Embed Size (px)
Citation preview

Exp. Eye Res. (1995) 61, 55-62
Dietary Restriction Delays Cataract and Reduces Ascorbate Levels in Emory Mice
A L L E N T A Y L O R " , ' ~ * , J E S S I C A J A H N G E N - H O D G E a, D O N A L D E. S M I T H L V I C T O R I A J . P A L M E R L G E R A R D E. D A L L A L L RUTH D. L I P M A N L N I T A P A D H Y E b
AND B A L Z FREIcl"
USDA Human Nutrition Research Center on Aging at Tufts University, Boston, MA 02111, b Center for Clinical Cataract Research, Boston, MA 02115, ~ Harvard School of Public Health, Boston,
MA 02115, d Department of Ophthalmology and Biochemistry, Tufts University School of Medicine, Boston, MA, U.S.A.
(Received Rochester 8 August 1994 and accepted 16 February 1995)
Dietary restriction can effectively extend lifespan and retard many age-related debilities. One hypothesis to explain the beneficial effects of dietary restriction is that it prolongs maintenance of cellular homeostasis by limiting endogenous oxidative stress and preserves oxidative defense mechanisms during aging. Ascorbate, a primary antioxidant, may play a major role in preventing oxidative damage. Ascorbate levels were determined in dietary restricted (R) and control (C) Emory mice, a strain which develops age-related cataract due in part to oxidative damage to lens proteins. Mice which consumed a diet restricted by 40% in calories had lower ascorbate concentrations in plasma, liver and kidney. Nevertheless, R animals showed significantly delayed progression of cataract which extended over the entire second half of life. The R diet did not result in different ascorbate levels in the lens. Aging was associated with a decrease in ascorbate in all the examined tissues except lens of both the R and C groups. It is not clear from these data that ascorbate is a prominent factor in the delay of cataract formation or other debilities in R Emory mice. However, it also appears unlikely that lens ascorbate is cataractogenic.
© 1995 Academic Press Limited Key words: dietary restriction ; mice; cataract; ascorbate; aging; age.
1. Introduction
Restriction of dietary calorie intake (R) is the only known means to extend mean and maximal life span (Masoro, 1988). It is also associated with retardation of many age-related debilities including tumors, cardiopathy, nephropathy and decline in immune function, as well as with extended fecundity when animals are returned to higher calorie diets (Masoro, 1988; Meydani et al., 1990: Taylor et al, 1995: Weindruch et al., 1986; Weindruch and Walford, 1988). In order to elucidate how R might extend life span, evidence in support of several theories of aging is being sought within the R paradigm. One of these, the free radical theory of aging, was first proposed by Harman (1956). This theory submits that oxidative stress is caused by free radicals generated in situ, and that these oxidatively damaging species compromise the function of many biomolecules and cellular systems. In partial corroboration of a role for free radical oxidants in aging are observations that: (1) oxidative stress is associated with shortened lifespan (Pryor, 1987); (2) some age-related diseases such as cataract are induced by oxygen exposure (Giblin et al.,
* For correspondence at: USDA Human Nutrition Research Center on Aging at Tufts University, 711 Washington Street, Boston, MA 02111, U.S.A.
"~ Current address: Boston University Medical Center, Whitaker Cardiovascular Institute, Boston, MA 02118, U.S.A.
1992; Schocket et al., 1972); (3) oxidative stress is greater in organelles in which oxygen flux is greatest (Ames et al., 1993); (4) several age-related diseases are less prevalent in persons or animals with high antioxidant nutrient intake and/or plasma anti- oxidant status (Ames, Shigenaga and Hagen, 1993; Taylor, 1992; Taylor, Jacques and Dorey, 1992); and (5) upon aging some (but not all) defenses against free radicals are compromised (Koizumi, Weindruch and Walford, 1987: Laganiere and Yu. 1989; Richer, Dreyer and Rose, 1993). However, attempts to extend life span by increasing levels of antioxidants and free radical scavenging drugs were only slightly effective (Masoro, Shinokawa and Yu, 1991). Since R is also effective in decreasing lipid peroxidation and delaying age-related decrements in, or in some cases en- hancing, antioxidant capabilities (Laganiere and Yu, 1989; Weindruch and Walford, 1988), it appeared plausible that augmented levels of antioxidants may be a factor in the beneficial effects of restriction.
Ascorbate has recently been characterized as a primary biological antioxidant (Frei, England and Ames, 1989). Support for this function include the following observations: (1) in many tissues ascorbate is concentrated as much as 400-fold with respect to plasma concentrations (Taylor et al., 1991; Tsao, Leung and Young, 1987; Varma, 198 7); (2) a variety of laboratory and epidemiological studies indicate extended function of biomolecules; or organs in
00144835/95/070055 +08 $12.00/0 © 1995 Academic Press Limited

56 A. T A Y L O R ET AL.
animals or people with elevated ascorbate status (Ames et al., 1993; Taylor, 1992; Blondin et al., 1986): ( 3 ) the reducing potential of other antioxidants may be related to levels of ascorbate 0acques, Chylack and Taylor, 1994: Winkler, Orselli and Rex, 1994; Mfirtensson and Meister, 1992); and (4) in diabetic cataract, now believed to be oxidation-related, ascorbate reduces cataract and protein leakage (Linklater et al., 1990). If the expression of life observed in R animals is due to enhanced antioxidant reserves, it might be anticipated that R animals would have enhanced levels of ascorbate.
A previous study showed that dietary restriction slowed the age-related loss of soluble 7-crystallin in CH3/C57BL mice but these mice do not get cataracts (Leveille et al., 1984). The relationship between calorie-restricted diets and delayed insolubilization of crystallins was corroborated in Emory mice, a strain which does get cataracts (Mura et al., 1993 ; Taylor et al., 1989). Moreover, preliminary work showed that mild dietary restriction (21%) delayed cataract pro- gression up to 14 months in Emory mice (Taylor et al., 1989). Recently we found that dietary restriction of 40% in Emory mice delayed cataract progression substantially more than was observed previously and also resulted in life extension, delays in age-related diseases, decreases in plasma glucose and glyco- hemoglobin, and increased DNA and RNA oxidation (Taylor et al., 1995). In the study described herein, we examined more extensively the progress of cataract in the two dietary groups. We also determined levels of ascorbate in tissues from R and control (C) Emory mice and we asked if these amounts of ascorbate were associated with cataract.
2. Materials and Methods
Animals and Diet
The principles of the Declaration of Helsinki and the Guiding Principles in the Care and Use of Animals have been followed, and the protocol approved by the Animal Care and Use Committee at Tufts University. Maintenance of these Emory mice, in vivo ophthal-
mological exams (using a 0 -5 cataract grading scale as described in Table I) and study design were as described (Taylor et al., 1989, 1995) unless indicated otherwise herein. Briefly, we wanted to see if late life, or senile, cataract could be delayed beyond early adulthood by feeding Emory mice a diet restricted by 40% in dietary calories. Animals were examined monthly by an ophthalmologist who was uninformed about study objectives and identity of the animals. Tissues taken at the time of death were frozen in liquid nitrogen within 2-5 min of removal, and stored at - 8 0 ° C . These mice were bred to develop a grade 3 cataract at approximately 12 months when allowed free access to normal mouse non-purified diet.
Diets were prepared from purified ingredients (Taylor et al., 1995) as adapted from Weindruch et al. (1986) except that AIN-76A vitamin mix without ascorbate was substituted for the ICN vitamin forti- fication mix and the AIN-76 mineral mix for the USP XIV mineral mixture. Diets were analysed for protein, moisture, fat, ash, carbohydrate and caloric content by Hazelton Laboratories America (Madison, WI, U.S.A.). The C diet composition was 17.5% protein, 13"4% fat and 60"5% carbohydrate; the R diet was 29.0% protein, 14'3 % fat and 43-4% carbohydrate. C mice were fed 3.1 g day 1 (which is close to the amount consumed by Emory mice given free access to food; R. Hopkins, A. Taylor and J. Jahngen-Hodge, unpublished observations) and the R mice were fed 1.9 g day -1. This resulted in a 40% calorie restriction and a 58 % reduction in carbohydrate in R mice.
Twenty-four hours prior to termination, food was removed from the cages. Accordingly, all plasma values presented are fasting values. For termination the mice were asphyxiated by CO 2 inhalation, and blood was collected by cardiac puncture and placed on ice. The blood was immediately centrifuged at 3000 rpm for 15 min, the resulting plasma decanted and immediately prepared for ascorbate analysis.
Statistical Analysis
The progress of cataracts in the 50 C and 50 R mice was compared using non-parametric repeated
TABLE [
Cataract grading scale
Grade Description of appearance of lens by slit lamp biomicroscope and ex vivo examination
0 1
2
3
4 5
Lens is entirely transparent or has very minor opacities that do not interfere with vision (both methods) Lens has a distinct hazy area visible with a slit-lamp biomicroscope but is transparent upon illumination of the ex vivo lens
Lens contains a distinct hazy translucent area visible with a slit-lamp biomicroscope and appears as a hazy, whitish area upon illumination of the ex vivo lens
Lens contains large, opaque areas against a translucent or transparent background by slit lamp and appears white upon illumination of the ex vivo lens
Lens is almost totally opacified with few remaining translucent or transparent areas (both methods) Lens appears as an opaque mass (both methods)

DIETARY RESTRICTION, CATARACT AND ASCORBATE LEVELS 57
measures growth curve analysis (Koziol et al. 1981) as implemented in Dallal et al. (1989) which ranks each mouse by its mean cataract grade at each time point and combines the ranks from different time points to construct a global test for common cataract growth curves. It was necessary to accommodate some missing values which occurred during the course of the study when mice died or were killed. The advantage of the Koziol method over repeated measures of analysis of variance is that the Koziol test accommodates missing values without difficulty. The proportions of C and R mice having grade 5 cataract at 11 and 22 months were compared using Pearson's chi-square test for homogeneity of proportions. Mul- tiple comparison techniques, such as the Bonferroni adjustment ,were used when many age groups were analysed separately. Effects were judged to be stat- istically significant if two-tailed observed significance levels (P-values) were no greater than 0"05,
Total and reduced ascorbate concentrations in blood, kidney, liver and lens were analysed using ANOVA with sex, diet, and age as study factors (Systat Version 5, Evanston, IL, U.S.A.). The logarithmic transformation was applied to the data prior to analysis because groups with larger mean levels also had larger within-group standard deviations. The data presented in the tables were not transformed. Because there were no significant sex differences, nor were there any significant interactions involving sex for any of the response variables, sex was ignored in the analyses. In addition, there was not a significant interaction between diet and age.
Ascorbate Analysis
Plasma samples were combined with equal volumes of cold 0.35 M perchloric acid containing 0"1 mg ml- ' EDTA (PCA/EDTA), vortexed and centrifuged at 15 000 g for 3 rain at 4°C. The deproteinized super- natant was isolated and then analysed in duplicate for total and reduced ascorbic acid. Liver and kidneys were thawed at 2°C, weighed and homogenized on ice in nine volumes of PCA/EDTA and centrifuged as described above. The resulting supernatants were also analysed in duplicate. Ascorbate was analysed as described in Berger et al. (1989): total ascorbate was determined by a modification of a colorimetric pro- cedure (Roe and Keuther, 1943), and reduced ascorbate was determined using an isocratic reverse phase high performance liquid chromatography pro- cedure with electrochemical detection according to Lee et al. (1982).
Lens ascorbate concentrations were determined as described by Frei et al. (1989). Lenses were stored at -80°C , thawed and homogenized in 5% meta- phosphoric acid (MPA) and 1 mM diethylene-triamine pentaacetic acid (DTPA) (1 mg lens per 10/zl MPA- DTPA) at 4°C. Lens total ascorbate was determined
after reduction of dehydroascorbate with 1% (w/v) homocysteine. Analysis of lens ascorbate was by paired-ion reversed-phase HPLC (LC,8 column, Supelco, Bellefonte, PA, U.S.A.) coupled with elec- trochemical detection at +0 .6 V using a LC-4B amperometric electrochemical detector (Bioanalytic Systems, West Lafayette, IN, U.S,A.). Ascorbate eluted as a single peak with a retention time of approximately 8 min.
3. Results
Cataract in Emory Mice Fed R and C Diets
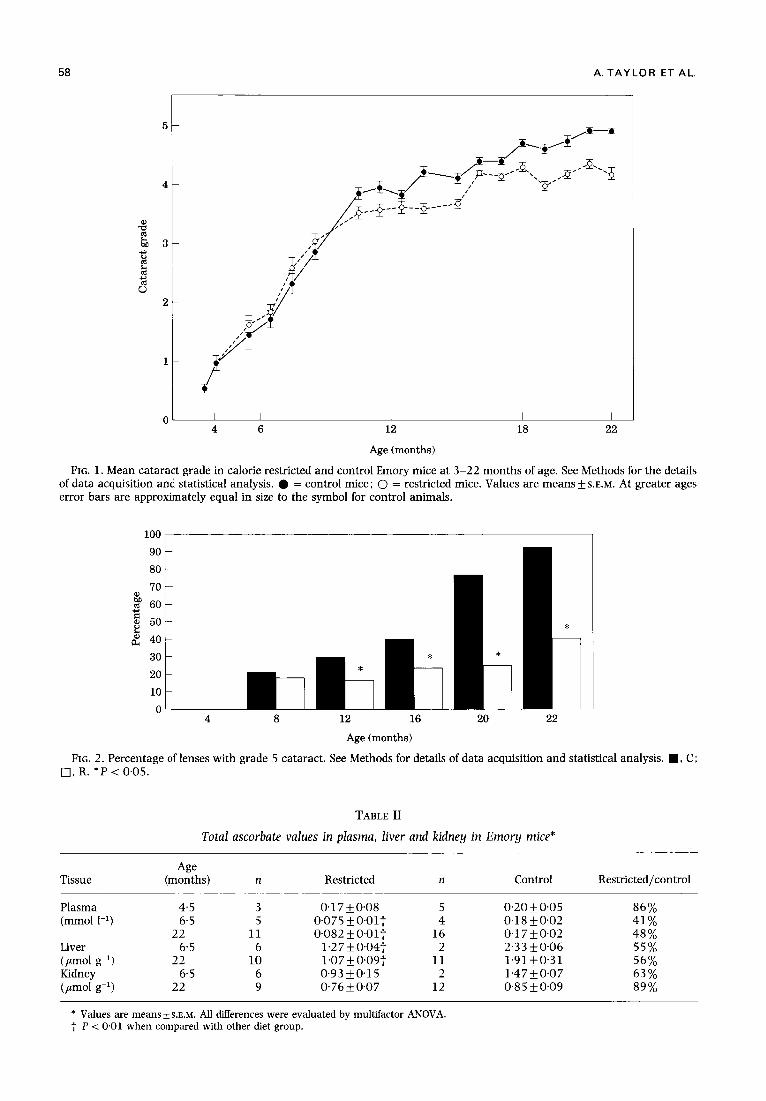
Breeding of these mice resulted in a colony of Emory mice which develops cataract at a uniform rate and in which advanced cataract is not observed to a substantial degree until after 12 months. As compared with the prior study in which calories were restricted by 21% (Taylor et al. 1989), the R group in this study consumed a diet restricted by 40% in calories. The data in Fig. 1 show that beginning at 11 months and continuing for the remainder of life, the grade of cataract in R animals was lower than in C animals (P < 0"001 for the difference between groups at all ages). Thus, at 12 months the mean grade of cataract in the R animals was 3.6 whereas in C animals it was 4'1. Greater differences in mean cataract grade were noted at other times. The diet-induced delay in rate of development of cataract becomes even clearer when the prevalence of lenses with grade 5 cataract is used as an indicator (Fig. 2). At 12 months 30% of the lenses from C mice, but only 17% of lenses from R mice had a grade 5 cataract (P = 0"05). By 22 months the discrepancy between the dietary groups widened: 93 % of the lenses from C mice had grade 5 cataract, whereas only 42 % of the lenses from R animals had such advanced opacities ( P = 0.001). Diet-related differences in cataract localization (i.e. nuclear, sub- capsular, cortical, mixed) were not obvious.
Ascorbate Concentrations in Emory Mouse Tissues
Levels of ascorbate in plasma, liver and kidney in C and R Emory mice are shown in Table II. The small sample size in the 4.5-month-old Emory mice made differences between diet groups at this age hard to distinguish. In 6.5- and 22-month-old R mice, plasma and liver ascorbate levels were only 4 1 - 5 6 % of the levels found in the C mice (Table II). Similar trends were also noted in kidney, but the difference was only marginally significant (P = 0.081).
Age-related decreases in ascorbate levels were also observed. This was particularly obvious in liver and kidney where levels of ascorbate at 22 months were approximately 16% (0.2/zmolg-1)t and 40% (0.4 #mol g-l) lower, respectively, than: at 6.5 months (Table II; liver, P = 0.068; kidney, P ~ 0.052).
58 A. TAYLOR ET AL.
~ 3
2
, ~ i ~ = / / "~'"
J /
0 I I I I I 4 6 12 18 22
Age (months)
FIG. 1. Mean cataract grade in calorie restricted and control Emory mice at 3-22 months of age. See Methods for the details of data acquisition and statistical analysis. • = control mice; O = restricted mice. Values are means_+ S.E.M. At greater ages error bars are approximately equal in size to the symbol for control animals.
100
90
80
7O
60 e. 50
40
3O
2O
10
0 4 8 12 16 20 22
Age (months)
FIG. 2. Percentage of lenses with grade 5 cataract. See Methods for details of data acquisition and statistical analysis. I , C; [2], R. *P < 0"05.
TABLE II
Total ascorbate values in plasma, liver and kidney in Emory mice*
Age Tissue (months) n Restricted n Control Restricted/control
Plasma 4.5 3 O. 17 + 0.08 5 0.20 + 0.05 869/o (mmol 1-1) 6"5 5 0-075 + 0.01"~ 4 0 '18 + 0 '02 41%
22 11 0-082 + O - O l t 16 0 . 1 7 + 0 - 0 2 48% Liver 6-5 6 1"27_+0'04"]" 2 2.33 _+0"06 55% (/zmol g 1) 22 10 1"07_+0"09t 11 1"91_+0"31 56% Kidney 6- 5 6 0-93 + O" 15 2 1"47 -+ 0"O 7 63 % (/,mol g 1) 22 9 0"76_+0"07 12 0 - 8 5 + 0 . 0 9 89%
* Values are means_+ S.mM. All differences were evaluated by multifactor ANOVA. P < 0.01 when compared with other diet group.

DIETARY RESTRICTION, CATARACT AND ASCORBATE LEVELS
TABLE III
Total and reduced ascorbate concentrations in Emory mouse lens*
59
Age Restricted (months) n (Fmol g-~)
Control (#mol g 1)
Total ascorbate 4-5 - - n.d. 9 18-29 13 0"91 +0'12 16
Reduced ascorbate 4.5 6 1-13 _ 0"15 9 18-29 13 0"77_+ 0'12"~ 16
1"08 ± 0"08 0"79 + O'06t 1-01 + 0"09 0'76_+0-07t
n.d. = not determined. *Values are means_s.E.M, in #mol (g tissue) 1. 1"P < 0.01 for comparison with other age greup.
Lens Ascorbate Levels in Emory Mice
Levels ofascorbate in the lens were five- to ten-fold greater than in plasma and were comparable to ascorbate levels in other tissues which concentrate ascorbate (compare Tables II and III; see Berger et al., 1989; Taylor et al., 1991, for data regarding other species). However, there was no statistically significant difference in lens ascorbate concentrations due to consumption of R or C diets by these mice. The data in Table III also indicate that there was a significant
"age-related decline in lens total ascorbate levels (P < 0-01). At 22 months ascorbate levels in lens were approximately 27% (0.28/~molg -1) lower than in lenses from 4.5-month-old mice. Reduced ascorbate values in Emory mouse lenses were at least 85 % of total ascorbate values and confirm the age-related decrease (P = 0"003).
4. Discussion
The Emory mouse provides one of the best models for h u m a n age-related cataract due to the gradual development of the lesion, the progression to full opacification late in life in a majority of the animals, and the many biochemical similarities to h u m a n cataract (Kuck, 1990; Taylor et al., 1989). The enhanced uniformity in rates of development of cataract achieved here increase the utility of Emory mice for intervention trials and for further studies into the mechanisms of cataractogenesis. A preliminary study, in which R Emory mice consumed a diet restricted only 21% in calories, indicated that cataract progression was delayed up to 14 months of age, at which time the study was terminated (Taylor et al. 1989). In order to maximize differences between groups, a 40% restriction resulted in a marked delay in the progress of advanced cataract until termination of the study at 22 months (Mura et al., 1993; Taylor et al., 1995). Prolonged solubility and delayed oxi- dation of crystallins in lenses from R mice also confirm the benefits of the R diet (Mura et al., 1993). It has been hypothesized that extended or enhanced anti- oxidant function is associated with the extension of life and many health advantages, including delayed cataract, observed in dietary calorie restricted animals.
Therefore it was of interest to determine if levels of ascorbate, a primary biological aqueous antioxidant, were enhanced.
Life extension of rodents of as much as 40% has been observed with 40% calorie-restricted diets (Weindruch and Walford, 1988), and the same was observed in these Emory mice (Taylor et al., 1995). Prior reports in other rodents showed no variation in ascorbate due to dietary calorie restriction (Laganiere and Yu, 1989). The major sites of murine ascorbate transport, synthesis and breakdown are plasma, liver, and kidney, respectively (Grollman and Lehninger, 1957). Levels of ascorbate in these tissues in C Emory mice (Table II) are in reasonable agreement with concentrations in liver and plasma of young Swiss Webster mice with free access to non-purified diet (Miele et al., 1989; Tsao and Young, 1989) and in plasma of guinea pigs fed a non-purified diet con- taining ascorbate (Berger et al., 1989). But in the R Emory mice, ascorbate levels in plasma, liver and kidney were considerably lower than in C animals. Thus, the extension of life and many health benefits observed in the R mice (Taylor et al., 1995) are associated with diminished rather than elevated ascorbate levels in these tissues. The decreased ascorbate levels in the R mice may reflect a limitation in the supply of carbohydrate precursors for ascorbate biosynthesis and /or enhanced utilization of ascorbate as a carbon source for synthesis of glucose (Mahler and Cordes, 1971).
Prior work indicated that ascorbate might be cataractogenic under certain circumstances (Bensch, Fleming and Lohmann, 1985). Since there were no differences in lens ascorbate levels between R and C mice at any age (Table III), it is unlikely that the prolonged lens clarity observed in the R mice is exclusively associated with the level of this moiety in the lens. A lack of association between lens ascorbate and cataract in mice is also suggested by the observation that ascorbate levels in these Emory mice were slightly higher than in eyes of Swiss Webster mice which do not develop opacities (Tsao et al., 1987) and by the absence of cataract in mice fed 8 % of their diet as ascorbate (Bensch et al., 1985; also see below). (While no specific lens measurement was made in the Swiss Webster mice, the lens, vitreous and

60 A. TAYLOR ET AL,
aqueous comprise the majority of the eye tissue, and ascorbate concentrations are usually comparable in all three compartments. Accordingly, it can be surmised that lens levels would be very similar to eye ascorbate levels.)
The significant age-related decline in lens total ascorbate (P = 0.008) is consistent with an attenu- ation in ascorbate levels in livers and kidneys. Comparable age-related decrements in ascorbate con- centrations were i~oted in lens and plasma in guinea pigs after consumption of diets which provided saturating levels of ascorbate (Berger et al., 1989) and in liver of Fisher 34:4 rats fed purified diets (Laganiere and Yu, 1989).
Lower ascorbate levels in R mice may also involve differential cycling of ascorbate and other anti- oxidants, including glutathione (GSH) and /or vitamin E. Recent reports indicate that elimination of dietary vitamin E enhanced the severity of cataracts in Emory mice (Varma et al., 1994). Unfortunately no lenticular vitamin E levels could be obtained in that or this study. GSH levels in kidney [ ~ 1"5/zmol (g protein) 1] and lens [ ~ 1 #mol (g protein) -1] were not significantly related to age or diet. The absence of a correlation between age and lens GSH data appear to be in contrast with the previously noted inverse relationship between cataract progress and GSH levels in lenses of Emory mice (Kack and Kuck, 1983) and pre- cataractous diabetic rats (Mitton et al., 1993). By 22 months, both reduced and total glutathione levels were 48 % higher in livers from R animals than from C mice (Taylor et al., 1995). The apparent main- tenance of liver GSH levels in R animals does not appear to be due to elevated ascorbate since ascorbate levels are lower in R than C mice. Recent data indicate that in liver and kidney, there are no significant diet- or age-related differences in the enzymatic activities of various antioxidant enzymes, such as catalase, gluta- thione peroxidase, Cu/Zn- and Mn-superoxide dis- mutase (Gong et al., manuscript in preparation).
Many health benefits (extended mean lifespan, delayed tumorigenesis and collagen cross-linking, prolonged immune function, diminished plasma glu- cose and glycohemoglobin, and continued regulation of plasma glucose upon aging, etc.) previously associated with calorie-restricted diets are also observed in R Emory mice (Taylor et al. 1995). Together with these data regarding extension of lens function, the dietary restriction paradigm becomes even more attractive and intriguing, and the need to elucidate the mechanisms by which these benefits are obtained becomes more compelling. Since ascorbate levels are either lower or not different in tissues of R versus C animal, it appears that the life extension observed within the calorie restriction paradigm in these animals is achieved by mechanisms other than enhanced ascorbate levels.
The lack of an association between lens ascorbate levels and delayed cataract in Emory mice is in
apparent contrast with a majority of epidemiologic studies (Jacques et al., 1994; Robertson et al., 1989: Taylor, 1992 ; Taylor et al., manuscript in preparation) which indicate relationships between enhanced ascorbate intake and/or plasma ascorbate status and diminished odds ratio for cataract. But none of these studies have correlated prevalence of cataract with lens levels of ascorbate. The present results are also not in agreement with many cell-flee and animal experiments which indicate that enhanced ascorbate levels confer protection against lens protein damage similar to that found in cataract (Taylor, 1992; Blondin et al., 1986; Blondin and Taylor, 1987).
These data beg questions as to why lens clarity is prolonged in the R animals. Recent studies indicate that glycation of lens proteins is associated with cataract, and that ascorbate as well as various sugars are involved in such glycation (Liang and Rossi, 1990; Nagaraj et al., 1991). That ascorbate is not a primary glycating agent in the Emory mouse is supported by the data noted above. The possibility that differential rates of cataract development in C versus R lenses may be due to glycation induced by higher glucose levels in C mice should be considered in future studies, since plasma glucose concentrations in these C mice were significantly higher than in R mice (C/R = 120%, Taylor et al., 1995). Future studies should also examine ascorbate and GSH levels in lenses with various grades of cataract in similarly aged animals from the R and C diet groups as well as interactions between these antioxidants.
Acknowledgements
The authors thank Drs Casilda Mura, Fu Shang and Martin Obin for critical review and comments, and Timi Mannion, Gayle Perrone, Doug Shepard, Gudbjorn Asmundsson and Matthew Wlodarczyk for technical as- sistance. ].].-H. thanks Dana C. Lipp for many helpful discussions.
This research was supported in part by U.S. Department of Agriculture, Agricultural Research Service contract #53- 3K06-0-1 (A.T.), National Eye Institute grant EY-086 (A. T.), as well as grant-in-aid 13-28-901 from the American Heart Association, Massachusetts Affiliate, Inc. (B.F.). The contents of this publication do not necessarily reflect the views or policies of the U.S. Department of Agriculture, nor does mention of trade names, commercial products, or organizations imply endorsement by the U.S. Government.
Portions of this data were presented by Taylor, A., Cyr, D., Lipman, R. D., Laxman, E., Smith, D., Palmer, V., Kidder, C., Shepard, D., Morrow, F. & Padhye, N. (1989). Effect of food restriction on incidence of cataracts, glycohemoglobin, and tail collagen integrity. Proc. Fifth Congress on U.S.-Japan Cooperative Cataract Research Group, 153.
References
Ames, B.N., Shigenaga, M.K. and Hagen, T.M. (1993). Oxidants, antioxidants, and the degenerative disease of aging. Proc. Natl. Acad. Sci., USA 90, 7915-22.

DIETARY RESTRICTION, CATARACT AND ASCORBATE LEVELS 61
Bensch, K. G., Fleming, J. E. and Lohmann, W. (1985). The role of ascorbic acid in senile cataract. Proc. Natl. Acad. Sci., USA 82, 7193-6.
Berger, J., Shepard, D., Morrow, F. and Taylor, A. (1989). Relationships between dietary intake and tissue levels of reduced and total vitamin C in the nonscorbutic guinea pig. ]. Nutr. 119, 7 3 4 4 0 .
Blondin, J., Baragi, V., Schwartz, E., Sadowski, ]. A. and Taylor, A. (1986). Delay of UV-induced eye lens protein damage in guinea pigs by dietary ascorbate. Free Radical Biol. Meal. 2, 275-81.
Blondin, ]. and Taylor, A. (1987). Measures of leucine aminopeptidase can be used to anticipate UV-induced age-related damage to lens proteins: ascorbate can delay this damage. Mech. Ageing Dev. 41, 39-46.
Dallal, G.E., Guo, S., Simon, R., Talmadge, J.E. and Klabansky, R.L. (1989). MUDIFT: multivariate distribution-free comparison of growth curves. The American Statistician 43, 125-34.
Frei, B., England, L. and Ames, B. N. (1989). Ascorbate is an outstanding antioxidant in human blood plasma. Proc. Natl. Acad. Sci., USA 86, 6377-81.
Giblin, F. J., Padgaonkar, V. A., Lin, L.-R., Dang, L., Leverenz, V. R., Lou, M. and Reddy, V. N. (1992). Hyperbaric 02 in vivo increases disulfide content and nuclear light scattering in lenses of guinea pigs. Invest. Ophthalmol. Vis. Sci. 33 (Suppl.), 779.
Grollman, A.P. and Lehninger, A.L. (1957). Enzymic synthesis of L-ascorbic acid in different animal species. Arch. Biochem. Biophys. 69, 458-67.
Harman, D. (1956). Aging: A theory based on free radical and radiation chemistry. J. Gerontol. 11, 298-300.
Jacques, P. F., Chylack, L. and Taylor, A. (1994). Relation- ships between cataract formation and natural anti- oxidants. In Natural Antioxidants in Human Health and Disease. (Ed. Frei, B.). Pp. 515-33. Academic Press: New York, U.S.A.
Koizumi, A., Weindruch, R. and Walford, R.L. (1987). Influences of dietary restriction and age on liver enzyme activities and lipid peroxidation in mice. ]. Nutr. 117, 361-7.
Koziol, ]. A., Maxwell, D. A., Fukushima, M., Colmerauer, M. E. M. and Pilch, Y. H. (1981). A distribution-free test for tumor-growth curve analyses with application to an animal tumor immunotherapy experiment. Biometrics 37, 383-90.
Kuck, J. F. R. (1990). Late onset hereditary cataract of the Emory mouse. A model for human senile cataract. Exp. Eye Res. 50, 659-64.
Kuck, ]. F. R. and Kuck, K.D. (1983). The Emory mouse cataract: loss of soluble protein, glutathione, protein sulfhydryl and other changes. Exp. Eye Res. 36, 352-62.
Laganiere, S. and Yu, B. P. (1989). Effect of chronic food restriction in aging rats. Mech. Ageing Dev. 48, 221-30.
Lee, W., Hamernyik, P., Hutchinson, M., Raisys, V. A. and Labbe, R. F. (1982). Ascorbic acid in lymphocytes: cell preparation and liquid chromatography assay. Clin. Chem. 28, 2165-9.
Leveille, P.J., Weindruch, R., Walford, R.L., Bok, D. and Horwitz, ]. (1984). Dietary restriction retards age- related loss of gamma crystallins in the mouse lens. Science 214, 1247-50.
Liang, J. N. and Rossi, M. T. (1990). In vitro non-enzymatic glycation and formation of browning products in the bovine lens ~-crystallin. Exp. Eye Res. 50, 367-71.
Linklater, H. A., Dzialoszynski, T., McLeod, H. L., Sanford, S.E. and Trevithick, J.R. (1990). Modelling cortical cataractogenesis. XI. Vitamin C reduces 7-crystallin leakage from lenses in diabetic rats. Exp. Eye Res. 51, 241-7.
Mahler, H. R. and Cordes, E. H. (1971). Biological Chemistry. Pp. 584-5. Harper and Row: New York, U.S.A.
M~rtensson, J. and Meister, A. (1992). Glutathione deficiency increases hepatic ascorbic acid synthesis in adult mice. Proc. Natl. Acad. Sci., USA 89, 11566-8.
Masoro, E.J. (1988). Food restriction in rodents: an evaluation of its role in the study of aging. ]. Gerontol. 43, B59-64.
Masoro, E.J., Shinokawa, I. and Yu, B.P. (1991). Re- tardation of the aging processes in rats by food restriction. In Physiological Senescence and its Post- ponement: Theoretical Approaches and Rational Inter- ventions. (Eds Pierpaoli, W. and Fabris, N.). Vol. 621. Pp. 337-52. New York Academy of Science: New York, U.S.A.
Meydani, S. N., Lipman, R., Blumberg, J. B. and Taylor, A. (1990). Dietary energy restriction decreases ex-vivo spleen prostaglandin E2 synthesis in Emory mice. J. Nutr. 120, 112-15.
Miele, M., Desole, M.S., Demontis, M.P., Enrice, P., DeNatale, G., Varoni, M.V. and Anania, V. (1989). Effect of adriamycin and d-amphetamine on endogen- ous levels of ascorbic acid in rodents. Pharm. Res. 21, 13-14.
Mitton, K. P., Dean, P. A. W., Dzialoszynski, T., Xiong, H., Sanford, S.E. and Trevithick, J.R. (1993). Modelling cortical cataractogenesis 13. Early effects on lens ATP/ADP and glutathione in the streptozotocin rat model of the diabetic cataract. Exp. Eye Res. 56, 187-98.
Mura, C. V., Roh, S., Smith, D., Palmer, V., Padhye, N. and Taylor, A. (1993). Cataract incidence and analysis of lens crystallins in water-, urea-, and SDS-soluble fractions of Emory mice fed a diet restricted 40 % in calories. Curr. Eye Res. 12, 1081-91.
Nagaraj, R. H., Sell, D. R., Prabhakaram, M., Ortwerth, B. ]. and Monnier, V. M. (1991). High correlation between pentosidine protein crosshnks and pigmentation impli- cates ascorbate oxidation in human lens senescence and cataractogenesis. Proc. Natl. Acad. Sci., USA 88, 10257-61.
Pryor, W.A. (1987). The free radical theory of aging revisited: a critique and a suggested disease-specific theory. In Modern Biological Theories of Aging. (Eds Warner, H. R. et al.). P. 93. Raven Press: New York, U.S.A.
Richer, S., Dreyer, R. and Rose, R.C. (1993). Spatial distribution and redox state of ascorbic acid in rabbit lens. Invest. Ophthalmol. Vis. Sei. 54 (Suppl.), 1283.
Robertson, J. M., Donner, A. P. and Trevithick, J. R. (1989). Vitamin E intake and risk of cataracts in humans. Ann. N.Y. Acad. Sci. 570, 372-82.
Roe, ]. H. and Keuther, C. A. (1943). The determination of ascorbic acid in whole blood and urine through the 2,4- dinitrophenylhydrazine derivative dehydroascorbic acid. J. Biol. Chem. 147, 399-403.
Schocket, S. S., Esterson, J., Bradford, B., Michaelis, M. and Richards, R. D. (1972). Induction of cataracts in mice by exposure to oxygen. Israel J. Med. 8, 1596-8.
Taylor, A. (1992). Role of nutrients in delaying cataracts. In Beyond Deficiency: New Views on the Function and Health Effects of Vitamins. (Eds Sauberlich, H. E. and Machlin, L. ].). Vol. 669. Pp. 111-24. New York Academy of Science: New York, U.S.A.
Taylor, A., Lipman, R. D., Jahngen-Hodge, J., Palmer, V., Smith, D., Padhye, N., Dallal, G.. E., Cyr, D. E., Laxman, E., Shepard, D., Morrow, F., Sallomon, R., Perrone, G., Asmundsson, G., Meydani, M., ]Mune, M., Harrison, D. E., Archer, J.R. and Shigenaga, M. (1995). Dietary calorie restriction in the Emory mouse: effects on

62 A. T A Y L O R ET AL.
lifespan, eye lens cataract prevalence and progression, levels of ascorbate, glutathione, glucose, and glyco- hemoglobin, tail collagen breaktime, DNA and RNA oxidation, skin integrity, fecundity, and cancer. Mecha- nisms of Ageing and Development (in press).
Taylor, A., Jacques, P. F. and Dorey, C. K. (1992). Oxidation and aging: impact on vision. In Antioxidants: Chemical, Physiological, Nutritional and Toxicological Aspects. (Ed. Williams, G.M.). Pp. 349-71. Princeton Scientific: Princeton, U.S.A.
Taylor, A., Jacques, P. F., Nadler, D., Morrow, F., Sulsky, S. I. and Shepard, D. (1991). Relationship in humans between ascorbic acid consumption and levels of total and reduced ascorbic acid in lens, aqueous humor, and plasma. Curr. Eye Res. 10, 751-9.
Taylor, A., Zuliani, A.M., Hopkins, R.E., Dallal, G.E., Treglia, P., Kuck, J. F. R. and Kuck, K. (1989). Moderate calorie restriction delays cataract formation in the Emory mouse. FASEB ] 3, 1741-6.
Tsao, C. S., Leung, P.Y. and Young, M. (1987). Effect of dietary ascorbic acid intake on tissue vitamin C in mice. ]. Nutr. 117, 291-7.
Tsao, C.S. and Young, M. (1989). Effect of exogenous ascorbic acid intake on biosynthesis of ascorbic acid in mice. Life Sci. 4, 1553-7.
Weindruch, R, and Walford, R. L. (1988). The Retardation of Aging and Disease by Dietary Restriction. Charles C. Thomas: Springfield, U.S.A.
Weindruch, R., Walford, R.L., Fligiel, S. and Guthrie, D. (1986). The retardation of aging in mice by dietary restriction: longevity, cancer, immunity and lifetime energy intake. ]. Nutr. 116, 641-54.
Winkler, B. S., Orselli, S. M. and Rex, T. S. (1994). The redox couple between glutathione and ascorbic acid: A chemical and physiological perspective. Free Radic. Biol. Med. 4, 3 3 3 4 9 .
Varma, S. D. (1987). Ascorbic acid and the eye with special reference to the lens. In Third Conference on Vitamin C. (Eds Burns, J.J., Rivers, I.M. and Machlin, L. J.). Vol. 498. Pp. 281-306. New York Academy of Sciences: New York, U.S.A.
Varma, S. D., Devamanoharan, P. S., Mansour, S. and Teter, B. (1994). Studies on Ernory mouse cataracts: oxidative factors. Ophthalmic Res. 26, 141-8.





























