Embed Size (px)
Citation preview

Pedosphere 24(2): 251–257, 2014
ISSN 1002-0160/CN 32-1315/P
c© 2014 Soil Science Society of China
Published by Elsevier B.V. and Science Press
Decomposition of Surface-Applied and Soil-Incorporated Bt Maize Leaf
Litter and Cry1Ab Protein During Winter Fallow in South Africa∗1
A. KAMOTA1, P. MUCHAONYERWA2,∗2 and P. N. S. MNKENI3
1Department of Crop Science, Bindura University of Science Education, Private Bag 1020, Bindura (Zimbabwe)2School of Agricultural, Earth and Environmental Sciences, University of KwaZulu-Natal, Private Bag X01, Scottsville 3209 (South
Africa)3Department of Agronomy, University of Fort Hare, Private Bag X1314, Alice 5700 (South Africa)
(Received April 17, 2013; revised January 21, 2014)
ABSTRACT
Unintended effects of genetic modification on chemical composition of Bt maize leaf litter may have impacts on its decomposition.
In most agricultural systems in South Africa, maize litter is either left on the soil surface or incorporated into the soil during tillage.
A litterbag experiment, using leaf litter of three maize hybrids (DKC80-12B, DKC80-10 and DKC6-125), was carried out at the
University of Fort Hare Research Farm, South Africa, to determine the effects of genetic modification on decomposition of maize leaf
litter when left on the soil surface under field conditions between July and November, the normal fallow period, in 2008. Another
litterbag experiment was conducted at the University of Fort Hare Research Farm and Zanyokwe Irrigation Scheme, South Africa,
using leaf litter of two maize hybrids genetically modified with the cry1Ab gene (MON810), DKC75-15B and PAN6Q-308B, and their
corresponding near isolines, CRN3505 and PAN6Q-121. The degradation of Cry1Ab protein in the litter, both surface-applied and
soil-incorporated, was also investigated. Decomposition of Bt maize litter was similar to that of non-Bt maize litter both when applied
on the surface and when incorporated into soil. Soil-incorporated litter, as well as its Cry1Ab protein, decomposed faster than that
applied on the surface. The leaf litter C:N ratios of PAN6Q-308B and PAN6Q-121 were similar throughout the study, whereas those of
DKC75-15B and CRN3505 declined by similar amounts during a 12-week period. These findings suggested that decomposition of leaf
litter of Bt maize, with the MON810 event, was not affected by maize genetic modification, and that the Cry1Ab protein broke down
together with plant leaf litter during the winter fallow regardless of whether the litter was applied on the soil surface or incorporated
into soil.
Key Words: genetic modification, litterbag experiment, maize hybrid, MON810 event, protein degradation
Citation: Kamota, A., Muchaonyerwa, P. and Mnkeni, P. N. S. 2014. Decomposition of surface-applied and soil-incorporated Bt
maize leaf litter and Cry1Ab protein during winter fallow in South Africa. Pedosphere. 24(2): 251–257.
INTRODUCTION
The cry1Ab gene in Bt crops may alter compo-
sitional quality with possible reduction in litter de-
composition and nutrient release. In most agricultural
practices in South Africa, maize litter is either left on
the surface as mulch in no-till systems or incorporated
into the soil to depths from 10 to 20 cm during mold-
board plough tillage operations. Leaving maize litter
on the surface, including that of Bt maize, is a com-
mon practice where conservation agriculture has been
adopted. It is estimated that dry matter of 6 and 2.5
t ha−1 is left in the field after harvesting maize grain
and plant materials for silage, respectively, and the re-
cycling of nutrients in this litter is governed by the rate
of its decomposition (Zwahlen et al., 2003).
Decomposition of litter is influenced by soil micro-
bial composition, activities of enzymes (Flores et al.,
2005), soil conditions, particularly moisture, tempera-
ture and pH (Donnelly et al., 1990) and lignin and
polyphenol contents and C:N ratio of the litter (Palm
et al., 1999). There are conflicting reports on the effect
of genetic modification on the chemical composition of
Bt maize plant materials.
Mungai et al. (2005) did not find any consistent
differences in the lignin content between Bt maize and
non-Bt near isolines. However, Saxena and Stotzky
(2001) observed a significantly higher lignin content in
the vascular bundle sheath and the surrounding cells
of Bt maize as opposed to their corresponding non-
Bt near isolines, with the highest lignin content being
found in Bt maize cultivars with event Bt11 followed
∗1Supported by the National Research Foundation of South Africa (NRF) and the Govan Mbeki Research and Development Center
(GMRDC) of the University of Fort Hare (No. GUN62299).∗2Corresponding author. E-mail: [email protected].
252 A. KAMOTA et al.
by MON810, with event 176 having the least. Flores
et al. (2005) and Daudu et al. (2009) also reported
significantly higher lignin contents in Bt maize.
In most studies on decomposition of Bt maize litter,
different hybrids with different C:N ratios and lignin
contents among other components are used. These vari-
ations in the background chemical compositions, due
to varietal differences, could be responsible for most of
the differences in decomposition. The effects of genetic
modification on decomposition of maize litter have,
however, not been consistent. Lehman et al. (2008)
and Daudu et al. (2009) reported similarities in de-
composition of litter of transgenic maize lines and their
near isolines incorporated in soil even though, in some
cases, lignin contents and C:N ratios were elevated in
Bt maize. On the other hand, Flores et al. (2005) re-
ported lower decomposition rates of Bt maize residues
than their non-Bt near isolines in a laboratory experi-
ment, which was attributed to differences in the lignin
content but not to C:N ratios or microbial activity.
Environmental conditions could also have an ef-
fect on decomposition of Bt maize litter (Lehman et
al., 2008), and this has generally been ignored. Don-
nelly et al. (1990) reported that microbial biomass and
the decomposition of lignin and cellulose in forest soils
increased with increases in soil moisture and tempe-
rature, with moisture being the dominant factor. In
South Africa, maize litter is either left on the surface
or incorporated into the soil during the fallow period,
which is characterized by cool dry soil conditions (May
to August) followed by a warm dry period. The pre-
sence of higher lignin contents in Bt maize, coupled
with unfavorable conditions during the fallow winter
period, could result in greater persistence of the litter
than reported in previous studies that were carried out
during warmer and wetter months of the year. This
could have significant implications on nutrient release
and availability for the subsequent crop, while provi-
ding an advantage of sufficient soil cover required in
conservation agriculture systems.
Some studies have investigated the decomposition
of Bt litter buried in the soil but little has been
done regarding surface-applied litter under field con-
ditions. With the increase in adoption of transgenic
crops in South Africa (James, 2007), it is imperative
to understand the fate of litter of locally grown Bt
maize hybrids when incorporated in the soil or left on
the soil surface. This study was carried out to deter-
mine the effect of genetic modification on decomposi-
tion of surface-applied and soil-incorporated Bt maize
(MON810) leaf litter and their Cry1Ab protein under
winter fallow field conditions.
MATERIALS AND METHODS
Site description
This study was carried out at the University of Fort
Hare (UFH) Research Farm (32◦47′ S; 26◦50′ E; 508 m
above sea level (asl)) in Alice and at the Zanyokwe Irri-
gation Scheme (ZIS) (32◦43′ and 37◦47′ S; 27◦01′ and
27◦07′ E; 520 m asl) in Eastern Cape Province, South
Africa. The UFH Research Farm experiences a semi-
arid climate, receiving a mean annual rainfall of 575
mm and a mean annual temperature of 18.1 ◦C (Van
Averbeke and Marais, 1991). The soil is classified as
an Oakleaf using the South African soil classification
system (Soil Classification Working Group, 1991) and
a Haplic Cambisol (Eutric) using the World Reference
Base for Soil Resources (Fey, 2010), with 64.2% sand,
16.0% silt, 19.8% clay, 8.4 g kg−1 organic carbon and a
pH of 6.2 (water) (Mandiringana et al., 2005). The ZIS
experiences a temperate to warm sub-humid climate
and receives an average annual rainfall of 590 mm. The
soil is a Valsrivier (the South African soil classification
system) (Soil Classification Working Group, 1991) or
a Haplic Luvisol (Chromic) (the World Reference Base
for Soil Resources) (Fey, 2010), with 66% sand, 22%
silt, 12% clay, 4.4 g kg−1 organic carbon and a pH
of 6.5 (water). Ambient temperatures and rainfall du-
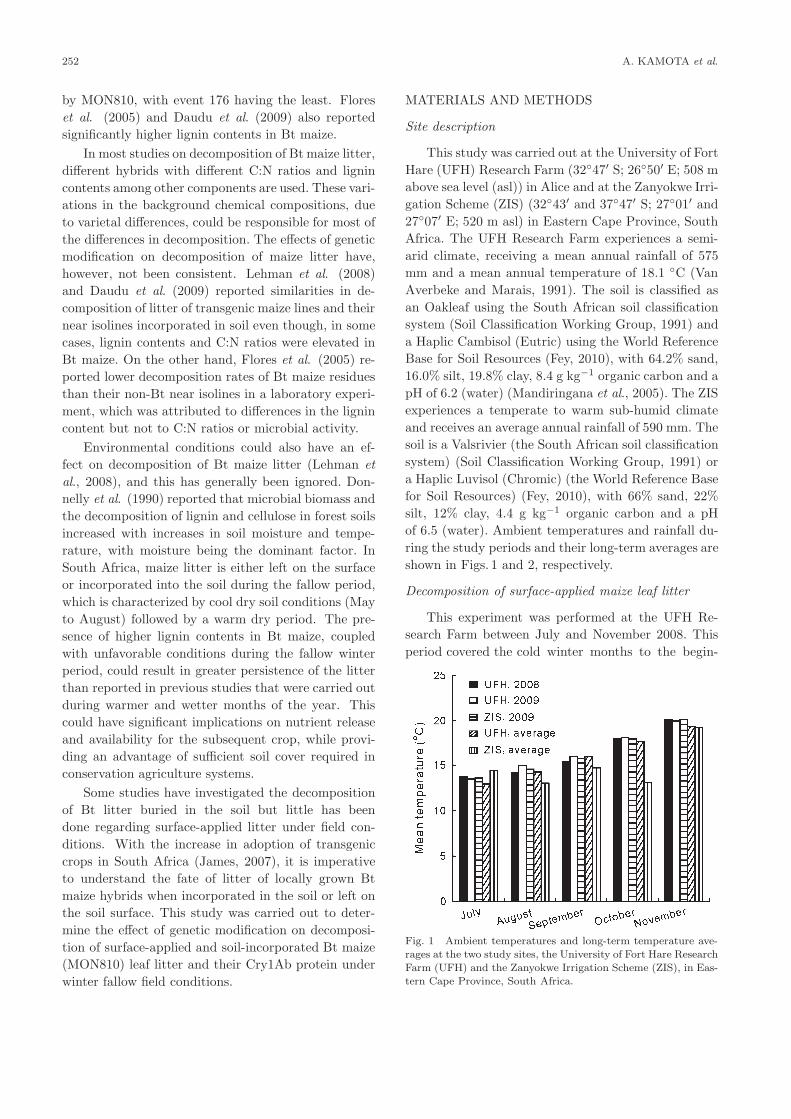
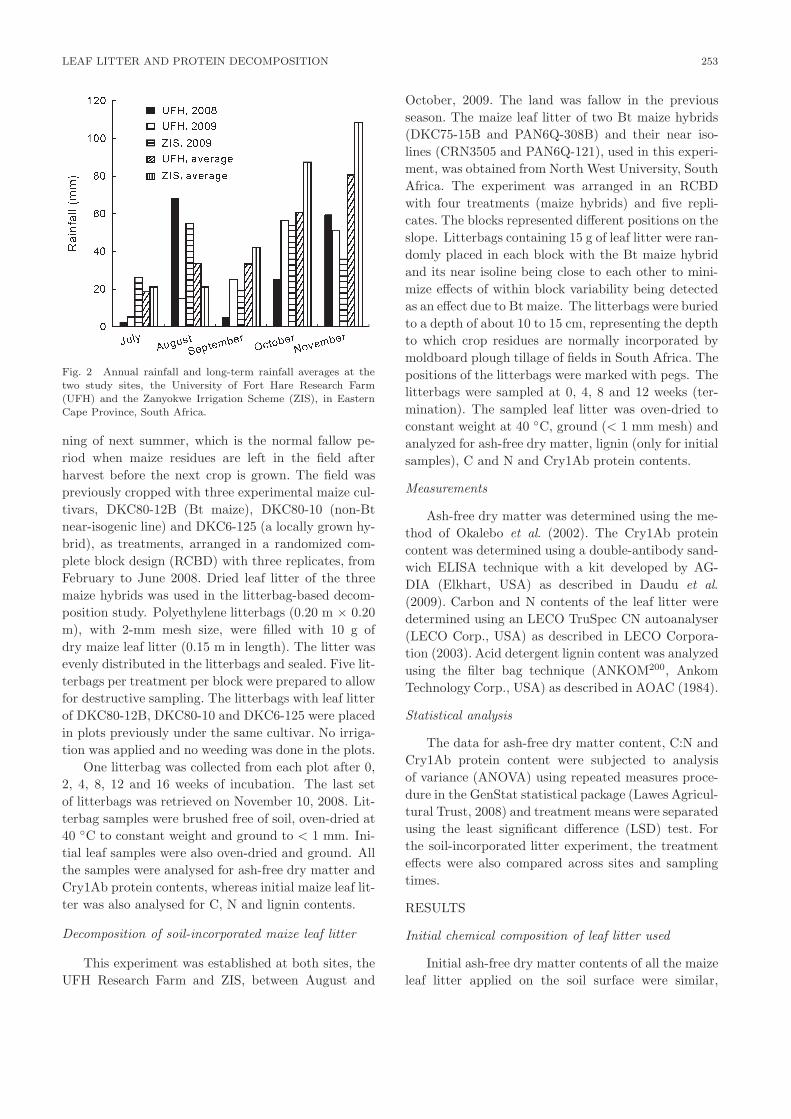
ring the study periods and their long-term averages are
shown in Figs. 1 and 2, respectively.
Decomposition of surface-applied maize leaf litter
This experiment was performed at the UFH Re-
search Farm between July and November 2008. This
period covered the cold winter months to the begin-
Fig. 1 Ambient temperatures and long-term temperature ave-
rages at the two study sites, the University of Fort Hare Research
Farm (UFH) and the Zanyokwe Irrigation Scheme (ZIS), in Eas-
tern Cape Province, South Africa.
LEAF LITTER AND PROTEIN DECOMPOSITION 253
Fig. 2 Annual rainfall and long-term rainfall averages at the
two study sites, the University of Fort Hare Research Farm
(UFH) and the Zanyokwe Irrigation Scheme (ZIS), in Eastern
Cape Province, South Africa.
ning of next summer, which is the normal fallow pe-
riod when maize residues are left in the field after
harvest before the next crop is grown. The field was
previously cropped with three experimental maize cul-
tivars, DKC80-12B (Bt maize), DKC80-10 (non-Bt
near-isogenic line) and DKC6-125 (a locally grown hy-
brid), as treatments, arranged in a randomized com-
plete block design (RCBD) with three replicates, from
February to June 2008. Dried leaf litter of the three
maize hybrids was used in the litterbag-based decom-
position study. Polyethylene litterbags (0.20 m × 0.20
m), with 2-mm mesh size, were filled with 10 g of
dry maize leaf litter (0.15 m in length). The litter was
evenly distributed in the litterbags and sealed. Five lit-
terbags per treatment per block were prepared to allow
for destructive sampling. The litterbags with leaf litter
of DKC80-12B, DKC80-10 and DKC6-125 were placed
in plots previously under the same cultivar. No irriga-
tion was applied and no weeding was done in the plots.
One litterbag was collected from each plot after 0,
2, 4, 8, 12 and 16 weeks of incubation. The last set
of litterbags was retrieved on November 10, 2008. Lit-
terbag samples were brushed free of soil, oven-dried at
40 ◦C to constant weight and ground to < 1 mm. Ini-
tial leaf samples were also oven-dried and ground. All
the samples were analysed for ash-free dry matter and
Cry1Ab protein contents, whereas initial maize leaf lit-
ter was also analysed for C, N and lignin contents.
Decomposition of soil-incorporated maize leaf litter
This experiment was established at both sites, the
UFH Research Farm and ZIS, between August and
October, 2009. The land was fallow in the previous
season. The maize leaf litter of two Bt maize hybrids
(DKC75-15B and PAN6Q-308B) and their near iso-
lines (CRN3505 and PAN6Q-121), used in this experi-
ment, was obtained from North West University, South
Africa. The experiment was arranged in an RCBD
with four treatments (maize hybrids) and five repli-
cates. The blocks represented different positions on the
slope. Litterbags containing 15 g of leaf litter were ran-
domly placed in each block with the Bt maize hybrid
and its near isoline being close to each other to mini-
mize effects of within block variability being detected
as an effect due to Bt maize. The litterbags were buried
to a depth of about 10 to 15 cm, representing the depth
to which crop residues are normally incorporated by
moldboard plough tillage of fields in South Africa. The
positions of the litterbags were marked with pegs. The
litterbags were sampled at 0, 4, 8 and 12 weeks (ter-
mination). The sampled leaf litter was oven-dried to
constant weight at 40 ◦C, ground (< 1 mm mesh) and
analyzed for ash-free dry matter, lignin (only for initial
samples), C and N and Cry1Ab protein contents.
Measurements
Ash-free dry matter was determined using the me-
thod of Okalebo et al. (2002). The Cry1Ab protein
content was determined using a double-antibody sand-
wich ELISA technique with a kit developed by AG-
DIA (Elkhart, USA) as described in Daudu et al.
(2009). Carbon and N contents of the leaf litter were
determined using an LECO TruSpec CN autoanalyser
(LECO Corp., USA) as described in LECO Corpora-
tion (2003). Acid detergent lignin content was analyzed
using the filter bag technique (ANKOM200, Ankom
Technology Corp., USA) as described in AOAC (1984).
Statistical analysis
The data for ash-free dry matter content, C:N and
Cry1Ab protein content were subjected to analysis
of variance (ANOVA) using repeated measures proce-
dure in the GenStat statistical package (Lawes Agricul-
tural Trust, 2008) and treatment means were separated
using the least significant difference (LSD) test. For
the soil-incorporated litter experiment, the treatment
effects were also compared across sites and sampling
times.
RESULTS
Initial chemical composition of leaf litter used
Initial ash-free dry matter contents of all the maize
leaf litter applied on the soil surface were similar,

254 A. KAMOTA et al.
whereas the C:N ratio of DKC80-10 leaf litter was lo-
wer than those of DKC80-12B and DKC6-125 leaf lit-
ter, which were similar (Table I). The lignin content of
DKC80-12B leaf litter was lower than that of its near-
isogenic line leaf litter. The initial ash-free dry matter
contents of all the soil-incorporated litter were similar
across the maize varieties (Table I). The leaf litter C:N
ratio of PAN6Q-308B was similar to that of PAN6Q-
121 and lower than that of CRN3505, which was also
lower than DKC75-15B. Only PAN6Q-121 leaf litter
had a higher lignin content than the leaf litter of the
other three varieties. The leaf litter of DKC75-15B had
a higher Cry1Ab protein content than that of PAN6Q-
308B.
Decomposition of Cry1Ab protein and maize leaf litter
applied on the soil surface
The decomposition of the leaf litter, as measured
by decline in ash-free dry matter content, was influ-
enced by incubation time but not by maize variety (Ta-
ble II). The decomposition of litter was similar across
all maize hybrids irrespective of whether they were ge-
netically modified or not (Table II). Relative to initial
levels, a significant decrease in ash-free dry matter con-
tent of 7.5% was observed after 4 weeks, following an
almost linear trend, with 91.4%, 84.5% and 75.4% of
initial ash-free dry matter remaining after 8, 12 and 16
weeks, respectively (Table III). The ash-free dry matter
levels were similar across the maize hybrids irrespective
of sampling time (Table II).
The Cry1Ab protein in the DKC80-12B litter sig-
nificantly degraded over time, firstly at a slow rate,
with 2.2% of the initial content lost during the first 4
weeks, followed by a faster decline to 74.8% (25.2% de-
cline) and 56% (43.9% decline) of the initial contents
after 12 and 16 weeks, respectively (Table III).
Decomposition of Cry1Ab protein and maize leaf litter
incorporated in soil
Decomposition of the maize leaf litter incorporated
into soil was affected by incubation time but not by
maize variety or site (Table IV). The leaf litter ash-
free dry matter contents decreased by 5.1% within 4
weeks and 69.8% of the initial ash-free dry matter re-
mained after 12 weeks (Table V). Ash-free dry matter
levels of leaf litter were all similar across the maize
hybrids irrespective of sampling time (Table IV).
The Cry1Ab protein content of the DKC75-15B
leaf litter (267.0 ng g−1) was greater than that of the
PAN6Q-308B leaf litter (253.0 ng g−1) during incuba-
tion. Degradation of the Cry1Ab protein was only sig-
nificantly influenced by incubation time. The Cry1Ab
protein content decreased by 12.6% and 60.6%, after 4
and 12 weeks of incubation, respectively (Table V).
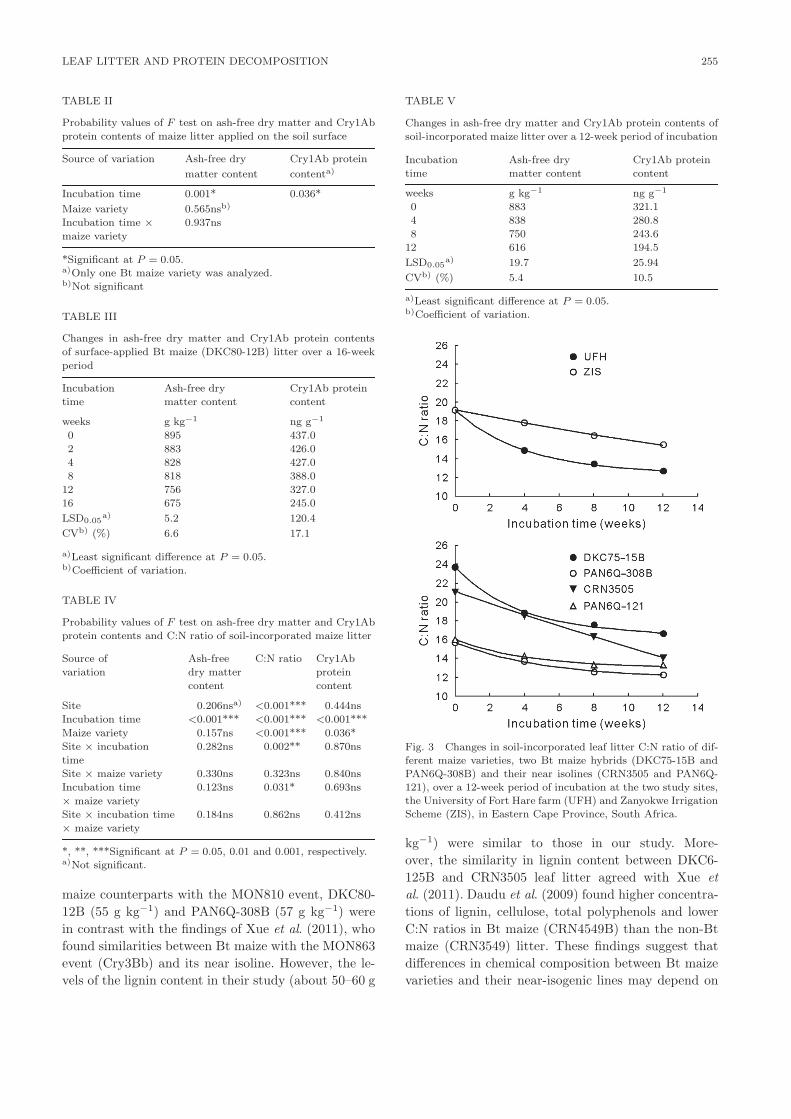
There were significant effects of site × incubation
time and maize variety × incubation time on changes
in C:N ratios of the leaf litter (Table IV). Main effects
of site, maize variety and incubation time were also
significant. At the UFH Research Farm, the C:N ra-
tios decreased at a faster rate, from 19.1% to 12.7%,
than at ZIS where a decline from 19.1% to 15.5% was
observed (Fig. 3). The leaf litter C:N ratios of PAN
6Q-308B and its near isoline PAN6Q-121 were simi-
lar, but lower than those of DKC75-15B and CRN3505
throughout the incubation period (Fig. 3). The leaf lit-
ter C:N ratio of DKC75-15B declined (from 23.7) at a
faster rate than CRN3505 (from 21.1) until the ratios
of the two varieties were similar after 4 and 8 weeks
of incubation. A slower decline in the leaf litter C:N
ratio of DKC75-15B, from 17.6 to 16.7, was observed
between 8 and 12 weeks of incubation, whereas that of
CRN3505 declined from 16.4 to 14.1 during the same
period. Overall, the decline in leaf litter C:N ratio was
similar between DKC75-15B (from 23.7 to 16.7) and
CRN3505 (from 21.1 to 14.1) over the 12-week incuba-
tion period.
DISCUSSION
The higher leaf litter lignin contents of DKC80-10
(63 g kg−1) and PAN6Q-121 (68 g kg−1) than their Bt
TABLE I
Initial ash-free dry matter, lignin and Cry1Ab protein contents and C:N ratios of maize leaf litter used in the study
Leaf litter Maize cultivar Ash-free dry matter content C:N ratio Lignin content Cry1Ab protein content
g kg−1 g kg−1 ng g−1
Surface-applied DKC80-12B (Bt) 898 20.6 55 437.0DKC80-10 886 19.5 63 ND
DKC6-125 900 21.0 NDa) ND
Soil-incorporated DKC75-15B (Bt) 889 23.7 58 328.2
PAN6Q-308B (Bt) 886 15.7 57 314.1
CRN3505 888 21.1 55 ND
PAN6Q-121 868 16.0 68 ND
a)Not determined.
LEAF LITTER AND PROTEIN DECOMPOSITION 255
TABLE II
Probability values of F test on ash-free dry matter and Cry1Ab
protein contents of maize litter applied on the soil surface
Source of variation Ash-free dry Cry1Ab protein
matter content contenta)
Incubation time 0.001* 0.036*
Maize variety 0.565nsb)
Incubation time × 0.937ns
maize variety
*Significant at P = 0.05.a)Only one Bt maize variety was analyzed.b)Not significant
TABLE III
Changes in ash-free dry matter and Cry1Ab protein contents
of surface-applied Bt maize (DKC80-12B) litter over a 16-week
period
Incubation Ash-free dry Cry1Ab protein
time matter content content
weeks g kg−1 ng g−1
0 895 437.0
2 883 426.0
4 828 427.0
8 818 388.0
12 756 327.0
16 675 245.0
LSD0.05a) 5.2 120.4
CVb) (%) 6.6 17.1
a)Least significant difference at P = 0.05.b)Coefficient of variation.
TABLE IV
Probability values of F test on ash-free dry matter and Cry1Ab
protein contents and C:N ratio of soil-incorporated maize litter
Source of Ash-free C:N ratio Cry1Ab
variation dry matter protein
content content
Site 0.206nsa) <0.001*** 0.444ns
Incubation time <0.001*** <0.001*** <0.001***
Maize variety 0.157ns <0.001*** 0.036*
Site × incubation 0.282ns 0.002** 0.870ns
time
Site × maize variety 0.330ns 0.323ns 0.840ns
Incubation time 0.123ns 0.031* 0.693ns
× maize variety
Site × incubation time 0.184ns 0.862ns 0.412ns
× maize variety
*, **, ***Significant at P = 0.05, 0.01 and 0.001, respectively.a)Not significant.
maize counterparts with the MON810 event, DKC80-
12B (55 g kg−1) and PAN6Q-308B (57 g kg−1) were
in contrast with the findings of Xue et al. (2011), who
found similarities between Bt maize with the MON863
event (Cry3Bb) and its near isoline. However, the le-
vels of the lignin content in their study (about 50–60 g
TABLE V
Changes in ash-free dry matter and Cry1Ab protein contents of
soil-incorporated maize litter over a 12-week period of incubation
Incubation Ash-free dry Cry1Ab protein
time matter content content
weeks g kg−1 ng g−1
0 883 321.1
4 838 280.8
8 750 243.6
12 616 194.5
LSD0.05a) 19.7 25.94
CVb) (%) 5.4 10.5
a)Least significant difference at P = 0.05.b)Coefficient of variation.
Fig. 3 Changes in soil-incorporated leaf litter C:N ratio of dif-
ferent maize varieties, two Bt maize hybrids (DKC75-15B and
PAN6Q-308B) and their near isolines (CRN3505 and PAN6Q-
121), over a 12-week period of incubation at the two study sites,
the University of Fort Hare farm (UFH) and Zanyokwe Irrigation
Scheme (ZIS), in Eastern Cape Province, South Africa.
kg−1) were similar to those in our study. More-
over, the similarity in lignin content between DKC6-
125B and CRN3505 leaf litter agreed with Xue et
al. (2011). Daudu et al. (2009) found higher concentra-
tions of lignin, cellulose, total polyphenols and lower
C:N ratios in Bt maize (CRN4549B) than the non-Bt
maize (CRN3549) litter. These findings suggest that
differences in chemical composition between Bt maize
varieties and their near-isogenic lines may depend on

256 A. KAMOTA et al.
the background genetic makeup of the maize. In all
these studies, the lignin contents were far lower than
the threshold of 150 g kg−1 to limit decomposition of
the surface-applied residues (Schroth, 2003).
Genetic modification of maize (MON810) did not
affect the decomposition of Bt maize litter under winter
fallow field conditions regardless of whether the litter
were surface-applied or soil-incorporated. Daudu et al.
(2009) found similar results with soil-incorporated lit-
ter in a study conducted during summer. They found
no effects of genetic modification (MON810) on de-
composition of stem and leaf litter of Bt maize incor-
porated in the same soil during the warmer and wetter
summer maize growing period, in the Central Eastern
Cape. The findings were also in agreement with those
of Tarkalson et al. (2008), in a study done over 23
months in Nebraska, and Lehman et al. (2008), in a
study carried out over 658 d in South Dakota (both
in USA), who did not find any differences in the de-
composition of Bt (events MON810 and MON863) and
non-Bt maize litter after burying them in the soil under
field conditions. In field studies conducted at the Cor-
nell University’s Musgrave Farm in Aurora, USA, Xue
et al. (2011) also found similar results with surface-
applied and soil-incorporated leaf, stem and root litter
of Bt maize with the MON863 event and its near iso-
line. Zwahlen et al. (2007) found similar results with
5 000-μm mesh size litterbags but reported faster Bt
maize decomposition when 125- and 20-μm mesh size
litterbags were used. While DKC75-15B had a higher
initial leaf litter C:N ratio than CRN3505 in our study,
the decline in C:N ratio (as a measure for decomposi-
tion of the leaf litter) was the same over a 12-week pe-
riod and the same was observed for PAN6Q-308B and
PAN6Q-121 leaf litter, which had similar C:N ratios.
These findings were in agreement with those of Daudu
et al. (2009), who reported no differences in litter de-
composition even though there were differences in C:N
ratio. A review article by Yanni et al. (2010) showed
that some researchers found differences in chemical
composition, particularly lignin content and C:N ra-
tio, between Bt maize varieties (with various trans-
formation events) and their near isolines, while oth-
ers reported no differences. However, where the diffe-
rences occurred, they did not translate to differences in
decomposition rates. Differences in C:N ratio did not
limit decomposition probably because the ratios were
below the threshold of 30 above which mineralization
of N could be limited (Schroth, 2003). These findings
suggest that genetic modification of maize with the
MON810 event may not affect decomposition and nu-
trient release from the maize litter. The faster decline
in C:N ratio of the maize litter at the UFH Research
Farm could be because of the higher clay content which
may have resulted in greater moisture storage. Howe-
ver, the effect was not significant for ash-free dry mat-
ter content.
About 84.4% of initial ash-free dry matter rema-
ined 12 weeks after surface application, whereas 69.8%
remained after the same period of soil incorporation ir-
respective of site, and this was in agreement with Xue
et al. (2011), who reported that maize litter incorpo-
rated into the soil decomposed at a faster rate. The
faster decomposition rate of incorporated litter could
be a result of greater proximity of the litter to soil
particles and moisture and hence greater exposure to
soil microbial activity. The initial lower decomposi-
tion of surface-applied litter was probably a result of
low temperature and soil moisture, as a result of low
rainfall in July, whereas a faster decomposition was
observed when temperatures and rainfall increased as
summer approached. Microbial activity is more pro-
nounced when temperatures are between 24 and 27◦C (Zwahlen et al., 2003). Daudu et al. (2009) ob-
served that 50% of the dry matter was decomposed
within a two-week period when the maize litter was
incorporated during moist summer months. The dif-
ferences in the findings could be explained by moisture
and temperature conditions. During the experimental
period of Daudu et al. (2009), 236.2 mm rainfall was
received and the temperature ranged from 16.4 to 23.2◦C, whereas in our study rainfall ranged from 98 to 128
mm and temperatures ranged from 15 to 18 ◦C. The
residues could have decomposed at a faster rate if the
study was extended into the summer.
The decrease in the Cry1Ab protein content fol-
lowed the same trend as that in the ash-free dry
matter content, with greater decomposition in soil-
incorporated litter than in litter applied on the soil sur-
face. Like in the decline in ash-free dry matter content,
these differences could be a result of better soil contact
which enhanced decomposition. The longer cool pe-
riod the surface-applied litter experienced (initiated in
July) than the incorporated litter (initiated in August)
may also have caused the difference. These results con-
tradicted those of Zwahlen et al. (2003), who observed
faster degradation of Cry1Ab protein in Bt maize lit-
ter left on the soil surface than that incorporated into
the soil. The difference in the findings could be be-
cause Zwahlen et al. (2003) used maize leaf litter at
the pollen-shed stage, earthworms were included and
the litter was applied on the soil surface, whereas we
used senescent maize leaf litter obtained after harvest
and no effort was made to include or exclude earth-

LEAF LITTER AND PROTEIN DECOMPOSITION 257
worms in any of the experiments. Moreover, activity
of earthworms in our study was minimal considering
the lower moisture and temperature during the study
periods.
The contents of Cry1Ab protein remained > 190
ng kg−1 (> 60% of initial values) after 12 weeks in this
study, whereas Daudu et al. (2009) observed that the
Cry1Ab protein approached undetectable levels within
14 d when Bt maize leaf litter was incorporated into the
soil. This difference could be explained by the higher
rainfall and temperature during the summer months of
February, March and April when the study by Daudu
et al. (2009) was conducted compared to the drier
cooler period for this study. If our study was allowed to
progress into summer, the Cry1Ab protein could have
further degraded to undetectable levels, thus reducing
exposure of the protein to none-target soil organisms.
CONCLUSIONS
The chemical composition of leaf litter of Bt maize
varieties producing Cry1Ab protein (MON810) tested
in this study might be slightly different from their near-
isogenic lines. Such differences did not affect their leaf
litter decomposition when the litter was applied on the
surface or incorporated into the soil during the fal-
low winter months in Eastern Cape Province, South
Africa. Bt maize litter did not limit decomposition and
nutrient cycling and, therefore, may not have an added
advantage of providing longer lasting soil cover in con-
servation agriculture systems or improving C seques-
tration in the soil.
REFERENCES
Association of Official Analytical Chemists (AOAC). 1984. Offi-
cial Methods of Analysis. 14th Ed. Association of Official
Analytical Chemists, Inc., Arlington.
Daudu, C. K., Muchaonyerwa, P. and Mnkeni, P. N. S. 2009.
Litterbag decomposition of genetically modified maize re-
sidues and their constituent Bacillus thuringiensis protein
(Cry1Ab) under field conditions in the central region of the
Eastern Cape, South Africa. Agr. Ecosyst. Environ. 134:
153–158.
Donnelly, P. K., Entry, J. A., Crawford, D. L. and Cromack, K.
Jr. 1990. Cellulose and lignin degradation in forest soils: Re-
sponse to moisture, temperature and acidity. Microb. Ecol.
20: 289–295.
Fey, M. 2010. Soils of South Africa. Cambridge University Press,
Cambridge.
Flores, S., Saxena, D. and Stotzky, G. 2005. Transgenic Bt
plants decompose less in soil than non-Bt plants. Soil Biol.
Biochem. 37: 1073–1082.
James, C. 2007. Global Status of Commercialized Biotech/GM
Crops: 2007. ISAAA Brief No. 37. ISAAA, Ithaca.
Lawes Agricultural Trust. 2008. GenStat Release 7.22 DE Re-
ference Manual. VSN International, Helmel Hempstead.
LECO Corporation. 2003. TRUSPEC C/N Determinator. In-
struction Manual. LECO Corporation, St. Joseph.
Lehman, M. R., Osborne, S. L. and Rosentrater, K. A. 2008. No
differences in decomposition rates observed between Bacillus
thuringiensis and corn residue incubated in the field. Agron.
J. 100: 163–168.
Mandiringana, O. T., Mnkeni, P. N. S., Mkile, Z., van Aver-
beke, W., Van Ranst, E. and Verplancke, H. 2005. Mine-
ralogy and Fertility Status of Selected Soils of the Eastern
Cape Province, South Africa. Commun. Soil Sci. Plan. 36:
2431–2446.
Mungai, N. W., Motavalli, P. P., Nelson, K. A. and Kremer,
R. J. 2005. Differences in yields, residue composition and
N mineralization dynamics of Bt and non-Bt maize. Nutr.
Cycl. Agroecosys. 73: 101–109.
Palm, C. A., Nziguheba, G., Gachengo, C. N., Gacheru, E. and
Rao, M. R. 1999. Organic materials as sources of phospho-
rus. Agrofor. Forum. 9: 30–33.
Okalebo, J. R., Gathua, K. W. and Woomer, P. L. 2002. Labora-
tory Methods of Soil and Pant Analysis: A Working Manual.
2nd Ed. TSBF-CIAT and SACRED Africa, Nairobi.
Saxena, D. and Stotzky, G. 2001. Bt corn has a higher lignin
content than non-Bt corn. Am. J. Bot. 88: 1704–1706.
Schroth, G. 2003. Decomposition and nutrient supply from bio-
mass. In Schroth, G. and Sinclair, P. F. (eds.) Trees, Crops
and Soil Fertility: Concepts and Research Methods. CABI
International, Wallingford. pp. 131–150.
Soil Classification Working Group. 1991. Soil Classification: A
Taxonomic System for South Africa. Memoirs on the Agri-
cultural Natural Resources of South Africa No. 15. Soil Cla-
ssification Working Group, Pretoria.
Tarkalson, D. D., Kachman, S. D., Knops, J. M. N., Thies, J. E.
and Wortmann, C. S. 2008. Decomposition of Bt and Non-
Bt corn hybrid residues in the field. Nutr. Cycl. Agroecosys.
80: 211–222.
Van Averbeke, W. and Marais, J. N. 1991. Evaluation of Ciskeian
Ecotopes for Rainfed Cropping: Final Report. University of
Fort Hare, Alice.
Xue, K., Serohijos, R. C., Devare, M. and Thies, J. E. 2011. De-
composition rates and residue-colonizing microbial commu-
nities of Bacillus thuringiensis insecticidal protein Cry3Bb-
expressing (Bt) and non-Bt corn hybrids in the field. Appl.
Environ. Microb. 77: 839–846.
Yanni, S. F., Whalen, J. K. and Ma, B. L. 2010. Crop residue
chemistry, decomposition rates, and CO2 evolution in Bt
and non-Bt corn agroecosystems in North America: a review.
Nutr. Cycl. Agroecosys. 87: 277–293.
Zwahlen, C., Hilbeck, A., Gugerli, P. and Nentwig, W. 2003.
Degradation of the Cry1Ab protein within transgenic Bacil-
lus thuringiensis corn tissue in the field. Mol. Ecol. 12: 765–
775.
Zwahlen, C., Hilbeck, A. and Nentwig, W. 2007. Field decom-
position of transgenic Bt maize residue and the impact on
non-target soil invertebrates. Plant Soil. 300: 245–257.





























