Embed Size (px)
Citation preview

www.elsevier.com/locate/ydbio
Developmental Biology 2
Development of the central nervous system in the larvacean
Oikopleura dioica and the evolution of the chordate brain
Cristian Canestro, Susan Bassham, John Postlethwait*
Institute of Neuroscience, University of Oregon, Eugene, OR 97403, USA
Received for publication 18 March 2005, revised 11 June 2005, accepted 17 June 2005
Available online 18 August 2005
Abstract
In non-vertebrate chordates, central nervous system (CNS) development has been studied in only two taxa, the Cephalochordata and a
single Class (Ascidiacea) of the morphologically diverse Urochordata. To understand development and molecular regionalization of the brain
in a different deeply diverging chordate clade, we isolated and determined the expression patterns of orthologs of vertebrate CNS markers
(otxa, otxb, otxc, pax6, pax2/5/8a, pax2/5/8b, engrailed, and hox1) in Oikopleura dioica (Subphylum Urochordata, Class Larvacea). The
three Oikopleura otx genes are expressed similarly to vertebrate Otx paralogs, demonstrating that trans-homologs converged on similar
evolutionary outcomes by independent neo- or subfunctionalization processes during the evolution of the two taxa. This work revealed that
the Oikopleura CNS possesses homologs of the vertebrate forebrain, hindbrain, and spinal cord, but not the midbrain. Comparing larvacean
gene expression patterns to published results in ascidians disclosed important developmental differences and similarities that suggest
mechanisms of development likely present in their last common ancestor. In contrast to ascidians, the lack of a radical reorganization of the
CNS as larvaceans become adults allows us to relate embryonic gene expression patterns to three subdivisions of the adult anterior brain. Our
study of the Oikopleura brain provides new insights into chordate CNS evolution: first, the absence of midbrain is a urochordate
synapomorphy and not a peculiarity of ascidians, perhaps resulting from their drastic CNS metamorphosis; second, there is no convincing
evidence for a homolog of a midbrain–hindbrain boundary (MHB) organizer in urochordates; and third, the expression pattern of ‘‘MHB-
genes’’ in the urochordate hindbrain suggests that they function in the development of specific neurons rather than in an MHB organizer.
D 2005 Elsevier Inc. All rights reserved.
Keywords: Appendicularia; Forebrain; Midbrain; Hindbrain; Larvacea; Organizer; Chordate evolution of development; Tunicate; Subfunctionalization;
Vertebrate; Urochordate; Rhombomere 4
Introduction
In vertebrate embryos, the expression of Otx2 and Hoxb1
in the dorsal epiblast reveals the nascent forebrain +
midbrain and hindbrain before these regions become
morphologically distinct (Lumsden and Krumlauf, 1996).
Organizing centers then emerge along the anterior–posterior
(AP) axis and play a crucial role in patterning the central
nervous system (CNS). The isthmic organizer patterns the
midbrain and hindbrain primordia (reviewed by Raible and
Brand, 2004; Rhinn and Brand, 2001; Wurst and Bally-Cuif,
2001), and the rhombomere-4 (r4) organizer patterns the
0012-1606/$ - see front matter D 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.ydbio.2005.06.039
* Corresponding author.
E-mail address: [email protected] (J. Postlethwait).
surrounding hindbrain, at least in zebrafish (Maves et al.,
2002; Walshe et al., 2002).
The isthmic (or midbrain–hindbrain boundary) organizer
(MHB) develops in three phases: positioning, establishment,
and maintenance (Rhinn and Brand, 2001). In the position-
ing phase, the MHB arises during gastrulation just anterior
to the tip of the notochord between the posterior limit of the
Otx2 expression domain and the anterior limit of the
Gbx2 expression domain (Rubenstein et al., 1998). In the
establishment phase, Pax2, Fgf8, and Wnt1 expression
initiates at the Otx2 –Gbx2 interface. During the mainte-
nance phase, genes already used in the establishment phase
and their downstream targets, including En1, En2, Pax5,
and Pax8, become mutually dependent for their continued
expression and maintain the boundary (Matsunaga et al.,
85 (2005) 298 – 315

C. Canestro et al. / Developmental Biology 285 (2005) 298–315 299
2000; Scholpp et al., 2003; Schwarz et al., 1999). Caudal to
the MHB, the presence of a second organizing activity in
zebrafish (Maves et al., 2002), and possibly in other
vertebrates (Graham et al., 1993, Graham and Lumsden,
1996, Marin and Charnay, 2000a,b) has been suggested by
transplantation and ectopic expression studies; this activity
is located in rhombomere 4 (r4) of the hindbrain, and
patterns the surrounding rhombomeres by FGF signaling in
the context of the Hox code.
Many genes with roles in the MHB subsequently play
new roles in the progressive refinement of AP subdivisions
and in the differentiation of specific cell populations in the
CNS (Lumsden and Krumlauf, 1996). Most genes involved
in CNS patterning belong to multigene families, many of
which arose during large-scale gene duplication events that
accompanied, and likely facilitated, early vertebrate evolu-
tion (Garcia-Fernandez and Holland, 1994; Holland et al.,
1994; Ohno, 1970). Understanding vertebrate CNS develop-
ment is complicated because duplicated genes generally
retain some redundancy but also acquire spatial and
temporal diversification of expression patterns (Force et
al., 1999; McClintock et al., 2002; Postlethwait et al., 2004).
Nervous systems of the non-vertebrate chordates—
cephalochordates and urochordates—though simpler than
the vertebrate CNS, share with vertebrates basic deve-
lopmental genetic mechanisms (reviewed in Holland and
Chen, 2001; Shimeld and Holland, 2000; Wada and Satoh,
2001). Understanding developmental mechanisms in non-
vertebrate chordates, which possess low gene redundancy
due to their divergence before the large-scale duplication
events, can facilitate inferences for the roles of the founding
members of CNS gene families in the last common ancestor
of chordates. The Subphylum Urochordata, or Tunicata, is
the sister taxon of the Cephalochordata + Vertebrata clade
(but see Graham, 2004), and includes three Classes:
ascidians, thaliaceans, and larvaceans. Among these, only
ascidians have been intensively studied at the embryological
and molecular level.
The application of molecular and genomic tools has led
to impressive progress in understanding ascidian develop-
ment (reviewed in Canestro et al., 2003; Corbo et al., 2001;
Holland and Gibson-Brown, 2003; Jeffery, 2002; Lemaire et
al., 2002; Meinertzhagen et al., 2004; Satoh, 2003; Sordino
et al., 2001). Ascidians and amphioxus share with verte-
brates the early expression of otx anteriorly and hox1
posteriorly, with an intervening gap. In vertebrates, the
MHB forms in this gap. The role and fate of this gap in non-
vertebrate chordates, however, are problematic (reviewed in
Holland and Holland, 1999; Wada and Satoh, 2001). The
discovery that pax2/5/8a is expressed in the otx –hox1 gap
in the ascidian Halocynthia roretzi led to the proposal that
the ‘‘neck’’ of the ascidian CNS is homologous to the MHB
(Wada et al., 1998). In amphioxus, however, homologs of
vertebrate MHB markers Pax2/5/8, Engrailed, andWnt1 are
not expressed in the otx –hox1 gap suggesting that
amphioxus lacks an MHB (Holland et al., 1997; Kozmik
et al., 1999). This apparent conflict was not resolved by
recent studies of additional markers in ascidians, which
revealed substantial variability in gene expression patterns
among different ascidian species (Imai et al., 2002; Jiang
and Smith, 2002; Mazet et al., 2003; Wada et al., 1998). The
absence of functional data leaves unclear the roles of these
genes during ascidian and cephalochordate CNS develop-
ment, or whether any portion of the ascidian CNS possesses
organizing activity.
Urochordate larvae possess the defining chordate fea-
tures of notochord, dorsal hollow nerve cord, gill slits, and a
muscular, post-anal tail. In ascidians, an elaborate meta-
morphosis erases most of these chordate features as the
notochord is reabsorbed and the CNS is dramatically
restructured. It is possible that selective pressures leading
to rapid metamorphosis in ascidians have obscured or
compromised some developmental mechanisms present in
stem chordates. In contrast, larvacean urochordates (Class
Appendicularia) retain a chordate body plan as adults.
Phylogenetic relationships of urochordate classes are not
yet known for certain, but the most accepted phylogeny
places larvaceans as the basal sister group of ascidians +
thaliaceans (Christen and Braconnot, 1998; Holland et al.,
1988; Swalla et al., 2000; Wada, 1998; Wada and Satoh,
1994; but see Stach and Turbeville, 2002). Genomic
analysis has uncovered substantial differences in genome
size and syntenic relationships between Oikopleura and
ascidians (Edvardsen et al., 2005; Seo et al., 2001; Seo et
al., 2004). For these reasons, the inclusion of larvaceans in
developmental genetic investigations is essential for obtain-
ing a balanced view of development in the urochordate
Subphylum.
In the work reported here, we present the first molecular
genetic analysis of larvacean brain regionalization. Signifi-
cant homologies and differences between larvacean and
ascidian CNS developmental mechanisms can be inferred
from the comparison of gene expression patterns. Knowl-
edge of Oikopleura CNS molecular regionalization provides
new insights into chordate CNS evolution, helps illuminate
previous conflicting interpretations of ascidian CNS expres-
sion data, extends within urochordates prior suggestions
about CNS organization that previously applied only to
ascidians, and provides a basis for functional genetic
analysis.
Materials and methods
Animal culture, embryo staging, and imaging
Oikopleura dioica individuals were collected with
plankton nets in the Pacific Ocean off Charleston, Oregon
(Oregon Institute of Marine Biology) and Vancouver Island,
B.C. (Bamfield Marine Sciences Centre). Animals were
cultured for several generations in 10-Am filtered seawater
in 4-l plastic jars and fed with algal concentrates (‘‘Coral
C. Canestro et al. / Developmental Biology 285 (2005) 298–315300
and Clam diet’’, Reed Mariculture). Embryos were grown at
13 T 3-C as described (Bassham and Postlethwait, 2000).
Because developmental rate varies strongly with temper-
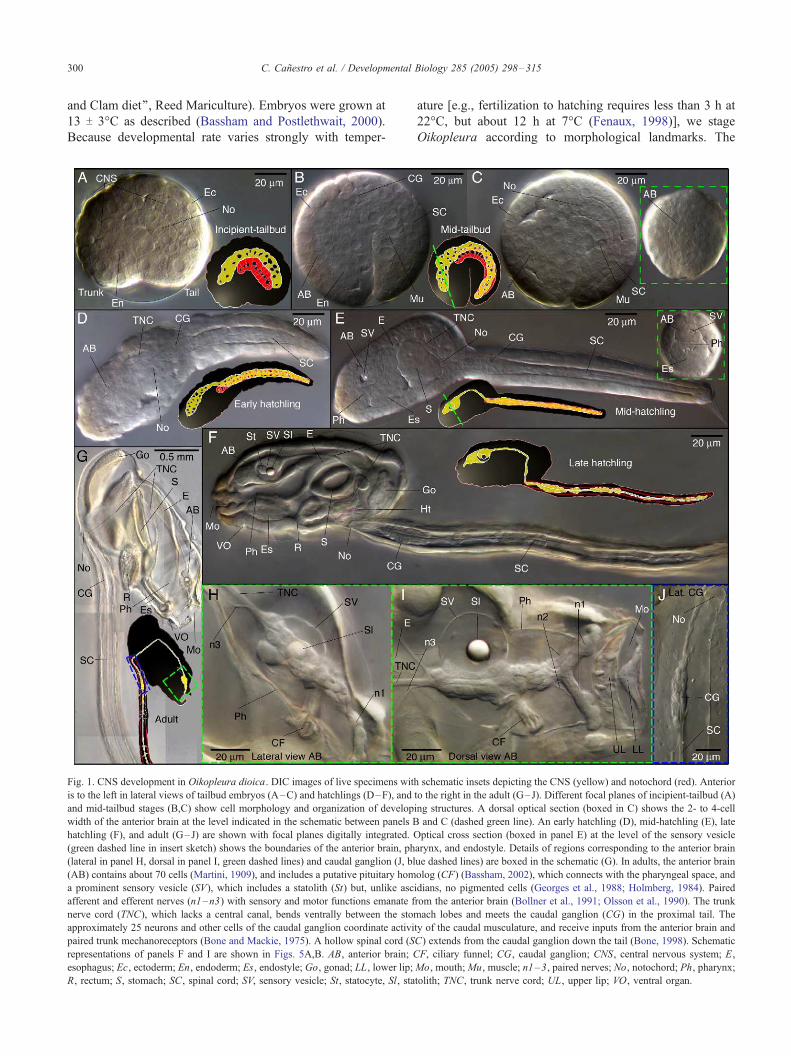
Fig. 1. CNS development in Oikopleura dioica. DIC images of live specimens wit
is to the left in lateral views of tailbud embryos (A–C) and hatchlings (D–F), and
and mid-tailbud stages (B,C) show cell morphology and organization of develop
width of the anterior brain at the level indicated in the schematic between panels B
hatchling (F), and adult (G–J) are shown with focal planes digitally integrated. O
(green dashed line in insert sketch) shows the boundaries of the anterior brain, ph
(lateral in panel H, dorsal in panel I, green dashed lines) and caudal ganglion (J, bl
(AB) contains about 70 cells (Martini, 1909), and includes a putative pituitary hom
a prominent sensory vesicle (SV), which includes a statolith (St) but, unlike asc
afferent and efferent nerves (n1–n3) with sensory and motor functions emanate f
nerve cord (TNC), which lacks a central canal, bends ventrally between the stom
approximately 25 neurons and other cells of the caudal ganglion coordinate activi
paired trunk mechanoreceptors (Bone and Mackie, 1975). A hollow spinal cord (S
representations of panels F and I are shown in Figs. 5A,B. AB, anterior brain; C
esophagus; Ec, ectoderm; En, endoderm; Es, endostyle; Go, gonad; LL, lower lip;
R, rectum; S, stomach; SC, spinal cord; SV, sensory vesicle; St, statocyte, Sl, sta
ature [e.g., fertilization to hatching requires less than 3 h at
22-C, but about 12 h at 7-C (Fenaux, 1998)], we stage
Oikopleura according to morphological landmarks. The
h schematic insets depicting the CNS (yellow) and notochord (red). Anterior
to the right in the adult (G–J). Different focal planes of incipient-tailbud (A)
ing structures. A dorsal optical section (boxed in C) shows the 2- to 4-cell
and C (dashed green line). An early hatchling (D), mid-hatchling (E), late
ptical cross section (boxed in panel E) at the level of the sensory vesicle
arynx, and endostyle. Details of regions corresponding to the anterior brain
ue dashed lines) are boxed in the schematic (G). In adults, the anterior brain
olog (CF) (Bassham, 2002), which connects with the pharyngeal space, and
idians, no pigmented cells (Georges et al., 1988; Holmberg, 1984). Paired
rom the anterior brain (Bollner et al., 1991; Olsson et al., 1990). The trunk
ach lobes and meets the caudal ganglion (CG) in the proximal tail. The
ty of the caudal musculature, and receive inputs from the anterior brain and
C) extends from the caudal ganglion down the tail (Bone, 1998). Schematic
F, ciliary funnel; CG, caudal ganglion; CNS, central nervous system; E,
Mo, mouth;Mu, muscle; n1–3, paired nerves; No, notochord; Ph, pharynx;
tolith; TNC, trunk nerve cord; UL, upper lip; VO, ventral organ.

C. Canestro et al. / Developmental Biology 285 (2005) 298–315 301
lack of a detailed blastomere fate map precludes an exact
description of expression patterns at cleavage stages
(Nishino and Satoh, 2001). Stages prior to hatching include:
(i) incipient-tailbud stage (beginning about 3.5 h post-
fertilization (pf) at 13-C), characterized by an indentation at
the initial demarcation of trunk and tail (Fig. 1A) and by an
irregular, rod-shaped notochord. (ii) Mid-tailbud stage
(beginning about 4 h pf at 13-C), in which a deep
indentation divides trunk and tail (Figs. 1B,C), and a rod-
shaped row of 20 notochord cells is easily distinguishable
by DIC microscopy (Fig. 1C). A twisting of the tail with
respect to the trunk (Delsman, 1910) is already pronounced
at this stage. This flexure makes it impossible to position
embryos for a true orthogonal image capture, but the
notochord and large muscle cells provide helpful landmarks
to orient samples. Nuclear staining (i.e., DAPI, Hoechst)
was routinely included in expression analysis at tailbud
stages to confirm notochord cell positions.
Stages after hatching conform to Fenaux’s (1976),
including: (i) early hatchling (Fig. 1D) (Fenaux stage I),
in which the animals are cylindrical, without substantial
demarcation between trunk and tail, and, although the trunk
still lacks obvious organs, the tail begins short bursts of
movement; (ii) mid-hatchling (Fenaux II) (Fig. 1E), in
which the beginning of organogenesis is obvious, and
animals can swim; (iii) late hatchling (Fenaux IV) (Fig. 1F),
in which movements of the heart and the cilia of the
digestive system and spiracles become visible, the trunk-tail
boundary is well defined, and coordinated swimming
improves; (iv) tailshift, characterized by the shift of the tail
to an acute angle relative to the trunk, signaling the end of
embryonic development, and competence to secrete and
inflate a filter-feeding house (Figs. 1G–J).
The transparency of Oikopleura embryos and adults
allows non-invasive study of internal morphology at the
level of individual cells. For some images, we merged DIC
optical sections using Adobe-Photoshop software to
integrate images of structures that spanned focal planes
(Figs. 1D–J).
Cloning and whole-mount in situ hybridization
Genomic DNA from about 50 Oikopleura individuals
from Bamfield Marine Station was used to construct an
arrayed fosmid library (Epicentre, CCFOS110). Complete
sequencing of targeted fosmid clones was performed by the
DOE Joint Genome Institute (Walnut Creek, CA). mRNA
from about 1500 embryos was used to synthesize cDNA as
described (Bassham and Postlethwait, 2000). Genes of
interest were amplified by PCR with degenerate primers
(see Table S1 in supplementary materials) using cDNA or
genomic DNA as template. Gene-specific primers were
designed for RACE PCR, and to screen the genomic library
using a pooling strategy. Coding sequence and gene
structures were inferred by sequence comparisons between
RACE products and fosmid genomic clones. We designate
O. dioica genes by a name (usually based on its human
ortholog) in italics and small-case letters (i.e., pax6), and
proteins by a name with the first letter in upper-case (i.e.,
Pax6). When multiple paralogs are found, we add Latin
letters in alphabetical order (i.e., otxa, otxb, otxc) with no
additional punctuation. Whole-mount in situ hybridization
was performed as described (Bassham and Postlethwait,
2000). Riboprobe information is provided in Table S2.
Results
Isolation of Oikopleura CNS markers
To study the development and regionalization of the
larvacean CNS, we characterized CNS markers that are
highly conserved across bilaterians and play a central
role in AP organization of the tripartite vertebrate brain
(Hirth et al., 2003; Reichert and Simeone, 2001). We
isolated eight O. dioica genes homologous to the Otx,
Pax6, Pax2/5/8, Engrailed, and Hox1 vertebrate gene
families. Sequence comparisons and phylogenetic analyses
unequivocally assigned each isolated Oikopleura gene
to its gene family. We isolated single copies for Oiko-
pleura pax6 (AY870650), engrailed (AY870647), and
hox1 (AY871214) genes, three duplicated copies of otx
(AY886542, AY897556, AY897557) and two copies of
pax2/5/8 (DQ020279, AY870648, AY870649).
During the preparation of this manuscript, a list of O.
dioica homeobox genes inferred from genome sequences
was reported (Edvardsen et al., 2005), and in the present
work, we have adopted the same names for the three
Oikopleura otx duplicate genes we independently isolated:
otxa, otxb, and otxc. Analysis of the predicted proteins for
the three Oikopleura genes revealed the presence of a
homeodomain, a single C-terminal hexapeptide motif (in
contrast with two hexapeptide motifs in vertebrate OTX
proteins), and a moderately conserved WSP motif (Fig.
S1A). The small amount of evolutionary information
provided by the conserved domains and variation in the
rest of the molecule hampered the construction of confident
protein alignments and phylogenetic inferences. Among the
three Oikopleura Otx duplicates, Otxa showed the highest
overall similarity to ascidian Otx. Analysis of exon–intron
organization of Oikopleura otx genes revealed multiple
introns in addition to the two introns conserved in all known
otx genes (Fig. S1A). Remarkably, all ascidian and
Oikopleura otx genes are characterized by the presence of
additional exons (yellow in Fig. S1A) between the
conserved exon containing the transcription origin and the
exon containing the N-terminal part of the homeobox. The
still detectable sequence similarities among these additional
exons in urochordates suggest that these exons were
originally gained before the separation of the ascidian and
larvacean lineages; thus, they are likely a synapomorphy of
urochordate otx genes. Analysis of fosmid sequences from

C. Canestro et al. / Developmental Biology 285 (2005) 298–315302
our Oikopleura genomic library (AY873983–AY873986)
shows that Oikopleura otxb and otxc are adjacent,
transcribed in opposite directions, and separated by a 17
kb stretch that contains no intervening putative genes.
This result suggests that Oikopleura otxb and otxc
probably arose by tandem gene duplication during the
evolution of larvaceans after they separated from the
ascidian lineage. Evidence from protein sequence similar-
ities, gene structures, and phylogenetic analysis (data not
shown and Edvardsen et al., 2005), shows that the
duplication event that produced the third Oikopleura otx
gene also probably occurred in the larvacean lineage after
it separated from the ascidian lineage, although an otx
gene lost during the evolution of the ascidian lineage
cannot be ruled out.
We isolated from Oikopleura two pax2/5/8 genes, only
one of which (pax2/5/8b) was reported by Edvardsen et al.
(2005). Sequence similarities and phylogenetic analysis
showed that one Oikopleura pax2/5/8 gene (AY870648,
DQ020279) is orthologous to ascidian pax2/5/8a and the
other (AY870649) is orthologous to ascidian pax2/5/8b
(Fig. S1B). These results show that the duplication event
that led to these two urochordate pax2/5/8 clades occurred
in the urochordate lineage after it diverged from the
cephalochordate + vertebrate lineage, but before the split
of the ascidian and larvacean lineages; therefore, the pax2/5/
8a+pax2/5/8b duplicates appear to be a urochordate
synapomorphy.
Development and AP regionalization of the Oikopleura
CNS
Whole-mount in situ hybridization experiments revealed
that the eight Oikopleura genes we isolated are transcribed
in the embryonic CNS, endoderm, and epidermis. Here, we
focus on expression patterns necessary to understand the
mechanisms of CNS development (Figs. 2–4). The complex
patterns of other expression domains will be described in
detail elsewhere.
Incipient-tailbud stage
Analysis of the expression of Oikopleura CNS markers
revealed that AP regionalization of the CNS begins early,
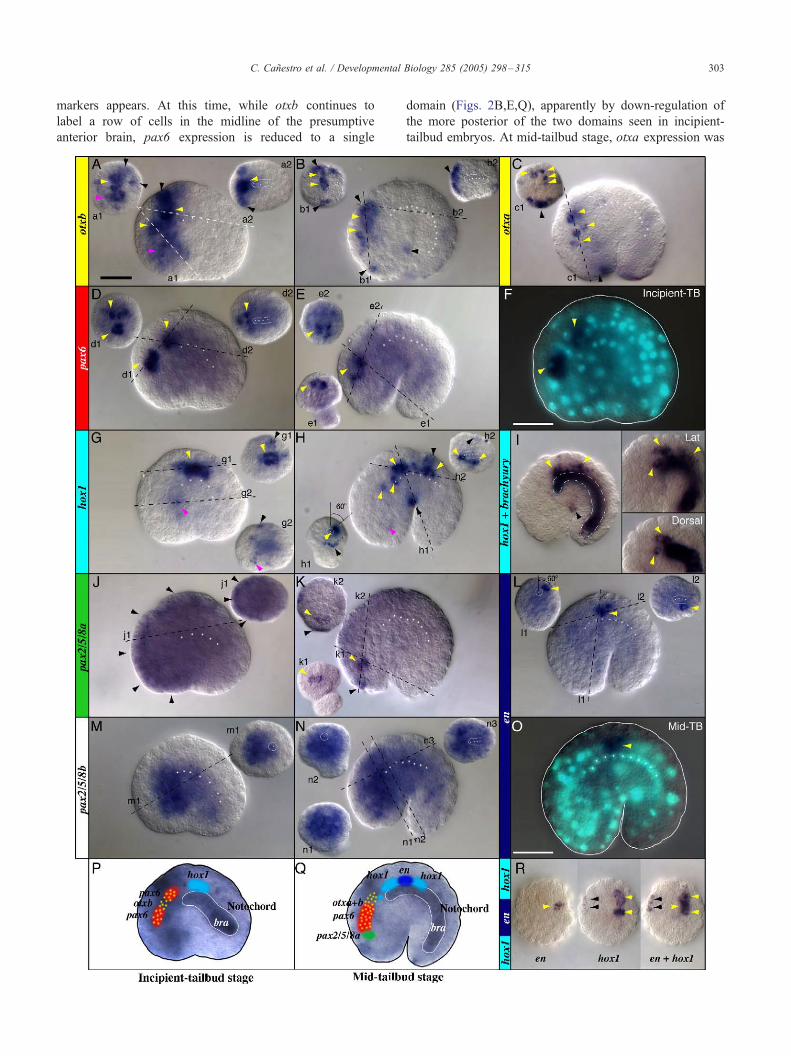
Fig. 2. Expression of CNS genes before hatching. Whole-mount in situ hybridizat
embryos (B,C,E,H,I,K,L,N,O,Q,R). otxb (yellow, A,B), otxa (yellow, C), pax6 (r
J,K), engrailed (dark blue, L,O), pax2/5/8b (M,N), and hox1 + engrailed (R). The
images (labeled with numbered lower case letters) are optical cross sections at the
line. In each panel, left insets are oriented with dorsal towards the top, and in right
expression shows the relative position between the Oikopleura hindbrain and the n
position of the anterior hox1 expression domain (lateral and dorsal views, squared
revealed by posterior CNS gene expression (i.e., H.h1,L.l1). Double in situ detectio
hox1 nested expression pattern (dorsal views oriented anterior towards the bottom
expression in the presumptive CNS (yellow), epidermis (black), and endoderm (pi
indicated by white dots. The cell number and size, and notochord position relati
exemplified in panels F and O, which are the same individuals shown in panels D
projected onto incipient-tailbud (P) and mid-tailbud (Q) embryos in which Brach
2000). Scale bar = 20 Am. TB, tailbud stage.
probably during gastrulation. Expression of otxb and hox1
is already detectable during cleavage stages (data not
shown), and probably provides the initial AP information
upon which subsequent AP regionalization is based.
At the incipient-tailbud stage, an anterior domain
expresses otxb and a posterior domain in the presumptive
CNS expresses hox1 (Figs. 2A,G,P). In addition to
epidermal expression, a broad otxb expression domain
may span the entire presumptive anterior brain (Fig. 2A).
At this stage, we did not detect expression of otxa or
otxc.
At the incipient-tailbud stage, the presumptive anterior
brain marked by otxb is subdivided by pax6 expression,
which appeared in two separate domains (Figs. 2D,F,P).
At least two bilateral pairs of cells located at the midline
constitute the most rostral pax6 domain (Fig. 2D.d1).
The caudal pax6 domain, anterior to the tip of the
notochord, also consists of at least two bilateral pairs of
cells (Fig. 2D.d1,d2).
While otxb and pax6 mark the anterior CNS, bilateral
rows of cells express hox1 dorsolaterally and posteriorly
to the anterior tip of the notochord, at the level of the
prospective caudal ganglion and anterior spinal cord
(Figs. 2G,P).
Because cells expressing otxb, pax6, and hox1 in the
presumptive CNS were already internal and close to the
midline at incipient-tailbud stage, we conclude that by this
stage, neurulation is at or nearing completion.
Contrary to expectations, neither of the two Oikopleura
pax2/5/8 genes is specifically expressed in the CNS. Pax2/
5/8a is expressed mainly in the trunk ectoderm (Fig. 2J).
Complementary to pax2/5/8a expression, pax2/5/8b was
detected in most of the internal portion of the trunk,
probably including some presumptive CNS cells (Fig. 2M).
These diffuse pax2/5/8 expression patterns are likely not
artifactual since different non-overlapping probes render
identical patterns, and the same probes reveal distinctive
tissue-specificity at later stages (Fig. 2K and data not
shown).
Mid-tailbud stage
At mid-tailbud stage, early broad gene expression
domains become refined and the expression of new CNS
ion in Oikopleura dioica incipient-tailbud (A,D,F,G,J,M,P) and mid-tailbud
ed, D–F), hox1 (light blue, G,H), hox1 + brachyury (I), pax2/5/8a (green,
central large image in each panel is a left lateral view, and the inset corner
levels of the dashed lines, viewed from the aspect of the label relative to the
insets, anterior is towards the left. Double detection of hox1 and brachyury
otochord (I). Inspection of different specimens reveals variability of the cell
insets in panel I). The degree of twisting of the trunk relative to the tail is
n of the expression of hox1 and engrailed (R) demonstrates the hox1 –en –
; comparable to panels H.h2 and L.l2 optical sections). Arrowheads label
nk). The position of notochord cells, sometimes in a different focal plane, is
ve to expression domains were routinely visualized by nuclear staining, as
and L, respectively. Schematized summaries of gene expression patterns are
yury expression (Bra) labels notochord (white) (Bassham and Postlethwait,
C. Canestro et al. / Developmental Biology 285 (2005) 298–315 303
markers appears. At this time, while otxb continues to
label a row of cells in the midline of the presumptive
anterior brain, pax6 expression is reduced to a single
domain (Figs. 2B,E,Q), apparently by down-regulation of
the more posterior of the two domains seen in incipient-
tailbud embryos. At mid-tailbud stage, otxa expression was

C. Canestro et al. / Developmental Biology 285 (2005) 298–315304
detected for the first time in the CNS, labeling asymmetri-
cally one cell in the right side and at least four cells in the
left side of the presumptive anterior brain (Fig. 2C). At
this stage, otxc expression was not yet detected within the
CNS.
The posterior CNS becomes subdivided in mid-tailbud
embryos, with the appearance of a hox1 –engrailed –hox1
nested pattern. Analysis of this nested pattern by single and
double in situ hybridization experiments, together with
notochord cell positioning by detection of brachyury
expression (Bassham and Postlethwait, 2000) and nuclear
staining (Figs. 2H,I,L,O,Q,R), revealed an anterior hox1
domain spanning approximately from the AP level of the
anterior tip of the notochord to the third notochord cell, a
central engrailed domain at the level of the fourth notochord
cell, and a posterior hox1 domain extending from the fifth to
seventh notochord cells (Figs. 2H,I). Analysis of several
dozen mid-tailbud embryos revealed slight variations in the
positions of cells forming the anterior hox1 domain with
respect to the tip of the notochord (Figs. 2H,I). At this stage,
frontal views of embryos probed for CNS markers revealed
an approximately 60- counterclockwise twist of the dorso-
ventral axis in the tail relative to the trunk, which causes the
posterior CNS to come to lie left of and somewhat dorsal to
the notochord (Fig. 2e1,h1,k1,l1). Double detection of
hox1 and brachyury expression showed that some of the
cells of the anterior hox1 domain come to lie in the midline
of the trunk, dorsal and sometimes anterior to the
notochord. We did not detect hox1 expression within the
notochord (Figs. 2H,I).
Although there is a gap between the posterior border of
the otxa + otxb expression domain and the hox1 expression
domain (see Figs. 2P,Q), we detected no pax2/5/8
expression specific for the CNS in that gap. This contrasts
with ascidians, in which pax2/5/8a expression fills the
otx–hox1 gap (Wada et al., 1998). In contrast, two other
tissues express Oikopleura pax2/5/8a in the trunk: a small
patch of epidermal cells in the prospective mouth region,
and a group of internal cells at the boundary between the
anterior brain and pharynx, rostral to the pax6 expression
domain (Fig. 2K, black, yellow arrowheads, respectively).
At this mid-tailbud stage, we could not discern the exact
anterior-most border of the anterior brain by morphology,
and fate mapping is needed to confirm whether individual
cells and their descendents in that region become part of
the anterior brain or part of the pharynx roof. Broad
expression of pax2/5/8b continues, restricted mainly to the
trunk (Fig. 2N).
Early and mid-hatchling stages
During early hatchling stages, larvacean embryos
rapidly elongate and organs related to movement, such
as the notochord, muscle cells, and CNS, are more mature
than the digestive system (Fig. 1D). Although the trans-
parency of living Oikopleura embryos allowed us to
distinguish much of the developing CNS from surrounding
tissues as early as incipient-tailbud stage (Figs. 1A–C), the
four main AP regions of the CNS—anterior brain (AB),
compact trunk nerve cord (TNC), caudal ganglion (CG),
and hollow tail nerve cord or spinal cord (SC)—do not
become obviously demarcated from each other until early
hatchling stages (Fig. 1D). This suggests that the primary
molecular definition of these regions is probably complete
before hatching, and therefore, the later expression of otx,
pax6, engrailed, and hox1 likely reflects secondary
functions related to cell fate maintenance or to specifica-
tion of neuron subpopulations, especially in the caudal
ganglion where motor neurons probably start to control tail
movements. Accordingly, while expression patterns of
these genes are largely continuous from mid-tailbud stage,
expression becomes restricted to specific cell subpopula-
tions (Fig. 3).
In early and mid-hatchlings, pax6 is expressed in the
anterior brain, outside the future sensory vesicle (Figs.
3A,B). A thin extension of expression reaches forward from
pax6-positive cells (rostral arrowhead in Fig. 3A), suggest-
ing the formation of neuronal processes. The internal pax2/
5/8a expression domain observed anterior to pax6 expres-
sion at mid-tailbud stage disappears by mid-hatchling stages
(data not shown).
Otxb expression narrows in hatchlings, from initially
spanning the entire presumptive anterior brain (Fig. 2A) to
comprising only a few cells (Fig. 3D). Interestingly, as otxb
expression declines in early hatchlings (Fig. 3D), the otxa
expression signal becomes intense (Fig. 3G), and for the
first time, otxc transcripts begin to accumulate in CNS cells
(Fig. 3J). This process, in which otxa + otxc expression
seems to replace otxb expression over time, is finished by
late hatchling stages when otxb transcription is almost
undetectable in the anterior brain (Fig. 3F).
During hatchling stages, the expression of the three otx
duplicates is restricted to the central portion of the
anterior brain, although there are differences in their
expression patterns; while some cells apparently co-
express multiple otx duplicates, other cells express only
one, and in some cases the expression patterns are asym-
metric (Figs. 3D,E,G,H,J,K).
In early hatchling stages, in addition to strong epidermal
hox1 expression at the trunk-tail transition (Fig. 4A, black
arrowheads), a row of hox1-expressing cells extends left of
the midline from anterior positions near the notochord tip
(within the trunk) caudally to at least the level of the fourth
notochord cell (Figs. 4A,B), probably spanning the
prospective posterior TNC and anterior caudal ganglion
(Fig. 1D). Additional hox1 signal with irregular shape and
thin diameter appears at the level of the fifth and sixth
notochord cells (Figs. 4A,B), and could mark caudal
cellular extensions from anterior hox1-expressing cells.
These observations broadly confirm and extend those for
hox1 in Seo et al. (2004). In early hatchlings, in addition to
the expression of hox1 in the anterior segment of the
caudal ganglion, we observed expression of engrailed (Fig.
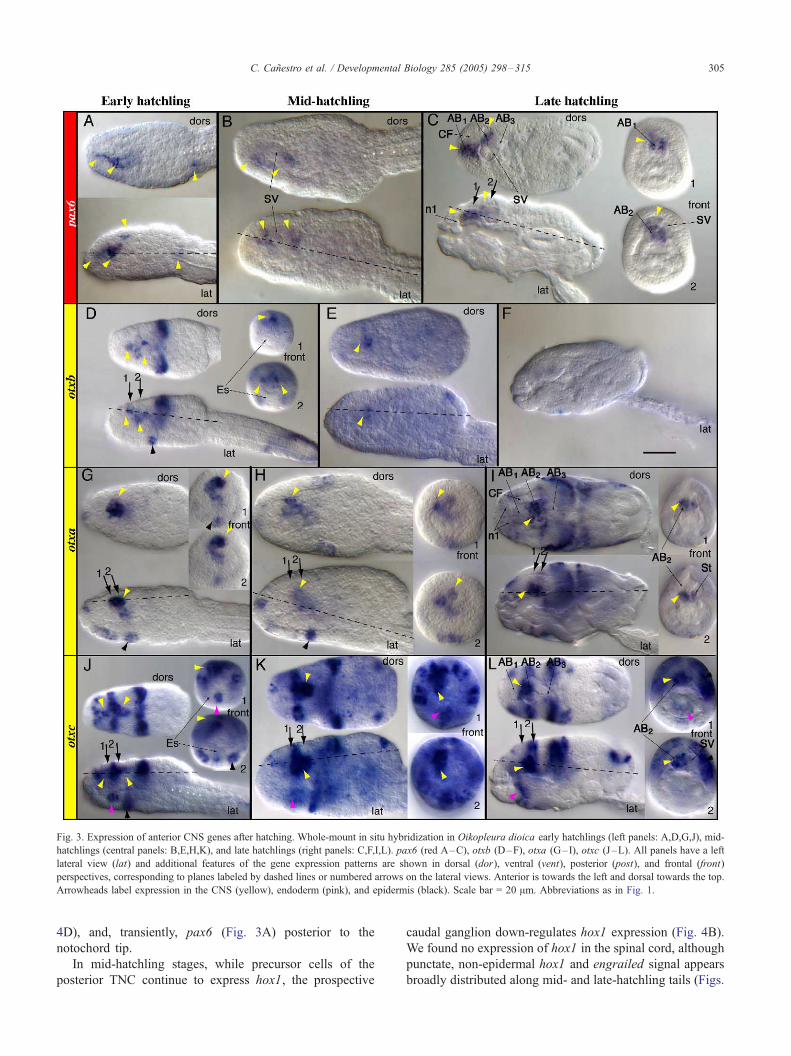
Fig. 3. Expression of anterior CNS genes after hatching. Whole-mount in situ hybridization in Oikopleura dioica early hatchlings (left panels: A,D,G,J), mid-
hatchlings (central panels: B,E,H,K), and late hatchlings (right panels: C,F,I,L). pax6 (red A–C), otxb (D–F), otxa (G– I), otxc (J–L). All panels have a left
lateral view (lat) and additional features of the gene expression patterns are shown in dorsal (dor), ventral (vent), posterior (post), and frontal (front)
perspectives, corresponding to planes labeled by dashed lines or numbered arrows on the lateral views. Anterior is towards the left and dorsal towards the top.
Arrowheads label expression in the CNS (yellow), endoderm (pink), and epidermis (black). Scale bar = 20 Am. Abbreviations as in Fig. 1.
C. Canestro et al. / Developmental Biology 285 (2005) 298–315 305
4D), and, transiently, pax6 (Fig. 3A) posterior to the
notochord tip.
In mid-hatchling stages, while precursor cells of the
posterior TNC continue to express hox1, the prospective
caudal ganglion down-regulates hox1 expression (Fig. 4B).
We found no expression of hox1 in the spinal cord, although
punctate, non-epidermal hox1 and engrailed signal appears
broadly distributed along mid- and late-hatchling tails (Figs.
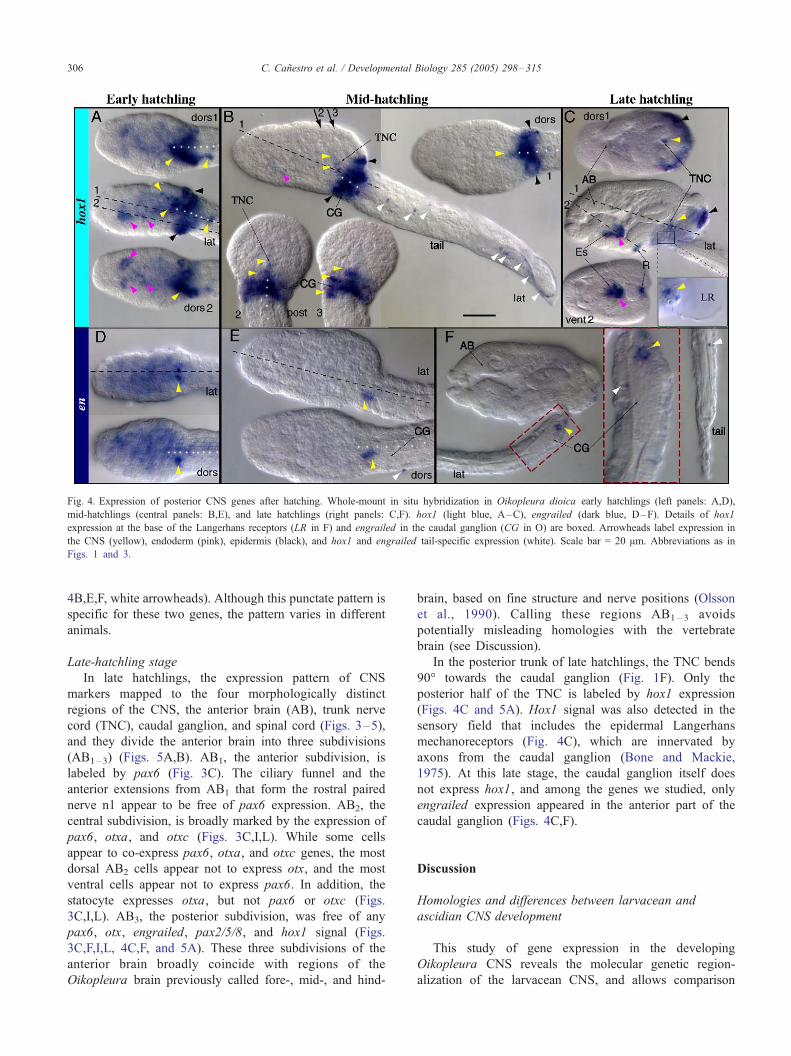
Fig. 4. Expression of posterior CNS genes after hatching. Whole-mount in situ hybridization in Oikopleura dioica early hatchlings (left panels: A,D),
mid-hatchlings (central panels: B,E), and late hatchlings (right panels: C,F). hox1 (light blue, A–C), engrailed (dark blue, D–F). Details of hox1
expression at the base of the Langerhans receptors (LR in F) and engrailed in the caudal ganglion (CG in O) are boxed. Arrowheads label expression in
the CNS (yellow), endoderm (pink), epidermis (black), and hox1 and engrailed tail-specific expression (white). Scale bar = 20 Am. Abbreviations as in
Figs. 1 and 3.
C. Canestro et al. / Developmental Biology 285 (2005) 298–315306
4B,E,F, white arrowheads). Although this punctate pattern is
specific for these two genes, the pattern varies in different
animals.
Late-hatchling stage
In late hatchlings, the expression pattern of CNS
markers mapped to the four morphologically distinct
regions of the CNS, the anterior brain (AB), trunk nerve
cord (TNC), caudal ganglion, and spinal cord (Figs. 3–5),
and they divide the anterior brain into three subdivisions
(AB1–3) (Figs. 5A,B). AB1, the anterior subdivision, is
labeled by pax6 (Fig. 3C). The ciliary funnel and the
anterior extensions from AB1 that form the rostral paired
nerve n1 appear to be free of pax6 expression. AB2, the
central subdivision, is broadly marked by the expression of
pax6, otxa, and otxc (Figs. 3C,I,L). While some cells
appear to co-express pax6, otxa, and otxc genes, the most
dorsal AB2 cells appear not to express otx, and the most
ventral cells appear not to express pax6. In addition, the
statocyte expresses otxa, but not pax6 or otxc (Figs.
3C,I,L). AB3, the posterior subdivision, was free of any
pax6, otx, engrailed, pax2/5/8, and hox1 signal (Figs.
3C,F,I,L, 4C,F, and 5A). These three subdivisions of the
anterior brain broadly coincide with regions of the
Oikopleura brain previously called fore-, mid-, and hind-
brain, based on fine structure and nerve positions (Olsson
et al., 1990). Calling these regions AB1– 3 avoids
potentially misleading homologies with the vertebrate
brain (see Discussion).
In the posterior trunk of late hatchlings, the TNC bends
90- towards the caudal ganglion (Fig. 1F). Only the
posterior half of the TNC is labeled by hox1 expression
(Figs. 4C and 5A). Hox1 signal was also detected in the
sensory field that includes the epidermal Langerhans
mechanoreceptors (Fig. 4C), which are innervated by
axons from the caudal ganglion (Bone and Mackie,
1975). At this late stage, the caudal ganglion itself does
not express hox1, and among the genes we studied, only
engrailed expression appeared in the anterior part of the
caudal ganglion (Figs. 4C,F).
Discussion
Homologies and differences between larvacean and
ascidian CNS development
This study of gene expression in the developing
Oikopleura CNS reveals the molecular genetic region-
alization of the larvacean CNS, and allows comparison

Fig. 5. (A) Schematized summary of gene expression patterns projected onto the CNS of a late hatchling (lateral view). pax6 (red), otxa + otxc (yellow), hox1
(light blue), and engrailed (dark blue). Notochord in pink. (B) Schematic representation of the anterior brain AP subdivisions (AB1– 3) in relation to neural
circuits (colored cells) in the adult brain described by Olsson et al. (1990). Afferent and efferent nerve pathways (paired nerves, n1–n3, and dorsal nerves, dn)
connecting the anterior brain (AB) with target cells (white: epidermal cells, EC; ciliary rings of spiracles, CR; ventral organ, VO; and upper and lower lips,
UL,LL) are shown. Abbreviations are as in Fig. 1. (C) Schematic comparison of CNS anteroposterior regionalization to gene expression patterns among
chordate Subphyla. Tree (bottom) indicates phylogenetic relationships. For larvaceans and ascidians, gene expression patterns are detailed during different
developmental stages (grey boxes). Gene expression domains labeled with an asterisk appear relatively late in development. Grey spheres represent proximal
notochord cells. Cephalochordate CNS regions are depicted relative to somite positions (s1–8). Vertebrate and urochordate data support homologies for
forebrain (black) and hindbrain (blue), but not for midbrain or MHB.
C. Canestro et al. / Developmental Biology 285 (2005) 298–315 307
with ascidians. The ascidian CNS has been divided into
four regions: the ‘‘brain’’ (which includes the sensory
vesicle housing the statocyte and ocellus), neck,
visceral ganglion, and spinal cord (Satoh, 2003). The
following section highlights evidence that defines
regional homologies in the CNS of two urochordate
classes, and discusses the differences observed along
the AP axis of the CNS, starting rostrally and moving
caudally.
The anterior CNS
The Oikopleura anterior brain and ascidian ‘‘brain’’ are
homologous. Expression of Oikopleura otxb and ascidian
otx demarcates the anterior CNS in cleavage stages
(Hinman and Degnan, 2000; Hudson and Lemaire, 2001;
Wada et al., 1996; Wada et al., 2004; and this study). Both
urochordate classes have two pax6 expression domains, the
posterior of which overlaps the expression of otx near the

C. Canestro et al. / Developmental Biology 285 (2005) 298–315308
anterior tip of the notochord (Figs. 2P and 5C.c,f) (Glardon
et al., 1997; Mazet et al., 2003). By the mid-tailbud stage
in Oikopleura, although the otxb posterior boundary does
not change relative to the notochord, pax6 is expressed
only in the former anterior domain, suggesting down-
regulation of pax6 expression in the posterior domain
(Figs. 2Q and 5C.d). Similarly, pax6 expression is absent
from the posterior part of the ascidian brain at mid-tailbud
stage (Glardon et al., 1997; Mazet et al., 2003). This
congruence of otx and pax6 expression patterns in the early
tailbud stages of Oikopleura and ascidians supports
homology of the larvacean anterior brain and the ascidian
‘‘brain’’.
Expression of otx and pax6 reveals differences between the
Oikopleura anterior brain and the ascidian ‘‘brain’’. Al-
though early expression of otx and pax6 is similar in the two
urochordate classes, at later stages, there are three major
differences. First, pax6, which is typically expressed in
photoreceptors (Callaerts et al., 1997), is expressed in the
photosensitive ocellus in the ascidian sensory vesicle
(Glardon et al., 1997), but not in that of Oikopleura, which
lacks pigmented photoreceptor cells. The second major
difference is that while pax6 is no longer expressed in the
ascidian ‘‘brain’’ by late larval stages (Mazet et al., 2003), in
Oikopleura, it continues to be expressed in the anterior brain
until at least the late hatchling stage (Figs. 3C and 5A,C.e).
This expression difference perhaps reflects the continuity of
embryonic and adult Oikopleura CNS patterning (see
discussion below), while the ascidian CNS is drastically
restructured during metamorphosis.
The third major difference is that the three Oikopleura
otx genes, in contrast to the single ascidian otx gene, show
‘‘phased’’ expression dynamics, differing in their temporal
and spatial expression patterns during CNS development.
These results suggest the hypothesis that the three otx
genes in Oikopleura have different developmental func-
tions. The broad and uniform expression of the single otx
gene described in the ascidian brain (Wada et al., 1996)
would mask potential multiple functions. Like Oikopleura,
tetrapods have three OTX paralogs (OTX1, OTX2, and
OTX5/CRX) due to independent duplications within the
vertebrate lineage. The vertebrate paralogs, like their
Oikopleura homologs, have also assumed separate early
and late developmental roles. First, the earliest expression
of Oikopleura otxb in the anterior neuroectoderm and
endoderm is comparable to the expression of OTX2 during
vertebrate gastrulation (Reichert and Simeone, 2001). And
second, the later appearance of Oikopleura otxa and otxb
expression in a reduced number of cells in the anterior
brain can be compared to the action of OTX1 and OTX2 in
specifying identity and fate of specific cell populations in
the vertebrate brain (Puelles et al., 2003; Puelles et al.,
2004). This parallelism between vertebrates and Oiko-
pleura provides an example of trans-homologs [genes
duplicated independently in different lineages from a
common ancestral pro-ortholog (Sharman, 1999)] converg-
ing on similar evolutionary outcomes. Functional analysis
of otx regulation in Oikopleura duplicates will test whether
this evolutionary convergence is due to parallel subfunc-
tionalization events in conserved regulatory modules (Force
et al., 1999) present in the last common ancestor of
vertebrates and urochordates, or whether phased otx
regulation has been gained independently in the two
chordate lineages.
Pax2/5/8 genes reveal differences between Oikopleura and
ascidian anterior CNS regions. In ascidians, the expres-
sion of pax2/5/8a in the gap between otx and hox1 domains
suggested homology with the vertebrate MHB (Wada et al.,
1998). Despite the expression similarities between Oiko-
pleura and ascidian pax2/5/8a and pax2/5/8b orthologs in
several domains (Mazet et al., 2005; Wada et al., 1998),
neither Oikopleura pax2/5/8 gene is expressed in the gap
between otx and hox1 domains. Therefore, pax2/5/8
expression data argue against the presence of an MHB
homolog in Oikopleura.
In mid-tailbud stages, ascidians lack pax2/5/8a expres-
sion similar to the Oikopleura pax2/5/8a internal domain at
the brain–pharynx border anterior to the pax6 domain
(Mazet et al., 2003; Wada et al., 1998). In amphioxus, pax2/
5/8 expression in the anterior cerebral vesicle has been
compared to vertebrate Pax2 expression in development of
the optic stalk and optic nerve (Fig. 5C.a) (Kozmik et al.,
1999; Krauss et al., 1991; Macdonald et al., 1997).
Additionally, it was proposed that pax2.1, in cooperation
with Fgf signaling, influences axon guidance and early
rostral midline development in the zebrafish forebrain
(Shanmugalingam et al., 2000). Because AB1, the rostral
subdivision of the anterior brain of Oikopleura, projects
axons rostrally via the paired nerves n1 and n2 to the ventral
organ and sensory receptors in the lips (Figs. 1I and 5B),
Oikopleura pax2/5/8a expression is consistent with a role in
axon guidance like its vertebrate homolog Pax2. Study of
more markers and gain- and loss-of-function experiments
are necessary to test these hypotheses for the role of the
Oikopleura pax2/5/8a.
Links between developmental expression patterns and the
adult larvacean brain. In contrast to ascidians, the
continuous transition of the Oikopleura CNS from hatch-
ling to adult provides direct links between embryonic gene
expression domains and the fine structure and function of
the adult Oikopleura brain (Olsson et al., 1990). Figs. 5A,B
show the major anatomical features of the anterior CNS in
the juvenile and adult Oikopleura. The rostral subdivision
of the anterior brain (AB1) labeled by pax6 expression
receives afferent pathways from anterior sensory cells,
including the ventral organ (via n1), and ciliated receptor
cells in the lips and pharynx (via n2) (Olsson et al., 1990;
and Fig. 5B). The efferent pathways described by Olsson et
al. (1990) may originate in the central and posterior

C. Canestro et al. / Developmental Biology 285 (2005) 298–315 309
subdivisions AB2 and AB3. AB2, which is labeled by pax6,
otxa, and otxc, probably includes three neurons that project
axons via n3, a postulated motor nerve carrying fibers to
the spiracle’s ciliary rings and to ventral epidermal cells
that might have a role in house secretion (Fig. 5B). The
posterior subdivision AB3, which does not express any of
the genes in this study during late development, sends one
process via the left n3 to the ciliary ring and another into
the TNC, probably towards the caudal ganglion. Therefore,
the AB1–3 subdivisions revealed by molecular markers
during development seem to correspond to different func-
tional areas of the brain in charge of integrating afferent
and efferent pathways.
The posterior CNS
The posterior urochordate CNS features a prominent
ganglion near the anterior tip of the notochord called the
caudal ganglion in Oikopleura and the visceral ganglion
in ascidians (Fig. 1) (Meinertzhagen et al., 2000). Variation
in gene expression and morphology among ascidian species,
however, hinders a straightforward assignment of homology
between larvacean and ascidian posterior CNS regions.
Larvaceans and ascidians share a two-domain expression
profile for hox1 (Fig. 5C.d,g): an anterior domain near the
anterior tip of the notochord [compare dorsal views of
Oikopleura in Fig. 4A to views of the ascidian in Fig. 5 of
Katsuyama et al. (1995)] and a posterior, transient domain in
the presumptive caudal/visceral ganglion and spinal cord
(Figs. 2P–Q and 5A,C) (Ikuta et al., 2004; Katsuyama et al.,
1995; Nagatomo and Fujiwara, 2003). In Oikopleura, we
can trace the anterior hox1 domain throughout development
to the posterior TNC. In ascidians, although the hox1
expression pattern is the same in different species, various
authors have differed in their interpretation of the fate of the
anterior domain, assigning it either to the neck or to the
visceral ganglion (Ikuta et al., 2004; Katsuyama et al., 1995;
Nagatomo and Fujiwara, 2003; Wada et al., 1998). This
controversy is probably due to the lack of a morphologically
distinguished neck in H. roretzi rather than a true fate
difference.
These expression data also indicate shared morphoge-
netic behaviors of larvacean and ascidian axial tissues. The
anterior hox1 expression domain shifts forward with
respect to the notochord during Oikopleura development,
as it does in ascidians (Katsuyama et al., 1995). This
domain shift probably explains the variability of the cell
positions of the anterior hox1 domain observed in
Oikopleura mid-tailbud stages as different individuals are
fixed at slightly different stages (Figs. 2H–I,R). In Ciona
intestinalis, neck precursor cells apparently move from
behind the tip of the notochord in tailbud embryos to an
anterior location in hatchlings, suggesting that the neck is
part of the visceral ganglion (Cole and Meinertzhagen,
2004; Meinertzhagen et al., 2000). This shift applies also to
H. roretzi pax2/5/8a-positive cells (Wada et al., 1998).
Therefore, despite the morphological differences between
the CNS of the two urochordate classes, similar morpho-
genetic processes seem to be conserved underlying the
development of the neural tube at the level of the anterior
tip of the notochord.
Engrailed expression in the posterior CNS is also similar
among urochordates. In Oikopleura, engrailed expression
begins at mid-tailbud stage between the two initial hox1
domains creating a hox1–engrailed–hox1 nested pattern
(Fig. 2R). Throughout development, engrailed expression
continues posterior to the anterior notochord tip, labeling the
anterior part of the caudal ganglion (Figs. 2L,O,R and 4D–
F). Expression of engrailed differs between the two
congeneric ascidian species in which it has been described.
In Ciona savignyi, a single bilateral pair of cells co-
expresses engrailed and pax2/5/8a (Jiang and Smith, 2002).
In the congener C. intestinalis, pax2/5/8a is also expressed
in a single pair of cells, but engrailed flanks the pax2/5/8
expression domain in an engrailed-pax2/5/8a-engrailed
nested pattern (Fig. 5C.g), in which the posterior engrailed
domain labels the anterior part of the visceral ganglion (Imai
et al., 2002). Although no direct comparison of hox1 and
engrailed expression has been published for any ascidian,
comparing the positions of hox1 and engrailed expression
domains with the notochord (Ikuta et al., 2004; Imai et al.,
2002; Nagatomo and Fujiwara, 2003) makes it likely that
the hox1 –engrailed –hox1 nested pattern is shared by
Oikopleura and at least some ascidians (Fig. 5C.g). The
fact that pax6 is expressed transiently at early hatchling
stages in the presumptive caudal/visceral ganglion approx-
imately at the same level as engrailed (Glardon et al.,
1997; Mazet et al., 2003; and our Fig. 3A) bolsters the
correspondence of larvacean and ascidian posterior
engrailed domains.
The similar expression of engrailed, pax6, and hox1
during development of the caudal ganglion in Oikopleura
and the visceral ganglion in ascidians suggests that these
two structures are homologous. The present expression data
are also consistent with the homology of the ascidian neck
and the posterior Oikopleura TNC. In agreement with the
interpretation that the ascidian neck is part of the visceral
ganglion (Meinertzhagen et al., 2000), we conclude that the
posterior TNC and the caudal ganglion derive from the same
embryonic CNS region labeled by the single hox1 domain at
incipient-tailbud stage (Fig. 2G). Because Oikopleura
engrailed expression remains in a constant position poste-
rior to the tip of the notochord, while the hox1 domain shifts
anteriorly, there may also be elongation or migration of
specific cells, or stationary cells turning on and off
expression, rather than simply a general anterior shift of
this entire part of the posterior CNS.
Homologies and differences between urochordate and
vertebrate CNS development
Our comparative analysis of Oikopleura and ascidian
urochordates has revealed homologies and differences in

C. Canestro et al. / Developmental Biology 285 (2005) 298–315310
developmental genetic pathways, which now can be
compared to other chordate Subphyla.
Which part of the vertebrate CNS is homologous to the
urochordate spinal cord?
In contrast to ascidians and vertebrates, Oikopleura and
cephalochordates (Glardon et al., 1998) lack pax6 expres-
sion along the length of their spinal cords. Because the
posterior Drosophila CNS expresses ey, which is the fly
ortholog of pax6 (Quiring et al., 1994), the most parsimo-
nious explanation for species-specific pax6 expression
patterns is that the last common ancestor of extant chordates
had pax6 expression in the spinal cord and it was
independently lost in the Oikopleura and cephalochordate
lineages.
The similar expression pattern between ascidian hox5
(Gionti et al., 1998) and the only Oikopleura central Hox-
subclass gene (called hox4, although equally related to
hox4/5/6/7) (Seo et al., 2004) suggests that these genes
may function to define the anterior boundary of the
urochordate spinal cord. Therefore, despite the variation
in pax6 expression patterns among chordates, we conclude
that the spinal cord of urochordates and vertebrates is
homologous.
Which part of the vertebrate CNS is homologous to the
urochordate TNC/neck and caudal ganglion/visceral
ganglion?
Since the caudal ganglion of Oikopleura and the
ascidian visceral ganglion are probably homologous, as
are the posterior TNC in Oikopleura and the ascidian
neck, and since these structures derive from a posterior
CNS region expressing hox1, we conclude that these
structures are homologous to at least part of the
vertebrate hindbrain (Fig. 5C.b,c). The presence of motor
function in the vertebrate hindbrain (Lumsden and
Krumlauf, 1996), the existence of motor neurons in the
caudal ganglion that coordinate muscular tail movements
in larvaceans (Bone, 1998), and the presence of motor
neurons in the neck and visceral ganglion of ascidians
(Katsuyama et al., 2005; Meinertzhagen et al., 2000;
Okada et al., 2002) are consistent with this proposed
homology; we will therefore refer to the posterior-TNC/
neck plus caudal/visceral ganglion as the ‘‘urochordate
hindbrain’’.
In ascidians, hox3 expression suggests that the anterior
limit of the visceral ganglion corresponds to the anterior
limit of r4 of the vertebrate hindbrain (Locascio et al.,
1999). Unexpectedly, despite other gene expression
similarities between the larvacean caudal ganglion and
the ascidian visceral ganglion (Fig. 5C.d,g), there does not
appear to be a hox3 ortholog in the Oikopleura genome
(Seo et al., 2004). Analysis of additional hindbrain
markers such as Kreisler and Krox20 could help us
understand the consequences of the loss of Oikopleura
hox3.
Which part of the vertebrate CNS is homologous to the
urochordate ‘‘anterior brain’’?
In addition to the ‘‘urochordate hindbrain’’, we designate
the anterior part of the urochordate CNS as ‘‘anterior brain’’,
rather than just ‘‘brain’’, because in vertebrates the term
‘‘brain’’ includes the forebrain, midbrain, and hindbrain.
Data from ascidians have led to conflicting interpreta-
tions concerning homologies between the ascidian ‘‘brain’’
and the vertebrate brain, sometimes because of differences
in morphology and gene expression among ascidian species
(Lemaire et al., 2002; Locascio et al., 1999; Meinertzhagen
and Okamura, 2001; Meinertzhagen et al., 2004; Satoh,
2003; Takahashi and Holland, 2004; Wada and Satoh,
2001). For example, the posterior part of the ascidian
‘‘brain’’ has inconsistently been proposed to be homologous
to (i) the vertebrate metencephalon [based on the co-
expression of Ci-fgf9/16/20 and Ci-engrailed (Meinertzha-
gen et al., 2004)]; (ii) the vertebrate midbrain [based on the
co-expression of Ci-otx and Ci-engrailed (Imai et al.,
2002)]; or (iii) the vertebrate forebrain [based on the
presence of Ci-otx expression (Hudson and Lemaire,
2001) and the absence of Ci-dmbx expression (Takahashi
and Holland, 2004)]. To address these conflicting conclu-
sions, we first integrated larvacean and ascidian CNS gene
expression patterns (see above), and now we compare that
result to vertebrate expression patterns.
In vertebrates, forebrain and midbrain are labeled by Otx
expression (Fig. 5C.b). The expression of Pax6 in two
domains, one in the posterior forebrain and one in the
anterior hindbrain, has been used to define the midbrain,
which develops in the intervening gap (‘‘pax6-gap’’) and is
regulated by Pax2 and Engrailed expression (Fig. 5C.b)
(Matsunaga et al., 2000; Scholpp et al., 2003; Schwarz et
al., 1999).
As in vertebrates, the prospective anterior brain of
urochordates is labeled by otx expression (Fig. 5C.c,f),
suggesting at first glance that the urochordate anterior brain
is homologous to the vertebrate forebrain + midbrain (Fig.
5C.b). In urochordates, however, the fact that the otx
domain is subdivided along the AP axis by two expression
domains of pax6 at early tailbud stage (Fig. 5C.c,f), leads to
an alternative interpretation. In this alternative, the anterior
pax6 domain in urochordates labels the homolog of the
vertebrate forebrain, while the posterior pax6 domain labels
the homolog of the anterior hindbrain, and the pax6-gap
could be the urochordate homolog of the vertebrate
midbrain. This interpretation, however, conflicts with two
facts. First, the pax6-gap of the urochordate anterior brain
fails to express the vertebrate midbrain markers pax2/5/8
and engrailed (Fig. 5C.b,c,f). And second, the posterior
expression domain of pax6 in the urochordate anterior brain
overlaps the otx expression domain, while Otx expression is
excluded from the vertebrate hindbrain (Fig. 5C.b,c,f).
Therefore, these data lead to the conclusion that the
urochordate anterior brain is homologous to the vertebrate
forebrain (Fig. 5C.b,c).

C. Canestro et al. / Developmental Biology 285 (2005) 298–315 311
New perspectives on chordate CNS evolution from an
Oikopleura point of view
Gbx gene loss, an ancestral event in urochordate evolution
In vertebrates, the action of Gbx excludes the posterior
boundary of Otx expression from the anterior hindbrain
labeled by Pax6 (Fig. 5C.b) (reviewed in Rhinn and Brand,
2001). We know that the boundary between Otx and Gbx
expression domains is crucial to define the limit between the
midbrain and the hindbrain in vertebrates because vertebrate
Gbx mutants show caudal expansion of Otx expression back
to the level of rhombomere 4. The finding that Gbx also
antagonizes Otx in Drosophila suggests that this gene
interaction is ancient in Bilateria (Hirth et al., 2003; Reichert
and Simeone, 2001). Interestingly, the apparent absence
of any Gbx homolog from the genome projects of C.
intestinalis (Wada et al., 2003) and C. savignyi (http://
www.broad.mit.edu/annotation/ciona/index.html), and the
failure to isolate a Gbx homolog from Oikopleura [despite
our unpublished efforts and the genome sequencing traces
(Edvardsen et al., 2005)], strongly suggest that the loss of
gbx is a urochordate synapomorphy. This gene loss might
have affected the evolution and regionalization of the
urochordate CNS, perhaps by permitting a caudal expansion
of the otx domain as observed in vertebrate Gbx mutants. A
caudal expansion of otx expression in an ancestral urochor-
date would be consistent with our comparative analysis of
Oikopleura and ascidian data, which shows that the
posterior boundary of otx expression overlaps with the
posterior pax6 expression domain in the anterior brain (see
discussion above and Fig. 5C.b,c,f). The study of additional
urochordate orthologs of genes upstream and downstream of
Gbx function in the vertebrate anterior hindbrain (e.g.,
Wnt1, Fgf8) will help assess the impact of the loss of gbx
from the urochordate genome.
Do urochordates have a homolog of the vertebrate
midbrain?
In the Oikopleura CNS, absence of engrailed and pax2/
5/8 expression anterior to the hox1 expression domain that
labels the hindbrain argues against the presence of midbrain
in the larvacean lineage. Thus, we conclude that the absence
of a midbrain in ascidians, recently proposed by T.
Takahashi and P.W.H. Holland (2004) based on the absence
of dmbx expression in the anterior brain, is not just a
peculiarity of the ascidian Class, resulting perhaps from its
drastic CNS metamorphosis, but is a synapomorphy of the
urochordate Subphylum.
Absence of evidence supporting the existence of a
midbrain in amphioxus (Holland et al., 1997) and ascidians
(Takahashi and Holland, 2004) led to the most parsimonious
conclusion that the absence of a distinct midbrain was the
ancestral condition for chordates, and that the midbrain is a
developmental innovation in the vertebrate lineage (Takaha-
shi and Holland, 2004). Although this conclusion continues
to be the most parsimonious explanation, our data on
Oikopleura CNS are also consistent with a different
hypothesis. The posterior overlap of otx and pax6 ex-
pression, the absence of gbx in urochordates, and the
variability or absence of the expression of orthologs of
vertebrate midbrain markers (such as engrailed and pax2/
5/8) in the urochordate anterior brain, suggest that the
midbrain was present in the last common ancestor of
extant chordates, but was modified or lost independently in
the urochordate and in the cephalochordate lineages. Study
of the developmental mechanisms underlying the posterior
subdivision of the anterior brain of Oikopleura (AB3)
could provide new data to better understand the evolution
of the urochordate brain, because AB3 is located just
anterior to the urochordate hindbrain, and its homology is
as yet uncertain due to the absence of expression of all
CNS markers so far analyzed.
Do urochordates have a midbrain–hindbrain organizer
(MHB)?
In vertebrates, the MHB forms at the border of Otx and
Gbx expression domains, and requires expression of Pax2,
Engrailed, Wnt11, and Fgf8 genes (reviewed in Rhinn and
Brand, 2001). The absence of a Gbx ortholog in urochor-
dates makes it difficult to recognize a putative MHB
organizer in this Subphylum. In Oikopleura, the absence
of engrailed and pax2/5/8 expression immediately posterior
to the otx expression domain argues against the presence of
an MHB organizer homolog in the larvacean CNS.
In ascidians, it was argued that because pax2/5/8
expression in the neck coincides with the gap between otx
and hox1 expression domains (Wada et al., 1998), and
because engrailed and fgf8/17/18 expression appears in the
anterior visceral ganglion (Imai et al., 2002), the neck region
of the ascidian CNS is homologous to the vertebrate
MHB (Imai et al., 2002; Jiang and Smith, 2002; Meinertz-
hagen et al., 2004; Satoh, 2003; Wada et al., 1998;
Wada and Satoh, 2001). Close inspection of Ci-pax2/5/8,
Ci-engrailed, and Ci-fgf8/17/18, however, reveals that they
are not co-expressed in C. intestinalis (Imai et al., 2002). If
these genes are not co-expressed, then they cannot have the
same genetic interactions their orthologs have in the
development of the vertebrate MHB. Even if Ci-fgf8/17/
18 signaling might provide organizer function in the
anterior part of the visceral ganglion in ascidians, that
function might not necessarily be homologous to the MHB
organizer specifically, because Fgf8 is also active in the r4
organizer in the hindbrain of the vertebrate Danio rerio
(Maves et al., 2002; Walshe et al., 2002). The interpretation
that Fgf expression in the ascidian CNS could be
homologous to the r4 organizer is compatible with our
definition of the ‘‘urochordate hindbrain’’, and with the
correspondence of the anterior part of the visceral ganglion
in ascidians with rhombomere 4 in vertebrates based on
expression of Ci-hox3 (Locascio et al., 1999).
Finally, the obvious differences in the expression patterns
of engrailed and pax2/5/8a between Oikopleura and

C. Canestro et al. / Developmental Biology 285 (2005) 298–315312
ascidians, and indeed among different ascidian species [i.e.,
even between the congeners C. savignyi and C. intestinalis
(Imai et al., 2002; Jiang and Smith, 2002)], are not expected
if the presence of an MHB organizer is fundamental for the
regionalization of at least the ascidian CNS. Therefore, in
the light of our data from Oikopleura and published results
in ascidians, and in the absence of any functional data about
the roles of engrailed, pax2/5/8, and fgf8/17/18 in ascidians,
we conclude that there is no convincing evidence for an
MHB homolog in urochordates. Characterization and func-
tional analysis of Fgf family members in Oikopleura and
ascidians would help to test whether organizer activity exists
in urochordates.
Alternative hypothesis for the function of ‘‘MHB genes’’ in
urochordates
Urochordate tailbud stage embryos may correspond to
much later developmental stages than the vertebrate gastrula
and neurula stages in which the MHB forms. In vertebrates,
many of the genes that are involved in the development of
the MHB are also expressed later in the hindbrain, where
they perform different functions than they do in the MHB
(Lumsden and Krumlauf, 1996). In the vertebrate hindbrain,
Pax2, Pax5, Pax8, En1, En2, Pax6, Hox1, Dmbx1, and Lim
transcription factors, in conjunction with graded FGF
signals, specify motor neurons and interneurons, and are
important for axon guidance (Burrill et al., 1997; Dasen et
al., 2003; Gavalas et al., 2003; Irving et al., 2002; Kawahara
et al., 2002; Pfeffer et al., 1998; Sapir et al., 2004; Segawa et
al., 2001). In cephalochordates, the expression of pax2/5/8
and en that appears during late developmental stages in the
amphioxus hindbrain, has been postulated to be related to
functions in neuron specification (Holland and Holland,
1999; Holland et al., 1997; Kozmik et al., 1999). The late
expression of ‘‘MHB genes’’ in the vertebrate and cepha-
lochordate hindbrains raises the hypothesis that tailbud stage
expression of the orthologs of these transcription factors in
the ‘‘urochordate hindbrain’’ is related to specification of
neuron fate and axon guidance rather than an organizer
function.
This alternative hypothesis is consistent with several
facts. First, shortly after hatching, Oikopleura and ascidians
probably have functional motor neurons because they show
coordinated tail movement. Second, motor neurons confined
to the ascidian CNS neck region begin to be specified as
early as late-gastrula stage (Katsuyama et al., 2005; Okada
et al., 2002). And third, the differences in expression
patterns of ‘‘MHB genes’’ in tailbud embryos between
different ascidian species, and between Oikopleura and
ascidians could merely reflect species-specific modifications
of developmental time and embryonic position of specific
neurons in the urochordate hindbrain rather than fundamen-
tal differences in CNS regionalization. Taken together, these
considerations suggest that the expression of ‘‘MHB genes’’
described so far in Oikopleura and ascidians reflects
development of specific neurons rather than action in an
MHB organizer. Functional experiments will be necessary
to disprove this conclusion.
Acknowledgments
We are grateful to Skipper B. Young of the ‘‘Charming
Polly’’ for help in larvacean collecting. We thank F. Mazet
for sharing with us her unpublished results on the expression
of Ci-pax2/5/8b in Ciona intestinalis. We thank L. Maves
and W. Cresko for helpful suggestions and discussion, and
A. Amores for providing Hox and Engrailed primers. We
thank undergraduate researchers T. Siriphatnaboon and T.
Keopuhiwa for help with animal care. We appreciate the
work of two anonymous reviewers for helping to improve
the manuscript. Complete fosmid sequencing was per-
formed under the auspices of the U.S. Department of
Energy, Office of Biological and Environmental Research,
in the University of California, Lawrence Berkeley National
Laboratory (contract DE-AC03-76SF00098). This material
is based on work supported by the National Science
Foundation under Grant No. IBN-0345203 and an IGERT
grant in Evolution of Development DGE-9972830. Any
opinions, findings, and conclusions or recommendations
expressed in this material are those of the author(s) and do
not necessarily reflect the views of the National Science
Foundation. We thank the Spanish Ministry of Education,
Culture and Sports for support for CC (EX2002-0059).
Appendix A. Supplementary data
Supplementary data associated with this article can be
found, in the online version, at doi:10.1016/j.ydbio.2005.
06.039.
References
Bassham, S., 2002. Molecular biology of a larvacean urochordate,
Oikopleura dioica, and the origin of chordate innovations. Ph.D.
Thesis. University of Oregon, Eugene, OR, USA.
Bassham, S., Postlethwait, J., 2000. Brachyury (T) expression in embryos
of a larvacean urochordate, Oikopleura dioica, and the ancestral role of
brachyury. Dev. Biol. 220, 322–333.
Bollner, T., Storm-Mathiesen, J., Ottersen, O.P., 1991. GABA-like
immunoreactivity in the nervous system of Oikopleura dioica (Appen-
dicularia). Biol. Bull. 180, 119–124.
Bone, Q., 1998. Nervous system, sense organs, and excitable epithelia. In:
Bone, Q. (Ed.), The Biology of Pelagic Tunicates. Oxford Univ. Press,
New York, pp. 55–80.
Bone, Q., Mackie, G.O., 1975. Skin impulses and locomotion in Oikopleura
(Tunicata: Larvacea). Biol. Bull. 149, 267–286.
Burrill, J.D., Moran, L., Goulding, M.D., Saueressig, H., 1997. PAX2 is
expressed in multiple spinal cord interneurons, including a population
of EN1 + interneurons that require PAX6 for their development.
Development 124, 4493–4503.
Callaerts, P., Halder, G., Gehring, W.J., 1997. PAX-6 in development and
evolution. Annu. Rev. Neurosci. 20, 483–532.

C. Canestro et al. / Developmental Biology 285 (2005) 298–315 313
Canestro, C., Bassham, S., Postlethwait, J.H., 2003. Seeing chordate
evolution through the Ciona genome sequence. Genome Biol. 4,
208–211.
Christen, R., Braconnot, J.-C., 1998. Molecular phylogeny of tunicates. A
preliminary study using 28S ribosomal RNA partial sequences:
important implications in terms of evolution and ecology. In: Bone, Q.
(Ed.), The Biology of Pelagic Tunicates. Oxford Univ. Press, New York,
pp. 265–271.
Cole, A.G., Meinertzhagen, I.A., 2004. The central nervous system of the
ascidian larva: mitotic history of cells forming the neural tube in late
embryonic Ciona intestinalis. Dev. Biol. 271, 239–262.
Corbo, J.C., Di Gregorio, A., Levine, M., 2001. The ascidian as a
model organism in developmental and evolutionary biology. Cell
106, 535–538.
Dasen, J.S., Liu, J.P., Jessell, T.M., 2003. Motor neuron columnar fate
imposed by sequential phases of Hox-c activity. Nature 425, 926–933.
Delsman, H.C., 1910. Beitrage zur Entwicklungsgeschichte von Oikopleura
dioica. Verh. Rijksinst. Orderz. Zee 3, 1–24.
Edvardsen, R.B., Seo, H.C., Jensen, M.F., Mialon, A., Mikhaleva, J.,
Bjordal, M., Cartry, J., Reinhardt, R., Weissenbach, J., Wincker,
P., Chourrout, D., 2005. Remodelling of the homeobox gene
complement in the tunicate Oikopleura dioica. Curr. Biol. 15,
R12–R13.
Fenaux, R., 1976. Cycle vital d’un appendiculaire: Oikopleura dioica Fol,
1872. Ann. Inst. Oceanogr., Paris 52, 89–101.
Fenaux, R., 1998. Life history of the Appendicularia. In: Bone, Q.
(Ed.), The Biology of Pelagic Tunicates. Oxford Univ. Press, Oxford,
pp. 151–159.
Force, A., Lynch, M., Pickett, F.B., Amores, A., Yan, Y.-L., Postlethwait, J.,
1999. Preservation of duplicate genes by complementary, degenerative
mutations. Genetics 151, 1531–1545.
Garcia-Fernandez, J., Holland, P.W., 1994. Archetypal organization of the
amphioxus Hox gene cluster. Nature 370, 563–566.
Gavalas, A., Ruhrberg, C., Livet, J., Henderson, C.E., Krumlauf, R., 2003.
Neuronal defects in the hindbrain of Hoxa1, Hoxb1 and Hoxb2 mutants
reflect regulatory interactions among these Hox genes. Development
130, 5663–5679.
Georges, D., Holmberg, K., Olsson, R., 1988. The ventral midbrain cells in
Oikopleura dioica (Appendicularia). Acta Embryol. Morphol. Exp. 9,
39–47.
Gionti, M., Ristoratore, F., Di Gregorio, A., Aniello, F., Branno, M., Di
Lauro, R., 1998. Cihox5, a new Ciona intestinalis Hox-related gene, is
involved in regionalization of the spinal cord. Dev. Genes Evol. 207,
515–523.
Glardon, S., Callaerts, P., Halder, G., Gehring, W.J., 1997. Conservation of
Pax-6 in a lower chordate, the ascidian Phallusia mammillata.
Development 124, 817–825.
Glardon, S., Holland, L.Z., Gehring, W.J., Holland, N.D., 1998. Isolation
and developmental expression of the amphioxus Pax-6 gene (Amphi-
Pax-6): insights into eye and photoreceptor evolution. Development
125, 2701–2710.
Graham, A., 2004. Evolution and development: rise of the little squirts.
Curr. Biol. 14, 956–958.
Graham, A., Lumsden, A., 1996. Interactions between rhombomeres
modulate Krox-20 and follistatin expression in the chick embryo
hindbrain. Development 122, 473–480.
Graham, A., Heyman, I., Lumsden, A., 1993. Even-numbered rhombo-
meres control the apoptotic elimination of neural crest cells from odd-
numbered rhombomeres in the chick hindbrain. Development 119,
233–245.
Hinman, V.F., Degnan, B.M., 2000. Retinoic acid perturbs Otx gene
expression in the ascidian pharynx. Dev. Genes Evol. 210, 129–139.
Hirth, F., Kammermeier, L., Frei, E., Walldorf, U., Noll, M., Reichert,
H., 2003. An urbilaterian origin of the tripartite brain: devel-
opmental genetic insights from Drosophila. Development 130,
2365–2373.
Holland, N.D., Chen, J., 2001. Origin and early evolution of the vertebrates:
new insights from advances in molecular biology, anatomy, and
paleontology. BioEssays 23, 142–151.
Holland, L.Z., Gibson-Brown, J., 2003. The Ciona intestinalis genome:
when the constraints are off. BioEssays 25, 529–532.
Holland, L.Z., Holland, N.D., 1999. Chordate origins of the vertebrate
central nervous system. Curr. Opin. Neurobiol. 9, 596–602.
Holland, L., Gorsky, G., Fenaux, R., 1988. Fertilization in Oikopleura
dioica (Tunicata, Appendicularia): acrosome reaction, cortical reaction
and sperm-egg fusion. Zoomorphology 108, 229–243.
Holland, P.W.H., Garcia-Fernandez, J., Williams, N.A., Sidow, A., 1994.
Gene duplications and the origins of vertebrate development. Develop-
ment, 125–133.
Holland, L.Z., Kene, M., Williams, N.A., Holland, N.D., 1997. Sequence
and embryonic expression of the amphioxus engrailed gene (AmphiEn):
the metameric pattern of transcription resembles that of its segment-
polarity homolog in Drosophila development. Development 124,
1723–1732.
Holmberg, K., 1984. A transmission electron microscopic investigation of
the sensory vesicle in the brain of Oikopleura dioica (Appendicularia).
Zoomorphology 104, 298–303.
Hudson, C., Lemaire, P., 2001. Induction of anterior neural fates in the
ascidian Ciona intestinalis. Mech. Dev. 100, 189–203.
Ikuta, T., Yoshida, N., Satoh, N., Saiga, H., 2004. Ciona intestinalis Hox
gene cluster: its dispersed structure and residual colinear expression in
development. Proc. Natl. Acad. Sci. U. S. A. 101, 15118–15123.
Imai, K.S., Satoh, N., Satou, Y., 2002. Region specific gene expressions in
the central nervous system of the ascidian embryo. Gene Expression
Patterns 2, 319–321.
Irving, C., Malhas, A., Guthrie, S., Mason, I., 2002. Establishing the
trochlear motor axon trajectory: role of the isthmic organiser and Fgf8.
Development 129, 5389–5398.
Jeffery, W.R., 2002. Ascidian gene-expression profiles. Genome Biol. 3
(10), 1030.1–1030.4.
Jiang, D., Smith, W.C., 2002. An ascidian engrailed gene. Dev. Genes
Evol. 212, 399–402.
Katsuyama, Y., Wada, S., Yasugi, S., Saiga, H., 1995. Expression of the
labial group Hox gene HrHox-1 and its alteration induced by retinoic
acid in development of the ascidian Halocynthia roretzi. Development
121, 3197–3205.
Katsuyama, Y., Okada, T., Matsumoto, J., Ohtsuka, Y., Terashima, T.,
Okamura, Y., 2005. Early specification of ascidian larval motor
neurons. Dev. Biol. 278, 310–322.
Kawahara, A., Chien, C.B., Dawid, I.B., 2002. The homeobox gene
mbx is involved in eye and tectum development. Dev. Biol. 248,
107–117.
Kozmik, Z., Holland, N.D., Kalousova, A., Paces, J., Schubert, N.,
Holland, L., 1999. Characterization of an amphioxus paired box
gene, AmphiPax2/5/8: developmental expression patterns in optic
support cells, nephridium, thyroid-like structures and pharyngeal gill
slits, but not in the midbrain boundary region. Development 126,
1295–1304.
Krauss, S., Johansen, T., Korzh, V., Fjose, A., 1991. Expression of the
zebrafish paired box gene pax [zf-b] during early neurogenesis.
Development 113, 1193–1206.
Lemaire, P., Bertrand, V., Hudson, C., 2002. Early steps in the formation of
neural tissue in ascidian embryos. Dev. Biol. 252, 151–169.
Locascio, A., Aniello, F., Amoroso, A., Manzanares, M., Krumlauf, R.,
Branno, M., 1999. Patterning and the ascidian nervous system:
structure, expression and transgenic analysis of the CiHox3 gene.
Development 126, 4737–4748.
Lumsden, A., Krumlauf, R., 1996. Patterning the vertebrate neuraxis.
Science 274, 1109–1115.
Macdonald, R., Scholes, J., Strahle, U., Brennan, C., Holder, N., Brand, M.,
Wilson, S.W., 1997. The Pax protein Noi is required for commissural
axon pathway formation in the rostral forebrain. Development 124,
2397–2408.
Marin, F., Charnay, P., 2000a. Hindbrain patterning: FGFs regulate Krox20

C. Canestro et al. / Developmental Biology 285 (2005) 298–315314
and mafB/kr expression in the otic/preotic region. Development 127,
4925–4935.
Marin, F., Charnay, P., 2000b. Positional regulation of Krox-20 and
mafB/kr expression in the developing hindbrain: potentialities of
prospective rhombomeres. Dev. Biol. 218, 220–234.
Martini, E., 1909. Studien uber die Konstanz histologischer Elemente: I.
Oikopleura longicauda. Z. Wiss. Zool. 92, 563–626.
Matsunaga, E., Araki, I., Nakamura, H., 2000. Pax6 defines the di-
mesencephalic boundary by repressing En1 and Pax2. Development
127, 2357–2365.
Maves, L., Jackman, W., Kimmel, C.B., 2002. FGF3 and FGF8 mediate a
rhombomere 4 signaling activity in the zebrafish hindbrain. Develop-
ment 129, 3825–3837.
Mazet, F., Hutt, J.A., Millard, J., Shimeld, S.M., 2003. Pax gene expression
in the developing central nervous system of Ciona intestinalis. Gene
Expression Patterns 3, 743–745.
Mazet, F., Hutt, J.A., Milloz, J., Millard, J., Graham, A., Shimeld, S.M.,
2005. Molecular evidence from Ciona intestinalis for the evolutionary
origin of vertebrate sensory placodes. Dev. Biol. 282, 494–508.
McClintock, J.M., Kheirbek, M.A., Prince, V.E., 2002. Knockdown of
duplicated zebrafish hoxb1 genes reveals distinct roles in hindbrain
patterning and a novel mechanism of duplicate gene retention.
Development 129, 2339–2354.
Meinertzhagen, I.A., Okamura, Y., 2001. The larval ascidian nervous
system: the chordate brain from its small beginnings. Trends Neurosci.
24, 401–410.
Meinertzhagen, I.A., Cole, A.G., Stanley, S., 2000. The central
nervous system, its cellular organisation and development, in the
tadpole larva of the ascidian Ciona intestinalis. Acta Biol. Hung. 51,
417–431.
Meinertzhagen, I.A., Lemaire, P., Okamura, Y., 2004. The neurobiology of
the ascidian tadpole larva: recent developments in an ancient chordate.
Annu. Rev. Neurosci. 27, 453–485.
Nagatomo, K., Fujiwara, S., 2003. Expression of Raldh2, Cyp26 and Hox-1
in normal and retinoic acid-treated Ciona intestinalis embryos. Gene
Expression Patterns 3, 273–277.
Nishino, A., Satoh, N., 2001. The simple tail of chordates: phylogenetic
significance of appendicularians. Genesis 29, 36–45.
Ohno, S., 1970. Evolution by Gene Duplication. Springer-Verlag,
New York.
Okada, T., Katsuyama, Y., Ono, F., Okamura, Y., 2002. The development of
three identified motor neurons in the larva of an ascidian, Halocynthia
roretzi. Dev. Biol. 244, 278–292.
Olsson, R., Holmberg, K., Lilliemark, Y., 1990. Fine structure of the brain
and brain nerves of Oikopleura dioica (Urochordata, Appendicularia).
Zool. Morphol. 110, 1–7.
Pfeffer, P.L., Gerster, T., Lun, K., Brand, M., Busslinger, M., 1998.
Characterization of three novel members of the zebrafish Pax2/5/8
family: dependency of Pax5 and Pax8 expression on the Pax2.1 (noi)
function. Development 125, 3063–3074.
Postlethwait, J., Amores, A., Cresko, W., Singer, A., Yan, Y.L., 2004.
Subfunction partitioning, the teleost radiation and the annotation of the
human genome. Trends Genet. 20, 481–490.
Puelles, E., Acampora, D., Lacroix, E., Signore, M., Annino, A., Tuorto, F.,
Filosa, S., Corte, G., Wurst, W., Ang, S.L., Simeone, A., 2003. Otx
dose-dependent integrated control of antero-posterior and dorso-ventral
patterning of midbrain. Nat. Neurosci. 6, 453–460.
Puelles, E., Annino, A., Tuorto, F., Usiello, A., Acampora, D., Czerny, T.,
Brodski, C., Ang, S.L., Wurst, W., Simeone, A., 2004. Otx2 regulates
the extent, identity and fate of neuronal progenitor domains in the
ventral midbrain. Development 131, 2037–2048.
Quiring, R., Walldorf, U., Kloter, U., Gehring, W.J., 1994. Homology of the
eyeless gene of Drosophila to the small eye gene in mice and Aniridia
in humans. Science 265, 785–789.
Raible, F., Brand, M., 2004. Divide et Impera—The midbrain–hindbrain
boundary and its organizer. Trends Neurosci. 27, 727–734.
Reichert, H., Simeone, A., 2001. Developmental genetic evidence for a
monophyletic origin of the bilaterian brain. Philos. Trans. R. Soc.
Lond., Ser. B Biol. Sci. 356, 1533–1544.
Rhinn, M., Brand, M., 2001. The midbrain–hindbrain boundary organizer.
Curr. Opin. Neurobiol. 11, 34–42.
Rubenstein, J.L., Shimamura, K., Martinez, S., Puelles, L., 1998. Region-
alization of the prosencephalic neural plate. Annu. Rev. Neurosci. 21,
445–477.
Sapir, T., Geiman, E.J., Wang, Z., Velasquez, T., Mitsui, S., Yoshihara, Y.,
Frank, E., Alvarez, F.J., Goulding, M., 2004. Pax6 and engrailed 1
regulate two distinct aspects of renshaw cell development. J. Neurosci.
24, 1255–1264.
Satoh, N., 2003. The ascidian tadpole larva: comparative molecular
development and genomics. Nat. Rev., Genet. 4, 285–295.
Scholpp, S., Lohs, C., Brand, M., 2003. Engrailed and Fgf8 act synergisti-
cally to maintain the boundary between diencephalon and mesencepha-
lon. Development 130, 4881–4893.
Schwarz, M., Alvarez-Bolado, G., Dressler, G., Urbanek, P., Busslinger,
M., Gruss, P., 1999. Pax2/5 and Pax6 subdivide the early neural tube
into three domains. Mech. Dev. 82, 29–39.
Segawa, H., Miyashita, T., Hirate, Y., Higashijima, S., Chino, N., Uyemura,
K., Kikuchi, Y., Okamoto, H., 2001. Functional repression of islet-2 by
disruption of complex with Ldb impairs peripheral axonal outgrowth in
embryonic zebrafish. Neuron 30, 423–436.
Seo, H.-C., Kube, M., Edvardsen, R.B., Jensen, M.F., Beck, A., Spriet, E.,
Gorsky, G., Thompson, E.M., Lehrach, H., Reinhardt, R., Chourrout,
D., 2001. Miniature genome in the marine chordate Oikopleura dioica.
Science 294, 2506.
Seo, H.C., Edvardsen, R.B., Maeland, A.D., Bjordal, M., Jensen, M.F.,
Hansen, A., Flaat, M., Weissenbach, J., Lehrach, H., Wincker, P.,
Reinhardt, R., Chourrout, D., 2004. Hox cluster disintegration with
persistent anteroposterior order of expression in Oikopleura dioica.
Nature 431, 67–71.
Shanmugalingam, S., Houart, C., Picker, A., Reifers, F., Macdonald, R.,
Barth, A., Griffin, K., Brand, M., Wilson, S.W., 2000. Ace/Fgf8 is
required for forebrain commissure formation and patterning of the
telencephalon. Development 127, 2549–2561.
Sharman, A.C., 1999. Some new terms for duplicated genes. Semin. Cell
Dev. Biol. 10, 561–563.
Shimeld, S.M., Holland, P.W., 2000. Vertebrate innovations. Proc. Natl.
Acad. Sci. U. S. A. 97, 4449–4452.
Sordino, P., Belluzzi, L., De Santis, R., Smith, W.C., 2001. Developmental
genetics in primitive chordates. Philos. Trans. R. Soc. Lond., Ser. B
Biol. Sci. 356, 1573–1582.
Stach, T., Turbeville, J.M., 2002. Phylogeny of Tunicata inferred from
molecular and morphological characters. Mol. Phylogenet. Evol. 25,
408–428.
Swalla, B., Cameron, C., Corley, L., Garey, J., 2000. Urochordates are
monophyletic within the deuterostomes. Syst. Biol. 49, 52–64.
Takahashi, T., Holland, P.W., 2004. Amphioxus and ascidian Dmbx
homeobox genes give clues to the vertebrate origins of midbrain
development. Development 131, 3285–3294.
Wada, H., 1998. Evolutionary history of free-swimming and sessile
lifestyles in urochordates as deduced from 18S rDNA molecular
phylogeny. Mol. Biol. Evol. 15, 1189–1194.
Wada, H., Satoh, N., 1994. Details of the evolutionary history from
invertebrates to vertebrates, as deduced from the sequences of 18S
rDNA. Proc. Natl. Acad. Sci U. S. A. 91, 1801–1804.
Wada, H., Satoh, N., 2001. Patterning the protochordate neural tube. Curr.
Opin. Neurobiol. 11, 16–21.
Wada, S., Katsuyama, Y., Sato, Y., Itoh, C.S., Aiga, H., 1996. Hroth, an
orthodenticle-related homeobox gene of the ascidian, Halocynthia
roretzi: its expression and putative roles in the axis formation during
embryogenesis. Mech. Dev. 60, 59–71.
Wada, H., Saiga, H., Satoh, N., Holland, P.W.H., 1998. Tripartite
organization of the ancestral chordate brain and the antiquity of
placodes: insights from ascidian Pax2/5/8, Hox and Otx genes.
Development 125, 1113–1122.

C. Canestro et al. / Developmental Biology 285 (2005) 298–315 315
Wada, S., Tokuoka, M., Shoguchi, E., Kobayashi, K., Di Gregorio, A.,
Spagnuolo, A., Branno, M., Kohara, Y., Rokhsar, D., Levine, M., Saiga,
H., Satoh, N., Satou, Y., 2003. A genomewide survey of developmen-
tally relevant genes in Ciona intestinalis: II. Genes for homeobox
transcription factors. Dev. Genes Evol. 213, 222–234.
Wada, S., Sudou, N., Saiga, H., 2004. Roles of Hroth, the ascidian otx gene,
in the differentiation of the brain (sensory vesicle) and anterior trunk
epidermis in the larval development of Halocynthia roretzi. Mech. Dev.
121, 463–474.
Walshe, J., Maroon, H., McGonnell, I.M., Dickson, C., Mason, I., 2002.
Establishment of hindbrain segmental identity requires signaling by
FGF3 and FGF8. Curr. Biol. 12, 1117–1123.
Wurst, W., Bally-Cuif, L., 2001. Neural plate patterning: upstream and
downstream of the isthmic organizer. Nat. Rev., Neurosci. 2, 99–108.