Embed Size (px)
Citation preview

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Feb. 2011, p. 1301–1308 Vol. 77, No. 40099-2240/11/$12.00 doi:10.1128/AEM.01662-10Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Different Substrate Recognition Requirements for Cleavage ofSynaptobrevin-2 by Clostridium baratii and Clostridium botulinum
Type F Neurotoxins�
Suzanne R. Kalb,1 Jakub Baudys,2 Christina Egan,3 Theresa J. Smith,4 Leonard A. Smith,4James L. Pirkle,1 and John R. Barr1*
Centers for Disease Control and Prevention, National Center for Environmental Health, Division of Laboratory Sciences,4770 Buford Hwy. N.E., Atlanta, Georgia 303411; Battelle Memorial Institute at the Centers for Disease Control and
Prevention, Atlanta, Georgia2; Wadsworth Center, New York State Department of Health, Albany,New York 122083; and Integrated Toxicology, United States Army Medical Research Institute of
Infectious Diseases (USAMRIID), Ft. Detrick, Maryland 217024
Received 12 July 2010/Accepted 4 December 2010
Botulinum neurotoxins (BoNTs) cause botulism, which can be fatal if it is untreated. BoNTs cleave proteinsnecessary for nerve transmission, resulting in paralysis. The in vivo protein target has been reported for allseven serotypes of BoNT, i.e., serotypes A to G. Knowledge of the cleavage sites has led to the development ofseveral assays to detect BoNT based on its ability to cleave a peptide substrate derived from its in vivo proteintarget. Most serotypes of BoNT can be subdivided into subtypes, and previously, we demonstrated that threeof the currently known subtypes of BoNT/F cleave a peptide substrate, a shortened version of synaptobrevin-2,between Q58 and K59. However, our research indicated that Clostridium baratii type F toxin did not cleave thispeptide. In this study, we detail experiments demonstrating that Clostridium baratii type F toxin cleavesrecombinant synaptobrevin-2 in the same location as that cleaved by proteolytic F toxin. In addition, wedemonstrate that Clostridium baratii type F toxin can cleave a peptide substrate based on the sequence ofsynaptobrevin-2. This peptide substrate is an N-terminal extension of the original peptide substrate used fordetection of other BoNT/F toxins and can be used to detect four of the currently known BoNT/F subtypes bymass spectrometry.
Botulism is caused by intoxication with a potent neurotoxinknown as botulinum neurotoxin (BoNT) and is a disease thatcan be fatal if it is untreated. Botulism can be contracted byingestion of food containing the toxin (11, 33), by colonizationof bacteria in the gastrointestinal tract in infants or susceptibleindividuals, by inhalation of the toxin, or by contact of bacteriawith a wound (11). Treatment of botulism involves administer-ing a therapeutic immunoglobulin product and is most effectivewhen performed within 24 h of exposure (11). Due to theextreme toxicity, availability, and ease of preparation of BoNT,it is considered a likely agent for bioterrorism (4).
The neurotoxin molecule is composed of three functionaldomains—the receptor-binding domain, the translocation do-main, and the enzymatic domain. The receptor-binding do-main interacts with ectoacceptors on target cell surfaces (25).The toxin is taken up into endosomes, where a conformationalchange in the translocation domain allows for the formation ofpores (24). These pores enable the enzymatic domain to enterthe cytoplasm of target neuronal cells, where docking proteins(SNARE proteins) necessary for nerve transmission are cleaved.This inhibits the nerve impulse, resulting in a flaccid paralysis
that can affect the lungs and may necessitate ventilator sup-port.
BoNTs are produced by some species of the genus Clos-tridium, in particular Clostridium botulinum, C. butyricum, C.baratii, and C. argentinense. These toxins are divided into se-rotypes A through G based on their serological properties.C. botulinum strains produce toxins of types A through F, andC. argentinense strains produce type G toxin. Some strains of C.butyricum produce type E toxin, and some C. baratii strainsproduce type F toxin. Four BoNT serotypes (A, B, E, and F)are commonly associated with human botulism. Each serotypecan be divided further into four to six toxin subtypes. WithinBoNT/F strains, proteolytic (23, 26, 44) and nonproteolytic (12,13, 26) BoNT/F strains and bivalent BoNT/Af toxin-producingstrains (15, 17) have been isolated from environmental samplesand food-borne botulism cases. To date, all cases associatedwith bivalent Bf strains have been in infants (5). BoNT/F-expressing Clostridium baratii strains have been responsible forboth infant and, presumably, food-borne botulism (18, 19).However, with C. baratii botulism, there is only tenuous evi-dence, if any at all, linking cases with causative foods, and whilemultiple members of a household may have consumed com-mon foodstuffs, only targeted members became ill (18). Thisinformation indicates that the immune status of the patientmay be a major factor in susceptibility to botulism due to C.baratii.
Different toxin serotypes have different enzymatic targets.BoNT/A, -C, and -E cleave synaptosome-associated protein
* Corresponding author. Mailing address: Centers for Disease Con-trol and Prevention, National Center for Environmental Health, Divi-sion of Laboratory Sciences, 4770 Buford Hwy. N.E., Atlanta, GA30341. Phone: (770) 488-7848. Fax: (770) 488-0509. E-mail: [email protected].
� Published ahead of print on 17 December 2010.
1301
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 01
Feb
ruar
y 20
22 b
y 2.
188.
218.
5.

(SNAP-25) (7, 8, 16, 28, 29, 43), whereas BoNT/B, -D, -F, and-G cleave synaptobrevin-2 (VAMP-2) (27, 31, 45, 46). OnlyBoNT/C is known to cleave more than one protein, i.e.,SNAP-25 and syntaxin (9, 16, 32). Each toxin serotype cleavesits target at a unique place in the molecule. Knowledge of thecleavage sites has led to the development of several assays todetect BoNT based on its ability to cleave a peptide substratederived from its in vivo protein target (3, 6, 10, 14, 20, 34–36,41, 42). Previously, our laboratory reported the developmentof an assay for BoNT detection and serotype differentiation,termed the Endopep-MS method (6, 10). This method detectsall BoNT serotypes, BoNT/A through BoNT/G, and involvesincubating BoNT with a peptide substrate that mimics BoNT’snatural, in vivo target. Each BoNT cleaves its peptide substratein a specific location, and that location is different for eachBoNT serotype. The reaction mixture is then introduced into amass spectrometer, which detects and accurately reports themasses of any peptides within the mixture. Detection of pep-tide cleavage products corresponding to the specific toxin-de-pendent location indicates the presence of a particular BoNTserotype.
Proteolytic BoNT/F (protBoNT/F) cleaves synaptobrevin-2between Q58 and K59 (30). The BoNT/F subtypes containsubstantial amino acid sequence differences from protBoNT/F,which may affect receptor binding and the cleavage of targetSNARE proteins. In previous work, we demonstrated thatthe enzymatic activities of most BoNT/A, -B, -E, and -Fsubtypes were equivalent in both location and potency (22).As part of that study, we confirmed that protBoNT/F, non-proteolytic BoNT/F (npBoNT/F), and several bivalent BoNT/F(bvBoNT/F) subtypes all cleaved their peptide substrate inthe same location (22). However, our research indicated thatBoNT/F from C. baratii strains (BoNT/FC. baratii) did not cleavethis peptide.
In the current study, we detail experiments which demon-strate that Clostridium baratii type F toxin cleaves synaptobre-vin-2 in the same location as that cleaved by proteolytic Ftoxin. Failure to cleave our standard synaptobrevin-2 pep-tide was found to be due to amino acid differences in andnear multiple exosites between protBoNT/F and BoNT/FC. baratii. Extension of the N terminus of the synaptobrevin-2peptide enabled cleavage of the substrate target by BoNT/FC. baratii subtypes. Use of this extended synaptobrevin-2 pep-tide allows detection of four of the currently known BoNT/Fsubtypes by mass spectrometry.
MATERIALS AND METHODS
Materials. Botulinum neurotoxin is very toxic and therefore requires appro-priate safety measures. All neurotoxins were handled in a level 2 biosafetycabinet equipped with HEPA filters. Commercially purified BoNT/B and prot-BoNT/F were purchased from Metabiologics (Madison, WI). Synaptobrevin-2recombinant protein was purchased from GenWay Biotech, Inc. (San Diego,CA). Monoclonal antibody 4E17.1 was obtained from James Marks at the Uni-versity of California at San Francisco. Protein G Dynabeads were purchasedfrom Invitrogen (Carlsbad, CA) at 1.3 g/cm3 in phosphate-buffered saline (PBS),pH 7.4, containing 0.1% Tween 20 and 0.02% sodium azide. All chemicalswere from Sigma-Aldrich (St. Louis, MO), except where indicated. Peptidesubstrates were synthesized by Los Alamos National Laboratory (LosAlamos, NM) (Fig. 1).
Production and characterization of Clostridium baratii type F culture super-natants. Clostridium baratii type F organisms from the Wadsworth Center Cul-ture Collection were cultured in 5 ml of Trypticase-peptone-glucose-yeast extract
(TPGY) broth prepared in-house at 35°C for approximately 3 days. Anaerobicconditions were created by using an Oxoid AnaeroGen atmosphere generationsystem (Basingstoke, United Kingdom). The supernatants were then filtered byusing an Acrodisc 0.22-�m syringe filter purchased from Pall Corporation (PortWashington, NY). Fifty microliters of filtered supernatant was cultured on sheepblood agar plates prepared in-house and incubated under the conditions de-scribed above to ensure that there were no viable organisms present in thesupernatants. The culture supernatants were tested at the Wadsworth Center byusing a mouse bioassay to determine the presence of type F toxin (38).
BoNT extraction. Monoclonal antibody 4E17.1 was immobilized and cross-linked to protein G Dynabeads as previously described (21, 22), by using 40 �gof antibody diluted in 500 �l of PBS for every 100 �l of protein G Dynabeads.Cross-linked IgG-coated Dynabeads were made fresh daily. An aliquot of 20 �lof antibody-coated beads was mixed for 1 h with a solution of 500 �l of culturesupernatant mixed with 50 �l of 10� phosphate-buffered saline with 0.01%Tween 20 (PBST), with 500 mouse 50% lethal doses (mLD50) of protBoNT/Fmixed with 500 �l of PBST, or with 24,000 mLD50 of BoNT/B mixed with 500 �lof PBST. After being mixed for 1 h with constant agitation at room temperature,the beads were washed twice with 1 ml (each time) of PBST and then once with100 �l of water. Negative controls consisted of PBST or blank culture superna-tant medium with no spiked toxin. The remainder of the extraction protocol wasas described above. The commercially purified toxins did not require antibodyextraction, but antibody extraction was performed with all samples to ensure thatall toxins were exposed to the same procedures to eliminate differential proce-dures as a source of inaccurate results. Levels of the commercially purified toxinswere chosen in order to achieve a reaction in which the cleavage products andunmodified substrate were clearly visible. The level of the C. baratii toxin in theculture used is not known.
Incubation of BoNT with peptide or protein target. The incubation of BoNTwith a target was performed as previously described (21), with a few modifica-tions. In all cases, a final reaction volume of 20 �l was added to the beads; thefinal concentration of the reaction buffer was 0.05 M HEPES (pH 7.3), 25 mMdithiothreitol, and 20 �M ZnCl2. For peptide reactions, the final concentrationof the peptide substrate was 50 pmol/�l, and the peptide sequences are listed inFig. 1. For recombinant synaptobrevin-2 reactions, the final concentration ofsynaptobrevin-2 was 250 ng/�l. All samples were incubated at 37°C for 4 h.
FIG. 1. Amino acid sequence of the recombinant synaptobrevin-2substrate utilized in and produced by cleavage of BoNT/F and -B. Theoriginal BoNT/F substrate is in red, with the new substrate in red andpurple. The observed m/z of each peptide or protein is also included.NT, N-terminal; CT, C-terminal.
1302 KALB ET AL. APPL. ENVIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 01
Feb
ruar
y 20
22 b
y 2.
188.
218.
5.

Peptide mass spectrometric detection. A 2-�l aliquot of each reaction super-natant was mixed with 18 �l of matrix solution consisting of alpha-cyano-4-hydroxycinnamic acid (CHCA) at 5 mg/ml in 50% acetonitrile, 0.1% trifluoro-acetic acid (TFA), and 1 mM ammonium citrate. A 0.5-�l aliquot of this mixturewas pipetted onto each spot of a 192-spot matrix-assisted laser desorption ion-ization (MALDI) plate (Applied Biosystems, Framingham, MA). Mass spectrafor each spot were obtained by scanning from 1,100 to 4,800 m/z in mass spec-trometry (MS) positive-ion reflector mode on an Applied Biosystems 4800 pro-teomic analyzer (Framingham, MA). The instrument uses an Nd-YAG laser at355 nm, and each spectrum is an average of 2,400 laser shots.
Protein mass spectrometric detection. All reaction mixtures were first sepa-rated by using a nanoACQUITY UltraPerformance LC (UPLC) system (Waters,Milford, MA). Mobile phases were 0.04% TFA with 0.06% formic acid (FA) inwater (mobile phase A) and 0.04% TFA and 0.06% FA in acetonitrile (mobilephase B). Synaptobrevin-2 and cleavage products were trapped at 500 ng on aPepswift PS-DVB monolithic trapping column (200 �m by 5 mm; Dionex, Sunny-vale, CA) and then washed for 4 min at a flow rate of 7.5 �l/min with 99% mobilephase A. Intact synaptobrevin-2 and cleavage products were eluted and sepa-rated by using a 70-min reverse-phase (RP) gradient at 750 nl/min (1 to 50%mobile phase B over 35 min) on a Pepswift PS-DVB monolithic nanoscale LCcolumn (100 �m by 5 cm; Dionex). The column temperature was set to 60°C.
A NanoMate TriVersa instrument (Ithaca, NY) was used for infusion andonline LC coupling analysis of the samples at a capillary spray voltage of 1.82 kV.The mass spectral data were acquired on a Synapt HDMS quadrupole time offlight (QTOF) instrument (Waters); the instrument was calibrated for a massrange of 550 to 4,550 m/z with cesium iodide through direct infusion. Thesampling and extraction cone voltages were optimized at 40 V and 4 V, respec-tively, for maximum intact synaptobrevin-2 sensitivity, by comparing on-columninjections. Source temperature was set to 150°C. A quadrupole RF transmissionprofile was defined for transmit masses from 800 to 5,000 Da. Trap and transfercollision energies were set to 6 V and 2 V, respectively, for maximum transmis-sion of the most abundant synaptobrevin-2 charge state. The data were acquiredin TOF V mode at a mass range of 700 to 2,500 m/z and a 2-scan/s acquisitiontime. All data were processed by using Waters MassLynx MaxEnt 1 software toobtain the deconvoluted mass at a range of 3,000 to 20,000 Da, with a massresolution of 0.5 Da. All spectra were processed with a uniform Gaussian damagemodel with an iterate to convergence option selected.
RESULTS
C. baratii type F does not cleave a peptide substrate recog-nized by other type F toxins. Three of the currently knownsubtypes of BoNT/F cleave a peptide substrate based on thesequence of synaptobrevin-2 (22). This peptide substrate hasthe sequence LQQTQAQVDEVVDIMRVNVDKVLERDQKLSELDDRADAL, with cleavage by BoNT/F occurring be-tween the bolded and underlined Q and K. These data (Fig. 2Bto D) and a negative control (Fig. 2A) are depicted by theirmass spectra in Fig. 2. Following toxin extraction and concen-tration of BoNT/FC. baratii, the peptide substrate remained un-altered as depicted in Fig. 2E, indicating that BoNT/FC. baratii
does not cleave this peptide substrate.C. baratii type F cleaves synaptobrevin-2 as other type F
toxins do. Because BoNT/FC. baratii did not cleave the peptidesubstrate as expected, its in vivo protein target was investi-gated. Schiavo and coauthors demonstrated that protBoNT/Fcleaved the protein synaptobrevin-2 when BoNT/F was incu-bated with rat synaptic vesicles (30). Because other testedsubtypes had the same cleavage site under in vitro conditions(22), we theorized that BoNT/FC. baratii had the same cleavagesite as the other type F toxins under in vitro conditions. To testthis theory, full-length recombinant synaptobrevin-2 needed tobe detected by mass spectrometry. Following extraction usingblank PBST buffer, antibody-coated beads were incubated withsynaptobrevin-2 in the presence of reaction buffer. The super-natant of this reaction was analyzed by QTOF mass spectrom-
etry. Figure 3 shows the deconvoluted mass spectrum of thisreaction. The peak at mass 13,824 corresponds to intact full-length recombinant synaptobrevin-2.
ProtBoNT/F was then extracted with antibody-coated beadsand incubated with full-length synaptobrevin-2. The massspectra obtained from this reaction are shown in Fig. 4.Figure 4A shows the deconvoluted mass spectrum producedon the QTOF instrument used to analyze larger-molecular-weight fragments. The peak at mass 13,823 corresponds tointact synaptobrevin-2, similar to that in Fig. 3. However, Fig.4A shows a second peak of interest at mass 10,343, whichcorresponds to cleavage of synaptobrevin-2 by protBoNT/F.
After examination of the amino acid sequence of recombi-nant synaptobrevin-2 (Fig. 1), it was determined that the peakat mass 10,343 in Fig. 4A corresponds to the N-terminal cleav-age product from the 95th residue of recombinant synaptobre-vin-2, or cleavage between glutamine 95 and lysine 96. Thisfinding confirms the cleavage location for protBoNT/F re-ported by Schiavo and coauthors (30). Figure 4B shows themass spectrum of protBoNT/F incubated with synaptobrevin-2.
FIG. 2. MALDI mass spectra of Endopep-MS BoNT/F reactionsfor no toxin (A), protBoNT/F (B), npBoNT/F (C), bvBoNT/F (D), andBoNT/FC. baratii (E) extracted using 4E17.1 antibody-coated beads. Thepeptide cleavage products indicating that BoNT/F is present are shownby peaks at m/z 1,345.8 and 3,168.4 (marked with asterisks), and thedoubly charged peptide substrate peak is present at m/z 2,248.9. Thepeak at m/z 1,345.8 is much larger than that at m/z 3,168.4, whichionizes poorly and as a result is less visible by mass spectrometry. %Int, % intensity.
VOL. 77, 2011 C. BARATII F CLEAVES SYNAPTOBREVIN-2 1303
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 01
Feb
ruar
y 20
22 b
y 2.
188.
218.
5.
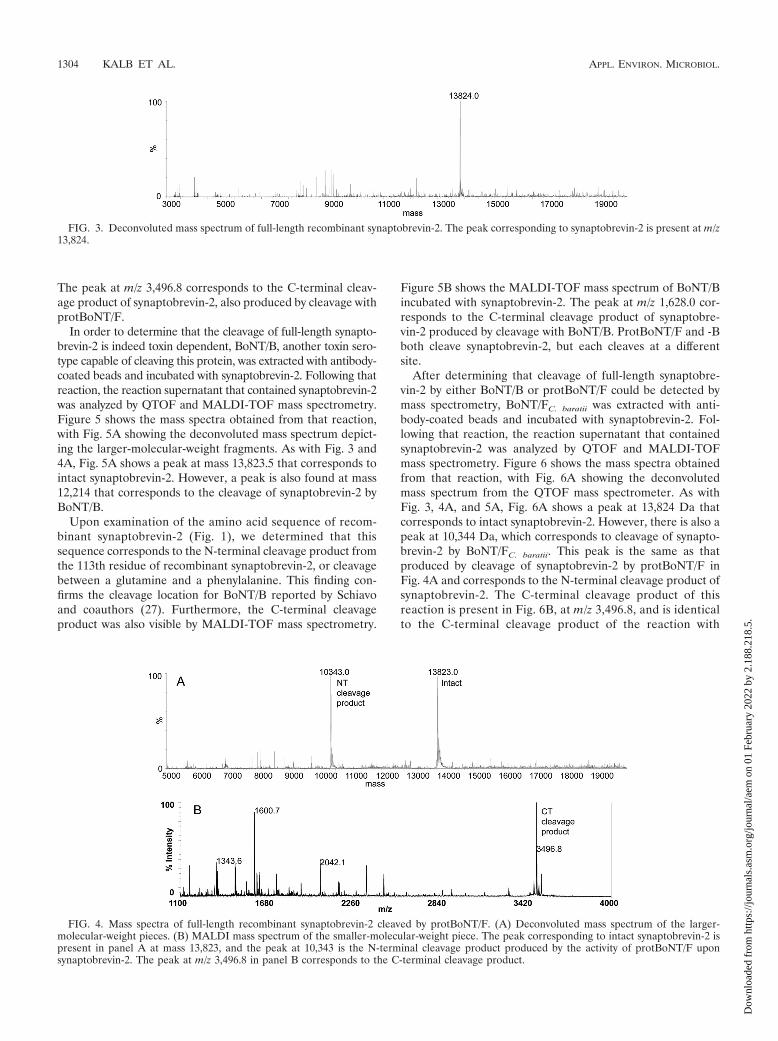
The peak at m/z 3,496.8 corresponds to the C-terminal cleav-age product of synaptobrevin-2, also produced by cleavage withprotBoNT/F.
In order to determine that the cleavage of full-length synapto-brevin-2 is indeed toxin dependent, BoNT/B, another toxin sero-type capable of cleaving this protein, was extracted with antibody-coated beads and incubated with synaptobrevin-2. Following thatreaction, the reaction supernatant that contained synaptobrevin-2was analyzed by QTOF and MALDI-TOF mass spectrometry.Figure 5 shows the mass spectra obtained from that reaction,with Fig. 5A showing the deconvoluted mass spectrum depict-ing the larger-molecular-weight fragments. As with Fig. 3 and4A, Fig. 5A shows a peak at mass 13,823.5 that corresponds tointact synaptobrevin-2. However, a peak is also found at mass12,214 that corresponds to the cleavage of synaptobrevin-2 byBoNT/B.
Upon examination of the amino acid sequence of recom-binant synaptobrevin-2 (Fig. 1), we determined that thissequence corresponds to the N-terminal cleavage product fromthe 113th residue of recombinant synaptobrevin-2, or cleavagebetween a glutamine and a phenylalanine. This finding con-firms the cleavage location for BoNT/B reported by Schiavoand coauthors (27). Furthermore, the C-terminal cleavageproduct was also visible by MALDI-TOF mass spectrometry.
Figure 5B shows the MALDI-TOF mass spectrum of BoNT/Bincubated with synaptobrevin-2. The peak at m/z 1,628.0 cor-responds to the C-terminal cleavage product of synaptobre-vin-2 produced by cleavage with BoNT/B. ProtBoNT/F and -Bboth cleave synaptobrevin-2, but each cleaves at a differentsite.
After determining that cleavage of full-length synaptobre-vin-2 by either BoNT/B or protBoNT/F could be detected bymass spectrometry, BoNT/FC. baratii was extracted with anti-body-coated beads and incubated with synaptobrevin-2. Fol-lowing that reaction, the reaction supernatant that containedsynaptobrevin-2 was analyzed by QTOF and MALDI-TOFmass spectrometry. Figure 6 shows the mass spectra obtainedfrom that reaction, with Fig. 6A showing the deconvolutedmass spectrum from the QTOF mass spectrometer. As withFig. 3, 4A, and 5A, Fig. 6A shows a peak at 13,824 Da thatcorresponds to intact synaptobrevin-2. However, there is also apeak at 10,344 Da, which corresponds to cleavage of synapto-brevin-2 by BoNT/FC. baratii. This peak is the same as thatproduced by cleavage of synaptobrevin-2 by protBoNT/F inFig. 4A and corresponds to the N-terminal cleavage product ofsynaptobrevin-2. The C-terminal cleavage product of thisreaction is present in Fig. 6B, at m/z 3,496.8, and is identicalto the C-terminal cleavage product of the reaction with
FIG. 3. Deconvoluted mass spectrum of full-length recombinant synaptobrevin-2. The peak corresponding to synaptobrevin-2 is present at m/z13,824.
FIG. 4. Mass spectra of full-length recombinant synaptobrevin-2 cleaved by protBoNT/F. (A) Deconvoluted mass spectrum of the larger-molecular-weight pieces. (B) MALDI mass spectrum of the smaller-molecular-weight piece. The peak corresponding to intact synaptobrevin-2 ispresent in panel A at mass 13,823, and the peak at 10,343 is the N-terminal cleavage product produced by the activity of protBoNT/F uponsynaptobrevin-2. The peak at m/z 3,496.8 in panel B corresponds to the C-terminal cleavage product.
1304 KALB ET AL. APPL. ENVIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 01
Feb
ruar
y 20
22 b
y 2.
188.
218.
5.
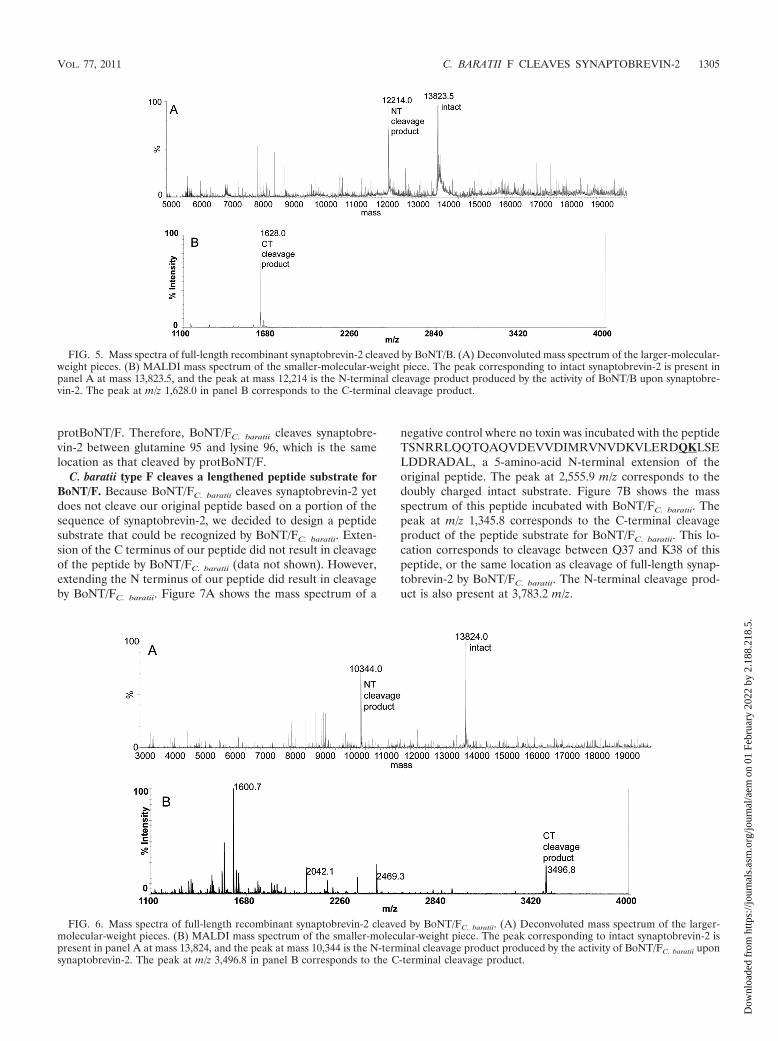
protBoNT/F. Therefore, BoNT/FC. baratii cleaves synaptobre-vin-2 between glutamine 95 and lysine 96, which is the samelocation as that cleaved by protBoNT/F.
C. baratii type F cleaves a lengthened peptide substrate forBoNT/F. Because BoNT/FC. baratii cleaves synaptobrevin-2 yetdoes not cleave our original peptide based on a portion of thesequence of synaptobrevin-2, we decided to design a peptidesubstrate that could be recognized by BoNT/FC. baratii. Exten-sion of the C terminus of our peptide did not result in cleavageof the peptide by BoNT/FC. baratii (data not shown). However,extending the N terminus of our peptide did result in cleavageby BoNT/FC. baratii. Figure 7A shows the mass spectrum of a
negative control where no toxin was incubated with the peptideTSNRRLQQTQAQVDEVVDIMRVNVDKVLERDQKLSELDDRADAL, a 5-amino-acid N-terminal extension of theoriginal peptide. The peak at 2,555.9 m/z corresponds to thedoubly charged intact substrate. Figure 7B shows the massspectrum of this peptide incubated with BoNT/FC. baratii. Thepeak at m/z 1,345.8 corresponds to the C-terminal cleavageproduct of the peptide substrate for BoNT/FC. baratii. This lo-cation corresponds to cleavage between Q37 and K38 of thispeptide, or the same location as cleavage of full-length synap-tobrevin-2 by BoNT/FC. baratii. The N-terminal cleavage prod-uct is also present at 3,783.2 m/z.
FIG. 5. Mass spectra of full-length recombinant synaptobrevin-2 cleaved by BoNT/B. (A) Deconvoluted mass spectrum of the larger-molecular-weight pieces. (B) MALDI mass spectrum of the smaller-molecular-weight piece. The peak corresponding to intact synaptobrevin-2 is present inpanel A at mass 13,823.5, and the peak at mass 12,214 is the N-terminal cleavage product produced by the activity of BoNT/B upon synaptobre-vin-2. The peak at m/z 1,628.0 in panel B corresponds to the C-terminal cleavage product.
FIG. 6. Mass spectra of full-length recombinant synaptobrevin-2 cleaved by BoNT/FC. baratii. (A) Deconvoluted mass spectrum of the larger-molecular-weight pieces. (B) MALDI mass spectrum of the smaller-molecular-weight piece. The peak corresponding to intact synaptobrevin-2 ispresent in panel A at mass 13,824, and the peak at mass 10,344 is the N-terminal cleavage product produced by the activity of BoNT/FC. baratii uponsynaptobrevin-2. The peak at m/z 3,496.8 in panel B corresponds to the C-terminal cleavage product.
VOL. 77, 2011 C. BARATII F CLEAVES SYNAPTOBREVIN-2 1305
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 01
Feb
ruar
y 20
22 b
y 2.
188.
218.
5.

This peptide substrate was also tested with other knownsubtypes of BoNT/F to ensure that they also cleave this peptidesubstrate. Figure 7C shows the mass spectrum of the reactionof protBoNT/F with the peptide substrate. The peak at m/z1,345.8 corresponds to the C-terminal cleavage product of thepeptide substrate for protBoNT/F and is identical to the C-terminal cleavage product produced by BoNT/FC. baratii. Thispeak is also present in Fig. 7D and E, which correspond to thereactions of npBoNT/F and bvBoNT/F, respectively. There-fore, four of the currently known subtypes of BoNT/F cleavethis new peptide substrate.
DISCUSSION
In previous work, our laboratory demonstrated that theactivities of most noncommercially available subtypes on apeptide substrate were the same as those of the commer-cially available toxins (22). For instance, we determined thatthe activities of BoNT/A2, -A3, and -A4 were the same as thatof BoNT/A1. This is important information, as the originalexperiments that discovered BoNT’s target proteins and cleav-age sites were performed with only one subtype of toxin perserotype, and it was not known if all subtypes would have the
same target. The current experiments detail that four subtypesof BoNT/F do indeed have the same protein target and cleav-age site, as those BoNT/F subtypes cleave synaptobrevin-2 inthe same location. However, differences in peptide substraterecognition sites (exosites) exist among the various BoNT/Fsubtypes, and these differences translate to an inability tocleave our 39-amino-acid peptide.
Several publications have addressed binding of synaptobre-vin-2 to protBoNT/F. James Schmidt et al. showed that apeptide including residues A37 to S75 of synaptobrevin-2 gen-erated effective cleavage by protBoNT/F, between 58Q and59K (35). Schmidt et al. later reported that one or more resi-dues from L32 to Q36 of synaptobrevin-2 are important forsubstrate binding and cleavage by protBoNT/F (37) and alsonoted that residues R66 to S75 are not required for substraterecognition and cleavage by protBoNT/F. Therefore, the opti-mal substrate for recognition and cleavage by protBoNT/F wasfound to be residues L32 to D65 of synaptobrevin-2. A 39-amino-acid peptide substrate consisting of residues L32 to L70of synaptobrevin-2 was tested in our laboratory and wasrecognized and cleaved by protBoNT/F, npBoNT/F, andbvBoNT/F but not by BoNT/FC. baratii.
Several publications have also analyzed the binding of syn-aptobrevin-2 to protBoNT/F through either structural analysisvia X-ray crystallography (1, 2) or point mutation of synapto-brevin-2 (40). Point mutations indicated that 14 amino acidsbetween residues T27 and Y88 are important for cleavage byprotBoNT/F, with critical amino acids beginning at residueQ33. X-ray crystallography results indicated that synaptobre-vin-2 makes contact with protBoNT/F mainly through residuesE41 to D65 (1). Residues Q33 to E41 of synaptobrevin-2 alsointeract with protBoNT/F (2) but are not critical for binding,as point mutations in this region do not affect the activity ofprotBoNT/F upon the peptide substrate (40). Additional in-teractions with protBoNT/F are reported for residues N25 toN29 of synaptobrevin-2; however, these interactions are re-ported as weak, and including these residues has little effect onincreasing the binding affinity (2).
It is clear from previously published studies and the exper-imental results presented here that the interaction of prot-BoNT/F with synaptobrevin-2 is different from that of BoNT/FC. baratii with synaptobrevin-2. Table 1 illustrates that BoNT/Fsubtypes differ by as much as 31.4% in overall amino acidcomposition. BoNT/FC. baratii is the most divergent, with aminoacid differences ranging from 26.3% to 31.4%, versus 7.6% to16.6% among the other subtypes (Table 1). Examination ofamino acid identities versus protBoNT/F by domain (Table 2)indicates that the translocation and receptor-binding domains
FIG. 7. MALDI mass spectra of the updated peptide substrate inEndopep-MS BoNT/F reactions for no toxin (A), BoNT/FC. baratii (B),protBoNT/F (C), npBoNT/F (D), and bvBoNT/F (E) extracted using4E17.1 antibody-coated beads. The peptide cleavage products indicat-ing that BoNT/F is present are shown by the peaks at m/z 1,345.8 and3,783.2 (marked with asterisks), and the doubly charged peptide sub-strate peak is present at m/z 2,555.9.
TABLE 1. BoNT/F subtype amino acid identities for four currentlyknown subtypes
Subtype
% Amino acid identity with subtype
Proteolytic F Nonproteolytic F C. baratii F BoNTAf-F
BoNTBf-F
Proteolytic F 87.4 73.7 92.4 83.4Nonproteolytic F 87.4 69.8 86.9 89.8C. baratii F 73.7 69.8 71.9 68.6BoNT Af-F 92.4 86.9 71.9 83.7BoNT Bf-F 83.4 89.8 68.6 83.7
1306 KALB ET AL. APPL. ENVIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 01
Feb
ruar
y 20
22 b
y 2.
188.
218.
5.

are both fairly conserved, with amino acid identities of 78.9%or better. The highest degree of divergence between BoNT/FC. baratii and protBoNT/F is seen within the enzymatic domain,where more than one-third of all amino acids differ, comparedwith fewer than 20% with any other BoNT/F subtype (Table 2).BoNT subtypes typically do not display such differences, asother subtypes have enzymatic domain identities of �81%(data not shown). Indeed, an amino acid identity of only 63%is quite close to the overall identity between differing sero-types, such as 63% between BoNT/E and -F and 57% betweenBoNT/B and -G. With such a high degree of divergence in theenzymatic domain, it is not surprising that BoNT/FC. baratii
interacts with its substrate in a different manner from that ofprotBoNT/F.
These large amino acid differences in BoNT/FC. baratii ac-count for its inability to bind a peptide substrate based on thesequence of synaptobrevin-2 from residues L32 to L70. Of theamino acids in protBoNT/F which are reported to be impor-tant for interactions with synaptobrevin-2, there are manywhich are mutated in BoNT/FC. baratii. Figure 8 lists the resi-
dues of synaptobrevin-2 from L32 to D57 and their reportedcontact with corresponding residues of protBoNT/F (2), withadded sequence alignments of npBoNT/F, bvBoNT/F, andBoNT/FC. baratii. Of the 26 amino acids in BoNT/FC. baratii pre-dicted to be contacted by residues L32 to D57 of synaptobre-vin-2, half are significantly altered from those in protBoNT/F.Six of the amino acids from L32 to D57 of synaptobrevin-2—Q33, V39, E41, V43, L54, and D57—reduce the cleavability ofsynaptobrevin-2 by more than 66% when mutated, demon-strating that these residues are critical for binding of synapto-brevin-2 to protBoNT/F (40). Five of those six residues contactmutated residues on BoNT/FC. baratii, which could explain whyresidues L32 to L70 of synaptobrevin-2 cannot bind BoNT/FC. baratii effectively.
All currently known subtypes of BoNT/F cleave a peptidesubstrate based on the sequence of synaptobrevin-2 fromresidues T27 to L70, and residues T27 to R31 (TSNRR) arenot necessary for binding to protBoNT/F, npBoNT/F, orbvBoNT/F. However, these residues are critical for binding toBoNT/FC. baratii. Residues T27 and N29 of TSNRR in synap-tobrevin-2 may play important roles in the binding of synap-tobrevin-2 to BoNT/FC. baratii. Residue T27 of synaptobrevin-2is reported to contact Y319 and W322 of protBoNT/F (2). Asindicated in Fig. 8, W322 is conserved in BoNT/FC. baratii
(W314), so T27 of synaptobrevin-2 could contact BoNT/FC. baratii via W314. In addition, N29 of synaptobrevin-2 isreported to contact Y316 of protBoNT/F (2). This residue isalso conserved in BoNT/FC. baratii, as Y308, so N29 of synap-tobrevin-2 could contact BoNT/FC. baratii via Y308. ResiduesR30 and R31 of synaptobrevin-2 could also play a role inbinding to BoNT/FC. baratii. Both of these residues are disor-dered in the crystal structure and therefore could not be mod-eled (2).
TABLE 2. Amino acid identities of three BoNT/F subtypes toprotBoNT/F for each domain of toxin, along with the strain
identification for each subtype listed
Subtype Strain
% Amino acid identity
Holotoxin Enzymaticdomain
Translocationdomain
Receptor-bindingdomain
Proteolytic F Langeland 100 100 100 100Nonproteolytic F Eklund 202F 87.4 94.3 87.0 80.9Clostridium
baratii FSullivan 73.7 63.3 79.2 78.9
BoNT Af-F 84 92.4 96.4 90.9 89.8BoNT Bf-F An436 83.4 81.8 86.3 82.3
FIG. 8. Amino acid contacts between residues 27 and 57 of synaptobrevin-2 and protBoNT/F. Sequence alignments with npBoNT/F, bv-BoNT/F, and BoNT/FC. baratii are also included, with mutations shown in red and conserved mutations shown in green. The figure includes datafrom reference 1.
VOL. 77, 2011 C. BARATII F CLEAVES SYNAPTOBREVIN-2 1307
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 01
Feb
ruar
y 20
22 b
y 2.
188.
218.
5.

The discovery that BoNT/FC. baratii requires a longer peptidesubstrate for cleavage than other BoNT/F subtypes is impor-tant to many assays that use activity to detect and differentiateBoNTs. Detection of BoNT via its activity upon a peptidesubstrate was first reported in 1994 (39), and many reportshave followed, some of which report lower limits of detectionthan those of the mouse bioassay, in a shorter time. Asubstrate consisting of residues L32 to D65 of synaptobre-vin-2 can be used to detect three of the currently knownsubtypes of BoNT/F. However, this peptide cannot be usedto detect BoNT/FC. baratii. Therefore, it is possible to use thedifferent responses of BoNT/FC. baratii toward the peptide sub-strates to identify the subtype of toxin as BoNT/FC. baratii basedon an activity assay.
ACKNOWLEDGMENT
The opinions, interpretations, and recommendations are those ofthe authors and are not necessarily those of the Centers for DiseaseControl and Prevention or the U.S. Army.
REFERENCES
1. Agarwal, R., T. T. Binz, and S. Swaminathan. 2005. Structural analysis ofbotulinum neurotoxin serotype F light chain: implications on substrate bind-ing and inhibitor design. Biochemistry 44:11758–11765.
2. Agarwal, R., J. J. Schmidt, R. G. Stafford, and S. Swaminathan. 2009. Modeof VAMP substrate recognition and inhibition of Clostridium botulinumneurotoxin F. Nat. Struct. Mol. Biol. 16:789–794.
3. Anne, C., F. Cornille, C. Lenoir, and B. P. Roques. 2001. High-throughputfluorogenic assay for determination of botulinum type B neurotoxin proteaseactivity. Anal. Biochem. 291:253–261.
4. Arnon, S. S., et al. 2001. Botulinum toxin as a biological weapon: medical andpublic health management. JAMA 285:1059–1070.
5. Barash, J. R., and S. S. Arnon. 2004. Dual toxin-producing strain of Clos-tridium botulinum type Bf isolated from a California patient with infantbotulism. J. Clin. Microbiol. 42:1713–1715.
6. Barr, J. R., et al. 2005. Botulinum neurotoxin detection and differentiationby mass spectrometry. Emerg. Infect. Dis. 11:1578–1583.
7. Binz, T. J., et al. 1994. Proteolysis of SNAP-25 by types E and A botulinalneurotoxins. J. Biol. Chem. 269:1617–1620.
8. Blasi, J., et al. 1993. Botulinum neurotoxin A selectively cleaves the synapticprotein SNAP-25. Nature 365:160–163.
9. Blasi, J., et al. 1993. Botulinum neurotoxin C1 blocks neurotransmitterrelease by means of cleaving HPC-1/syntaxin. EMBO J. 12:4821–4828.
10. Boyer, A. E., et al. 2005. From the mouse to the mass spectrometer: detectionand differentiation of the endoproteinase activities of botulinum neurotoxinsA-G by mass spectrometry. Anal. Chem. 7:3916–3924.
11. Centers for Disease Control and Prevention. 1998. Botulism in the UnitedStates: 1899–1996. Handbook for epidemiologists, clinicians and laboratoryworkers. Centers for Disease Control and Prevention, Atlanta, GA.
12. Condit, P. K., and H. A. Renteln. 1966. Epidemiologic notes and reports,botulism—California. MMWR Morb. Mortal. Wkly. Rep. 15:355–356.
13. Eklund, M. W., F. T. Poysky, and D. I. Wieler. 1967. Characteristics ofClostridium botulinum type F isolated from the Pacific Coast of the UnitedStates. Appl. Microbiol. 15:1316–1323.
14. Ekong, T. A. N., I. M. Feavers, and D. Sesardic. 1997. RecombinantSNAP-25 is an effective substrate for Clostridium botulinum type A toxinendopeptidase activity in vitro. Microbiology 143:2227–2247.
15. Fernandez, R. A., A. S. Ciccarelli, G. N. Arenas, and D. F. Gimenez. 1986.First outbreak of botulism caused by Clostridium botulinum subtype Af. Rev.Argent. Microbiol. 18:29–31.
16. Foran, P., G. W. Lawrence, C. C. Shone, K. A. Foster, and J. O. Dolly. 1996.Botulinum neurotoxin C1 cleaves both syntaxin and SNAP-25 in intact andpermeabilized chromaffin cells: correlation with its blockade of catechol-amine release. Biochemistry 35:2630–2636.
17. Gimenez, D. F., and A. S. Ciccarelli. 1978. New strains of Clostridium botu-linum subtype Af. Zentralbl. Bakteriol. Orig. A 240:215–220.
18. Gupta, A., C. J. Sumner, M. Castor, S. Maslanka, and J. Sobel. 2005. Adultbotulism type F in the United States, 1981–2002. Neurology 65:1694–1700.
19. Hall, J. D., L. M. McCroskey, B. J. Pincomb, and C. L. Hatheway. 1985.Isolation of an organism resembling Clostridium baratii which produces type
F botulinal toxin from an infant with botulism. J. Clin. Microbiol. 21:654–655.
20. Hallis, B., B. A. H. James, and C. C. Shone. 1996. Development of novelassays for botulinum type A and B neurotoxins based on their endopeptidaseactivities. J. Clin. Microbiol. 34:1934–1938.
21. Kalb, S. R., et al. 2006. The use of Endopep-MS for the detection ofbotulinum neurotoxins A, B, E, and F in serum and stool samples. Anal.Biochem. 351:84–92.
22. Kalb, S. R., et al. 2008. The use of Endopep-MS to detect multiple subtypesof botulinum neurotoxins A, B, E, and F. Int. J. Mass Spectrom. 278:101–108.
23. Moller, V., and I. Scheibel. 1960. Preliminary report on the isolation of anapparently new type of Cl. botulinum. Acta Pathol. Microbiol. Scand. 48:80.
24. Montal, M. S., R. Blewitt, J. M. Tomich, and M. Montal. 1992. Identificationof an ion channel-forming motif in the primary structure of tetanus andbotulinum neurotoxins. FEBS Lett. 313:12–18.
25. Poulain, B., et al. 1988. Neurotransmitter release is blocked intracellularly bybotulinum neurotoxin, and this requires uptake of both toxin polypeptides bya process mediated by the larger chain. Proc. Natl. Acad. Sci. U. S. A.85:4090–4094.
26. Raphael, B. H., M. J. Choudoir, C. Luquez, R. Fernandez, and S. E.Maslanka. 2010. Sequence diversity of genes encoding botulinum neurotoxintype F. Appl. Environ. Microbiol. 76:4805–4812.
27. Schiavo, G., et al. 1992. Tetanus and botulinum B neurotoxins block neuro-transmitter release by proteolytic cleavage of synaptobrevin. Nature 359:832–835.
28. Schiavo, G., et al. 1993. Identification of the nerve terminal targets ofbotulinum neurotoxin serotypes A, D, and E. J. Biol. Chem. 268:23784–23787.
29. Schiavo, G., et al. 1993. Botulinum neurotoxins serotypes A and E cleaveSNAP-25 at distinct COOH-terminal peptide bonds. FEBS Lett. 335:99–103.
30. Schiavo, G., C. C. Shone, O. Rosetto, F. C. Alexander, and C. Montecucco.1993. Botulinum neurotoxin serotype F is a zinc endopeptidase specific forVAMP/synaptobrevin. J. Biol. Chem. 268:11516–11519.
31. Schiavo, G., et al. 1994. Botulinum G neurotoxin cleaves VAMP/synapto-brevin at a single Ala-Ala peptide bond. J. Biol. Chem. 269:20213–20216.
32. Schiavo, G., C. C. Shone, M. K. Bennett, R. H. Scheller, and C. Montecucco.1995. Botulinum neurotoxin type C cleaves a single Lys-Ala bond within thecarboxyl-terminal region of syntaxins. J. Biol. Chem. 270:10566–10570.
33. Schiavo, G., M. Matteoli, and C. Montecucco. 2000. Neurotoxins affectingneuroexocytosis. Physiol. Rev. 80:717–766.
34. Schmidt, J. J., and K. A. Bostian. 1995. Proteolysis of synthetic peptides bytype A botulinum neurotoxin. J. Protein Chem. 14:703–708.
35. Schmidt, J. J., R. G. Stafford, and C. B. Millard. 2001. High-throughputassays for botulinum neurotoxin proteolytic activity: serotypes A, B, D, andF. Anal. Biochem. 296:130–137.
36. Schmidt, J. J., and R. G. Stafford. 2003. Fluorogenic substrates for theprotease activities of botulinum neurotoxins, serotypes A, B, and F. Appl.Environ. Microbiol. 69:297–303.
37. Schmidt, J. J., and R. G. Stafford. 2005. Botulinum neurotoxin serotype F:identification of substrate recognition requirements and development ofinhibitors with low nanomolar affinity. Biochemistry 44:4067–4073.
38. Shayegani, M., W. B. Stome, and G. E. Hannett. 1984. An outbreak ofbotulism in waterfowl and fly larvae in New York State. J. Wildl. Dis.20:86–89.
39. Shone, C. C., and A. K. Roberts. 1994. Peptide substrate specificity andproperties of the zinc-endopeptidase activity of botulinum type B neuro-toxin. Eur. J. Biochem. 225:263–270.
40. Sikorra, S., T. Henke, T. Galli, and T. Binz. 2008. Substrate recognitionmechanism of VAMP/synaptobrevin-cleaving clostridial neurotoxins. J. Biol.Chem. 283:21145–21152.
41. Wictome, M., et al. 1999. Development of in vitro assays for the detection ofbotulinum toxin in foods. FEMS Immunol. Med. Microbiol. 24:319–323.
42. Wictome, M., et al. 1999. Development of an in vitro bioassay for Clostridiumbotulinum type B neurotoxin in foods that is more sensitive than the mousebioassay. Appl. Environ. Microbiol. 65:3787–3792.
43. Williamson, L. C., J. L. Halpern, C. Montecucco, J. E. Brown, and E. A.Neale. 1996. Clostridial neurotoxins and substrate proteolysis in intact neu-rons: botulinum neurotoxin C acts on synaptosomal-associated protein of 25kDa. J. Biol. Chem. 271:7694–7699.
44. Williams-Walls, N. J. 1968. Clostridium botulinum type F: isolation fromcrabs. Science 162:375–376.
45. Yamasaki, S., et al. 1994. Cleavage of members of the synaptobrevin/VAMPfamily by types D and F botulinal neurotoxins and tetanus toxin. J. Biol.Chem. 269:12764–12772.
46. Yamasaki, S., et al. 1994. Botulinum neurotoxin type G proteolyses theAla81-Ala82 bond of rat synaptobrevin 2. Biochem. Biophys. Res. Commun.200:829–835.
1308 KALB ET AL. APPL. ENVIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 01
Feb
ruar
y 20
22 b
y 2.
188.
218.
5.