Embed Size (px)
Citation preview





Dengue Bulletin – Volume 31, 2007 iii
Acknowledgements
Editor, Dengue Bulletin, WHO/SEARO, gratefully thanks the following for peer reviewing manuscriptssubmitted for publication.
In-house Review:
Nand L. Kalra: Reviewed the manuscripts in respect of format check, content, conclusions drawn,including condensation of tabular and illustrative materials for clear, concise and focused presentationand bibliographic references. He was also involved in the final stages of printing of the Bulletin.
Peer reviewers
1. Duane J. GublerAsia-Pacific Institute of Tropical Medicine and Infectious DiseasesJohn A. Burns School of MedicineHonolulu, HI 96813, USAE-mail: [email protected]
2. John G. AaskovWHO Collaborating Centre for Arbovirus Reference and ResearchQueensland University of TechnologyBrisbane, QueenslandAustraliaE-mail: [email protected]
3. Eng Eong OoiDefense Medical and Environmental Research InstituteDSO National Laboratories27 Medical Drive, #09-01Singapore 117510E-mail: [email protected]
4. V.K. SaxenaCentre for Medical Entomology and Vector ManagementNational Institute of Communicable Diseases22, Shamnath MargNew Delhi 110054, IndiaE-mail: [email protected]
5. Philip McCallVector Research GroupLiverpool Tropical School of MedicinePembroke PlaceLiverpool, UKE-mail: [email protected]
6. Henry WildeDivision of Research AffairsFaculty of MedicineChulalongkorn UniversityRama IV Road, BangkokThailand 10330E-mail: [email protected]
7. Kate L. McElroyMolecular Virology and Surveillance LaboratoryCDC Dengue Branch1324 Calle CañadaSan Juan, PR 00920E-mail: [email protected]
8. Pierre F. GuilletVector Control & PreventionGlobal Malaria ProgrammeWHO, 27 Avenue AppiaCH-1211 Geneva 27SwitzerlandE-mail: [email protected]

iv Dengue Bulletin – Volume 31, 2007
Acknowledgements
9. Brian KayAustralian Centre for International and Tropical Health and NutritionQueensland Institute of Medical ResearchPost Office Royal Brisbane HospitalBrisbane, QueenslandAustralia 4029E-mail: [email protected]
10. Audrey LenhartVector GroupLiverpool School of Tropical MedicinePembroke PlaceLiverpool L3 5QA, UKE-mail: [email protected]
11. Craig WilliamsSansom InstituteSchool of Pharmacy & Medical SciencesUniversity of South AustraliaGPO Box 2471 AdelaideSouth Australia 5000E-mail: [email protected]
12. Gavin ScreatonImperial CollegeHammersmith HospitalDu Cane RoadLondon W12 0NNE-mail: [email protected]
13. Linda S. LloydPublic Health Consultant3443 Whittier St.San Diego, CA 92106E-mail: [email protected]
14. Veerle VanlerbergheEpidemiology and Disease Control UnitDepartment of Public HealthInstitute of Tropical MedicineNationalestraat 1552000 AntwerpBelgiumE-mail: [email protected]
15. Rosemary C. SangArbovirology/VHF LaboratoryCentre for Virus ResearchKenya Medical Research InstituteP. O. Box 54628Nairobi, KenyaE-mail: [email protected]
16. Goro KunoArboviral Diseases BranchDivision of Vector-Borne Infectious DiseasesNational Center for Infectious DiseasesCenters for Disease Control and PreventionFort Collins, ColoradoUSAE-mail: [email protected]
17. John D. EdmanProfessor EmeritusDepartment of EntomologyUC DavisOne Shields AvenueDavis, CA 95616-8584USAE-mail: [email protected]
18. Cameron SimmonsOxford University Clinical Research UnitHospital for Tropical Diseases190 Ben Ham Tu, District 5Ho Chi Minh City, Viet NamE-mail: [email protected]
19. Alan L. RothmanCenter for Infectious Disease and Vaccine ResearchUniversity of Massachusetts Medical School55 Lake Avenue NorthWorcester, MA 01655USAE-mail: [email protected]

Acknowledgements
Dengue Bulletin – Volume 31, 2007 v
20. Daniel StrickmanVeterinary, Medical, and Urban EntomologyAgricultural Research ServiceU.S. Department of AgricultureGeorge Washington Carver Center5601 Sunnyside AvenueBeltsville, Maryland, 20705USAE-mail: [email protected]
21. Timothy P. EndyInfectious Disease DivisionDepartment of MedicineSUNY Upstate Medical University725 Irving Avenue, Suite 304Syracuse, NY 13210USAE-mail: [email protected]
22. Wej ChoochoteDepartment of ParasitologyFaculty of MedicineChiang Mai UniversityChiang Mai 50200ThailandE-mail: [email protected]
23. Ng Lee-ChingEnvironmental Health InstituteNational Environment Agency40 Scotts RoadEnvironment Building #13-00Singapore 228231E-mail: [email protected]
24. John T. RoehrigArboviral Diseases BranchDVBID, NCZVED, CCIDU.S. Centers for Disease Control and Prevention3150 Rampart RoadFort Collins, CO 80521USAE-mail: [email protected]; [email protected]
25. Gerardo ChowellSchool of Human Evolution and Social ChangeArizona State UniversityBox 872402Tempe, AZ 85287, USAE-mail: [email protected]
26. Amy MorrisonNMRCDClinica NavalAv. La Marina con C/Trujillo #951Punchana, PeruE-mail: [email protected]
27. Lourdes EstevaDepartamento de MatemáticasFacultad de CienciasUNAM 04510 México, D.F.MexicoE-mail: [email protected]
28. Dana A. FocksInfectious Disease Analysis, LLCP.O. Box 12852Gainesville, FL 32604or7409 NW 23rd AvenueGainesville, FL 32606, USAE-mail: [email protected]
29. Neal AlexanderInfectious Diseases Epidemiology UnitLondon School of Hygiene and Tropical MedicineLondon, UKE-mail: [email protected]
30. Siripen KalyanaroojWHO Collaborating Centre for Case Management of Dengue/DHF/DSSQueen Sirikit National Institute of Child Health (Children’s Hospital)Bangkok, ThailandE-mail: [email protected]

vi Dengue Bulletin – Volume 31, 2007
Acknowledgements
31. Morteza ZaimWHO Pesticide Evaluation Scheme (WHOPES)Vector Ecology & ManagementDepartment of Control of Neglected Tropical DiseasesWorld Health Organization20 Avenue AppiaCH-1211 Geneva 27SwitzerlandE-mail: [email protected]
32. Michael NathanWorld Health OrganizationHeadquarters20 Avenue Appia1211 Geneva 27SwitzerlandE-mail: [email protected]
The quality and scientific stature of the Dengue Bulletin is largely due to the conscientiousefforts of the experts and also due to the positive response of contributors to comments andsuggestions.

Dengue Bulletin – Volume 31, 2007 1
Epidemic situation of dengue fever in Guangdongprovince, China, 1990-2005
He Jian-feng* , Luo Hui-ming*, Liang Wenj-jia, Zheng Kui, Kang Minand Liu Li-ping
Guangdong Provincial Center for Disease Control and Prevention,WHO Collaborating Centre for Surveillance, Research and Training of Emerging Infectious Diseases,
Guangzhou, China
Abstract
During the period 1990 to 2005, a total of 11 844 cases of dengue fever (DF), with 3 deaths, werereported in Guangdong province. The average attack rate was 1.27/100 000 pop. The disease affected17 out of 21 cities in the province. DF occurred throughout the year and the epidemic phase extendedfrom July to December. DENV-1, that affected all age groups, appeared to be the predominant circulatingvirus, although DENV-2, -3 and -4 were also involved. Aedes albopictus was the only vector speciesresponsible for the DF outbreaks.
The results of molecular epidemiological studies showed that dengue fever epidemics in Guangdongprovince were initiated by imported cases from South-east Asia. Control measures comprised ofinsecticidal fogging and elimination of man-made breeding places of Ae. albopictus.
Keywords: Dengue fever; Epidemiological surveillance; Guangdong; China.
E-mail: [email protected]; Tel.: 86-20-84195466, Fax: 86-20-84193323*These authors contributed equally to this work.
Introduction
Dengue fever (DF), an acute febrile viraldisease characterized by sudden onset of feverfor 2–7 days (sometimes biphasic), intenseheadache, myalgia, arthralgia, retro-orbital pain,anorexia, nausea, vomiting and rash, is one ofthe most common and widespread vector-borne arboviral infections in the world. Theviruses of dengue fever belong to the familyFlaviridae and include four serotypes (DENV-1, -2, -3, -4), all of which can classically causeundifferentiated fever, dengue fever and itssevere form, dengue haemorrhagic fever
(DHF). The global prevalence of dengue hasincreased dramatically in recent decades. Asper WHO estimates, DF/DHF is now prevalentin over 100 countries, posing a threat to morethan 2.5 billion people in the tropics andsubtropics.[1,2]
In China, in the 1980s, Aedes aegypti wasreported in Hainan island and Leizhoupeninsula, and caused an epidemic affectingone million people. However, Aedes albopictus,another common species of mosquito, isknown to be the principal vector of DF inGuangdong province since 1990.[3-5] Guangdong

2 Dengue Bulletin – Volume 31, 2007
Dengue in Guangdong Province, China
has been a major province in China affectedby dengue fever outbreaks in addition to Fujian,Zhejiang and Jiangsu provinces since 1990.[6]
Guangdong province had a population of85 million (in 2000).[7] It is located in southernChina in the subtropical zone, which is suitablefor mosquito breeding. The first confirmedoutbreak of dengue in the province was reportedin 1978.[8-9] Since then, dengue fever hasbecome a major public health concern. Thepresent study focuses on the epidemiologicalfeatures and molecular characterization ofdengue viruses during 1990–2005.
Materials and methods
Data sources
In China, DF is a notifiable disease. Hence, allhealth care facilities like hospitals and clinicsin the country are required to report clinicaland confirmed cases of dengue to the Centersfor Disease Control and Prevention (CDC)system: first to the county CDC, which thenreports to the provincial CDC. Once thesenotifications are received, the CDC performslaboratory verification, then conducts furtherepidemiological investigation, and searches forother dengue cases or clusters.
Based on mandatory notification, wecollected and analysed all notified cases ofdengue in Guangdong province from 1990 to2005. However, it is required by law to classifythese notifications of dengue into differentclinical manifestations {DF, DHF, dengue shocksyndrome (DSS)}. Both clinical cases andconfirmed cases are reported.[10]
Case definitions
A clinical case of dengue is defined as follows:
• Clinical symptoms, such as abruptonset of fever, severe headache,myalgia, arthralgia, nausea, vomiting,rash, etc.
• When only sporadic cases are foundor no local cases have been reported:clinical manifestations of dengue (DF,DHF or DSS) with positive IgG or IgMin a single serum specimen.
• When local cases have been reported:clinical manifestations of dengue (DF,DHF or DSS) with decreased whitecell count (<4×109/l) and lowerthrombocytopenia count (<100×109/l).
• The isolation of DENV, by cell cultureand virus antigen preparation fromculture supernatants of DENV-1,DENV-2, DENV-3 and DENV-4-infected C6/36 cells.[11] The culturesupernatants were used as the sourceof E/M and NS1 antigens for ELISA.The control antigen was prepared bythe same procedure from vero cellsculture without viral infection.
• Positive test of real-time one-step RT-PCR: The nucleotide sequence ofDENV-1 E/NS1 gene segment wasisolated. Viral RNA was extracted foruse in one-step reverse transcriptasepolymerase chain reaction (RT-PCR).Following amplification by RT-PCR, thepartial nucleotide fragments of the E/NS1 gene junction were then clonedinto the plasmid pbluescript II SK forsequencing. The following primerswere used for sequencing:
Forward primer:5’–GTCGAGCTCGGATCACAAGAAGGAG–3’
Reverse primer:5’–TGATGGTACCGAGACGAGTGGCTGA–3’
The sequence results were analysed usingthe DNASTAR software.
Dengue in Guangdong Province, China
Dengue Bulletin – Volume 31, 2007 3
• Positive seroconversion or four-foldincrease in dengue-specific IgM or IgGantibody from appropriately timedpaired serum (with acute-phase seracollected during day 1–7 after theonset of symptoms, and early and lateconvalescent sera collected during day8–13 and day 14–30, respectively).
• High-titre dengue-specific IgM and IgGantibody in a single serum specimenwhere cross-reaction to Japaneseencephalitis (JE) had been excluded.
Vector surveillance
In Guangdong, surveillance of Aedes albopictuswas carried out throughout the year, particularlyin areas reporting active cases. Theinvestigations included search for Aedesbreeding places and determining theentomological indices, viz. Breteau index (BI)(number of positive containers for Aedes per100 houses) and Container index (CI)(percentage of containers positive for Aedesbreeding) until two weeks after the last localcase was reported.
Results
Epidemiological information
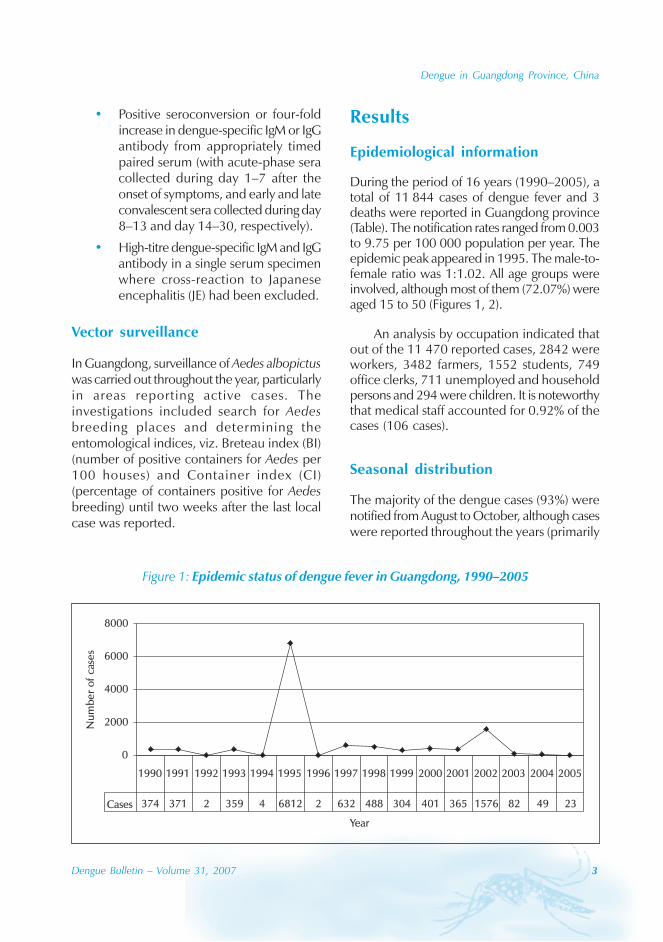
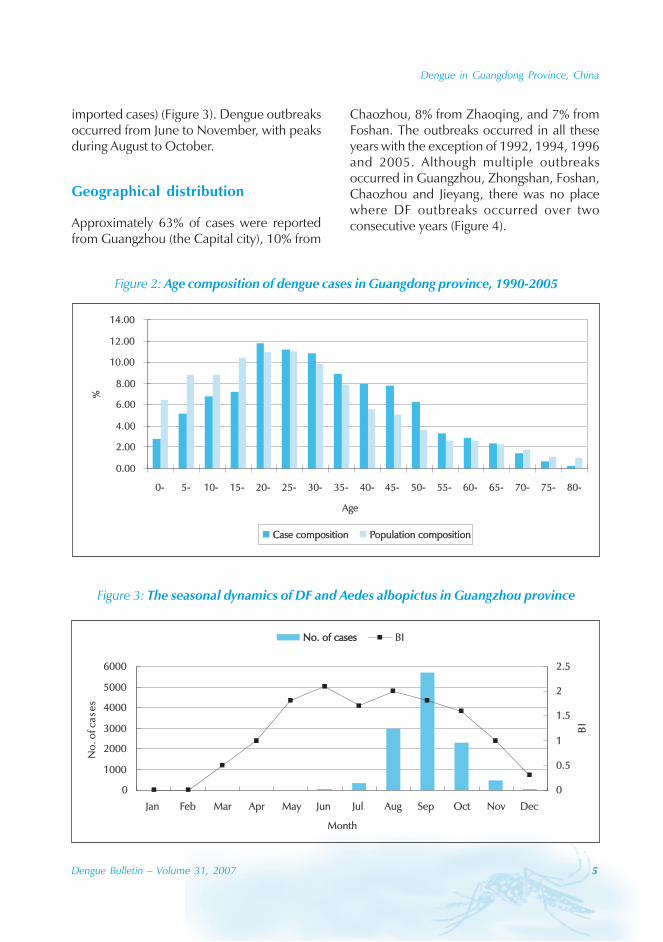
During the period of 16 years (1990–2005), atotal of 11 844 cases of dengue fever and 3deaths were reported in Guangdong province(Table). The notification rates ranged from 0.003to 9.75 per 100 000 population per year. Theepidemic peak appeared in 1995. The male-to-female ratio was 1:1.02. All age groups wereinvolved, although most of them (72.07%) wereaged 15 to 50 (Figures 1, 2).
An analysis by occupation indicated thatout of the 11 470 reported cases, 2842 wereworkers, 3482 farmers, 1552 students, 749office clerks, 711 unemployed and householdpersons and 294 were children. It is noteworthythat medical staff accounted for 0.92% of thecases (106 cases).
Seasonal distribution
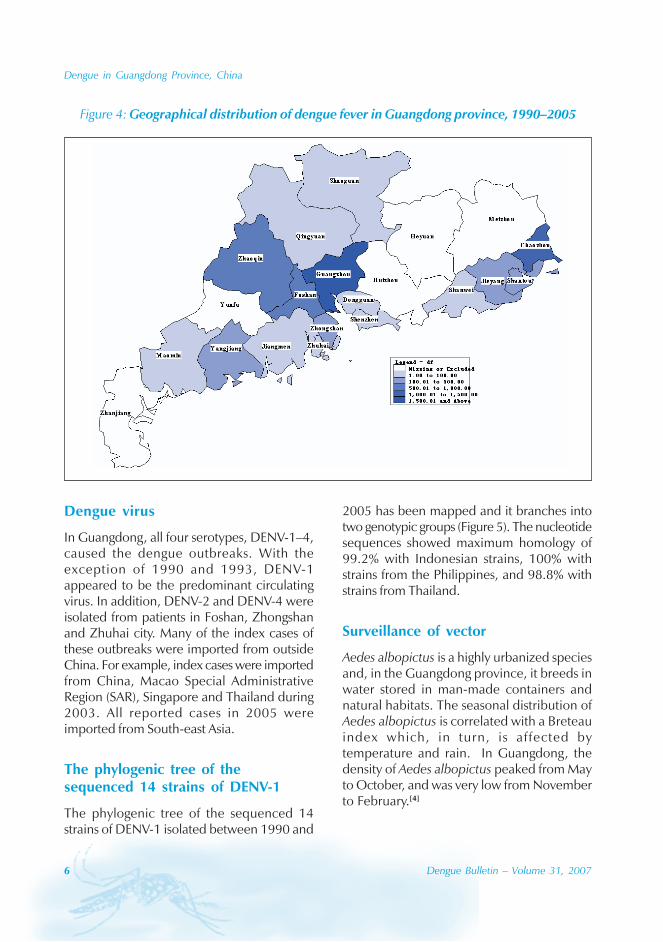
The majority of the dengue cases (93%) werenotified from August to October, although caseswere reported throughout the years (primarily
Figure 1: Epidemic status of dengue fever in Guangdong, 1990–2005
0
2000
4000
6000
8000
Year
Num
ber
of
case
s
Cases 374 371 2 359 4 6812 2 632 488 304 401 365 1576 82 49 23
1990 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000 2001 2002 2003 2004 2005
4 Dengue Bulletin – Volume 31, 2007
Dengue in Guangdong Province, China
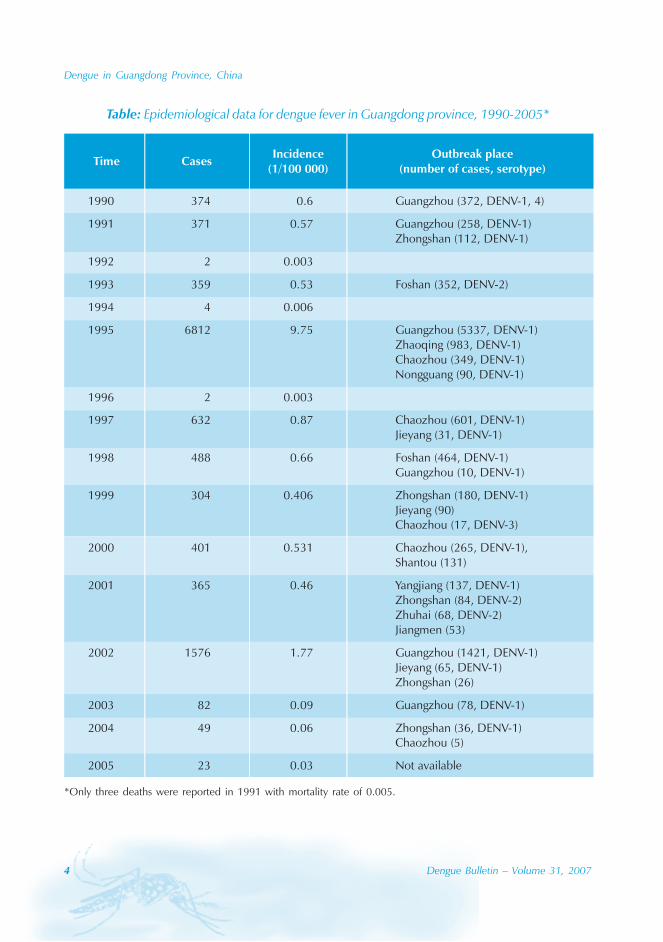
Table: Epidemiological data for dengue fever in Guangdong province, 1990-2005*
*Only three deaths were reported in 1991 with mortality rate of 0.005.
Dengue in Guangdong Province, China
Dengue Bulletin – Volume 31, 2007 5
imported cases) (Figure 3). Dengue outbreaksoccurred from June to November, with peaksduring August to October.
Geographical distribution
Approximately 63% of cases were reportedfrom Guangzhou (the Capital city), 10% from
Chaozhou, 8% from Zhaoqing, and 7% fromFoshan. The outbreaks occurred in all theseyears with the exception of 1992, 1994, 1996and 2005. Although multiple outbreaksoccurred in Guangzhou, Zhongshan, Foshan,Chaozhou and Jieyang, there was no placewhere DF outbreaks occurred over twoconsecutive years (Figure 4).
Figure 2: Age composition of dengue cases in Guangdong province, 1990-2005
0.00
2.00
4.00
6.00
8.00
10.00
12.00
14.00
0- 5- 10- 15- 20- 25- 30- 35- 40- 45- 50- 55- 60- 65- 70- 75- 80-
Age
%
Case compositionCase composition Population compositionPopulation composition
Figure 3: The seasonal dynamics of DF and Aedes albopictus in Guangzhou province
0
1000
2000
3000
4000
5000
6000
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Month
No
.ofc
ase
s
0
0.5
1
1.5
2
2.5
BI
No. of casesNo. of cases BI
6 Dengue Bulletin – Volume 31, 2007
Dengue in Guangdong Province, China
Figure 4: Geographical distribution of dengue fever in Guangdong province, 1990–2005
Dengue virus
In Guangdong, all four serotypes, DENV-1–4,caused the dengue outbreaks. With theexception of 1990 and 1993, DENV-1appeared to be the predominant circulatingvirus. In addition, DENV-2 and DENV-4 wereisolated from patients in Foshan, Zhongshanand Zhuhai city. Many of the index cases ofthese outbreaks were imported from outsideChina. For example, index cases were importedfrom China, Macao Special AdministrativeRegion (SAR), Singapore and Thailand during2003. All reported cases in 2005 wereimported from South-east Asia.
The phylogenic tree of thesequenced 14 strains of DENV-1
The phylogenic tree of the sequenced 14strains of DENV-1 isolated between 1990 and
2005 has been mapped and it branches intotwo genotypic groups (Figure 5). The nucleotidesequences showed maximum homology of99.2% with Indonesian strains, 100% withstrains from the Philippines, and 98.8% withstrains from Thailand.
Surveillance of vector
Aedes albopictus is a highly urbanized speciesand, in the Guangdong province, it breeds inwater stored in man-made containers andnatural habitats. The seasonal distribution ofAedes albopictus is correlated with a Breteauindex which, in turn, is affected bytemperature and rain. In Guangdong, thedensity of Aedes albopictus peaked from Mayto October, and was very low from Novemberto February.[4]

Dengue in Guangdong Province, China
Dengue Bulletin – Volume 31, 2007 7
Discussion
We have provided the epidemiologicalinformation of dengue spanning over a periodof 16 years from 1990 to 2005 in Guangdongprovince. Our data sources are based onmandatory notification, which have the intrinsicinadequacies such as incomplete reporting, orfalse positive results due to cross reaction ofdengue viruses with Japanese encephalitisvaccine viruses. In addition, denguenotifications in China are not classified byclinical manifestations (DF, DHF or DSS), amajority of them are known to be presentedas classical dengue fever. For example, a totalof 758 cases of dengue were reported in 1995,of which 12 (1.58%) were DHF.[6] Out of the978 hospitalized dengue cases in an infectiousdisease hospital of Guangzhou in 2002, onlytwo were DHF.[12]
A secular trend of increased reporting ofdengue outbreak was observed. The developingeconomy and rapid urbanization withinGuangdong have contributed towards increasedopportunities for the development andtransmission of DF in the province, especiallyin the Pearl delta region. Risk factors for DFepidemic included: (i) the changing lifestyle ofpeople; for example, more people now growbonsai and raise fish in tanks near their homes;(ii) more construction sites with poor sanitationconditions; (iii) frequent travel to and fromSouth-east Asia resulting in importation ofviruses, especially the Chaozhou and Shantou;(iv) delayed detection and treatment of activedisease due to lack of knowledge of DF amongprimary medical staff; and (v) high densities ofvector mosquitoes. The vector control mainlycovered adult Aedes mosquitoes control, whichremains largely an ineffective intervention.
Figure 5: Phylogenetic tree of DENV-1 isolated in different years and regions fromGuangdong province

8 Dengue Bulletin – Volume 31, 2007
Dengue in Guangdong Province, China
References
[1] World Health Organization. Strengtheningimplementation of the global strategy for denguefever/dengue haemorrhagic fever. report of theinformal consultation, 18-20 October 1999.Geneva: 2001. Document WHO/CDS/(DEN)/IC/2000.1.
[2] Monath TP. Dengue: the risk to developed anddeveloping countries. Proc Natl Acad Sci U SA. 1994 Mar 29; 91(7): 2395-400.
[3] Baolin L. The problem of the method selectionin mosquito control. Chinese Journal of VectorBiology and Control, 1997, 8: I-V.
[4] Huiming L, editor. Manual of dengue feverprevention, control and treatment. StandardPress of China, 2002.
[5] Guijiao L, Youchan G, Bao-yan W, Ye T. Ananalysis of epidemic of dengue fever inZhongshan city. Chin J Dis Control Prev. 2004;8(5): 396-398.
[6] Wenjie W. Control of dengue/denguehaemorrhagic fever in China. Dengue Bulletin.1997; 21: 25-29.
The evidences from epidemiologicalinvestigation and molecular epidemiologysupported the hypothesis that the identifieddengue cases were imported from othercountries into Guangdong. The circulating virusstrains in an area were usually different indifferent years, and different virus serotypesprevailed in different areas in one year. From1979 to 1999, the circulating DENV-1 strainbelonged to two geno-subtypes. It is likely thatmost of these strains were imported fromneighbouring countries such as the Philippines,Indonesia and Thailand, where indigenousdengue epidemics have been confirmed. Theresults of nucleotide sequencing showed thatDENV-1 was closely related to viruses identifiedin these regions. High mosquito density andfavourable natural conditions (such as optimaltemperature and rainfall) are responsible forlocalized outbreaks, once dengue viruses areimported into the province.[13]
For building up an early response tooutbreaks, it is necessary to improve diagnosticcapacity and strengthen training and reportingawareness of local physicians, particularly inearly detection, diagnosis, notification andisolating patients. Secondly, it is important toestablish a vector monitoring system tounderstand population dynamics of Aedes
albopictus. Thirdly, comprehensive measuresof prevention and control includingenvironmental solid waste disposal andchemical, biological and ecological controlshould be developed. Lastly, community healtheducation needs to be strengthened to improveawareness of the dengue virus transmissionthrough Aedes albopictus.
From our experience, source reduction anduse of non-chemical methods supported byintersectoral and community participation arethe most potential methods for control ofdengue during interepidemic periods. TheBreteau index is the most important indicatorto predict impending dengue outbreaks in thisprovince. Maintaining lower BI levels (below5) is a very effective measure to prevent andcontrol dengue outbreaks. Further studies arerequired to determine the BI cut-off value formosquito control to prevent DF outbreaks.
Acknowledgment
The authors thank local health departmentemployees who provided epidemiological andlaboratory data. The authors also thank thesupport of Chin-Kei Lee and Ms AndreaBoudvill of the World Health Organization.

Dengue in Guangdong Province, China
Dengue Bulletin – Volume 31, 2007 9
[7] Population Census Office of Guangdongprovince. Tabulation of the 2000 populationcensus of Guangdong province. Beijing: ChinaStatistics Office, 2002.
[8] Zhao HL, Luo QH, Shen G. The epidemiologyof dengue outbreak in Shiwanjheu, Nanhaicounty, Guangdong province., China. MedJour. 1981; 61(8): 466-469.
[9] Zao ZG, Lin HZ. Report and analysis of dengueand dengue haemorrhagic fever 1980-86.Special publication on dengue. Hainan Medi.1986; 36: 60-66. Chinese.
[10] Hao R, Luo H, Peng W, et al. Diagnosis criteriaand treatment principles for dengue fever.Beijing: Publishing House of China; 2002.p728.
[11] Tesh RB. A method for the isolation andidentification of dengue viruses, using mosquitocell cultures. Am J Trop Med Hyg. 1979 Nov;28(6): 1053-9.
[12] Luo H, He J, Zheng K, Li L, Jiang L. Analysis onthe epidemiologic features of dengue fever inGuangdong province, 1990-2000. ZhonghuaLiu Xing Bing Xue Za Zhi. 2002 Dec; 23(6):427-30. Chinese.
[13] Yanqing C, Fuchun Z, Jian W. Clinicalcharacteristics analysis of 978 patients withdengue fever in Guangzhou areas. J TropicalMedicine. 2003 Jun; 3(2): 190-192. Chinese.

10 Dengue Bulletin – Volume 31, 2007
A clinical, epidemiological and virological study ofa dengue fever outbreak in Guangzhou, China –
2002-2006
Fuchun Zhanga , Xiaoping Tanga, Xuchu Hub, Yecheng Lua, Yanqing Chena,Jian Wanga, Wanshan Chena and Haolan Hea
aDepartment of Infectious Diseases, Guangzhou No. 8 People’s Hospital, Guangzhou, 510060, China
bDepartment of Parasitology, Zhongshan Medical College of Sun Yat-sen University, Guangzhou, 510060,China
Abstract
We analysed the clinical and epidemiological characteristics of dengue fever (DF) during the denguevirus (DENV)-1 outbreak in Guangzhou, China. Clinical and epidemiological data of 1342 patientswith DF from May 2002 to November 2006 were analyzed retrospectively. The average age was34.7±13.2 years. The ratio of male to female was 1.05:1. The peak time of the epidemic lasted fromAugust to October. The most common manifestations included fever (100%), headache (85.9%), myalgia(64.5%), bone soreness (46.6%), fatigue (78.2%), skin rash (65.9%) and positive tourniquet test (51.3%).Leukopenia, thrombocytopenia, elevated alanine aminotransferase (ALT), elevated aspartateaminotransferase (AST) and hypopotassemia were found in 66%, 61.3%, 69%, 85.7% and 28.4% of thepatients respectively. Only 2(0.15%) patients fulfilled the WHO case definition criteria for denguehaemorrhagic fever (DHF), the others were all diagnosed as classic DF. However, 64(4.8%) patients hadsevere clinical manifestations (internal haemorrhage, shock, marked thrombocytopenia, myocarditis,sepsis, pneumonia and encephalopathy). Anti-dengue IgM was detected in 90% of patients. Dengueviruses were isolated using the Aedes albopictus C6/36 cell line and identified as DENV-1 by RT-PCR. A346bp fragment from RT-PCR product of every isolate was sequenced to compare with publishedsequences of other DENV-1 viruses. The nucleotide homology were 97%, 97% and 98% comparedwith those of DENV-1 strains of dengue fever outbreak in Cambodia, in 1997 and 1999 in China,respectively. In conclusion, the epidemic of dengue fever was caused by DENV-1 infection from 2002to 2006 in Guangzhou. Patients with severe clinical manifestations are few, but in some of them, thediagnosis of DHF may be missed if the WHO classification is strictly applied, especially in adults.
Keywords: Dengue fever; DENV-1, Severe dengue; Epidemiological and clinical characteristics.
E-mail: [email protected]
Introduction
Dengue fever (DF) is an acute febrile viraldisease frequently presenting with headaches,bone or joint and muscular pains, rash andleukopenia. DF has become one of the most
important emerging public health problems inthe tropics and subtropics. In the past 20 years,there has been a dramatic increase in thenumber of cases and the severity of illness incountries in South-East Asia, the Caribbean andLatin America.[1,2] An estimated 50-100 million

Dengue fever outbreak in Guangzhou, China – 2002-2006
Dengue Bulletin – Volume 31, 2007 11
people across the globe contract dengueannually, with about 500 000 persons developingthe more severe dengue haemorrhagic fever/dengue shock syndrome (DHF/DSS) andresulting in about 21 000 deaths.[3]
WHO classifies symptomatic dengue virusinfection into three categories: undifferentiatedfever, classic dengue fever, and DHF.[3]
According to WHO criteria, DHF is defined bythe presence of fever, a haemorrhagic tendency,thrombocytopenia, and some evidence ofplasma leakage. DHF is further subdivided, withmost severe cases categorized as DSS, whencirculatory failure is present. The WHOclassification system has been widely applied inresearch settings and publications. In recentyears, several studies reported difficulties withclassification, especially in adults with manysevere cases being missed: more than two thirdsof all adults with severe dengue manifestationswere not classified as having DHF.[4-7]
The first dengue case in Guangdong wasreported in 1978. Infections with all 4 denguevirus serotypes (DENV-1, 2, 3, and 4) werereported in Guangdong.[8] The first dengueoutbreak was caused by DENV-3 in 1980. Sincethen, increasing larger outbreaks have beendocumented throughout the 1980s and the1990s, caused by DENV-1, DENV-2, and DENV-4. In this paper, we present a retrospectiveanalysis of 1342 patients to analyse theepidemiological, clinical, laboratory andvirological presentations of the dengue infectionfrom 2002 to 2006 in Guangzhou, China.
Materials and methods
Patients and investigative methods
The study was conducted from May 2002 toNovember 2006 at the Guangzhou No. 8People’s Hospital. A total of 1360 patients were
initially enrolled in this study. Eighteen patientswhich were imported cases in 2004-2005 wereexcluded. Thus, 1342 patients (978 in 2002,54 in 2003, and 310 in 2006) were analysedin Guangzhou where dengue was endemic.
In all patients, a detailed history was takenand clinical examination was done at the timeof admission and subsequently during the stayat the hospital. The hospital records for allpatients were supported by serological and/orvirological confirmation. Serology wasperformed by indirect ELISA, and/orimmunochromatographic test (ICT), and dotimmunobinding assay (DIBA) for specific IgM.
Virus isolation
We collected 85 blood samples within five daysafter onset (36 samples were obtained in 2002,18 samples in 2003, and 41 samples in 2006).Virus was isolated by Aedes albopictus mosquitoC6/36 cell micromethod. The patient bloodserum was diluted with RPMI 1640 containing2% FBS to an appropriate concentration andinoculated into the C6/36 cells. The C6/36 cellswere incubated at 28 °C, 5% CO2 for 21 days.Cultures were monitored for cytopathic effect(CPE) daily under the inverted phase contrastmicroscope. The inoculated cells werepassaged once a week. Those with CPE werejudged as positive for dengue virus, while thosewithout CPE after passaging three times weredeclared negative.[9]
RT-PCR, DNA purification andsequencing
Viral RNA was extracted from the culturesupernatants of infected C6/36 with animproved sodium iodide method. Nucleotidesencoding a fragment of the NS2 gene wereamplified using RT-PCR. The amplificationparameter was as follows: 93° 40s, 55° 45s,

12 Dengue Bulletin – Volume 31, 2007
Dengue fever outbreak in Guangzhou, China – 2002-2006
72° 60s, thirty cycles. The products of RT-PCRwere then analysed by 1.5% agarose (includingethidium bromide) electrophoresis. Theuniversal primer sequences were P 5’-G A C AT G G G G TAT T G G AT- 3 ’ , P 5 ’ -TCCATCCCATACCAGCA-3’, which result in anamplification product of 413bp. DENV-1-typespecific primer sequences were D1S 5’-G T T G T TC C G C A G A C TA- 3 ’ , D 1 C 5 ’ -CATGGTTAGTGGTTTG -3’, and the expectedsize of the amplification product was 346bp.The DENV and DENV-1, 2, 3, 4-type specificprimers were provided by Institute of MilitaryMedicine in Joint-Service Department ofGuangzhou Military Region. The bands ofpredicted size were cut and purified using aGolden Bead Product Purification Kit (Songon,China) according to the manufacturers’instructions. Purified PCR products were clonedon to pMD18-T Vector (TaKaRa, Japan) anddelivered to TaKaRa Biotechnology Co. Ltd.for sequencing.[10]
Sequences analysis
Sequence homology searches within theGenBank gene database were performed usingNCBI BLASTn. In addition, CLUSTALX andTREEVIEW software were used to analysesequence homology, and constructed its systemcladogram. The DENV-1 international anddomestic reference strains are as follows:Singapore strain(S275/90), Cambodia strain(Cambodia), West Pacific Ocean strain(WestPac), Thai strain (TH-Sman), Peruvianstrain (Peru), Nauru strain (Nauru), Hawaianstandard strain (Hawaii), and Guangdong 1995strain (GD23/95), 1997 strain (GD14/97), 1999epidemic strain (GD05/99).
Statistics analyses
All data were analysed using the SPSS10.0software package.
Result
Patients’ characteristics andgeographical and time distributionof dengue outbreak
During the study period, we observed a total of1342 patients (687 male and 655 female, male/female ratio = 1.05:1). The involvement of allage groups, especially an adult predominance,was observed. The mean age of the patientswas 34.4±13.1 years and most of them belongedto the 20~29-year age group; less than 15%patients were children (Figure 1).
Figure 1: Age groups distribution in 1342patients with dengue fever
8.2
14.3
23.4
20.2
16.2
9.8
5.4
2.5
0
5
10
15
20
25
0- 10- 20- 30- 40- 50- 60- 70-
Age groupsAge groups
Perc
ent
(%)
Perc
ent
(%)
Geographical and time distributionof dengue outbreak
Dengue fever cases were observed to bedistributed in all 12 districts of Guangzhou.There were 872(65%) patients in thepredominant outbreak spots where the numberof DF cases was above 10.
The peak time of the epidemics was fromAugust to October each year. The number ofcases in September reached its maximum, with587 hospitalized cases, which constituted43.7% of the total cases.
Dengue fever outbreak in Guangzhou, China – 2002-2006
Dengue Bulletin – Volume 31, 2007 13
Figure 2: Clinical presentation in 1342 patients with dengue fever
fever
headache
fatigue
skin rash
myalgia
positive tourniquet test
bone pain
nausea and vomiting
petechiae
lymphadenectasis
diarrhoea
splenohepatomegalia
cough
retro-orbital pain,
internal haemorrhage
pleural effusion
0 200 400 600 800 1000 1200 1400 1600
The number of patients of every clinical presentation
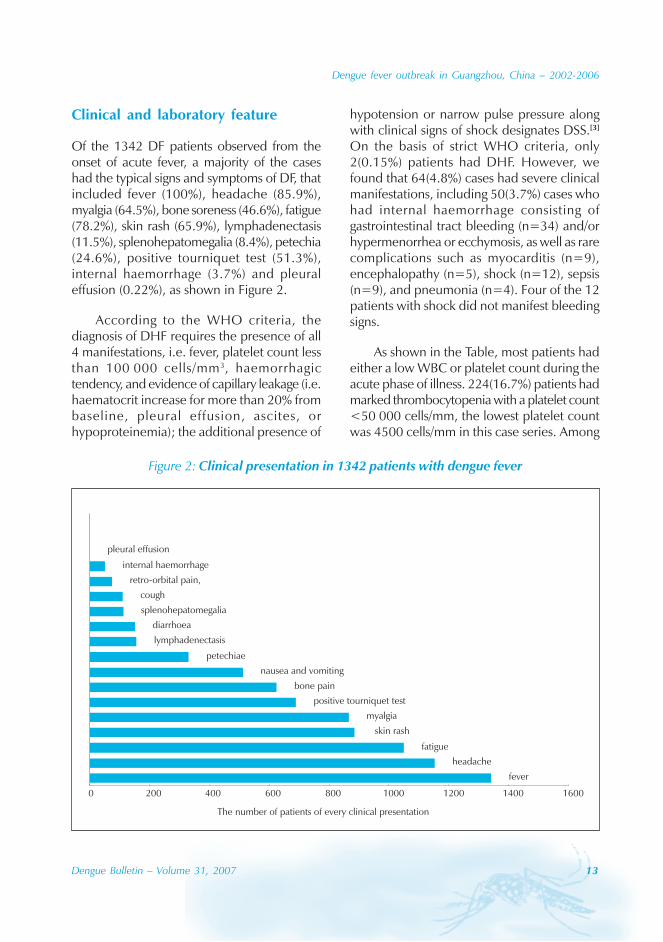
Clinical and laboratory feature
Of the 1342 DF patients observed from theonset of acute fever, a majority of the caseshad the typical signs and symptoms of DF, thatincluded fever (100%), headache (85.9%),myalgia (64.5%), bone soreness (46.6%), fatigue(78.2%), skin rash (65.9%), lymphadenectasis(11.5%), splenohepatomegalia (8.4%), petechia(24.6%), positive tourniquet test (51.3%),internal haemorrhage (3.7%) and pleuraleffusion (0.22%), as shown in Figure 2.
According to the WHO criteria, thediagnosis of DHF requires the presence of all4 manifestations, i.e. fever, platelet count lessthan 100 000 cells/mm3, haemorrhagictendency, and evidence of capillary leakage (i.e.haematocrit increase for more than 20% frombaseline, pleural effusion, ascites, orhypoproteinemia); the additional presence of
hypotension or narrow pulse pressure alongwith clinical signs of shock designates DSS.[3]
On the basis of strict WHO criteria, only2(0.15%) patients had DHF. However, wefound that 64(4.8%) cases had severe clinicalmanifestations, including 50(3.7%) cases whohad internal haemorrhage consisting ofgastrointestinal tract bleeding (n=34) and/orhypermenorrhea or ecchymosis, as well as rarecomplications such as myocarditis (n=9),encephalopathy (n=5), shock (n=12), sepsis(n=9), and pneumonia (n=4). Four of the 12patients with shock did not manifest bleedingsigns.
As shown in the Table, most patients hadeither a low WBC or platelet count during theacute phase of illness. 224(16.7%) patients hadmarked thrombocytopenia with a platelet count<50 000 cells/mm, the lowest platelet countwas 4500 cells/mm in this case series. Among

14 Dengue Bulletin – Volume 31, 2007
Dengue fever outbreak in Guangzhou, China – 2002-2006
Item Cases (%)White blood cell count<×109/L 886 (66.0)4~10×109/L 376 (28.0)>×109/L 80 (6.0)Blood platelets count<100×109/L 828 (61.3)<50×109/L 224 (16.7)ALT elevation 926 (69.0)
40~200 U/L 782 (89.3)>200 U/L 144 (10.7)
AST elevation 1150 (85.7)40~200 U/L 992 (88.2)>200 U/L 158 (11.8)
CK elevation 404 (30.1)LDH elevation 611 (45.5)Hypopotassemia (<3.6 umol/L) 276 (28.4)Total bilirubin elevation 72 (5.4)BUN elevation (>7.1 umol/L) 15 (1.1)
Table: Laboratory parameters in 1342patients with dengue fever
those patients tested, over two thirds of themhad increased liver enzyme levels, 10.7% and11.8% of the patients had a 5-fold increase inALT and AST levels respectively. Hypopotassemiawere found in 276 (28.4%) patients.
Diagnosis of dengue virus infectionand identification of DENV-isolatedstrains and sequence analysis
In the serological analysis, 1208(90%) patientswere positive for IgM antibody while467(34.8%) were positive for anti-dengue IgG.
Forty-four virus isolations (51.7%) weremade from the blood samples of 85 caseswhich were inoculated in C6/36 cells. RT-PCR amplification from the supernatants ofthe infected C6/36 cells samples wasperformed using universal primers, which
Figure 3: Agarose gel analysis of the cDNA products from RT-PCR samples isolated from thesupernatants of the infected C6/36 cells. The expected size of the amplification product
was 346bp with DENV-1 type specific primers.
346bp
1 2 3 4 5
2000bp
1000bp750bp500bp250bp100bp
Lane 1: Sample 1Lane 2: DL 2000 marker (GeneRuler)Lane 3: Sample 2Lane 4: Sample 3Lane 5: Sample 4

Dengue fever outbreak in Guangzhou, China – 2002-2006
Dengue Bulletin – Volume 31, 2007 15
yielded the 413bp fragment, which was thenused for DENV identification and sequencing.All the results were found to be confirmedDENV-1 positive by DENV-1-specific primerand amplified fragment was 346bp locatedon DENV-1 gene group’s NS2a~NS2b site,see Figure 3. The results confirmed that all
dengue viruses belonged to the DENV-1 virus.The nucleotide homology of Guangdongepidemic strain in 2003 were 97%ÿ97% and98% when compared with that of DENV-1strain of dengue fever outbreak in Cambodia,in 1997 and 1999 in Guangdong respectively(see Figure 4).
Figure 4: Sequence comparison of four DENV-1 isolated strains from Guangdong and Cambodia
GD29/2003 : GTTGTTCCGCAGACTAACATCCAGAGAAGTTCTTCTTCTAACAATTGGATTGAGTC :56
GD14/97 : .....................T.................T................: .....................T.................T................ :56
GD05/99 : A..A............................C.: A..A............................C....................... :56
Cambodia : a..a.................................................... :56: a..a.................................................... :56
GD29/2003 : TAGTGGCATCTGTGGAGTTACCAAATTCCTTGGAGGAGCTGGGGGATGGACTTGCA: TAGTGGCATCTGTGGAGTTACCAAATTCCTTGGAGGAGCTGGGGGATGGACTTGCA :112
GD14/97 : .............................C.......................... :112
GD05/99 : ........................................................ :112
Cambodia : ........................................................ :112
GD29/2003 : ATGGGCATCATGATTTTAAAATTATTGACTGACTTTCAATCACATTAGTTGTGGGC :168: ATGGGCATCATGATTTTAAAATTATTGACTGACTTTCAATCACATTAGTTGTGGGC :168
GD14/97 : ........T............: ........T.............................G...T..C..C....... :168.................G...T..C..C....... :168
GD05/99 : .............................................C.......... :168: .............................................C.......... :168
Cambodia : ........t....................................c..c....... :168: ........t....................................c..c....... :168
GD29/2003 : TACCTTGCTGTCCTTGACATTTATCAAAACAACGTTTTCCTTGCACTATG: TACCTTGCTGTCCTTGACATTTATCAAAACAACGTTTTCCTTGCACTATGCATGGA :224CATGGA :224
GD14/97 : ........................................................ :224: ........................................................ :224
GD05/99 : ........................................................ :224: ........................................................ :224
Cambodia : ........................................................ :224: ........................................................ :224
GD29/2003 : AGACAA: AGACAATAGCTATGGTACTGTCAATTGTATCTCTCTTCCCCTTATGCCTGTCCACG :280TAGCTATGGTACTGTCAATTGTATCTCTCTTCCCCTTATGCCTGTCCACG :280
GD14/97 : .......G................................................ :280: .......G................................................ :280
GD05/99 : ........................................................ :280: ........................................................ :280
Cambodia : .......g................................................ :280: .......g................................................ :280
GD29/2003 : ACCTCCCAAAAAACAACATGGCTTCCGGTGCTGTTGGGATCTCTTGGATGCAAACC: ACCTCCCAAAAAACAACATGGCTTCCGGTGCTGTTGGGATCTCTTGGATGCAAACC :336
GD14/97 : ................................A......................: ................................A....................... :336
GD05/99 : ..........................: ........................................................ :336
Cambodia : ................................a...................: ................................a....................... :336
GD29/2003 : ACTAACCATG :346: ACTAACCATG :346
GD14/97 : .......... :346: .......... :346
GD05/99 : .......... :346: .......... :346
Cambodia : .......... :346: .......... :346

16 Dengue Bulletin – Volume 31, 2007
Dengue fever outbreak in Guangzhou, China – 2002-2006
Discussion
From May 2002 to November 2006, Guangzhouexperienced a comparatively large-scaledengue fever outbreak. The case specimenscollected at different times over four years wereanalysed by virus isolation, RT-PCR geneidentification and sequencing, confirming thatthis epidemic was caused by DENV-1. Thenucleotide homology of the Guangdongepidemic strain in 2003 were 97%, 97%, and98% compared with those of DENV-1 strain ofdengue fever outbreak in Cambodia, in 1997and 1999 in China, respectively, and thesestrains belonged to the identical gene type.The initial cases were imported during theepidemic period and subsequentlydisseminated locally. Although four serotypeshad been involved in several episodes of thecomparatively large-scale DENV-1 outbreakssince 1995, [7,8] DENV-1 was the mostpredominant serotype in Guangdong.
DF, as a mosquito-borne infection, wouldbe expected to spread rapidly in populousareas, so the presence of a large proportion ofpatients in downtown is consistent with theexperience elsewhere of dengue infection asan urban phenomenon.[11,12] In our study, 65%of 1342 cases presented in the main outbreakspots in downtown Guangzhou. This DENV-1epidemic occurred in the rainy seasons fromAugust to October when the abundant rainfalland the temperature favoured high rates ofmosquito’s reproduction. Rainfall is consideredto be a risk factor for dengue outbreak,especially in urban districts with poor sanitaryenvironment, providing optimal breeding sitesfor mosquitoes.[13] The main mosquito vectorfor dengue viruses is Ae. albopictus inGuangdong, which breeds primarily in relativelyclean, stagnant domestic water containersduring the rainy season.
Our data showed that there was no obviousdifference in the patients’ gender distribution.
All age groups were affected, but over 80% ofthem were adults. Children aged 0~10 yearscomprised only 8.2% of the sample.
In our study, the patients showed most ofthe typical clinical features of dengue fever,with only 2(0.15%) patients developing DHFaccording to the WHO criteria. The incidenceof DHF was much lower in this study,compared with the reported 2%~6% inpopulations in the areas where dengue isendemic.[14,15] In an in-vitro experiment, DENV-1 was isolated from samples of DF patients in2002. DENV-1 is not as virulent as the othertypes of DENV and DENV-1 infection seemsto have mild clinical presentations.[16,17]
In the present study, a few unusual clinicalobservations and complications such asfulminant hepatitis, myocarditis,encephalopathy, and ophthalmic disease wereobserved.[18-23] We found that 64(4.8%) of caseshad severe clinical manifestations includinginternal haemorrhage like gastrointestinal tractbleeding and/or hypermenorrhea orecchymosis, as well as rare complications suchas myocarditis, encephalopathy, shock, sepsis,and pneumonia. 16.7% of the patients hadmarked thrombocytopenia with a platelet count<50 000 cells/mm. More than 10% of thepatients had a 5-fold increased liver enzymes.Our result is consistent with other reports.[12,18]
In recent years, the validity of the WHOclassification scheme to define severe denguedisease has been found to be inconsistent.Several studies have found that the WHOclassification is difficult to apply, with manysevere adult cases being missed.[4-6] Because,according to the WHO criteria, haemorrhagicmanifestations without capillary leakage do notconstitute DHF, the DSS refers to a conditionin which shock is present in addition to all fourDHF-defining conditions. The WHOclassification is mainly based on studies ofchildren in South-east Asia in the 1960s and

Dengue fever outbreak in Guangzhou, China – 2002-2006
Dengue Bulletin – Volume 31, 2007 17
may therefore not be applicable topredominantly adult subjects. Shock and plasmaleakage appear to be more common inchildren, whereas internal haemorrhage is amore frequent manifestation in adults. Inaddition, the DHF/DSS classification excludessevere dengue disease associated with“unusual manifestations”. Therefore, a newdefinition for severe dengue is urgentlyneeded.[24] A large multicentre study shouldbe performed to establish a more robustdengue classification scheme for use byclinicians, epidemiologists and scientistsinvolved in dengue pathogenesis research.[6]
In brief, the DF epidemic in Guangzhoufrom 2002 to 2006 was caused by DENV-1.Most cases had the typical clinical manifestation
of dengue fever and were diagnosed as typicalDF. But in some patients, the diagnosis of DHFmay be missed if the WHO classification isstrictly applied, especially in adults. A newdefinition for severe dengue may be needed.
Acknowledgements
We are grateful to the CDC of Guangzhou andthe CDC of Guangdong province for theircontinuing epidemiological survey andvirological laboratory support. This work waspartially supported by grants from theFoundation of Guangzhou Municipal Scienceand Technology Bureau. We thank AlfredoGarzino-Demo for critically reading thismanuscript.
References
[1] Gubler DJ. Dengue and dengue haemorrhagicfever: its history and resurgence as a globalpublic health problem. In: Gubler DJ, Kuno G,editors. Dengue and dengue haemorrhagic fever.New York: CAB International; 1997. p.1-23.
[2] Gubler DJ. Dengue and dengue hemorrhagicfever. Clin Microbiol Rev. 1998 Jul; 11(3): 480-96.
[3] World Health Organization. Denguehaemorrhagic fever: diagnosis, treatment,prevention and control. 2nd edn. Geneva;WHO, 1997.
[4] Balmaseda A, Hammond SN, Pérez MA,Cuadra R, Solano S, Rocha J, Idiaquez W,Harris E. Short report: assessment of the WorldHealth Organization scheme for classificationof dengue severity in Nicaragua. Am J TropMed Hyg. 2005 Dec; 73(6): 1059-62.
[5] Phuong CX, Nhan NT, Kneen R, Thuy PT, vanThien C, Nga NT, Thuy TT, Solomon T,Stepniewska K, Wills B; Dong Nai StudyGroup. Clinical diagnosis and assessment ofseverity of confirmed dengue infections inVietnamese children: is the World Health
Organization classification system helpful? AmJ Trop Med Hyg 2004 Feb;70(2):172-9.
[6] Deen JL, Harris E, Wills B, Balmaseda A,Hammond SN, Rocha C, Dung NM, HungNT, Hien TT, Farrar JJ. The WHO dengueclassification and case definitions: time for areassessment. Lancet. 2006 Jul 8; 368(9530):170-3.
[7] Luo H, He J, Zheng K, Li L, Jiang L. [Analysis onthe epidemiologic features of Dengue fever inGuangdong province, 1990-2000]. ZhonghuaLiu Xing Bing Xue Za Zhi. 2002 Dec; 23(6):427-30. Chinese.
[8] Zhang FC, Chen YQ, Lu YC, Wang J, ChenWS, Hong WX. [Analysis on clinical andepidemiological characteristics of 1032patients with Dengue fever in Guangzhou].Zhonghua Liu Xing Bing Xue Za Zhi. 2005 Jun;26(6): 421-3. Chinese.
[9] Gubler DJ, Kuno G, Sather GE, Velez M, OliverA. Mosquito cell cultures and specificmonoclonal antibodies in surveillance fordengue viruses. Am J Trop Med Hyg. 1984 Jan;33(1): 158-65.

18 Dengue Bulletin – Volume 31, 2007
Dengue fever outbreak in Guangzhou, China – 2002-2006
[10] Deubel V, Laille M, Hugnot JP, Chungue E,Guesdon JL, Drouet MT, Bassot S, Chevrier D.Identification of dengue sequences by genomicamplification: rapid diagnosis of dengue virusserotypes in peripheral blood. J Virol Methods.1990 Oct; 30(1): 41-54.
[11] Kuno G. Factors influencing the transmissionof dengue viruses. In: Gubler DJ, Kuno G,editors. Dengue and dengue haemorrhagic fever.New York: CAB International; 1997. p. 61-88.
[12] Brown TU, Babb K, Nimrod M, CarringtonCVF, Salas RA, Monteil MA. A retrospectivestudy of the 1996 DENV-1 epidemic inTrinidad: demographic and clinical features.Dengue Bulletin. 2004; 28: 7-19.
[13] Focks DA, Chadee DD. Pupal survey: anepidemiologically significant surveillancemethod for Aedes aegypti: an example usingdata from Trinidad. Am J Trop Med Hyg. 1997Feb; 56(2): 159-67.
[14] Guzmán MG, Kouri G, Valdes L, Bravo J,Alvarez M, Vazques S, Delgado I, Halstead SB.Epidemiologic studies on Dengue in Santiagode Cuba, 1997. Am J Epidemiol. 2000 Nov 1;152(9): 793-9; discussion 804.
[15] Shepard DS, Suaya JA, Halstead SB, NathanMB, Gubler DJ, Mahoney RT, Wang DN,Meltzer MI. Cost-effectiveness of a pediatricdengue vaccine. Vaccine. 2004 Mar 12;22(9-10):1275-80.
[16] Zhang J, Jian R, Wan YJ, Peng T, An J.Identification and phylogenetic analysis ofDENV-1 virus isolated in Guangzhou, China,in 2002. Dengue Bulletin. 2004; 28: 135-144.
[17] Kalayanarooj S, Nimmannitya S. Clinical andlaboratory presentations of dengue patientswith different serotypes. Dengue Bulletin.2000; 24:53-59.
[18] Wichmann O, Gascon J, Schunk M, PuenteS, Siikamaki H, Gjørup I, Lopez-Velez R, ClerinxJ, Peyerl-Hoffmann G, Sundøy A, Genton B,Kern P, Calleri G, de Górgolas M, MühlbergerN, Jelinek T; European Network onSurveillance of Imported Infectious Diseases.Severe dengue virus infection in travelers: riskfactors and laboratory indicators. J Infect Dis.2007 Apr 15; 195(8): 1089-96.
[19] Sumarmo, Wulur H, Jahja E, Gubler DJ,Sutomenggolo TS, Saroso JS. Encephalopathyassociated with dengue infection. Lancet. 1978Feb 25; 1(8061): 449-50.
[20] Seet RC, Lim EC, Wilder-Smith EP. Acutetransverse myelitis following dengue virusinfection. J Clin Virol. 2006 Mar; 35(3): 310-2.
[21] Misra UK, Kalita J, Syam UK, Dhole TN.Neurological manifestations of dengue virusinfection. J Neurol Sci. 2006 May 15; 244(1-2): 117-22.
[22] Poovorawan Y, Hutagalung Y, ChongsrisawatV, Boudville I, Bock HL. Dengue virus infection:a major cause of acute hepatic failure in Thaichildren. Ann Trop Paediatr. 2006 Mar; 26(1):17-23.
[23] Chan DP, Teoh SC, Tan CS, Nah GK,Rajagopalan R, Prabhakaragupta MK, CheeCK, Lim TH, Goh KY; The Eye Institute Dengue-Related Ophthalmic ComplicationsWorkgroup. Ophthalmic complications ofdengue. Emerg Infect Dis. 2006 Feb; 12(2):285-9.
[24] Rigau-Pérez JG. Severe dengue: the need fornew case definitions. Lancet Infect Dis. 2006May; 6(5): 297-302.

Dengue Bulletin – Volume 31, 2007 19
Circulation of dengue serotypes in five provinces ofnorthern Thailand during 2002-2006
Punnarai Veeraseatakul , Boonrat Wongchompoo, Somkhid Thichak,Yuddhakarn Yananto, Jarurin Waneesorn and Salakchit Chutipongvivate
Clinical Pathology Section, Regional Medical Sciences Centre Chiangmai, Department of Medical Sciences,Ministry of Public Health, 191 M.8 T. Donkaew, Maerim District, Chiangmai 50180, Thailand
Abstract
Dengue haemorrhagic fever is an epidemic infectious diseases caused by dengue virus. It is a majordisease prevalent in all provinces of Thailand. This study was to determine the circulating dengueserotypes by reverse transcription polymerase chain reaction (RT-PCR). A total of 1116 seropositiveacute samples were analysed from DF/DHF patients in five provinces of northern Thailand (Chiangmai,Lampang, Lamphun, Mae Hong Son and Phrae) during the period January 2002 to December 2006.Five hundred and fifty-nine samples were found positive, of which 47.2%, 30.6%, 18.4% and 3.8%were affected with DENV-2, DENV-1, DENV-4 and DENV-3 respectively. From 2002 to 2005, thepredominant dengue serotype was DENV-2, whereas DENV-1 was predominant in 2006. There was anapparent increase in the percentage of DENV-4 from 2005 to 2006. Our results indicated that all fourdengue serotypes were circulating in this region and the annual change of predominant serotypes wasthe cause of the severity of the disease.
Keywords: Dengue haemorrhagic fever; Dengue serotype; Northern Thailand.
E-mail: [email protected], [email protected]; Fax: 66-53-112192
Introduction
Dengue is a mosquito-borne viral infectioncaused by four distinct dengue virus serotypesDENV-1–4. It is the most prevalent arbovirusin tropical and subtropical regions of Africa, theAmericas, the Eastern Mediterranean, theWestern Pacific and South-East Asia.[1,2] InThailand, the first dengue outbreak occurredin Bangkok in 1958, initially in a pattern with a2-year cycle, and subsequently in irregularcycles, as the disease spread throughout thecountry. The largest outbreak was reported in1987, with an incidence rate of 325 cases/100 000 population. In recent years,
increasingly larger dengue outbreaks haveoccurred. There were 99 410, 127 189 and114 800 cases of dengue reported to theBureau of Epidemiology in 1997, 1998 and2002 respectively.[3] In 1999, the Ministry ofPublic Health initiated a dengue controlprogramme to reduce the incidence rate toless than 50 cases/100 000 population.[4] Innorthern Thailand, many provinces are locatedapproximately 250–300 metres above the sealevel, with some urban areas and large ruralareas. There were 13 915, 11 092, 6147, 6992and 6914 dengue cases reported during thefive-year period 2002–2006, with incidencerates of 119.4, 91.5, 51.3, 58.9 and 58.1 cases/

20 Dengue Bulletin – Volume 31, 2007
Circulation of dengue serotypes in Thailand
100 000 population respectively,[5-8] with 120deaths.
Gubler et al.[9] and Lam et al.[10] reportedthat virological surveillance, which involvesmonitoring of dengue virus infection in humans,has been used as an early warning system topredict outbreaks. Such surveillance, based onthe isolation and identification of dengueviruses infecting the human population,provides an important means of early detectionof any change in the prevalence of denguevirus serotypes. Each dengue serotype hascharacteristics that affect the nature of dengueepidemic and disease severity. Nisalak et al.[11]
reported that the predominant dengue serotypein the outbreaks in Bangkok during 1997–1998was DENV-3. Anantapreecha et al.[12] detectedthe predominant serotypes DENV-1 and DENV-2 in six provinces across Thailand during 2001–2002. However, dengue serotypes in fiveprovinces of northern Thailand have not yetbeen well elucidated, thus the present study,which is aimed at clarifying the pattern ofcirculating dengue serotypes in this region witha view to better understand theepidemiological complexities of the epidemicsof dengue infection.
Materials and methods
Patients
A total of 1116 seropositive acute samples withpositive anti-dengue IgM antibody wereanalysed for dengue serotype by RT-PCR atthe Regional Medical Sciences Centre,Chiangmai (RMSc_CM), Thailand. Thesesamples were collected from DF/DHF patientsin five northern provinces that includedChiangmai, Lamphun, Lampang, Mae HongSon and Phrae during 2002–2006 (Figure 1).
Figure 1: Map of five provinces of northernThailand
Laos
North
Myanmar
Central
Northeast
East
South
Malaysia
Mac Hong Son
Chiang MaiLampang
LamphunPhrae
RNA extraction
Viral RNA was isolated by using QIAamp viralRNA Mini Kit (QIAGEN, Cat. no. 52904).Briefly, the serum (100 μl) was added andmixed with 400 μl of AVL/RNA carrier solution(lysis buffer). After incubation at roomtemperature for 10 min, 400 μl of absoluteehtanol was added to the solution. All thesolutions were then transferred to a spincolumn and were spun at 8000 rpm for 1 min.The RNA was then washed by adding 500 μlof AW1 (washing buffer 1) and spun at 8000rpm for 1 min and following the sameprocedure with AW2 (washing buffer 2). Finally,the RNA was eluted by adding 60 μl of elutionbuffer and spun at 8000 rpm for 1 min. Theeluted RNA was kept in –70 °C until use.
RT-PCR
RT-PCR method was performed as previouslydescribed by Yenchitsomanus et al.[13] Briefly,

Circulation of dengue serotypes in Thailand
Dengue Bulletin – Volume 31, 2007 21
5 μl of RNA solution was mixed with reagentsof one step RT-PCR kit (QIAGEN, Cat. no.210212) and specific oligonucleotide primersof dengue-envelope (E) gene; DUL and DUR.The cDNA synthesis was performed at 50 °Cfor 30 min in a thermal cycler (MJ researchPTC-100, USA) followed by 40 cycles of PCRstep including 94 °C for 5 min, 94 °C for 1 min,45 °C for 1 min and 72 °C (5 min for the lastcycle). 1 μl of the primary PCR product wasused as the template for the second PCR withfour serotype-specific primer pairs; D1L, D1R,D2L, D2R, D3L, D3R, D4L and D4R (Table1). The PCR step was the same as above withthe annealing temperature set at 62 °C.Negative and positive dengue controls wereused. The secondary PCR products wereanalysed in 2% agarose gel electrophoresis andthen visualized by ethidium bromide staining.
Results and discussion
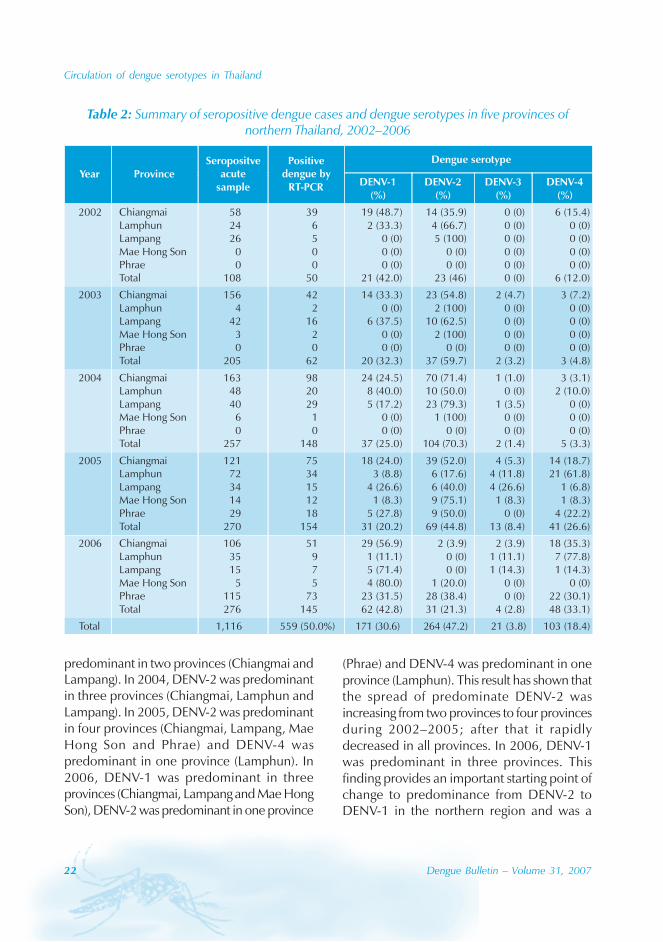
The number of seropositive acute cases anddengue serotypes in the five provinces during2002-2006 are shown in Table 2. Five hundredand fifty-nine dengue viral cases were detectedwith an average positivity rate of 50.0% by RT-PCR. All the four dengue serotypes weredetected during this study. The total numbersof positive dengue cases were analysed. DENV-
Table 1: Primer sequences of dengue virusand serotyping by RT-PCR
Primer Sequence 5’- 3’DUL GCTGTGTCACCCAGAGTGGCCATDUR TGGCTGGTGCACAGACAATGGTTD1L GGGGCTTCAACATCCCAAGAGD1R GCTTAGTTTCAAAGCTTTTTCACD2L ATCCAGATGTCATCAGGAAACD2R CCGGCTCTACTCCTATGATGD3L CAATGTGCTTGAATACCTTTGTD3R GGACAGGCTCCTCCTTCTTGD4L GGACAACAGTGGTGAAAGTCAD4R GGTTACACTGTTGGTATTCTCA
2 was the predominant serotype (47.2% cases),followed by DENV-1 (30.6% cases), DENV-4(18.4% cases) and DENV-3 (3.8% cases).
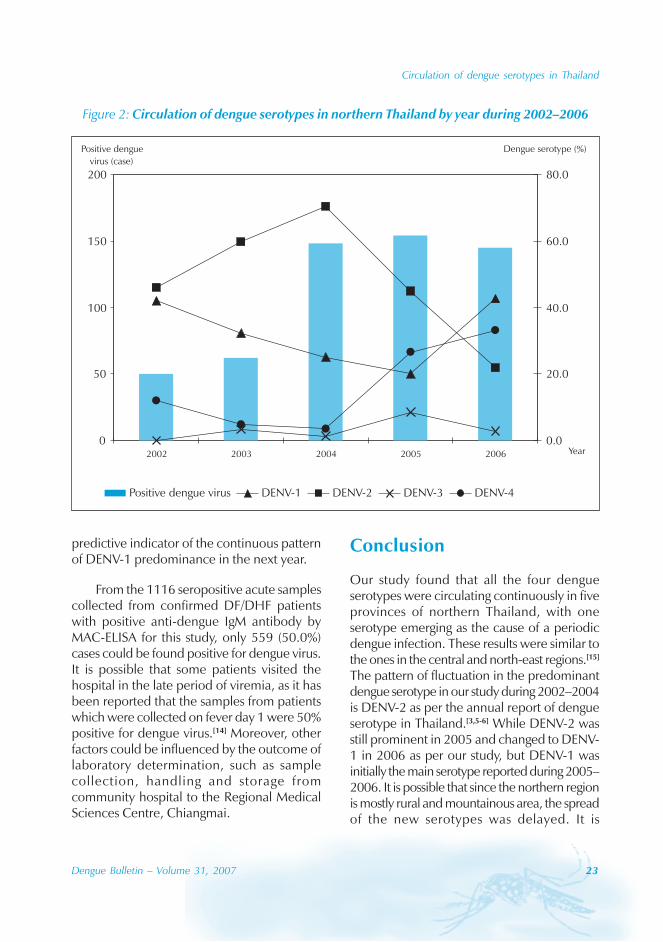
The circulation of dengue serotypes in northernThailand by year during 2002-2006 is shown inFigure 2. From 2002 to 2005, the predominantserotype was DENV-2 as 46.0%, 59.7%, 70.3%and 44.8% of cases were of this serotype,respectively, followed by DENV-1 (42.0%,32.3% and 25.0% cases) from 2002 to 2004respectively and DENV-4 (26.6% cases) in 2005.In 2006, the predominant serotype hadchanged to DENV-1 (42.8% cases), followedby DENV-4 (33.1% cases). DENV-3 was foundto be the least circulating serotype during 2003to 2006 and not found at all in 2002.
The distribution of the predominant dengueserotypes by provinces were analysed whenmore than five positive dengue virus sampleswere detected. In Chiangmai province, thepredominant serotypes were DENV-1 (48.7%and 56.9% cases) in 2002 and 2006, and DENV-2 (54.8%, 71.4% and 52.0% cases) in 2003,2004 and 2005 respectively. In Lampangprovince, the predominant serotypes wereDENV-2 (100.0%, 62.5%, 79.3% and 40.0%cases) from 2002 to 2005 respectively andDENV-1 (72.0% cases) in 2006. In Lamphunprovince, the predominant serotypes wereDENV-2 (66.7% and 50.0% cases) in 2002 and2004, and DENV-4 (61.8% and 77.8% cases)in 2005 and 2006. In Mae Hong Son province,the predominant serotypes were DENV-2(75.1% cases) in 2005 and DENV-1 (80.0%cases) in 2006. In Phrae province, thepredominant serotype was DENV-2 (50.0% and38.4% cases) in 2005 and 2006.
The comparison of the predominant dengueserotypes among the five provinces by yearwas analysed. In 2002, DENV-1 waspredominant in one province (Chiangmai) andDENV-2 was predominant in two provinces(Lamphun and Lampang). In 2003, DENV-2 was
22 Dengue Bulletin – Volume 31, 2007
Circulation of dengue serotypes in Thailand
predominant in two provinces (Chiangmai andLampang). In 2004, DENV-2 was predominantin three provinces (Chiangmai, Lamphun andLampang). In 2005, DENV-2 was predominantin four provinces (Chiangmai, Lampang, MaeHong Son and Phrae) and DENV-4 waspredominant in one province (Lamphun). In2006, DENV-1 was predominant in threeprovinces (Chiangmai, Lampang and Mae HongSon), DENV-2 was predominant in one province
(Phrae) and DENV-4 was predominant in oneprovince (Lamphun). This result has shown thatthe spread of predominate DENV-2 wasincreasing from two provinces to four provincesduring 2002–2005; after that it rapidlydecreased in all provinces. In 2006, DENV-1was predominant in three provinces. Thisfinding provides an important starting point ofchange to predominance from DENV-2 toDENV-1 in the northern region and was a
Table 2: Summary of seropositive dengue cases and dengue serotypes in five provinces ofnorthern Thailand, 2002–2006
Circulation of dengue serotypes in Thailand
Dengue Bulletin – Volume 31, 2007 23
predictive indicator of the continuous patternof DENV-1 predominance in the next year.
From the 1116 seropositive acute samplescollected from confirmed DF/DHF patientswith positive anti-dengue IgM antibody byMAC-ELISA for this study, only 559 (50.0%)cases could be found positive for dengue virus.It is possible that some patients visited thehospital in the late period of viremia, as it hasbeen reported that the samples from patientswhich were collected on fever day 1 were 50%positive for dengue virus.[14] Moreover, otherfactors could be influenced by the outcome oflaboratory determination, such as samplecollection, handling and storage fromcommunity hospital to the Regional MedicalSciences Centre, Chiangmai.
Figure 2: Circulation of dengue serotypes in northern Thailand by year during 2002–2006
0
50
100
150
200
2002 2003 2004 2005 2006 Year
Positive dengue
virus (case)
0.0
20.0
40.0
60.0
80.0
Dengue serotype (%)
Positive dengue virus DENV-1 DENV-2 DENV-3 DENV-4
Conclusion
Our study found that all the four dengueserotypes were circulating continuously in fiveprovinces of northern Thailand, with oneserotype emerging as the cause of a periodicdengue infection. These results were similar tothe ones in the central and north-east regions.[15]
The pattern of fluctuation in the predominantdengue serotype in our study during 2002–2004is DENV-2 as per the annual report of dengueserotype in Thailand.[3,5-6] While DENV-2 wasstill prominent in 2005 and changed to DENV-1 in 2006 as per our study, but DENV-1 wasinitially the main serotype reported during 2005–2006. It is possible that since the northern regionis mostly rural and mountainous area, the spreadof the new serotypes was delayed. It is

24 Dengue Bulletin – Volume 31, 2007
Circulation of dengue serotypes in Thailand
concordant that DENV-4 was found increasingduring 2005–2006, which was first detected inAugust 2005 (data not shown), whereas areported case with DENV-4 was initially detectedin Bangkok in April 2005. Our study found thatDENV-3 was the least reported serotype.Nisalak et al. reported that DENV-3 was theleast predominant dengue serotype in Bangkokin 1997–1998.[11]
Our study has shown the pattern of denguevirus serotypes in five provinces of northernThailand from year to year and provided someinsight into the dengue epidemic situation inthis region. This information should bebeneficial in the long-term dengue surveillance,
and future work can focus on using this patternof dengue serotype circulation to develop apredictive model of DF/DHF in Thailand.
Acknowledgements
We thank Ms Surapee Anantapreecha, Chiefof Arbovirus Laboratory at Thai NIH forproviding dengue control isolate. We also thankMs Sutheewan Sriuprayo, Director of theRegional Medical Sciences Centre, Chiangmai,for her support. This work was supported by agrant from the Regional Medical SciencesCentre, Chiangmai, Department of MedicalSciences, Ministry of Public Health, Thailand.
References
[1] Gubler DJ. Dengue and dengue hemorrhagicfever. Clin Microbiol Rev. 1998 Jul; 11(3): 480-96.
[2] Chusak P, Andjaparidze AG, Kumar V. Currentstatus of DF/DHF in WHO South-East AsiaRegion. Dengue Bulletin. 1993; 22: 1-11.
[3] Ministry of Public Health, Thailand. Annualepidemiological surveillance report 2002.Nonthaburi: Bureau of Epidemiology,Department of Disease Control, 2002.
[4] Kantachuvessiri A. Dengue hemorrhagic feverin Thai society. Southeast Asian J Trop MedPublic Health. 2002 Mar;33(1): 56-62.
[5] Ministry of Public Health, Thailand. Annualepidemiological surveillance report 2003.Nonthaburi: Bureau of Epidemiology,Department of Disease Control, 2003.
[6] Ministry of Public Health, Thailand. Annualepidemiological surveillance report 2004.Nonthaburi: Bureau of Epidemiology,Department of Disease Control, 2004.
[7] Ministry of Public Health, hailand. Annualepidemiological surveillance report 2005.
Nonthaburi: Bureau of Epidemiology,Department of Disease Control, 2005.
[8] Ministry of Public Health, Department ofDisease Control, Bureau of Epidemiology,Thailand. Weekly epidemiological surveillancereport. 2006; 37: 898-99.
[9] Gubler DJ. Surveillance for dengue anddengue hemorrhagic fever. Bull Pan Am HealthOrgan. 1989; 23(4): 397-404.
[10] Lam SK. Two decades of dengue in Malaysia.Trop Med. 1993; 35: 195-200.
[11] Nisalak A, Endy TP, Nimmannitya S,Kalayanarooj S, Thisayakorn U, Scott RM,Burke DS, Hoke CH, Innis BL, Vaughn DW.Serotype-specific dengue virus circulation anddengue disease in Bangkok, Thailand from1973 to 1999. Am J Trop Med Hyg. 2003 Feb;68(2): 91-202.
[12] Anantapreecha S, Chanama S, A-nuegoonpipat A, Naemkhunthot S, Sa-ngasang A, Sawanpanyalert P, Kurane I. Annualchanges of predominant dengue virusserotypes in six regional hospitals in Thailandfrom 1999 to 2002. Dengue Bulletin. 2004;28: 1-6.

Circulation of dengue serotypes in Thailand
Dengue Bulletin – Volume 31, 2007 25
[13] Yenchitsomanus PT, Sricharoen P, JaruthasanaI, Pattanakitsakul SN, Nitayaphan S,Mongkolsapaya J, Malasit P. Rapid detectionand identification of dengue viruses bypolymerase chain reaction (PCR). SoutheastAsian J Trop Med Public Health. 1996 Jun;27(2): 228-36.
[14] Sa-ngasang A, Wibulwattanakij S, ChanamaS, O-rapinpatipat A, A-nuegoonpipat A,
Anantapreecha S, Sawanpanyalert P, KuraneI. Evaluation of RT-PCR as a tool for diagnosisof secondary dengue virus infection. Jpn J InfectDis. 2003 Oct-Dec; 56(5-6): 205-9.
[15] Dengue serotype in Thailand (cited at 5 August2007). Available from: URL: http://www.cclts.org/dengue/index.asp

26 Dengue Bulletin – Volume 31, 2007
Clinical diagnostic delays and epidemiology of denguefever during the 2002 outbreak in Colima, Mexico
Gerardo Chowella,b , Porfirio Díaz-Dueñasc, Diego Chowelld, Sarah Hewse,Gabriel Ceja-Espírituf, James M. Hymanb and Carlos Castillo-Chaveze
aSchool of Human Evolution and Social Change, Arizona State University, Box 872402,Tempe, AZ 85287, USA
bCenter for Nonlinear Studies & Mathematical Modeling and Analysis (MS B284),Los Alamos National Laboratory, Los Alamos, NM 87545, USA
cHospital General de Medicina Familiar No. 1. Instituto Mexicano del Seguro Social (IMSS),Colima, Col., Mexico
dSchool of Sciences, Universidad de Colima, Bernal Díaz del Castillo No. 340,C.P. 28045 Colima, Col., Mexico
eDepartment of Mathematics and Statistics, Arizona State University, P.O. Box 871804,Tempe, AZ 85287-1804, USA
fSchool of Medicine, Universidad de Colima, Colima, Col., Mexico
Abstract
Dengue fever is a re-emergent and challenging public health problem in the world. Here, we assessretrospectively the epidemiological and clinical characteristics of the 2002 dengue epidemic in thestate of Colima, Mexico. This study is carried out by analysing a database containing demographic,epidemiological and clinical information. Of the 4040 clinical dengue cases diagnosed in the hospitalsof the Mexican Institute of Public Health in the state of Colima, 548 cases were confirmed by laboratorytests, and 495 cases presented at least one haemorrhagic manifestation. Of the total clinically diagnosedcases, the most common symptoms observed were: fever (99.6%), headache (92.4%), myalgia (89.4%)and arthralgia (88.6%). The most common haemorrhagic manifestations were: petechiae (7.1%), gingivitis(3.4%) and epistaxis (3.6%). The median time between the onset of illness and visit to the health careclinic (diagnostic delay) was 1 day (interquartile range [IQR]: 0-3). For cases presenting haemorrhagicmanifestations, the diagnostic delay was higher (median: 2 days, IQR: 0-4) than for non-haemorrhagiccases (median: 1 day, IQR: 0-3). The proportion of males presenting haemorrhagic manifestations washigher than females (Fisher Exact test; p<0.01). Moreover, the age group 0-5 years presented a lowerproportion of cases with haemorrhagic manifestations compared with the age group of 6 years andolder (p=0.0281). No significant differences were found between the diagnostic delays in the case ofmales and females.
Keywords: Dengue fever; Clinical diagnostic delay; Haemorrhagic; Colima; Mexico.
E-mail: [email protected] Phone: 480-965-4730 Fax: 480-965-7671

Clinical diagnostic delays and dengue fever in Colima, Mexico (2002)
Dengue Bulletin – Volume 31, 2007 27
Introduction
Annually there are approximately 100 millioncases of dengue worldwide.[1] It is endemic inAfrica, the Americas, the EasternMediterranean, South-East Asia and theWestern Pacific regions. The challenges seemdaunting. For example, in Singapore, dengueis a major public health problem despite thefact that a set of extraordinary control effortshave been put into place over the past fewyears.[2] The etiological agent is a Flavivirus withfour different serotypes (DENV-1–4). Theprimary vectors of dengue are mosquitoes ofthe species Aedes aegypti and Aedes albopictus.Humans are infected when bitten by feedinginfectious females. Those who recover maybecome permanently immune to the serotypeinvolved and partially immune to the otherserotypes.[3] Susceptible vectors acquire theinfection when feeding on infectious humans.Female mosquitoes are responsible for thetransmission of the virus since males are non-blood suckers and feed primarily on plants andflowers.[4]
Cases of dengue are classified asasymptomatic, clinically non-specific flu-likesymptoms, dengue fever (DF), denguehaemorrhagic fever (DHF) and dengue shocksyndrome (DSS).[5] DHF and DSS are the severeforms of the disease. Dengue attack rates varyfrom 40% to 50% but may be as high as 80%to 90%.[6]
Mexico was declared dengue-free whenthe principal vector, Ae. aegypti, was eliminatedin 1963. However, Ae. aegypti reappeared twoyears later and the disease returned.[7] All thefour dengue serotypes are now circulating inMexico since 1980.[7] DHF has become apublic health problem in the country since1994.[8] In 2002, over 30 Latin Americancountries reported a total of over a million casesof classical dengue as well as more than 17 000
cases of DHF.[9] We report the results of aretrospective study on the epidemiological andclinical characteristics of the 2002 (serotypeDENV-2) dengue epidemic in the state ofColima, Mexico.[10]
Materials and methods
The epidemiological and clinical characteristicsof dengue fever cases recorded during the2002 epidemic (January through December)in the state of Colima have been studied. Thestate of Colima is located on the Pacific coast,has a tropical climate, and has a population of488 028 (Figure 1).[11] The data used includecases diagnosed at the hospitals of the MexicanInstitute of Public Health (IMSS). The IMSS area collection of state hospitals that provideprimary health services to 60% of thepopulation in the state. The remainingpopulation receives health care services fromthe hospitals of the Mexican Health Ministryand the private sector.
Figure 1: Map of the state of Colima,Mexico, with divisions by municipalities
Armería (1), Colima (2), Comala (3),Coquimatlán (4), Cuauhtémoc (5),
Ixtlahuacán (6), Manzanillo (7), Minatitlán(8), Tecomán (9), Villa de Alvarez (10)

28 Dengue Bulletin – Volume 31, 2007
Clinical diagnostic delays and dengue fever in Colima, Mexico (2002)
Patient record data are stratified bymunicipality where the dengue cases werediagnosed (Figure 1); the week of symptomonset and diagnosis; the IgM antibody testresult; patient’s age and gender; and diagnosticdelay (as presented later in the paper).Furthermore, non-haemorrhagic recordedsymptoms included: fever, headache, myalgia,arthralgia, retro-orbital pain, exanthema,diarrhoea, vomit, nausea, pruritus, chills,photophobia, abdominal pain, conjunctivitis,nasal decongestion, cough, hepatomegaly andsplenomegaly. Haemorrhagic recordedsymptoms included: petechiae, ecchymosis,ascites, pleural effusion, gingivitis, epistaxis,haematemesis and melena. We classified apatient as “delayed” when clinical diagnosis wasmade two days after the onset of symptoms.
The World Health Organization (WHO)case definition of probable dengue cases[3]
requires the presence of fever or chills and atleast two symptoms from: myalgia, arthralgia,retro-orbtial pain, headache, rash, or somehaemorrhagic manifestation (e.g. petechiae,haematuria, haematemesis, melena). Thelaboratory testing was only carried out in a smallsubset of the clinically diagnosed dengue casesthrough anti-dengue IgM antibody tests (ELISA).
We classified cases of DHF following asclosely as possible all the four requirementsfor the WHO definition of DHF (fever,haemorrhage, thrombocytopenia, and signs ofplasma leakage).[6] Difficulties arose becausebasic measurements, such as platelet counts,were conducted in only 1227 (30%) of thecases and signs of plasma leakage were assessedonly clinically (pleural effusion and/or ascites).The difficulties in characterizing DHF casesusing the WHO system have led someinvestigators to use modified classificationschemes.[12,13] Here, we used the number ofhaemorrhagic symptoms as a measure ofdisease severity and classify dengue patientsas haemorrhagic whenever one haemorrhagicsymptom was reported. Dengue cases that did
not exhibit haemorrhagic manifestations wereclassified as non-haemorrhagic.
We characterize variations using the meanand standard deviation (SD). Proportions arecompared using Fisher’s exact test ofindependence, which uses a hypergeometricsampling distribution for cell frequencies.[14]
Population distributions are compared using thenon-parametric Wilcoxon test. Results aredeemed significant when the p value is lessthan 0.05. Some records are not complete.For example, the date of symptom onset wasonly recorded in 2242 patient records. Hence,the number of records (denoted by N) used isincluded in the analyses.
Results
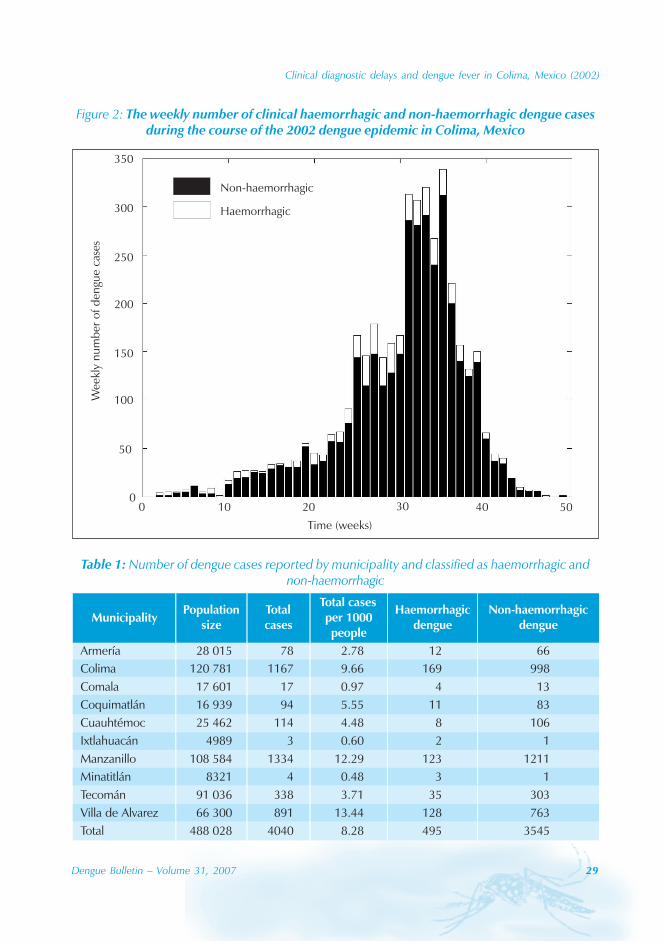
The 2002 dengue epidemic in Colima, Mexico,began in January, peaked in September anddied out in December (Figure 2). Fourthousand and forty cases were clinicallydiagnosed, including 495 cases withhaemorrhagic manifestations. A total of 555clinical dengue cases (14%) were subjectedto ELISA test and 548 cases were positive foranti-dengue IgM antibodies. Thrombocytopeniawas detected in 528 cases but platelet countswere only recorded for 1227 cases (30%). Boththrombocytopenia and haemorrhagicsymptoms were present in 203 cases (17%,N=1227), but only 8 cases could be classifiedas DHF under the WHO classification. Thedistribution of dengue cases by municipality isgiven in Table 1. The mean age of a denguecase was 24.61 ± 16.30 (SD) years (Figure3A). The attack rate was about 9.5 denguecases per 1000 persons. This rate was reportedamong two age groups of 5-14 and 25-34 years.The male/female ratio was about 1:1 with 2045(50.6%) males and 1993 (49.4%) females. Wefound no significant difference between theage distribution of non-haemorrhagic andhaemorrhagic cases (Wilcoxon test, p=0.5018,N=4007).
Clinical diagnostic delays and dengue fever in Colima, Mexico (2002)
Dengue Bulletin – Volume 31, 2007 29
Figure 2: The weekly number of clinical haemorrhagic and non-haemorrhagic dengue casesduring the course of the 2002 dengue epidemic in Colima, Mexico
Non-haemorrhagic
Haemorrhagic
350
300
250
200
150
100
50
Wee
kly
nu
mb
erof
den
gue
case
s
Time (weeks)
10 20 30 40 5000
Table 1: Number of dengue cases reported by municipality and classified as haemorrhagic andnon-haemorrhagic
30 Dengue Bulletin – Volume 31, 2007
Clinical diagnostic delays and dengue fever in Colima, Mexico (2002)
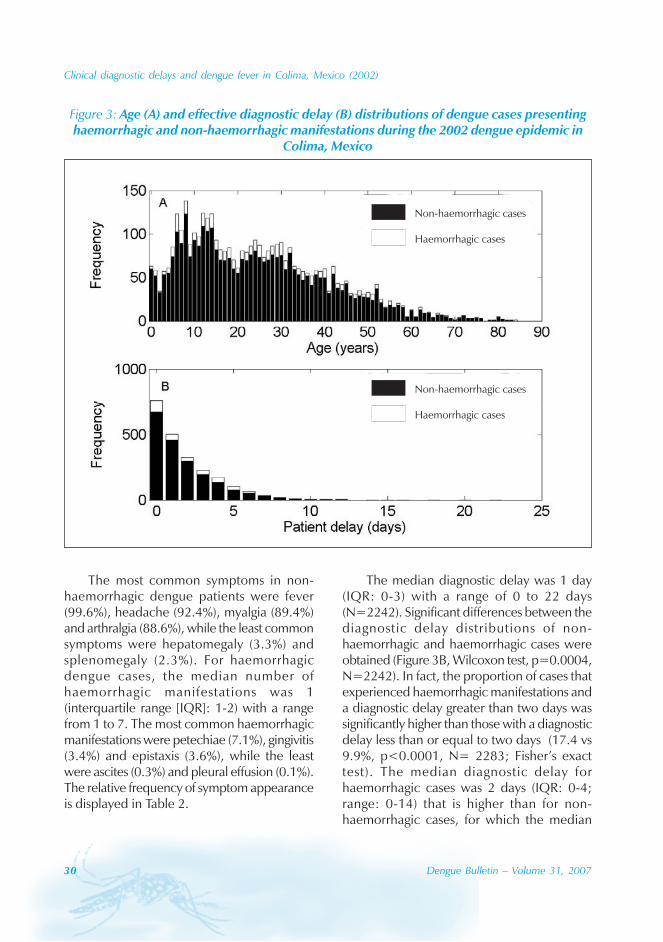
Figure 3: Age (A) and effective diagnostic delay (B) distributions of dengue cases presentinghaemorrhagic and non-haemorrhagic manifestations during the 2002 dengue epidemic in
Colima, Mexico
The most common symptoms in non-haemorrhagic dengue patients were fever(99.6%), headache (92.4%), myalgia (89.4%)and arthralgia (88.6%), while the least commonsymptoms were hepatomegaly (3.3%) andsplenomegaly (2.3%). For haemorrhagicdengue cases, the median number ofhaemorrhagic manifestations was 1(interquartile range [IQR]: 1-2) with a rangefrom 1 to 7. The most common haemorrhagicmanifestations were petechiae (7.1%), gingivitis(3.4%) and epistaxis (3.6%), while the leastwere ascites (0.3%) and pleural effusion (0.1%).The relative frequency of symptom appearanceis displayed in Table 2.
The median diagnostic delay was 1 day(IQR: 0-3) with a range of 0 to 22 days(N=2242). Significant differences between thediagnostic delay distributions of non-haemorrhagic and haemorrhagic cases wereobtained (Figure 3B, Wilcoxon test, p=0.0004,N=2242). In fact, the proportion of cases thatexperienced haemorrhagic manifestations anda diagnostic delay greater than two days wassignificantly higher than those with a diagnosticdelay less than or equal to two days (17.4 vs9.9%, p<0.0001, N= 2283; Fisher’s exacttest). The median diagnostic delay forhaemorrhagic cases was 2 days (IQR: 0-4;range: 0-14) that is higher than for non-haemorrhagic cases, for which the median
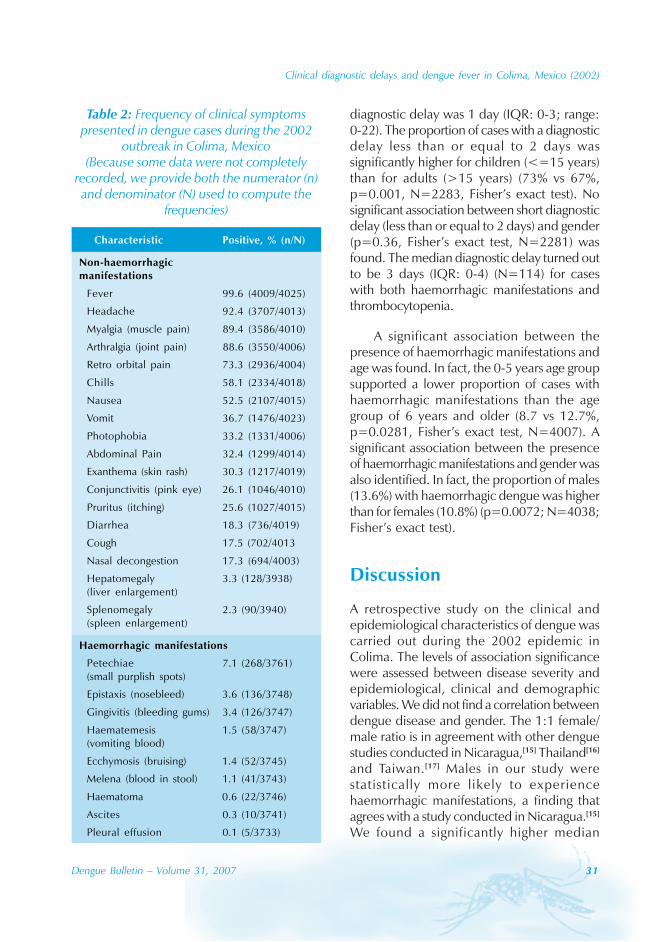
Clinical diagnostic delays and dengue fever in Colima, Mexico (2002)
Dengue Bulletin – Volume 31, 2007 31
diagnostic delay was 1 day (IQR: 0-3; range:0-22). The proportion of cases with a diagnosticdelay less than or equal to 2 days wassignificantly higher for children (<=15 years)than for adults (>15 years) (73% vs 67%,p=0.001, N=2283, Fisher’s exact test). Nosignificant association between short diagnosticdelay (less than or equal to 2 days) and gender(p=0.36, Fisher’s exact test, N=2281) wasfound. The median diagnostic delay turned outto be 3 days (IQR: 0-4) (N=114) for caseswith both haemorrhagic manifestations andthrombocytopenia.
A significant association between thepresence of haemorrhagic manifestations andage was found. In fact, the 0-5 years age groupsupported a lower proportion of cases withhaemorrhagic manifestations than the agegroup of 6 years and older (8.7 vs 12.7%,p=0.0281, Fisher’s exact test, N=4007). Asignificant association between the presenceof haemorrhagic manifestations and gender wasalso identified. In fact, the proportion of males(13.6%) with haemorrhagic dengue was higherthan for females (10.8%) (p=0.0072; N=4038;Fisher’s exact test).
Discussion
A retrospective study on the clinical andepidemiological characteristics of dengue wascarried out during the 2002 epidemic inColima. The levels of association significancewere assessed between disease severity andepidemiological, clinical and demographicvariables. We did not find a correlation betweendengue disease and gender. The 1:1 female/male ratio is in agreement with other denguestudies conducted in Nicaragua,[15] Thailand[16]
and Taiwan.[17] Males in our study werestatistically more likely to experiencehaemorrhagic manifestations, a finding thatagrees with a study conducted in Nicaragua.[15]
We found a significantly higher median
Table 2: Frequency of clinical symptomspresented in dengue cases during the 2002
outbreak in Colima, Mexico(Because some data were not completely
recorded, we provide both the numerator (n)and denominator (N) used to compute the
frequencies)
Characteristic Positive, % (n/N)
Non-haemorrhagicmanifestations
Fever 99.6 (4009/4025)
Headache 92.4 (3707/4013)
Myalgia (muscle pain) 89.4 (3586/4010)
Arthralgia (joint pain) 88.6 (3550/4006)
Retro orbital pain 73.3 (2936/4004)
Chills 58.1 (2334/4018)
Nausea 52.5 (2107/4015)
Vomit 36.7 (1476/4023)
Photophobia 33.2 (1331/4006)
Abdominal Pain 32.4 (1299/4014)
Exanthema (skin rash) 30.3 (1217/4019)
Conjunctivitis (pink eye) 26.1 (1046/4010)
Pruritus (itching) 25.6 (1027/4015)
Diarrhea 18.3 (736/4019)
Cough 17.5 (702/4013
Nasal decongestion 17.3 (694/4003)
Hepatomegaly 3.3 (128/3938)(liver enlargement)
Splenomegaly 2.3 (90/3940)(spleen enlargement)
Haemorrhagic manifestations
Petechiae 7.1 (268/3761)(small purplish spots)
Epistaxis (nosebleed) 3.6 (136/3748)
Gingivitis (bleeding gums) 3.4 (126/3747)
Haematemesis 1.5 (58/3747)(vomiting blood)
Ecchymosis (bruising) 1.4 (52/3745)
Melena (blood in stool) 1.1 (41/3743)
Haematoma 0.6 (22/3746)
Ascites 0.3 (10/3741)
Pleural effusion 0.1 (5/3733)

32 Dengue Bulletin – Volume 31, 2007
Clinical diagnostic delays and dengue fever in Colima, Mexico (2002)
diagnostic delay for cases presentinghaemorrhagic manifestations than for non-haemorrhagic cases.
Not surprisingly, this finding indicates animportant correlation between the disease’sclinical evolution and diagnostic delay.Diagnostic delays may complicate the clinicalstate of the patient while facilitating thetransmission of dengue in the population. Inother words, although interventions haveshown to be incapable of reverting a dengueepidemic,[18] reductions in the diagnostic delayscan reduce the final epidemic size.Mathematical models of dengue transmissionhave shown that educational campaigns aimingat shortening the diagnostic delays can lead tosignificant reductions in the final epidemicsize.[19] Control strategies that benefit from aprompt identification of dengue cases includethe use of nets and screens, the application ofinsecticides to clothing and the application ofmosquito repellents. Moreover, strategiesaiming at the elimination or reduction in thenumber of breeding sites through the use oflarvicidal control including ovitraps[20] andmalathion spraying for adult control can alsoreduce the dengue burden.
A fever alone is usually not enough tomotivate patients to seek medical care. It is onlythe presence of severe symptoms (e.g.haemorrhagic manifestations) that motivate toseek medical care. This observation may explainthe significant association that we foundbetween diagnostic delays and the presence ofhaemorrhagic manifestations. Specifically, wefound that the proportion of cases with adiagnostic delay of less than or equal to 2 dayswas significantly higher for children (<=15 years)than for adults (>15 years). This is in agreementwith the findings by Ahorlu et al.[21] who studiedthe socio-cultural determinants of treatmentdelay for childhood malaria in southern Ghana.Ahorlu et al.[21] found that families with similareconomic situations who have sick children will
be more likely to seek treatment. Genderexhibited no correlation with the length of thediagnostic delays. The most commonexplanations for diagnostic delays in malariastudies in sub-Saharan Africa include poverty andthe inability to pay for treatment.[22] Thisperspective could not be assessed here as mostof the patients comprised only the insuredindividuals by IMSS. In summary, a combinationof cultural factors, the inability to distinguishdengue illnesses from traditional fevers at theearly stages of the disease, and the differentiatedtreatment are the main factors behinddifferences in diagnostic delays.
The impact of dengue diagnostic delaydistributions and their association with clinical,demographic and epidemiological factors havenot received much attention. This contrasts, forexample, with epidemiological studies dealingwith other infectious diseases such as pulmonarytuberculosis.[23] However, there are somerelevant studies. For example, an estimate ofthe median diagnostic delay of 5 days has beenreported for the 1996 dengue epidemic innorth-eastern Brazil,[24] that is, a significantlylonger median diagnostic delay than ourestimate of 1 day. Guzman et al.[25] found thatthe average time from fever onset tohospitalization associated with the 1997 dengueoutbreak in Cuba was about 2.9 days. On theother hand, a median diagnostic delay of 2 dayswas reported for malaria-stricken travellersreturning to Sweden during 1994 to 2001.[26]
We found a lower proportion ofhaemorrhagic cases in the age group 0-5 yearsthan those in the age group of 6 years and older.Secondary dengue infections have beenassociated with the presence of haemorrhagicmanifestations, a phenomenon explained by thetheory of antibody-dependent enhancement.[5]
Seasonal effects are also critical. The peakof the epidemic in Colima occurred in mid-September, which correlates well with the

Clinical diagnostic delays and dengue fever in Colima, Mexico (2002)
Dengue Bulletin – Volume 31, 2007 33
peak in the rainfall.[27] A similar pattern hasbeen reported for other dengue outbreaks{Nicaragua (1998),[15] El Salvador (2000)[28] andBangladesh (2000)[29]}.
In contrast to dengue hyperendemic areaswhere dengue haemorrhagic fever is primarilya disease of children under 15 years, we founda lower proportion of haemorrhagic cases inthe age group 0-5 years compared to the agegroup of 6 years and older.
The number of dengue infections permunicipality was determined by the location ofthe patient’s address albeit the actual dengueinfection may have occurred at work becausedengue mosquitoes are daytime feeders.[30]
This study was limited by the lack ofcomplete medical records. Difficulties wereencountered in rigorously following the WHOclassification scheme for DHF, because thesigns of plasma leakage were assessed onlyclinically (pleural effusion and/or ascites). Thenumber of DHF cases may in fact be as highas 203, that is, the number of cases for whichthe other three requirements of the WHOclassification scheme for DHF (fever,haemorrhage and thrombocytopenia) weresatisfied. On the other hand, dengue cases withsevere haemorrhage, not accompanied byincreased vascular permeability, have beenreported from several regions of the world
including Indonesia, China, India, Philippines,Thailand, South Pacific and Latin America.[13]
To improve the tracking of dengue, workers atIMSS hospitals would need to collect ascomplete patient information as possible. Effortsto follow the WHO classification guidelines asclosely as possible would be quite helpful.Dengue symptoms can be confused in theepidemic and non-epidemic situations withother exanthematous and non-exanthematousviral diseases such as measles, rubella,enteroviruses and influenza.[31] We recognizethat there are limited resources for laboratorytesting. In the 2002 dengue epidemic inColima, only 14% of the clinical dengue caseswere tested in the laboratory for anti-dengueIgM. The low proportion of clinical denguecases tested serologically make it difficult tovalidate some of our findings.
Our findings indicate that an educationalcampaign in the community with the objectiveof informing the population about earlysymptoms of dengue infection could lead tonot only reductions in diagnostic delays but alsoto reduce the final size of a dengue epidemic.
Acknowledgement
Gerardo Chowell was supported by a Director’sPost-doctoral Fellowship from Los AlamosNational Laboratory.
References
[1] Kurane I, Takasaki T. Dengue fever and denguehaemorrhagic fever: challenges of controllingan enemy still at large. Rev Med Virol. 2001Sep-Oct; 11(5): 301-11.
[2] Singapore Ministry of Health. Thecommunicable disease surveillance inSingapore 2004. Available from: URL:http://www.moh.gov.sg/cmaweb/attachments/pub l i ca t ion /36c375c7%9a8t /Vec tor-Borne\_Diseases.pdf
[3] World Health Organization. Clinical diagnosisof dengue. Available from: URL:http://www.who.int/csr/resources/publications/dengue/012-23.pdf
[4] Scott TW, Chow E, Strickman D, KittayapongP, Wirtz RA, Lorenz LH, Edman JD. Blood-feeding patterns of Aedes aegypti (Diptera:Culicidae) collected in a rural Thai village. JMed Entomol. 1993 Sep; 30(5): 922-7.

34 Dengue Bulletin – Volume 31, 2007
Clinical diagnostic delays and dengue fever in Colima, Mexico (2002)
[5] Vaughn DW, Green S, Kalayanarooj S, InnisBL, Nimmannitya S, Suntayakorn S, Endy TP,Raengsakulrach B, Rothman AL, Ennis FA,Nisalak A. Dengue viremia titer, antibodyresponse pattern, and virus serotype correlatewith disease severity. J Infect Dis. 2000Jan;181(1):2-9.
[6] World Health Organization. Dengue anddengue hemorrhagic fever. Geneva: WHO,2008. Fact sheet No. 117. (http://www.who.int/mediacentre/factsheets/fs117/en/ - accessed 30 December 2008).
[7] Díaz FJ, Black WC 4th, Farfán-Ale JA, Loroño-Pino MA, Olson KE, Beaty BJ. Dengue viruscirculation and evolution in Mexico: aphylogenetic perspective. Arch Med Res. 2006Aug; 37(6): 760-73.
[8] Navarrete-Espinosa J, Gómez-Dantés H, Celis-Quintal JG, Vázquez-Martínez JL. Clinicalprofile of dengue hemorrhagic fever cases inMexico. Salud Publica Mex. 2005 May-Jun;47(3): 193-200.
[9] Guzman MG, Kouri G. Dengue and denguehemorrhagic fever in the Americas: lessons andchallenges. J Clin Virol. 2003 May; 27(1): 1-13.
[10] Espinoza-Gómez F, Díaz-Dueñas P, Torres-Lepe C, Cedillo-Nakay RA, Newton-SánchezOA. Clinical pattern of hospitalized patientsduring a dengue epidemic in Colima, Mexico.Dengue Bull. 2005; 29: 8-17.
[11] INEGI. National Institute of Geography andInformatics. 1995.
[12] Deen JL, Harris E, Wills B, Balmaseda A,Hammond SN, Rocha C, Dung NM, HungNT, Hien TT, Farrar JJ. The WHO dengueclassification and case definitions: time for areassessment. Lancet. 2006 Jul 8; 368(9530):170-3.
[13] Bandyopadhyay S, Lum LC, Kroeger A.Classifying dengue: a review of the difficultiesin using the WHO case classification fordengue haemorrhagic fever. Trop Med IntHealth. 2006 Aug; 11(8): 1238-55.
[14] Agresti A. An introduction to categorical dataanalysis. New York: Wiley, 1996.
[15] Harris E, Videa E, Pérez L, Sandoval E, Téllez Y,Pérez ML, Cuadra R, Rocha J, Idiaquez W,Alonso RE, Delgado MA, Campo LA, AcevedoF, Gonzalez A, Amador JJ, Balmaseda A.Clinical, epidemiologic, and virologic featuresof dengue in the 1998 epidemic in Nicaragua.Am J Trop Med Hyg. 2000 Jul-Aug; 63(1-2): 5-11.
[16] Wichmann O, Hongsiriwon S,Bowonwatanuwong C, Chotivanich K,Sukthana Y, Pukrittayakamee S. Risk factorsand clinical features associated with severedengue infection in adults and children duringthe 2001 epidemic in Chonburi, Thailand.Trop Med Int Health. 2004 Sep; 9(9): 1022-9.
[17] Chao DY, Lin TH, Hwang KP, Huang JH, LiuCC, King CC. 1998 dengue hemorrhagic feverepidemic in Taiwan. Emerg Infect Dis. 2004Mar; 10(3): 552-4.
[18] Gubler DJ. Aedes aegypti and Aedes aegypti-borne disease control in the 1990s: top downor bottom up. Charles Franklin Craig Lecture.Am J Trop Med Hyg. 1989 Jun; 40(6): 571-8.
[19] Chowell G, Diaz-Dueñas P, Miller JC, Alcazar-Velazco A, Hyman JM, Fenimore PW, Castillo-Chavez C. Estimation of the reproductionnumber of dengue fever from spatial epidemicdata. Math Biosci. 2007 Aug; 208(2): 571-89.
[20] Ooi EE, Goh KT, Gubler DJ. Dengueprevention and 35 years of vector control inSingapore. Emerg Infect Dis. 2006 Jun; 12(6):887-93.
[21] Ahorlu CK, Koram KA, Ahorlu C, de SavignyD, Weiss MG. Socio-cultural determinants oftreatment delay for childhood malaria insouthern Ghana. Trop Med Int Health. 2006Jul;11(7):1022-31.
[22] Williams HA, Jones CO. A critical review ofbehavioral issues related to malaria control insub-Saharan Africa: what contributions havesocial scientists made? Soc Sci Med. 2004 Aug;59(3): 501-23.

Clinical diagnostic delays and dengue fever in Colima, Mexico (2002)
Dengue Bulletin – Volume 31, 2007 35
[23] Chowell G, Diaz-Dueñas P, Chowell D. Thedynamics of pulmonary tuberculosis inColima, Mexico (1999-2002). Scand J InfectDis. 2005; 37(11-12): 858-62.
[24] Flannery B, Pereira MM, Velloso L de F,Carvalho C de C, De Codes LG, Orrico G deS, Dourado CM, Riley LW, Reis MG, Ko AI.Referral pattern of leptospirosis cases during alarge urban epidemic of dengue. Am J TropMed Hyg. 2001 Nov; 65(5): 657-63.
[25] Guzmán MG, Alvarez M, Rodríguez R, RosarioD, Vázquez S, Vald s L, Cabrera MV, Kourí G.Fatal dengue hemorrhagic fever in Cuba, 1997.Int J Infect Dis. 1999 Spring; 3(3): 130-5.
[26] Askling HH, Ekdahl K, Janzon R, HenricBraconier J, Bronner U, Hellgren U, Rombo L,Tegnell A. Travellers returning to Sweden withfalciparum malaria: pre-travel advice,behaviour, chemoprophylaxis and diagnosticdelay. Scand J Infect Dis. 2005; 37(10): 760-5.
[27] Chowell G, Sanchez F. Climate-baseddescriptive models of dengue fever: the 2002epidemic in Colima, Mexico. J Environ Health.2006 Jun; 68(10): 40-4, 55.
[28] Hayes JM, García-Rivera E, Flores-Reyna R,Suárez-Rangel G, Rodríguez-Mata T, Coto-Portillo R, Baltrons-Orellana R, Mendoza-Rodríguez E, De Garay BF, Jubis-Estrada J,Hernández-Argueta R, Biggerstaff BJ, Rigau-Pérez JG. Risk factors for infection during asevere dengue outbreak in El Salvador in 2000.Am J Trop Med Hyg. 2003 Dec; 69(6): 629-33.
[29] Rahman M, Rahman K, Siddque AK, ShomaS, Kamal AH, Ali KS, Nisaluk A, Breiman RF.First outbreak of dengue hemorrhagic fever,Bangladesh. Emerg Infect Dis. 2002 Jul; 8(7):738-40.
[30] Effler PV, Pang L, Kitsutani P, Vorndam V, NakataM, Ayers T, Elm J, Tom T, Reiter P, Rigau-PerezJG, Hayes JM, Mills K, Napier M, Clark GG,Gubler DJ; Hawaii Dengue OutbreakInvestigation Team. Dengue fever, Hawaii,2001-2002. Emerg Infect Dis. 2005 May;11(5): 742-9.
[31] George R, Lum LCS. Clinical spectrum ofdengue infection. In: Gubler DG and KunoG, editors. Dengue and dengue hemorrhagicfever. New York: CABI Publishing; 1997.

36 Dengue Bulletin – Volume 31, 2007
Exposure to the risk of dengue virus infection in anurban setting: ecological versus individual heterogeneity
Maria Glória Teixeiraa , Maurício Lima Barretoa, Maria da Conceição N. Costaa,Leila Denise Alves Ferreirab, Vanessa Moratoa, Pedro Fernando Vasconcelosc
and Sandy Cairncrossd
aInstituto de Saúde Coletiva/UFBA – Rua Basílio da Gama, s/n Campus Universitário, Canela,Salvador –Bahia CEP:41.110-170
bInstituto de Matemática da Universidade Federal da BahiacInstituto Evandro Chagas – SVS/Ministério da Saúde do Brasil
dLondon School of Hygiene and Tropical Medicine
Abstract
The dynamics of dengue virus circulation in the intra-urban spaces of large cities and the risk factors forthe occurrence of such infections are still not well known. Although it has been established that povertyis one of the determinants of the majority of infectious and parasitic diseases, in the case of dengue thisis still a matter of some controversy. This study had the objective of describing the distribution of dengueseroprevalence and seroincidence in different intra-urban spaces within a large and complex city innorth-eastern Brazil. The study investigated whether there is any relationship between the intensity ofvirus circulation and the population’s living conditions or between group immunity and Aedes aegyptiinfestation rates. The variability in the risk of such infections was also examined. A prospective study wasconducted by means of serological investigations among a sample of people living in 30 differentspaces (“sentinel areas”) in the city of Salvador, which was selected according to extreme differences inliving conditions. High rates of seroprevalence (67.7%) and seroincidence (70.6%) were found for thecirculating serotypes (DENV-1 and DENV-2). Similar to what has been occurring in south-east Asia, theseroincidence was high (55%) even when the group immunity had already been partially established(42%) and the Ae. aegypti infestation rates were relatively low (<3%). Contrary to the ecological analysis,at the individual level, substantial heterogeneity in dengue exposure was observed. This paper discussesthis apparent contradiction, highlighting its implications for the effectiveness of vector control strategies.
Keywords: Dengue; Prospective study; Herd immunity; Seroprevalence; Seroincidence; Spatial distribution;Aedes aegypti; Epidemiology.
E-mail: [email protected]; Tel (71) 32837412; Fax 32837460
Introduction
The dynamics and determinants of dengue viruscirculation in urban areas, particularly in largecities, are not well-established. Epidemiological
studies of dengue infections have beenneglected,[1] even though they are importantfor developing dengue prevention and controlstrategies, as has recently been recognized inthe dengue research agenda of WHO.[2]

Exposure to the risk of dengue virus infection in an urban setting
Dengue Bulletin – Volume 31, 2007 37
The re-emergence of this disease in severalcontinents and its potential virulence is of greatconcern to public health worldwide. Moreover,the available control strategies are based onvector control, and these have not always beeneffectively implemented to preventtransmission of the disease.[3,4] Unlike yellowfever, restrictions on human movementthrough quarantine are not applicable for thecontrol of dengue for lack of vaccines.
In the large urban centres that are infestedwith Aedes aegypti, large numbers ofsusceptible individuals and high populationdensity[3] probably facilitate more extensivetransmission of dengue virus as mosquitovectors, because of multiple probing/feedingbehaviours are likely to infect multipleindividuals in different households. However,the risk of exposure to dengue virus infectionin relation to the range of social and economicconditions in big cities remains unclear. Theserisk factors are related both to low income areasand areas with more favourable conditions.[5,6]
In Salvador, a large city in north-easternBrazil, the dengue virus was first detected inJanuary 1995, and, by the end of 1999, therehad been two recorded epidemics. The meanannual incidence over that period reached 691reported cases per 100 000 inhabitants.[7] Wecarried out a serological study of the prevalenceand incidence of dengue virus infection in thiscity,[8] at a time when only DENV-1 and DENV-2 were circulating.[7] The present studyattempts to examine the relationship betweenthe spatial distribution of dengue viruscirculation and the population’s livingconditions, group immunity and Ae. aegyptiinfestation rates, and also to find out anyvariability in the risk of such infections.
Materials and methods
This prospective study on dengueseroprevalence and seroincidence was carried
out in Salvador, Bahia state, Brazil. This cityhad more than 2.3 million inhabitants in 1998and presented marked differences betweenspecific areas with regard to socioeconomicconditions and environmental sanitation. Thepresent study considered 30 spatial units ofanalysis that were named “sentinel areas”.These were selected by stratified samplingusing data obtained from the Brazilian Institutefor Geography and Statistics[9] regardingsanitation system coverage and income levels,which were taken to be estimators of livingconditions, using the following strata:
(1) High: more than 80% of the homeswere connected to the sanitationsystem and more than 50% of thefamilies had an income of more thanfive minimum salaries (80 US dollarsat the time of the study) (6 areas).
(2) Medium: 50% to 80% of the homeswere connected to the sanitationsystem and more than 50% of thefamilies had an income of betweenone and four minimum salaries (19areas).
(3) Low: less than 50% of the homes wereconnected to the sanitation systemand more than 50% of the familieshad an income of less than oneminimum salary (5 areas).
This selection strategy has been describedin more detail elsewhere.[10]
To determine the number of individualsfor the serological surveys, a seroprevalenceof 50% was assumed because this was themean observed in previous surveys in Brazilianstate capitals.[6,11] With this, and assumingprecision of less than or equal to 3% and aconfidence level of 5%, the sample size wasestimated to be 1503 individuals. After adding30% to compensate for possible losses, theresult was 2149 individuals. Using the databaseof a demographic census carried out in 30

38 Dengue Bulletin – Volume 31, 2007
Exposure to the risk of dengue virus infection in an urban setting
sentinel areas in 1997, when 68 749 individualswere counted, a random draw for theparticipants was made (without replacements).These individuals were then grouped in areasby taking the home address intoconsideration.[12]
Two surveys were carried out. The first, in1998, is referred to here as “the seroprevalencesurvey”. The second, one year later in 1999,is termed “the seroincidence survey.”
After the approval of the study protocol bythe Ethics Committee for Scientific Research ofthe Gonçalo Moniz Research Centre (OswaldoCruz Foundation, Bahia), a structuredquestionnaire was formulated during Maythrough July 1998. The data sought in thequestionnaire included: name, address, sex, age,educational level and history of vaccinationagainst yellow fever. Just after the interview,these individuals were provided withclarifications regarding the nature of the studyand they were asked to sign an informed-consentform. Following this, the first blood sample wascollected. Three individuals previouslyvaccinated against yellow fever were excludedin order to avoid false positive serological testresults due to cross-reactions. A second bloodsample was collected a year later, from theindividuals who were negative during the firstserological survey, or had positive reactions toonly one serotype of the dengue virus.
Blood samples were collected in 10 mlsterilized vacuum tubes, and the serum wasseparated by centrifugation and stored at–20 °C. These samples were sent in thermalboxes containing ice to the arbovirus laboratoryof the Evandro Chagas Institute. There, thehaemagglutination inhibition (HI) test,[13] asmodified by Shope,[14] was carried out usingantigens for the four serotypes of the denguevirus and four other flaviviruses (YF, Rocio,Ilhéus and St. Louis encephalitis), althoughthese do not circulate in Salvador. There is a
controversy regarding the interpretation of theserological response to flaviviruses, whichdiffers between the first (primary) infection andany subsequent (secondary) infection withanother flavivirus or serotype (flaviviruses showan increasingly strong response on subsequentinfection and serological cross-reactions arefrequently observed).
Thus, for the interpretation of serologicalresponse, the WHO[15] criteria was followed,i.e. HI titres of 1:20 or higher, exclusively for aspecific dengue serotype, or titres four timeshigher for one serotype than for another (DENV-1 or DENV-2), were considered positive andspecific for that serotype (primary response).Titres indicative of secondary response alsofollowed the WHO criteria.[15] These were alsoconfirmed by IgG enzyme-linkedimmunosorbent assay[16] and were consideredpositive to both serotypes, meaning that infectionwith both DENV-1 and DENV-2 had occurred.
For each sentinel area, the seroprevalenceand seroincidence rates of dengue infection wereestimated both unadjusted (crude) and withstandardization by age using the indirect method(Rothman),[17] and the total composition of thestudy sample as the reference population. Sincethe interval between the two surveys was oneyear, the seroincidence was expressed as anannual rate per cent. The prevalence ratio (PR)and relative risk (RR) of dengue virus infectionwith 95% confidence intervals (CI) wereestimated by taking as the reference standardthe sentinel areas with the lowestseroprevalence (area 427) and seroincidence(area 7), respectively, among the areas in thehighest socio-sanitary stratum. For the threestrata according to living conditions, therespective seroprevalence and infectionincidence were calculated after Rothman,[17] andthe chi-squared test for trend was applied.
From the information collected using thequestionnaire, the frequency indicators were

Exposure to the risk of dengue virus infection in an urban setting
Dengue Bulletin – Volume 31, 2007 39
estimated for each area by taking intoconsideration the proportions of individualsaccording to sex, age greater than or equal to15 years, schooling (assuming that individualswere at risk if they were 15 or more years oldand had not completed elementary schooling)and mean family income less than or equal totwo minimum monthly salaries. The meanpopulation density was obtained from the 1996census.[9] Calculations of Pearson’s correlationcoefficient were used to investigate theexistence of associations between the variablesof interest.
In April 1999, health workers trained andsupervised by the research team visited andinspected all the houses in the 30 sentinel areas.The existence of foci of Ae. aegypti was checkedand records were made of these visits. The unitof analysis for these data was the sentinel area.Every building with one or more breeding sitescontaining the larvae of this mosquito wasconsidered to be positive, and the PremisesIndex (PI) was estimated as the percentage ofpositive buildings. The infection incidence wascalculated for different PI bands (less than orequal to 3%; 3.1% to 5%; 5.1% to 10%; andover 10%) using covariance analysis,[18] withadjustments for age and mean seroprevalence(herd immunity indicator). The preventablefraction was estimated by considering individualswho lived in areas with PI less than or equal to3% to be “non-exposed”.
The data were entered using Epi-Info 6.0and analysed using SAS and STATA.
Results
Among the 1515 individuals who took part inthe seroprevalence survey, 58% were femaleand 71% were aged 15 years or older, mainlyin the age groups 15 to 29 (33%) and 30 to 49(29%). The majority (68%) had had schoolingfor eight years or less. Around 25% reported
that their family income was less than twominimum salaries and 50% earned from twoto less than five minimum salaries. There were595 individuals in the seroincidence survey, outof 860 who were eligible according to thecriteria established, which represented a lossof 31%. The great majority of these losseswere due to changes of address, and it wasnot appropriate to locate these losses since thiswas an ecological study of sentinel areas.Nonetheless, the social and demographicstructure of the sample remained similar towhat was found in the first survey. Among thesentinel areas, the population density variedwidely, from a maximum of 49 980 to aminimum of 1834 inhabitants per km2.
The mean seroprevalence was 69%(ranging from 16% to 98% among the sentinelareas), and this distribution was little changedafter standardization for age. The prevalenceratio (PR) indicated a risk of positive findingsthat ranged from 0.36 in sentinel area 1011 to2.20 in area 1054, which were both in themedium stratum of living conditions (Table 1).
As shown in Table 2, the stratum of lowestliving conditions had the highest meanseroprevalence (74.0%), and this trend wasstatistically significant (χ2 = 8.386; p = 0.004).It can be seen from Table 3 that theseroprevalence presented a positive correlation(r = 0.4914; p = 0.006) with population densityand a weak negative correlation (r = –0.2778;p = 0.137) with the proportion of individualsaged 15 years or over who had had less thaneight years of schooling. No statisticallysignificant association was found with meanincome (r = 0.0571; p = 0.764).
The mean seroincidence was 71.0% peryear. Three areas in which the sample size wasless than four were not taken into consideration;in the others, the seroincidence ranged from50% (area 323) to 90% (area 678) (Table 1). Inthe areas in which the seroprevalence was
40 Dengue Bulletin – Volume 31, 2007
Exposure to the risk of dengue virus infection in an urban settingTa
ble
1: S
erop
reva
lenc
e an
d se
roin
cide
nce
(cru
de a
nd st
anda
rdiz
ed) f
or d
engu
e, st
udy
popu
latio
n, p
opul
atio
n de
nsity
and
pre
mise
sin
dex
(PI)
acco
rdin
g to
sent
inel
are
a an
d st
rata
of l
ivin
g co
nditi
ons i
n Sa
lvad
or –
Bah
ia, B
razi
l, 19
98-1
999
*: 1
998;
**:
199
9; H
: H
igh
livin
g co
nditi
ons;
M:
Mid
dle
livin
g co
nditi
ons;
L:
Low
liv
ing
cond
ition
s
Exposure to the risk of dengue virus infection in an urban setting
Dengue Bulletin – Volume 31, 2007 41
lower, the incidence of infection indicated bya change in serological status was high, exceptin area 571. There was only one area (575) inwhich no individuals seroconverted in thesecond survey, although the number of initiallynegative individuals was only two and this areahad the lowest PI (0.27%) (Table 1). Therelative risk of seroincidence among the areasin the second survey ranged from 0.64 to 1.52,excluding area 575, in which there were nonew cases. In addition, there was a statisticallysignificant negative correlation between theincidence of infection and the proportion ofindividuals aged 15 years or over, who had notcompleted elementary schooling. For meanincome and population density, a negativecorrelation was also found, but withoutstatistical significance (Table 4).
Table 2: Seroprevalence (%) and seroincidence (%) of dengue, prevalence ratio (PR), relative risk(RR) and confidence interval (CI: 95%) according to living condition strata in 30 sentinel areas of
Salvador – Bahia, Brazil, 1998-1999
Table 3: Seroprevalence and seroincidence for two serotypes of dengue virus in Salvador, Brazil,1998-1999
Differing from the seroprevalence survey,the highest unadjusted or standardizedseroincidence rates were in the stratum ofhighest living conditions (Table 2), but the chi-squared test for trend did not show statisticalsignificance (χ2 = 1.332; p = 0.248).
A comparison between the incidenceadjusted for age and the mean seroprevalencein the sentinel areas (herd-immunity indicator),when grouped according to the PI rangesconsidered (Figure), revealed that the lowestseroincidence (55%) was in the group with PIless than or equal to 3% and the highest (77%)was in the group from 3% to 5%. Thedifferences were statistically significant at the5% level only between the first and second PIgroups (p < 0.01) and between the first andfourth groups (p = 0.02).
Test for trend: *χ2 = 8.386, p= 0.004; ** χ2 = 1.332, p= 0.2484
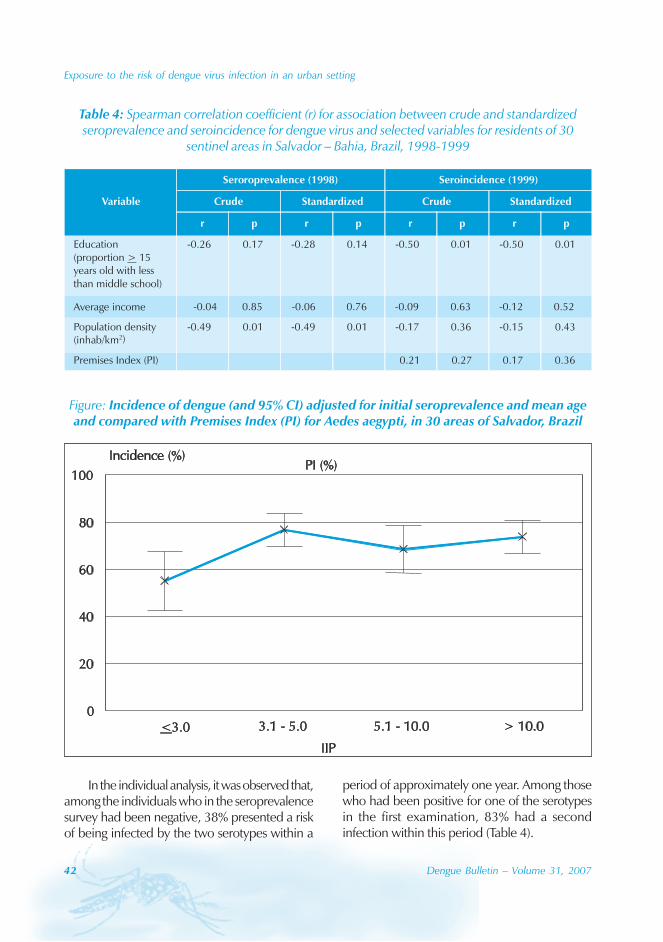
42 Dengue Bulletin – Volume 31, 2007
Exposure to the risk of dengue virus infection in an urban setting
Table 4: Spearman correlation coefficient (r) for association between crude and standardizedseroprevalence and seroincidence for dengue virus and selected variables for residents of 30
sentinel areas in Salvador – Bahia, Brazil, 1998-1999
Figure: Incidence of dengue (and 95% CI) adjusted for initial seroprevalence and mean ageand compared with Premises Index (PI) for Aedes aegypti, in 30 areas of Salvador, Brazil
In the individual analysis, it was observed that,among the individuals who in the seroprevalencesurvey had been negative, 38% presented a riskof being infected by the two serotypes within a
period of approximately one year. Among thosewho had been positive for one of the serotypesin the first examination, 83% had a secondinfection within this period (Table 4).

Exposure to the risk of dengue virus infection in an urban setting
Dengue Bulletin – Volume 31, 2007 43
Discussion
The high seroprevalence level for dengue virusinfection found in this study (69%), and thesubsequent high incidence level (71%), weresurprising. This was particularly so given the shortperiod of time (around four years) that hadelapsed since the virus was introduced intoSalvador. These findings demonstrate the forceof the transmission of this virus in Salvador, giventhat surveys carried out in other Brazilian statecapitals after similar periods of time of viruscirculation revealed lower mean seroprevalence(41% to 44%).[6,19] One plausible explanationfor the rates that were so high in Salvador mayrelate to the almost complete lack of vectorcontrol measures at the outset of virustransmission in this city. Such measures wereonly[7] had waned. On the other hand, in othercities[6,19] such measures had already beenimplemented routinely even before theintroduction of the dengue virus.
It is worth emphasizing that the resultsfrom the present investigation have made itpossible to estimate that, between 1995 and1999, around two million individuals in Salvadorwere infected by the dengue virus. This meansthat the population was at a higher risk forhaemorrhagic dengue, given that DENV-3started circulating in 2002.
The association found in 1998 betweendengue seroprevalence and precarious livingconditions (Table 2) disappeared in theseroincidence for the subsequent year. In otherwords, the risk of being infected by the denguevirus became practically the same between thedifferent economic strata of the population asviral transmission became established inSalvador. This distribution, which was relativelyhomogeneous between the strata, has alsobeen observed in Fortaleza and São Luís doMaranhão. The risk in Fortaleza was even foundto be slightly higher in the districts with bettersocioeconomic indices.[6,19] These facts strongly
suggest that dengue in Brazil is a disease thataffects all social classes.
Despite the statistically significantassociations found between dengueseroprevalence and population density and alsobetween seroincidence and degree of PI andschooling, it can be seen that the risk of infectionwas high in almost all the areas, including in theareas with good living conditions. It is likely thatthis dynamics is at least partially due to the factthat in Salvador high population density and PIare found both in areas with precarious livingconditions and in those where economicallymore favoured populations live, even if somepeople may have been infected away fromhome, for instance at school or at work.
This possible similarity of exposure to therisk of being infected by the dengue virus, indifferent intra-urban environments,differentiates this agent from those of the greatmajority of infectious and parasitic diseases. Inparticular, it differentiates dengue from themicroorganisms whose transmission routes areconnected with the environment and whichpredominantly affect poor populations.
In contrast to the uniformity of exposuresuggested by the ecological comparisondescribed above, our results indicate substantialheterogeneity in exposure to dengue at theindividual level. Since there is no evidence ofvariation in the susceptibility of naive individualsto dengue infection, our finding of a higherincidence of seropositivity (83%) amongindividuals who were initially seropositive toone serotype, is strongly indicative ofheterogeneity in the degree of exposure amongthe individuals of the population of this city.The lack of variation in exposure betweenneighbourhoods, as described above, suggeststhat the main differences are betweenindividual households or people.
These results suggest puzzling andapparently contradictory conclusions regarding

44 Dengue Bulletin – Volume 31, 2007
Exposure to the risk of dengue virus infection in an urban setting
the degree to which exposure to dengueinfection varies across the population, unrelatedto the observed Ae. aegypti density in eacharea. The lack of association between mosquitodensity and dengue seroincidence is particularlyremarkable in view of the way in whichdifferentials existing between individuals canbe expected to be amplified in ecologicalcomparisons.[20] It is hard to see how people’smovement about the city could explain thisphenomenon; by bringing each individual intocontact with various local environments, onewould expect mobility to render their exposuremore homogeneous rather than the contrary.
The larval survey was carried out by theresearch team in 100% of the households,which is grounds for confidence in this indicator,but the use of only one measure of the vectorpopulation constitutes one of the limitationsof this study. Perhaps an option to refine ourresults might have been to count viable pupae,or adult females, per person.[21,22] However,this was not possible, for reasons of cost andoperational complexity. For these same reasons,most vector control programmes use only larvalindicators in their routine monitoring. Thoughthe larval densities vary widely during the courseof a year,[23] the geographical patterns remainconsistent through time.[24] So, our results havedisturbing implications for the effectiveness ofvector control strategies.
An approach that takes into account thelifestyles that potentially favour greaterexposure to the risk of being infected by thedengue virus ought to form one of the lines ofresearch for elucidating this question. This isbecause the differences may be related bothto the public and the private domain, sincethe environment of the home and itssurroundings have a decisive influence on thetransmission of dengue virus.
Another important finding from this studyrelates to the high incidence of dengue virusinfection even when the infestation indices were
relatively low and group immunity had alreadybeen partially established (42%), which alsoexpresses the transmission strength of this agent.It was found that seroincidence did not varywith PI when this was greater than 3%. Thisfinding has strong implications for Ae. aegypticontrol programmes, since it suggests that it isnecessary to intensify vector control and bringAedes densities under 3%, before an observableimpact can be registered. This finding is inagreement with the observation in Singaporethat from the early 1990s the incidence ofdengue rose considerably, even when the PIwas less than 2%.[2] The lack of a safe andeffective vaccine against the virus increases theneed for improved strategies and technologiesfor vector control, and for epidemiologicalstudies to identify changes in infection patterns,such as the locations of transmission foci, theage group of peak incidence, and falling herdimmunity. The last of these was one of themost important factors in the re-establishmentin recent years of intense dengue viruscirculation in Singapore.[25]
It can be understood, therefore, that theresults from this investigation should beconsidered at the time of defining denguecontrol policies and improving vector controlmeasures. On the one hand, observation thatthe dengue virus in our environment does notrespect social spaces strengthens the principlethat vector control measures must always beuniversally applied in each territory. On theother hand, the identification of specific riskfactors in the domestic domain may indicate aneed for other evidence-based interventionswhich can help to eliminate the disease fromcities such as Salvador.
Acknowledgements
The authors acknowledge the financial andtechnical support of the National Centre ofEpidemiology, Ministry of Health, Brazil,especially its Director, Dr Jarbas Barbosa daSilva Junior.

Exposure to the risk of dengue virus infection in an urban setting
Dengue Bulletin – Volume 31, 2007 45
References
[1] Kuno G. Review of the factors modulatingdengue transmission. Epidemiol Rev. 1995;17(2): 321-35.
[2] Farrar J, Focks D, Gubler D, Barrera R,Guzman MG, Simmons C, Kalayanarooj S,Lum L, McCall PJ, Lloyd L, Horstick O, Dayal-Drager R, Nathan MB, Kroeger A. WHO/TDRDengue Scientific Working Group. Towardsa global dengue research agenda. Trop MedInt Health. 2007 Jun; 12(6): 695-9.
[3] Newton EA, Reiter P. A model of thetransmission of dengue fever with anevaluation of the impact of ultra-low volume(ULV) insecticide applications on dengueepidemics. Am J Trop Med Hyg. 1992 Dec;47(6): 709-20.
[4] Gubler DJ. Dengue and dengue hemorragicfever: its history and resurgence as a globalhealth problem. In: Gubler DJ, Kuno G.,editors. Dengue and dengue hemorragic fever.New York: CAB International, 1997. p. 1-22.
[5] Rodhain F, Rosen L. Mosquito vectors anddengue virus-vector relationships. In: GublerDJ, Kuno G. Editors. Dengue and denguehemorragic fever. New York: CAB International,1997. p. 45- 60.
[6] Vasconcelos PF, Lima JW, da Rosa AP, TimbóMJ, da Rosa ES, Lima HR, Rodrigues SG, daRosa JF. [Dengue epidemic in Fortaleza, Ceará:randomized seroepidemiologic survey] RevSaude Publica. 1998 Oct; 32(5): 447-54.Portuguese.
[7] Teixeira MG, Costa MC, Barreto ML, BarretoFR. [Epidemiology of dengue in Salvador-Bahia, 1995-1999] Rev Soc Bras Med Trop.2001 May-Jun; 34(3): 269-74. Portuguese.
[8] Teixeira Mda G, Barreto ML, Costa Mda C,Ferreira LD, Vasconcelos PF, Cairncross S.Dynamics of dengue virus circulation: a silentepidemic in a complex urban area. Trop MedInt Health. 2002 Sep; 7(9): 757-62.
[9] Censo Demográfico. Fundação IBGE. Rio deJaneiro, Brasil: Censo demográfico, 1991.
[10] Teixeira Md Mda G, Barreto ML, Costa MdaC, Strina A, Martins D Jr, Prado M. Sentinelareas: a monitoring strategy in public health.Cad Saude Publica. 2002 Sep-Oct; 18(5):1189-95.
[11] Figueiredo LT, Cavalcante SM, Simões MC.Dengue serologic survey of schoolchildren inRio de Janeiro, Brazil, in 1986 and 1987. BullPan Am Health Organ. 1990; 24(2): 217-25.
[12] Cochran WG, Sampling techniques. 3rd
edition, New York: Wiley & Sons, 1977.
[13] Clarke DH, Casals J. Techniques forhemagglutination and hemagglutination-inhibition with arthropod-borne viruses. Am JTrop Med Hyg. 1958 Sep; 7(5): 561-73.
[14] Shope RE. The use of a microhemagglutination-inhibition test to followantibody response after arthropod-borne virusinfection in a community of forest animals.Anais de microbiologia (Rio de Janeiro) v.11(parte A) 1963 Sep: 167-171.
[15] World Health Organization. Denguehaemorrhagic fever: diagnosis, treatment,prevention and control. 2nd edition, Geneva:WHO, 1997. p.84.
[16] Chungue E, Marché G, Plichart R, Boutin JP,Roux J. Comparison of immunoglobulin Genzyme-linked immunosorbent assay (IgG-ELISA) and haemagglutination inhibition (HI)test for the detection of dengue antibodies.Prevalence of dengue IgG-ELISA antibodies inTahiti. Trans R Soc Trop Med Hyg. 1989 Sep-Oct; 83(5): 708-11.
[17] Rothman KJ. Standardization of rates. In:Modern epidemiology. Ed. Little Brown andCompany, Boston-Toronto: Little Brown & Co,,1986. p. 41-49.
[18] Montgomery DC. Design and analysis ofexperiments. 3rd edition. New York: Wiley,1991.
[19] Vasconcelos PF, Lima JW, Raposo ML,Rodrigues SG, da Rosa JF, Amorim SM, da Rosa

46 Dengue Bulletin – Volume 31, 2007
Exposure to the risk of dengue virus infection in an urban setting
ES, Moura CM, Fonseca N, da Rosa AP. [Aseroepidemiological survey on the island ofSão Luis during a dengue epidemic inMaranhão] Rev Soc Bras Med Trop. 1999 Mar-Apr; 32(2): 171-9. Portuguese.
[20] Koopman JS, Longini IM Jr. The ecologicaleffects of individual exposures and nonlineardisease dynamics in populations. Am J PublicHealth. 1994 May; 84(5): 836-42.
[21] Focks DA, Brenner RJ, Hayes J, Daniels E.Transmission thresholds for dengue in termsof Aedes aegypti pupae per person withdiscussion of their utility in source reductionefforts. Am J Trop Med Hyg. 2000 Jan; 62(1):11-8.
[22] Strickman D, Kittayapong P. Dengue and itsvectors in Thailand: calculated transmissionrisk from total pupal counts of Aedes aegypti
and association of wing-length measurementswith aspects of the larval habitat. Am J TropMed Hyg. 2003 Feb; 68(2): 209-17.
[23] Secretaria Municipal de Saúde. Relatório doPlano de Erradicação do Aedes aegypti noMunicípio de Salvador. Prefeitura Municipalde Salvador. Centro de Controle de Zoonozes,1999.
[24] Morrison AC, Gray K, Getis A, Astete H,Sihuincha M, Focks D, Watts D, Stancil JD,Olson JG, Blair P, Scott TW. Temporal andgeographic patterns of Aedes aegypti (Diptera:Culicidae) production in Iquitos, Peru. J MedEntomol. 2004 Nov; 41(6): 1123-42.
[25] Ooi EE, Goh KT, Gubler DJ. Dengueprevention and 35 years of vector control inSingapore. Emerg Infect Dis. 2006 Jun; 12(6):887-93.

Dengue Bulletin – Volume 31, 2007 47
Detection of transovarial dengue virus from field-caughtAedes aegypti and Ae. albopictus larvae using C6/36 cell
culture and reverse transcriptase-polymerase chainreaction (RT-PCR) techniques
A. Rohania , I. Zamreea, H.L. Leea, I. Mustafakamalb, M.J. Norjaizab
and D. Kamilanc
aMedical Entomology Unit, Infectious Disease Research Centre, Institute for Medical Research, Kuala Lumpur
bVector Borne Disease Centre Programme, Kuala Terengganu, Terengganu
cVector Borne Disease Centre Programme, Kuantan, Pahang
Abstract
Larvae of Aedes aegypti and Aedes albopictus were collected from a wide variety of artificial containers.Most samples were collected from used tyres and water-holding containers located in residential urbanor rural areas. The identified mosquito larvae were pooled according to the species, date and localityand stored at –70 °C. A total of 378 pools of Ae. aegypti and 553 pools of Ae. albopictus were collected.Virus isolation was carried out using cell culture (C6/36 clone) of Ae. albopictus and virus detection byreverse-transcriptase polymerase chain reaction (RT-PCR). Transovarial transmission of dengue viruswas demonstrated in both Ae. aegypti and Ae. albopictus in nature. Infected larvae were recovered from16 localities (10 in Terengganu; 5 in Kuala Lumpur and 1 in Pahang). The study showed that both thecell culture and RT-PCR techniques can be used to detect dengue virus from mosquito larvae.
Keywords: Dengue; Aedes aegypti; Aedes albopictus; Transovarial transmission; Malaysia.
E-mail: [email protected]
Introduction
Dengue is an acute arboviral human diseasecaused by four serotypes of dengue viruses,which are closely related but antigenicallydistinct. Dengue viruses are transmitted tohumans through the bite of infectedmosquitoes. For many years, members of thesubgenus Stegomyia, especially Aedes aegypti(Linn.) and Aedes albopictus (Skuse.) have beenrecognized as the primary vectors of
dengue.[1,2,3] Ae. aegypti breeds in clean watercollected in and around human settlements.Ae. albopictus can be found not only aroundand near human habitation, like Ae. aegypti,but also in forests and plantations.
The emergence of dengue has been linkedto economic and ecological changes thatcaused dramatic expansion of the urbanizedvector of dengue viruses. Industrialization andrapid human population growth led to

48 Dengue Bulletin – Volume 31, 2007
Detection of transovarial dengue virus from field-caught larvae of Ae. aegypti and Ae. albopictus
uncontrolled urbanization in Malaysia. In theabsence of proper water supply (for examplein the squatter (slum) areas), residents have tostore water for domestic use, creating the idealecological niche for Ae. aegypti and Ae.albopictus. These mosquitoes prefer to feedon humans and to lay eggs in artificialcontainers. The increase in air travel alsocontributed to the expansion of dengue,providing the means for viremic people to movevery rapidly from one place to another.[4]
The prevention and control of dengueoutbreaks depends upon the surveillance ofcases and mosquito vectors. Vector surveillanceallows timely implementation of emergencymosquito control measures to limit animpending outbreak from spreading.
In Malaysia, a comprehensive mosquitocontrol programme administered by the Vector-Borne Disease Control Programme, Ministryof Health, incorporates source reduction, publichealth education, community participation andlaw enforcement against mosquito breeding.The Aedes control strategy has focused mainlyon surveillance for the elimination of Aedeslarval breeding habitats and emergency controlof adults using insecticidal fogging duringoutbreaks. Although this strategy hassuccessfully reduced the Aedes mosquitopopulation to a relatively low level, as indicatedby the overall House index (HI), it has notprevented the emergence of progressivelylarger outbreaks in recent years.[5]
Despite extensive research on vaccinedevelopment, there are at present no knownmethods of controlling dengue except bylimiting the mosquito vectors. Virologicalsurveillance, which involves the monitoring ofdengue virus infection in humans, has beenused as an early warning system to predictoutbreaks.[6,7] Such surveillance is based onisolation of dengue virus from human serumby cell culture or mosquito inoculation and
type-specific identification by immuno-flourescence or multiplex RT-PCR. Thisapproach is less effective since the virus isalready circulating in the human population. Amore effective approach is to detect the virusin mosquitoes before it is introduced into thehuman population. This way, preventive vectorcontrol measures can be undertakenimmediately to offset an outbreak. Surveillanceof mosquitoes infected with dengue virusesprovides an early warning sign for risk oftransmission in an area and the specificpredominant circulating serotype in the vectorpopulation.[8] Control programmes can beprioritized and focused more effectively inspecific localities.
The objective of this study was to detectdengue virus from field mosquitoes using C6/36cell culture and reverse transcriptase-polymerase chain reaction (RT-PCR)techniques.
Materials and methods
Larval collections
Third and fourth instar Aedes aegypti and Aedesalbopictus larvae were collected from the fieldin major towns in Malaysia, i.e. Terengganu,Pahang and Kuala Lumpur. The larvae werecollected during dengue outbreaks for eachstate in 2005. Mosquito larvae collected in thesurvey were identified using standardtaxonomic keys. Identified mosquito larvaewere segregated according to species, site anddate. Mosquito larvae were stored in pools of10 larvae per pool in cryogenic vials at –70 °Cfor future virus isolation studies. The mosquitopools were selected randomly and dengue virusdetection was carried out by RT-PCR and C6/36 cell culture technique. A total of 931 poolsof Aedes mosquitoes were separated equallyand assayed by C6/36 cell culture and RT-PCR.

Detection of transovarial dengue virus from field-caught larvae of Ae. aegypti and Ae. albopictus
Dengue Bulletin – Volume 31, 2007 49
Detection of dengue virus usingreverse transcriptase-polymerasechain reaction (RT-PCR)
Extraction of RNA
The larval pools were homogenized in a sterilehomogenizer and RNA was extracted usingHigh Pure Nucleic Acid Kit (Roche, Germany).The procedures of RNA extraction were asfollows: 200 μl sample were addedsubsequently into 1.5 ml eppendorf tubescontaining 50 μl proteinase K. This was brieflyvortexed and incubated at 27 °C for 10minutes. Subsequently, 100 μl of isopropanolwas added and vortexed briefly. The solutionwas then pipetted into the high pure filtertubes, placed into collection tubes and werethen centrifuged at 14 000 rpm for 1 minute.The flow-through was discarded and filter tubeswere placed into new collection tubes. Fivehundred microlitres of inhibitor-remover washbuffer were added into the upper reservoir andcentrifuged at 14 000 rpm for 1 minute. Theflow-through was discarded and again the filtertubes were placed into new collection tubes.A volume of 450 μl wash buffer was added tothe upper reservoir and centrifuged at 14 000rpm for 1 minute. The flow-through wasdiscarded and again the filter tubes were placedinto new collection tubes. The wash step wasrepeated and continued with a short spin for15 seconds. The collection tube was discardedand the filter tubes were inserted into clean1.5 ml eppendorf tubes. RNA was eluted byadding 35 μl elution buffer into the middle spotof the filter tubes and centrifuged at 14 000rpm for 2 minutes. The extracted RNA waskept at –70 °C until used.
Detection of viral RNA using consensusprimers
RNA isolated from each pool was subjected tothe RT-PCR assay using Lanciotti’s dengue
consensus primers (TCAATATGCTGAAACGCGCAGAAACCG and TTGCACCAACAGTCAATGTCTTCAGGTTC).[9] The primers correspond togenome positions 131 to 161 and 616 to 644of dengue virus respectively. Master mix wasprepared using Titan One Tube RT-PCR Kit.Each reaction required 9.75 μl of doubledistilled water, 2 μl of dNTP mix, 1.25 μl ofDTT, 0.5 μl RNAse inhibitor, 5.0 μl of RT-PCRbuffer, 0.5 μl of enzyme mix, 0.5 μl of dengueuniversal sense primer and 0.5 μl of dengueuniversal anti-sense primer to reach 20 μl totalvolume of master mix.
RNA products were prepared by heatingthe tubes at 65 °C for 5 minutes by block heater.Five microlitres of each RNA product wereadded to the master mix and then centrifugedat 8000 rpm. The RT step was carried out at51 °C for one 30 minutes to produce cDNAwhich was then amplified by the following PCRsteps: 92 °C for 3 minutes as initial denaturation,92 °C for 30 seconds as denaturation steps, 51 °Cfor 45 seconds as annealing step and 72 °C for1 minutes as extension step. The cycle wasrepeated 41 times before final extension at 72 °Cfor 5 minutes. For every RT-PCR run, a positivecontrol (a confirmed dengue isolate) andnegative control (C6/36 cell culture withoutdengue virus) were included.
The PCR products were analysed byperforming electrophoresis in a 2.0% NusievePCR gel (FC Bio, USA) stained with ethidiumbromide and run at about 100 volts. The gelwas viewed under ultraviolet illuminator (UltraLum Ins., California, USA) and the image ofthe resulting band was captured with a Polaroidcamera.
Dengue virus detection using Ae.albopictus clone C6/36 cells[10]
The method employed was modified fromManeekarn.[11] The C6/36 cells were grown in

50 Dengue Bulletin – Volume 31, 2007
Detection of transovarial dengue virus from field-caught larvae of Ae. aegypti and Ae. albopictus
minimum essential medium (MEM) growthmedia supplemented with 10% fetal calf serum(FCS) for 2 days until cell monolayers wereformed in culture tubes. Pooled mosquitoeswere homogenized on ice in 1.5 ml MEMmedium with 5% fetal calf serum. 100 ul eachof the homogenate were passed through 0.2um filters and inoculated into respective culturetubes. The culture tubes were then vortexedand left to incubate for 2 hours at ambienttemperature for adsorption. Maintenancemedium containing 2% FCS was added andthe culture tubes were incubated at 28 °C for7 days.
Smear preparation
The culture tubes were vortexed andcentrifuged after 7 days’ incubation period.Cells from the sediments were transferred ontothe Teflon-coated, 12 well slides where eachslide would have a maximum of 7 test smears,4 positive control and a negative control. Thesmears were left to dry at 28 °C for 4 hours ina class II biohazard cabinet with the air bloweron. The smears were then fixed with coldacetone for 20 minutes. The cultures werestored at –20 °C for confirmation by a secondor third passage in cell culture if only the initialresults were positive.
Peroxidase-antiperoxidase (PAP) staining
PAP staining procedures were performed aspreviously described by Igarashi.[12] The cellsthat were fixed with cold acetone reacted withdengue anti-serum at 1:1000 at roomtemperature for 40 minutes. The antibody isuniversally reactive to all 4 dengue serotypes.The cells were rinsed in PBS and reacted withrabbit anti-mouse IgG at 1:1000 for 40 minutes.The cells were rinsed in PBS again and thenreacted with goat-anti rabbit IgG at 1:1000 for40 minutes. The cells were then rinsed in PBSand exposed to peroxidase-rabbit anti-
peroxidase complex at 1:1000 for 40 minutesfollowed by washing and peroxidase reaction,using 0.2 mg/ml of 3,3 diaminobenzidine(DAB) and 0.2% H2O2 as the substrate. Thecells were observed under a normal lightcompound microscope. The positive sampleswere reconfirmed by a second passage in cellculture.
Minimum Infection Rate (MIR)
The MIR was used to compare virus infectionrates in Ae. aegypti and Ae. albopictus larvaeat the sampling area. The MIR was calculatedas (number of positive pools by species ÷ totalnumber of that species tested) X 1000.[13]
Virus Infection Rate (VIR)
The VIR was calculated as (number ofmosquitoes by species infected with denguevirus ÷ total number of that species tested) X100.[14]
Results and discussion
In urban areas of Malaysia, artificial containersare the major larval habitats of Aedesmosquitoes in and near human habitation. Thetype and location, presence of shade and waterconditions usually associated with waterreceptacles are known to affect breeding ofAedes mosquitoes.[15]
The dominant mosquito larvae collectedin the present survey were Ae. albopictus,whereas only very low number of containerswere positive for Ae. aegypti larvae. Ae. aegyptitend to be more selective in nature comparedto Ae. albopictus. Ae. aegypti larvae preferredheavily shaded areas. This preferential choiceof shaded breeding area was due to the stabilityof water temperature in the artificial containersand therefore more conducive to breeding.[15]
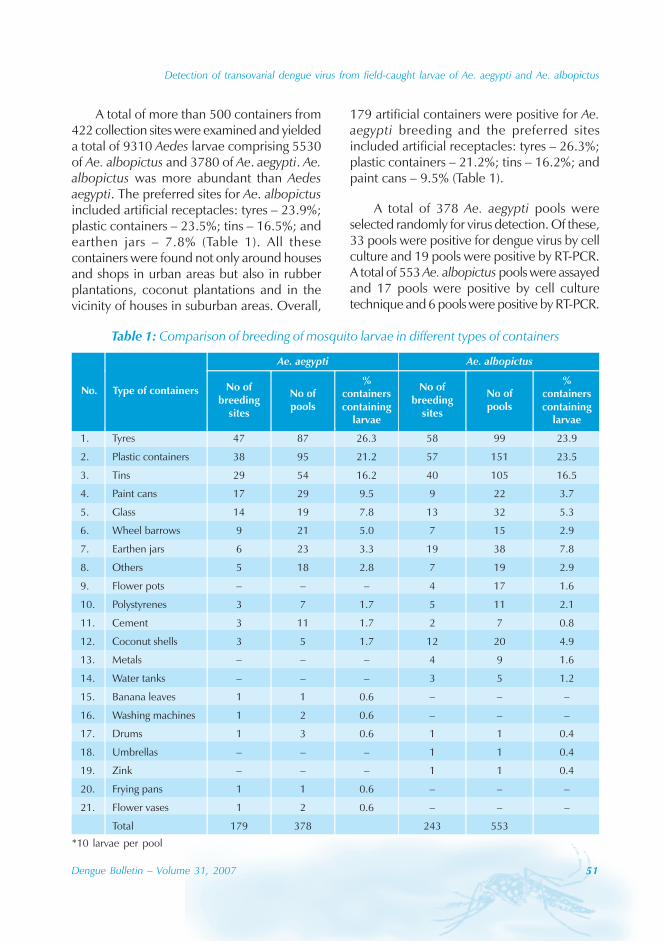
Detection of transovarial dengue virus from field-caught larvae of Ae. aegypti and Ae. albopictus
Dengue Bulletin – Volume 31, 2007 51
A total of more than 500 containers from422 collection sites were examined and yieldeda total of 9310 Aedes larvae comprising 5530of Ae. albopictus and 3780 of Ae. aegypti. Ae.albopictus was more abundant than Aedesaegypti. The preferred sites for Ae. albopictusincluded artificial receptacles: tyres – 23.9%;plastic containers – 23.5%; tins – 16.5%; andearthen jars – 7.8% (Table 1). All thesecontainers were found not only around housesand shops in urban areas but also in rubberplantations, coconut plantations and in thevicinity of houses in suburban areas. Overall,
179 artificial containers were positive for Ae.aegypti breeding and the preferred sitesincluded artificial receptacles: tyres – 26.3%;plastic containers – 21.2%; tins – 16.2%; andpaint cans – 9.5% (Table 1).
A total of 378 Ae. aegypti pools wereselected randomly for virus detection. Of these,33 pools were positive for dengue virus by cellculture and 19 pools were positive by RT-PCR.A total of 553 Ae. albopictus pools were assayedand 17 pools were positive by cell culturetechnique and 6 pools were positive by RT-PCR.
Table 1: Comparison of breeding of mosquito larvae in different types of containers
*10 larvae per pool
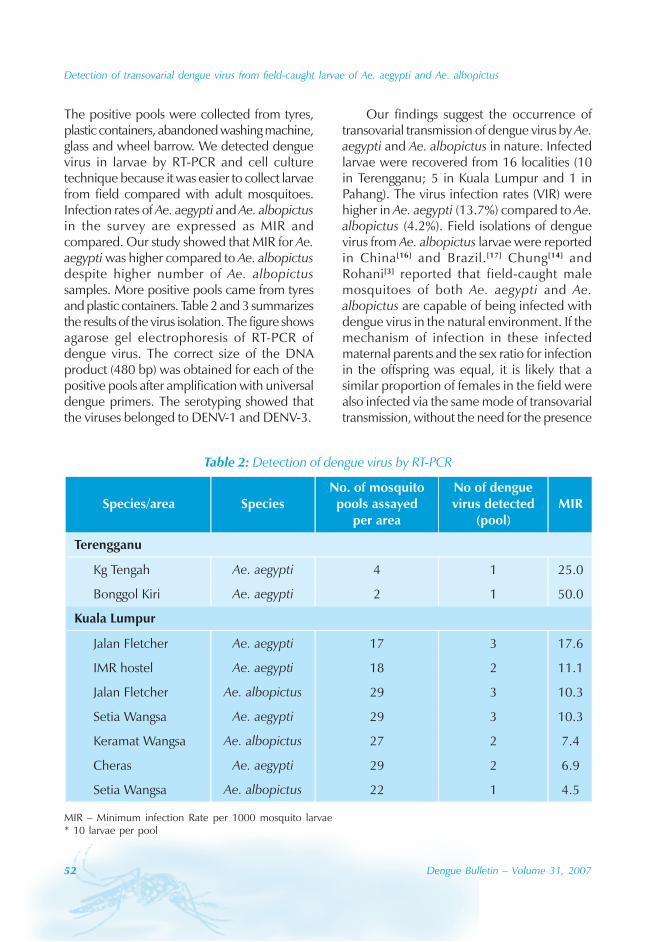
52 Dengue Bulletin – Volume 31, 2007
Detection of transovarial dengue virus from field-caught larvae of Ae. aegypti and Ae. albopictus
The positive pools were collected from tyres,plastic containers, abandoned washing machine,glass and wheel barrow. We detected denguevirus in larvae by RT-PCR and cell culturetechnique because it was easier to collect larvaefrom field compared with adult mosquitoes.Infection rates of Ae. aegypti and Ae. albopictusin the survey are expressed as MIR andcompared. Our study showed that MIR for Ae.aegypti was higher compared to Ae. albopictusdespite higher number of Ae. albopictussamples. More positive pools came from tyresand plastic containers. Table 2 and 3 summarizesthe results of the virus isolation. The figure showsagarose gel electrophoresis of RT-PCR ofdengue virus. The correct size of the DNAproduct (480 bp) was obtained for each of thepositive pools after amplification with universaldengue primers. The serotyping showed thatthe viruses belonged to DENV-1 and DENV-3.
Our findings suggest the occurrence oftransovarial transmission of dengue virus by Ae.aegypti and Ae. albopictus in nature. Infectedlarvae were recovered from 16 localities (10in Terengganu; 5 in Kuala Lumpur and 1 inPahang). The virus infection rates (VIR) werehigher in Ae. aegypti (13.7%) compared to Ae.albopictus (4.2%). Field isolations of denguevirus from Ae. albopictus larvae were reportedin China[16] and Brazil.[17] Chung[14] andRohani[3] reported that field-caught malemosquitoes of both Ae. aegypti and Ae.albopictus are capable of being infected withdengue virus in the natural environment. If themechanism of infection in these infectedmaternal parents and the sex ratio for infectionin the offspring was equal, it is likely that asimilar proportion of females in the field werealso infected via the same mode of transovarialtransmission, without the need for the presence
Table 2: Detection of dengue virus by RT-PCR
MIR – Minimum infection Rate per 1000 mosquito larvae* 10 larvae per pool
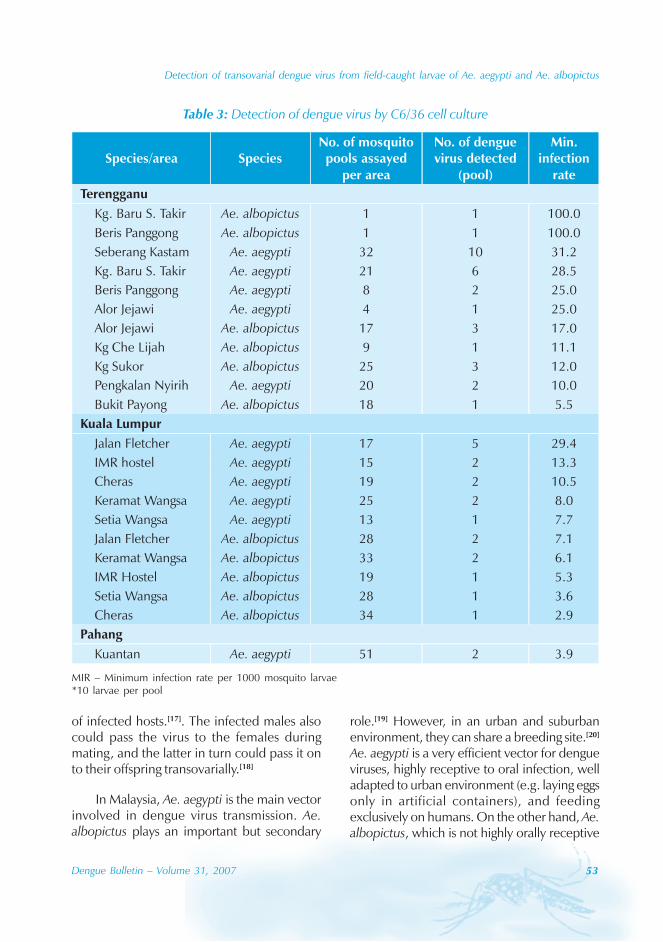
Detection of transovarial dengue virus from field-caught larvae of Ae. aegypti and Ae. albopictus
Dengue Bulletin – Volume 31, 2007 53
Table 3: Detection of dengue virus by C6/36 cell culture
MIR – Minimum infection rate per 1000 mosquito larvae*10 larvae per pool
of infected hosts.[17]. The infected males alsocould pass the virus to the females duringmating, and the latter in turn could pass it onto their offspring transovarially.[18]
In Malaysia, Ae. aegypti is the main vectorinvolved in dengue virus transmission. Ae.albopictus plays an important but secondary
role.[19] However, in an urban and suburbanenvironment, they can share a breeding site.[20]
Ae. aegypti is a very efficient vector for dengueviruses, highly receptive to oral infection, welladapted to urban environment (e.g. laying eggsonly in artificial containers), and feedingexclusively on humans. On the other hand, Ae.albopictus, which is not highly orally receptive
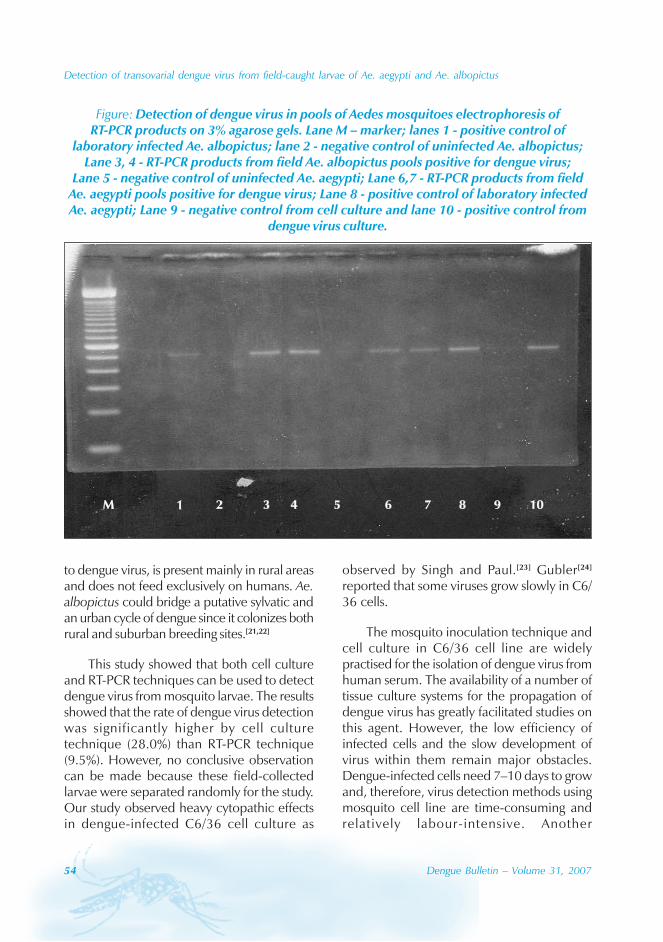
54 Dengue Bulletin – Volume 31, 2007
Detection of transovarial dengue virus from field-caught larvae of Ae. aegypti and Ae. albopictus
Figure: Detection of dengue virus in pools of Aedes mosquitoes electrophoresis ofRT-PCR products on 3% agarose gels. Lane M – marker; lanes 1 - positive control of
laboratory infected Ae. albopictus; lane 2 - negative control of uninfected Ae. albopictus;Lane 3, 4 - RT-PCR products from field Ae. albopictus pools positive for dengue virus;
Lane 5 - negative control of uninfected Ae. aegypti; Lane 6,7 - RT-PCR products from fieldAe. aegypti pools positive for dengue virus; Lane 8 - positive control of laboratory infectedAe. aegypti; Lane 9 - negative control from cell culture and lane 10 - positive control from
dengue virus culture.
to dengue virus, is present mainly in rural areasand does not feed exclusively on humans. Ae.albopictus could bridge a putative sylvatic andan urban cycle of dengue since it colonizes bothrural and suburban breeding sites.[21,22]
This study showed that both cell cultureand RT-PCR techniques can be used to detectdengue virus from mosquito larvae. The resultsshowed that the rate of dengue virus detectionwas significantly higher by cell culturetechnique (28.0%) than RT-PCR technique(9.5%). However, no conclusive observationcan be made because these field-collectedlarvae were separated randomly for the study.Our study observed heavy cytopathic effectsin dengue-infected C6/36 cell culture as
observed by Singh and Paul.[23] Gubler[24]
reported that some viruses grow slowly in C6/36 cells.
The mosquito inoculation technique andcell culture in C6/36 cell line are widelypractised for the isolation of dengue virus fromhuman serum. The availability of a number oftissue culture systems for the propagation ofdengue virus has greatly facilitated studies onthis agent. However, the low efficiency ofinfected cells and the slow development ofvirus within them remain major obstacles.Dengue-infected cells need 7–10 days to growand, therefore, virus detection methods usingmosquito cell line are time-consuming andrelatively labour-intensive. Another

Detection of transovarial dengue virus from field-caught larvae of Ae. aegypti and Ae. albopictus
Dengue Bulletin – Volume 31, 2007 55
disadvantage to this method of flavivirusdetection in field-caught mosquitoes has beenthe requirement to keep captured mosquitoesalive or frozen fresh to detect viral infection.
RT-PCR for the detection of flaviviruses inhuman clinical samples has been used fornearly a decade now.[25] The accuracy andspeed of the RT-PCR assay makes it anappealing test for the diagnosis of dengue andepidemiologic surveillance. The present studyindicates that PCR could detect dengue virusin mosquitoes using larvae collected from thefield. Similar findings were also reported byChao et al. (2007).[26] These techniquesconstitute practical molecular diagnostic andepidemiological tools for the virologicalsurveillance of dengue virus-infected Aedesmosquitoes to serve as an early warning systemfor dengue outbreaks.
In typical infections many types ofmosquito tissues are eventually infected andthe virus can persist in these tissues throughoutthe life of the vector. It is generally assumedthat once a mosquito becomes infected withdengue it retains that infection for life.[27]
Therefore, the collection of wild-caughtmosquitoes would appear to be a reasonableapproach to virus detection compared withattempts at virus recovery in human
populations, which typically have very shortperiods (4-5 days) of detectable viremia.
Active surveillance for dengue virus infectedmosquitoes can be an effective way to predictthe risk of dengue infection in a given area. Aneed exists for better techniques to conducttimely, accurate and meaningful vector surveysand virus detection for estimating transmissionrisk among susceptible population in endemicareas. PCR could be considered for processinglarge numbers of samples in operationalprogrammes, mapping areas with different levelsof endemicity and stratification of areas, providedthe technique is further refined to have higherspecificity and sensitivity. A better understandingof dengue virus-vector relationships in the fieldand their association with human disease canhelp establish reliable vector population targetlevels for initiating measures to control andreduce virus transmission.
Acknowledgements
The authors wish to thank the Director, Institutefor Medical Research, Kuala Lumpur, forpermission to publish. Thanks are also due tothe staff of the Medical Entomology Unit/IDRC,IMR, and the Vector-Borne Disease CentreProgramme, Terengganu and Pahang.
References
[1] Boromisa RD, Rai KS, Grimstad PR. Variationin the vector competence of geographic strainsof Aedes albopictus for dengue 1 virus. J AmMosq Control Assoc. 1987 Sep; 3(3): 378-86.
[2] Gubler DJ. Dengue and dengue hemorrhagicfever in the Americas. P R Health Sci J. 1987Aug;6(2):107-11.
[3] Ahmad R, Ismail A, Saat Z, Lim LH. Detectionof dengue virus from field Aedes aegypti andAedes albopictus adults and larvae. Southeast
Asian J Trop Med Public Health. 1997 Mar;28(1): 138-42.
[4] Gubler DJ. The global pandemic of dengue/dengue haemorrhagic fever: current status andprospects for the future. Ann Acad MedSingapore. 1998 Mar; 27(2): 227-34.
[5] VBDCP. Annual report vector borne diseasecontrol programme, Ministry of Health,Malaysia. 2004: 177.

56 Dengue Bulletin – Volume 31, 2007
Detection of transovarial dengue virus from field-caught larvae of Ae. aegypti and Ae. albopictus
[6] Lam SK. Two decades of dengue in Malaysia.Trop Med. 1993; 35(4): 195-200.
[7] Seah CL, Chow VT, Tan HC, Can YC. Rapid,single-step RT-PCR typing of dengue virusesusing five NS3 gene primers. J Virol Methods.1995 Feb; 51(2-3): 193-200.
[8] Loroño-Pino MA, Cropp CB, Farfán JA,Vorndam AV, Rodríguez-Angulo EM, Rosado-Paredes EP, Flores-Flores LF, Beaty BJ, GublerDJ. Common occurrence of concurrentinfections by multiple dengue virus serotypes.Am J Trop Med Hyg. 1999 Nov; 61(5): 725-30.
[9] Lanciotti RS, Calisher CH, Gubler DJ, ChangGJ, Vorndam AV. Rapid detection and typingof dengue viruses from clinical samples byusing reverse transcriptase-polymerase chainreaction. J Clin Microbiol. 1992 Mar; 30(3):545-51.
[10] Igarashi A. Isolation of a Singh’s Aedesalbopictus cell clone sensitive to dengue andchikungunya viruses. J Gen Virol. 1978 Sep;40(3): 531-44.
[11] Maneekarn N, Morita K, Tanaka M, IgarashiA, Usawattanakul W, Sirisanthana V, Innis BL,Sittisombut N, Nisalak A, Nimmanitya S.Applications of polymerase chain reaction foridentification of dengue viruses isolated frompatient sera. Microbiol Immunol. 1993; 37(1):41-7.
[12] Igarashi A, Fujita N, Okura Y. Isolation ofdengue viruses from patients with denguehemorrhagic fever (DHF) and those with feverof unknown origin (FUO) in Jakarta, Indonesiain the year of 1981 and 1982. Annals ofInternational Council for Medical Research1992;2:7-17.
[13] Chow VT, Chan YC, Yong R, Lee KM, Lim LK,Chung YK, Lam-Phua SG, Tan BT. Monitoringof dengue viruses in field-caught Aedes aegyptiand Aedes albopictus mosquitoes by a type-specific polymerase chain reaction and cyclesequencing. Am J Trop Med Hyg. 1998 May;58(5): 578-86.
[14] Kow CY, Koon LL, Yin PF. Detection of dengueviruses in field caught male Aedes aegypti andAedes albopictus (Diptera: Culicidae) inSingapore by type-specific PCR. J MedEntomol. 2001 Jul; 38(4): 475-9.
[15] Rohani A, Abdullah AG, Ong YF, Saadiyah I,Zamree I, Lee H L. Survey of mosquito larvaedistribution in container habitats collectedfrom urban and rural areas in major towns ofMalaysia. Trop Biomed. 2001; 18(1): 41-49.
[16] Zuo L, Shu LP. Isolation, identification, andphylogenetic analysis of a dengue virus strainfrom Aedes albopictus collected in Mawei townin Guizhou Province, China. Chin Med J (Engl).2004 Dec; 117(12): 1847-9.
[17] Mitchell CJ, Miller BR. Vertical transmission ofdengue viruses by strains of Aedes albopictusrecently introduced into Brazil. J Am MosqControl Assoc. 1990 Jun; 6(2): 251-3.
[18] Tu WC, Chen CC, Hou RF. Ultrastructuralstudies on the reproductive system of maleAedes aegypti (Diptera: Culicidae) infectedwith dengue 2 virus. J Med Entomol. 1998 Jan;35(1): 71-6.
[19] Lee HL. A nation wide resurvey of the factorsaffecting the breeding of Aedes aegypti (L.) andAedes albopictus (Skuse) (Diptera:Culicidae)in urban town of Peninsular Malaysia 1988-1999. Trop Biomed. 1991;8:185-189.
[20] Lee HL. Environmental friendly approachesto mosquito control. Symposium on “TowardsA mosquito-safe environment” 29-30 June1999. Legend Hotel, Kuala Lumpur, Malaysia.
[21] Rudnick A. Ecology of dengue virus. Asian JInfectious Disease. 1978; 2: 156-160.
[22] Vazeille M, Rosen L, Mousson L, Failloux AB.Low oral receptivity for dengue type 2 virusesof Aedes albopictus from Southeast Asiacompared with that of Aedes aegypti. Am JTrop Med Hyg. 2003 Feb; 68(2): 203-8.
[23] Singh KR, Pavri KM. Experimental studies withchikungunya virus in Aedes aegypti and Aedesalbopictus. Acta Virol. 1967 Nov; 11(6): 517-26.

Detection of transovarial dengue virus from field-caught larvae of Ae. aegypti and Ae. albopictus
Dengue Bulletin – Volume 31, 2007 57
[24] Gubler DJ, Kuno G, Sather GE, Velez M, OliverA. Mosquito cell cultures and specificmonoclonal antibodies in surveillance fordengue viruses. Am J Trop Med Hyg. 1984 Jan;33(1): 158-65.
[25] Eldadah ZA, Asher DM, Godec MS, PomeroyKL, Goldfarb LG, Feinstone SM, Levitan H,Gibbs CJ Jr, Gajdusek DC. Detection offlaviviruses by reverse-transcriptase polymerasechain reaction. J Med Virol. 1991 Apr; 33(4):260-7.
[26] Chao DY, Davis BS, Chang GJ. Developmentof multiplex real-time reverse transcriptase PCRassays for detecting eight medically importantflaviviruses in mosquitoes. J Clin Microbio. 2007Feb; 45(2): 584-9.
[27] Rodhain F and Rosen L. Mosquito vectors anddengue virus relationships. In: DJ Gubler andG Kuno, editors. Dengue and denguehemorrhagic fever. Wallingford (UK): CAB;1997. p. 45-60.

58 Dengue Bulletin – Volume 31, 2007
E-mail: [email protected]; Tel.: 603-7967-6818; Fax: 603-7967-4579
A new approach to classify risk in dengue infectionusing bioelectrical impedance analysis
Fatimah Ibrahima , Wan Abu Bakar Wan Abasa, Mohd Nasir Taibb,Chan Chong Guanc and Saadiah Sulaimand
aDepartment of Biomedical Engineering, Faculty of Engineering, University of Malaya, 50603, Kuala Lumpur,Malaysia
bFaculty of Electrical Engineering, Universiti of Teknologi MARA, Malaysia
cDepartment of Cardiology, Hospital Pantai Putri Ipoh, Perak, Malaysia
dHealthmedic Research/Universiti Kebangsan, Malaysia
Abstract
The study on risk classification was conducted on 183 hospitalized dengue patients (4 DF, 179 DHF).The severity of the risk criteria was determined based on three blood investigations, namely, platelet(PLT) count (less than or equal to 30 000 cells per mm3), haematocrit (HCT) (increase by more than orequal to 20%), and either aspartate aminotransferase (AST) level (raised by 5-fold the normal upperlimit) or alanine aminotransferase (ALT) level (raised by 5-fold the normal upper limit). The patientswere divided into three groups based on their risk factors and the corresponding bioelectrical impedanceanalysis (BIA) (i.e. bioelectrical tissue conductivity (BETC) parameters), namely, resistance (R), phaseangle (α), body capacitance (BC) and capacitive reactance (XC) were obtained and quantified. Usinglogistic regression analysis, XC was found to be the best predictor in predicting the risk and severity indengue patients. Subsequent two-way analysis of variance (ANOVA) found that there was a statisticallysignificant relationship between the group categories and the change in XC.
Hence, it was shown that BIA, as reflected by the XC value, can effectively segregate between thelower-risk (female mean XC = 60.70Ω, range 58.09Ω–63.43Ω; male mean XC = 62.17Ω, range 59.92Ω–64.46Ω) and the higher-risk dengue patients (female mean XC = 43.99Ω, range 42.05Ω–45.97Ω; malemean XC = 50.65Ω range 49.25Ω–52.09Ω), with the control data ((female mean XC =69.41Ω, range67.09Ω–71.74Ω), and (male mean XC =64.19Ω, range 61.37Ω–67.15Ω)).
Keywords: Dengue infections; Bioelectrical impedance; Classify; Risk; Reactance.
Introduction
Dengue fever was first reported in Malaya (theformer name for Malaysia) after an epidemicin Penang in 1902.[1,2] Since then the disease
has become endemic in the country with casesreported throughout the year. In 1998, therewere 27 373 dengue cases with 58 deathsreported, as compared to 19 544 cases with50 deaths in 1997. This has shown an increase

Risk measurement of dengue infection using bioelectrical impedance analysis
Dengue Bulletin – Volume 31, 2007 59
of 7829 cases or 40.1% in one year.[3]
However, the dengue haemorrhagic fever(DHF) case-fatality rate (CFR) increased to1.7%.[4] In 2001, there was a sharp increase indengue cases in the country according to thevector-borne disease unit of the Ministry ofHealth, Malaysia.[5] In 2003, the serologicallyconfirmed dengue cases totaled 14 170, whichnumber was slightly lower than the previousyear (15 493). The CFR dropped to 4.5% ascompared to 5.05% in 2002. The most deathswere seen in the 5–9-year-old age group, withadults mainly in the 20–35 years age groups.The dengue cases have seen a rise of 16%every year since 2003.[6] Fatalities related todengue cases reached record levels in 2004,when 102 fatalities were reported.[6] In January2005, the number of infections in parts ofMalaysia rose significantly, with about 250suspected dengue cases reported each weekin Kuala Lumpur, compared to an average of100 cases weekly in December 2003. Anincrease of 34.97%, representing 1416 cases,was recorded during the second week (Jan 9-15) of the year compared to the previous week(1049 cases).[7] Forty-four dengue patients diedin the first four months of 2007 out of 16 214cases reported, compared to 21 deaths and10 244 cases in the same period last year.[6]
Due to the constantly high fatality rate, anaccurate determination of the prognosis of thedisease plays a very important role in the patientmanagement, so that the high risk patients canbe treated more intensively in their early phaseto improve survival. A non-invasive prognosticsystem for dengue infection is yet to bedeveloped and deployed in Malaysia. Due tothe endemic dengue phenomenon in thecountry, there is a real and obvious need forsuch a prognostic system to be made availableas soon as possible.
This paper describes a novel non-invasiveapproach to monitor and classify the daily riskin dengue patients using the single-frequency
bioelectrical impedance technique. Theresearch findings illustrate that any change inthe value of reactance (XC) will indicate thechanges in electrical conductivity of the bodyand this can be used to monitor and classifythe risk severity in dengue patients.
Principles of bioelectricalimpedance in humans
Impedance (Z) is a measurement of the abilityof a medium to conduct current. All conductivematerials have impedance, including livingtissues. The impedance of various organismsis known as bioelectrical impedance, whichmay vary seasonally, diurnally, and whenstimulated with an assortment of stimuli. Anexample of the said stimulus includesphysiological changes that occur when a bodyis affected by fever caused by bacterial andviral infections.
Bioelectrical impedance analysis (BIA) isthe assessment of changes in electrical tissueconductivity that indicate altered bodycomposition. The impedance of the body ismade up of two components. The resistance(R) is the index of conductivity determined byenergy-dissipating characteristics of the body.The reactance (XC) is the index of conductivitydetermined by the energy-storingcharacteristics of the body.
In the healthy living body, the cellmembrane consists of a layer of non-conductivelipid material sandwiched between two layersof conductive protein molecules. The structureof cell membranes makes them capacitivereactive elements which behave as capacitorswhen exposed to an alternating current (Figures1a & 1b).[8] Although the total body water andextracellular water offer resistance to electricalcurrent, only cell membranes offer capacitivereactance. Since fat tissue cells are notsurrounded by cell membranes, reactance is

60 Dengue Bulletin – Volume 31, 2007
Risk measurement of dengue infection using bioelectrical impedance analysis
not affected by the quantity of body fat. Typicaltotal body BIA measurements display thevectors of resistance and reactance, which areintrinsically based on a series network ofresistors and capacitors (Figure 2).
Biologically, the cell membrane functionsas a selectively permeable barrier separatingthe intracellular and extracellular fluidcompartments. It protects the interior of the
cell while allowing passage of some materialsto which it is permeable. The cell membranemaintains a fluid osmotic pressure and ionconcentration gradient between theintracellular and extracellular compartments.This gradient creates an electrical potentialdifference across the membrane which isessential to cell survival. Damage to the cellmembrane, and its functions, is as lethal tothe cell as direct damage to the nucleus itself.
Figure 1(a): The plasma membrane of cell
Extacellular spaceResistor
Protein channelCytoplasma
Pumpprotein
ProteinDielectric
Capacitor
Lipid bilayer
Intracellularspace
Figure 1(b): Plasma cell membrane electrical equivalent circuit
Dielectric
Resistor
Capacitor
Resistor
Capacitor
Capacitor
Extracellular Fluid
Intracellular FluidCell Membrane
Cell Nucleous
Dielectric
Dielectric
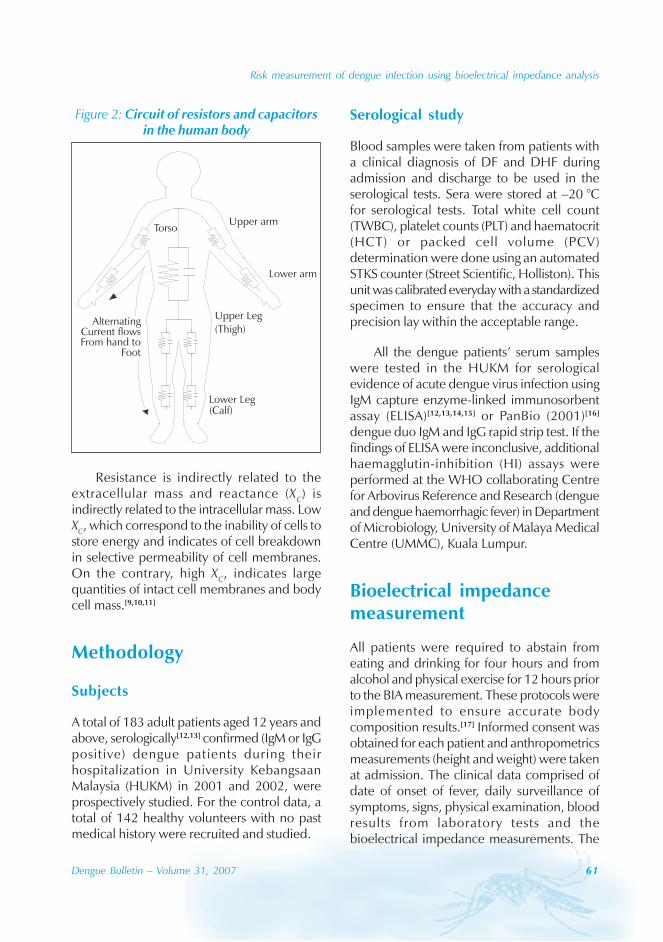
Risk measurement of dengue infection using bioelectrical impedance analysis
Dengue Bulletin – Volume 31, 2007 61
Resistance is indirectly related to theextracellular mass and reactance (XC) isindirectly related to the intracellular mass. LowXC, which correspond to the inability of cells tostore energy and indicates of cell breakdownin selective permeability of cell membranes.On the contrary, high XC, indicates largequantities of intact cell membranes and bodycell mass.[9,10,11]
Methodology
Subjects
A total of 183 adult patients aged 12 years andabove, serologically[12,13] confirmed (IgM or IgGpositive) dengue patients during theirhospitalization in University KebangsaanMalaysia (HUKM) in 2001 and 2002, wereprospectively studied. For the control data, atotal of 142 healthy volunteers with no pastmedical history were recruited and studied.
Serological study
Blood samples were taken from patients witha clinical diagnosis of DF and DHF duringadmission and discharge to be used in theserological tests. Sera were stored at –20 °Cfor serological tests. Total white cell count(TWBC), platelet counts (PLT) and haematocrit(HCT) or packed cell volume (PCV)determination were done using an automatedSTKS counter (Street Scientific, Holliston). Thisunit was calibrated everyday with a standardizedspecimen to ensure that the accuracy andprecision lay within the acceptable range.
All the dengue patients’ serum sampleswere tested in the HUKM for serologicalevidence of acute dengue virus infection usingIgM capture enzyme-linked immunosorbentassay (ELISA)[12,13,14,15] or PanBio (2001)[16]
dengue duo IgM and IgG rapid strip test. If thefindings of ELISA were inconclusive, additionalhaemagglutin-inhibition (HI) assays wereperformed at the WHO collaborating Centrefor Arbovirus Reference and Research (dengueand dengue haemorrhagic fever) in Departmentof Microbiology, University of Malaya MedicalCentre (UMMC), Kuala Lumpur.
Bioelectrical impedancemeasurement
All patients were required to abstain fromeating and drinking for four hours and fromalcohol and physical exercise for 12 hours priorto the BIA measurement. These protocols wereimplemented to ensure accurate bodycomposition results.[17] Informed consent wasobtained for each patient and anthropometricsmeasurements (height and weight) were takenat admission. The clinical data comprised ofdate of onset of fever, daily surveillance ofsymptoms, signs, physical examination, bloodresults from laboratory tests and thebioelectrical impedance measurements. The
Figure 2: Circuit of resistors and capacitorsin the human body
Lower arm
Upper armTorso
Upper Leg(Thigh)
Lower Leg(Calf)
AlternatingCurrent flowsFrom hand to
Foot

62 Dengue Bulletin – Volume 31, 2007
Risk measurement of dengue infection using bioelectrical impedance analysis
clinical data, weight and bioelectricalimpedance measurement were taken daily, onadmission, and upon discharge. This in vivotechnique involved the application of a smallaverage constant current of less than 1 mA ata single frequency of 50 kHz through the humanbody, and measuring the body’s bioelectricaltissue conductivity (BETC) parameters, namely,R, phase angle (α), body capacitance (BC), andcapacitive reactance (XC) via four surfaceelectrodes.
Two electrodes were placed on thepatient’s right hand, one at the base of theknuckles and another slightly above the wristjoint. Another two electrodes were placed onthe right foot; one near the base of the toesand the other slightly above the ankle joint. Aconstant current was applied to the base ofthe knuckles and base of the toes, and thesignal was picked up by the other two sensorelectrodes (slightly above the ankle and wristjoint) as shown in Figure 3.
Study protocol and riskclassification
The clinical diagnosis and severity of denguehaemorrhagic fever grade I to IV were basedon WHO recommendations.[18] Since thepatients were admitted at different stages oftheir illness, the daily progress of the patientswas dated with reference to the day the feversubsided (‘Fever day +0’). Hence, ‘Fever day+0’ is defined as the day the fever subsided,i.e. when the body temperature fell below37.5 °C. Days prior to the fever day +0 weredesignated as ‘Fever day –1’ (one day beforethe fever subsided), etc. Days after the feversubsided are designated as ‘Fever day +1’ (oneday after the fever subsided) and so on.
In this study, the severity of dengue riskcriteria was determined based on the earlierresearch findings[18,19,20,21,22,23,24] and confirmedhaematological profile (PLT, HCT, aspartate
Figure 3: Bioelectrical impedance measurement test position andplacement of sensor electrodes

Risk measurement of dengue infection using bioelectrical impedance analysis
Dengue Bulletin – Volume 31, 2007 63
aminotransferase (AST), alanine amino-transfansferase (ALT)) analysis finding in thisresearch. An exhaustive literature reviewconducted by Bandyopadhya et al.[24] revealsthe difficulties of using the WHO caseclassification for DHF. It found that mostclinicians reported difficulties in meeting all thefour criteria: (i) fever; (ii) haemorrhagictendencies; (iii) thrombocytopenia (100 000cells per mm3); and (iv) HCT >20% set byWHO. Therefore, most clinicians used themodified classification in their study. This studyalso faced the same problem and the criteriaof the risk were not based on WHOclassification solely; therefore, somemodification was made. Thus, the severity ofdengue risk criteria was determined based onthe following blood investigations: (i) platelet(PLT) count was less than or equal to 30 000cells per mm3;[19,20] (ii) haematocrit (HCT)increase by more than or equal to 20%;[18] and(iii) aspartate aminotransferase (AST) rose by5-fold the normal upper limit for AST[21,22,23] andalanine aminotransfansferase (ALT) rose by 5-fold the normal upper limit for ALT.[21,22,23] Thepatients were divided into three groups basedon their risk factors: (i) Group 1 (lower riskgroup) accounted for DHF patients who didnot experience any of the defined risk criteria,or experienced only one of the three riskcriteria; (ii) Group 2 (higher risk group)accounted for DHF patients who experiencedtwo or more risk criteria; and (iii) Group 3(control data) was the control group for healthyfemale and male subjects. Once the denguepatients were classified according to theirgroups, the corresponding BETC parameterswere subsequently obtained and quantified.
Statistical analysis
Data were analysed using the SPSS statisticalpackage version 10.01 for Windows 1998.Kolmogorov-Smirnov test and simple linearregression were used for examining the
normality assumption and for tests ofsignificance. If the variable was not normallydistributed, natural logarithm was used totransform the variables. Using logistic regressionanalysis, XC was found to be the best predictorin predicting the risk factors in denguepatients.[23] Subsequent two-way analysis ofvariance (ANOVA) found that there was astatistically significant relationship between thegroup categories and the change in XC. Themean of HCT and XC in the Groups 1 and 2with fluid infusion and without were comparedto find any significant changes.
Results and discussions
Clinical dengue patients
The 142 healthy controls (91 females and 51males) had their BIA measured. The clinical dataand BIA measurements were made on 97 maleand 86 female DF and DHF patients during theirhospitalization. The ages of the patients variedbetween 12 and 83 years (mean of 30.8 years).The length of hospital stay ranged from 2 to 18days (mean of 4.7 days). Three female and onemale patients were clinically diagnosed as DF,83 as DHF I (39 females and 44 males), 92 asDHF II (40 females and 52 males) and 4 femalepatients as dengue shock syndrome (DSS). NoDSS or fatal case was reported in the male DHFpatients.
The detailed descriptive analysis ofhaematological profile (97 males and 86 femaledengue patients) for PLT, HCT, AST and ALTare tabulated in Table 1. For example, theminimum platelet count recorded on the dayof fever defervescence (‘Fever day 0’) was8000 cell/mm3, while the maximum was136 000 cell/mm3, with a mean of 37 400 ±25 cell/mm3 for the male patients. The females’minimum and maximum documented plateletcounts were 7000 cell/mm3 and 162 000 cell/mm3 respectively with a mean of 48 000 ± 35
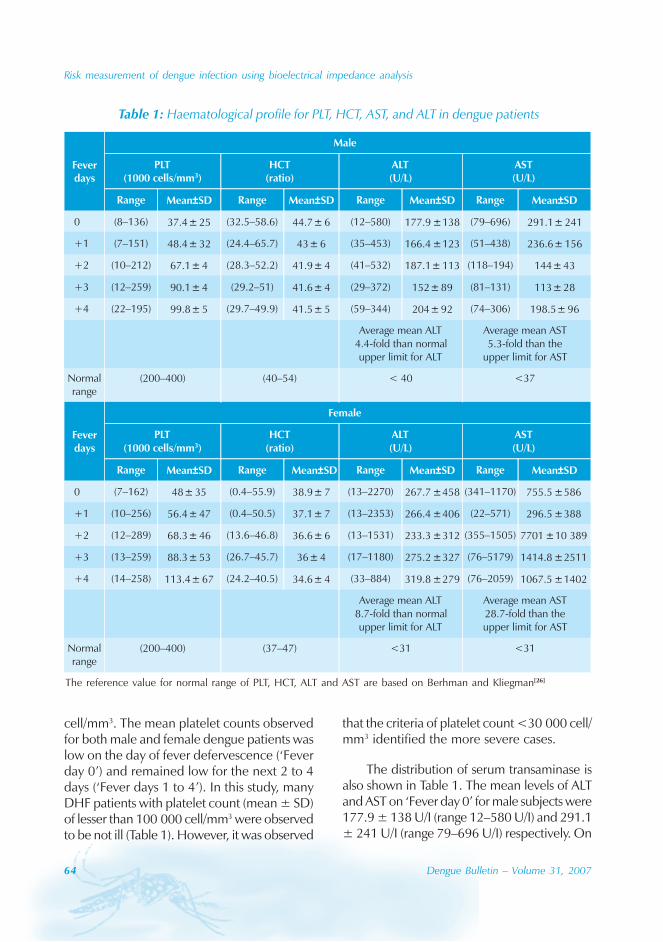
64 Dengue Bulletin – Volume 31, 2007
Risk measurement of dengue infection using bioelectrical impedance analysis
cell/mm3. The mean platelet counts observedfor both male and female dengue patients waslow on the day of fever defervescence (‘Feverday 0’) and remained low for the next 2 to 4days (‘Fever days 1 to 4’). In this study, manyDHF patients with platelet count (mean ± SD)of lesser than 100 000 cell/mm3 were observedto be not ill (Table 1). However, it was observed
that the criteria of platelet count <30 000 cell/mm3 identified the more severe cases.
The distribution of serum transaminase isalso shown in Table 1. The mean levels of ALTand AST on ‘Fever day 0’ for male subjects were177.9 ± 138 U/I (range 12–580 U/I) and 291.1± 241 U/I (range 79–696 U/I) respectively. On
Table 1: Haematological profile for PLT, HCT, AST, and ALT in dengue patients
The reference value for normal range of PLT, HCT, ALT and AST are based on Berhman and Kliegman[26]

Risk measurement of dengue infection using bioelectrical impedance analysis
Dengue Bulletin – Volume 31, 2007 65
the other hand, the female mean levels of ALTand AST on ‘Fever day 0’ were 267.7 ± 458 U/I (range 13–2270 U/I) and 755.5 ± 586 U/I(range 341–1170 U/I) respectively. The resultsindicate that both male and female denguepatients had liver involvement (hepatocyteinjury), ranging from mild to moderate degrees,as indicated by the elevation in the transaminaselevels (>5-fold greater than the normal upperlimit of AST and ALT).
It was also observed that the AST level washigher than that of ALT level in female denguepatients, while in the male patients, it onlyoccurred on fever days 0 and +1. The femalemean levels of AST and ALT were higher (theaverage for fever days 0 to +4 was >9-foldgreater than the normal upper limit of AST andALT) than the value for male subjects for all feverdays (the average for fever days 0 to 4 was <5-fold greater than the normal upper limit for theAST and ALT), as illustrated in Table 1. Thesefindings are in agreement with those of Nguyenet al.[25] and Kuo et al.,[21] who documentedthat the elevation in the transaminase levels were<5-fold and >10-fold greater than the normalupper limit for AST and ALT respectively. Thus,the criteria of 5-fold rise above the normal upperlimit for AST and ALT were chosen.
The mean HCT levels for both female andmale dengue patients were not much elevatedwith subsequent fever days. For example, on‘Fever day 0’ the female and male HCT valueswere 38.9 ± 7% and 44.7 ± 6% respectively.The overall descriptive analysis of haematologicalprofile (Table 1) showed that constant
abnormalities occur in PLT, AST and ALT, whileHCT only has a moderate elevation.
In the group’s risk categorization, 91 (49.7%)dengue patients were in the lower risk group,while 92 (50.3%) were in the higher risk group.The detailed dengue diagnosis breakdown isshown in Table 2. The higher risk group consistsof 92 DHF patients, which was made up of 32(17.5%) DHF I, 56 (30.6%) DHF II, and 4 (2.2%)DHF IV/DSS patients. The detailed descriptiveanalysis of haematological profile for PLT, HCT,AST and ALT in both lower and higher-risk patientgroups in the duration of ‘Fever day 0’ to ‘Feverday +4’ are tabulated in Table 3.
It can be observed that both groupsproduced abnormal PLT, HCT, ALT and ALTlevels. Patients in the higher-risk group (Group2) produced the lowest PLT values with anaverage of 7000 cells/mm3 for both genders.The distribution of serum transaminases (ASTand ALT) indicate that the females in Group 2had their ALT levels raised by 76-fold (2353/31)the normal upper limit (<31),[26] whereas theAST levels were raised by 167-fold (5179/31)the normal upper limit (<31).[26] On the otherhand, the ALT and AST values for males in Group2 were only raised by 14.5-fold and 18.8-foldthe normal upper limits of ALT and AST,respectively (Table 3). These results show thatpatients in the higher-risk group (Group 2) hada high degree of liver involvement. In addition,the HCT values for female (55.9) and male(65.7) subjects in Group 2 had exceeded theupper normal values (37–47) of HCT at 47 and54 respectively (see Table 3).
Table 2: Dengue diagnosis breakdown according to risk group
Groups DF DHF I DHF II DHF IV/DSS Total cases
Lower risk (Group 1) 4 51 36 Nil 91
Higher risk (Group 2) Nil 32 56 4 92
Dengue diagnosis cases 4 83 92 4 183
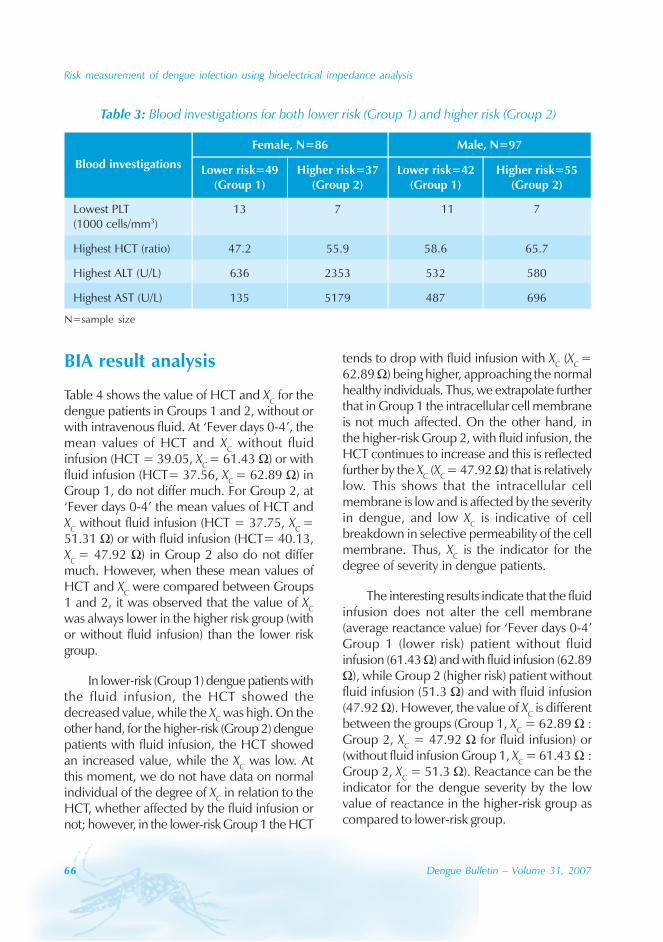
66 Dengue Bulletin – Volume 31, 2007
Risk measurement of dengue infection using bioelectrical impedance analysis
BIA result analysis
Table 4 shows the value of HCT and XC for thedengue patients in Groups 1 and 2, without orwith intravenous fluid. At ‘Fever days 0-4’, themean values of HCT and XC without fluidinfusion (HCT = 39.05, XC = 61.43 Ω) or withfluid infusion (HCT= 37.56, XC = 62.89 Ω) inGroup 1, do not differ much. For Group 2, at‘Fever days 0-4’ the mean values of HCT andXC without fluid infusion (HCT = 37.75, XC =51.31 Ω) or with fluid infusion (HCT= 40.13,XC = 47.92 Ω) in Group 2 also do not differmuch. However, when these mean values ofHCT and XC were compared between Groups1 and 2, it was observed that the value of XCwas always lower in the higher risk group (withor without fluid infusion) than the lower riskgroup.
In lower-risk (Group 1) dengue patients withthe fluid infusion, the HCT showed thedecreased value, while the XC was high. On theother hand, for the higher-risk (Group 2) denguepatients with fluid infusion, the HCT showedan increased value, while the XC was low. Atthis moment, we do not have data on normalindividual of the degree of XC in relation to theHCT, whether affected by the fluid infusion ornot; however, in the lower-risk Group 1 the HCT
tends to drop with fluid infusion with XC (XC =62.89 Ω) being higher, approaching the normalhealthy individuals. Thus, we extrapolate furtherthat in Group 1 the intracellular cell membraneis not much affected. On the other hand, inthe higher-risk Group 2, with fluid infusion, theHCT continues to increase and this is reflectedfurther by the XC (XC = 47.92 Ω) that is relativelylow. This shows that the intracellular cellmembrane is low and is affected by the severityin dengue, and low XC is indicative of cellbreakdown in selective permeability of the cellmembrane. Thus, XC is the indicator for thedegree of severity in dengue patients.
The interesting results indicate that the fluidinfusion does not alter the cell membrane(average reactance value) for ‘Fever days 0-4’Group 1 (lower risk) patient without fluidinfusion (61.43 Ω) and with fluid infusion (62.89Ω), while Group 2 (higher risk) patient withoutfluid infusion (51.3 Ω) and with fluid infusion(47.92 Ω). However, the value of XC is differentbetween the groups (Group 1, XC = 62.89 Ω :Group 2, XC = 47.92 Ω for fluid infusion) or(without fluid infusion Group 1, XC = 61.43 Ω :Group 2, XC = 51.3 Ω). Reactance can be theindicator for the dengue severity by the lowvalue of reactance in the higher-risk group ascompared to lower-risk group.
Table 3: Blood investigations for both lower risk (Group 1) and higher risk (Group 2)
N=sample size
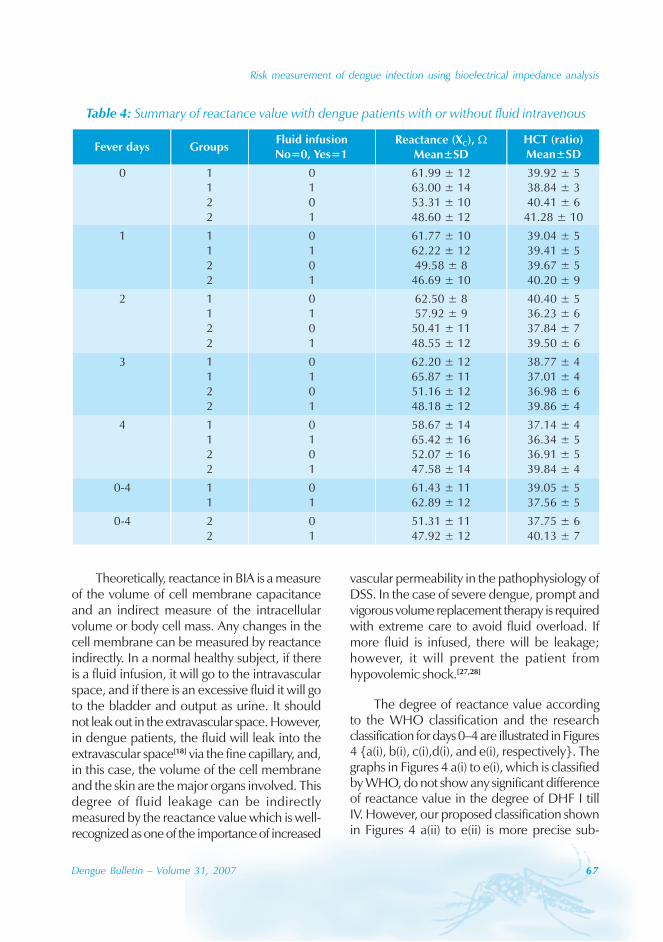
Risk measurement of dengue infection using bioelectrical impedance analysis
Dengue Bulletin – Volume 31, 2007 67
Theoretically, reactance in BIA is a measureof the volume of cell membrane capacitanceand an indirect measure of the intracellularvolume or body cell mass. Any changes in thecell membrane can be measured by reactanceindirectly. In a normal healthy subject, if thereis a fluid infusion, it will go to the intravascularspace, and if there is an excessive fluid it will goto the bladder and output as urine. It shouldnot leak out in the extravascular space. However,in dengue patients, the fluid will leak into theextravascular space[18] via the fine capillary, and,in this case, the volume of the cell membraneand the skin are the major organs involved. Thisdegree of fluid leakage can be indirectlymeasured by the reactance value which is well-recognized as one of the importance of increased
vascular permeability in the pathophysiology ofDSS. In the case of severe dengue, prompt andvigorous volume replacement therapy is requiredwith extreme care to avoid fluid overload. Ifmore fluid is infused, there will be leakage;however, it will prevent the patient fromhypovolemic shock.[27,28]
The degree of reactance value accordingto the WHO classification and the researchclassification for days 0–4 are illustrated in Figures4 {a(i), b(i), c(i),d(i), and e(i), respectively}. Thegraphs in Figures 4 a(i) to e(i), which is classifiedby WHO, do not show any significant differenceof reactance value in the degree of DHF I tillIV. However, our proposed classification shownin Figures 4 a(ii) to e(ii) is more precise sub-
Table 4: Summary of reactance value with dengue patients with or without fluid intravenous
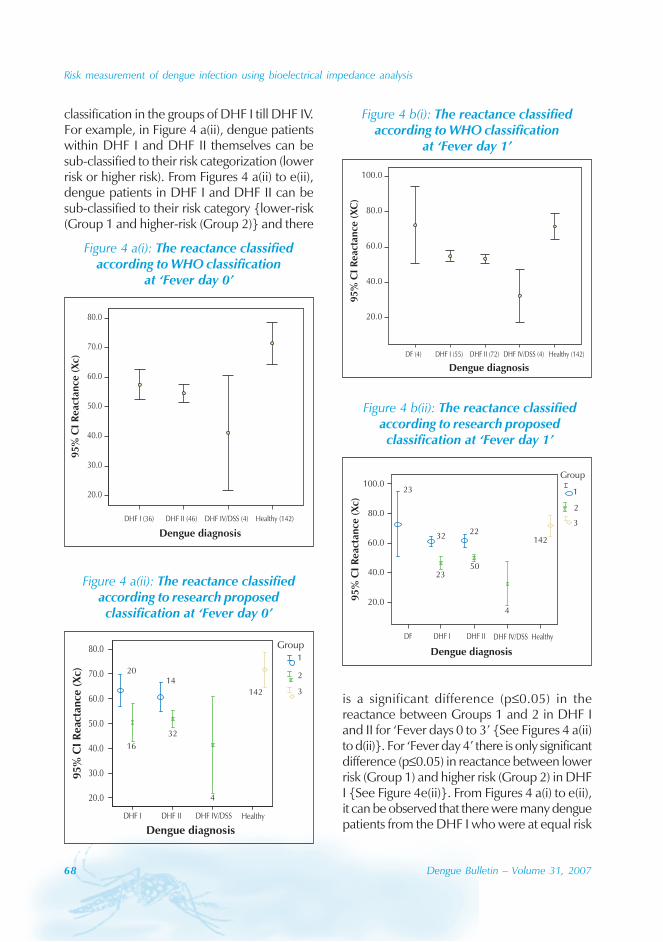
68 Dengue Bulletin – Volume 31, 2007
Risk measurement of dengue infection using bioelectrical impedance analysis
classification in the groups of DHF I till DHF IV.For example, in Figure 4 a(ii), dengue patientswithin DHF I and DHF II themselves can besub-classified to their risk categorization (lowerrisk or higher risk). From Figures 4 a(ii) to e(ii),dengue patients in DHF I and DHF II can besub-classified to their risk category {lower-risk(Group 1 and higher-risk (Group 2)} and there
is a significant difference (p≤0.05) in thereactance between Groups 1 and 2 in DHF Iand II for ‘Fever days 0 to 3’ {See Figures 4 a(ii)to d(ii)}. For ‘Fever day 4’ there is only significantdifference (p≤0.05) in reactance between lowerrisk (Group 1) and higher risk (Group 2) in DHFI {See Figure 4e(ii)}. From Figures 4 a(i) to e(ii),it can be observed that there were many denguepatients from the DHF I who were at equal risk
Figure 4 a(i): The reactance classifiedaccording to WHO classification
at ‘Fever day 0’
DHF I (36) DHF II (46) DHF IV/DSS (4) Healthy (142)
Dengue diagnosis
20.0
30.0
40.0
50.0
60.0
70.0
80.0
95
%C
IR
eact
ance
(Xc)
Figure 4 a(ii): The reactance classifiedaccording to research proposedclassification at ‘Fever day 0’
95
%C
IR
eact
ance
(Xc)
Dengue diagnosis
DHF I DHF II DHF IV/DSS Healthy
20.0
30.0
40.0
50.0
60.0
70.0
80.0
16
32
4
142
2014
Group1
2
3
Figure 4 b(i): The reactance classifiedaccording to WHO classification
at ‘Fever day 1’
DF (4) DHF I (55) DHF II (72) DHF IV/DSS (4) Healthy (142)
Dengue diagnosis
20.0
40.0
60.0
80.0
100.0
95%
CI
Rea
ctan
ce(X
C)
Figure 4 b(ii): The reactance classifiedaccording to research proposedclassification at ‘Fever day 1’
95
%C
IR
eac
tan
ce(X
c)
1
2
3
23
3222
142
4
5023
20.0
40.0
60.0
80.0
100.0
DF DHF I DHF II DHF IV/DSS Healthy
Dengue diagnosis
Group
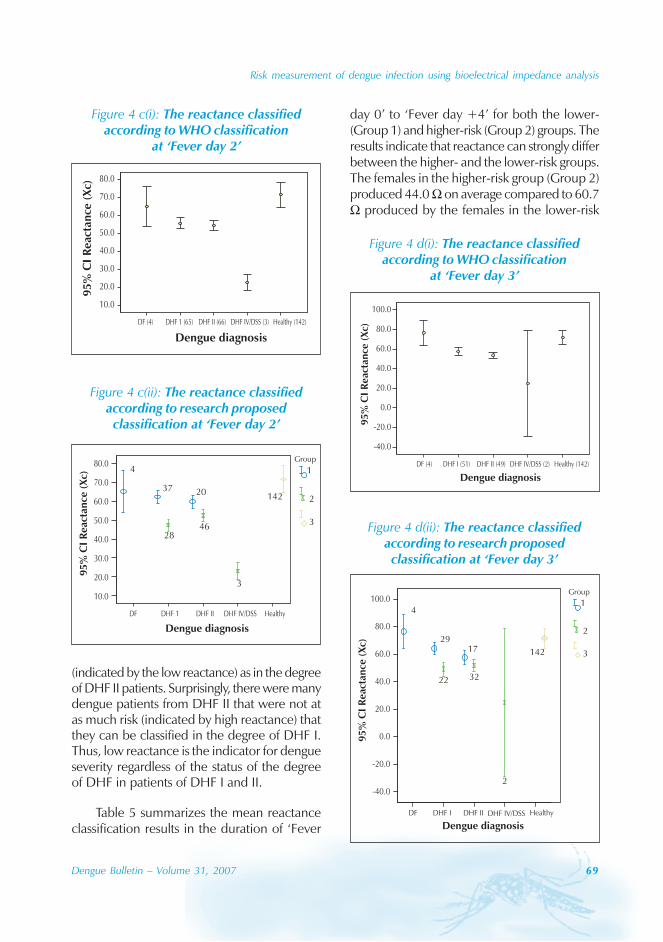
Risk measurement of dengue infection using bioelectrical impedance analysis
Dengue Bulletin – Volume 31, 2007 69
DF (4) DHF 1 (65) DHF II (66) DHF IV/DSS (3) Healthy (142)
Dengue diagnosis
10.0
20.0
30.0
40.0
50.0
60.0
70.0
80.0
95%
CI
Reac
tan
ce(X
c)
Figure 4 c(i): The reactance classifiedaccording to WHO classification
at ‘Fever day 2’
Figure 4 c(ii): The reactance classifiedaccording to research proposedclassification at ‘Fever day 2’
DF DHF 1 DHF II DHF IV/DSS Healthy
Dengue diagnosis
10.0
20.0
30.0
40.0
50.0
60.0
70.0
80.0
95
%C
IR
eact
ance
(Xc)
46
3
142
4
37 20
28
Group
1
2
3
Figure 4 d(i): The reactance classifiedaccording to WHO classification
at ‘Fever day 3’
DF (4) DHF I (51) DHF II (49) DHF IV/DSS (2) Healthy (142)
Dengue diagnosis
-40.0
-20.0
0.0
20.0
40.0
60.0
80.0
100.0
95%
CI
Reac
tan
ce(X
c)
Figure 4 d(ii): The reactance classifiedaccording to research proposedclassification at ‘Fever day 3’
DF DHF I DHF II DHF IV/DSS Healthy
-40.0
-20.0
0.0
20.0
40.0
60.0
80.0
100.0
32
2
142
22
1729
4
Group
1
2
3
95
%C
IR
eact
ance
(Xc)
Dengue diagnosis
(indicated by the low reactance) as in the degreeof DHF II patients. Surprisingly, there were manydengue patients from DHF II that were not atas much risk (indicated by high reactance) thatthey can be classified in the degree of DHF I.Thus, low reactance is the indicator for dengueseverity regardless of the status of the degreeof DHF in patients of DHF I and II.
Table 5 summarizes the mean reactanceclassification results in the duration of ‘Fever
day 0’ to ‘Fever day +4’ for both the lower-(Group 1) and higher-risk (Group 2) groups. Theresults indicate that reactance can strongly differbetween the higher- and the lower-risk groups.The females in the higher-risk group (Group 2)produced 44.0 Ω on average compared to 60.7Ω produced by the females in the lower-risk
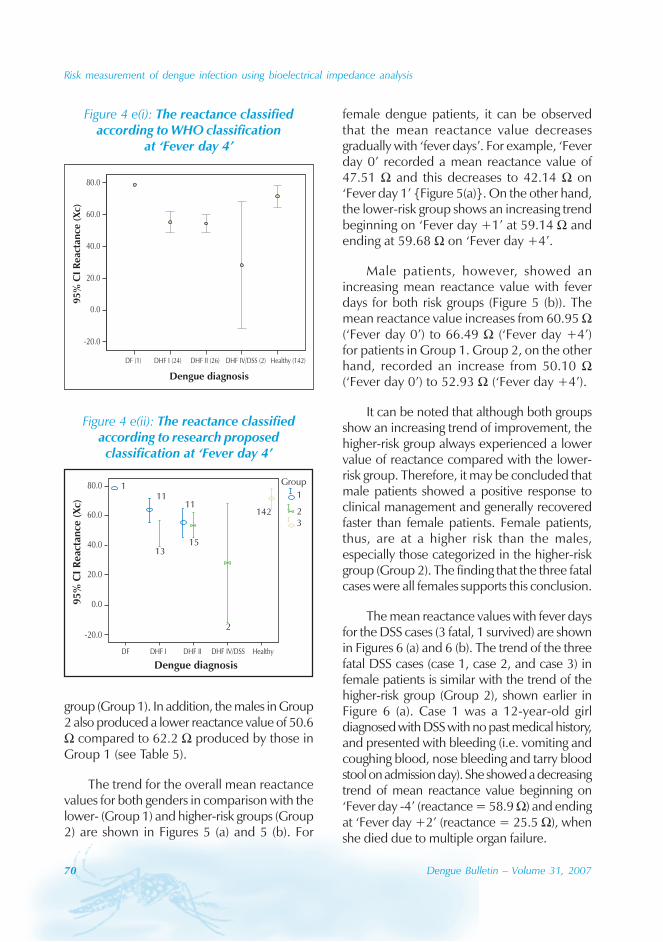
70 Dengue Bulletin – Volume 31, 2007
Risk measurement of dengue infection using bioelectrical impedance analysis
group (Group 1). In addition, the males in Group2 also produced a lower reactance value of 50.6Ω compared to 62.2 Ω produced by those inGroup 1 (see Table 5).
The trend for the overall mean reactancevalues for both genders in comparison with thelower- (Group 1) and higher-risk groups (Group2) are shown in Figures 5 (a) and 5 (b). For
female dengue patients, it can be observedthat the mean reactance value decreasesgradually with ‘fever days’. For example, ‘Feverday 0’ recorded a mean reactance value of47.51 Ω and this decreases to 42.14 Ω on‘Fever day 1’ {Figure 5(a)}. On the other hand,the lower-risk group shows an increasing trendbeginning on ‘Fever day +1’ at 59.14 Ω andending at 59.68 Ω on ‘Fever day +4’.
Male patients, however, showed anincreasing mean reactance value with feverdays for both risk groups (Figure 5 (b)). Themean reactance value increases from 60.95 Ω(‘Fever day 0’) to 66.49 Ω (‘Fever day +4’)for patients in Group 1. Group 2, on the otherhand, recorded an increase from 50.10 Ω(‘Fever day 0’) to 52.93 Ω (‘Fever day +4’).
It can be noted that although both groupsshow an increasing trend of improvement, thehigher-risk group always experienced a lowervalue of reactance compared with the lower-risk group. Therefore, it may be concluded thatmale patients showed a positive response toclinical management and generally recoveredfaster than female patients. Female patients,thus, are at a higher risk than the males,especially those categorized in the higher-riskgroup (Group 2). The finding that the three fatalcases were all females supports this conclusion.
The mean reactance values with fever daysfor the DSS cases (3 fatal, 1 survived) are shownin Figures 6 (a) and 6 (b). The trend of the threefatal DSS cases (case 1, case 2, and case 3) infemale patients is similar with the trend of thehigher-risk group (Group 2), shown earlier inFigure 6 (a). Case 1 was a 12-year-old girldiagnosed with DSS with no past medical history,and presented with bleeding (i.e. vomiting andcoughing blood, nose bleeding and tarry bloodstool on admission day). She showed a decreasingtrend of mean reactance value beginning on‘Fever day -4’ (reactance = 58.9 Ω) and endingat ‘Fever day +2’ (reactance = 25.5 Ω), whenshe died due to multiple organ failure.
Figure 4 e(i): The reactance classifiedaccording to WHO classification
at ‘Fever day 4’
DF (1) DHF I (24) DHF II (26) DHF IV/DSS (2) Healthy (142)
-20.0
20.0
40.0
60.0
80.0
95
%C
IR
eact
ance
(Xc)
Dengue diagnosis
0.0
Figure 4 e(ii): The reactance classifiedaccording to research proposedclassification at ‘Fever day 4’
95
%C
IR
eact
ance
(Xc)
DF DHF I DHF II DHF IV/DSS Healthy
Dengue diagnosis
-20.0
20.0
40.0
60.0
80.0
0.0
142
15
2
13
111
111
23
Group
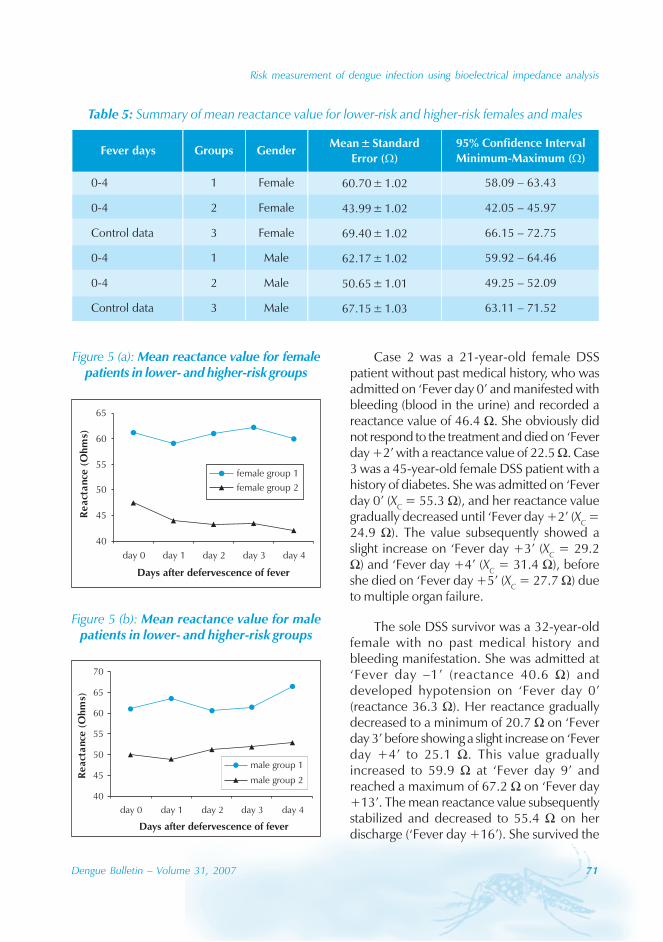
Risk measurement of dengue infection using bioelectrical impedance analysis
Dengue Bulletin – Volume 31, 2007 71
Case 2 was a 21-year-old female DSSpatient without past medical history, who wasadmitted on ‘Fever day 0’ and manifested withbleeding (blood in the urine) and recorded areactance value of 46.4 Ω. She obviously didnot respond to the treatment and died on ‘Feverday +2’ with a reactance value of 22.5 Ω. Case3 was a 45-year-old female DSS patient with ahistory of diabetes. She was admitted on ‘Feverday 0’ (XC = 55.3 Ω), and her reactance valuegradually decreased until ‘Fever day +2’ (XC =24.9 Ω). The value subsequently showed aslight increase on ‘Fever day +3’ (XC = 29.2Ω) and ‘Fever day +4’ (XC = 31.4 Ω), beforeshe died on ‘Fever day +5’ (XC = 27.7 Ω) dueto multiple organ failure.
The sole DSS survivor was a 32-year-oldfemale with no past medical history andbleeding manifestation. She was admitted at‘Fever day –1’ (reactance 40.6 Ω) anddeveloped hypotension on ‘Fever day 0’(reactance 36.3 Ω). Her reactance graduallydecreased to a minimum of 20.7 Ω on ‘Feverday 3’ before showing a slight increase on ‘Feverday +4’ to 25.1 Ω. This value graduallyincreased to 59.9 Ω at ‘Fever day 9’ andreached a maximum of 67.2 Ω on ‘Fever day+13’. The mean reactance value subsequentlystabilized and decreased to 55.4 Ω on herdischarge (‘Fever day +16’). She survived the
Table 5: Summary of mean reactance value for lower-risk and higher-risk females and males
Figure 5 (a): Mean reactance value for femalepatients in lower- and higher-risk groups
40
45
50
55
60
65
day 0 day 1 day 2 day 3 day 4
Days after defervescence of fever
Re
act
ance
(Oh
ms)
female group 1
female group 2
Figure 5 (b): Mean reactance value for malepatients in lower- and higher-risk groups
40
45
50
55
60
65
70
day 0 day 1 day 2 day 3 day 4
Days after defervescence of fever
Re
act
ance
(Oh
ms)
male group 1
male group 2
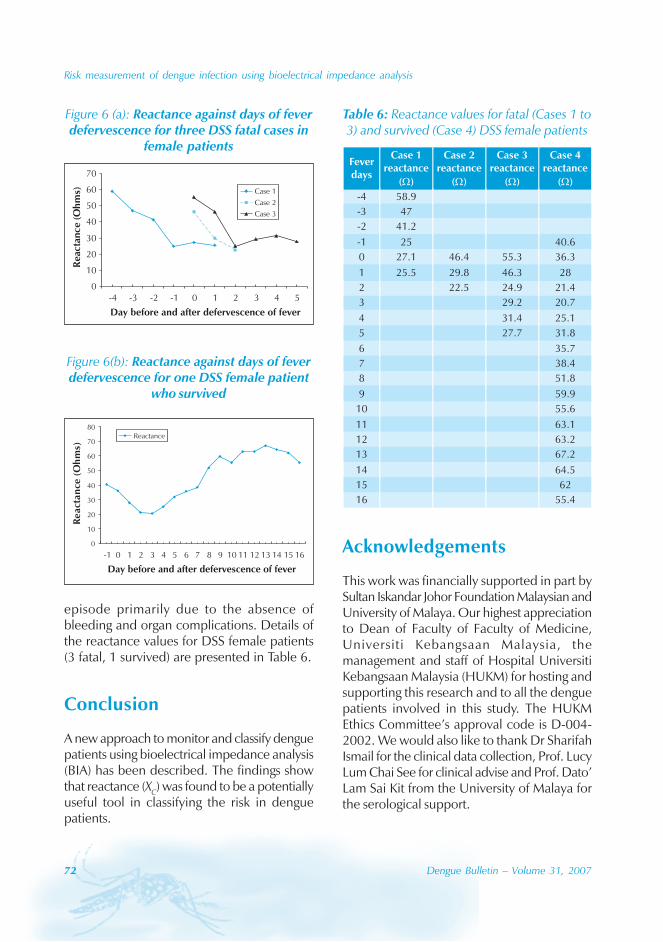
72 Dengue Bulletin – Volume 31, 2007
Risk measurement of dengue infection using bioelectrical impedance analysis
episode primarily due to the absence ofbleeding and organ complications. Details ofthe reactance values for DSS female patients(3 fatal, 1 survived) are presented in Table 6.
Conclusion
A new approach to monitor and classify denguepatients using bioelectrical impedance analysis(BIA) has been described. The findings showthat reactance (XC) was found to be a potentiallyuseful tool in classifying the risk in denguepatients.
Figure 6 (a): Reactance against days of feverdefervescence for three DSS fatal cases in
female patients
0
10
20
30
40
50
60
70
-4 -3 -2 -1 0 1 2 3 4 5
Day before and after defervescence of fever
Rea
ctan
ce(O
hm
s) Case 1
Case 2
Case 3
Figure 6(b): Reactance against days of feverdefervescence for one DSS female patient
who survived
0
10
20
30
40
50
60
70
80
-1 0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
Day before and after defervescence of fever
React
an
ce(O
hm
s)
Reactance
Table 6: Reactance values for fatal (Cases 1 to3) and survived (Case 4) DSS female patients
Acknowledgements
This work was financially supported in part bySultan Iskandar Johor Foundation Malaysian andUniversity of Malaya. Our highest appreciationto Dean of Faculty of Faculty of Medicine,Universiti Kebangsaan Malaysia, themanagement and staff of Hospital UniversitiKebangsaan Malaysia (HUKM) for hosting andsupporting this research and to all the denguepatients involved in this study. The HUKMEthics Committee’s approval code is D-004-2002. We would also like to thank Dr SharifahIsmail for the clinical data collection, Prof. LucyLum Chai See for clinical advise and Prof. Dato’Lam Sai Kit from the University of Malaya forthe serological support.

Risk measurement of dengue infection using bioelectrical impedance analysis
Dengue Bulletin – Volume 31, 2007 73
References
[1] Skae FM. Dengue fever in Penang. Br Med J.1902; 2:1581-1582.
[2] Gordon SCE. Dengue: An introduction. In:Rudnick A, Lim TW, Editors. Dengue feverstudies in Malaysia Institute for MedicalResearch. Malaysia Bulletin. 1986; 23:1-5.
[3] World Health Organization. Annual Report(1998) of the WHO Collaborating Centre forArbovirus Reference and Research (dengueand dengue haemorrhagic fever) byDepartment of Medical Microbiology, Facultyof Medicine, University of Malaya, 50603Kuala Lumpur; 1998.
[4] World Health Organization. Annual Report(1999/2000) of the WHO CollaboratingCentre for Arbovirus Reference and Research(dengue and dengue haemorrhagic fever) byDepartment of Medical Microbiology, Facultyof Medicine, University Malaya, 50603 KualaLumpur; 2000.
[5] World Health Organization. Annual Report(2001) of the WHO Collaborating Centre forArbovirus Reference and Research (dengueand dengue haemorrhagic fever) byDepartment of Medical Microbiology, Facultyof Medicine, University Malaya, 50603 KualaLumpur; 2001.
[6] Data released by the Health Ministry’s Directorof Disease Control, Hasan Abdul Rahman inNew Straits Times Newspaper (2 May 2007).
[7] Dengue and dengue haemorrhagic fever.Nation Pg.29, The Star Newspaper 2005 (15January 2005).
[8] Liedtke RJ. (1997). Principles of bioelectricalimpedance analysis. Available from:URL:http://www.rohan.sdsu.edu/~ens314/bia.tm
[9] Máttar JA. Application of total bodybioimpedance to the critically ill patient.Brazilian Group for Bioimpedance Study.New Horiz. 1996 Nov; 4(4): 493-503.
[10] Ott M, Fischer H, Polat H, Helm EB, Frenz M,Caspary WF, Lembcke B. Bioelectrical
impedance analysis as a predictor of survivalin patients with human immunodeficiencyvirus infection. J Acquir Immune Defic SyndrHum Retrovirol. 1995 May 1; 9(1): 20-5.
[11] Baumgartner RN. Electrical impedance andtotal body electrical conductivity. In: RocheAF, Heymsfield SB, Lohman TG Editors. Humanbody composition. Champaign, IL: HumanKinetics; 1996. p. 79-107.
[12] Chungue E, Boutin JP, Roux J. [Significance ofIgM titration by an immunoenzyme technicfor the serodiagnosis and epidemiologicalsurveillance of dengue in French Polynesia].Res Virol. 1989 May-Jun; 140(3): 229-40.French.
[13] Lam SK, Devi S, Pang T. Detection of specificIgM in dengue infection. Southeast Asian J TropMed Public Health 1987 Dec;18(4):532-8.
[14] Lam SK, Devine PL. Evaluation of captureELISA and rapid immunochromatographic testfor the determination of IgM and IgGantibodies produced during dengue infection.Clin Diagn Virol 1998 May 1;10(1):75-81.
[15] Lam SK. Application of rapid laboratorydiagnosis in dengue control. Asia Pac J MolBiol Biotechnol. 1995; 3: 351-355
[16] PanBio. Dengue duo IgM and IgG rapid striptest. Catalogue No. DEN-25S, 2001.
[17] Biodynamics Model 450 BioimpedanceAnalyzer user’s guide, “Basic Principles ofBioimpedance Testing”, 1st ed. Copyright ©Biodynamics Corporation; 2000. p. 1-22.
[18] World Health Organization. Denguehaemorrhagic fever, diagnosis, treatment,prevention and control. 2nd edn., Geneva:WHO, 1997.
[19] Chua MN, Molanida R, de Guzman M,Laberiza F. Prothrombin time and partialthromboplastin time as a predictor of bleedingin patients with dengue hemorrhagic fever.Southeast Asian J Trop Med Public Health.1993; 24 Suppl 1: 141-3.

74 Dengue Bulletin – Volume 31, 2007
Risk measurement of dengue infection using bioelectrical impedance analysis
[20] Tai DYH, Chee YC, Chan KW. The naturalhistory of dengue illness based on a study ofhospitalized patients in Singapore. SingaporeMed J. 1999; 40: 238-242.
[21] Kuo CH, Tai DI, Chang-Chien CS, Lan CK,Chiou SS, Liaw YF. Liver biochemical tests anddengue fever. Am J Trop Med Hyg. 1992 Sep;47(3): 265-70.
[22] Ibrahim F, Mohd NT, Wan Abas WAB,Sulaiman S, Chan CG. A novel approach toclassify risk in dengue haemorrhagic feverusing bioelectrical impedance analysis. IEEETransactions on Instrumentation andMeasurement. 2005 Feb; 54(1):237-244.
[23] Ibrahim F. Prognosis of dengue fever and denguehaemorrhagic fever using bioelectricalimpedance. PhD thesis. Department ofBiomedical Engineering , Faculty ofEngineering, University of Malaya, July 2005.
[24] Bandyopadhyay S, Lum LC, Kroeger A.Classifying dengue: a review of the difficultiesin using the WHO case classification fordengue haemorrhagic fever. Trop Med IntHealth. 2006 Aug;11(8):1238-55.
[25] Nguyen TL, Nguyen TH, Tieu NT. The impactof dengue haemorrhagic fever on liverfunction. Res Virol. 1997 Jul-Aug; 148(4):273-7.
[26] Berhman RE, Kliegman RM. Nelson essentialsof paediatrics. 4th edn. Philadelphia: WBSaunders Company, 2002.
[27] Ganong WF. Review of Medical Physiology. 21st
edn. New York: McGraw Hill, 2003.
[28] McPhee SJ, Lingappa VR, Ganong WF, LangeJD. Pathophysiology of disease: an introductionto clinical medicine. 2nd edn. Connecticut:Appleton & Lange Prentice Hall, 1997.

Dengue Bulletin – Volume 31, 2007 75
Utilization of a geographical information systemfor surveillance of Aedes aegypti and
dengue haemorrhagic fever in north-eastern Thailand
Anun Chaikoolvatanaa , Pratap Singhasivanonb and Peter Haddawyc
aDepartment of Pharmaceutical Sciences, Ubon Rajathanee University, Warinchumrab, Ubon Ratchathani,Thailand 34190
bFaculty of Tropical Medicine, Mahidol University, Thailand
cSchool of Advanced Technology, Asian Institute of Technology, Thailand
Abstract
The study aims to develop a geographical information system (GIS) for surveillance of Aedes aegypti anddengue haemorrhagic fever (DHF) in north-eastern Thailand. There are three steps in the developmentof the GIS – collecting primary and secondary data, analysing the data and searching the target location,and presenting the results via figures on a map. Two sub-districts in each of the five districts in UbonRatchathani province with high incidences of DHF cases in the last three years were investigated.Primary {e.g. House Index (HI), Container Index (CI) and Breteau Index (BI)} and secondary data (e.g.number of DHF cases/100 000 population) were collected. The time period was divided into twophases, a low disease incidence phase (February–March 2007) and a high disease incidence phase(June–July 2007). The GIS was developed via ArcView programme 3.2a®. The primary data of Ae.aegypti indices including HI, CI and BI indicated a rise in the rainy season period compared with thedry weather period, and the secondary data showed a similar rise and fall in the number of DHF casesin the rainy and dry weather periods respectively. GIS technology can help in planning, implementationand evaluation of the dengue control measures.
Keywords: Geographical information system (GIS); Ae. aegypti indices; Geographical positioning system (GPS);Dengue haemorrhagic fever (DHF).
E-mail: [email protected], [email protected]; Tel.: 00-66-45-353670; Fax: 00-66-45-288-384
Introduction
Dengue haemorrhagic fever (DHF) caused bythe mosquito species Aedes aegypti is one ofthe world’s major health problems.[1] InThailand, the reported cases of DHF vary from20 000 cases to more than 100 000 casesannually.[2] Recently, the incidence of DHF hasincreased in many provinces in the north-
eastern part of the country, including UbonRatchathani, Si Sa Ket, Yasothon and AmnatCharoen.[3,4] Ubon Ratchathani is ranked as theprovince with the fourth highest incidence ofDHF in Thailand with 172.20 cases per 100 000population (Figure 1).[5,6]
A geographical information system (GIS)is an initiative to support dengue control inThailand. It is an automated computer-based

76 Dengue Bulletin – Volume 31, 2007
Use of GIS for DHF surveillance in Thailand
system with the ability to capture, retrieve,manage, display and analyse large quantitiesof spatial and temporal data in a geographicalcontext. Roads, residential buildings and otherrelevant data can be obtained and mapped toform the base map layer using the softwareArcview GIS®. Other layers, such as patientpopulations, mosquito breeding, dengue caseincidences, addresses, sensitive areas andcontainers, can also be mapped into the GIS.[7-
11] Among the publications on the use of GISin DHF surveillance[12-18] is included oneconcerning the development of a databasesystem of mosquito breeding habitats. This ledto the development of a system called“Mosquito Information Retrieval System(MIRS)”.[13] Another study conducted at theNortre Dame University developed a computerprogramme called “MODABUND” that aimedto provide basic information about mosquitoessuch as breeding habitats, life cycles, types,epidemiology, diseases transmitted by
mosquitoes and means of surveillance andcontrol. [12] However, there have beenlimitations that include lack of professionals,restricted budget, inadequate knowledge of GISapplications and a lack of updated data. As aresult, significant benefits of the use of a GISin dengue surveillance and control have notbeen as successful as anticipated.[10,11,18,19]
Because of these limitations, researchers aretrying to develop a GIS programme to helppublic health officers gather data andinformation on DHF to assist in the control ofthis health problem.
This study is an attempt to develop a GISfor surveillance of Ae. aegypti and DHF in north-eastern Thailand.
Materials and methods
Study area
Ubon Ratchathani is located in north-easternThailand, about 629 kilometres from Bangkok(Figure 1). The terrain is generally flat with theriver Moon flowing through the central area. Thetotal area of the province is 15 517 sq. km. withhighlands and mountains in the east of theprovince. The total population is about1 600 000. There are three distinct seasons, dry(March-May), rainy (June-September) and cold(October-February). Average temperatures arefrom 25 °C to 35 °C depending on the season.The average rainfall is 1300–1800 mm per year.
Implementing an ArcViewprogramme 3.2a®
The GIS requires a language commandprogramme called “Avenue” with the DialogDesign function of ArcView software 3.2a®.Users follow the menu and extract the requireddata. The overall structural functions include:
Figure 1: Map of Thailand indicatinglocation of Ubon Ratchathani province
ThailandUbonRatchathani
Use of GIS for DHF surveillance in Thailand
Dengue Bulletin – Volume 31, 2007 77
Figure 2: Development programme of a geo-database for surveillance of Ae. aegypti anddengue haemorrhagic fever in Ubon Ratchathani province
(a)
(b)

78 Dengue Bulletin – Volume 31, 2007
Use of GIS for DHF surveillance in Thailand
• Installation of primary and secondarydata
• Analysis of data and a search of thetarget location
• Presentation of the results via figureson a map
In this study, epidemiological, digital,satellite and Global Positioning System (GPS)data were incorporated into GIS databases tobetter understand the spatial distribution ofDHF cases {see Figures 2(a) and 2(b)}.
Process
Process of GIS development
The project started in February 2007 andconcluded in July 2007. The species of denguemosquito studied was Ae. aegypti. UbonRatchathani province in the lower north-easternpart of Thailand was selected as the study sitedue to its high incidence of DHF cases. Thestudy focused on two sub-districts in each ofthe five districts in the province with the highestincidences of DHF cases in the last three years.Within each sub-district, one village wasselected as part of the study, making a total of10 villages.
Two categories of data were collected:
Primary data included:
• dengue vector indices
• water storage containers
To quantify the relative density ofmosquitoes, House Index (HI), ContainerIndex (CI), and Breteau Index (BI) were takenas indicators of the density. For CI, all naturaland artificial indoor and outdoor containers,such as water jars, tyres, cement tanks and antguard jars, etc. in every house, were inspectedto determine the presence or absence of Ae.aegypti. The primary data were collected by
the visual larval survey.[7] The positions of thehouses in the ten villages were mapped usinga GPS tool. Also, data including village names,house addresses, demographic primary data,breeding sites and containers were mappedand imported into a GIS software for furtherconstruction (ArcView 3.2a®).
Secondary data included:
• number of reported DHF cases/100 000 population
Secondary data on patient cases werereported by a primary care unit (PCU), localhospital or central hospital and collected bythe staff of the Office of Disease Preventionand Control Department 7 (DPC), UbonRatchathani, using WHO case definition. Bothprimary and secondary data were collected bythe same process. Data collections weredivided into low disease incidence (February–March 2007; dry weather) and high diseaseincidence (June–July 2007; rainy season).
Method of data analysis
Analysis of the primary data
Dengue vector indexes were calculated intothree variables:
• House Index (HI): Percent housespositive for containers with Ae. aegyptilarvae or pupae
• Container Index (CI): Percent of water-holding containers positive for Ae.aegypti larvae or pupae
• Breteau Index: Number of containerspositive for Ae. aegypti larvae or pupaeper 100 houses.
Analysis of secondary data
The total number of DHF patients waspresented as the number of reported cases ofDHF/100 000 population of the province.
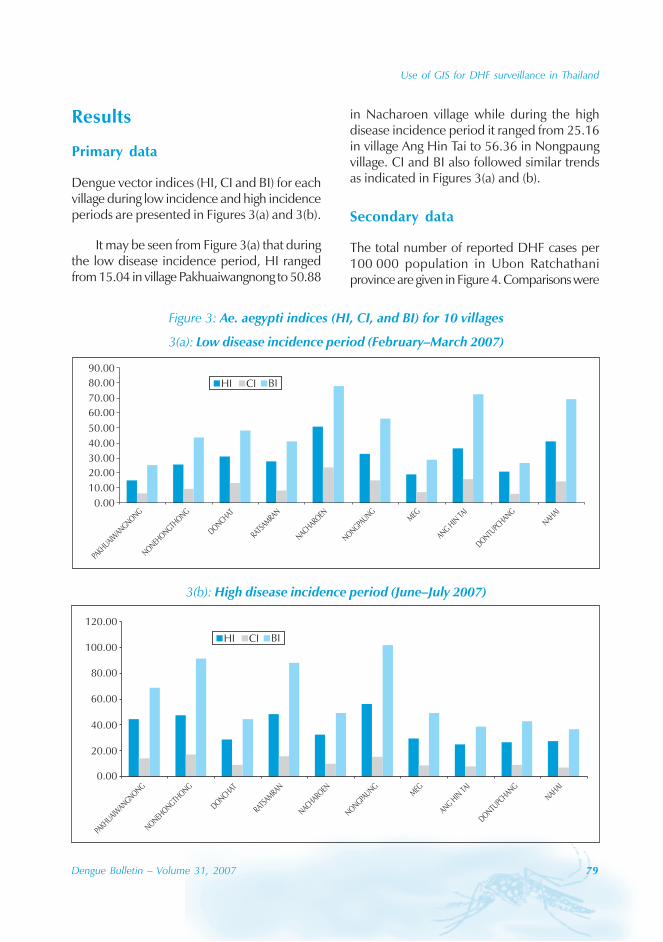
Use of GIS for DHF surveillance in Thailand
Dengue Bulletin – Volume 31, 2007 79
Results
Primary data
Dengue vector indices (HI, CI and BI) for eachvillage during low incidence and high incidenceperiods are presented in Figures 3(a) and 3(b).
It may be seen from Figure 3(a) that duringthe low disease incidence period, HI rangedfrom 15.04 in village Pakhuaiwangnong to 50.88
in Nacharoen village while during the highdisease incidence period it ranged from 25.16in village Ang Hin Tai to 56.36 in Nongpaungvillage. CI and BI also followed similar trendsas indicated in Figures 3(a) and (b).
Secondary data
The total number of reported DHF cases per100 000 population in Ubon Ratchathaniprovince are given in Figure 4. Comparisons were
Figure 3: Ae. aegypti indices (HI, CI, and BI) for 10 villages
3(a): Low disease incidence period (February–March 2007)
0.00
10.00
20.00
30.00
40.00
50.00
60.00
70.00
80.00
90.00
HI CI BI
NAHAI
DONTUPC
HANG
ANGHIN
TAI
MEG
NONGPAUNG
NACHAROEN
RATSA
MRAN
DONCHAT
NONEHONGTH
ONG
PAKH
UAIWANGNONG
3(b): High disease incidence period (June–July 2007)
0.00
20.00
40.00
60.00
80.00
100.00
120.00
NAHAI
DONTUPC
HANG
ANGHIN
TAI
MEG
NONGPAUNG
NACHAROEN
RATSA
MRAN
DONCHAT
NONEHONGTH
ONG
PAKH
UAIWANGNONG
HI CI BI

80 Dengue Bulletin – Volume 31, 2007
Use of GIS for DHF surveillance in Thailand
made between the low disease incidence periodand high disease incidence period (Figure 4).
Discussion
The results show that the number of DHF casesincreased during the high disease incidenceperiod compared to the low disease incidenceperiod, suggesting a positive correlationbetween the peak rainfall period in June-Julyand the high density of Ae. aegypti mosquitoesand high incidence of DHF cases. This findingis similar to that identified in other South-EastAsian countries.[20,21] Since the spread of thedisease is facilitated by the movement ofviraemic people, source-reduction methodsshould get priority in vulnerable areas (hotels,hospitals and other places of congregations).Generally, such practices undertaken in the lowdisease incidence period lead to a reduction
in the vector receptivity (breeding potential)at the beginning of the wet season and thiswill have an impact on the number of DHFcases.
In Ubon Ratchathani, Ae. aegyptipopulation density remains high in most areasand the incidence of DHF normally tends toincrease every other year. This indicates thatwhen an outbreak occurs, it spreads rapidly tonearby areas. The clustering of DHF cases insome big centres such as Muang, Detudon andWarinchamrap has also been attributed to thecharacteristics of Ae. aegypti because of itsmultiple feeding probe to complete one bloodmeal. In other south-east Asian countries, ithas been reported that most infections occuramong children who spend most of their timeindoors and are bitten by the usually day-bitingAe. aegypti. The high incidence of DHFcontinues to occur due to lack of standard
Low disease incidence period High disease incidence period
Number of cases
Figure 4: Total number of reported dengue haemorrhagic fever cases in Ubon Ratchathaniprovince during low and high disease incidence periods

Use of GIS for DHF surveillance in Thailand
Dengue Bulletin – Volume 31, 2007 81
mosquito control policies, poor sanitation andpoor preventive measures.
The introduction and implementation of aneffective remote-sensing tool such as ageographical information system (GIS) into theroutine activities of the Disease Prevention andControl Programme staff needs to beconsidered. This tool can address somelimitations regarding Ae. aegypti and DHFsurveillance like updating, collecting, editing,locating and analysing dengue indices and DHFcase data. By this process, rapid and effectiveprevention and control strategies can bedeveloped prior to the onset of the disease thatwould eventually reduce both the morbidity andmortality rates of dengue infection.
Acknowledgements
The authors express their sincere thanks to theThai Research Funds (TRF) and the World HealthOrganization (WHO), Thailand, for their financialsupport. They also thank Mr ChawalitTantinimitkul of WHO for his kindness. All datasupport was kindly provided by the Office ofDisease Prevention and Control Department 7(DPC), Ubon Ratchathani. Thanks also go toBob Tremayne, Division of InternationalCooperation, Ubon Rajathanee University(UBU), for his English language assistance.Finally, our appreciation goes to Mr WacharapongSaengnill, College of Medicine and PublicHealth, UBU, for his technical support.
References
[1] Thara A. Dengue vector in hemorrhagic fever.National library report. National Institute ofHealth, Department of Medical Science,Thailand. 2001. p. 1-36.
[2] Bureau of Epidemiology, Department ofDisease Control, Ministry of Public Health,Thailand. An annual report of diseasesurveillance. Bangkok: Veteran DonatingOrganization Press, 1997. p. 106-07.
[3] Srijakrawanwong A, Popanya S, RojanasuphujS, Warachit P. A diagnostic test confirmeddengue hemorrhagic fever between 1986 and1989. J Med Sci. 1993; 35: 13-23.
[4] Sathimai W, Laithaweewat L, Wonghiranraj P.A diagnostic confirmation of dengue virusamong unknown fever patients at localhospitals; Khon Kaen and Roi-Et provinces.Infectious Disease Journal. 1998; 24(4): 538-45.
[5] Bureau of Epidemiology, Department ofDisease Control, Ministry of Public Health. Anannual report of disease control. Bangkok:Veteran Donating Organization Press, 2002.p. 40-56.
[6] Research strategy in health care. An annualreport from medical science committee.Research Council. Bangkok: ChulalongkornUniversity Press, 2000. p.240-95.
[7] Goh KT. Dengue - A re-emerging infectiousdisease in Singapore. In: Dengue in Singapore,technical monograph series, no 2. Singapore:Institute of Environmental Epidemiology,Ministry of the Environment, 1993. p.33-40.
[8] Clarke KC, McLafferty SL, Tempalski BJ. Onepidemiology and geographic informationsystems: a review and discussion of futuredirections. Emerg Infect Dis. 1996 Apr-Jun;2(2): 85-92.
[9] Gratz NG, Knudsen AB. The rsie and spread ofdengue, dengue haemorrhagic fever and itsvectors: a historical review (up to 1995).Geneva: World Health Organization, 1996.Document WHO CTD/FIL (DEN) 96/07.
[10] Ai-leen GT, Song RJ. The use of GIS ovitrapsmonitoring for dengue control in Singapore,Dengue Bulletin. 2000; 24: 110-16.

82 Dengue Bulletin – Volume 31, 2007
Use of GIS for DHF surveillance in Thailand
[11] World Health Organization. Division ofControl of Tropical Disease - Health Map -Using GIS in public health 2006. Availablefrom: URL: http://www.who.int/ctd/html/hmapph.html
[12] Crovello TJ. MODABUND. The computerizedmosquito data bank at University of NortreDame. Mosq News. 1972; 32(4): 548-54.
[13] Russo RJ, McCain TL. The use of computerizedinformation retrieval in mosquito control.Mosq News. 1979; 39(2): 333-38.
[14] Russo RJ, McCausland S. Strategies ofcomputer use in mosquito control. MosqNews. 1983; 43(3): 311-14.
[15] World Health Organization. Pesticideapplication equipment for vector control. WHOTechnical Report Series No. 720. Geneva:WHO, 1985. p.36-37.
[16] Chandaeng A, Chansaeng J, Benjapong N. Theclassification of mosquito vectors in Thailandvia a computerized multimedia program. JHealth Sci. 1997; 6(3): 545-52.
[17] Sithiprasasna R, Linthicum KJ, Lerdthusnee K,Brewer TG. Use of geographical informationsystem to study the epidemiology of denguehaemorrhagic fever in Thailand. DengueBulletin. 1997; 21: 68-72.
[18] Sithiprasasna R, Linthicum KJ, Liu GJ, JonesJW, Singhasivanon P. Use of GIS-based spatialmodeling approach to characterize the spatialpatterns of malaria mosquito vector breedinghabitats in northwestern Thailand. SoutheastAsian J Trop Med Public Health. 2003 Sep;34(3): 517-28.
[19] Focks DA, Chadee DD. Pupal survey: anepidemiologically significant surveillancemethod for Aedes aegypti: an example usingdata from Trinidad. Am J Trop Med Hyg. 1997Feb; 56(2): 159-67.
[20] World Health Organization. Vector ecology.WHO Technical Report Series No 501.Geneva: WHO, 1972: p.30-31.
[21] Gould DJ, Mount GA, Scanlon JE, Ford HR,Sullivan MF. Ecology and control of denguevectors on an island in the Gulf of Thailand. JMed Entomol. 1970 Aug 25; 7(4): 499-508.
Dengue Bulletin – Volume 31, 2007 83
Transmission thresholds and pupal/demographic surveysin Yogyakarta, Indonesia for developing a dengue controlstrategy based on targeting epidemiologically significant
types of water-holding containers
Dana A. Focksa , Michael J. Bangsb, Cole Churchc, Mohammad Juffried
and Sustriayu Nalime
aInfectious Disease Analysis, P.O. Box 12852, Gainesville, FL, USA
bFreeport Indonesia, Public Health and Malaria Control-Kuala Kencana, P.O. Box 616, Cairns 4870Australia
cOuachita Parish Mosquito Abatement District, Monroe, LA, USA
dDepartment of Pediatrics, Gadjah Mada University, Yogyakarta, Indonesia
eTahija Dengue Project, Jl. Pandega Sakti 159 Kaliurang Street KM 6, 2, Yogyakarta 55283, Indonesia
Abstract
All water-holding containers (ca. 3000) associated with approximately 320 residences in Yogyakarta,Indonesia, were examined for the presence of Aedes aegypti (L.), Aedes albopictus Skuse, and Culexquinquefasciatus Say pupae in four replicate surveys conducted during two dry seasons (1996 and1998) and two wet seasons (1997 and 1999). Less than 6% of these receptacles had pupae. Ae. aegyptipupae collected were ten times more than Ae. albopictus (ca. 1600 vs. 160 respectively); Cx.quinquefasciatus was rarely encountered. From a dengue perspective, the epidemiological significanceof a particular type of container is a function of the number of Ae. aegypti pupae per person –calculated simply as the ratio of total number of Ae. aegypti pupae recovered in that type and thenumber of residents, ca. 2800. Overall, there was an average of 0.57 Ae. aegypti pupae per person.Assuming an overall herd immunity of 33% and an average temperature of 29 °C, we estimate thetransmission threshold in Yogyakarta to be approximately 0.43 Ae. aegypti pupae per person. Byeliminating mosquito production in two common household containers – wells and used tyres thenumber of Ae. aegypti pupae would be reduced from 0.57 to 0.29 per person, below our estimate ofthe transmission threshold. An assessment of the effectiveness of this strategy is currently being conductedin a multi-year study in Yogyakarta using an insect growth regulator (IGR) for mosquito control.
Keywords: Dengue; Epidemiology; Prevention and control; Risk assessment; Targeted source reduction andcontrol; Sustainable; Community-based; Transmission threshold.
E-mail: [email protected]; Fax: +1-352-372-1838

84 Dengue Bulletin – Volume 31, 2007
Targeted source reduction/control strategy for dengue control
Introduction
Dengue in Indonesia
Dengue and dengue haemorrhagic fever (DF/DHF) was first observed in Indonesia in 1968in Surabaya and Jakarta, the two largestmetropolitan cities on the island of Java.[1]
While this initial epidemic involved less than60 cases, the case-fatality rate exceeded 40%.Since then the incidence of DF/DHF hasincreased dramatically in Indonesia and hasspread geographically to all regions of thecountry. The DHF incidence fluctuates monthlyand typically reaches its peak in December andJanuary every year, except in large cities suchas Jakarta, Bandung and Surabaya, where thehighest incidence is reported in April and May.Currently, dengue is the eighth leading causeof hospitalization among Indonesian children.[2]
Similar trends of progressively larger epidemicsinterspersed with quieter, inter-epidemic yearsin the neighbouring countries of Cambodia,Myanmar, Laos, Thailand and Viet Nam reflectwaxing and waning human population “herd”immunity, urbanization, the movement ofpeople, and the influence of weatheranomalies associated with El Niño/SouthernOscillation (ENSO) events. In the face of themost significant ENSO event of the century,1998 witnessed the largest epidemic on recordin Indonesia with 72 133 reported cases and1414 deaths; the case-fatality rate (CFR) in thisepidemic was 2.0%, reflecting several decadesof improved clinical management.[2] In 2001,the total number of cases (DF and DHF) anddeaths reported were almost 20 000 and 180respectively.[3] In early 2004, DF/DHF made adramatic rebound with over 58 000 cases and658 deaths reported in the first four months.
Past dengue control efforts
The World Health Organization’s RegionalOffice for South-East Asia (WHO/SEARO)
conducted an external review of the dengue/dengue haemorrhagic fever prevention andcontrol programme of Indonesia in June2000.[2] The following brief history of theIndonesian dengue prevention and controlprogramme reflects this report.
The Indonesian Ministry of Health hasconsiderably modified its strategies to controldengue over the past three decades. Initially,adult control using perifocal space spraying ofinsecticides with portable and vehicle-mountedthermal fogging and ultra low volume (ULV)machines was the government’s recommendedmethod and response for most areas. Theprotocol specified treatment within a 100-metreradius of reported DHF cases. In the 1980s,the strategy changed to include the addition ofextensive larviciding using temephos (1% sandgranules). The policy was to treat all breedingsites in dengue-endemic urban areas a singletime each year, timed ideally to precede theonset of the transmission season. The Ministryof Health subsequently modified this strategyto target only those urban areas reporting DHFfor three consecutive years, wherein re-treatments were scheduled with a frequencyof three months. This selective larvicidingprogramme was implemented between 1986and 1991. Beginning in 1992 and continuinguntil the present, the national strategic emphasishas been larval control involving communityefforts, health education and intersectoralcoordination. Currently, national efforts havefocused on organizing working groups at thevillage level under the general guidance of localhealth centre personnel. This programme, calledBulan Gerakan (or 3M), emphasizes intensivehealth education using mass media, women’sgroups and schoolchildren, community-basedbreeding source reduction, and door-to-doorhouse inspections to monitor for larvae, and toclean containers and apply temephos asnecessary. An important member of the 3Mprogramme is the Family Welfare EducationWomen’s Movement (Pendidikan Kesejahteraan

Targeted source reduction/control strategy for dengue control
Dengue Bulletin – Volume 31, 2007 85
Keluarga or PKK). The role of the PKK iseducation of the house-owner about larvalinspections, methods to store water safely, andthe elimination or cleaning of breedingcontainers. An additional role of the PKK involvescommunity-based group education andmonitoring programme results. Finally, authoritiesdeveloped health education programmes forelementary schoolchildren and for use with themass media.
Unfortunately, with only a few notableexceptions, these efforts have not beensuccessful in controlling dengue.[2] However, apilot project in the city of Purwokerto, aidedwith funding from Rotary International hasshown promise. The project organized a strategyof community partnership based on dasa wisma(ten houses) in several villages in the area. Theresults were encouraging to the extent thatRotary and others funded an extension to thiseffort that targeted 11 major urban areas inIndonesia. Patterned after the Purwokertoproject, it included educational programmes forthe public and medical personnel. The extendedproject had the endorsement and support ofthe Indonesian government, WHO, the USPublic Health Service, and importantly, mayors’offices. While funding was available and keypublic officials remained prominently associatedwith the project publically, e.g., the mayor’swife, the effort did reduce dengue cases(Sustriayu Nalim, personal communication).
Proposed targeted source reductionand control efforts
The goal of the present work is to build on thefoundation of the 3M and PKK programmes byreducing the number of types of breedingcontainers that must be controlled or eliminatedto only a select few that are responsible for mostof the adult vector production. We believe it ispossible to estimate the degree of reductionrequired and to identify the types of containers
that are particularly important by some recentdevelopments outlined below. This strategytargets only the most epidemiologicallyimportant types of breeding containers. Wemeasure the epidemiological importance ofeach type of container in the environment usingthe statistic the total standing crop of Ae. aegypti(L.) pupae per hectare or per person associatedwith each particular type.
Results from prior research
Transmission models
Recently, there has been a movement in theepidemiological community to recognize thepervasive influence of the environment andclimate on various vector-borne diseases. Theefforts of Martens et al.[4] and Patz et al.,[5] forexample, have documented substantial ties ofdisease activity to environmental features andclimate trends for dengue, schistosomiasis andmalaria. The work of Bouma et al.[6] establishingstatistical relationships between weatheranomalies associated with El Niño and malariain Colombia is especially encouraging in thecontext of developing early warning/mitigationsystems for weather-driven infectious diseases.
Recognizing these ties, mathematicalepidemiologists and public health specialists arebeginning to construct disease modelsincorporating environment and climateparameters. A number of researchers haverecently been involved in these types of studieson the dengue system and have developedsimulation models and estimates of transmissionthresholds.[7,8,9,10,11] These results have had adegree of success and are being evaluated bythe public health community. The algorithms ofthe dengue models[7,9] take into account keyfactors known to influence dengueepidemiology; the result is a software tool usedby researchers and public health practitionersthat is orientated toward site-specific simulation.

86 Dengue Bulletin – Volume 31, 2007
Targeted source reduction/control strategy for dengue control
Validation studies compared model output withfield and laboratory observations at sites in Asiaand the Americas.[8] Funding from WHO andthe US National Institutes of Health (NIH) hasled to ongoing evaluations of transmissionthresholds derived from the models in Viet Namand Peru respectively. The hope is that thepublished thresholds[11] can be used in tropicallocations to predict disease vulnerability, assesscontrol measures and provide guidance intargeting the especially important classes ofbreeding containers. The recent developmentof the pupal/demographic survey, coupled withestimates of transmission thresholds, make theresults of simulation studies with the denguemodels available to operational controlprogrammes in the developing world.[10,11,12] Themodels are currently being re-written andextended through funding from the Tahija FamilyFoundation (Jakarta) and the Innovative VectorControl Consortium (IVCC) funded by the Billand Melinda Gates Foundation (the citation forIVCC/dengue is: Hemingway J, Beaty BJ,Rowland M, Scott TW, Sharp BL. The InnovativeVector Control Consortium: Improved Controlof mosquito-borne diseases. Trends inParasitology 2006 (22): 308-312.)
Transmission thresholds and thepupal/demographic survey
The expense and ineffectiveness of drift-basedinsecticide aerosols to either prevent or controldengue epidemics has led to suppressionstrategies based on eliminating larval breedingsites.[13] With the notable, albeit short-lived,exceptions of Cuba and Singapore, these source-reduction efforts have met with littledocumented success. Public health workersattribute failure to two factors: inadequateparticipation of the communities, and a strategythat entailed destruction or treatment of virtuallyevery breeding container in the environment.The transmission thresholds for dengue basedon the standing crop of Ae. aegypti pupae per
person[11] were developed for use in theassessment of risk of transmission and to providetargets for the actual degree of suppression bytype of breeding container required to preventor eliminate transmission in source-reductionprogrammes. When coupled with fieldobservations from pupal/demographic surveys(as reported herein for Yogyakarta), it is possiblefor the first time for control specialists to knowhow important the various types of containersin the environment are in terms of contributingto the transmission threshold.[10,11] This strategyof concentrating on only the types of containersmost responsible for the majority of adult vectorproduction and hence transmission, e.g. outdoordrums and tyres vs typically low-producing indoorvases and domestic containers, was recentlyevaluated in a 9-country study in the Americasand SE Asia with WHO/TDR funding (citationis: Focks DA, Alexander N. Multicountry studyof Aedes aegypti pupal productivity surveymethodology. Findings and recommendations.2006. TDR/IRM/Den/06.1)). The WHO’sTropical Diseases Research (TDR) programmehas commissioned a review article on the currentstate of the science for entomological surveyingfor dengue risk assessment and control.[14]
Central in this document are the concepts ofthe pupal/demographic survey, transmissionthresholds and targeted source reduction andcontrol of especially productive containers.There is a growing recognition that adherenceto the current strategy of attempting to eliminateor control all containers, irrespective ofproductivity or time of the year, is doomed tocontinued failure.[10]
Dengue early warning system (EWS)
In the last several years, there has been a callby the directors of national anti-dengueprogrammes in Indonesia, Thailand and VietNam for the operational need of an early warningsystem (EWS) that would provide sufficient leadtime (1 to 3 months) to permit mobilization of

Targeted source reduction/control strategy for dengue control
Dengue Bulletin – Volume 31, 2007 87
control operations. In response to this call,preliminary EWSs for Yogyakarta and Bangkokwere developed; they are based on logisticregression analysis.[15] The predictor variables aresea surface temperature (SST) anomalies overthe tropical Pacific and monthly cases of denguein each city. The predicted variable is theprobability of an epidemic year forecast 1 to 3months before the peak transmission season.The Java EWS was sufficiently accurate to beoperational. The Yogyakarta EWS gave perfect1- and 2-month forecasts; the 3-month forecastincorrectly classified one year in the 14-yearperiod of record.
Biological control agents
A very promising recent measure is theapplication of biological agents like Mesocyclops,Micronecta and larvivorous fish. In Viet Nam,Mesocyclops, a tiny copepod crustacean, havebeen found to be good predators of Ae. aegyptilarvae. Since February 1993, Mesocyclops havebeen released into water containers of 400houses of a hamlet in My Van district, Hai Hungprovince. In March 1994, this measure wassupplemented by a campaign to eliminatediscarded containers (which are too small forMesocyclops to survive), and education abouthow to maintain adequate populations ofMesocyclops in domestic water containers. After17 months, Ae. aegypti had been completelyeliminated and this result has been sustaineduntil today. Since July 1995, mobilization of thecommunity for peridomestic hygiene and useof Mesocyclops to control Aedes larvae wasimplemented for 1600 houses in one communein Thuong Tin district, Ha Tay province. Workingtogether with the network of healthcollaborators, primary-school pupils, localauthorities and health staff, and using the systemof local communication, health educationcampaigns were organized in order to improvepeople’s knowledge and to mobilize everymember of the community. Almost all big water
containers in the commune receivedMesocyclops or larvivorous fish; it is now verydifficult to find discarded containers, andmosquito indices have been reduced almost tozero. From 1996, this model has been extendedto three provinces, Nam Ha, Hai Hung and HaiPhong in northern Viet Nam.[16]
Goals of the present work
The purpose of this report is to provide ananalysis of four annual pupal/demographicsurveys conducted in Yogyakarta between 1996-1999 highlighting the epidemiologicalsignificance of the twenty-some types of Ae.aegypti-breeding containers in the environment.From this analysis, and using the estimates oftransmission thresholds for dengue, we proposeto develop a targeted source-reduction/controlstrategy for Yogyakarta that will requiresubstantially less effort than the traditionalcommunity-based efforts without targetingwhere the goal is to control or eliminate allcontainers irrespective of their contribution tothe adult population of Ae. aegypti.
Methods
Study site
Yogyakarta, a city of over 520 000 people, isthe provincial capital of Yogyakarta, located incentral Java. The province is divided intoadministrative districts called kabupatens witheach district divided into progressively smallerunits beginning with sub-districts calledkecamatans, and these, in turn, divided intokelurahans, and further divided into rukun warga(RW), and finally into rukun tetangga (RT), thesmallest administrative unit composed ofapproximately 50-80 families each. The studysite was located in kecamatan Gondokusuman,within Yogyakarta city. The actual study areacovered approximately 6.34 ha, from which

88 Dengue Bulletin – Volume 31, 2007
Targeted source reduction/control strategy for dengue control
323 houses were selected and subsequentlysampled over four time periods. Disease anddemographic data were derived from a recentstudy conducted in the same area.[17] Anaverage of five houses were chosen from eachRW where a dengue case had occurred andincluded 64 RWs distributed among fivekelurahans (Kota Baru = 4 RWs; Terban = 11RWs; Baciro = 21 RWs; Klitren = 16 RWs,and Demangan = 12 RWs). The location ofeach sampled house was provided coordinatesin 1999 using a hand-held geographicalpositioning device (Magellan GPS 300tm, SanDimas, CA). Demography data were takenfrom each house, including number ofpermanent residents, house size and land area.Climatological data (daily rainfall and maximum/minimum ambient temperatures) wereobtained from the local Meteorology andGeophysics Agency (Station Bulaksumar,University of Gadjah Mada).
The pupal/demographic survey
Each premise was sampled consecutively forimmature stages of container-breedingmosquitoes during four different time periodsbetween 1996 and 1999, with two samplesurveys during wet seasons and two during thedry periods of the year.# A team of three orfour people would visit each house and carefullyinspect the inside and around the outsideperimeter all natural and artificial containers forpreimaginal stages of mosquitoes. The numberand type of containers present at each housewere recorded as well as the number ofpermanent residents. The presence or absenceof mosquito larvae in each container wasrecorded without regard to the number present.Collections concentrated on quantifying pupal
abundance by type of container. With the aidof a flashlight, hand-held fine-meshed nettingdevices and pipettes were used to remove allpupae from the container with capturedspecimens placed in white trays for easyobservation. While in the field, all pupae wouldbe first immobilized using hot water suppliedfrom a thermos and immediately placed inlabelled plastic bags containing 70% ethyl alcoholand sealed. Bag labels included house number,container type, location (indoor or outdoor) andnumber of pupae collected. Additionally, allinformation, including lot and house size, washand-recorded in a field logbook and latertranscribed into a computerized database. Theonly water-holding containers not surveyed inthis study were residential wells, which areknown to harbour Ae. aegypti and Culexquinquefasciatus Say larvae.[18]
Specimen identification
Preserved pupae were returned to thelaboratory for identification. Using an illustratedkey developed for this purpose, pupae wereexamined using a stereomicroscope and easilyidentified for species and sex.[19] For thepurposes of this study, pupae were identifiedas either Ae. aegypti, Aedes albopictus Skuse,or Cx. quinquefasciatus, with all other pupaeidentified only to genera.
Results
With very few exceptions, the four pupal/demographic surveys conducted in thekecamatan Gondokusuman returned to thesame ca. 316 houses each year. The residentsassociated with these houses numberedapproximately 2800 (Table 1). Each exhaustivesurvey collected pupae from the ca. 3000water-filled containers associated with the studyhouses. Approximately 5.5% of the containerswere positive for one or more pupae of Ae.
#Collection dates: Dry season: 8–22 May 1996,Wet: 23 January–7 February 1997,Dry: 15–30 September 1998, andWet: 30 March–19 April 1999.
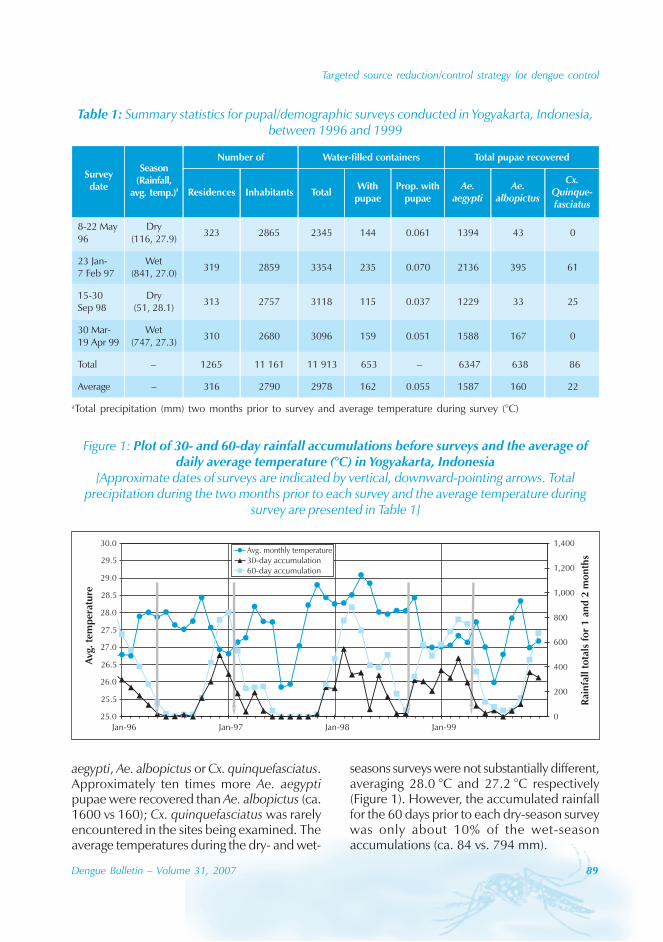
Targeted source reduction/control strategy for dengue control
Dengue Bulletin – Volume 31, 2007 89
aegypti, Ae. albopictus or Cx. quinquefasciatus.Approximately ten times more Ae. aegyptipupae were recovered than Ae. albopictus (ca.1600 vs 160); Cx. quinquefasciatus was rarelyencountered in the sites being examined. Theaverage temperatures during the dry- and wet-
Table 1: Summary statistics for pupal/demographic surveys conducted in Yogyakarta, Indonesia,between 1996 and 1999
aTotal precipitation (mm) two months prior to survey and average temperature during survey (°C)
Figure 1: Plot of 30- and 60-day rainfall accumulations before surveys and the average ofdaily average temperature (°C) in Yogyakarta, Indonesia
[Approximate dates of surveys are indicated by vertical, downward-pointing arrows. Totalprecipitation during the two months prior to each survey and the average temperature during
survey are presented in Table 1]
25.0
25.5
26.0
26.5
27.0
27.5
28.0
28.5
29.0
29.5
30.0
Jan-96 Jan-97 Jan-98 Jan-990
200
400
600
800
1,000
1,200
1,400Avg. monthly temperature30-day accumulation60-day accumulation
Rai
nfa
llto
tals
for
1an
d2
mo
nth
s
Avg
.te
mp
erat
ure
seasons surveys were not substantially different,averaging 28.0 °C and 27.2 °C respectively(Figure 1). However, the accumulated rainfallfor the 60 days prior to each dry-season surveywas only about 10% of the wet-seasonaccumulations (ca. 84 vs. 794 mm).
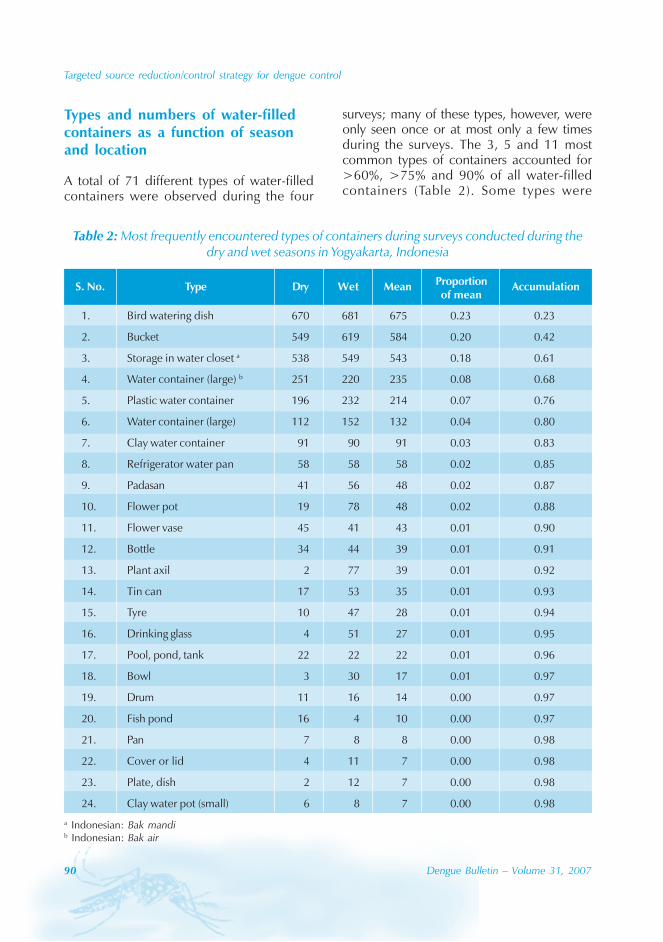
90 Dengue Bulletin – Volume 31, 2007
Targeted source reduction/control strategy for dengue control
Types and numbers of water-filledcontainers as a function of seasonand location
A total of 71 different types of water-filledcontainers were observed during the four
surveys; many of these types, however, wereonly seen once or at most only a few timesduring the surveys. The 3, 5 and 11 mostcommon types of containers accounted for>60%, >75% and 90% of all water-filledcontainers (Table 2). Some types were
Table 2: Most frequently encountered types of containers during surveys conducted during thedry and wet seasons in Yogyakarta, Indonesia
a Indonesian: Bak mandib Indonesian: Bak air
S. No. Type Dry Wet Mean Proportion Accumulationof mean
1. Bird watering dish 670 681 675 0.23 0.23
2. Bucket 549 619 584 0.20 0.42
3. Storage in water closet a 538 549 543 0.18 0.61
4. Water container (large) b 251 220 235 0.08 0.68
5. Plastic water container 196 232 214 0.07 0.76
6. Water container (large) 112 152 132 0.04 0.80
7. Clay water container 91 90 91 0.03 0.83
8. Refrigerator water pan 58 58 58 0.02 0.85
9. Padasan 41 56 48 0.02 0.87
10. Flower pot 19 78 48 0.02 0.88
11. Flower vase 45 41 43 0.01 0.90
12. Bottle 34 44 39 0.01 0.91
13. Plant axil 2 77 39 0.01 0.92
14. Tin can 17 53 35 0.01 0.93
15. Tyre 10 47 28 0.01 0.94
16. Drinking glass 4 51 27 0.01 0.95
17. Pool, pond, tank 22 22 22 0.01 0.96
18. Bowl 3 30 17 0.01 0.97
19. Drum 11 16 14 0.00 0.97
20. Fish pond 16 4 10 0.00 0.97
21. Pan 7 8 8 0.00 0.98
22. Cover or lid 4 11 7 0.00 0.98
23. Plate, dish 2 12 7 0.00 0.98
24. Clay water pot (small) 6 8 7 0.00 0.98
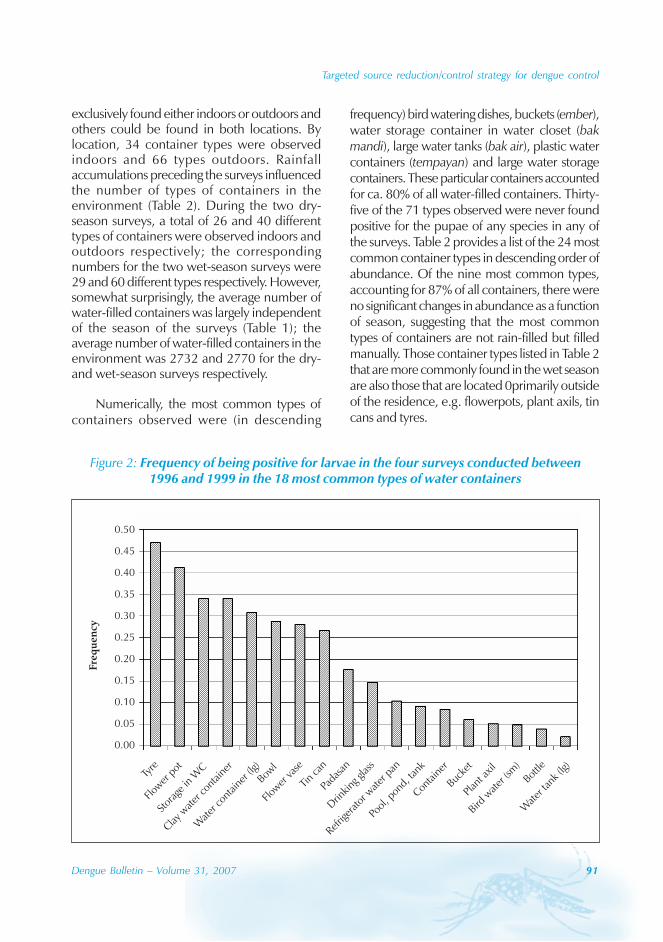
Targeted source reduction/control strategy for dengue control
Dengue Bulletin – Volume 31, 2007 91
exclusively found either indoors or outdoors andothers could be found in both locations. Bylocation, 34 container types were observedindoors and 66 types outdoors. Rainfallaccumulations preceding the surveys influencedthe number of types of containers in theenvironment (Table 2). During the two dry-season surveys, a total of 26 and 40 differenttypes of containers were observed indoors andoutdoors respectively; the correspondingnumbers for the two wet-season surveys were29 and 60 different types respectively. However,somewhat surprisingly, the average number ofwater-filled containers was largely independentof the season of the surveys (Table 1); theaverage number of water-filled containers in theenvironment was 2732 and 2770 for the dry-and wet-season surveys respectively.
Numerically, the most common types ofcontainers observed were (in descending
frequency) bird watering dishes, buckets (ember),water storage container in water closet (bakmandi), large water tanks (bak air), plastic watercontainers (tempayan) and large water storagecontainers. These particular containers accountedfor ca. 80% of all water-filled containers. Thirty-five of the 71 types observed were never foundpositive for the pupae of any species in any ofthe surveys. Table 2 provides a list of the 24 mostcommon container types in descending order ofabundance. Of the nine most common types,accounting for 87% of all containers, there wereno significant changes in abundance as a functionof season, suggesting that the most commontypes of containers are not rain-filled but filledmanually. Those container types listed in Table 2that are more commonly found in the wet seasonare also those that are located 0primarily outsideof the residence, e.g. flowerpots, plant axils, tincans and tyres.
Figure 2: Frequency of being positive for larvae in the four surveys conducted between1996 and 1999 in the 18 most common types of water containers
0.50
0.45
0.40
0.35
0.30
0.25
0.20
0.15
0.10
0.05
0.00
Fre
qu
en
cy
Tyre
Flow
erpo
t
Stor
age
inW
C
Claywater
cont
aine
r
Water
cont
aine
r (lg)
Bowl
Flow
erva
se
Tin
can
Pada
san
Drin
king
glass
Refrig
erator
water
pan
Pool
, pon
d,tank
Contain
er
Bucke
t
Plan
t axil
Birdwater
(sm)
Bottle
Water
tank
(lg)
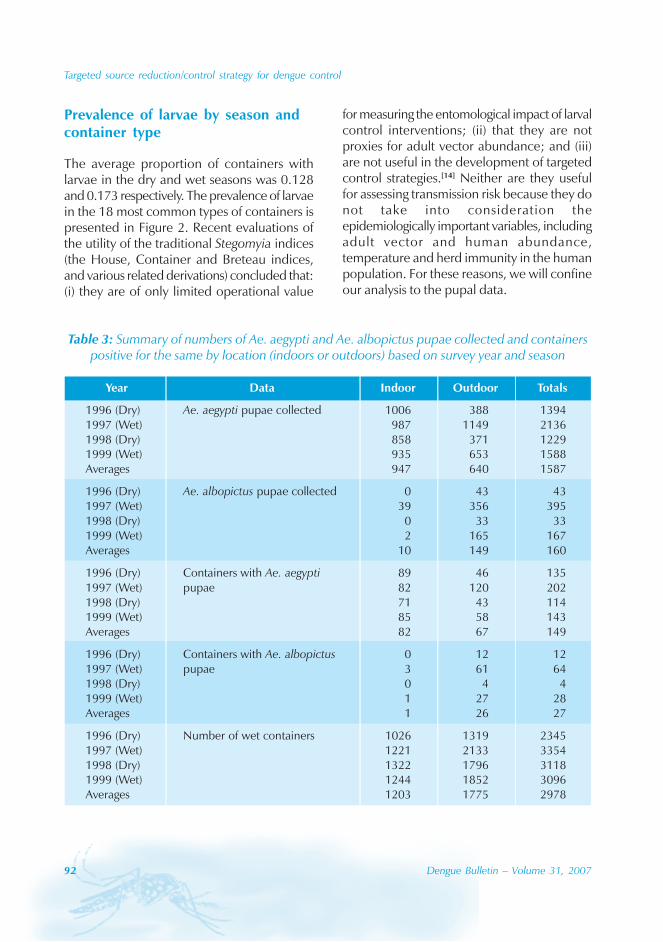
92 Dengue Bulletin – Volume 31, 2007
Targeted source reduction/control strategy for dengue control
Prevalence of larvae by season andcontainer type
The average proportion of containers withlarvae in the dry and wet seasons was 0.128and 0.173 respectively. The prevalence of larvaein the 18 most common types of containers ispresented in Figure 2. Recent evaluations ofthe utility of the traditional Stegomyia indices(the House, Container and Breteau indices,and various related derivations) concluded that:(i) they are of only limited operational value
for measuring the entomological impact of larvalcontrol interventions; (ii) that they are notproxies for adult vector abundance; and (iii)are not useful in the development of targetedcontrol strategies.[14] Neither are they usefulfor assessing transmission risk because they donot take into consideration theepidemiologically important variables, includingadult vector and human abundance,temperature and herd immunity in the humanpopulation. For these reasons, we will confineour analysis to the pupal data.
Table 3: Summary of numbers of Ae. aegypti and Ae. albopictus pupae collected and containerspositive for the same by location (indoors or outdoors) based on survey year and season
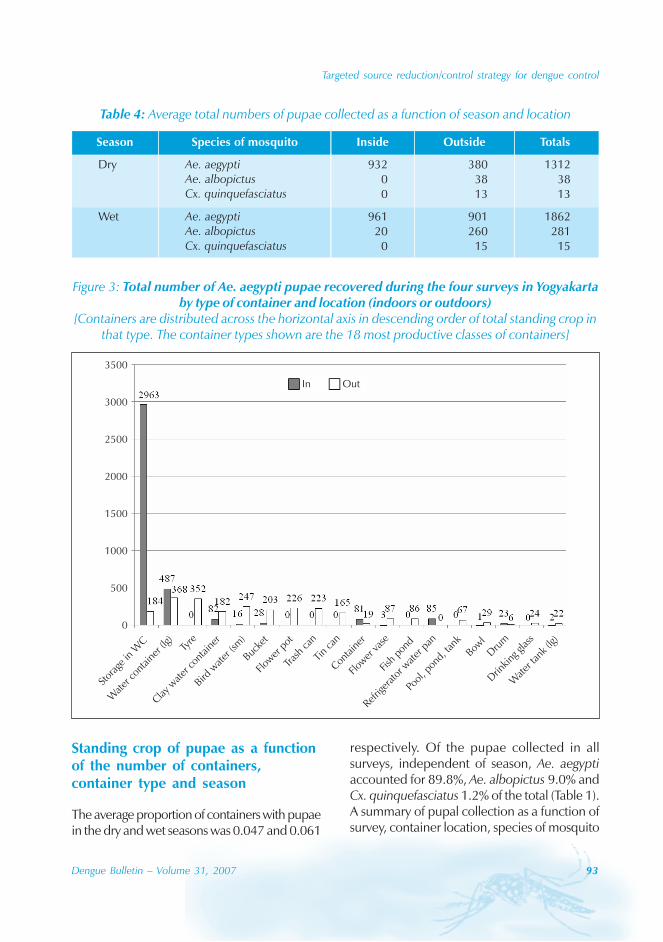
Targeted source reduction/control strategy for dengue control
Dengue Bulletin – Volume 31, 2007 93
Table 4: Average total numbers of pupae collected as a function of season and location
Figure 3: Total number of Ae. aegypti pupae recovered during the four surveys in Yogyakartaby type of container and location (indoors or outdoors)
[Containers are distributed across the horizontal axis in descending order of total standing crop inthat type. The container types shown are the 18 most productive classes of containers]
Tyre
Flow
erpo
t
Stor
age
inW
C
Claywater
cont
aine
r
Water
cont
aine
r (lg)
Bowl
Flow
erva
se
Tin
can
Tras
hca
n
Drin
king
glass
Refrig
erator
water
pan
Pool
, pon
d,tank
Contain
er
Bucke
t
Fish
pond
Birdwater
(sm)
Dru
m
Water
tank
(lg)
3500
3000
2500
2000
1500
1000
500
0
In Out
Standing crop of pupae as a functionof the number of containers,container type and season
The average proportion of containers with pupaein the dry and wet seasons was 0.047 and 0.061
respectively. Of the pupae collected in allsurveys, independent of season, Ae. aegyptiaccounted for 89.8%, Ae. albopictus 9.0% andCx. quinquefasciatus 1.2% of the total (Table 1).A summary of pupal collection as a function ofsurvey, container location, species of mosquito
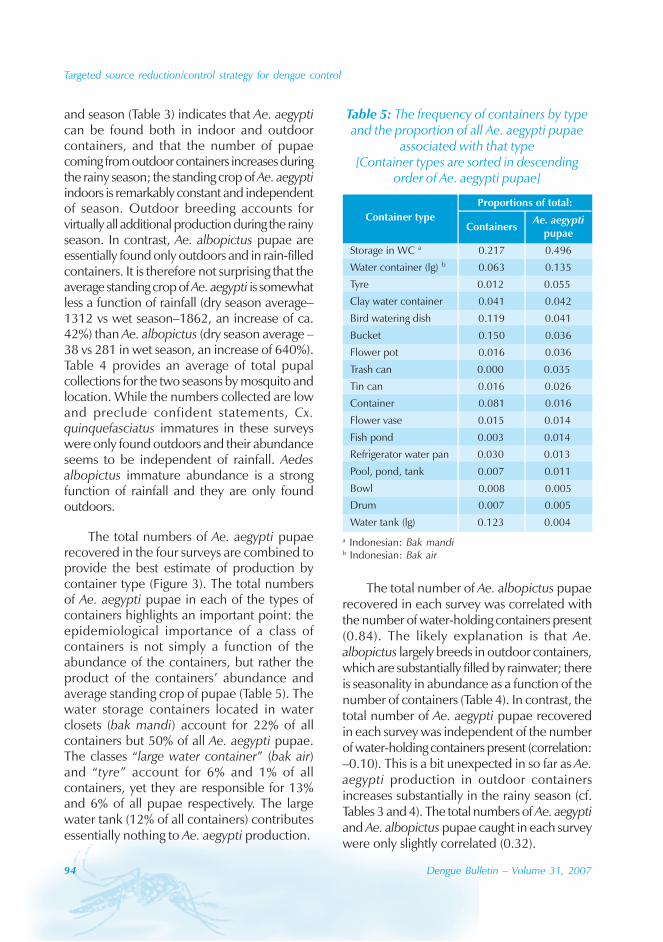
94 Dengue Bulletin – Volume 31, 2007
Targeted source reduction/control strategy for dengue control
and season (Table 3) indicates that Ae. aegyptican be found both in indoor and outdoorcontainers, and that the number of pupaecoming from outdoor containers increases duringthe rainy season; the standing crop of Ae. aegyptiindoors is remarkably constant and independentof season. Outdoor breeding accounts forvirtually all additional production during the rainyseason. In contrast, Ae. albopictus pupae areessentially found only outdoors and in rain-filledcontainers. It is therefore not surprising that theaverage standing crop of Ae. aegypti is somewhatless a function of rainfall (dry season average–1312 vs wet season–1862, an increase of ca.42%) than Ae. albopictus (dry season average –38 vs 281 in wet season, an increase of 640%).Table 4 provides an average of total pupalcollections for the two seasons by mosquito andlocation. While the numbers collected are lowand preclude confident statements, Cx.quinquefasciatus immatures in these surveyswere only found outdoors and their abundanceseems to be independent of rainfall. Aedesalbopictus immature abundance is a strongfunction of rainfall and they are only foundoutdoors.
The total numbers of Ae. aegypti pupaerecovered in the four surveys are combined toprovide the best estimate of production bycontainer type (Figure 3). The total numbersof Ae. aegypti pupae in each of the types ofcontainers highlights an important point: theepidemiological importance of a class ofcontainers is not simply a function of theabundance of the containers, but rather theproduct of the containers’ abundance andaverage standing crop of pupae (Table 5). Thewater storage containers located in waterclosets (bak mandi) account for 22% of allcontainers but 50% of all Ae. aegypti pupae.The classes “large water container” (bak air)and “tyre” account for 6% and 1% of allcontainers, yet they are responsible for 13%and 6% of all pupae respectively. The largewater tank (12% of all containers) contributesessentially nothing to Ae. aegypti production.
The total number of Ae. albopictus pupaerecovered in each survey was correlated withthe number of water-holding containers present(0.84). The likely explanation is that Ae.albopictus largely breeds in outdoor containers,which are substantially filled by rainwater; thereis seasonality in abundance as a function of thenumber of containers (Table 4). In contrast, thetotal number of Ae. aegypti pupae recoveredin each survey was independent of the numberof water-holding containers present (correlation:–0.10). This is a bit unexpected in so far as Ae.aegypti production in outdoor containersincreases substantially in the rainy season (cf.Tables 3 and 4). The total numbers of Ae. aegyptiand Ae. albopictus pupae caught in each surveywere only slightly correlated (0.32).
Table 5: The frequency of containers by typeand the proportion of all Ae. aegypti pupae
associated with that type[Container types are sorted in descending
order of Ae. aegypti pupae]
a Indonesian: Bak mandib Indonesian: Bak air
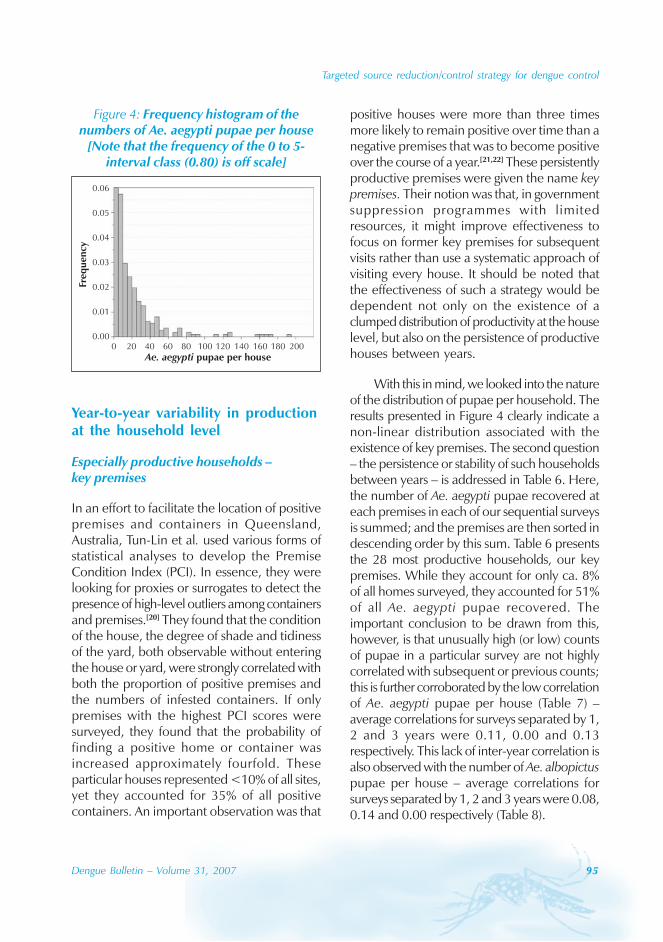
Targeted source reduction/control strategy for dengue control
Dengue Bulletin – Volume 31, 2007 95
positive houses were more than three timesmore likely to remain positive over time than anegative premises that was to become positiveover the course of a year.[21,22] These persistentlyproductive premises were given the name keypremises. Their notion was that, in governmentsuppression programmes with limitedresources, it might improve effectiveness tofocus on former key premises for subsequentvisits rather than use a systematic approach ofvisiting every house. It should be noted thatthe effectiveness of such a strategy would bedependent not only on the existence of aclumped distribution of productivity at the houselevel, but also on the persistence of productivehouses between years.
With this in mind, we looked into the natureof the distribution of pupae per household. Theresults presented in Figure 4 clearly indicate anon-linear distribution associated with theexistence of key premises. The second question– the persistence or stability of such householdsbetween years – is addressed in Table 6. Here,the number of Ae. aegypti pupae recovered ateach premises in each of our sequential surveysis summed; and the premises are then sorted indescending order by this sum. Table 6 presentsthe 28 most productive households, our keypremises. While they account for only ca. 8%of all homes surveyed, they accounted for 51%of all Ae. aegypti pupae recovered. Theimportant conclusion to be drawn from this,however, is that unusually high (or low) countsof pupae in a particular survey are not highlycorrelated with subsequent or previous counts;this is further corroborated by the low correlationof Ae. aegypti pupae per house (Table 7) –average correlations for surveys separated by 1,2 and 3 years were 0.11, 0.00 and 0.13respectively. This lack of inter-year correlation isalso observed with the number of Ae. albopictuspupae per house – average correlations forsurveys separated by 1, 2 and 3 years were 0.08,0.14 and 0.00 respectively (Table 8).
Figure 4: Frequency histogram of thenumbers of Ae. aegypti pupae per house
[Note that the frequency of the 0 to 5-interval class (0.80) is off scale]
0.06
0.05
0.04
0.03
0.02
0.01
0.00
Freq
uen
cy
Ae. aegypti pupae per house0 20 40 60 80 100 120 140 160 180 200
Year-to-year variability in productionat the household level
Especially productive households –key premises
In an effort to facilitate the location of positivepremises and containers in Queensland,Australia, Tun-Lin et al. used various forms ofstatistical analyses to develop the PremiseCondition Index (PCI). In essence, they werelooking for proxies or surrogates to detect thepresence of high-level outliers among containersand premises.[20] They found that the conditionof the house, the degree of shade and tidinessof the yard, both observable without enteringthe house or yard, were strongly correlated withboth the proportion of positive premises andthe numbers of infested containers. If onlypremises with the highest PCI scores weresurveyed, they found that the probability offinding a positive home or container wasincreased approximately fourfold. Theseparticular houses represented <10% of all sites,yet they accounted for 35% of all positivecontainers. An important observation was that
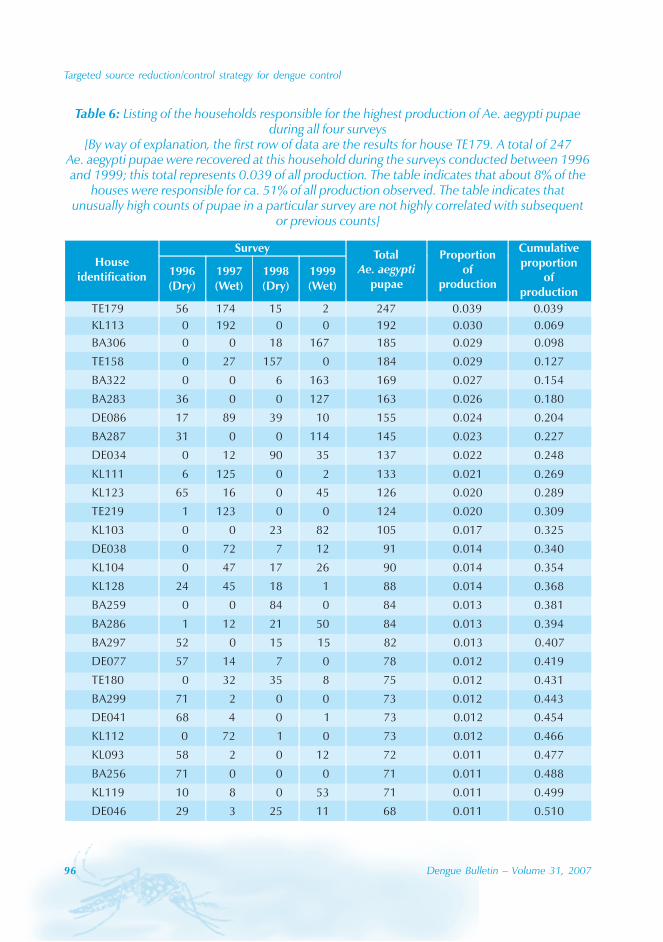
96 Dengue Bulletin – Volume 31, 2007
Targeted source reduction/control strategy for dengue control
Table 6: Listing of the households responsible for the highest production of Ae. aegypti pupaeduring all four surveys
[By way of explanation, the first row of data are the results for house TE179. A total of 247Ae. aegypti pupae were recovered at this household during the surveys conducted between 1996and 1999; this total represents 0.039 of all production. The table indicates that about 8% of the
houses were responsible for ca. 51% of all production observed. The table indicates thatunusually high counts of pupae in a particular survey are not highly correlated with subsequent
or previous counts]
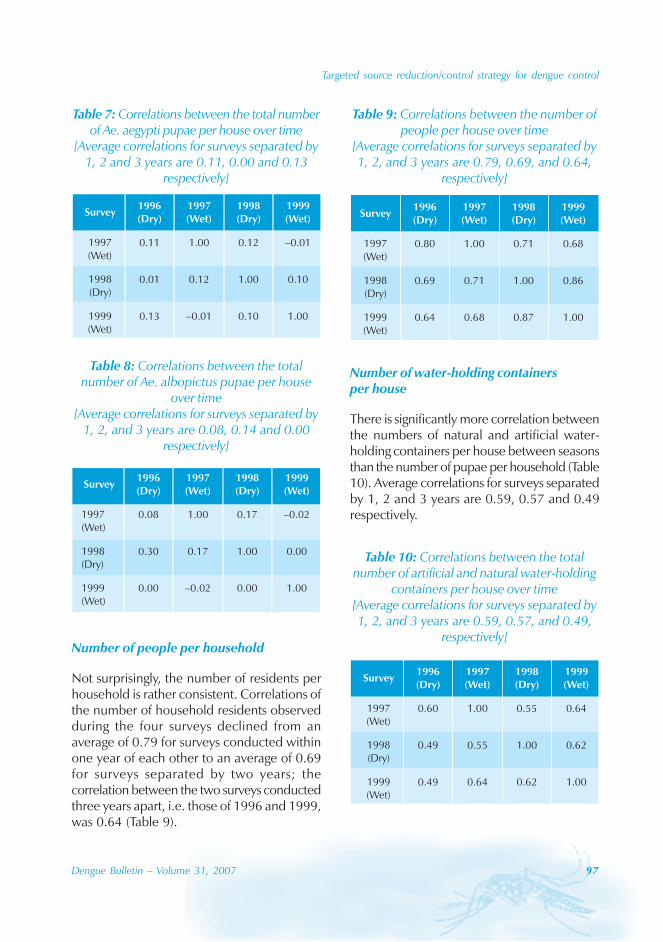
Targeted source reduction/control strategy for dengue control
Dengue Bulletin – Volume 31, 2007 97
Number of people per household
Not surprisingly, the number of residents perhousehold is rather consistent. Correlations ofthe number of household residents observedduring the four surveys declined from anaverage of 0.79 for surveys conducted withinone year of each other to an average of 0.69for surveys separated by two years; thecorrelation between the two surveys conductedthree years apart, i.e. those of 1996 and 1999,was 0.64 (Table 9).
Number of water-holding containersper house
There is significantly more correlation betweenthe numbers of natural and artificial water-holding containers per house between seasonsthan the number of pupae per household (Table10). Average correlations for surveys separatedby 1, 2 and 3 years are 0.59, 0.57 and 0.49respectively.
Table 7: Correlations between the total numberof Ae. aegypti pupae per house over time
[Average correlations for surveys separated by1, 2 and 3 years are 0.11, 0.00 and 0.13
respectively]
Table 8: Correlations between the totalnumber of Ae. albopictus pupae per house
over time[Average correlations for surveys separated by
1, 2, and 3 years are 0.08, 0.14 and 0.00respectively]
Table 9: Correlations between the number ofpeople per house over time
[Average correlations for surveys separated by1, 2, and 3 years are 0.79, 0.69, and 0.64,
respectively]
Table 10: Correlations between the totalnumber of artificial and natural water-holding
containers per house over time[Average correlations for surveys separated by1, 2, and 3 years are 0.59, 0.57, and 0.49,
respectively]
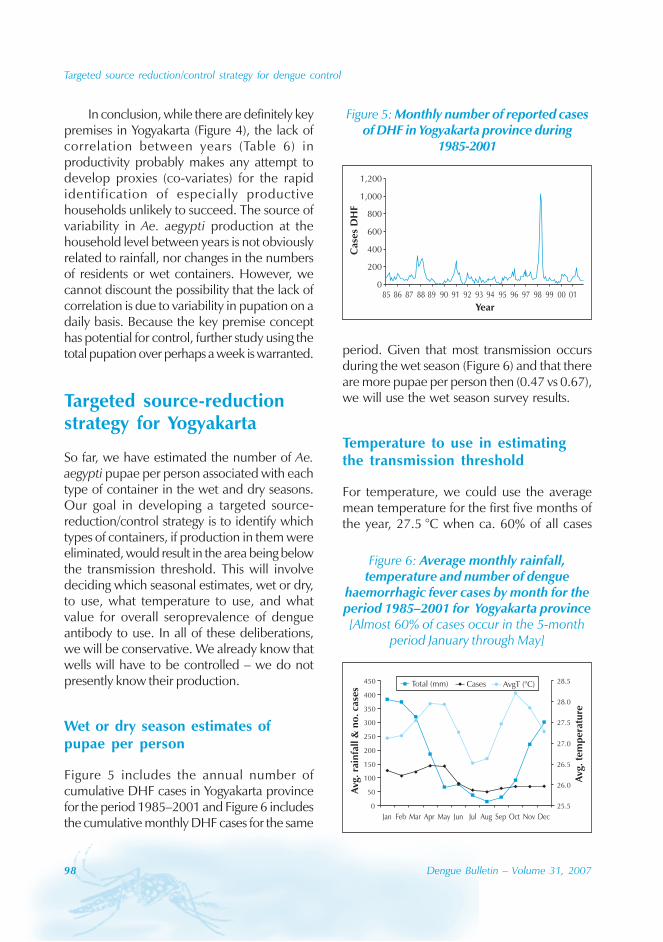
98 Dengue Bulletin – Volume 31, 2007
Targeted source reduction/control strategy for dengue control
Figure 6: Average monthly rainfall,temperature and number of dengue
haemorrhagic fever cases by month for theperiod 1985–2001 for Yogyakarta province[Almost 60% of cases occur in the 5-month
period January through May]
In conclusion, while there are definitely keypremises in Yogyakarta (Figure 4), the lack ofcorrelation between years (Table 6) inproductivity probably makes any attempt todevelop proxies (co-variates) for the rapididentification of especially productivehouseholds unlikely to succeed. The source ofvariability in Ae. aegypti production at thehousehold level between years is not obviouslyrelated to rainfall, nor changes in the numbersof residents or wet containers. However, wecannot discount the possibility that the lack ofcorrelation is due to variability in pupation on adaily basis. Because the key premise concepthas potential for control, further study using thetotal pupation over perhaps a week is warranted.
Targeted source-reductionstrategy for Yogyakarta
So far, we have estimated the number of Ae.aegypti pupae per person associated with eachtype of container in the wet and dry seasons.Our goal in developing a targeted source-reduction/control strategy is to identify whichtypes of containers, if production in them wereeliminated, would result in the area being belowthe transmission threshold. This will involvedeciding which seasonal estimates, wet or dry,to use, what temperature to use, and whatvalue for overall seroprevalence of dengueantibody to use. In all of these deliberations,we will be conservative. We already know thatwells will have to be controlled – we do notpresently know their production.
Wet or dry season estimates ofpupae per person
Figure 5 includes the annual number ofcumulative DHF cases in Yogyakarta provincefor the period 1985–2001 and Figure 6 includesthe cumulative monthly DHF cases for the same
period. Given that most transmission occursduring the wet season (Figure 6) and that thereare more pupae per person then (0.47 vs 0.67),we will use the wet season survey results.
Temperature to use in estimatingthe transmission threshold
For temperature, we could use the averagemean temperature for the first five months ofthe year, 27.5 °C when ca. 60% of all cases
Figure 5: Monthly number of reported casesof DHF in Yogyakarta province during
1985-2001
0
200
400
600
800
1,000
1,200
85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 00 01
Year
Cas
esD
HF
0
50
100
150
200
250
300
350
400
450
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
25.5
26.0
26.5
27.0
27.5
28.0
28.5Total (mm) Cases AvgT (°C)
Avg
.ra
infa
ll&
no
.ca
ses
Avg
.te
mp
erat
ure

Targeted source reduction/control strategy for dengue control
Dengue Bulletin – Volume 31, 2007 99
are observed. Yet, epidemic transmission isoften associated with anomalously hightemperatures, almost 29 °C, for the Yogyakartatime series (data not shown). Therefore, to beconservative, we will use a value of 29 °C.
The value of seroprevalence to beused in estimating the transmissionthreshold
This one is a bit more difficult and we willhave to make a crude estimate. Graham et al.conducted a serosurvey in the late 1990s inYogyakarta and estimated that the annualseroconversion rate among children aged 4 to9 years was, for serotypes 1–4, 0.048, 0.077,0.042 and 0.034 respectively.[16] In theendemic situation, the rate of seroconversiondeclines with age as those susceptible toinfection become increasingly rare. Weestimate the age-specific antibody prevalencerate up through age 80 by multiplying the 4–9-year-olds’ rates for each year by one less theprevalence in the previous year. Now, toestimate the average overall prevalences foreach of the dengue strains, we need apopulation estimate of the human agedistribution in Yogyakarta. For this we used theWHO demographic data for Indonesia. Takingthe average of the product of the age-specificage and prevalence proportions gives us theoverall population seroprevalence for eachserotype – 0.54, 0.66, 0.51 and 0.45 forserotypes 1–4 respectively (average: 0.54). Thetransmission thresholds for 0.33 and 0.67 for29 °C from the table of transmission thresholdsgiven in Focks et al.[11] are 0.43 and 0.96respectively. Again, to be conservative, we willuse the value associated with a seroprevalenceof 0.33, i.e., a strategy to bring the area-wideabundance of Ae. aegypti to somewhat below0.43 Ae. aegypti pupae per person.
Identification of the targetedclasses of containers
In Table 11 are listed the most productiveclasses of containers observed in the wetseason surveys. We see that if we controlledthe wells and a single type of container, theStorage in WC (Bak mandi), we would achievea standing crop of Ae. aegypti of 0.37 pupaeper person. With the elimination of just thesetwo types of containers, we are already belowour transmission threshold. Again, to beconservative, if we add to our list of types ofcontainers to be controlled – Wells, Watercontainer (lg) (Bak air) – a fourth type, Tyre, wecan anticipate being at levels of 0.29 and 0.23pupae per person respectively, if all of thetargeted types are completely controlled.
Discussion
The three targeted types of containers accountfor 45% of all production, they also accountfor 65% of all containers. Is this much of astrategy, given the number of containers weare trying to control? We think so for thefollowing reasons: wells, bak mandis and bakairs are in known locations within the houseand are typically masonry in construction. Thismeans that they are easy to find, identify andcontrol with several methods currently availablein Indonesia, e.g. IGRs (Altocid and Sumilarv)and temephos (Abate). Tyres are similarly easyto locate and identify.
Research topics
Most containers are negative for larvaeand pupae
Why is this so? Is it because they are useddomestically and frequently used, cleaned oremptied? Is it because of natural biological
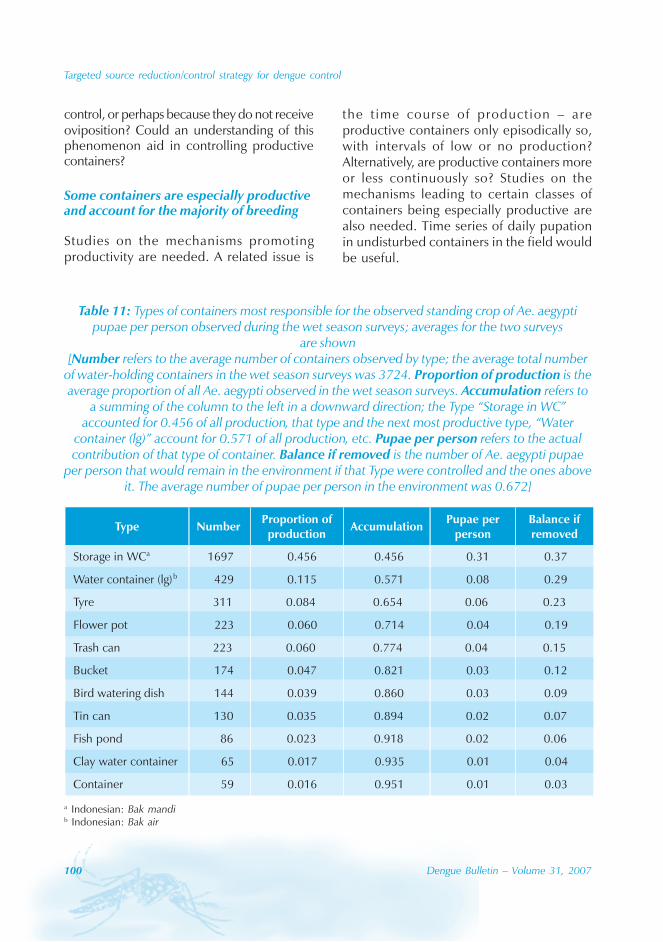
100 Dengue Bulletin – Volume 31, 2007
Targeted source reduction/control strategy for dengue control
control, or perhaps because they do not receiveoviposition? Could an understanding of thisphenomenon aid in controlling productivecontainers?
Some containers are especially productiveand account for the majority of breeding
Studies on the mechanisms promotingproductivity are needed. A related issue is
the t ime course of production – areproductive containers only episodically so,with intervals of low or no production?Alternatively, are productive containers moreor less continuously so? Studies on themechanisms leading to certain classes ofcontainers being especially productive arealso needed. Time series of daily pupationin undisturbed containers in the field wouldbe useful.
Table 11: Types of containers most responsible for the observed standing crop of Ae. aegyptipupae per person observed during the wet season surveys; averages for the two surveys
are shown[Number refers to the average number of containers observed by type; the average total number
of water-holding containers in the wet season surveys was 3724. Proportion of production is theaverage proportion of all Ae. aegypti observed in the wet season surveys. Accumulation refers to
a summing of the column to the left in a downward direction; the Type “Storage in WC”accounted for 0.456 of all production, that type and the next most productive type, “Water
container (lg)” account for 0.571 of all production, etc. Pupae per person refers to the actualcontribution of that type of container. Balance if removed is the number of Ae. aegypti pupae
per person that would remain in the environment if that Type were controlled and the ones aboveit. The average number of pupae per person in the environment was 0.672]
a Indonesian: Bak mandib Indonesian: Bak air

Targeted source reduction/control strategy for dengue control
Dengue Bulletin – Volume 31, 2007 101
Seasonality in transmission and theutility of an early warning system
A recent National Research Council (USA)publication investigating the feasibility ofdeveloping practical and sustainable earlywarning systems (EWSs) for infectious diseasesconcluded that EWSs would provide significantutility where mitigation methods wereavailable.[23] Their primary value lies in theability temporally to focus scarce resources forcontrol in those periods when epidemics werelikely. Cases of dengue and DHF occur invirtually every province and during every monthof the year in Indonesia. Does the seasonalityof dengue transmission in Indonesia suggestthat there would be merit in gaining the abilityto time suppression activities to precede peaktransmission periods based on an EWS? Initialattempts to develop an EWS for dengue inYogyakarta have been promising and will bepursued.
Given that the first five months of the yearaccount for an average of 60% of all cases andthe remaining months report about ca. 5%
each, it seems likely that control would typicallybe continuous. Perhaps the utility of validatedEWSs for Indonesia would be to forecastepidemic years as an aid to the nationalprogramme in securing adequate funding foranticipated epidemic years.
Acknowledgement
This work was partially supported by a numberof institutions including the Jean and JuliusTahija Family Foundation, the Office of GlobalProgrammes, National Oceanographic andAtmospheric Administration (NOAA), GadjahMada University, and the Navy MedicalResearch Center, Silver Spring, MD. We thankall of the staff who dedicated long hours in thefield to carefully collecting mosquito specimensand the data used in this study. We areespecially thankful to Lely Sianturi, SaptoroRusmiarto, Yoyo R. Gionar, Dwiko Susapto, andthe health staff of Gondokusuman for theirdiligence and hard work during the years ofinvestigation. We are also grateful to IqbalElyazar for data set preparation.
References
[1] Suroso T, Holani A, Ali I. Dengue haemorrhagicfever outbreaks in Indonesia 1997-1998.Dengue Bulletin. 1998(22): 45-48.
[2] World Health Organization, Regional Officefor South-East Asia, New Delhi. Report of anexternal review: dengue/dengue haemorrhagicfever prevention and control programme inIndonesia. New Delhi: WHO-SEARO. 2001.Document SEA-Haem Fever-75, SEA-VBC-79.
[3] Kusriastuti R. Internal report. Jakarta,Indonesia: Arbovirus Sub-directorate,Directorate of Vector-Born Disease Control,Ministry of Health 2002.
[4] Martens WJM, Jetten TH, Focks DA. Sensitivityof malaria, schistosomiasis and dengue to
global warming. Climate Change. 1997(35):145-156.
[5] Patz JA, Martens WJ, Focks DA, Jetten TH.Dengue fever epidemic potential as projectedby general circulation models of global climatechange. Environ Health Perspect. 1998 Mar;106(3): 147-53.
[6] Bouma MJ, Poveda G, Rojas W, Chavasse D,Quiñones M, Cox J, Patz J. Predicting high-riskyears for malaria in Colombia using parametersof El Niño Southern Oscillation. Trop Med IntHealth. 1997 Dec; 2(12): 1122-7.
[7] Focks DA, Haile DG, Daniels E, Mount GA.Dynamic life table model for Aedes aegypti(Diptera: Culicidae): analysis of the literature

102 Dengue Bulletin – Volume 31, 2007
Targeted source reduction/control strategy for dengue control
and model development. J Med Entomol.1993 Nov; 30(6): 1003-17.
[8] Focks DA, Haile DG, Daniels E, Mount GA.Dynamic life table model for Aedes aegypti(diptera: Culicidae): simulation results andvalidation. J Med Entomol. 1993 Nov; 30(6):1018-28.
[9] Focks DA, Daniels E, Haile DG, Keesling JE. Asimulation model of the epidemiology ofurban dengue fever: literature analysis, modeldevelopment, preliminary validation, andsamples of simulation results. Am J Trop MedHyg. 1995 Nov; 53(5): 489-506.
[10] Focks DA, Chadee DD. Pupal survey: anepidemiologically significant surveillancemethod for Aedes aegypti: an example usingdata from Trinidad. Am J Trop Med Hyg. 1997Feb; 56(2): 159-67.
[11] Focks DA, Brenner RJ, Hayes J, Daniels E.Transmission thresholds for dengue in terms ofAedes aegypti pupae per person with discussionof their utility in source reduction efforts. Am JTrop Med Hyg. 2000 Jan; 62(1): 11-8.
[12] Focks DA, Brenner RJ, Chadee DD, Trosper J.The use of spatial analysis in the control andrisk assessment of vector-borne diseases. AmEntomol. 1998; (45): 173-183.
[13] World Health Organization. Key Issues indengue vector control towards theoperationalization of a global strategy: reportof consultation. Geneva: WHO; 1995.Document CTD/FIL (Den)/IC.96.1).
[14] Focks DA. A review of entomological samplingmethods and indicators for dengue vectors.Geneva: World Health Organiation, 2004.Document TDR/IDE/Den/03.1.
[15] Focks DA, Lele SR, Juffrie M, SuvannadabbaS, Sobel AL, Trahan MW. Early warning systemsfor dengue in Indonesia and Thailand. ProcUnified Science & Tech Reducing BiologicalThreats & Countering Terrorism.Albuquerque: New Mexico. 2002.
[16] Nam VS, Yen NT, Holynska M, Reid JW, KayBH. National progress in dengue vector control
in Vietnam: survey for Mesocyclops(Copepoda), Micronecta (Corixidae), and fishas biological control agents. Am J Trop MedHyg. 2000 Jan; 62(1): 5-10.
[17] Graham RR, Juffrie M, Tan R, Hayes CG,Laksono I, Ma’roef C, Erlin, Sutaryo, PorterKR, Halstead SB. A prospectiveseroepidemiologic study on dengue inchildren four to nine years of age in Yogyakarta,Indonesia I. studies in 1995-1996. Am J TropMed Hyg. 1999 Sep; 61(3): 412-9.
[18] Gionar YR, Rusmiarto S, Susapto D, Bangs MJ.Use of a funnel trap for collecting immatureAedes aegypti and copepods from deep wellsin Yogyakarta, Indonesia. J Am Mosq ControlAssoc. 1999 Dec; 15(4): 576-80.
[19] Bangs MJ, Focks DA. Abridged pupaidentification key to the common container-breeding mosquitoes in urban Southeast Asia.J Am Mosq Control Assoc, 2006 Sep; 22(3);565-572.
[20] Tun-Lin W, Kay BH, Barnes A. The PremiseCondition Index: a tool for streamliningsurveys of Aedes aegypti. Am J Trop Med Hyg.1995 Dec; 53(6): 591-4.
[21] Barker-Hudson P, Jones R, Kay BH.Categorization of domestic breeding habitatsof Aedes aegypti (Diptera: Culicidae) inNorthern Queensland, Australia. J MedEntomol. 1988 May; 25(3): 178-82.
[22] Tun-Lin W, Kay BH, Barnes A. Understandingproductivity, a key to Aedes aegyptisurveillance. Am J Trop Med Hyg. 1995 Dec;53(6): 595-601.
[23] Burke D, Carmichael A, Focks D, Grimes D,Harte J, Lele S, Martens P, Mayer J, Means L,Pulwarty R, Real L, Ropelewski C, Rose J,Shope R, Simpson J, Wilson M. Under theweather: exploring the linkages betweenclimate, ecosystems, infectious disease, andhuman health. Washington, DC.: NationalResearch Council, National Academy Press,2001.

Dengue Bulletin – Volume 31, 2007 103
Elevated levels of soluble tumour necrosis factorreceptor 1, thrombomodulin and soluble endothelial
cell adhesion molecules in patients withdengue haemorrhagic fever
Beti Ernawati Dewia,b, Tomohiko Takasakia, T. Mirawati Sudirob, R.H. Nelwanc
and Ichiro Kuranea
aLaboratory of Vector-Borne Viruses, Department of Virology 1, National Institute of Infectious Diseases,1-23-1 Toyama, Shinjuku-ku, Tokyo, 163-8640, Japan
bDepartment of Microbiology, Medical Faculty, University of Indonesia, Jalan Pegangsaan Timur No. 16,Jakarta 10320, Indonesia
cDepartment of Internal Medicine, Medical Faculty, University of Indonesia, Jakarta, Indonesia
Abstract
The pathological mechanism of dengue haemorrhagic fever (DHF) is still poorly understood. Previousstudies have suggested that immune responses contribute to an increase in capillary permeability. Weexamined the levels of cytokines and activated endothelial substances in serum samples collected fromDHF patients in Indonesia. We measured the levels of soluble TNF-R1, MCP-1, GM-CSF, IL-17, TNF-α,soluble thrombomodulin, soluble E-selectin, soluble ICAM-1 and soluble VCAM-1. The levels ofactivated endothelial substances such as sE-selectin, sICAM-1 and sVCAM-1 were higher in DHF patientsthan in healthy controls. High levels of soluble activated endothelial substances suggest that endothelialcells are highly activated. When compared with patients with other febrile illness (OFI) or healthycontrols, the levels of sTNF-R1 were higher in DHF patients. A similar trend was observed in the level ofthrombomodulin. MCP-1, GM-CSF and IL-17 were not detected in serum samples from any patientsand healthy controls. TNF-α was detected in 5(9%) of 53 patients. The results suggest that endothelialcells are highly activated in DHF patients and TNF-α is one of the factors which contributes to theactivation.
Keywords: Tumor necrosis factor receptor-1; Thrombomodulin; Vascular cell adhesion molecule-1; Intercellularadhesion molecule-1.
E-mail: [email protected]
Introduction
Dengue virus infection continues to increasein tropical and sub-tropical countries. Denguevirus causes a spectrum of clinical illnesses
ranging from a mild disease as dengue fever(DF) to dengue haemorrhagic fever (DHF) anddengue shock syndrome (DSS).[1] It is estimatedthat up to 100 million individuals are infectedwith dengue virus, and 500 000 cases of DHF

104 Dengue Bulletin – Volume 31, 2007
Elevated level of cytokines on DHF patients
and 25 000 deaths occur annually.[1] Thepathological mechanisms of DHF are still poorlyunderstood. The role of host immune responseshave been examined in a number of studies.These studies have suggested that differenttypes of cytokines and abnormalities incoagulation may cause an increase in thecapillary permeability, the hallmark of DHF.[2,3]
The absence of massive structural damage,a short-lived nature of the plasma leakagesyndrome and a remarkably rapid recovery ofchildren with DSS all suggest that alteredpermeability is induced by soluble mediators.[4]
It has been reported that serum levels oftumour necrosis factor alpha (TNF-α) areelevated in patients with dengue haemorrhagicfever.[5,6] The role of TNF-α in the increasedpermeability of vascular endothelial cells hasbeen postulated.[7,8]
TNF-α starts to affect cell function bybinding to the specific, high-affinity receptorson the surface; TNF-R1 (p55 TNFR) and TNF-R2 (p75 TNFR).[9,10] Soluble forms of TNF-R1and TNF-R2 are present in the serum[11] andappear to play a role as modulators of thebiological function of TNF-α, modifying thefunction and availability of TNF-α. [12,13]
Measuring sTNFRs in serum offers someadvantages compared with direct qualificationof TNF-α, because TNF-α is rapidly clearedfrom circulation, and some assays are unableto detect TNF-α bound to soluble TNFRs.[14]
Soluble TNFRs are very stable and their serumconcentrations correlate well with those ofTNF-α.[15,16]
Most of the biological effects induced byTNF-α have been shown to be mediated byTNF-R1.[17] TNF-alpha induces expression ofadhesion molecules such as ICAM-1, VCAM-1and E-selectin under the control of TNF-R1.[18]
TNF-R1 also plays a key role in TNF-alpha-dependent responses that include proliferation,differentiation and apoptosis.[19] Wachtel et al.
(2001)[20] showed that down regulation of tightjunction components such as occluding by TNFis mediated via TNF-R1.
In the present study, we examined thelevel of sTNF-R1 in serum samples from DHFpatients. In addition, we also measured thelevels of thrombomodulin and activationmolecules released from endothelial cells inthe serum samples.
Materials and methods
Study population
Blood specimens were collected from 53patients admitted to Cipto MangunkusumoHospital, Jakarta, Indonesia, between May andAugust 2002. After informed consent wasobtained, blood specimens were obtained fromadult patients with suspected dengue infection.The patients were diagnosed as dengue virusinfection by clinical findings and laboratoryconfirmation – IgM-capture ELISA (PanBio,Brisbane, Australia) and IgG-capture ELISA(PanBio, Brisbane, Australia). The patients wereclinically classified as DHF according to thecriteria of the World Health Organization;[21]
grade I: fever associated with non-specificconstitutional symptom and positive torniquettest as the only haemorrhagic manifestation;grade II: the same symptom with grade I withthe spontaneous haemorrhagic manifestation;grade III: circulatory failure manifested by rapid,weak pulse with narrowing of the pulsepressure (<20 mm of Hg) or hypotension; gradeIV: profound shock with undetectable bloodpressure and pulse.
Fifty-three patients are included in thisstudy – 25 females and 28 males, with anaverage age of 26.75±10.77 years. Eighteenpatients were classified as DHF grade II, and24 patients as DHF grade I; 11 patients as non-dengue infection (other febrile illness, OFI).

Elevated level of cytokines on DHF patients
Dengue Bulletin – Volume 31, 2007 105
Plasma samples were also obtained from 20healthy donors with a mean age of 37.29±9.67years as controls.
Serum sample
Peripheral blood (5–10 ml) was drawn into steriletubes containing EDTA. Blood samples werecollected everyday from the day of admissionuntil the days when body temperature andthrombocyte count became normal. Bloodspecimens were centrifuged at 800 rpm for 10minutes and the plasma was separated and thenstored at –80 °C until use. The plasma sampleswere transported to the NIID, Tokyo, Japan, indry ice and stored at –80 °C until use.
Measurement of level of cytokines,chemokines and activatedendothelial substance bystandard ELISA
Levels of tumour necrosis factor-α (TNF-α),monocyte chemoattractant protein-1 (MCP-1),interleukin 17 (IL-17), granulocyte macrophagecolony stimulating factor (GM-CSF), soluble E-selectin, soluble vascular cell adhesionmolecule-1 (sVCAM-1) and ICAM-1 wereassessed with ELISA kits, according to the manualof the manufacturer (BioSource InternationalInc., USA). Levels of soluble tumour necrosisfactor receptor 1 (s TNF-R1) were measuredusing a human sTNF-R1 immunoassay (R&DSystems, Inc., USA). Levels of thrombomodulinwere measured with soluble CD141 ELISA kits(Diaclone Research, France).
Statistical analysis
Differences were statistically analysed byMann-Whitney U-test. The difference wasconsidered to be significant, when p value wasless than 0.05.
Results
Level of TNF-R1
Levels of soluble TNF-R1 in DHF patients, OFIpatients during disease days 1–15 and healthycontrols are shown in Figure 1. The highestmean levels were 230.51±67.07 pg/ml on day4 after onset of fever in patients with grade IIand 245.02±47.72 pg/ml on day 10 after onsetof fever in patients with grade I. The highestmean level was 184±77.64 pg/ml in patientswith OFI, and the mean level in healthycontrols was 59.37±15.12 pg/ml. The levelsof soluble TNF-R1 in patients with grades I andII were significantly higher those in healthycontrols, and those in patients with OFI ondays 7, 8 and 9 after onset of fever.
Figure 1: Variation of level of soluble TNF-R1 in the plasma of DHF patients, otherfebrile illness (OFI) patients and healthy
controls on days after onset of fever[The symbol represents the soluble TNF-R1
value obtained from individual patients]
DHF-II DHF-I OFI Healthy600
500
400
300
200
100
0
pg/m
l
Days after onset of fever
3 5 7 9 11 13 15 1 3 5 7 9 11 5 7 9 11 13 15
Level of thrombomodulin, sE-selectin,sVCAM-1 and sICAM-1
The levels of thrombomodulin, soluble E-selectin, soluble VCAM-1 and soluble ICAM-1were assessed as markers for activation of
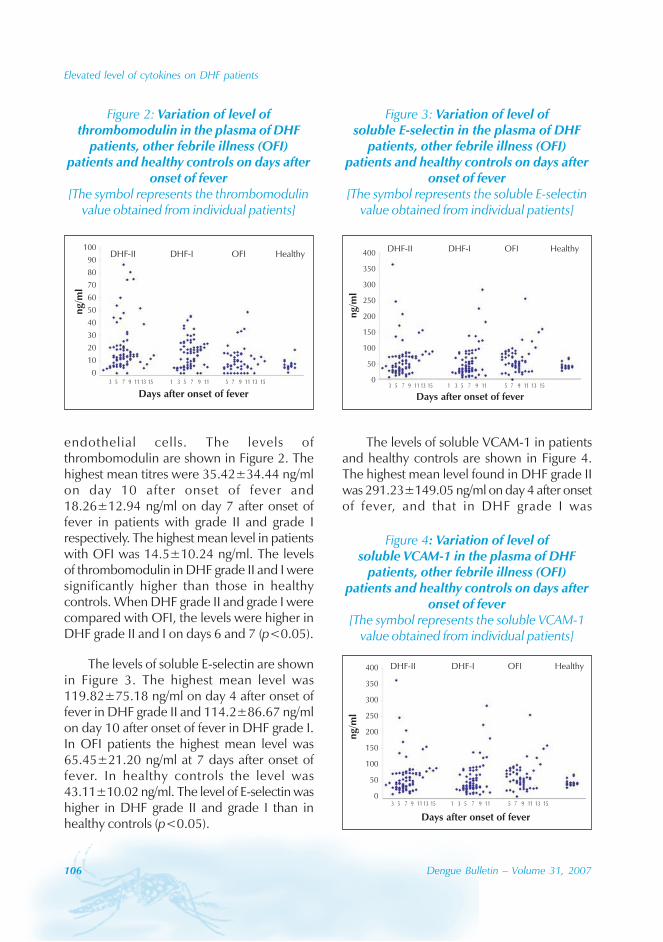
106 Dengue Bulletin – Volume 31, 2007
Elevated level of cytokines on DHF patients
endothelial cells. The levels ofthrombomodulin are shown in Figure 2. Thehighest mean titres were 35.42±34.44 ng/mlon day 10 after onset of fever and18.26±12.94 ng/ml on day 7 after onset offever in patients with grade II and grade Irespectively. The highest mean level in patientswith OFI was 14.5±10.24 ng/ml. The levelsof thrombomodulin in DHF grade II and I weresignificantly higher than those in healthycontrols. When DHF grade II and grade I werecompared with OFI, the levels were higher inDHF grade II and I on days 6 and 7 (p<0.05).
The levels of soluble E-selectin are shownin Figure 3. The highest mean level was119.82±75.18 ng/ml on day 4 after onset offever in DHF grade II and 114.2±86.67 ng/mlon day 10 after onset of fever in DHF grade I.In OFI patients the highest mean level was65.45±21.20 ng/ml at 7 days after onset offever. In healthy controls the level was43.11±10.02 ng/ml. The level of E-selectin washigher in DHF grade II and grade I than inhealthy controls (p<0.05).
The levels of soluble VCAM-1 in patientsand healthy controls are shown in Figure 4.The highest mean level found in DHF grade IIwas 291.23±149.05 ng/ml on day 4 after onsetof fever, and that in DHF grade I was
Figure 2: Variation of level ofthrombomodulin in the plasma of DHF
patients, other febrile illness (OFI)patients and healthy controls on days after
onset of fever[The symbol represents the thrombomodulin
value obtained from individual patients]
Days after onset of fever
ng/m
l
100
90
80
70
60
50
40
30
20
10
0
DHF-II DHF-I OFI Healthy
3 5 7 9 11 13 15 1 3 5 7 9 11 5 7 9 11 13 15
Figure 3: Variation of level ofsoluble E-selectin in the plasma of DHF
patients, other febrile illness (OFI)patients and healthy controls on days after
onset of fever[The symbol represents the soluble E-selectin
value obtained from individual patients]
Days after onset of fever3 5 7 9 11 13 15 1 3 5 7 9 11 5 7 9 11 13 15
ng/m
l
400
350
300
250
200
150
100
50
0
DHF-II DHF-I OFI Healthy
Figure 4: Variation of level ofsoluble VCAM-1 in the plasma of DHF
patients, other febrile illness (OFI)patients and healthy controls on days after
onset of fever[The symbol represents the soluble VCAM-1
value obtained from individual patients]
DHF-II DHF-I OFI Healthy
ng/m
l
400
350
300
250
200
150
100
50
0
Days after onset of fever
3 5 7 9 11 13 15 1 3 5 7 9 11 5 7 9 11 13 15

Elevated level of cytokines on DHF patients
Dengue Bulletin – Volume 31, 2007 107
109.79±27.71 ng/ml on day 10 after onset offever. The highest level of sVCAM-1 was81.46±17.15 ng/ml in OFI patients, and thelevel was 7.01±2.51 ng/ml in healthy controls.The levels of sVCAM-1 were significantly higherin DHF grades II and I than in healthy controls.When DHF grade II patients were comparedwith OFI, significant differences were observedfrom day 4 to day 9 after onset of fever.
The levels of soluble ICAM-1 are shownin Figure 5. Low level was observed in healthycontrols (12.4±3.11 ng/ml). The highest meanlevel in DHF grade II was 32.99±17.67 ng/mlon day 4 after onset of fever, and in DHF gradeI, the highest mean level was 21.23±6.52 ng/ml on day 10 after onset of fever. The highestmean level in OFI patients was 21.63±9.6 ng/ml at day 10 after onset of fever. The levels ofsICAM-1 were significantly higher in patientswith DHF than in healthy controls, whichshowed a significant difference (p<0.05).
A correlation was not found among plasmalevel sTNF-R1 with the level of activated
endothelial substances. The highest levels ofcytokines and activated endothelial substanceswere detected on the same days after onsetof fever. For example, in DHF grade II patients,the highest mean levels of sTNF-R1 sE-selectin,sVCAM-1 and sICAM-1 were detected on day4 after onset of fever. In DHF grade I, thehighest levels of sTNF-R1, sE-selectin, sVCAM-1 and sICAM-1 were detected on day 10 afteronset of fever.
Level of other cytokines
The plasma samples were also examined forthe levels of TNF-α, MCP-1, GM-CSF and IL-17. TNF-α was detected in 5(9%) of 53 patientswith DHF, but were not detected in any of thehealthy controls. MCP-1, GM-CSF and IL-17were not detected in plasma samples from anyof the DHF patients or healthy controls.
Discussion
It has been hypothesized that DHF is causedby soluble mediator induced in immuneresponses. Recent studies have demonstratedthe elevation of multiple mediators in DHFpatients, including IL-6,[22] tumour necrosisfactor alpha (TNF-α),[6] IL-10,[23] IFN-gamma(IFN-γ),[24] and IL-2.[25] In the present study wedid not find high levels of TNF-α; however,levels of TNF-R1 were higher in DHF patientsthan in healthy controls or patients with OFI.Direct qualification of TNF-α is often difficultbecause TNF-α is rapidly cleared from thecirculation and some assays cannot detect TNF-α bound to soluble TNFRs.[14]
It has been known that TNF-α has manyeffects on the endothelial cell barrier functions.TNF-α induces the release of cytokines andchemokines,[26] the production of enzymes,such as cyclooxygenases-2[27] and nitric oxidesynthesis. [28] TNF-α also increases the
Figure 5: Variation of level ofsoluble ICAM-1 in the plasma of DHF
patients, other febrile illness (OFI)patients and healthy controls on days after
onset of fever[The symbol represents the soluble ICAM-1
value obtained from individual patients]
Days after onset of fever
ng/m
l
140
120
100
80
60
40
20
0
DHF-II DHF-I OFI Healthy
3 5 7 9 11 13 15 1 3 5 7 9 11 5 7 9 11 13 15

108 Dengue Bulletin – Volume 31, 2007
Elevated level of cytokines on DHF patients
expression of adhesion molecules onendothelial cells.[29] Ferrero et al. 2001[30]
showed that TNF induced alteration ofendothelial permeability in vitro and in vivovia TNF-R1. High levels of TNF-R1 in DHFpatients suggest that TNF-α may play importantroles in the development of DHF, because thelevel of sTNFRs in serum correlate well withthe level of TNF-α[15,16] and because most ofthe biological effects of TNF-α have beenattributed to TNF-R1.[17]
Endothelial cells line the inner surface ofblood vessels, and play an important role invascular functions as a barrier between bloodand interstitial compartments.[31] It is knownthat hormones, cytokines and neurotransmittersregulate endothelial cell functions.[32] We foundthat levels of activated endothelial substances– sICAM-1, sVCAM-1 and sE-selectin – wereelevated in DHF patients. Elevation of thesesubstances in serum indicates that endothelialcells are activated during dengue virus infection.
In addition, we measured the levels of anendothelial cell injure marker, thrombomodulin.We found that mean levels of thrombomodulinwere elevated in DHF patients compared tothose in healthy controls and in patients withOFI. Thrombomodulin is an endothelial cellsurface glycoprotein and possesses an
anticoagulant property. This protein wasimmobilized onto polytetrafluorethylenesurfaces to create biomaterials with enhancedhaemocompatibility.[33] Thrombomodulin as athrombin-binding protein serves to initiate theprotein C anticoagulant pathway.[34] High levelsof thrombomodulin in the microcirculationsuggest that thrombin might accumulate andultimately lead to microvascular occlusion.[35]
It is well-known that thrombin increases thepermeability of endothelial cells.[32] We feelthat high levels of thrombomodulin in DHFpatients reflect the functional changes ofvascular endothelial cells in patients with DHF.
Acknowledgments
We thank Muhareva Raekiansyah MS, S. TajimaPhD, Mr A. Kotaki, R. Nerome PhD, Laboratoryof Vector-Borne Viruses, Department ofVirology 1, National Institute of InfectiousDiseases, for their technical support. This studywas supported by the grant-in-aid (No.12877045) from the Ministry of Education,Sciences, Sports, and Culture, Japan, and thegrant for Research on Emerging and Re-Emerging Infectious Diseases from Ministry ofHealth, Labour and Welfare, Japan, (H18-Shinkou-Ippan-009).
References
[1] Gubler DJ. Resurgent vector-borne diseases asa global health problem. Emerg Infect Dis. 1998Jul-Sep; 4(3): 442-50.
[2] Bosch I, Xhaja K, Estevez L, Raines G, MelicharH, Warke RV, Fournier MV, Ennis FA, RothmanAL. Increased production of interleukin-8 inprimary human monocytes and in humanepithelial and endothelial cell lines afterdengue virus challenge. J Virol. 2002 Jun;76(11): 5588-97.
[3] Libraty DH, Pichyangkul S, Ajariyakhajorn C,Endy TP, Ennis FA. Human dendritic cells areactivated by dengue virus infection:enhancement by gamma interferon andimplications for disease pathogenesis. J Virol.2001 Apr; 75(8): 3501-8.
[4] Innis BL. Dengue and dengue hemorrhagicfever. In: Exotic viral infection. Poterfield JS. Ed.London: Chapman & Hall; 1995. p.103.

Elevated level of cytokines on DHF patients
Dengue Bulletin – Volume 31, 2007 109
[5] Iyngkaran N, Yadav M, Sinniah M. Augmentedinflammatory cytokines in primary dengueinfection progressing to shock. Singapore MedJ. 1995 Apr; 36(2): 218-21.
[6] Hober D, Delannoy AS, Benyoucef S, DeGroote D, Wattré P. High levels of sTNFR p75and TNF alpha in dengue-infected patients.Microbiol Immunol. 1996; 40(8): 569-73.
[7] Brett J, Gerlach H, Nawroth P, Steinberg S,Godman G, Stern D. Tumor necrosis factor/cachectin increases permeability ofendothelial cell monolayers by a mechanisminvolving regulatory G proteins. J Exp Med.1989 Jun 1;169(6):1977-91.
[8] Jacobs M, Levin M. An improved endothelialbarrier model to investigate denguehaemorrhagic fever. J Virol Methods. 2002 Jul;104(2): 173-85.
[9] Loetscher H, Pan YC, Lahm HW, Gentz R,Brockhaus M, Tabuchi H, Lesslauer W.Molecular cloning and expression of thehuman 55 kd tumor necrosis factor receptor.Cell. 1990 Apr 20; 61(2): 351-9.
[10] Smith CA, Davis T, Anderson D, Solam L,Beckmann MP, Jerzy R, Dower SK, Cosman D,Goodwin RG. A receptor for tumor necrosisfactor defines an unusual family of cellularand viral proteins. Science. 1990 May 25;248(4958): 1019-23.
[11] Pinckard JK, Sheehan KC, Arthur CD,Schreiber RD. Constitutive shedding of bothp55 and p75 murine TNF receptors in vivo. JImmunol. 1997 Apr 15; 158(8): 3869-73.
[12] Peetre C, Thysell H, Grubb A, Olsson I. A tumornecrosis factor binding protein is present inhuman biological fluids. Eur J Haematol. 1988Nov; 41(5): 414-9.
[13] Aderka D, Engelmann H, Shemer-Avni Y,Hornik V, Galil A, Sarov B, Wallach D. Variationin serum levels of the soluble TNF receptorsamong healthy individuals. LymphokineCytokine Res. 1992 Jun; 11(3): 157-9.
[14] Engelberts I, Möller A, Schoen GJ, van derLinden CJ, Buurman WA. Evaluation ofmeasurement of human TNF in plasma by
ELISA. Lymphokine Cytokine Res. 1991 Apr;10(1-2): 69-76.
[15] Aukrust P, Liabakk NB, Müller F, Lien E, EspevikT, Frøland SS. Serum levels of tumor necrosisfactor-alpha (TNF alpha) and soluble TNFreceptors in human immunodeficiency virustype 1 infection–correlations to clinical,immunologic, and virologic parameters. JInfect Dis. 1994 Feb; 169(2): 420-4.
[16] Zangerle R, Gallati H, Sarcletti M, Wachter H,Fuchs D. Tumor necrosis factor alpha andsoluble tumor necrosis factor receptors inindividuals with human immunodeficiencyvirus infection. Immunol Lett. 1994 Jul; 41(2-3): 229-34.
[17] Weiss T, Grell M, Hessabi B, Bourteele S, MüllerG, Scheurich P, Wajant H. Enhancement ofTNF receptor p60-mediated cytotoxicity byTNF receptor p80: requirement of the TNFreceptor-associated factor-2 binding site. JImmunol. 1997 Mar 1; 158(5): 2398-404.
[18] Wolf D, Hallmann R, Sass G, Sixt M, Küsters S,Fregien B, Trautwein C, Tiegs G. TNF-alpha-induced expression of adhesion molecules inthe liver is under the control of TNFR1–relevance for concanavalin A-inducedhepatitis. J Immunol. 2001 Jan 15; 166(2):1300-7.
[19] Doan JE, Windmiller DA, Riches DW.Differential regulation of TNF-R1 signaling:lipid raft dependency of p42mapk/erk2activation, but not NF-kappaB activation. JImmunol. 2004 Jun 15; 172(12): 7654-60.
[20] Wachtel M, Bolliger MF, Ishihara H, Frei K,Bluethmann H, Gloor SM. Down-regulationof occludin expression in astrocytes by tumournecrosis factor (TNF) is mediated via TNF type-1 receptor and nuclear factor-kappaBactivation. J Neurochem. 2001 Jul; 78(1): 155-62.
[21] World Health Organization. Denguehemorrhagic fever: diagnosis, treatment andcontrol. Geneva: WHO, 1986.
[22] Juffrie M, Meer GM, Hack CE, Haasnoot K,Sutaryo, Veerman AJ, Thijs LG. Inflammatory

110 Dengue Bulletin – Volume 31, 2007
Elevated level of cytokines on DHF patients
mediators in dengue virus infection inchildren: interleukin-6 and its relation to C-reactive protein and secretory phospholipaseA2. Am J Trop Med Hyg. 2001 Jul;65(1):70-5.
[23] Green S, Vaughn DW, Kalayanarooj S,Nimmannitya S, Suntayakorn S, Nisalak A,Rothman AL, Ennis FA. Elevated plasmainterleukin-10 levels in acute dengue correlatewith disease severity. J Med Virol. 1999 Nov;59(3): 329-34.
[24] Azeredo EL, Zagne SM, Santiago MA, GouveaAS, Santana AA, Neves-Souza PC, NogueiraRM, Miagostovich MP, Kubelka CF.Characterisation of lymphocyte response andcytokine patterns in patients with dengue fever.Immunobiology. 2001 Dec; 204(4): 494-507.
[25] Kurane I, Innis BL, Nimmannitya S, Nisalak A,Meager A, Janus J, Ennis FA. Activation of Tlymphocytes in dengue virus infections. Highlevels of soluble interleukin 2 receptor, solubleCD4, soluble CD8, interleukin 2, andinterferon-gamma in sera of children withdengue. J Clin Invest. 1991 Nov; 88(5): 1473-80.
[26] Mantovani A, Sozzani S, Introna M.Endothelial activation by cytokines. Ann N YAcad Sci. 1997 Dec 15; 832: 93-116.
[27] Mark KS, Trickler WJ, Miller DW. Tumornecrosis factor-alpha inducescyclooxygenase-2 expression andprostaglandin release in brain microvesselendothelial cells. J Pharmacol Exp Ther. 2001Jun;297(3):1051-8.
[28] Bove K, Neumann P, Gertzberg N, Johnson A.Role of ecNOS-derived NO in mediating TNF-
induced endothelial barrier dysfunction. AmJ Physiol Lung Cell Mol Physiol. 2001 May;280(5): L914-22.
[29] Nakao S, Kuwano T, Ishibashi T, Kuwano M,Ono M. Synergistic effect of TNF-alpha insoluble VCAM-1-induced angiogenesisthrough alpha 4 integrins. J Immunol. 2003Jun 1; 170(11): 5704-11.
[30] Ferrero E, Zocchi MR, Magni E, Panzeri MC,Curnis F, Rugarli C, Ferrero ME, Corti A. Rolesof tumor necrosis factor p55 and p75 receptorsin TNF-alpha-induced vascular permeability.Am J Physiol Cell Physiol. 2001 Oct; 281(4):C1173-9.
[31] Crone C. Modulation of solute permeabilityin microvascular endothelium. Fed Proc. 1986Feb; 45(2): 77-83.
[32] Rabiet MJ, Plantier JL, Rival Y, Genoux Y,Lampugnani MG, Dejana E. Thrombin-induced increase in endothelial permeabilityis associated with changes in cell-to-celljunction organization. Arterioscler Thromb VascBiol. 1996 Mar; 16(3): 488-96.
[33] Sperling C, Salchert K, Streller U, Werner C.Covalently immobilized thrombomodulininhibits coagulation and complementactivation of artificial surfaces in vitro.Biomaterials. 2004 Sep; 25(21): 5101-13.
[34] Esmon CT. Coagulation and inflammation. JEndotoxin Res. 2003; 9(3): 192-8.
[35] Esmon CT. The roles of protein C andthrombomodulin in the regulation of bloodcoagulation. J Biol Chem. 1989 Mar 25;264(9): 4743-6.

Dengue Bulletin – Volume 31, 2007 111
Increased dengue virus-infected endothelialcell apoptosis caused by antibodies against
nonstructural protein 1
Chiou-Feng Lin, Shu-Wen Wan, Mei-Chun Chen, Huan-Yao Leiand Yee-Shin Lin
Department of Microbiology and Immunology, National Cheng Kung University Medical College,Tainan, Taiwan, Republic of China
Abstract
Vascular dysfunction is a hallmark of severe dengue haemorrhagic fever (DHF) in dengue virus (DENV)infection. Both viral infection and immunopathogenesis are involved in the vasculopathy of DHF. Wepreviously showed that antibodies against DENV nonstructural protein 1 (NS1) may cross-react withendothelial cells. A further study demonstrated the pathogenic role of anti-NS1 antibodies on endothelialdysfunction by inducing cell death and inflammation. In this study, we investigated the effect of anti-NS1 antibodies on DENV-infected human endothelial cells. We found increased binding activity ofanti-NS1 antibodies to endothelial cells after DENV infection. Either DENV infection or anti-NS1treatment caused endothelial cell apoptosis. Importantly, co-treatment with anti-NS1 antibodies increasedDENV-induced endothelial cell apoptosis. The generation of nitric oxide (NO) could be detected inanti-NS1-stimulated, but not DENV-infected, endothelial cells. Furthermore, anti-NS1-inducedendothelial cell apoptosis was NO-dependent, whereas DENV infection-induced apoptosis was NO-independent. These results suggest an additive effect but distinct mechanisms between anti-NS1 antibodiesand DENV in endothelial cell damage. These findings indicate that both viral infection and cross-reactive antibodies may be involved in dengue pathogenesis by causing endothelial dysfunction.
Keywords: Dengue virus (DENV); Autoantibody; NS1; Endothelial cells; Apoptosis.
E-mail: [email protected]
Introduction
Dengue virus (DENV) causes human diseaseslike mild dengue fever (DF) and severe denguehaemorrhagic fever and dengue shocksyndrome (DHF/DSS) in most tropical andsubtropical areas of the world. Sequentialinfection with different DENV serotypes mayinfluence the severity of the disease.[1,2]
Antibody-dependent enhancement (ADE) of
DENV infection is responsible for DHF in thesecondary infection due to the presence ofcross-reactive and non-neutralizing antibodiesfrom prior infection.[3-7] In addition to ADE, viralvariation and immunopathogenesis are involvedin the progression of DHF.[5,6,8-10] We previouslyfound that anti-DENV nonstructural protein 1(NS1) antibodies cross-reacted with endothelialcells and induced these cells to undergoapoptosis.[11-14] Autoimmunity-mediatedendothelial cell damage may, therefore, also

112 Dengue Bulletin – Volume 31, 2007
Anti-NS1 enhances DENV-induced endothelial cell apoptosis
contribute to the pathogenesis of denguedisease.
Vasculopathy, coagulopathy andthrombocytopenia are the hallmarks of severedengue haemorrhagic syndrome.[1,5,6] Previousstudies showed the manifestation ofvasculopathy characterized by endothelialdysfunction caused by direct viral cytotoxicityas well as immune-mediated inflammatoryresponses and cellular damage.[15,16] In addition,we demonstrated that anti-NS1 antibodiespresent in dengue patient sera causedendothelial cell apoptosis.[11,12] Furthermore,NS1 protein can be expressed in a glycosyl-phosphatidylinositol (GPI)-linked form on thesurface of DENV-infected cells.[17] In thepresent study, we investigated the apoptoticeffect of DENV infection in human endothelialcells in the presence of anti-NS1 antibodies.
Materials and methods
Cell cultures
Human umbilical cord vein endothelial cells(HUVECs) were cultured in modified M-199medium as previously described.[11] Humanmicrovascular endothelial cell line-1 (HMEC-1) was passed in culture plates containingendothelial cell growth medium M200 (CascadeBiologics) composed of LSGS (2% fetal bovineserum, 1 μg/ml hydrocortisone, 10 ng/mlepidermal growth factor, 3 ng/ml basic fibroblastgrowth factor, 10 μg/ml heparin, and antibiotics.C6/36 cells were cultured in DMEM mediumcontaining 10% fetal bovine serum andantibiotics. Fetal bovine serum was pre-incubated at 56 °C for 30 minutes to inactivatecomplement before cell culture. Forexperiments, 1000 U/ml trypsin and 0.5 mMethylenediaminetetraacetic acid (EDTA) wereused to detach cells.
Viruses
DENV-2 (PL046, Taiwan-isolated) wasmaintained in C6/36 cells. Briefly, a monolayerof C6/36 cells was incubated with DENV-2 ata multiplicity of infection (MOI) of 0.01 andincubated at 26 °C in 5% CO2 for 5 days. Thecultured medium was harvested and cell debriswas removed by centrifugation at 900 × g for10 minutes. After further centrifugation at16 000 × g for 10 minutes, the virussupernatant was collected and stored at -70 °Cuntil use. The virus titre was determined usinga plaque assay.
Generating anti-NS1 antibodies
Recombinant DENV-2 NS1 was expressed andpurified as previously described.[13] To generateantibodies against DENV NS1, we immunizedBALB/c mice intraperitoneally, once with 25μg of recombinant NS1 protein emulsified incomplete Freund’s adjuvant and then four timeswith 25 μg of recombinant NS1 proteinemulsified in incomplete Freund’s adjuvant. Serawere obtained 3 or 4 days after the lastimmunization. The immunoglobulin G (IgG)fractions from hyperimmunized mouse sera andnormal mouse sera used for the control werepurified using a protein G-Sepharose affinitychromatography column (AmershamPharmacia, Uppsala, Sweden).
Detecting binding activity
Endothelial cells (5 × 105) were washed withphosphate-buffered saline (PBS) and fixed with1% formaldehyde in PBS at room temperaturefor 10 minutes, then washed again with PBS.Mouse anti-NS1 IgG or control IgG were thenincubated with cells at 4 °C for 1 hour. Afterthe cells had been washed three times with

Anti-NS1 enhances DENV-induced endothelial cell apoptosis
Dengue Bulletin – Volume 31, 2007 113
PBS, they were incubated with 1 μl of 1 mg/mlfluorescein isothiocyanate (FITC)-conjugatedgoat anti-mouse IgG (Jackson ImmunoResearchLaboratories, West Grove, PA) at 4 °C for 1 hourand then washed again with PBS. The bindingactivity of mouse anti-NS1 or control IgG toendothelial cells was analysed using flowcytometry (FACSCalibur; BD Biosciences, SanJose, CA) with excitation set at 488 nm.
Detecting cell death
Cells were fixed with 70% ethanol in PBS for aterminal deoxynucleotidyl transferase-mediateddUTP nick-end-labeling (TUNEL) reaction usingthe ApoAlert DNA Fragmentation Assay Kit(Clontech, Palo Alto, CA) according to themanufacturer’s instructions, and then analysedusing flow cytometry.
Detecting caspase-3 activity
Caspase-3 activation was determined using theApoAlert caspase-3 colorimetric assay kit(Clontech) according to the manufacturer’sinstructions. Relative optical density (OD)measurements were performed using amicroplate reader (Molecular Devices).
Detecting nitric oxide production
The production of nitric oxide (NO) wasassessed as the accumulation of nitrite (NO2
-)in the medium using a colorimetric reactionwith the Griess reagent. Briefly, the culturesupernatants were collected and mixed withan equal (1:1) volume of Griess reagent (0.1%N - ( 1 - n a p h t h y l ) e t h y l e n e d i a m i n edihydrochloride, 1% sulfanilamide, and 2.5%H3PO4). The absorbance was measured at 540nm using a microplate reader. NaNO2 wasdissolved in double-distilled H2O and used forthe standard control (from 1 to 50 μM).
Statistical analysis
Using the Student’s unpaired t-tests inSigmaPlot version 8.0 for Windows (CytelSoftware Corporation, Cambridge, MA), P-values were determined for all data from threeindependent experiments. Statisticalsignificance was set at P < 0.05.
Results
Binding activity of anti-NS1 toDENV-infected humanendothelial cells
Previous studies in our laboratory showed thepresence of anti-endothelial cell autoantibodiesin dengue patient sera and that the levels ofanti-endothelial cell autoantibodies in DHF/DSSpatient sera were higher than those in DFpatient sera.[11,12] Further studies demonstratedthat anti-NS1 antibodies accounted for thecross-reactivity and the induction of endothelialcell death. After DENV infection, theexpression of NS1 protein was detected inpatient plasma and on cell surface.[17-19] In thisexperiment, HUVECs were infected withDENV-2 at MOIs of 1 and 10 for 12, 24, and48 hours, and the binding ability of anti-NS1antibodies was analysed using flow cytometry.We found greater anti-NS1 time-dependentendothelial cell binding activity in DENV-infected cells than in mock-infected cells(Figure 1).
Effect of anti-NS1 in DENV-inducedendothelial cell apoptosis
Our previous studies[11-14] showed that anti-NS1antibodies induced endothelial cell apoptosis.Another study[15] showed that DENV infectioncaused endothelial cell cytotoxicity. In thisexperiment, we further investigated the effect
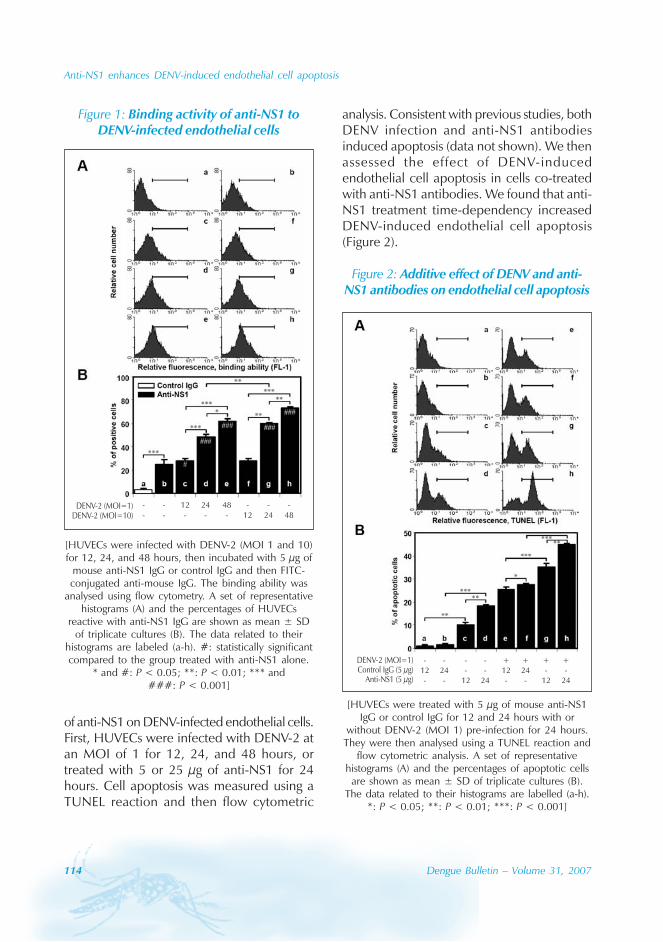
114 Dengue Bulletin – Volume 31, 2007
Anti-NS1 enhances DENV-induced endothelial cell apoptosis
of anti-NS1 on DENV-infected endothelial cells.First, HUVECs were infected with DENV-2 atan MOI of 1 for 12, 24, and 48 hours, ortreated with 5 or 25 μg of anti-NS1 for 24hours. Cell apoptosis was measured using aTUNEL reaction and then flow cytometric
Figure 1: Binding activity of anti-NS1 toDENV-infected endothelial cells
DENV-2 (MOI=1)DENV-2 (MOI=10)
- - 12 24 48 - - -- - - - - 12 24 48
[HUVECs were infected with DENV-2 (MOI 1 and 10)for 12, 24, and 48 hours, then incubated with 5 μg of
mouse anti-NS1 IgG or control IgG and then FITC-conjugated anti-mouse IgG. The binding ability was
analysed using flow cytometry. A set of representativehistograms (A) and the percentages of HUVECs
reactive with anti-NS1 IgG are shown as mean ± SDof triplicate cultures (B). The data related to their
histograms are labeled (a-h). #: statistically significantcompared to the group treated with anti-NS1 alone.
* and #: P < 0.05; **: P < 0.01; *** and###: P < 0.001]
analysis. Consistent with previous studies, bothDENV infection and anti-NS1 antibodiesinduced apoptosis (data not shown). We thenassessed the effect of DENV-inducedendothelial cell apoptosis in cells co-treatedwith anti-NS1 antibodies. We found that anti-NS1 treatment time-dependency increasedDENV-induced endothelial cell apoptosis(Figure 2).
DENV-2 (MOI=1)μμ
Control IgG (5 g)Anti-NS1 (5 g)
- - - - + + + +12 24 - - 12 24 - -- - 12 24 - - 12 24
Figure 2: Additive effect of DENV and anti-NS1 antibodies on endothelial cell apoptosis
[HUVECs were treated with 5 μg of mouse anti-NS1IgG or control IgG for 12 and 24 hours with or
without DENV-2 (MOI 1) pre-infection for 24 hours.They were then analysed using a TUNEL reaction and
flow cytometric analysis. A set of representativehistograms (A) and the percentages of apoptotic cells
are shown as mean ± SD of triplicate cultures (B).The data related to their histograms are labelled (a-h).
*: P < 0.05; **: P < 0.01; ***: P < 0.001]
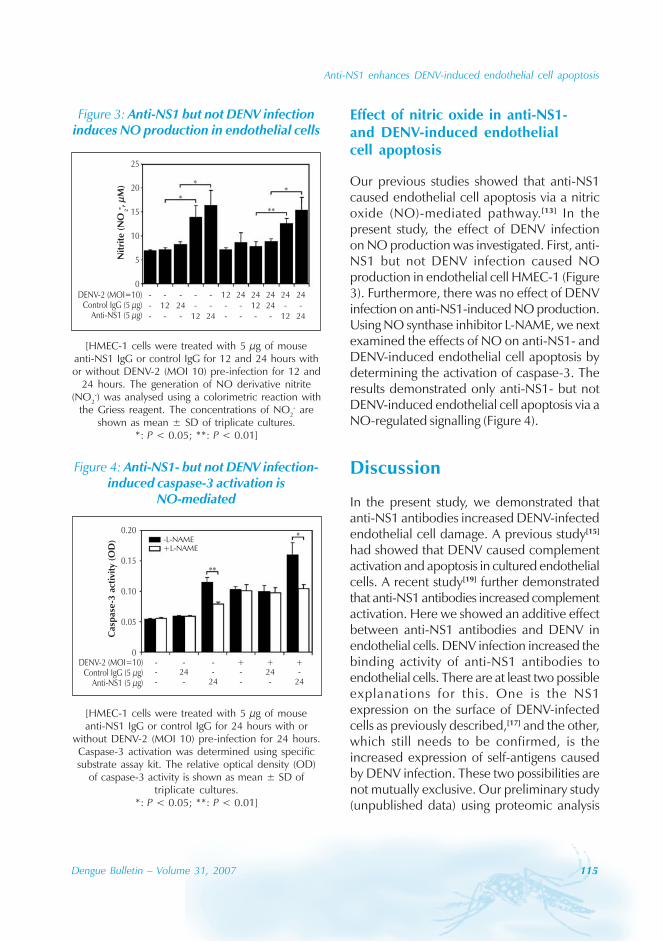
Anti-NS1 enhances DENV-induced endothelial cell apoptosis
Dengue Bulletin – Volume 31, 2007 115
Effect of nitric oxide in anti-NS1-and DENV-induced endothelialcell apoptosis
Our previous studies showed that anti-NS1caused endothelial cell apoptosis via a nitricoxide (NO)-mediated pathway.[13] In thepresent study, the effect of DENV infectionon NO production was investigated. First, anti-NS1 but not DENV infection caused NOproduction in endothelial cell HMEC-1 (Figure3). Furthermore, there was no effect of DENVinfection on anti-NS1-induced NO production.Using NO synthase inhibitor L-NAME, we nextexamined the effects of NO on anti-NS1- andDENV-induced endothelial cell apoptosis bydetermining the activation of caspase-3. Theresults demonstrated only anti-NS1- but notDENV-induced endothelial cell apoptosis via aNO-regulated signalling (Figure 4).
Discussion
In the present study, we demonstrated thatanti-NS1 antibodies increased DENV-infectedendothelial cell damage. A previous study[15]
had showed that DENV caused complementactivation and apoptosis in cultured endothelialcells. A recent study[19] further demonstratedthat anti-NS1 antibodies increased complementactivation. Here we showed an additive effectbetween anti-NS1 antibodies and DENV inendothelial cells. DENV infection increased thebinding activity of anti-NS1 antibodies toendothelial cells. There are at least two possibleexplanations for this. One is the NS1expression on the surface of DENV-infectedcells as previously described,[17] and the other,which still needs to be confirmed, is theincreased expression of self-antigens causedby DENV infection. These two possibilities arenot mutually exclusive. Our preliminary study(unpublished data) using proteomic analysis
Figure 3: Anti-NS1 but not DENV infectioninduces NO production in endothelial cells
DENV-2 (MOI=10)μμ
Control IgG (5 g)Anti-NS1 (5 g)
- - - - - 12 24 24 24 24 24- 12 24 - - - - 12 24 - -- - - 12 24 - - - - 12 24
Nit
rite
(NO
- ,μ
M)
2
25
20
15
10
5
0
[HMEC-1 cells were treated with 5 μg of mouseanti-NS1 IgG or control IgG for 12 and 24 hours withor without DENV-2 (MOI 10) pre-infection for 12 and
24 hours. The generation of NO derivative nitrite(NO2
-) was analysed using a colorimetric reaction withthe Griess reagent. The concentrations of NO2
- areshown as mean ± SD of triplicate cultures.
*: P < 0.05; **: P < 0.01]
Figure 4: Anti-NS1- but not DENV infection-induced caspase-3 activation is
NO-mediated
DENV-2 (MOI=10)μμ
Control IgG (5 g)Anti-NS1 (5 g)
- - - + + +- 24 - - 24 -- - 24 - - 24
Cas
pas
e-3
acti
vity
( OD
)
0.20
0.15
0.10
0.05
0
-L-NAME+L-NAME
[HMEC-1 cells were treated with 5 μg of mouseanti-NS1 IgG or control IgG for 24 hours with or
without DENV-2 (MOI 10) pre-infection for 24 hours.Caspase-3 activation was determined using specificsubstrate assay kit. The relative optical density (OD)
of caspase-3 activity is shown as mean ± SD oftriplicate cultures.
*: P < 0.05; **: P < 0.01]

116 Dengue Bulletin – Volume 31, 2007
Anti-NS1 enhances DENV-induced endothelial cell apoptosis
identified several candidate autoantigenspresent on the surface of endothelial cells. Weare currently investigating whether DENVinfection may upregulate the expression ofthese candidate autoantigens.
Although anti-NS1 binding activity wasincreased after DENV infection, furthermechanistic studies indicated that the causesof endothelial cell apoptosis by anti-NS1antibodies and DENV infection were distinct.Anti-NS1 treatment without DENV infectioncaused cell apoptosis through a NO-regulatedpathway. DENV infection-induced endothelialcell apoptosis was, however, not mediated byNO. There are at least two possibilities for thedistinct mechanisms of apoptotic inductionbetween anti-NS1 and DENV infection. Oneis that DENV infection may induce apoptosisvia a NO-independent manner as evidencedin Figure 4. The other, which remains to beinvestigated, is that GPI-linked NS1 protein[17]
on the surface of DENV-infected cells does notinvolve a NO-mediated pathway by anti-NS1stimulation. Because anti-NS1 but not DENVcauses a NO-mediated apoptotic pathway, itis likely that DENV-increased autoantigen
expression is not involved in the additive effectof endothelial cell apoptosis after anti-NS1binding. This yet needs to be confirmed.
In this study, the induction of endothelialcell cytotoxicity was demonstrated. We previouslyshowed that anti-NS1 antibodies causedinflammatory activation that was mediated bythe production of cytokines, chemokines andadhesion molecules.[20,21] A previous study[15]
also showed that DENV infection led tochemokine production in endothelial cells.Whether the combination of DENV and anti-NS1 antibodies may increase the inflammatoryresponse requires further investigation.
Acknowledgements
The authors thank Bill Franke for editorialassistance. This work was supported by grant(NSC94-3112-B-006-007) from the NationalResearch Program of Genomic Medicine,National Science Council, Taiwan. Dr Chiou-Feng Lin was a post-doctoral fellow supportedby a grant (PD9403) from the National HealthResearch Institutes, Taiwan.
References
[1] Gubler DJ. Dengue and dengue hemorrhagicfever. Clin Microbiol Rev. 1998 Jul; 11(3): 480-96.
[2] Rothman AL. Dengue: defining protectiveversus pathologic immunity. J Clin Invest. 2004Apr; 113(7): 946-51.
[3] Halstead SB. Pathogenesis of dengue:challenges to molecular biology. Science. 1988Jan 29; 239(4839): 476-81.
[4] Halstead SB. Neutralization and antibody-dependent enhancement of dengue viruses.Adv Virus Res. 2003; 60: 421-67.
[5] Rothman AL, Ennis FA. Immunopathogenesisof dengue hemorrhagic fever. Virology. 1999Apr 25; 257(1): 1-6.
[6] Lei HY, Yeh TM, Liu HS, Lin YS, Chen SH, LiuCC. Immunopathogenesis of dengue virusinfection. J Biomed Sci. 2001 Sep; 8(5): 377-88.
[7] Huang KJ, Yang YC, Lin YS, Huang JH, Liu HS,Yeh TM, Chen SH, Liu CC, Lei HY. The dual-specific binding of dengue virus and targetcells for the antibody-dependentenhancement of dengue virus infection. JImmunol. 2006 Mar 1; 176(5): 2825-32.
[8] Diamond MS, Edgil D, Roberts TG, Lu B, HarrisE. Infection of human cells by dengue virus ismodulated by different cell types and viralstrains. J Virol. 2000 Sep; 74(17): 7814-23.

Anti-NS1 enhances DENV-induced endothelial cell apoptosis
Dengue Bulletin – Volume 31, 2007 117
[9] Leitmeyer KC, Vaughn DW, Watts DM, SalasR, Villalobos I, de Chacon, Ramos C, Rico-Hesse R. Dengue virus structural differencesthat correlate with pathogenesis. J Virol. 1999Jun; 73(6): 4738-47.
[10] Vaughn DW, Green S, Kalayanarooj S, InnisBL, Nimmannitya S, Suntayakorn S, Endy TP,Raengsakulrach B, Rothman AL, Ennis FA,Nisalak A. Dengue viremia titer, antibodyresponse pattern, and virus serotype correlatewith disease severity. J Infect Dis. 2000 Jan;181(1): 2-9.
[11] Lin CF, Lei HY, Shiau AL, Liu CC, Liu HS, YehTM, Chen SH, Lin YS. Antibodies from denguepatient sera cross-react with endothelial cellsand induce damage. J Med Virol. 2003 Jan;69(1): 82-90.
[12] Lin CF, Lei HY, Liu CC, Liu HS, Yeh TM, ChenSH, Lin YS. Autoimmunity in dengue virusinfection. Dengue Bulletin. 2004; 28:51-57.
[13] Lin CF, Lei HY, Shiau AL, Liu HS, Yeh TM, ChenSH, Liu CC, Chiu SC, Lin YS. Endothelial cellapoptosis induced by antibodies againstdengue virus nonstructural protein 1 viaproduction of nitric oxide. J Immunol. 2002Jul 15; 169(2): 657-64.
[14] Lin YS, Lin CF, Lei HY, Liu HS, Yeh TM, ChenSH, Liu CC. Antibody-mediated endothelialcell damage via nitric oxide. Curr Pharm Des.2004; 10(2): 213-21.
[15] Avirutnan P, Malasit P, Seliger B, Bhakdi S,Husmann M. Dengue virus infection of humanendothelial cells leads to chemokineproduction, complement activation, andapoptosis. J Immunol. 1998 Dec 1; 161(11):6338-46.
[16] Falconar AK. The dengue virus nonstructural-1 protein (NS1) generates antibodies tocommon epitopes on human blood clotting,
integrin/adhesin proteins and binds to humanendothelial cells: potential implications inhaemorrhagic fever pathogenesis. Arch Virol.1997; 142(5): 897-916.
[17] Jacobs MG, Robinson PJ, Bletchly C,Mackenzie JM, Young PR. Dengue virusnonstructural protein 1 is expressed in aglycosyl-phosphatidylinositol-linked form thatis capable of signal transduction. FASEB J. 2000Aug; 14(11): 1603-10.
[18] Libraty DH, Young PR, Pickering D, Endy TP,Kalayanarooj S, Green S, Vaughn DW, NisalakA, Ennis FA, Rothman AL. High circulatinglevels of the dengue virus nonstructural proteinNS1 early in dengue illness correlate with thedevelopment of dengue hemorrhagic fever. JInfect Dis. 2002 Oct 15; 186(8): 1165-8.
[19] Avirutnan P, Punyadee N, Noisakran S,Komoltri C, Thiemmeca S, AuethavornananK, Jairungsri A, Kanlaya R, TangthawornchaikulN, Puttikhunt C, Pattanakitsakul SN,Yenchitsomanus PT, Mongkolsapaya J,Kasinrerk W, Sittisombut N, Husmann M,Blettner M, Vasanawathana S, Bhakdi S,Malasit P. Vascular leakage in severe denguevirus infections: a potential role for thenonstructural viral protein NS1 andcomplement. J Infect Dis. 2006 Apr 15; 193(8):1078-88.
[20] Lin CF, Chiu SC, Hsiao YL, Wan SW, Lei HY,Shiau AL, Liu HS, Yeh TM, Chen SH, Liu CC,Lin YS. Expression of cytokine, chemokine,and adhesion molecules during endothelialcell activation induced by antibodies againstdengue virus nonstructural protein 1. JImmunol. 2005 Jan 1; 174(1): 395-403.
[21] Lin CF, Wan SW, Cheng HJ, Lei HY, Lin YS.Autoimmune pathogenesis in dengue virusinfection. Viral Immunol. 2006; 19(2): 127-32.

118 Dengue Bulletin – Volume 31, 2007
Role of nitric oxide in the pathogenesis of dengue
Germán Añeza , Nereida Valeroa and Jesús Mosquerab
aSección de Virología, Instituto de Investigaciones Clínicas “Dr. Américo Negrette”, Facultad de Medicina,Universidad del Zulia, Apartado Postal 23, Maracaibo 4001-A, Estado Zulia, Venezuela
bSección de Inmunología y Biología Celular, Instituto de Investigaciones Clínicas “Dr. Américo Negrette”,Facultad de Medicina, Universidad del Zulia, Apartado Postal 23, Maracaibo 4001-A, Estado Zulia, Venezuela
Abstract
The free radical nitric oxide (NO) has emerged in recent years as a fundamental signalling molecule forthe maintenance of homeostasis, as well as a potent cytotoxic effector involved in the pathogenesis ofa wide range of human diseases. The presence of NO during dengue infection as well as its experimentalantiviral and apoptotic effects have been documented. In this regard, increased serum NO levels indengue fever and basal levels in the haemorrhagic form of dengue have been reported. Clinical andexperimental data suggest that NO could act as a beneficial factor during dengue infection by itsantiviral and apoptotic effects; however, more intense investigations are required.
Keywords: Dengue virus; Nitric oxide; Dengue haemorrhagic fever, Pathogenesis.
E-mail: [email protected]; Tel.: +58 416 6658687; Fax: +58 261 7597247
Introduction
Dengue fever (DF) is the most importantarboviral disease affecting public health in thedeveloping world. The disease is endemic inmany countries and causes epidemicsfrequently. These epidemics involve anestimated 100 million cases annuallyworldwide, causing great health, social andeconomic burden.[1,2] Dengue virus (DENV), aRNA virus belonging to genus, Flavivirus, familyFlaviviridae, has four serotypes (DENV-1 to 4).Clinically, the disease may be asymptomaticor may range from a mild febrile illness, DF, tothe severe, life-threatening form, denguehaemorrhagic fever/dengue shock syndrome(DHF/DSS), which can be caused indistinctlyby any serotype.[3-5]
The discovery that mammalian cells havethe ability to synthesize the free radical nitricoxide (NO) has stimulated an extraordinaryimpetus for scientific research in all fields ofbiology and medicine. NO is produced bythree isoforms of nitric oxide synthase (NOS),the inducible form (iNOS), and the twoconstitutive isoforms endothelial NOS (eNOS)and neuronal NOS (nNOS). Since its earlydescription as an endothelial-derived relaxingfactor, NO has emerged as a fundamentalsignalling device regulating virtually every criticalcellular function, as well as a potent mediatorof cellular damage in a wide range of conditions.Recent evidence indicates that most of thecytotoxicity attributed to NO is rather due toperoxynitrite, produced from the diffusion-controlled reaction between NO and another

Role of nitric oxide in the pathogenesis of dengue
Dengue Bulletin – Volume 31, 2007 119
free radical, the superoxide anion. Peroxynitriteinteracts with lipids, DNA, and proteins viadirect oxidative reactions or via indirect, radical-mediated mechanisms. These reactions triggercellular responses ranging from subtlemodulations of cell signalling to overwhelmingoxidative injury, committing cells to necrosisor apoptosis. In vivo, peroxynitrite generationrepresents a crucial pathogenic mechanism inconditions such as stroke, myocardial infarction,chronic heart failure, diabetes, circulatoryshock, chronic inflammatory diseases, cancer,neurodegenerative disorders and viralinfections.[6-9]
Previous reports have suggested a possiblerole of NO in DF and DHF/DSSpathogenesis; [10-17] however, very littleinformation is available with regard to its roleduring the human dengue infection. Therefore,evidence from in vivo and in vitro conditionssuggesting the possible role of NO in dengueinfection is presented in this review.
Nitric oxide in dengueinfection
NO is ubiquitous physiological-free radical thatis responsible for many pathological disorders.[6-
9] Its presence in human dengue infection wasreported for the first time by Valero andcollaborators.[10]
In this regard, increased serum NO levelsin DF and normal levels in DHF patients werereported. There was no relationship betweenthe viral serotypes and the increased values ofNO. Interaction of dengue virus with primaryhuman platelet cultures did not induce furtherregulation of NO production.[10] These datasuggest that increased levels of NO in DFpatients could have a protector effect duringthe disease, and, on the contrary, decreasedlevels could be the cause of the deleterious
effects observed in DHF. In addition, it seemsto be that virus-platelets interaction is notinvolved in the increased production of NO,suggesting other sources for this molecule. Inthis regard, monocyte/macrophages are themost important targets of DENV,[18] and arecapable of producing high amounts of NO;thus, Mo/MΦ-DENV interaction may induceincreased amount of NO. In accordance withthat suggestion, increased expression ofinducible nitric oxide synthase in monocytesfrom DF patients has been reported,[13] whichtranslates into sustained NO production,[13,15,16]
suggesting the possible source of NO found inpatients with DF.
Since endothelium is an important NOproducer, the direct or indirect interaction ofDENV with the endothelial cells could inducean increased amount of serum NO in thesepatients.[6-8,16,17] However, increased levels ofNO in patients with dengue seem to becontroversial. Other investigators[19] havereported levels of serum NO significantly lowerthan those of normal controls in Asiatic childrenwith DF and DHF, suggesting that theendothelial damage renders the endotheliumincapable of producing NO. These data aredifferent from the increased amount of serumNO found in patients with DF reported byothers.[10] This different response to the virusinfection could be related to the widespreadprevalence of human dengue resistance genesin the Americas’ populations compared to Asianpopulations. The people of countries in South-East Asia have high rates of severe denguedisease compared to consistently low case-fatality rates in American countries reportingDHF.[20] Virtually, nothing is known aboutdengue resistance gene(s) in the pathogenesisof dengue. Are the manifestations of DFsuppressed by this gene(s), or does the gene(s)selectively dampen dengue vasculopathy?[20]
The expression of this gene(s) could involvedifferent cell targets during the disease leading

120 Dengue Bulletin – Volume 31, 2007
Role of nitric oxide in the pathogenesis of dengue
to differential production of NO. Futureresearch on dengue infections shouldemphasize population-based designs to shedlight on the heterogeneity of dengue inpopulations living in the Western and Easternhemispheres.
Anti-viral effect of nitric oxide
The beneficial effect of NO in DF could berelated to an antiviral effect of the molecule.Previous studies have shown that NO couldhave antiviral effect on dengue-infected cells.In this regard, decreased expression of denguevirus antigens in NO-producing mononuclearcells has been reported. In addition, treatmentof those cultures with an iNOS inhibitor (NG-methyl-L-Arginine) induced increased viralantigens detection.[13] The effect of NOexogenous donor (S-nitroso-N-acethylpennicillamine or SNAP) on DENVreplication in neuroblastoma cell cultures hasalso been reported. DENV-2-infected cellstreated with SNAP showed suppression of viralRNA synthesis and, consequently, decreasedviral proteins and viral progeny production.[12]
This effect was also observed in DENV-1-infected C6/36 cell cultures treated with sodiumnitroprussiate, a NO donor.[13] Takhampunyaet al.[14] showed the inhibitory effect of NOon DENV-infected LLC-MK2 cell cultures usingSNAP. This report found a diminished cellularaccumulation of viral RNA during viral RNAsynthesis. This inhibitory effect of NO seemsto be related to the inhibition of DENV-2 RNA-dependent RNA polymerase (RdRp) domainof NS5, with further suppression of viral RNAsynthesis, viral structural proteins, anddecreased number of infectious particles in theculture medium.
Recently, Ubol et al.[21] reported DENV-2clinical isolates derived from DF and DHF cases,which are NO-sensitive and NO-resistant,
respectively, to endogenous NO produced byTHP-1 cells (human monocytic leukemia cellline). These findings were also related withamino acids changes in the RdRp andMethyltransferase (MTase) domains of NS5.Additionally, when compared the geneexpression on DENV-2 infected THP-1 cells,NO-resistant but no NO-sensitive isolates,induced the immune-related genes IL-6, IL-7,IL-8, RANTES, and MCP-3, all of which arepotent mediators of tissue damage as well ascoagulopathy. These findings need to becarefully studied to determine if this effect isparticularly restricted to DENV-2 or to its Asiangenotype, or if NO-resistant strains occurs inthe nature for the other serotypes.
All the previous data suggest a possibleantiviral mechanism of NO in vivo, and suggesta beneficial role of increased serum levels ofNO reported in DF.[10] Lower levels of NOfound in patients with DHF could be relatedto the described DENV action on target cells.In this regard, Chareonsirisuthigul et al.[11]
showed a different antiviral response inexperiments using DENV-infected THP-1 cellcultures untreated and treated with an anti-DENV-enhancing antibodies (a model ofinfection via ADE). These experiments showedsuppression of NO production in ADE culturesdue to disruption of transcription of the IRF-1(an iNOS gene transcription factor) and blockingof the activation of STAT-1. These findings couldbe related to the pathogenesis of DHF/DSSand could suggest a mechanism for basal serumNO levels in DHF patients.[10]
This antiviral effect of NO is not restrictedto the dengue virus. Several studies havereported a NO antiviral effect on an extensivelist of viruses including SARS-coronavirus,hantavirus, Ross River virus, vesicular stomatitisvirus, Crimean-Congo haemorrhagic fever virus,and members of the genus Flavivirus, includingJapanese encephalitis.[22-29]

Role of nitric oxide in the pathogenesis of dengue
Dengue Bulletin – Volume 31, 2007 121
Nitric oxide and apoptosis
The dengue virus may be an apoptosis inducerby direct or indirect mechanisms. Infection ofMo/MΦ or endothelial cells with DENV resultsin apoptosis;[30-32] however, the mechanismsof apoptosis induction remain unclear. The freeradical NO has emerged in recent years as afundamental signalling molecule for themaintenance of homeostasis, as well as apotent cytotoxic effector. Under normalconditions, NO produced in low concentrationacts as a messenger and cytoprotective(antioxidant) factor; however, when thecircumstances allow the formation of substantialamounts of NO, this molecule could induceboth oxidative and nitrosative stresses, whichform the basis of the apoptosis generallyattributed to NO.[7-9]
Previous investigations have shown thepresence of apoptosis in patients both withdengue and in experimental dengueinfection. [30-34] However, there is littleinformation about the role of NO in apoptosisduring dengue infection. In this regard,antibodies against dengue virus nonstructuralprotein 1 (NS1), which has cross-reactivity withendothelial cells (AECA), induce apoptosis viaa NO-mediated mechanism in experimentaldengue virus infection. Endothelial cellsundergo apoptosis via the mitochondria-dependent pathway that is regulated by NOproduction, suggesting that NO-regulatedendothelial cell injury may play a role in thedisruption of vessel endothelium and contributeto the pathogenesis of vasculopathy.[16,17] Otherinvestigators have shown increased apoptosisin dengue virus-replicating Kupffer cellsassociated with increased expression ofinducible NO synthase and production ofNO.[15] Apoptosis could avoid the release ofviral particles and, together with the
phagocytosis and digestion of apoptotic cells,represent mechanisms to prevent viralprogeny.[32] However, damage of endothelialcells and monocytes/macrophages couldinduce severe forms of dengue infection.Further investigation is required to determinethe modulation of NO in order to provide thetherapeutic strategies for dengue infection bypreventing the AECA-mediated endothelial cellapoptosis and to determine the in vivomechanism of NO-mediated apoptosisinduced by DENV during the course of humandengue infection.
Conclusion and futureperspective
NO, on the basis of its physiological chemistry,provides a conceptual framework, which helpsto distinguish between the beneficial and toxicconsequences of NO, and to envision potentialtherapeutic strategies for the future. The highlevels of NO may be beneficial during dengueinfection by its antiviral and apoptotic effects;however it could also induce severe forms ofthe disease by damage to endothelial cells.Further investigations are required to determinethe source of NO during human dengueinfection, its role in the pathogenesis of DFand DHF/DSS, its role in dengue infectionaccording to the prevalence of human dengueresistance genes, and the presence of NO-resistant strains in all the DENV serotypes.
Acknowledgments
This work was supported by the Consejo parael Desarrollo Científico y Humanístico de laUniversidad del Zulia (CONDES) (CC-0789-04).

122 Dengue Bulletin – Volume 31, 2007
Role of nitric oxide in the pathogenesis of dengue
References
[1] Añez G, Balza R, Valero N, Larreal Y. Economicimpact of dengue and dengue hemorrhagicfever in the State of Zulia, Venezuela, 1997-2003. Rev Panam Salud Publica. 2006 May;19(5): 314-20. Spanish.
[2] Gubler DJ. Epidemic dengue/denguehemorrhagic fever as a public health, socialand economic problem in the 21st century.Trends Microbiol. 2002 Feb; 10(2): 100-3.
[3] World Health Organization. Denguehaemorrhagic fever: diagnosis, treatment andcontrol. 2nd edn. Geneva: WHO; 1997.
[4] Halstead SB. Pathogenesis of dengue:challenges to molecular biology. Science. 1988Jan 29; 239(4839): 476-81.
[5] Halstead SB. Observations related topathogensis of dengue hemorrhagic fever. VI.Hypotheses and discussion. Yale J Biol Med.1970 Apr; 42(5): 350-62.
[6] Moncada S, Higgs EA. The discovery of nitricoxide and its role in vascular biology. Br JPharmacol. 2006 Jan; 147 Suppl 1: S193-201.
[7] Liaudet L, Soriano FG, Szabó C. Biology ofnitric oxide signaling. Crit Care Med. 2000Apr; 28(4 Suppl): N37-52.
[8] Pacher P, Beckman JS, Liaudet L. Nitric oxideand peroxynitrite in health and disease. PhysiolRev. 2007 Jan; 87(1): 315-424.
[9] Nagy G, Clark JM, Buzás EI, Gorman CL, CopeAP. Nitric oxide, chronic inflammation andautoimmunity. Immunol Lett. 2007 Jul 31;111(1): 1-5.
[10] Valero N, Espina LM, Añez G, Torres E,Mosquera JA. Short report: increased level ofserum nitric oxide in patients with dengue.Am J Trop Med Hyg. 2002 Jun; 66(6): 762-4.
[11] Chareonsirisuthigul T, Kalayanarooj S, UbolS. Dengue virus (DENV) antibody-dependentenhancement of infection upregulates theproduction of anti-inflammatory cytokines,
but suppresses anti-DENV free radical and pro-inflammatory cytokine production, in THP-1cells. J Gen Virol. 2007 Feb; 88(Pt 2): 365-75.
[12] Charnsilpa W, Takhampunya R, Endy TP,Mammen MP Jr, Libraty DH, Ubol S. Nitricoxide radical suppresses replication of wild-type dengue 2 viruses in vitro. J Med Virol.2005 Sep; 77(1): 89-95.
[13] Neves-Souza PC, Azeredo EL, Zagne SM, Valls-de-Souza R, Reis SR, Cerqueira DI, NogueiraRM, Kubelka CF. Inducible nitric oxidesynthase (iNOS) expression in monocytesduring acute dengue Fever in patients andduring in vitro infection. BMC Infect Dis. 2005Aug 18; 5:64.
[14] Takhampunya R, Padmanabhan R, Ubol S.Antiviral action of nitric oxide on dengue virustype 2 replication. J Gen Virol. 2006 Oct; 87(Pt10): 3003-11.
[15] Marianneau P, Steffan AM, Royer C, DrouetMT, Jaeck D, Kirn A, Deubel V. Infection ofprimary cultures of human Kupffer cells bydengue virus: no viral progeny synthesis, butcytokine production is evident. J Virol. 1999Jun; 73(6): 5201-6.
[16] Lin CF, Lei HY, Shiau AL, Liu HS, Yeh TM, ChenSH, Liu CC, Chiu SC, Lin YS. Endothelial cellapoptosis induced by antibodies againstdengue virus nonstructural protein 1 viaproduction of nitric oxide. J Immunol. 2002Jul 15;169(2):657-64.
[17] Lin YS, Lin CF, Lei HY, Liu HS, Yeh TM, ChenSH, Liu CC. Antibody-mediated endothelialcell damage via nitric oxide. Curr Pharm Des.2004; 10(2): 213-21.
[18] Blackley S, Kou Z, Chen H, Quinn M, RoseRC, Schlesinger JJ, Coppage M, Jin X. Primaryhuman splenic macrophages, but not T or Bcells, are the principal target cells for denguevirus infection in vitro. J Viro.l 2007 Dec;81(24): 13325-34.

Role of nitric oxide in the pathogenesis of dengue
Dengue Bulletin – Volume 31, 2007 123
[19] Trairatvorakul P, Chongsrisawat V,Ngamvasinont D, Asawarachun D, NantasookJ, Poovorawan Y. Serum nitric oxide inchildren with dengue infection. Asian Pac JAllergy Immunol. 2005 Jun-Sep; 23(2-3): 115-9.
[20] Halstead SB. Dengue in the Americas andSoutheast Asia: do they differ? Rev PanamSalud Publica. 2006 Dec; 20(6): 407-15.
[21] Ubol S, Chareonsirisuthigul T, Kasisith J,Klungthong C. Clinical isolates of dengue viruswith distinctive susceptibility to nitric oxideradical induce differential gene responses inTHP-1 cells. Virology. 2008 Jul 5; 376(2): 290-6.
[22] Keyaerts E, Vijgen L, Chen L, Maes P,Hedenstierna G, Van Ranst M. Inhibition ofSARS-coronavirus infection in vitro by S-nitroso-N-acetylpenicillamine, a nitric oxidedonor compound. Int J Infect Dis. 2004 Jul;8(4): 223-6.
[23] Klingström J, Akerström S, Hardestam J, StoltzM, Simon M, Falk KI, Mirazimi A, RottenbergM, Lundkvist A. Nitric oxide and peroxynitritehave different antiviral effects against hantavirusreplication and free mature virions. Eur JImmunol. 2006 Oct; 36(10): 2649-57.
[24] Lidbury BA, Mahalingam S. Specific ablationof antiviral gene expression in macrophagesby antibody-dependent enhancement of RossRiver virus infection. J Virol. 2000 Sep; 74(18):8376-81.
[25] Chesler DA, Dodard C, Lee GY, Levy DE, ReissCS. Interferon-gamma-induced inhibition ofneuronal vesicular stomatitis virus infection isSTAT1 dependent. J Neurovirol. 2004 Feb;10(1): 57-63.
[26] Bi Z, Reiss CS. Inhibition of vesicular stomatitisvirus infection by nitric oxide. J Virol. 1995Apr; 69(4): 2208-13.
[27] Simon M, Falk KI, Lundkvist A, Mirazimi A.Exogenous nitric oxide inhibits Crimean
Congo hemorrhagic fever virus. Virus Res. 2006Sep; 120(1-2): 184-90.
[28] Saxena SK, Singh A, Mathur A. Antiviral effectof nitric oxide during Japanese encephalitisvirus infection. Int J Exp Pathol. 2000 Apr; 81(2):165-72.
[29] Lin YL, Huang YL, Ma SH, Yeh CT, Chiou SY,Chen LK, Liao CL. Inhibition of Japaneseencephalitis virus infection by nitric oxide:antiviral effect of nitric oxide on RNA virusreplication. J Virol. 1997 Jul; 71(7): 5227-35.
[30] Espina LM, Valero NJ, Hernández JM,Mosquera JA. Increased apoptosis andexpression of tumor necrosis factor-alphacaused by infection of cultured humanmonocytes with dengue virus. Am J Trop MedHyg. 2003 Jan; 68(1): 48-53.
[31] Avirutnan P, Malasit P, Seliger B, Bhakdi S,Husmann M. Dengue virus infection of humanendothelial cells leads to chemokineproduction, complement activation, andapoptosis. J Immunol. 1998 Dec 1; 161(11):6338-46.
[32] Mosquera JA, Hernandez JP, Valero N, EspinaLM, Añez GJ. Ultrastructural studies on denguevirus type 2 infection of cultured humanmonocytes. Virol J. 2005 Mar 31; 2: 26.
[33] Jan JT, Chen BH, Ma SH, Liu CI, Tsai HP, WuHC, Jiang SY, Yang KD, Shaio MF. Potentialdengue virus-triggered apoptotic pathway inhuman neuroblastoma cells: arachidonicacid, superoxide anion, and NF-kappaB aresequentially involved. J Virol. 2000 Sep;74(18): 8680-91.
[34] Myint KS, Endy TP, Mongkolsirichaikul D,Manomuth C, Kalayanarooj S, Vaughn DW,Nisalak A, Green S, Rothman AL, Ennis FA,Libraty DH. Cellular immune activation inchildren with acute dengue virus infections ismodulated by apoptosis. J Infect Dis. 2006Sep 1; 194(5): 600-7.

124 Dengue Bulletin – Volume 31, 2007
Comparative oviposition preferences ofAedes (Stegomyia) aegypti (L.) to water fromstorm water drains and seasoned tap water
C.D. Chena , W.A. Naznia, B. Seleenaa, J.Y. Moob, M. Azizahc and H.L. Leea
aMedical Entomology Unit, Institute for Medical Research, Jalan Pahang, 50588 Kuala Lumpur, Malaysia
bCollege of Tunku Abdul Rahman, Jalan Genting Kelang, Setapak, 53100 Kuala Lumpur, Malaysia
cBacteriology Unit, Institute for Medical Research, Jalan Pahang, 50588 Kuala Lumpur, Malaysia
Abstract
The comparative oviposition preferences of Aedes aegypti to water from storm-water drains* and seasonedtap water were evaluated in the laboratory. The sample was collected from concrete storm-water drainswith stagnant clear water in a dengue-endemic site, Taman Samudera, Selangor. Ae. aegypti adults weregiven a blood meal and released into the cage. Gravid females were given a choice between drain waterand seasoned tap water for egg deposition. In a no-choice test, there was no significant difference in thenumbers of eggs, larvae, pupae and adults colonized from the drain water and seasoned tap water(p>0.05), indicating that Ae. aegypti oviposit their eggs on a substrate which is readily available. In achoice test, the number of eggs laid by Ae. aegypti in drain water (1630.67 ± 204.26) was significantlymore than that in seasoned tap water (221.33 ± 53.18) (p<0.05). The number of eggs was 6-fold higherin the drain water compared to seasoned tap water. The oviposition activity index was 0.71, indicatingthat the drain water was more attractive compared to seasoned tap water as an oviposition substrate. ThepH and biochemical oxygen demand (BOD) values of both drain water and seasoned tap water were notsignificantly different (p>0.05), indicating that water from the drain did not contain high organic content.Significant water conductivity (p<0.05) and the presence of bacteria could have contributed to the siteselection for egg laying by Ae. aegypti. The drain water successfully supported the colonization of theimmatures, with the emergence of 824.33 ± 13.96 adult mosquitoes. The ratio of male and femalemosquitoes was 1:1. This study concluded that the concrete drainage system with clear stagnant waterprovides a suitable medium for the colonization of dengue vector Ae. aegypti.
Keywords: Oviposition; Aedes aegypti; Drain water, Seasoned tap water; Malaysia.
E-mail: [email protected]*Water from storm-water drains will be mentioned as ‘drain water’ throughout the text.
Introduction
During the 19th century, dengue was considereda sporadic disease, causing epidemics at longintervals. However, dramatic changes in thispattern have occurred and currently, dengue
ranks as the most important mosquito-borneviral disease in the world. In the past 50 years,its incidence has increased 30-fold, with largeoutbreaks occurring in five of the six WorldHealth Organization (WHO) regions. Atpresent, dengue is endemic in 112 countriesin the world.[1]

Comparative oviposition preferences of Aedes (Stegomyia) aegypti (L.) to water from storm water drains andseasoned tap water
Dengue Bulletin – Volume 31, 2007 125
Dengue has remained endemic in Malaysiasince the first case was documented in 1901–1902.[2] The disease was made notifiable in1973.[3]
Aedes aegypti is a domestic mosquito whileAedes albopictus is semi-domestic in urban areas.Ae. aegypti breeds exclusively in artificialcontainers such as earthen jars and plasticcontainers which contain relatively clean waterin and around houses.[4] The typical habitats ofAe. albopictus are artificial containers, tree holesand bamboo stumps near human dwellings.[5]
The ability of gravid mosquito females todistinguish among potential oviposition sites thatwill or will not support the growth, developmentand survival of their offspring is critical for themaintenance of the mosquito population. Oncethe blood meal large enough to initiate ovariandevelopment has been obtained and thetemperature-dependent egg maturation periodhas been completed, mosquitoes must locatesuitable oviposition sites.[6]
In Malaysia, rapid urbanization has resultedin the creation of suitable habitats for Ae.aegypti. The present study was conducted toexamine the suitability of drain water for thebreeding of Ae. aegypti mosquitoes.
Materials and Methods
Mosquito colony
Ae. aegypti mosquitoes** (F 967) used in thisstudy came from the colony maintained in theinsectarium of the Medical Entomology Unit,Institute for Medical Research, Kuala Lumpur,Malaysia. The colony, established 30 years ago,produced test mosquitoes that are pathogen-free and homogenous.
Collection of drain water fromstudy site
The drain water was collected from TamanSamudera (N03°13.987’, E101°41.918’),Selangor state, Malaysia, which is a dengue-endemic area, located 16 km from KualaLumpur city centre. Taman Samudera covers 30ha of land which consists of approximately 400double storey terrace brick houses.
The ovitrap surveillance conducted in aprevious study by Chen et al.[7] indicated thatAe. aegypti was the principal mosquito, bothindoors and outdoors, as reflected by ovitrapcollections.
Taman Samudera is an urban residentialsite, generally clean. This site had minimumartificial larval habitats such as containers andtyres. The major larval habitat appeared to bethe concrete drain with clear stagnant water.The drain water was collected from fivedifferent drains in Taman Samudera for thisstudy. All samples of the drain water werepooled together for colonization.
Preparation of seasoned tap water
Seasoned tap water (de-chlorinated water) wasprepared by collecting the tap water directlyfrom pipe into a plastic container and seasonedfor several days.
Measurement of water parameters
The pH, conductivity and biochemical oxygendemand (BOD) of the drain and seasoned tapwaters were determined. The bacterial fauna ofthe waters were determined by plating the watersample on Nutrient Agar. Each bacteria colonywas isolated and identified at the BacteriologyUnit, Institute for Medical Research.**Test insects were from a 30-years-old colony and as
such may not necessarily reflect behaviour patterns thatare comparable to those of field populations – Editor

126 Dengue Bulletin – Volume 31, 2007
Comparative oviposition preferences of Aedes (Stegomyia) aegypti (L.) to water from storm water drains andseasoned tap water
Laboratory evaluation of ovipositionpreference
Preparation of ovitrap
Ovitrap as described by Lee[8] was used in thisstudy. Each ovitrap consisted of a 300 ml plasticcontainer with straight, slightly tapered sides.A filter paper in cone shape was placed in theovitrap to serve as a resting and ovipositionsite. Each ovitrap was filled with drain water orseasoned tap water to a level of 5.5 cm.
No-choice test
The ovitrap described as above was filled withdrain water and placed into a cage (30 cm X 30cm X 30 cm). A total of 30 laboratory-bred 5-days-old Ae. aegypti were given a blood mealby feeding on a guinea pig. The gravid femaleswere released into the cage and left for a week.For all tests, 10% sugar solution was provided.At the end of the week, the ovitrap wasremoved. The filter paper was dried and theeggs were counted. The filter paper was thenplaced into a tray holding drain water for theeggs to hatch. The larvae, pupae and adultemergence were counted and recorded. Threereplicates were conducted. The same procedurewas repeated by using seasoned tap water.
Choice test
Two ovitraps each holding drain water andseasoned tap water were placed in a cage (30cm X 30 cm X 30 cm). The experimentalprocedure was then carried out according tothe no-choice test.
Data analysis
The relative attractiveness of test sample wasexpressed as an oviposition activity index (OAI)calculated according to Kramer and Mulla,[9]
where
Oviposition Activity Index (OAI) =
[NT denotes the number of eggs laid on test water(drain water) and NS denotes the number of eggs laidin control water (seasoned tap water). Index valuesrange from +1 to –1. Positive values indicate thatmore eggs were deposited in the test water than inthe control water, and that the test water wasattractive. On the other hand, if more eggs weredeposited in the control water than in the test water,negative index values will be obtained indicating thatthe test water acted as repellent]
Statistical analysis
T-test analysis (SPSS v.10) was used todetermine the significant difference of thephysical measures and the number of eggs,larvae, pupae and adult obtained betweendrain water and seasoned tap water. All levelsof statistical significance were determined atp<0.05. The hatchability and emergence rateof Ae. aegypti in drain water and seasoned tapwater were also calculated.
Results and discussion
The oviposition preference of Ae. aegypti to drainwater and seasoned tap water from the no-choice test (Table 1) shows that there was nosignificant difference in the numbers of eggs,larvae, pupae and adults colonized from drainwater and seasoned tap water (p>0.05),indicating that Ae. aegypti oviposit their eggs ona substrate which is readily available. However,in the choice test (Table 2), the number of eggslaid by Ae. aegypti in drain water (1630.67 ±204.26) was significantly more than in seasonedtap water (221.33 ± 53.18) (p<0.05). Thenumber of eggs was 6-fold higher in drain watercompared to seasoned tap water. Furthermore,the oviposition activity index (OAI) was 0.71,indicating that drain water was more attractivecompared to seasoned tap water as anoviposition substrate.
NT–NSNT+NS
Comparative oviposition preferences of Aedes (Stegomyia) aegypti (L.) to water from storm water drains andseasoned tap water
Dengue Bulletin – Volume 31, 2007 127
The hatchability and emergence rates of Ae.aegypti in drain water and seasoned tap water(Figure) indicated that there was no completehatchability and emergence in all the preferredwater. The majority of eggs placed in drain waterand seasoned tap water did hatch, with 69.0–82.0% hatchability in drain water and 63.0–74.0% in seasoned tap water. In both choiceand no-choice studies, drain water and seasonedtap water successfully supported the colonizationof Ae. aegypti immatures. In the choice test, atotal of 824.33 ± 13.96 adult mosquitoesemerged from the population that was colonizedin the drain water; and a total of 125.67 ± 44.95adult mosquitoes emerged from the colony inthe seasoned water (Tables 1 and 2).
Figure: Hatchability rate and emergencerate of Ae. aegypti in drain water and
seasoned tap water
76.3381.8481.84
68.70
89.98
73.59
87.15
63.10
0
10
20
30
40
50
60
70
80
90
100
No choice test Choice test No choice test Choice test
Hatchability rate Emergence rate
Drain water Seasonal water
Per
cen
tage
( %)
Table 1: Mean numbers of eggs, larvae, pupae and adult obtained from 30 gravid femaleAe. aegypti in no-choice test
Table 2: Mean numbers of egg, larvae, pupae and adult obtained from 30 gravid femaleAe. aegypti in choice test
p<0.05 = Significant; p>0.05 = Not significant; SE = Standard error
* = p<0.05 (Significant); ** = p<0.01 (Highly significant); SE = Standard error
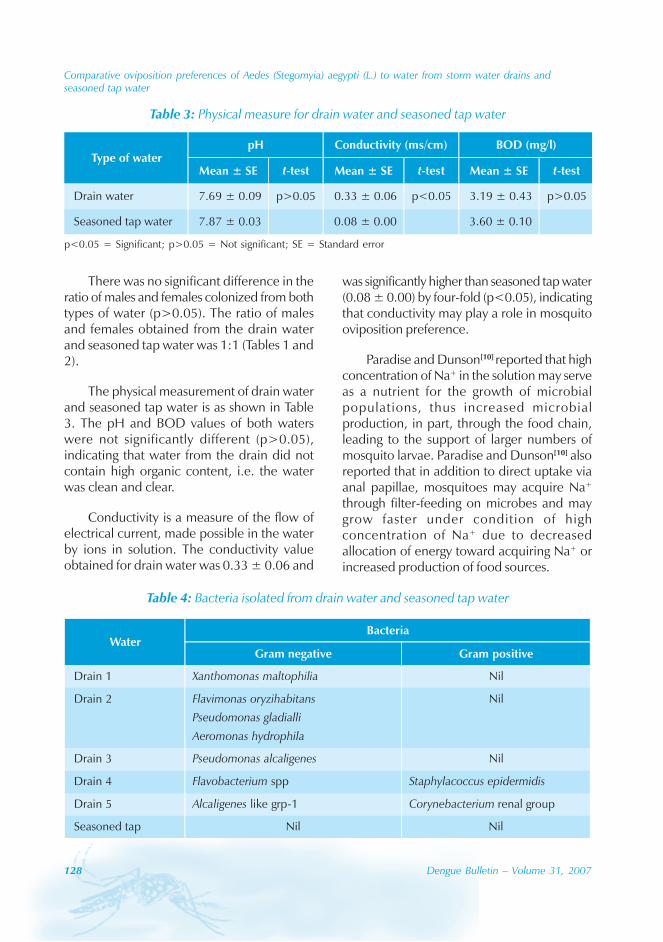
128 Dengue Bulletin – Volume 31, 2007
Comparative oviposition preferences of Aedes (Stegomyia) aegypti (L.) to water from storm water drains andseasoned tap water
There was no significant difference in theratio of males and females colonized from bothtypes of water (p>0.05). The ratio of malesand females obtained from the drain waterand seasoned tap water was 1:1 (Tables 1 and2).
The physical measurement of drain waterand seasoned tap water is as shown in Table3. The pH and BOD values of both waterswere not significantly different (p>0.05),indicating that water from the drain did notcontain high organic content, i.e. the waterwas clean and clear.
Conductivity is a measure of the flow ofelectrical current, made possible in the waterby ions in solution. The conductivity valueobtained for drain water was 0.33 ± 0.06 and
was significantly higher than seasoned tap water(0.08 ± 0.00) by four-fold (p<0.05), indicatingthat conductivity may play a role in mosquitooviposition preference.
Paradise and Dunson[10] reported that highconcentration of Na+ in the solution may serveas a nutrient for the growth of microbialpopulations, thus increased microbialproduction, in part, through the food chain,leading to the support of larger numbers ofmosquito larvae. Paradise and Dunson[10] alsoreported that in addition to direct uptake viaanal papillae, mosquitoes may acquire Na+
through filter-feeding on microbes and maygrow faster under condition of highconcentration of Na+ due to decreasedallocation of energy toward acquiring Na+ orincreased production of food sources.
Table 3: Physical measure for drain water and seasoned tap water
p<0.05 = Significant; p>0.05 = Not significant; SE = Standard error
Table 4: Bacteria isolated from drain water and seasoned tap water

Comparative oviposition preferences of Aedes (Stegomyia) aegypti (L.) to water from storm water drains andseasoned tap water
Dengue Bulletin – Volume 31, 2007 129
Microbacterial examination from drainwater and seasoned tap water is presented inTable 4. A total of 9 bacteria were found in 5different drain water samples collected fromTaman Samudera. In the seasoned tap water,no bacteria were found growing on theNutrient Agar.
Since bacteria are important as nutrientsfor Ae. aegypti,[11] it is reasonable that microbesand microbial metabolites influence siteselection for egg laying.[12] Certain species ofbacteria have been shown to influenceoviposition behaviour. Trexler et al.[13] foundthat water containing Psychrobacter immobilis(from larval-rearing water), Sphingobacteriummultivorum (from soil-contaminated cottontowels) and an undetermined Bacillus species(from oak leaf infusion) elicited significantlyhigher oviposition than control water withoutbacteria. Benzon and Apperson[14] reported thattwo dominant species of bacteria, Acinetobactercalcoaceticus and Enterobacter cloacae, wereidentified in holding water, which contributedto the oviposition response of gravid Ae.aegypti. Hazard et al.[15] also reported isolatingbacterial species from an infusion of alfalfa haythat produced chemical stimulants ofoviposition in Ae. aegypti and Culexquinquefasciatus.
Ikeshoji et al.[16] isolated Pseudomonasaeruginosa and found that it produced anoviposition stimulant or attractant for Ae. aegyptiand Cx. pipens molestus Forskal in vitro fromdecanoic acid. Two bacteria of the familyPseudomonadaceae were isolated in this study,i.e. Pseudomonas gladialli and Pseudomonasalcaligenes. In studies by Ikeshoji et al.,[16] itwas shown that Ae. aegypti preferred waterwhich had this family of bacteria.
Trexler et al.[13] also reported that acombination of the bacterial species would elicita stronger oviposition response, increasing thepercentage of eggs that are deposited in thetest containers.
Larvae of many mosquito species eatbacteria or depend on them to condition organicmatter into suitable food.[11] Our study indicatesthat at least 9 species of bacteria were isolatedfrom the drain water; however, the speciesthat commonly attract Ae. aegypti to oviposittheir eggs have not been studied. The survivalrates of offspring might be increased if anovipositing female has the ability to discriminateamong habitats based on bacterial quantity andspecies composition. Thus, further studies arerequired to explain this phenomenon.
Vythilingam et al.[17] also reported that Ae.aegypti in the field is able to lay eggs in outdoordrain water. Hence, dengue vector controlmeasures should now target the concrete drainsystem as well, being a potential source of Aedesbreeding. In this study, we did attempt to detectthe presence of Aedes breeding in the drainsunder field conditions but it was not possiblebecause most portion of the drainage systemwas covered with concrete slabs. In Singapore,preventive dengue control measures includelarviciding of drains in the residential sites, asAe. aegypti and Ae. albopictus have beenconsistently found breeding in the drains (RamaChandramogan, SWRO–NEA, per. comm.).
Acknowledgments
The authors wish to thank the Director of theInstitute for Medical Research, Kuala Lumpur,for permission to publish this paper.

130 Dengue Bulletin – Volume 31, 2007
Comparative oviposition preferences of Aedes (Stegomyia) aegypti (L.) to water from storm water drains andseasoned tap water
References
[1] Malavige GN, Fernando S, Fernando DJ,Seneviratne SL. Dengue viral infections.Postgrad Med J 2004. Oct; 80(948): 588-601.
[2] Skae FMT. Dengue fever in Penang. Brit MedJ. 1902; 2: 1581-1582.
[3] Lam SK. Two decades of dengue in Malaysia.Tropical Biomedicine. 1993; 10: 195-200.
[4] Ishak H, Miyagi I, Toma T, Kamimura K.Breeding habitats of Aedes aegypti (L) andAedes. albopictus (Skuse) in villages of Barru,South Sulawesi, Indonesia. Southeast Asian JTrop Med Public Health. 1997 Dec; 28(4): 844-50.
[5] Hawley WA. The biology of Aedes albopictus.J Am Mosq Control Assoc. Suppl 1988 Dec; 1:1-39.
[6] Bentley MD, Day JF. Chemical ecology andbehavioral aspects of mosquito oviposition.Annu Rev Entomol. 1989; 34: 401-21.
[7] Chen CD, Seleena B, Nazni WA, Lee HL, MasriSM, Chiang YF, Sofian-Azirun M. Denguevectors surveillance in endemic areas in KualaLumpur City Centre and Selangor State,Malaysia. Dengue Bulletin. 2006; 30: 197-203.
[8] Lee HL. Aedes ovitrap and larval survey inseveral suburban communities in Selangor,Malaysia. Mosquito Borne Diseases Bulletin.1992; 9(1): 9-15.
[9] Kramer WL, Mulla MS. Oviposition attractantsand repellents of mosquitoes: ovipositionresponses of Culex mosquitoes to organicinfusions. Environ Entomo.l 1979; 8: 1111-1117.
[10] Paradise CJ, Dunson WA. Effects of sodiumconcentration on Aedes triseriatus (Diptera:Culicidae) and microorganisms in treeholes. JMed Entomol. 1998 Sep; 35(5): 839-44.
[11] Rozeboom LE. The relation of bacteria andbacterial filtrates to the development ofmosquito larvae. Amer J Hyg. 1935; 21: 167-179.
[12] Reiter P. [Oviposition and dispersion of Aedesaegypti in an urban environment]. Bull SocPathol Exot. 1996; 89(2): 120-2. French.
[13] Trexler JD, Apperson CS, Zurek L, Gemeno C,Schal C, Kaufman M, Walker E, Watson DW,Wallace L. Role of bacteria in mediating theoviposition responses of Aedes albopictus(Diptera: Culicidae). J Med Entomol. 2003Nov; 40(6): 841-8.
[14] Benzon GL, Apperson CS. Reexamination ofchemically mediated oviposition behavior inAedes aegypti (L.) (Diptera: Culicidae). J MedEntomol. 1988 May; 25(3): 158-64.
[15] Hazard EI, Mayer MS, Savage KE. Attractionand oviposition stimulation of gravid femalemosquitoes by bacteria isolated from hayinfusions. Mosquito News. 1967; 27: 133-136.
[16] Ikeshoji T, Saito K, Yano A. Bacterial productionof the ovipositional attractants for mosquitoeson fatty acid substrates. Appl Entomol Zool.1975; 10: 302-308.
[17] Vythilingam I, Novi S, Seleena P. Ovipositionresponse of Aedes aegypti and Aedesalbopictus in different types of water. TropicalBiomedicine. 1999; 16: 23-28.

Dengue Bulletin – Volume 31, 2007 131
Evaluation of a grass infusion-baited autocidal ovitrapfor the monitoring of Aedes aegypti (L.)
Choon Siang Tanga , Sai Gek Lam-Phuab, Youne Kow Chunga
and Andrew D. Gigerc
aEnvironmental Health Department, National Environment Agency, Environment Building,40 Scotts Road, #21-00, Singapore 228231
bEnvironmental Health Institute, National Environment Agency, Environment Building,40 Scotts Road, #21-00, Singapore 228231
cSingapore Science Centre, Singapore
Abstract
Autocidal ovitraps used for the monitoring of Aedes (Stegomyia) aegypti (L.) (Diptera: Culicidae) inSingapore are routinely stocked with cow-grass (Axonopus commpressus) infusion. The optimumconcentration of this infusion was determined to be a 1:4 dilution of a stock infusion prepared byfermenting dried cow-grass in tap water at a rate of 10g/l. Aedes aegypti females confined in an emptyroom and offered an ovitrap baited with cow-grass in competition with a range of common ovipositionsites (plastic bottle, paper cup, plastic pail, flower pot plate, Coca-Cola can, and vase) laid significantlymore eggs in the ovitrap than in the next most competitive container. The number of eggs laid in theovitrap did not change with the competitiveness of the other sites offered, nor with the number of othersites, but it did change – albeit non-linearly – with the number of mosquitoes released. Implications forthe interpretation of quantitative ovitrap data for the monitoring of mosquito populations are discussed.
Keywords: Aedes aegypti; Autocidal ovitrap; Mosquito surveillance; Hay infusion; Oviposition site.
E-mail: [email protected]; Tel: 65-67319314; Fax: 65-67319749
Introduction
Aedes aegypti (L.) is a container-inhabitingmosquito that is commonly found in urbanareas. It is the primary vector of the denguevirus which infects hundreds of thousands ofpeople every year, causing dengue fever (DF)and the potentially fatal, dengue haemorrhagicfever (DHF). No effective drug or vaccineagainst dengue has been developed despiteseveral decades of research. Hence, past and
current dengue control efforts are focused onreducing the population of Ae. aegypti.
Many control methods have beendeveloped for Ae. aegypti, including adulticidingand larviciding with insecticides and/orbreeding-source reduction. These interventionsare adequately supported by health educationand legislation. The commonly employedmethods for the surveillance of Ae. aegyptiinclude the inspection of premises for larvaeand pupae and the use of ovitraps. Most

132 Dengue Bulletin – Volume 31, 2007
Autocidal ovitrap for monitoring Ae. aegypti
operational surveillance systems depend onsuch surveys to gather their house (premise)(the percentage of houses infested with larvaeor pupae), Breteau (the number of positivecontainers per 100 houses) and larval-densityindices.[1] However, Focks and Chadee[2] foundno correlation between these traditional indicesand the measured absolute densities of Ae.aegypti and stated that the Stegomyia indicesused as epidemiological indicators of denguetransmission should be viewed with caution.They also concluded that pupal survey couldprovide an indication or estimation of the vectordensity. Direct sampling of the adult populationrequires very labour-intensive manual catchingbecause the diurnal and essentially domesticadult Ae. aegypti do not respond well to lighttraps, even if the traps are supplemented withdry ice.[3] A more detailed review of Stegomyiasampling methods was being carried out byFocks (2003),[4] and it is not necessary for thispaper to re-visit it.
Ovitraps had been used to provide usefuldata for Aedes control operation and had beenreported to be more sensitive than thetraditional Stegomyia indices in detecting lowmosquito populations.[5] Ovitraps are devicesdesigned to attract female mosquitoes to layeggs which can then be counted and identified.The original ovitrap, developed in the UnitedStates for the surveillance of Ae. aegypti, is adark-coloured, water-filled containersupplemented with a wooden paddle or velourpaper strip as oviposition substrate.[6] Thisdesign was first used in Singapore in 1969 as asupplementary measure for the control of Ae.aegypti in the Paya Lebar International Airport,Singapore.[7] Subsequently, the design wasmodified by adding a floating plastic ring withfine wire mesh to prevent the escape ofemerged adults. Chan et al.[8] demonstratedthat the resulting autocidal ovitrap was superiorto common domestic containers in attractingAe. aegypti to oviposit.
Ovitraps are now commonly used as asurveillance or monitoring tool in the field.Lee[9] and Rawlins et al.[10] found that Ae. aegyptisurveillance using ovitraps is more sensitivethan visual inspection for larvae. In the case ofAe. albopictus, studies by Ang et al.[11] and Yapet al.[12] indicated that the ovitraps they usedattracted more ovipositioning females of thatspecies than natural and other artificialcontainers.
In situations where the vector populationshave been reduced to such low levels that larvalsearches are largely unproductive, ovitraps maybe a useful indicator of the Aedes population.However, a distinction has to be madebetween using ovitraps to detect the presenceof a species in a given area, and usingquantitative ovitrap data as an indicator of thesize of a mosquito population. The latter mayonly be feasible under certain conditions, sinceovitrap catches are likely to be affected byfactors other than the number of femalemosquitoes, such as availability of alternativebreeding sites, predation on the eggs, thephysical surroundings of the ovitrap, etc. In thePhilippines, Schultz[13] found that ovitrapcatches did not reflect population changes.Studies conducted in Singapore showed thatsimultaneous adulticiding and source reductionresulted in no change in ovitrap catches (Giger,unpublished data). This may suggest that anyeffect of a reduction in the mosquito populationwas cancelled out by the effect of a reductionin the number of alternative breeding sites.Alternatively, the house index in Singapore isas low as less than 1% and this may also havean effect on ovitrap catches.
The ability of gravid mosquito females todistinguish among potential oviposition sitesthat will or will not support the growth,development and survival of their progeny iscritical because a wrong choice may result inthe death of many larvae and affect the

Autocidal ovitrap for monitoring Ae. aegypti
Dengue Bulletin – Volume 31, 2007 133
propagation of the species. Gravid females ofmany species show a high degree ofpreference in selecting specific ovipositionsites.[14,15] Aedes aegypti uses both physical andchemical cues in its oviposition site selection.[16]
These cues include the colour of the container,the texture of the oviposition substrate, andthe water quality. Over the years, variousaspects of the ovitrap have been improved,such as the structure of the oviposition substrateand the ovitrap as a whole, the water used,etc. Different types of organic materials –leaves, grass, sod, etc. – have been fermentedto create infusions attractive to gravid Ae.aegypti.[17,18,19,20] Recent reports have shownthat hay infusions were more attractive toovipositioning Ae. aegypti females than cleanwater.[10,18]
However, Chadee et al.[19] indicated thatthere were no significant preferences by gravidAe. aegypti mosquitoes to lay eggs in tap wateror various concentrations of hay infusion. Theysuggested that this could be due to thevariability in oviposition response in mosquitoesfrom different areas or variability in the type ofhay used and differences in the methodologyof preparation. Organic infusions are easy tomake but different mosquito species will needdifferent optimal infusions. The mass of theorganic material and the fermentation time willdetermine the properties of the infusion.
In Singapore, ovitraps are routinely baitedwith cow-grass infusion that has been shownin the laboratory to be more attractive to femaleAedes mosquitoes than clean water (Lam-Phua,unpublished data). Cow-grass (Axonopuscompressus) is growing throughout the countryin great abundance. It is cut on a monthly basisand usually disposed of as waste, making iteasily available for this alternative use.
The study presented here consisted ofthree experiments, aiming to determine theconcentration of cow-grass infusion that worksbest in the field (“Optimum concentration ofinfusion”); evaluate the competitiveness of theautocidal ovitrap baited with cow-grass infusionagainst various artificial containers commonlyassociated with Ae. aegypti breeding (“Ovitrapcompetitiveness”); and ascertain the effects ofmosquito population size and availability ofalternative breeding sites on the ovitrap catch(“Effects on ovitrap catch”).
Materials and methods
Mosquito colony origin andmaintenance
Aedes aegypti larvae were collected inSingapore between January and August 2002and reared to adults. Their offspring werereared in plastic trays (25 cm × 31 cm × 6cm) with aged tap water, limiting the numberof larvae to 800 per tray to avoid overcrowding.The colonies were maintained at 28 ± 1 °C,79 ± 3% RH, and a photoperiod ofapproximately 9:15 hours (L:D). First instarlarvae were initially fed with instant yeast (BakeKing, Singapore). Subsequent instars were fedwith dog chow (Glen Forrest Stockfeeders,Western Australia). The adults were kept inPlexiglas cages (30 cm × 30 cm × 30 cm)fitted with cotton cloth on two sides and werecontinuously provided with a 10% sucrosesolution. The F1 and F2 generations were usedin the experiments. The females were blood-fed on guinea pigs 3–5 days after emergenceand used for the experiments two days later.This timing ensured that the oviposition peakfour days after the blood meal[21] fell withinthe one-week duration of each replicate.

134 Dengue Bulletin – Volume 31, 2007
Autocidal ovitrap for monitoring Ae. aegypti
Preparation of infusion
Fresh cow-grass was collected by a contractor,cleaned of foreign matter, dried under the sunand packed in zip-loc bags until further use.Following a modification of the method ofReiter et al.,[18] stock infusions were preparedby fermenting dried cow-grass in tap water ata rate of 10 g/l. Grass and tap water werecontained in a 50-litre plastic trough and werefermented for one week at a temperature of28 ± 1 °C. The infusion was then strainedthrough a screen to remove the cow-grass, andkept in plastic canisters at 28 ± 1 °C for notmore than 8 weeks.
Study site
All experiments were carried out in 16 emptyrooms (3.5 m × 3.0 m, 2.5 m high; concretewalls, floor and ceiling) in the same block offlats. All windows and openings were sealedwith fine netting to prevent the mosquitoesfrom escaping while providing sufficientventilation within the rooms. Throughout thestudy, the relative humidity and temperatureinside the rooms was 78 ± 11% RH and 30 ±1 °C.
Experiment 1: Optimumconcentration of infusion
The stock cow-grass infusion was diluted withweek-old aged tap water to obtain a range ofconcentrations. For each of the 48 replicates,four autocidal ovitraps[8] (height 155 mm,internal diameter 103 mm) with two hardboardoviposition paddles each (82 mm × 32 mm ×4 mm) and a floating plastic ring with fine wiremesh to prevent the escape of emerged adultswere used (Figure 1). Each ovitrap was filledwith 700 ml of cow-grass infusion at four
different concentrations (0%, 25%, 50% and100%) and randomly assigned to the fourcorners of a room. Ten gravid females werethen released into the room and leftundisturbed for 7 days. At the end of thatperiod, larvae found in the ovitraps werecounted and discarded, while the ovipositionpaddles were packed in individual plastic bagsand brought back to the laboratory forexamination. Eggs laid on the paddles werecounted under a stereomicroscope, excludingempty egg shells. The total number of larvaeand unhatched eggs were tabulated for eachovitrap. Data analysis was carried out on thetotal number of the eggs laid during the 7-dayperiod, i.e. the number of larvae plus eggs laidon the paddles. The differences between datapoints were tested for significance usingANOVA (SPSS Ver 15.0).
Figure 1: Autocidal ovitrap. The twowooden paddles allow mosquitoes to layeggs and the float plastic ring with fine
wire mesh helps to prevent the escape ofemerged adult mosquitoes
Adult
Pupa
Larva
Wire mesh Cardboardpaddle
Overflow hole

Autocidal ovitrap for monitoring Ae. aegypti
Dengue Bulletin – Volume 31, 2007 135
Experiment 2: Ovitrapcompetitiveness
Autocidal ovitraps baited with the optimuminfusion determined in experiment 1 weretested against six different artificial containersthat are commonly found in households andare known to attract oviposition by Ae aegyptiin the field. These containers were: plasticbottle (height 204 mm, base diameter 60 mm,transparent); paper cup (height 115 mm,opening diameter 80 mm, white with greenpattern); plastic pail (3 litres, blue); flower potplate (plastic, diameter 130 mm) under a pottedmoney plant (Scindapsus aureus); aluminiumCoca-Cola drink can (height 80 mm, diameter58 mm, red and silver); vase (height 200 mm,opening diameter 65 mm, pink) with two luckybamboo plants (Dracaena sp.). All the artificialcontainers were filled with week-old aged tapwater to simulate field conditions.
For each of the 48 replicates, threedifferent artificial containers and one ovitrapwere randomly assigned to the four corners ofa vacant room. The selection of containers foreach replicate was random, with the conditionthat each container was included 24 times. Tengravid females were then released into theroom for 7 days, after which any larvae in thecontainers and ovitraps were counted, and thecontainers and oviposition paddles were broughtback to the laboratory and thoroughlyexamined for eggs. The total number of larvaeand unhatched eggs were tabulated for eachcontainer and ovitrap. Data analysis was carriedout as in Experiment 1.
Experiment 3: Effects onovitrap catch
In this experiment, one ovitrap was placed ina vacant room together with either 1, 3 or 9pails as alternative breeding sites, and in each
of these situations either 1, 3 or 9 gravidmosquitoes were released and left undisturbedfor 7 days. Thus, there were nine differentdesigns, each of which was replicated fourtimes. Data collection and analysis were carriedout as in Experiments 1 and 2.
Results
Experiment 1: Optimumconcentration of infusion
In order to determine the optimumconcentration of cow-grass infusion, two roundsof replicates were conducted. In the first round,the four concentrations offered were 0% (i.e.aged tap water only), 25%, 50% and 100% ofthe stock infusion. In this round, significantlymore eggs were laid in the ovitraps with 25%infusion than in ovitraps containing any otherconcentration (p<0.01; Figure 2). There wasno significant difference in the amount of eggslaid in the ovitraps with 0%, 50% and 100%infusion. In the second round of replicates, therange of concentrations was narrowed downto 0–45%. Here the numbers of eggs laid in15%, 30% and 45%, respectively, were notsignificantly different, but all three of theseconcentrations received significantly more eggs
Figure 2: Mean number of eggs laid inovitraps containing different
concentrations of cow-grass infusion(Error bars: standard error)
300
250
200
150
100
50
0
Mea
nnu
mbe
rof
eggs
laid
Infusion Concentration [%]
0 10 20 30 40 50 60 70 80 90 100
1 round2 round
st
nd

136 Dengue Bulletin – Volume 31, 2007
Autocidal ovitrap for monitoring Ae. aegypti
than 0% (p<0.01). Hence, concentrationsbetween 15% and 45% can be regarded asoptimum concentrations. For practical reasons(ease of dilution of stock infusion by vectorcontrol officers in the field) the concentrationof 20% stock infusion was selected for thestandard infusion to be used in the remainingexperiments and subsequent application in thefield.
Experiment 2: Ovitrapcompetitiveness
The second experiment aimed to determinehow well the autocidal ovitrap with cow-grassinfusion competes with other types of potentialbreeding sites commonly found in the field.Pitted against three different containers at atime, the ovitrap attracted an average of 77.1%of the eggs laid in each replicate (Figure 3),which is significantly more than any othercontainer (p<0.01). Among the remainingcontainers, the pail attracted significantly moreeggs than any one of the others (p<0.01).There were no significant differences in theegg count among the other containers.
To determine the effect of the presence orabsence of the most competitive alternativebreeding site (the pail) on the absolute egg countin the ovitrap and the total egg count in eachreplicate, the replicates with a pail present werecompared with the replicates without a pail. Thepresence of a pail did not have any significanteffect on the absolute number of eggs laid inthe ovitrap (mean absolute egg count ± standarderror: with pail 538.5 ± 37.4; without pail 506.8± 32.3; p>0.05), but the average total eggcount in each replicate was significantly higherwith the pail present (with pail 764.1 ± 32.9;without pail 595.5 ± 29.4; p<0.01).
Experiment 3: Effects onovitrap catch
The third experiment was designed todetermine the respective effects of themosquito population size and the number ofalternative breeding sites on the number ofeggs collected from ovitraps. The mostcompetitive container in experiment 2 (thepail) was used as an alternative breeding site.
Figure 3: Mean percentage of eggs laid inthe ovitrap (n = 48) and the six alternative
containers (n = 24) in each of 48replicates offering one ovitrap and
three different alternatives.(Error bars: standard error)
Mea
npe
rcen
tage
ofeg
gsla
id
100
80
60
40
20
0
Ovitrap77.1
Pail19.8
Vase8.9
Plate5.7
Can5.6
Cup4.9
Bottle1.0
Figure 4: Mean number of eggs laid in theovitrap and alternative oviposition sites as
a function of the number of alternativesites and the number of mosquitoes
released. n = 4 for each column
a, b: p<0.01c, d, e: p<0.05
Ovitrap
All alternativebreeding sites
Mea
nnu
mbe
rof
eggs
laid
700
600
500
400
300
200
100
0
Number ofalt. breeding sites
93
1 13
9
Number ofMosquitoes

Autocidal ovitrap for monitoring Ae. aegypti
Dengue Bulletin – Volume 31, 2007 137
The number of alternative breeding siteshad no significant effect on the number of eggslaid in the ovitrap, but the number ofmosquitoes did (Figure 4, significantly differentvalues are marked with a, b: p<0.01; c, d, e:p<0.05). However, the ovitrap catch did notincrease linearly with the number ofmosquitoes released into the room; with ninemosquitoes, the number of eggs in the ovitrapswas, on average, less than twice that withthree mosquitoes.
Discussion
As a product of fermentation, grass infusionmay vary greatly in its attractive properties,depending on the quality and relative amountsof grass and water used, duration offermentation, ambient temperature duringfermentation, subsequent dilution and possiblyother factors. However, if grass infusion is tobe used in ovitraps for the purpose ofquantitative monitoring of mosquitopopulations, its properties should ideally beconstant and reproducible in order to allowcomparisons over time and between ovitraps.In an attempt to standardize our cow-grassinfusion as much as possible, we used driedgrass and controlled the ratio of grass to water,ambient temperature and duration offermentation to prepare a stock infusion.Experiment 1, carried out to determine theoptimum dilution of the stock infusion,demonstrated that ovipositing femalespreferred infusions prepared from 15–45%stock infusion. Taking pragmatic considerationsinto account, the optimum concentration forthe subsequent experiments as well as theapplication in operational monitoring was thusdefined as a 1:4 dilution (to obtain aconcentration of 20%) of the stock infusion.
The competitiveness of the autocidalovitrap against a range of other containers had
previously been tested for Ae. albopictus withall containers in close proximity.[7] In Experiment2 we attempted to evaluate thatcompetitiveness for Ae. aegypti and in a settingmore representative of the situation in the field,where ovitraps are not placed immediately nextto competing breeding sites. However, due tooperational constraints, our experiments hadto be conducted within the confines of singlerooms, which is still very different from thefield situation, where the closest water-bearingcontainer may be far away from the ovitrapand may be in a different room of a building orotherwise physically separated. Furthermore,the mosquitoes in our experiments wereconfined for one week in the vicinity of thesame four breeding sites, which is unlikely tobe the case in the field. Nevertheless, bothexperiments 2 and 3 demonstrate that, in asetting which provides the mosquitoes withample opportunity to investigate all availableoviposition sites, the ovitrap was much morecompetitive than any of the other containerstested. This high acceptability of the ovitrapwith grass infusion to ovipositing females –even in the presence of other potentialbreeding sites – means that the ovitrap is wellsuited for the application as a monitoring toolin the field.
Even a very competitive ovitrap might beexpected to receive fewer eggs in the presenceof alternative oviposition sites. However, theresults of experiment 2 reveal that the presenceof a pail (the most competitive alternativecontainer) did not decrease the absolutenumber of eggs laid in the ovitrap but, rather,increased the total number of eggs laid.Similarly, in experiment 3, the number ofalternative breeding sites had no bearing onthe ovitrap catch. These findings may beexplained by the fact that all availableoviposition sites were offered within one roomand the mosquitoes were confined to thatroom. Ae. aegypti females are known to

138 Dengue Bulletin – Volume 31, 2007
Autocidal ovitrap for monitoring Ae. aegypti
distribute their eggs among a number ofoviposition sites (“skip oviposition”).[22,23] In thefield, mosquitoes would encounter differentoviposition sites sequentially, and the qualityof an alternative breeding site encounteredbefore an ovitrap may well affect themosquito’s subsequent exploration and, thus,its chances of finding the ovitrap in the firstplace. On the other hand, after laying a clutchof eggs in an ovitrap in the field, the mosquitomay well move out of the vicinity of the ovitrapand not return, a behaviour which would havebeen prevented in our experiment, leading torepeated oviposition in the ovitrap. Thesespeculative arguments require furtherinvestigation, but they illustrate that the resultsof the experiments reported here do not ruleout that the catches of ovitraps set in the fieldare affected by the number of alternativeoviposition sites in the area.
In Experiment 3, the number of eggs laidin the ovitrap increased with the number ofthe mosquitoes present. This increase, however,was non-linear, with fewer eggs laid permosquito when more mosquitoes were present.While the total number of eggs laid in eachreplicate increased more or less linearly withthe number of mosquitoes, the ovitrapcollected a decreasing share of these eggs. Inother words, each mosquito, on average, laidfewer eggs in the ovitrap (the most attractivecontainer) when there were more mosquitoescompeting for the same oviposition sites. Thisfinding, as well as the result of experiment 2
where more eggs were laid, in total, with twoattractive containers available (ovitrap and pail)than with only one (ovitrap), suggests that thereis a limit to the number of eggs an ovitrap canaccommodate. The nature of such a restrictionto oviposition remains unclear, but this findingagrees well with studies that demonstrated thatfemale Ae. aegypti lay fewer eggs in containerswith higher existing egg density.[24,25]
This study has established that theautocidal ovitrap baited with a standardizedcow-grass infusion is highly competitive againstoviposition sites commonly encountered in thefield. Consequently, this ovitrap is well suitedfor Aedes surveillance in Singapore as far asthe choice of container and infusion isconcerned. The present knowledge isinsufficient, however, to conclude whetherquantitative data collected with these ovitrapswould be able to provide a measure of themosquito population in the field. To determinethat, more research is required into thecompetitiveness of the ovitrap under true fieldconditions, as well as the nature of the apparentlimit to the capacity of the ovitrap.
Acknowledgment
We would like to thank Mr Khoo Seow Poh,Director-General for Public Health, for hispermission to publish this paper. Our greatestthank go to our lab attendant, Mr Semawi BinSyed, for all the hard work done by him.
References
[1] Chan YC, Ho BC, Chan KL. Aedes aegypti (L.)and Aedes albopictus (Skuse) in Singapore City.5. Observations in relation to denguehaemorrhagic fever. Bull World Health Organ.1971; 44(5): 651-7.
[2] Focks DA, Chadee DD. Pupal survey: anepidemiologically significant surveillancemethod for Aedes aegypti: an example usingdata from Trinidad. Am J Trop Med Hyg. 1997Feb; 56(2): 159-67.

Autocidal ovitrap for monitoring Ae. aegypti
Dengue Bulletin – Volume 31, 2007 139
[3] Chan KL. Methods and indices used in thesurveillance of dengue vectors. Mosquito-BorneDisease Bulletin. 1985; 1: 79-88.
[4] Focks DA. A review of entomological samplingmethods and indicators for dengue vectors.Geneva: World Health Organization, 2003.Document TDR/IDE/Den/03.1.
[5] Micks DW, Moon WB. Aedes aegypti in aTexas coastal county as an index of denguefever receptivity and control. Am J Trop MedHyg. 1980 Nov; 29(6): 1382-8.
[6] Schliessmann DJ. Aedes aegypti eradicationprogram of the United States–progress report1965. Am J Public Health Nations Health. 1967Mar; 57(3): 460-5.
[7] Chan KL. The eradication of Aedes aegypti atthe Singapore Paya Lebar International Airport.In: Chan YC, Chan KL, Ho BC, editors. VectorControl in Southeast Asia. Singapore, 1973. p.85-88.
[8] Lok CK, Kiat NS, Koh TK. An autocidal ovitrapfor the control and possible eradication ofAedes aegypti. Southeast Asian J Trop MedPublic Health. 1977 Mar; 8(1): 56-62.
[9] Lee HL. Aedes ovitrap and larval survey inseveral suburban communities in Selangor,Malaysia. Mosquito-Borne Disease Bulletin.1992; 9(1): 9-15.
[10] Rawlins SC, Martinez R, Wiltshire S, Legall G.A comparison of surveillance systems for thedengue vector Aedes aegypti in Port of Spain,Trinidad. J Am Mosq Control Assoc. 1998 Jun;14(2): 131-6.
[11] Ang WK, Chan KL, Lee, K.M. Field evaluationof the attractiveness of an ovitrap to Aedesalbopictus (diptera, culicidae). Proceedings,Science Research Congress, Singapore; 1995.
[12] Yap HH, Lee CY, Chong NL, Foo AE, Lim MP.Oviposition site preference of Aedes albopictusin the laboratory. J Am Mosq Control Assoc.1995 Mar; 11(1): 128-32.
[13] Schultz GW. Seasonal abundance of denguevectors in Manila, Republic of the Philippines.Southeast Asian J Trop Med Public Health. 1993Jun; 24(2): 369-75.
[14] Zahiri N, Rau ME. Oviposition attraction andrepellency of Aedes aegypti (Diptera:Culicidae) to waters from conspecific larvaesubjected to crowding , confinement,starvation, or infection. J Med Entomol. 1998Sep; 35(5): 782-7.
[15] Roberts D. Overcrowding of Culex sitiens(Diptera: Culicidae) larvae: populationregulation by chemical factors or mechanicalinterference. J Med Entomol. 1998 Sep; 35(5):665-9.
[16] Bentley MD, Day JF. Chemical ecology andbehavioral aspects of mosquito oviposition.Annu Rev Entomol. 1989; 34: 401-21.
[17] Holck AR, Meek CL, Holck JC. Attractantenhanced ovitraps for the surveillance ofcontainer breeding mosquitoes. J Am MosqControl Assoc. 1988 Mar; 4(1): 97-8.
[18] Reiter P, Amador MA, Colon N. Enhancementof the CDC ovitrap with hay infusions for dailymonitoring of Aedes aegypti populations. J AmMosq Control Assoc. 1991 Mar; 7(1): 52-5.
[19] Chadee DD, Lakhan A, Ramdath WR, PersadRC. Oviposition response of Aedes aegyptimosquitoes to different concentrations of hayinfusion in Trinidad, West Indies. J Am MosqControl Assoc. 1993 Sep; 9(3): 346-8.
[20] Allan SA, Kline JD. Evaluation of organicinfusions and synthetic compounds mediatingoviposition in Aedes albopictus and Aedesaegypti (Diptera: Culicidae). J Chemical Ecol.1995; 21: 1847-1860.
[21] Trexler JD, Apperson CS, Schal C. Dieloviposition patterns of Aedes albopictus (Skuse)and Aedes triseriatus (Say) in the laboratoryand the field. J Vector Eco.l 1997 Jun;22(1):64-70.

140 Dengue Bulletin – Volume 31, 2007
Autocidal ovitrap for monitoring Ae. aegypti
[22] Gillett JD. Contributions to the oviposition-cycle by the individual mosquitoes in apopulation. Journal of Insect Physiology. 1962;8: 665-681.
[23] Reiter P, Amador MA, Anderson RA, Clark GG.Short report: dispersal of Aedes aegypti in anurban area after blood feeding asdemonstrated by rubidium-marked eggs. AmJ Trop Med Hyg. 1995 Feb; 52(2): 177-9.
[24] Apostol BL, Black WC 4th, Reiter P, Miller BR.Use of randomly amplified polymorphic DNAamplified by polymerase chain reactionmarkers to estimate the number of Aedesaegypti families at oviposition sites in San Juan,Puerto Rico. Am J Trop Med Hyg. 1994 Jul;51(1): 89-97.
[25] Chadee DD, Corbet PS and Greenwood JJD.Egg-laying yellow fever mosquitoes avoid sitescontaining eggs laid by themselves orconspecifics. Entomologia Experimentalis etApplicata. 1990; 57:295-298.

Dengue Bulletin – Volume 31, 2007 141
Development sites of Aedes aegypti and Ae. albopictusin Nakhon Si Thammarat, Thailand
S. Wongkoon, M. Jaroensutasinee , K. Jaroensutasinee and W. Preechaporn
Computational Science Graduate Programme, School of Science, Walailak University, Thasala,Nakhon Si Thammarat 80161, Thailand
Abstract
This study investigated how the seasons affect the development sites of Aedes larvae in three topographicalareas: mangrove, rice paddy fields and mountainous areas. We examined how the number of Aedeslarvae varied in different types of water containers. Water containers were categorized into the followinggroups: indoor/outdoor containers, artificial/natural containers, earthen/plastic containers, containerswith/without lids and dark/light-coloured containers. Samples were collected from 300 households inboth the wet and dry seasons from three topographical areas in Nakhon Si Thammarat province with100 households per topographical area. The results showed that in the wet season, there were highernumbers of Ae. aegypti and Ae. albopictus larvae than in the dry season. Moreover, the number of Ae.albopictus larvae was higher in mountainous areas than in mangrove and rice paddy areas, in both thewet and dry seasons. The number of positive containers was higher in outdoor containers than inindoor containers, higher in artificial containers than natural containers, higher in earthen containersthan plastic containers, higher in containers without lids than containers with lids, and higher in thedark-coloured containers than light-coloured containers in the three topographical areas in both wetand dry seasons. The number of positive earthen containers was higher in mangrove areas than in ricepaddy and mountainous areas. The number of positive plastic containers was higher in rice paddyareas than mountainous and mangrove areas.
Keywords: Aedes larvae; Season; Topography; Larval development site.
E-mail: [email protected]
Introduction
Dengue fever is caused by dengue viruses ofthe family Flaviviridae, transmitted principallyby Aedes aegypti, and, possibly Ae. albopictus,in the tropical and subtropical regions of theworld.[1,2] Two clinical features, namely, denguehaemorrhagic fever (DHF) and dengue shocksyndrome (DSS) may lead to death.[3,4] Noeffective vaccine or chemotherapy is currentlyavailable for the prevention or treatment of
dengue fever; therefore, prevention and controlof the disease depend on vector surveillanceand control measures.[5] Transmission cycles ofthe dengue virus depend on theinterrelationship between the virus and itsmosquito vector, which is influenced byenvironmental conditions.[6] Adult female Aedesmosquitoes acquire the dengue virus by bitinginfected humans during the viremic phase,which usually lasts for 4–5 days, although itmay last up to 12 days. The virus is transmitted

142 Dengue Bulletin – Volume 31, 2007
Development sites of Ae. aegypti and Ae. albopictus in Thailand
to other persons via bites from infectedmosquitoes.[7] The mosquitoes that adverselyaffect people in southern Thailand are primarilyAe. aegypti L. and Ae. albopictus Skuse.[8-12]
An epidemic of DHF occurred in the SamuiIslands in 1966 and 1967[13] where Ae. aegyptiand Ae. albopictus were abundant, and wereresponsible for the transmission of the denguevirus.[14-16] Ae. aegypti breeds in a wideassortment of domestic containers, whereasAe. albopictus is more likely to be found innatural containers, such as bamboo stumps andcoconut shells, or in artificial containers outsidethe houses such as tyres, opened cans andplastic bottles.[12,17-19] In southern Thailand, Ae.aegypti and Ae. albopictus have been found inforested habitats as well as in a variety of otherhabitats in rural and suburban areas.[11-12,18-19]
Since most Thai households store waterfor cooking and bathing in a variety of jars and
cisterns, Ae. aegypti is a more important threatfor DHF.[20] Ae. albopictus is encountered inthe peripheral areas of towns where it replacesAe. aegypti populations.[21]
This study investigated the number ofmosquito larvae (i.e. Ae. aegypti and Ae.albopictus) in different types of containers (i.e.indoor/outdoor containers, artificial/naturalcontainers, earthen/plastic containers,containers with/without lids and dark/lightcoloured containers) in the wet and dry seasonsin three topographical areas in Nakhon SiThammarat, Thailand.
Materials and methods
Data collection
The mosquito larval survey was conducted inNakhon Si Thammarat province (8° 32' 16.5"
Figure 1: (a) Map of Thailand; (b) Map of three topographical areas of study
Thailand
Nakhon Si Thammarat
(a) (b)
123412341234
123412341234
1234123412341234Mangrove, Rice paddy and Mountainous areas

Development sites of Ae. aegypti and Ae. albopictus in Thailand
Dengue Bulletin – Volume 31, 2007 143
N latitude, 99° 56' 50.7" E longitude) in twoseasons: (i) dry season (March-April 2006) and(ii) wet season (October-November 2006). Weconducted the mosquito larval survey in threetopographical areas: mangrove, rice paddy andmountainous areas (Figure 1). People in thesethree topographical areas differed in terms oftheir main occupations. In the mangrove areas,the main occupations of the people are fishingand aquaculture, while in the rice paddy andmountainous areas, the main occupations aremostly agriculture and farm-related.
Samples were collected in households inall sub-districts in the nine districts comprisingthe study area using the stratified simplerandom sampling technique. Nine districtswere divided into three categories: mangrove,rice paddy and mountainous areas. Themangrove areas included Khanom, Sichon andPakpanang districts; the rice paddy areasincluded Praprom, Chalermprakiet and Huasaidistricts; and the mountainous areas includedThungyai, Bangkhan and Thungsong districts(Figures 1a, 1b). One hundred sampling unitswere assigned to each topographical area. Thetopographical areas were assigned as stratums,one collected household identified as a sampleunit. A structured questionnaire was constructedto obtain the information by personal interview.There were a total of 300 households sampledin the dry season. The same 300 householdswere re-sampled in the wet season in order tocompare the difference in the number of Aedeslarvae in various water containers between thetwo seasons.
Entomological studies
We collected all mosquito larvae from bothindoor and outdoor containers using fine-meshed fishnets. The outdoor larval surveyswere conducted within 15 m of thehouses.[10,12,18-19] Very small water containerswere emptied out through the fishnet. Largerwater containers were sampled by dipping the
net in the water, starting at the top of thecontainer and continuing to the bottom in aswirling motion that sampled all edges of thecontainer[12,18-19,22]. All live mosquito larvae werecollected in plastic bags, taken to the 11.2Vector-Borne Disease Control Centrelaboratory, Nakhon Si Thammarat. All mosquitolarvae were preserved and identified up to thespecies level using Rattanarithikul andPanthusiri’s[23] keys. In this study, the first andsecond instars and pupae were discardedbecause immature mosquitoes at these stagescould not be identified.
There were a total of 29 containercategories in this study. Water containers werecategorized into the following groups: indoor/outdoor containers, artificial/natural containers,earthen/plastic containers, containers with/without lids and dark/light-coloured containers.Indoor containers were all containers inside thehouse that held some water such as earthenjars, cement tanks, plastic containers, flowervases, ant guards, ornamental pots andrefrigerators with plates. Outdoor containerswere all containers within 15 m of the house,such as earthen jars, cement tanks, plasticcontainers, ornamental pots and naturalcontainers. Artificial containers included earthenjars, flower vases, used cans, discarded tyres,metal boxes, preserved areca jars and foamcontainers. The preserved areca jars wereearthen jars used to preserve areca nuts. Naturalcontainers were composed of areca/coconuthusks, banana trees, coconut shells, tree holesand bamboo clumps. Earthen containers werecomposed of small/large earthen jars, cementtanks, plant pot saucers, plant pots andpreserved areca jars. Plastic containers werecomposed of plastic containers, plastic bottlesand plastic buckets. Lids were made of cement,plastic, wood and metal. The dark-colouredcontainers were containers that were black,brown, dark grey, deep green, deep blue andred. On the other hand, the light colouredcontainers were containers that were white, lightpink, light blue and yellow.
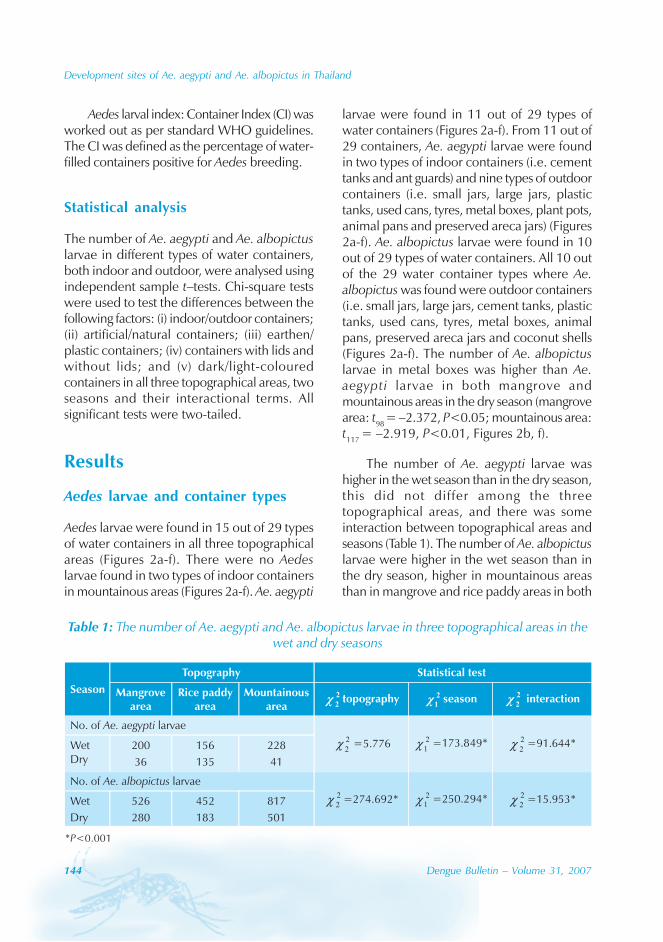
144 Dengue Bulletin – Volume 31, 2007
Development sites of Ae. aegypti and Ae. albopictus in Thailand
Aedes larval index: Container Index (CI) wasworked out as per standard WHO guidelines.The CI was defined as the percentage of water-filled containers positive for Aedes breeding.
Statistical analysis
The number of Ae. aegypti and Ae. albopictuslarvae in different types of water containers,both indoor and outdoor, were analysed usingindependent sample t–tests. Chi-square testswere used to test the differences between thefollowing factors: (i) indoor/outdoor containers;(ii) artificial/natural containers; (iii) earthen/plastic containers; (iv) containers with lids andwithout lids; and (v) dark/light-colouredcontainers in all three topographical areas, twoseasons and their interactional terms. Allsignificant tests were two-tailed.
Results
Aedes larvae and container types
Aedes larvae were found in 15 out of 29 typesof water containers in all three topographicalareas (Figures 2a-f). There were no Aedeslarvae found in two types of indoor containersin mountainous areas (Figures 2a-f). Ae. aegypti
larvae were found in 11 out of 29 types ofwater containers (Figures 2a-f). From 11 out of29 containers, Ae. aegypti larvae were foundin two types of indoor containers (i.e. cementtanks and ant guards) and nine types of outdoorcontainers (i.e. small jars, large jars, plastictanks, used cans, tyres, metal boxes, plant pots,animal pans and preserved areca jars) (Figures2a-f). Ae. albopictus larvae were found in 10out of 29 types of water containers. All 10 outof the 29 water container types where Ae.albopictus was found were outdoor containers(i.e. small jars, large jars, cement tanks, plastictanks, used cans, tyres, metal boxes, animalpans, preserved areca jars and coconut shells(Figures 2a-f). The number of Ae. albopictuslarvae in metal boxes was higher than Ae.aegypti larvae in both mangrove andmountainous areas in the dry season (mangrovearea: t98 = –2.372, P<0.05; mountainous area:t117 = –2.919, P<0.01, Figures 2b, f).
The number of Ae. aegypti larvae washigher in the wet season than in the dry season,this did not differ among the threetopographical areas, and there was someinteraction between topographical areas andseasons (Table 1). The number of Ae. albopictuslarvae were higher in the wet season than inthe dry season, higher in mountainous areasthan in mangrove and rice paddy areas in both
Table 1: The number of Ae. aegypti and Ae. albopictus larvae in three topographical areas in thewet and dry seasons
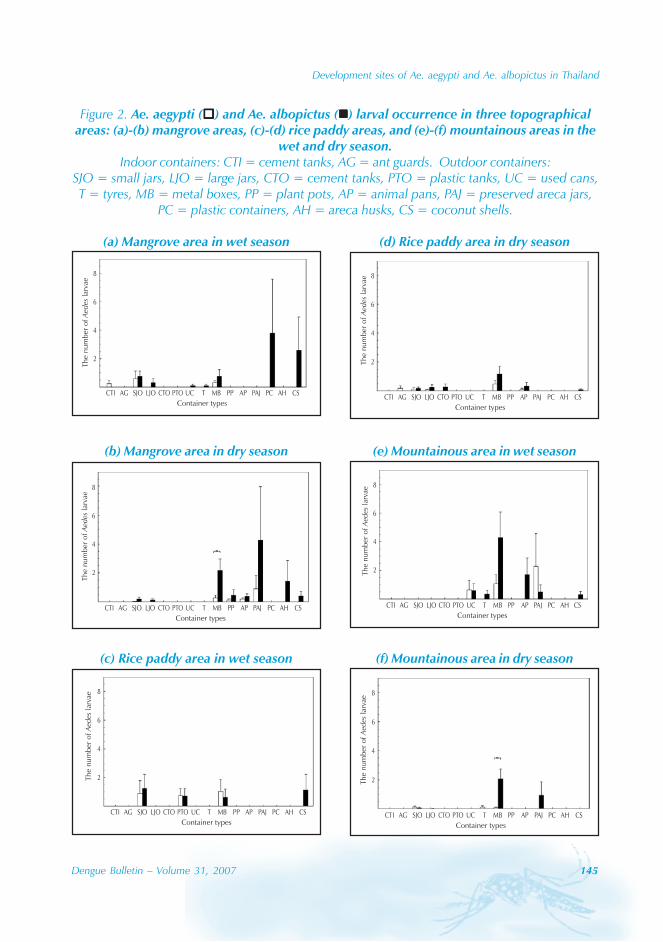
Development sites of Ae. aegypti and Ae. albopictus in Thailand
Dengue Bulletin – Volume 31, 2007 145
Figure 2. Ae. aegypti ( ) and Ae. albopictus ( ) larval occurrence in three topographicalareas: (a)-(b) mangrove areas, (c)-(d) rice paddy areas, and (e)-(f) mountainous areas in the
wet and dry season.Indoor containers: CTI = cement tanks, AG = ant guards. Outdoor containers:
SJO = small jars, LJO = large jars, CTO = cement tanks, PTO = plastic tanks, UC = used cans,T = tyres, MB = metal boxes, PP = plant pots, AP = animal pans, PAJ = preserved areca jars,
PC = plastic containers, AH = areca husks, CS = coconut shells.
The
nu
mb
erof
larv
aeA
ed
es
8
6
4
2
Container types
CTI AG SJO LJO CTO PTO UC T MB PP AP PAJ PC AH CS
(a) Mangrove area in wet season
(b) Mangrove area in dry season
The
nu
mb
erof
larv
aeA
ed
es
8
6
4
2
Container types
CTI AG SJO LJO CTO PTO UC T MB PP AP PAJ PC AH CS
(c) Rice paddy area in wet season
The
nu
mb
erof
larv
aeA
ed
es
8
6
4
2
Container types
CTI AG SJO LJO CTO PTO UC T MB PP AP PAJ PC AH CS
(d) Rice paddy area in dry season
The
nu
mb
erof
larv
aeA
ed
es
8
6
4
2
Container types
CTI AG SJO LJO CTO PTO UC T MB PP AP PAJ PC AH CS
(e) Mountainous area in wet season
The
nu
mb
erof
larv
aeA
ed
es
8
6
4
2
Container types
CTI AG SJO LJO CTO PTO UC T MB PP AP PAJ PC AH CS
(f) Mountainous area in dry season
The
nu
mb
erof
larv
aeA
ed
es
8
6
4
2
Container types
CTI AG SJO LJO CTO PTO UC T MB PP AP PAJ PC AH CS
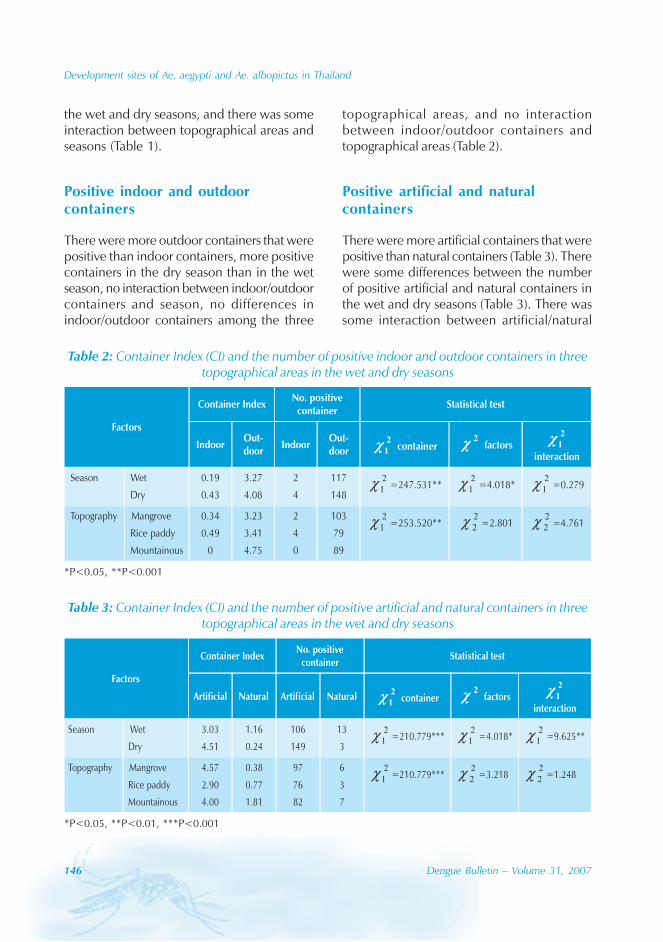
146 Dengue Bulletin – Volume 31, 2007
Development sites of Ae. aegypti and Ae. albopictus in Thailand
the wet and dry seasons, and there was someinteraction between topographical areas andseasons (Table 1).
Positive indoor and outdoorcontainers
There were more outdoor containers that werepositive than indoor containers, more positivecontainers in the dry season than in the wetseason, no interaction between indoor/outdoorcontainers and season, no differences inindoor/outdoor containers among the three
Table 3: Container Index (CI) and the number of positive artificial and natural containers in threetopographical areas in the wet and dry seasons
Table 2: Container Index (CI) and the number of positive indoor and outdoor containers in threetopographical areas in the wet and dry seasons
topographical areas, and no interactionbetween indoor/outdoor containers andtopographical areas (Table 2).
Positive artificial and naturalcontainers
There were more artificial containers that werepositive than natural containers (Table 3). Therewere some differences between the numberof positive artificial and natural containers inthe wet and dry seasons (Table 3). There wassome interaction between artificial/natural
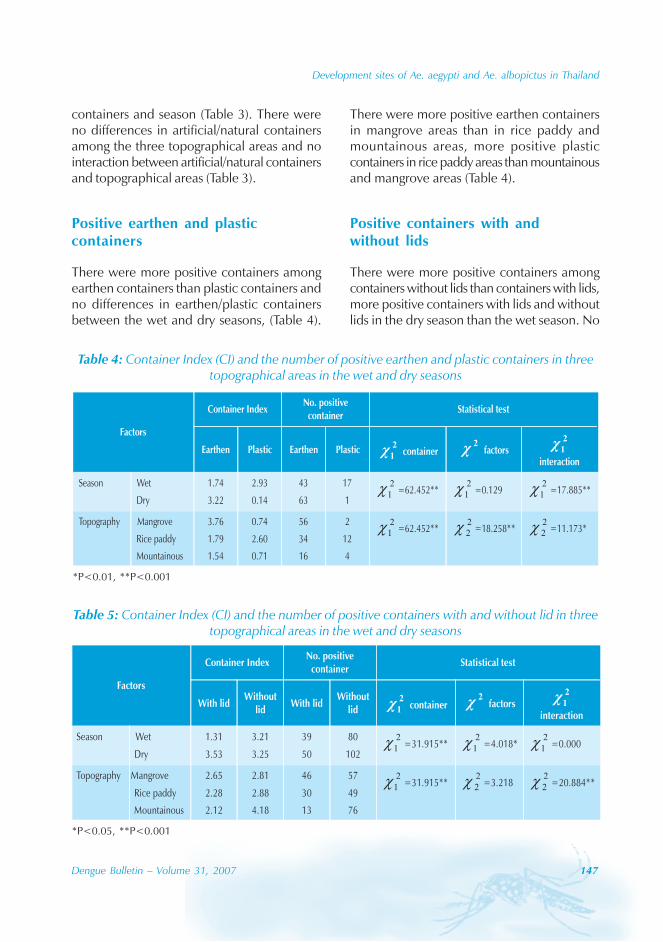
Development sites of Ae. aegypti and Ae. albopictus in Thailand
Dengue Bulletin – Volume 31, 2007 147
containers and season (Table 3). There wereno differences in artificial/natural containersamong the three topographical areas and nointeraction between artificial/natural containersand topographical areas (Table 3).
Positive earthen and plasticcontainers
There were more positive containers amongearthen containers than plastic containers andno differences in earthen/plastic containersbetween the wet and dry seasons, (Table 4).
Table 5: Container Index (CI) and the number of positive containers with and without lid in threetopographical areas in the wet and dry seasons
Table 4: Container Index (CI) and the number of positive earthen and plastic containers in threetopographical areas in the wet and dry seasons
There were more positive earthen containersin mangrove areas than in rice paddy andmountainous areas, more positive plasticcontainers in rice paddy areas than mountainousand mangrove areas (Table 4).
Positive containers with andwithout lids
There were more positive containers amongcontainers without lids than containers with lids,more positive containers with lids and withoutlids in the dry season than the wet season. No
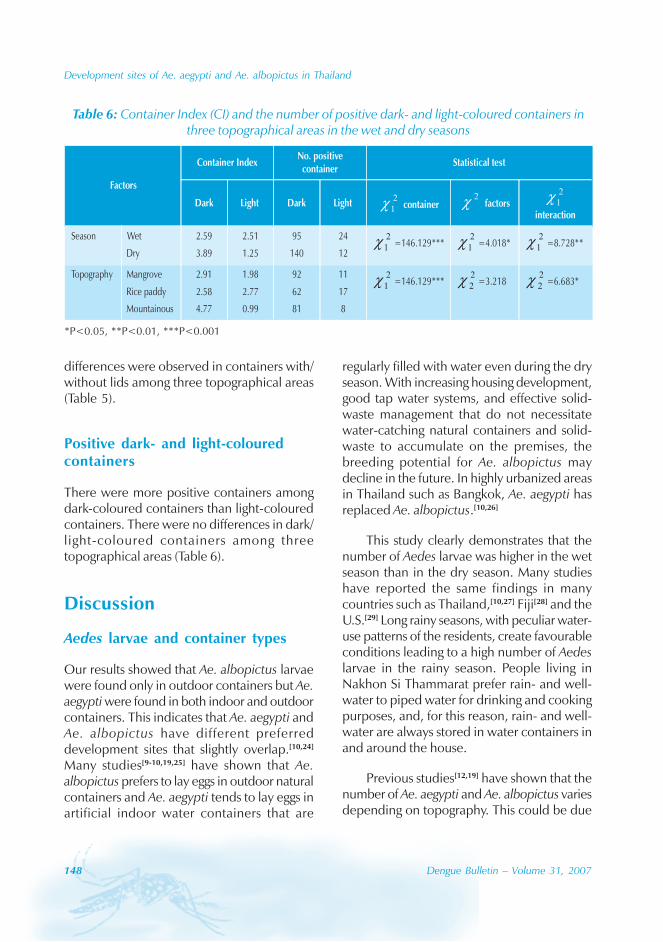
148 Dengue Bulletin – Volume 31, 2007
Development sites of Ae. aegypti and Ae. albopictus in Thailand
differences were observed in containers with/without lids among three topographical areas(Table 5).
Positive dark- and light-colouredcontainers
There were more positive containers amongdark-coloured containers than light-colouredcontainers. There were no differences in dark/light-coloured containers among threetopographical areas (Table 6).
Discussion
Aedes larvae and container types
Our results showed that Ae. albopictus larvaewere found only in outdoor containers but Ae.aegypti were found in both indoor and outdoorcontainers. This indicates that Ae. aegypti andAe. albopictus have different preferreddevelopment sites that slightly overlap.[10,24]
Many studies[9-10,19,25] have shown that Ae.albopictus prefers to lay eggs in outdoor naturalcontainers and Ae. aegypti tends to lay eggs inartificial indoor water containers that are
regularly filled with water even during the dryseason. With increasing housing development,good tap water systems, and effective solid-waste management that do not necessitatewater-catching natural containers and solid-waste to accumulate on the premises, thebreeding potential for Ae. albopictus maydecline in the future. In highly urbanized areasin Thailand such as Bangkok, Ae. aegypti hasreplaced Ae. albopictus.[10,26]
This study clearly demonstrates that thenumber of Aedes larvae was higher in the wetseason than in the dry season. Many studieshave reported the same findings in manycountries such as Thailand,[10,27] Fiji[28] and theU.S.[29] Long rainy seasons, with peculiar water-use patterns of the residents, create favourableconditions leading to a high number of Aedeslarvae in the rainy season. People living inNakhon Si Thammarat prefer rain- and well-water to piped water for drinking and cookingpurposes, and, for this reason, rain- and well-water are always stored in water containers inand around the house.
Previous studies[12,19] have shown that thenumber of Ae. aegypti and Ae. albopictus variesdepending on topography. This could be due
Table 6: Container Index (CI) and the number of positive dark- and light-coloured containers inthree topographical areas in the wet and dry seasons

Development sites of Ae. aegypti and Ae. albopictus in Thailand
Dengue Bulletin – Volume 31, 2007 149
to the different needs for water-storagecontainers in topographic types. There was notap water available in mangrove areas; peoplein this area have to store water for drinkingand washing in large water containers, especiallyin the dry season. Our results showed thatthere were more Aedes larvae in the mangroveareas than in other areas in the dry season.However, our results showed that there werehigher numbers of Aedes larvae in mountainousareas than in other areas in the wet season.This could be because there was a higheramount of rainfall in mountainous areas duringthe wet season. Waste products such as cans,bottles, tyres, etc., might collect rain-watercreating many suitable larval development sitesfor Aedes larvae in the area. In addition, therewere many rubber plantations around thehuman dwellings in the mountainous areas. Itis a common practice to suspend tappingtemporarily in rubber plantations leading toaccumulation of rainwater in sap-collectingcontainers and small shallow ponds betweenrows of rubber trees.[30] Therefore, thesewould provide an ideal developmental site forAedes mosquitoes, especially Ae. albopictus.Sumodan’s[31] study showed that Ae. albopictuslaid eggs in sap-collecting containers and theireggs could fully develop into mature adultsduring that period.
Positive indoor and outdoorcontainers
Our results support previous findings inKhonkean, Thailand,[32] and Pattani, Thailand,[11]
that the number of positive containers washigher in outdoor than indoor containers.However, Kittayapong and Strickman[33] foundthat containers placed inside houses had morelarvae than those placed outdoors and undereaves or in the bathroom. Most indoor water-storage containers are cleaned regularly oneto three times per week. This kind of routine
cleaning practice helps to eliminate Aedeslarvae in the indoor containers, especially Ae.aegypti larvae.
Positive earthen and plasticcontainers
Our results showed that the number ofpositive containers was higher among earthencontainers than among plastic containers. Thisresult supports Luemoh et al.’s[11] study thatthere were a lower number of Aedes larvae inplastic water containers, both with lids andwithout lids than clay or cement containers.This could be for two possible reasons. First,people in Nakhon Si Thammarat prefer to storetheir water in earthen containers than plasticcontainers. This may lead to mosquitoes thatprefer to oviposit eggs in earthen watercontainers over time. Second, previous studiesshow that female mosquitoes prefer to ovipositin darker coloured and less transparentcontainers over lighter coloured and moreopaque containers. Earthen containers usuallyare dark brown colour and completely dark,while plastic containers are more often lightcoloured and transmit light. Third, temperaturesin earthen containers tend to vary less than inplastic containers. This might promote fullmosquito larval development more than inhighly variable temperatures.
Positive containers with andwithout lids
Our results support previous findings[11,27,34] thatthe number of positive containers was higherin containers without lids than containers withlids. This could be for two possible reasons.First, lids could prevent gravid mosquitofemales from ovipositing in water storagecontainers. From a behavioural study,[27] Ae.aegypti females actively sought the narrow gaps

150 Dengue Bulletin – Volume 31, 2007
Development sites of Ae. aegypti and Ae. albopictus in Thailand
between the lid and the lip of the watercontainers to reach oviposition sites, althoughthe number of eggs deposited was less incontainers with lids. Second, lids may preventleaf litter, insects and organic materials fallinginto the water containers. These watercontainers with lids will have fewer nutrientsavailable. High nutrient water is a signal of goodfood in terms of quality and quantity known toattract ovipositioning females.[35-37] Therefore,water containers with lids might attract lessovipositioning females due to less nutrientsbeing available for mosquito larvae requiredduring larval development.
Positive dark- and light-colouredcontainers
A significant amount of research has been doneon colour cues for oviposition behaviour inmosquitoes.[38] For example, Toxorhynchites r.septentroinalis, Tx. r. rutilus, Tx. moctezuma andTx. Amboinensis prefer to oviposit in black ratherthan white containers.[38-41] In addition,Yonoviak’s[42] study investigated how the colourof water containers affects mosquito diversityand abundance and found that black- and red-
coloured artificial treeholes attracted highermosquito species diversity and abundance thangreen-coloured artificial containers. Our resultssupport previous findings that the number ofAedes larvae was higher in dark-colouredcontainers. Therefore, in order to minimize thenumber of Aedes larvae in water storagecontainers, people might use light-colouredwater storage containers in their houses.
Acknowledgements
This work was supported in part by the ThailandResearch Fund through the Royal Golden JubileePh.D. Programme (Grant No. PHD/0201/2548), the Institute for the Promotion ofTeaching Science and Technology, andCXKURUE, the Institute of Research andDevelopment, Walailak University. We thankJohn A. Endler and David Harding for theircomments on the previous versions of thismanuscript. We also thank the 11.2 Vector-Borne Disease Control Centre, Nakhon SiThammarat, for mosquito larval identificationand students from Yothinbamrung School forfield surveys in this study.
References
[1] Hay SI, Myers MF, Burke DS, Vaughn DW,Endy T, Ananda N, Shanks GD, Snow RW,Rogers DJ. Etiology of interepidemic periodsof mosquito-borne disease. Proc Natl Acad SciUSA. 2000 Aug 1; 97(16): 9335-9.
[2] Stephenson I, Roper J, Fraser M, Nicholson K,Wiselka M. Dengue fever in febrile returningtravellers to a UK regional infectious diseasesunit. Travel Med Infect Dis. 2003 May; 1(2):89-93.
[3] National Institute of Communicable Diseases.Investigation & control of outbreaks denguefever and dengue haemorrhagic fever. New
Delhi: Ministry of Health and Family Welfare,2001.
[4] Jacobs M, Levin M. An improved endothelialbarrier model to investigate denguehaemorrhagic fever. J Virol Methods. 2002 Jul;104(2): 173-85.
[5] Morrison AC, Astete H, Chapilliquen F,Ramirez-Prada C, Diaz G, Getis A, Gray K, ScottTW. Evaluation of a sampling methodologyfor rapid assessment of Aedes aegyptiinfestation levels in Iquitos, Peru. J MedEntomol. 2004 May; 41(3): 502-10.

Development sites of Ae. aegypti and Ae. albopictus in Thailand
Dengue Bulletin – Volume 31, 2007 151
[6] Thongrungkiat S, Jirakanjanakit N,Apiwathnasorn C, Prummongkol S, SamungY. Comparative susceptibility to oral infectionwith dengue viruses among local strains ofAedes aegypti (Diptera: Culicidae) collectedat different seasons of the year. J Vector Ecol.2003 Dec; 28(2): 166-70.
[7] Ali M, Wagatsuma Y, Emch M, Breiman RF.Use of a geographic information system fordefining spatial risk for dengue transmission inBangladesh: role for Aedes albopictus in anurban outbreak. Am J Trop Med Hyg. 2003Dec; 69(6): 634-40.
[8] Winter PE, Yuill TM, Udomsakdi S, Gould D,Nantapanich S, Russell PK. An insular outbreakof dengue hemorrhagic fever. I. Epidemiologicobservations. Am J Trop Med Hyg. 1968 Jul;17(4): 590-9.
[9] Thavara U, Tawatsin A, Phan-Urai P, NgamsukW, Chansang C, Liu M, Li Z. Dengue vectormosquitoes at a tourist attraction, Ko Samui,in 1995. Southeast Asian J Trop Med PublicHealth. 1996 Mar; 27(1): 160-3.
[10] Thavara U, Tawatsin A, Chansang C, Kong-ngamsuk W, Paosriwong S, Boon-Long J,Rongsriyam Y, Komalamisra N. Larvaloccurrence, oviposition behavior and bitingactivity of potential mosquito vectors of dengueon Samui Island, Thailand. J Vector Ecol. 2001Dec; 26(2): 172-80.
[11] Luemoh A, McNeil D, Kuning M. Waterconsumption and distribution of denguelarvae in Pattani villages. Songklanakarind MedJ. 2003; 21(3): 209-16.
[12] Wongkoon S, Jaroensutasinee M,Jaroensutasinee K. Larval Infestations of Aedesaegypti and Ae. albopictus in Nakhon SiThammarat, Thailand. Dengue Bull. 2005; 29:169-75.
[13] Thavara U, Tawatsin A, Chompoosri J.Evaluation of attractants and egg-layingsubstrate preference for oviposition by Aedesalbopictus (Diptera: Culicidae). J Vector Ecol.2004 Jun; 29(1): 66-72.
[14] Gould DJ, Yuill TM, Moussa MA, SimasathienP, Rutledge LC. An insular outbreak of dengue
hemorrhagic fever. 3. Identification of vectorsand observations on vector ecology. Am J TropMed Hyg. 1968 Jul; 17(4): 609-18.
[15] Russell PK, Yuill TM, Nisalak A, Udomsakdi S,Gould DJ, Winter PE. An insular outbreak ofdengue hemorrhagic fever. II. Virologic andserologic studies. Am J Trop Med Hyg. 1968Jul; 17(4): 600-8.
[16] Russell PK, Gould DJ, Yuill TM, Nisalak A, andWinter PE. Recovery of Dengue-4 viruses frommosquito vectors and patients during anepidemic of Dengue Haemorrhagic Fever. AmJ Trop Med Hyg. 1969; 18: 580–3.
[17] Lester PJ, Pike AJ. Container surface area andwater depth influence the populationdynamics of the mosquito Culex pervigilans(Diptera: Culicidae) and its associatedpredators in New Zealand. J Vector Ecol. 2003Dec; 28(2): 267-74.
[18] Promprou S, Jaroensutasinee M,Jaroensutasinee K. Climatic factors affectingdengue haemorrhagic fever incidence insouthern Thailand. Dengue Bull. 2005; 29:41-8.
[19] Preechaporn W, Jaroensutasinee M,Jaroensutasinee K. The larval ecology of Aedesaegypti and Ae. albopictus in threetopographical areas of southern Thailand.Dengue Bull. 2006; 30: 204-13.
[20] Scott TW, Chow E, Strickman D, KittayapongP, Wirtz RA, Lorenz LH, Edman JD. Blood-feeding patterns of Aedes aegypti (Diptera:Culicidae) collected in a rural Thai village. JMed Entomol. 1993 Sep; 30(5): 922-7.
[21] Kalra NL, Kaul SM, Rastogi RM. Prevalence ofAedes aegypti and Aedes albopictus vectors ofdengue and dengue haemorrhagic fever innorth, north-East and central India. DengueBull. 1997; 21: 84-92.
[22] Eshita Y, Kurihara T. Studies on the habitats ofAedes albopictus, Aedes riversi in theSouthwestern part of Japan. Japn J Sanit Zool.1978; 30: 181-6.

152 Dengue Bulletin – Volume 31, 2007
Development sites of Ae. aegypti and Ae. albopictus in Thailand
[23] Rattanarithikul R, Panthusiri P. Illustrated keysto the medically important mosquitoes ofThailand. Southeast Asian J Trop Med PublicHealth. 1994; 25 Suppl 1: 1-66.
[24] Gould DJ, Mount GA, Scanlon JE, Ford HR,Sullivan MF. Ecology and control of denguevectors on an island in the Gulf of Thailand. JMed Entomol. 1970 Aug 25; 7(4): 499-508.
[25] Eamchan P, Nisalak A, Foy HM, ChareonsookOA. Epidemiology and control of dengue virusinfections in Thai villages in 1987. Am J TropMed Hyg. 1989 Jul; 41(1): 95-101.
[26] Hammon WM, Rudnick A, Sather GE. Virusesassociated with epidemic hemorrhagic feversof the Philippines and Thailand. Science. 1960Apr 15; 131: 1102-3.
[27] Strickman D, Kittayapong P. Dengue and itsvectors in Thailand: calculated transmissionrisk from total pupal counts of Aedes aegyptiand association of wing-length measurementswith aspects of the larval habitat. Am J TropMed Hyg. 2003 Feb; 68(2): 209-17.
[28] Raju AK. Community mobilization in Aedesaegypti control programme by sourcereduction in Peri-Urban district of Lautoka,Viti Levu, Fiji Islands. Dengue Bull. 2003; 27:149-55.
[29] Hoeck PA, Ramberg FB, Merrill SA, Moll C,Hagedorn HH. Population and parity levels ofAedes aegypti collected in Tucson. J VectorEcol. 2003 Jun; 28(1): 65-73.
[30] Dieng H, Mwandawiro C, Boots M, MoralesR, Satho T, Tuno N, Tsuda Y, Takagi M. Leaflitter decay process and the growthperformance of Aedes albopictus larvae(Diptera: Culicidae). J Vector Ecol. 2002 Jun;27(1): 31-8.
[31] Sumodan PK. Potential of rubber plantationsas breeding source for Aedes albopictus inKerala, India. Dengue Bull. 2003; 27: 197-8
[32] Chareonsook O, Setsaputra S, Singklang K,Yawa T, Purahung S, Suwankiri P. Prevalenceof Aedes mosquito in big cement jars and rainwater tanks. Commun Dis J. 1985; 11: 247–63.
[33] Kittayapong P, Strickman D. Distribution ofcontainer-inhabiting Aedes larvae (Diptera:Culicidae) at a dengue focus in Thailand. JMed Entomol. 1993 May; 30(3): 601-6.
[34] Schneider JR, Morrison AC, Astete H, Scott TW,Wilson ML. Adult size and distribution of Aedesaegypti (Diptera: Culicidae) associated withlarval habitats in Iquitos, Peru. J Med Entomol.2004 Jul; 41(4): 634-42.
[35] Wilton DP. Oviposition site selection by thetree-hole mosquito, Aedes triseriatus (Say). JMed Entomol. 1968 Jun 10; 5(2): 189-94.
[36] Beehler J, Lohr S, Defoliart G. Factorsinfluencing oviposition in Aedes triseriatus(Diptera: Culicidae). Great Lakes Entomol.1992; 25: 259-64.
[37] Paradise CJ, Kuhn KL. Interactive effects of pHand leaf litter on a shredder, the scirtid beetle,Helodes pulchella, inhabiting tree-holes.Freshwater Biol. 1999; 41: 43-9.
[38] Collins LE, Blackwell A. Electroantennogramstudies of potential oviposition attractants forToxorhynchites moctezuma and Toxorhynchitesamboinensis mosquitoes. Physiol Entomol.1998; 23: 214-9.
[39] Slaff ME, Reilly JJ, Crans WJ. Colonization ofthe predaceous mosquito, Toxorhynchitesrutilus septentroinalis (Dyar and Knab). Proc NJ Mosq Contr Assoc. 1975; 62: 146-8.
[40] Hilburn LR, Willis NL, Seawright JA. An analysisof preference in the color of oviposition sitesexhibited by female Toxorhynchites. rutilus inthe laboratory. Mosq News. 1983; 43: 302-6.
[41] Linley JR. Laboratory tests of the effects of p-cresol and 4-methylcyclohexanol onoviposition by three species of Toxorhynchitesmosquitoes. Med Vet Entomol. 1989 Oct; 3(4):347-52.
[42] Yanoviak SP. Container color and locationaffect macroinvertebrate community structurein artificial treeholes in Panama. FloridaEntomol. 2001; 84: 265–71.

Dengue Bulletin – Volume 31, 2007 153
Behavioural responses of deltamethrin-and permethrin-resistant strains of Aedes aegypti
when exposed to permethrin in anexcito-repellency test system
P. Paeporna , K. Supaphathoma, S. Sathantriphopa, T. Chareonviriyaphapb
and R. Yaicharoenc
aChemical Control Section, Department of Medical Sciences, Ministry of Public Health,Nonthaburi 11000, Thailand
bDepartment of Entomology, Faculty of Agriculture, Kasetsart University, Bangkok 10900, Thailand
cDepartment of Parasitology, Faculty of Medical Technology, Mahidol University, Bangkok 10700, Thailand
Abstract
This study compared the behavioural avoidance responses of the permethrin-resistant and deltamethrin-resistant strains of Aedes aegypti, a primary vector of dengue haemorrhagic fever (DHF) in Thailand. Thebackground of biochemical-based resistance mechanism assay of these two strains revealed a significantincrease of esterase activity and monooxygenase levels when compared with a laboratory-susceptiblestrain. Glutathione-S-transferase activity was found to increase only in the permethrin-resistant strain.The DNA sequence of knockdown resistance (kdr) mutation in the voltage-gated sodium channel (IIS6region) was determined but the leucine to phenylalanine amino acid substitution, which is commonlyassociated with resistance to pyrethroids in many insect species, was not found in either strain. Thebehavioural escape response of both contact irritancy and non-contact repellency when exposed topermethrin at standard field dose (0.25 g/m2) was observed by using an excito-repellency test chamber.The results showed that in contact trials, the permethrin-resistant strain showed a lower irritancyresponse when compared with the deltamethrin-resistant strain. This was probably due to the higherlevels of resistance to this insecticide for the permethrin resistance strain. For the repellency test by non-contact trials, the response was not significantly different between the two strains. This may be becausethe repellency effect was much weaker than that of the irritancy effect. This study indicated that thebehavioural response of mosquitoes differs according to different pyrethroid compounds and to thephysiological resistance mechanism of the mosquitoes. However, further work is necessary to understandhow these responses are mediated.
Keywords: Behavioural response; Aedes aegypti; Insecticide resistance; Excito-repellency test chamber.
E-mail: [email protected]

154 Dengue Bulletin – Volume 31, 2007
Behavioural responses of deltamethrin- and permethrin-resistant strains of Aedes aegypti
Introduction
Insecticide resistance in Aedes aegypti, a primaryvector of dengue haemorrhagic fever (DHF),has been reported in many parts of the world.In Thailand, fenitrothion, deltamethrin andpermethrin are the primary insecticides usedin thermal fogging and ultra-low volume spraysfor the control of adult Aedes aegypti mosquitoesduring DHF outbreaks. The widespread use ofinsecticides has led to selective insecticideresistance in vector mosquitoes reducing theefficacy of control measures.
Insecticide resistance can be grouped withinfour categories:[1] (i) behavioural resistance wherethe insect behaviour is modified to avoid contactwith the insecticide; (ii) penetration resistance,where the composition of the insect’sexoskeleton becomes modified in a way thatinhibits insecticide penetration; (iii) target-siteinsensitivity, where the chemical site of actionfor the insecticide is modified, limiting the affinityof the insecticide for the target site and reducingits ability to disrupt the function of the targetsite; and (iv) metabolic resistance, wheredetoxification enzymes rapidly break down theinsecticide so it is no longer toxic to mosquitoes.Although several reports of insecticide resistancein Ae. aegypti have been published, most of themdescribe biochemical resistance. The impact ofthese compounds on Ae. aegypti in terms ofbehavioural resistance has not been studied.
In this paper, permethrin anddeltamethrin-resistant strains were examinedfor target-site insensitivity and for behaviouralresistance.
Materials and methods
Test populations
A susceptible colony of Ae. aegypti from theDepartment of Medical Sciences, Ministry of
Public Health, Thailand, was used as alaboratory-susceptible strain.
Two selected strains of Ae. aegypti, i.e.one deltamethrin-selected strain which showedsignificant elevation of esterase andmonooxygenase activity as compared withlaboratory-susceptible strains, and the other thepermethrin-resistant strain which had asignificant increase in glutathione-S-transferaseactivity when compared with the deltamethrin-resistant strain and the susceptible strain. Thedetails to obtain these two resistant strainswere described elsewhere.[2]
PCR and sequencing of partialgenomic DNA of sodium channelgene
Adult female mosquitoes, from permethrin-and deltamethrin-selected strains were usedfor the detection of point mutation in thesegment 6 of domain II (IIS6) of sodiumchannel gene. [3] The mosquitoes werehomogenized individually and total genomicDNA was extracted by phenol chloroformmethod.[4] The DNA pellet was diluted in TEbuffer before using as PCR template. PCR wasperformed by using AegF, (5’ AAC TTA CTCATT TCC ATC ATG G3’)[5] and Dg2, (5’ GC (T/G/A) AT (C/T) TT (A/G) TT(G/A/T/C) GT (G/A)TC (G/A) TT (G/A) TC 3’) primers. Nested PCRwas carried out using primers Dg1, (5’ TGG AT(T/C/A) G (A/C) (A/G) (T/A) (C/G) (A/C/T) ATGTGG GA (T/C) TG 3’) and Dip2, (5’TTG GACAAA AGC AA (G/A) GCT AAG 3’) primers.[6]
The PCR products were subjected to DNAsequencing (Bioservice Unit, Thailand).
Behavioural resistance test
Tests were carried out to compare thebehavioural responses of the two strains of Ae.aegypti exposed to 0.25 g/m2 permethrin by

Behavioural responses of deltamethrin- and permethrin-resistant strains of Aedes aegypti
Dengue Bulletin – Volume 31, 2007 155
using excito-repellency test chambers (Figures1 and 2) developed by Chareonviriyaphap etal.[7] Generally, behavioural responses, orinsecticide avoidance, can be categorized ascontact irritancy and non-contact repellency.Irritancy results from physical contact with
chemical-treated surfaces whereas repellencyis a response from a distinct distance withoutphysical contact with insecticides.
The test system consists of two treated testchambers and two paired control chambers. Priorto the exposure, mosquitoes were starved forat least 24 hours. Twenty-five female mosquitoeswere carefully transferred into each of the 4test chambers (Figure 3). Mosquitoes wereallowed a 3-min resting period to permitadjustment to the chamber conditions.Observations for behavioural responses weretaken at 1 min intervals for 30 mins. After eachtest was completed, the number of dead orknockdown specimens was recorded separatelyfor each exposure chamber, paired controlchamber and external holding cage, which wasthe receiving box connected to the exit portalfor collecting escaped mosquitoes. Escapedspecimens and those remaining inside thechambers, for each treatment, were held
Figure 2: Excito-repellency test chamber 2
3. Contact control4. Contact treatment
1. Non-contact control2. Non-contact
treatment
Repellencysystem
Irritabilitysystem
Figure 1: Excito-repellency test chamber 1
1
2
3
4
5
6
7
1. Rear door cover
2. Plexiglass panel with rubbered door
3. Plexiglass holding frame
4. Screened inner chamber
5. Outer chamber
6. Front door
7. Exit portal

156 Dengue Bulletin – Volume 31, 2007
Behavioural responses of deltamethrin- and permethrin-resistant strains of Aedes aegypti
separately in small holding containers with cottonpads soaked in 10% sugar solution as food.Mortality counts were made at the end of the24-hour observation period. The test wasreplicated 4 times for each insecticide strain andeach arm (permethrin treatment, control).
Results
PCR and sequencing of thepartial genomic DNA ofsodium channel gene
To investigate whether alteration of the sodiumchannel gene was involved in resistancemechanism, a 400 bp of genomic DNAsequence comprising the S6 trans-membranesegment of domain II (IIS6) in the sodiumchannel gene was PCR-amplified andsequenced. The result revealed that the
leucine (L, CTC) to phenylalanine (F, TTC)substitution was not found from themosquitoes sequenced, although allmosquitoes showed the highest resistanceagainst insecticide from each strain.
Behavioural test
Even though this was the preliminary study, asthe sample size was not sufficient for statisticalevaluation, it can be seen that in contact trialsthe deltamethrin-resistant strain showed highercontact escape response (27.3% within 30mins) compared with the permethrin-selectedstrain. The difference in escape response wasnot observed in non-contact trials. Thepercentage of the mosquitoes remaining in theexposure chambers under contact and non-contact conditions are shown in Figures 4 and5 respectively.
Figure 3: Excito-repellency test chamber 3
External holdingcage
Exposure-testedchamber
Behavioural responses of deltamethrin- and permethrin-resistant strains of Aedes aegypti
Dengue Bulletin – Volume 31, 2007 157
Figure 4: Escape pattern of permethrin- and deltamethrin-resistant populations incontact trial with 0.25 g/m2 permethrin
60
70
80
90
100
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30
Time (minutes)
%M
osq
uit
oes
rem
ainin
gin
exp
osu
rech
amb
ers
Permethrin-resistant Deltamethrin-resistant Lab-susceptable
Figure 5: Escape pattern of permethrin- and deltamethrin-resistant populations innon-contact trial with 0.25 g/m2 permethrin
60
70
80
90
100
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30
Time (minutes)
%M
osq
uito
esre
mai
nin
gin
exp
osu
rech
amb
ers
Permethrin-resistant Deltamethrin-resistant Lab-susceptable
158 Dengue Bulletin – Volume 31, 2007
Behavioural responses of deltamethrin- and permethrin-resistant strains of Aedes aegypti
The percentage of mortality of the escapedmosquitoes was a little lower than thoseremaining in the test chamber for the two strains,both in contact and non-contact trials (Table).
Discussion
PCR and sequencing of thepartial genomic DNA ofsodium channel gene
The results suggested that other mechanisms,such as the increase of detoxification enzymes,may be involved in the resistance, or anothersite of point mutation which is not commonlyassociated with resistance to pyrethroids inmany insect species may be involved in Ae.aegypti resistance. This requires further studies.
Behavioural test
In our experiment, the study was designed toknow the contact and non-contact responsesof the permethrin-resistant strain and thedeltamethrin-resistant strain of mosquitoes topermethrin. So, no bait or similar attractant wasused to avoid any confusion in the results.
From this study the lower irritancy responsein the permethrin-selected strain was probablydue to its higher levels of resistance to thisinsecticide, which may have influenced theescape response because of increased durationof exposure time with this insecticide. Similarresults were observed with Anopheles gambiaeby Chandre et al.[8] These authors demonstratedthat resistant mosquitoes could tolerate higher-dose permethrin and stayed longer thansusceptibles.
For a repellency test by non-contact trials,the response was not significantly differentbetween the two strains. This may be due torepellency effect which was much weaker thanthat of the irritancy effect.
Even though the permethrin-resistant strainof Ae. aegypti, which has increased metabolicdetoxification enzymes involved in itsresistance, does not change the target-siteinsensitivity but it showed a behaviouralresistance to permethrin. This study providesimportant information because the biochemicalresistance in mosquito vectors can mediatetheir behavioural response to that insecticidewhen applied as a space spray or on surface-treated areas for adult control.
Table: Escape response and percent mortalities of permethrin- and deltamethrin-resistantpopulations when exposed to 0.25 g/m2 in contact and non-contact trials
Treatedchamber
Controlchamber
% Mortality
Treatmentchamber
Controlchamber
Testcondition
StrainNo.
tested%
escapedNo.
tested%
escapedEscaped
Didnot
escapeEscaped
Didnot
escape
Contact Permethrin-resistant
Deltamethrin-resistant
Lab-susceptible
99
99
96
11.1
27.3
25.0
98
97
93
7.1
10.3
43.0
0
1
8.3
1
2
44.8
0
0
1.1
1
3.1
5.4
Non-contact Deltamethrin-resistant
Lab-susceptible
99
99
95
6
2
35.8
100
100
95
6
6
26.3
1
0
0
2
3.1
20
2
0
4.2
2
1
6.3

Behavioural responses of deltamethrin- and permethrin-resistant strains of Aedes aegypti
Dengue Bulletin – Volume 31, 2007 159
References
[1] Miller TA. Mechanism of resistance topyrethroids insecticides. Parasitology Today.1988; 4(7): S8-S12.
[2] Paeporn P, Supaphathom K, Srisawat R,Komalamisra N, Deesin V, Ya-umphan P,Leeming Sawat S. Biochemical detection ofpyrethroid resistance mechanism in Aedesaegypti in Ratchaburi province, Thailand. TropBiomed. 2004 Dec; 21(2): 145-51.
[3] Soderlund DM, Knipple DC. The molecularbiology of knockdown resistance to pyrethroidinsecticides. Insect Biochem Mol Biol. 2003Jun; 33(6): 563-77.
[4] Ballinger-Crabtree ME, Black WC 4th, MillerBR. Use of genetic polymorphisms detectedby the random-amplified polymorphic DNApolymerase chain reaction (RAPD-PCR) fordifferentiation and identification of Aedesaegypti subspecies and populations. Am J TropMed Hyg. 1992 Dec; 47(6): 893-901.
[5] Brengues C, Hawkes NJ, Chandre F, McCarrollL, Duchon S, Guillet P, Manguin S, Morgan
JC, Hemingway J. Pyrethroid and DDT cross-resistance in Aedes aegypti is correlated withnovel mutations in the voltage-gated sodiumchannel gene. Med Vet Entomol. 2003 Mar;17(1): 87-94.
[6] Martinez-Torres D, Chandre F, Williamson MS,Darriet F, Bergé JB, Devonshire AL, Guillet P,Pasteur N, Pauron D. Molecularcharacterization of pyrethroid knockdownresistance (kdr) in the major malaria vectorAnopheles gambiae s.s. Insect Mol Biol. 1998May; 7(2): 179-84.
[7] Chareonviriyaphap T, Prabaripai A,Sungvornyothrin S. An improved excito-repellency test chamber for mosquitobehavioral tests. J Vector Ecol. 2002 Dec; 27(2):250-2.
[8] Chandre F, Darriet F, Duchon S, Finot L,Manguin S, Carnevale P, Guillet P.Modifications of pyrethroid effects associatedwith kdr mutation in Anopheles gambiae. MedVet Entomol. 2000 Mar; 14(1): 81-8.

160 Dengue Bulletin – Volume 31, 2007
Perceived self-efficacy to plan and execute anenvironmental action plan for dengue control among
Filipino University students
Jeffrey L. Lennon
Liberty University, Department of Health Sciences and Kinesiology, Lynchburg, VA 24502, USA
Abstract
This study sought to evaluate the change in the perceived self-efficacy in students exposed to a trainingmodule with field exercise to conduct an environmental action plan for dengue control. Eighteeneducation students at a Filipino university participated in the study. A pretest-post-test design wasemployed. Evaluation was conducted through a nine-point self-efficacy scale. Field exercises wereconducted on the grounds of the university campus. The results demonstrated statistically significantincreases in perceived self-efficacy scores.
Keywords: Dengue control; Environmental action plan; Self-efficacy; Students.
E-mail: [email protected]
Introduction
Dengue fever/dengue haemorrhagic fever (DF/DHF) is at present the leading mosquito-borneviral disease worldwide.[1] Dengue especiallyaffects school-age children and youth.[2]
Schools, especially in the tropical andsubtropical dengue-endemic areas, arefrequently the breeding sites for Aedes aegypti,the carrier of the dengue virus.[3,4] Teachersand students therefore potentially provide keylinks between the schools, the students andthe community in dengue control.[5] Teachershave the potential not only to be trainers totheir students in dengue control but also canserve as role models on their school campusgrounds. In addition, teachers can serve asfacilitators for community action, especially as
their students are able to carry out denguecontrol activities, such as environmental actionplans for their own homes and neighbourhoods.
Mosquito larval control through sourcereduction of mosquito breeding sites remainsa primary emphasis in dengue control.[6] Variousinternational projects lay stress on sourcereduction for dengue control at the communitylevel and at schools.[7]
A number of studies have described themobilization of schoolchildren throughinvolvement of teachers. In the English-speaking Caribbean, teachers trained childrenas community-change agents through anenvironmental health model.[8] In Colombia,eleventh graders, after 20 hours of training,conducted community-wide dengue control

Perceived self-efficacy to plan and execute environmental plan for dengue control
Dengue Bulletin – Volume 31, 2007 161
communication. One creative approach wasthe use of calendars in classrooms byteachers. [9] In Malaysia, schoolchildrendistributed weekly worksheets tohouseholds.[10] In Puerto Rico, fourth gradershad a dengue educational booklet integratedinto class work. Their teachers received specifictraining as a part of the programme.[11] InHonduras, primary-school children, afterreceiving training by teachers, conductedhousehold mosquito control surveys. Also inthe study, parents and teachers had an increasedawareness about mosquito control. [12]
However, no study has been done to showthat teachers, or students who had beentrained, had undergone field exercises toprepare them in dengue control, let aloneexamine the participants’ confidence levelsthrough their experience.
The health behaviour theory provides aframework to conduct health education andhealth promotion programmes. [13] Self-efficacy, a construct of Social Cognitive Theory,is a powerful predictor of behaviour.[14,15]
One’s efficacy beliefs are central to thisconstruct. According to Bandura, “Perceivedself-efficacy refers to beliefs in one’scapabilities to organize and execute thecourses of action required to produce givenattainments.”[15] Self-efficacy significantlyincreased in two dengue-related studies, onein Thailand, [16] and the other in thePhilippines.[17] In Thailand, self-efficacy tocarry out source-reduction measures to reducemosquito breeding sites increased through acommunity-based programme.[16] Self-efficacyto perform dengue prevention and controlmeasures significantly increased as a result ofthe play of a board game among Filipinoelementary and high-school students.[17]
No study has yet demonstrated the use of anenvironmental action plan for teachers or
future teachers to increase self-efficacy ofmosquito larval prevention and controlmeasures to control dengue. Therefore, thisstudy sought to examine whether universityeducation students (future teachers) mayincrease their perceived self-efficacy toconduct dengue-related mosquito controlmeasures through the use of environmentalaction plan training and experiential fieldactivity to control mosquito breeding sites ina university setting. In this study, perceivedself-efficacy is operationalized to be definedas the beliefs one has in one’s capability toexecute an environmental action plan as laterdescribed in this study. This refers to the self-reported self-efficacy without validationthrough behavioural indicators.
Materials and methods
Participants
Eighteen students from the FoundationUniversity College of Education course entitled,“Personal and Community Hygiene” inDumaguete city, Philippines, a dengue-endemic city participated in the study in 2005.The student group included 14 females andfour males and had no previous exposure tothe environmental action plan. Participation inthe evaluation of the study activities wasvoluntary. All the students participated till thecompletion of the exercise. The majority ofthe students were primary-school traineeswhile the remainder were secondary-schooltrainees. The breakdown of the number of first-,second- and third-year students was seven,nine and two respectively. There were nofourth-year students, as all of them had previousexposure to the environmental action plan. Theaverage age of the students was approximately20 years.

162 Dengue Bulletin – Volume 31, 2007
Perceived self-efficacy to plan and execute environmental plan for dengue control
Materials
A nine-item self-efficacy questionnaire wasdistributed to the participants. Thisquestionnaire covered such topics asconfidence in the following: regular removalof disposable containers, cover reusablecontainers, change water in flower vases,removal of used tyres, turn over open coconutshells, participation in environmental clean-up,as well as identification of mosquito breedingsites. It was previously distributed to elementaryand high school student trainees,[17] as well asstudent-teachers.[18] This questionnaire was pre-tested with student-teachers at a seminar.Similarly, an environmental action-plan formatwas also utilized by the students. This formatwas also previously conducted by educationstudents[19] as well as discussed in training forstudent-teachers.[18]
Procedures
Prior to attending a teaching session on theuse of an environmental action plan fordengue-related mosquito control, studentswere given the self-efficacy questionnaire. Theteaching session opened with a learner-centred, problem-posing self-discovery, action-oriented module addressing mosquito larvalcontrol, and thus the need of sourcereduction. The training included visual surveyand area mapping of potential mosquitobreeding sites, followed by instruction oncreation of environmental action plans for thecontrol of potential mosquito breeding sites.The action plan steps included: identificationof mosquito breeding sites, number ofmosquito breeding sites, action to be taken,how frequently the control activity should beundertaken, and by whom. The following daythe students were assigned in groups toconduct visual surveys, mapping and creationof environmental action plans in and arounddesignated buildings and areas around the
campus. Each designated campus area had itsown environmental action plan. However,there was a core set of environmentalproblems common to all environmental actionplans in this study. This included such topicsas the identification of disposable watercontainers and covering of open containers.Three days later the students received a post-test of the self-efficacy questionnaire.
Data analysis
For reliability, a Cronbach’s alpha was calculatedon the questions of the pretest questionnaireby SPSS 15.0 for Windows. The possibledifferences between the pretest and post-testself-efficacy scores were analysed by paired t-test. All t-tests were two tailed. Statisticalsignificance was set at the .05 level. Theanalyses were calculated on MS-Excel.
ResultsThe Cronbach’s alpha for reliability for the self-efficacy questionnaire was .92. This scoreindicated that the study’s results were reliable.Behavioural capability (knowledge) wasassessed through the successful completion ofall steps of the environmental action plan byall groups. This included identification of thecore environmental problem sets in allenvironmental action plans.
The results of the two-tailed paired t-testwith a critical t of 2.111 revealed a significantdifference between mean self-efficacy scores,t(17)=2.517; p<0.0222. The sample meanself-efficacy score for the post-test wassignificantly higher than the pretest (for pretest,mean=5.574, variance=2.296; for post-test,mean=6.439, variance=.419). The increaseddifference in the mean scores was .865. Thisresulted in a 15.52% increase of mean scores,from pretest to post-test.

Perceived self-efficacy to plan and execute environmental plan for dengue control
Dengue Bulletin – Volume 31, 2007 163
Discussion
This study’s results demonstrated a significantincrease in the perceived self-efficacy meanscores of dengue-related environmental controlmeasures from the pretest to the post-test. Thissuggested that the training accompanied withfield exercise to conduct the environmentalaction plan for the control of mosquito breedingsites was effective as a means to increaseperceived self-efficacy to carry outenvironmental control activities through the useof environmental action plans.
This study had its limitations in its smallsize, the absence of a control group, as well asthe absence of environmental clean-upbehavioural observations. However, sincehigher-level education students at thisuniversity had undergone training in theenvironmental action plan for dengue control,the group of students in this study remained avaluable sample of education studentspreviously unexposed to the environmentalaction plan for dengue control.
Why the concern about perceived self-efficacy or efficacy beliefs related to denguecontrol? Typically, sustained self-efficacy, and,additionally, sustained positive environmentalbehavioural change, ought to be the concern.However, before one can get to theseimportant issues it is desirable to get aprogramme’s start on a firm footing. Accordingto Bandura, “Efficacy beliefs influence goalsand aspirations. The stronger the perceived self-efficacy, the higher the goals people set forthemselves and the firmer their commitmentto them. Self-efficacy beliefs shape theoutcomes people expect their efforts toproduce.”[14] That is why Bandura stresses theimportance of self-efficacy as an indicator ofbehavioural intent. A new or special activitymay heighten enthusiasm or spark interest, but
not necessarily elevate self-efficacy. Someactivities may demonstrate initial observablebehavioural changes. However, without a highlevel of perceived self-efficacy, the sustainabilityof such activities may falter. Consequently,increasing perceived self-efficacy may beimportant before investing time, energy andactivities in a dengue control programme.
This study demonstrated that universityeducation students were able to increase theirperceived self-efficacy to perform denguecontrol activities through a single training andsimple environmental action plan activity.Education students eventually become student-teachers, and, in time, teachers. Increasingone’s perceived self-efficacy to perform specificdengue-related mosquito control activitiesduring the university course may aid futureteachers to potentially increase their skills tocarry out dengue control activities. This willput future teachers in a better position tobecome school and community leaders for thecontrol of dengue.
Acknowledgements
The author thanks the administration of theFoundation University for permission toconduct this study. Thanks are also due to DrThelma Florendo, Dean of FoundationUniversity College of Education, for hercooperation, and to Dr Paplito dela Rama forstatistical assistance. Also, thanks to Chona F.Lennon and Daniel A. Lennon for theirtechnical assistance. Comments andsuggestions from Dr David Coombs weregreatly appreciated. The support for this studyby the International Technical Assistance Groupis gratefully acknowledged. The author held afaculty appointment at the FoundationUniversity, and was course professor while thisstudy was conducted.

164 Dengue Bulletin – Volume 31, 2007
Perceived self-efficacy to plan and execute environmental plan for dengue control
References
[1] World Health Organization. Dengueprevention and control. Report by theSecretariat of Fifty-fifth World Health Assembly,AA/19, Provisional Agenda Item 13.14 dated4 March 2002.
[2] Gubler DJ. Dengue and dengue hemorrhagicfever. Clin Microbiol Rev. 1998 Jul; 11(3): 480-96.
[3] Strickman D, Kittayapong P. Dengue and itsvectors in Thailand: introduction to the studyand seasonal distribution of Aedes larvae. AmJ Trop Med Hyg. 2002 Sep; 67(3): 247-59.
[4] Kusumawathie PHD. Larval infestation ofAedes aegypti and Ae. albopictus in six typesof institutions in a dengue transmission area inKandy, Sri Lanka. Dengue Bulletin. 2005; 29:165-168.
[5] Lennon JL. Students’ perceptions aboutmosquito larval control in a dengue-endemicPhilippine City. Dengue Bulletin. 2004; 28:196-206.
[6] World Health Organization. Scientific WorkingGroup on Dengue, 3-4 April 2000, Geneva,Switzerland: Scientific Working Group onDengue, Meeting. Geneva: WHO, 2000.Document TDR/DEN/SWG/00.1.
[7] Parks WJ, Lloyd LS, Nathan MB, Hosein E,Odugleh A, Clark GG, Gubler DJ, PrasittisukC, Palmer K, San Martin J-L, Siversen SR,Dawkins Z, Renganathan E. Internationalexperiences in social mobilization andcommunication for dengue prevention andcontrol . Dengue Bul let in. 2004; 28:Suppl.1-7
[8] Nathan MB, Lloyd L, Wiltshire A. Communityparticipation in environmental managementfor dengue vector control experiences fromthe English-speaking Carribbean. DengueBulletin. 2004; 28: Suppl.13-16.
[9] Luna JE, Chain I, Hernandez J, Clark GG,Bueno A, Escalante R, Angarita S, Martinez A.Social mobilization using strategies ofeducation and communication to preventdengue fever in Bucaramanga, Colombia.Dengue Bulletin. 2004, 28: Suppl.17-21.
[10] Suhaili MR, Hosein E, Mokhtar Z, Ali N, PalmerK, Isa MM. Applying communication-for-behavioral-impact (COMBI) in the preventionand control of dengue in Johor, Bahru, Johore,Malaysia. Dengue Bulletin. 2004; 28:Suppl.39-43.
[11] Clark GG, Gubler DJ. Seda H, Perez C.Development of pilot programmes for dengueprevention in Puerto Rico: a case study.Dengue Bulletin. 2004; 28: Suppl.48-52.
[12] Avila Montes GA, Martínez M, Sherman C,Fernández Cerna E. [Evaluation of aneducational module on dengue and Aedesaegypti for schoolchildren in Honduras]. RevPanam Salud Publica. 2004 Aug; 16(2): 84-94. Spanish.
[13] Glanz K, Lewis FM, Rimer BK. Linking theory,research and practice. In Glanz K, Lewis FM,Rimer BK, eds. Health behavior and healtheducation: theory, research and practice. 2nd
edn. San Francisco: Jossey-Bass Publishers,1997. p. 19-35.
[14] Bandura A. Health promotion by socialcognitive means. Health Educ Behav. 2004 Apr;31(2): 143-64.
[15] Bandura A. Self-efficacy: the exercise of control.New York: WH Freeman and Company, 1997.p. 1-34.
[16] Therawiwat M, Fungladda W, KaewkungwalJ, Imamee N, Steckler A. Community-basedapproach for prevention and control ofdengue hemorrhagic fever in KanchanaburiProvince, Thailand. Southeast Asian J Trop MedPublic Health. 2005 Nov; 36(6): 1439-49.

Perceived self-efficacy to plan and execute environmental plan for dengue control
Dengue Bulletin – Volume 31, 2007 165
[17] Lennon JL, Coombs DW. The utility of a boardgame for dengue haemorrhagic fever healtheducation. Health Education. 2007; 107: 290-306.
[18] Lennon JL. Dengue control in the Philippines:health education strategies for dengue controlfor schools and communities. 2001,
University of Alabama at Birmingham, John J.Sparkman Center for International PublicHealth Education.
[19] Lennon JL. Knowledge of denguehaemorrhagic fever by Filipino UniversityStudents. Dengue Bulletin. 1996; 20: 82-86.

166 Dengue Bulletin – Volume 31, 2007
Short note
Dengue presenting as Guillain Barre Syndrome
Ira Shah
Department of Paediatrics, B.J. Wadia Hospital for Children, Parel, Mumbai-400012, India
E-mail: [email protected]; Tel.: 91-22-23693509
Dengue fever (DF) has been increasinglyreported among children in Mumbai. Thecommon clinical and laboratory features seenin dengue-infected children are: fever,thrombocytopenia, elevated serumtransaminases, elevated partial thromboplastintime (PTT), hypotension, vomiting ,haemoconcentration, leukopenia andhepatomegaly.[1] Though dengue virus (DENV)is a member of the Flaviviridae group of viruseswhich include a number of neutropic viruses,[2]
immune-mediated central nervous system’s(CNS) involvement following dengue infectionhas been rarely reported. Reports of only fourchildren of Guillain Barre Syndrome (GBS) dueto DENV have been published.[3-6] We reporta 2-year-old boy who presented with GBS dueto dengue infection.
Case report
A 2-year-old boy born of non-consanguineousmarriage presented in the monsoons withweakness of lower limbs since day 1. Theweakness was progressive and now had eveninvolved the upper limb. He had fever withdiarrhoea four days back. There was no historyof any immunization or injections taken in therecent past. He had been immunized till age.
On examination, he was conscious andoriented. He had hypotonia with reducedpower in all four limbs (lower limb involvedmore as compared to the upper limb). He haddiaphragmatic weakness though respirationwas not laboured. Superficial and deep tendonreflexes were absent though gag reflex waspresent. There was no cranial nerveinvolvement or sensory involvement. Othersystemic examination was normal. Thus, hewas diagnosed as a case of ascendingprogressive motor polyneuropathy, most likelyGuillain Barre Syndrome. His haemogramrevealed thrombocytopenia [Platelet =1 04 000/cumm] and normal haemoglobin andWBC count. His serum electrolytes liverfunction tests and renal function tests werenormal. HIV ELISA and HBsAg were negative.Nerve conduction velocity showed absent ‘F’waves in lower limbs suggestive of axonalradiculopathy. Dengue IgM by Panbio Kit waspositive. [1.33 AI (Positive = > 0.9 AI)]. Thechild was given intravenous immunoglobulin(IVIg) (2 gm/kg) following which his weaknessimproved and he had regained full power inupper limb and power of grade 3 in lowerlimbs after 15 days of IVIg . Histhrombocytopenia resolved within seven daysof presentation. Thus, this child had denguepolyneuropathy.

Dengue presenting as Guillain Barre Syndrome
Dengue Bulletin – Volume 31, 2007 167
Discussion
Dengue is a common arboviral infection affectingpatients in South-East Asia, including India, andSouth America. CNS involvement is known withthis viral infection though its actual frequency isunknown.[2,4] The common CNS manifestationsare altered sensorium and seizures[2,7] andthought to be due to prolonged denguehaemorrhagic fever (DHF) with fluidextravasation, cerebral oedema, hyponatremia,liver failure, renal failure and direct neurotropiceffect of dengue virus.[2,8] Immune-mediatednervous system involvement has been describedin the form of GBS and post-infectious-disseminated encephalitis rarely.[4] GBS is a post-infectious polyradiculopathy known to occurpost-gastrointestinal infection withCampylobacter jejuni and other infective agents,like Mycoplasma, human cytomegalovirus(CMV), Ebstein-Barr virus and herpes virus. Itrarely occurs due to demyelination post-dengueinfection. The nerve injury in GBS is mediatedby immunological mechanisms. DHF anddengue shock syndrome (DSS) occur due toimmunopathological mechanisms wherebysecondary exposure to dengue viruses lead to amore severe disease. Immune enhancement isthought to play a major role in the pathogenesis
of enhanced dengue infection, leading to cross-reacting antibodies and demyelination.[2,7] GBS,in the form of polyradiculopathy of a primarilydemyelinating nature with an associated axonalcomponent,[3,4] has been found as was seen inour patient who had absent “F” waves in lowerlimbs suggestive of axonal radiculopathy.
DHF and DSS are characterized bythrombocytopenia. We found that 92.3% ofpatients infected with DF havethrombocytopenia.[9] Similarly, our patient hadfever prior to the episode of GBS andthrombocytopenia at the time of presentationin the monsoon season (a time when denguetends to peak in Mumbai), which made ussuspect dengue-associated GBS which wasconfirmed by positive dengue IgM test.
Previous reports have found completerecovery of dengue-associated GBS withintravenous immunoglobulin (IVIg) as was alsofound in our patient, suggesting that cross-reacting antibodies from a previous dengueinfection may be responsible for the illness.
Thus, in conclusion, one should rule outdengue-associated GBS in areas endemic fordengue whether, or not they have other featuresof the disease.
References
[1] Cam BV, Fonsmark L, Hue NB, Phuong NT,Poulsen A, Heegaard ED. Prospective case-control study of encephalopathy in childrenwith dengue hemorrhagic fever. Am J Trop MedHyg 2001 Dec; 65(6): 848-51.
[2] Sulekha C, Kumar S, Philip J. Guillain-Barresyndrome following dengue fever. IndianPediatr. 2004 Sep; 41(9): 948-50.
[3] Palma-da Cunha-Matta A, Soares-Moreno SA,Cardoso-de Almeida A, Aquilera-de Freitas V,Carod-Artal FJ. Neurological complicationsarising from dengue virus infection. Rev Neurol.2004 Aug 1-15; 39(3): 233-7. Spanish.
[4] Esack A, Teelucksingh S, Singh N. The Guillain-Barré syndrome following dengue fever. WestIndian Med J. 1999 Mar; 48(1): 36-7.
[5] Sainte-Foie S, Niel L, Moreau JP, Ast-Ravallec,Chippaux A. A case of polyradiculoneuritisassociated with dengue in a patient native toFrench Guiana. Bull Soc Pathol Exot. 1993;86(2): 117-8; discussion 119. French.
[6] Cardosa MJ. Dengue haemorrhagic fever:questions of pathogenesis. Curr Opin InfectDis. 2000 Oct; 13(5): 471-475.

168 Dengue Bulletin – Volume 31, 2007
Dengue presenting as Guillain Barre Syndrome
[7] Lum LC, Lam SK, Choy YS, George R, Harun F.Dengue encephalitis: a true entity? Am J TropMed Hyg. 1996 Mar; 54(3): 256-9.
[8] Chaturvedi UC, Dhawan R, Khanna M, MathurA. Breakdown of the blood-brain barrierduring dengue virus infection of mice. J GenVirol. 1991 Apr; 72 (Pt 4): 859-66.
[9] Shah I, Deshpande GC, Tardeja PN. Outbreakof dengue in Mumbai and predictive markersfor dengue shock syndrome. J Trop Pediatr.2004 Oct; 50(5): 301-5.

Dengue Bulletin – Volume 31, 2007 169
Short note
An unusual case of brainstem encephalitis
Ira Shah
Department of Paediatrics, B.J. Wadia Hospital for Children, Parel, Mumbai-400012, India
E-mail: [email protected]; Tel.: 91-22-23693509
Dengue is the commonest arboviral infectionthat affects mankind. Infection by any of thefour serological types of dengue viruses (DENV-1 to 4) cause dengue fever (DF), denguehaemorrhagic fever (DHF) or dengue shocksyndrome (DSS). In recent years, there havebeen reports of neurological complicationsassociated with dengue in Asia and SouthAmerica, though their frequency still remainsunknown.[1-7] The encephalopathy associatedwith dengue infection has been postulated tobe due to prolonged DHF with fluidextravasation, cerebral oedema, hyponatremia,liver failure, renal failure and a possible directneurotropic effect of dengue virus.[1,4,8,9] Thoughcentral nervous system (CNS) manifestationshave been reported in adults, reports in childrenare rare and have only been reported in threestudies,[1,3,5] and none from India. We studieda 10-year-old girl who presented with brainstemencephalitis, thrombocytopenia, elevated livertransaminases and positive dengue IgM withcomplete resolution of her symptoms withinthree days of the onset of the disease.
Case report
A 10-year-old girl born of non-consanguineousmarriage presented in monsoons with fever sinceeight days, projectile vomiting since five days,
altered sensorium, inability to stand and squintsince day 1. Her younger brother was alsoadmitted with fever and was diagnosed to havedengue. She had no convulsions. Onexamination, she was conscious and orientedand had meningeal signs with positiveMacewan’s sign. Her vital parameters werenormal. Fundus showed papilledema. She hadright lateral palsy and tone and power werenormal. Deep tendon reflexes were brisk andright planter was extensor. She had ataxia withintention tremors. Other systemic examinationwas normal. Thus, she was suspected to havebrainstem encephalitis or posterior fossa tumorwith raised ICT. Her haemogram revealedthrombocytopenia (platelet 84 000/cumm) withnormal haemoglobin and WBC count. Livertransaminases were deranged (SGOT = 121 IU/L), SGPT = 60 IU/L); serum electrolytes, renalfunction tests and blood sugar was normal. MRIbrain did not show any space-occupying lesionand showed only inflammatory changes in rightmaxillary, ethmoid and sphenoid sinuses. WithMRI ruling out posterior fossa tumor,cerebrospinal fluid (CSF) analysis was done whichshowed aseptic meningitic picture (3polymorphs, 19 lymphocytes/cumm; 52 mg/dl– sugar; proteins = 10.3 mg %). CSF culturewas negative. In view of thrombocytopenia,deranged liver enzymes and meningitis and herbrother suffering from dengue, her dengue IgM

170 Dengue Bulletin – Volume 31, 2007
An unusual case of brainstem encephalitis
by capture ELISA was done, which was positive{1.2 OD units (positive = > 0.9 OD units)}.Blood culture and leptospira IgM ELISA werenegative. Thus, this child was diagnosed as acase of dengue brainstem encephalitis. She wastreated with Mannitol and all her symptomsresolved within three days. Her repeat plateletcount and liver transaminases were normal.Thus, here is a case of dengue presenting asbrainstem encephalitis.
Discussion
Dengue virus is a member[2] of the Flaviviridaegroup of viruses, which include a number ofneurotropic viruses such as Japanese encephalitisvirus, St. Louis encephalitis virus and tick-borneencephalitis virus. In recent years, we and othershave reported neurological manifestations inpatients with DHF and DSS characterized bydepression of consciousness with normal CSFanalysis[1,10] and complete recovery in survivors.In fact, in a study published by us, we foundthat almost 48.7% of patients had alteredsensorium on presentation with higherpredominance in the DSS group as comparedto the DHF group (77.8% vs 25%, P = 0.0035).However, most of the patients had additionalfeatures such as bleeding manifestations,hepatomegaly and serositis. Isolated CNSinvolvement is a rarity as seen in our patient.
The encephalopathy was thought to be dueto cerebral oedema, anoxia, haemorrhage,hyponatremia, hepatic failure, microcapillaryhaemorrhage and release of toxic substances.[1]
However, recent reports have demonstrated a
possible direct neutropic effect of dengue virus[9]
and localized invasion of the CNS. Similarlocalized CNS manifestation in the form ofbrainstem encephalitis with raised intracranialtension and CNS leukocorrhia was seen in ourpatient. A case report by Lum et al.[4] has isolateddengue virus (DENV-2 and 3) from the CSF ofaffected patients, suggesting direct invasion ofthe brain and neurovirulent properties of thedengue virus.
Cam et al.[1] have determined that patientswith dengue encephalitis have significantlyelevated liver enzymes with cerebral oedema.Similar findings were noted in our patient wherethe child had elevated liver transaminases andcomplete CNS recovery with Mannitol. In fact,in our patient, the presence ofthrombocytopenia with history of her brothersuffering from dengue made us suspect dengueencephalitis. In a previous study, we had foundthat 92.3% of patients affected by dengue hadthrombocytopenia, and this combination ofthrombocytopenia, elevated liver transaminasesand fever is very characteristic of dengue inMumbai.[10]
Thus, it would be wiser to investigatepatients with encephalitis and encephalopathyin dengue-endemic areas for dengue infection,irrespective of the fact whether they have otherfeatures of the disease or not.
In conclusion, it can be stated that dengueencephalitis is rare but neutropism of the virusis known. It can masquerade as other types ofviral encephalitis, but its clinical course andoutcome is usually favourable.
References
[1] Cam BV, Fonsmark L, Hue NB, Phuong NT,Poulsen A, Heegaard ED. Prospective case-control study of encephalopathy in childrenwith dengue hemorrhagic fever. Am J Trop MedHyg. 2001 Dec; 65(6): 848-51.
[2] Solomon T, Dung NM, Vaughn DW, Kneen R,Thao LT, Raengsakulrach B, Loan HT, Day NP,Farrar J, Myint KS, Warrell MJ, James WS,Nisalak A, White NJ. Neurological

An unusual case of brainstem encephalitis
Dengue Bulletin – Volume 31, 2007 171
manifestations of dengue infection. Lancet.2000 Mar 25; 355(9209): 1053-9.
[3] Kankirawatana P, Chokephaibulkit K,Puthavathana P, Yoksan S, Apintanapong S,Pongthapisit V. Dengue infection presentingwith central nervous system manifestation. JChild Neurol. 2000 Aug; 15(8): 544-7.
[4] Lum LC, Lam SK, Choy YS, George R, Harun F.Dengue encephalitis: a true entity? Am J TropMed Hyg. 1996 Mar; 54(3): 256-9.
[5] Palma-da Cunha-Matta A, Soares-Moreno SA,Cardoso-de Almeida A, Aquilera-de Freitas V,Carod-Artal FJ. Neurological complicationsarising from dengue virus infection. Rev Neurol.2004 Aug 1-15; 39(3): 233-7. Spanish.
[6] Hommel D, Talarmin A, Deubel V, Reynes JM,Drouet MT, Sarthou JL, Hulin A. Dengueencephalitis in French Guiana. Res Virol. 1998Jul-Aug; 149(4): 235-8.
[7] Ramos C, Sánchez G, Pando RH, Baquera J,Hernández D, Mota J, Ramos J, Flores A,Llausás E. Dengue virus in the brain of a fatalcase of hemorrhagic dengue fever. J Neurovirol.1998 Aug; 4(4): 465-8.
[8] Miagostovich MP, Ramos RG, Nicol AF,Nogueira RM, Cuzzi-Maya T, Oliveira AV,Marchevsky RS, Mesquita RP, Schatzmayr HG.Retrospective study on dengue fatal cases. ClinNeuropathol. 1997 Jul-Aug; 16(4): 204-8.
[9] Chaturvedi UC, Dhawan R, Khanna M, MathurA. Breakdown of the blood-brain barrierduring dengue virus infection of mice. J GenVirol. 1991 Apr; 72 (Pt 4): 859-66.
[10] Shah I, Deshpande GC, Tardeja PN. Outbreakof dengue in Mumbai and predictive markersfor dengue shock syndrome. J Trop Pediatr.2004 Oct; 50(5): 301-5.

172 Dengue Bulletin – Volume 31, 2007
Short note
Dengue presenting as viral myocarditis
Ira Shah
Department of Paediatrics, B.J. Wadia Hospital for Children, Parel, Mumbai-400012, India
E-mail: [email protected]; Tel.: 91-22-23693509
Dengue fever (DF) is endemic in India andcauses dengue fever, its severe form denguehaemorrhagic fever (DHF) and dengue shocksyndrome (DSS). Hypotension due tointravascular depletion due to capillary leak iscommon in DHF and DSS. However,myocardial dysfunction is also seen with DHF/DSS and could be responsible for hypotensionand shock.[1] Cardiac involvement in the formof decreased left ventricular performance[2]
and arrhythmias[3,4] are known. However,isolated myocarditis has rarely been seen withdengue virus.[5] We report an 11-year-old boywho presented with viral myocarditis due todengue infection, with complete recoverywithin three months.
Case report
An eleven-year-old boy born of non-consanguineous marriage presented withvomiting since two days and fever since dayone. He had no skin rash or bleeding from anysite. On examination, he had tachycardia (heartrate = 110/min) with hypotension (BP = 85/50 mm of Hg) with prolonged capillary refilland bilateral basal crepitations. There were nosigns of dehydration. Other systemicexamination was normal. He was immediatelytreated with Dobutamine and IV fluids. He was
suspected to have viral myocarditis withcardiogenic shock. His investigations showedlymphopenia with thrombocytopenia(haemoglobin = 12.9 gm/dl, WBC = 7200/cumm, absolute lymphocyte count = 720/cumm, platelet count = 1 27 000/cumm) withabsent C-reactive protein and no growth onblood culture. His serum CPK levels wereelevated (344 IU/L) and CPK-MB fraction wasalso elevated {71 units/L (normal = upto 25units/L)}. Serum LDH was also elevated (972IU/L). His renal function tests, liver functiontests and blood sugar were normal.Echocardiography showed left ventriculardilatation with systolic dysfunction andfractional shortening of 22%. His dengue IgMby ELISA was positive {0.95 AI (Positive = >0.9 AI)} and leptospira IgM and peripheralsmear for malaria were negative. He was thusdiagnosed as a case of dengue myocarditis.Intravenous immunoglobulin (2 gm/kg) wasgiven following which his left ventricularfunctions improved with fractional shorteningof 31% and ejection fraction of 62%. He wasgradually tapered off the ionotropic support anddischarged. On follow up after three months,he was asymptomatic and a repeatechocardiography showed normal leftventricular dimensions and functions withfractional shortening of 37% and ejectionfraction of 74%.

Dengue presenting as viral myocarditis
Dengue Bulletin – Volume 31, 2007 173
Discussion
Viral myocarditis may lead to cardiogenic shockdue to fulminant cardiac failure, recurrentwheezy episodes (mistaken as bronchialasthma), bronchiolitis and rhythmdisturbance.[5] Serum enzymes such as SGOT,CPK, LDH are elevated and echocardiographyshows left ventricular dilatation with globaldyskinesia.[5] Our patient also presented withcardiogenic shock due to cardiac failure, hadelevated CPK and LDH, and echocardiographyshowed left ventricular dilatation with systolicdysfunction confirming the diagnosis ofmyocarditis. Viral myocarditis is known to occurcommonly with Coxsackie virus B1 and B4 andrarely with dengue virus[5,6]. Our patient had apositive dengue IgM test, confirming thediagnosis of dengue myocarditis. Cardiac
dysfunction in the form of reduced ejectionfraction and decreased end-diastolic volumeare noted in patients with dengue.[2] Similarechocardiographic findings were noted in ourpatient. Viral myocarditis may respondfavourably to routine anti-failure measures,steroid therapy and intravenousimmunoglobulins (IVIG), with completereversal of echocardiographic findings withinmonths. [5] Similarly, our patient had afavourable response to IVIG and anti-failuremeasures and had complete recovery afterthree months of the onset of symptoms.
In conclusion, viral myocarditis may be amanifestation of dengue, though relativelyuncommon as an isolated feature, and itsresponse to therapy is favourable.
References
[1] Wali JP, Biswas A, Chandra S, Malhotra A,Aggarwal P, Handa R, Wig N, Bahl VK. Cardiacinvolvement in Dengue Haemorrhagic Fever.Int J Cardiol. 1998 Mar 13; 64(1): 31-6.
[2] Khongphatthanayothin A, Suesaowalak M,Muangmingsook S, Bhattarakosol P,Pancharoen C. Hemodynamic profiles ofpatients with dengue hemorrhagic fever duringtoxic stage: an echocardiographic study.Intensive Care Med. 2003 Apr; 29(4): 570-4.
[3] Horta Veloso H, Ferreira Júnior JA, Braga dePaiva JM, Faria Honório J, Junqueira BelleiNC, Vicenzo de Paola AA. Acute atrialfibrillation during dengue hemorrhagic fever.Braz J Infect Dis. 2003 Dec; 7(6): 418-22.
[4] Promphan W, Sopontammarak S,Pruekprasert P, Kajornwattanakul W,Kongpattanayothin A. Dengue myocarditis.Southeast Asian J Trop Med Public Health. 2004Sep; 35(3): 611-3.
[5] Gowrishankar K, Rajajee S. Variedmanifestations of viral myocarditis. Indian JPediatr 1994 Jan-Feb; 61(1): 75-80.
[6] Kabra SK, Juneja R, Madhulika, Jain Y, SinghalT, Dar L, Kothari SS, Broor S. Myocardialdysfunction in children with denguehaemorrhagic fever. Natl Med J India. 1998Mar-Apr; 11(2): 59-61.

174 Dengue Bulletin – Volume 31, 2007
Short note
Defrost-water-collection trays of refrigerators –A potential breeding habitat of Aedes aegypti in dengue-
and chickungunya-infested areasof southern India
R. Srinivasan , T. Mariappan and P. Jambulingam
Vector Control Research Centre (Indian Council of Medical Research), Puducherry 605006, India
E-mail: [email protected]
Arthropod-borne viral diseases like denguefever (DF) and its severe form, denguehaemorrhagic fever (DHF), and chikungunyafever are known to occur throughout the tropicsthe world over. These diseases are increasingalarmingly in the South-East Asian countries.[1]
Dengue epidemics are being reported fromdifferent parts of India since 1963.[2,3,4,5] Forthe control of dengue outbreaks, managementof Aedes aegypti, the vector species, throughsource reduction remains the mainstay. Hence,an assessment of the breeding sources of thismosquito is essential in order to devise anddevelop a suitable control strategy.
Ae. aegypti is mainly a domestic speciesof urban areas, but it has now spread to rural,suburban and urban areas as well. This speciesprefers to breed in man-made containersholding stored water or rainwater in domesticand peri-domestic environments.
During epidemic investigations inPuducherry (earlier known as Pondicherry) andCuddalore during 2004-05, a rare phenomenonof Ae. aegypti breeding was observed in the
defrost trays of refrigerators in both these towns.To understand the importance of this habitat inthe proliferation of this vector mosquito duringdifferent seasons, a total of three surveys werecarried out: one each during the south-westmonsoon season (June-August 2004), north-east monsoon period (October-December2004) and during summer (March-May 2005).In houses ranging from 746–792, the numberof refrigerators examined ranged from 29–335.Water was emptied into an enamel trayemploying either a siphon or pipette andabsolute count of larvae and pupae, if any, wasmade, brought to laboratory, reared to adultsand identified to species, following a standardkey.[8] The breeding of Aedes mosquitoes wasfound in 39 trays (1.6–8.4%) in Puducherry and8 trays (4.8–6.9%) in Cuddalore (Table). Aedesaegypti was the only species obtained from thepupae collected from defrost-water-collectiontrays in refrigerators from both the areas. Thelarvae and pupae recorded were significantlyhigh (p>0.05) during the summer period in boththe towns as compared to the other twoseasons.
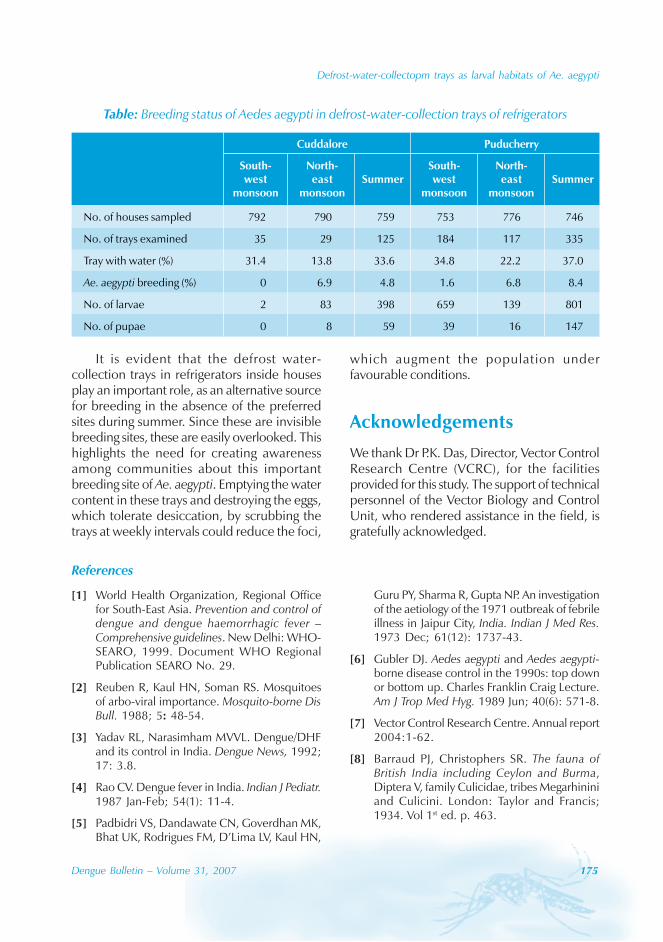
Defrost-water-collectopm trays as larval habitats of Ae. aegypti
Dengue Bulletin – Volume 31, 2007 175
It is evident that the defrost water-collection trays in refrigerators inside housesplay an important role, as an alternative sourcefor breeding in the absence of the preferredsites during summer. Since these are invisiblebreeding sites, these are easily overlooked. Thishighlights the need for creating awarenessamong communities about this importantbreeding site of Ae. aegypti. Emptying the watercontent in these trays and destroying the eggs,which tolerate desiccation, by scrubbing thetrays at weekly intervals could reduce the foci,
Table: Breeding status of Aedes aegypti in defrost-water-collection trays of refrigerators
which augment the population underfavourable conditions.
Acknowledgements
We thank Dr P.K. Das, Director, Vector ControlResearch Centre (VCRC), for the facilitiesprovided for this study. The support of technicalpersonnel of the Vector Biology and ControlUnit, who rendered assistance in the field, isgratefully acknowledged.
References
[1] World Health Organization, Regional Officefor South-East Asia. Prevention and control ofdengue and dengue haemorrhagic fever –Comprehensive guidelines. New Delhi: WHO-SEARO, 1999. Document WHO RegionalPublication SEARO No. 29.
[2] Reuben R, Kaul HN, Soman RS. Mosquitoesof arbo-viral importance. Mosquito-borne DisBull. 1988; 5: 48-54.
[3] Yadav RL, Narasimham MVVL. Dengue/DHFand its control in India. Dengue News, 1992;17: 3.8.
[4] Rao CV. Dengue fever in India. Indian J Pediatr.1987 Jan-Feb; 54(1): 11-4.
[5] Padbidri VS, Dandawate CN, Goverdhan MK,Bhat UK, Rodrigues FM, D’Lima LV, Kaul HN,
Guru PY, Sharma R, Gupta NP. An investigationof the aetiology of the 1971 outbreak of febrileillness in Jaipur City, India. Indian J Med Res.1973 Dec; 61(12): 1737-43.
[6] Gubler DJ. Aedes aegypti and Aedes aegypti-borne disease control in the 1990s: top downor bottom up. Charles Franklin Craig Lecture.Am J Trop Med Hyg. 1989 Jun; 40(6): 571-8.
[7] Vector Control Research Centre. Annual report2004:1-62.
[8] Barraud PJ, Christophers SR. The fauna ofBritish India including Ceylon and Burma,Diptera V, family Culicidae, tribes Megarhininiand Culicini. London: Taylor and Francis;1934. Vol 1st ed. p. 463.
Cuddalore Puducherry
South- North- South- North-west east Summer west east Summer
monsoon monsoon monsoon monsoon
No. of houses sampled 792 790 759 753 776 746
No. of trays examined 35 29 125 184 117 335
Tray with water (%) 31.4 13.8 33.6 34.8 22.2 37.0
Ae. aegypti breeding (%) 0 6.9 4.8 1.6 6.8 8.4
No. of larvae 2 83 398 659 139 801
No. of pupae 0 8 59 39 16 147

176 Dengue Bulletin – Volume 31, 2007
Short note
Point-of-care diagnostics for dengue, chikungunya,Japanese encephalitis and West Nile virus infection
Subhash C. Arya and Nirmala Agarwal
Sant Parmanand Hospital, 18 Alipore Road, Delhi-110054
E-mail: [email protected]
The global resurgence of dengue in otherwisenaive locations has been associated withconcurrent dissemination of identical vector-borne viral diseases. Concurrent infection bydengue virus (DENV) and chikungunya virus(CHIKV) has been known for severaldecades.[1] Nevertheless, recent global CHIKVdissemination[2] or its local re-emergence[3] aftera gap has been intriguing . Increasedintercontinental travel has blown up the ghostof the traditional endemic foci of CHIKV andhas resulted in CHIKV patients being found inthe United States.[4] Moreover, there couldeven be coincidental episodes of the Japaneseencephalitis virus (JEV) infection. During theearly 1940s, there were dengue outbreaks inGuam, followed, during 1947, by theconcurrent epidemics of mumps virus and JEV.Serological investigations in children aged 10years or less during 1953 had revealedneutralizing antibody to JEV, and DENV-1 and -2 among those born before 1947.[5]
In India, during 1997, concurrent infectionwas detected in Haryana state in the north ofthe country. In a follow-up study of a dengueoutbreak, out of 30 serum samples collected,eight cases were found positive for dengue IgMantibodies, two were positive for all the threeinfections, viz. DENV, JEV and WNV, while one
sample was positive for two infections, viz. JEVand West Nile virus (WNV).[6]
There has been no chemotherapy availablefor patients with JEV, DENV, CHIKV or WNV.Such cases require appropriate clinicalmanagement and public health response. Duringthe initial stage of illness the clinical presentationsare ambiguous and are accompanied by viremia.These cases are highly infectious with their bloodteeming with viral RNA. If bitten by the mosquitovector, they would contribute to diseasedissemination. Moreover, they are likely to beoffered empirical doses of antibiotics by theclinicians. Any point-of-care indication about JEV,DENV, CHIKV or WNV would be imperativefrom the clinical and public health perspective.That would necessitate initiating appropriate anti-vector measures attuned to vector biology.Vectors transmitting JE and dengue/chikungunya/West Nile fever would be controlled by entirelydifferent strategies.
A commercial diagnostic kit for the fielddiagnosis of Japanese encephalitis (JE) antiviralIgM has been evaluated recently.[7] Rather thana multi-step, time-consuming format, the on-the-spot point-of-care diagnostic would addressthe ground realities in the JE-endemic areas.Without prejudice to the conventional enzyme-

Dengue, chikungunya, Japanese encephalitis and West Nile virus
Dengue Bulletin – Volume 31, 2007 177
linked immunosorbent assay (ELISA), a simpleron-the-spot diagnostic would assist health careproviders better. Obviously, immunoglobulin M(IgM)-based assays would be adequate for bothJE and chikungunya. Nevertheless, for dengue,additional markers would be essential. Theimmunological response during the secondaryor tertiary dengue episodes is secondary ratherthan a primary one. The initial immunologicalresponse would be towards IgG productionrather than IgM. Dengue non-structural antigen,NS1, has been reported to be better than IgMin cases of primary or secondary infections.[8]
Immunochromatographic assay kits for a point-of-care dengue IgG and IgM detection havebeen popular for a while, namely, the PanbioDengue Duo IgM and IgG Rapid Strip test, and
the Bio-Check Plus Dengue G/M Cassette Test(Brittney). Recent introduction of a rapid,immunochromatographic kit for chikungunya hasbeen a positive feature.[9]
International organizations including theWorld Health Organization, the Programme forAppropriate Technology in Health (PATH) andthe Centers for Disease Control and Prevention(CDC) would be obliged to encouragestandardization of a concurrent point-of-carediagnostic for JE, dengue, chikungunya andWest Nile fever. A multifaceted but cost-effective rapid assay format would be a valuablearmour for clinicians and public healthpersonnel to diagnose dengue, chikungunya,Japanese encephalitis and West Nile fever.
References
[1] Myers RM, Carey DE. Concurrent isolationfrom patient of two arboviruses, Chikungunyaand dengue type 2. Science. 1967 Sep 15;157(794): 1307-8.
[2] Simon F, Parola P, Grandadam M, FourcadeS, Oliver M, Brouqui P, Hance P, Kraemer P, AliMohamed A, de Lamballerie X, Charrel R,Tolou H. Chikungunya infection: an emergingrheumatism among travelers returned fromIndian Ocean islands. Report of 47 cases.Medicine (Baltimore). 2007 May; 86(3): 123-37.
[3] AbuBakar S, Sam IC, Wong PF, MatRahim N,Hooi PS, Roslan N. Reemergence of endemicChikungunya, Malaysia. Emerg Infect Dis. 2007Jan; 13(1): 147-9.
[4] Centers for Disease Control and Prevention(CDC). Update: chikungunya fever diagnosedamong international travelers–United States,2006. MMWR Morb Mortal Wkly Rep. 2007Mar 30; 56(12): 276-7.
[5] Hammon WM, Tigertt WD, Sather GE, BergeTO, Meiklejohn G. Epidemiologic studies ofconcurrent virgin epidemics of Japanese Bencephalitis and of mumps on Guam, 1947-1948, with subsequent observations including
dengue, through 1957. Am J Trop Med Hyg.1958 Jul; 7(4): 441-67.
[6] Katyal R, Bhardwaj M, Harit AK, Sharma SK,Kumar K, Gill KS. Dengue, Japaneseencephalitis and West Nile flaviviral infectionsdetected during a dengue outbreak in Sonepatdistrict, Haryana state, India. Dengue Bulletin.2004 Dec; 28: 24-28.
[7] Jacobson JA, Hills SL, Winkler JL, MammenM, Thaisomboonsuk B, Marfin AA, GibbonsRV. Evaluation of three immunoglobulin Mantibody capture enzyme-linkedimmunosorbent assays for the diagnosis ofJapanese encephalitis. Am J Trop Med Hyg.2007 Jul; 77(1): 164-8.
[8] Kumarasamy V, Chua SK, Hassan Z, WahabAH, Chem YK, Mohamad M, Chua KB.Evaluating the sensitivity of a commercialdengue NS1 antigen-capture ELISA for earlydiagnosis of acute dengue virus infection.Singapore Med J. 2007 Jul; 48(7): 669-73.
[9] CTK Biotech. Onsite Rapid Test for thedetection of Chikungunya fever. San Diego, CA:CTK Biotech. (http://www.ctkbiotech.com/Default.aspx?tabid=118 – accessed 11 August2008).

178 Dengue Bulletin – Volume 31, 2007
Short note
Role of helplines for dissemination of informationduring an outbreak of dengue fever in Delhi, India,
in 2006: An experience
Bir Singh, Anil Goswami, Neena Chawla and Sosamma Shyam
Centre for Community Medicine, All India Institute of Medical Sciences, New Delhi-110029, India
Outbreaks of dengue fever (DF) and denguehaemorrhagic fever (DHF) have been reportedin various parts of India during the last fourdecades.[1] Delhi has been endemic for DF/DHFfor quite some time and still maintains highvulnerability due to a large number of internalmigrants and international tourists visiting theCapital and the extensive breeding potential thecity provides to Aedes aegypti mosquitoes.[2]
Delhi recorded epidemics of dengue feverduring 1967, 1970, 1982, 1988, 1991, 1992and 1996.[1,3] During the 1988 outbreak of DF/DHF, about 33% mortality was reported amongchildren admitted in hospitals,[4] while during1996, 10 252 cases were hospitalized and 423deaths were recorded.[3] This epidemic peakedin September when an Aedes aegypti larvalhouse index (HI) of 43.70% was recorded.
In 2001, there were 3306 cases with 53deaths; in 2002, the number was 1926 casesand 33 deaths. In 2003, there was a largeoutbreak with 12 754 cases and 215 deaths.[5]
The 2006 outbreak started in August andpeaked in October. As of end-November 2006,it was estimated that 10 344 cases and 162deaths due to severe forms of dengue (i.e.DHF/DSS) had taken place.[6]
As is generally the case, there waswidespread panic and apprehension in the general
public about the outbreak. Anybody having feverrushed to a health facility or the local privatepractitioner thinking that the fever may be dueto dengue. Reports of chikungunya fever fromthe states of Tamil Nadu and Kerala in south Indiamade the situation worse. A few cases ofchikungunya were reported in Delhi too.
Though some public informationadvertisements issued by the MunicipalCorporation of Delhi, the New Delhi MunicipalCouncil and the Ministry of Health and FamilyWelfare appeared in the national and regionaldailies and on radio and television fordissemination of information on dengue feverand chikungunya fever and their prevention andmanagement, there was a clear gap in meetingthese information needs – especially on themanagement of fever. As a result, a largeattendance of patients with fever andsuspected dengue fever was recorded at majorhospitals of the city, particularly the All IndiaInstitute of Medical Sciences (AIIMS) whichbore the brunt of this patient load.[7]
AIIMS is considered to be a premiermedical institute in India. Though envisagedto be a tertiary care/referral hospital when itwas established in 1956, but owing to itsgrowing reputation for providing quality health

Role of helplines during the 2006 Delhi outbreak
Dengue Bulletin – Volume 31, 2007 179
care, it has been attracting a large number ofpatients in its outpatient department (OPD)and Emergency services. For example, in 2005,the attendance in its OPD was a whopping16 30 479 patients.[8] This included a largepercentage of patients who don’t qualify to becalled as “referred patients”.
In the dengue outbreak of 2006, eventhough the Government of Delhi had madearrangements for the management of fever/suspected dengue cases in about 33 hospitalsof the city, a large number of patients – about1000 to 1200 per day – reported at AIIMSstarting from the first week of October to mid-November,[7] stretching its human resourcesand infrastructural facilities to the maximum.
One important method of containment ofdengue outbreak was considered to bedissemination of correct information to thegeneral pubic on the preventive aspects ofthese diseases as well as on the managementof fevers. Allaying the fear and panic setting inthe general public was the real need. AIIMStook several important steps in this direction.Apart from distributing 18 000 informationbooklets, 10 000 handbills and pamphlets andputting up banners, it started two helplines toaugment public education efforts:
• Telephone helpline: It was a mannedtelephone service that functioned from10 AM to 10 PM every day – includingholidays. A trained public healtheducator, supported by a qualifiedmedical doctor and a public healthspecialist, manned this service. Thishelpline (Phone no. 011-26589712)was subsequently converted into a 24-hour helpline in 2007.
• Internet (E-mail) helpline: To helpthose who are net savvy, an Internet(E-mail) helpline ([email protected]) was also started. Questions
received were responded to within atime frame of 6-8 hours.
Observations
Both these services were operated for sixweeks starting from the first week of October2006 to the second week of November 2006.By mid-November, the outbreak was almostover. Some of the highlights of these helplinesare as follows:
• No. of calls/queries received: Duringthe six weeks of the operations, thetelephone helpline received 378 calls,while 79 queries were received on theInternet.
• Geographical distribution of thecallers: 263(69.5%) of the callers werefrom the Capital and 98(26%) werefrom outside Delhi. Rest of the callersdid not reveal their place of residence.Residents of south and west Delhidominated (50%) among those whocalled from Delhi. The details aregiven in Table 1.
Table 1: Geographical distribution oftelephone callers
Area No. of callers (%)
South Delhi 84(22.22%)
West Delhi 105(27.77%)
East Delhi 46(12.16%)
North Delhi 19(5.02%)
Central Delhi 09(2.38%)
Outside of Delhi 98(25.92%)
Didn’t specify 17(4.49%)
Total 378(100%)
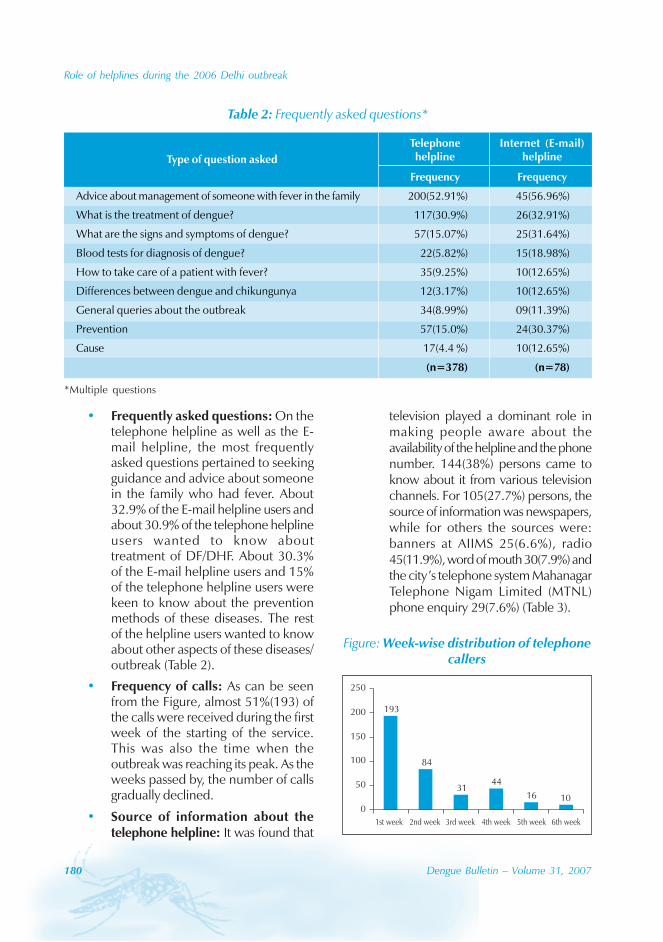
180 Dengue Bulletin – Volume 31, 2007
Role of helplines during the 2006 Delhi outbreak
• Frequently asked questions: On thetelephone helpline as well as the E-mail helpline, the most frequentlyasked questions pertained to seekingguidance and advice about someonein the family who had fever. About32.9% of the E-mail helpline users andabout 30.9% of the telephone helplineusers wanted to know abouttreatment of DF/DHF. About 30.3%of the E-mail helpline users and 15%of the telephone helpline users werekeen to know about the preventionmethods of these diseases. The restof the helpline users wanted to knowabout other aspects of these diseases/outbreak (Table 2).
• Frequency of calls: As can be seenfrom the Figure, almost 51%(193) ofthe calls were received during the firstweek of the starting of the service.This was also the time when theoutbreak was reaching its peak. As theweeks passed by, the number of callsgradually declined.
• Source of information about thetelephone helpline: It was found that
television played a dominant role inmaking people aware about theavailability of the helpline and the phonenumber. 144(38%) persons came toknow about it from various televisionchannels. For 105(27.7%) persons, thesource of information was newspapers,while for others the sources were:banners at AIIMS 25(6.6%), radio45(11.9%), word of mouth 30(7.9%) andthe city’s telephone system MahanagarTelephone Nigam Limited (MTNL)phone enquiry 29(7.6%) (Table 3).
Table 2: Frequently asked questions*
*Multiple questions
193
84
3144
16 100
50
100
150
200
250
1st week 2nd week 3rd week 4th week 5th week 6th week
Figure: Week-wise distribution of telephonecallers
Telephone Internet (E-mail)Type of question asked helpline helpline
Frequency Frequency
Advice about management of someone with fever in the family 200(52.91%) 45(56.96%)
What is the treatment of dengue? 117(30.9%) 26(32.91%)
What are the signs and symptoms of dengue? 57(15.07%) 25(31.64%)
Blood tests for diagnosis of dengue? 22(5.82%) 15(18.98%)
How to take care of a patient with fever? 35(9.25%) 10(12.65%)
Differences between dengue and chikungunya 12(3.17%) 10(12.65%)
General queries about the outbreak 34(8.99%) 09(11.39%)
Prevention 57(15.0%) 24(30.37%)
Cause 17(4.4 %) 10(12.65%)
(n=378) (n=78)

Role of helplines during the 2006 Delhi outbreak
Dengue Bulletin – Volume 31, 2007 181
Discussion
It is evident that these types of helplines do fulfilla felt need of the community in a crisis situation.
Table 3: Source of information about thetelephone helpline (first response)
One of the major problems in an outbreak of acommunicable disease that can be fatal is thepanic and apprehension that sets in in the generalpopulace – mainly due to lack of adequate andeasily available and technically correctinformation. As most of the helpline-userswanted to know about “What to do and whereto go because someone in their family hadfever”, it is important that all available means ofmass communication should be effectivelyutilized by health care managers to provide correctinformation. Helplines, especially on telephone,can be very effective because of their easy,personalized and reassuring approach. This cansupplement other methods of disseminatinginformation effectively. Hence, such helplinesmust be operated in crisis situations. Locallanguage should be the mode of communicationon this type of service.
References
[1] Yadava RL, Narsimham MVVL. Dengue/denguehaemorrhagic fever and its control in India.Dengue Newsletter. 1992; 17: 3-8.
[2] Kalra NL, Kaul SM, Rastogi RM. Prevalence ofAedes aegypti and Aedes albopictus – Vectorsof dengue and dengue haemorrhagic fever innorth, north-east and central India. DengueBulletin .1997; 21: 84-92.
[3] Kaul SM, Sharma RS, Sharma SN, PanigrahiN, Phukan PK, Shiv Lal. Preventing dengueand DHF: The role of entomologicalsurveillance. J Comm Dis. 1998; 30: 187-192.
[4] Kabra SK, Verma IC, Arora NK, Jain Y, Kalra V.Dengue haemorrhagic fever in children inDelhi. Bull World Health Organ. 1992; 70(1):105-8.
[5] Ministry of Health and Family WelfareGovernment of India. Annual Report 2005-06. New Delhi: MOH&FW, 2006. p. 59-74.
[6] Personal communication: PL Joshi, Director,National Vector-borne Diseases ControlProgramme, Government of India, 24November 2006.
[7] All India Institute of Medical Sciences. Pressreleases on dengue updates from AIIMS fromOctober 2-18 October 2006 and publishedin the Times of India and other dailies fromNew Delhi.
[8] All India Institute of Medical Sciences. AnnualReport 2005-06: New Delhi, AIIMS, 2006.
Source No. (%)
Television 144(38.0%)
Newspapers 105(27.7%)
MTNL phone enquiry 29(7.6%)
Banners at AIIMS 25(6.6%)
Word of mouth 30(7.9%)
Radio 45 (11.9%)
Total 378 (100%)

182 Dengue Bulletin – Volume 31, 2007
Book review
Report of the WHO consultation on integrated vectormanagement (IVM)
WHO Headquarters, Geneva, Switzerland, 1-4 May 2007
WHO/CDS/NTD/VEM/2007.1
The consultation on integrated vectormanagement (IVM) took place at theheadquarters of the World Health Organization(WHO) in Geneva, Switzerland, from 1 to 4 May2007. Its purpose was to bring together expertsin IVM from country, regional and global levelsto advance the development and promotion ofthe IVM approach, as set out in the Globalstrategic framework for integrated vectormanagement,[1] by reviewing current status anddeveloping a practical strategic plan for the nextthree years.
The four key recommendations of theconsultation were:
(1) In order to fully embed IVM in all controlprogrammes dealing with vector-bornediseases, concerted efforts should bemade, through advocacy and socialmobilization, to articulate the need forand benefits of IVM and to mobilizeresources for implementation.
(2) The current human resource andinfrastructure base for IVM is inadequateto meet increasing demands. This mustbe addressed urgently, through capacity-building, and both needs-based trainingand institutional development, to ensurethose trained receive continuedmentoring and support, and can functionin systems where their skills are mosteffectively applied.
(3) Action must be evidence-based, so it isessential to intensify applied researchand monitoring and evaluation to assessthe impact and performance of vectorcontrol, both to refine implementationof IVM and to provide data for advocacy.
(4) Develop an institutional framework topromote, further develop, implementand scale up IVM, including nationallegislative frameworks.
The main conclusion of the meeting wasthat now is the time to tap the preventive powerof vector control, given the serious risks ofincreasing transmission of vector-borne diseasesrelated to climate change, population movementand environmental degradation, and the majoropportunities for financial support.
The next steps will be to promote andimplement the strategic plan, propose WHOappoint a panel of experts on IVM to guide itsdevelopment, organize a follow-up meeting withwider representation of other sectors and diseasecontrol programmes and develop the IVMconcept through the homepage of the VEM website.
Reference
[1] World Health Organization. Global strategicframework for integrated vector management.Geneva, WHO, 2004. Document WHO/CDS/CPE/PVC/2004.10.

Dengue Bulletin – Volume 31, 2007 183
Book review
Report of the Scientific Working Group meetingon dengue
Geneva, 1-5 October 2006
TDR/SWG/08
Dengue is the most rapidly spreading vectorborne disease. An estimated 50 million dengueinfections occur annually and approximately 2.5billion people live in dengue endemiccountries. Because of the rapidly increasingpublic health importance of this disease, in1999 dengue was incorporated in the portfolioof the UNICEF, UNDP, World Bank, WHOSpecial Programme for Research and Trainingin Tropical Diseases (TDR). The 2002 WorldHealth Assembly Resolution WHA55.17 urgedgreater commitment to dengue amongMember States and WHO; of particularsignificance is the 2005 Revision of theInternational Health Regulations (WHA58.3),which includes dengue as an example of adisease that may constitute a public healthemergency of international concern.
It was against this background that theDengue Scientific Working Group of 60 expertsfrom 20 countries including WHO staff fromfour Regions and Headquarters met in Genevain October 2006 to review existing knowledgeon dengue and establish priorities for futuredengue research aimed at improving denguetreatment, prevention and control. The goalof the Scientific Working Group was to outlinea research agenda by identifying bottlenecksand making detailed and specific researchrecommendations. The SWG wanted toidentify areas of research that could lead to
tangible benefits for people in disease endemiccountries within the coming years as well asoutline a strategic vision for applied and basicresearch from which benefits would be felt inthe medium to long term.
As a result of major demographic changes,rapid urbanization on a massive scale, globaltravel and environmental change, the world,particularly the tropical world, faces enormousfuture challenges from emerging infectiousdiseases. Dengue epitomizes these challenges.In the early years of the 21st Century we arecollectively failing to meet the challenge posedby dengue as the disease spreads unabatedand almost 40% of the world’s population nowlives at risk of contracting the disease. There iscurrently no specific clinically useful diagnostictest, no drugs, and no vaccine, and we havefailed to widely or effectively implementexisting vector control and clinical managementmeasures that we know would help to reducethe vector population and reduce case fatalityrates. Yet there has never been a moreoptimistic time to be involved in dengue anddengue research, and interest in the diseasehas attracted a new generation of talented andcommitted clinicians and scientists. Modernscience, from clinical medicine to basic researchon pathophysiology, drug and vaccine discovery,through to the social and behavioural sciencesand vector biology and control, offers a unique

184 Dengue Bulletin – Volume 31, 2007
Report of the Scientific Working Group on dengue, 2006
opportunity to make a tangible and substantialimpact on dengue over the next decade. Butin order to achieve what is possible, a paradigmshift is required in our current approach. Thedengue research community needs to: pushfor much greater implementation of existingknowledge to reduce case fatality rates, extendbasic and clinical research to understand theunderlying pathophysiology, aid diagnostics anddrug discovery and further improve clinicaloutcome, speed up the development ofvaccine candidates including moving as quicklyas possible to efficacy trials, and gatherevidence for implementing best practices forcontrol of the vector.
All of this is possible in the next ten years.But to achieve this, dengue needs a muchstronger voice within dengue endemic countriesand within the global public health communityto persuade society, funding agencies andpolicy-makers of the importance of the disease.We are at a critical epidemiological juncture ininfectious, particularly viral, emerging diseasesat the start of the 21st Century, and in manyways dengue serves as a model for how wemight meet that challenge. The lessons learnedfrom dengue will have implications for anumber of other diseases and our approach totheir control. The implementation of the bestof existing knowledge and practicesupplemented by future research applied inan integrated, holistic fashion can be expectedto significantly change the lives of individualsliving in dengue-endemic countries in thecoming years. The Scientific Working Grouphopes this research agenda will help provide astrategic plan for how we might collectivelyachieve the aims of reducing morbidity andmortality based on better understanding of thepathophysiology associated with dengue, onimplementation strategy, and on reduction ofvirus transmission.
Global dengue researchagenda
The priority dengue research areas are organizedaround four major research streams, which willprovide evidence and information for policy-makers and control programmes and lead tomore cost-effective strategies that will reversethe epidemiological trend.
Stream 1: Research related toreducing disease severity andcase fatality
Optimization of clinical management
An efficient out-patient system and clinical andlaboratory indicators of early dengue virusinfection, plasma leakage and shock, as wellas an effective and safe method of managingsevere haemorrhage, dengue in pregnancy, andpatients with co-morbidity, need to be validatedin order to scale up the use of improvedtreatment guidelines.
It is recommended to analyse:
• New methods and guidelines for triageand out-patient care.
• The validity, role and accessibility ofavailable and new diagnostics.
• The predictive value of prognosticmarkers (host and viral, non-invasivemeasurement of vascular leakage).
• Standardized approaches todetermining and documenting severedisease and response to treatment.
• Best practices for the treatment ofdengue including early treatment toreduce severity, treatment of

Report of the Scientific Working Group on dengue, 2006
Dengue Bulletin – Volume 31, 2007 185
established shock, and effective andsafe management of haemorrhage.
• The impact of co-morbidities ondisease severity, and the effect ofpregnancy.
• The causes of dengue deaths(including treatment failures).
Process and impact evaluation ofstaff training
Training programmes in case management canhelp to rapidly reduce case fatality. However,training has to be standardized and adapted tothe prevailing local health care system and bestpractice has to be identified and implementedin dengue-endemic countries.
It is recommended to analyse:
• The process and impact of existing/future training programmes.
Critical issues in dengue pathogenesis
A better understanding of dengue pathogenesiswill provide a foundation for future rationalclinical interventions. In particular we need tounderstand the changes underlying: endothelialpermeability in plasma leakage, dengue virusdiversity, and immune response to dengueviruses.
It is recommended to analyse:
• The physiological and molecularmechanisms leading to vascular leakand haemorrhage.
• The host genetic factors associatedwith dengue severity.
• The dengue viral factors and antigenicsubtypes related to tropism, epidemicpotential, and virulence.
• The mechanisms of antibody-mediatedenhancement and protection.
• The mechanisms of virus entry andcellular/tissue tropism.
• T and B cell responses and theirrelation to immunopathology andprotection.
• Genetic predisposition to dengue.
Stream 2: Research related totransmission control throughimproved vector management
Development and evaluation of vectorcontrol tools and strategies
The effectiveness of powerful vector controltools has been compromised by issues ofdelivery, coverage and acceptability. Promisingnew tools and approaches need to beevaluated.
It is recommended to analyse:
• The efficacy of new vector controltools and strategies in differentcontexts.
• Combinations of new and/or existingtools in different contexts.
• The scaling up of successful pilotprojects to state or national level.
Surveillance and response
Strengthening of surveillance systems throughdevelopment and validation of reliable riskindicators and the application of informationtechnology is needed for improved decision-making.
It is recommended to analyse:
• Improved methods and theirapplication/standardization inoperational contexts.
• The development and utilization ofearly warning and response systems.

186 Dengue Bulletin – Volume 31, 2007
Report of the Scientific Working Group on dengue, 2006
• The triggers that will allow effectiveresponse to incipient epidemics.
• The contribution of informationtechnology to decision-making.
Stream 3: Research related toprimary and secondary prevention
Vaccines
Vaccines offer the greatest hope for dengueprevention and there are several candidates inclinical development. The identification ofvaccine components that are suitably safe andimmunogenic, and of immune correlates ofprotection, should accelerate successful vaccinedevelopment and regulatory approval.
It is recommended to analyse:
• New vaccine candidates, adjuvants,and vaccination strategies.
• Correlates of protective immunity foruse as an endpoint in vaccine trials.
• Immune responses in vaccine trialsand natural infections.
• Prospects for phase 3 and 4 vaccineevaluation trials in multiple field sites.
• Issues associated with future vaccineusage and coverage, including cost-effectiveness studies ofimplementation.
Drugs
Anti-dengue drugs may have prophylactic (e.g.outbreak prevention) and therapeutic (e.g.prevention of severe disease) uses, includingpotential for impact on incidence and severityof ensuing disease. Anti-viral drug discovery fordengue has accelerated in recent years alongwith our knowledge of ‘drugable’ targets in thevirus.
It is recommended to analyse:
• Viral-encoded proteins for drug,diagnostics and vaccine design.
• New (including natural) products orexisting licensed drugs.
Stream 4: Health policy researchcontributing to adequate publichealth response
There is a contradiction between the highpriority afforded to dengue at political level andthe low level of resources allocated to dengueprevention and control programme activities.Health policy research will facilitate a redressof this imbalance.
It is recommended to analyse:
• Tools for rational decision-making andadequate prioritization of dengue, suchas studies of dengue burden and costsof illness.
• Factors leading to success or failure ofnational programmes.
• Decision-making that results indeclaration of state of emergency.
• The importance and burden of denguein less studied regions (e.g. Africa).
Conclusion
The Scientific Working Group hopes thisresearch agenda will help provide a strategicplan for how we might collectively achieve theaims of reducing dengue morbidity andmortality and its negative socioeconomicimpact. Donors and the research communityare encouraged to take part in this majorprogramme and to contribute through timelyinformation to the database TDR is establishingfor keeping track of research activities andrelevant findings.

Dengue Bulletin – Volume 31, 2007 187
Book review
Outbreak and spread of chikungunya
Weekly Epidemiological Record, No. 47, 2007, 82, 409-416
http://www.who.int/wer
Chikungunya is a mosquito-borne disease firstdescribed during an outbreak in the southernpart of the United Republic of Tanzania in1952. The name chikungunya derives from aroot verb in the Kimakonde language meaning“to become contorted” and it describes thestooped appearance of sufferers with arthralgia.The causative agent is an alphavirus of the familyTogaviridae. Since the first description ofchikungunya, there have been numerousoutbreaks in Africa, India and South-East Asia,the principal vectors being Aedes mosquitoes,of which Aedes albopictus and Ae. aegypti arethe most important. Chikungunya oftenproduces a mild illness that may be confusedwith dengue, since symptoms include fever,headache, arthralgia, myalgia and rash. Althoughserious complications are uncommon, arthralgiacan be debilitating and may persist for monthsor even years after infection; in elderly people,chikungunya can be a contributing factor todeath. In immunologically naive populations,where mosquito vectors are numerous,epidemic outbreaks may occur affecting manythousands of people. Outside of epidemics andwherever serological surveillance is lacking, thediagnosis of chikungunya may easily be missed.Chikungunya circulated in West and East Africaat relatively low levels until 1999–2000 whenaround 50 000 people became infected duringan outbreak in the Democratic Republic of theCongo. Evidence from India suggested that
outbreaks during the 1960s, were followed byrelatively little transmission after 1973.Chikungunya outbreaks are often separated byperiods of >10 years when infection is notapparent. For example, in Indonesia,chikungunya occurred sporadically until 1985after which there were no reports until a seriesof outbreaks between 2001 and 2007.
Recent outbreaks, 2001–2007
Starting in February 2005, a major outbreakoccurred among islands in the western part ofthe Indian Ocean, affecting the Comoros,Madagascar, Mayotte, Mauritius, La Réunionand the Seychelles (Figure). In La Réunion, byJune 2006 there had been an estimated266 000 cases, accounting for roughly one thirdof the population. Chikungunya continued tocirculate in La Réunion in 2007, albeit at amuch reduced level. The major vector in LaRéunion is Ae. albopictus, which appears tohave displaced Ae. aegypti on much of theisland as it has also in some other regions.During this period, there were an estimated9000 cases of chikungunya in the Seychelles,7290 in Mayotte and about 6000 in Mauritius.Associated with this outbreak in the IndianOcean islands, there were a large number ofimported cases of chikungunya in Europe,occurring in returning tourists and visitors,

188 Dengue Bulletin – Volume 31, 2007
Outbreak and spread of chikungunya
particularly those returning from La Réunion.Metropolitan France experienced the largestnumber of cases: between April 2005 andAugust 2006, there were 808 imported casesof chikungunya confirmed by serology. Thetemporal change in numbers reflected the riseand decline of the epidemic in La Réunion,with a peak of 178 cases in March 2006.Elsewhere in Europe, cases were reported fromGermany, Italy, Norway and Spain.
Ae. albopictus has been inadvertentlyintroduced into several European countries duringthe past 30 years; these countries include Albania,Belgium, Bosnia, Croatia, France, Greece, Italy,Montenegro, the Netherlands, Serbia, Slovenia,Spain and Switzerland. Although Ae. albopictusoccurs in the departments of Alpes-Maritimes andVar in France, where cases of chikungunya thatoriginated in La Réunion have been diagnosed,there has been no evidence of local transmission.
During 2006, there was a large outbreak ofchikungunya in India, with 1.39 million officiallyreported cases spread over 16 states; attack rates
were estimated at 45% in some areas. Theoutbreak was first noticed in Andhra Pradeshand it subsequently spread to Tamil Nadu.Thereafter, Kerala and Karnataka were affected;subsequently, the outbreak spread northwardsas far as Delhi. During 2007, up until 12October, a further 37 683 cases had beenreported by national authorities, largely in areasnot involved in the previous year’s epidemic.In India, the persistent and disabling arthralgiafollowing infection has been of particularconcern. In some areas, the outbreak wasassociated with high densities of Ae. albopictus,but Ae. aegypti is thought to be the major vectorelsewhere. During 2006, there were alsooutbreaks in the Andaman and Nicobar islands,Malaysia, and by November 2006, chikungunyahad appeared in Sri Lanka. Between January2001 and April 2007, Indonesia reported 15 207chikungunya cases from 7 provinces, with apeak in 2003. During 2006, there were 9imported cases of chikungunya in the Caribbean,involving Martinique, French Guyana andGuadeloupe; these mainly occurred in travellers
Figure: Geographical distribution of chikungunya cases 2001–2007 (data are presented asreporting period followed by the estimated number of cases, where data are available)

Outbreak and spread of chikungunya
Dengue Bulletin – Volume 31, 2007 189
from the Indian Ocean islands. The Caribbeanis an area of active dengue transmission, thevectors of which are also vectors of chikungunya,hence the region is vulnerable to localtransmission. In 2006, in the United States, 37cases of chikungunya were confirmed inreturning travellers, 32 from India, 3 from SriLanka, 1 from La Réunion and 1 from Zimbabwe.In 2006, in the United States, 35 cases ofchikungunya were confirmed in returningtravellers, 2 returning from La Réunion and 33from India. The United States is also vulnerableto local transmission owing to the widedistribution of Ae. albopictus. Large areas ofCentral and South America are similarlyvulnerable owing to the presence of abundantpopulations of both Ae. aegypti and Ae.albopictus.
In 2007, there was an outbreak ofchikungunya in Gabon, mainly affectingLibreville. Symptoms included headache, severearthralgia and skin rash. Up until 24 June, a totalof 17 618 cases had been reported, with 808hospitalizations. During a mosquito survey inLibreville in November 2006, Ae. albopictus wasfound for the first time in Gabon, and virus wassubsequently isolated from field populations ofthis species collected at the time of theoutbreak. Ae. aegypti is also common in the city,but none of the specimens collected at the sametime were positive for chikungunya virus.
In August 2007, the first cases ofindigenous transmission of chikungunya inEurope were reported from a largely rural areain the provinces of Ravenna and Forlì-Cesenain the Emilia-Romagna region of north-east Italy.Ae. albopictus is widespread and numerouswithin much of Italy. Between 15 June and 21September 2007, 292 suspected cases ofchikungunya were identified in the regionthrough a programme of active surveillance;many have been laboratory-confirmed. Theindex case is presumed to be a person fromthe region who travelled to a chikungunya-
affected area of Kerala (India) and returned toItaly in mid-June, developing symptoms shortlythereafter. The peak of the outbreak occurredin the third week of August. Epidemiologicaldata suggest that transmission took place at 4different localities in Emilia-Romagna, and thevirus has been isolated from local Ae. albopictusmosquitoes.
It has been confirmed that European Ae.albopictus are susceptible to infection withchikungunya virus from the Indian Oceanoutbreak. Samples of this mosquito, collectedas eggs from the Alpes-Maritimes, France, havebeen reared in the laboratory and the adultfemales experimentally infected by feeding onblood containing a strain of chikungunya virusderived from La Réunion. When Ae. albopictuswere examined 12–14 days after engorgement,27/35 (77%) showed evidence of infection inthe head. Such a high infection rate supportsthe view that this strain of the virus is well adaptedto transmission by this mosquito species. In thesame experiments, 33/49 (67%) Ae. detritusbecame infected. This species commonly biteshumans and has a wide distribution in thePalaearctic Region, breeding in coastal and inlandsaline water. Ae. caspius, another species fromthe Palaearctic Region found in brackish water,showed an infection rate of 4/16 (25%). A singlefemale Ae. vittatus survived the incubation periodand was found to be infected. This study raisesthe possibility that these other mosquito speciesmay play a role in transmission during an outbreakof chikungunya.
Genotyping of viral isolates has provideduseful information on the possible origins of theoutbreak in the Indian Ocean islands as well asevidence for the circulation in that outbreak ofviruses with a high evolutionary potential. Duringoutbreaks, arboviruses such as chikungunyadiversify due to mutation, leading to severalidentifiable strains. Chikungunya virus has itsorigins in continental Africa and is related to thevirus responsible for o’nyong-nyong. The

190 Dengue Bulletin – Volume 31, 2007
Outbreak and spread of chikungunya
chikungunya virus circulating in the outbreak inthe Indian Ocean islands represents a distinctclade within the East-Central-South Africanphylogroup, compatible with its recent origin inthe African region. However, evidence fromgenotyping suggests that some time after thevirus reached La Réunion, before transmissionrates rose steeply, a mutation occurred thatenabled more effective transmission by Ae.albopictus. If this hypothesis were confirmed,it could help explain the rapid spread anddissemination of chikungunya after it reachedLa Réunion. The viral genotype involved in thelarge outbreak in India is from the same lineageas that seen in La Réunion, whereas previousoutbreaks in India have involved an Asiangenotype of the virus.
Although the 2 major mosquito species,Ae. aegypti and Ae. albopictus, have beenimplicated in the larger outbreaks ofchikungunya, within Africa, several othermosquito vectors have been implicated,including species of the Ae. furcifer-taylori groupand Ae. luteocephalus. In this region, variousprimates have been implicated as reservoirhosts. Elsewhere in Africa, little is known ofthe possible role animals have as reservoirs orof other mosquitoes as vectors.
Whereas Ae. aegypti is confined to thetropics and subtropics (approximately within the10° isotherms), Ae. albopictus also occursextensively in temperate and even cold–temperate regions, thereby extending thegeographical range over which chikungunyatransmission may occur. Ae. albopictus exploitsa wider range of breeding sites than Ae. aegypti.These include bromeliads, coconut husks, cocoapods, bamboo stumps, tree holes and rock poolsin addition to outdoor artificial containers,including vehicle tyres, various discardedcontainers and saucers beneath plant pots. Thisdiversity of larval habitats explains the abundanceof Ae. albopictus in rural as well as periurban
areas and in shady city parks. Ae. aegypti is moreclosely associated with human habitation andexploits indoor breeding sites, including flowervases, water storage vessels and concrete watertanks in bathrooms, as well as the same artificialhabitats outdoors as Ae. albopictus.
Control measures
A key factor in the increasing geographical rangeand prevalence of Ae. albopictus has been theglobalization of trade, particularly thetransportation of used tyres between countriesand continents. Both Ae. aegypti and Ae.albopictus have drought-resistant eggs thatremain viable over periods of several weeks,enabling them to survive lengthy journeys withsea-borne freight, and on aircraft or long-distance road transport. For these reasons, somecountries have introduced a ban on theimportation of used tyres. Both mosquitospecies are competent vectors of otherarboviruses, including dengue, so there arewider public health concerns about theaccidental transfer of these species betweencountries and continents.
It will be impossible to eliminate Ae.albopictus with current technologies in areaswhere it has become well established (theAmericas, Cameroon, Italy and Nigeria). Insuch regions, efforts should concentrate onreducing vector density in order to reduce therisk of transmission. Where a newly introducedAe. albopictus population is detectedsufficiently early, concerted efforts, includingthe use of insecticide applications, are requiredto prevent this species becoming established;there are examples of the successful use ofthis approach. In areas at high risk for theintroduction of Ae. albopictus, good vectorsurveillance is required since there is only ashort window of opportunity to eliminate newfoci before they become established.

Outbreak and spread of chikungunya
Dengue Bulletin – Volume 31, 2007 191
Chikungunya poses considerable problemsfor public health authorities once an outbreakhas commenced. The major environmentalmeasures used to reduce sources of mosquitobreeding may not be fully implemented withinthe timescale of an outbreak. Few public healthprogrammes are up to the task of respondingto large epidemics of arboviruses, but thosecountries with good routine programmes ofvector surveillance and control are better ableto mitigate the effects of an outbreak.
Measures to reduce the number of water-containing receptacles may contributesignificantly to a reduction in emergentmosquitoes, but such action needs to be basedon sound knowledge of the ecology of theimmature stages of the mosquito in that specificlocation. During an arbovirus outbreak, areduction in transmission may require theapplication of insecticides as space sprays, asresidual applications on surfaces in and aroundcontainer habitats where mosquitoes alight, andas larvicides to kill the immature stages. Sourcereduction will be particularly difficult in ruralareas where numerous natural, as well as man-made, breeding sites may occur. Nevertheless,reducing the number of containers in andaround the home is to be encouraged, andthe most important health messages for thepublic relate to actions that can be taken toreduce the number of habitats.
In urban areas, used-tyre dumps enablelarge-scale breeding, particularly of Ae.albopictus, and require focused controlmeasures, including the application ofinsecticides. In the longer term, investment inrecycling programmes for tyres should beencouraged. In hospitals and other health-caresettings where chikungunya (and dengue)patients may be treated, vector controlmeasures should be routinely applied andintensified during outbreaks. Viraemic patientsshould be nursed in screened wards or under
mosquito nets. The potential risks oftransmission through donation of blood ororgans should also be considered.
Personal protection
For individual protection during outbreaks ofchikungunya, clothing that minimizes skinexposure to the day-biting vectors is advised.Repellents may be applied to exposed skin orto clothing. Repellents should contain DEET(N, N-diethyl- 3-methylbenzamide), IR3535 (3-[N-acetyl-N-butyl]-aminopropionic acid ethylester) or icaridin (1-piperidinecarboxylic acid,2-[2-hydroxyethyl]-1-methylpropylester). Theyshould be applied in strict accordance with labelinstructions. Insecticide-treated mosquito netsafford good protection for those who sleepduring the day, particularly young children.Where indoor biting occurs, mosquito coils orother insecticide vaporizers may also reducebiting.
Surveillance
Accurate data on the extent of infection in thehuman population requires the use ofimmunological testing, such as enzyme-linkedimmunosorbent assays (ELISA) for the detectionof immunoglobulin M and immunoglobulin Ganti-chikungunya antibodies. Various reversetranscriptase–polymerase chain reaction (RT-PCR) methods are available; they are of variablesensitivity and some will detect very low levelsof viral transcripts. Some are suited to clinicaldiagnosis and others to surveys. RT-PCR productsfrom clinical samples may also be used forgenotyping of the virus, allowing comparisonwith virus from various geographical sources.
In areas of particular risk, routine vectorsurveillance is essential for effective vectorcontrol, as identified above.

192 Dengue Bulletin – Volume 31, 2007
Book review
Multicountry study of Aedes aegypti pupal productivitysurvey methodology: Findings and recommendations#
Dana A. Focksa and Neal Alexanderb
aInfectious Disease Analysis, Gainesville, Florida, USA
bInfectious Diseases Epidemiology Unit, London School of Hygiene and Tropical Medicine, London, UK
Rationale
Dengue fever and the potentially lethalhaemorrhagic form of dengue are a fast growingpublic health problem worldwide (50 millioncases with 500 000 manifesting shock and/orhaemorrhage each year), and particularlyimportant in the Americas and Asia. The onlyknown strategy for reducing dengue transmissionis to reduce the vector population, which isachieved through interventions in domestic andperi-domestic water containers. However, littleis known about the efficacy of interventionstargeted to specific classes of water-holdingcontainer, and there is uncertainty about thebest indicators to use for measuring the successof interventions on vector populations.
TDR financed a multicountry studyinvolving nine Latin American, Asian, andAfrican countries, based on the rationale thatcertain water containers are particularlyproductive of the dengue vector, Aedes aegypti(L.), and that, therefore, an interventiontargeted to these containers would bring vectordensities below the threshold for epidemictransmission. This pupal/demographic surveymethod was described recently.[1]
The objective of this multicountry study wasto evaluate the practicality of the survey methodand whether it can consistently identify andclassify particularly productive classes ofcontainer, and so provide guidance ondevelopment of targeted control strategies. Aswill be discussed in greater detail, containersmay be classified on any number of criteria suchas descriptive name, volume, use,abandonment, location indoors or out, etc. Theresults from this study will have importantconsequences for the development of improveddengue control strategies.
It is important to emphasize thatoperationally, pupal/demographic surveys areexpected to be conducted infrequently,perhaps only at the beginning of a process toidentify the epidemiologically importantcontainer classes to target. When targetedinterventions via community participationmethods are under way, the primary surveygoal would be simply to determine the actualamount of intervention in place without larvaeor pupae being counted.
This document summarizes the largeamount of data created by the nine study teams
# For complete details, please access the original document TDR/IRM/DEN/06.1

Multicountry study of Aedes aegypti pupal productivity survey methodology: Findings and recommendations
Dengue Bulletin – Volume 31, 2007 193
with the goal of serving as a reference text forother researchers as well as for controlprogrammes which are expected to adopt thetargeted intervention technique. The documentreflects the fact that today, most denguecontrol efforts are based on suppression of Ae.aegypti and not on eradication; these effortswould benefit from answers to the followingquestions:
• How much suppression is adequateto be prophylactic for dengueoutbreaks in a particular location (Table2)?
• How do we monitor the degree ofsuppression achieved in ongoingprogrammes?
Some have recently been advocating anadditional question:
• Given that the epidemiologicalimportance of a particular class ofcontainer is the product of the averageadult productivity and the abundanceof that class of container in theenvironment, how do we selectsubsets of containers for interventionsuch that we minimize labour andcosts while maximizing reduction ofadults?
And finally,
• In developing a targeted strategy, howmany households must be surveyedto be confident that the importantcontainer classes have beenidentified?
Answering the first question involves anunderstanding of transmission thresholds –what they represent and how they areestimated. Thresholds are a function of manyfactors, but a key factor is the ratio of thenumbers of adult Ae. aegypti and people, hencethe need for a survey method that permits
estimation of this important variable.[2] Becausepupae can be counted and are highly correlatedwith the standing crop of adults, a surveymethod involving counting pupae and peoplehas been developed.[3] Mechanistic andmathematical models of dengue transmissionhave been developed that allow estimation oftransmission thresholds in terms of Ae. aegyptipupae per person as a function of temperatureand herd immunity; it is only this statistic thatcan provide estimates of how much reductionis necessary to be prophylactic for dengueoutbreaks (Table 2). The practical challengesof obtaining this statistic and its usefulness indeveloping targeted strategies are the subjectof this paper.
Several authors have recently made thecase that use of the traditional Stegomyiaindices as epidemiologic indicators of denguetransmission risk should be abandoned as theyhave a number of serious shortcomings.[1] Theargument is that a pupal and demographicsurvey that provides an estimate of the numberof pupae per person in a community by classof container, e.g. drums, flower vases and pots,cisterns, abandoned containers, discarded tyres,is more appropriate for assessing risk anddirecting control operations. The ratio of pupaeper person is used for several reasons:
• Unlike any of the other mosquito life-cycle stages, it is possible to actuallycount the number of Ae. aegypti pupaein most domestic environments (forexceptions, see summary of the VietNam study below).
• Container-inhabiting Stegomyia pupaeare easily and inexpensively separatedfrom other genera and identified tospecies as pupae or adults.
• Because pupal mortality is slight andwell-characterized, the number ofpupae is highly correlated with thenumber of adults.

194 Dengue Bulletin – Volume 31, 2007
Multicountry study of Aedes aegypti pupal productivity survey methodology: Findings and recommendations
• The statistic of pupae per person canbe related to transmission risk andprovide target levels of reductionrequired in control efforts.[2]
The survey method being evaluated in thesestudies is called the pupal/demographic survey,so named because we count both pupae andthe number of people associated with them. Inpractice, such a survey involves visiting 100 ormore residences (see section on survey sizerequirements below), usually by two inspectorsequipped with nothing more than a few litresof clean water, a sieve,* some largemouthpipettes, a white enamel pan, and small shellvials. The inspectors request permission toexamine the water-holding containers in ahouse, and enquire as to the number of peopleliving in the house (or sleeping there the previousnight). With permission, they proceed to strainthe contents of each container at the location,re-suspending the sieved contents in a smallamount of clean water in the enamel pan, fromwhere the pupae from each container arepipetted into a labelled vial. If there are othercontainer-inhabiting species in the area besidesAe. aegypti, the contents of each vial aretransferred to small cups covered with bridalveil secured with a rubber band; these are heldin the lab until the adults emerge, when theycan be identified. As a minimum, the datarecorded for each container include the type ofcontainer (drum, bottle, etc.), location (indoors,outdoors, under vegetation, etc.), method offilling (manually, by rain, roof runoff, etc.), andthe number of pupae by species. These inputsare used to develop a classification scheme (seecontainer classification section below). Data areusually summarized by container class on aspreadsheet; for each container class found,simply add up the number of pupae associatedwith that type in the survey, and divide by the
sum of the number of people living in theresidences inspected. Sorting the list of containerclasses by their associated number of pupae perperson quickly identifies the productive and non-productive classes of container.
While exhaustive counts of pupae in everycontainer are certainly more labour intensivethan simply noting whether a container haslarvae and/or pupae (as in the various traditionalStegomyia indices), the pupal/demographicsurvey permits us to know the number ofpupae per person associated with the variousclasses of container in the environment. Thisstatistic, Ae. aegypti pupae per person by classof container, is not an index after the fashionof the Stegomyia indices, rather it is theepidemiologically significant statistic, the ratioof hosts and vectors; published transmissionthresholds permit us to appreciate this statisticin the context of estimates of herd immunityand ambient temperatures.[2] What each of thestudies summarized below demonstrates is thatsome classes of container are more importantby virtue of their production of Ae. aegyptiadults than others, and that this leads logicallyto the development of targeted interventions.An earlier publication provides a discussion ofthe utility of other indices for dengue riskassessment and control.[2]
The objectives of this document are to:
• Summarize the key findings from thenine study sites of the multicountryproject on pupal/demographic surveymethodology.
• Outline the prospects and limitationsof this survey methodology.
• Draw conclusions and propose furtherresearch and implications for dengueprevention and control programmes.
*USA standard sieve series, number 30 sieve(equivalent to ASTM designation E11, 600 μm[0.0243"] opening)

Multicountry study of Aedes aegypti pupal productivity survey methodology: Findings and recommendations
Dengue Bulletin – Volume 31, 2007 195
Key findings of themulticountry study
• The pupal/demographic surveyidentifies epidemiologically importantand unimportant types of container
• Containers and sites can be classifiedusing different schema
• Distribution of Ae. aegypti pupae percontainer is strongly clumped
• A potentially important class ofcontainer, the rare but extremelyproductive container
• Survey size requirements to developa targeted control strategy
Prospects and limitations ofthe pupal productivity surveymethodology
• Results are useful in developingtargeted source reduction/controlstrategy for dengue
• Difficulty of surveying large containers
Conclusions, further research,and implications for dengueprevention and controlprogrammes
Conclusions
The studies have confirmed that the pupal/demographic survey provides the necessarydata for developing targeted strategies. Themethod certainly requires more labour than dothe traditional Stegomyia indices, but on a cost/benefit analysis, the traditional measures failbecause, regardless of cost, they cannot informtargeted strategy nor can they be used fortransmission risk assessment. Unpublished
longterm (>5 years) and repeated longitudinalpupal/demographic surveys in Iquitos, Peru,[3]
and Yogyakarta, Indonesia, indicate that pupalproduction by container type is remarkablystable over time and suggest, as stated above,that only an initial, high quality survey may berequired.
Calibrated sweep net surveys may providea method for estimating absolute numbers ofpupae for some types of container. However,calibration factors determined in one locationshould be used with great caution in anotherarea, where subtle differences in climate,elevation, methods of handling water, etc., mayinfluence productivity, and hence accuracy.
Further research
Evaluate dengue control strategiesusing transmission thresholds andpupal/demographic surveymethodology
Since the appearance of dengue haemorrhagicfever in South-East Asia in the 1960s and itssubsequent spread to the Americas and Oceania,several control strategies have been advocated.Ultra low volume (ULV) spray applications appliedthrough national control programmes were givenprominence for several decades. While it wasdocumented that the sprays certainly killed adultmosquitoes, their use in controlling epidemicswas harder to demonstrate. Transitory butinfrequent depressions in adult mosquito survivalsimply could not suppress dengue transmissionin endemic locations. As appreciation of theinadequacies of ULV grew in the public healthcommunity, the focus of control efforts turnedto involving affected communities, and much wasmade of community participation for the purposeof source reduction and/or control. Yetexperience, with the exception of the two verticaland national programmes of Singapore and Cuba,has been disappointing. Some thought the

196 Dengue Bulletin – Volume 31, 2007
Multicountry study of Aedes aegypti pupal productivity survey methodology: Findings and recommendations
problem lay with an inadequate understandingon the part of the communities and that, withtime and education, dengue control could beachieved. Others suggested that another factorwas at work—that the public was in factconducting a cost–benefit analysis with regard todengue—and simply concluded that preventingmosquito breeding in all containers every dayindefinitely was, as indicated by practice, simplynot worthwhile from a labour/time perspective.The promise of targeted source reduction andcontrol, building on the significantaccomplishments to date in the area of communityparticipation, lies with reducing the labour andcosts in the cost–benefit calculation. With fewexceptions, evaluations of community-basedinterventions were based on sociological evidencesuch as an increase in knowledge, change inpractice, etc., and not on measurements oftransmission or disease prevalence. Perhaps thisis understandable in light of the time and effortrequired to change human behaviour with regardto cigarettes, seat belts, etc.
In light of the foregoing comments, thechief research recommendation of thissummary is to conduct targeted interventionswith the dependent variable being somemeasure of transmission. Studies withsignificant funding can measure transmissiondirectly using longitudinal serosurveys of infantsand adolescents using the prevalence of anti-dengue IgM as a proxy; other studies will haveto rely on case data. In addition, evaluationsof interventions should monitor the actualimpact on pupal production in the targetedcontainers.** If a targeted intervention effort
fails, it is critical to know whether failure isattributable to inadequate control in targetedcontainers or if the problem lies with the notionsof targeted intervention and transmissionthreshold.
Estimate the amount of suppressionrequired in interventions usingtransmission thresholds
Mechanistic models of the dynamics of denguehave been used to estimate transmissionthresholds (the number of Ae. aegypti pupaeper person) as a function of pre-existingantibody levels in the human population (herdimmunity) and ambient air temperature (Table1).[2] The threshold, an epidemiological conceptcentral to infectious disease control, is simplythe breaking or tipping point; at levels belowthreshold, infection rates and viral persistencedecline in the human population. Thresholdlevels were estimated to range between 0.5and 1.5 Ae. aegypti pupae per person for anambient air temperatures of 28 °C and initialseroprevalence of 0% to 67%. The concept oftransmission threshold has given us a new andhopefully useful tool for monitoring targets forsource reduction/control efforts. Moreover, interms of risk assessment, transmissionthresholds provide estimates of the level ofelimination or control necessary to precludetransmission (Table 2). In targetedinterventions, attempts should be made toreduce the number of Ae. aegypti pupae perperson to below the threshold estimate. Notein table 2 that several locations would require>70% reduction in adult production to beprophylactic.
It goes without saying that we seeexceeding the threshold as being a necessarybut not sufficient cause of transmission. Thetable of transmission thresholds (Table 1) takesinto account the degree of susceptibility in thehuman population (and indicates the possible
**A current evaluation in Yogyakarta, Indonesia, usingpyriproxyfen to prevent adult emergence involves:weekly visual inspection of targeted containers forthe presence or absence of the compound;laboratory bio-assay of water collected from treatedcontainers in the field; and laboratory observation offield- collected pupae for emergence success anddeformities. Transmission is monitored using: casedata, and seroprevalence of IgM in children andinfants measured every three months.
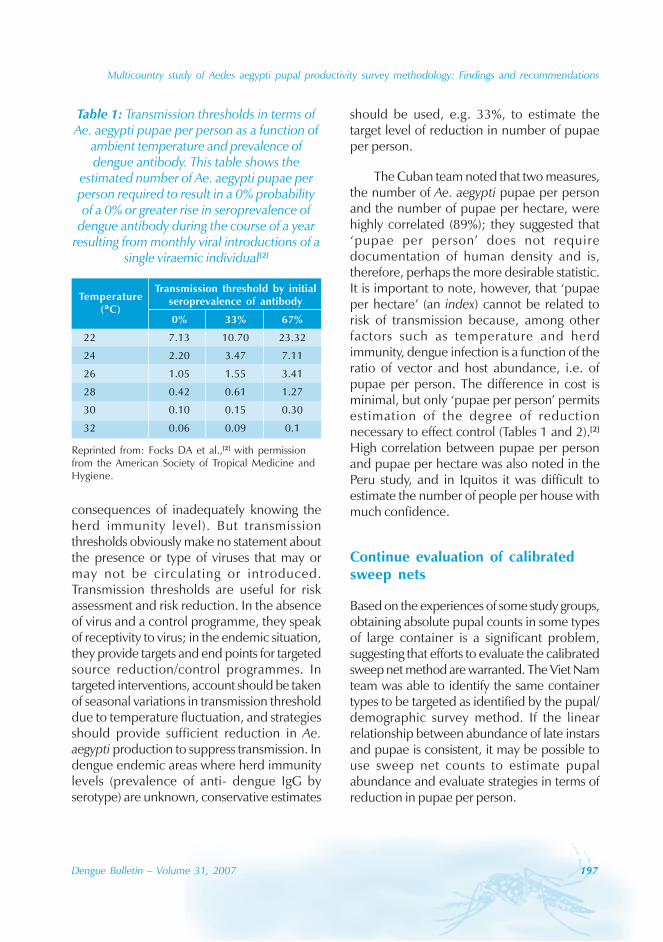
Multicountry study of Aedes aegypti pupal productivity survey methodology: Findings and recommendations
Dengue Bulletin – Volume 31, 2007 197
consequences of inadequately knowing theherd immunity level). But transmissionthresholds obviously make no statement aboutthe presence or type of viruses that may ormay not be circulating or introduced.Transmission thresholds are useful for riskassessment and risk reduction. In the absenceof virus and a control programme, they speakof receptivity to virus; in the endemic situation,they provide targets and end points for targetedsource reduction/control programmes. Intargeted interventions, account should be takenof seasonal variations in transmission thresholddue to temperature fluctuation, and strategiesshould provide sufficient reduction in Ae.aegypti production to suppress transmission. Indengue endemic areas where herd immunitylevels (prevalence of anti- dengue IgG byserotype) are unknown, conservative estimates
Table 1: Transmission thresholds in terms ofAe. aegypti pupae per person as a function of
ambient temperature and prevalence ofdengue antibody. This table shows the
estimated number of Ae. aegypti pupae perperson required to result in a 0% probabilityof a 0% or greater rise in seroprevalence of
dengue antibody during the course of a yearresulting from monthly viral introductions of a
single viraemic individual[2]
Reprinted from: Focks DA et al.,[2] with permissionfrom the American Society of Tropical Medicine andHygiene.
should be used, e.g. 33%, to estimate thetarget level of reduction in number of pupaeper person.
The Cuban team noted that two measures,the number of Ae. aegypti pupae per personand the number of pupae per hectare, werehighly correlated (89%); they suggested that‘pupae per person’ does not requiredocumentation of human density and is,therefore, perhaps the more desirable statistic.It is important to note, however, that ‘pupaeper hectare’ (an index) cannot be related torisk of transmission because, among otherfactors such as temperature and herdimmunity, dengue infection is a function of theratio of vector and host abundance, i.e. ofpupae per person. The difference in cost isminimal, but only ‘pupae per person’ permitsestimation of the degree of reductionnecessary to effect control (Tables 1 and 2).[2]
High correlation between pupae per personand pupae per hectare was also noted in thePeru study, and in Iquitos it was difficult toestimate the number of people per house withmuch confidence.
Continue evaluation of calibratedsweep nets
Based on the experiences of some study groups,obtaining absolute pupal counts in some typesof large container is a significant problem,suggesting that efforts to evaluate the calibratedsweep net method are warranted. The Viet Namteam was able to identify the same containertypes to be targeted as identified by the pupal/demographic survey method. If the linearrelationship between abundance of late instarsand pupae is consistent, it may be possible touse sweep net counts to estimate pupalabundance and evaluate strategies in terms ofreduction in pupae per person.
TemperatureTransmission threshold by initial
(°°°°°C)seroprevalence of antibody
0% 33% 67%
22 7.13 10.70 23.32
24 2.20 3.47 7.11
26 1.05 1.55 3.41
28 0.42 0.61 1.27
30 0.10 0.15 0.30
32 0.06 0.09 0.1

198 Dengue Bulletin – Volume 31, 2007
Multicountry study of Aedes aegypti pupal productivity survey methodology: Findings and recommendations
Develop better classificationschemes for containers, andmethods to locate rare butespecially productive containersat house and block scales
When rare but especially productive containers(REPCs) are encountered, priority should begiven to:
• elucidating the factors responsible forthe anomalous production
• identifying the common featuresamong REPCs discovered during thesurvey (location, association withvegetation, type of container withrespect to material, use/non-use, etc.)
• developing a classification schemethat includes factors to facilitate thelocating of REPCs without exhaustivesurvey.
The fact that REPCs are infrequently found,that they appear as anomalies and are nottypical, should not mean serious considerationis not given to this potentially important classof container. Rare they may be, but mostreasonably sized surveys discover one or moreREPCs often accounting for a significantproportion of production. In addition todiscovering common factors associated withREPCs which might be useful in developing asearch strategy targeting them, anotherimportant question is whether their productionis stable over time. If it is stable (the likelycase) then we should probably always includean REPC class in our surveys to keep us mindfulof their existence and importance, helping usto think in terms of answering the three pointsabove.
The container classification schemesdeveloped in the studies in Peru, Puerto Rico,and other countries, involving the concepts ofabandonment, location with respect tovegetation or placement, etc., are very good
examples of the utility of thinking in terms ofnew covariates of production and should bestudied and applied to local surveys. Rapidlocation of especially productive classes ofcontainer could serve to further reduce thelabour associated with targeted strategies.
Implications for dengueprevention and controlprogrammes
The results of these studies are encouraging—they suggest that control of transmission maybe possible with the treatment or eliminationof only a small portion of the large number ofclasses of water-holding container found in theenvironment. Each study identified theimportant classes of container to be controlled,and also the classes that could safely be ignored.Commonly, the more productive classes werelarger containers that were stable through time,a feature that may make them more amenableto control with biological control agents orchemicals. Given that all control programmesare resource limited, this new and quantitativeunderstanding of productivity can lead totargeted interventions that may be sustainablebecause control that relies on targeted strategyrequires substantially less labour. A TDRsupported multicentre study on the cost-effectiveness of targeted vs. non-targetedinterventions will provide answers to thisquestion.
Reducing the number of breedingcontainers will not cause an increasein productivity of the remainingcontainers
A common concern regarding targeted strategiesis the notion that, immediately following theelimination of one or more classes of container,more oviposition, and hence more adult
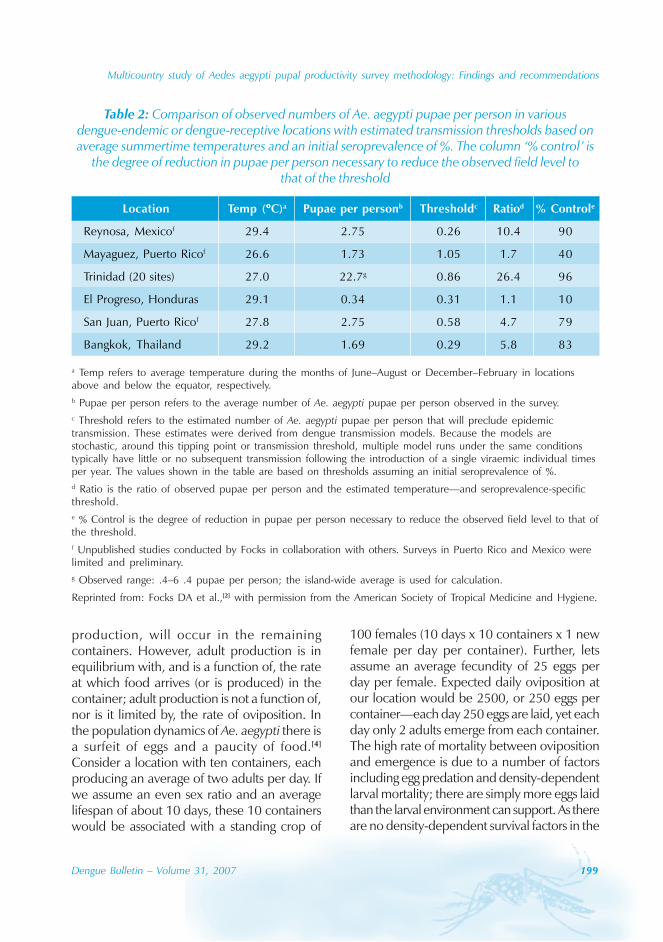
Multicountry study of Aedes aegypti pupal productivity survey methodology: Findings and recommendations
Dengue Bulletin – Volume 31, 2007 199
Table 2: Comparison of observed numbers of Ae. aegypti pupae per person in variousdengue-endemic or dengue-receptive locations with estimated transmission thresholds based onaverage summertime temperatures and an initial seroprevalence of %. The column ‘% control’ is
the degree of reduction in pupae per person necessary to reduce the observed field level tothat of the threshold
a Temp refers to average temperature during the months of June–August or December–February in locationsabove and below the equator, respectively.b Pupae per person refers to the average number of Ae. aegypti pupae per person observed in the survey.c Threshold refers to the estimated number of Ae. aegypti pupae per person that will preclude epidemictransmission. These estimates were derived from dengue transmission models. Because the models arestochastic, around this tipping point or transmission threshold, multiple model runs under the same conditionstypically have little or no subsequent transmission following the introduction of a single viraemic individual timesper year. The values shown in the table are based on thresholds assuming an initial seroprevalence of %.d Ratio is the ratio of observed pupae per person and the estimated temperature—and seroprevalence-specificthreshold.e % Control is the degree of reduction in pupae per person necessary to reduce the observed field level to that ofthe threshold.f Unpublished studies conducted by Focks in collaboration with others. Surveys in Puerto Rico and Mexico werelimited and preliminary.g Observed range: .4–6 .4 pupae per person; the island-wide average is used for calculation.
Reprinted from: Focks DA et al.,[2] with permission from the American Society of Tropical Medicine and Hygiene.
production, will occur in the remainingcontainers. However, adult production is inequilibrium with, and is a function of, the rateat which food arrives (or is produced) in thecontainer; adult production is not a function of,nor is it limited by, the rate of oviposition. Inthe population dynamics of Ae. aegypti there isa surfeit of eggs and a paucity of food.[4]
Consider a location with ten containers, eachproducing an average of two adults per day. Ifwe assume an even sex ratio and an averagelifespan of about 10 days, these 10 containerswould be associated with a standing crop of
100 females (10 days x 10 containers x 1 newfemale per day per container). Further, letsassume an average fecundity of 25 eggs perday per female. Expected daily oviposition atour location would be 2500, or 250 eggs percontainer—each day 250 eggs are laid, yet eachday only 2 adults emerge from each container.The high rate of mortality between ovipositionand emergence is due to a number of factorsincluding egg predation and density-dependentlarval mortality; there are simply more eggs laidthan the larval environment can support. As thereare no density-dependent survival factors in the
Location Temp (°°°°°C)a Pupae per personb Thresholdc Ratiod % Controle
Reynosa, Mexicof 29.4 2.75 0.26 10.4 90
Mayaguez, Puerto Ricof 26.6 1.73 1.05 1.7 40
Trinidad (20 sites) 27.0 22.7g 0.86 26.4 96
El Progreso, Honduras 29.1 0.34 0.31 1.1 10
San Juan, Puerto Ricof 27.8 2.75 0.58 4.7 79
Bangkok, Thailand 29.2 1.69 0.29 5.8 83

200 Dengue Bulletin – Volume 31, 2007
Multicountry study of Aedes aegypti pupal productivity survey methodology: Findings and recommendations
adult stage, elimination of 25% or 50% of adultproduction results in a similar reduction in thestanding crop of adults.[5]
In a targeted source reduction campaign,in addition to removing the waterholdingcontainers, the associated Ae. aegypti larvalfood is removed from the environment andproductivity of the environment goes down,as does the standing crop of adults. Anytransient rise in oviposition in untreatedcontainers is met with increased density-dependent larval mortality as productivity is afunction of food availability—recall the largeratio of daily oviposition and emergence. Incontrast, in an intervention involving larvicides,
biological control agents, or insect growthregulators, neither the number of ovipositionsites nor the amount of food declines. Ourtreatment has eliminated the production insome containers, thereby lowering the standingcrop of adults and oviposition. One mightwonder if productivity would go down inuntreated containers, oviposition rates in thembeing somewhat lower, as a lower number offemales is associated with the same numberof containers. Simulation and field studies inKenya suggest that until adult populations arereduced by >40%, container productivity willnot be limited by lowered oviposition ratesbecause food-based density-dependent larvalmortality decreases.[4]
References
[1] Focks DA. A review of entomological samplingmethods and indicators for dengue vectors.Geneva, World Health Organization, 2003.Document TDR/IDE/Den/03.1.
[2] Focks DA et al. Transmission thresholds fordengue in terms of Aedes aegypti pupae perperson with discussion of their utility in sourcereduction efforts. American Journal of TropicalMedicine& Hygiene. 2000; 62:11–18.
[3] Focks DA, Chadee DD. Pupal survey: Anepidemiologically significant surveillancemethod for Aedes aegypti: An example usingdata from Trinidad. American Journal ofTropical Medicine and Hygiene. 1997; 56: 159–167.
[4] Subra R, Mouchet J. The regulation ofpreimaginal populations of Aedes aegypti,Diptera: Culicidae, on the Kenya coast: 2.Food as a main regulatory factor. Annals ofTropical Medicine and Parasitology. 1984; 78:63–70.
[5] Focks DA et al. Dynamic life table model forAedes aegypti (L.) (Diptera: Culicidae). Analysisof the literature and model development.Journal of Medical Entomology. 1993; 30:1003–1017.

Dengue Bulletin – Volume 31, 2007 201
Instructions for contributors
Dengue Bulletin welcomes all original researchpapers, short notes, review articles, letters tothe Editor and book reviews which have a director indirect bearing on dengue fever/denguehaemorrhagic fever prevention and control,including case management. Papers should notcontain any political statement or reference.
Manuscripts should be typewritten inEnglish in double space on one side of whiteA4-size paper, with a margin of at least oneinch on either side of the text and should notexceed 15 pages. The title should be as shortas possible. The name of the author(s) shouldappear after the title, followed by the name ofthe institution and complete address. The e-mail address of the corresponding author shouldalso be included and indicated accordingly.
References to published works should belisted on a separate page at the end of thepaper. References to periodicals should includethe following elements: name and initials ofauthor(s); title of paper or book in its originallanguage; complete name of the journal,publishing house or institution concerned; andvolume and issue number, relevant pages anddate of publication, and place of publication(city and country). References should appearin the text in the same numerical order (Arabicnumbers in parenthesis) as at the end of thearticle. For example:
(1) Nimmannitaya S. Clinical spectrum andmanagement of dengue haemorrhagicfever. The Proceedings of theInternational Conference on DengueHaemorrhagic Fever, Kuala Lumpur,September 1-3, 1983:16-26.
(2) Gubler DJ. Dengue and denguehaemorrhagic fever: Its history andresurgence as a global public healthproblem. In: Gubler DJ, Kuno G (ed.),
Dengue and dengue haemorrhagic fever.CAB International, New York, NY,1997, 1-22.
(3) Nguyen Trong Lan, Nguyen ThanhHung, Do Quang Ha, Bui Thi MaiPhuong, Le Bich Lien, Luong Anh Tuan,Vu Thi Que Huong, Lu Thi Minh Hieu,Tieu Ngoc Tran, Le Thi Cam andNguyen Anh Tuan. Treatment ofdengue haemorrhagic fever atChildren’s Hospital N.1, Ho Chi MinhCity, 1991-1996. Dengue Bulletin.1997; 22: 150-161.
Figures and tables (Arabic numerals), withappropriate captions and titles, should beincluded on separate pages, numberedconsecutively, and included at the end of thetext with instructions as to where they belong.Abbreviations should be avoided or explainedat the first mention. Graphs or figures shouldbe clearly drawn and properly labelled,preferably using MS Excel, and all data clearlyidentified.
Articles should include a self-explanatoryabstract at the beginning of the paper of notmore than 300 words explaining the need/gapin knowledge and stating very briefly the areaand period of study. The outcome of the researchshould be complete, concise and focused,conveying the conclusions in totality. Appropriatekeywords and a running title should also beprovided.
Articles submitted for publication shouldbe accompanied by a statement that they havenot already been published, and, if acceptedfor publication in the Bulletin, will not besubmitted for publication elsewhere withoutthe agreement of WHO, and that the right ofrepublication in any form is reserved by theWHO Regional Offices for South-East Asia(SEARO) and the Western Pacific (WPRO).

202 Dengue Bulletin – Volume 31, 2007
Instructions for contributors
One hard copy of the manuscript withoriginal and clear figures/tables and a computerdiskette/CD-ROM indicating the name of thesoftware should be submitted to:
The EditorDengue BulletinWHO Regional Office for South-East AsiaIndraprastha EstateMahatma Gandhi RoadNew Delhi 110002, IndiaTelephone: 91-11-23370804Fax: 91-11-23379507, 23370972E-mail: [email protected]
Manuscripts received for publication aresubjected to in-house review by professionalexperts and are peer-reviewed by experts inthe respective disciplines. Papers are acceptedon the understanding that they are subject toeditorial revision, including, where necessary,condensation of the text and omission oftabular and illustrative material.
Original copies of articles submitted forpublication will not be returned. The principalauthor will receive 10 reprints of the articlepublished in the Bulletin. A pdf file can besupplied on request.

![Dengue Fever/Severe Dengue Fever/Chikungunya Fever · Dengue fever and severe dengue (dengue hemorrhagic fever [DHF] and dengue shock syndrome [DSS]) are caused by any of four closely](https://img.pdfslide.net/doc/110x75/5e87bf3e7a86e85d3b149cd7/dengue-feversevere-dengue-feverchikungunya-dengue-fever-and-severe-dengue-dengue.jpg)














