Embed Size (px)
Citation preview

Introduction
Feeding characterizations are key to understand different biological aspects such as: the ecological organization in communities and assemblages (Toft, 1980; Toft, 1981; Lima and Magnuson, 1998; Parmelee, 1999; Muñoz-Guerrero et al., 2007; Arroyo et al., 2008), the phylogenetic relationships among species (Santos et al., 2003; Darst, 2005; Grant et al., 2006), and their behavior and physiology (Toft, 1980; Toft, 1981; Lima and Magnuson, 1998). For instance, the diet composition has been showed to be important in
the early evolutionary history of some higher taxa (Vitt and Pianka, 2005), influencing the present organization of the communities (Greene, 2005). Diet has important consequences in the occupation of habitats, being one of the most important dimensions in the resource partitioning (Schoener, 1974; Toft, 1985).
There are many factors influencing the feeding habits in amphibians, such as seasonal abundance of food (Toft, 1980), ecological tolerances, and morphological constraints (Duellman andTrueb, 1994; Pough et al., 2004). In spite of the fact that the dietary specialization is common in dendrobatids (Toft, 1995; Caldwell, 1996), most of the amphibians are considered generalist carnivores, whose diets are composed mainly by arthropods and other invertebrates (Duellman and Trueb, 1994; Pough et al., 2004).
Herein, we report the diet and trophic ecology of Leptodactylus fragilis (Brochi, 1977) and Dendrosophus columbianus (Boettger, 1892) that inhabit open and disturbed environments. Leptodactylus fragilis (Fig. 1A) is widely distributed from Texas in the United States until the north of South America (Colombia and Venezuela). In Colombia, the species is distributed primarily in the north of the country (Heyer, 1978), as well as in the valleys of the Río Magdalena and Río Cauca (Heyer et al., 2006; Mendez-Narvaez et al.,
Herpetology Notes, volume 7: 299-305 (2014) (published online on 26 May 2014)
Diet and trophic ecology of Leptodactylus fragilis (Leptodactylidae) and Dendropsophus columbianus (Anura:
Hylidae) in a disturbed area in southwestern Colombia
Javier Méndez-Narváez1,4,* , Jhon Jairo Ospina-Sarria,2,4 and Wilmar Bolívar-G2,4
1 Grupo de Ecofisiología, Comportamiento y Herpetología.
Department of Biology, Universidad de los Andes, Bogotá, Colombia, A.A. 4976
2 Instituto de Ciencia Naturales, Universidad Nacional de Colombia, Bogotá, Colombia
3 Grupo de investigación en Ecología Animal, Departamento de Biología, Universidad del Valle, Cali-Colombia. A. A. 25360, Cali, Colombia.
4 Fundación Calima, fundación para la investigación de la biodiversidad y la conservación
*Corresponding author; e-mail: [email protected]
Abstract. Here we described the diet and trophic ecology of Leptodactylus fragilis and Dendropsophus columbianus in a disturbed habitat in the Cordillera Central, Colombia. We identified the food items to the lowest possible taxonomic category and analyzed the trophic niche breadth and niche overlap of L. fragilis and D. columbianus. The prey categories with the highest importance in the diet of L. fragilis were Coleoptera and Formicidae, whereas for D. columbianus were Araneae and Lepidoptera larvae. The trophic niche breadth was broader in D. columbianus than in L. fragilis. The trophic niche overlap between both species was 68%. The differences in the composition of the diet and in the trophic niche could be related to the foraging strategies of each species in a continuum of generalist-specialist forager; i.e., L. fragilis shows an active foraging behavior, whereas D. columbianus a sit and wait behavior.
Keywords: Leptodactylus fragilis, Dendropsophus columbianus, trophic niche, foraging strategy, disturbed habitat, Colombia

Javier Mendez-Narváez et al.300
2009). On the other hand, D. columbianus (Fig. 1B) is an endemic species from Colombia distributed on the western slopes of the Cordillera Central and eastern and western slopes of the Cordillera Occidental (Bolivar and Rengifo, 2004). Unfortunately, studies in natural history for these species are scarce (see Heyer et al., 20606 for L. fragilis) or practically non-existent. We herein described the diet and feeding ecology of these two species and analyze resource partitioning between them in a disturbed area.
Materials and Methods
We collected individuals of both species in the Municipality of Caloto, Vereda Morales, Department of Cauca, Colombia (02º 59` 49.1” N, 76º 24` 25.2” W; 1300 m). The study site is a tropical dry forest (bs-T, Holdridge’s classification, 1967), under intensive development of livestock and agricultural crops, mainly rice, with a few small relicts of native forest. The study was undertaken in a livestock area, where both species were active at night, characterized by both dry soil and small temporary ponds.
Sampling was carry out in October 2006 and March 2007 (four days duration in each one), using the method of visual encounter survey (Lips et al., 2001). We searched for specimens at night between 18:00 and 00:00 hours. The specimens were fixed in 10% formalin and preserved in 70% ethanol, the stomachs were removed and were preserved in 70% ethanol, and the stomach content was analyzed with a stereoscope. The identification of prey items were done to the lowest
taxonomy category. We measured the preys (length and width) using a digital caliper (0.01mm). We calculated the volume, occurrence (percentage of stomachs containing each prey category) and the numerical and volumetric percentage of each prey category. Volume of each prey item was calculated by means of the formula for an ellipsoid (Dunhan, 1983):
V = 4/3π (1/2 L) (1/2 W)2, where V is volume, L is the length and W is width.
We used the taxonomic category of Order to estimate the trophic niche breadth of the two species and the trophic niche overlap between them; since this category was the lowest common category for the food items in both species. Estimation of niche breadth was employed using the reciprocal index of Simpson (1949):
B = (1/ ∑ pi2), where “i” is the alimentary resource category, “p” is the proportion of the category used by the species (in relation to the number of consumed prey items). The value of niche breadth ranges between 1 (when only one category is consumed) and n (when all the prey categories are consumed equally).
To estimate trophic niche overlap between the two species, we used the index overlap proposed by Pianka (1973):
Where, pij and pik represent the proportion of the ith resource used by the jth and the kth species, respectively.
Figure 1. Species studied in the locality of Caloto, Cauca. A) Leptodactylus fragilis; B) Dendropsophus columbianus.
Oik= Okj = ∑ pij pik
∑ pij2∑ pik
2
Diet and trophic ecology of Leptodactylus fragilis and Dendropsophus columbianus 301
Additionally, the contribution of each prey category in the diet was determined through the index of relative importance (IRI) (Pinkas et al., 1971):
IRI = % FO x (% N + % V), where % FO is the frequency occurrence of each category, %N the numeric percentage and %V the volumetric percentage
The specimens of both species were deposited in the Museo de Herpetología at the Universidad del Valle, Cali-Colombia (UV-C 15672-15686) and in the collection of Docencia (CD 1601-1605, 1785, 1782-1784, 1786, 1791-1804, 1934-1939), Zoology section, Cali, Colombia.
Results
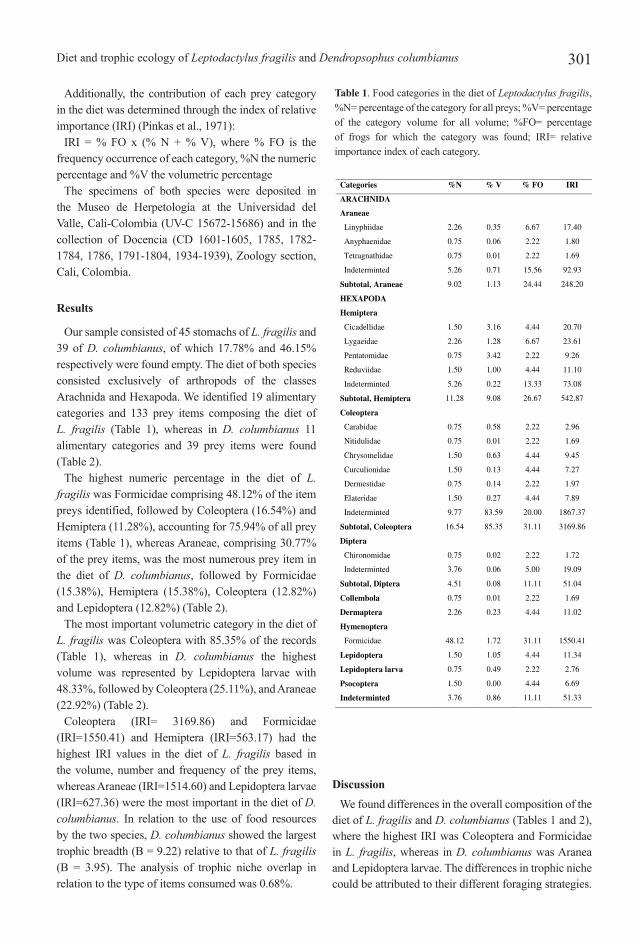
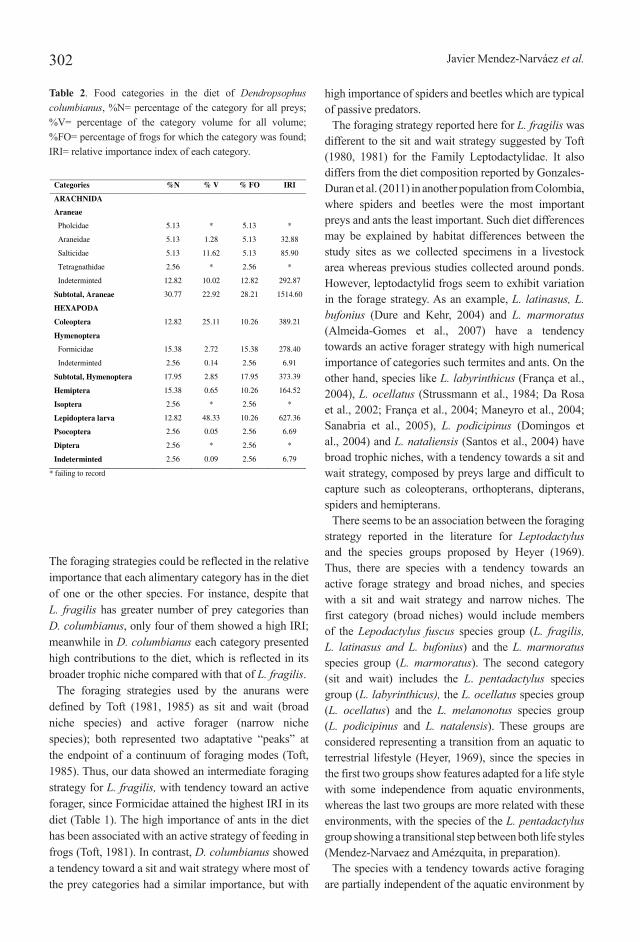
Our sample consisted of 45 stomachs of L. fragilis and 39 of D. columbianus, of which 17.78% and 46.15% respectively were found empty. The diet of both species consisted exclusively of arthropods of the classes Arachnida and Hexapoda. We identified 19 alimentary categories and 133 prey items composing the diet of L. fragilis (Table 1), whereas in D. columbianus 11 alimentary categories and 39 prey items were found (Table 2).
The highest numeric percentage in the diet of L. fragilis was Formicidae comprising 48.12% of the item preys identified, followed by Coleoptera (16.54%) and Hemiptera (11.28%), accounting for 75.94% of all prey items (Table 1), whereas Araneae, comprising 30.77% of the prey items, was the most numerous prey item in the diet of D. columbianus, followed by Formicidae (15.38%), Hemiptera (15.38%), Coleoptera (12.82%) and Lepidoptera (12.82%) (Table 2).
The most important volumetric category in the diet of L. fragilis was Coleoptera with 85.35% of the records (Table 1), whereas in D. columbianus the highest volume was represented by Lepidoptera larvae with 48.33%, followed by Coleoptera (25.11%), and Araneae (22.92%) (Table 2).
Coleoptera (IRI= 3169.86) and Formicidae (IRI=1550.41) and Hemiptera (IRI=563.17) had the highest IRI values in the diet of L. fragilis based in the volume, number and frequency of the prey items, whereas Araneae (IRI=1514.60) and Lepidoptera larvae (IRI=627.36) were the most important in the diet of D. columbianus. In relation to the use of food resources by the two species, D. columbianus showed the largest trophic breadth (B = 9.22) relative to that of L. fragilis (B = 3.95). The analysis of trophic niche overlap in relation to the type of items consumed was 0.68%.
DiscussionWe found differences in the overall composition of the
diet of L. fragilis and D. columbianus (Tables 1 and 2), where the highest IRI was Coleoptera and Formicidae in L. fragilis, whereas in D. columbianus was Aranea and Lepidoptera larvae. The differences in trophic niche could be attributed to their different foraging strategies.
Categories %N % V % FO IRI
ARACHNIDA
Araneae
Linyphiidae 2.26 0.35 6.67 17.40
Anyphaenidae 0.75 0.06 2.22 1.80
Tetragnathidae 0.75 0.01 2.22 1.69
Indeterminted 5.26 0.71 15.56 92.93
Subtotal, Araneae 9.02 1.13 24.44 248.20
HEXAPODA
Hemiptera
Cicadellidae 1.50 3.16 4.44 20.70
Lygaeidae 2.26 1.28 6.67 23.61
Pentatomidae 0.75 3.42 2.22 9.26
Reduviidae 1.50 1.00 4.44 11.10
Indeterminted 5.26 0.22 13.33 73.08
Subtotal, Hemiptera 11.28 9.08 26.67 542.87
Coleoptera
Carabidae 0.75 0.58 2.22 2.96
Nitidulidae 0.75 0.01 2.22 1.69
Chrysomelidae 1.50 0.63 4.44 9.45
Curculionidae 1.50 0.13 4.44 7.27
Dermestidae 0.75 0.14 2.22 1.97
Elateridae 1.50 0.27 4.44 7.89
Indeterminted 9.77 83.59 20.00 1867.37
Subtotal, Coleoptera 16.54 85.35 31.11 3169.86
Diptera
Chironomidae 0.75 0.02 2.22 1.72
Indeterminted 3.76 0.06 5.00 19.09
Subtotal, Diptera 4.51 0.08 11.11 51.04
Collembola 0.75 0.01 2.22 1.69
Dermaptera 2.26 0.23 4.44 11.02
Hymenoptera
Formicidae 48.12 1.72 31.11 1550.41
Lepidoptera 1.50 1.05 4.44 11.34
Lepidoptera larva 0.75 0.49 2.22 2.76
Psocoptera 1.50 0.00 4.44 6.69
Indeterminted 3.76 0.86 11.11 51.33
Table 1. Food categories in the diet of Leptodactylus fragilis, %N= percentage of the category for all preys; %V= percentage of the category volume for all volume; %FO= percentage of frogs for which the category was found; IRI= relative importance index of each category.
The foraging strategies could be reflected in the relative importance that each alimentary category has in the diet of one or the other species. For instance, despite that L. fragilis has greater number of prey categories than D. columbianus, only four of them showed a high IRI; meanwhile in D. columbianus each category presented high contributions to the diet, which is reflected in its broader trophic niche compared with that of L. fragilis.
The foraging strategies used by the anurans were defined by Toft (1981, 1985) as sit and wait (broad niche species) and active forager (narrow niche species); both represented two adaptative “peaks” at the endpoint of a continuum of foraging modes (Toft, 1985). Thus, our data showed an intermediate foraging strategy for L. fragilis, with tendency toward an active forager, since Formicidae attained the highest IRI in its diet (Table 1). The high importance of ants in the diet has been associated with an active strategy of feeding in frogs (Toft, 1981). In contrast, D. columbianus showed a tendency toward a sit and wait strategy where most of the prey categories had a similar importance, but with
high importance of spiders and beetles which are typical of passive predators.
The foraging strategy reported here for L. fragilis was different to the sit and wait strategy suggested by Toft (1980, 1981) for the Family Leptodactylidae. It also differs from the diet composition reported by Gonzales-Duran et al. (2011) in another population from Colombia, where spiders and beetles were the most important preys and ants the least important. Such diet differences may be explained by habitat differences between the study sites as we collected specimens in a livestock area whereas previous studies collected around ponds. However, leptodactylid frogs seem to exhibit variation in the forage strategy. As an example, L. latinasus, L. bufonius (Dure and Kehr, 2004) and L. marmoratus (Almeida-Gomes et al., 2007) have a tendency towards an active forager strategy with high numerical importance of categories such termites and ants. On the other hand, species like L. labyrinthicus (França et al., 2004), L. ocellatus (Strussmann et al., 1984; Da Rosa et al., 2002; França et al., 2004; Maneyro et al., 2004; Sanabria et al., 2005), L. podicipinus (Domingos et al., 2004) and L. nataliensis (Santos et al., 2004) have broad trophic niches, with a tendency towards a sit and wait strategy, composed by preys large and difficult to capture such as coleopterans, orthopterans, dipterans, spiders and hemipterans.
There seems to be an association between the foraging strategy reported in the literature for Leptodactylus and the species groups proposed by Heyer (1969). Thus, there are species with a tendency towards an active forage strategy and broad niches, and species with a sit and wait strategy and narrow niches. The first category (broad niches) would include members of the Lepodactylus fuscus species group (L. fragilis, L. latinasus and L. bufonius) and the L. marmoratus species group (L. marmoratus). The second category (sit and wait) includes the L. pentadactylus species group (L. labyrinthicus), the L. ocellatus species group (L. ocellatus) and the L. melanonotus species group (L. podicipinus and L. natalensis). These groups are considered representing a transition from an aquatic to terrestrial lifestyle (Heyer, 1969), since the species in the first two groups show features adapted for a life style with some independence from aquatic environments, whereas the last two groups are more related with these environments, with the species of the L. pentadactylus group showing a transitional step between both life styles (Mendez-Narvaez and Amézquita, in preparation).
The species with a tendency towards active foraging are partially independent of the aquatic environment by
Table 2. Food categories in the diet of Dendropsophus columbianus, %N= percentage of the category for all preys; %V= percentage of the category volume for all volume; %FO= percentage of frogs for which the category was found; IRI= relative importance index of each category.
Categories %N % V % FO IRI
ARACHNIDA
Araneae
Pholcidae 5.13 * 5.13 *
Araneidae 5.13 1.28 5.13 32.88
Salticidae 5.13 11.62 5.13 85.90
Tetragnathidae 2.56 * 2.56 *
Indeterminted 12.82 10.02 12.82 292.87
Subtotal, Araneae 30.77 22.92 28.21 1514.60
HEXAPODA
Coleoptera 12.82 25.11 10.26 389.21
Hymenoptera
Formicidae 15.38 2.72 15.38 278.40
Indeterminted 2.56 0.14 2.56 6.91
Subtotal, Hymenoptera 17.95 2.85 17.95 373.39
Hemiptera 15.38 0.65 10.26 164.52
Isoptera 2.56 * 2.56 *
Lepidoptera larva 12.82 48.33 10.26 627.36
Psocoptera 2.56 0.05 2.56 6.69
Diptera 2.56 * 2.56 *
Indeterminted 2.56 0.09 2.56 6.79
* failing to record
Javier Mendez-Narváez et al.302

placing their foam nests in terrestrial burrows (Heyer, 1969; Haddad and Prado, 2005), while those showing a tendency for the sit and wait strategy have higher dependence on aquatic or semi-aquatic environments. The type of foraging and trophic niche could be also related to structural differences in habitat heterogeneity. If this is true, we would expect that the areas with water sources have higher structural heterogeneity compared with dry areas. Those specimens feeding on the first area could have greater availability of prey items, because the positive correlation between heterogeneity and diversity of arthropods (Tews et al., 2004), and thus they can exhibit a sit and wait strategy and a broad trophic niche. Meanwhile, those predators in grasslands, which is characterized for low diversity of invertebrate and the dominance of ants (Chacón de Ulloa et al., 2006; Kamura et al., 2007), which live in colonies spaced along the habitat, should be active for detection and capturing (Dure and Kehr, 2004).
The sit and wait foraging strategy with broad trophic niche exhibited by D. columbianus and its generalist diet are similar to studies performed previously in other species of the genus Dendropsophus: e.g. Dendropsophus minutus (Van Sluys and Rocha, 1998), Dendropsophus sanborni and Dendropsophus nanus (Menin et al., 2004). The variety in prey categories reported here for D. columbianus have been reported in studies of diet in other members of the genus Dendropsophus (Basso, 1990; Van Sluys and Rocha, 1998; Peltzer and Lajmanovich, 2000; Menin et al., 2004; Santos et al., 2004; Muñoz-Guerrero et al., 2007). However, there was variation in the relative importance of the alimentary categories in the diet of the different species, as well as in populations of the same species (Van Sluys and Rocha, 1998; Peltzer and Lajmanovich, 2000; Santos et al., 2004; Basso, 1990; Menin et al., 2004). Those results can be attributed to differences in food availability in their habitats as has been suggested by Toft (1980) in tropical communities, since these frogs get their food passively and do not make distinctions among prey, accessing what is available in the environment.
The level of overlap in alimentary categories exhibited between D. columbianus and L. fragilis suggest a high similarity in the resource utilization in this habitat, which could indicate a competitive scenario between them for the food. However, if the resources being utilized by two species are not scarce, they could be shared without detriment to them (Pianka, 1974). In the same way, some studies have shown that other factors than diet, like
differences in foraging strategy (Lima and Magnuson, 1998; França et al., 2004), differential utilization of microhabitat (Van Sluys and Rocha, 1998; França et al., 2004; Dure and Kehr, 2004), and environmental structure (Menin et al., 2005) among others, have an influence in the differential distribution among sympatric species. This agrees with the idea, that there is not just one, but an interaction of various mechanisms that allow the coexistence among species in amphibian communities (Toft, 1985). Additionally, the taxonomic level to which prey were identified may underestimate differences in the trophic resource utilization between sympatric species (Menin et al., 2005). Despite their similarity in trophic resource utilization there were differences in the relative importance of each category in the species diet in this study. This could coincide with the regular spacing that presumably is present along one niche dimension (Schoener, 1974) being in this case the selectively of a particular type of prey: slow-moving arthropods, hard bodied (ants), or mobile arthropods soft bodied prey (spiders, crickets, beetles), based on their activity and capture cost (Toft, 1980).
In summary, both L. fragilis and D. columbianus showed a generalist diet, with differences in the relative importance of their alimentary categories. They also varied in the foraging strategies with a tendency towards one or another adaptative “peak” of foraging, when compared with other species in the genera Leptodactylus and Dendropsophus. Nevertheless, additional studies in different populations are needed, especially when living in sympatry with congeneric species. Also, it would be interesting to evaluate if some aspects related to the diet can reveal useful information to solve phylogenetic relationships among species, as has been shown in other anuran studies (Grant et al., 2006; Darst, 2005). Our study reveals some concordance between behavioral characters (foraging strategies) and taxonomic groups (Leptodactylus species group). But it is necessary to obtain more data from other species within the genus, and from species with sympatric distributions that belong to different species groups, all carried out within a phylogenic framework.
Acknowledgements. We are grateful to the academic program of Biology of the Universidad del Valle and its course of ecology. We thank to J. Cabra, A. Vasquez, V. Rodriguez and N. Baena for the help with the prey identification. We also thank to W.R. Heyer, R. de Sá, and M.N. de C. Kokubum for their comments in the early versions of the manuscript, which greatly improved its quality.
Diet and trophic ecology of Leptodactylus fragilis and Dendropsophus columbianus 303

References
Almeida-Gomes, M.M., Van Sluys, M., Duarte, C.F. (2007): Ecological observations on the leaf-litter frog Adenomera marmorata in an Atlantic rainforest area of southeastern Brazil. The Herpetololgical Journal 17: 81-85.
Arroyo S.B., Serrano-Cardozo, V.H., Ramírez, M.P. (2008): Diet, microhabitat and time of activity in a Pristimantis (Anura: Strabomantidae) assemblage. Phyllomedusa 7: 109-119.
Basso, N.G. (1990): Estrategias adaptativas en una comunidad subtropical de anuros. Cuadernos de Herpetologia, Serie Monografias 1: 1-70.
Caldwell, J.P. (1996): The evolution of myrmecophagy and its correlates in poison frogs (Family Dendrobatidae). Journal of Zoology 240: 75-101.
Chacón De Ulloa, P., Jaramillo G.I., Lozano, M.M. (2006): Hormigas urbanas en el departamento del Valle del Cauca, Colombia. Revista de la Academia Colombiana de Ciencias Naturales y Exactas 30: 435-441.
Da Rosa I, Canavero, A., Maneyro R., Naya D.E., Camargo A. (2002): Diet of four sympatric anuran species in a temperate environment. Boletín de la Sociedad Zoológica del Uruguay 13: 12-20.
Darst, C.R., Menendez-Guerrero P.A., Coloma L.A., Cannatella D.C. (2005): Evolution of dietary specialization and chemical defense in poison frogs (Dendrobatidae): a comparative analysis. Amererican Naturalist 165: 56-69.
Díaz-Páez, H., Ortiz J.C. (2003): Hábitos alimentarios de Pleurodema thaul (Anura, Leptodactylidae), en Concepción, Chile. Gayana 67: 25-32.
Rodrigues, D.J., Uetanabaro M., Prado C.P.A. (2004): Seasonal and ontogenetic variation in diet composition of Leptodactylus podicipinus (Anura, Leptodactylidae) in the southern Pantanal, Brazil. Revista Española de Herpetología 18: 19-28.
Duellman, W.E., Trueb, L. (1994): Biology of Amphibians. Baltimord, The Johns Hopkins University Press.
Duré, M.I., Kehr A.I. (2004): Influence of Microhabitat on the Trophic Ecology of Two Leptodactylids from Northeastern Argentina. Herpetologica, 60: 295-303.
França, L.F., Facure K.G., Giaretta A.A. (2004): Trophic and Spatial Niches of two Large-sized Species of Leptodactylus (Anura) in Southeastern Brazil. Studies on Neotropical Fauna and Environment 39: 243-248.
Gonzales–Duran, G.A., Gutierrez-Cardenas, P.D., Escobar-Laso, S. (2011): Leptodactylus fragilis (Mexican White-lipped Frog) Diet. Herpetological Review 42: 583-584.
Grant, T., Frost, D.R., Caldwell, J.P., Gagliardo, R., Haddad, C.F.B., Kok, P.J.R., Means, B.D. B., Noonan, P., Schargel, W., Wheeler, W.C. (2006): Phylogenetic systematics of dart-poison frogs and their relatives (Anura: Athesphatanura: Dendrobatidae). Bulletin of the American Museum of Natural History 299: 1-262.
Greene, H.W. (2005): Historical influences on community ecology. Proceedings of the National Academy of Sciences 102: 8395-8396.
Haddad, C.F.B., Prado. C.P.A. (2005): Reproductive Modes in Frogs and Their Unexpected Diversity in the Atlantic Forest of Brazil. BioScience 55: 207-217.
Heyer, W.R. (1969): The adaptive ecology of the species groups of the frog genus Leptodactylus (Amphibia, Leptodactylidae). Evolution 23: 421-428.
Heyer, W.R. (1978): Systematics of the fuscus group of the frog genus Leptodactylus (Amphibia, Leptodactylidae). Natural History Museum, Los Angeles Co. 29: 1-85.
Heyer, M.M., Heyer, W.R., De Sá, R.O. (2006): Leptodactylus fragilis. Catalogue of American Amphibians and Reptiles 830: 1-26.
Kamura, C.M., Morini, M., Figueiredo, C.J., Bueno, O.C., Campos-Farinha, A. (2007): Ant communities (Hymenoptera: Formicidae) in an urban ecosystem near the Atlantic Rainforest. Brazilian Journal of Biology 67: 635-641.
Lima, A.P., Magnusson, W.E. (1998): Partitioning seasonal time: interactions among size, foraging activity and diet in leaf-litter frogs. Oecologia 116: 259-266
Maneyro, R., Naya, D.E., Da Rosa, I., Canavero, A., Camargo, A. (2004): Diet of the South American frog Leptodactylus ocellatus (Anura, Leptodactylidae) in Uruguay. Iheringia, Série Zoologia 94: 57-61.
Mendez-Narvaez, J., Ospina-Sarria, J.J., Bolivar-G., W. (2009): Amphibia, Anura, Leptodactylidae, Leptodactylus fragilis: Distribution extension, Colombia. Check List 5:460-462.
Menin, M., Rossa-Feres, D.C., Giaretta, A. (2005): Resource use and coexistence of two syntopic hylid frogs (Anura, Hylidae). Revista Brasileira de Zoologia 22: 61-72.
Parmelee, J.R. (1999): Trophic Ecology of a Tropical Anuran Assemblage. Natural History Museum, University of Kansas 11: 1-59.
Peltzer, P.M., Lajmanovich, R.C. (2000): Dieta de Hyla nana (Anura: Hylidae) en charcas temporarias de la llanura aluvial del río Paraná, Argentina. Boletín de la Asociación Herpetológica Española 11: 71-73.
Pianka, E.R. (1973): The structure of lizard communities. Annual Review of Ecology and Systematics 4: 53-74.
Pianka E.R. (1974): Niche Overlap and Diffuse Competition. Proceedings of the National Academy of Sciences 71: 2141-2145.
Pinkas, L., Oliphant, M.S., Iverson, I.L., (1971): Food Habitat of albacore bluefin, tuna and bonito in California waters. Fish Bulletin 152: 1-105.
Pough, F.H., Andrews, R.M., Cadle, J.E., Crump, M.L., Savitzky, A.H., Wells, K.D. (2004): Herpetology. USA, Prentice Hall.
Sanabria, E.A., Quiroga, L.B., Acosta, J.C. (2005): Dieta de Leptodactylus ocellatus (Linnaeus, 1758) (Anura: Leptodactylidae) en un humedal del oeste de Argentina. Revista Peruana de Biología 12: 472- 477.
Santos, E.M., Almeida, A.V., Vasconcelos, S.D. (2004): Feeding habits of six anuran (Amphibia: Anura) species in a rainforest fragment in Northeastern Brazil. Iheringia, Série Zoologia 94: 433-438.
Schoener, T.W. (1974): Resource partitioning in ecological communities. Science 185: 27-39.
Simpson, E.H. (1949): Measurements of diversity. Nature 163: 688.
Strussmann, C., Ribeiro Do Vale, M.B., Meneghini, M.H., Magnusson, W.E. (1984): Diet and Foraging Mode of Bufo
Javier Mendez-Narváez et al.304

Accepted by Philip de Pous
marinus and Leptodactylus ocellatus. Journal of Herpetology 18: 138-146.
Tews, J., Brose, U., Grimm, V., Tielborger, K., Wichmann, M.C., Schwager, M. Jeltsch, F. (2004): Animal species diversity driven by habitat heterogeneity, diversity: the importance of keystone structures. Journal of biogeography 31: 79-92.
Toft, C.A. (1980): Feeding ecology of thirteen species of syntopic species. Oecologia 45: 131-141.
Toft, C.A. (1981): Feeding ecology of Panamanian litters anurans: patterns in diet and foraging mode. Journal of herpetology 15: 139-144.
Toft, C.A. (1995): Evolution of Diet Specialization in Poison-Dart Frogs (Dendrobatidae). Herpetologica 51: 202-216.
Van Sluys, M., Rocha, C.F.D. (1998): Feeding habits and microhabitat utilization by two syntopic brazilian amazonian frogs (Hyla minuta and Pseudopaludicola sp. (gr. falcipes). Revista Brasileira de Biologia 58: 559-562.
Vitt, L.J., Pianka, E. R. (2005): Deep history impacts present-day ecology and biodiversity. Proceedings of the National Academy of Sciences 102: 7877-7881.
Diet and trophic ecology of Leptodactylus fragilis and Dendropsophus columbianus 305





























