Embed Size (px)
Citation preview

Cell Tiss. Res. 189, 109-130 (1978) Cell and Tissue Research �9 by Springer-Verlag 1978
Different Types of Small Granule-Containing Cells and Neurons in the Guinea-Pig Adrenal Medulla*
K. Unsicker, O. Habura-F1/Jh and U. Zwarg
Department of Anatomy, University of Kiel, F.R.G.
Summary. An electron microscopic, histo- and biochemical study was carried out on the adrenal medulla of newborn and adult guinea-pigs giving special emphasis to small granule-containing (SGC) cells. Adrenaline (A) was the predominating catecholamine (CA) both in newborn (70-90 ~ of total CA) and adult (85-90 ~o) guinea-pig adrenals. In analogy to the biochemical findings electron microscopy revealed a high predominance of A cells, which contained large granular vesicles with an average diameter of 180 nm. Most noradrenaline (NA) storing cells showed granular vesicles of a considerably smaller average diameter (80 nm) and had a higher nuclear-cytoplasmic ratio. These cells were termed SGC-NA cells. NA cells with large granular vesicles (average diameter 170nm) were extremely rare. Another type of SGC cells contained granular vesicles with cores of low to medium electron-density (SGC-NA-negative cells). Biochemical determinations made it unlikely that these cells contained predominantly dopamine (DA). SGC cells were scarcely innervated by cholinergic nerves. They formed processes, which were found both in the adrenal cortex and medulla contacting blood vessels including sinusoid capillaries, steroid producing cells of the reticularis and fasciculata zone and processes, which were interpreted to belong to medullary nerve cells.
Two types of neurons were present in the guinea-pig adrenal medulla, one resembling the principal neurons in sympathetic ganglia, the other, which, according to its morphology, occupied an intermediate position between principal neurons and SGC cells.
In adrenomedullary grafts under the kidney capsule, which were studied three weeks after transplantation, "ordinary" A cells resembled SGC-NA negative cells with respect to their ultramorphology. Processes of transplanted principal neurons showed uptake of 5-hydroxydopamine and, hence, were
Send offprint requests to: Professor Dr. Klaus Unsicker, Anatomisches Institut der Universit~it Kid, Olshausenstr. 40-60, 2300 Kiel, F.R.G.
* Supported by a grant from Deutsche Forschungsgemeinschaft (Un 34/4)
Dedicated to Professor H. Leonhardt in honor of his 60th birthday.
0302-766X/78/0189/0109/$04.40

110 K. Unsicker et al.
considered to be adrenergic. Despite the lack of extrinsic nerves to the transplants, few principal neurons received cholinergic synapses, the origin of which is uncertain to date.
Key words: Guinea-pig adrenal medulla - Small granule-containing cells - Neurons - Electron and fluorescence microscopy - Catecholamines.
Introduction
Small granule-containing (SGC) cells have been described to occur in a large number of sympathetic ganglia (Grillo, 1966; Williams, 1967; Elfvin, 1968; Siegrist et al., 1968; Jacobowiz, 1970; Watanabe, 1971 ; Yokota, 1973). They are most likely identical to a catecholamine (CA)-storing population of small intensely fluorescent (SIF) cells (see Grillo et al., 1974) first reported by Er/ink6 and H/irkonen (1963). Several types of SGC cells appear to exist, as judged from their ultramorphology (Watanabe, 1971; Dail et al., 1975; Elfvin et al., 1975; Lu et al., 1976), histochemistry (Santer et al., 1975), and response to certain drug treatments (Heym, 1975, 1976; Heym and Grube, 1975). Although the functional significance of SGC cells is still under discussion, one type, which gives rise to processes making efferent synapses with presumed dendrites of principal sympathetic neurons (Matthews and Raisman, 1969), has been proposed to modulate synaptic transmission causing the inhibitory postsynaptic potentials in sympathetic ganglia (Libet and Owman, 1974). Dopamine (DA) has been identified in some SIF cells (Bj6rklund et al., 1970), and it has been suggested that DA is the amine involved in intraganglionic inhibition (Libet and Tosaka, 1970) by activating a DA-sensitive adenyl cyclase (Kebabian and Greengard, 1971). The adrenal medulla is a modified sympathetic ganglion, and, hence, the occurrence of cell types similar to those in sympathetic ganglia might be expected. There are two types of cells in the mammalian adrenal medulla, which are related specifically to the two medullary hormones adrenaline (A) and noradrenaline (NA) (Er/ink6, 1955), and, in addition, a few neurons (Unsicker, 1967; Diner, 1968). Cells with the features of SGC cells have not been described in the adrenal medulla of adult mammals until recently (Coupland et al., 1977; Kobayashi and Coupland, 1977). However, adrenal glands of lower vertebrates display a larger variety of cell types, some of which resemble SGC cells in mammals (Unsicker, 1973a--c; 1976). This report deals with SGC cells and neurons nerve cells in the adrenal medulla of the guinea-pig both in situ and in transplants under the kidney capsule and includes some biochemical data which might be useful for an interpretation of these cells.
Materials and Methods
Electron Microscopy. Newborn and adult guinea-pigs (weighing 250-400 gm) were anesthetized with ether and perfused via the heart or descending aorta with 150 ml of phosphate-buffered (0.1 M) 3.5 % glutaraldehyde. Adrenal glands were removed, sliced, kept in the same solution for up to 2 h and rinsed in phosphate buffer for at least 2 h. After postfixation in 2 % aqueous OsO4, slices were either block-stained in a saturated aqueous solution of uranyl acetate or directly dehydrated through a graded series of alcohols. Araldite-embedded tissues were sectioned on a Reichert-Sitte OmU 2 ultramicrotome. Thin sections were stained with lead citrate, sometimes in combination with uranyl acetate, and viewed under Zeiss EM 9A and Siemens 101 electron microscopes.

SGC-CelIs in Guinea-Pig Adrenals 111
Histochemistry. The argentaffin reaction was applied to semithin sections of Araldite-embedded, glutaraldehyde fixed postosmicated material (Gorgas and B6ck, 1976a) using the ammoniacal silver nitrate solution described by Singh (1964). This method allows identification of NA-storing cells in the adrenal medulla, since NA forms insoluble precipitates with glutaraldehyde (Hopwood, 1971). A and uncontrolled amounts of DA are lost during the fixation procedure, and, accordingly, cannot be shown with this method.
Adrenal glands were processed for formaldehyde-induced fluorescence according to the method of Falck and Hillarp (Falck et al., 1962). The formaldehyde treatment was performed at 80~ for 1 h. The paraformaldehyde used had previously been equilibrated in air at 7 0 ~ relative humidity. After vacuum embedding in paraffin, specimens were sectioned and mounted in liquid paraffin. They were analyzed in a Zeiss fluorescence microscope fitted with BG 3 or BG 12 (Schott) primary and Zeiss 47 + 50 secondary filters. For further technical details, see Falck et al. (1962) and Bjrrklund, Falck and Owman (1972).
15 ~t thick native frozen sections of 4 adrenals were processed according to the glyoxylic acid method for the demonstration of biogenic amines (de la Torre and Surgeon, 1976).
Drug Treatments. Animals were injected with 5- or 6-hydroxydopamine (OHDA). The schedule of treatment was as follows: 5-OHDA: each animal received 3 injections of 200 mg/kg in 12 h intervals; 6- OHDA: newborn animals received 3 injections of 100 mg/kg in 24 h intervals; adult animals received 4 injections of 100 mg/kg each on days 1, 2, 7 and 8, the last one 12 h before sacrifice. Drugs were dissolved in 1-2 ml 0,9 ~ NaCl-solution with 0.2 700 ascorbic acid and injected intraperitoneally. Tissues from treated animals were processed in the same way for electron microscopy as described above.
Transplant Experiments were performed on adult guinea-pigs. Under ether anesthesia the left adrenal gland was removed, freed from cortical tissue and grafted under the left kidney capsule. Three weeks later the animals were perfused with phosphate-buffered glutaraldehyde and transplants and adrenal glands were processed for electron microscopy. A few animals were pretreated with reserpine or received 5-OHDA prior to sacrifice as described above.
Determination of Phenylethanolamine-N-methyltransferase (PNMT) Activity and Catecholamines ~. Adrenal glands (fight or left) of newborn or adult guinea-pigs were homogenized in 6 ml of 5 mM tris- HC1 buffer (pH 7.0), containing 0.2 ~ Triton X-100 and 0.2 ~o bovine serum albumin (w/v both). The homogenate was divided into two parts for the estimation of PNMT activity and catecholamines.
PNMT activity was measured by the method of Axelrod (1962) with slight modifications. The homogenate was centrifuged for 30 rain at 20.000 rpm in a Sorvall Superspeed RC2-B centrifuge. 20 p.l of the supernatant fluid was incubated at 37~ for 5 min in a total volume of 200 p.l containing 80 I~1 5 mM tris-HC1 buffer (pH 7.0), 50 ~tl OF L N tris-HC1 buffer (pH 8.6), 1.8 mM phenylethanolamine (diluted in 25 ial 0.001 N HCI), 20 lal 0.001 N HCI and 5 p.l S-adenosylmethionine solution (total 0.7 nmol, 17 ~ 14C- labelled, specific activity 56.7 mCi nmol- ~). Throughout the incubation period the initial rate of enzyme activity was constant. The reaction was stopped by addition of 500 lal 0.5 M borate buffer (pH 10.0). Blanc values were prepared by reversed addition of S-adenosylmethionine and borate buffer. The ~4C- labelled N-methylated reaction product was extracted in 6 ml of 97 ~ toluene- 3 ~o isoamylalcohol (v/v), which was added to the reaction mixture. The tube was shaken for 5 min and centrifuged. A 4-ml aliquot of the organic solvent was used for liquid scintillation counting (Packard model 3380) after addition of 10ml scintillation fluid.
Noradrenalirte and adrenaline were extracted from 2 ml of the homogenate by addition of 2 ml 10 trichloroacetic acid for 20 rain at 4 ~ C. After centrifugation at 5000 rpm for 10 min at 4 ~ C, the sediment was reextracted with 6 ml 5 ~o trichloroactic acid and centrifuged as described above. The supernatants were pooled and the catecholamines of an aliquot (2 ml) were adsorbed on and eluted from aluminia. Noradrenaline and adrenaline were measured by differential fluorometry (Unsicker et al., 1977a).
Drugs Used. Phenylethanolamine (ICN), S-Adenosyl-methionine (Boehringer Mannheim), 14C-S- Adenosyl-methionine (NEN, Dreieichenhain).
Biochemical determinations were carried out by Drs R. Lindmar, K. L6ffelholz and U. Wolf, Department of Pharmacology, University of Mainz.

112 K. Unsicker et al.
Results
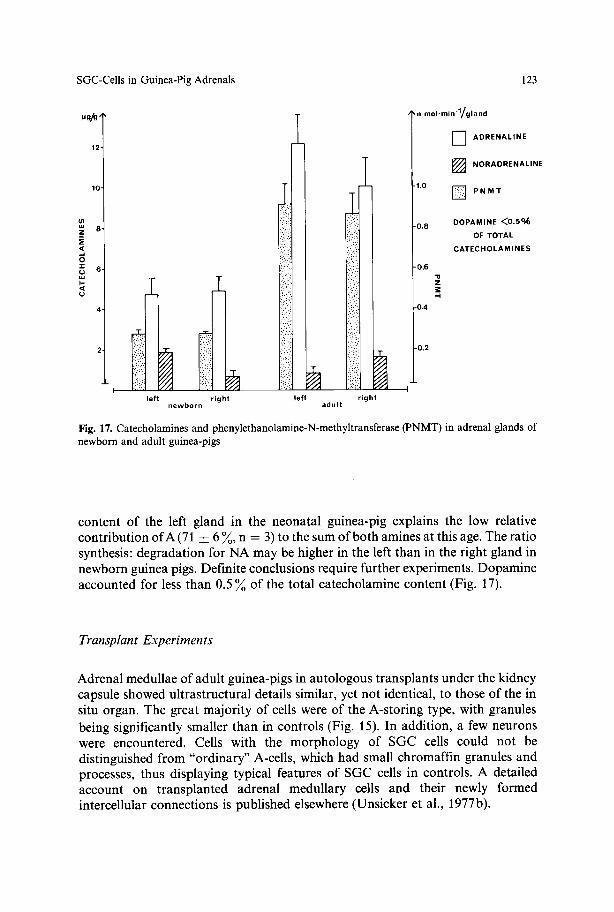
A-Cells (Fig. 1). Cells containing granular vesicles with cores of medium electron densities and an average diameter of 180 nm represented the vast majority of CA- storing cells in the adrenal medulla of both newborn and adult guinea-pigs. In biochemical determinations A was also identified to be the predominating CA, although there were significant differences in the proportion of A and NA, when newborn and adult animals were compared (Fig. 17).
There was a wide range in electron densities of granular cores in A-cells (Fig. 1). The overall ultrastructural picture of the cells including their organelles corresponded to former descriptions of mammalian chromaffin cells as reviewed by Coupland (1972) and Grynszpan-Winograd (1975).
NA-cells (Fig. 2) with highly electron-dense cored vesicles having roughly the size of A-granules (average diameter 170 nm) (Fig. 16) occurred very rarely and were clearly outnumbered by one type of SGC-cells, which also stored NA (see below).
SGC-Cells differed from ,,ordinary" A- and NA-storing cells described above with respect to their smaller cell body, a higher nucleus-cytoplasm ratio, a smaller average size of membrane-bounded cytoplasm granules and a pronounced capacity to form processes. By correlating adjacent semithin sections stained with an ammoniacal silver nitrate solution (Sing, 1964) and ultrathin sections it was possible to distinguish at least two different types of SGC-cells. Cells staining deeply brown to black in semithin sections contained NA (Figs. 3, 4). On an ultrastructural level (Fig. 3) a large majority of these cells had granular vesicles with highly electron-dense cores measuring 20 to 140 nm in diameter (average 80 nm). Thus, the granular vesicles were significantly smaller than in"ordinary" NA-storing cells, which constituted only a minority among NA-cells in the guinea pig-adrenal medulla (see above).
A second SGC-cell type was characterized by a population of membrane- bounded granules of variable moderate to low electron-density with few highly electron-dense cores (average diameter 80-90 nm) present in a few vesicles (Fig. 5). These cells were not stained in semithin sections treated according to Singh (Fig. 6, inset). Both types of SGC-cells had small (40-70 nm) clear and dense-cored vesicles in the cytoplasm resembling those in adrenergic nerve varicosities (Fig. 12). Loosely arranged rough ER was not particularly prominent. Profiles suggestive of granule formation in the Golgi apparatus were consistently observed. Granular vesicles were less tightly packed than in "ordinary" A- and NA-cells. There were spacious areas within the cells which exhibited only tubules and scanty profiles of ER embedded into electron-lucent ground cytoplasm.
SGC-cells occurred single or in small groups comprising up to 6 cells per semithin section. Most SGC-cells were in contact with one another or with A-cells their cell membranes being closely apposed. Extensive desmosomes were consistently observed at such sites. Narrow profiles of satellite cells were often apposed to SGC-cells, but mostly did not intervene where the cells approached the pericapillary space (Fig. 12). Administration of 5- or 6-OHDA did not cause ultramorphological changes in the cell bodies of SGC-cells.

Fig. 1. A cell with large storage granules. The storage granules have granular cores, which display a wide range of different electron densities. The vast majority of chromaffin cells in the adrenal medulla of the guinea-pig belongs to this cell type. • 18,000
Fig. 2. NA cell with large storage granules, which show excentrically located cores of high electron densities. This type of cell is extremely rare. x 18,000

Fig. 3. SGC-NA-positive cell, which stores small granules of high electron densities. Note the larger granules in neighbouring A cells! x 18,000
Fig. 4. Low power electron micrograph of the cell seen in Fig. 3. In an adjacent semithin section (inset) this cell reacts strongly for NA after incubation in an ammoniacal silver nitrate solution, x 5400, x 250

Fig. 5. SGC-NA-negative cell. The storage granules have cores of low to medium electron densities
Fig. 6. Same cell as shown in Fig. 5 at a lower magnification and in a semithin section incubated with an ammoniacal silver nitrate solution (inset). No positive-reaction is seen. x 5400, x 250

116 K. Unsicker et al.
While "ordinary" A- and NA-storing cells had a rich supply of cholinergic nerve endings with typical synapses, synaptic endings with pre- and postsynaptic membrane specializations were extremely rare with SGC-cells of both types.
Neurons. Large numbers of nerve cells were present in the adrenal. At an ultrastructural level two kinds of neurons could be distinguished in the medulla of newborn and adult guinea-pigs. Type I (Fig. 7) was a large multipolar cell with a rounded, lucent nucleus and 1-2 very prominent nucleoli. Cytoplasm was abundant containing densely packed patches of Nissl substance. In the Golgi region there were a few membrane-bounded granules, which were different from those in SGC- cells with respect to size (80-120 nm in diameter). These nerve cells received both axo-somatic and axo-dendritic synapses. The terminals contained small clear (about 50 nm) and large dense-cored (80-120 nm) vesicles, neither of which showed uptake of 5-OHDA, and, hence, were considered to be cholinergic. Type II (Fig. 8) of medullary neurons comprised a more heterogeneous population of what appeared to be transitional stages between nerve- and SGC-cells. These cells were usually smaller than type I cells and contained varying amounts of granular vesicles, which had the size and electron densities of those in SGC-cells.
Processes of SGC and Neurons Cells
A common feature of these processes were varying amounts of small vesicles (40- 70 nm), which were mostly clear and rarely exhibited an excentrically located highly electron dense core. This implies that profiles of processes which exhibited only these small vesicles could not be classified as belonging to one of the respective cell types. Processes of medullary neurons contained additional larger vesicles (80- 120 nm) with an opaque core and were found in the following locations: (1) along muscular and non-muscular blood vessels approaching smooth muscle, smooth muscle-like (Fig. 10) (see Unsicker and Gr6schel-Stewart, 1977) and endothelial cells at distances up to 50 nm. Their processes were found both in the medulla and in the cortex, where they were concentrated in the reticularis and glomerulosa zone. With the glomerulosa zone it was not established whether axons belonged to medullary or extra-adrenal nerve cells. Most of the neuronal processes appeared to be axons as judged by their microtubule content and the lack of rough ER and free ribosomes. Bundles of axons were invested by Schwann cells in the usual manner, but became satellite cell free at varicose regions, where vesicles were concentrated. Often processes of SGC cells were found admixed to the nerve cell axons.
(2) Another location, where nerve cell axons and their varicosities were found, was adjacent to cortical steroid producing cells (Fig. 11).
(3) Very rarely profiles, which might be varicosities of nerve cell axons were observed in contact with chromaffin cells (Fig. 12, inset). Vesicles of nerve cell axons showed uptake of 5-OHDA (Fig. 10). After application of 6-OHDA varicosities were destroyed and exhibited the ultrastructural changes, which are known to be caused by this drug (Tranzer and Thoenen, 1967; Thoenen, 1972).
Processes of SGC cells contained both small (about 50nm) vesicles with occasional dense cores and the large granular vesicles characteristic for this cell
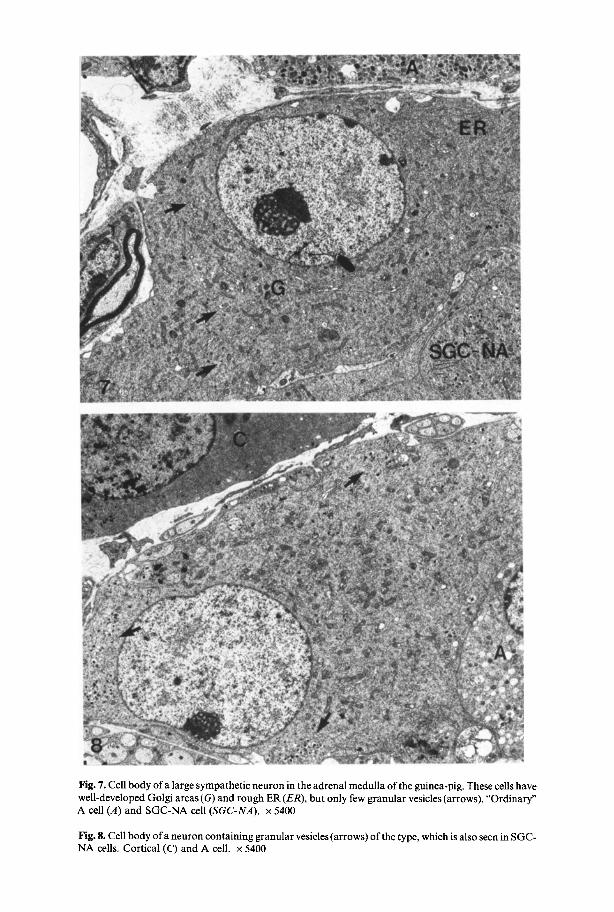
Fig. 7. Cell body of a large sympathetic neuron in the adrenal medulla of the guinea-pig. These cells have well-developed Golgi areas (G) and rough ER (ER), but only few granular vesicles (arrows). "Ordinary" A cell (A) and SGC-NA cell (SGC-NA). x 5400
Fig. 8. Cell body of a neuron containing granular vesicles (arrows) of the type, which is also seen in SGC- NA cells. Cortical (C) and A cell. x 5400

Fig. 9. Processes (P) of SGC-NA-negative cells containing large granular and small, mostly clear vesicles (arrow). Two processes are seen to contact a steroid producing cell of the fasciculata zone (arrowheads). x 18,000
Fig. 10. Two adrenergic axons (a) without a satellite cell sheath in close contact with an arteriolar vessel in the adrenal cortex. After 5-OHDA treatment most vesicles in the axons display highly electron dense cores. It might be assumed that the axons belong to adrenergic neurons and no t to SGC cells, since large storage granules are absent. • 18,000
Fig. 11. Adrenergic axons (a) containing vesicles, which have taken up 5-OHDA, in close contact with a fasciculata (FC) and a capillary endothelial cell (E). • 30,000

SGC-Cells in Guinea-Pig Adrenals 119
Fig. 12. Processes of SGC-NA cells containing both the large storage granules (arrows) and clusters of microvesicles (arrowheads). One process contacts a sinusoid endothelial cell (E). x 18,000. Inset: Adrenergic axon (a) in contact with an A storing cell (A). The identity of the axons in terms of its origin (adrenergic sympathetic neuron or SGC cell) is not clear, x 18,000
Fig. 13. Processes of SGC-NA cells (P) in close associations with what might be profiles of dendrites originating from nearby neurons. The contact areas bear membrane specializations (arrowheads). x 18,000

120 K. Unsicker et al.
Fig. 14. Nerve fibres in the reticularis zone (R) showing a strong CA-specific fluorescence (arrows). Groups of medullary chromaffin cells (CH). x 410
type. These processes devoid of satellite cells were found in close association with cortical endocrine cells (Fig. 9) mainly in the fasciculata and less frequently in the reticularis zone. In addition, they were observed apposed to the surface of sinusoid vessels in medulla and cortex. Some SGC processes terminated with synaptic endings showing vesicle accumulation and synaptic membrane specializations upon profiles, which could be dendrites of medullary neurons (Fig. 13), 5-OHDA uptake was observed in the small vesicle population of a few, but not all SGC cell processes.
Fluorescence Histochemistry
Treatment of frozen sections of adrenal glands according to the glyoxylic acid technique or sections of freeze-dried and paraffin-embedded glands (Falck-Hillarp method) revealed catecholamine-specific fluorescence in medullary cells and in fibres, which were most prominent (Fig. 14) in the reticularis and glomerulosa zones. In the medulla chromaffin and neurons could be distinguished by means of the different fluorescence intensities (or colours, respectively) of their cytoplasm.
Biochemical Results
Determinations ofcatecholamines on adrenal glands of newborn and adult guinea- pigs revealed a high predominance of A. The contribution of A to the sum of both, A

SGC-Cells in Guinea-Pig Adrenals 121
Fig. 15. Adrenal chromaffin cell from a transplant under the kidney capsule three weeks after operation. Granular vesicles are less numerous (cf. Fig. 1) and smaller (cf. inset) than in controls. Note the process (P), which extends from the cell body. • 18,000
and NA, was a r o u n d 90 ~ . In the right adrena l gland, the relat ive conten t o f A did no t change f rom the neona ta l to the adu l t age, since the abso lu te con ten t per g land o f bo th A and N A increased by the same rate. The abso lu te conten t o f A o f the left gland showed s imilar ly an a b o u t 2-fold increase, whereas the N A conten t decreased f rom 1 . 9 • (n = 3) to 0.84-0.3 ~tg per g land (n = 5). The r e m a r k a b l y high N A

122 K. Unsicker et al.
400
3 0 0
200
100
40 80 120
m
m
A D U L T A - C E L L S
100 2 0 0 2 4 0 2 8 0 40 80 120
- - m
m
160
N E W B O R N A - C E L L S
200 240 280
70(
000
500
400
300
200
100
Y 4O 120 160
A D U L T S G C - N A C E L L S
size i n n m
4O
m
m
80
N E W B O R N S G C - N A
C E L L S
120 160
600
500.
r tll
400-
,.~ 300-
E
200-
100-
m
m
A D U L T S G C - N A - N E G A T I V E
C E L L S
40 80 120 160
i
4O
N E W B O R N
S G C - N A - N E G A T I V E
C E L L S
L 120
Fig. 16. Size distribution of granules in "normal" adrenaline-storing (A) and small granule-containing noradrenaline (NA) and NA-negative cells in newborn and adult guinea-pig adrenal medulla
SGC-CelIs in Guinea-Pig Adrenals 123
12-
10-
~ 8' Z
- i 0 ~ e I-
u
, n mol-min-1/gland
I i__] ,O.E.,L,N E O.O.ENA',NE
-1.0
"7!!: !i';i! ! DOPAMINE <0. S ~ ::'!:: j - 0 . 8
i i~.ii i% OF TOT*' ::;:i'i" CATECHOLAMINES !i!iii: '5 :j,.. " ::. - O, 6
�9 : : ' r : i}S i~:i.~: :::.% 7)."; ~ i:}i~ -o,, �9 S.% .:?.:..
,.':..:. :?): - 0 , 2
left right left right newborn adult
Fig. 17. Catecholamines and phenylethanolamine-N-methyltransferase (PNMT) in adrenal glands of newborn and adult guinea-pigs
content of the left gland in the neonatal guinea-pig explains the low relative contribution of A (71 • 6 ~ , n = 3) to the sum of both amines at this age. The ratio synthesis: degradation for NA may be higher in the left than in the right gland in newborn guinea pigs. Definite conclusions require further experiments. Dopamine accounted for less than 0.5 ~o of the total catecholamine content (Fig. 17).
Transplant Experiments
Adrenal medullae of adult guinea-pigs in autologous transplants under the kidney capsule showed ultrastructural details similar, yet not identical, to those of the in situ organ. The great majority of cells were of the A-storing type, with granules being significantly smaller than in controls (Fig. 15). In addition, a few neurons were encountered. Cells with the morphology of SGC cells could not be distinguished from "ordinary" A-cells, which had small chromaffin granules and processes, thus displaying typical features of SGC cells in controls. A detailed account on transplanted adrenal medullary cells and their newly formed intercellular connections is published elsewhere (Unsicker et al., 1977b).

124 K. Unsicker et al.
�9 t
Fig. 18. Semi-schematic drawing showing the main results of the present study. Medullary neurons (N), SGC cells (S) and chromaffin cells (CH) receive preganglionic cholinergic inputs (arrows). Two types of SGC cells (NA positive = S + and NA negative = S - ) may be distinguished. Their processes contact neurons, cortical cells (C) and medullary and cortical blood vessels (Is)

SGC-CelIs in Guinea-Pig Adrenals
Preganglionic
cholinergic nerve
Preganglionic
cholinergic nerve
125
V e s s e l
Corte~ ce J
i i m i
Vessel (smooth muscle ] endothelium) ]
Fig. 19. Afferent and efferent connections of chromaffin, SGC and nerve cells and their presumptive targets in the guinea-pig adrenal gland
Discussion
The observations reported in this study give morphological evidence for the presence of SGC cells in the adrenal medulla of newborn and adult guinea-pigs. These cells may be distinguished from "ordinary" A- and NA-storing cells in the adrenal medulla by smaller granules, more polyribosomes, a higher nucleus- cytoplasm ratio, and a great capacity to form processes. Very similar cells have been described by Coupland and co-workers (Coupland et al., 1977; Kobayashi and Coupland, 1977) and by Gorgas and B6ck (1976b) in the adrenal medulla of the

126 K. Unsicker et al.
mouse. With respect to many ultrastructural features adrenal SGC cells resemble their counterparts in sympathetic ganglia, which also have small cell bodies (6- 12 ~m)(Matthews and Raisman, 1969), an abundance of ribosomes (Matthews and Raisman, 1969), processes of variable length (Williams et al., 1975), and granular vesicles, which are usually smaller than those found in the chromaffin cells of the adrenal medulla (rat superior cervical ganglion: 60-120nm (H6kfelt, 1969; Matthews and Raisman, 1969; Taxi et al., 1969; Williams and Palay, 1969); rabbit inferior mesenteric ganglion: 90-170 nm (Elfvin, 1968).
As is the case with SGC cells in sympathetic ganglia adrenal SGC cells do not appear to be a homogeneous population. The majority of SGC cells in the guinea- pig adrenal medulla has highly electron-dense granules and stains with ammoni- acal silver solutions indicating the presence of a primary amine, most probably NA. This would be in accordance with the biochemical findings showing that about 10~ of the total amine content is NA, an amount, which the extremely rare "ordinary" NA-cells with large granular vesicles could not account for. With respect to electron density of granules, but not granular size, these NA containing SGC cells would correspond to type II SGC-cells in rat superior cervical and coeliac- mesenteric ganglia (Lu et al., 1976), type I cells in the guinea-pig hypogastric ganglion (Watanabe, 1971) and one type of cell described by Elfvin et al. (1975) in the guinea-pig inferior mesenteric ganglion. A minority of SGC cells in the guinea- pig adrenal medulla has granular vesicles with cores of low to medium electron density, which could indicate the presence of A or might be due to fixation-mediated outwash of a primary amine, DA or NA. Very likely, DA is not the amine stored in these cells, since it accounts for less than 0.5 ~ of the total catecholamine content in the guinea-pig adrenal medulla. This suggests that DA is probably not stored in a separate type of cell. Thus, the second type of SGC cell may contain NA or A.
What may be the functional significance of adrenal SGC cells ? Could a specific function different from that of "ordinary" chromaffin cells be assigned to them? The mere presence of granules which are smaller than those in other chromaffin cells would certainly not allow per se to establish a new functional type of cell. The results of our transplant (Unsicker et al., 1977 and this study) and tissue culture experiments (Unsicker and Chamley, 1977) suggest that the lack of environmental factors, among which corticosteroid hormones and innervation may play a crucial role, or the failure to respond to circulating agents such as corticosteroids, cause a reduction in the size of the granules in "ordinary" chromaffin cells. Accordingly, SGC cells in the adrenal medulla might be interpreted as relatively immature elements, that are either not reached by factors, which are required for further differentiation, or, more likely, which do not respond to them. The high nucleus- cytoplasm ratio and the abundance of ribosomes would be in line with an interpretation of SGC cells as immature chromaffin cells.
On the other hand, a large number of adrenal SGC cells display clear neuronal characteristics. They receive cholinergic synapses, although not many, they have processes, which contain the 40-60 nm clear and dense cored vesicles typical of adrenergic varicosities, and their processes contact various target cell candidates such as smooth muscle, cortical endocrine and neurons. Processes of both types of adrenal SGC cells were seen to extend over considerable length. They were found in the medulla, in the reticularis, the fasciculata and in the glomerulosa zone.

SGC-CelIs in Guinea-Pig Adrenals 127
Such processes were observed to terminate with typical pre- and postsynaptic membrane specializations and vesicle accumulations on the presynaptic side upon profiles, which we interpreted to be dendrites of large nerve cells, since identical profiles could be traced back to the cell bodies and since these synapses were consistently found in the immediate vicinity of nerve cells. SGC cells in the rat superior cervical ganglion have been described to give rise to efferent synapses (Siegrist et al., 1968; Matthews and Raisman, 1969; Williams and Palay, 1969), but only in a few cases has the post-junctional element been identified as a principle neuron (Taxi et al., 1969; Yokota, 1973). These findings have largely contributed to center the speculation about the role of the SGC cell in sympathetic ganglia on the possibility that they might be an interneuron intercalated between preganglionic cholinergic axons and adrenergic neurons. In analogy with this interpretation it would be tempting to ascribe an interneuron function at least to some of the adrenal SGC cells. Other possibilities for a functional interpretation of these cells are provided by their intimate relationships with smooth muscle, cortical steroid producing cells and even endothelial cells. It has been suggested that in lower vertebrates chromaffin cells may directly act upon vascular smooth muscle (Unsicker, 1976). Moreover, in recent culture experiments with adrenal chromaffin cells and dissociated smooth muscle from vas deferens we were able to show that chromaffin cell processes formed long-lasting intimate associations with the muscle cells as compared to those, which occurred with ureter muscle and which were only short-lasting (Unsicker, unpublished). Thus, the results were identical to those obtained by Chamley et al. (1972) with sympathetic nerve cells suggesting that adrenal chromaffin cells (which turn into SGC-like cells in culture, see Unsicker and Chamley, 1977) and sympathetic neurons may establish identical relationships with smooth muscle from a densely innervated organ.
An innervation ofcorticosteroid producing cells has been suggested on the basis of morphological findings, although convincing experimental evidence has not been obtained to day (see Unsicker, 1973d, for review). Among the agents, which are known to influence corticosteroid secretion from the adrenal cortex, are catecholamines (Vogt, 1944; Brudieux and Delost, 1968).
On the other hand, corticosteroids are known to increase the activity of various adrenal medullary enzymes, in particular PNMT, which are involved in catecholamine synthesis (Wurtman and Axelrod, 1966). Thus, SGC cells, by influencing corticosteroid secretion, would act as modulators on CA synthesis in the adrenal medulla.
However, the possibility of a more direct influence of SGC cells on CA secretion should also be considered. A selective innervation of SGC cells rising local NA levels could inhibit further release ofcatecholamines from adjacent chromaffin cells by acting upon their ~-adrenoreceptors (Gutman and Boonyaviroy, 1974; Starke et al., 1974).
Small and large neurons exist in the guinea-pig adrenal medulla. While a continuous line leads from the small elements to SGC cells via many intermediate stages, the large cells seem to constitute a separate entity. Small neurons, not all of which may correspond to SIF cells, have also been found in the superior cervical ganglion of cat and man and in the adrenal gland of lower vertebrates (Unsicker, 1973 a, 1976). They were interpreted to be elements retarded in their development.

128 K. Unsicker et al.
The large neurons o f the adrena l medul la p r o b a b l y co r r e spond to the pr inc ipa l neurons o f sympathe t i c ganglia. They receive chol inergic inputs as judged f rom the lack o f effect o f 5 - O H D A on the small ag ranu la r vesicles in the pregangl ionic nerve endings (see Gri l lo , 1966; Tranzer and Thoenen, 1967; Chiba , 1973). Processes o f nerve cells con ta in ing 80-120 nm g ranu la r vesicles in add i t i on to small (40-60 nm) ag ranu la r and g ranu la r vesicles were found at medu l l a ry and cor t ical b lood vessels. These f indings suggest tha t there are adrenergic neurons in the adrena l medul la , which might affect regional b lood flow. The classif icat ion o f ad rena l neurons as adrenergic elements receives suppor t f rom our t r ansp lan t studies, where 5 - O H D A - labeled processes could di rect ly be t raced back to the s o m a t a o f large ne rve cells. However , these exper iments also revealed the presence o f non-adrenerg ic synapses u p o n t r ansp lan ted nerve cells (Unsicker et al., 1977). Since extr insic nerves d id not invade the t ransplants , these synapses mus t be fo rmed by intrinsic, i.e. ad rena l medul la ry nerve cells. This opens the possibi l i ty tha t the adrena l medul la conta ins a second ca tegory o f poss ib ly chot inergic neurons , a l though it canno t be ruled out tha t a few t r ansp lan ted adrenergic nerve cells switched their t ransmi t te r f rom N A to acetylchol ine as sympathe t i c adrenergic neurons m a y do under cer ta in condi t ions in cul ture (Pat terson et al., 1975; O 'Lague et al., 1975).
References
Axelrod, J.: Purification and properties of phenylethanolamine-N-methyltransferase. J. biol. Chem. 237, 1657-1660 (1962)
Bj6rklund, A., Cegrell, C., Falck, B., Ritz6n, M., Rosengren, E.: Dopamine-containing cells in sympathetic ganglia. Acta physiol, scand. 78, 334-338 (1970)
Brudieux, R., Delost, P.: Secretion des corticosteroids apr6s autogreffe surr6nalienne chez le rat. J. Physiol. (Paris) 60, Suppl. 2, 411 (1968)
Chamley, J.H., Mark, G.E., Campell, G.R., Burnstock, G.: Sympathetic ganglia in culture. I. Neurons. Z. Zellforsch. 135, 287-314 (1972)
Chiba, T.: Electron microscopic and histochemical studies on the synaptic vesicles in mouse vas deferens and atrium after 5-hydroxydopamine administration. Anat. Rec. 175, 35-48 (1973)
Coupland, R.E.: The chromaffin system. In: Handbook of Experimental Pharmacology, Vol. XXXIII, Catecholamines (H. Blaschko and E. Muscholl, eds.), pp. 16-45. Berlin-Heidelberg-New York: Springer 1972
Coupland, R.E., Kobayashi, S., Tomlinson, A.: On the presence of small granule chromaffin (SGC) cells in the rodent adrenal medulla. Proc. Anat. Soc. Great Britain and Ireland 24/25, 3 (1977)
Dail, W.G., Evan, A.P., Eason, H.R.: The major ganglion in the pelvic plexus of the male rat. A histochemical and ultrastructural study. Cell Tiss. Res. 159, 49-62 (1975)
Diner, O.: Presence d'amas de v6sieules de type synaptique du niveau des p~ricaryons des cellules nerveuses des la m6dullosurr6nale du Hamster. C.R. Acad. Sci. (Paris) 266, 2441-2443 (1968)
Elfvin, L.G.: A new granule containing nerve cell in the inferior mesenteric ganglion of the rabbit. J. Ultrastruct. Res. 22, 37-44 (1968)
Elfvin, L.G., H6kfelt, T., Goldstein, M.: Fluorescence microscopical, immunohistochemical and ultrastructural studies on sympathetic ganglia of the guinea-pig, with special reference to the SIF- cells and their catecholamine content. J. Ultrastruct. Res. 51, 377-396 (1975)
Er~ink6, O.: Distribution of fluorescing islets, adrenaline and noradrenaline in the adrenal medulla of the hamster. Acta endocr. (Kbh.) 18, 180-188 (1955)
Er/ink6, O., H/irkonen, M.: Histochemical demonstration of fluorogenic amines in the cytoplasm of sympathetic ganglion cells of the rat. Acta physiol, scand. 58, 285-286 (1963)
Falk, B., H illarp, N.-A., Thieme, G., Torp, A.: Fluorescence of catecholamines and related compounds condensed with formaldehyde. J. Histochem. Cytochem. 10, 348-354 (1962)

SGC-Cells in Guinea-Pig Adrenals 129
Gorgas, K., B6ck, P.: Identification of chromaffin and enterochromaffin cells in semithin sections by means of the argentaffin reaction. Mikroskopie 32, 57-63 (1976a)
Gorgas, K., BSck, P.: Morphology and histochemistry of the adrenal medulla I. Various types of primary catecholamine-storing cells in the mouse adrenal medulla. H istochemistry 50,17-31 (1976 b)
Grillo, M.A.: Electron microscopy of sympathetic tissues. Pharmacol. Rev. 18, 387-399 (1966) Grillo, M.A., Jacobs, L., Conroe, J.H., Jr.: A combined fluorescence histochemical and electron
microscopic method for studying special monoamine-containing cells (S.I.F. cells). J. comp. Neurol. 153, 1-14 (1974)
Grynszpan-Winograd, O, : Ultrastructure of the chromaffin cell. In: Handbook of Physiology, Sect. 7, Vol. VI, Adrenal gland, pp. 295-308. Washington D.C.: Americ. Physiol. Soc. 1975
Gutmann, Y., Boonyaviroy, P.: Suppression by noradrenaline of catecholamine secretion from adrenal medulla. Europ. J. Pharmacol. 28, 384-386 (1974)
Heym, Ch. : Ganglion cells and paraganglionic cells in the developing superior cervical ganglion of normal and p-chlorophenylalanine treated rats. Cell Tiss. Res. 163, 249-262 (1975)
Heym, Ch.: Monoamine storage sites in the rat superior cervical ganglion following synthesis inhibition. Cell Tiss. Res. 165, 239-248 (1976)
Heym, Ch., Grube, D.: Effects of guanethidine on paraganglionic cells in the superior cervical ganglion of the rat. Anat. Embryol. 148, 89-97 (1975)
H6kfelt, T.: Distribution of noradrenaline storing particles in peripheric adrenergic neurons as revealed by electron microscopy. Acta physiol, scand. 76, 427-440 (1969)
Hopwood, D.: The histochemistry and electron histochemistry of chromaffin tissue. Progr. Histochem. Cytochem. 3, No. 1 (1971)
Jacobowitz, D.: Catecholamine fluorescence studies of adrenerglc neurons and chromaffin cells in sympathetic ganglia. Fed. Proc. 29, 1929 (1970)
Kebabian, J.W., Greengard, P.: Dopamine-sensitive adenylcyclase: possible role in sympathetic transmission. Science 174, 1346 (1971)
Kobayashi, S., Coupland, R.E.: Two populations of microvesicles in the SGC (small granule chromaffin) cells of the mouse adrenal medulla. Arch. histol, jap. 40, 251-259 (1977)
Libet, B., Owman, Ch.: Concomitant changes in formaldehyde-induced fluorescence of dopamine interneurons and in slow inhibitory postsynaptic potentials of the rabbit superior cervical ganglion, induced by stimulation of the preganglionic nerve or by a muscarinic agent. J. Physiol. (Lond.) 237, 635-662 (1974)
Libet, B., Tosaka, T.: Dopamine as a sympathetic transmitter and modulator in sympathetic ganglia: A different mode of synaptic action. Proc. nat. Acad. Sci. (Wash.) 67, 667-673 (1970)
Lu, K.-S., Lever, J.D., Santer, R.M., Presley, R.: Small granulated cell types in rat superior cervical and coeliac-mesenteric ganglia. Cell Tiss. Res. 172, 331-343 (1976)
Matthews, M.R., Raisman, G. : The ultrastructure and somatic efferent synapses of small granule- containing cells in the superior cervical ganglion. J. Anat. (Lond.) 1115, 255-282 (1969)
O'Lague, P.H., MacLeish, P.R., Nurse, C.A., Claude, P., Furshpan, E.J., Potter, D.D.: Physiological and morphological studies on developing sympathetic neurons in dissociated cell culture. Cold Spr. Harb. Symp. quant. Biol. 40, 399~t07 (1975)
Patterson, P.H., Reichhardt, L.F., Chun, L.L.Y.: Biochemical studies on the development of primary sympathetic neurons in cell culture. Cold Spr. Harb. Symp. quant. Biol. 40, 389-397 (1975)
Santer, R.M., Lu, K.-S., Lever, J.D., Presley, R.: A study of the distribution of chromaffin-positive (CH + ) and small intensely fluorescent (SIF) cells in sympathetic ganglia of the rat at various ages. J. Anat. (Lond.) 119, 589-599 (1975)
Siegrist, G., Dolivo, M., Dunant, Y., Foroglou-Kerannens, C., Ribaupierre, F. de, Rouiller, Ch.: Ultrastructure and junction of the chromaffin cells in the superior cervical ganglion of the rat. J. Ultrastruct. Res. 25, 381~,07 (1968)
Singh, J.: A modification of the Masson-Hamperl method for staining of argentaffin cells. Anat. Anz. 115, 81-82 (1964)
Starke, K., Gorlitz, B.D., Montel, H., Schiimann, H.J.: Local ct-adrenoreceptor mediated feed-back inhibition of catecholamine release from the adrenal medulla? Experientia (Basel)311, 1170-1171 (1974)
Taxi, J., Gantron, J., L'Hermite, P.: Donne ultrastructurale sur une eventuelle modulation adrenergique de l'activite du ganglion cervical superieur du rat. C.R. Acad. Sci. (Paris) 269, 1281-1284 (1969)

130 K. Unsicker et al.
Thoenen, H.: Surgical immunological and chemical sympathectomy. Their application in the investigation of the physiology and pharmacology of the sympathetic nervous system. In: Handbook of Experimental Pharmacology, Vol. XXXIII, Catecholamines (H. Blaschko and E. Muscholl, eds.), pp. 813-844. Berlin-Heidelberg-New York: Springer 1972
Tranzer, J.P., Thoenen, H.: Ultramorphologische Ver/inderungen der synaptischen Nervenendigungen der Katze nach Vorbehandlung mit 5- und 6-Hydroxy-Dopamin. Naunyn-Schmiedebergs Arch. Pharmak. exp. Path. 257, 343-344 (1967)
Torre, J.C. de la, Surgeon, J.W.: A methodological approach to rapid and sensitive monoamine histofluorescence using a modified glyoxylic acid technique. The SPG method. Histochemistry 49, 81-93 (1976)
Unsicker, K.: Ober die Ganglienzellen in Nebennierenmark des Goldhamsters (Mesocrieetus auratus). Ein Beitrag zur Frage der peripheren Neurosekretion. Z. Zellforsch. 76, 187-219 (1967)
Unsicker, K.: Fine structure and innervation of the avian adrenal gland. I. Fine structure of adrenal chromaffin cells and ganglion cells. Z. Zellforsch. 145, 389-416 (1973a)
Unsicker, K.: Fine structure and innervation of the avian adrenal gland. II. Cholinergic innervation of adrenal chromaffin cells. Z. Zellforsch. 145, 417-442 (1973b)
Unsicker, K.: Fine structure and innervation of the avian adrenal gland. III. Non-cholinergic nerve fibres. Z. Zellforsch. 145, 557-575 (1973c)
Unsicker, K.: Fine structure and innervation of the avian adrenal gland. V Innervation of interrenal cells. Z. Zellforsch. 176, 403-416 (1973d)
Unsicker, K.: Chromaffin, small granule-containing and ganglion cells in the adrenal gland of reptiles. Cell Tiss. Res. 165, 477-508 (1976)
Unsicker, K., Chamley, J.H.: Growth characteristics of postnatal rat adrenal medulla in culture. Cell Tiss. Res, 177, 247-268 (1977)
Unsicker, K., Gr6schel-Stewart, U.: Distribution of contractile proteins and adrenergic nerves in the adrenal gland of guinea-pig, rat and ox as revealed by immunofluorescence and the glyoxylic acid technique (1977, in press)
Unsicker, K., Polonius, T., Lindmar, R., L6ffelholz, K., Wolf, U.: Catecholamines and 5- hydroxytryptamine in corpuscles of Stannius of the salmonid, Salmo irideus L. A study correlating electron microscopical, histochemical and chemical findings. Gen. comp. Endocr. 31, 121-132 (1977a)
Unsicker, K., Zwarg, U., Habura, O.: Electron microscopic evidence for the formation of synapses and synaptoid contacts in adrenal medullary grafts. Brain Res. 120, 533-539 (1977b)
Vogt, M.: Observations on some conditions affecting the rate of hormone output by the suprarenal cortex. J. Physiol. (Lond.) 103, 317-332 (1944)
Watanabe, H.: Adrenergic nerve elements in the hypogastric ganglion of the guinea-pig. Amer. J. Anat. 130, 305-330 (1971)
Williams, T.H.W.: Electron microscopic evidence for an autonomic interneuron. Nature (Lond.) 214, 309-310 (1967)
Williams, T.H.W., Black, A.C. Jr., Chiba, T., Bhalla, R.C.: Morphology and biochemistry of small intensely fluorescent cells of sympathetic ganglia. Nature (Lond.) 256, 315-317 (1975)
Williams, T.H.W., Paley, S.L.: Ultrastructure of the small neurons in the superior cervical ganglion. Brain Res. 15, 17-34 (1969)
Wurtman, R.J., Axelrod, J.: Control of enzymatic synthesis of adrenaline in the adrenal medulla by adrenal cortical steroids. Biol. Chemistry 241, 2301-2305 (1966)
Yokota, R.: The granule containing cell somata in the superior cervical ganglion of the rat, as studied by a serial sampling method for electron microscopy. Z. Zellforsch. 141, 331-345 (1973)
Accepted January 22, 1978