Embed Size (px)
Citation preview

i
ABSTRACT
DEVELOPMENT OF GRAFTING PROTOCOLS FOR SALT-TOLERANT CACTUS PEAR (OPUNTIA FICUS-INDICA)
Protocols for the production of grafted salt-tolerant cactus pear (Opuntia
ficus-indica) were developed as follows:
Four different surface sterilization protocols and four different shoot
induction media were tested for the establishment of sterile cultures of three
patented salt-tolerant cactus pear (Opuntia ficus-indica) cultivars grown in central
California. 6-Benzylaminopurine (BA) levels were varied to assess the effect of
BA concentration on shoot induction in one of the media. A harsh protocol and a
gentle protocol both provided adequate surface sterilization. Media with 5mg/l BA
(El Finti, El Boullani, El Ayadi, Ait Aabd, & El Mousadik, 2012) and 2.5 mg/l BA
(Estrada-Luna, 1988) were the most effective at shoot induction.
Shoot proliferation, root induction and in vitro grafting were examined in
three cultivars of salt-tolerant cactus pear for the development of an in vitro
production system. Lower concentrations of BA in shoot proliferation media were
shown to eliminate the production of abnormal phenotype explants (p = 0.028). In
vitro grafting results were inconclusive because all grafts showed death of
rootstock, death of scion, or contamination.
Parallel Wedge Grafts, Horizontal Grafts, and Perpendicular Wedge Grafts
were tested in two different scion/rootstock combinations of salt-tolerant cactus
pear. Perpendicular Wedge Grafting was the most successful of the three
techniques with a success rate of 66.67%. Overall, a successful in situ grafting
protocol was developed and progress was made in the development of an in vitro
production system.
Jacob Hurst December 2016

ii

i
DEVELOPMENT OF GRAFTING PROTOCOLS FOR SALT-
TOLERANT CACTUS PEAR (OPUNTIA FICUS-INDICA)
by
Jacob Hurst
A thesis
submitted in partial
fulfillment of the requirements for the degree of
Master of Science in Plant Science
in the Jordan College of Agricultural Sciences and Technology
California State University, Fresno
December 2016

ii
© 2016 Jacob Hurst

iii
APPROVED
For the Department of Plant Science:
We, the undersigned, certify that the thesis of the following student meets the required standards of scholarship, format, and style of the university and the student's graduate degree program for the awarding of the master's degree. Jacob Hurst
Thesis Author
John Bushoven (Chair) Plant Science
Dave Goorahoo Plant Science
Gary Bañuelos USDA-ARS Parlier
For the University Graduate Committee:
Dean, Division of Graduate Studies

iv
AUTHORIZATION FOR REPRODUCTION
OF MASTER’S THESIS
X I grant permission for the reproduction of this thesis in part or in
its entirety without further authorization from me, on the
condition that the person or agency requesting reproduction
absorbs the cost and provides proper acknowledgment of
authorship.
Permission to reproduce this thesis in part or in its entirety must
be obtained from me.
Signature of thesis author:

v
ACKNOWLEDGMENTS
I thank Dr. John Bushoven for serving as my academic advisor and the
chair of my thesis committee. I thank Dr. Gary Bañuelos for being my research
mentor and a member of my thesis committee, and for allowing me unhindered
access to his patented Opuntia ficus-indica material. I also thank Dr. Dave
Goorahoo for serving on my thesis committee and for providing support and
advice during the planning and execution of my research projects.
I thank Irvin Arroyo for showing me the experimental field site, for
assisting with cladode harvest, and for providing Opuntia cultivation advice. I
thank Jerry Serimian for his advice on in situ grafting and for sharing his
knowledge about the Horizontal grafting technique.
I thank Calliope Correia for providing me with greenhouse space, pots, and
potting soil. I thank Eshan Bhardwaj for providing invaluable help in the lab with
media preparation and establishment of sterile cultures, and for providing sound
advice on sterilizing agents. I thank Mauro Trujillo and Brian Coelho for help with
initial in vitro work.
I also thank my parents, Mark and Linda Hurst, for their love and support
(both emotional and financial) throughout this endeavor.

vi
TABLE OF CONTENTS
Page
LIST OF TABLES ................................................................................................ viii
JUSTIFICATION OF STUDY AND LITERATURE REVIEW ............................ 1
Statement of Problem ........................................................................................ 1
Justification and Significance of Study ............................................................. 1
Literature Review .............................................................................................. 4
CHAPTER 1: ESTABLISHMENT OF ASEPTIC CULTURES AND SHOOT INDUCTION IN THREE CULTIVARS OF SALT-TOLERANT CACTUS PEAR (OPUNTIA FICUS-INDICA) .......................................... 15
Introduction ..................................................................................................... 15
Materials and Methods .................................................................................... 15
Results ............................................................................................................. 22
Discussion ....................................................................................................... 25
CHAPTER 2: IN VITRO SHOOT PROLIFERATION, ROOTING AND MICROGRAFTING OF THREE CULTIVARS OF SALT-TOLERANT CACTUS PEAR (OPUNTI FICUS-INDICA) ............................................. 28
Introduction ..................................................................................................... 28
Materials and Methods .................................................................................... 29
Results ............................................................................................................. 30
Discussion ....................................................................................................... 31
CHAPTER 3: IN SITU GRAFTING OF SALT-TOLERANT CACTUS PEAR (OPUNTIA FICUS-INDICA) ...................................................................... 33
Introduction ..................................................................................................... 33
Materials and Methods .................................................................................... 33
Results ............................................................................................................. 37
Discussion ....................................................................................................... 40
CHAPTER 4: CONCLUSIONS ............................................................................. 41

Page
vii vii
In Situ Grafting ................................................................................................ 41
In Vitro Production and Grafting .................................................................... 42
REFERENCES ....................................................................................................... 44

viii
LIST OF TABLES
Page
Table 1 Summary of Experimental Conditions..................................................... 21
Table 2 Contamination and Shoot Induction Rates by Sterilization Method, Media, Origin and Age of Cladodes Used for Initiation of Sterile Cultures. Shoot Initiation Rates Calculated for Clean Explants of Patented Cultivars Only .......................................................................... 23
Table 3 Shoot Proliferation and Incidence of Normal and Abnormal Phenotypes .............................................................................................. 31
Table 4 Root Production by Phenotype ................................................................ 31
Table 5 Graft Failure by Type .............................................................................. 31
Table 6 Success Rates for Three Grafting Techniques ........................................ 40

ix
LIST OF FIGURES
Page
Figure 1. Ideal in vitro production system ............................................................... 3
Figure 2. Ideal in situ production system ................................................................. 3
Figure 3. Explants, shoot initiation, growth, and proliferation .............................. 18
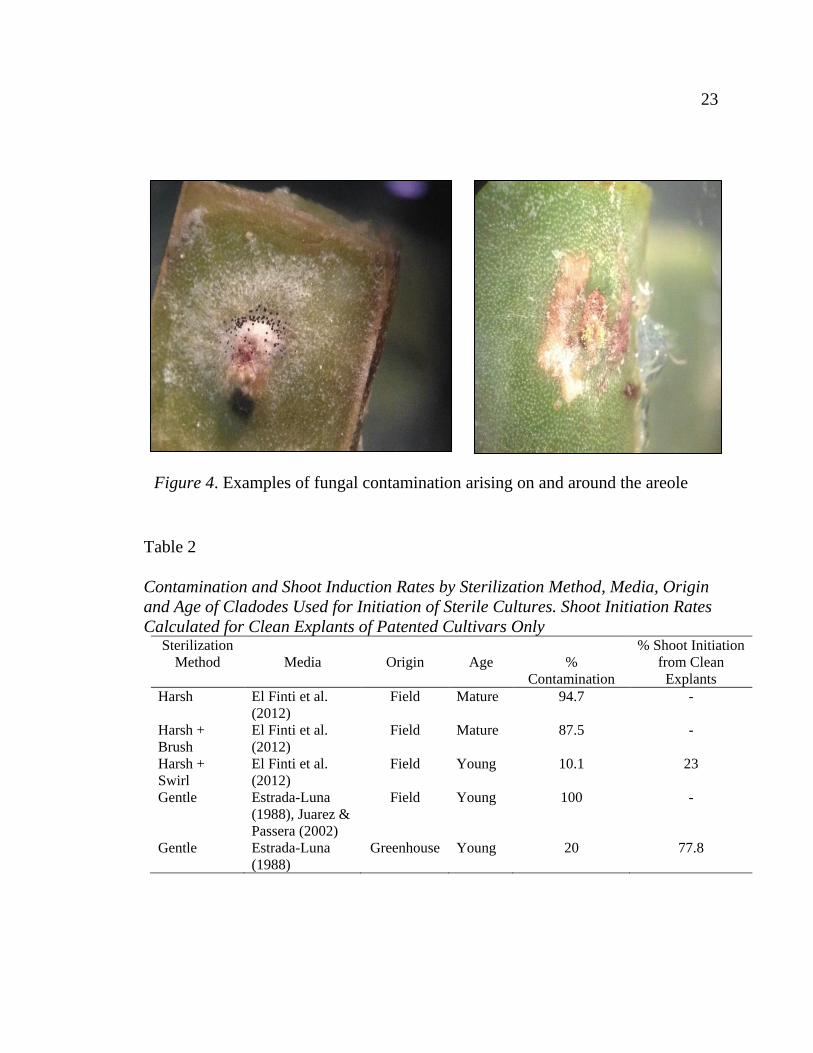
Figure 4. Examples of fungal contamination arising on and around the areole .... 23
Figure 5. Parallel wedge graft progression from graft initiation to graft failure ... 35
Figure 6. Completed horizontal grafts ................................................................... 38
Figure 7. Initial stage of perpendicular wedge graft and successful
perpendicular wedge graft ...................................................................... 39
Figure 8. A production system for field-grafted plants ......................................... 41

1
JUSTIFICATION OF STUDY AND LITERATURE REVIEW
Statement of Problem
The drought in California from 2012-2014 was the worst in recorded
history and, based on tree-ring data, was the most severe period of drought in the
past 1200 years (Griffin & Anchukaitis, 2014). Unfortunately, drought is predicted
to become more frequent and more severe in the coming decades (Diffenbaugh,
Swain, & Touma, 2015). Drought in California has a pronounced negative effect
on irrigated agriculture in the western San Joaquin Valley. Farmers there rely on
surface water to grow crops and importantly, to leach naturally-occurring salts
from their soil. With a shortage of water, the range of crops that can be grown is
severely limited and farmers are forced to fallow large tracts of land because most
conventional crops are relatively salt-sensitive and few salt-tolerant alternatives
are available (Glenn, Brown, & Blumwald, 1999).
Justification and Significance of Study
In response to these conditions, researchers with the USDA-ARS Parlier
and California State University, Fresno have developed the “Seleno” series of salt-
tolerant prickly pear cactus (Opuntia ficus-indica) (Banuelos, Freeman, & Diener,
2013a-d). This alternative crop can be grown on land that would be unsuitable for
other crops and produces unique antioxidant-rich fruit with potential anti-
carcinogenic properties (Banuelos et al., 2012) when grown on high-selenium soils
such as those in the western San Joaquin Valley. Growing this crop could allow
farmers to produce a value-added crop on land that might otherwise be left out of
production. There are four varieties in the series, named for the color of their fruit
(Seleno-Red, -Green, -Orange, and -Purple). Seleno-Orange and -Green are more
tolerant of saline conditions than the other two varieties, but their fruit are less
desirable than the larger, more globular red or purple fruit from Seleno-Red and

2 2
-Purple. Can Seleno-Red or -Purple be grafted onto Seleno-Green or -Orange to
allow for the production of desirable fruits under poor growing conditions?
Cacti have been grafted for many years, but mainly in ornamental varieties
that display a columnar growth habit. Opuntia species have not been widely
grafted for two reasons: desiccation and pathogenic fungal growth (Maldonado &
Zapien-Barragan, 1977). Opuntia spp. cladodes are flat and wide, which increases
the surface area exposed to the air relative to a cylindrical stem. This physical
characteristic leads to greater water loss and increased risk of exposure to
pathogens when the cladodes are grafted. An optimized grafting process should
reduce the negative impacts of desiccation and fungal pathogens and subsequently
allow for the production of grafted salt-tolerant O. ficus-indica plants. These
grafted plants could be a new crop for use in poor quality soils on the West Side of
the San Joaquin Valley. The purpose of this study is to develop systems for the
production of grafted cactus plants in vitro and in situ.
In an ideal in vitro production system (Figure 1), plant material would be
sterilized and then go through initial shoot induction. Thereafter, in vitro shoot
proliferation would provide a continuous supply of sterile rootstock and scion
material for in vitro grafting and allow for rapid production of grafted plants that
would be acclimated to ex vitro conditions and eventually moved to the field.
An ideal in situ production system schematically looks very similar to the
in vitro system (Figure 2), but skips sterilization and acclimation stages because it
all occurs ex vitro. Perhaps the most important difference is that the in situ system
can only be utilized during periods of active growth and stops during the winter,
while the in vitro system can be implemented year-round. However, larger plants
can be grafted in situ, which could have implications for survival in the field
(Turner, Alcorn, & Olin, 1969).
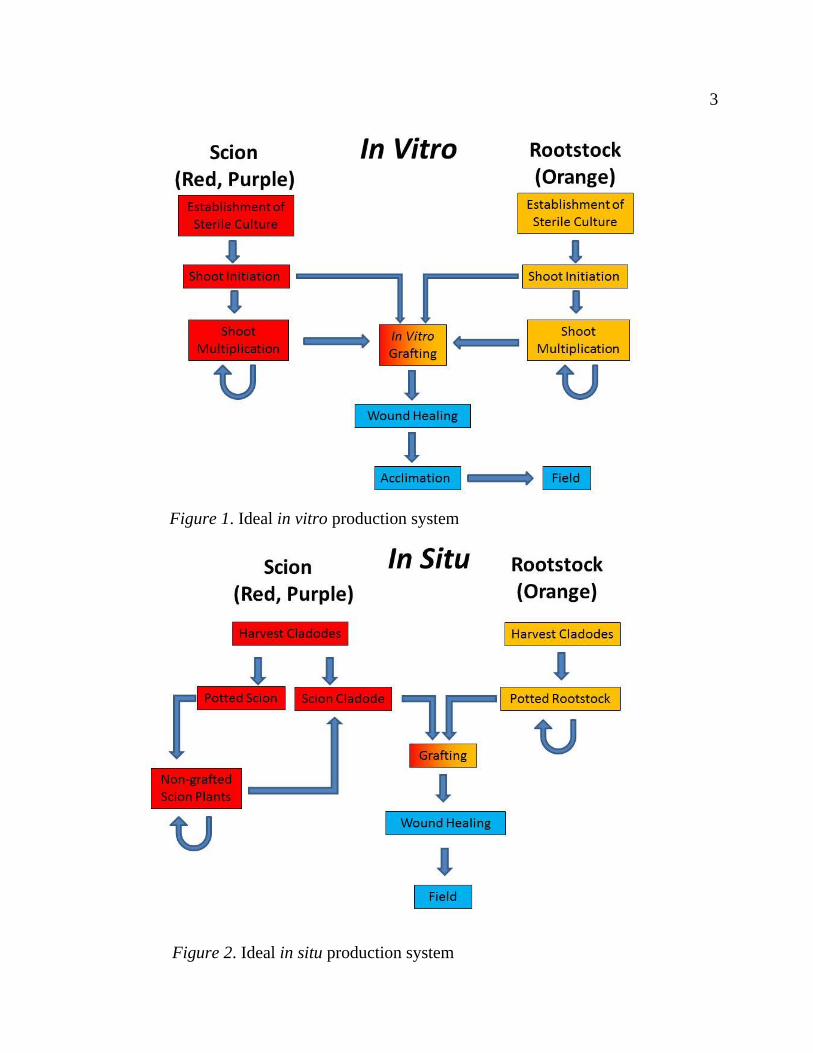
3 3
Figure 1. Ideal in vitro production system
Figure 2. Ideal in situ production system

4 4
Literature Review
The Physiology of Grafting: Graft Union Formation and Rootstock-Scion Interactions
Grafting is a well-established technique of plant propagation that has been
practiced at least as far back as 1560 BC (Mudge et al., 2009). The reasons that
justify grafting are many, but the central appeal of grafting is that it allows for the
combination of traits from two or more types of plants in a relatively short time
without the need to breed all the traits into a single plant. This technique allows for
the rapid production of specimens that are difficult to replicate through traditional
breeding and it makes it possible for certain grafted crops to thrive under
conditions that would normally have a negative impact on yield (Hartmann,
Kester, Davies, & Geneve, 2010). Grafting is conceptually straightforward, but
physiologically is a complicated process that merely begins with the physical act
of placing a scion on a rootstock. This paper will review literature relevant to the
physiology of graft union formation and the subsequent interactions between
rootstock and scion.
Graft union formation. Graft union formation is generally divided into five
stages: 1) lining up vascular cambiums of the rootstock and scion; 2) wounding
response; 3) callus bridge formation; 4) differentiation of vascular cambium across
the callus bridge, and 5) production of secondary xylem and phloem (Hartmann et
al., 2010). First, the vascular cambiums need to be lined up between the rootstock
and scion to maximize contact. Next, the damaged cells on the cut ends of the
rootstock and scion die and form a necrotic layer. This layer of dead cells contains
pectin and other sticky compounds from the cytoplasm, which act like glue and
may help keep the graft union moist (Moore & Walker, 1981a,b). Eventually, the

5 5
material from the necrotic layer is absorbed back into the living cells and the
proliferation of undifferentiated callus begins. Much like the construction of the
Transcontinental Railroad, the rootstock and scion begin forming callus at their
respective ends of the graft junction and then meet in the middle to complete the
callus bridge (Williams, 1996). After the callus forms a continuous line, a similar
ends-to-middle differentiation of xylem and phloem initials begins in the callus
cells. Once the xylem and phloem initials from the rootstock and scion come into
contact, the vascular cambiums are connected and secondary xylem and phloem
begin to form. Estrada-Luna, Lopez-Peralst, and Cardenas-Soriano (2002) include
healing of the epidermis as an additional final stage in graft union formation. It is
important to consider this step for practical settings because the epidermis serves
as the main barrier to pathogens, and therefore the vascular system is vulnerable to
infection until the exterior of the wound is sealed. Their study of Opuntia spp.
shows that under ideal conditions, the process of graft union formation takes
around 30 days from grafting to a successfully healed graft union (Estrada-Luna et
al., 2002).
Factors Affecting Rate of Successful Graft Union Formation
Provided that grafting is conducted with disease-free materials, there are
four main factors that can have an impact on successful formation of the graft
union: cambial layer contact, temperature, plant hormones, and incompatibility of
the two species being grafted. Each of these reasons will be described below.
Cambial layer contact. As previously mentioned, the vascular cambiums of
the rootstock and scion need to be appropriately aligned for graft union formation
to proceed. Appropriate alignment can mean different things in different cases. If

6 6
the rootstock and scion are of different diameters, proper alignment may involve
only one side of the vascular cambiums (Hartmann et al., 2010). To maximize the
likelihood of successful graft union formation, the cambial layers of the rootstock
and scion should have the highest degree of contact possible. Shimomura and
Fujihara (1978) showed that while graft unions successfully formed even when the
rootstock and scion vasculatures were misaligned by 1 mm, only 13% of plants
grafted in this manner formed successful graft unions. Plants that were grafted
with direct contact between the rootstock and scion vascular bundles had virtually
100% successful graft union formation (Shimomura & Fujihara, 1978).
Temperature. Temperature affects the success of graft union formation by
regulating the speed of metabolic processes within the rootstock and scion.
Extreme temperatures can also damage cells or proteins. Generally, warmer
temperatures increase the speed of graft union formation, while cooler
temperatures can delay the process (Hartmann et al., 2010). If conditions get too
hot or too cold, graft union formation will not be successful. In black walnut
(Juglans nigra), Sitton (1931) demonstrated that the optimal temperature for graft
union formation is around 75 to 80º F.
Hormones. Plant hormones play an important role in almost every aspect of
plant growth and development. The two hormones that play an important role in
the formation of graft unions are auxins and cytokinins, with the majority of
research focusing on auxins.
Auxin (IAA, or indole acetic acid) that is produced in shoot apices exhibits
a one-way basipetal flow down to the roots (Aloni, Cohen, Karni, Aktas, &
Edelstein, 2010). This flow is the product of mixed passive and active transport of
auxin molecules within plant cells. Auxin enters a cell through diffusion or active

7 7
transport and is held inside the cell by acid trapping, in which the pH of the
cytoplasm causes auxin molecules to deprotonate and lose their ability to pass
through cell membranes. Auxin then travels via basipetal transport. Once the
auxin reaches the basipetal region of the cell, active transport proteins send the
hormone out into the top of the cell below, where the process begins anew until
auxin reaches the roots. Auxin is typically associated with root formation and
differentiation of vascular tissues.
Cytokinins are generally produced in the roots and are translocated up to
shoots (Aloni et al., 2010). They are typically associated with shoot initiation and
cell division. Cytokinins are involved in budbreak after dormancy and encourage
branching by activating lateral buds.
Hormone applications for grafting. Applying hormones to graft junctions or
(in the case of auxins) to the tip of the scion material has been shown to influence
graft union formation success rates. In the Shimomura and Fujihara cactus study
(1978), the successful graft union formation rate was improved by applications of
auxin even in treatments where there was less-than-optimal rootstock-scion
cambial contact. Cummins (1997) demonstrated that Malus trees treated with
auxin at the time of grafting were more likely to successfully form a graft union
and reach a saleable size during the next season than their untreated counterparts.
Similar results have been obtained in cactus. Moghadam, Ardebili, and Rezaie
(2014) showed that applications of indole-3-butyric acid (IBA) could improve the
number of successful grafts in an intergeneric rootstock/scion combination. The
hormone applications also improved the health and vigor of the grafted plants.
Measures of vigor, such as number of buds, diameter of scion, thickness of
cambial layer, and number of activated buds, were all improved when grafted

8 8
plants received three applications of 100 ppm IBA over the course of their study.
Lower measures of vigor were exhibited in treatments with lower levels of IBA
and treatments with fewer that the maximum of three applications of IBA during
their study. There also appears to be evidence that auxin concentrations above the
100 ppm treatment begin to negatively impact graft union success and plant vigor.
Single applications of 150 ppm IBA and two applications of 150 ppm IBA had a
lower percentage of successful graft unions than the same numbers of applications
of 100 ppm IBA.
Köse and Güleryüz (2006) showed that cytokinin applications can influence
the success of graft union formation. In Vitis vinifera, application of cytokinins to
the grafting site was shown to improve the number of successful grafts compared
to a control, yielding a 100% success rate under a 250 mg/l kinetin application.
Auxins improved rooting of grafted cuttings, but were detrimental to grafting
success, reducing the success rate to 0% under many auxin treatments (Köse &
Güleryüz, 2006).
Incompatibility
Some rootstock-scion pairings do not form graft unions, while others will
successfully form a union but exhibit poor growth and eventually die. In these
cases, the rootstock and scion are said to exhibit incompatibility (Andrews &
Marquez, 1993). The likelihood of having an incompatible rootstock-scion
combination increases as the rootstock and scion become more distantly related to
each other. Intra-specific grafts tend to be more successful than interspecific
grafts, and intra-generic grafts tend to be more successful than intergeneric grafts
(Hartmann et al., 2010). Estrada-Luna et al. (2002) reported such differences in
cactus. Interspecific in vitro micrografts of Opuntia ficus-indica on O. cochinera

9 9
and O. leucotricha showed 36% to 43% less growth than conspecific micrografts,
which was thought to be due to reduced physiological compatibility. Lohar &
VandenBosch (2005) demonstrated that intergeneric grafts between Lotus
japonicas and Medicago truncatula formed one third the number of successful
graft unions compared to self-grafts of L. japonicas.
Incompatibility is caused by a physiological intolerance between different
cells, but the mechanism by which this intolerance occurs is not clear. There are
three main mechanisms (barring the presence of pathogens) proposed to explain
incompatibility: 1) toxicity between the rootstock and scion; 2) phenolics that
inhibit proper union formation; and 3) hormonal abnormalities.
Incompatibility between pear (Pyrus communis) cultivars on quince
rootstocks has been attributed to rootstock toxicity (Gur, Samish, & Lifshitz,
1968). Quince (Cydonia oblonga) produces prunasin, which breaks down to
produce cyanide, an inhibitor of cellular respiration that can lead to the death of
phloem cells. Among pear cultivars, ‘Bartlett’ is incompatible with quince, while
‘Old Home’ can be successfully grafted onto quince rootstocks. This difference in
compatibility is the result of a physiological difference; ‘Old Home’ produces an
enzyme that detoxifies prunasin, while ‘Bartlett’ lacks the enzyme and dies from
cyanide poisoning. However, cyanide-containing compounds are not present in
many species of plants, so this cannot be the sole reason for graft incompatibility.
The presence of phenolic compounds has also been linked to graft
incompatibility (Evans & Rassmussen, 1972). Phenolics are produced in response
to stress and wounding, both of which occur during grafting. Additionally, they
are involved in lignification, which is an important step in the formation of a
successful graft union.

10 10
A growing body of evidence also supports the hormonal abnormality
hypothesis. Auxin in particular is implicated in delayed incompatibility (Aloni et
al., 2008).
Rootstock-Scion Interaction
Once the graft union has successfully formed, there is a complete vascular
connection between the rootstock and scion. This allows for the transfer of
materials between the rootstock and scion via the xylem and phloem, and also via
the parenchyma cells surrounding the vasculature. It is clear that rootstock and
scion tissues communicate (Hartmann et al., 2010), as demonstrated by so-called
“rootstock effects.” Certain rootstocks are associated with increased vigor or
dwarfing when combined with certain scions, which is thought to be hormonally
mediated. Recent research has demonstrated a variety of molecules are exchanged
between rootstock and scion, including genetic material (Fuentes, Stegemann,
Golczyk, Karcher, & Bock, 2014).
Hormone signaling. Hormones are exchanged between rootstocks and
scions of grafted plants. Auxins from the shoot apices of the scion travel down to
the roots of the rootstock, cytokinins from the rootstock travel upward into the
scion, and both have effects on growth and morphology in their respective
destinations. This “hormone message concept” has been extensively tested and
now provides a working model for the examination of the many processes that
result from grafting (Aloni et al., 2010). Bangerth (1994) found that cytokinin and
auxin levels in plants are regulated through a feedback loop. As levels of IAA
(indole acetic acid) decrease, cytokinin production in the roots is stimulated. Once
cytokinin levels in the xylem rise to sufficient levels, IAA production and
translocation are stimulated. The rise in IAA then causes a reduction in cytokinin

11 11
levels in the xylem. Through this constant ebb and flow of antagonistic hormones,
an un-grafted plant maintains a balanced cytokine/auxin ratio (Sorce, Massai,
Picciarelli, & Lorenzi, 2002). However, in a grafted plant the hormone levels may
be unbalanced, which could explain the resulting “rootstock effects” seen in
grafted trees. Rootstocks that confer vigor would produce higher levels of
cytokinin relative to the scion’s auxin levels (which would also reduce overall
auxin levels), resulting in greater shoot growth and increased branching. In Van
Hooijdonk, Woolley, Warrington, and Tustin (2010), rootstocks of differing vigor
were demonstrated to affect the architecture of commercial scions of ‘Royal Gala’
apple. The dwarfing rootstock, ‘M.9’, had a similar effect on branching and shoot
growth as an application of auxin transport inhibitor. Applications of a cytokinin
(BAP) reversed the dwarfing effects of the ‘M.9’ rootstock. This effect suggests
that dwarfing rootstocks produce low concentrations of cytokinin relative to auxin
levels in the scion, perhaps due to inhibition of auxin transport. Inhibition of auxin
transport has been shown to prevent symptoms of graft incompatibility in
normally incompatible watermelon/pumpkin combinations (Aloni et al., 2008).
Genetic exchange. That grafted plants might be able to hybridize without
sexual reproduction is a long held idea, perhaps first articulated by Darwin (1868)
in his The Variation of Animals and Plants Under Domestication. Other notable
proponents of the “hybridization by grafting” theory include Hans Winkler and
Ivan Michurin. Winkler conducted work on Solanum spp. and demonstrated that
polyploid plants could be produced from grafting (Zhou & Liu, 2015). Michurin
invented the process of “mentor grafting,” in which seedlings are grafted to mature
plants (Liu, Wang, & Li, 2011; Zhou & Liu, 2015). He demonstrated that
seedlings grafted in this method developed characteristics similar to the mentor

12 12
specimen. Small pieces of genetic material (mRNA, nucleic acids) have been
observed passing between rootstocks and scions (Gahan, 2013), so it is
conceivable that more substantial translocations of genes or groups of genes could
happen. However, conclusive evidence of this phenomenon did not exist until
2014, when Fuentes et al. showed that two grafted plants of different species can
produce material that represents a new allopolyploid species, with a genome that
has the same number of chromosomes as the sum of its two parent species. In
Fuentes et al.’s study, two species of tobacco (Niccotiana glauca and N. tabacum)
were grafted together. Each plant had been transformed to express resistance to a
different antibiotic. Once a successful graft union had formed, pieces of the graft
junction were placed on a medium that contained both antibiotics. The pieces of
graft junction that successfully grew on the test medium were grown into plants
and allowed to flower. The plants derived from the dual resistant plant material
exhibited many traits that were intermediate between the two parent species. Leaf
color and shape, flower color, flower shape, and flower size all appeared as a
blend between the character states of N. glauca and N. tabacum. Plant height and
chromosome count were larger than either parent species, which is often seen in
polyploidy. However, most polyploid specimens show a straightforward doubling
of the genome and therefore contain twice the number of chromosomes as the
parent species. Since the new tobacco species (dubbed N. tabauca) is the result of
combining two different species, its chromosome count reflects that fact. N.
glauca has 24 chromosomes, N. tabacum has 48 chromosomes, and the N. tabauca
contains 72 chromosomes. This finding shows that the new species contains the
full nuclear genomes of the two parent species, rather than just the sections of
DNA that confer antibiotic resistance.

13 13
Practical Implications
Grafting is an ancient technique that remains valuable in modern
agriculture. Recent evidence has furthered our understanding of how graft unions
form (successfully and otherwise) and how rootstocks and scions communicate to
influence each other during and after graft union formation. Very recent work has
confirmed the long-held belief that grafting can produce hybrids without sexual
reproduction. Understanding the physiological underpinnings of grafting will
allow for greater manipulation of grafted plants in the field. Because much of
rootstock/scion interactions are mediated by plant hormones that are commercially
available, manipulations of grafting physiology should be relatively feasible to
implement on a large scale. Manipulation of grafting physiology has already been
conducted in apple with an eye toward commercial production (Cummins, 1997)
and other examples are sure to appear as academic findings make their way out
into industry.
Cactus Pear (Opuntia ficus-indica) Origin, Cultivation, and Uses
Opuntia ficus-indica originated in central Mexico (Casas & Barbera, 2002)
and has since spread throughout the globe and is found on all continents except
Antarctica (Inglese, Basile, & Schirra, 2002). The plant species has been used by
humans for millennia (Casas & Barbera, 2002). The succulent pads (cladodes) are
eaten as a vegetable when young and tender (Colunga Garcia-Marin, 1984), while
older cladodes have shown promise as supplemental fiber in baked goods (Ayadi,
Abdelmaksoud, Ennouri, & Attia, 2009). Cladodes of any age can be used as
forage for livestock, and can help livestock survive droughts by providing much-
needed moisture (Mondragón-Jacobo & Pérez-González, 2001). O. ficus-indica
has also been used to treat a wide range of ailments, including inflammation

14 14
(Casas & Barbera, 2002), and diabetes (Frati, Jiménez, & Ariza, 1990). The
mucilage produced from cut cladodes can be used as an adhesive and the dried
skeletons of the plants can be used as fire wood (Bravo-Hollis, 1978; Colunga
Garcia-Marin, 1984). The fruit is eaten fresh or processed into jellies, candies, and
beverages (Lee, Pyo, Ahn, & Kim, 2005; Sawaya, Khatchadourian, Safi, & Al-
Mouhammad, 1983). The total world crop production was estimated at 300,000
tons of fruit. Mexico is the largest producer, with an estimated production of
200,000 tons of fruit on an area of 50,000 ha (Flores & Gallegos, 1994). Italy,
Spain, Egypt, Morocco, and Israel are other important producers, with minor
production in South America, South Africa, Southwestern Asia, the U.S., and
Australia. Fruits are typically sold locally, although Mexico, Italy, and Chile also
provide fruit for export (Tous & Ferguson, 1996). Within the United States, a
majority of cactus pear is produced in California. Fruit production can be a very
profitable business for growers; for example, 450 acres in Monterey County
grossed around $2 million in 1998 (Crop Profile for Cactus Pear in California,
2000). Recent studies of the fruit from cacti grown on poor quality soils (e.g., high
in salinity and selenium) show that the fruit accumulates selenium in the form of
selenoamino acids, which have been suggested to possess anti-carcinogenic
properties (Banuelos et al., 2012).

15
CHAPTER 1: ESTABLISHMENT OF ASEPTIC CULTURES AND SHOOT INDUCTION IN THREE CULTIVARS OF
SALT-TOLERANT CACTUS PEAR (OPUNTIA FICUS-INDICA)
Introduction
Opuntia ficus-indica has a well-documented history of successful in vitro
propagation. However, salt-tolerant cultivars of cactus have yet to be evaluated for
in vitro propagation potential. The “Seleno” series of salt-tolerant cactus are a
potential new crop for salt-affected agricultural lands (Banuelos & Lin, 2010). In
vitro production of these unique cultivars will allow for large-scale production of
plants for field plantings and can facilitate other in vitro techniques, like
micrografting or physiological studies. The initial stages of such production
include establishment of sterile culture and shoot induction.
Materials and Methods
Plant Material
Mature overwintered cladodes and young tender cladodes of an unnamed
Opuntia ficus-indica cultivar were collected from a private residence in Del Rey,
CA, located 12 miles from the USDA-ARS in Parlier, CA for use in initial
sterilization experiments. The remaining experiments used young tender cladodes
that were collected from USDA-ARS in Parlier during the period of active growth
or produced in the greenhouses at the California State University, Fresno
Horticulture Unit from the Parlier material.
Experiment 1-Harsh Sterilization
The first relatively harsh sterilization method was as follows: whole
cladodes were immersed for 10 min in 20% bleach, 30 min in benzalkonium

16 16
chloride (1.1856%) Lysol®, and were rinsed four times with distilled water
(Mohamed-Yasseen, Barringer, Splittstoesser, & Schnell, 1995). After surface
sterilization, cladodes were cut into approximately 2cm x 2cm squares including at
least one areole, with one square per magenta box. Nineteen magenta boxes were
used, with 50 ml of medium in each box. The medium (El Finti, El Boullani, El
Ayadi, Ait Aabd, & El Mousadik, 2012) was MS (Murashige & Skoog, 1962)
supplemented with 50 g/l sucrose, 50mg/l monosodium phosphate, 40 mg/l
adenine sulfate, 3g/l agar, 5mg/l BA, adjusted to pH 5.7 and autoclaved at 121º C
for 30 min after pouring into culture vessels. Plants were maintained in a growth
room under a 16-h day. Light intensity was 500 µmol m-2 sec-1 PPFD provided by
fluorescent lamps and temperature was maintained at 25º C ± 2º C.
Contamination was assessed and recorded for 2 weeks.
Experiment 2-Harsh + Brush Sterilization
Harsh + Brush Sterilization was a modified version of Harsh Sterilization.
All steps remain the same, but whole mature cladodes were brushed with a 9:1
ratio of 90% ethanol to Lysol mixture for 5 min with a toothbrush and rinsed
under tap water before proceeding to the bleaching step. After surface sterilization,
the cladodes were cut into approximately 3 x 3 cm squares that included at least
one areole. Forty-eight culture jars with 20 ml of medium per jar were used. The
medium is of the composition laid out by El Finti et al. (2012). The medium was
adjusted to pH 5.7 and autoclaved at 121º C for 30 min after pouring into culture
vessels. Each culture jar received one square of cladode. Contamination was
assessed and recorded each week for 4 weeks.

17 17
Experiment 3-Harsh + Swirl Sterilization
Harsh + Swirl Sterilization was a modified version of Harsh + Brush
Sterilization. Instead of brushing the cladodes with a toothbrush for 5 min, the
whole cladodes were gently swirled in the 9:1 mixture of 90% ethanol to Lysol for
30 s. Tender young cladodes of the unnamed cultivar that were 1-4 inches in
length were used. After surface sterilization, the cladodes were cut into
approximately 3 x 3 cm squares with at least 1 areole. Twenty-five magenta boxes
with 40 ml of medium per box were used as culture vessels. The shoot induction
medium was made using the formulation in El Finti et al. (2012). The medium was
adjusted to pH 5.7 and autoclaved at 121º C for 30 min after pouring into culture
vessels. One piece of cladode was placed in each magenta box. Contamination
was observed and recorded each week for 4 weeks.
Experiment 4- Different BA Concentrations on El Finti et al. (2012) Medium
A second trial of Harsh + Swirl Sterilization was conducted using patented
O. ficus-indica cultivars. Tender young cladodes from cultivars Seleno-Red (252),
Seleno-Purple(248), and Seleno-Orange(255) were used. After surface
sterilization, the cladodes were cut into approximately 3 x 3 cm squares with at
least one areole. Four pieces of cladode were placed in each culture vessel (Figure
3), and 158 magenta boxes with approximately 40 ml of medium per box were
used. Fifty boxes each were allotted to cultivar 252 and 255, with 58 boxes
allotted to cultivar 248. MS medium with 50g/l sucrose, 50mg/l monosodium
phosphate, 40 mg/l adenine sulfate, and 3 g/l agar, adjusted to pH 5.7 (El Finti et
al., 2012) was used with 4 different BA concentrations: 0 mg/l (control), 5mg/l
(low), 7.5 mg/l (medium), and 10 mg/l (high). Ten magenta boxes of each of the

18 18
Figure 3. Explants, shoot initiation, growth, and proliferation

19 19
control, medium and high levels of BA were allotted to each cultivar. Twenty
boxes each of low BA were allotted to cultivars 255 and 252, while 28 boxes of
low were allotted to cultivar 248. Explants were placed on shelves under lights in
a growth room. A 16-h day was maintained, with a light intensity of 500 µmol m-2
sec-1 PPFD provided by fluorescent lamps and the temperature was maintained at
25º C ± 2º C. Contamination, health of explants, and shoot initiation were
assessed and recorded each week for 7 weeks.
Gentle Sterilization
Gentle Sterilization was as follows: whole tender young cladodes were
washed in soapy water for 5 min, then swirled in 70% ethanol for 30 s, then placed
in 10% bleach and Tween 20 for 10 min and then rinsed 3 times with sterile DI
water (Kitchen Culture Technologies Inc., 2010). This method was used 3 times as
part of alternative media formulation experiments.
Experiment 5- medium with GA3 vs medium without GA3. Surface
sterilization followed the Gentle Sterilization protocol. After surface sterilization,
young tender cladodes were cut into approximately 3 x 3 cm squares with at least
one areole. Two media, one containing gibberellic acid (Llamoca-Zarate, Aguiar,
Landsmann, & Campos, 1999) and one without gibberellic acid (El Finti et al.,
2012) were prepared for use on greenhouse-grown cladodes of cultivars 255 and
252. Specifically, the Llamoca-Zarate et al. medium was MS medium with 50 g/l
sucrose, solidified with 8 g/1 agar and supplemented with 2.22 µM BA and 1.44
µM gibberellic acid (GA3) (Llamoca-Zarate et al., 1999). The medium without
GA3 was the El Finti et al. medium. All media were adjusted to pH 5.7 and
autoclaved at 121º C for 30 min after pouring into culture vessels. Two pieces of
cladode were placed in each culture vessel. Three magenta boxes of each media

20 20
were prepared for each cultivar for a total of six boxes. Contamination and shoot
induction were observed and recorded each week for 4 weeks.
Experiment 6-medium with IBA vs medium without IBA. Surface
sterilization followed Gentle Sterilization protocol. After surface sterilization,
cladodes were cut into approximately 3 x 3 cm squares with at least one areole.
Two other media were prepared for field-grown cladodes of cultivars 248, 252, and
255. Media with indole-3-butyric acid (IBA) (Juárez & Passera, 2002) and without
IBA (Estrada-Luna, 1988) were compared. Specifically, the medium from Juárez
and Passera contained MS medium with 2.25 mg/l BA, 2.03 mg/l IBA, 30 g/l
sucrose and 8g/l agar. The Estrada-Luna (1988) medium contained MS medium
supplemented with 50 g/l sucrose, 7 g/l agar, and 2.5 g/l agar. All media were
adjusted to pH 5.7 and autoclaved at 121º C for 30 min after pouring into culture
vessels. Two to three squares were placed in each culture vessel and 30 magenta
boxes of each medium were prepared. Ten boxes of each medium were allotted to
cultivar 255, five boxes of each medium were allotted to cultivar 248, and five
boxes for each medium were allotted to cultivar 252. Contamination and shoot
induction were observed and recorded each week for 4 weeks.
Experiment 7-greenhouse-grown explants on medium without IBA.
Fifteen magenta boxes of the Estrada-Luna (1988) medium were prepared for
greenhouse-grown cladodes of cultivars 255 and 248. Ten boxes with 50 ml of
medium per box were allotted to cultivar 255 and five boxes were allotted to cultivar
248. Surface sterilization followed gentle sterilization, after which the cladodes were
cut into approximately 3 x 3 cm squares with at least one areole. Contamination and
shoot induction were observed and recorded each week for 4 weeks. Table 1
summarizes all experimental conditions.

21
Table 1
Summary of Experimental Conditions
Experiment
Sterilization
Protocol
Media
Cultivars Used
Number of Replicates
PGR Concentration
Age of
Cladodes
Origin of
Cladodes
1 Harsh El Finti et al. (2012) unnamed 19 5mg/l BA Mature Field
2 Harsh + Brush El Finti et al. (2012) unnamed 25 5mg/l BA Mature Field
3 Harsh + Swirl El Finti et al. (2012) unnamed 48 5mg/l BA Young Field
4
Harsh + Swirl
El Finti et al. (2012)
248
252
255
10 per cultivar 0mg/l BA Young Field
4
Harsh + Swirl
Modified El Finti et al.
(2012)
248
252
255
28 for 248
20 for 252
20 for 255
5mg/l BA Young Field
4
Harsh + Swirl Modified El Finti et al.
(2012)
248
252
255
10 per cultivar 7.5mg/l BA Young Field
4
Harsh + Swirl Modified El Finti et al.
(2012)
248
252
255
10 per cultivar 10mg/l BA Young Field
5
Gentle El Finti et al. (2012) 252
255
3 per cultivar 5mg/l BA Young Greenhouse
5
Gentle Llamoca-Zarate et al.
(1999)
252
255
3 per cultivar 0.5mg/l BA, 0.5 mg/l
GA3
Young Greenhouse
6
Gentle Juarez & Passera (2002) 248
252
255
5 for 248
5 for 252
10 for 255
2.25mg/l BA, 2.03 mg/l
IBA
Young Field
6
Gentle
Estrada-Luna (1988)
248
252
255
5 for 248
5 for 252
10 for 255
2.5 mg/l BA Young Field
7
Gentle Estrada-Luna (1988) 248
255
5 for 248
10 for 255
2.5 mg/l BA Young Greenhouse

22 22
Statistical Analysis
Statistical analysis was performed with IBM SPSS Version 24 software.
Contamination and shoot initiation data were recorded in a present/absent format
with present coded as ‘1’ and absent coded as ‘0’. The data were not normally
distributed, so Kruskal-Wallis and Mann-Whitney U tests were used to detect
differences between treatments.
Results
Contamination
For Harsh Sterilization, 18 boxes were contaminated (94.7%
contamination) after 2 weeks. Some explants produced shoots in contaminated
culture vessels. Fungal pathogens were the primary contaminants.
After 4 weeks, the 42 jars that had been treated with Harsh + Brush
Sterilization were contaminated (87.5% contamination). Fungal contamination
appeared in three locations within a culture vessel: 1) isolated spots on the
medium away from any explants, 2) on the edges or sides of explants with contact
to the medium, and 3) on or around the areoles of the explants without contact
with the medium. Contamination originating in the areole was noticed starting at 2
weeks, and it was the main source of new contamination at 4 weeks. At 4 weeks,
71.4% of jars that with newly documented contamination had fungal growth from
the areole (Figure 4). There was no significant difference in contamination rate
between Harsh Sterilization and Harsh + Brush Sterilization (Table 2).
Contamination for Harsh + Swirl Sterilization was 10.1%. Harsh + Swirl
Sterilization was observed to be significantly different from Harsh Sterilization
(p<0.001) and Harsh + Brush Sterilization (p<0.001). A significant difference
23 23
Table 2
Contamination and Shoot Induction Rates by Sterilization Method, Media, Origin
and Age of Cladodes Used for Initiation of Sterile Cultures. Shoot Initiation Rates
Calculated for Clean Explants of Patented Cultivars Only Sterilization
Method
Media
Origin
Age
%
Contamination
% Shoot Initiation
from Clean
Explants
Harsh El Finti et al.
(2012)
Field Mature 94.7 -
Harsh +
Brush
El Finti et al.
(2012)
Field Mature 87.5 -
Harsh +
Swirl
El Finti et al.
(2012)
Field Young 10.1 23
Gentle Estrada-Luna
(1988), Juarez &
Passera (2002)
Field Young 100 -
Gentle Estrada-Luna
(1988)
Greenhouse Young 20 77.8
Figure 4. Examples of fungal contamination arising on and around the areole

24 24
in contamination level was detected between mature and young field-grown
cladodes (p < 0.001). Overall, there was no significant difference in
contamination level between the patented cultivars. Shoot induction for
successfully sterilized boxes was observed to differ significantly (p<0.001)
between Harsh + Swirl Sterilization (23%) and Gentle Sterilization (77.8%). A
significant difference in contamination rate (p<0.001) was observed between field-
grown material and greenhouse-grown material treated with Gentle Sterilization.
There was no significant difference in the rate of shoot induction between the
medium with GA3 (Llamoca-Zarate et al., 1999) and the medium without GA3 (El
Finti et al., 2012). No meaningful comparison between the medium with IBA
(Juárez & Passera, 2002) and the medium without IBA (Estrada-Luna, 1988)
could be made because all culture vessels were contaminated, but a single shoot of
255 was produced on the medium without IBA before the culture vessel
succumbed to contamination.
In the trials using patented cultivars, two distinct morphologies of shoot
were produced: normal, elongated shoots that resembled cactus seedlings and
abnormal shoots that seemed to increase more in diameter than in length as they
grew. Some shoot multiplication occurred after the first shoots were produced.
Axillary buds on the initiated shoots were activated and produced additional
smaller side shoots. Abnormal explants from cultivar 248 were observed in
abnormal shoots on El Finti et al. (2012) medium, but not on Estrada-Luna (1988)
medium.
Different BA Concentrations in El Finti et al. (2012) Medium
Total Shoot induction for cultivars 248, 252, and 255 on El Finti et al.
(2012) medium over all BA levels was 29.3%, 26 %, and 2%, respectively. There

25 25
was no significant difference in shoot induction between cultivar 248 and cultivar
252. Cultivar 255 did differ significantly from the other two cultivars (p<0.001).
There was no significant difference in shoot induction between any of the cultivars
at the control and high BA levels. A significant difference between cultivar 255
and both the 248 and 252 cultivars was detected for the low and medium BA
concentrations (p = 0.001 &0.049, p = 0.009 & 0.029, respectively). A significant
difference was detected between cultivars 248 and 255 at the low BA
concentration (p = 0.038), but not at the medium BA level.
Discussion
Harsh sterilization and Harsh + Brush Sterilization did not provide a high
degree of surface sterilization. Harsh + Swirl Sterilization is almost identical to
Harsh Sterilization and Harsh + Brush Sterilization, yet Harsh + Swirl Sterilization
had a much lower rate of contamination. The important difference appears to be
the age of the cladodes at the time of sterilization. The mature cladodes from
Harsh Sterilization and Harsh + Brush Sterilization had been growing in the field
for at least 1 year before they were harvested, while the young tender cladodes
used in the Harsh + Swirl Sterilization protocol were less than 6 months old. The
young cladodes spent less time exposed to potential contaminants and probably
had a lower overall contaminant load. Additionally, morphological differences
between mature and young cladodes may have contributed to differences in
contamination rate. In mature cladodes, the areole is covered in densely-packed
hairs and the surrounding cuticle has visible pits that house the stomata scattered
across its surface. Both of these structures appear to harbor fungal pathogens
(Figure 4) and may protect them during surface sterilization (Garcıa-Saucedo,
Valdez-Morales, Valverde, Cruz-Hernandez, & Paredes-Lopez, 2005). These

26 26
structures are not well-developed on young cladodes and likely provide a less-
secure refuge for pathogens. Source location also appeared to play a role in
successful surface sterilization in Gentle Sterilization. Field grown plants were
universally contaminated, but 80% of greenhouse-grown cladodes were
successfully sterilized. This indicates that greenhouse-grown plants are inherently
cleaner than field-grown plants and so can be successfully sterilized with a more
gentle method.
BA concentration was an important factor in shoot induction. BA
concentrations that were too low failed to induce shooting, while BA
concentrations that were too high seemed to inhibit shoot induction. For cultivars
252 and 248, treatment with 5 mg/l BA resulted in a good rate of shoot induction,
but many of the explants produced were of abnormal morphology per culture
vessel (30.6±12.1 % and 49.5.5±15.6%, respectively). The Estrada-Luna (1988)
medium utilizes a BA level of 2.5 mg/l. This medium showed an even higher rate
of shoot induction than El Finti et al. (2012) medium and there was no incidence
of abnormal morphology in any of the induced shoots for cultivar 248. Cultivar
252 was not successfully tested on Estrada-Luna (1988) medium, but it has been
shown to respond to BA in a similar manner to cultivar 248. For cultivar 255, all
levels of BA tested using El Finti et al. medium appeared to be too high and
inhibited shoot induction. Shoots were induced on the Estrada-Luna (1988)
medium, but a very high incidence of abnormal shoots (89.3±2.1 %) was
observed. This suggests that cultivar 255 requires lower BA concentrations than
the other cultivars for successful shoot induction and to produce normal shoots.
The low rate of shoot induction on El Finti et al. medium does not agree with
findings in the literature (El Finti, 2012; Khalafalla, Abdellatef, Mohameed-
Ahmed, & Osman 2007). The contrary findings may be due to cultivar differences,

27 27
differences in harvest technique, or differences in cultural practices for source
plants between studies. Shoot induction on the Estrada-Luna (1988) medium
supported previous findings (Estrada-Luna, 1988). Abnormal shoots have been
observed in Mammilaria pectinifera, Pelecyphora aselliformis (Giusti et al.,
2002), and Cereus peruvianus (Maria de Fátima & Prioli, 1996) explants placed
on shoot induction medium, but the rate of abnormal shoot formation was very
low.
The findings of this study are instructive in the construction of an in vitro
production method for salt-tolerant cactus pear. Greenhouse-grown cladodes
should be used as the source material and sterilized with Gentle Sterilization.
Shoots could be induced in cultivars 252 and 248 on Estrada-Luna et al. (1988)
medium. The exact concentration of BA needed to induce mostly normal shoots in
cultivar 255 is as yet unknown, but will be less than 2.5 mg/l.

28
CHAPTER 2: IN VITRO SHOOT PROLIFERATION, ROOTING AND MICROGRAFTING OF THREE CULTIVARS OF SALT-
TOLERANT CACTUS PEAR (OPUNTI FICUS-INDICA)
Introduction
In vitro production of cactus pear (Opuntia ficus-indica) has been examined
for its potential in large-scale production of plants for field planting and further in
vitro studies (El Finti et al., 2012) This study was specifically aimed at developing
a cultivar-specific production system for three unique cultivars developed and
patented by the USDA and California State University, Fresno. The “Seleno”
series of prickly pear cactus (Orange, Red, and Green, with accession numbers
255, 252, and 248, respectively) were selected for salt-tolerance and have the
additional characteristic of accumulating selenium-based antioxidants in their fruit
when grown on soils contaminated with selenium. They can produce a value
added crop on saline soils and tolerate minimal irrigation with poor-quality water.
These qualities make them an attractive option for farmers in the western San
Joaquin Valley, who are facing increasing shortages of surface water for irrigation
and/or leaching of naturally-occurring salts from the soil (Diffenbaugh et al.,
2014). However, there are differences between cultivars (Banuelos et al., 2012),
with higher salt-tolerance in cultivar 255, and desired fruiting characteristics in
cultivars 252 and 248. One way to quickly combine desired characteristics is to
graft. Preliminary attempts at in situ grafting suggested that desiccation and
contamination are major obstacles to grafting success. To circumvent these issues
and produce large numbers of grafted plants in a short time, in vitro techniques
were employed. The previous section described the development of protocols for
establishment of sterile cultures, so this chapter will delve deeper into shoot
proliferation, root induction and in vitro grafting.

29 29
Materials and Methods
Shoot Proliferation
In vitro-produced shoots from the three cultivars were placed on one of two
shoot induction media. Each culture vessel received one shoot. Shoots that were
approximately 3 cm long were cut in half longitudinally and placed on the media.
Cultivars 248 (n= 3) and 255 (n=4) were placed on MS medium (Murashige &
Skoog, 1962) supplemented with 50 g/l sucrose, 7 g/l agar, and 2.5 g/l agar
(Estrada-Luna, 1988). Cultivars 248 (n= 3) and 252 (n=4) were placed on MS
medium supplemented with 50 g/L sucrose, 50mg/L monosodium phosphate, 40
mg/L adenine sulfate, 3g/L agar, 5mg/l BA. All media were adjusted to pH 5.7 and
autoclaved at 121º C for 30 min after pouring into culture vessels. Total number of
shoots produced, number of normal shoots produced, and number of abnormal
shoots produced was observed after 18 weeks.
Root Induction
In vitro-produced shoots from cultivar 255 were used for root induction.
Five explants of normal phenotype and six of abnormal phenotype shoots were
placed on a rooting medium consisting of ½ strength MS medium supplemented
with 30 g/l sucrose, 3g/l agar, and 0.5 mg/l IBA (El Finti et al., 2012). The
medium was adjusted to pH 5.7 and autoclaved at 121º C for 30 min after pouring
into culture vessels. Each culture vessel received one explant. The number of
shoots produced by each explant was recorded after 13 weeks.
In vitro Grafting
In vitro-produced shoots of cultivars 255 and 252 were selected. Six
representatives each of normal and abnormal phenotype shoots from cultivar 255
were used as rootstock material. All scion material was normal phenotype shoots

30 30
from cultivar 252. A horizontal graft was used to join the rootstock and scion. The
medium used was of the same composition as that from the root induction
experiment (El Finti et al., 2012) and was adjusted to pH 5.7 and autoclaved at
121º C for 30 min after pouring into culture vessels. Each culture vessel received
one grafted explant and was placed in specific lighting conditions.
Statistical Analysis
Statistical Analysis was performed with IBM SPSS version 24 software.
ANOVA and LSD means separation were performed on shoot proliferation data
and an Independent Samples T-test was used for the rooting data. The Mann-
Whitney U test was performed on the in vitro grafting data.
Results
No significant difference in total shoot production was observed between
treatments. No significant difference in number of normal or abnormal shoots was
observed between the cultivars on El Finti et al. (2012) medium. Significant
differences were observed in number of normal shoots, number of abnormal
shoots, and percent abnormal shoots between cultivars 248 and 255 when on
Estrada-Luna (1988) medium (p =0.028 ). Those same variables were found to be
significantly different between the cultivars on El Finti et al. medium and cultivars
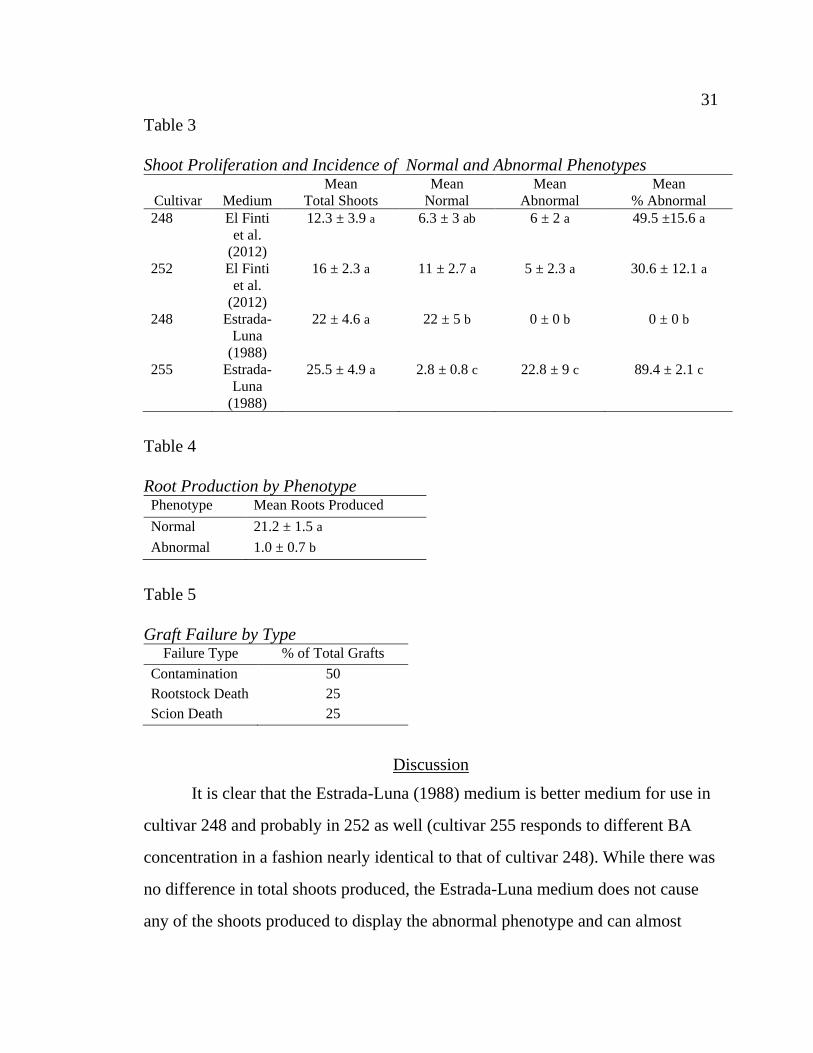
on Estrada-Luna medium (p =0.004 ) (Table 3). A significant difference in roots
produced was observed between normal and abnormal shoots of cultivar 255 (p
<0.001) (Table 4).
All in vitro micrografts failed. Distribution of failure type was not
significantly different between rootstock phenotype. 50% of grafts failed due to
contamination, 25% failed due to death of the rootstock material, and 25% failed
due to death of the scion material (Table 5).
31 31
Table 3
Shoot Proliferation and Incidence of Normal and Abnormal Phenotypes
Cultivar
Medium
Mean
Total Shoots
Mean
Normal
Mean
Abnormal
Mean
% Abnormal
248 El Finti
et al.
(2012)
12.3 ± 3.9 a 6.3 ± 3 ab 6 ± 2 a 49.5 ±15.6 a
252 El Finti
et al.
(2012)
16 ± 2.3 a 11 ± 2.7 a 5 ± 2.3 a 30.6 ± 12.1 a
248 Estrada-
Luna
(1988)
22 ± 4.6 a 22 ± 5 b 0 ± 0 b 0 ± 0 b
255 Estrada-
Luna
(1988)
25.5 ± 4.9 a 2.8 ± 0.8 c 22.8 ± 9 c 89.4 ± 2.1 c
Table 4
Root Production by Phenotype Phenotype Mean Roots Produced
Normal 21.2 ± 1.5 a
Abnormal 1.0 ± 0.7 b
Table 5
Graft Failure by Type Failure Type % of Total Grafts
Contamination 50
Rootstock Death 25
Scion Death 25
Discussion
It is clear that the Estrada-Luna (1988) medium is better medium for use in
cultivar 248 and probably in 252 as well (cultivar 255 responds to different BA
concentration in a fashion nearly identical to that of cultivar 248). While there was
no difference in total shoots produced, the Estrada-Luna medium does not cause
any of the shoots produced to display the abnormal phenotype and can almost

32 32
double the number of normal shoots produced compared to the El Finti et al.
(2012) medium. Cultivar 255 has previously been shown to respond poorly to El
Finti et al. medium and had a high incidence of abnormal shoots on Estrada-Luna
medium. This observation suggests that a lower concentration of BA might be
necessary to eliminate production of abnormal shoots. If so, it should be possible
to replicate the success of cultivar 248 in cultivar 255 and produce 100% normal
shoots. Normal shoots are preferred to abnormal shoots because they are more
vigorous in vitro. Normal shoots of cultivar 255 produced more roots than
abnormal shoots by a factor of almost 20. Unfortunately, it was impossible to
determine if the differences between normal and abnormal shoots extended to in
vitro grafting. All of the attempted grafts failed. Contamination was the cause of
failure for half of the grafts, while death of either rootstock or scion material
doomed the other half of the grafts. The contamination was most likely introduced
during grafting due to the multiple transfers from separate vessels into a shared
vessel. The low success rate of in vitro grafts reported here is contrary to what is
found in the literature. Previous research demonstrated a grafting success rate of
90% in horizontal micrografts (Estrada-Luna et al., 2002). The discrepancy in
success rate could be due to cultivar difference between studies or because
abnormal explants were included. The results of this study provide information
that is important for the design of a successful in vitro production system for
cultivars 248, 252, and 255. Abnormalities in shoot phenotype are largely the
product of the shoot induction medium and can be eliminated by lowering the
level of BA in the medium. In vitro grafting should utilize only normal phenotype
shoots should be closely monitored for contamination and death of rootstock or
scion material in the future.

33
CHAPTER 3: IN SITU GRAFTING OF SALT-TOLERANT CACTUS PEAR (OPUNTIA FICUS-INDICA)
Introduction
The “Seleno” series of Opuntia ficus-indica (which includes cultivars 248
(Seleno-Purple), 252 (Seleno-Red), and 255(Seleno-Orange)), was selected for
salt-tolerance and has the added benefit of producing nutraceutical fruit on
selenium-contaminated soils. However, all desired characteristics do not occur on
the same plant, so grafting is necessary.
Cacti are frequently grafted, but Opuntia-on-Opuntia grafts are not
commonly practiced. These types of grafts have a reputation for being difficult to
perform due to desiccation and contamination (Maldonado & Zapien-Barragan,
1977). Despite these problems, two methods have been developed specifically for
interspecific grafting of Opuntia (Huffmann, 2003; Serimian, personal
communication, May, 6, 2016). However, they have not been attempted in the
“Seleno” series. These two techniques, along with a new technique design
specifically for this experiment, will be evaluated for their effectiveness in
248/255 and 252/255 grafts for the purpose of developing an in situ production
system.
Materials and Methods
Plant Material
Plant material was collected from USDA-ARS in Parlier California. Large
mature cladodes were collected during summer. Cultivar 255 served as the
rootstock, with cultivars 252 and 248 serving as scions.

34 34
Parallel Wedge Graft
Rootstock cladodes were cured indoors at approximately 25º C for 2 weeks
prior to rooting. Rootstock cladodes were rooted in damp sand and then
transferred into 3.8 liter nursery pots filled with a well-drained potting medium.
Grafting was performed inside a closed room. Rootstock and scion
cladodes were surface sterilized with 70% ethanol before grafting. For the parallel
wedge graft, a wedge of tissue was cut from the apical portion of the rootstock
cladode. A corresponding wedge of tissue was cut from the scion material. Cutting
tools were sprayed between cuts with 70% ethanol and a flamed with a butane
torch. The cut edges of the scion wedge were secured in place against the cut
surfaces of the rootstock cladode using sterilized toothpicks. Care was taken to
ensure that a high degree of contact between rootstock and scion cut surfaces was
achieved. The graft wound was then sealed with a quick-curing silicone sealant
(Figure 5). After the silicone had cured and was no longer tacky to the touch,
grafted plants were moved to a greenhouse. Plants were watered once the potting
soil had dried completely. Five grafts were made of each scion/rootstock
combination (252/255 and 248/255). The condition of the graft union and overall
plant health were observed for 4 weeks.
Horizontal Graft
Unrooted cladodes were used for ease of handling. Grafting was performed
in a closed windowless room. Work surfaces were sprayed with 70% ethanol prior
to grafting. Both rootstock and scion cladodes were dipped in 2% bleach solution
and allowed to dry before any cuts were made. Guide lines were marked on scion
cladodes and used to determine where to cut on a corresponding rootstock cladode
to best match cladode width and thickness at the site of the graft wound and were
marked on the rootstock cladodes. Cladodes were cut horizontally in a straight line

35 35
Figure 5. Parallel wedge graft progression from graft initiation to graft
failure

36 36
across the widest face of the cladode. The cut was made in a single stroke using a
long sharp knife to get the cleanest cut possible. Between uses, the knife was
sprayed with 70% ethanol and flamed with a butane torch. The horizontal graft
used the apical piece of scion cladodes and the basal piece of each rootstock
cladode. After cutting, the cut surfaces of the rootstock and scion cladodes were
aligned to maximize physical contact. Hot melt glue for use on rubber, nylon, PVC
and metal (Ace Hardware) was used to secure the two cladode pieces together and
to seal the graft. Hot glue was dispensed from a dual heat glue gun set to “high”.
Hot glue separates from cactus cuticle if the cuticle is not heat-treated (Serimian,
personal communication, May, 6, 2016), so the heating element of the hot glue
gun was pressed against the cuticle of the cladodes in areas that would receive the
hot glue. Hot glue was not applied into the graft wound, but rather applied in a
continuous layer that extended between the rootstock and scion. Additional
support struts of glue were applied to maintain stability of the graft before
successful union formation. One to two support struts extended from the main area
of hot glue application up and down each side of the grafted cladode (Figure 6).
After the hot glue had cooled, the grafted cladodes were potted into nursery pots
filled with a well-drained potting medium. Five 252/255 grafts and six 248/255
grafts were made. One 248/255 graft utilized a much younger piece of scion
material than the other grafts. Graft union success or failure and overall cladode
health were observed for 3 weeks.
Perpendicular Wedge Graft
Grafting was performed in a closed windowless room. This protocol used
tender cladodes that still had vestigial leaves or had recently lost them. Scion
material was obtained from potted mother cladodes maintained in a greenhouse.

37 37
Scion material was grafted onto young rootstock cladodes that had emerged from
potted mother cladodes. Both rootstock and scion were sprayed with 70% ethanol
and allowed to dry before cutting. All cuts were made with a scalpel that was
sprayed with 70% ethanol and allowed to dry between grafts. A scion cladode was
cut from the mother cladode. The bottom was then trimmed into a tapered wedge
shape. The cuticle was removed to expose the photosynthetic chlorenchyma where
the trimmed section of the scion cladode was thickest (at the top of the trimmed
portion). In the rootstock, a vertical cut of the same length as the trimmed portion
of the cladode was made down the center of the rootstock cladode. The scion piece
was inserted into the vertical cut in the rootstock rotated 90º relative to the
orientation of the rootstock cladode, so that from above, the two cladodes would
appear to form a “+” shape (Figure 7). Scion material was secured in place with a
few drops of super glue at the top of both sides of the graft and an elastic band was
tied around the graft wound to maintain constant pressure. Grafted cladodes were
then moved to a greenhouse to heal. Graft health and success or failure was
observed and recorded for 2 weeks.
Statistical Analysis
Statistical Analysis was performed with IBM SPSS version 24 software.
Due to the non-parametric nature of success/failure experiments, data were
analyzed with the Kruskal-Wallis Test and Mann-Whitney U Test.
Results
Parallel Wedge Graft
After 1 week, the 252/255 grafts showed yellowing at cut edges, with little
or no yellowing on cut edges of 248/255 grafts. Necrosis was apparent in all grafts

38 38
after 2 weeks (Figure 5). Interestingly, three of the 248/255 grafts did not show
yellowing, despite the necrosis. Three weeks after grafting, all but three grafts
showed substantial necrosis and had begun to desiccate. Only one 252/255 graft
and two 248/255 grafts were still alive. After 4 weeks, all grafts had failed due to
necrosis and desiccation. A fungal pathogen was identified as the cause of the
widespread necrosis. This grafting method apparently provided two extremes of
suboptimal conditions. The sealed graft wound remained too humid and promoted
fungal growth, which caused necrosis. As the necrosis progressed past the bounds
of the silicone sealant, the plant tissue began to desiccate.
Horizontal Graft
After 1 week, one 248/255 graft had failed because the rootstock and scion
material separated. This problem continued into week 2, with a total of three failed
Figure 6. Completed horizontal grafts

39 39
248/255 grafts and one failed 252/255 graft. By week 3, all but a single 248/255
graft had failed and scion pieces were shriveled. The surviving graft successfully
formed a graft union and began to grow.
Perpendicular Wedge Graft
One graft of each scion/rootstock combination died after 2 weeks. All scion
pieces showed noticeable water loss as evidenced by their wrinkled appearance,
but those that had successfully formed a graft union eventually became turgid and
began to grow (Figure 7).
No significant difference was observed between cultivars with respect to
grafting success, so all graft types were analyzed as one group. There was a
significant association between grafting method and graft success (p = 0.001), but
no significant difference in graft success between Parallel Wedge graft and
Figure 7. Initial stage of perpendicular wedge graft and successful perpendicular
wedge graft

40 40
Horizontal graft. A significant difference in graft success was observed between
Perpendicular Wedge graft and Parallel Wedge graft (p = 0.007), and between
Perpendicular Wedge graft and Horizontal graft (p = 0.006).
Table 6
Success Rates for Three Grafting Techniques Grafting Method % Success
Parallel Wedge Graft 0.0 a
Horizontal Graft 9.09 a
Perpendicular Wedge Graft 66.67 b
Discussion
Both Parallel Wedge grafts and Horizontal grafts had displayed successful
grafting in other Opuntia species and cultivars (Huffmann, 2003; Serimian,
personal communication, May, 6, 2016). However, they do not appear to work
well on mature cladodes of the “Seleno” series cultivars. The only successful
Horizontal graft used much younger scion material than the other Horizontal
grafts, so perhaps Horizontal and Parallel Wedge grafts would work well on
younger material. However, they have drawbacks independent of graft success.
Both of these methods make it difficult to observe the graft union to monitor its
health and the sealants are not able to contract as the scion material shrinks prior
to graft union formation. The Perpendicular Wedge graft does not rely on sealants,
but rather on a snug fit between rootstock and scion pieces. This technique allows
for easy monitoring of the graft union. The lack of sealant material also allows for
good air circulation around the graft wound, which may be the reason why
Perpendicular Wedge grafts had no contamination issues. Overall, the
Perpendicular Wedge graft was superior to the other grafting techniques tested
with respect to the “Seleno” Series (Table 6). This grafting technique can now be
used as the basis for developing grafted cactus pear for field plantings.

41
CHAPTER 4: CONCLUSIONS
In Situ Grafting
Based on the results of these three studies, in situ grafting via Perpendicular
Wedge graft is the most immediately applicable technique for production of
grafted salt-tolerant cacti and is most likely to be useful for commercial
production. The Perpendicular Wedge graft is easy to perform, requires very little
in the way of equipment and supplies, and should translate well to field settings
(Figure 8). The graft site is easy to observe and graft success can be ascertained
within a week.
Figure 8. A production system for field-grafted plants

42 42
In Vitro Production and Grafting
In vitro techniques have yet to be optimized for successful large-scale
production of grafted salt-tolerant cacti. The Estrada-Luna (1988) medium (2.5
mg/l BA) would be used for the scion cultivars (248 and 252), but the rootstock
cultivar (255) needs lower levels of BA than were used in these studies to avoid
the production of abnormal shoots. Once that medium formulation is determined,
shoot production of all varieties can commence and in vitro grafting can be
conducted. In vitro grafting had three major problems in this study that impeded
successful graft union formation- death of rootstock, death of scion, and
contamination.
Rootstock death may be prevented by avoiding abnormal shoots. Avoiding
abnormal shoots should be simple because abnormal shoots were not produced in
scion material at lower levels of BA. Reduction in the rate of abnormal shoot
formation should occur for the rootstock material once an appropriate BA
concentration is determined.
Scion death may be prevented by selecting large healthy shoots for scion
material. The scion material is not in contact with the growth medium and cannot
receive nutrients from the rootstock until the vascular systems are connected, so
the grafted scion material needs to contain sufficient reserves of nutrients to
survive for the 30 days required to successfully form a graft union (Estrada-Luna
et al., 2002).
Contamination was either introduced during the grafting process or was
present-but-dormant on the parent material for the in vitro-produced shoots. If
contamination is not present after an abundantly cautious sterile technique is
followed, then the contamination was introduced during the grafting process.
Contamination that persists even after additional steps are taken to avoid external

43 43
sources of contamination indicates that dormant fungal spores may be present in
the apparently sterile cultures. This mode of contamination would be an extreme
version of the delayed-onset contamination that was observed in the first two
surface sterilization trials. Fungal spores may be present on the parent material and
lay dormant until disturbed by the grafting process. If this is determined to be the
case, in vitro–produced shoots would need to undergo further surface sterilization
and a further shoot proliferation step to ensure that contamination from the parent
material is no longer present before grafting. If cultures display contamination
after multiple rounds of surface sterilization, endogenous contamination is most
likely present. In vitro cultures would need to be screened for the pathogen and
infected explants would need to be disinfested or discarded (Leifert, Ritchie, &
Waites, 1991). Overall, the in vitro grafting system demonstrated in Estrada-
Luna (2002) does not work well for the “Seleno” series of Opuntia ficus-indica
cultivars, but can be adapted to meet their specific hormone requirements.
However, since an effective in situ grafting technique has been developed, an in
vitro production system is unnecessary until such time that the in situ production
system is rendered inefficient due to disease or unmet demand for grafted plants.

44
REFERENCES
Aloni, B., Karni, L., Deventurero, G., Levin, Z., Cohen, R., Katzir, N., Lotan-
Pompan, M., Edelstein, M., Akta, H., Turham, E., Joel, D.M., Horev, C., &
Kapulnik, Y. (2008). Physiological and biochemical changes at the rootstock-
scion interface in graft combinations between Cucurbita rootstocks and a
melon scion. The Journal of Horticultural Science & Biotechnology, 83(6),
777-783.
Aloni, B., Cohen, R., Karni, L., Aktas, H., & Edelstein, M. (2010). Hormonal
signaling in rootstock–scion interactions. Scientia Horticulturae, 127(2),
119-126.
Andrews, P. K., & Marquez, C. S. (1993). Graft incompatibility. Horticultural
Reviews, 15, 183-232.
Ayadi, M. A., Abdelmaksoud, W., Ennouri, M., & Attia, H. (2009). Cladodes
from Opuntia ficus indica as a source of dietary fiber: Effect on dough
characteristics and cake making. Industrial Crops and Products, 30(1), 40-
47.
Bangerth, F. (1994). Response of cytokinin concentration in the xylem exudate of
bean (Phaseolus vulgaris L.) plants to decapitation and auxin treatment, and
relationship to apical dominance. Planta, 194, 439-442.
Banuelos, G. S., & Lin, Z. Q. (2010). Cultivation of the Indian fig Opuntia in
selenium‐rich drainage sediments under field conditions. Soil Use and
Management, 26(2), 167-175.
Banuelos, G. S. et al. (2012). Biofortified, selenium enriched, fruit and cladode
from three Opuntia Cactus pear cultivars grown on agricultural drainage
sediment for use in nutraceutical foods. Food Chemistry, 135(1), 9-16.
Banuelos, G., Freeman, L., & Diener, J. (2013a). Opuntia plant named ‘Seleno-
Green’.U.S. Patent No. US PP24,053 P3. Washington, DC: U.S. Patent and
Trademark Office.
Banuelos, G., Freeman, L., & Diener, J. (2013b). Opuntia plant named ‘Seleno-
Orange’.U.S. Patent No. US PP24,039 P3. Washington, DC: U.S. Patent and
Trademark Office.

45 45
Banuelos, G., Freeman, L., & Diener, J. (2013c). Opuntia plant named ‘Seleno-
Purple’.U.S. Patent No. US PP24,072 P3. Washington, DC: U.S. Patent and
Trademark Office.
Banuelos, G., Freeman, L., & Diener, J. (2013d). Opuntia plant named ‘Seleno-
Red’.U.S. Patent No. US PP24,052 P3. Washington, DC: U.S. Patent and
Trademark Office.
Bravo-Hollis, H. (1978). Las cactáceas de México. Mexico City, Mexico:
Universidad Autónoma de México.
Casas, A., & Barbera, G. (2002). Mesoamerican domestication and diffusion. In
P.S. Nobel (Ed.), Cacti: biology and uses (pp. 143-162). Los Angeles, CA:
University of California Press.
Colunga García-Marín, S. P. (1984). Variación morfológica, manejo agrícola y
gradod de domesticación de Opuntia spp. en el Bajío Guanajuatense.
(Master’s Thesis). Colegio de Postgraduados, Chapingo, Mexico.
Crop Profile for Cactus Pear in California. (2000). Washington, DC: United
States Department of Agriculture.
Cummins, J. (1997). Rooting hormones may increase grafting success. Pomona,
2(34). Retrieved from http://www.rooting-hormones.com/cummins.htm
Darwin, C. 1868. The variation of animals and plants under domestication. John
Murray, London, UK.
Diffenbaugh, N. S., Swain, D. L., & Touma, D. (2015). Anthropogenic warming
has increased drought risk in California. Proceedings of the National
Academy of Sciences, 112(13), 3931-3936.
Estrada-Luna, A. A., Lopez-Peralta, C., & Cardenas-Soriano, E. (2002). In vitro
micrografting and the histology of graft union formation of selected species
of prickly pear cactus (Opuntia spp.). Scientia Horticulturae, 92(3), 317-327.
Estrada-Luna, A. A. (1988). Producción de brotes e injertación in vitro de seis
especies de nopal (Opuntia spp) originarias del altiplano Potosino-
Zacatecano (MS Thesis). Colegio de Postgraduados, Campus Montecillo,
Institución en Enseñanza e Investigación en Ciencias Agrícolas.
El Finti, A., El Boullani, R., El Ayadi, F., Ait Aabd, N., & El Mousadik, A.
(2012). Micropropagation in vitro of Opuntia ficus-indica in south of
Morocco. IJCBS, 1, 6-10.

46 46
Evans, G. E., & Rasmussen, H. P. (1972). Anatomical changes in developing graft
unions of Juniperus. Journal of the American Society of Horticultural
Science, 97, 228-232.
Flores, C. A., & Gallegos, C. (1994). The production of prickly pear in the north-
central region of Mexico. In P. Felker and J.R. Moss (Eds.), Proceedings of
the Fifth Annual Texas Prickly Pear Council (13-28). Kingsville, TX: Texas
A&M University.
Frati, A. C., Jiménez, E., & Ariza, C. R. (1990). Hypoglycemic effect of Opuntia
ficus indica in non insulin‐dependent diabetes mellitus patients. Phytotherapy
Research, 4(5), 195-197.
Fuentes, I., Stegemann, S., Golczyk, H., Karcher, D., & Bock, R. (2014).
Horizontal genome transfer as an asexual path to the formation of new
species. Nature, 511(7508), 232-235.
Gahan, P. (2013). Circulating nucleic acids: Possible inherited effects. Biology
Journal of the Linnean Society, 110, 931-948.
Garcıa-Saucedo, P. A., Valdez-Morales, M., Valverde, E.M. Cruz-Hernandez, A.,
& Paredes-Lopez, O. (2005). Regeneration of three Opuntia genotypes used
as human food. Plant Cell, Tissue and Organ Culture, 80, 215-219.
Giusti, P., Vitti, D., Fiocchetti, F., Colla, G., Saccardo, F., & Tucci, M. (2002). In
vitro propagation of three endangered cactus species. Scientia Horticulturae,
95(4), 319-332.
Glenn, E. P., Brown, J. J., & Blumwald, E. (1999). Salt tolerance and crop
potential of halophytes. Critical reviews in plant sciences, 18(2), 227-255.
Griffin, D., & Anchukaitis, K. J. (2014). How unusual is the 2012–2014 California
drought? Geophysical Research Letters, 41(24), 9017-9023.
Gur, A., Samish, R. M., & Lifshitz, E. (1968). Role of cyanogenic glycoside of
quince in incompatibility between pear cultivars and quince rootstocks.
Horticultural Research, 8(2), 113-134.
Hartmann, H. T., Kester, D. E., Davies, F. T., & Geneve, R. (2010). Plant
propagation: principles and practices (8th ed.). Upper Saddle River, NJ:
Prentice-Hall.
Huffmann, M. (2003). Cactus grafting methods. Journal of the Professional
Association for Cactus Development, 5(1), 106-114.

47 47
Inglese, P., Basile, F., & Schirra, M. (2002). Cactus pear fruit production. In P.S.
Nobel (Ed.), Cacti: biology and uses (pp. 143-162). Los Angeles: University
of California Press.
Juárez, M. C. & C. B. Passera (2002). In vitro propagation of Opuntia ellisiana
Griff. and acclimatization to field conditions. BIOCELL, 26(3), 319-324.
Khalafalla, M. M., Abdellatef, E., Mohameed-Ahmed, M. M., & Osman, M.
(2007). Micropropagation of cactus (Opuntia ficus-indica) as strategic tool
to combat desertification in arid and semi arid regions. International
Journal of Sustainable Crop Production, 2(4), 1-8.
Kitchen Culture Technologies Inc. (2010). Plant Tissue Culture for the Serious
Hobbyist, Teacher, Nurseryman and All Plant Lovers. Brush Prairie, WA:
Stiff, Carol.
Köse, C., & Güleryüz, M. (2006). Effects of auxins and cytokinins on graft union
of grapevine (Vitis vinifera). New Zealand journal of crop and horticultural
science, 34(2), 145-150.
Lee, Y. C., Pyo, Y. H., Ahn, C. K., & Kim, S. H. (2005). Food functionality of
Opuntia ficus-indica var. cultivated in Jeju Island. Preventive Nutrition and
Food Science, 10(1), 103-110.
Leifert, C., Ritchie, J. Y., & Waites, W. M. (1991). Contaminants of plant-tissue
and cell cultures. World Journal of Microbiology and Biotechnology, 7(4),
452-469.
Liu, Y., Wang, G., & Li, X. (2011). Michurin’s legacy to biological science.
Journal of biosciences, 36(1), 13-16.
Lohar, D. P., & VandenBosch, K. A. (2005). Grafting between model legumes
demonstrates roles for roots and shoots in determining nodule type and
host/rhizobia specificity. Journal of Experimental Botany, 56(416), 1643-
1650.
Llamoca-Zarate, R. M., Aguiar, L. F., Landsmann, J., & Campos, F. A. P. (1999).
Whole plant regeneration from the shoot apical meristem of Opuntia ficus-
indica Mill. (Cactaceae). Angewandte Botanik, 73(3-4), 83-85.
Maldonado, L. J., & Zapien-Barragan, M. (1977). Nopal en Mexico. Ciencia
forestal, 5(2), 36-53.

48 48
Maria de Fátima, P. S., & Prioli, J. (1996). Micropropagation of Cereus
peruvianus Mill (Cactaceae) by areole activation. In Vitro Cellular &
Developmental Biology-Plant, 32(3), 199-203.
Moghadam, A. R. L., Ardebili, Z. O., & Rezaie, L. (2014). Effect of indole butyric
acid on micrografting of cactus. African Journal of Biotechnology, 11(24),
6484-6493.
Mohamed-Yasseen, Y., Barringer, S. A., Splittstoesser, W. E., & Schnell, R. J.
(1995). Rapid propagation of tuna (Opuntia ficus-indica) and plant
establishment in soil. Plant Cell, Tissue and Organ Culture, 42(1), 117-119.
Mondragón-Jacobo, C., & Pérez-González, S. (Eds.). (2001). Cactus (Opuntia
spp.) as forage. Rome, Italy: Food & Agriculture Organization of the United
Nations.
Moore, R., & Walker, D. B. (1981a). Studies of vegetative compatibility-
incompatibility in higher plants. I. A structural study of a compatible
autograft in Sedum telephoides (Crassulaceae). American Journal of Botany.
68(6), 820-830.
Moore, R., & Walker, D. B. (1981b). Studies of vegetative compatibility-
incompatibility in higher plants. II. A structural study of an incompatible
heterograft between Sedum telephoides (Crassulaceae) and Solanum penellii
(Solanaceae).American Journal of Botany. 68(6), 831-842.
Mudge, K., Janick, J., Scofield, S., & Goldschmidt, E. E. (2009). A history of
grafting. Horticultural Reviews. 35, 437-493.
Murashige, T., & Skoog, F. (1962). A revised medium for rapid growth and bio
assays with tobacco tissue cultures. Physiologia plantarum, 15(3), 473-497.
Sawaya, W. N., Khatchadourian, H. A., Safi, W. M., & Al‐Muhammad, H. M.
(1983). Chemical characterization of prickly pear pulp, Opuntia ficus‐indica,
and the manufacturing of prickly pear jam. International Journal of Food
Science & Technology, 18(2), 183-193.
Shimomura, T., & Fujihara, K. (1978). Prevention of auxin-induced vascular
differentiation in wound callus by surface-to-surface adhesion between
calluses of stock and scion in cactus grafts. Plant and Cell Physiology, 19(5),
877-886.
Sitton, B. G. (1931). Vegetative propagation of the black walnut. Michigan
Agricultural Experiment Station Technical Bulletin. 119.

49 49
Sorce, C., Massai, R., Picciarelli, P., & Lorenzi, R. (2002). Hormonal relationships
in xylem sap of grafted and ungrafted Prunus rootstock. Scientia
Horticulturae, 93, 333-342.
Tous, J., & Ferguson, L. (1996). Mediterranean fruits. In J. Janick (Ed.), Progress
in New Crops (pp. 416-430). Arlington, VA: American Society for
Horticultural Science Press.
Turner, R. M., Alcorn, S. M., & Olin, G. (1969). Mortality of transplanted saguaro
seedlings. Ecology, 50(5), 835-844.
Van Hooijdonk, B.M., Woolley, D.J., Warrington, I.J., & Tustin, D.S. (2010).
Initial alteration of scion architecture by dwarfing apple rootstocks may
involve shoot–root–shoot signaling by auxin, gibberellin, and cytokinin.
Journal of Horticultural Science and Biotechnology. 85(1), 59-65.
Williams, J. H. (1996). A great and shining road: The epic story of the
transcontinental railroad. Lincoln, NE: University of Nebraska Press.
Zhou, X., & Liu, Y. (2015). Hybridization by grafting: A new perspective?
HortScience, 50(4), 520-521.

Fresno State
Non-Exclusive Distribution License
(to archive your thesis/dissertation electronically via the library’s eCollections database)
By submitting this license, you (the author or copyright holder) grant to Fresno State Digital
Scholar the non-exclusive right to reproduce, translate (as defined in the next paragraph), and/or
distribute your submission (including the abstract) worldwide in print and electronic format and
in any medium, including but not limited to audio or video.
You agree that Fresno State may, without changing the content, translate the submission to any
medium or format for the purpose of preservation.
You also agree that the submission is your original work, and that you have the right to grant the
rights contained in this license. You also represent that your submission does not, to the best of
your knowledge, infringe upon anyone’s copyright.
If the submission reproduces material for which you do not hold copyright and that would not be
considered fair use outside the copyright law, you represent that you have obtained the
unrestricted permission of the copyright owner to grant Fresno State the rights required by this
license, and that such third-party material is clearly identified and acknowledged within the text
or content of the submission.
If the submission is based upon work that has been sponsored or supported by an agency or
organization other than Fresno State, you represent that you have fulfilled any right of review or
other obligations required by such contract or agreement.
Fresno State will clearly identify your name as the author or owner of the submission and will not
make any alteration, other than as allowed by this license, to your submission. By typing your
name and date in the fields below, you indicate your agreement to the terms of this
distribution license.
Embargo options (fill box with an X).
Make my thesis or dissertation available to eCollections immediately upon
submission.
Embargo my thesis or dissertation for a period of 2 years from date of graduation. After
two years, I understand that my work will automatically become part of the university’s
public institutional repository.
Type full name as it appears on submission
Date
Jacob Hurst
December 8, 2016
x





























