Embed Size (px)
DESCRIPTION
adiccion
Citation preview

B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4
ava i l ab l e a t www.sc i enced i rec t . com
www.e l sev i e r. com/ loca te /b ra in res rev
Review
Functional neuroimaging of reward processing anddecision-making: A review of aberrant motivational andaffective processing in addiction and mood disorders
Esther Kristina Diekhof ⁎, Peter Falkai, Oliver GruberSystems Neuroscience Unit, Department of Psychiatry and Psychotherapy, Georg August University,Von-Siebold-Strasse 5, D-37075 Goettingen, Germany
A R T I C L E I N F O
⁎ Corresponding author. Fax: +49 551 39 2798.E-mail addresses: [email protected] (E.K.
0165-0173/$ – see front matter © 2008 Elsevidoi:10.1016/j.brainresrev.2008.07.004
A B S T R A C T
Article history:Accepted 11 July 2008Available online 21 July 2008
The adequate integration of reward- and decision-related information provided by theenvironment is critical for behavioral success and subjective well being in everyday life.Functional neuroimaging researchhas alreadypresented a comprehensivepictureonaffectiveand motivational processing in the healthy human brain and has recently also turned itsinterest to the assessment of impaired brain function in psychiatric patients. This articlepresents an overview on neuroimaging studies dealing with reward processing and decision-makingby combiningmost recent findings from fundamental and clinical research. It providesan outline on the neural mechanisms guiding context-adequate reward processing anddecision-making processes in the healthy brain, and also addresses pathophysiologicalalterations in the brain's reward systemthathavebeenobserved in substance abuse andmooddisorders, two highly prevalent classes of psychiatric disorders. The overall goal is to criticallyevaluate the specificity of neurophysiological alterations identified in these psychiatricdisorders and associated symptoms, and to make suggestions concerning future research.
© 2008 Elsevier B.V. All rights reserved.
Keywords:Substance abuse disorderMajor depressionBipolar disorderfMRIPETALE meta-analysis
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1652. Learning to predict reward . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1653. Representation of reward and punishment—‘relevance detectors’ in the brain . . . . . . . . . . . . . . . . . . . . . . 1664. Parsing reward—Affective value and motivational significance . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1665. Deciding advantageously—Reversal-learning, prospective planning and impulsive choice . . . . . . . . . . . . . . . 1676. Implications for psychiatric disorders . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 167
6.1. Activation likelihood estimation (ALE) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1687. Substance (ab)use disorders . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 169
7.1. Regional distribution of activation associated with drug-related stimulation in addiction—Activation LikelihoodEstimation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 171
7.2. Substance (ab)use disorders—Conclusions and outstanding questions . . . . . . . . . . . . . . . . . . . . . . 171Likelihood Estimation
Diekhof), [email protected] (P. Falkai), [email protected] (O. Gruber).
er B.V. All rights reserved.

165B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4
8. Mood disorders—Changes in motivational relevance and affective processing . . . . . . . . . . . . . . . . . . . . . 1768.1. Regional distribution of activation associated with mood-congruent processing in MDD—Activation Likelihood
Estimation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1788.2. Mood disorders—Conclusions and outstanding questions . . . . . . . . . . . . . . . . . . . . . . . . . . . . 178
9. General conclusion. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17910. Limitations of ALE . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 180Appendix A. Supplementary data . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 180References. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 180
Likelihood Estimation
1. Introduction
Reward is no unitary concept, but subsumes several compo-nents including learning-related, motivational, affective anddecisional aspects (Chau et al., 2004). Organisms formpredictions, compare different reward opportunities withregard to their relative value and balance immediate rewardwith long-term utility to guarantee overall behavioral success.In the healthy brain the integration of reward-relatedinformation is ensured by several cortical and subcorticalregions that form the brain's reward circuit, which is alsoinvolved in the selection and initiation of goal-directedbehavior (O'Doherty, 2004). Adaptive decision-making furtherrelies on regions that represent higher-order cognitive controlprocesses (e.g., planning) and help to balance affectivelyguided impulsive choice and rational future-oriented con-siderations in order to maximize long-term behavioralsuccess (Trepel et al., 2005).
Disturbances within the reward system have detrimentaleffects on behavior. Especially mood disorders and substanceabuse disorders have already been reported to involve overtbehavioral deficits in emotional processing leading to abnor-mal motivational and affective processing. Recent neuroima-ging studies have provided a useful means that allowed torelate these overt deficits to their underlying neural substratesand to identify specific regional disturbances within thereward system of psychiatric patients. Nevertheless, tocompletely understand pathophysiological alterations it isalso necessary to be aware of the neurophysiology of rewardprocessing and decision-making in the healthy brain. Thisreview will therefore provide a brief outline on brain regionsthat in the healthy brain predict rewards, sense rewardingoutcomes, process them in a context-sensitive way and finallydecide upon the actions that lead to the direct acquisition ofreward by also balancing it with long-term reward utility. Wewill thereby briefly address recent functional neuroimagingstudies on healthy human subjects, before in a second stepwewill deal with their relevance for the pathophysiology ofpsychiatric disorders. The focus will be on substance abusedisorders and mood disorders, two highly prevalent classes ofpsychiatric disorders, which can be characterized by overtmotivational deficits, disordered affective processing andimpairments in decision-making. The aim of this article is toreview and evaluate neuroimaging findings from the lastdecade with regard to their contribution to the understandingof pathophysiological processes and their disorder-relatedspecificity regarding alterations in the reward system. Fromthis, we will be able to derive useful suggestions for futureneuroimaging research by also pointing to potential intercon-
nections with other experimental approaches (e.g., psycho-pharmacological research).
2. Learning to predict reward
The prospective organization of goal-directed behavior requiresthe correct prediction of when and where rewards will occur.Decisive information for the formation of such predictions hasthereby been assumed to be mainly derived from the observa-tion of environmental regularities (e.g., the contingencybetween a certain action and its outcome; O'Doherty, 2004). Asthe major source for the release of a neural teaching signalproviding the organism with important information on chan-ging reward contingencies, midbrain dopamine projections tothe striatum and frontal cortex have been found to commu-nicate an error in reward prediction. Accordingly, in animalsunexpected or bigger than expected rewards and also unex-pected reward-predicting stimuli elicited a positive response inthese neurons, whereas the same dopaminergic neuronsremained unresponsive to a completely expected reward(Schultz, 1998). In addition, dopaminergic neurons have alsobeen observed to be responsive to a large category of salient andarousing events, including aversive, high intensity and novelstimuli without any rewarding property (see Horvitz, 2000 for acomprehensive review). It has hence been suggested thatdopamine neurons represent an essential component in theprocess of attentional andbehavioral switchingbypreparing theorganism for the appropriate reaction to all kinds of salientenvironmental changes regardless of their affective valence(Redgrave et al., 1999; Horvitz, 2000), even though this view hasnot remained undisputed (Ungless, 2004).
In humans, activations observed in subcortical dopaminetarget sites like the ventral striatum resembled the rewardprediction error signal found in nonhuman primates (O'Do-herty et al., 2003a, 2004; Davidson et al., 2004; Abler et al. 2006;Wrase et al., 2007a), but also represented salient events ingeneral (Zink et al., 2003, 2006). Dopaminergic function maythereby have a direct modulatory influence on striatal activityand behavioral choice, which has recently been demonstratedthrough pharmacological probe (Pessiglione et al., 2006).Within this context, a further distinction has to be madebetween prediction error-related activation in the ventralstriatum that coded the difference between the actual andexpected outcome during Pavlovian conditioning and activa-tion in dorsal striatal areas that occurred preferentially duringinstrumental conditioning and positive reinforcement learn-ing (O'Doherty et al., 2004). This observation indicated that thedorsal striatum supposedly serves a central function in the

166 B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4
establishment of stimulus–response(–reward) contingencies,while ventral striatal activity represents outcome-relatedpredictions, even independent of affective value (see alsoWrase et al., 2007a). These complementary functions ofventral and dorsal striatal regions may well accord withmodels of reinforcement learning and instrumental condi-tioning (e.g., the actor–critic model; Sutton and Barto, 1998;Holroyd and Coles, 2002). Accordingly, the prediction errorresponse in the ventral striatum may allow organisms toupdate predictions of expected reward or punishment,whereas the response in the dorsal striatum supposedlyoptimizes reward-related action choice and thus helps theorganism to maximize expected long-term reward (O'Dohertyet al., 2004; Wrase et al., 2007a).
Other regions involved in both predictive learning ofpositive and aversive events in humans were the amygdala(pain relief; Seymour et al., 2005; monetary loss; e.g., Yacubianet al., 2006) and frontal dopamine target sites, whose activitywas associated with reward-related learning processes. Thesefrontal areas included the medial orbitofrontal cortex (OFC;Berns et al., 2001; O'Doherty et al., 2003a; Tobler et al., 2006),but also lateral parts of the OFC (Tobler et al., 2007). Other partsof the medial prefrontal cortex like the anterior cingulatecortex (ACC)were activated by successfully obtained reward inamanner that scaled to the amount of the respective outcome(Knutson et al., 2003), or responded to unexpected events thatrequired the withholding of an already prepared response,which indicated that this region may have been involved inguiding behavioral choice based on highly salient environ-mental events (Davidson et al., 2004).
After learning has occurred, reward-related neural res-ponses have been observed to shift from the actual reward tothe reward-predicting stimulus or response (O'Doherty, 2004).Activation in the ventral striatum (in particular in the nucleusaccumbens) has thereby been reported to convey the motiva-tional significance of emotionally laden stimuli. Ventralstriatal responses represented either the magnitude of ananticipated gain predicted by a visual cue (Breiter et al., 2001;Knutson et al., 2001; Galvan et al., 2005; Knutson et al., 2005), orthe anticipation of an aversive stimulus (Jensen et al., 2003).Similarly, the OFC also exhibited increased activation inresponse to the anticipation of a sweet taste reward (O'Dohertyet al., 2002) and to a rewarding outcome occurring contingenton a manual response (Galvan et al., 2005). Amygdala activa-tion has more often been observed during the anticipation ofaversive events (e.g., Breiter et al., 2001; Glaescher and Büchel,2005), even though there is also evidence for its involvement inthe prediction of positive events (Gottfried et al., 2003). Thisalso underlines the important role of these regions inconditioning and stimulus–reward association learning, inwhich the reward value of a reinforcer is transferred to areward-predicting cue (e.g., Cox et al., 2005; Gottfried et al.,2003).
3. Representation of reward and punishment—‘relevance detectors’ in the brain
Besides the establishment of predictions of when and whererewards will occur, adaptive behavior requires the ability to
represent the current motivational value of available rewards,which permits context-sensitive behavioral choices betweendifferent rewards.Within the human brain's reward circuit, theOFC has thereby beenmost consistently found to represent therewarding outcomes and the relative value of reinforcers(O'Doherty, 2004). Particularly, activations in posterior parts ofthe OFCwere associatedwith the positivemotivational value ofnatural reinforcers (e.g., food) depending on current need state(O'Doherty et al., 2000; Morris and Dolan, 2001; Gottfried et al.,2003; Kringelbach et al., 2003), which was independent ofstimulus identity (e.g., both verbal feedback and monetaryreward activated an identical region, but to a different degree;Kirsch et al., 2003). Thus, activity in these brain regions mayprovide a collective motivational currency for adaptive (beha-vioral) decisions (Montague and Berns, 2002; Kringelbach, 2005).Additionally, some studies further proposed a similar role forthe amygdala, the ventral striatumand the ACC (Gottfried et al.,2003). However, in contrast to the OFC, amygdala activationsfailed to parallel the decision for or against a reinforcer (Aranaet al., 2003), and the ACC and the ventral striatal responsesupposedly rather seemed to play an important role in theresponse selection process (Gottfried et al., 2003).
Interestingly, activity in most of these regions was alsoobservedduringprocessing of aversive andpunishing outcomes.In theamygdala increasedactivationalsooccurred inassociationwith fear- or threat-cues, especially facial expressions (Whalen,1998; Calder et al., 2001; Williams, 2006), and the posterior OFCwas also responsive to aversive events like unpleasant odors(e.g., Gottfried et al., 2002 but see Rolls et al., 2003). This indicatedthat the amygdala and at least posterior parts of the OFC mayhave rather coded the emotional relevance than the actualaffective valence of environmental stimuli, which was alsosupported by the findings of a valence-independent representa-tion of all motivationally significant events in the amygdala(Garavan et al., 2001; Liberzon et al., 2003; Phan et al., 2004;Fitzgerald et al., 2006) and a general representation of motiva-tional significance in the posterior OFC (unpublished data fromour laboratory). These ‘relevance detectors’ may therefore bedevoted to the processing of a broader category of the motiva-tional relevanceof biologically significant stimuli independent ofaffective valence (see also Sander et al., 2003).
4. Parsing reward—Affective value andmotivational significance
The existence of valence-independent ‘relevance detectors’implies that it may be critical to make a clear distinctionbetween the affective value of a stimulus (i.e., positive ornegative) and its actual motivational significance for theorganism. Although these two components usually occurcollectively in the environment, at least in animals there isevidence that they recur on different neural substrates.Dopamine has been assumed to represent the motivational‘wanting’ component of reward (eliciting approach or avoid-ance responses), while the hedonic or pleasure componentremained widely unaffected by manipulations of the dopami-nergic system. Instead affective ‘liking’ supposedly recurs onopioid neurotransmission onto GABAergic spiny neurons inthe nucleus accumbens (for an elaborate review on the neuro-

167B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4
biological basis of the concepts of motivation and pleasureplease refer to Berridge and Robinson, 2003). Neuroimagingevidence for a regional dissociation of the two reward com-ponents is sparse, but there may be some hints for candidateregions that supposedly (exclusively) represent positive hedo-nic value (e.g., themedial orbitofrontal cortex; O'Doherty et al.,2001).
5. Deciding advantageously—Reversal-learning, prospective planning andimpulsive choice
Decision-making refers to the process of forming preferences,selecting and executing actions, as well as evaluating (possibleor anticipated) outcomes (Ernst and Paulus, 2005). It requiresthe ability to accurately evaluate elements of outcome,including motivational value, outcome predictability, andrisk. On the most basic level, organisms base their behavioraldecisions on simple stimulus–response–outcome associa-tions. Significant changes in themotivational value of environ-mental stimuli should thereby directly modulate behavioralchoice, because alternative options might provide the chanceto increase overall profit or to avoid future failure. Accordingly,in humans, cortical activation related to rapid behavioraladjustments in response to environmental change havemainly been observed in parts of the orbitofrontal and thecingulate cortex. Interestingly, the lateral OFC was therebyexclusively responsive to negative feedback that was followedby a subsequent change in or an immediate optimization ofbehavior, but remained unresponsive to negative feedback perse (O'Doherty et al., 2001; Cools et al., 2002; Kringelbach andRolls, 2003; O'Doherty et al., 2003b; Wrase et al., 2007a). Thisindicated that the lateral OFC may play an important role inflexible guidance of behavior by supporting inhibitory controlprocesses and helping organisms to overcome perseverativeresponding (Elliott and Deakin, 2005). The active response-se-lection component of reward-based decision-making in thereversal-learning paradigm has further been assigned to theACC and associated parts of the medial frontal cortex (Bushet al., 2002; Kringelbach and Rolls, 2003; see also Kringelbach,2005) and activation of the ACC has also been observed duringlearning frompunishment in that it correlatedwith behavioraladjustments following punishment-related information (e.g.,Wrase et al., 2007a). In addition, striatal subregions may alsoserve important functions during goal-directed decision-making based on simple stimulus–response–outcome asso-ciations. The specific topography of focal projections fromvarious parts of the frontal cortex may thereby predisposecorresponding striatal regions for differential, but neverthe-less complementary functions during reward-related deci-sion-making (Haber et al., 2000, 2006). Accordingly, the ventralstriatum and adjacent parts of the dorsomedial and centralstriatum, which are densely connected with ACC, OFC andventromedial frontal cortex, may integrate various aspects ofreward evaluation including reward predictability, expecta-tion and salience and may thus coordinate behavioral guidingrules (see also Section 2 of this review). In contrast, dorsal androstral aspects of the striatum receive dense projections fromthe dorsolateral prefrontal cortex, which partly converge with
inputs from ACC and OFC, suggesting a central role for dorsalstriatal compartments in the synchronization of differentaspects of reward-related learning operations. Through theseconnections the dorsal striatum may establish the linkagebetween reward and subsequent behavior, integrate incentivedrive with long-term planning or facilitate habit formation(Haber et al., 2000, 2006; see also Balleine et al., 2007).
Although on many occasions direct stimulus–outcome orreward-related information may suffice to guide adaptivebehavior (e.g., in reversal-learning or reinforcer devaluationparadigms), there are many decisions that neverthelessrequire subjects to deliberate about future consequences ofcurrent choices to maximize overall future prospects. Thisnecessitates the ability to balance the direct emotional‘wanting’ of immediate gratification and cognitive future-oriented strategic planning to achieve optimal choice beha-vior. For instance, in the Iowa Gambling Task (IGT) subjectshave to abstain from a short-term benefit to maximize long-termmonetary profit (Bechara et al., 1996). On the neural level,especially the dorsolateral prefrontal cortex (DLPFC) has beenassumed to organize the temporal integration of decision-related information in humans in that particular task (Trepelet al., 2005). Resting-state cerebral blood flow in this regionshowed a significant correlation with IGT performance (Adin-off et al., 2003; Northoff et al., 2006), while patients with lesionsof the DLPFC failed to choose optimally in this task (Fellowsand Farah, 2005). In addition, the ventromedial prefrontalcortex (including the OFC) has also been reported to beassociated with success in the IGT (Northoff et al., 2006).Nevertheless, this association may rather be assigned to thedirect reversal-learning component of the task (i.e., to theinability to shift from the initially preferred ‘bad’ decks to the‘good’ decks; Fellows and Farah, 2005) that may equallyaccount for the observed relation between activation inother medial frontal areas and behavioral success in the IGT(Fukui et al., 2005).
6. Implications for psychiatric disorders
Overt behavioral impairments of motivational and affectiveprocessing like an overvaluing of some reward categories (e.g.,drugs in patients with substance use disorders) at the expenseof others or a mood-congruent bias towards a certain categoryof affective stimuli (e.g., sad faces in depressives) have beenreported to significantly compromise everyday life of psychia-tric patients. Despite small sample sizes and obvious con-founds in clinical populations in comparison to healthycontrol subjects (e.g., medication effects or differential com-pliance), neuroimaging studies provide a powerful means forrevealing the underlying pathophysiological alterations thatrelate to behavioral impairments in psychiatric populations,which may further help to identify disorder-specific markersin the brain.
The first part of this article provided a brief outline onneuroimaging findings from studies on reward processing anddecision-making in healthy human subjects. Evidence fromfundamental research thereby suggested that partially mis-cellaneous but nevertheless overlapping circuits allow theprediction and anticipation of positive or negative outcomes

168 B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4
andhelp to guide adaptive behavioral adjustments in responseto changing reward contingencies allowing maximization of(expected) long-term profit during decision-making and rein-forcement learning. In the second part of this article we willprovide a comprehensive review of the results from recentneuroimaging studies using either the method of functionalmagnetic resonance imaging (fMRI) or positron emissiontomography (PET) to reveal the underlying pathophysiologyof affective, motivational and decision-making deficits inaddiction and mood disorders. The focus thereby lies onneuropsychiatric studies that dealt with reward prediction,general processing of reward feedback or motivational cues(e.g., mood-congruent stimuli in depression or drug-relatedstimulation in addiction) or with reward-related decision-making. The specific aims of our review of the psychiatricneuroimaging literature from the last decade are (1.) to providea systematic, comprehensive overview of (disorder-specific)characteristics of neural aberrations during motivational andaffective processing in psychiatric patients with mood dis-orders or addiction, with a direct reference to overt symptomsand behavioral alterations in these disorders, and (2.) toachieve a critical appraisal of the literature that allows us tohighlight factual confounds and shortcomings of previousneuropsychiatric research. Finally, we also would like to (3.)draw conclusions that help us to point out currently unan-swered questions that (4.) allow us to present suggestions forfuture neuroimaging research.
For this purpose we pursued a systematic MEDLINE searchfor “substance abuse disorders” (or “addiction”, “drug abuse” or“craving”), “major (or unipolar) depression” and “bipolar(affective) disorder” by including the name of the respectivedisorder in the search termini and combining it (“AND”) with“reward”, “decision-making”or “emotion” in combinationwith“fMRI” or “PET”. We thereby restricted our search to relevantarticles published within the last decade (between 1997 and2007). Since we wanted to focus on functional neuroimagingresearch in adults with mood disorders or substance abusedisorders, studies that dealt with pediatric or adolescent casesor reported results from structural imaging were excluded.Also, neuroimaging studies assessing the neural correlates of“behavioral” addictions (e.g., pathological gambling) were notincluded in our systematic literature review.
In all, combination of the search termini and subsequentapplication of the exclusion criteria yielded 57 articles. Of the26 neuroimaging studies on addiction, 17 articles dealt withcue-induced craving in subjects during various states ofabstinence (5 articles dealt with alcohol abuse, 6 with cocaineabuse, 1withopiate abuseand5 testedhealthy smokers). In yetanother 5 studies subjects were tested with paradigmsassessing the effects of monetary reward or feedback (1 inalcohol abusers, 2 in cocaine abusers, 1 in opiate addicts and 1in healthy smokers). The IGT was employed by 3 studies onreward-related decision-making in addiction (2 in cocaine and1 inmarijuana abusers) and the Cambridge Risk Taskwas usedby one study (in heroin abusers). For the search term “major (orunipolar) depression” we found 18 studies that used motiva-tionally relevant cues of different affective value in mostlymedicated patients (9 studies employed either emotionalpictures orwords, 5 used emotional facial displays, 1 presentedperformance-related feedback, 1 used a reward expectation
paradigm and 2 studies employed a mood-provocation task).Finally, thirteen neuroimaging studies dealt withmotivationalprocessing in medicated patients with bipolar disorder (in 8 ofthese studies the sample was in the state of mania or a mixedstate, four assessed only euthymic patients and 1 dealt withpatients in adepressive state).Of these studies, 5usedaffectivefacial displays of varying value to assess explicit and implicitaffect processing, 6 presented emotional words or pictures, 1employed a reward expectation paradigm and 1 assessed risk-taking performance in bipolar patients in the state of mania.
The results of these functional imaging studies are listed inTables 1–3, which delineate affected brain regions and reportneural activation levels in relation to the specific experimentaltasks employed (in comparison to either a healthy controlgroup or to neutral stimulation). Supplementary Tables 1 to 5provide additional and more detailed information on indivi-dual studies (e.g., information on imaging method, samplesize, medication status and subtraction-contrasts). In thefollowing sections we will summarize these findings toachieve a comprehensive and conclusive picture on currentevidence from neuroimaging research in mood disorders andsubstance abuse.
6.1. Activation likelihood estimation (ALE)
In addition, to our systematic literature review we also pur-sued a meta-analysis using activation likelihood estimation(ALE) as implemented in the GingerALE software (Laird et al.,2005a; see also Turkeltaub et al., 2002). This voxel-basedapproach allows the analysis of the spatial distribution andconcordance of coordinates collected from a predefined set ofstudies that dealt with a specific domain or psychiatricdisorder in human brain mapping. ALE models report a focusof activation as the center of a Gaussian probability distribu-tion and create a whole-brain statistical map that estimatesthe likelihood of activation for each voxel for that task asdetermined by the complete set of studies (see also Laird et al.,2005b). Pursuing a quantitative meta-analysis was intended tocomplement the systematic literature review, which wasmainly based on anatomical labels assigned by authors, byproviding an increased spatial level of distinction. In parti-cular, we wanted to examine the consistency within theregional distribution of drug-cue-related hyperactivity inaddiction and further assess the spatial concordance of hyper-activation to mood-congruent or negative stimuli in majordepression (MDD) when compared to the healthy population.This meta-analysis is understood as an initial attempt toidentify disorder-specific alterations in the pathophysiologicalmechanisms leading to a bias in salience attribution towards acertain category of stimuli in two highly prevalent neuropsy-chiatric disorders and to derive suggestions as well as possiblehypotheses for future research.
For this purpose we extracted Talairach-coordinatesreported by the relevant studies. Coordinates reported inthe stereotactic space of the template from the MontrealNeurological Institute (MNI) were converted to Talairachspace using the Lancaster transform (icbm2tal) in GingerALE(Laird et al. 2005a). We only selected activation maxima thateither occurred in patients in association with a relevantcondition-of-interest (e.g. drug-related cues in addiction)

169B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4
compared to a neutral control condition and/or showed asignificant difference in patients and healthy control sub-jects during processing of a condition-of-interest (seeSupplementary Tables 6 and 7). In case both comparisonswere reported, only the latter comparison was used. If astudy reported results from both direct subtraction-contrastsand correlation analyses, only the former were included inthe meta-analysis. From studies that reported neighboringpeaks that lay maximally 2 voxels apart we selected only thepeak with the highest p-value reported within that particularregion. This was intended to avoid a bias of the meta-analytic results in favor of a subset of articles whenincluding multiple contrasts from the same article and alsoto increase the reliability of activation by not allowingstudies that report more peaks within one region tocontribute disproportionately. Studies that reported separateresults for male and female subjects were entered ascombined data.
Studies were excluded if they only reported anatomicallabels without coordinates (Addiction: Wang et al., 1999;Wexler et al., 2001; McClernon et al., 2005; MDD: Sheline etal., 2001; Siegle et al., 2002), or only assessed medicationeffects on brain function (MDD: Tremblay et al., 2005). Withregard to MDD studies were also not included if they onlypresented positive stimuli (Elliott et al., 1998; Kumari et al.,2003; Schaefer et al., 2006), or reported no differentialactivation in the direct comparison of controls and patientsduring negative stimulation (Lawrence et al. 2004; Keedwell etal., 2005a).
ALE maps were created according to the proceduredescribed by Turkeltaub et al. (2002). Reported foci of maximalanatomical difference were modeled as the peaks of three-dimensional Gaussian probability density functions with afull-width half-maximum (FWHM) of 10 mm. These probabil-ities were combined to construct a map of the ALE score ateach voxel. A permutation test (5000 permutations) wasperformed in order to determine the statistical significanceof randomly generated foci using the same FWHM value andan identical number of foci as in the computation of ALEvalues. Probability maps were then corrected for multiplecomparisons using the false discovery rate (FDR) and thre-sholded at p<0.05 with a minimal cluster extent of 100 voxels(see also Laird et al., 2005a). Separate ALE maps werecomputed for addiction and MDD in order to assess regionalconcordance in activation patterns within groups.
After application of the exclusion criteria 14 of 17 studiesthat reported hyperactivation in association with drug-relatedstimulation in addiction remained in the sample, yielding atotal of 122 foci (see Supplementary Table 6). Displaying thesefoci in Talairach space with the Java-based BrainMap applica-tion Sleuth (Laird et al., 2005c) revealed a diffuse pattern ofactivation across the whole-brain with some convergence infrontal areas (see Fig. 1). This pattern appeared to be quitesimilar for different drugs of abuse.
For MDD, 10 out of 17 studies could be included in themeta-analysis comprising 74 foci of hyperactivation relatedto negative stimulation (see Supplementary Table 7). InTalairach space peaks were distributed diffusely withsomewhat higher density in anterior parts of the brain(see Fig. 2).
7. Substance (ab)use disorders
Addiction may be best described as a pathology of incentivemotivation and behavioral control. As the most prominentbehavioral feature, patients commonly exhibit a decreasedability to control the desire to obtain drugs, despite knowledgeabout the aversive consequences following drug intake or thelow expectation of actual pleasure expected from the drug (seeSchoenbaum et al., 2006). The compulsive character of drugseeking, the obvious lack of inhibitory control and the lackingability to integrate information on aversive outcomes intobehavioral choice may therefore be assigned to a pathologicalattribution of motivational relevance to the drug of abuse or tocues predicting the drug, which makes the drug maximallysalient at the expense of other available (naturally) rewardingstimuli affecting all stages of reward processing (see Robinsonand Berridge, 2003).
Aberrant brain activations appear to play a role in severalcomponents of maladaptive behavior observed in substanceabuse, including the motivational bias in reward expectancy,reward evaluation and craving as well as impaired decision-making abilities (London et al., 2000). Learned, drug-relatedcues thereby produced brain activations comparable to thoseseen with non-drug evocative reward stimuli in healthycomparison subjects and affected roughly the same brainregions that in healthy subjects had been found to be involvedin reward evaluation andprediction (see above). Inmore detail,addiction to various kinds of substances including alcohol,nicotine, cocaine and opiates was accompanied by abnormalblood flow and altered activation patterns in reward-sensitivedopamine-dependent sites in the striatum, the amygdala aswell as in parts of the frontal cortex like the OFC, the ACC andthe DLPFC (see Goldstein and Volkow, 2002). These brainregions showed an activity bias towards the drug of abuse ordrug-related stimuli and exhibited increased activation todrug-related stimuli compared to neutral stimuli in abstinentindividuals, especially during the state of craving or acutewithdrawal (e.g., Wrase et al., 2002; see also: Goldstein andVolkow, 2002; Dom et al., 2005; see also Table 1 and Sup-plementary Table 1). Since addiction to different kinds ofsubstances (nicotine: Franklin et al., 2007; alcohol: Hermann etal., 2006; and partly also opiate and cocaine: Sell et al., 2000;Risinger et al., 2005) appeared to lead to similar activationincreases in the ‘relevance detectors’ OFC and amygdala aswell as other regions of the medial and dorsal frontal lobeduring drug-related stimulation (see Fig. 1), the neural mecha-nism of the motivational bias in addiction may be bestcharacterized as a general neural reward evaluation deficit.
Apart from an overvaluing of drug-reward, patients alsoexhibited a decreased sensitivity to the relative value of otheravailable non-drug rewards, which was also obvious on theneural level. Despite the limited number of neuroimagingstudies that directly assessed reward valuation independentlyof drug-related stimuli (cf., Table 1 and Supplementary Table2), evidence suggests that at least some substance abusersmay be compromised in their sensitivity for different amountsof monetary reward and verbal feedback. The deficit observedin some patients was reflected in either reduced or evenabsent activations of the ‘relevance detectors' OFC, amygdala

Table 1 – Activations in regions of the brain's reward circuit in patients with substance abuse disorders or in healthysmokers observed during performance of emotional and reward-processing tasks
Region Direction ofactivation⁎
Task Study
MPFC Activation increase Passive perception of craving-related stimuli Garavan et al. (2000)2; Grüsser et al. (2004)1;Heinz et al. (2004)1; Hermann et al. (2006)1;McBride et al. (2006)4
Monetary feedback Martin-Soelch et al. (2001a)5
Activation decrease Passive perception of craving-related stimuli Bonson et al. (2002)2
Passive viewing of sex-related stimuli Garavan et al. (2000)2
Decision-making in IGT Bolla et al. (2003)2
ACC Activation increase Passive perception of craving-related stimuli Brody et al. (2002)4; Brody et al. (2004)4;Garavan et al. (2000)2, Grüsser et al. (2004)1;McBride et al. (2006)4; Wrase et al. (2002)1
Cocaine-related imagery Kilts et al. (2004)2
Craving for cocaine self-administration Risinger et al. (2005)2
Active perception of emotional stimuli Wexler et al. (2001)2
Implicit perception of smoking-related stimuliin a target detection task
McClernon et al. (2005)4
Decision-making in IGT Adinoff et al. (2003)2
Mid cingulategyrus
Activation increase Passive perception of craving-related stimuli Hermann et al. (2006)1
Verbal feedback Martin-Soelch et al. (2001b)4
Activation decrease Craving for cocaine self-administration Risinger et al. (2005)2
PCC Activation increase Passive perception of craving-related stimuli Franklin et al. (2007)4; Garavan et al. (2000)2;McBride et al. (2006)4; Sell et al. (2000)5
Cocaine-related imagery Kilts et al. (2004)2
Interview about cocaine themes Wang et al. (1999)2
Activation decrease Passive viewing of sex-related stimuli Garavan et al. (2000)2
OFC Activation increase Passive perception of craving-related stimuli Bonson et al. (2002)2; Brody et al. (2002)4 ;Franklin et al. (2007)4; Hermann et al. (2006)1;Wrase et al. (2002)1; McBride et al. (2006)4;Sell et al. (2000)5
Craving for cocaine self-administration Risinger et al. (2005)2
Verbal feedback Martin-Soelch et al. (2001a)5
Decision-making in IGT Bolla et al. (2003)2
Activation decrease Reward evaluation Goldstein et al. (2007a)2;Goldstein et al. (2007b)2
Verbal feedback Martin-Soelch et al. (2001b)4
Decision-making in IGT Bolla et al. (2005)3
Cambridge Risk Task Ersche et al. (2006)5
LPFC6 Activation increase Passive perception of craving-related stimuli Bonson et al. (2002)2; Brody et al. (2002)4;Franklin et al. (2007)4; Garavan et al. (2000)2;Hermann et al. (2006)1; McBride et al. (2006)4 ;Sell et al. (2000)5; Wrase et al. (2002)1
Craving for cocaine self-administration Risinger et al. (2005)2
Implicit perception of smoking-related stimuliin a target detection task
McClernon et al. (2005)4
Reward evaluation Goldstein et al. (2007b)2
Monetary reward Martin-Soelch et al. (2001a)5
Verbal feedback Martin-Soelch et al. (2001a)5
Anticipation of monetary loss Wrase et al. (2007b)1
Decision-making in IGT Adinoff et al. (2003)2
Activation decrease Passive perception of craving-related stimuli Bonson et al. (2002)2
Passive viewing of sex-related stimuli Garavan et al. (2000)2
Reward evaluation Goldstein et al. (2007a)2
Verbal feedback Martin-Soelch et al. (2001b)2
Decision-making in IGT Bolla et al. (2003)2;Bolla et al. (2005)3
Amygdala Activation increase Passive perception of craving-related stimuli Bonson et al. (2002)2; Franklin et al. (2007)4;Schneider et al. (2001)1
Activation decrease Reward evaluation Goldstein et al. (2007b)2
Midbrain Activation increase Verbal feedback Martin-Soelch et al. (2001b)4
Striatum7 Activation increase Passive perception of craving-related stimuli Franklin et al. (2007)4; Garavan et al. (2000)2;Grüsser et al. (2004)1; Heinz et al. (2004)1;McBride et al. (2006)4; Wrase et al. (2002)1
170 B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4

Table 1 (continued)
Region Direction ofactivation⁎
Task Study
Striatum7 Activation increase Cocaine-related imagery Kilts et al. (2004)2
Craving for cocaine-self-administration Risinger et al. (2005)2
Implicit perception of smoking-related stimuliin a target detection task
McClernon et al. (2005)4
Decision-making in IGT Bolla et al. (2003)2
Activation decrease Anticipation of monetary gain Wrase et al. (2007b)1
Anticipation of monetary loss Wrase et al. (2007b)1
⁎Either in the between-group comparison (versus controls) or within group in comparison with neutral stimuli; 1alcohol; 2cocaine; 3marijuana;4nicotine; 5opiates; 6including DLPFC, VLPFC and APFC; 7including putamen, nucleus caudatus, pallidum and nucleus accumbens.List of abbreviations; ACC — anterior cingulate cortex; APFC — anterior prefrontal cortex; DLPFC — dorsolateral prefrontal cortex; IGT —Iowa gambling task; LPFC — lateral prefrontal cortex; MPFC — medial prefrontal cortex; OFC — orbitofrontal cortex; PCC — posteriorcingulate cortex; VLPFC — ventrolateral prefrontal cortex.
171B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4
and striatum as well as of sensory processing regions duringprocessing of monetary feedback (in opiate abusers andhealthy smokers: Martin-Soelch et al., 2001a,b; in abstinentcocaine abusers: Goldstein et al., 2007a). In one study abs-tinent alcohol-dependent subjects further exhibited aninverse correlation between craving severity during anticipa-tion of monetary gain and activation in the ventral striatum(Wrase et al., 2007b). In addition, sex-related stimuli were alsoassociated with a reduced responsiveness in the brain'sreward circuit in abstinent cocaine abusers when comparedto responses to cocaine-related stimulation or responsesobserved in healthy controls, despite similar ratings in bothgroups, which ruled out that decreases in brain activationswere simply a function of decreased task engagement(Garavan et al., 2000). However, especially monetary rewardmay not be the best stimulus to test reward-sensitivity outsideof the drug context, because money may as well represent adrug-related cue that has the propensity to elicit craving,which should be expected to also impair the ability to correctlyevaluate varying amounts. In fact, in one study there wasevidence that the ability to correctly evaluate different rewardamounts negatively correlated with the degree of craving-likeresponses in OFC and amygdala during evaluation of differentmonetary amounts (Goldstein et al., 2007b). This observationfurther complicates the picture of the underlying pathophy-siology of addiction as it underlines that in some patientpopulations increased activation within reward-sensitiveregions are not necessarily indicative of a preserved rewardprocessing ability when dealing with monetary gain.
There is evidence that deficits in reward-sensitive regionswere also related to impulsive choice and temporal discount-ing of the future in patients with substance abuse disorders,particularly when tested with drug stimuli (Coffey et al., 2003)or in a drug-deprived context (Giordano et al., 2002). Forinstance, Bechara and colleagues (2002) reported a significantdecrease in IGT performance in substance abusers similar tothe one observed in patients with orbitofrontal and dorso-lateral frontal damage (e.g., Fellows and Farah, 2005), whichwas accompanied by decreased activation in both right lateralOFC and DLPFC in one study (in abstinent marijuana abusers;Bolla et al., 2005). Patients who did not show such a clearbehavioral deficit instead displayed an increased activation in
the right anterior OFC, which positively correlated with taskperformance and thus could have represented a counter-vailing response that compensated for reduced activation inboth right DLPFC and left MPFC (abstinent cocaine abusers;Bolla et al., 2003; see also Table 1 and Supplementary Table 3).Performance on this task was further found to be positivelycorrelated with DLPFC and ACC metabolism at rest (Adinoff etal., 2003). It therefore seems to be highly likely that adisturbed balance between activation in limbic regions,favoring immediate gratification, and frontal regions, inte-grating future-oriented deliberation into behavioral choice,may have caused the observed decision-making deficitswhereby a pathologically increased impulsivity may haveled to a preference for immediate monetary gratificationregardless of detrimental long-term effects.
7.1. Regional distribution of activation associated withdrug-related stimulation in addiction—Activation LikelihoodEstimation
The results from the ALE meta-analysis widely supported theresults of our systematic literature review, which had alreadyindicated that the overvaluing of drug-related stimuli with thepropensity to elicit craving is associated with activation in abrain circuit that comprises orbitofrontal, limbic (i.e. cingulatecortex and amygdala) as well as striatal subregions (see Table4A; Fig. 3A). The highest ALE valuewithin regions of the brain'sreward system was thereby observed in right ACC with acluster extent of 3720mm3. In addition, we also found clustersin the left striatum, in the amygdalae, the left OFC and ACC aswell as in the left PCC. Outside of the reward system, themeta-analysis yielded a cluster in the left precuneus with thehighest ALE value of all clusters and a cluster extent of1552mm3. In addition, another 4 clusters were observed thatappeared in the superior temporal gyrus bilaterally, the leftmiddle temporal gyrus and the right superior frontal gyrus.
7.2. Substance (ab)use disorders—Conclusions andoutstanding questions
Drug addiction appears to affect a wide variety ofmotivationaland decision-making processes of every day life. The best
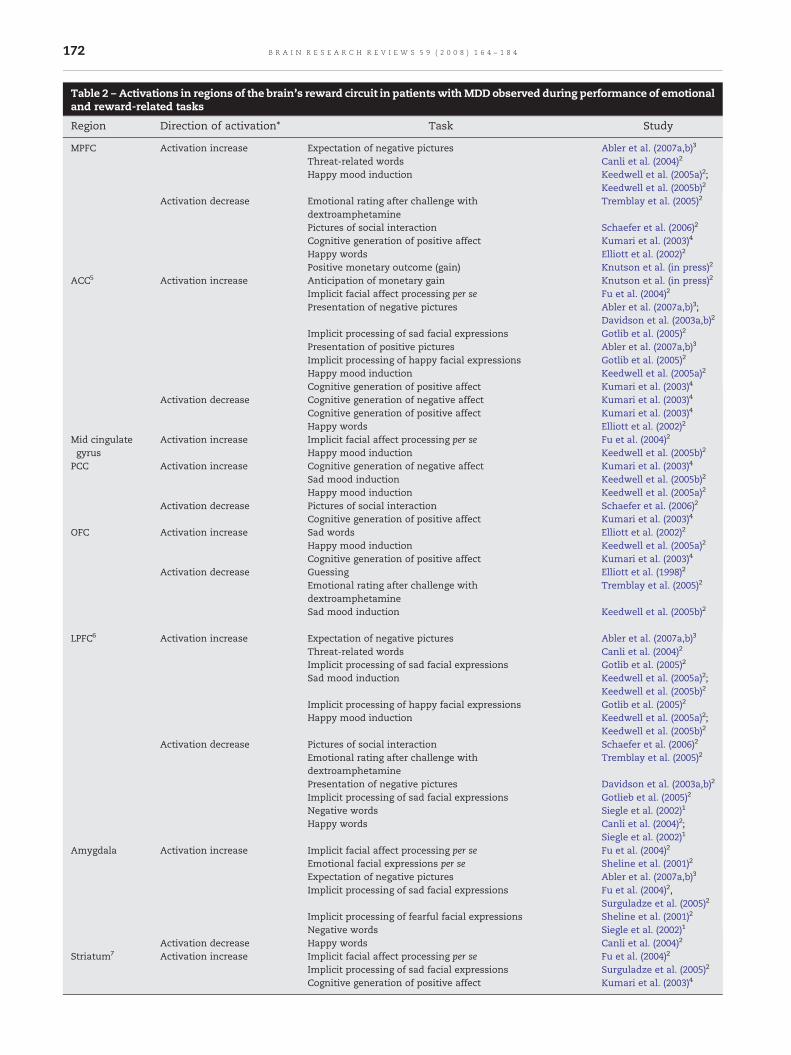
Table 2 – Activations in regions of the brain's reward circuit in patientswithMDDobserved during performance of emotionaland reward-related tasks
Region Direction of activation⁎ Task Study
MPFC Activation increase Expectation of negative pictures Abler et al. (2007a,b)3
Threat-related words Canli et al. (2004)2
Happy mood induction Keedwell et al. (2005a)2;Keedwell et al. (2005b)2
Activation decrease Emotional rating after challenge withdextroamphetamine
Tremblay et al. (2005)2
Pictures of social interaction Schaefer et al. (2006)2
Cognitive generation of positive affect Kumari et al. (2003)4
Happy words Elliott et al. (2002)2
Positive monetary outcome (gain) Knutson et al. (in press)2
ACC5 Activation increase Anticipation of monetary gain Knutson et al. (in press)2
Implicit facial affect processing per se Fu et al. (2004)2
Presentation of negative pictures Abler et al. (2007a,b)3;Davidson et al. (2003a,b)2
Implicit processing of sad facial expressions Gotlib et al. (2005)2
Presentation of positive pictures Abler et al. (2007a,b)3
Implicit processing of happy facial expressions Gotlib et al. (2005)2
Happy mood induction Keedwell et al. (2005a)2
Cognitive generation of positive affect Kumari et al. (2003)4
Activation decrease Cognitive generation of negative affect Kumari et al. (2003)4
Cognitive generation of positive affect Kumari et al. (2003)4
Happy words Elliott et al. (2002)2
Mid cingulategyrus
Activation increase Implicit facial affect processing per se Fu et al. (2004)2
Happy mood induction Keedwell et al. (2005b)2
PCC Activation increase Cognitive generation of negative affect Kumari et al. (2003)4
Sad mood induction Keedwell et al. (2005b)2
Happy mood induction Keedwell et al. (2005a)2
Activation decrease Pictures of social interaction Schaefer et al. (2006)2
Cognitive generation of positive affect Kumari et al. (2003)4
OFC Activation increase Sad words Elliott et al. (2002)2
Happy mood induction Keedwell et al. (2005a)2
Cognitive generation of positive affect Kumari et al. (2003)4
Activation decrease Guessing Elliott et al. (1998)2
Emotional rating after challenge withdextroamphetamine
Tremblay et al. (2005)2
Sad mood induction Keedwell et al. (2005b)2
LPFC6 Activation increase Expectation of negative pictures Abler et al. (2007a,b)3
Threat-related words Canli et al. (2004)2
Implicit processing of sad facial expressions Gotlib et al. (2005)2
Sad mood induction Keedwell et al. (2005a)2;Keedwell et al. (2005b)2
Implicit processing of happy facial expressions Gotlib et al. (2005)2
Happy mood induction Keedwell et al. (2005a)2;Keedwell et al. (2005b)2
Activation decrease Pictures of social interaction Schaefer et al. (2006)2
Emotional rating after challenge withdextroamphetamine
Tremblay et al. (2005)2
Presentation of negative pictures Davidson et al. (2003a,b)2
Implicit processing of sad facial expressions Gotlieb et al. (2005)2
Negative words Siegle et al. (2002)1
Happy words Canli et al. (2004)2;Siegle et al. (2002)1
Amygdala Activation increase Implicit facial affect processing per se Fu et al. (2004)2
Emotional facial expressions per se Sheline et al. (2001)2
Expectation of negative pictures Abler et al. (2007a,b)3
Implicit processing of sad facial expressions Fu et al. (2004)2,Surguladze et al. (2005)2
Implicit processing of fearful facial expressions Sheline et al. (2001)2
Negative words Siegle et al. (2002)1
Activation decrease Happy words Canli et al. (2004)2
Striatum7 Activation increase Implicit facial affect processing per se Fu et al. (2004)2
Implicit processing of sad facial expressions Surguladze et al. (2005)2
Cognitive generation of positive affect Kumari et al. (2003)4
172 B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4

Table 2 (continued)
Region Direction of activation⁎ Task Study
Striatum7 Activation decrease Feedback per se Elliott et al. (1998)2
Positive monetary outcome (gain) Knutson et al. (in press)2
Pictures of social interaction Schaefer et al. (2006)2
Emotional rating after challenge with dextroamphetamine Tremblay et al. (2005)2
Positive words Epstein et al. (2006)2
Implicit processing of positive facial expressions Surguladze et al. (2005)2
⁎Either in the between-group comparison (versus controls) or within group in comparison with neutral stimuli; 1remission; 2depressive state;3sub- or postacute; 4treatment-resistant depression; 5including the subgenual cingulate cortex; 6including DLPFC, VLPFC and APFC; 7includingputamen, nucleus caudatus, pallidum and nucleus accumbens.List of abbreviations; ACC — anterior cingulate cortex; APFC — anterior prefrontal cortex; DLPFC — dorsolateral prefrontal cortex;IGT — Iowa gambling task; LPFC — lateral prefrontal cortex; MDD — major depression; MPFC — medial prefrontal cortex; OFC —orbitofrontal cortex; PCC — posterior cingulate cortex; VLPFC — ventrolateral prefrontal cortex.
173B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4
replicated finding is that patients showed a motivational biastowards the drug-of-abuse and related stimuli, which wasparticularly evident in increased responses for drug-relatedstimulation in theACC, the ‘relevancedetector’OFC, but also inthe amygdala and other regions of the brain's (dopaminergic)reward system, which could be inferred from both thesystematic literature review and the results of the ALE-meta-analysis. Further, a reduced neural sensitivity to drug-unre-lated reward could also be observed in some patients, whichmay in part be a result of a shift of activation towards the drugof abuse at the expense of other drug-unrelated rewards.Alternatively, diminished responses in reward-sensitiveregions could have also occurred as a function of successfulsuppression of craving-related responses when patients triedto correctly evaluate different amounts of monetary rewards(Goldstein et al., 2007b). Future research needs to morecarefully test the behavioral and neural sensitivity to drug-unrelated reward by also including primary reinforcers (e.g.,food) that have no potential to elicit craving, and compare it toactivation induced by drug-related stimulation.
Nevertheless, similar neural responses to drug-related (andpartly also to drug-unrelated) stimuli in addiction for varioustypes of substances (see Fig. 1) and high ALE values in reward-sensitive regions,which reflected strong concordance betweenstudies, indicate that especially the neurobiological mechan-isms underlying the processing of drug-related cues thattrigger craving or relapse may recur on similar neuralsubstrates independent of the drug of abuse. In fact, the neuralmechanisms that accompany the acquisition of drug use andits maintenance have been assumed to be similar for differentdrug classes. Reward circuits including the ventral striatumand the amygdala may thereby be crucial for the initiation ofdrug self-administration (see Volkow and Fowler, 2000). Bycontrast, the compulsive aspects of drug seeking have beenassumed to be secondary to intermittent drug-induced dopa-minergic activation and an overall decrease in dopaminergicfunction that affects other regions like the orbitofrontal andcingulate cortices. This disrupts adaptive incentive processingby eventually leading to compulsive drug behaviors thatpersist even in the presence of adverse reactions (see Kalivaset al., 2006; Volkow et al., 2003, 2007). As drug seeking becomeswell established there is also evidence for a progressive shiftfrom motivated seeking of goals dependent on the ventralstriatum to stimulus–response habits mediated by the dorsal
striatum (see also Hyman et al., 2006). It should therefore bepromising to further investigate similarities betweenmechan-ismsmediating different aspects of substance abuse and thoseinvolved in other “addiction-like” disorders that have similarlyshown to be associated with a biased response to a certaincategory of motivationally significant events in the environ-ment. Behavioral and substance addictions share commoncore clinical features and also exhibit phenomenological,genetic and neurobiological links that relate to the impulsiveas well as the compulsive aspects of addiction (see Grant et al.,2006). Evidence for a correspondence between neural deficitsobserved in substance abuse and pathological gamblingassociated with a deficient ventral frontal and striatalresponse to rewarding stimuli already exists (Reuter et al.,2005; see also Potenza et al. 2006). Further, pathologically obesesubjects also show similarities to substance abusers (e.g., areduction in striatal dopamineD2-receptors;Wang et al., 2004),which recently led to a redefinition of obesity as a substancedependence disorder (James et al., 2004; see also Trinko et al.,2007). Neuroimaging approaches that compare these motiva-tional disorders might be useful to further understand theneurophysiological basis of dependency from a more generalperspective and are expected to provide new directions fortreating the growing epidemic of eating-disorders. Also, theneural basis of reward-related processing in substance depen-dence for drugs that directly affect the dopamine system (e.g.,cocaine) compared to those with indirect effects (marijuana)remain to be further elucidated, as marijuana dependenceseemed to be underrepresented by current neuroimagingresearch, especially when regarding the frequency of mar-ijuana abuse in the population.
Another important question that cannot be answered bycurrent research is why some patients want to become cleanand others do not and whether these patient populationsdiffer in their affective processing ability. Although, it has alsobeen proposed by some researchers (e.g. Robinson andBerridge, 2003) that the motivational component of drugseeking behavior (“drug wanting”) may be completelydetached from affective processing, a reduction of hedonicdrug effects (i.e., the increasing tolerance towards the hedoniceffects of the drug and thus the reduction in feelings ofpleasure) may be another potential reason for an increasedmotivation to seek the effects of cocaine, which reverse thehedonic deficit (e.g., Ahmed, 2004). In humans, there are at

Table 3 – Activations in regions of the brain's reward circuit in patients with BD observed during performance of emotionaland reward-related tasks
Region Direction ofactivation⁎
Task Study
MPFC Activation increase Cognitive generation of positive affect Malhi et al. (2004a)3;Malhi et al. (2007)1
Implicit processing of mild happy facial expressions Lawrence et al. (2004)1,3
ACC Activation increase Decision-making/Risk-taking Rubinsztein et al. (2001)2
Happy distractor words Elliott et al. (2004)2
Implicit and explicit processing of happy facial expressions Chen et al., (2006)3
Cognitive generation of positive affect Malhi et al. (2007)1
Implicit processing of intense sad facial expressions Lawrence et al. (2004)1,3
Activation decrease Explicit facial affect processing per se Lennox et al. (2004)2
Mid cingulate gyrus Activation decrease Explicit facial affect processing per se Lennox et al. (2004)2
PCC Activation increase Explicit facial affect processing per se Lennox et al. (2004)2
Implicit and explicit processing of happy facial expressions Chen et al., (2006)3
Cognitive generation of negative affect Malhi et al. (2007)1
OFC Activation increase Happy distractor words Elliott et al. (2004)2
Activation decrease Implicit facial affect processing per se Altshuler et al. (2005)2
Implicit processing of mild sad facial expressions Lawrence et al. (2004)1,3
LPFC4 Activation increase Emotional words per se Elliott et al. (2004)2
Cognitive generation of affect per se Malhi et al. (2005)1
Happy distractor words Elliott et al. (2004)2
Cognitive generation of positive affect Malhi et al. (2004a3,b)2;Malhi et al. (2007)1
Implicit and explicit processing of happy facial expressions Chen et al., (2006)3
Implicit processing of mild happy expressions Lawrence et al. (2004)1,3
Sad distractor words Elliott et al. (2004)2
Implicit processing of sad expressions Lawrence et al. (2004)1,3
Cognitive generation of negative affect Malhi et al. (2004a3,b)2;Malhi et al. (2007)1
Implicit and explicit processing of fearful facial expressions Lawrence et al. (2004)1,3
Activation decrease Emotional words per se Elliott et al. (2004)2
Decision-making/Risk-taking Rubinsztein et al. (2001)2
Implicit and explicit processing of (mild) fearful facialexpressions
Yurgelun-Todd et al. (2000)1;Lawrence et al. (2004)1,3
Implicit processing of sad facial expressions Lawrence et al. (2004)1,3
Emotional prosody Mitchell et al. (2004)1
Midbrain Activation increase Implicit and explicit processing of fearful facial expressions Chen et al., (2006)2,3
Activation decrease Implicit processing of intense happy facial expressions Lawrence et al. (2004)1,3
Amygdala Activation increase Implicit facial affect processing per se Altshuler et al. (2005)2
Cognitive generation of positive affect Malhi et al. (2004a)3
Implicit processing of mild happy facial expressions Lawrence et al. (2004)1,3
Implicit and explicit processing of (intense) fearful facialexpressions
Yurgelun-Todd et al. (2000)1;Lawrence et al. (2004)1,3
Activation decrease Implicit processing of happy facial expressions Lawrence et al. (2004)1,3
Implicit processing of mild fearful facial expressions Lawrence et al. (2004)1,3
Emotional prosody Mitchell et al. (2004)1
Striatum5 Activation increase Explicit facial affect processing per se Lennox et al. (2004)2
Implicit and explicit processing of happy faces Chen et al., (2006)3;Lawrence et al. (2004)1,3
Cognitive generation of negative affect Malhi et al. (2004b)2
Implicit and explicit processing of fearful facialexpressions
Chen et al., (2006)2,3;Lawrence et al. (2004)1,3
Activation decrease Implicit processing of mild sad facial expressions Lawrence et al. (2004)1,3
Reward anticipation and receipt of monetary reward Abler et al. (2007b)2
⁎Either in the between-group comparison (versus controls) or within group in comparison with neutral stimuli;1mixed or euthymic; 2(hypo-)manic; 3depressive; 4including DLPFC, VLPFC and APFC; 5including putamen, nucleus caudatus, pallidum and nucleus accumbens.List of abbreviations; ACC — anterior cingulate cortex; APFC — anterior prefrontal cortex; BD — bipolar disorder; DLPFC — dorsolateralprefrontal cortex; LPFC — lateral prefrontal cortex; MPFC — medial prefrontal cortex; OFC — orbitofrontal cortex; PCC — posteriorcingulate cortex; VLPFC — ventrolateral prefrontal cortex.
174 B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4
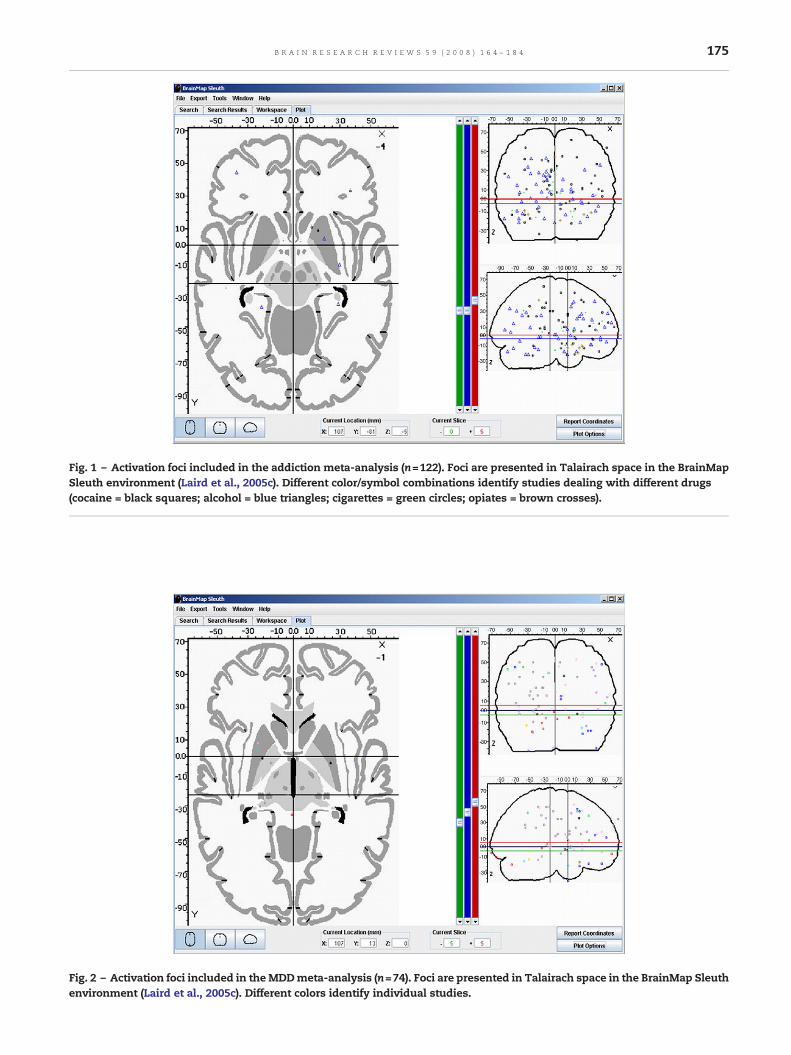
Fig. 1 – Activation foci included in the addiction meta-analysis (n=122). Foci are presented in Talairach space in the BrainMapSleuth environment (Laird et al., 2005c). Different color/symbol combinations identify studies dealing with different drugs(cocaine = black squares; alcohol = blue triangles; cigarettes = green circles; opiates = brown crosses).
Fig. 2 – Activation foci included in theMDDmeta-analysis (n=74). Foci are presented in Talairach space in the BrainMap Sleuthenvironment (Laird et al., 2005c). Different colors identify individual studies.
175B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4

176 B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4
least some indications for a physiologic link between cocaine-cue responses and normal dysphoric states, as craving-relatedactivations in one study resembled responses to sad stimulinormally observed in healthy controls (Wexler et al., 2001).Further, during pharmacological challenge, the magnitude ofdopamine release in the anterior ventral striatum, but not inthe dorsal caudate, was positively correlated with the hedonicresponse (euphoria) to dextroamphetamine (Drevets et al.,2001), which implied that stimulant-associated “drug want-ing” may not be entirely uncoupled from a positive affectiveexperience of “hedonic liking” and may be particularlyimportant for initiation of drug self-administration (see alsoVolkow and Fowler, 2000; Grant et al., 2006). Patients withcomorbid affective disorders should thus be of high interestfor future research. In fact, some psychiatric symptoms mayalso be a cause for comorbid chronic drug abuse (e.g., drug-taking as self-medication in depression) and chronic drugabuse may in term exacerbate the symptoms of other pre-existing mental disorders (Bruijnzeel et al., 2004; Currie et al.,2005). It thus remains to be tested how comorbid depressionalters the neural response to different kinds of rewards andwhether this response is similar to the one observed indepression per se or in addiction without depressive symp-toms or whether it is rather unique. Knowledge about thoseinteractions may help to identify disorder-specific neural traitmarkers for addiction or may alternatively lead to a redefini-tion of disorders (e.g., through an introduction of an ‘addic-tion-subtype' in schizophrenia; Mancini-Marïe et al., 2006),which could even help to guide subtype-specific treatmentinterventions in the future.
Table 4 – Results of the ALEmeta-analyses of hyperaction relatenegative stimuli in MDD
Anatomical region Brodmann area
(A) Regional concordance in hyperactivation related to stimulation by drug-cuesR ACC 32L/R striatum subcorticalL amygdala subcorticalR amygdala subcortical
L OFC 47L PCC 23L ACC 24L precuneus 31L superior temporal gyrus 38R superior temporal gyrus 41L middle temporal gyrus 21R superior frontal gyrus 10
(B) Regional concordance in hyperactivation related to negative stimuli in MDDL striatum subcortical
R pregenual ACC 32L subthalamic nucleus subcorticalL thalamus subcorticalL insula 13L middle frontal gyrus 8L inferior parietal lobe 40L inferior temporal cortex/uncus 20R lingual gyrus 17
8. Mood disorders—Changes in motivationalrelevance and affective processing
Mood disorders likemajor depressive and bipolar disorder (BD)have been characterized by significant changes in bothmotivational and affective processing (see Davidson et al.,2003a,b; Leppänen, 2006). Dominated by persistent dysphoricemotions and thoughts (e.g., anhedonia) – even in the absenceof acute stressors – depressive patients have been reported toexhibit a decreasedmotivation to seek and a reduced ability toexperience reward (Drevets, 2001; Chau et al., 2004). Incontrast, episodes of mania in patients with BD have beentypically characterized by elated or irritable mood, disinhibi-tion and a compromised judgment leading to damaging futureconsequences (Malhi et al., 2004a,b). On the behavioral level,the most prominent features of both MDD and BD observed inexperimental studies on emotional processing were (1) anemotional bias (Leppänen et al., 2004), (2) an impaired re-cognition accuracy in recognizing emotional expressions (Getzet al., 2003; Surguladze et al., 2004), and (3) an enhancedmemory for negative valent stimuli in the depressive state(Moritz et al., 2005; Leppänen, 2006).
In accordance with behavioral findings, patients withmood disorders showed significant alterations in brainactivity ‘at rest’ that mainly affected the brain's rewardsystem including the amygdala, regions of the mesolimbicdopamine system and frontal cortices (see Drevets, 1999;Davidson et al., 2002 for comprehensive reviews). In addition,regions that commonly processed stimuli with positiveemotional content in healthy human subjects (e.g., Elliott et
d to either stimulation by drug-related cues in addiction or to
Coordinates ALE Volume (mm3)
in addiction6 44 −4 0.0135 3720−6 8 6 0.0125 4616−20–2 −12 0.0118 56020–6 −12 0.0093 42430 0 −20 0.0076 200−32 18 −16 0.0093 368−6–38 24 0.0090 392−8 22 22 0.0079 320−10–58 30 0.0159 1552−56 12 −20 0.0133 51248–34 8 0.0106 952−36–2 −22 0.0088 56818 52 14 0.0099 392
−28 2 16 0.0132 1136−10–4 16 0.0131 1008−20 16 16 0.0067 1126 38 16 0.0122 1224−12–10 −8 0.0141 2728−20–30 2 0.0109 608−40 0 −2 0.0097 840−44 14 44 0.0071 336−26–40 50 0.0067 248−30 0 −42 0.0067 14416–102 −8 0.0066 112

Fig. 3 – ALE maps for cue-related hyperactivation inaddiction and hyperactivation related to negative stimulationin MDD at p<0.05. (A) Addiction: in the ALE meta-analysissignificant activation likelihood was seen in the right ACC, inthe striatum (right putamen, right nucleus accumbens andleft ventral caudate), in the amygdalae and in the leftposterior OFC. (B) MDD: the ALE meta-analysis revealedsignificant activation likelihood in the right pregenual ACC,somewhat above the cluster observed in addiction, in the leftlateral striatum (putamen), but neither in the amygdalae norin the posterior OFC.
177B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4
al., 2002; Surguladze et al., 2005) also showed an affective biasof varying extent towards negative stimulation (e.g., sadexpressions). For instance, the ‘relevance detector' amygdalaexhibited a shift in activation in favor of mood-congruentaffective stimuli, which was particularly evident in thedepressive or untreated state of MDD (e.g., Fu et al., 2004; seealso: Table 2 and Supplementary Table 4). In addition,increased responses during negative emotional stimulationhave also been observed in other reward-sensitive regions(e.g., the ACC), although not always consistently (Table 2).Differences in the emotional stimulation between studies (seebelow) and the possibility that different subtypes of depres-sion may exist on the neural level can supposedly account forthese inconsistencies. There is evidence that patients with atreatment-resistant form of MDD were characterized by an
overall reduced anterior cingulate responsiveness to emo-tional stimuli per se (Kumari et al., 2003), and to affectivelynegative stimulation (Davidson et al., 2003a,b) pointing to apossible subtype of MDD. In patients with a clear negative biasprior to treatment, treatment success was accompanied by asignificant attenuation of activation in the affected regions(e.g., Fu et al., 2004; Sheline et al., 2001) and also led to anincreased responsiveness to other emotional stimuli (Schaeferet al., 2006), which suggested that these alterations may haverepresented a disorder-related state measure in this subgroupof depressive patients.
At the same time, the neural responsiveness of reward-sensitive regions (e.g., of the ventral striatumor the amygdala)to positive valent emotional stimuli and monetary outcomewas found to be diminished inmany previous studies onMDD(e.g., Canli et al., 2004, Elliott et al., 2002; Knutson et al., inpress). It became obvious that the lacking emotional inputfrom the amygdala and associated regions supposedly alsocompromised sensory processing regions that similarlyexhibited a bias against positive stimuli (Surguladze et al.,2005; see also Vuilleumier and Pourtois, 2006). The bias there-by appeared to be directly associated to depression severity orthe degree of anhedonia (Keedwell et al., 2005a,b; Surguladzeet al., 2005), which again suggested that these alterations alsoreflected another state-marker of depression.
However, some studies also reported an increase inactivation to implicit processing of happy facial expressions(Gottlib et al., 2005) and pictures (Abler et al., 2007a,b), in thedirect comparison of happy and neutral autobiographicalprompts intended to induce a mood state (Keedwell et al.,2005a) or during anticipation of a potential monetary gain(Knutson et al., in press), when compared to activation inhealthy controls or activity evoked by neutral stimulation.These stimuli affected different parts of the regions that alsoresponded to negative stimulation (e.g., the anterior cingulatecortex and the amygdala; see also Table 2). Partly discrepantfindings may thereby be directly assigned to differentialsalience levels of the stimulusmaterial used. Onemay assumethat especially positive emotionalwords (e.g., Canli et al., 2004)were not salient enough to evoke an emotional response in thedepressive state, while emotional facial displays (Gottlib et al.,2005) with a closer relation to the underlying neural circuitry,outcome-uncertainty during anticipation (Knutson et al., inpress), or positive autobiographical prompts (Keedwell et al.,2005a,b) with a direct linkage to the individual's personalemotional experiences, instead may have been perceived ashighly salient and were therefore supposedly more deeplyprocessed leading to the observed increase in activation.
In contrast to patients with MDD, bipolar patients being inthe manic phase exhibited no such bias for negative valentstimuli, but either showed an impaired recognition of negativefacial affect, an increased responsiveness to positive stimuli orexhibited a general disability to process emotional stimuli atall, which was also expressed in the neural response pattern(cf., Table 3 and Supplementary Table 5). For example, patientsin the manic state exhibited a reduced sensitivity for reward-related and motivationally salient information, which wasaccompanied by a decreased behavioral sensitivity for differ-ences in reward value and a lack of relative activationalincrease in the nucleus accumbens during both reward

178 B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4
anticipation and receipt when compared to expectation andreceipt of no reward (Abler et al., 2007b). In addition, patientswith bipolar mania rated sad facial expressions as less intensethen healthy controls (e.g., Lennox et al., 2004). Concurrently,these patients showed an attenuation of activation in thesubgenual ACC and the bilateral amygdalae, which wasaccompanied by an increased activation in theposterior cingu-late and the posterior insula when compared to healthy con-trols, but at the same time they showed an unaltered neuralresponse to happy facial expressions (Lennox et al., 2004). Inanother study, evidence was found for a positive affectivebias in manic patients showing an enhanced ventral andmedial prefrontal response to positive emotional distractors(Elliott et al., 2004). The same patients, however, lacked adifferential response to happy and sad targets, which may beassigned to both a general bias in emotional processing and aheightened distractibility observed during mania that couldhave impaired processing of explicit targets and may haveinstead increased responses to emotional distractors (Elliottet al., 2004). Considering that these effects were mediated bydistinct functional regions when compared to patients withMDDperforming the same task as indicatedby the results from3 previous studies (Elliott et al., 2002; 2004; Lawrence et al.,2004), the pathophysiological basis of altered processing ofemotional stimuli appears to differ in these two disorders. Inthe depressive and euthymic state of BD, patients partlyshowed unique responses to both negative and positive facialexpressions when compared to MDD patients (Lawrence et al.,2004), although overlapping changes in activation patternscould also be observed (e.g., the hippocampal response tomildsadness correlated with depression severity; Lawrence et al.,2004). This led to the question on whether some alterationsoccurring in both MDD and BD were actually trait markers ofthe disease and therefore independent of mood state. Inpatients with severe MDD there is already evidence for state-dependency of some neural deficits, in that for instance thehyperactivation of the amygdala and the striatum in responseto sad faces receded in the state of remission (Fu et al., 2004).Similarly, in bipolar patients hyperactivation of subcorticalregions in both depressed bipolar patients when processinghappy and fearful faces (Lawrence et al., 2004) and in hypo-manic patients in response to negative stimuli (Malhi et al.,2004b) supposedly represented a specific marker for BD in theacute state, which apparently disappeared in euthymicpatients (Malhi et al., 2005, 2007). In addition, some studieswere already able to show similar activationswhen comparingdifferent BD patients in the depressive and in the manic state(e.g., Chen et al., 2006), providing indications for functionaltrait markers in BD. Still, to date further evidence fromdifferent phases of BD observed in the same patients remainsto be acquired.
8.1. Regional distribution of activation associated withmood-congruent processing in MDD—Activation LikelihoodEstimation
The results from the ALEmeta-analysis indicated that the biastowards mood-congruent or negative stimulation may bemediated by a network that comprises parts of themesolimbicreward system (striatum and pregenual ACC), but also
includes regions involved in cognitive and sensory processing(see Table 4B; Fig. 3B). The highest ALE value in regions of thereward systemwas observed in the left striatumwith a clusterextent of 1136mm3 followed by one cluster in the rightpregenual ACC and 2 further clusters in the left striatum. Inaddition, we also observed clusters in the left thalamus andsubthalamic nucleus, the left insula adjacent to the posteriorOFC, the left middle frontal gyrus, left inferior parietal lobe,left inferior temporal cortex as well as the right lingual gyrus.These results in part confirm the observation from thesystematic review (Table 2) and indicate that some of theregions, which contribute in the attribution of motivationalsalience and goal-directed behavior in the healthy brain, maybe biased towards negative stimulation in MDD. In addition,attentional and sensory processing areas also showed con-cordant hyperactivation for negative stimulation in patients inthe depressive state. Surprisingly, the ALE meta-analysisfound no evidence for a dysfunction of the ‘relevance detector’amygdala during mood-congruent stimulation, albeit 3 stu-dies reported an increased amygdala response during negativestimulation (Fu et al., 2004; Surguladze et al., 2005; Abler et al.,2007a,b; see also Sheline et al., 2001 and Siegle et al., 2002, whoalso reported activation in the amygdala, but did not reportpeak coordinates). However, one has to be aware of the factthat the limited number of peaks included in the analysis(<100) as well as variations in depression severity of patientsassessed by different studies could have severely affected thepresent results. The meta-analytic results may thus onlyprovide initial evidence that in MDD the observable bias insalience attribution may in part be associated with distur-bances within the mesolimbic dopaminergic system, but mayalso be related to a dysfunctional attentional control system.Future research is definitely necessary to provide a broaderdata base that allows one to achieve more conclusive results.
8.2. Mood disorders—Conclusions and outstandingquestions
Mood disorders may be best characterized by an overalldisturbed ability to process motivationally relevant stimuliin a context-appropriate way (i.e., according to their ‘objective'affective valence). Depending on current mood state, somepatients exhibited a kind of ‘motivational blindness’ formood-incongruent stimuli that was associated with decreasedactivity in brain regions involved in emotional salienceattribution and sometimes even in regions of general stimulusprocessing, while mood-congruent stimulation in mostpatients led to enhanced activation in similar brain regions(see Table 4B). Important to note, it could be ruled out thatthese differences in neural response patterns were simply afunction of decreased effort or task engagement, as they wereboth evident during implicit and explicit perception ofemotional stimulation (especially facial expressions) andalso occurred in patients with normal behavioral performance(e.g., Epstein et al., 2006). It nevertheless remains to be furtherelucidated, how the observed neural deficits may affect thosestages of the decision-making process, that rely on unequi-vocal affective signals as guidance for adaptive behavioralchoice (e.g., reward-related reversals or the decision for theadvantageous deck in the IGT). Patients with BD have already

179B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4
been reported to show a lack of inhibitory control, supposedlydue to an insufficient integration of negative affective signalsinto behavioral choice leading to overall disadvantageousbehavior (Christodoulou et al., 2006). In addition, patients withMDD showed normal performance in the IGT, when advanta-geous decks provided immediate large punishment, and evenlarger future reward, but were significantly impaired in thenormal version of the IGT with low immediate rewardfollowed by even lower punishment in advantageous decks(Must et al., 2006). This observation indicated that depressiveswere supposedly only able to use reward feedback in anadaptive way, if it was (1.) considerable high, (2.) infrequentand (3.) also highly variable in value, which opens newperspectives for future research in depression. Accordingly,the impairment in processing of positive valent stimuli couldhave simply been a function of salience level and not of ageneral impairment in processing of positive or affectivestimuli per se, which means that depressives should suppo-sedly be still able to process positive stimuli, but only at highsalience levels.
9. General conclusion
Context-sensitive reward processing and adaptive decision-making require the intact interplay between various corticaland subcortical regions that form the brain's reward circuit.Disturbances within this network have been reported in all ofthe psychiatric disorders reviewed above, even in patientswithout overt behavioral deficits. Both patients with sub-stance dependence and mood disorders thereby showed amore or less consistent motivational bias towards certainstimulus categories (in mood disorders depending on currentmood state) that also affected the neural response in the‘relevance detectors' amygdala and OFC as well as in otherregions of the brain's reward system. In addiction, regions ofthe reward system that most consistently showed a motiva-tional bias towards the drug of abuse appeared to be the ACC,the striatum, the amygdala and the OFC (Table 4A), while inMDD therewas evidence for concordant activity inmesolimbicsites of the dopaminergic system during processing of mood-congruent or negative stimuli across the studies reviewed(Table 4B). Still, when considering the results of the systematicreview of studies on MDD (Table 2) it cannot be ruled out thatother regions like the ‘relevance detector’ amygdala may alsobe important in a mechanism that leads to the observed biasin salience attribution during the depressive state.
Nevertheless, despite some similarities in the neuralresponses to “biased stimuli” (e.g., the observation ofincreased mesocorticolimbic responses to drug stimuli inaddiction or sad/negative stimulation in MDD; see also Fig. 3),the underlying neural mechanisms may be different. Inparticular studies that did not exclusively assess activationdifferences in patient populations, but also examined theunderlying neurotransmitter systems in humans providefurther evidence on pathophysiological (and maybe evenetiological) differences between these disorders. In addiction,most drugs have shown to trigger a phasic increase indopamine release in the “wanting” system of the brain. Ithas been proposed that this increase may produce a false,
fixed reward prediction error in dopamine neurons (Redish etal., 2004), which leads to a sensitization of certain drug effects,by increasing the relative reward value of drugs and drug-related stimuli through a simple Pavlovian stimulus–stimulusassociation learning in the OFC and associated areas like thenucleus accumbens (Robinson and Berridge, 2003). In combi-nation with a significant reduction of dopamine receptordensity or lowered presynaptic dopamine production andrelease (e.g., Heinz et al., 2004; Chang and Haning, 2006), thiscould significantly contribute to the overvaluing of drug-related stimuli, leading to a persistence of neural ‘craving-responses' even in the state of abstinence and may thus alsoimpair the ability to adequately respond to drug-unrelatedreward (Wrase et al., 2007b). Conversely, inmooddisorders, themotivational bias appears to depend on current mood state infavor of mood-congruent stimulation, and there are only fewindications for neural deficits that persisted over differentstates. The underlying mechanism leading to this bias may atleast in part be assigned to external stressors leading toincreases in cortisol and norepinephrine levels affectingactivation of the amygdala and other reward-sensitive regions(Drevets, 2001). The loss of pleasure and motivational driveduring the state of depression, which may be partly related toan underlying dysfunction of regions sensitive to dopaminer-gic modulation (e.g. Willner et al., 2005; Nutt et al., 2007; seealso Willner, 2002), may further be assumed to lead to areduced responsiveness to salient stimuli in general, whichwould favor those stimuli that have a per se higher saliencelevel and importance due to their direct relevance for theorganism's survival, namely fear- or threat-related cues,especially when displayed in human facial expressions. Inhealthy humans, the extent of dopamine transmission hasshown to bemaximal during processing of salient reward (Zaldet al., 2004), and activations in regions like the striatumincreased with rising salience, even outside of the rewardcontext (Zink et al., 2006). Converselyduring the state ofmania,it may be possible that the assumed hyperdopaminergic statecould have increased the ‘noise’ in the reward system, whichcould lead to an overall inability to focus on relevant emotionalevents at all (see also Juckel et al., 2006, who found evidence forsuch a process taking place in never-medicated schizophrenics).A combination of functional neuroimagingmethods (fMRI andPET) and pharmacological challenge of selective neurotrans-mitter systems may thus represent one promising route forfuture neuropsychiatric research, as indicated by recentfindings in healthy controls (e.g., Pessiglione et al., 2006) andin patients with MDD (Tremblay et al. 2005).
In the direct comparison of disorders, it will further benecessary to unify experimental approaches to achieve abetter discriminatory power regarding disorder-specificalterations in neurophysiologic processes. This means thatfuture research will certainly gain from the examination ofdifferent patient populations with similar and not onlydisease-specific experimental paradigms and will thus allowa more fundamental understanding of the underlying patho-physiology. So far most neuroimaging studies in substanceabuse presented either drug-related and neutral stimuli inpassive viewing tasks or used monetary reward and thusmainly assessed responses during the state of craving or theactual reception of positive feedback contingent on a manual

180 B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4
response. Conversely, in mood disorders subjects ratherperformed emotional evaluation tasks, which allowed directcomparisons between neural responses to happy and sadstimulation. Emotional content was thereby often conveyedthrough facial displays, which makes it possible that thepathophysiological role of the amygdala in depression hassimply been overestimated, when compared to other dis-orders, because also in healthy humans this region plays asuperior role in processing of emotional faces (Vuilleumierand Pourtois, 2006). The future use of similar, if not identicalbehavioral paradigms from fundamental research may havethe overall advantage that it allows a clear differentiation ofdisorder-specific aberrations in brain function.
10. Limitations of ALE
ALE provides a useful tool that has many advantages overlabel-based reviews (see Laird et al., 2005b for a detaileddiscussion), but nevertheless suffers from some potentiallimitations that have not been overcome yet. For instance, ALEneither provides a weighting factor for the number of subjectsin each study nor for the different voxel heights (z-values) andspatial extents of clusters in the studies included as data basis.In addition, thismethod does not account for the proportion ofstudies that activate in a certain region, but instead looks forcorrespondence in the clustering of peaks regardless ofwhether a certain cluster comprises only foci from a singlestudy (Wager et al., 2007). We have tried to overcome thisproblem by selecting only peaks with the highest statisticalsignificance from a subset of neighboring peaks and byrestricting the analysis to one relevant contrast per study(see above). Still, one cannot rule out that for example the 27foci from the study by Fu et al. (2004) may have contributeddisproportionally to the results of the ALE meta-analysis onMDD, albeit reported peaks in that particular study weredistributed across the entire brain. Different meta-analyticapproaches will always suffer from one or the other limitation(see Laird et al., 2005a; Wager et al., 2007) and thus meta-analytic results always need to be treated with care.
Appendix A. Supplementary data
Supplementary data associated with this article can be found,in the online version, at doi:10.1016/j.brainresrev.2008.07.004.
R E F E R E N C E S
Abler, B., Walter, H., Erk, S., Kammerer, H., Spitzer, M., 2006.Prediction error as a linear function of reward probability iscoded in human nucleus accumbens. Neuroimage 31, 790–795.
Abler, B., Erk, S., Herwig, U., Walter, H., 2007a. Anticipation ofaversive stimuli activates extended amygdala in unipolardepression. J. Psychiatr. Res. 41, 511–522.
Abler, B., Greenhouse, I., Ongur, D., Walter, H., Heckers, S., 2007b.Abnormal Reward System Activation in Mania.Neuropsychopharmacology (Electronic publication ahead ofprint).
Adinoff, B., Devous, M.D., Cooper, D.B., Best, S.E., Chandler, P.,Harris, T., Cervin, C.A., Cullum, C.M., 2003. Resting regionalcerebral blood flow and gambling task performance incocaine-dependent subjects and healthy comparison subjects.Am. J. Psychiatr. 160, 1892–1894.
Ahmed, S.H., 2004. Addiction as compulsive reward prediction.Science 306, 1901–1902.
Altshuler, L., Bookheimer, S., Proenza, M.A., Townsend, J., Sabb, F.,Firestine, A., Bartzokis, G., Mintz, J., Mazziotta, J., Cohen, M.S.,2005. Increased amygdala activation during mania: afunctionalmagnetic resonance imaging study. Am. J. Psychiatr.162, 1211–1213.
Arana, F.S., Parkinson, J.A., Hinton, E., Holland, A.J., Owen, A.M.,Roberts, A.C., 2003. Dissociable contributions of the humanamygdala and orbitofrontal cortex to incentive motivation andgoal selection. J. Neurosci. 23, 9632–9638.
Balleine, B.W., Delgado, M.R., Hikosaka, O., 2007. The role of thedorsal striatum in reward and decision-making. J. Neurosci. 27,8161–8165.
Bechara, A., Tranel, D., Damasco, H., Damasco, A.R., 1996. Failureto respond autonomically to anticipated future outcomesfollowing damage to prefrontal cortex. Cerebral Cortex 6,215–225.
Bechara, A., Dolan, S., Hindes, A., 2002. Decision-making andaddiction (part II): myopia for the future or hypersensitivity toreward? Neuropsychologia 40, 1690–1705.
Berns, G.S., McClure, S.M., Pagnoni, G., Montague, P.R., 2001.Predictability modulates human brain response to reward.J. Neurosci. 21, 2793–2798.
Berridge, K.C., Robinson, T.E., 2003. Parsing reward. Trends inNeurosciences 26, 507–513.
Bolla, K.I., Eldreth, D.A., London, E.D., Kiehl, K.A., Mouratidis, M.,Contoreggi, C., Matochik, J.A., Kurian, V., Cadet, J.L., Kimes, A.S.,Funderburk, F.R., Ernst, M., 2003. Orbitofrontal cortexdysfunction in abstinent cocaine abusers performing adecision-making task. Neuroimage 19, 1085–1094.
Bolla, K.I., Eldreth, D.A., Matochik, J.A., Cadet, J.L., 2005. Neuralsubstrates of faulty decision-making in abstinent marijuanausers. Neuroimage 26, 480–492.
Bonson, K.R., Grant, S.J., Contoreggi, C.S., Links, J.M., Metcalfe, J.,Weyl, H.L., Kurian, V., Ernst, M., London, E.D., 2002. Neuralsystems and cue-induced cocaine craving.Neuropsychopharmacology 26, 376–386.
Breiter, H.C., Aharon, I., Kahneman, D., Dale, A., Shizgal, P., 2001.Functional imaging of neural responses to expectancy andexperience of monetary gains and losses. Neuron 30,619–639.
Brody, A.L., Mandelkern, M.A., London, E.D., Childress, A.R., Lee,G.S., Bota, R.G., Ho, M.L., Saxena, S., Baxter, L.R., Madsen, D.,Jarvik, M.E., 2002. Brain metabolic changes during cigarettecraving. Arch. Gen. Psychiatry 59, 1162–1172.
Brody, A.L., Mandelkern, M.A., Lee, G., Smith, E., Sadeghi, M.,Saxena, S., Jarvik, M.E., London, E.D., 2004. Attenuation ofcue-induced cigarette craving and anterior cingulate cortexactivation in bupropion-treated smokers: a preliminary study.Psychiatry Res. 130, 269–281.
Bruijnzeel, A.W., Repetto, M., Gold, M.S., 2004. Neurobiologicalmechanisms in addictive and psychiatric disorders. ThePsychiatric clinics of North America 27, 661–674.
Bush, G., Vogt, B.A., Holmes, J., Dale, A.M., Greve, D., Jenike, M.A.,Rosen, B.R., 2002. Dorsal anterior cingulate cortex: a role inreward-based decision making. Proc. Natl. Acad. Sci., U S A 99,523–528.
Calder, A.J., Lawrence, A.D., Young, A.W., 2001. Neuropsychologyof fear and loathing. Nat. Rev. Neurosci. 2, 352–363.
Canli, T., Sivers, H., Thomason, M.E., Whitfield-Gabrieli, S.,Gabrieli, J.D., Gotlib, I.H., 2004. Brain activation to emotionalwords in depressed vs healthy subjects. Neuroreport 15,2585–2588.

181B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4
Chang, L., Haning, W., 2006. Insights from recent positronemission tomographic studies of drug abuse and dependence.Curr. Opin. Psychiatry 19, 246–252.
Chau, D.T., Roth, R.M., Green, A.I., 2004. The neural circuitry ofreward and its relevance to psychiatric disorders. Curr.Psychiatry Rep. 6, 391–399.
Chen, C.H., Lennox, B., Jacob, R., Calder, A., Lupson, V.,Bisbrown-Chippendale, R., Suckling, J., Bullmore, E., 2006.Explicit and implicit facial affect recognition in manic anddepressed States of bipolar disorder: a functional magneticresonance imaging study. Biol. Psychiatry 59, 31–39.
Christodoulou, T., Lewis, M., Ploubidis, G.B., Frangou, S., 2006. Therelationship of impulsivity to response inhibition anddecision-making in remitted patients with bipolar disorder.Eur. Psychiatr. 21, 270–273.
Coffey, S.F., Gudleski, G.D., Saladin, M.E., Brady, K.T., 2003.Impulsivity and rapid discounting of delayed hypotheticalrewards in cocaine-dependent individuals. Exp. Clin.Psychopharmacol. 11, 18–25.
Cools, R., Clark, L., Owen, A.M., Robbins, T.W., 2002. Defining theneural mechanisms of probabilistic reversal learning usingevent-related functional magnetic resonance imaging.J. Neurosci. 22, 4563–4567.
Cox, S.M., Andrade, A., Johnsrude, I.S., 2005. Learning to like: a rolefor human orbitofrontal cortex in conditioned reward. J.Neurosci. 25, 2733–2740.
Currie, S.R., Patten,S.B.,Williams, J.V.,Wang, J., Beck,C.A., El-Guebaly,N., Maxwell, C., 2005. Comorbidity of major depression withsubstance use disorders. Can. J. Psychiatry 50, 660–666.
Davidson, R.J., Pizzagalli, D., Nitschke, J.B., Putnam, K., 2002.Depression: perspectives from affective neuroscience. Annu.Rev. Psychol. 53, 545–574.
Davidson, M.C., Horvitz, J.C., Tottenham, N., Fossella, J.A., Watts,R., Ulug, A.M., Caseya, B.J., 2003a. Differential cingulate andcaudate activation following unexpected nonrewardingstimuli. NeuroImage 23, 1039–1045.
Davidson, R.J., Irwin, W., Anderle, M.J., Kalin, N.H., 2003b. Theneural substrates of affective processing in depressed patientstreated with venlafaxine. Am. J. Psychiatry 160, 64–75.
Davidson,M.C., Horvitz, J.C., Tottenham,N., Fossella, J.A.,Watts, R.,Uluğ, A.M., Casey, B.J., 2004. Differential cingulate and caudateactivation following unexpected nonrewarding stimuliNeuroimage 23, 1039–1045.
Dom, G., Sabbe, B., Hulstijn, W., van den Brink,W., 2005. Substanceuse disorders and the orbitofrontal cortex: systematic reviewof behavioural decision-making and neuroimaging studies.Br. J. Psychiatry 187, 209–220.
Drevets, W.C., 1999. Prefrontal cortical-amygdalar metabolism inmajor depression. Ann. N. Y. Acad. Sci. 877, 614–637.
Drevets, W.C., 2001. Neuroimaging and neuropathological studiesof depression: implications for the cognitive-emotionalfeatures of mood disorders. Curr. Opi. Neurobiol. 11, 240–249.
Drevets, W.C., Gautier, C., Price, J.C., Kupfer, D.J., Kinahan, P.E.,Grace, A.A., Price, J.L., Mathis, C.A., 2001. Amphetamine-induced dopamine release in human ventral striatumcorrelates with euphoria. Biol. Psychiatry 49, 81–96.
Elliott, R., Sahakian, B.J., Michael, A., Paykel, E.S., Dolan, R.J., 1998.Abnormal neural response to feedback on planning andguessing tasks in patients with unipolar depression. Psychol.Med. 28, 559–571.
Elliott, R., Rubinsztein, J.S., Sahakian, B.J., Dolan, R.J., 2002. Theneural basis of mood-congruent processing biases indepression. Arch. Gen. Psychiatry 59, 597–604.
Elliott, R., Ogilvie, A., Rubinsztein, J.S., Calderon, G., Dolan, R.J.,Sahakian, B.J., 2004. Abnormal ventral frontal response duringperformance of an affective go/no go task in patients withmania. Biol. Psychiatry 55, 1163–1170.
Elliott, R., Deakin, B., 2005. Role of the orbitofrontal cortex inreinforcement processing and inhibitory control: evidence
from functional magnetic resonance imaging studies inhealthy human subjects. Int. Rev. Neurobiol. 65, 89–116.
Epstein, J., Pan, H., Kocsis, J.H., Yang, Y., Butler, T., Chusid, J.,Hochberg, H., Murrough, J., Strohmayer, E., Stern, E.,Silbersweig, D.A., 2006. Lack of ventral striatal response topositive stimuli in depressed versus normal subjects. Am. J.Psychiatry 163, 1784–1790.
Ernst, M., Paulus, M.P., 2005. Neurobiology of decision making: aselective review from a neurocognitive and clinicalperspective. Biol. Psychiatry 58, 597–604.
Ersche, K.D., Fletcher, P.C., Roiser, J.P., Fryer, T.D., London, M.,Robbins, T.W., Sahakian, B.J., 2006. Differences in orbitofrontalactivation during decision-making betweenmethadone-maintained opiate users, heroin users and healthyvolunteers. Psychopharmacology 188, 364–373.
Fellows, L.K., Farah, M.J., 2005. Different underlying impairmentsin decision-making following ventromedial and dorsolateralfrontal lobe damage in humans. Cerebral Cortex 15, 58–63.
Fitzgerald, D.A., Angstadt, M., Jelsone, L.M., Nathan, P.J., Phan,K.L., 2006. Beyond threat: amygdala reactivity acrossmultiple expressions of facial affect. Neuroimage 30,1441–1448.
Franklin, T.R., Wang, Z., Wang, J., Sciortino, N., Harper, D., Li, Y.,Ehrman, R., Kampman, K., O'Brien, C.P., Detre, J.A., Childress,A.R., 2007. Limbic Activation to Cigarette Smoking CuesIndependent of Nicotine Withdrawal: a Perfusion fMRI Study.Neuropsychopharmacology (Electronic publication ahead ofprint).
Fu, C.H., Williams, S.C., Cleare, A.J., Brammer, M.J., Walsh, N.D.,Kim, J., Andrew, C.M., Pich, E.M., Williams, P.M., Reed, L.J.,Mitterschiffthaler, M.T., Suckling, J., Bullmore, E.T., 2004.Attenuation of the neural response to sad faces in majordepression by antidepressant treatment: a prospective,event-related functional magnetic resonance imaging study.Arch. Gen. Psychiatry 61, 877–889.
Garavan, H., Pankiewicz, J., Bloom, A., Cho, J.K., Sperry, L., Ross,T.J., Salmeron, B.J., Risinger, R., Kelley, D., Stein, E.A., 2000.Cue-induced cocaine craving: neuroanatomical specificityfor drug users and drug stimuli. Am. J. Psychiatry. 157,1789–1798.
Garavan, H., Pendergrass, J.C., Ross, T.J., Stein, E.A., Risinger, R.C.,2001. Amygdala response to both positively and negativelyvalenced stimuli. Neuroreport 12, 2779–2783.
Galvan, A., Hare, T.A., Davidson, M., Spicer, J., Glover, G., Casey,B.J., 2005. The role of ventral frontostriatal circuitry inreward-based learning in humans. J. Neurosci. 25, 8650–8656.
Getz, G.E., Shear, P.K., Strakowski, S.M., 2003. Facial affectrecognition deficits in bipolar disorder. J. Int. Neuropsychol.Soc. 9, 623–632.
Giordano, L.A., Bickel, W.K., Loewenstein, G., Jacobs, E.A., Marsch,L., Badger, G.J., 2002. Mild opioid deprivation increases thedegree that opioid-dependent outpatients discount delayedheroin and money. Psychopharmacology (Berl) 163,174–182.
Glascher, J., Buchel, C., 2005. Formal learning theory dissociatesbrain regions with different temporal integration. Neuron 47,295–306.
Goldstein, R.Z., Volkow, N.D., 2002. Drug addiction and itsunderlying neurobiological basis: neuroimaging evidence forthe involvement of the frontal cortex. Am. J. Psychiatry 159,1642–1652.
Goldstein, R.Z., Alia-Klein, N., Tomasi, D., Zhang, L., Cottone, L.A.,Maloney, T., Telang, F., Caparelli, E.C., Chang, L., Ernst, T.,Samaras, D., Squires, N.K., Volkow, N.D., 2007a. Is decreasedprefrontal cortical sensitivity to monetary reward associatedwith impaired motivation and self-control in cocaineaddiction? Am. J. Psychiatry 164, 43–51.
Goldstein, R.Z., Tomasi, D., Alia-Klein, N., Cottone, L.A., Zhang, L.,Telang, F., Volkow, N.D., 2007b. Subjective sensitivity to

182 B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4
monetary gradients is associated with frontolimbic activationto reward in cocaine abusers. Drug andAlcohol Dependence 87,233–240.
Gotlib, I.H., Sivers, H., Gabrieli, J.D.E., Whitfield-Gabrieli, S., Goldin,P., Minor, K.L., Canli, T., 2005. Subgenual anterior cingulateactivation to valenced emotional stimuli in major depression.Neuroreport 16, 1731–1734.
Gottfried, J.A., O'Doherty, J., Dolan, R.J., 2002. Appetitive andaversive olfactory learning in humans studied usingevent-related functional magnetic resonance imaging.J. Neurosci. 22, 10829–10837.
Gottfried, J.A., O'Doherty, J., Dolan, R.J., 2003. Encoding predictivereward value in human amygdala and orbitofrontal cortex.Science 301, 1104–1107.
Grant, J.E., Brewer, J.A., Potenza, M.N., 2006. The neurobiology ofsubstance and behavioral addictions. CNS Spectrums 11,924–930.
Grusser, S.M., Wrase, J., Klein, S., Hermann, D., Smolka, M.N., Ruf,M., Weber-Fahr, W., Flor, H., Mann, K., Braus, D.F., Heinz, A.,2004. Cue-induced activation of the striatum and medialprefrontal cortex is associated with subsequent relapse inabstinent alcoholics. Psychopharmacology (Berl). 175, 296–302.
Haber, S.N., Fudge, J.L., McFarland, N.R., 2000. Striatonigrostriatalpathways in primates form an ascending spiral from the shellto the dorsolateral striatum. J. Neurosci. 20, 2369–2382.
Haber, S.N., Kim, K.S., Mailly, P., Calzavara, R., 2006.Reward-related cortical inputs define a large striatal regionin primates that interface with associative corticalconnections, providing a substrate for incentive-basedlearning. J. Neurosci. 26, 8368–8376.
Heinz, A., Siessmeier, T., Wrase, J., Hermann, D., Klein, S., Grusser,S.M., Flor, H., Braus, D.F., Buchholz, H.G., Grunder, G.,Schreckenberger, M., Smolka, M.N., Rosch, F., Mann, K.,Bartenstein, P., 2004. Correlation between dopamine D(2)receptors in the ventral striatum and central processing ofalcohol cues and craving. Am. J. Psychiatry 161, 1783–1789.
Hermann, D., Smolka, M.N., Wrase, J., Klein, S., Nikitopoulos, J.,Georgi, A., Braus, D.F., Flor, H., Mann, K., Heinz, A., 2006.Blockade of cue-induced brain activation of abstinentalcoholics by a single administration of amisulpride asmeasured with fMRI. Alcoholism: Clin. Exp. Res. 30,1349–1354.
Holroyd, C.B., Coles, M.G.H., 2002. The neural basis of human errorprocessing: reinforcement learning, dopamine, and theerror-related negativity. Psychol. Rev. 109, 679–709.
Horvitz, J.C., 2000. Mesolimbocortical and nigrostriatal dopamineresponses to salient non-reward events. Neuroscience 96,651–656.
Hyman, S.E., Malenka, R.C., Nestler, E.J., 2006. Neural mechanismsof addiction: the role of reward-related learning and memory.Annu. Rev. Neurosci. 29, 565–598.
James, G.A., Gold, M.S., Liu, Y., 2004. Interaction of satiety andreward response to food stimulation. J. Addict. Disord. 23, 23–37.
Jensen, J., McIntosh, A.R., Crawley, A.P., Mikulis, D.J., Remington,G., Kapur, S., 2003. Direct activation of the ventral striatum inanticipation of aversive stimuli. Neuron 40, 1251–1257.
Juckel, G., Schlagenhauf, F., Koslowski, M., Wustenberg, T.,Villringer, A., Knutson, B., Wrase, J., Heinz, A., 2006.Dysfunction of ventral striatal reward prediction inschizophrenia. Neuroimage 29, 409–416.
Kalivas, P.W., Peters, J., Knackstedt, L., 2006. Animal models andbrain circuits in drug addiction. Mol. Interv. 6, 339–344.
Keedwell, P.A., Andrew, C., Williams, S.C., Brammer, M.J., Phillips,M.L., 2005a. The neural correlates of anhedonia in majordepressive disorder. Biol. Psychiatry 58, 843–853.
Keedwell, P.A., Andrew, C., Williams, S.C., Brammer, M.J., Phillips,M.L., 2005b. A double dissociation of ventromedial prefrontalcortical responses to sad and happy stimuli in depressed andhealthy individuals. Biol. Psychiatry 58, 495–503.
Kilts, C.D., Gross, R.E., Ely, T.D., Drexler, K.P., 2004. The neuralcorrelates of cue-induced craving in cocaine-dependentwomen. Am. J. Psychiatry 161, 233–241.
Kirsch, P., Schienle, A., Stark, R., Sammer, G., Blecker, C., Walter, B.,Ott, U., Burkart, J., Vaitl, D., 2003. Anticipation of reward in anonaversive differential conditioning paradigm and the brainreward system: an event-related fMRI study. Neuroimage 20,1086–1095.
Knutson, B., Adams, C.M., Fong, G.W., Hommer, D., 2001.Anticipation of increasing monetary reward selectivelyrecruits nucleus accumbens. J. Neurosci. 21, 159.
Knutson, B., Fong, G.W., Bennett, S.M., Adams, C.M., Hommer, D.,2003. A region of mesial prefrontal cortex tracks monetarilyrewarding outcomes: characterizationwith rapid event-relatedfMRI. Neuroimage 18, 263–272.
Knutson, B., Taylor, J., Kaufman, M., Peterson, R., Glover, G., 2005.Distributed neural representation of expected value.J. Neurosci. 25, 4806–4812.
Knutson, B., Bhanji, J.P., Cooney, R.E., Atlas, L.Y., Gotlib, I.H., inpress. Neural responses to monetary incentives in majordepression. Biol. Psychiatry.
Kringelbach, M.L., 2005. The human orbitofrontal cortex: linkingreward to hedonic experience. Nat. Rev., Neurosci. 6, 691–702.
Kringelbach, M.L., Rolls, E.T., 2003. Neural correlates of rapidreversal learning in a simple model of human socialinteraction. Neuroimage 20, 1371–1383.
Kringelbach, M.L., O'Doherty, J., Rolls, E.T., Andrews, C., 2003.Activation of the human orbitofrontal cortex to a liquid foodstimulus is correlated with its subjective pleasantness.Cerebral Cortex 13, 1064–1071.
Kumari, V., Mitterschiffthaler, M.T., Teasdale, J.D., Malhi, G.S.,Brown, R.G., Giampietro, V., Brammer, M.J., Poon, L.,Simmons, A., Williams, S.C., Checkley, S.A., Sharma, T.,2003. Neural abnormalities during cognitive generation ofaffect in treatment-resistant depression. Biol. Psychiatry 54,777–791.
Laird, A.R., Fox, M., Price, C.J., Glahn, D.C., Uecker, A.M., Lancaster,J.L., Turkeltaub, P.E., Kochunov, P., Fox, P.T., 2005a. ALEmeta-analysis: controlling the false discovery rate andperforming statistical contrasts. Hum. Brain Mapp. 25, 155–164.
Laird, A.R., McMillan, K.M., Lancaster, J.L., Kochunov, P.,Turkeltaub, P.E., Pardo, J.V., Fox, P.T., 2005b. A comparison oflabel-based review and ALE meta-analysis in the Stroop task.Hum. Brain Mapp. 25, 6–21.
Laird, A.R., Lancaster, J.L., Fox, P.T., 2005c. BrainMap: the socialevolution of a functional neuroimaging database.Neuroinformatics 3, 65–78.
Lawrence, N.S., Williams, A.M., Surguladze, S., Giampietro, V.,Brammer, M.J., Andrew, C., Frangou, S., Ecker, C., Phillips, M.L.,2004. Subcortical and ventral prefrontal cortical neuralresponses to facial expressions distinguish patients withbipolar disorder and major depression. Biol. Psychiatry 55,578–587.
Lennox, B.R., Jacob, R., Calder, A.J., Lupson, V., Bullmore, E.T., 2004.Behavioural and neurocognitive responses to sad facial affectare attenuated in patients with mania. Psychol. Med. 34,795–802.
Leppänen, J.M., Milders, M., Bell, J.S., Terriere, E., Hietanen, J.K.,2004. Depression biases the recognition of emotionally neutralfaces. Psychiatry Res. 128, 123–133.
Leppänen, J.M., 2006. Emotional information processing in mooddisorders: a review of behavioral and neuroimaging findings.Curr. Opin. Psychiatry 19, 34–39.
Liberzon, I., Phan, K.L., Decker, L.R., Taylor, S.F., 2003. Extendedamygdala and emotional salience: a PET activation studyof positive and negative affect. Neuropsychopharmacology 28,726–733.
Malhi, G.S., Lagopoulos, J., Ward, P.B., Kumari, V., Mitchell, P.B.,Parker, G.B., Ivanovski, B., Sachdev, P., 2004a. Cognitive

183B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4
generation of affect in bipolar depression: an fMRI study.European J. Neurosci. 19, 741–754.
Malhi, G.S., Lagopoulos, J., Sachdev, P., Mitchell, P.B., Ivanovski, B.,Parker, G.B., 2004b. Cognitive generation of affect inhypomania: an fMRI study. Bipolar Disord. 6, 271–285.
Malhi, G.S., Lagopoulos, J., Sachdev, P.S., Ivanovski, B., Shnier, R.,2005. An emotional Stroop functional MRI study of euthymicbipolar disorder. Bipolar Disord. 7 (Suppl 5), 58–69.
Malhi, G.S., Lagopoulos, J., Owen, A.M., Ivanovski, B., Shnier, R.,Sachdev, P., 2007. Reduced activation to implicit affectinduction in euthymic bipolar patients: an fMRI study. J. Affect.Disord. 97, 109–122.
Mancini-Marie, A., Potvin, S., Fahim, C., Beauregard, M., Mensour,B., Stip, E., 2006. Neural correlates of the affect regulationmodel in schizophrenia patients with substance use history: afunctional magnetic resonance imaging study. J. Clin.Psychiatry 67, 342–350.
Martin-Soelch, C., Chevalley, A.F., Kunig, G., Missimer, J., Magyar,S., Mino, A., Schultz, W., Leenders, K.L., 2001a. Changes inreward-induced brain activation in opiate addicts. European J.Neurosci. 14, 1360–1368.
Martin-Soelch, C., Magyar, S., Kunig, G., Missimer, J., Schultz, W.,Leenders, K.L., 2001b. Changes in brain activation associatedwith reward processing in smokers and nonsmokers. A positronemission tomography study. Exp. Brain Res. 139, 278–286.
McBride, D., Barrett, S.P., Kelly, J.T., Aw, A., Dagher, A., 2006. Effectsof expectancy and abstinence on the neural response tosmoking cues in cigarette smokers: an fMRI study.Neuropsychopharmacology 31, 2728–2738.
McClernon, F.J., Hiott, F.B., Huettel, S.A., Rose, J.E., 2005.Abstinence-induced changes in self-report craving correlatewith event-related FMRI responses to smoking cues.Neuropsychopharmacology 30, 1940–1947.
Mitchell, R.L., Elliott, R., Barry, M., Cruttenden, A., Woodruff, P.W.,2004. Neural response to emotional prosody in schizophreniaand in bipolar affective disorder. Br. J. Psychiatry 184, 223–230.
Montague, P.R., Berns, G.S., 2002. Neural economics and thebiological substrates of valuation. Neuron 36, 265–284.
Moritz, S., Glascher, J., Brassen, S., 2005. Investigation ofmood-congruent false and true memory recognition indepression. Depress. Anxiety 21, 9–17.
Morris, J.S., Dolan, R.J., 2001. Involvement of human amygdala andorbitofrontal cortex in hunger-enhanced memory for foodstimuli. J. Neurosci. 21, 5304–5310.
Must, A., Szabo, Z., Bodi, N., Szasz, A., Janka, Z., Keri, S., 2006.Sensitivity to reward and punishment and the prefrontalcortex in major depression. J. Affect. Disord. 90, 209–215.
Northoff, G., Grimm, S., Boeker, H., Schmidt, C., Bermpohl, F.,Heinzel, A., Hell, D., Boesiger, P., 2006. Affective judgment andbeneficial decision making: ventromedial prefrontal activitycorrelates with performance in the Iowa Gambling Task. Hum.Brain Mapp. 27, 572–587.
Nutt, D., Demyttenaere, K., Janka, Z., Aarre, T., Bourin, M.,Canonico, P.L., Carrasco, J.L., Stahl, S., 2007. The other face ofdepression, reduced positive affect: the role of catecholaminesin causation and cure. J. Psychopharmacol. 21, 461–471.
O'Doherty, J.P., 2004. Reward representations and reward-relatedlearning in the human brain: insights from neuroimaging.Curr. Opin. Neurobiol. 14, 769–776.
O'Doherty, J., Rolls, E.T., Francis, S., Bowtell, R., McGlone, F., Kobal,G., Renner, B., Ahne, G., 2000. Sensory-specific satiety-relatedolfactory activation of the human orbitofrontal cortex.Neuroreport 11, 893–897.
O'Doherty, J., Kringelbach, M.L., Rolls, E.T., Hornak, J., Andrews, C.,2001. Abstract reward and punishment representationsin the human orbitofrontal cortex. Nat. Neurosci. 4, 95–102.
O'Doherty, J.P., Deichmann, R., Critchley, H.D., Dolan, R.J., 2002.Neural responses during anticipation of a primary tastereward. Neuron 33, 815–826.
O'Doherty, J.P., Dayan, P., Friston, K., Critchley, H., Dolan, R.J.,2003a. Temporal difference models and reward-relatedlearning in the human brain. Neuron 38, 329–337.
O'Doherty, J., Critchley, H., Deichmann, R., Dolan, R.J., 2003b.Dissociating valence of outcome from behavioral control inhuman orbital and ventral prefrontal cortices. J. Neurosci. 23,7931–7939.
O'Doherty, J., Dayan, P., Schultz, J., Deichmann, R., Friston, K.,Dolan, R.J., 2004. Dissociable roles of ventral and dorsalstriatum in instrumental conditioning. Science 304,452–454.
Pessiglione, M., Seymour, B., Flandin, G., Dolan, R.J., Frith, C.D.,2006. Dopamine-dependent prediction errors underpinreward-seeking behaviour in humans. Nature 442,1042–1045.
Phan, K.L., Wager, T.D., Taylor, S.F., Liberzon, I., 2004. Functionalneuroimaging studies of human emotions. CNS Spectrums 9,258–266.
Potenza, M.N., 2006. Should addictive disorders includenon-substance-related conditions? Addiction 101 (suppl 1),142–151.
Redgrave, P., Prescott, T.J., Gurney, K., 1999. Is the short-latencydopamine response too short to signal reward error? TrendsNeurosci. 22, 146–151.
Redish, A.D., 2004. Addiction as a computational process goneawry. Science 306, 1944–1947.
Reuter, J., Raedler, T., Rose, M., Hand, I., Glascher, J., Buchel, C.,2005. Pathological gambling is linked to reduced activation ofthe mesolimbic reward system. Nat. Neurosci. 8, 147–148.
Risinger, R.C., Salmeron, B.J., Ross, T.J., Amen, S.L., Sanfilipo, M.,Hoffmann, R.G., Bloom, A.S., Garavan, H., Stein, E.A., 2005.Neural correlates of high and craving during cocaineself-administration using BOLD fMRI. Neuroimage 26,1097–1108.
Robinson, T.E., Berridge, K.C., 2003. Addiction. Annu. Rev. Psychol.54, 25–53.
Rolls, E.T., Kringelbach, M.L., de Araujo, I.E., 2003. Differentrepresentations of pleasant and unpleasant odours in thehuman brain. Eur. J. Neurosci. 18, 695–703.
Rubinsztein, J.S., Fletcher, P.C., Rogers, R.D., Ho, L.W., Aigbirhio, F.I.,Paykel, E.S., Robbins, T.W., Sahakian, B.J., 2001.Decision-making in mania: a PET study. Brain 124, 2550–2563.
Sander, D., Grafman, J., Zalla, T., 2003. The human amygdala: anevolved system for relevance detection. Rev. Neurosci. 14,303–316.
Schaefer, H.S., Putnam, K.M., Benca, R.M., Davidson, R.J., 2006.Event-related functional magnetic resonance imagingmeasures of neural activity to positive social stimuli inpre- and post-treatment depression. Biol. Psychiatry 60,974–986.
Schneider, F., Habel, U., Wagner, M., Franke, P., Salloum, J.B., Shah,N.J., Toni, I., Sulzbach, C., Honig, K., Maier, W., Gaebel, W.,Zilles, K., 2001. Subcortical correlates of craving in recentlyabstinent alcoholic patients. Am. J. Psychiatry 158, 1075–1083.
Schoenbaum, G., Roesch, M.R., Stalnaker, T.A., 2006. Orbitofrontalcortex, decision-making and drug addiction. Trends Neurosci.29, 116–124.
Schultz, W., 1998. Predictive reward signal of dopamine neurons.J. Neurophysiol. 80, 1–27.
Sell, L.A., Morris, J.S., Bearn, J., Frackowiak, R.S.J., Friston, K.J.,Dolan, R.J., 2000. Neural responses associated with cue evokedemotional states and heroin in opiate addicts. Drug AlcoholDepend. 60, 207–216.
Seymour, B., O'Doherty, J.P., Koltzenburg, M., Wiech, K.,Frackowiak, R., Friston, K., Dolan, R., 2005. Opponentappetitive–aversive neural processes underlie predictivelearning of pain relief. Nat. Neurosci. 8, 1234–1240.
Sheline, Y.I., Barch, D.M., Donnelly, J.M., Ollinger, J.M., Snyder, A.Z.,Mintun, M.A., 2001. Increased amygdala response to masked

184 B R A I N R E S E A R C H R E V I E W S 5 9 ( 2 0 0 8 ) 1 6 4 – 1 8 4
emotional faces in depressed subjects resolves withantidepressant treatment: an fMRI study. Biol. Psychiatry 50,651–658.
Siegle, G.J., Steinhauer, S.R., Thase, M.E., Stenger, V.A., Carter, C.S.,2002. Can't shake that feeling: event-related fMRI assessmentof sustained amygdala activity in response to emotionalinformation in depressed individuals. Biol. Psychiatry 51,693–707.
Surguladze, S.A., Young, A.W., Senior, C., Brebion, G., Travis, M.J.,Phillips, M.L., 2004. Recognition accuracy and response bias tohappy and sad facial expressions in patients with majordepression. Neuropsychology 18, 212–218.
Surguladze, S., Brammer, M.J., Keedwell, P., Giampietro, V., Young,A.W., Travis, M.J., Williams, S.C., Phillips, M.L., 2005. Adifferential pattern of neural response toward sad versushappy facial expressions in major depressive disorder. Biol.Psychiatry 57, 201–209.
Sutton, R.S., Barto, A.G., 1998. Reinforcement Learning: AnIntroduction. MIT Press, Cambridge, MA.
Tobler, P.N., O'Doherty, J.P., Dolan, R.J., Schultz, W., 2006. Humanneural learning depends on reward prediction errors in theblocking paradigm. J. Neurophysiol. 95, 301–310.
Tobler, P.N., O'Doherty, J.P., Dolan, R.J., Schultz, W., 2007. Rewardvalue coding distinct from risk attitude-related uncertaintycoding in human reward systems. J. Neurophysiol. 97,1621–1632.
Tremblay, L.K., Naranjo, C.A., Graham, S.J., Herrmann, N.,Mayberg, H.S., Hevenor, S., Busto, U.E., 2005. Functionalneuroanatomical substrates of altered reward processing inmajor depressive disorder revealed by a dopaminergic probe.Arch. Gen. Psychiatry 62, 1228–1236.
Trepel, C., Fox, C.R., Poldrack, R.A., 2005. Prospect theory on thebrain? Toward a cognitive neuroscience of decision under risk.Brain Res. Cogn. Brain Res. 23, 34–50.
Trinko, R., Sears, R.M., Guarnieri, D.J., DiLeone, R.J., 2007. Neuralmechanisms underlying obesity and drug addiction. Physiol.Behav. 91, 499–505.
Turkeltaub, P.E., Eden, G.F., Jones, K.M., Zeffiro, T.A., 2002.Meta-analysis of the functional neuroanatomy of single-wordreading: method and validation. Neuroimage 16, 765–780.
Ungless, M.A., 2004. Dopamine: the salient issue. Trends Neurosci.27, 702–706.
Volkow, N.D., Fowler, J.S., 2000. Addiction, a disease of compulsionand drive: involvement of the orbitofrontal cortex. CerebralCortex 10, 318–325.
Volkow, N.D., Fowler, J.S., Wang, G.J., 2003. The addicted humanbrain: insights from imaging studies. J. Clin. Invest. 111,1444–1451.
Volkow, N.D., Fowler, J.S., Wang, G.J., Swanson, J.M., Telang, F.,2007. Dopamine in drug abuse and addiction: results ofimaging studies and treatment implications. Archives ofNeurology 64, 1575–1579.
Vuilleumier, P., Pourtois, G., 2006. Distributed and interactivebrain mechanisms during emotion face perception: evidencefrom functional neuroimaging. Neuropsychologia 45,174–194.
Wager, T.D., Lindquist, M., Kaplan, L., 2007. Meta-analysis offunctional neuroimaging data: Current and future directions.Soc., Cogn., Affect. Neurosci. 2, 150–158.
Wang, G.J., Volkow, N.D., Fowler, J.S., Cervany, P., Hitzemann, R.J.,Pappas, N.R., Wong, C.T., Felder, C., 1999. Regional brainmetabolic activation during craving elicited by recall ofprevious drug experiences. Life Sciences 64, 775–784.
Wang, G.J., Volkow, N.D., Thanos, P.K., Fowler, J.S., 2004. Similaritybetween obesity and drug addiction as assessed byneurofunctional imaging: a concept review. J. Addict. Disord.23, 39–53.
Wexler, B.E., Gottschalk, C.H., Fulbright, R.K., Prohovnik, I.,Lacadie, C.M., Rounsaville, B.J., Gore, J.C., 2001. Functionalmagnetic resonance imaging of cocaine craving. Am. J.Psychiatry 158, 86–95.
Whalen, P.J., 1998. Fear, vigilance, and ambiguity: initialneuroimaging studies of the human amygdala. Curr. Dir.Psychol. Sci. 7, 177–188.
Williams, L.M., 2006. An integrative neuroscience model of“significance” processing. J. Integr. Neurosci. 5, 1–47.
Willner, P., 2002. Dopamine and depression. In: Di Chiara, G. (Ed.),Handbook of Experimental Pharmacology: Dopamine in theCNS. Springer, Berlin, pp. pp. 387–416.
Willner, P., Hale, A.S., Argyropoulos, S., 2005. Dopaminergicmechanism of antidepressant action in depressed patients.J. Affect. Disord. 86, 37–45.
Wrase, J., Grusser, S.M., Klein, S., Diener, C., Hermann, D., Flor, H.,Mann, K., Braus, D.F., Heinz, A., 2002. Development ofalcohol-associated cues and cue-induced brain activation inalcoholics. Eur. Psychiatr. 17, 287–291.
Wrase, J., Kahnt, T., Schlagenhauf, F., Beck, A., Cohen, M.X.,Knutson, B., Heinz, A., 2007. Different neural systems adjustmotor behavior in response to reward and punishment.Neuroimage 36, 1253–1262.
Wrase, J., Schlagenhauf, F., Kienast, T.,Wustenberg, T., Bermpohl, F.,Kahnt, T., Beck, A., Strohle, A., Juckel, G., Knutson, B., Heinz, A.,2007b. Dysfunction of reward processing correlates with alcoholcraving in detoxified alcoholics. Neuroimage 35, 787–794.
Yacubian, J., Glascher, J., Schroeder, K., Sommer, T., Braus, D.F.,Buchel, C., 2006. Dissociable systems for gain- and loss-relatedvalue predictions and errors of prediction in the humanbrain. J. Neurosci. 26, 9530–9537.
Yurgelun-Todd, D.A., Gruber, S.A., Kanayama, G., Killgore, W.D.,Baird, A.A., Young, A.D., 2000. fMRI during affectdiscrimination in bipolar affective disorder. Bipolar Disord. 2,237–248.
Zald, D.H., Boileau, I., El-Dearedy, W., Gunn, R., McGlone, F.,Dichter, G.S., Dagher, A., 2004. Dopamine transmission in thehuman striatum during monetary reward tasks. J. Neurosci. 24,4105–4112.
Zink, C.F., Pagnoni, G., Martin, M.E., Dhamala, M., Berns, G.S., 2003.Human striatal response to salient nonrewarding stimuli.J. Neurosci. 23, 8092–8097.
Zink, C.F., Pagnoni, G., Chappelow, J., Martin-Skurski, M., Berns,G.S., 2006. Human striatal activation reflects degree ofstimulus saliency. Neuroimage 29, 977–983.






![Diseny 2008 Calendar [MMS911© 2008] · Title: Diseny 2008 Calendar [MMS911© 2008] Author: MMS911© 2008 Subject: 2008 Calendar Keywords: Disney; Calendar Created Date: 1/13/2008](https://img.pdfslide.net/doc/110x75/6059ae5486c210065a54d497/diseny-2008-calendar-mms911-2008-title-diseny-2008-calendar-mms911-2008.jpg)






















