Embed Size (px)
Citation preview

Da
ST
a
ARRA
KACNHED
1
nhgaftt
baia1r
0d
Journal of Neuroscience Methods 182 (2009) 17–24
Contents lists available at ScienceDirect
Journal of Neuroscience Methods
journa l homepage: www.e lsev ier .com/ locate / jneumeth
evelopment and optimization of a high-throughput electrophysiologyssay for neuronal �4�2 nicotinic receptors
erguei S. Sidach, Nikolai B. Fedorov, Patrick M. Lippiello, Merouane Bencherif ∗
argacept, Inc., 200 East First Street, Suite 300 Winston-Salem, NC 27101, United States
r t i c l e i n f o
rticle history:eceived 4 March 2009eceived in revised form 20 May 2009ccepted 20 May 2009
eywords:cetylcholineholinergicicotinic receptorsigh-throughput screeninglectrophysiologyrug discovery
a b s t r a c t
Historically, the identification of �4�2 nicotinic acetylcholine receptor ligands has been based on high-throughput radioligand binding, rubidium efflux assays and Ca++ flux assays using a fluorometric imagingplate reader (FLIPR). Among other approaches, low-throughput electrophysiological assays in Xenopusoocytes and two channel application “liquid filament” systems for mammalian cells have been commonlyused. More recent technical innovations that have been introduced into the field of electrophysiol-ogy allow for automated simultaneous multi-channel operation. Here we report the development andoptimization of a high-throughput electrophysiological assay for identifying functionally active �4�2nicotinic receptor ligands using such a system. Characterization of the test system yielded results compa-rable to those obtained by other investigators using conventional electrophysiological assays. For example,the concentration–response relationships obtained for �4�2 receptor activation by acetylcholine andnicotine were best described by biphasic Hill equations, and the inhibition of �4�2 receptor currents by
the nicotinic antagonist dihydro-�-erythroidine was consistent with previously published results. Func-tional up-regulation of �4�2 receptors by prolonged exposure to nicotine or lower temperature was alsoconfirmed. Using this methodology we were able to characterize the activation of �4�2 receptors bymultiple compounds in a mammalian cell expression system, exemplifying its utility for rapid identifi-cation of novel nicotinic ligands within a screening cascade. Our results demonstrate the utility of thiselectrophysiological tool for the discovery of �4�2 nicotinic acetylcholine receptor ligands with potentialapplications in numerous clinical indications.. Introduction
The high affinity �4�2 receptor subtype, one of the primaryeuronal nicotinic acetylcholine receptor subtypes in the brain,as been identified as a key therapeutic target in several neurode-enerative and cognitive disorders such as Alzheimer’s disease,ttention-deficit/hyperactivity disorder (ADHD) and cognitive dys-unction in schizophrenia. Consequently, in the last two decadeshe number of �4�2-selective compounds being assessed in clinicalrials has increased substantially (Arneric et al., 2007).
Traditionally the identification of nicotinic receptor ligands haseen based on high-throughput radioligand binding (Houghtling etl., 1995) and low-throughput functional assays, including rubid-
um efflux assays (Lukas and Cullen, 1988), electrophysiologicalssays in Xenopus oocytes (Harvey et al., 1997; Zwart and Vijverberg,998) and two channel application “liquid filament” systems foreceptors expressed in mammalian cell lines (Buisson and Bertrand,∗ Corresponding author. Tel.: +1 336 480 2100; fax: +1 336 480 2107.E-mail address: [email protected] (M. Bencherif).
165-0270/$ – see front matter © 2009 Elsevier B.V. All rights reserved.oi:10.1016/j.jneumeth.2009.05.014
© 2009 Elsevier B.V. All rights reserved.
2001). In recent years, a novel high-throughput assay measuringCa++ flux with a fluorometric imaging plate reader (FLIPR) was intro-duced that has proven to be a useful screening tool for identifyingnicotinic ligands (Quik et al., 1997; Fitch et al., 2003; Dunlop et al.,2007). However, fluorescence-based high-throughput screening isnot able to achieve the precision required to identify compoundsthat preferentially bind to open, closed or inactivated ion channels.The whole-cell voltage clamp technique, described almost threedecades ago (Hamill et al., 1981) has been considered to be thegold standard method for recording ion channel currents carriedthrough ligand- and voltage-gated channels. Patch clamp offersa real time direct measurement of the effects of chemical com-pounds on ion flow through these channels. It also allows currentmeasurements under conditions where repetitive stimulation (aphenomenon that naturally occurs in excitable tissues) is appliedto the test system.
Although informative and precise, traditional electrophysio-logical methods are labor and cost intensive, require a skilledinvestigator and generate very few data points per experimentalday. Several years ago, the Dynaflow® system was introduced byCellectricon (Sinclair et al., 2003; Olofsson et al., 2004). This sys-

1 roscie
ts4ii(mfbgDtfHtnp
2
2
n(gMpIFfr2eBs1gtwia2rcaio
2
o(pcEro(bbNS
8 S.S. Sidach et al. / Journal of Neu
em still required all of the components of a manual patch clampetup, but included a fast solution exchange feature using up to8 channels, with low throughput capacity. More recent technical
nnovations that have been introduced into the field of electrophys-ology include the PatchXpress® 7000A (Tao et al., 2004) and QPatchMathes, 2006). Both devices are medium-throughput fully auto-
ated multi-channel systems that allow simultaneous recordingrom 16 (PatchXpress® 7000A) or 48 (QPatch) channels. Althoughoth platforms have been used extensively for studying voltage-ated ion channels (Dubin et al., 2005; Guo and Guthrie, 2005;unlop et al., 2007; Trepakova et al., 2007; Korsgaard et al., 2009),
here are few reports of their implementation in screening cascadesor ligand-gated channels (Dunlop et al., 2007; Friis et al., 2009).ere we describe the development and optimization of a high-
hroughput assay for screening and identifying �4�2 neuronalicotinic receptor-selective ligands using a PatchXpress® 7000Aarallel patch–clamp system.
. Methods
.1. Cell culture and harvesting
The human epithelial SH-EP1 cell line stably expressing �4�2euronal nicotinic receptors was obtained from Dr. Ronald J. LukasBarrow Neurological Institute, Phoenix, AZ). Cells were routinelyrown at 37 ◦C under a 95% O2/5% CO2 atmosphere, in Dulbecco’sodified Eagle’s Medium (DMEM) (Invitrogen, Carlsbad, CA) sup-
lemented with 10% heat-inactivated fetal bovine serum (FBS,nvitrogen) and 130 ng/mL Geneticin (MediaTech, Manassas, VA).or low-temperature up-regulation studies plates were incubatedor 24 h at 29 ◦C prior to the experiments. For nicotine-induced up-egulation studies, 1 �M nicotine was added to the growth medium4 h prior to the experiments. Prior to harvesting, cell monolay-rs were washed twice with pre-warmed Dulbecco’s Phosphateuffered Saline (DPBS, Invitrogen) without calcium and magne-ium. After aspiration of the solution, plates were incubated for5 min at 37 ◦C with 5 mL of Accumax (Innovative Cell Technolo-ies, San Diego, CA). After incubation, 10 mL of DMEM was addedo the plates and the cell suspensions were triturated several timesith a serological pipette. Cell suspensions were then transferred
nto a plastic 15 mL centrifuge tube and left undisturbed for 15 mint 37 ◦C. Prior to experiments, cell suspensions were centrifuged formin at 158 × g, the supernatant was discarded and the pellet was
e-suspended in 10 mL of recording medium. After another round ofentrifugation for 2 min at 158 × g, the supernatant was discardednd the cell pellet was finally re-suspended in 100 �L of record-ng medium. All experimental results presented in this study werebtained from cells with a low-passage number (10–20).
.2. Solutions
All chemicals were obtained from Sigma (St. Louis, MO) and weref ASC purity or higher. The extracellular solution was composed ofmM): 120 NaCl, 3 KCl, 2 MgCl2, 2 CaCl2, 25 d-glucose, 10 HEPES,H adjusted to 7.4 with TRIS-base. The intracellular solution wasomposed of (mM): 110 TRIS phosphate dibasic, 28 TRIS base, 11GTA, 2 MgCl2, 0.1 CaCl2, 4 Na-ATP, pH 7.3 with H3PO4. As previously
eported, this solution may help to eliminate functional rundownf �4�2 receptors (Wu et al., 2006). Test solutions of acetylcholineACh) and nicotine were prepared fresh daily in the extracellularuffer described above. Test concentrations were loaded into flat-ottomed 96-well plastic plates (Fisher Scientific, Pittsburgh, PA).on-sterile PatchXpress® 7000A pipette tips (Molecular Devices,unnyvale, CA) were used in all experiments.nce Methods 182 (2009) 17–24
2.3. Electrophysiology
All recordings were performed at room temperature (22 ± 3 ◦C).SealChipTM electrode arrays (manufactured by AVIVA Biosciencesand distributed by Molecular Devices Corporation) were loadedand automatically prepared according to the built-in procedure.Cell suspensions were triturated 4 times by an on-board Carvopipetting robot and 3 �L of cell suspension was added to eachwell. After applying the initial attraction pressure, pressure rampswere repeatedly applied until the whole-cell configuration wasestablished. After a 2 min stabilization period, the experimentalsequence was initiated. Cells were held at −60 mV and a compound-triggered voltage protocol was applied. ACh and nicotine solutionswere delivered at 100 �L/s for 2 s. Based on the published literature,the times employed for ACh and nicotine applications vary signifi-cantly (from 200 ms to 8 s). It is unlikely that peak current will bedependent on changes in duration of application within this timerange. It was also found empirically by others (Wu et al., 2004)and confirmed in the present studies that 2 s applications with theinternal and external solution compositions used did not producefunctional rundown of receptors. After the 2 s application the ago-nist was continuously washed out for 1 min by buffer infusion intothe recording chamber using a peristaltic pump, and removal bysuction. Following a 10 s wait period, the next application of drugwas performed. Test compounds were applied in half-log incre-ments, at 0.01–3000 �M for ACh and 0.01–1000 �M for nicotine.Cells with at least 50 pA of peak current and Rm ≥ 200 M� wereused for analyses. Data analyses were performed using DataXpress2.0 (Molecular Devices). Statistical analyses and curve fitting wereperformed with Prism 5.0 (GraphPad Software).
3. Results
3.1. Acetylcholine concentration–response
Typical electrophysiological responses in SH-EP1 cells stablytransfected with human �4�2 receptors are shown in Fig. 1A. Thebest fit of the concentration–response curve was obtained usingtwo Hill equations (Fig. 1B, solid line), consistent with the presenceof two known stoichiometries of the �4�2 nicotinic receptor sub-type, one having high sensitivity ([�4]2[�2]3) and the other lowsensitivity ([�4]3[�2]2) to ligand activation (Moroni et al., 2006;Briggs et al., 2006). EC50 values for the high-sensitivity and low-sensitivity receptor isoforms were 3.4 ± 0.03 �M and 40 ± 0.32 �M,respectively. These values do not differ significantly from the dataobtained by other investigators using similar test systems (Buissonet al., 1996; Nelson et al., 2003; Wu et al., 2004; Vallejo et al.,2005; Spivak et al., 2007). The maximal averaged current ampli-tude, observed in the presence of 300 �M ACh, was 0.38 ± 0.03 nA(mean ± SEM, n = 68).
The experimental paradigm used for electrophysiological stud-ies of nicotinic receptor ligands typically involves application ofACh to the same cell. Since responses to the ligand are normal-ized to the maximal response evoked by ACh, the absolute currentamplitude is rarely taken into consideration, as long as it meets theacceptance criteria adopted in a particular laboratory. In addition,when an electrophysiological experiment is performed manually,the choice of individual cells is always at the investigator’s dis-cretion. As a result, the particular population of cells (selectedon the basis of their size, shape, appearance, etc.) evaluated dur-
ing the course of an experiment may not necessarily reflect thebehavior of a more diverse population of cells. In contrast, cellselection in automated electrophysiology is not controlled by theinvestigator and the experimental data are obtained from a trulyrandomized population of cells. Evaluation of the SH-EP1/�4�2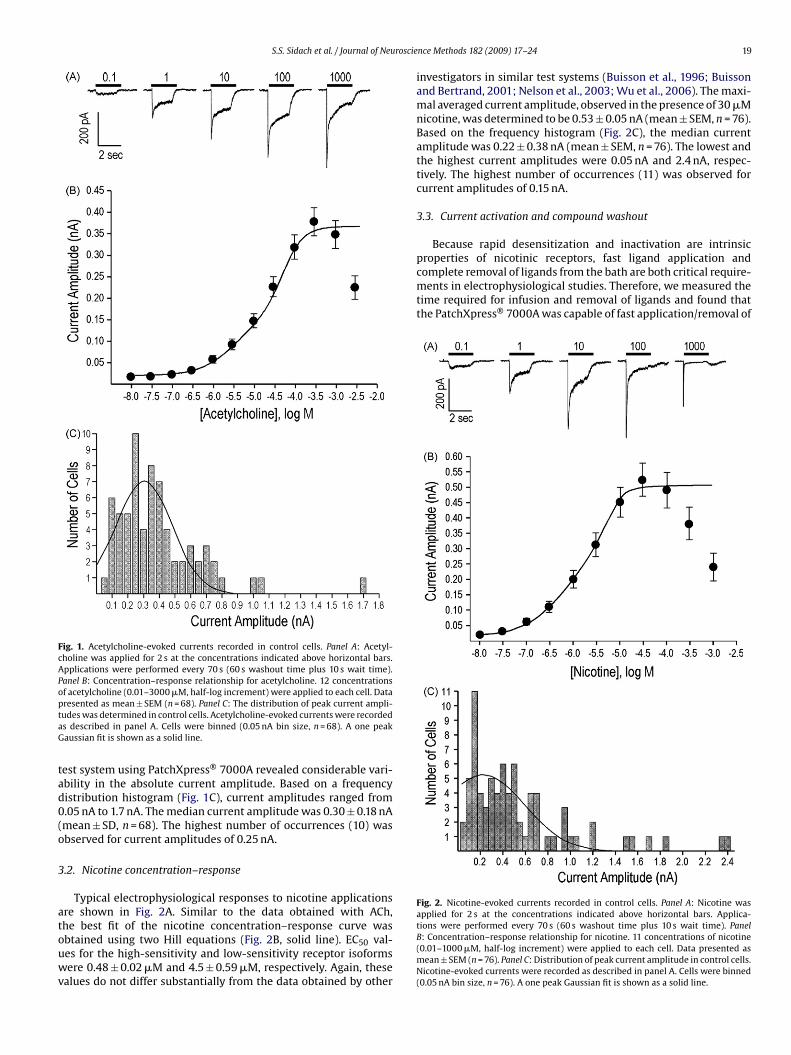
S.S. Sidach et al. / Journal of Neuroscie
Fig. 1. Acetylcholine-evoked currents recorded in control cells. Panel A: Acetyl-choline was applied for 2 s at the concentrations indicated above horizontal bars.Applications were performed every 70 s (60 s washout time plus 10 s wait time).Panel B: Concentration–response relationship for acetylcholine. 12 concentrationsof acetylcholine (0.01–3000 �M, half-log increment) were applied to each cell. DataptaG
tad0(o
3
atouwv
properties of nicotinic receptors, fast ligand application andcomplete removal of ligands from the bath are both critical require-ments in electrophysiological studies. Therefore, we measured thetime required for infusion and removal of ligands and found thatthe PatchXpress® 7000A was capable of fast application/removal of
Fig. 2. Nicotine-evoked currents recorded in control cells. Panel A: Nicotine wasapplied for 2 s at the concentrations indicated above horizontal bars. Applica-
resented as mean ± SEM (n = 68). Panel C: The distribution of peak current ampli-udes was determined in control cells. Acetylcholine-evoked currents were recordeds described in panel A. Cells were binned (0.05 nA bin size, n = 68). A one peakaussian fit is shown as a solid line.
est system using PatchXpress® 7000A revealed considerable vari-bility in the absolute current amplitude. Based on a frequencyistribution histogram (Fig. 1C), current amplitudes ranged from.05 nA to 1.7 nA. The median current amplitude was 0.30 ± 0.18 nAmean ± SD, n = 68). The highest number of occurrences (10) wasbserved for current amplitudes of 0.25 nA.
.2. Nicotine concentration–response
Typical electrophysiological responses to nicotine applicationsre shown in Fig. 2A. Similar to the data obtained with ACh,
he best fit of the nicotine concentration–response curve wasbtained using two Hill equations (Fig. 2B, solid line). EC50 val-es for the high-sensitivity and low-sensitivity receptor isoformsere 0.48 ± 0.02 �M and 4.5 ± 0.59 �M, respectively. Again, thesealues do not differ substantially from the data obtained by other
nce Methods 182 (2009) 17–24 19
investigators in similar test systems (Buisson et al., 1996; Buissonand Bertrand, 2001; Nelson et al., 2003; Wu et al., 2006). The maxi-mal averaged current amplitude, observed in the presence of 30 �Mnicotine, was determined to be 0.53 ± 0.05 nA (mean ± SEM, n = 76).Based on the frequency histogram (Fig. 2C), the median currentamplitude was 0.22 ± 0.38 nA (mean ± SEM, n = 76). The lowest andthe highest current amplitudes were 0.05 nA and 2.4 nA, respec-tively. The highest number of occurrences (11) was observed forcurrent amplitudes of 0.15 nA.
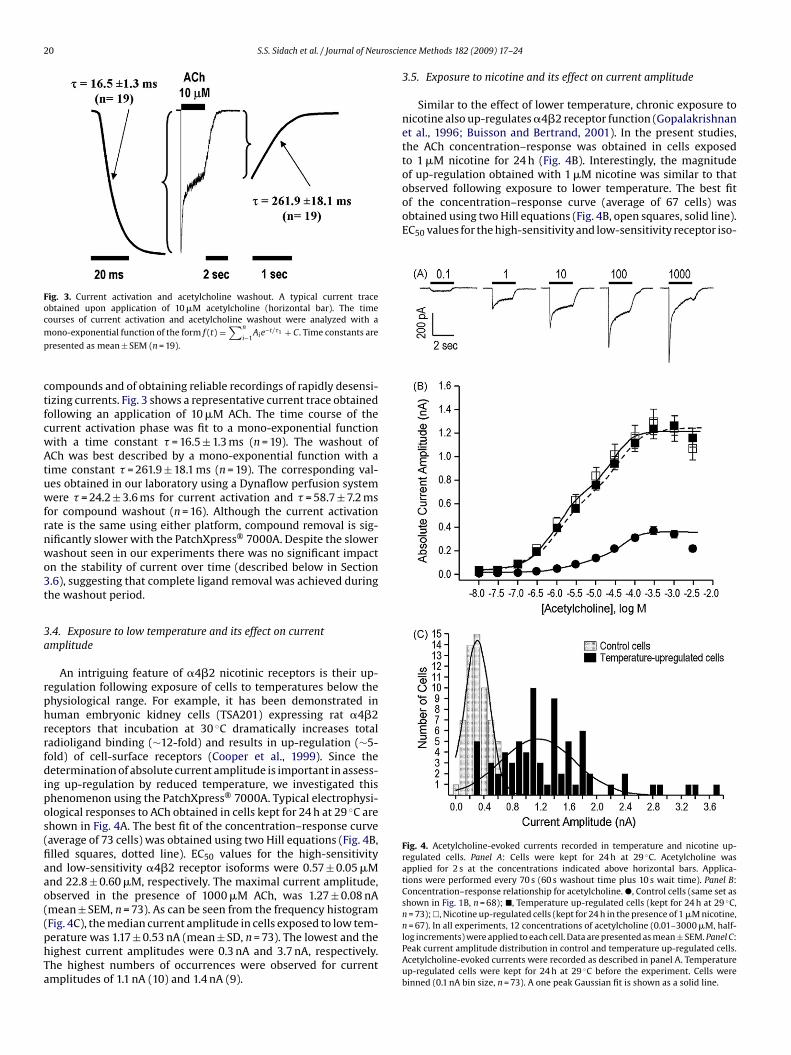
3.3. Current activation and compound washout
Because rapid desensitization and inactivation are intrinsic
tions were performed every 70 s (60 s washout time plus 10 s wait time). PanelB: Concentration–response relationship for nicotine. 11 concentrations of nicotine(0.01–1000 �M, half-log increment) were applied to each cell. Data presented asmean ± SEM (n = 76). Panel C: Distribution of peak current amplitude in control cells.Nicotine-evoked currents were recorded as described in panel A. Cells were binned(0.05 nA bin size, n = 76). A one peak Gaussian fit is shown as a solid line.
20 S.S. Sidach et al. / Journal of Neuroscie
Fig. 3. Current activation and acetylcholine washout. A typical current traceobtained upon application of 10 �M acetylcholine (horizontal bar). The timecm
p
ctfcwAtuwfrnwo3t
3a
rphrrfdipos(fiaao((phTa
observed following exposure to lower temperature. The best fitof the concentration–response curve (average of 67 cells) wasobtained using two Hill equations (Fig. 4B, open squares, solid line).EC50 values for the high-sensitivity and low-sensitivity receptor iso-
Fig. 4. Acetylcholine-evoked currents recorded in temperature and nicotine up-regulated cells. Panel A: Cells were kept for 24 h at 29 ◦C. Acetylcholine wasapplied for 2 s at the concentrations indicated above horizontal bars. Applica-tions were performed every 70 s (60 s washout time plus 10 s wait time). Panel B:Concentration–response relationship for acetylcholine. �, Control cells (same set asshown in Fig. 1B, n = 68); �, Temperature up-regulated cells (kept for 24 h at 29 ◦C,n = 73);�, Nicotine up-regulated cells (kept for 24 h in the presence of 1 �M nicotine,n = 67). In all experiments, 12 concentrations of acetylcholine (0.01–3000 �M, half-
ourses of current activation and acetylcholine washout were analyzed with aono-exponential function of the form f (t) =
∑n
i−1Aie−t/�1 + C. Time constants are
resented as mean ± SEM (n = 19).
ompounds and of obtaining reliable recordings of rapidly desensi-izing currents. Fig. 3 shows a representative current trace obtainedollowing an application of 10 �M ACh. The time course of theurrent activation phase was fit to a mono-exponential functionith a time constant � = 16.5 ± 1.3 ms (n = 19). The washout ofCh was best described by a mono-exponential function with aime constant � = 261.9 ± 18.1 ms (n = 19). The corresponding val-es obtained in our laboratory using a Dynaflow perfusion systemere � = 24.2 ± 3.6 ms for current activation and � = 58.7 ± 7.2 ms
or compound washout (n = 16). Although the current activationate is the same using either platform, compound removal is sig-ificantly slower with the PatchXpress® 7000A. Despite the slowerashout seen in our experiments there was no significant impactn the stability of current over time (described below in Section.6), suggesting that complete ligand removal was achieved duringhe washout period.
.4. Exposure to low temperature and its effect on currentmplitude
An intriguing feature of �4�2 nicotinic receptors is their up-egulation following exposure of cells to temperatures below thehysiological range. For example, it has been demonstrated inuman embryonic kidney cells (TSA201) expressing rat �4�2eceptors that incubation at 30 ◦C dramatically increases totaladioligand binding (∼12-fold) and results in up-regulation (∼5-old) of cell-surface receptors (Cooper et al., 1999). Since theetermination of absolute current amplitude is important in assess-
ng up-regulation by reduced temperature, we investigated thishenomenon using the PatchXpress® 7000A. Typical electrophysi-logical responses to ACh obtained in cells kept for 24 h at 29 ◦C arehown in Fig. 4A. The best fit of the concentration–response curveaverage of 73 cells) was obtained using two Hill equations (Fig. 4B,lled squares, dotted line). EC50 values for the high-sensitivitynd low-sensitivity �4�2 receptor isoforms were 0.57 ± 0.05 �Mnd 22.8 ± 0.60 �M, respectively. The maximal current amplitude,bserved in the presence of 1000 �M ACh, was 1.27 ± 0.08 nAmean ± SEM, n = 73). As can be seen from the frequency histogramFig. 4C), the median current amplitude in cells exposed to low tem-
erature was 1.17 ± 0.53 nA (mean ± SD, n = 73). The lowest and theighest current amplitudes were 0.3 nA and 3.7 nA, respectively.he highest numbers of occurrences were observed for currentmplitudes of 1.1 nA (10) and 1.4 nA (9).nce Methods 182 (2009) 17–24
3.5. Exposure to nicotine and its effect on current amplitude
Similar to the effect of lower temperature, chronic exposure tonicotine also up-regulates �4�2 receptor function (Gopalakrishnanet al., 1996; Buisson and Bertrand, 2001). In the present studies,the ACh concentration–response was obtained in cells exposedto 1 �M nicotine for 24 h (Fig. 4B). Interestingly, the magnitudeof up-regulation obtained with 1 �M nicotine was similar to that
log increments) were applied to each cell. Data are presented as mean ± SEM. Panel C:Peak current amplitude distribution in control and temperature up-regulated cells.Acetylcholine-evoked currents were recorded as described in panel A. Temperatureup-regulated cells were kept for 24 h at 29 ◦C before the experiment. Cells werebinned (0.1 nA bin size, n = 73). A one peak Gaussian fit is shown as a solid line.
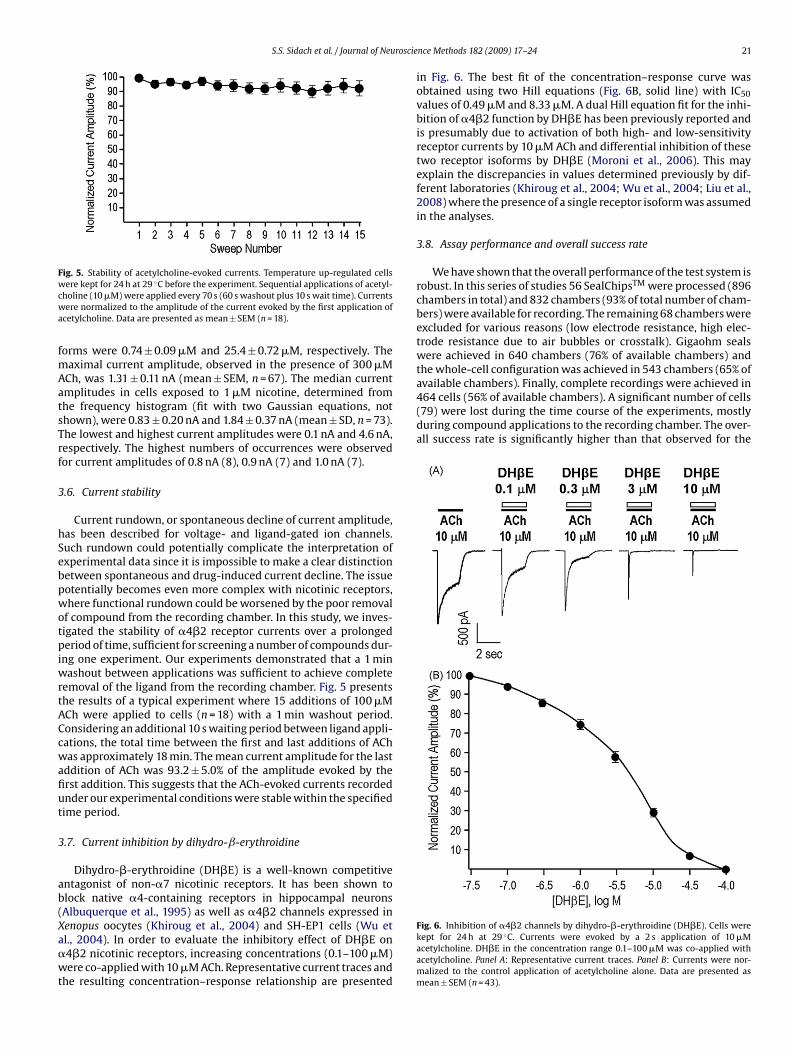
S.S. Sidach et al. / Journal of Neuroscie
Fig. 5. Stability of acetylcholine-evoked currents. Temperature up-regulated cellswcwa
fmAatsTrf
3
hSebpwotpiwrtACcwafiut
3
ab(Xa�wt
available chambers). Finally, complete recordings were achieved in464 cells (56% of available chambers). A significant number of cells(79) were lost during the time course of the experiments, mostlyduring compound applications to the recording chamber. The over-all success rate is significantly higher than that observed for the
ere kept for 24 h at 29 ◦C before the experiment. Sequential applications of acetyl-holine (10 �M) were applied every 70 s (60 s washout plus 10 s wait time). Currentsere normalized to the amplitude of the current evoked by the first application of
cetylcholine. Data are presented as mean ± SEM (n = 18).
orms were 0.74 ± 0.09 �M and 25.4 ± 0.72 �M, respectively. Theaximal current amplitude, observed in the presence of 300 �M
Ch, was 1.31 ± 0.11 nA (mean ± SEM, n = 67). The median currentmplitudes in cells exposed to 1 �M nicotine, determined fromhe frequency histogram (fit with two Gaussian equations, nothown), were 0.83 ± 0.20 nA and 1.84 ± 0.37 nA (mean ± SD, n = 73).he lowest and highest current amplitudes were 0.1 nA and 4.6 nA,espectively. The highest numbers of occurrences were observedor current amplitudes of 0.8 nA (8), 0.9 nA (7) and 1.0 nA (7).
.6. Current stability
Current rundown, or spontaneous decline of current amplitude,as been described for voltage- and ligand-gated ion channels.uch rundown could potentially complicate the interpretation ofxperimental data since it is impossible to make a clear distinctionetween spontaneous and drug-induced current decline. The issueotentially becomes even more complex with nicotinic receptors,here functional rundown could be worsened by the poor removal
f compound from the recording chamber. In this study, we inves-igated the stability of �4�2 receptor currents over a prolongederiod of time, sufficient for screening a number of compounds dur-
ng one experiment. Our experiments demonstrated that a 1 minashout between applications was sufficient to achieve complete
emoval of the ligand from the recording chamber. Fig. 5 presentshe results of a typical experiment where 15 additions of 100 �MCh were applied to cells (n = 18) with a 1 min washout period.onsidering an additional 10 s waiting period between ligand appli-ations, the total time between the first and last additions of AChas approximately 18 min. The mean current amplitude for the last
ddition of ACh was 93.2 ± 5.0% of the amplitude evoked by therst addition. This suggests that the ACh-evoked currents recordednder our experimental conditions were stable within the specifiedime period.
.7. Current inhibition by dihydro-ˇ-erythroidine
Dihydro-�-erythroidine (DH�E) is a well-known competitiventagonist of non-�7 nicotinic receptors. It has been shown tolock native �4-containing receptors in hippocampal neuronsAlbuquerque et al., 1995) as well as �4�2 channels expressed in
enopus oocytes (Khiroug et al., 2004) and SH-EP1 cells (Wu etl., 2004). In order to evaluate the inhibitory effect of DH�E on4�2 nicotinic receptors, increasing concentrations (0.1–100 �M)ere co-applied with 10 �M ACh. Representative current traces andhe resulting concentration–response relationship are presented
nce Methods 182 (2009) 17–24 21
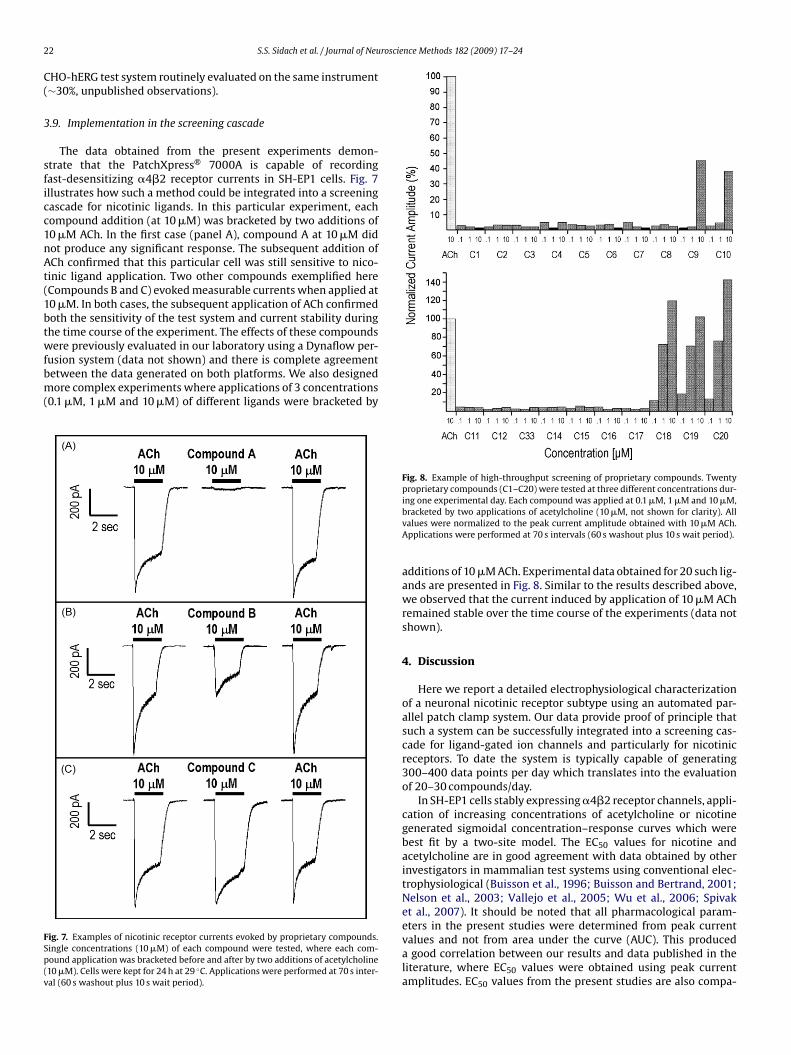
in Fig. 6. The best fit of the concentration–response curve wasobtained using two Hill equations (Fig. 6B, solid line) with IC50values of 0.49 �M and 8.33 �M. A dual Hill equation fit for the inhi-bition of �4�2 function by DH�E has been previously reported andis presumably due to activation of both high- and low-sensitivityreceptor currents by 10 �M ACh and differential inhibition of thesetwo receptor isoforms by DH�E (Moroni et al., 2006). This mayexplain the discrepancies in values determined previously by dif-ferent laboratories (Khiroug et al., 2004; Wu et al., 2004; Liu et al.,2008) where the presence of a single receptor isoform was assumedin the analyses.
3.8. Assay performance and overall success rate
We have shown that the overall performance of the test system isrobust. In this series of studies 56 SealChipsTM were processed (896chambers in total) and 832 chambers (93% of total number of cham-bers) were available for recording. The remaining 68 chambers wereexcluded for various reasons (low electrode resistance, high elec-trode resistance due to air bubbles or crosstalk). Gigaohm sealswere achieved in 640 chambers (76% of available chambers) andthe whole-cell configuration was achieved in 543 chambers (65% of
Fig. 6. Inhibition of �4�2 channels by dihydro-�-erythroidine (DH�E). Cells werekept for 24 h at 29 ◦C. Currents were evoked by a 2 s application of 10 �Macetylcholine. DH�E in the concentration range 0.1–100 �M was co-applied withacetylcholine. Panel A: Representative current traces. Panel B: Currents were nor-malized to the control application of acetylcholine alone. Data are presented asmean ± SEM (n = 43).
2 roscience Methods 182 (2009) 17–24
C(
3
sficc1nAt(1btwfbm(
FSp(v
2 S.S. Sidach et al. / Journal of Neu
HO-hERG test system routinely evaluated on the same instrument∼30%, unpublished observations).
.9. Implementation in the screening cascade
The data obtained from the present experiments demon-trate that the PatchXpress® 7000A is capable of recordingast-desensitizing �4�2 receptor currents in SH-EP1 cells. Fig. 7llustrates how such a method could be integrated into a screeningascade for nicotinic ligands. In this particular experiment, eachompound addition (at 10 �M) was bracketed by two additions of0 �M ACh. In the first case (panel A), compound A at 10 �M didot produce any significant response. The subsequent addition ofCh confirmed that this particular cell was still sensitive to nico-inic ligand application. Two other compounds exemplified hereCompounds B and C) evoked measurable currents when applied at0 �M. In both cases, the subsequent application of ACh confirmedoth the sensitivity of the test system and current stability duringhe time course of the experiment. The effects of these compoundsere previously evaluated in our laboratory using a Dynaflow per-
usion system (data not shown) and there is complete agreementetween the data generated on both platforms. We also designedore complex experiments where applications of 3 concentrations
0.1 �M, 1 �M and 10 �M) of different ligands were bracketed by
ig. 7. Examples of nicotinic receptor currents evoked by proprietary compounds.ingle concentrations (10 �M) of each compound were tested, where each com-ound application was bracketed before and after by two additions of acetylcholine10 �M). Cells were kept for 24 h at 29 ◦C. Applications were performed at 70 s inter-al (60 s washout plus 10 s wait period).
Fig. 8. Example of high-throughput screening of proprietary compounds. Twentyproprietary compounds (C1–C20) were tested at three different concentrations dur-ing one experimental day. Each compound was applied at 0.1 �M, 1 �M and 10 �M,
bracketed by two applications of acetylcholine (10 �M, not shown for clarity). Allvalues were normalized to the peak current amplitude obtained with 10 �M ACh.Applications were performed at 70 s intervals (60 s washout plus 10 s wait period).additions of 10 �M ACh. Experimental data obtained for 20 such lig-ands are presented in Fig. 8. Similar to the results described above,we observed that the current induced by application of 10 �M AChremained stable over the time course of the experiments (data notshown).
4. Discussion
Here we report a detailed electrophysiological characterizationof a neuronal nicotinic receptor subtype using an automated par-allel patch clamp system. Our data provide proof of principle thatsuch a system can be successfully integrated into a screening cas-cade for ligand-gated ion channels and particularly for nicotinicreceptors. To date the system is typically capable of generating300–400 data points per day which translates into the evaluationof 20–30 compounds/day.
In SH-EP1 cells stably expressing �4�2 receptor channels, appli-cation of increasing concentrations of acetylcholine or nicotinegenerated sigmoidal concentration–response curves which werebest fit by a two-site model. The EC50 values for nicotine andacetylcholine are in good agreement with data obtained by otherinvestigators in mammalian test systems using conventional elec-trophysiological (Buisson et al., 1996; Buisson and Bertrand, 2001;Nelson et al., 2003; Vallejo et al., 2005; Wu et al., 2006; Spivaket al., 2007). It should be noted that all pharmacological param-eters in the present studies were determined from peak current
values and not from area under the curve (AUC). This produceda good correlation between our results and data published in theliterature, where EC50 values were obtained using peak currentamplitudes. EC50 values from the present studies are also compa-
roscie
rDusmnusAp
spnoIpfa(dewcsoipm
oeobttehdo
pfsAaettftaltaecctai3alws
S.S. Sidach et al. / Journal of Neu
able to those obtained previously in our own laboratory using aynaflow perfusion system where peak current amplitudes weresed for calculations. Although beyond the scope of the presenttudies, there may be some situations where an AUC approachay be more appropriate, such as with fast-desensitizing alpha7
AChR channels (Friis et al., 2009). For this channel type EC50 val-es obtained from peak current calculations appear to display aignificant rightward shift. In general, EC50 values generated withUC typically correlate well with published values obtained fromeak current amplitudes.
We did not explore the effects of temperature in the presenttudies, primarily because the PatchXpress does not have a tem-erature control function and thus all experiments were byecessity performed at ambient temperature. The same is truef other automated electrophysiology platforms (e.g., QPatch andonWorks Quattro). To the best of our knowledge, there are noublished recordings of acetylcholine-evoked currents obtainedor neuronal �4�2 receptors at physiological temperature in vitro,lthough some limited work has been done on muscle-type nAChRsDemazumder and Dilger, 2008). Information on the temperature-ependence of drug effects for other types of ion channels isxtremely limited. For example, the effects of 15 compoundsere evaluated on cloned hERG channels expressed in HEK-293
ells and channel blockade by 2 of the drugs was temperature-ensitive (Kirsch et al., 2004). We would speculate that the effectsf most compounds tested in the present studies are temperature-nsensitive. Nevertheless, we cannot exclude the possibility that theotency of some compounds may be affected if the same experi-ents were performed at physiological temperature.We also evaluated the effects of the nicotinic antagonist DH�E
n �4�2 receptor channels stably expressed in SH-EP1 cells. DH�Exhibited biphasic concentration–response curves with IC50 valuesf 0.49 �M and 8.33 �M. Interestingly, these IC50s are compara-le to the two different values reported for a SH-EP1 – �4�2est system (Liu et al., 2008) and for a Xenopus oocytes – �4�2est system (Khiroug et al., 2004), where fitting to single-site Hillquations likely reflected the relative proportions of low- andigh-sensitivity receptors present. Our results are consistent withifferential inhibition of high- and low-sensitivity stoichiometriesf �4�2 nicotinic receptors by DH�E (Moroni et al., 2006).
Because neuronal nicotinic receptors exhibit a significantropensity for desensitization, ligand applications must be very
ast in order to elicit the best response. In manual approaches, fastolution exchange can be achieved using a “U-tube” (Alkondon andlbuquerque, 1993; Barbosa et al., 1996), a “puffer” pipette (Zhongnd Nurse, 1997) or a laminar flow system (Dynaflow) (Sinclairt al., 2003). By comparison, in the present experiments, solu-ion exchange in the recording chamber could be controlled byhe rate of ligand delivery (i.e., how fast the solution is extrudedrom the delivery pipette), as well as by the position of the pipetteip relative to the cell. Although it is appealing to use the fastestllowable delivery rate, we found that such an approach is greatlyimited by reduced cell survival. The faster the solution injection,he greater are the chances that the cell will be lost during the lig-nd application. Considering that during the time course of a typicalxperiment at least 12 additions are made to the recording chamber,ell loss could be a significant factor in determining the overall suc-ess rate. Therefore, we developed experimental conditions wherehe rate of solution exchange is comparable to that used in manualpproaches (Dynaflow system) and the percentage of cells lost dur-ng the experiments was relatively low. As indicated above (Section
.3, Fig. 3), �4�2 currents recorded under such conditions exhibitn activation rate comparable to the Dynaflow system. Althoughigand removal from the recording chamber is significantly slowerith the PatchXpress® 7000A, we did not see any impact on thetability of recording or the overall performance of the test system.
nce Methods 182 (2009) 17–24 23
With repeated applications of acetylcholine we found that a 1 minwashout is sufficient for its complete removal from the record-ing chamber, as judged by the current stability observed withinan ∼18 min time period.
Using the present approach we demonstrated functional up-regulation of �4�2 receptors resulting from prolonged exposure tonicotine or lower temperature. It should be noted that the uniquehigh-throughput ability of the automated multi-channel systemis extremely valuable for experimental models that utilize abso-lute current amplitude. As indicated above, we found significantvariability of the peak current amplitude when recordings wereobtained in a truly randomized population of cells. In control cells,the peak current amplitude for responses to acetylcholine rangedfrom 0.05 nA to 1.7 nA, whereas maximal nicotine-induced currentswere in the range of 0.05–2.4 nA. Although the average currentamplitude was significantly higher in the cells exposed to nicotineor lower temperature, there was a significant overlap with controlcells. In temperature up-regulated cells the peak current ampli-tudes in response to acetylcholine ranged from 0.3 nA to 3.7 nA,whereas in nicotine up-regulated cells the peak current ampli-tudes ranged from 0.1 nA to 4.6 nA. Because of the high-throughputcapability of PatchXpress® 7000A we were able to record currentsfrom relatively large populations of cells (67–76) and to investigatethe distribution of peak current amplitudes. As mentioned above,prolonged exposure to nicotine or lower temperature caused func-tional up-regulation of �4�2 receptors, as judged by an increase inabsolute current amplitude.
We have shown that the overall performance of the test system isrobust. In this series of studies 56 SealChipsTM were processed (896chambers in total) and 832 chambers (93% of total number of cham-bers) were available for recording. The remaining 68 chambers wereexcluded for various reasons (low electrode resistance, high elec-trode resistance due to air bubbles or crosstalk). Gigaohm sealswere achieved in 640 chambers (76% of available chambers) andthe whole-cell configuration was achieved in 543 chambers (65% ofavailable chambers). Finally, complete recordings were achieved in464 cells (56% of available chambers). A significant number of cells(79) were lost during the time course of the experiments, mostlyduring compound applications to the recording chamber. The over-all success rate is significantly higher than that observed for theCHO-hERG test system routinely evaluated on the same instrument(∼30%, unpublished observations).
Based on the present results we can conclude that availablemulti-channel automated patch–clamp technology can be appliedas a high-throughput electrophysiological tool for studying fast-desensitizing neuronal nicotinic �4�2 receptors. Our experimentalresults clearly demonstrate that agonist EC50 values are compa-rable to corresponding values obtained by conventional manualelectrophysiology assays. Using this novel high-throughput elec-trophysiological approach, the discovery and development of novelnicotinic receptor ligands should be greatly accelerated.
References
Albuquerque EX, Pereira EF, Castro NG, Alkondon M, Reinhardt S, Schroder H, et al.Nicotinic receptor function in the mammalian central nervous system. Ann N YAcad Sci 1995;757:48–72.
Alkondon M, Albuquerque EX. Diversity of nicotinic acetylcholine receptors in rathippocampal neurons. I. Pharmacological and functional evidence for distinctstructural subtypes. J Pharmacol Exp Ther 1993;265:1455–73.
Arneric SP, Holladay M, Williams M. Neuronal nicotinic receptors: a perspective ontwo decades of drug discovery research. Biochem Pharmacol 2007;74:1092–101.
Barbosa CT, Alkondon M, Aracava Y, Maelicke A, Albuquerque EX. Ligand-gated ionchannels in acutely dissociated rat hippocampal neurons with long dendrites.Neurosci Lett 1996;210:177–80.
Briggs CA, Gubbins EJ, Putman CB, Thimmapaya R, Meyer MD, Surowy CS. High-and low-sensitivity subforms of alpha4beta2 and alpha3beta2 nAChRs. J MolNeurosci 2006;30:11–2.

2 roscie
B
B
C
D
D
D
F
F
G
G
H
H
H
K
K
K
4 S.S. Sidach et al. / Journal of Neu
uisson B, Bertrand D. Chronic exposure to nicotine upregulates the human(alpha)4((beta)2 nicotinic acetylcholine receptor function. J Neurosci2001;21:1819–29.
uisson B, Gopalakrishnan M, Arneric SP, Sullivan JP, Bertrand D. Human alpha4beta2neuronal nicotinic acetylcholine receptor in HEK 293 cells: a patch–clamp study.J Neurosci 1996;16:7880–91.
ooper ST, Harkness PC, Baker ER, Millar NS. Up-regulation of cell-surfacealpha4beta2 neuronal nicotinic receptors by lower temperature and expressionof chimeric subunits. J Biol Chem 1999;274:27145–52.
emazumder D, Dilger JP. The kinetics of competitive antagonism of nicotinic acetyl-choline receptors at physiological temperature. J Physiol 2008;586:951–63.
ubin AE, Nasser N, Rohrbacher J, Hermans AN, Marrannes R, Grantham C, et al.Identifying modulators of hERG channel activity using the PatchXpress planarpatch clamp. J Biomol Screen 2005;10:168–81.
unlop J, Roncarati R, Jow B, Bothmann H, Lock T, Kowal D, et al. In vitro screeningstrategies for nicotinic receptor ligands. Biochem Pharmacol 2007;74:1172–81.
itch RW, Xiao Y, Kellar KJ, Daly JW. Membrane potential fluorescence: a rapid andhighly sensitive assay for nicotinic receptor channel function. Proc Natl Acad SciUSA 2003;100:4909–14.
riis S, Mathes C, Sunesen M, Bowlby MR, Dunlop J. Characterization of compoundson nicotinic acetylcholine receptor alpha7 channels using higher throughputelectrophysiology. J Neurosci Methods 2009;177:142–8.
opalakrishnan M, Monteggia LM, Anderson DJ, Molinari EJ, Piattoni-Kaplan M,Donnelly-Roberts D, et al. Stable expression, pharmacologic properties and reg-ulation of the human neuronal nicotinic acetylcholine alpha 4 beta 2 receptor. JPharmacol Exp Ther 1996;276:289–97.
uo L, Guthrie H. Automated electrophysiology in the preclinical evaluation of drugsfor potential QT prolongation. J Pharmacol Toxicol Methods 2005;52:123–35.
amill OP, Marty A, Neher E, Sakmann B, Sigworth FJ. Improved patch–clamp tech-niques for high-resolution current recording from cells and cell-free membranepatches. Pflugers Arch 1981;391:85–100.
arvey SC, McIntosh JM, Cartier GE, Maddox FN, Luetje CW. Determinants of speci-ficity for alpha-conotoxin MII on alpha3beta2 neuronal nicotinic receptors. MolPharmacol 1997;51:336–42.
oughtling RA, vila-Garcia MI, Kellar KJ. Characterization of (+/−)(−)[3H]epibatidinebinding to nicotinic cholinergic receptors in rat and human brain. Mol Pharmacol1995;48:280–7.
hiroug SS, Khiroug L, Yakel JL. Rat nicotinic acetylcholine receptor alpha2beta2channels: comparison of functional properties with alpha4beta2 channels inXenopus oocytes. Neuroscience 2004;124:817–22.
irsch GE, Trepakova ES, Brimecombe JC, Sidach SS, Erickson HD, Kochan MC,et al. Variability in the measurement of hERG potassium channel inhibition:effects of temperature and stimulus pattern. J Pharmacol Toxicol Methods2004;50:93–101.
orsgaard MP, Strobaek D, Christophersen P. Automated planar electrode elec-trophysiology in drug discovery: examples of the use of QPatch in basic
nce Methods 182 (2009) 17–24
characterization and high content screening on Na(v), K(Ca)2.3, and K(v)11.1channels. Comb Chem High Throughput Screen 2009;12:51–63.
Liu Q, Yu KW, Chang YC, Lukas RJ, Wu J. Agonist-induced hump current produc-tion in heterologously-expressed human alpha4beta2-nicotinic acetylcholinereceptors. Acta Pharmacol Sin 2008;29:305–19.
Lukas RJ, Cullen MJ. An isotopic rubidium ion efflux assay for the functional charac-terization of nicotinic acetylcholine receptors on clonal cell lines. Anal Biochem1988;175:212–8.
Mathes C. QPatch: the past, present and future of automated patch clamp. ExpertOpin Ther Targets 2006;10:319–27.
Moroni M, Zwart R, Sher E, Cassels BK, Bermudez I. alpha4beta2 nicotinic recep-tors with high and low acetylcholine sensitivity: pharmacology, stoichiometry,and sensitivity to long-term exposure to nicotine. Mol Pharmacol 2006;70:755–68.
Nelson ME, Kuryatov A, Choi CH, Zhou Y, Lindstrom J. Alternate stoichiometries ofalpha4beta2 nicotinic acetylcholine receptors. Mol Pharmacol 2003;63:332–41.
Olofsson J, Pihl J, Sinclair J, Sahlin E, Karlsson M, Orwar O. A microfluidics approach tothe problem of creating separate solution environments accessible from macro-scopic volumes. Anal Chem 2004;76:4968–76.
Quik M, Philie J, Choremis J. Modulation of alpha7 nicotinic receptor-mediated cal-cium influx by nicotinic agonists. Mol Pharmacol 1997;51:499–506.
Sinclair J, Olofsson J, Phil J, Orwar O. Stabilization of high-resistance seals inpatch–clamp recordings by laminar flow. Anal Chem 2003;75:6718–22.
Spivak CE, Lupica CR, Oz M. The endocannabinoid anandamide inhibits thefunction of alpha4beta2 nicotinic acetylcholine receptors. Mol Pharmacol2007;72:1024–32.
Tao H, Santa AD, Guia A, Huang M, Ligutti J, Walker G, et al. Automated tight sealelectrophysiology for assessing the potential hERG liability of pharmaceuticalcompounds. Assay Drug Dev Technol 2004;2:497–506.
Trepakova ES, Malik MG, Imredy JP, Penniman JR, Dech SJ, Salata JJ. Applicationof PatchXpress planar patch clamp technology to the screening of new drugcandidates for cardiac KCNQ1/KCNE1 (I Ks) activity. Assay Drug Dev Technol2007;5:617–27.
Vallejo YF, Buisson B, Bertrand D, Green WN. Chronic nicotine exposure upregulatesnicotinic receptors by a novel mechanism. J Neurosci 2005;25:5563–72.
Wu J, Kuo YP, George AA, Xu L, Hu J, Lukas RJ. beta-Amyloid directly inhibitshuman alpha4beta2-nicotinic acetylcholine receptors heterologously expressedin human SH-EP1 cells. J Biol Chem 2004;279:37842–51.
Wu J, Liu Q, Yu K, Hu J, Kuo YP, Segerberg M, et al. Roles of nicotinic acetylcholinereceptor beta subunits in function of human alpha4-containing nicotinic recep-
tors. J Physiol 2006;576:103–18.Zhong H, Nurse CA. Nicotinic acetylcholine sensitivity of rat petrosal sensory neuronsin dissociated cell culture. Brain Res 1997;766:153–61.
Zwart R, Vijverberg HP. Four pharmacologically distinct subtypes of alpha4beta2nicotinic acetylcholine receptor expressed in Xenopus laevis oocytes. Mol Phar-macol 1998;54:1124–31.