Embed Size (px)
Citation preview

Developmental Brain Research 141 (2003) 15–24www.elsevier.com/ locate/devbrainres
Research report
D ifferential expression of fibroblast growth factor receptors in thedeveloping murine choroid plexus
*Sarah Reid, Patrizia FerrettiDevelopmental Biology Unit, Institute of Child Health, UCL, 30, Guilford Street, London WC1N 1EH, UK
Accepted 26 November 2002
Abstract
The choroid plexuses (CPs) are specialised secretory organs situated within the ventricles of the brain involved in the production ofcerebrospinal fluid (CSF) and the maintenance of the blood–CSF barrier. Abnormal function of the CPs can lead to hydrocephalus andraised intracranial pressure, pathologies frequently observed in certain craniofacial syndromes caused by single point mutations infibroblast growth factor receptors (FGFRs). At present, relatively little is known about the embryonic CPs in terms of gene or proteinexpression, function as the brain develops or on the potential role of FGFRs within this context. Given the limited information availableon the regulation of FGFRs during development of the CPs and periventricular tissues, we have carried out a detailed analysis of thelocalisation of FGFR1, 2, 3 and 4 proteins in these regions of the murine embryo from the time of formation of the CP in the thirdventricle at E12.5 throughout the second half of gestation, and examined the expression of different FGFR isoforms at E12.5 by RT-PCR.We show here that FGFR1 and FGFR4 are expressed in murine CPs at E12.5 but not at E15.5 or E18.5, suggesting a role for the signalingpathways transduced by these receptors at early stages of CP development. In contrast, FGFR2 expression is maintained throughout CPdevelopment, indicating that this receptor may play a role in the function of immature and mature CP. Also FGFR3 is detected at eachdevelopmental stage studied, but surprisingly its expression appears confined to the nuclei of CP cells, suggesting that FGFR3 in the CPdoes not respond to extracellular FGFs but may act in intracrine fashion. 2002 Elsevier Science B.V. All rights reserved.
Theme: Cellular and molecular biology/Development and regeneration
Topic: Blood–brain barrier /Glia and other non-neuronal cells
Keywords: Choroid plexus; Craniosynostosis; Development; Fibroblast growth factor receptor; Mouse
1 . Introduction The mammalian choroid plexus originates from twodistinct embryonic lineages. The choroid plexus epithelium
The choroid plexuses (CPs) are specialised secretory (CPe) is derived from the neuroepithelium of the neuralorgans situated within the ventricles of the brain. CPs have tube, whereas the underlying tissue, choroid plexus mesen-several functions, which include the production of cere- chyme (CPm) is derived from primitive paraxial cranialbrospinal fluid (CSF) and the maintenance of the blood– mesoderm [47]. Signs of CP formation are first observed inCSF barrier. Under healthy conditions, the CPs continu- the fourth ventricle between E10 and E10.5 in the mouseously secrete CSF, and thus influence intracranial pressure [C. Johanson, personal communication], followed by thedirectly. In the embryo, CPs develop early, and exhibit lateral ventricle CPs at approximately E11 [39]. The thirdaspects of adult functional characteristics prior to neuronal ventricle CP develops shortly afterwards, and becomesdifferentiation. As such, embryonic CPs may directly continuous with the lateral ventricle CPs [25,4]. Asinfluence the extracellular environment of the brain as it development proceeds, CPe thins rapidly [39,5] and ex-develops and matures. hibits many of the characteristic ultrastructural and mor-
phological features of adult CPs, such as intercellular tightjunctions [33,4,12] and enzymes involved in CSF secretion*Corresponding author. Tel.:144-20-7829-8894; fax:144-20-7831-[3]. CPs are highly vascular at a time when the blood4366.
E-mail address: [email protected](P. Ferretti). supply to the developing brain is still very limited.
0165-3806/02/$ – see front matter 2002 Elsevier Science B.V. All rights reserved.doi:10.1016/S0165-3806(02)00635-1

16 S. Reid, P. Ferretti / Developmental Brain Research 141 (2003) 15–24
Combined, these factors indicate that embryonic CPs by immunohistochemistry during the second half of gesta-influence the extracellular environment of the immature tion, and to compare the spatial and temporal differences inbrain during neurogenesis. Abnormal function of the FGFR protein expression between the CPe, the CPm, thedeveloping CPs may therefore have a substantial effect cells of the ependyma, and the periventricular CNS. Weupon the development of the central nervous system have found that FGFRs are differently regulated during CP(CNS). development suggesting that some may play a role only in
A high incidence of raised intracranial pressure and early CP development, whereas others may also be re-hydrocephalus is observed in individuals with syndromic quired for the function of mature CPs.craniosynostosis, a group of disorders characterised by thepremature fusion of one or more cranial sutures [17]. Manyof these syndromes are caused by single point mutations in2 . Materials and methodsfibroblast growth factor receptors (FGFRs), the highaffinity tyrosine kinase receptors that transduce FGF All reagents were purchased from Sigma–Aldrichsignaling. FGF signaling plays multiple roles in vertebrate (Dorset, UK) unless stated otherwise.development, including the CNS. At early stages of CNSdevelopment, FGFs have a role in neural induction [36,11], 2 .1. Animalsand in early patterning of the neural tube [38,8]. At laterstages, FGF signaling is thought to play a role in determin- Out-bred CD-1 albino mice were supplied by Charlesing certain neuronal cell types [49] and in stimulating the River Mouse Farms (Kent, UK), and housed under Homeproliferation of subsets of cells of the CNS during neuro- Office regulations. The morning of detection of a vaginalgenesis [31]. To date, five FGFRs (FGFR1–5) have been plug was designated as embryonic day 0.5 (E0.5). Preg-described, but only FGFR1, 2 and 3 have so far been nant time-mated mice were killed by cervical dislocation.implicated in syndromic craniosynostoses [2,16]. There are In each case, the uterus containing between 9 and 21numerous FGFR isoforms which can alter receptor func- embryos was removed whole and placed in ice-coldtion [40,41]. Alternative splicing of the third extracellular phosphate-buffered saline (PBS). The embryos were dis-immunoglobulin-like loop has been shown to confer sected free of uterine and amniotic tissues and decapitateddifferent ligand-binding specificity, and is often charac- immediately.teristic for a specific cell type [20,44,7,10,1].
Although many groups have studied FGF and FGFR 2 .2. Immunohistochemistryexpression in the adult and the developing CNS, there isconsiderably less direct or precise information regarding Heads of embryos between E12.5 and E16.5 were fixedexpression of either of these families of molecules in the in 4% paraformaldehyde (PFA) in phosphate bufferedCPs. It is clear, however, that in the adult mammal, CPs saline (PBS) at 48C overnight. For E18.5 embryos, theexpress FGF2 [15,23,9] and FGFR2 [48], but not FGFR1 overlying skin was removed prior to fixation in 4% PFA at[48]. In the developing embryo, there are several reports of 48C for 2 days. Fixed samples were immersed in a seriesFGF/FGFR expression in the CPs [31,6,14], and low- of graded alcohols, Histoclear (National Diagnostics, At-affinity heparan sulphate proteoglycan (HSPG) binding lanta, USA) and embedded in paraffin wax (Raymondsites for FGF2 have been identified in the murine CPs at Lamb, Sussex, UK). Serial sections (6mm) were cut andE15.5 [13]. These published data suggest that CPs express transferred to glass slides precoated with 10ml undilutedsome FGFs and FGFRs during various stages of develop- poly-L-lysine. Several slides (two to three) were processedment, and that expression of certain FGF/FGFRs may for immunohistochemistry at the same time for eachpersist in the adult. receptor. Every slide had between three and five sections
The expression patterns of FGFR proteins in the CPs, from the same animal on them, and each one washowever, are generally poorly described, as many reports individually assessed. The experimental series was re-do not distinguish between forebrain and hindbrain CPs; peated at least twice on the same animal, and at least twicefew authors describe differences in expression patterns on animals from different litters. Sections were dewaxedbetween the epithelia and stroma/mesenchymal component for 20 min in two fresh changes of Histoclear andof the CP, and in the literature CPs may be referred to at rehydrated. Endogenous peroxidase activity was quenchedone developmental stage, and then not referred to at other for 15 min in 3% hydrogen peroxide in PBS. Samples werestages analysed. Consequently systematic expression data pretreated by boiling in citric acid buffer at pH 6.0 for 8regarding the FGFRs in the CPs that would allow one to min in a 700-W microwave oven at full power and then leftstart to address the issue of the potential roles played by in running water to cool. In order to eliminate nonspecificdifferent FGFRs in development and function of the CPs antibody interaction with the sample, blocking solutionare not currently available. The aim of our research was to (0.5% bovine serum albumin–10% foetal calf serum inperform a detailed analysis of the expression of FGFR 1, 2, PBS) was applied for 1 h. Sections were incubated in3 and 4 in developing murine forebrain and hindbrain CPs primary anti-FGFR1, anti-FGFR2, anti-FGFR3 or anti-

S. Reid, P. Ferretti / Developmental Brain Research 141 (2003) 15–24 17
FGFR4 affinity-purified rabbit polyclonal antibodies (Santa ACTGGAGCCTTA antisense TGGTTGGCCTGCCCTAT-Cruz Biotechnology, CA, USA), each diluted 1:100 in ATA, FGFR2 IIIc sense AAACCAGCACTGGAGCCTTA,blocking solution at 48C overnight in a humidified antisense CGTGATCTCCTTCTCTCTCACA, FGFR3 IIIbchamber, then washed thoroughly in 0.05% Tween20–PBS sense AGTGGAGCTTGGTCATGGAA antisense TGA-three times, and plain PBS twice. A horseradish per- ACACGCAGCCAAAAG FGFR3 IIIc sense AGTG-oxidase-conjugated goat anti-rabbit antibody (Dako, Cam- GAGCTTGGTCATGCAA antisense ACGCAGAGTGAT-bridge, UK) was applied to each section at a concentration GGGAAAAC, FGFR4 sense AAGTCATCCGTGGC-of 1:200, diluted in blocking solution, for 1 h and washed CACTAC antisense GTCTGCCAAATCCTTGTCGG,off as before. The peroxidase substrate, diaminobenzidine GAPDH sense TTCCAGTATGACTCCACTCACG anti-was applied to each slide for 2 min, and then removed by sense GGATGCAGGGATGATGTTCT.rinsing in distilled water. Slides were counterstained in PCR products were amplified with Taq polymerasemethyl green nuclear stain for 10 min, dehydrated by (Promega) using the manufacturer’s protocol, each at animmersion in butanol for 10 min, then placed in Histoclear annealing temperature of 568C. PCR products were visual-for 20 min prior to mounting in DPX permanent mounting ised on a 1.5% agarose gel containing ethidium bromidemedium. In order to confirm staining specificity, antibody under UV light, and were captured usingpreabsorption for 1 h prior to application of the primary AlphaImager�1200 computer software (Alpha Innotech,antibody was performed for each FGFR using an equal USA). Embryonic heart mRNA was used as a control.dilution of the corresponding peptides mapping to the Detection of GAPDH was used as a positive control. Watercarboxy-terminus used to raise each antibody (Santa Cruz was used in place of cDNA as a negative control.Biotechnology). The specificity of each FGFR antibodywas further confirmed by preabsorption of one FGFRantibody sample with a noncorresponding FGFR peptide. 3 . Results
In order to describe the variation in immunohistochemi-cal staining between tissue structures, and FGFRs, we In order to study the developmental regulation ofallocated an ‘intensity score’ to the CPe, CPm, ependyma FGFRs, we assessed the reactivity of anti-FGFR affinity-and neuronal tissue in each section (Fig. 1). The scoring purified polyclonal antibodies on brain sections containingsystem, ranging from2 (no expression) to111 (strong CP from E12.5, E15 and E18.5 mouse embryos. Theimmunoreactivity), was based on a visual inspection of the specificity of each antibody was confirmed by preabsorp-staining of each of these tissues, at the same magnification, tion with the relevant FGFR peptides.in comparison to one another on the same slide, and incomparison to other slides which had been processed at the3 .1. FGFR1 protein expressionsame time.
At E12.5, the epithelial cells (CPe) and mesenchymal2 .3. Photography cells (CPm) of the CPs were immunoreactive for FGFR1
(Fig. 1A and B). There was no detectable difference inDigital images were captured electronically using a intensity between lateral ventricle CP, and fourth ventricle
Zeiss Kontron ProgRes 3012 digital camera (Imaging CP in any of the sections examined. Similarly, theAssociates, Oxford, UK) and version 2.0 of the associated ependymal cells lining the ventricles expressed FGFR1software. Images were stored asTIFF files using ADOBE protein, as did the neuronal cells of the developing CNS,PHOTOSHOPv6.0 (Adobe Systems, Edinburgh, UK). particularly in the regions closest to the ventricles. Staining
was obliterated when the FGFR1 antibody was preincu-2 .4. RT-PCR bated with FGFR1 peptide (data not shown). At E15.5,
immunoreactivity for FGFR1 was barely detectable in theTotal RNA was extracted from fourth ventricle CPs and CPe and CPm of the lateral and fourth ventricles, com-
homogenised in 1 ml TRI Reagent� (Sigma–Aldrich). pared with the levels observed at E12.5 (Fig. 1C and D,RNA was phenol–chloroform extracted as directed by the respectively). In addition, the ependyma and surroundingmanufacturer. cDNA was synthesised in accordance with CNS did not express FGFR1 at this stage. By contrast, thethe manufacturers instructions using M-MLV reverse tran- developing eye demonstrated positive FGFR1 immuno-scriptase (Promega, Southampton, UK) from 1mg of reactivity, particularly in the cells of the corneal epi-RNA, using custom synthesised oligo-dT primers (Gen- thelium, the mesenchyme of the eyelids, and the neuralosys, Cambridge, UK). PCR was performed using the retina (Fig. 1E). This result confirmed that the immuno-following primer pairs, shown 59 to 39: histochemical technique could reliably detect the FGFR1
FGFR1 IIIb sense GCCACCTGGAGCATCATAAT antigen, but that the cells of the CPe, CPm, CNS andantisense GCTTCAGCCACTGAATGTGAT, FGFR1 IIIc ependyma did not express FGFR1 protein at this stage. Atsense GCCACCTGGAGCATCATAAT antisense CTTC- E18.5 the CPm was not immunoreactive to FGFR1. ThereCAGAACGGTCAACC, FGFR2 IIIb sense AAACCAGC- was slight immunoreactivity to FGFR1 in the CPe of both
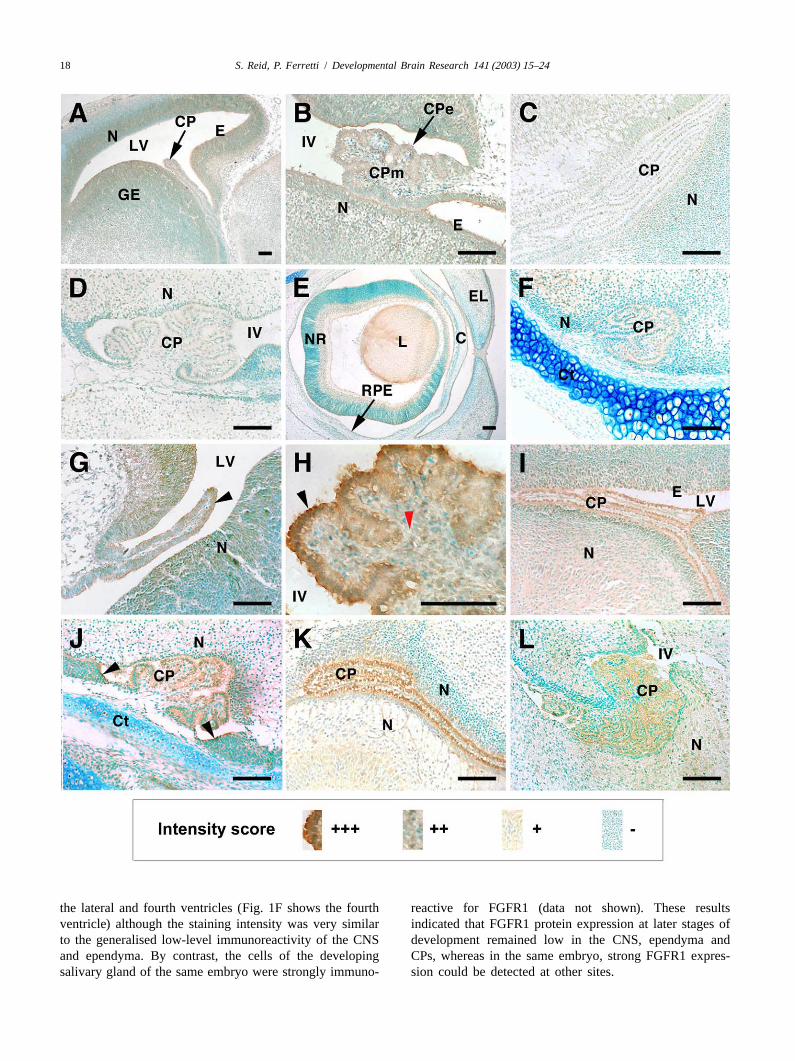
18 S. Reid, P. Ferretti / Developmental Brain Research 141 (2003) 15–24
the lateral and fourth ventricles (Fig. 1F shows the fourth reactive for FGFR1 (data not shown). These resultsventricle) although the staining intensity was very similar indicated that FGFR1 protein expression at later stages ofto the generalised low-level immunoreactivity of the CNS development remained low in the CNS, ependyma andand ependyma. By contrast, the cells of the developing CPs, whereas in the same embryo, strong FGFR1 expres-salivary gland of the same embryo were strongly immuno- sion could be detected at other sites.

S. Reid, P. Ferretti / Developmental Brain Research 141 (2003) 15–24 19
3 .2. FGFR2 protein expression specificity of the FGFR3 antibody. Extranuclear stainingwas observed only in the cytoplasm of cells forming the
At E12.5 the CPe of the lateral and fourth ventricles salivary gland in the developing mandible (data notwere highly immunoreactive for FGFR2 (Fig. 1G–H). The shown). As seen at E12.5, all nuclei were labeled forCPm was less immunoreactive than the CPe. In addition to FGFR3 in the embryonic head at E15.5 including the CPethe CPs, the ependymal cells lining the ventricles and the and CPm (Fig. 2E and F). At E18.5 nuclear labelling ofneuronal cells of the CNS were also positive for FGFR2 FGFR3 protein was again predominant in the CNS, theprotein at this stage. At E15.5 there was a change in ependyma, and the CPs in the lateral and fourth ventriclesFGFR2 immunoreactivity in the region of the CPs. The (not shown).CPe of the lateral ventricle remained strongly immuno-reactive for FGFR2 (Fig. 1I). The most intense staining for 3 .4. FGFR4 protein expressionFGFR2 was located at the apical surfaces of the CPe. Inthe fourth ventricle, FGFR2 immunoreactivity was de- At E12.5 FGFR 4 protein expression was present in thetected in all CPe cells, including the ‘stalk’, at the junction CPe, CPm and ependymal cells in the lateral ventriclesbetween the ependyma and CPe (Fig. 1J). FGFR2 im- (Fig. 2G). FGFR4 expression was distributed throughoutmunoreactivity was less intense in the CPm of the lateral the cytoplasm of the CPe cells, with strong immuno-and fourth ventricle CPs than the CPe. The ependymal reactivity on both apical and basal sides of the nuclei. Thecells and the neuronal cells in the CNS did not express ependymal cells were most immunoreactive in the regionFGFR2 protein strongly at E15.5. Although a low level of of their ventricular surfaces. The neuronal cells were lessperoxidase staining was apparent in these tissues, theimmunoreactive than the CP and ependyma. The CPm wasintensity of staining was considerably less than that also positively stained for FGFR4 in the lateral ventricles.observed in the CPe. At E18.5, there was strong immuno- The fourth ventricle CP showed similar immunoreactivityreactivity for FGFR2 in the lateral and fourth ventricle to the CPe of the lateral ventricles, with FGFR4 immuno-CPe, particularly towards the apical surfaces of the cells reactivity throughout the cytoplasm of the CPe (Fig. 2H).(Fig. 1K–L). The CNS tissue closest to the ventricles was There was intense staining for FGFR4 on the apicalnegative for FGFR2 at this later stage of gestation. surfaces of some CPe. Similar to the results for LV
ependymal cells, the ependymal cells in the fourth ventri-cle were strongly immunoreactive for FGFR4 on their
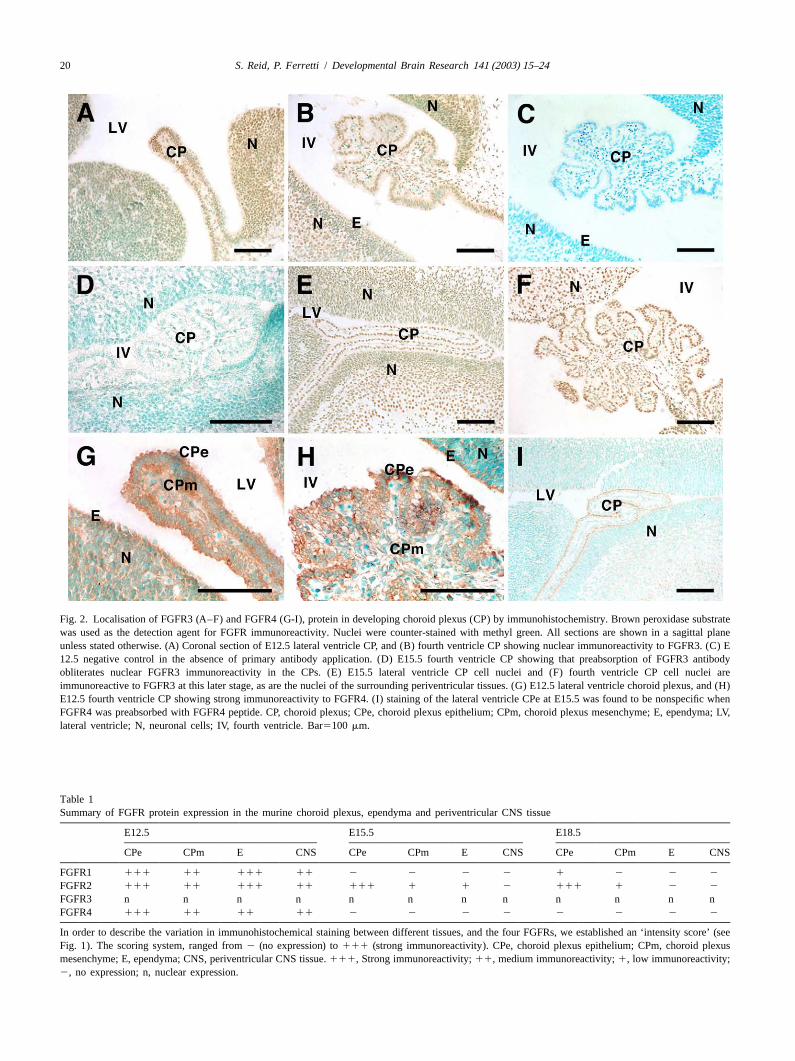
3 .3. FGFR3 protein expression apical surfaces. At E15.5 the expression of FGFR4 proteinwas not detected in periventricular tissues of the CNS,
FGFR3 immunoreactivity was predominantly localised including the CPe, CPm and ependyma. Apical surfaceto the nuclei of all cells in the CNS, including the staining of the CPe at E15.5 (Fig. 2I) and E18.5 (notependyma, and the CPe and CPm at E12.5 (Fig. 2A andshown) was found to be nonspecific when the antibody wasB). Fig. 2C is a negative control, which has no detectable preabsorbed with FGFR4 peptide. Discrete immuno-levels of peroxidase reaction, indicating that there was no reactivity for FGFR4 protein however, was detected inimmunoreactivity in the absence of the primary anti- outer neuronal layers of the frontal cortex at these stagesFGFR3 antibody. Cytoplasmic immunoreactivity was not and was abolished by preabsorption with the FGFR4observed in any of these regions at this stage in develop-peptide.ment. This nuclear FGFR3 staining however, was com- The results of our immunohistochemical analyses show-pletely blocked by preabsorption of the antibody with ing changes in FGFR expression with development areFGFR3 peptide (Fig. 2D), demonstrating the binding summarised in Table 1.
Fig. 1. Localisation of FGFR1 (A–F) and FGFR2 (G–L), protein in developing choroid plexus (CP) by immunohistochemistry. Brown peroxidasesubstrate was used as the detection agent for FGFR immunoreactivity. Nuclei were counter-stained with methyl green. All sections are shown in a sagittalplane unless stated otherwise. (A) E12.5 lateral ventricle CP, and (B) fourth ventricle CP showing immunoreactivity to FGFR1. (C) E15.5 lateral ventricleCP, and (D) fourth ventricle CP are not immunoreactive to FGFR1 at this later stage. (E) Developing eye at E15.5 identifies FGFR1 immunoreactivitypresent in layers of the neural retina, the corneal epithelium, and in the mesenchymal tissue of the eyelid. (F) E18.5, fourth ventricle CP and surroundingCNS tissues showing very low levels of immunoreactivity to FGFR1. (G) Coronal section of lateral ventricle CP at E12.5 demonstrates FGFR2immunoreactivity, particularly at distal CPe regions (arrowhead). (H) High-power micrograph of E12.5 fourth ventricle CP also shows immunoreactivity toFGFR2 particularly in distal CPe cells (black arrowhead). The CPm (red arrowhead) is less immunoreactive for FGFR2. (I) E15.5 lateral ventricle CPshowing FGFR2 immunoreactivity persists in the CP, and to a lesser degree in the surrounding ependyma. CNS tissues are not immunoreactive to FGFR2at this stage. (J) E15.5 fourth ventricle CP is immunoreactive to FGFR2, including the CP ‘stalks’, where the ependymal cells and the CPe cells form acontinuum (arrowheads). At E18.5, the CPe is immunoreactive to FGFR2 in both the lateral (K) and fourth ventricle (L) CPe but not in the CPm,ependyma or periventricular CNS. C, cornea; CP, choroid plexus; CPe, choroid plexus epithelium; CPm, choroid plexus mesenchyme; Ct, cartilage; E,ependyma; EL, eyelid; L, lens; LV, lateral ventricle; N, neuronal cells; NR, neural retina; RPE, retinal pigmented epithelium (detached), IV, fourth ventricle;bars5100mm. The box at the bottom contains examples of our immunoreactivity intensity scores described in the materials and methods section and usedto evaluate changes in FGFR expression;111, strong immunoreactivity;11, medium immunoreactivity;1, low immunoreactivity;2, no expression.
20 S. Reid, P. Ferretti / Developmental Brain Research 141 (2003) 15–24
Fig. 2. Localisation of FGFR3 (A–F) and FGFR4 (G-I), protein in developing choroid plexus (CP) by immunohistochemistry. Brown peroxidase substratewas used as the detection agent for FGFR immunoreactivity. Nuclei were counter-stained with methyl green. All sections are shown in a sagittal planeunless stated otherwise. (A) Coronal section of E12.5 lateral ventricle CP, and (B) fourth ventricle CP showing nuclear immunoreactivity to FGFR3. (C) E12.5 negative control in the absence of primary antibody application. (D) E15.5 fourth ventricle CP showing that preabsorption of FGFR3 antibodyobliterates nuclear FGFR3 immunoreactivity in the CPs. (E) E15.5 lateral ventricle CP cell nuclei and (F) fourth ventricle CP cell nuclei areimmunoreactive to FGFR3 at this later stage, as are the nuclei of the surrounding periventricular tissues. (G) E12.5 lateral ventricle choroid plexus, and (H)E12.5 fourth ventricle CP showing strong immunoreactivity to FGFR4. (I) staining of the lateral ventricle CPe at E15.5 was found to be nonspecific whenFGFR4 was preabsorbed with FGFR4 peptide. CP, choroid plexus; CPe, choroid plexus epithelium; CPm, choroid plexus mesenchyme; E, ependyma; LV,lateral ventricle; N, neuronal cells; IV, fourth ventricle. Bar5100 mm.
Table 1Summary of FGFR protein expression in the murine choroid plexus, ependyma and periventricular CNS tissue
E12.5 E15.5 E18.5
CPe CPm E CNS CPe CPm E CNS CPe CPm E CNS
FGFR1 111 11 111 11 2 2 2 2 1 2 2 2
FGFR2 111 11 111 11 111 1 1 2 111 1 2 2
FGFR3 n n n n n n n n n n n nFGFR4 111 11 11 11 2 2 2 2 2 2 2 2
In order to describe the variation in immunohistochemical staining between different tissues, and the four FGFRs, we established an ‘intensity score’ (seeFig. 1). The scoring system, ranged from2 (no expression) to111 (strong immunoreactivity). CPe, choroid plexus epithelium; CPm, choroid plexusmesenchyme; E, ependyma; CNS, periventricular CNS tissue.111, Strong immunoreactivity;11, medium immunoreactivity;1, low immunoreactivity;2, no expression; n, nuclear expression.

S. Reid, P. Ferretti / Developmental Brain Research 141 (2003) 15–24 21
3 .5. Expression of FGFR isoforms by RT-PCR 4 . Discussion
In order to assess whether specific FGFR isoforms were The data presented here clearly show that FGFRs arepreferentially expressed in the CP, we used RNA extracts differently regulated during development of the CP, sug-of E12.5 fourth ventricle CPs and primers that specifically gesting that FGF signaling is important for CP develop-amplify the different spliced immunoglobulin III domains, ment, and that different FGFRs may play different roles inIIIb and IIIc, of FGFR1, FGFR2 and FGFR3. Expression this process.of FGFR4, which does not have such spliced forms, wasalso assessed in the E12.5 CP.
Consistent with the immunohistochemical analysis, 4 .1. FGFR1 and FGFR4 expression is downregulatedFGFR1, FGFR2, FGFR3 and FGFR4 mRNAs were de- with CP developmenttected at this stage of development (Fig. 3). Interestingly,both IIIb and IIIc isoforms of FGFR2 were expressed In agreement with our findings in the mouse, FGFR1strongly, whereas only the IIIc isoforms of FGFR1 and immunoreactivity has been identified in the CNS of theFGFR3 were detected. FGFR4 mRNA was also clearly early rat embryo, particularly in periventricular regions ofdetected, but the band intensity was always weaker than the developing CNS at E11.5 [31]. This group described athose for the other receptors, suggesting a lower level of gradual downregulation of FGFR1 mRNA transcripts inexpression of FGFR4 in the CP. the CNS of the embryonic rat by E17.5 [31]. A similar
The lateral ventricle CPs were not analysed for mRNA downregulation of FGFR1 mRNA transcripts in the CPsexpression using RT-PCR. Whilst we cannot rule out the and CNS was also reported in a study of human FGFR1possibility that there may be differences in the levels of mRNA expression between 10 and 18 weeks gestation [6].FGFR mRNA expression between the lateral and fourth Complementary to these findings, we found that FGFR1ventricle CPs, this seemed unlikely, as our immunohistoch- immunoreactivity declined in the CNS and CPs during theemical results indicated that the intensity of FGFR anti- second half of gestation. This immunohistochemical databody reactivity was comparable between forebrain and for FGFR1 expression agrees well with previously pub-hindbrain CPs. lished reports, supporting the conclusions of this study that
Fig. 3. Expression of FGFR isoforms in E12.5 choroid plexus (CP) from the fourth ventricle analysed by RT-PCR. Embryonic heart (Ht) mRNA was usedas a control. Bands of the predicted sizes for FGFR1 IIIb (243 bp) and IIIc (450 bp), FGFR2 IIIb (422 bp) and IIIc (480 bp), FGFR3 IIIb (471 bp) and IIIc(428 bp), and FGFR4 (397 bp) were detected in the E12.5 CP. GAPDH (491 bp) was used as a positive control. Water was used in place of cDNA asnegative controls for each primer pair on each occasion (2ve).

22 S. Reid, P. Ferretti / Developmental Brain Research 141 (2003) 15–24
there is a reduction in FGFR1 immunoreactivity in the CPs isoform of FGFR3 was more discrete. We did not detectduring the second half of murine gestation. In addition we cytoplasmic staining for FGFR3 in the developing CP atfound that the predominant FGFR1 isoform expressed in any of the stages tested. Cell signaling via exogenousthe CP at E12.5 is the IIIc variant. FGFs requires transmembranous FGFR3, and so it is most
There are conflicting reports regarding FGFR4 expres- likely that the cells of the CP would not respond tosion in the embryo. One group isolated and cloned FGFR4, extracellular FGFs via FGFR3, due to its nuclear location.and found that mRNA expression was confined to the gut, FGFR3 mRNA transcripts encoding the transmembranemyotomes and skeletal muscles at E14.5 whereas the CNS and juxtamembrane portions of FGFR3 have been detecteddid not express FGFR4 [37]. More recently, mRNA in the germinal epithelium of the murine forebrain andtranscripts have been identified using in situ hybridisation, hindbrain at E14.5 [29], however we did not detect FGFR3in the ‘cells of the ventricular region’ and outer cortical protein in this region using immunohistochemistry.cells of the CNS in the embryonic mouse brain at E16.5,but no reference was made to FGFR4 expression in theCPs in this report [26]. In our study, immunohistochemical 4 .3. FGFR2 protein expression is maintained throughoutanalysis suggested that protein expression was consider-developmentable in the CPs at E12.5, when the transcript could also bedetected, but not at E15.5. We found that FGFR4 was not FGFR2 expression in the CPs differed from the expres-expressed by the CPs at later stages of gestation. In sion profiles of the other FGFRs examined. We found thataddition, FGFR4 mRNA has not been detected using both isoforms of FGFR2, IIIb and IIIc, were expressed inRT-PCR analysis in the CNS tissues of the adult rat [48], the E12.5 CP, and that FGFR2 protein was present in theconsistent with our finding that FGFR4 is not expressed by CPe and CPm throughout the second half of gestation. Itthe CPs beyond midgestation in the mouse. was particularly interesting to us that FGFR2 immuno-
reactivity became restricted to the CPe cells as gestationproceeded. In agreement with our findings, one group
4 .2. FGFR3 protein expression in the CP is confined to identified discrete FGFR2 transcripts in mouse CP but notnuclei in the CNS tissue, at E16.5 [28]. Similarly, FGFR2 IIIb
and IIIc transcripts in the CP of the human embryoThe nuclear reactivity that we observed with the anti- between 10 and 18 weeks gestation have also been
body raised against FGFR3 in the brain was unexpected, described [6]. Using in situ hybridisation, these authorsgiven that FGFRs are transmembrane proteins transducing reported that FGFR2 IIIb and FGFR2 IIIc mRNA signalssignals from the extracellular environment to the nucleus. were particularly strong in the CPe. Our immunohistoch-Despite numerous attempts to vary the immunohistochemi- emical results complement these findings, and these datacal method, there was always similar nuclear expression of combined provide strong evidence that FGFR2 expressionthis protein in the CP at all stages tested, though cyto- is a characteristic feature of CPe at later gestational stages.plasmic staining was observed in other tissues. The fact FGFR2 mRNA expression has also been described in CPsthat the FGFR3 peptide used to raise this antibody of adult rats [48], adding to the hypothesis that the CPscompletely blocked its binding and that we could detect have the capacity to respond to FGF stimulation through-expression of the IIIc isoform of FGFR3 by RT-PCR in out life via the FGFR2 receptor. It is likely therefore thatE12.5 CPs, strongly indicate that FGFR3 is indeed ex- expression of this receptor molecule in the CPs persistspressed in the developing CP. Interestingly, there are other throughout life and may be necessary for the normalinstances where nuclear localisation of FGFR3 protein has functionality of the CPs during postnatal life as well asbeen reported, such as in breast epithelial cells [21]. during development.Further analysis by this group revealed that there are two The expression pattern of FGFR2 in the CPs duringisoforms of FGFR3, of 135- and 110-kDa. The 110-kDa development is particularly interesting in the context ofisoform was found to lack exons 7 and 8, which resulted in syndromic craniosynostosis. The FGFR2 gene has pointthe deletion of the transmembrane domain of FGFR3, but mutations at various locations in children with Crouzon,an intact kinase domain. This was the isoform located in Pfeiffer, Jackson Weiss and Beare Stevenson cutis gyratathe nucleus of cultured breast epithelial cells, although its syndromes [32,24,46,18,2]. Mutations in FGFR1 andfunction, if any, is unknown at present. By contrast, the FGFR3 can also occur in Pfeiffer syndrome and Crouzon135-kDa isoform of FGFR3 was located primarily at the patients with acanthosis nigricans, respectively [24,35,30].cell surface. The antibody we used for FGFR3 analysis Out of all individuals with syndromic craniosynostosis,was raised against the intracellular kinase domain of those with a Crouzon and Pfeiffer phenotype due toFGFR3, and so would not distinguish between the two mutations in FGFR2 appear to be most at risk of raiseddifferent isoforms. It is possible that the immunological intracranial pressure [43,45,22]. As FGFR2 receptor ex-technique detected the 110-kDa isoform in the nuclei of pression is maintained in the CPe throughout the secondthe cells in the CNS, whereas the signal for the 135-kDa half of gestation and adult, we hypothesise that abnormal

S. Reid, P. Ferretti / Developmental Brain Research 141 (2003) 15–24 23
with the cerebrospinal fluid in man and other species. Part I: TheFGFR2 signaling in the CPs during development inventricular system, meninges and choroid plexuses, Arch. Anat.individuals with syndromic craniosynostosis might influ-Cytol. Pathol. 46 (1998) 153–169.
ence development and/or function of the CPs, and contrib- [5] J.G. Chamberlain, Analysis of developing ependymal and choroidalute to the pathology of raised ICP in these individuals. surfaces in rat brains using scanning electron microscopy, Dev. Biol.
The expression of FGFRs during development of the 31 (1973) 22–30.[6] C.T.J. Chan, P. Thorogood, Plieotropic features of syndromicCPs suggests that these receptors are important factors in
craniosynostosis correlate with differential expression of fibroblastthe normal ontogeny of the murine CP, particularly at earlygrowth factor receptors 1 and 2 during human craniofacial develop-stages, when the CPs emerge and become morphologicallyment, Pediatr. Res. 45 (1999) 46–53.
distinct, tight junctions are forming at CPe borders, and [7] A.T. Chellaiah, D.G. McEwen, S. Werner, J. Xu, D.M. Ornitz,cell–cell interactions in various regions of the ependyma Fibroblast growth factor receptor (FGFR) 3. Alternative splicing in
immunoglobulin-like domain III creates a receptor highly specificare changing. We found that expression of FGFRs 1 and 4for acidic FGF/FGF-1, J. Biol. Chem. 269 (15) (1994) 11620–in the CP was short-lived whereas FGFR2 expression11627.persisted to late gestational stages, suggesting a more
[8] P.H. Crossley, S. Martinez, G.R. Martin, Midbrain developmentlong-term function for this receptor. Though strong im- induced by FGF8 in the chick embryo, Nature 380 (1996) 66–68.munoreactivity for FGF2 has been reported in the rat CPe [9] P. Cuevas, C. Revilla, O. Herreras, C. Largo, G. Gimenez-Gallego,
Neuroprotective effect of acidic fibroblast growth factor on seizure-at E17.5 [31], the physiological role of FGF signaling inassociated brain damage, Neurol. Res. 16 (1994) 365–369.the developing CPs has yet to be determined. In adult
[10] L. De Moerlooze, B. Spencer-Dene, J. Revest, M. Hajihosseini, I.models, exogenous infusion of FGF2 into the cerebrospinalRosewell, C. Dickson, An important role for the IIIb isoform of
fluid of rats [19] and common marmosets [27] causes fibroblast growth factor receptor 2 (FGFR2) in mesenchymal–hydrocephalus. This may point to a role of FGF signaling epithelial signaling during mouse organogenesis, Development 127
(3) (2000) 483–492.in the control of secretory function by the CPe. However,[11] T. Doniach, Basic FGF as an inducer of anteroposterior neuralcontrary to this hypothesis, it has been found that FGF2
pattern, Cell 83 (1995) 1067–1070.infused into the lateral ventricles of rats at high doses[12] K.M. Dziegielewska, G.W. Knott, N.R. Saunders, The nature and
reduces the rate of CSF reabsorption into the bloodstream, composition of the internal environment of the developing brain,by the formation of collagen deposits around CSF drainage Cell Mol. Neurobiol. 20 (2000) 41–56.
[13] N.A. Fayein, Y. Courtios, J.-C. Jeanny, Basic fibroblast growthsites [19]. It remains unclear as to what function FGFfactor high and low affinity binding sites in developing mouse brain,signaling might have in the embryonic CPe. Recently, wehippocampus and cerebellum, Biol. Cell 76 (1992) 1–13.used an in vitro CPe culture model [42] to show that in
[14] P.W. Finch, G.R. Cunha, J.S. Rubin, J. Wong, D. Ron, Pattern ofvitro FGF2 affects the growth of fluid filled CPe vesicles keratinocyte growth factor and keratinocyte growth factor receptor[34]. Work is in progress to characterise the nature of this expression during mouse fetal development suggests a role in
mediating morphogenetic mesenchymal–epithelial interactions, Dev.response in more detail.Dyn. 203 (1995) 223–240.
[15] K. Fuxe, B. Tinner, M. Zoli, R.F. Pettersson, A. Baird, G. Biagini,G. Chadi, L.F. Agnati, Computer-assisted mapping of basic fi-broblast growth factor immunoreactive nerve cell populations in the
A cknowledgements rat brain, J. Chem. Neuroanat. 11 (1996) 13–35.[16] K. Gaudenz, E. Roessler, S. Vainikka, K. Alitalo, M. Muenke,
Analysis of patients with craniosynostosis syndromes for aWe thank Conrad Johanson for reading the manuscriptpro246Arg mutation of FGFR4, Mol. Genet. Metab. 64 (1) (1998)and sharing unpublished observations and Andrew Copp76–79.for useful discussions.This study was supported by Great
[17] R.J. Gorlin, M.M. Cohen, L.S. Levin, Syndromes with craniosynos-Ormond Street Hospital for Children NHS Trust / Institute tosis: general aspects and well known syndromes, in: Syndromes ofof Child Health Science Development Initiative. the Head and Neck, Oxford University Press, Oxford, 1990, pp.
519–521.[18] E.W. Jabs, L. Xiang, A.F. Scott, G. Meyers, W. Chen, M. Eccles,
L.R. Charnas, C.E. Jackson, M. Jaye, Jackson-Weiss and Crouzonsyndromes are allelic with mutations in fibroblast growth factor
R eferences receptor 2, Nat. Genet. 8 (1994) 275–278.[19] C.E. Johanson, J. Szmydynger-Chodobska, A. Chodobski, A. Baird,
[1] H.D. Beer, L. Vindevoghel, M.J. Gait, J.M. Revest, D.R. Duan, I. P. McMillan, E.G. Stopa, Altered formation and bulk absorption ofMason, C. Dickson, S. Werner, Fibroblast growth factor (FGF) cerebrospinal fluid in FGF-2-induced hydrocephalus, Am. J. Physiol.receptor 1–IIIb is a naturally occurring functional receptor for FGFs 277 (1999) R263–R271.that is preferentially expressed in the skin and the brain, J. Biol. [20] D.E. Johnson, J. Lu, H. Chen, S. Werner, L.T. Williams, The humanChem. 275 (2000) 16091–16097. fibroblast growth factor receptor genes: a common structural ar-
[2] D. Burke, D. Wilkes, T.L. Blundell, S. Malcolm, Fibroblast growth rangement underlies the mechanisms for generating receptor formsfactor receptors: lessons from the genes, Trends Biol. Sci. 23 (1998) that differ in their third immunoglobulin domain, Mol. Cell Biol. 1159–62. (9) (1991) 4627–4634.
[3] M. Catala, Carbonic anhydrase activity during development of the [21] C.L. Johnston, H.C. Cox, J.J. Gomm, R.C. Coombes, Fibroblastchoroid plexus in the human fetus, Childs Nerv. Syst. 13 (1997) growth factor receptors (FGFRs) localize in different cellular364–368. compartments. A splice variant of FGFR-3 localizes to the nucleus,
J. Biol. Chem. 270 (51) (1995) 30643–30650.[4] M. Catala, Embryonic and fetal development of structures associated

24 S. Reid, P. Ferretti / Developmental Brain Research 141 (2003) 15–24
[22] S.H. Kan, N. Elanko, D. Johnson, L. Cornejo-Roldan, J. Cook, E.W. [36] J. Slack, Role of fibroblast growth factors as inducing agents inReich, S. Tomkins, A. Verloes, S.R. Twigg, S. Rannan-Eliya, D.M. early embryonic development, Mol. Reprod. Dev. 39 (1994) 118–McDonald-McGinn, E.H. Zackai, S.A. Wall, M. Muenke, A.O. 125.Wilkie, Genomic screening of fibroblast growth-factor receptor 2 [37] K.L. Stark, J.A. McMahon, A.P. McMahon, FGFR-4, a new memberreveals a wide spectrum of mutations in patients with syndromic of the fibroblast growth factor receptor family, expressed in thecraniosynostosis, Am. J. Hum. Genet. 70 (2) (2002) 472–486. definitive endoderm and skeletal muscle lineages of the mouse,
[23] H. Kataoka, E. Yamada, F. Hazama, Increased basic fibroblast Development 113 (1991) 641–651.growth factor immunoreactivity in the brain of stroke-prone sponta- [38] K.G. Storey, A. Goriely, C.M. Sargent, J.M. Brown, H.D. Burns,neously hypertensive rats, Acta Neuropathol. (Berl) 88 (1994) 7–13. H.M. Abud, J.K. Heath, Early posterior neural tissue is induced by
[24] M. Muenke, U. Schell, A. Hehr, N.H. Robin, H.W. Losken, A. FGF in the chick embryo, Development 125 (1998) 473–484.Schinzel, L.J. Pulleyn, P. Rutland, W. Reardon, S. Malcolm et al., A [39] R.R. Sturrock, A morphological study of the development of thecommon mutation in the fibroblast growth factor receptor 1 gene in mouse choroid plexus, J. Anat. 129 (1979) 777–793.Pfeiffer syndrome, Nat. Genet. 8 (1994) 269–274. [40] G. Szebenyi, J.F. Fallon, Fibroblast growth factors as multifunction-
[25] M.G. Netsky, S. Shuangshoti, in: The Choroid Plexus in Health and al signaling factors, Int. Rev. Cytol. 185 (1999) 45–106.Disease, Wright, Bristol, 1975, pp. 3–18. [41] S. Takaishi, M. Sawada, Y. Morita, H. Seno, H. Fukuzawa, T. Chiba,
[26] K. Ozawa, T. Uruno, K. Miyakawa, M. Seo, T. Imamura, Expres- Identification of a novel alternative splicing of human FGF receptorsion of the fibroblast growth factor family and their receptor family 4: soluble-form splice variant expressed in human gastrointestinalgenes during mouse brain development, Brain Res. Mol. Brain Res. epithelial cells, Biochem. Biophys. Res. Commun. 267 (2) (2000)41 (1996) 279–288. 658–662.
[27] R.K. Pearce, P. Collins, P. Jenner, C. Emmett, C.D. Marsden,[42] T. Thomas, E. Stadler, M. Dziadek, Effects of the extracellular
Intraventricular infusion of basic fibroblast growth factor (bFGF) inmatrix on fetal choroid plexus epithelial cells: changes in morpholo-
the MPTP-treated common marmoset, Synapse 23 (1996) 192–200.gy and multicellular organization do not affect gene expression,
[28] K.G. Peters, S. Werner, G. Chen, L.T. Williams, Two FGF receptorExp. Cell Res. 203 (1992) 198–213.
genes are differentially expressed in epithelial and mesenchymal[43] D. Thompson, W. Harkness, B. Jones, S. Gonzalez, U. Andar, R.
tissues during limb formation and organogenesis in the mouse,Hayward, Subdural intracranial pressure monitoring in craniosynos-
Development 114 (1992) 233–243.tosis: its role in surgical management, Child’s Nerv. Syst. 11 (1995)
[29] K. Peters, D. Ornitz, S. Werner, L. Williams, Unique expression269–275.
pattern of the FGF receptor 3 gene during mouse organogenesis,[44] S. Werner, D.S. Duan, C. de Vries, K.G. Peters, D.E. Johnson, L.T.Dev. Biol. 155 (1993) 423–430.
Williams, Differential splicing in the extracellular region of fi-[30] K.A. Przylepa, W. Paznekas, M. Zhang, M. Golabi, W. Bias, M.J.broblast growth factor receptor 1 generates receptor variants withBamshad, J.C. Carey, B.D. Hall, R. Stevenson, S. Orlow, M.M.-J.different ligand-binding specificities, Mol. Cell Biol. 12 (1) (1992)Cohen, E.W. Jabs, Fibroblast growth factor receptor 2 mutations in82–88.Beare-Stevenson cutis gyrata syndrome, Nat. Genet. 13 (1996)
[45] A.O. Wilkie, Craniosynostosis: genes and mechanisms, Hum. Mol.492–494.Genet. 6 (10) (1997) 1647–1656.[31] R. Raballo, J. Rhee, R. Lyn-Cook, J.F. Leckman, M.L. Schwartz,
[46] A.O. Wilkie, S.F. Slaney, M. Oldridge, M.D. Poole, G.J. Ashworth,F.M. Vaccarino, Basic fibroblast growth factor (Fgf2) is necessaryA.D. Hockley, R.D. Hayward, D.J. David, L.J. Pulleyn, P. Rutland etfor cell proliferation and neurogenesis in the developing cerebralal., Apert syndrome results from localized mutations of FGFR2 andcortex, J. Neurosci. 20 (2000) 5012–5023.is allelic with Crouzon syndrome, Nat. Genet. 9 (2) (1995) 165–[32] W. Reardon, R.M. Winter, P. Rutland, L.J. Pulleyn, B. Jones, S.172.Malcolm, Mutations in the fibroblast growth factor receptor 2 gene
[47] J. Wilting, B. Christ, An experimental and ultrastructural study oncause Crouzon syndrome, Nat. Genet. 8 (1994) 98–103.the development of the avian choroid plexus, Cell Tissue Res. 255[33] S.L. Reid, A. Copp, P. Ferretti, The effect of FGF-2 on developing(1989) 487–494.choroid plexus in vitro. Abstract, Eur. J. Neurosci. 12 (Suppl. 11)
[48] N. Yazaki, Y. Hosoi, K. Kawabata, A. Miyake, M. Minami, M.(2000) 163.2.Satoh, M. Ohta, T. Kawasaki, N. Itoh, Differential expression[34] N.R. Saunders, M.D. Habgood, K.M. Dziegielewska, Barrier mecha-patterns of mRNAs for members of the fibroblast growth factornisms in the brain II: immature brain, Clin. Exp. Pharmacol. Physiolreceptor family, FGFR-1–FGFR-4, in rat brain, J. Neurosci. Res. 3726 (1999) 85–91.(1994) 445–452.[35] U. Schell, A. Hehr, G.J. Feldman, N.H. Robin, E.H. Zackai, C.
Die-Smulders, D.H. Viskochil, J.M. Stewart, G. Wolff, H. Ohashi, [49] W. Ye, K. Shimamura, J.L. Rubenstein, M.A. Hynes, A. Rosenthal,Mutations in FGFR1 and FGFR2 cause familial and sporadic FGF and Shh signals control dopaminergic and serotonergic cell fatePfeiffer syndrome, Hum. Mol. Genet. 4 (1995) 323–328. in the anterior neural plate, Cell 93 (1998) 755–766.





























