Embed Size (px)
Citation preview

Deep-Sea Research II 58 (2011) 2477–2488
Contents lists available at ScienceDirect
Deep-Sea Research II
0967-06
doi:10.1
n Corr
E-m
journal homepage: www.elsevier.com/locate/dsr2
Organic geochemistry of submarine canyons: The Portuguese Margin
Kostas Kiriakoulakis a,b, Sabena Blackbird a, Jeroen Ingels c, Ann Vanreusel c, George A. Wolff a,n
a School of Environmental Sciences, University of Liverpool, 4 Brownlow Street, Liverpool L69 3GP, UKb School of Natural Sciences and Psychology, Liverpool John Moores University, James Parsons, Byrom Street, Liverpool, Merseyside, L3 3AF, UKc Marine Biology Department, Ghent University, Krijgslaan 281 S8, 9000 Ghent, Belgium
a r t i c l e i n f o
Available online 16 April 2011
Keywords:
Portuguese Continental Margin
Organic geochemistry
Submarine canyons
Organic matter preservation
Terrestrial organic matter
Lipid biomarkers
45/$ - see front matter & 2011 Elsevier Ltd. A
016/j.dsr2.2011.04.010
esponding author. Tel.: þ44 151 794 4094; fa
ail address: [email protected] (G.A. Wolff).
a b s t r a c t
The organic geochemistry of the Portuguese Margin of the North-Eastern Atlantic Ocean reveals a
highly heterogeneous environment that is strongly influenced by canyons that incise the continental
margin. Suspended particulate organic matter (sPOM) is funnelled through the canyons to the deep sea,
particularly in the Nazare Canyon where there are high concentrations of sPOM even at 42000 m
water depth. The nature of the sPOM through the water column varies, with that transported through
the canyons having higher contributions of terrestrial organic matter (higher C/N and larger contribu-
tion of land plant-derived lipids) than sPOM in overlying waters and close to the seafloor on the
adjacent slope. Zooplankton-derived lipids dominate sPOM associated with the upper and lower
boundaries of the Mediterranean Overflow Water (MOW�600 and 1500 m, respectively). Canyon
sediments are enriched in organic carbon when compared to slope sediments, but sedimentary organic
matter also appears to derive from multiple sources and undergoes significant alteration prior to
deposition. On the open slope, low sedimentation rates and long oxygen exposure times lead to
intensive oxidation of organic matter.
& 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Submarine canyons are kilometre-scale, deep, narrow valleyscut into the continental margin. They are important conduits forthe transport of sediment from shelf depths to the deep sea(Gardner, 1989a, b). Field and modelling studies have shown thatwhere canyons cut the slope, fluid-dynamic complexity increases,creating locally enhanced current velocities that impact particu-late-matter and sediment transport (Boyer et al., 2004; Perenneet al., 2001a, b). The presence or absence of canyons should,therefore, strongly influence the magnitude of organic carbontransport across the shelf break (de Haas et al., 2002). Observa-tions of large episodic particulate flux events (Canals et al., 2006;de Stigter et al., 2007) confirm that canyons can act as quantita-tively important pathways for carbon transport to the deep ocean.In the Gulf of Lyons, a single dense shelf water cascading event(DSWC) was estimated to export 0.6 Tg of fresh marine organicmatter (C/N�6.5) to the deep sea over �40 days. Such largeevents lead to significant erosion and redistribution of sediments,and associated organic matter (OM) fluxes that are likely tosignificantly impact canyon and deep-sea ecosystems.
However, questions remain as to the nature, provenance andfate of OM transported through canyons. For example, at theCalifornian continental margin, macroalgae from near-shore
ll rights reserved.
x: þ44 151 794 5196.
growth are transported to canyon heads by long-shore transport.Strong tidal and gravity currents transport such material enrich-ing the deeper canyon system. In the Scripps and La Jolla canyons,detrital mats are found down to 550 m and the more resistantalgal remains down to �900 m attracting an abundant and variedmegafauna (Vetter and Dayton, 1999). The enrichment of OMin canyon sediments leads to enhanced sediment communityoxygen consumption rates (Accornero et al., 2002; Duineveldet al., 2001), relative to the adjacent open slope; these enhancedrates are correlated with meiofaunal density (Duineveld et al.,2001; Ingels et al., 2009). At the same time, enhanced burial oforganic carbon in canyon sediments suggests that a part of thisOM is not biologically labile; higher C/N ratios in canyon sedi-ments relative to the open slope in the Lacaze-Duthier Canyon, inthe Gulf of Lions, support the hypothesis that there is an elevatedcontribution of terrestrial material to these sediments (Schmiedlet al., 2004). This is reflected in the depleted d13C of benthicforaminiferal carbonate in the canyon, relative to the open slope.Conversely, sediments in the Whittard Canyon, which are notdirectly associated with a river system, are enriched in labilephytodetrital organic matter, particularly at the canyon head(Duineveld et al., 2001). Thus there is variability in the processescausing OM enrichment in seafloor sediments in canyons, but atleast part of shelf/slope productivity is focussed there.
One approach to assessing the importance of canyons in thetransfer of OM from shelf seas to the deep ocean is to study theirorganic geochemistry. While there have been a number of suchstudies of shelf and slope sediments to assess, for example, the
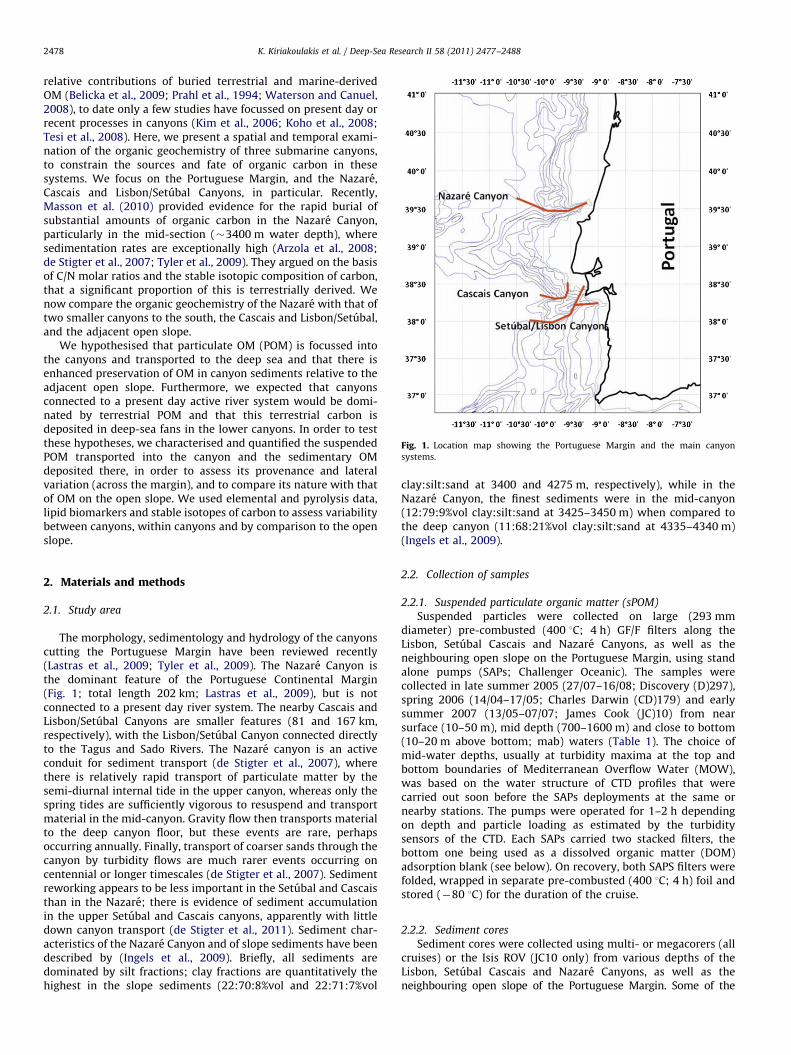
Fig. 1. Location map showing the Portuguese Margin and the main canyon
systems.
K. Kiriakoulakis et al. / Deep-Sea Research II 58 (2011) 2477–24882478
relative contributions of buried terrestrial and marine-derivedOM (Belicka et al., 2009; Prahl et al., 1994; Waterson and Canuel,2008), to date only a few studies have focussed on present day orrecent processes in canyons (Kim et al., 2006; Koho et al., 2008;Tesi et al., 2008). Here, we present a spatial and temporal exami-nation of the organic geochemistry of three submarine canyons,to constrain the sources and fate of organic carbon in thesesystems. We focus on the Portuguese Margin, and the Nazare,Cascais and Lisbon/Setubal Canyons, in particular. Recently,Masson et al. (2010) provided evidence for the rapid burial ofsubstantial amounts of organic carbon in the Nazare Canyon,particularly in the mid-section (�3400 m water depth), wheresedimentation rates are exceptionally high (Arzola et al., 2008;de Stigter et al., 2007; Tyler et al., 2009). They argued on the basisof C/N molar ratios and the stable isotopic composition of carbon,that a significant proportion of this is terrestrially derived. Wenow compare the organic geochemistry of the Nazare with that oftwo smaller canyons to the south, the Cascais and Lisbon/Setubal,and the adjacent open slope.
We hypothesised that particulate OM (POM) is focussed intothe canyons and transported to the deep sea and that there isenhanced preservation of OM in canyon sediments relative to theadjacent open slope. Furthermore, we expected that canyonsconnected to a present day active river system would be domi-nated by terrestrial POM and that this terrestrial carbon isdeposited in deep-sea fans in the lower canyons. In order to testthese hypotheses, we characterised and quantified the suspendedPOM transported into the canyon and the sedimentary OMdeposited there, in order to assess its provenance and lateralvariation (across the margin), and to compare its nature with thatof OM on the open slope. We used elemental and pyrolysis data,lipid biomarkers and stable isotopes of carbon to assess variabilitybetween canyons, within canyons and by comparison to the openslope.
2. Materials and methods
2.1. Study area
The morphology, sedimentology and hydrology of the canyonscutting the Portuguese Margin have been reviewed recently(Lastras et al., 2009; Tyler et al., 2009). The Nazare Canyon isthe dominant feature of the Portuguese Continental Margin(Fig. 1; total length 202 km; Lastras et al., 2009), but is notconnected to a present day river system. The nearby Cascais andLisbon/Setubal Canyons are smaller features (81 and 167 km,respectively), with the Lisbon/Setubal Canyon connected directlyto the Tagus and Sado Rivers. The Nazare canyon is an activeconduit for sediment transport (de Stigter et al., 2007), wherethere is relatively rapid transport of particulate matter by thesemi-diurnal internal tide in the upper canyon, whereas only thespring tides are sufficiently vigorous to resuspend and transportmaterial in the mid-canyon. Gravity flow then transports materialto the deep canyon floor, but these events are rare, perhapsoccurring annually. Finally, transport of coarser sands through thecanyon by turbidity flows are much rarer events occurring oncentennial or longer timescales (de Stigter et al., 2007). Sedimentreworking appears to be less important in the Setubal and Cascaisthan in the Nazare; there is evidence of sediment accumulationin the upper Setubal and Cascais canyons, apparently with littledown canyon transport (de Stigter et al., 2011). Sediment char-acteristics of the Nazare Canyon and of slope sediments have beendescribed by (Ingels et al., 2009). Briefly, all sediments aredominated by silt fractions; clay fractions are quantitatively thehighest in the slope sediments (22:70:8%vol and 22:71:7%vol
clay:silt:sand at 3400 and 4275 m, respectively), while in theNazare Canyon, the finest sediments were in the mid-canyon(12:79:9%vol clay:silt:sand at 3425–3450 m) when compared tothe deep canyon (11:68:21%vol clay:silt:sand at 4335–4340 m)(Ingels et al., 2009).
2.2. Collection of samples
2.2.1. Suspended particulate organic matter (sPOM)
Suspended particles were collected on large (293 mmdiameter) pre-combusted (400 1C; 4 h) GF/F filters along theLisbon, Setubal Cascais and Nazare Canyons, as well as theneighbouring open slope on the Portuguese Margin, using standalone pumps (SAPs; Challenger Oceanic). The samples werecollected in late summer 2005 (27/07–16/08; Discovery (D)297),spring 2006 (14/04–17/05; Charles Darwin (CD)179) and earlysummer 2007 (13/05–07/07; James Cook (JC)10) from nearsurface (10–50 m), mid depth (700–1600 m) and close to bottom(10–20 m above bottom; mab) waters (Table 1). The choice ofmid-water depths, usually at turbidity maxima at the top andbottom boundaries of Mediterranean Overflow Water (MOW),was based on the water structure of CTD profiles that werecarried out soon before the SAPs deployments at the same ornearby stations. The pumps were operated for 1–2 h dependingon depth and particle loading as estimated by the turbiditysensors of the CTD. Each SAPs carried two stacked filters, thebottom one being used as a dissolved organic matter (DOM)adsorption blank (see below). On recovery, both SAPS filters werefolded, wrapped in separate pre-combusted (400 1C; 4 h) foil andstored (�80 1C) for the duration of the cruise.
2.2.2. Sediment cores
Sediment cores were collected using multi- or megacorers (allcruises) or the Isis ROV (JC10 only) from various depths of theLisbon, Setubal Cascais and Nazare Canyons, as well as theneighbouring open slope of the Portuguese Margin. Some of the
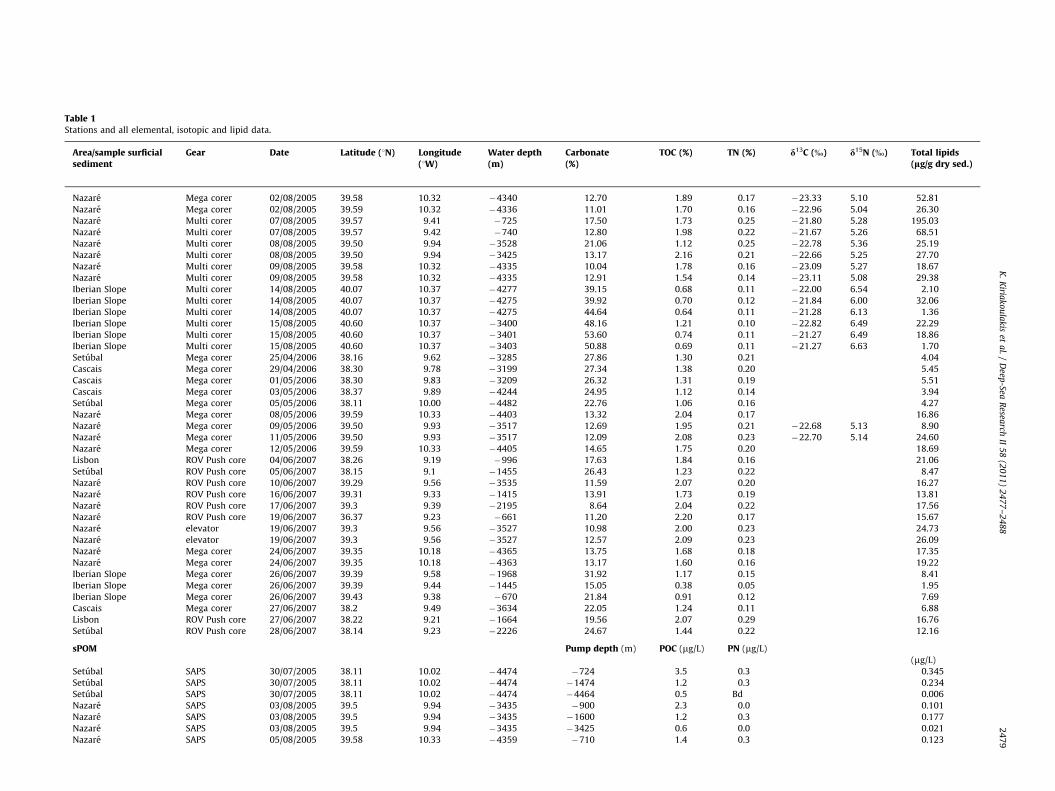
Table 1Stations and all elemental, isotopic and lipid data.
Area/sample surficialsediment
Gear Date Latitude (1N) Longitude(1W)
Water depth(m)
Carbonate(%)
TOC (%) TN (%) d13C (%) d15N (%) Total lipids(lg/g dry sed.)
Nazare Mega corer 02/08/2005 39.58 10.32 �4340 12.70 1.89 0.17 �23.33 5.10 52.81
Nazare Mega corer 02/08/2005 39.59 10.32 �4336 11.01 1.70 0.16 �22.96 5.04 26.30
Nazare Multi corer 07/08/2005 39.57 9.41 �725 17.50 1.73 0.25 �21.80 5.28 195.03
Nazare Multi corer 07/08/2005 39.57 9.42 �740 12.80 1.98 0.22 �21.67 5.26 68.51
Nazare Multi corer 08/08/2005 39.50 9.94 �3528 21.06 1.12 0.25 �22.78 5.36 25.19
Nazare Multi corer 08/08/2005 39.50 9.94 �3425 13.17 2.16 0.21 �22.66 5.25 27.70
Nazare Multi corer 09/08/2005 39.58 10.32 �4335 10.04 1.78 0.16 �23.09 5.27 18.67
Nazare Multi corer 09/08/2005 39.58 10.32 �4335 12.91 1.54 0.14 �23.11 5.08 29.38
Iberian Slope Multi corer 14/08/2005 40.07 10.37 �4277 39.15 0.68 0.11 �22.00 6.54 2.10
Iberian Slope Multi corer 14/08/2005 40.07 10.37 �4275 39.92 0.70 0.12 �21.84 6.00 32.06
Iberian Slope Multi corer 14/08/2005 40.07 10.37 �4275 44.64 0.64 0.11 �21.28 6.13 1.36
Iberian Slope Multi corer 15/08/2005 40.60 10.37 �3400 48.16 1.21 0.10 �22.82 6.49 22.29
Iberian Slope Multi corer 15/08/2005 40.60 10.37 �3401 53.60 0.74 0.11 �21.27 6.49 18.86
Iberian Slope Multi corer 15/08/2005 40.60 10.37 �3403 50.88 0.69 0.11 �21.27 6.63 1.70
Setubal Mega corer 25/04/2006 38.16 9.62 �3285 27.86 1.30 0.21 4.04
Cascais Mega corer 29/04/2006 38.30 9.78 �3199 27.34 1.38 0.20 5.45
Cascais Mega corer 01/05/2006 38.30 9.83 �3209 26.32 1.31 0.19 5.51
Cascais Mega corer 03/05/2006 38.37 9.89 �4244 24.95 1.12 0.14 3.94
Setubal Mega corer 05/05/2006 38.11 10.00 �4482 22.76 1.06 0.16 4.27
Nazare Mega corer 08/05/2006 39.59 10.33 �4403 13.32 2.04 0.17 16.86
Nazare Mega corer 09/05/2006 39.50 9.93 �3517 12.69 1.95 0.21 �22.68 5.13 8.90
Nazare Mega corer 11/05/2006 39.50 9.93 �3517 12.09 2.08 0.23 �22.70 5.14 24.60
Nazare Mega corer 12/05/2006 39.59 10.33 �4405 14.65 1.75 0.20 18.69
Lisbon ROV Push core 04/06/2007 38.26 9.19 �996 17.63 1.84 0.16 21.06
Setubal ROV Push core 05/06/2007 38.15 9.1 �1455 26.43 1.23 0.22 8.47
Nazare ROV Push core 10/06/2007 39.29 9.56 �3535 11.59 2.07 0.20 16.27
Nazare ROV Push core 16/06/2007 39.31 9.33 �1415 13.91 1.73 0.19 13.81
Nazare ROV Push core 17/06/2007 39.3 9.39 �2195 8.64 2.04 0.22 17.56
Nazare ROV Push core 19/06/2007 36.37 9.23 �661 11.20 2.20 0.17 15.67
Nazare elevator 19/06/2007 39.3 9.56 �3527 10.98 2.00 0.23 24.73
Nazare elevator 19/06/2007 39.3 9.56 �3527 12.57 2.09 0.23 26.09
Nazare Mega corer 24/06/2007 39.35 10.18 �4365 13.75 1.68 0.18 17.35
Nazare Mega corer 24/06/2007 39.35 10.18 �4363 13.17 1.60 0.16 19.22
Iberian Slope Mega corer 26/06/2007 39.39 9.58 �1968 31.92 1.17 0.15 8.41
Iberian Slope Mega corer 26/06/2007 39.39 9.44 �1445 15.05 0.38 0.05 1.95
Iberian Slope Mega corer 26/06/2007 39.43 9.38 �670 21.84 0.91 0.12 7.69
Cascais Mega corer 27/06/2007 38.2 9.49 �3634 22.05 1.24 0.11 6.88
Lisbon ROV Push core 27/06/2007 38.22 9.21 �1664 19.56 2.07 0.29 16.76
Setubal ROV Push core 28/06/2007 38.14 9.23 �2226 24.67 1.44 0.22 12.16
sPOM Pump depth (m) POC (mg/L) PN (mg/L)
(mg/L)
Setubal SAPS 30/07/2005 38.11 10.02 �4474 �724 3.5 0.3 0.345
Setubal SAPS 30/07/2005 38.11 10.02 �4474 �1474 1.2 0.3 0.234
Setubal SAPS 30/07/2005 38.11 10.02 �4474 �4464 0.5 Bd 0.006
Nazare SAPS 03/08/2005 39.5 9.94 �3435 �900 2.3 0.0 0.101
Nazare SAPS 03/08/2005 39.5 9.94 �3435 �1600 1.2 0.3 0.177
Nazare SAPS 03/08/2005 39.5 9.94 �3435 �3425 0.6 0.0 0.021
Nazare SAPS 05/08/2005 39.58 10.33 �4359 �710 1.4 0.3 0.123
K.
Kiria
ko
ula
kis
eta
l./
Deep
-SeaR
esearch
II5
8(2
01
1)
24
77
–2
48
82
47
9
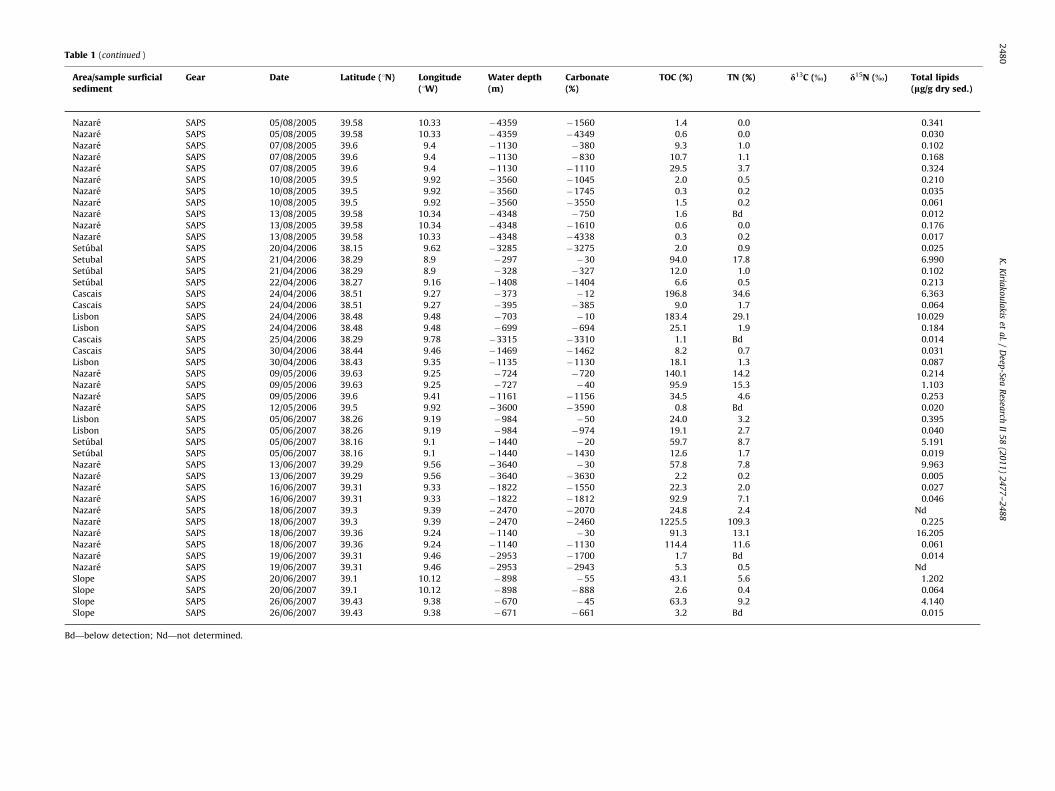
Table 1 (continued )
Area/sample surficialsediment
Gear Date Latitude (1N) Longitude(1W)
Water depth(m)
Carbonate(%)
TOC (%) TN (%) d13C (%) d15N (%) Total lipids(lg/g dry sed.)
Nazare SAPS 05/08/2005 39.58 10.33 �4359 �1560 1.4 0.0 0.341
Nazare SAPS 05/08/2005 39.58 10.33 �4359 �4349 0.6 0.0 0.030
Nazare SAPS 07/08/2005 39.6 9.4 �1130 �380 9.3 1.0 0.102
Nazare SAPS 07/08/2005 39.6 9.4 �1130 �830 10.7 1.1 0.168
Nazare SAPS 07/08/2005 39.6 9.4 �1130 �1110 29.5 3.7 0.324
Nazare SAPS 10/08/2005 39.5 9.92 �3560 �1045 2.0 0.5 0.210
Nazare SAPS 10/08/2005 39.5 9.92 �3560 �1745 0.3 0.2 0.035
Nazare SAPS 10/08/2005 39.5 9.92 �3560 �3550 1.5 0.2 0.061
Nazare SAPS 13/08/2005 39.58 10.34 �4348 �750 1.6 Bd 0.012
Nazare SAPS 13/08/2005 39.58 10.34 �4348 �1610 0.6 0.0 0.176
Nazare SAPS 13/08/2005 39.58 10.33 �4348 �4338 0.3 0.2 0.017
Setubal SAPS 20/04/2006 38.15 9.62 �3285 �3275 2.0 0.9 0.025
Setubal SAPS 21/04/2006 38.29 8.9 �297 �30 94.0 17.8 6.990
Setubal SAPS 21/04/2006 38.29 8.9 �328 �327 12.0 1.0 0.102
Setubal SAPS 22/04/2006 38.27 9.16 �1408 �1404 6.6 0.5 0.213
Cascais SAPS 24/04/2006 38.51 9.27 �373 �12 196.8 34.6 6.363
Cascais SAPS 24/04/2006 38.51 9.27 �395 �385 9.0 1.7 0.064
Lisbon SAPS 24/04/2006 38.48 9.48 �703 �10 183.4 29.1 10.029
Lisbon SAPS 24/04/2006 38.48 9.48 �699 �694 25.1 1.9 0.184
Cascais SAPS 25/04/2006 38.29 9.78 �3315 �3310 1.1 Bd 0.014
Cascais SAPS 30/04/2006 38.44 9.46 �1469 �1462 8.2 0.7 0.031
Lisbon SAPS 30/04/2006 38.43 9.35 �1135 �1130 18.1 1.3 0.087
Nazare SAPS 09/05/2006 39.63 9.25 �724 �720 140.1 14.2 0.214
Nazare SAPS 09/05/2006 39.63 9.25 �727 �40 95.9 15.3 1.103
Nazare SAPS 09/05/2006 39.6 9.41 �1161 �1156 34.5 4.6 0.253
Nazare SAPS 12/05/2006 39.5 9.92 �3600 �3590 0.8 Bd 0.020
Lisbon SAPS 05/06/2007 38.26 9.19 �984 �50 24.0 3.2 0.395
Lisbon SAPS 05/06/2007 38.26 9.19 �984 �974 19.1 2.7 0.040
Setubal SAPS 05/06/2007 38.16 9.1 �1440 �20 59.7 8.7 5.191
Setubal SAPS 05/06/2007 38.16 9.1 �1440 �1430 12.6 1.7 0.019
Nazare SAPS 13/06/2007 39.29 9.56 �3640 �30 57.8 7.8 9.963
Nazare SAPS 13/06/2007 39.29 9.56 �3640 �3630 2.2 0.2 0.005
Nazare SAPS 16/06/2007 39.31 9.33 �1822 �1550 22.3 2.0 0.027
Nazare SAPS 16/06/2007 39.31 9.33 �1822 �1812 92.9 7.1 0.046
Nazare SAPS 18/06/2007 39.3 9.39 �2470 �2070 24.8 2.4 Nd
Nazare SAPS 18/06/2007 39.3 9.39 �2470 �2460 1225.5 109.3 0.225
Nazare SAPS 18/06/2007 39.36 9.24 �1140 �30 91.3 13.1 16.205
Nazare SAPS 18/06/2007 39.36 9.24 �1140 �1130 114.4 11.6 0.061
Nazare SAPS 19/06/2007 39.31 9.46 �2953 �1700 1.7 Bd 0.014
Nazare SAPS 19/06/2007 39.31 9.46 �2953 �2943 5.3 0.5 Nd
Slope SAPS 20/06/2007 39.1 10.12 �898 �55 43.1 5.6 1.202
Slope SAPS 20/06/2007 39.1 10.12 �898 �888 2.6 0.4 0.064
Slope SAPS 26/06/2007 39.43 9.38 �670 �45 63.3 9.2 4.140
Slope SAPS 26/06/2007 39.43 9.38 �671 �661 3.2 Bd 0.015
Bd—below detection; Nd—not determined.
K.
Kiria
ko
ula
kis
eta
l./
Deep
-SeaR
esearch
II5
8(2
01
1)
24
77
–2
48
82
48
0

K. Kiriakoulakis et al. / Deep-Sea Research II 58 (2011) 2477–2488 2481
cores were frozen on recovery, extruded when still frozen, wrappedin pre-combusted (400 1C; 4 h) foil and stored (�80 1C) for theduration of the cruise. Others were sliced down to 10 cm (usuallyevery cm down to 6 cm and every 2 cm down to 10 cm) soon afterrecovery and then frozen (�80 1C) for the rest of the cruise.
2.3. Analytical methods
2.3.1. Organic C and N
After freeze-drying, punched circles (�133–346 mm2) weretaken from the top SAPS filters to measure suspended particulateorganic carbon and nitrogen (sPOC, sPN). Care was taken to avoidareas of the filter that appeared to be heterogeneous (i.e. largedarker/lighter or coloured spots were avoided). Due to the design ofthe SAPs, water cannot pass through the whole area of the filter andthis can lead to small but variable underestimation of values if filterweight is used in the calculation. Therefore effective filter areas(i.e. corrected for non-pumped surface of the filter) were used tonormalise all concentrations (relative to volume of water pumped,mg L�1). sPOC values were obtained after de-carbonation of thefilters (Yamamuro and Kayanne, 1995), whereas sPN values weredetermined without de-carbonation. Adsorption of DOM onto GF/Ffilters can lead to the overestimation of sPOC and sPN concentra-tions, particularly when these are low (Liu et al., 2005; Turnewitschet al., 2007). Therefore the bottom filters were used to correct forthis and used as ‘DOM blanks’. The measurements were carried outas for sPOC and sPN (non-decarbonated values), and it was assumedthat the measured organic carbon (OC) and N in the ‘DOM-blank’bottom filters were adsorbed DOC (ad-DOC) and DN (ad-DN). OCand N values of the top filters that were close to the analyticaldetection limit were often lower than adsorbed values. In thesecases sPOC and sPN values were considered nil. Quoted sPOC andsPN values are corrected for ad-DOM unless otherwise stated(Table 1). Aliquots of freeze-dried sediments (5–10 mg) were alsodecarbonated (Yamamuro&Kayanne, 1995) in order to determinetotal organic carbon (TOC) content (%) of the dry sediment. Non-decarbonated aliquots were used to determine total N and C (TN andTC, respectively). The difference between TC and TOC was used tocalculate carbonate contents of the sediments [(TC�TOC)�8.33],assuming that all carbonate was CaCO3. All carbon and nitrogenelemental analyses were carried out using a CEInstruments NC 2500CHN analyser in duplicate (mean value used; all values were within10% of the mean). TOC and TN values (% of dry sediment) of wholesediment cores (10 cm) were calculated according to the formula:[S(dry slice weight� TOC slice%)]/S(dry slice weight).
2.3.2. Organic C and N stable isotopes
Organic carbon and nitrogen isotopic analyses on selected samplealiquots were carried out, with and without de-carbonation respec-tively, using a Thermo Finnigan Delta-Plus Advantage isotope ratiomass spectrometer (IRMS), coupled to a Costech Instruments ECS4010 elemental combustion system (‘‘EA’’) via a Thermo FinniganConflo III open-split interface. The EA was equipped with a CostechZero Blank autosampler, utilising a 32-position, large-capacity sam-ple wheel. Sample d13C and d15N ratios were measured relative tointernationally accepted standards (Pee Dee Belemnite and atmo-spheric nitrogen, respectively), calibrated against a variety of primarystandards, and normalised for weight-to-d bias and calibration offsetusing a secondary isotope ratio standard (acetanilide).
2.3.3. Rock-Eval pyrolysis
To determine hydrocarbons (HC) and CO2 generated from thesediment samples, a Rock-Eval 6 Pyrolysis device fitted withflame ionisation and thermal conductivity detectors was used. HCwere quantified as S1 and S2, and CO2 as S3 peaks (mg HC/g or
mg/g CO2 per sample). The hydrogen index (HI) and oxygen index(OI) were calculated using the equations HI¼S2�100/TOC andOI¼S3�100/TOC, with TOC contents being determined accordingto Section 2.2.1. The temperatures of maximum HC generation,Tmax (in 1C), were also recorded.
2.3.4. Lipids
Lipid analyses were carried out on portions of the pump filters(0.8–1 g; circular sectors) and sediment aliquots (�1 g) after themethod modified from Kiriakoulakis et al. (2004). Briefly, filterand sediment samples were extracted by sonication (15 min; �3)in a solvent mixture (dichloromethane:methanol, 9:1) and trans-methylated in the dark with 5% methanolic acetyl chloride (40 1C;12 h; (Christie, 1982)). After complete removal of the metha-nolic acetyl chloride under a stream of nitrogen the sampleswere derivatised with bis-trimethylsilyltrifluoroacetamide (1%trimethylsilylchloride; 30–50 mL; 40 1C; 0.5–1 h) prior to GC-MSanalyses. Aliquots of the transmethylated and derivatised extractswere injected into a Trace 2000 Series gas chromatograph fittedwith an on-column injector and a fused high-temperature silicacolumn (60 m�0.25 mm i.d.; 5%; DB5-HT equivalent; J&W) withhelium as the carrier gas (ca. 1.6 mL min�1). A retention gap ofdeactivated fused silica (1 m�0.32 mm i.d.) was used at the frontof the column. The oven was programmed from 60 1C to 170 1C at6 1C min�1 after 1 min, then up to 315 1C at 2.5 1C min�1 and heldfor 10 min. The GC column was fed directly into the EI source of aThermoquest Finnigan TSQ7000 mass spectrometer (ionisationpotential 70 eV; source temperature 215 1C; trap current 300 mA),operated in Full Data Acquisition mode, (50–600 Thompsonscycled every second) and data was processed using Xcalibursoftware. Compounds were identified either by comparison oftheir mass spectra and relative retention indices with thoseavailable in the literature and/or by comparison with authenticstandards. Quantitative data were calculated by comparison ofpeak area of the internal standard (5a(H)-cholestane spiked ontothe samples before extraction) with those of the compounds ofinterest, using the total ion current chromatogram. The relativeresponse factors of the analytes were determined individually for36 representative compounds using authentic standards. Foranalytes for which authentic standards were not available, theresponse factors for similar compounds of the same class and/orsimilar structure were used. Reproducibility of similar lipidanalyses was determined to be o715% by Kiriakoulakis et al.(2000). Data quality was checked regularly with proceduralblanks for each extracted sample batch and organic contamina-tion (which when detected, consisted mainly of C16 and C18 fattyacids and cholesterol) was subtracted from the sample values,although in most cases it was insignificant (o1% of the samplevalues). Total lipids were calculated as the sum of all identifiedcompounds from the total ion chromatograms (TIC).
2.4. Statistical analyses
Principal component analyses (PCA) were conducted on lipidbiomarkers, concentrations being normalised to organic carbon(OC). The analyses were carried out separately for sPOM andsurficial sediments. For sPOM, 83 variables were chosen; thesewere individual identified lipids that occurred in at least one ofthe 45 samples. Where there were zero values, a minimumdetection limit was chosen of one half of the lowest recordedconcentration, namely 0.1 mg g OC�1 (Yunker et al., 2005). In thesame way for surficial sediments 123 variables were chosen thatoccurred in at least one of the 38 samples. Data were fourth roottransformed and screened to confirm a normal distribution usinga Draftsman’s plot for all variables prior to PCA, which was carried
K. Kiriakoulakis et al. / Deep-Sea Research II 58 (2011) 2477–24882482
out using PRIMER software (Primer-E Ltd., UK). All quantitativedata can be found in the Supplementary Material.
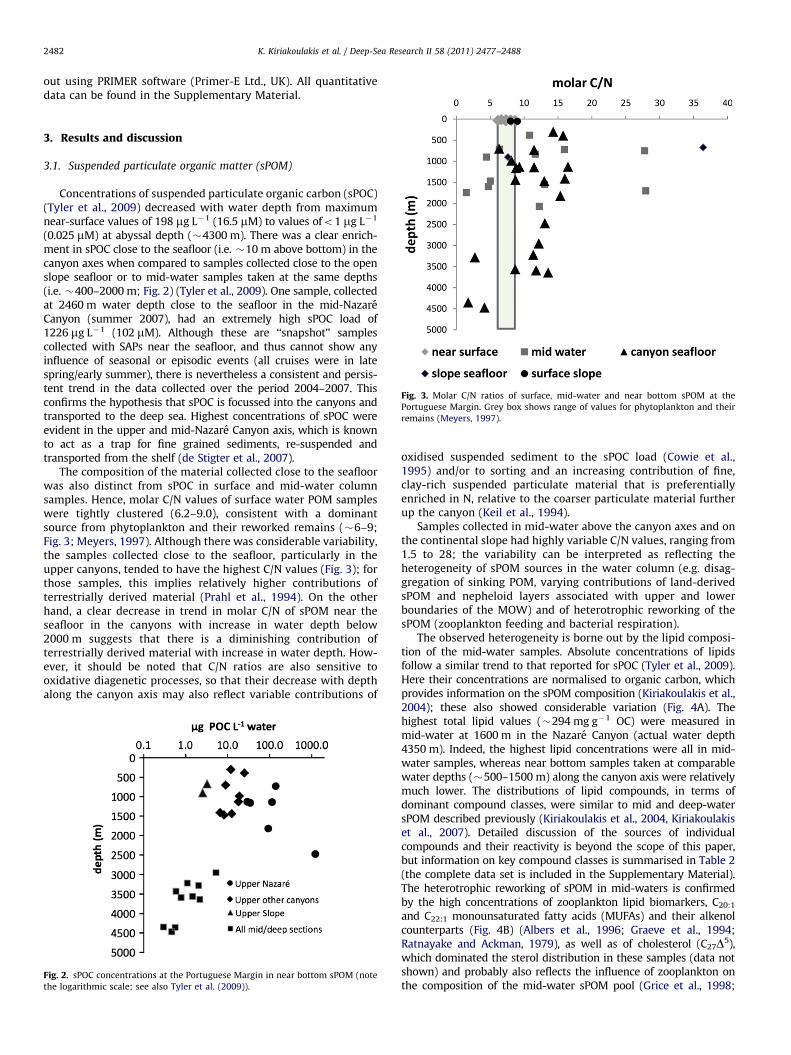
Fig. 3. Molar C/N ratios of surface, mid-water and near bottom sPOM at the
Portuguese Margin. Grey box shows range of values for phytoplankton and their
remains (Meyers, 1997).
3. Results and discussion
3.1. Suspended particulate organic matter (sPOM)
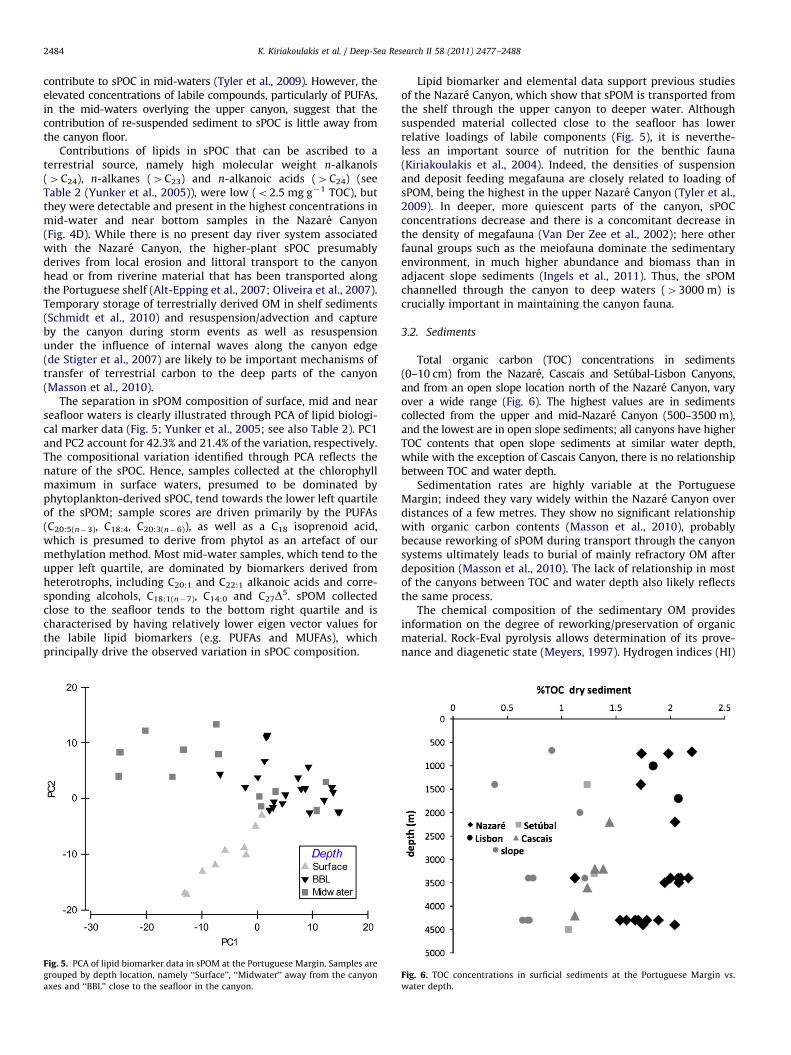
Concentrations of suspended particulate organic carbon (sPOC)(Tyler et al., 2009) decreased with water depth from maximumnear-surface values of 198 mg L�1 (16.5 mM) to values ofo1 mg L�1
(0.025 mM) at abyssal depth (�4300 m). There was a clear enrich-ment in sPOC close to the seafloor (i.e. �10 m above bottom) in thecanyon axes when compared to samples collected close to the openslope seafloor or to mid-water samples taken at the same depths(i.e. �400–2000 m; Fig. 2) (Tyler et al., 2009). One sample, collectedat 2460 m water depth close to the seafloor in the mid-NazareCanyon (summer 2007), had an extremely high sPOC load of1226 mg L�1 (102 mM). Although these are ‘‘snapshot’’ samplescollected with SAPs near the seafloor, and thus cannot show anyinfluence of seasonal or episodic events (all cruises were in latespring/early summer), there is nevertheless a consistent and persis-tent trend in the data collected over the period 2004–2007. Thisconfirms the hypothesis that sPOC is focussed into the canyons andtransported to the deep sea. Highest concentrations of sPOC wereevident in the upper and mid-Nazare Canyon axis, which is knownto act as a trap for fine grained sediments, re-suspended andtransported from the shelf (de Stigter et al., 2007).
The composition of the material collected close to the seafloorwas also distinct from sPOC in surface and mid-water columnsamples. Hence, molar C/N values of surface water POM sampleswere tightly clustered (6.2–9.0), consistent with a dominantsource from phytoplankton and their reworked remains (�6–9;Fig. 3; Meyers, 1997). Although there was considerable variability,the samples collected close to the seafloor, particularly in theupper canyons, tended to have the highest C/N values (Fig. 3); forthose samples, this implies relatively higher contributions ofterrestrially derived material (Prahl et al., 1994). On the otherhand, a clear decrease in trend in molar C/N of sPOM near theseafloor in the canyons with increase in water depth below2000 m suggests that there is a diminishing contribution ofterrestrially derived material with increase in water depth. How-ever, it should be noted that C/N ratios are also sensitive tooxidative diagenetic processes, so that their decrease with depthalong the canyon axis may also reflect variable contributions of
Fig. 2. sPOC concentrations at the Portuguese Margin in near bottom sPOM (note
the logarithmic scale; see also Tyler et al. (2009)).
oxidised suspended sediment to the sPOC load (Cowie et al.,1995) and/or to sorting and an increasing contribution of fine,clay-rich suspended particulate material that is preferentiallyenriched in N, relative to the coarser particulate material furtherup the canyon (Keil et al., 1994).
Samples collected in mid-water above the canyon axes and onthe continental slope had highly variable C/N values, ranging from1.5 to 28; the variability can be interpreted as reflecting theheterogeneity of sPOM sources in the water column (e.g. disag-gregation of sinking POM, varying contributions of land-derivedsPOM and nepheloid layers associated with upper and lowerboundaries of the MOW) and of heterotrophic reworking of thesPOM (zooplankton feeding and bacterial respiration).
The observed heterogeneity is borne out by the lipid composi-tion of the mid-water samples. Absolute concentrations of lipidsfollow a similar trend to that reported for sPOC (Tyler et al., 2009).Here their concentrations are normalised to organic carbon, whichprovides information on the sPOM composition (Kiriakoulakis et al.,2004); these also showed considerable variation (Fig. 4A). Thehighest total lipid values (�294 mg g�1 OC) were measured inmid-water at 1600 m in the Nazare Canyon (actual water depth4350 m). Indeed, the highest lipid concentrations were all in mid-water samples, whereas near bottom samples taken at comparablewater depths (�500–1500 m) along the canyon axis were relativelymuch lower. The distributions of lipid compounds, in terms ofdominant compound classes, were similar to mid and deep-watersPOM described previously (Kiriakoulakis et al., 2004, Kiriakoulakiset al., 2007). Detailed discussion of the sources of individualcompounds and their reactivity is beyond the scope of this paper,but information on key compound classes is summarised in Table 2(the complete data set is included in the Supplementary Material).The heterotrophic reworking of sPOM in mid-waters is confirmedby the high concentrations of zooplankton lipid biomarkers, C20:1
and C22:1 monounsaturated fatty acids (MUFAs) and their alkenolcounterparts (Fig. 4B) (Albers et al., 1996; Graeve et al., 1994;Ratnayake and Ackman, 1979), as well as of cholesterol (C27D5),which dominated the sterol distribution in these samples (data notshown) and probably also reflects the influence of zooplankton onthe composition of the mid-water sPOM pool (Grice et al., 1998;
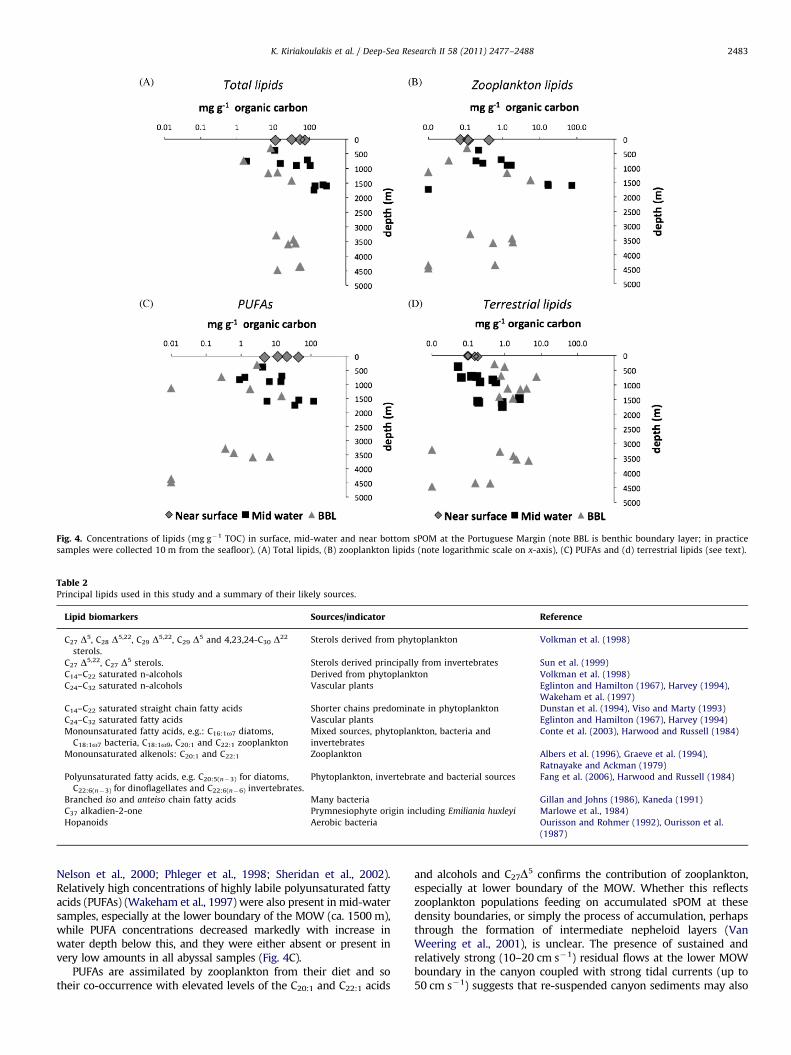
Fig. 4. Concentrations of lipids (mg g�1 TOC) in surface, mid-water and near bottom sPOM at the Portuguese Margin (note BBL is benthic boundary layer; in practice
samples were collected 10 m from the seafloor). (A) Total lipids, (B) zooplankton lipids (note logarithmic scale on x-axis), (C) PUFAs and (d) terrestrial lipids (see text).
Table 2Principal lipids used in this study and a summary of their likely sources.
Lipid biomarkers Sources/indicator Reference
C27 D5, C28 D5,22, C29 D5,22, C29 D5 and 4,23,24-C30 D22
sterols.
Sterols derived from phytoplankton Volkman et al. (1998)
C27 D5,22, C27 D5 sterols. Sterols derived principally from invertebrates Sun et al. (1999)
C14–C22 saturated n-alcohols Derived from phytoplankton Volkman et al. (1998)
C24–C32 saturated n-alcohols Vascular plants Eglinton and Hamilton (1967), Harvey (1994),
Wakeham et al. (1997)
C14–C22 saturated straight chain fatty acids Shorter chains predominate in phytoplankton Dunstan et al. (1994), Viso and Marty (1993)
C24–C32 saturated fatty acids Vascular plants Eglinton and Hamilton (1967), Harvey (1994)
Monounsaturated fatty acids, e.g.: C16:1o7 diatoms,
C18:1o7 bacteria, C18:1o9, C20:1 and C22:1 zooplankton
Mixed sources, phytoplankton, bacteria and
invertebrates
Conte et al. (2003), Harwood and Russell (1984)
Monounsaturated alkenols: C20:1 and C22:1 Zooplankton Albers et al. (1996), Graeve et al. (1994),
Ratnayake and Ackman (1979)
Polyunsaturated fatty acids, e.g. C20:5(n�3) for diatoms,
C22:6(n�3) for dinoflagellates and C22:6(n�6) invertebrates.
Phytoplankton, invertebrate and bacterial sources Fang et al. (2006), Harwood and Russell (1984)
Branched iso and anteiso chain fatty acids Many bacteria Gillan and Johns (1986), Kaneda (1991)
C37 alkadien-2-one Prymnesiophyte origin including Emiliania huxleyi Marlowe et al., 1984)
Hopanoids Aerobic bacteria Ourisson and Rohmer (1992), Ourisson et al.
(1987)
K. Kiriakoulakis et al. / Deep-Sea Research II 58 (2011) 2477–2488 2483
Nelson et al., 2000; Phleger et al., 1998; Sheridan et al., 2002).Relatively high concentrations of highly labile polyunsaturated fattyacids (PUFAs) (Wakeham et al., 1997) were also present in mid-watersamples, especially at the lower boundary of the MOW (ca. 1500 m),while PUFA concentrations decreased markedly with increase inwater depth below this, and they were either absent or present invery low amounts in all abyssal samples (Fig. 4C).
PUFAs are assimilated by zooplankton from their diet and sotheir co-occurrence with elevated levels of the C20:1 and C22:1 acids
and alcohols and C27D5 confirms the contribution of zooplankton,especially at lower boundary of the MOW. Whether this reflectszooplankton populations feeding on accumulated sPOM at thesedensity boundaries, or simply the process of accumulation, perhapsthrough the formation of intermediate nepheloid layers (VanWeering et al., 2001), is unclear. The presence of sustained andrelatively strong (10–20 cm s�1) residual flows at the lower MOWboundary in the canyon coupled with strong tidal currents (up to50 cm s�1) suggests that re-suspended canyon sediments may also
K. Kiriakoulakis et al. / Deep-Sea Research II 58 (2011) 2477–24882484
contribute to sPOC in mid-waters (Tyler et al., 2009). However, theelevated concentrations of labile compounds, particularly of PUFAs,in the mid-waters overlying the upper canyon, suggest that thecontribution of re-suspended sediment to sPOC is little away fromthe canyon floor.
Contributions of lipids in sPOC that can be ascribed to aterrestrial source, namely high molecular weight n-alkanols(4C24), n-alkanes (4C23) and n-alkanoic acids (4C24) (seeTable 2 (Yunker et al., 2005)), were low (o2.5 mg g�1 TOC), butthey were detectable and present in the highest concentrations inmid-water and near bottom samples in the Nazare Canyon(Fig. 4D). While there is no present day river system associatedwith the Nazare Canyon, the higher-plant sPOC presumablyderives from local erosion and littoral transport to the canyonhead or from riverine material that has been transported alongthe Portuguese shelf (Alt-Epping et al., 2007; Oliveira et al., 2007).Temporary storage of terrestrially derived OM in shelf sediments(Schmidt et al., 2010) and resuspension/advection and captureby the canyon during storm events as well as resuspensionunder the influence of internal waves along the canyon edge(de Stigter et al., 2007) are likely to be important mechanisms oftransfer of terrestrial carbon to the deep parts of the canyon(Masson et al., 2010).
The separation in sPOM composition of surface, mid and nearseafloor waters is clearly illustrated through PCA of lipid biologi-cal marker data (Fig. 5; Yunker et al., 2005; see also Table 2). PC1and PC2 account for 42.3% and 21.4% of the variation, respectively.The compositional variation identified through PCA reflects thenature of the sPOC. Hence, samples collected at the chlorophyllmaximum in surface waters, presumed to be dominated byphytoplankton-derived sPOC, tend towards the lower left quartileof the sPOM; sample scores are driven primarily by the PUFAs(C20:5(n�3), C18:4, C20:3(n�6)), as well as a C18 isoprenoid acid,which is presumed to derive from phytol as an artefact of ourmethylation method. Most mid-water samples, which tend to theupper left quartile, are dominated by biomarkers derived fromheterotrophs, including C20:1 and C22:1 alkanoic acids and corre-sponding alcohols, C18:1(n�7), C14:0 and C27D5. sPOM collectedclose to the seafloor tends to the bottom right quartile and ischaracterised by having relatively lower eigen vector values forthe labile lipid biomarkers (e.g. PUFAs and MUFAs), whichprincipally drive the observed variation in sPOC composition.
Fig. 5. PCA of lipid biomarker data in sPOM at the Portuguese Margin. Samples are
grouped by depth location, namely ‘‘Surface’’, ‘‘Midwater’’ away from the canyon
axes and ‘‘BBL’’ close to the seafloor in the canyon.
Lipid biomarker and elemental data support previous studiesof the Nazare Canyon, which show that sPOM is transported fromthe shelf through the upper canyon to deeper water. Althoughsuspended material collected close to the seafloor has lowerrelative loadings of labile components (Fig. 5), it is neverthe-less an important source of nutrition for the benthic fauna(Kiriakoulakis et al., 2004). Indeed, the densities of suspensionand deposit feeding megafauna are closely related to loading ofsPOM, being the highest in the upper Nazare Canyon (Tyler et al.,2009). In deeper, more quiescent parts of the canyon, sPOCconcentrations decrease and there is a concomitant decrease inthe density of megafauna (Van Der Zee et al., 2002); here otherfaunal groups such as the meiofauna dominate the sedimentaryenvironment, in much higher abundance and biomass than inadjacent slope sediments (Ingels et al., 2011). Thus, the sPOMchannelled through the canyon to deep waters (43000 m) iscrucially important in maintaining the canyon fauna.
3.2. Sediments
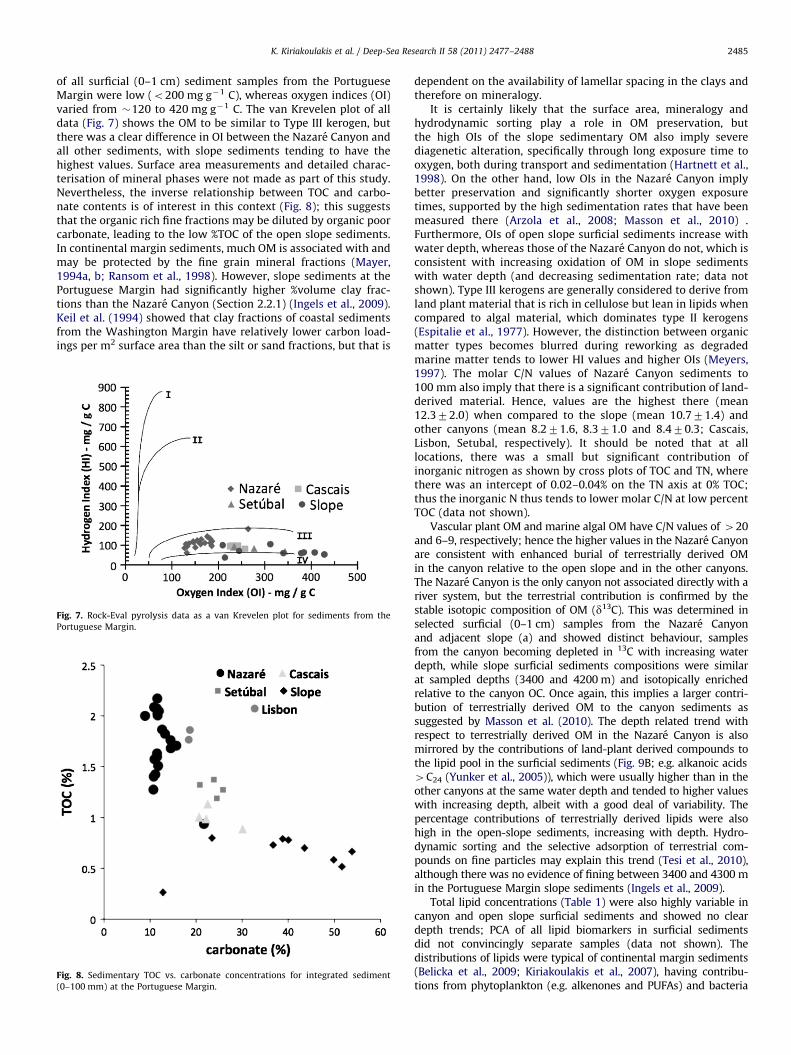
Total organic carbon (TOC) concentrations in sediments(0–10 cm) from the Nazare, Cascais and Setubal-Lisbon Canyons,and from an open slope location north of the Nazare Canyon, varyover a wide range (Fig. 6). The highest values are in sedimentscollected from the upper and mid-Nazare Canyon (500–3500 m),and the lowest are in open slope sediments; all canyons have higherTOC contents that open slope sediments at similar water depth,while with the exception of Cascais Canyon, there is no relationshipbetween TOC and water depth.
Sedimentation rates are highly variable at the PortugueseMargin; indeed they vary widely within the Nazare Canyon overdistances of a few metres. They show no significant relationshipwith organic carbon contents (Masson et al., 2010), probablybecause reworking of sPOM during transport through the canyonsystems ultimately leads to burial of mainly refractory OM afterdeposition (Masson et al., 2010). The lack of relationship in mostof the canyons between TOC and water depth also likely reflectsthe same process.
The chemical composition of the sedimentary OM providesinformation on the degree of reworking/preservation of organicmaterial. Rock-Eval pyrolysis allows determination of its prove-nance and diagenetic state (Meyers, 1997). Hydrogen indices (HI)
Fig. 6. TOC concentrations in surficial sediments at the Portuguese Margin vs.
water depth.
K. Kiriakoulakis et al. / Deep-Sea Research II 58 (2011) 2477–2488 2485
of all surficial (0–1 cm) sediment samples from the PortugueseMargin were low (o200 mg g�1 C), whereas oxygen indices (OI)varied from �120 to 420 mg g�1 C. The van Krevelen plot of alldata (Fig. 7) shows the OM to be similar to Type III kerogen, butthere was a clear difference in OI between the Nazare Canyon andall other sediments, with slope sediments tending to have thehighest values. Surface area measurements and detailed charac-terisation of mineral phases were not made as part of this study.Nevertheless, the inverse relationship between TOC and carbo-nate contents is of interest in this context (Fig. 8); this suggeststhat the organic rich fine fractions may be diluted by organic poorcarbonate, leading to the low %TOC of the open slope sediments.In continental margin sediments, much OM is associated with andmay be protected by the fine grain mineral fractions (Mayer,1994a, b; Ransom et al., 1998). However, slope sediments at thePortuguese Margin had significantly higher %volume clay frac-tions than the Nazare Canyon (Section 2.2.1) (Ingels et al., 2009).Keil et al. (1994) showed that clay fractions of coastal sedimentsfrom the Washington Margin have relatively lower carbon load-ings per m2 surface area than the silt or sand fractions, but that is
Fig. 7. Rock-Eval pyrolysis data as a van Krevelen plot for sediments from the
Portuguese Margin.
Fig. 8. Sedimentary TOC vs. carbonate concentrations for integrated sediment
(0–100 mm) at the Portuguese Margin.
dependent on the availability of lamellar spacing in the clays andtherefore on mineralogy.
It is certainly likely that the surface area, mineralogy andhydrodynamic sorting play a role in OM preservation, butthe high OIs of the slope sedimentary OM also imply severediagenetic alteration, specifically through long exposure time tooxygen, both during transport and sedimentation (Hartnett et al.,1998). On the other hand, low OIs in the Nazare Canyon implybetter preservation and significantly shorter oxygen exposuretimes, supported by the high sedimentation rates that have beenmeasured there (Arzola et al., 2008; Masson et al., 2010) .Furthermore, OIs of open slope surficial sediments increase withwater depth, whereas those of the Nazare Canyon do not, which isconsistent with increasing oxidation of OM in slope sedimentswith water depth (and decreasing sedimentation rate; data notshown). Type III kerogens are generally considered to derive fromland plant material that is rich in cellulose but lean in lipids whencompared to algal material, which dominates type II kerogens(Espitalie et al., 1977). However, the distinction between organicmatter types becomes blurred during reworking as degradedmarine matter tends to lower HI values and higher OIs (Meyers,1997). The molar C/N values of Nazare Canyon sediments to100 mm also imply that there is a significant contribution of land-derived material. Hence, values are the highest there (mean12.372.0) when compared to the slope (mean 10.771.4) andother canyons (mean 8.271.6, 8.371.0 and 8.470.3; Cascais,Lisbon, Setubal, respectively). It should be noted that at alllocations, there was a small but significant contribution ofinorganic nitrogen as shown by cross plots of TOC and TN, wherethere was an intercept of 0.02–0.04% on the TN axis at 0% TOC;thus the inorganic N thus tends to lower molar C/N at low percentTOC (data not shown).
Vascular plant OM and marine algal OM have C/N values of 420and 6–9, respectively; hence the higher values in the Nazare Canyonare consistent with enhanced burial of terrestrially derived OMin the canyon relative to the open slope and in the other canyons.The Nazare Canyon is the only canyon not associated directly with ariver system, but the terrestrial contribution is confirmed by thestable isotopic composition of OM (d13C). This was determined inselected surficial (0–1 cm) samples from the Nazare Canyonand adjacent slope (a) and showed distinct behaviour, samplesfrom the canyon becoming depleted in 13C with increasing waterdepth, while slope surficial sediments compositions were similarat sampled depths (3400 and 4200 m) and isotopically enrichedrelative to the canyon OC. Once again, this implies a larger contri-bution of terrestrially derived OM to the canyon sediments assuggested by Masson et al. (2010). The depth related trend withrespect to terrestrially derived OM in the Nazare Canyon is alsomirrored by the contributions of land-plant derived compounds tothe lipid pool in the surficial sediments (Fig. 9B; e.g. alkanoic acids4C24 (Yunker et al., 2005)), which were usually higher than in theother canyons at the same water depth and tended to higher valueswith increasing depth, albeit with a good deal of variability. Thepercentage contributions of terrestrially derived lipids were alsohigh in the open-slope sediments, increasing with depth. Hydro-dynamic sorting and the selective adsorption of terrestrial com-pounds on fine particles may explain this trend (Tesi et al., 2010),although there was no evidence of fining between 3400 and 4300 min the Portuguese Margin slope sediments (Ingels et al., 2009).
Total lipid concentrations (Table 1) were also highly variable incanyon and open slope surficial sediments and showed no cleardepth trends; PCA of all lipid biomarkers in surficial sedimentsdid not convincingly separate samples (data not shown). Thedistributions of lipids were typical of continental margin sediments(Belicka et al., 2009; Kiriakoulakis et al., 2007), having contribu-tions from phytoplankton (e.g. alkenones and PUFAs) and bacteria
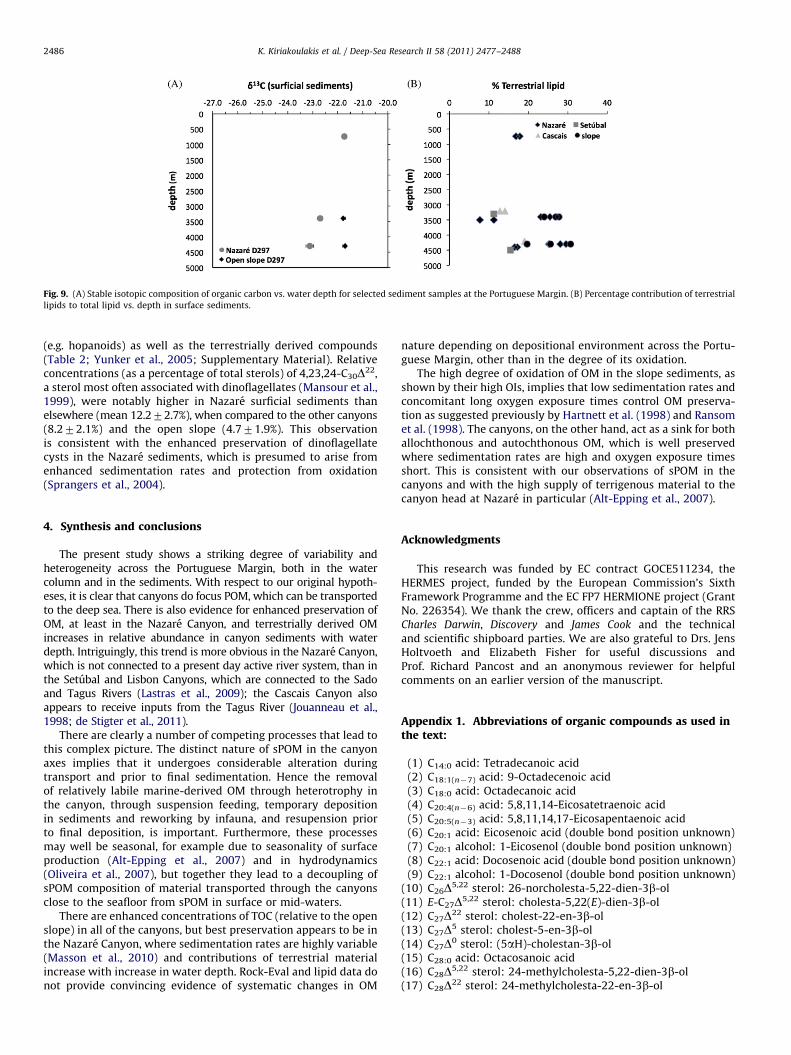
Fig. 9. (A) Stable isotopic composition of organic carbon vs. water depth for selected sediment samples at the Portuguese Margin. (B) Percentage contribution of terrestrial
lipids to total lipid vs. depth in surface sediments.
K. Kiriakoulakis et al. / Deep-Sea Research II 58 (2011) 2477–24882486
(e.g. hopanoids) as well as the terrestrially derived compounds(Table 2; Yunker et al., 2005; Supplementary Material). Relativeconcentrations (as a percentage of total sterols) of 4,23,24-C30D22,a sterol most often associated with dinoflagellates (Mansour et al.,1999), were notably higher in Nazare surficial sediments thanelsewhere (mean 12.272.7%), when compared to the other canyons(8.272.1%) and the open slope (4.771.9%). This observationis consistent with the enhanced preservation of dinoflagellatecysts in the Nazare sediments, which is presumed to arise fromenhanced sedimentation rates and protection from oxidation(Sprangers et al., 2004).
4. Synthesis and conclusions
The present study shows a striking degree of variability andheterogeneity across the Portuguese Margin, both in the watercolumn and in the sediments. With respect to our original hypoth-eses, it is clear that canyons do focus POM, which can be transportedto the deep sea. There is also evidence for enhanced preservation ofOM, at least in the Nazare Canyon, and terrestrially derived OMincreases in relative abundance in canyon sediments with waterdepth. Intriguingly, this trend is more obvious in the Nazare Canyon,which is not connected to a present day active river system, than inthe Setubal and Lisbon Canyons, which are connected to the Sadoand Tagus Rivers (Lastras et al., 2009); the Cascais Canyon alsoappears to receive inputs from the Tagus River (Jouanneau et al.,1998; de Stigter et al., 2011).
There are clearly a number of competing processes that lead tothis complex picture. The distinct nature of sPOM in the canyonaxes implies that it undergoes considerable alteration duringtransport and prior to final sedimentation. Hence the removalof relatively labile marine-derived OM through heterotrophy inthe canyon, through suspension feeding, temporary depositionin sediments and reworking by infauna, and resupension priorto final deposition, is important. Furthermore, these processesmay well be seasonal, for example due to seasonality of surfaceproduction (Alt-Epping et al., 2007) and in hydrodynamics(Oliveira et al., 2007), but together they lead to a decoupling ofsPOM composition of material transported through the canyonsclose to the seafloor from sPOM in surface or mid-waters.
There are enhanced concentrations of TOC (relative to the openslope) in all of the canyons, but best preservation appears to be inthe Nazare Canyon, where sedimentation rates are highly variable(Masson et al., 2010) and contributions of terrestrial materialincrease with increase in water depth. Rock-Eval and lipid data donot provide convincing evidence of systematic changes in OM
nature depending on depositional environment across the Portu-guese Margin, other than in the degree of its oxidation.
The high degree of oxidation of OM in the slope sediments, asshown by their high OIs, implies that low sedimentation rates andconcomitant long oxygen exposure times control OM preserva-tion as suggested previously by Hartnett et al. (1998) and Ransomet al. (1998). The canyons, on the other hand, act as a sink for bothallochthonous and autochthonous OM, which is well preservedwhere sedimentation rates are high and oxygen exposure timesshort. This is consistent with our observations of sPOM in thecanyons and with the high supply of terrigenous material to thecanyon head at Nazare in particular (Alt-Epping et al., 2007).
Acknowledgments
This research was funded by EC contract GOCE511234, theHERMES project, funded by the European Commission’s SixthFramework Programme and the EC FP7 HERMIONE project (GrantNo. 226354). We thank the crew, officers and captain of the RRSCharles Darwin, Discovery and James Cook and the technicaland scientific shipboard parties. We are also grateful to Drs. JensHoltvoeth and Elizabeth Fisher for useful discussions andProf. Richard Pancost and an anonymous reviewer for helpfulcomments on an earlier version of the manuscript.
Appendix 1. Abbreviations of organic compounds as used inthe text:
(1)
C14:0 acid: Tetradecanoic acid (2) C18:1(n�7) acid: 9-Octadecenoic acid (3) C18:0 acid: Octadecanoic acid (4) C20:4(n�6) acid: 5,8,11,14-Eicosatetraenoic acid (5) C20:5(n�3) acid: 5,8,11,14,17-Eicosapentaenoic acid (6) C20:1 acid: Eicosenoic acid (double bond position unknown) (7) C20:1 alcohol: 1-Eicosenol (double bond position unknown) (8) C22:1 acid: Docosenoic acid (double bond position unknown) (9) C22:1 alcohol: 1-Docosenol (double bond position unknown)(10)
C26D5,22 sterol: 26-norcholesta-5,22-dien-3b-ol (11) E-C27D5,22 sterol: cholesta-5,22(E)-dien-3b-ol (12) C27D22 sterol: cholest-22-en-3b-ol (13) C27D5 sterol: cholest-5-en-3b-ol (14) C27D0 sterol: (5aH)-cholestan-3b-ol (15) C28:0 acid: Octacosanoic acid (16) C28D5,22 sterol: 24-methylcholesta-5,22-dien-3b-ol (17) C28D22 sterol: 24-methylcholesta-22-en-3b-ol
K. Kiriakoulakis et al. / Deep-Sea Research II 58 (2011) 2477–2488 2487
(18)
C29D5,22 sterol: 24-ethylcholesta-5,22-dien-3b-ol (19) C29D5 sterol: 24-methylcholesta-5-en-3b-ol (20) C29D0 sterol: (5aH)-24-methylcholestan-3b-ol (21) 4,23,24-C30D22 sterol: (5aH)-4,23,24-trimethylcholest-22-en-3b-ol, dinosterol
Appendix A. Supplementary material
Supplementary data associated with this article can be foundin the online version at doi:10.1016/j.dsr2.2011.04.010.
References
Accornero, A., Picon, P., de Bovee, F., 2002. Biogeochemical transformations ofdissolved organic matter at the sediment–water interface: results from in situincubations within benthic chambers. Oceanologica Acta 25, 171–178.
Albers, C.S., Kattner, G., Hagen, W., 1996. The compositions of wax esters,triacylglycerols and phospholipids in Arctic and Antarctic copepods: evidenceof energetic adaptations. Marine Chemistry 55, 347–358.
Alt-Epping, U., Mil-Homens, M., Hebbeln, D., Abrantes, F., Schneider, R.R., 2007.Provenance of organic matter and nutrient conditions on a river- andupwelling influenced shelf: a case study from the Portuguese Margin. MarineGeology 243, 169–179.
Arzola, R.G., Wynn, R.B., Lastras, G., Masson, D.G., Weaver, P.P.E., 2008. Sedimen-tary features and processes in the Nazare and Setubal submarine canyons,west Iberian margin. Marine Geology 250, 64–88.
Belicka, L.L., Macdonald, R.W., Harvey, H.R., 2009. Trace element and molecularmarkers of organic carbon dynamics along a shelf-basin continuum insediments of the western Arctic Ocean. Marine Chemistry 115, 72–85.
Boyer, D.L., Haidvogel, D.B., Perenne, N., 2004. Laboratory-numerical modelcomparisons of canyon flows: a parameter study. Journal of Physical Oceano-graphy 34, 1588–1609.
Canals, M., Puig, P., Durrieu du Madron, X., Heussner, S., Palanques, A., Fabres, J.,2006. Flushing submarine canyons. Nature 444, 354–357.
Christie, W.W., 1982. Esterification of fatty-acids in adipose-tissue. Journal of theScience of Food and Agriculture 33, 809.
Conte, M.H., Dickey, T.D., Weber, J.C., Johnson, R.J., Knap, A.H., 2003. Transientphysical forcing of pulsed export of bioreactive material to the deep SargassoSea. Deep-Sea Research I 50, 1157–1187.
Cowie, G.L., Hedges, J.I., Prahl, F.G., Delange, G.J., 1995. Elemental and majorbiochemical-changes across on oxidation front in a relict turbidite—an oxygeneffect. Geochimica Et Cosmochimica Acta 59, 33–46.
de Haas, H., Van Weering, T.C.E., de Stigter, H.C., 2002. Organic carbon in shelfseas: sinks or sources, processes and products. Continental Shelf Research 22,691–717.
de Stigter, H.C., Boer, W., Mendes, P., Jesus, C.C., Thomsen, L., Van Den Bergh, G.D.,Van Weering, T.C.E., 2007. Recent sediment transport and deposition in theNazare Canyon, Portuguese continental margin. Marine Geology 246,144–164.
de Stigter, H.C., Jesus, C.C., Boer, W., Richter, T.O., Costa, A., van Weering, T.C.E.,2011. Recent sediment transport and deposition in the Lisbon-Setubal andCascais Canyon, Portuguese continental margin. Deep-Sea Research II 58,2321–2344.
Duineveld, G., Lavaleye, M., Berghuis, E., de Wilde, P., 2001. Activity and composi-tion of the benthic fauna in the Whittard Canyon and the adjacent continentalslope (NE Atlantic). Oceanologica Acta 24, 69–83.
Dunstan, G.A., Volkman, J.K., Barrett, S.M., Leroi, J.M., Jeffrey, S.W., 1994. Essentialpolyunsaturated fatty-acids from 14 species of diatom (bacillariophyceae).Phytochemistry 35, 155–161.
Eglinton, G., Hamilton, R.J., 1967. Leaf epicuticular waxes. Science 156, 1322–1335.Espitalie, J., Laporte, J.L., Madec, M., Marquis, F., Leplat, P., Paulet, J., Boutefeu, A.,
1977. Rapid method for source rocks characterization and for determination ofpetroleum potential and degree of evolution. Revue De L Institut Francais DuPetrole 32, 23–42.
Fang, J.S., Uhle, M., Billmark, K., Bartlett, D.H., Kato, C., 2006. Fractionation ofcarbon isotopes in biosynthesis of fatty acids by a piezophilic bacteriumMoritella japonica strain dsk1. Geochimica Et Cosmochimica Acta 70,1753–1760.
Gardner, W.D., 1989a. Baltimore canyon as a modern conduit of sediment to thedeep-sea. Deep-Sea Research I 36, 323–358.
Gardner, W.D., 1989b. Periodic resuspension in baltimore canyon by focusing ofinternal waves. Journal of Geophysical Research—Oceans 94, 18185–18194.
Gillan, F.T., Johns, R.B., 1986. Chemical markers for marine bacteria: fatty acids andpigments. In: Johns, R.B. (Ed.), Biological Markers in the Sedimentary Environ-ment. Elsevier Press, New York.
Graeve, M., Hagen, W., Kattner, G., 1994. Herbivorous or omnivorous—on thesignificance of lipid compositions as trophic markers in antarctic copepods.Deep-Sea Research I 41, 915–924.
Grice, K., Breteler, W., Schouten, S., Grossi, V., de Leeuw, J.W., Damste, J.S.S., 1998.Effects of zooplankton herbivory on biomarker proxy records. Paleoceanogra-phy 13, 686–693.
Hartnett, H.E., Keil, R.G., Hedges, J.I., Devol, A.H., 1998. Influence of oxygenexposure time on organic carbon preservation in continental margin sedi-ments. Nature 391, 572–574.
Harvey, H.R., 1994. Fatty-acids and sterols as source markers of organic-matter insediments of the North-Carolina continental-slope. Deep-Sea Research II 41,783–796.
Harwood, J.L., Russell, N.L., 1984. Lipids in Plants and Microorganisms. GeorgeAllen and Unwin, London.
Ingels, J., Billett, D.S.M., Kiriakoulakis, K., Wolff, G.A., Vanreusel, A., 2011. Structuraland functional diversity of Nematoda in relation with environmental variablesin the Setubal and Cascais canyons, Western Iberian Margin. Deep-SeaResearch II 58, 2354–2368.
Ingels, J., Kiriakoulakis, K., Wolff, G.A., Vanreusel, A., 2009. Nematode diversity and itsrelation to the quantity and quality of sedimentary organic matter in the deepNazare Canyon, western Iberian margin. Deep-Sea Research I 56, 1521–1539.
Jouanneau, J.M., Garcia, C., Oliveira, A., Rodrigues, A., Dias, J.A., Weber, O., 1998.Dispersal and deposition of suspended sediment on the shelf off the Tagus andSado estuaries, SW Portugal. Progress in Oceanography 42, 233–257.
Kaneda, T., 1991. Iso-fatty and anteiso-fatty acids in bacteria—biosynthesis,function, and taxonomic significance. Microbiological Reviews 55, 288–302.
Keil, R.G., Tsamakis, E., Fuh, C.B., Giddings, J.C., Hedges, J.I., 1994. Mineralogical andtextural controls on the organic composition of coastal marine-sediments—
hydrodynamic separation using splitt-fractionation. Geochimica Et Cosmochi-mica Acta 58, 879–893.
Kim, J.H., Schouten, S., Buscail, R., Ludwig, W., Bonnin, J., Damste, J.S.S., Bourrin, F.,2006. Origin and distribution of terrestrial organic matter in the NWMediterranean (Gulf of Lions): exploring the newly developed bit index.Geochemistry Geophysics Geosystems, 7.
Kiriakoulakis, K., Bett, B.J., White, M., Wolff, G.A., 2004. Organic biogeochemistry ofthe Darwin Mounds, a deep-water coral ecosystem, of the NE Atlantic. Deep-Sea Research I 51, 1937–1954.
Kiriakoulakis, K., Freiwald, A., Fisher, E., Wolff, G.A., 2007. Organic matter qualityand supply to deep-water coral/mound systems of the NW European con-tinental margin. International Journal of Earth Sciences 96, 159–170.
Kiriakoulakis, K., Marshall, J.D., Wolff, G.A., 2000. Biomarkers in a lowerJurassic concretion from Dorset (UK). Journal of the Geological Society 157,207–220.
Koho, K.A., Garcia, R., de Stigter, H.C., Epping, E., Koning, E., Kouwenhoven, T.J., VanDer Zwaan, G.J., 2008. Sedimentary labile organic carbon and pore water redoxcontrol on species distribution of benthic foraminifera: a case study fromLisbon–Setubal canyon (southern Portugal). Progress in Oceanography 79,55–82.
Lastras, G., Arzola, R.G., Masson, D.G., Wynn, R.B., Huvenne, V.A.I., Huhnerbach, V.,Canals, M., 2009. Geomorphology and sedimentary features in the centralPortuguese submarine canyons, western Iberian margin. Geomorphology 103,310–329.
Liu, Z.F., Stewart, G., Cochran, J.K., Lee, C., Armstrong, R.A., Hirschberg, D.J., Gasser,B., Miquel, J.C., 2005. Why do POC concentrations measured using niskin bottlecollections sometimes differ from those using in-situ pumps? Deep-SeaResearch I 52, 1324–1344.
Mansour, M.P., Volkman, J.K., Jackson, A.E., Blackburn, S.I., 1999. The fatty acid andsterol composition of five marine dinoflagellates. Journal of Phycology 35,710–720.
Marlowe, I.T., Green, J.C., Neal, A.C., Brassell, S.C., Eglinton, G., Course, P.A., 1984.Long-chain (n�C37–C39) alkenones in the prymnesiophyceae—distribution ofalkenones and other lipids and their taxonomic significance. British Phycolo-gical Journal 19, 203–216.
Masson, D.G., Huvenne, V.A.I., de Stigter, H.C., Wolff, G.A., Kiriakoulakis, K., Arzola,R.G., Blackbird, S., 2010. Efficient burial of carbon in a submarine canyon.Geology 38, 831–834.
Mayer, L.M., 1994a. Relationships between mineral surfaces and organic-carbonconcentrations in soils and sediments. Chemical Geology 114, 347–363.
Mayer, L.M., 1994b. Surface-area control of organic-carbon accumulation incontinental-shelf sediments. Geochimica Et Cosmochimica Acta 58,1271–1284.
Meyers, P.A., 1997. Organic geochemical proxies of paleoceanographic, paleolim-nologic, and paleoclimatic processes. Organic Geochemistry 27, 213–250.
Nelson, M.M., Phleger, C.F., Mooney, B.D., Nichols, P.D., 2000. Lipids of gelatinousAntarctic zooplankton: Cnidaria and Ctenophora. Lipids 35, 551–559.
Oliveira, A., Santos, A.I., Rodrigues, A., Vitorino, J., 2007. Sedimentary particledistribution and dynamics on the Nazare Canyon system and adjacent shelf(Portugal). Marine Geology 246, 105–122.
Ourisson, G., Rohmer, M., 1992. Hopanoids. 2. Biohopanoids—a novel class ofbacterial lipids. Accounts of Chemical Research 25, 403–408.
Ourisson, G., Rohmer, M., Poralla, K., 1987. Prokaryotic hopanoids and otherpolyterpenoid sterol surrogates. Annual Review of Microbiology 41, 301–333.
Perenne, N., Haidvogel, D.B., Boyer, D.L., 2001a. Laboratory-numerical modelcomparisons of flow over a coastal canyon. Journal of Atmospheric andOceanic Technology 18, 235–255.
Perenne, N., Lavelle, J.W., Smith, D.C., Boyer, D.L., 2001b. Impulsively startedflow in a submarine canyon: comparison of results from laboratory andnumerical models. Journal of Atmospheric and Oceanic Technology 18,1698–1718.

K. Kiriakoulakis et al. / Deep-Sea Research II 58 (2011) 2477–24882488
Phleger, C.F., Nichols, P.D., Virtue, P., 1998. Lipids and trophodynamics of Antarcticzooplankton. Comparative Biochemistry and Physiology B—Biochemistry andMolecular Biology 120 (311-323).
Prahl, F.G., Ertel, J.R., Goni, M.A., Sparrow, M.A., Eversmeyer, B., 1994. Terrestrialorganic-carbon contributions to sediments on the Washington margin. Geo-chimica Et Cosmochimica Acta 58, 3035–3048.
Ransom, B., Kim, D., Kastner, M., Wainwright, S., 1998. Organic matter preserva-tion on continental slopes: importance of mineralogy and surface area.Geochimica Et Cosmochimica Acta 62, 1329–1345.
Ratnayake, W.N., Ackman, R.G., 1979. Fatty alcohols in capelin, herring andmackerel oils and muscle lipids. 2. Comparison of fatty-acids from wax esterswith those of triglycerides. Lipids 14, 804–810.
Schmidt, F., Hinrichs, K.-U., Elvert, M., 2010. Sources, transport, and partitioning oforganic matter at a highly dynamic continental margin. Marine Chemistry 118,37–55.
Schmiedl, G., Pfeilsticker, M., Hemleben, C., Mackensen, A., 2004. Environmentaland biological effects on the stable isotope composition of recent deep-seabenthic foraminifera, from the western Mediterranean Sea. Marine Micro-paleontology 51, 129–152.
Sheridan, C.C., Lee, C., Wakeham, S.G., Bishop, J.K.B., 2002. Suspended particleorganic composition and cycling in surface and midwaters of the equatorialPacific ocean. Deep-Sea Research I 49, 1983–2008.
Sprangers, M., Dammers, N., Brinkhuis, H., Van Weering, T.C.E., Lotter, A.F., 2004.Modern organic-walled dinoflagellate cyst distribution offshore NW Iberia;tracing the upwelling system. Review of Palaeobotany and Palynology 128,97–106.
Sun, M.Y., Aller, R.C., Lee, C., Wakeham, S.G., 1999. Enhanced degradation of algallipids by benthic macrofaunal activity: effect of Yoldia limatula. Journal ofMarine Research 57, 775–804.
Tesi, T., Langone, L., Goni, M.A., Turchetto, M., Miserocchi, S., Boldrin, A., 2008.Source and composition of organic matter in the Bari Canyon (Italy): densewater cascading versus particulate export from the upper ocean. Deep-SeaResearch I 55, 813–831.
Tesi, T., Puig, P., Palanques, A., Goni, M.A., 2010. Lateral advection of organic matterin cascading-dominated submarine canyons. Progress in Oceanography 84,185–203.
Turnewitsch, R., Springer, B.M., Kiriakoulakis, K., Vilas, J.C., Aristegui, J., Wolff, G.,Peine, F., Werk, S., Graf, G., Waniek, J.J., 2007. Determination of particulateorganic carbon (POC) in seawater: the relative methodological importance ofartificial gains and losses in two glass-fiber-filter-based techniques. MarineChemistry 105, 208–228.
Tyler, P., Amaro, T., Arzola, R., Cunha, M.R., de Stigter, H., Gooday, A., Huvenne, V.,Ingels, J., Kiriakoulakis, K., Lastras, G., Masson, D., Oliveira, A., Pattenden, A.,Vanreusel, A., Van Weering, T., Vitorino, J., Witte, U., Wolff, G., 2009. Europe’sGrand Canyon: Nazare submarine canyon. Oceanography 22, 46–57.
Van Der Zee, C., Slomp, C.P., Van Raaphorst, W., 2002. Authigenic p formation andreactive p burial in sediments of the Nazare Canyon on the Iberian margin (NEAtlantic). Marine Geology 185, 379–392.
Van Weering, T.C.E., de Stigter, H.C., Balzer, W., Epping, E.H.G., Graf, G., Hall, I.R.,Helder, W., Khripounoff, A., Lohse, L., Mccave, I.N., Thomsen, L., Vangriesheim,A., 2001. Benthic dynamics and carbon fluxes on the NW European continentalmargin. Deep-Sea Research II 48, 3191–3221.
Vetter, E.W., Dayton, P.K., 1999. Organic enrichment by macrophyte detritus, andabundance patterns of megafaunal populations in submarine canyons. MarineEcology Progress Series 186, 137–148.
Viso, A.C., Marty, J.C., 1993. Fatty-acids from 28 marine microalgae. Phytochem-istry 34, 1521–1533.
Volkman, J.K., Barrett, S.M., Blackburn, S.I., Mansour, M.P., Sikes, E.L., Gelin, F.,1998. Microalgal biomarkers: a review of recent research developments.Organic Geochemistry 29, 1163–1179.
Wakeham, S.G., Lee, C., Hedges, J.I., Hernes, P.J., Peterson, M.L., 1997. Molecularindicators of diagenetic status in marine organic matter. Geochimica EtCosmochimica Acta 61, 5363–5369.
Waterson, E.J., Canuel, E.A., 2008. Sources of sedimentary organic matter in theMississippi river and adjacent Gulf of Mexico as revealed by lipid biomarkerand delta(13) c-toc analyses. Organic Geochemistry 39, 422–439.
Yamamuro, M., Kayanne, H., 1995. Rapid direct determination of organic-carbonand nitrogen in carbonate-bearing sediments with a Yanaco mt-5 CHNanalyzer. Limnology and Oceanography 40, 1001–1005.
Yunker, M.B., Belicka, L.L., Harvey, H.R., Macdonald, R.W., 2005. Tracing the inputsand fate of marine and terrigenous organic matter in Arctic Ocean sediments:a multivariate analysis of lipid biomarkers. Deep-Sea Research II 52,3478–3508.