokards bei Insuffizienz 329
a stare of normal competence. This s ta te can be measured by determina .
t ion of the cardiac reserve capacity at different levels of venous inflow,
the normal competence index being 1,0 to 0,8. If, however, the hear ts were damaged by infusion of phenylbu tazone or fluoroacetic acid crea- t ine phosphate was di lninished more and more in parallel to the extent of the myoeardia l weakness. Therefore in hearts with a competence index ranging from 0,70 to 0,30 ereatine phosphate was found to be redueed to 2/3 of the normal , whereas the creatine phosphate eontent was as low as 1/3 of the normal if the ihll stare of ineompetenee had developped. I n this ease A T P too was clearly affeeted showing a reduct ion to 7/10 of the normal .
Literatur
BEGO¥IC, S., u. P . STERN: ])aS Ca rd io -Pu ]mona l -P räpa ra t der Ra t t e . Naunyn- Schmiedeberg's Arch. exp. Path. Pharmak. 219, 333 (1953).- BERENRLV~ä, I., and E. CHAI~: An improved method for the colorimetric determination of phosphate. Biochem. J. 82, 295 (1938). - - BOGATZKI, M., U. H. STAUB: ATP-System und übri- ger Phosphatstoffwechsel im normalen und akut-insuffizienten Herzmuskel unter der Behandlung mit Herzglykosiden. Z. ges. exp. Med. 127, 425 (1956). - - BRAVl% W., H. ALTHOFF U. 1~. TAUGNER: Bubble fiow mete r mi t au tomat i sche r Registr ie- rung. Naunyn-Schmiedeberg's Arch. exp. Path. Pharmak. 222, 529 (1954). - - BVRNS, W., and E. W. H. CRUICKSttANK: Changes in creatine, phosphagen and adenylpyrophospate in relation to the gaseons metabolism of the heart J. Physiol. (Lond.) 91, 314 (1937). - - EGGLETON, G. P., and P. EGOnETON: A method of esti- mating phosphagen and some other phosphorns eompounds in muscle tissue. J. Physiol. (Lond.) 68, 193 (1929).-- F~wAz, G., and E. S. HAWA: Phosphoerea~ine con- tent of mammalian eardiac musc]e. Proc. Soc. exp. Biol. (N. Y.) 84, 277 (1953). - - FLECKENSTEIN, A., u. E. GERLACH: Papierchromatographische Trennung von Adenosintriphosphat, Adenosindiphosphat und anderen Phosphor-Verbindungen bei Säugetier- und Taubenerythrocyten verschiedenen K+-Gehalts. Naunyn- Sehmiedeberg's Arch. exp. Path. Pharmak. 219, 531 (t953). - - FLECKENSTEIN, A., u. J. JAI~~KE: Papierchromatographische Trennung von ATP, ADP und anderen Phosphor-Verbindungen im kontrahierten und erschlafften Froschmuskel. Pflügers Arch. ges. Physiol. 258, 177 (1953) . - FLECKENSTEIN, A.: Der Kalium-Natrium- Austausch als Energieprinzip in Muskel und Nerv. Berlin, Göttingen, Heidelberg: Springer-Verlag 1955. - - FLECKENSTEIN, A. : Der Herzmuskelstoffwechsel und seine Abhängigkeit von der Coronardurchbhitung. Referat Internat. Symposion über Pro- bleme um die Coronarthrombose, Wien 21.--22.6. 1957. Wien. Z. inn. Med. 89, 69--83 (1958). - - GERLACK, E., E. WEBER U. ~-[. J. DÖRING: Einige neueLösungsmi t t e l für die Papierchromatographie von Phosphorsäure-Estern. Naunyn Schmiedeberg's Arch. exp. Path. Pharmak. 226, 9 (1955). -- GERL~C~r, E., H. J. DöRING U. A. FLECKEN- STEIN: Papierchromatographische Studien über die Adenin- und Guanin-Nucleo- tide sowie andere säurelösliche Phosphor-Verbindungen des Gehirns bei Narkose, Ischämie und in Abhängigkeit von der Technik der Gewebsentnahme. Pflügers Arch. ges. Physio] 266, 266 (1958). - - GIAJA, J. : Sur l'asphyxie par confinement ~ ]a neu- tralité thermique. Bull. Acad. Sei. math. e~ natur., B. Sei. n~tur. 6, 215 (1940). - - HAI~ES, C. S., and F. A. ISHERWOOD: Separation of the phosphoric esters on the filter paper chromatogram. Nature (Lond.) 1949, 1107. - - I{OCltREI~I, ]{., H. J. ~TILKE U. ~V~. STRAPAZAKIS ." Experimentelle Insuffizienz am Herz-Lungen-Präparat des Meerschweinchens. Naunyn-Schmiedeberg's Areh. exp. Path. Pharmak. 232,
Pflügers Arch. ges. Physiol.. Bd. 267 23

330 H. HOC~R~r~ u. H. J . Dö~I~G: Phosphat-Verblndungen des Myokards
535 (1958). - - KRiY~I~, O. : Versuche am insufilzienten Herzen. I~aunyn-Schmiede- berg's Arch. exp. Path. Pharmak. 162, 1 (1931). - - LIEG, G., and R. W. GERARD : The membrane potential and metabolism of musc]e fibers. J. cell. comp. Physiol. 34, 413 (1949). - - M~GG~, H. : Eine Erweiterung der Methode zur Bestimmung der Phosphorfraktionen im Muskelgewebe, unter besonderer Berücksichtigung der Phosphagenbestimmung im Warmblüterherzen. lk!aunyn-Schmiedeberg's Arch. exp. Path. Pharmak. 173, 333 (1933). - - NACn~A~SO~N, D., R. T. Cox, C. W. COATES and A. L. ~Acm~»o: Aetion potential and enzyme activity in the electric organ of Electrophorus eleetricus. I I . Phosphocreatine as energy source of the action poten- tial. J. Neurophysio]. 6, 383 (1943). - - POLLACK., E. FLOCK, A. E. ESS~X and J. L. BOLL~L~~: Phosphorus compounds in the perfused heart of the dog. Amer. J. Phy- sio]. l lO, 97 (1943). - - W~ICXÊR, B. : Stoffwechselwirkung des Strophanthins am Warmblüterherzen. I~aunyn-Schmiedeberg's Arch. exp. Path. Pharmak. 178, 524 (1935). - - WOLLE~BV.RaV.~, A. : On the energy-rich phosphate supply of the fafling heart. Amer. J. Physiol. 150, 733 (1947).
Dr. H. I-Ioc~REIN u. Dr. H, J. I)ÖRING, Physio]og. Insti tut der Albert-Ludwigs- Universität, Freiburg i. Br., Katharinenstral]e 29