Embed Size (px)
Citation preview

0014-2980/00/0606-1562$17.50+.50/0 © WILEY-VCH Verlag GmbH, D-69451 Weinheim, 2000
Differential desensitization of lipopolysaccharide-inducible chemokine gene expression in humanmonocytes and macrophages
Andreas Kaufmann1, Diethard Gemsa1 and Hans Sprenger2
1 Institute of Immunology, Philipps University, Marburg, Germany2 Institute of Laboratory Medicine, Leopoldina-Hospital, Schweinfurt, Germany
Bacterial lipopolysaccharide (LPS) has been shown to induce a wide variety of pro-inflammatory cytokines and chemokines. An initial challenge with minute amounts of LPScauses tolerance to later LPS effects which is characterized by a much lower or abrogatedrelease of pro-inflammatory cytokines. To explore the relationship between the production ofchemokines and the induction of LPS tolerance, we pretreated human monocytes withincreasing LPS doses and thereafter restimulated with LPS. The re-expression of the CCchemokines macrophage inflammatory protein (MIP)-1 § , MIP-1 g and RANTES was substan-tially suppressed after pre-incubation with low LPS doses. In striking contrast, the re-expression of neutrophil-attracting IL-8 and melanoma growth stimulatory activity- § and ofthe monocyte-attracting monocyte chemotactic protein-1 remained high and was, in part,initially increased after restimulation with LPS. The corresponding gene expression patternas determined by Northern blot analyses correlated closely with the release of chemokinesand cytokines. Thus, a basic set of chemotactic mediators that are still produced by other-wise LPS-desensitized monocytes/macrophages may ensure the continuing recruitment ofmonocytes and neutrophils into an inflammatory process caused by gram-negative bacteria.
Key words: Chemokine / Inflammation / Lipopolysaccharide / Monocyte/Macrophage
Received 10/1/00Revised 3/3/00Accepted 9/3/00
[I 20479]
Abbreviations: GRO: Melanoma growth stimulatory activ-ity MCP: Monocyte chemotactic protein MIP: Macrophageinflammatory protein RANTES: Regulated on activation,normal T cell expressed and secreted
1 Introduction
Infection with gram-negative bacteria is characterized bythe induction of inflammatory and chemotactic cytokines.The major membrane component LPS stimulates mono-cytes/macrophages to release a cascade of mediators,which can finally lead to septic shock [1]. However, a sin-gle sublethal injection of LPS induces a transient refrac-tory state to a subsequent LPS challenge [2, 3]. This phe-nomenon, referred to as endotoxin tolerance, was foundboth in vivo and in vitro [4–7] and may be the result ofa decreased capacity of monocytes to produce pro-inflammatory cytokines such as TNF- § , IL-1 g and IL-6.
In an endotoxin-tolerant state, only little data are avail-able about LPS-inducible chemokines. Chemokines arepotent chemoattractant cytokines [8, 9] that attract andactivate distinct leukocyte populations. The chemokinescan be broadly divided into two major subfamilies. TheCC chemokines, such as monocyte chemotactic protein(MCP)-1, macrophage inflammatory protein (MIP)-1 § ,MIP-1 g and regulated on activation, normal T cellexpressed and secreted (RANTES) preferentially attractand activate mononuclear cells [10]. The recruitment ofneutrophils is mediated by members of the CXC subfam-ily which contain an ELR motif preceding the first cyste-ine, such as IL-8 and melanoma growth stimulatoryactivity (GRO)- § [11, 12].
In the present study, we observed a marked and perma-nent down-regulation of the chemokines MIP-1 § , MIP-1 g and RANTES after LPS restimulation whereas, in con-trast, the release of the neutrophil-attracting chemokinesIL-8 and GRO- § and the major monocyte-attracting che-mokine MCP-1 remained unaffected.
1562 A. Kaufmann et al. Eur. J. Immunol. 2000. 30: 1562–1567

Fig. 1. Differential desensitization of cytokine and chemokine release by LPS. Human monocytes (0.75 × 106/ml) were pretre-ated with increasing doses of LPS (0.01 to 1 ng/ml) for 48 h and then restimulated with a second LPS challenge (10 ng/ml) for16 h. Cell cultures without restimulation were used as controls. Pretreatment with LPS induced a substantial suppression of cyto-kine release (A) as well as a marked suppression of a distinct set of chemokines (B). The release of the CXC chemokines IL-8 andGRO- § and the CC chemokine MCP-1 was not suppressed by pretreatment with LPS (C). Representative data from differentexperiments using cells from eight donors are shown. Data are means ± SD. *Significantly different from the restimulated cellswithout pretreatment (p X 0.05).
2 Results
2.1 Differential dose-dependent desensitizationof human monocytes by LPS
Pre-incubation of human monocytes with LPS reducedthe capacity to release the cytokines TNF- § and IL-6after restimulation with a second LPS challenge(Fig. 1 A). Similar results were obtained for the release ofthe chemokines MIP-1 § , MIP-1 g and RANTES (Fig. 1 B).The suppression correlated in a dose-dependent mannerwith the amount of LPS used in the primary culture: thehighest LPS dose induced the strongest state of unre-sponsiveness. The lysates of the restimulated cells didnot contain detectable amounts of chemokines andcytokines (data not shown), which excluded the pres-ence of intracellular stores and an impeded secretionafter desensitization. In striking contrast to these obser-vations, the production of the chemokines IL-8, GRO- §and MCP-1 was even enhanced in a manner dependenton the LPS dose used for pretreatment (Fig. 1 C). A con-tinuing release and accumulation after the primary LPSstimulus was not responsible for these results since con-trol cells without a LPS rechallenge did no longer releasechemokines and cytokines.
When we restimulated LPS-pretreated monocytes with10 ng/ml PMA or zymosan (15 particles/cell), the cellswere again fully capable of responding and releasedlarge amounts of cytokines, independently of the LPSdosis used for primary stimulation (Fig. 2).
2.2 Kinetics of the differential desensitizationby LPS
To analyze the opposing results obtained for the re-inducible release of chemokines in more detail, we per-formed different kinetic analyses. In a first set of experi-ments, primary cultures were treated with the initial LPSstimulus from 2 h to 5 days and were then restimulatedwith the second LPS challenge for 16 h. As shown inFig. 3 A, pretreatment of monocytes for 4–12 h wasalready sufficient to induce a significant reduction of asecondary MIP-1 § , MIP-1 g and RANTES release. Therelease of these chemokines and TNF- § and IL-6 (datanot shown) remained permanently suppressed. In strik-ing contrast, LPS-rechallenged monocytes were stillcapable of producing the chemokines IL-8, GRO- § andMCP-1 over the entire time period (Fig. 3 B). Even pre-
Eur. J. Immunol. 2000. 30: 1562–1567 Differential chemokine desensitization by LPS 1563

Fig. 2. Unaffected cytokine and chemokine release afterrestimulation with PMA and zymosan. LPS-pretreatedmonocytes were restimulated with 10 ng/ml PMA (A) orzymosan (15 particles/cell) (B) for 20 h. As exemplarilyshown for IL-8, MIP-1 § and TNF- § , pre-incubation with LPSdid not influence the release of cytokines and chemokinesinduced by PMA or zymosan. Data shown are from one outof four independent experiments with cells from differentdonors. *Significant difference to the restimulated control atp X 0.05.
Fig. 3. Kinetics of LPS pretreatment. Human monocytes were pretreated with increasing amounts of LPS (0.1 to 10 ng/ml) forthe indicated times (2–120 h), followed by a second LPS challenge (10 ng/ml) for another 16 h. Pretreatment with LPS for 4–12 hwas sufficient to induce the suppression of the chemokines MIP-1 § , MIP-1 g and RANTES (A). Significant differences to the con-trol (p X 0.05) were already observed after 4–12 h of LPS pretreatment. In contrast, pretreatment of human monocytes with LPSdid not induce significant differences for the re-inducible expression of the chemokines IL-8, GRO- § and MCP-1 (B). Data repre-sent the mean ± SD of a representative experiment out of five.
treatment with LPS up to 10 ng/ml was not able toreduce the release of these chemokines.
In a second set of kinetic experiments, monocyte cul-tures were pretreated with the first stimulus for 48 h andthereafter restimulated with the second LPS stimulusfrom 4 to 48 h. We found a significant reduction of thesecondary release of the cytokines TNF- § and IL-6 andof the chemokines MIP-1 § , MIP-1 g and RANTES (exem-plarily shown for TNF- § and MIP-1 § in Fig. 1 A). In strik-ing contrast, pre-incubation with LPS led to a fast andearly dose-dependent induction of the chemokines IL-8,GRO-a and MCP-1 (Fig. 4 B). At the beginning of the LPSrechallenge, the highest LPS dose used in the primarycultures induced the strongest responsiveness. Thereaf-ter, the release of IL-8, GRO- § and MCP-1 was lower,depending on the LPS dose. These data were alsoverified by Northern blot analyses which showed thatMCP-1 and IL-8 gene expression was strongly enhancedat the beginning of the LPS rechallenge: the stronger thefirst stimulus, the stronger was the reinduced geneexpression (data not shown).
3 Discussion
Infection with gram-negative bacteria is characterized byan enhanced release of inflammatory and chemotacticcytokines (chemokines). These mediators play a pivotalrole in the regulation of host response and recruitment ofleukocytes by extravasation into infected tissue [13, 14].The excessive production of inflammatory mediatorsmay result in fever, intravascular coagulation and septicshock. This is predominantly mediated by the activationof monocytes/macrophages in response to LPS, a major
1564 A. Kaufmann et al. Eur. J. Immunol. 2000. 30: 1562–1567
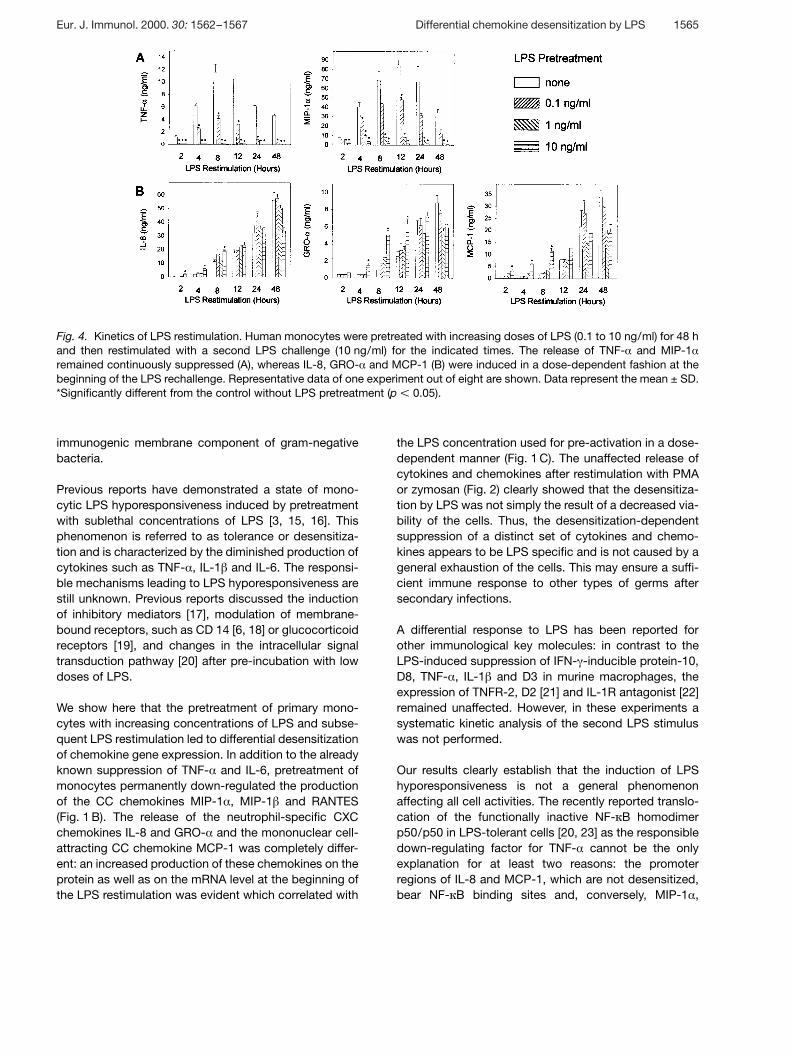
Fig. 4. Kinetics of LPS restimulation. Human monocytes were pretreated with increasing doses of LPS (0.1 to 10 ng/ml) for 48 hand then restimulated with a second LPS challenge (10 ng/ml) for the indicated times. The release of TNF- § and MIP-1 §remained continuously suppressed (A), whereas IL-8, GRO- § and MCP-1 (B) were induced in a dose-dependent fashion at thebeginning of the LPS rechallenge. Representative data of one experiment out of eight are shown. Data represent the mean ± SD.*Significantly different from the control without LPS pretreatment (p X 0.05).
immunogenic membrane component of gram-negativebacteria.
Previous reports have demonstrated a state of mono-cytic LPS hyporesponsiveness induced by pretreatmentwith sublethal concentrations of LPS [3, 15, 16]. Thisphenomenon is referred to as tolerance or desensitiza-tion and is characterized by the diminished production ofcytokines such as TNF- § , IL-1 g and IL-6. The responsi-ble mechanisms leading to LPS hyporesponsiveness arestill unknown. Previous reports discussed the inductionof inhibitory mediators [17], modulation of membrane-bound receptors, such as CD 14 [6, 18] or glucocorticoidreceptors [19], and changes in the intracellular signaltransduction pathway [20] after pre-incubation with lowdoses of LPS.
We show here that the pretreatment of primary mono-cytes with increasing concentrations of LPS and subse-quent LPS restimulation led to differential desensitizationof chemokine gene expression. In addition to the alreadyknown suppression of TNF- § and IL-6, pretreatment ofmonocytes permanently down-regulated the productionof the CC chemokines MIP-1 § , MIP-1 g and RANTES(Fig. 1 B). The release of the neutrophil-specific CXCchemokines IL-8 and GRO- § and the mononuclear cell-attracting CC chemokine MCP-1 was completely differ-ent: an increased production of these chemokines on theprotein as well as on the mRNA level at the beginning ofthe LPS restimulation was evident which correlated with
the LPS concentration used for pre-activation in a dose-dependent manner (Fig. 1 C). The unaffected release ofcytokines and chemokines after restimulation with PMAor zymosan (Fig. 2) clearly showed that the desensitiza-tion by LPS was not simply the result of a decreased via-bility of the cells. Thus, the desensitization-dependentsuppression of a distinct set of cytokines and chemo-kines appears to be LPS specific and is not caused by ageneral exhaustion of the cells. This may ensure a suffi-cient immune response to other types of germs aftersecondary infections.
A differential response to LPS has been reported forother immunological key molecules: in contrast to theLPS-induced suppression of IFN- + -inducible protein-10,D8, TNF- § , IL-1 g and D3 in murine macrophages, theexpression of TNFR-2, D2 [21] and IL-1R antagonist [22]remained unaffected. However, in these experiments asystematic kinetic analysis of the second LPS stimuluswas not performed.
Our results clearly establish that the induction of LPShyporesponsiveness is not a general phenomenonaffecting all cell activities. The recently reported translo-cation of the functionally inactive NF- ‹ B homodimerp50/p50 in LPS-tolerant cells [20, 23] as the responsibledown-regulating factor for TNF- § cannot be the onlyexplanation for at least two reasons: the promoterregions of IL-8 and MCP-1, which are not desensitized,bear NF- ‹ B binding sites and, conversely, MIP-1 § ,
Eur. J. Immunol. 2000. 30: 1562–1567 Differential chemokine desensitization by LPS 1565

which is strongly desensitized, does not. Therefore, inaddition to the p50 homodimer, other negative regulatorymechanisms must exist.
In conclusion, our data demonstrate that repeated LPSstimulation of primary monocytes leads to a selectivedesensitization of chemokine gene expression. The LPS-unaffected release of the chemokines IL-8, GRO- § andMCP-1 may significantly contribute to the continuousrecruitment and activation of new monocytes and neu-trophils within the center of an inflammation, even in thepresence of high endotoxin concentrations. Freshlyimmigrated cells are strong producers of immunoregula-tory molecules, such as TNF- § , IL-6 and chemokines,which may further augment an ongoing inflammationand support a successfull eradication of invaded gram-negative bacteria.
4 Materials and methods
4.1 Cell preparation, culture and treatment with LPS
Human monocytes were isolated from the buffy coat ofblood from healthy donors provided by the Department ofTransfusion Medicine, University of Marburg, Germany andcultured (0.75 × 106/ml) in RPMI 1640 medium plus 2 %heat-inactivated AB serum as previously described [24, 25].
Monocytes were pretreated for the indicated time periodswith different concentrations of LPS (from Escherichia coliO127:B8, Difco, Detroit, MI). Restimulation was performedwith 10 ng/ml LPS for the indicated times. Thereafter, super-natants were harvested and stored in aliquots at − 70 °C untilfurther use.
All reagents were essentially free of contaminating endo-toxin as determined by the Limulus test (LAL; BioWhittaker,Inc., Walkersville, MD) and the absence of spontaneous che-mokine and cytokine production when LPS was omitted.
4.2 Determination of chemokines and cytokines
Chemokine and cytokine levels were determined by specificsandwich ELISA developed in our laboratory [26]. Briefly, 96-well microtiter plates (Maxisorp; Nunc, Wiesbaden, Ger-many) were coated with mAb specific for IL-8 (IC Chemika-lien, Ismaning, Germany), GRO- § (Sigma, München, Ger-many), MCP-1, IL-6, TNF- § (all from PharMingen, Hamburg,Germany) or MIP-1 § , MIP-1 g and RANTES (all from R&DSystems, Wiesbaden, Germany) in PBS. Plates wereblocked with 2 % BSA in PBS. Samples (100 ? l/well) wereincubated at room temperature for 1 h. After three washeswith 0.05 % Tween-20 in PBS, a specific polyclonal antibodywas added and incubated for another hour. The polyclonalantibodies were purchased from IC Chemikalien (IL-8) , R&D
Systems (GRO- § , MIP-1 § , MIP-1 g , RANTES) or from Phar-Mingen (MCP-1, IL-6, TNF- § ). Detection was performed witha peroxidase-conjugated third antibody (donkey anti-goat ordonkey anti-rabbit, both from Dianova, Hamburg, Germany)or a streptavidin-peroxidase conjugate (Boehringer Mann-heim, Mannheim, Germany) and subsequent conversionof o-phenylenediamine dihydrochloride substrate (Sigma,München, Germany). The absorbance was determined pho-tometrically at 490 nm and plotted against a standard curveperformed with the respective recombinant chemokines andcytokine purchased from IC Chemikalien (IL-8, MCP-1, MIP-1 § , RANTES), R&D Systems (GRO- § , MIP-1 g ), PharMingen(TNF- § ) or PBH, Hannover, Germany (IL-6). The sensitivitiesof the established ELISA were X 20 pg/ml for IL-8, MCP-1,RANTES, MIP-1 g and IL-6, X 50 p/ml for GRO- § andX 100 pg/ml for TNF- § and MIP-1 § . Intra- and interassay
variances were less than 5 %.
4.3 RNA preparation and Northern blot analysis
After restimulation with the second LPS challenge for differ-ent times, total RNA was prepared by a modified guanidinethiocyanate method as previously described in detail [27].Analyses of chemokine- und cytokine-specific mRNA levelswere performed as previously described [28].
4.4 Statistics
The results of the ELISA were expressed as means ± SD ofthree independently performed cultures of cells derived froma single donor. They were compared with control values byStudent’s t-test and considered significantly different at pvalues of X 0.05.
Acknowledgements: The authors gratefully acknowledgethe expert technical help of E. Rischkowsky. This work wassupported by grants of the Deutsche Forschungsgemein-schaft (Sp 395/2-2).
References
1 Dinarello, C. A., Proinflammatory and anti-inflammatory cyto-kines as mediators in the pathogenesis of septic shock. Chest1997. 112 (6 Suppl): 321–324.
2 Henricson, B. E., Benjamin, W. R. and Vogel, S. N., Differentialcytokine induction by doses of lipopolysaccharide and mono-phosphoryl lipid A that result in equivalent early endotoxin toler-ance. Infect. Immun. 1990. 58: 2429–2437.
3 Goto, M., Yoshioka, T., Young, R. L., Battelino, T., Anderson,C. L. and Zeller, W. P., A sublethal dose of LPS to pregnant ratsinduces TNF-alpha tolerance in their 0-day-old offspring. Am. J.Physiol. 1997. 273: R1158–1162.
4 Virca, G. D., Kim, S. Y., Glaser, K. B. and Ulevitch, R. J., Lipo-polysaccharide induces hyporesponsiveness to its own action inRAW 264.7 cells. J. Biol. Chem. 1989. 264: 21951–21956.
1566 A. Kaufmann et al. Eur. J. Immunol. 2000. 30: 1562–1567

5 Ziegler-Heitbrock, H. W. L., Frankenberger, M. and Wedel, A.,Tolerance to lipopolysaccharide in human blood monocytes.Immunobiology 1995. 193: 217–223.
6 Fahmi, H. and Chaby, R., Desensitization of macrophages toendotoxin effects is not correlated with a down-regulation oflipopolysaccharide-binding sites. Cell. Immunol. 1993. 150:219–229.
7 Seatter, S. C., Li, M. H., Bubrick, M. P. and West, M. A., Endo-toxin pretreatment of human monocytes alters subsequentendotoxin-triggered release of inflammatory mediators. Shock1995. 3: 252–258.
8 Baggiolini, M., Dewald, B. and Moser, B., Interleukin-8 andrelated chemotactic cytokines-CXC and CC chemokines. Adv.Immunol. 1994. 55: 97–179.
9 Baggiolini, M., Dewald, B. and Moser, B., Human chemokines:an update. Annu. Rev. Immunol. 1997. 15: 675–705.
10 Schall, T. J., Biology of the RANTES/SIS cytokine family. Cyto-kine 1991. 3: 165–183.
11 Matsushima, K., Morishita, K., Yoshimura, T., Lavu, S.,Kobayashi, Y., Lew, W., Appella, E., Kung, H. F., Leonard, E. J.and Oppenheim, J. J., Molecular cloning of a human monocyte-derived neutrophil chemotactic factor (MDNCF) and the inductionof MDNCF mRNA by interleukin 1 and tumor necrosis factor.J. Exp. Med. 1988. 167: 1883–1893.
12 Schröder, J. M., Mrowietz, U. and Christophers, E., Purifica-tion and partial biologic characterization of a human lymphocyte-derived peptide with potent neutrophil-stimulating activity.J. Immunol. 1988. 140: 3534–3540.
13 Luster, A. D., Chemokines-chemotactic cytokines that mediateinflammation. N. Engl. J. Med. 1998. 338: 436–445.
14 Imhof, B. A. and Dunon, D., Leukocyte migration and adhesion.Adv. Immunol. 1995. 58: 345–416.
15 Ziegler-Heitbrock, H. W. L., Blumenstein, M., Kafferlein, E.,Kieper, D., Petersmann, I., Endres, S., Flegel, W. A., Northoff,H., Riethmüller, G. and Haas, J. G., In vitro desensitizationto lipopolysaccharide suppresses tumour necrosis factor,interleukin-1 and interleukin-6 gene expression in a similar fash-ion. Immunology 1992. 75: 264–268.
16 Erroi, A., Fantuzzi, G., Mengozzi, M., Sironi, M., Orencole, S.F., Clark, B. D., Dinarello, C. A., Isetta, A., Gnocchi, P. and Gio-varelli, M., Differential regulation of cytokine production in lipo-polysaccharide tolerance in mice. Infect. Immun. 1993. 61:4356–4359.
17 Fahmi, H. and Chaby, R., Selective refractoriness of macro-phages to endotoxin-induced production of tumor necrosis fac-tor, elicited by an autocrine mechanism. J. Leukoc. Biol. 1993.53: 45–52.
18 Haas, J. G., Meyer, N., Riethmüller, G. and Ziegler-Heitbrock,H. W. L., Molecular mechanism in down-regulation of tumornecrosis factor expression. Proc. Natl. Acad. Sci. USA 1990. 87:9563–9567.
19 Salkowski, C. A. and Vogel, S. N., Lipopolysaccharide increasesglucocorticoid receptor expression in murine macrophages. Apossible mechanism for glucocorticoid-mediated suppression ofendotoxicity. J. Immunol. 1992. 149: 4041–4047.
20 Ziegler-Heitbrock, H. W. L., Wedel, A., Schraut, W., Strobel,M., Wendelgass, P., Sternsdorf, T., Bäuerle, P. A., Haas, J. G.and Riethmüller, G., Tolerance to lipopolysaccharide involvesmobilization of nuclear factor kappa B with predominance of p50homodimers. J. Biol. Chem. 1994. 269: 17001–17004.
21 Henricson, B. E., Manthey, C. L., Perera, P. Y., Hamilton, T. A.and Vogel, S. N., Dissociation of lipopolysaccharide (LPS)-inducible gene expression in murine macrophages pretreatedwith smooth LPS versus monophosphoryl lipid A. Infect. Immun.1993. 61: 2325–2333.
22 Randow, F., Syrbe, U., Meisel, C., Krausch, D., Zuckermann,H., Platzer, C. and Volk, H. D., Mechanism of endotoxin desensi-tization: involvement of interleukin 10 and transforming growthfactor g . J. Exp. Med. 1995. 181: 1887–1892.
23 Kastenbauer, S. and Ziegler-Heitbrock, H. W. L., NF- O B1 (p50)is upregulated in lipopolysaccharide tolerance and can blocktumor necrosis factor gene transcription. Infect. Immun. 1999.67: 1553–1559.
24 Kaufmann, A., Mühlradt, P. F., Gemsa, D. and Sprenger, H.,Induction of Cytokines and chemokines in human monocytesby Mycoplasma fermentans-derived lipoprotein MALP-2. Infect.Immun. 1999. 67: 6303–6308.
25 Sprenger, H., Meyer, R. G., Kaufmann, A., Bussfeld, D., Risch-kowsky, E. and Gemsa, D., Selective induction of monocyte andnot neutrophil-attracting chemokines after influenza A virus infec-tion. J. Exp. Med. 1996. 184: 1191–1196.
26 Sprenger, H., Rösler, A., Tonn, P., Braune, H. J., Huffmann, G.and Gemsa, D., Chemokines in the cerebrospinal fluid ofpatients with meningitis. Clin. Immunol. Immunopathol. 1996. 80:155–161.
27 Sprenger, H., Konrad, L., Rischkowsky, E. and Gemsa, D.,Background reduction in Northern analysis by preabsorptionof digoxigenin-labeled riboprobes. Biotechniques 1995. 19:334–336.
28 Sprenger, H., Krause, A., Kaufmann, A., Priem, S., Fabian, D.,Burmester, G. R., Gemsa, D. and Rittig, M. G., Borrelia burg-dorferi induces chemokines in human monocytes. Infect. Immun.1997. 65: 4384–4388.
Correspondence: Andreas Kaufmann, Institute of Immu-nology, Philipps University Marburg, Robert-Koch-Str. 17,D-35037 Marburg, GermanyFax: +49-6421-286-6813e-mail: kaufmana — mailer.uni-marburg.de
Eur. J. Immunol. 2000. 30: 1562–1567 Differential chemokine desensitization by LPS 1567
![chemokine/chemokine receptor pair ccL20/ccR6 in human ... · pancreas, stomach, prostate, testis, uterine cervix and skin[11]. The chemokine receptor CCR6 was originally described](https://img.pdfslide.net/doc/110x75/5f9ac7b0798b75658905651c/chemokinechemokine-receptor-pair-ccl20ccr6-in-human-pancreas-stomach-prostate.jpg)




























