Embed Size (px)
Citation preview

Defensins in the immunology of bacterial infectionsAlfredo Menendez and B Brett Finlay
Defensins are a component of the host response against
bacterial infections. Multiple studies suggest a linked
upregulation of b-defensins and pro-inflammatory cytokines
expression in various tissues, as well as the possibility of
mutual induction. Recent data demonstrate the importance of
nucleotide-binding oligomerization proteins for the expression
of defensins, and associate low levels of a-defensins
expression by intestinal Paneth cells with susceptibility to
Crohn’s disease of the ileum. A novel anti-toxin activity has
been identified for several a- and u-defensins, expanding the
repertoire of the antimicrobial functions of defensins. It has
been shown that bacterial proteins can inactivate the action of
defensins, and that pathogen type III secretion systems (T3SS)
manipulate defensins expression via T3SS-mediated inhibition
of the NF-kB pathway.
Addresses
Michael Smith Laboratories, The University of British Columbia, 301-
2185 East Mall, Vancouver, BC, Canada V6T 1Z4
Corresponding author: Brett Finlay, B. ([email protected])
Current Opinion in Immunology 2007, 19:385–391
This review comes from a themed issue on
Host–pathogen interactions
Edited by Louis Schofield and Ed Pearce
0952-7915/$ – see front matter
# 2007 Elsevier Ltd. All rights reserved.
DOI 10.1016/j.coi.2007.06.008
IntroductionDefensins are one of the two most common types of
antimicrobial peptides (AMPs). The term ‘defensin’ was
coined by Ganz et al. [1] and Selsted et al. [2], to refer to
three short peptides with antimicrobial activity isolated
from human neutrophil granules [1,2]. Since then, a large
number of defensins and defensin-like peptides have
been identified in many organisms. As of February 2007,
304 entries had been recorded in a defensin database
compiled by the Bioinformatics Institute of Singapore
(http://defensins.bii.a-star.edu.sg) [3]. The ubiquitous dis-
tribution of defensins and defensin-like peptides across the
biological world is testimony to their crucial role in protec-
tion against pathogens.
Defensins from vertebrates are small in size, are cationic,
and have three intra-molecular disulfide bonds, mediated
by six conserved cysteines [4�]. There are two main
subfamilies: the a- and the b-defensins, based on the
www.sciencedirect.com
cysteine pairing and the length of the peptide fragments
between the cysteines. All members of these two sub-
families whose structure has been analyzed show a
similar, distinctive ‘defensin fold’ composed of a predo-
minant three-stranded b-sheet [5–7]. A third group of
defensins, the u-defensins are only found in non-human
primates, and are structurally unrelated to the a and b
subfamilies [8,9]. A paradoxical feature of a number of
defensins is the lack of in vitro antimicrobial activity in
physiological conditions or at the observed physiological
concentrations, but in addition to direct microbial killing,
defensins perform other functions related to both the
innate and adaptive immune responses (reviewed in [4�]).
Herein, recent advances in the understanding of the
biology of defensins (mainly human defensins) are
described in the context of bacterial infections. Most of
the work reviewed has been published during the last two
years, and centers on the immunoregulatory functions of
defensins, rather than on their bactericidal properties. We
highlight new findings regarding the expression of defen-
sins as components of the overall innate immune
response, as well as their role in protection from bacterial
infections and their possible connection with suscepti-
bility to disease. In addition, several mechanisms used by
bacteria to antagonize their antimicrobial functions are
also examined.
Defensins expression can be induced inresponse to bacterial infectionDefensins are expressed in multiple tissues in the body,
most notably leukocytes and epithelial surfaces. In
humans, six a- and four b-defensins have been charac-
terized in some detail. The expression and release of
defensins is differentially regulated, depending on the
defensin molecule, the cell type, and the microenviron-
mental stimuli [10–14]. Detection of bacteria may trigger
the induction and release of defensins, mediated either
by the engagement of toll-like receptors (TLR) by their
cognate bacterial pathogen-associated molecular patterns
(PAMP), or by any of several TLR-independent path-
ways (reviewed in [15�]). Bactericidal concentrations of a-
defensins are found in the phagocytic vacuoles of human
neutrophils after being delivered from cytoplasmic gran-
ules, and in the crypts of the small intestine following
their secretion by Paneth cells. There is no indication
thus far that the expression of the myeloid a-defensins
human neutrophil protein (HNP)-1-3 vary in any signifi-
cant way [4�]. Moreover, enteric mouse a-defensins
(cryptdins) levels are generally invariant ([16] and refer-
ences therein), although the normal, constitutive expres-
sion of cryptdin-4 and cryptdin-10 requires signaling
Current Opinion in Immunology 2007, 19:385–391

386 Host–pathogen interactions
mediated by nucleotide-binding oligomerization domain-
2 (Nod2), as has been demonstrated in Nod2-deficient
mice [17��], indicating some level of regulation mediated
by this PAMP receptor.
The human b-defensin 2 (hBD-2) is a well-known
example of a defensin induced by bacterial products,
and by pro-inflammatory cytokines normally produced
in response to infection [18,19,20�]. Human vaginal
epithelial cells respond to bacterial LPS and peptido-
glycan by inducing the expression of TNF-a and hBD-
2 [21], as do pulmonary epithelial cells upon infection
with live Mycobacterium bovis [22]. Infection of the
intestinal epithelial cell lines Caco-2 and HT-29 with
C. jejuni induces the expression of hBD-2 [23]. Sig-
naling through the IL-1 receptor (activated either by
IL-1a or IL-1b), or treatment with TNF-a, induce the
synthesis of hBD-2 by keratinocytes [24], epidermal
cultures [11], and pulmonary [18,25], uterine [26], gin-
gival [19], intestinal [23] and middle ear epithelial cells
[27]. The NF-kB pathway has been recognized as a
key component in the induction of hBD-2 expression
[18,20�,21,22], however, other studies have observed
induction mediated by the mitogen-activated protein
kinase (MAPK) pathways [19,27]. Thus, increased
expression of hBD-2 in epithelial cells is associated
with the pro-inflammatory response, a concept sup-
ported by the finding that the anti-inflammatory cyto-
kines IL-10 and IL-13 downregulate the synthesis of
hBD-2 in atopic dermatitis [28].
Expression of hBD-3 is induced in gastric epithelial cells
infected with Helicobacter pylori [29�], as well as in
primary human keratinocytes infected with Staphylococ-cus aureus [30] and in epidermal cultures treated with
supernatants from mononuclear blood leukocytes stimu-
lated with LPS or peptidoglycan [11]. The NF-kB path-
way does not appear to play such a prominent role in
promoting the expression of hBD-3, but instead, induc-
tion seems to be mainly mediated by the MAPK/p38 [30]
and the EGFR/ERK pathways [11,29�]. The regulation
of hBD-1 and hBD-4 synthesis is less well understood;
hBD-1 has been largely considered a constitutively
expressed defensin, but a recent observation of a rela-
tively low degree of induction in epidermal cultures,
suggests some sort of regulation by LPS and supernatants
from stimulated mononuclear blood leukocytes [11].
Moreover, hBD-1 upregulation by albumin, arginine
and isoleucine has been observed in human colon tumor
cells, by a pathway involving c-myc [31], and induction
by glucose has been reported in human renal cells [32].
Similar to hBD-2 and -3, hBD-4 expression is induced by
Ca2+, TNFa, IL-1b, phorbol 12-myristate 13-acetate and
heat-killed P. aeruginosa in primary keratinocytes [10].
The above indicates a general trend of b-defensin induc-
tion in response to bacteria, or the inflammatory response
they induce.
Current Opinion in Immunology 2007, 19:385–391
Defensins are tough; they can handlebacteriaThe role of defensins in bacterial infections is not limited
to bacterial killing; in fact, the immunomodulatory reper-
toire of defensins is likely as important as their killing
capabilities. It has been demonstrated that physiological
concentrations of b-defensins can increase the expression
of pro-inflammatory cytokines and chemokines by pur-
ified human peripheral blood mononuclear cells (PBMC)
[33�] and by human primary keratinocytes [34], in the
absence of infection or stimulation with bacterial pro-
ducts. In PBMC, hBD-1 or hBD-2 but not hBD-3, were
strong cytokine inducers [33�], whereas in keratinocytes
hBD-2, -3 and -4 but not hBD-1 upregulated most of the
genes tested [34]. Interestingly, hBD-2, -3 and -4 but not
hBD-1 increased phosphorylation of EGFR, STAT1 and
STAT3, and keratinocyte proliferation and migration
[34]. These results strongly suggest that b-defensins
are critical to expand the initial cytokine response of
inflammation and wound healing at sites of injury or
infection (Figure 1). Another recent report showed that
hBD-3 and hBD-4 induced chemotaxis and degranulation
of rat and human mast cells, as well as phosphorylation of
MAPK/p38 and ERK1/2 [35].
Unambiguous proof of specific functions of a particular
defensin in vivo are difficult to obtain, mainly because
of the overlap of antibacterial mechanisms operating
simultaneously in most tissues, and the redundancy of
defensins. Nonetheless, altered levels of defensins at sites
of active infection and inflammation have generally been
interpreted as an indication of their role in antagonizing
bacterial infections in vivo [36–40]. Reduced expression of
Paneth cell a-defensins HD-5 and HD-6 has been
observed in ileal Crohn’s disease [37��], while low hBD-
2 gene copy number resulting from genetic polymorphism
at the defensin locus in chromosome 8, is associated with
Crohn’s disease of the colon [41�]. Although a causal
relation was not established in either of these studies, they
support the interpretation that decreased expression of
defensins affects the innate immunity of the bowel and
the control of the microbiota, thereby contributing to the
establishment and/or maintenance of the chronic inflam-
mation characteristic of Crohn’s disease.
Increasing the levels of porcine (p) BD-1 in the lungs of
piglets (by administration of synthetic pBD-1) provided
protection from challenge with B. pertussis [42]. Over-
expression of rat BD-2 in the lungs of rats has conferred
protection against P. aeruginosa pneumonia and sepsis-
induced lung injury [43]. However, other studies using
BD-1-deficient mice have found only a modest role of
BD-1 in protection, in the form of delayed clearance of H.influenzae from the lungs of knockout mice [44], and
increased numbers of Staphylococcus sp. in the bladder,
without any difference in the clearance of S. aureus from
the lungs of infected animals [45].
www.sciencedirect.com

Defensins in the immunology of bacterial infections Menendez and Finlay 387
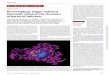
Figure 1
Amplification of b-defensin expression may occur through local autocrine/paracrine loops. At sites of infection, bacteria are detected by
phagocytes (A) and induce the synthesis and secretion of pro-inflammatory mediators (e.g. IL-1 and TNF-a). Infected cells (e.g. a keratinocyte, B)
also detect bacteria through interactions with LPS and peptidoglycan (PG), resulting in synthesis of defensins and more pro-inflammatory
mediators, which can act in an autocrine fashion (dashed black arrows), or in a paracrine fashion (solid black arrows) on neighboring cells (C),
and promote further induction of defensins and cytokines.
Experiments with defensin-deficient animals have
suggested that a-defensins are involved in protection
from enteric bacterial infections in vivo. Loss of Paneth
cell defensins due to a knockout of the processing enzyme
matrilysin (MMP-7) gene improved the survival of
luminal E. coli, and rendered the deficient mice more
susceptible to infection and killing by S. typhimurium [46].
MMP-7-deficient mice also had higher bacterial burden
than controls in the small intestine but not in spleen, lung
and kidneys, in a model of vaginal infection with C.trachomatis, suggesting again that the loss of Paneth cell
defensins has a severe influence on the intestinal innate
immunity [47]. The strongest evidence yet of a defensin-
mediated protection in vivo has been provided by Salz-
man et al. [48], who found that mice transgenic for the
enteric HD-5 were markedly resistant to oral challenge
with S. typhimurium [48].
Recently, a novel toxin inhibition function has been
described for the human a-defensins HNP-1-3
www.sciencedirect.com
[49��,50��], and for retrocyclins (RC), a group of synthetic
u-defensins [51��]. Lethal toxin (LT) from B. anthracis is a
virulence factor with zinc-dependent metalloproteinase
activity, which targets selected MAPK kinases of host
cells. HNP-1-3 are capable of neutralizing LT and protect
from toxin-induced death in vitro and in vivo [49��]; HNP-
1-3 also inhibit diphtheria toxin (DT) and Pseudomonasendotoxin A (ETA), two bacterial toxins with mono-
ADP-ribosyltransferase activity, and protect cells from
DT-induced or ETA-induced cell death [50��]. More-
over, Wang et al. demonstrated that RC-1, -2 and -3, as
well as HNP-1 not only killed B. anthracis bacilli, but also
inhibited the enzymatic activity of LT [51��].
But bacteria are also tough, andthey retaliateDefensins, and in general AMPs, are a pervasive threat to
microbes. Thus, not surprisingly, a significant number of
pathogen-encoded virulence mechanisms function to
antagonize the action and effects of AMPs (see Table 1
Current Opinion in Immunology 2007, 19:385–391

388 Host–pathogen interactions
Table 1
Recently described examples of bacterial antagonism of the antimicrobial actions of defensins
Bacterium Virulence factor Mechanism Reference
Streptococcus pyogenes Streptolysin O Degranulation of neutrophils [70]
Streptococcus pyogenes Streptococal inhibitor of complement (SIC) Inhibition of HNP-1, hBD-1, hBD-2, hBD-3 [52,53]
Streptococcus pyogenes Distantly related to SIC (DRS) Inhibition of HNP-1, hBD-2, hBD-3 [53]
Streptococcus agalactiae Penicillin-binding Protein 1a (PBP1a) Not determined [58]
Salmonella typhimurium Alternative sigma factor sE Induction of alternative electron transport pathway(s) [71]
Listeria monocytogenes VirR (part of the two components
system VirR/VirS)
Positive regulation of mprF and the dlt operon
(involved in cell surface charge modification)
[72]
Listeria monocytogenesa Peptidoglycan N-deacetylase (PgdA) N-deacetylation of peptidoglycan [59]
Bordetella bronchiseptica T3SS Inhibition of the NFk-B pathway [60]
a Effect on defensin expression not experimentally demonstrated, but very likely.
for recently reported examples). Several highly virulent
strains of Streptococcus pyogenes (group A streptococci)
secrete proteins that have a direct effect on the host
immune function. For example, the streptococcal inhibitor
of complement (SIC) binds to and inhibits the antimicro-
bial activities of HNP-1, hBD-1, -2 and -3 in vitro [52,53].
Mutants of SIC have shown significant reduction of throat
colonization in a mouse infection model, suggesting an
important role of SIC on S. pyogenes virulence [54]. Another
S. pyogenes protein, DRS (Distantly Related to SIC) also
inactivates HNP-1, hBD-2 and hBD-3 [53]. In addition, S.pyogenes secretes streptolysin O (SLO), a cholesterol-de-
pendent cytolytic toxin that promotes degranulation of
neutrophils and the release of their a-defensins [55].
Alpha-defensins stored in the azurophil granules of neu-
trophils are normally released into phagosomes, where they
(and other AMPs) reach high concentrations that are effec-
tively antimicrobial. Thus, the SLO-induced premature
release of a-defensins may function as a mechanism to
disarm the neutrophils from a safe distance, much the same
as the staphylokinase of S. aureus [56]. Other cholesterol-
dependent cytolysins are expressed by pathogenic Bacillus,Clostridium and Listeria [57]. It will be interesting to deter-
mine whether their toxins have a similar neutrophil degra-
nulation function.
S. agalactiae (group B streptococci) produces a surface-
associated penicillin-binding protein (PBP1a), which is
essential for bacterial virulence. PBP1a promotes resist-
ance of S. agalactiae to HNP-1 and other AMPs [58] by a
yet unknown mechanism that does not involve cell
envelope charge modification such as those promoted
by dltABCD and mprF . Recently, Boneca et al. demon-
strated that peptidoglycan N-deacetylation functions as
an important virulence mechanism in L. monocytogenesinfections [59�]. This type of cell surface modification
makes Listeria more resistant to killing by lysozyme, and
impairs the detection of peptidoglycan by Nod1 and
Nod2. Since Nod proteins are required for expression
of some cryptdins in the mouse ileum [17��] and for
induction of human b-defensin synthesis [20�,29�], it is
conceivable that peptidoglycan N-deacetylation is also
involved in preventing defensin expression.
Current Opinion in Immunology 2007, 19:385–391
T3SS effector-mediated inhibition of the MAPK and NFk-
B pathways is becoming a recognized bacterial strategy
to interfere with the induction of the innate immune
response. A connection between T3SS effectors and
antagonism of defensin function has been recently uncov-
ered [60��]. A T3SS from Bordetella bronchiseptica has been
implicated in inhibiting the expression of the bovine b-
defensin Tracheal Antimicrobial Peptide (TAP), in
primary bovine tracheal epithelial cells [60��]. This down-
regulation occurred through the inhibition of NFk-B-
mediated induction of TAP expression, at some point
along the pathway that leads to the nuclear translocation
of NFk-B [60��]. From these results, it can be predicted
that T3SS effectors recently found to inactivate NFk-B,
such as VP1686 (V. parahaemolyticus), YopJ (Yersinia), AopP
(Aeromonas) and OspF (Shiguella) [61��–65��], may also
mediate the inhibition of defensin synthesis.
ConclusionsAn increasing number of bacterial virulence factors are
being associated with antagonizing the effects of AMPs; it
is also apparent that some mechanisms of resistance are
shared by pathogens from different taxonomical groups. A
recent report showing acquisition of adaptive resistance to
hBD-1-4 by Porphyromonas gingivalis [66] is especially
disturbing, since such a mechanism could have a pro-
found impact in microbial persistence. The transcription
factor NF-kB is an essential component of the AMP
induction pathways, as well as for many other aspects
of the inflammatory response; therefore, the involvement
of a T3SS in resistance to AMPs via the inhibition of NF-
kB function is not unexpected; in fact, we predict that
more of such examples will be reported shortly.
The findings that hBD-2, -3 and -4 can activate the
MAPK/p38 and EGFR pathways [34,35] suggests the
existence of positive feedback regulation in the control
of defensin expression, since these pathways can also be
involved in the induction of hBD-2 and hBD-3 in various
cell types. In addition, pro-inflammatory cytokines can
induce the synthesis of defensins whereas, defensins in
turn may upregulate the synthesis of cytokines and che-
mokines, and influence the expression of other AMPs
www.sciencedirect.com

Defensins in the immunology of bacterial infections Menendez and Finlay 389
[67]. Such regulatory loops would act in an autocrine or
paracrine manner, and most likely, involve the participa-
tion of several cell types (Figure 1). Little is known about
how defensin expression is turned down and their homeo-
stasis restored, although it may be anticipated that some
of the mechanisms turning off other elements of the
inflammatory response may act directly or indirectly to
downregulate the expression of defensins. This is an
important area to investigate since defensins can reach
cytotoxic levels in vivo [68], and sustained overexpression
of mBD-6 induces muscle degeneration in a transgenic
mouse model [69]. In addition, studies of possible con-
nections between altered defensin expression and chronic
inflammatory conditions will be interesting. In summary,
the expression and functions of defensins are intimately
associated with other aspects of the innate immunity; and
in many respects, defensins emulate the behavior of other
elements of the inflammatory response. As we learn more
about their functions and how their expression is regulated,
it becomes apparent that (at least in vertebrates) defensins
should be considered an integral component of the host
inflammatory response to bacterial infection.
AcknowledgementsThis work was supported by operating grants to B.B.F. from theCanadian Institutes of Health Research (CIHR), the Howard HughesMedical Institute (HHMI) and the Foundation for NIH, as part ofthe Bill and Melinda Gates Grand Challenge program. AM is a recipientof postdoctoral fellowships from the Michael Smith Foundation forHealth Research (MSFHR), and the Natural Sciences and EngineeringResearch Council of Canada. B.B.F. is a CIHR Distinguished Investigator,an HHMI International Research Scholar and the University of BritishColumbia Peter Wall Distinguished Professor. We would like to thankthank M. Chow and members of the Finlay laboratory for critical readingof the manuscript.
References and recommended readingPapers of particular interest, published within the annual period ofreview, have been highlighted as:
� of special interest
�� of outstanding interest
1. Ganz T, Selsted ME, Szklarek D, Harwig SS, Daher K, Bainton DF,Lehrer RI: Defensins. Natural peptide antibiotics of humanneutrophils. J Clin Invest 1985, 76:1427-1435.
2. Selsted ME, Harwig SS, Ganz T, Schilling JW, Lehrer RI: Primarystructures of three human neutrophil defensins. J Clin Invest1985, 76:1436-1439.
3. Seebah S, Suresh A, Zhuo S, Choong YH, Chua H, Chuon D,Beuerman R, Verma C: Defensins knowledgebase: a manuallycurated database and information source focused on thedefensins family of antimicrobial peptides. Nucleic Acids Res2007, 35:D265-D268.
4.�
Bowdish DME, Davidson DJ, Hancock REW: Immunomodulatoryproperties of defensins and cathelicidins. Curr Top MicrobiolImmunol 2006, 306:27-66.
An excellent review on the topic.
5. Hill CP, Yee J, Selsted ME, Eisenberg D: Crystal structure ofdefensin HNP-3, an amphiphilic dimer: mechanisms ofmembrane permeabilization. Science 1991, 251:1481-1485.
6. Zimmermann GR, Legault P, Selsted ME, Pardi A: Solutionstructure of bovine neutrophil beta-defensin-12: the peptidefold of the beta-defensins is identical to that of the classicaldefensins. Biochemistry 1995, 34:13663-13671.
www.sciencedirect.com
7. Szyk A, Wu Z, Tucker K, Yang D, Lu W, Lubkowski J: Crystalstructures of human alpha-defensins HNP4, HD5 and HD6.Protein Sci 2006, 15:2749-2760.
8. Tang YQ, Yuan J, Osapay G, Osapay K, Tran D, Miller CJ,Ouellette AJ, Selsted ME: A cyclic antimicrobial peptideproduced in primate leukocytes by the ligation of twotruncated alpha-defensins. Science 1999, 286:498-502.
9. Selsted ME: Theta-defensins: cyclic antimicrobial peptidesproduced by binary ligation of truncated alpha-defensins.Curr Protein Pept Sci 2004, 5:365-371.
10. Harder J, Meyer-Hoffert U, Wehkamp K, Schwichtenberg L,Schroder JM: Differential gene induction of human beta-defensins (hBD-1, -2, -3 and -4) in keratinocytes is inhibited byretinoic acid. J Invest Dermatol 2004, 123:522-529.
11. Sorensen OE, Thapa DR, Rosenthal A, Liu L, Roberts AA, Ganz T:Differential regulation of beta-defensin expression in humanskin by microbial stimuli. J Immunol 2005, 174:4870-4879.
12. Schauber J, Dorschner RA, Yamasaki K, Brouha B, Gallo RL:Control of the innate epithelial antimicrobial response is cell-type specific and dependent on relevant microenvironmentalstimuli. Immunology 2006, 118:509-519.
13. Veldhuizen EJ, Hendriks HG, Hogenkamp A, van Dijk A,Gaastra W, Tooten PC, Haagsman HP: Differential regulation ofporcine beta-defensins 1 and 2 upon Salmonella infection inthe intestinal epithelial cell line IPI-2I. Vet ImmunolImmunopathol 2006, 114:94-102.
14. Veerayutthwilai O, Byers MR, Pham TT, Darveau RP, Dale BA:Differential regulation of immune responses by odontoblasts.Oral Microbiol Immunol 2007, 22:5-13.
15.�
Froy O: Regulation of mammalian defensin expression byToll-like receptor-dependent and independent signallingpathways. Cell Microbiol 2005, 7:1387-1397.
An excellent review on the topic.
16. Tanabe H, Ayabe T, Bainbridge B, Guina T, Ernst RK, Darveau RP,Miller SI, Ouellette AJ: Mouse paneth cell secretory responsesto cell surface glycolipids of virulent and attenuatedpathogenic bacteria. Infect Immun 2005, 73:2312-2320.
17.��
Kobayashi KS, Chamaillard M, Ogura Y, Henegariu O, Inohara N,Nunez G, Flavell RA: Nod2-dependent regulation of innate andadaptive immunity in the intestinal tract. Science 2005,307:731-734.
Using a knockout mouse model, this paper shows that Nod2 deficientmice are more susceptible than wild type mice to intragastric infectionwith Listeria (but not to intravenous or intraperitoneal infections), pointingto an important role for Nod2 in intestinal defenses. It also shows thatNod2 is necessary for the expression of cryptdins 4 and 10 in the mouseileum. Together with Refs. [20�,29�], this establishes a role for Nodproteins in the expression of defensins.
18. Kao CY, Chen Y, Thai P, Wachi S, Huang F, Kim C, Harper RW,Wu R: IL-17 markedly upregulates beta-defensin-2 expressionin human airway epithelium via JAK and NF-kappaB signalingpathways. J Immunol 2004, 173:3482-3491.
19. Ouhara K, Komatsuzawa H, Shiba H, Uchida Y, Kawai T,Sayama K, Hashimoto K, Taubman MA, Kurihara H, Sugai M:Actinobacillus actinomycetemcomitans outer membraneprotein 100 triggers innate immunity and production of beta-defensin and the 18-kilodalton cationic antimicrobial proteinthrough the fibronectin-integrin pathway in human gingivalepithelial cells. Infect Immun 2006, 74:5211-5220.
20.�
Voss E, Wehkamp J, Wehkamp K, Stange EF, Schroder JM,Harder J: NOD2/CARD15 mediates induction of theantimicrobial peptide human beta-defensin-2. J Biol Chem2006, 281:2005-2011.
See annotation for ref. [17��].
21. Pivarcsi A, Nagy I, Koreck A, Kis K, Kenderessy-Szabo A, Szell M,Dobozy A, Kemeny L: Microbial compounds induce theexpression of pro-inflammatory cytokines, chemokines andhuman beta-defensin-2 in vaginal epithelial cells.Microbes Infect 2005, 7:1117-1127.
22. Mendez-Samperio P, Miranda E, Trejo A: Mycobacteriumbovis Bacillus Calmette-Guerin (BCG) stimulates human
Current Opinion in Immunology 2007, 19:385–391

390 Host–pathogen interactions
beta-defensin-2 gene transcription in human epithelial cells.Cell Immunol 2006, 239:61-66.
23. Zilbauer M, Dorrell N, Boughan PK, Harris A, Wren BW, Klein NJ,Bajaj-Elliott M: Intestinal innate immunity to Campylobacterjejuni results in induction of bactericidal human beta-defensins 2 and 3. Infect Immun 2005, 73:7281-7289.
24. Liu L, Roberts AA, Ganz T: By IL-1 signaling, monocyte-derivedcells dramatically enhance the epidermal antimicrobialresponse to lipopolysaccharide. J Immunol 2003,170:575-580.
25. Tsutsumi-Ishii Y, Nagaoka I: Modulation of human beta-defensin-2 transcription in pulmonary epithelial cells bylipopolysaccharide-stimulated mononuclear phagocytes viaproinflammatory cytokine production. J Immunol 2003,170:4226-4236.
26. Pioli PA, Weaver LK, Schaefer TM, Wright JA, Wira CR, Guyre PM:Lipopolysaccharide-induced IL-1 beta production by humanuterine macrophages upregulates uterine epithelial cellexpression of human beta-defensin 2. J Immunol 2006,176:6647-6655.
27. Moon SK, Lee HY, Pan H, Takeshita T, Park R, Cha K, Andalibi A,Lim DJ: Synergistic effect of interleukin 1 alpha on nontypeableHaemophilus influenzae-induced upregulation of human beta-defensin 2 in middle ear epithelial cells. BMC Infect Dis 2006,6:12.
28. Howell MD, Novak N, Bieber T, Pastore S, Girolomoni G,Boguniewicz M, Streib J, Wong C, Gallo RL, Leung DY:Interleukin-10 downregulates anti-microbial peptideexpression in atopic dermatitis. J Invest Dermatol 2005,125:738-745.
29.�
Boughan PK, Argent RH, Body-Malapel M, Park JH, Ewings KE,Bowie AG, Ong SJ, Cook SJ, Sorensen OE, Manzo BA et al.:Nucleotide-binding oligomerization domain-1 and epidermalgrowth factor receptor: critical regulators of beta-defensinsduring Helicobacter pylori infection. J Biol Chem 2006,281:11637-11648.
See annotation for Ref. [17��].
30. Menzies BE, Kenoyer A: Signal transduction and nuclearresponses in Staphylococcus aureus-induced expression ofhuman beta-defensin 3 in skin keratinocytes. Infect Immun2006, 74:6847-6854.
31. Sherman H, Chapnik N, Froy O: Albumin and amino acidsupregulate the expression of human beta-defensin 1.Mol Immunol 2006, 43:1617-1623.
32. Malik AN, Al-Kafaji G: Glucose regulation of beta-defensin-1mRNA in human renal cells. Biochem Biophys Res Commun2007, 353:318-323.
33.�
Boniotto M, Jordan WJ, Eskdale J, Tossi A, Antcheva N,Crovella S, Connell ND, Gallagher G: Human beta-defensin 2induces a vigorous cytokine response in peripheral bloodmononuclear cells. Antimicrob Agents Chemother 2006,50:1433-1441.
This paper provides a detailed set of data showing the cytokine synth-esis response of PBMC after exposure to hBD-1, -2 and -3. It alsodelineates differences in the cytokine expression patterns induced bythese three defensins. These are interesting findings that may alsocontribute to explain the diversity of the defensins gene pool withinhumans and mice.
34. Niyonsaba F, Ushio H, Nakano N, Ng W, Sayama K, Hashimoto K,Nagaoka I, Okumura K, Ogawa H: Antimicrobial peptides humanbeta-defensins stimulate epidermal keratinocyte migration,proliferation and production of proinflammatory cytokinesand chemokines. J Invest Dermatol 2007, 127:594-604.
35. Chen X, Niyonsaba F, Ushio H, Hara M, Yokoi H, Matsumoto K,Saito H, Nagaoka I, Ikeda S, Okumura K et al.: Antimicrobialpeptides human beta-defensin (hBD)-3 and hBD-4 activatemast cells and increase skin vascular permeability.Eur J Immunol 2007, 37:434-444.
36. Harada K, Ohba K, Ozaki S, Isse K, Hirayama T, Wada A,Nakanuma Y: Peptide antibiotic human beta-defensin-1 and -2contribute to antimicrobial defense of the intrahepatic biliarytree. Hepatology 2004, 40:925-932.
Current Opinion in Immunology 2007, 19:385–391
37.��
Wehkamp J, Salzman NH, Porter E, Nuding S, Weichenthal M,Petras RE, Shen B, Schaeffeler E, Schwab M, Linzmeier R et al.:Reduced Paneth cell alpha-defensins in ileal Crohn’s disease.Proc Natl Acad Sci U S A 2005, 102:18129-18134.
A clear association between Crohn’s disease of the ileum and a specificdecrease of defensins synthesis had not been previously demonstrated.Here, the levels of HD-5 and HD-6 in intestinal specimens from diseasedhuman subjects were analyzed and found to be lower than in non-diseased controls. The reduction on HD-5 and HD-6 was further exa-cerbated in some specimens also harboring mutations in the NOD2 gene.So far the clearest indication of defensins alteration in Crohn’s. Alsodocuments some fundamental differences between Crohn’s disease ofthe ileum and the colon (see also annotation to Ref. [41]).
38. Rivas-Santiago B, Sada E, Tsutsumi V, Aguilar-Leon D,Contreras JL, Hernandez-Pando R: beta-Defensin geneexpression during the course of experimental tuberculosisinfection. J Infect Dis 2006, 194:697-701.
39. Valore EV, Wiley DJ, Ganz T: Reversible deficiency ofantimicrobial polypeptides in bacterial vaginosis. Infect Immun2006, 74:5693-5702.
40. Yanagi S, Ashitani J, Imai K, Kyoraku Y, Sano A, Matsumoto N,Nakazato M: Significance of human beta-defensins in theepithelial lining fluid of patients with chronic lower respiratorytract infections. Clin Microbiol Infect 2007, 13:63-69.
41.�
Fellermann K, Stange DE, Schaeffeler E, Schmalzl H, Wehkamp J,Bevins CL, Reinisch W, Teml A, Schwab M, Lichter P et al.: Achromosome 8 gene-cluster polymorphism with low humanbeta-defensin 2 gene copy number predisposes to Crohndisease of the colon. Am J Hum Genet 2006, 79:439-448.
A genetic explanation to the hBD-2 deficiency observed in colonicCrohn’s disease.
42. Elahi S, Buchanan RM, Attah-Poku S, Townsend HG, Babiuk LA,Gerdts V: The host defense peptide beta-defensin 1 confersprotection against Bordetella pertussis in newborn piglets.Infect Immun 2006, 74:2338-2352.
43. Shu Q, Shi Z, Zhao Z, Chen Z, Yao H, Chen Q, Hoeft A, Stuber F,Fang X: Protection against Pseudomonas aeruginosapneumonia and sepsis-induced lung injury by overexpressionof beta-defensin-2 in rats. Shock 2006, 26:365-371.
44. Moser C, Weiner DJ, Lysenko E, Bals R, Weiser JN, Wilson JM:beta-Defensin 1 contributes to pulmonary innate immunity inmice. Infect Immun 2002, 70:3068-3072.
45. Morrison G, Kilanowski F, Davidson D, Dorin J: Characterizationof the mouse beta defensin 1, Defb1, mutant mouse model.Infect Immun 2002, 70:3053-3060.
46. Wilson CL, Ouellette AJ, Satchell DP, Ayabe T, Lopez-Boado YS,Stratman JL, Hultgren SJ, Matrisian LM, Parks WC: Regulation ofintestinal alpha-defensin activation by the metalloproteinasematrilysin in innate host defense. Science 1999,286:113-117.
47. Pal S, Schmidt AP, Peterson EM, Wilson CL, de la Maza LM: Roleof matrix metalloproteinase-7 in the modulation of aChlamydia trachomatis infection. Immunology 2005,117:213-219.
48. Salzman NH, Ghosh D, Huttner KM, Paterson Y, Bevins CL:Protection against enteric salmonellosis in transgenic miceexpressing a human intestinal defensin. Nature 2003,422:522-526.
49.��
Kim C, Gajendran N, Mittrucker HW, Weiwad M, Song YH,Hurwitz R, Wilmanns M, Fischer G, Kaufmann SH: Human alpha-defensins neutralize anthrax lethal toxin and protect againstits fatal consequences. Proc Natl Acad Sci U S A 2005,102:4830-4835.
In this series of excellent papers (Refs. [49��–51��]), a novel biologicalfunction of a- and u-defensins is characterized. These defensins showedefficient inhibition of the enzymatic activity of two different types ofbacterial toxins, and by virtue of it, protection from the toxins lethaleffects, in vitro and in vivo. Breakthrough papers that expand the knownantimicrobial repertoire of defensins.
50.��
Kim C, Slavinskaya Z, Merrill AR, Kaufmann SH: Humanalpha-defensins neutralize toxins of the mono-ADP-ribosyltransferase family. Biochem J 2006, 399:225-229.
www.sciencedirect.com

Defensins in the immunology of bacterial infections Menendez and Finlay 391
51.��
Wang W, Mulakala C, Ward SC, Jung G, Luong H, Pham D,Waring AJ, Kaznessis Y, Lu W, Bradley KA et al.: Retrocyclins killbacilli and germinating spores of Bacillus anthracis andinactivate anthrax lethal toxin. J Biol Chem 2006,281:32755-32764.
52. Binks MJ, Fernie-King BA, Seilly DJ, Lachmann PJ, Sriprakash KS:Attribution of the various inhibitory actions of thestreptococcal inhibitor of complement (SIC) to regions withinthe molecule. J Biol Chem 2005, 280:20120-20125.
53. Fernie-King BA, Seilly DJ, Lachmann PJ: Inhibition ofantimicrobial peptides by group A streptococci: SIC and DRS.Biochem Soc Trans 2006, 34:273-275.
54. Lukomski S, Hoe NP, Abdi I, Rurangirwa J, Kordari P, Liu M,Dou SJ, Adams GG, Musser JM: Nonpolar inactivation of thehypervariable streptococcal inhibitor of complement gene(sic) in serotype M1 Streptococcus pyogenes significantlydecreases mouse mucosal colonization. Infect Immun 2000,68:535-542.
55. Nilsson M, Sorensen OE, Morgelin M, Weineisen M, Sjobring U,Herwald H: Activation of human polymorphonuclearneutrophils by streptolysin O from Streptococcus pyogenesleads to the release of proinflammatory mediators. ThrombHaemost 2006, 95:982-990.
56. Jin T, Bokarewa M, Foster T, Mitchell J, Higgins J, Tarkowski A:Staphylococcus aureus resists human defensins byproduction of staphylokinase, a novel bacterial evasionmechanism. J Immunol 2004, 172:1169-1176.
57. Palmer M: The family of thiol-activated, cholesterol-bindingcytolysins. Toxicon 2001, 39:1681-1689.
58. Hamilton A, Popham DL, Carl DJ, Lauth X, Nizet V, Jones AL:Penicillin-binding protein 1a promotes resistance of group Bstreptococcus to antimicrobial peptides. Infect Immun 2006,74:6179-6187.
59.�
Boneca IG, Dussurget O, Cabanes D, Nahori MA, Sousa S,Lecuit M, Psylinakis E, Bouriotis V, Hugot JP, Giovannini M et al.: Acritical role for peptidoglycan N-deacetylation in Listeriaevasion from the host innate immune system. Proc Natl AcadSci U S A 2007, 104:997-1002.
Description of an essential virulence function in Listeria, mediated by abacterial peptidoglycan modification that confers the ability to resistkilling by lysozyme, and escape detection by Nod proteins. Even thougha connection with the expression or functions of defensins was not tested,it is likely to exist since there is evidence that Nod proteins may regulatethe expression of defensins (see Refs. [17��,20�,29�]).
60.��
Legarda D, Klein-Patel ME, Yim S, Yuk MH, Diamond G:Suppression of NF-kappaB-mediated beta-defensin geneexpression in the mammalian airway by the Bordetella type IIIsecretion system. Cell Microbiol 2005, 7:489-497.
This is the only report (in a surprisingly unexplored area) that associatesthe presence of a functional T3SS with a decreased ability to producedefensins (or any other AMPs, for that matter). This suggests thateffectors translocated to the host cell can block the inducible AMPscomponent of the host response, a very attractive virulence trait, whichwill be probably found on many pathogens. This study brings togetherthe facts that NFk-B is a key player in the expression of some AMPs,and the inhibition of this pathway by T3SS effectors. We are onlybeginning to understand how these effectors do it (see Refs. [61��–65��]).
www.sciencedirect.com
61.��
Bhattacharjee RN, Park KS, Kumagai Y, Okada K, Yamamoto M,Uematsu S, Matsui K, Kumar H, Kawai T, Iida T et al.: VP1686, aVibrio type III secretion protein, induces toll-like receptor-independent apoptosis in macrophage through NF-kappaBinhibition. J Biol Chem 2006, 281:36897-36904.
The mechanism(s) by which T3SS effectors inactivate the NFk-B andMAPK pathways were not characterized, until now. As is demonstrated inthis series of papers (Refs. [61��–65��]), bacteria have evolved severalways to do it. The end point is the suppression of the pro-inflammatorygene expression, whether is achieved by inactivation of MAPKs throughacetylation or dephosphorylation, or direct interaction of the T3SS effec-tor with NFk-B. In any case, these are novel functions described for T3SSeffectors, which likely are a common virulence trait (see also annotationfor Ref. [60��]).
62.��
Mukherjee S, Keitany G, Li Y, Wang Y, Ball HL, Goldsmith EJ,Orth K: Yersinia YopJ acetylates and inhibits kinase activationby blocking phosphorylation. Science 2006, 312:1211-1214.
63.��
Mittal R, Peak-Chew SY, McMahon HT: Acetylation of MEK2 and Ikappa B kinase (IKK) activation loop residues by YopJ inhibitssignalling. Proc Natl Acad Sci U S A 2006, 103:18574-18579.
64.��
Fehr D, Casanova C, Liverman A, Blazkova H, Orth K,Dobbelaere D, Frey J, Burr SE: AopP, a type III effector protein ofAeromonas salmonicida, inhibits the NF-kappaB signallingpathway. Microbiology 2006, 152:2809-2818.
65.��
Arbibe L, Kim DW, Batsche E, Pedron T, Mateescu B, Muchardt C,Parsot C, Sansonetti PJ: An injected bacterial effector targetschromatin access for transcription factor NF-kappaB to altertranscription of host genes involved in immune responses. NatImmunol 2007, 8:47-56.
66. Shelburne CE, Coulter WA, Olguin D, Lantz MS, Lopatin DE:Induction of beta-defensin resistance in the oral anaerobePorphyromonas gingivalis. Antimicrob Agents Chemother 2005,49:183-187.
67. Stroinigg N, Srivastava MD: Modulation of toll-like receptor 7and LL-37 expression in colon and breast epithelial cells byhuman beta-defensin-2. Allergy Asthma Proc 2005, 26:299-309.
68. Wencker M, Brantly ML: Cytotoxic concentrations of alpha-defensins in the lungs of individuals with alpha(1)-antitrypsindeficiency and moderate to severe lung disease. Cytokine2005, 32:1-6.
69. Yamaguchi Y, Nagase T, Tomita T, Nakamura K, Fukuhara S,Amano T, Yamamoto H, Ide Y, Suzuki M, Teramoto S et al.: {beta}-defensin overexpression induces progressive muscledegeneration in mice. Am J Physiol Cell Physiol 2007,292:C2141-C2149.
70. Kraus D, Peschel A: Molecular mechanisms of bacterialresistance to antimicrobial peptides. Curr Top MicrobiolImmunol 2006, 306:231-250.
71. Crouch ML, Becker LA, Bang IS, Tanabe H, Ouellette AJ, Fang FC:The alternative sigma factor sigma is required for resistanceof Salmonella enterica serovar Typhimurium to anti-microbialpeptides. Mol Microbiol 2005, 56:789-799.
72. Mandin P, Fsihi H, Dussurget O, Vergassola M, Milohanic E,Toledo-Arana A, Lasa I, Johansson J, Cossart P: VirR, a responseregulator critical for Listeria monocytogenes virulence. MolMicrobiol 2005, 57:1367-1380.
Current Opinion in Immunology 2007, 19:385–391