Embed Size (px)
Citation preview

Dendrit ic Secret ion: A Way for the Neuron to C o m m u n i c a t e wi th the Vasculature
G. W. K r e u t z b e r g and L. T d t h *
M a x - P l a n c k - I n s t i t u t fiir Psychia t r ic , Mflnchen
W e are i nves t iga t ing neurop lasmic t r a n s p o r t in t he dendr i tes of m a m m a l i a n m o t o n e u r o n s [t, 2]. W e found t h a t acetyl- chol ines terase (ACHE) is an excel lent m a r k e r subs t ance for th i s t ype of t r a n s p o r t [3]. I t s ac t iv i ty can be d e m o n s t r a t e d cy to- chemical ly a t the e lec t ron microscopic level [4]. The specif ic i ty of t he reac t ion is e s tab l i shed b y u s i ng inhib i tors of non- specific chol inesferases , e.g. e thopropazine , i so-OMPA, and b y control expe r i men t s wi th t he specific A C h E inhib i tor B W 2 8 4 C 5 1 or t he i r reversible inh ib i to r D F P . The occur- rence of diffusion a r t e fac t s du r ing f ixa t ion is exc luded b y the use of c ryos t a t sect ions of unf ixed mater ia l . In addi t ion, A C h E ac t iv i ty is d e m o n s t r a t e d l igh t microscopical ly b y two o ther t e chn iques [5, 6] and i t is found in the s ame vascu la r and neurona l loca t ions as wi th the Lewis and Shu te techni- que.
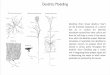
In nerve cells the enzyme first occurs in the rough endo- p lasmic r e t i cn lum (r-ER) and is t h e n t r a n s p o r t e d in to the processes of t he neurons . I n t he nuc leus of the facial nerve in ra t s and gu inea pigs, e n z y m e ac t iv i ty in the dendr i tes of the m o t o n e u r o n s can be seen associa ted wi th tubu la r -ves i cu la r s t ruc tu res . These s t ruc tu re s represen t mie ro tubn les as well as c is ternae of t he s m o o t h endop lasmic re t i cu lum. As judged f rom the locat ion of t he reac t ion product , t he enzym e ap- pears b o u n d to t he ex te rna l surface of the dendr i t ic m e m b r a n e , par t icu lar ]y the p o s t s y n a p t i c site. The reac t ion p roduc t can be found f r equen t ly in the synap t i c cleft (Fig. 1 a). U n d e r cer ta in expe r imen ta l condi t ions, e.g. 2 or 3 weeks af ter a x o t o m y of t he facial nerve ill gu inea pigs, e n z y m e ac t iv i ty also appears in the capillaries of t h e facial nuc leus [7]. E lec t ron mic roscopy ha s d e m o n s t r a t e d t h a t t he e n z y m e ac t iv i ty in t he capillaries is located in t he b a s e m e n t m e m - branes . There is no A C h E ac t iv i ty in the r - E R of the endo- thel ial cells, ind ica t ing t h a t t hese cells are no t capable of syn thes i z ing ACHE. Thus , t he e n z y m e m u s t or iginate f rom a source o ther t h a n the endothel ia l ceils. In the area u n d e r s t u d y the facial m o t o n e u r o n s are the on ly source of ACHE. Therefore, it m u s t be pos tu l a t ed t h a t the e n z y m e is re leased f rom the dendr i tes by m e a n s of a h i the r to u n k n o w n m e c h a n i s m which we could like to call " d e n d r i t i c sec re t ion" . On t h e basis of our e lectron microscopic obse rva t ions i t is ev iden t tha t , following a x o t o m y , A C h E appears ill t h e ext race l lu lar space a n d t h e n arr ives a t the b a s e m e n t m e m b r a n e s of the capillaries. The b a s e m e n t m e m b r a n e s are composed of poly- ionic g lycoprote ins and mucopolysacchar ides , which are p robab ly able to ca tch charged macromolecu les f loa t ing in the ext race l lu lar space, e.g. ACHE. Once the macromolecu les are t r apped in the b a s e m e n t m e m - b rane t h e y can be t a k e n up by microp inocy tos i s in to the endothel ia l cells. We have observed va r ious t ypes of ACHE- posi t ive p inocyto t ic vesicles on the endothe l ia l in ter face of the b a s e m e n t m e m b r a n e . All t r ans i t iona l s tages can be seen, f rom b u d s to s ta lked vesicles and p inched-off vesicles. This obse rva t ion indica tes t he capabi l i ty of the endothe l ia l cells to incorpora te mate r ia l p roduced and released by the neurons . The p h e n o m e n o n observed here in an expe r imen t a l s i t ua t i on seems also to occur unde r n o r m a l condi t ions . AChE-pos i t ive capil laries can be seen no rma l ly in cer ta in a reas of t he brain, e.g. in the cerebel lum and t he inferior olive of the r abb i t [8], in t he c ingula te region and in t he media l t h a l a m u s of ra t s [9] and in t he area s u b p o s t r e m a of t he cat [t0]. In s u m m a r y , we h a v e observed a complex t r ans loca t ion of ace ty lchol ines te rase f rom the neurona l pe r ika ryon to the dendri tes . After pas s ing the dendr i t ic m e m b r a n e , t he e n z y m e enters t he extracelIular space and reaches the b a s e m e n t m e m b r a n e s of t he capillaries, f rom where i t can be t a k e n up b y microp inocy tos i s into the endothel ia l cy top la sm. ACh E m a y no t be the on ly subs t ance m o v i n g a long th is p a t h w a y , as it is conceivable t h a t i t m a y also be used by o ther specific subs t ances which could ac t as signals. Thus , i t seems likely t h a t we have d e m o n s t r a t e d a p a t h w a y f rom the nerve cell to the endothel ia l cells t h a t enables the neu ron to c o m m u n i c a t e wi th the vascu la tu re .
Fig. t . a Two axon t e rmina l s m a k i n g synap t i c con tac t s on a dendr i t e (D). A C h E ac t i v i t y is d e m o n s t r a t e d by the electron- dense reac t ion p roduc t in tubu lo-c i s te rna l s t ruc tu re s inside t he dendr i t e and in the synap t i c cleft. 43 000 • b Capi l lary in t he facial nuc leus of gu inea pig. 2-3 weeks af ter facial ne rve t r ansec t ion t he b a s e m e n t m e m b r a n e s show increas ing A C h E ac t iv i ty , d e m o n s t r a t e d here by t he black reac t ion product . No ac t iv i ty is seen in t he endop lasmic r e t i cu lum of the endothel ia l cells, sugges t i ng a non -vascu l a r origin of the enzyme. Note A C h E con ta in ing p inocy to t i c vesicles (arrow heads) . 12000 •
Received N o v e m b e r 7, 1973
* On leave of absence f rom the Ana tomica l I n s t i t u t e of t h e NIedicaI Univers i ty , Szeged, H u n g a r y .
t . Schuber t , P., Lux , H. D., Kreu tzberg , G. W. : A c t s Neuro- pa th . (Berl.) Suppl. 5, t79 ( t97t)
2. Schuber t , P., Kreu tzberg , G. W., Lux , H. D.: Bra in Res. 47, 331 (t972)
3. Kreu tzberg , G. W., et al.: ibid. 62, 399 (1973) 4. Lewis, P. R., Shute , C. C.: J. Microsc. 89, t g l (t969) 5- Gomori , G.: Microscopic H i s tochemis t ry . Univ. Chicago
Press 1952 6. K a r n o v s k y , M. J., Roots , L.: J. H i s tochem. Cytochem.
12, 219 (~964) 7. Kreu tzberg , G. W., et al., in: I n t e rna t . Sympos . on The
P a t h o l o g y of Microcirculat ion (J. Cervbs -Navar ro and F. Matakas , eds.), 3. 5 -Sept . t973. Berl in: De G ru y t e r (in press)
8. Crook, J. C. : N a t u r e 199, 41 (1963) 9- F lumerfe l t , B. A., Lewis, P. R., Gwyn, D. G. : H i s tochem.
J. 5, 67 (1973) 10. Gwyn, D. G., Wols tencrof t , J. H. : J. Comp. Neurol . 133,
289 (1968)
Na tu rw i s senscha f t en 61 (1974) �9 by Spr inger-Ver lag t974 37