Embed Size (px)
Citation preview

CELL STRUCTURE AND FUNCTION 25: 195–204 (2000)
© 2000 by Japan Society for Cell Biology
195
Differential Expressions of BMP Family Genes during Chondrogenic Differentiation of Mouse ATDC5 Cells
Haruhiko Akiyama1, Chisa Shukunami2, Takashi Nakamura1, and Yuji Hiraki2, �
1Department of Orthopedic Surgery, Graduate School of Medicine, and 2Department of Molecular
Interaction and Tissue Engineering, Institute for Frontier Medical Sciences, Kyoto University, Kyoto 606-
8507, Japan
ABSTRACT. Clonal cell line ATDC5 enables the monitoring of the early- and late-phase chondrogenic
differentiation in a single culture. Undifferentiated ATDC5 cells differentiate into type II collagen expressing
chondrocytes through a cellular condensation stage (early-phase differentiation) and then to type X collagen-
expressing hypertrophic chondrocytes (late-phase differentiation). Progression of cellular differentiation was
accelerated by the activation of bone morphogenetic protein (BMP) signaling. ATDC5 cells expressed transcripts
for at least four members of the BMP family. The BMP-4 transcripts were expressed in all stages of differentiation,
as were transcripts for BMP type IA receptor (ALK-3) and BMP type II receptor. In contrast, transcripts for Growth/
Differentiation factor-5 (GDF-5) were induced during a cellular condensation, and those for BMP-6 were induced
during the formation of cartilage nodules, and declined as the differentiated ATDC5 cells became hypertrophic,
and BMP-7 transcripts were only detected after cells became calcified. Exogenously added BMP-4 indeed
promoted the early-phase differentiation. Late-phase differentiation of cells was also stimulated by BMP-4 and
BMP-6. Thus, the cumulative increase in BMP signaling promoted the sequential transitions of differentiation
steps of cells. These results indicate that the coordinated expressions of endogenous BMPs are involved in the
progression of chondrogenic differentiation in ATDC5 cells.
Key words: cartilage differentiation/BMP/ATDC5/endochondral bone formation
The bone morphogenetic protein (BMP) family plays a vital
role in skeletal development and are capable of inducing
ectopic bone formation through an endochondral pathway
which includes a cascade of events such as the recruitment
of progenitor cells, cellular condensation to form cartilagi-
nous bone precursors, vascular invasion, the replacement of
cartilage by bone, and marrow development (Rosen and
Thies, 1992). Several BMP family genes are expressed
at sites of bone formation with a unique spatiotemporal
pattern, suggesting that the coordinated expression of these
molecules is crucial for the cascade of events underlying
endochondral bone formation. Transcripts for BMP-2
and BMP-4 genes are localized to precartilaginous
mesenchymal condensation, and then the perichondrium
(Lyons, et al., 1990). Growth/differentiation factor (GDF)-5
is expressed predominantly in the precartilaginous
mesenchymal condensation and the cartilaginous cores of
the developing long bone (Storm, et al., 1994). In contrast,
BMP-6 and BMP-7 genes are expressed in mature
chondrocytes (Carey and Liu, 1995; Vortkamp, et al., 1996;
Vukicevic, et al., 1994).
In our previous studies (Shukunami, et al., 1997;
Shukunami, et al., 1996), we demonstrated that the clonal
cell line ATDC5 enables the monitoring of the multistep
chondrogenic differentiation in a single culture. In the
presence of insulin, chondrogenic differentiation is induced
to form cartilage nodules through cellular condensation
(early-phase differentiation). When the formation of carti-
lage nodules is completed, the cells are then converted
to type X collagen-expressing hypertrophic chondrocytes
followed by mineralization (late-phase differentiation).
However, insulin by itself induced only a cellular conden-
sation in the cultures of undifferentiated ATDC5 cells.
* To whom correspondence should be addressed.Tel and Fax: �81�75�751�4633E-mail: [email protected]: ActR, activin receptor; ALP, alkaline phosphatase; BMP,
bone morphogenetic protein; FBS, fetal bovine serum; GDF, growth/differ-entiation factor; G3PDH, glyceradehyde 3- phosphate dehydrogenase; OP,osteogenic protein; PTH, parathyroid hormone; RT-PCR, reverse transcrip-tion-polymerase chain reaction.

H. Akiyama et al.
196
Instead, exogenously added BMP-2 directly induced
chondrogenesis by skipping out the condensation stage
(Shukunami, et al., 1998). In order to decipher mechanism
underlying the sequential differentiation of ATDC5 cells,
we studied the temporal patterns during differentiation of
cells in the presence of insulin and the actions of BMP
signaling molecules.
Materials and Methods
Materials
Human recombinant BMP-6 was a generous gift from Dr. J. M.
Wozney (Genetics Institute Inc., Cambridge, MA, USA). Xenopus
recombinant BMP-4 homodimer and BMP-4/7 heterodimer were
kindly provided by Takeda Chemical Industries Ltd. (Osaka,
Japan).
Cell culture
ATDC5 cells were maintained in a medium consisting of a 1:1
mixture of DME and Ham’s F-12 (DME/F12) medium (Flow
Laboratories, Irvine, U.K.) containing 5% fetal bovine serum
(FBS; JRH Biosciences, Lenexa, KS, USA), and ITS, i.e., 10 �g/
ml bovine insulin (I: Wako Pure Chemical, Osaka, Japan), 10 �g/
ml human transferrin (T: Boehringer Mannheim, Gaithersburg,
MD), and 3�10�8 M sodium selenite (S: Sigma, St. Louis,
MO, USA), as previously described (Shukunami, et al., 1997;
Shukunami, et al., 1996). In brief, the inoculum size of the cells
was 6�104 cells/well in 6-multiwell plates (Corning Glass,
Corning, NY, USA) at 37�C under 5% CO2 in air. On day 21, the
culture medium was switched to �MEM containing 5% FBS plus
ITS, and the CO2 concentration was shifted to 3% for the facili-
tation of cellular hypertrophy and mineralization in culture.
RNA extraction and hybridization analysis
Total RNA was prepared from the cultures by the single-step meth-
od of Chomczynski and Sacchi (Chomczynski and Sacchi, 1987).
For northern hybridization, 20 �g of total RNA was denatured with
6% formaldehyde, separated by 1% agarose gel (Seakem GTG,
FMC Bioproducts, Rockland, ME, USA) electrophoresis, and
transferred onto Nytran membranes with Turboblotter (Schleicher
& Schuell, Dassel, Germany). Hybridization was performed over-
night at 42�C with an appropriate cDNA probe (106 cpm/ml) in a
solution containing 50% formamide, 6�SSPE (0.9 M NaCl, 20
mM NaH2PO4, 6 mM EDTA at pH 7.4), 0.2% BSA (Calbiochem-
Novabiochem Co., La Jolla, CA, USA), 0.2% Ficoll 400
(Pharmacia, Uppsala, Sweden), 0.2% polyvinylpyrolidone (Wako
Pure Chemical, Osaka, Japan), 0.1% SDS, and 200 �g/ml dena-
tured salmon sperm DNA. Hybridization probes were prepared by
the random-primer method with a BcaBEST labeling kit (Takara,
Shiga, Japan) using the appropriate cDNA fragments: a 1.4 kb
EcoRI fragment of pKT1180 as a probe for �1(II) collagen mRNA
(Kimura et al., 1987); a 0.65 kb Hind III fragment of pSAm10h as
a probe for �1(X) collagen mRNA (Apte et al., 1992); a 2.5-Kb
EcoRI fragment of rAP54 (Noda et al., 1987) as a probe for
alkaline phosphatase (ALP) mRNA. The following cDNA frag-
ments were amplified by reverse transcription-polymerase chain
reaction (RT-PCR) with a specific primer set as described below,
and used as a probe for northern hybridization: a 0.48 kb fragment
of GDF-5 cDNA; a 0.6 kb fragment of BMP-4 cDNA; a 0.42 kb
fragment of BMP-6 cDNA; a 0.34 kb fragment of ALK-3 cDNA; a
0.93 kb fragment of BMP type II receptor cDNA. After hybridiza-
tion, the filters were washed for 30 min at 55�C in 2�SSPE and
0.1% SDS, then washed for 30 min at 55�C in 0.1�SSPE, 0.1%
SDS, and then exposed to X-OMAT film (Eastman Kodak Co.,
Rochester, NY, USA) at �80�C with a Cronex lightening plus
intensifying screen (DuPont, Boston, MA, USA).
RT-PCR
The RT-PCR was performed as described previously (Shukunami,
et al., 1996). Briefly, first-strand cDNA was synthesized using
SuperScript II RNase H� reverse transcriptase (GIBCO BRL,
Gaithersburg, MD, USA) with total RNA extracted from ATDC5
cells. The PCR conditions were 94�C for 30 sec, 60�C for 1 min,
72�C for 1 min for 30 cycles, and final extension at 72�C for 5
min. The primers specific for mouse glyceraldehyde 3- phosphate
dehydrogenase (G3PDH) gene (Clontech, Palo Alto, CA, USA)
were used as a positive control for cDNA amplification. The fol-
lowing specific primers were used: 5'-CTCTTCAACCTCAG-
CAGCATCC-3' (sense primer) and 5'-TGGCAGTAGAAGGC-
CTGGTAGC-3' (antisense primer) for the detection of BMP-4
mRNA; 5'-TGACACGCAGACACATCTCC-3' (sense primer) and
5'-AGAAGGCACCTCCACAGAGC-3' (antisense primer) for
BMP-6 mRNA; 5'-GCTGAACTGCGGATCTTACG-3' (sense
primer) and 5'-AGCCTTGAGGTTCTTGCTGG-3' (antisense
primer) for GDF-5 mRNA; 5'-GTCGGACCTCTTCTTGCTGG-3'
(sense primer) and 5'-GACCGGATACTACGGAGATGG-3' (anti-
sense primer) for BMP-7 mRNA; 5'-GGCCTTGTCTGCT-
GCTAGTGG-3' (sense primer) and 5'-CTTCACGTCTTCCTG-
CACGC-3' (antisense primer) for type IB activin receptor (ActR-IB
or ALK-4) mRNA; 5'-GGAGACGATGTCACGAGGCC-3' (sense
primer) and 5'-CAGGTCCACATTGGTGACGG-3' (antisense
primer) for type IIB activin receptor (ActR-IIB) mRNA. For the
detection of mRNAs for the Smad family members, the following
primers were used: 5'- CCGTCCAACAATAAGAACCGC-3'
(sense primer) and 5'-ATCGTGGCTCCTTCGTCAGG-3' (anti-
sense primer) for the detection of Smad1 mRNA; 5'-ACCAG-
GTCTCTTGATGGCCG-3' (sense primer) and 5'-TGACGCATG-
GAAGGTCTCTCC-3' (antisense primer) for Madr2 mRNA; 5'-
AGACATTCCACGCCTCACAGC-3' (sense primer) and 5'-CA-
CACTGGAACAGCGGATGCT-3' (antisense primer) for Smad3
mRNA; 5'-CACCAGCACCACCAACTTCC-3' (sense primer) and
5'-CAACCTCGCTCTCTCAATCGC-3' (antisense primer) for
DPC4 mRNA; 5'-GCCGTGAAGCGATTGTTGG-3' (sense prim-
er) and 5'-GCGGTTCATTGTGGCTCAGG-3' (antisense primer)
for Smad5 mRNA; 5'-ACAAGCCACTGGATCTGTCCG-3' (sense
primer) and 5'-GGAGGTGATGAACTGTCGCG-3' (antisense
primer) for Smad6 mRNA; 5'-ATGCTGTACCTTCCTCCGCG-3'
Coordinated Bmp Expression during Chondrogenesis
197
(sense primer) and 5'-CACAGCCGATCTTGCTCCG-3' (antisense
primer) for Smad7 mRNA, and 5'-CAAGTTCCGAAGTGC-
CTCGCT-3' (sense primer) and 5'-GATCTTGCAGACAGTGGC-
CG-3' (antisense primer) for Smad8 mRNA. Aliquots (8 �l) of
each PCR reaction products were resolved on 3% NuSieve 3:1
agarose gels (FMC Bioproducts) alongside the markers. The
amplified products were stained with ethidium bromide. The
amplified PCR products were subcloned pCRII (Invitrogen, San
Diego, CA, USA), and the nucleotide sequences of all cDNA frag-
ments were verified by an ALP Express Sequencer (Amersham-
Pharmacia, Uppsala, Sweden). No products were observed when
the reverse transcriptase was omitted during the synthesis of
cDNA (data not shown).
Alizarin red staining
ATDC5 cells were cultured in DME/F12 medium containing 5%
FBS and ITS for 21 days. Then the culture medium was switched
to �MEM containing 5% FBS and ITS, and the cultures were con-
tinued for an additional 21 days. After a total of 42 days, the cul-
tures were incubated for an additional 8 days with or without 1,000
ng/ml of BMP-7 in �MEM containing 5% FBS and ITS. Cells
were rinsed three times with PBS and fixed with 95% methanol for
30 min. They were stained with 1% alizarin red S (Wako Pure
Chemicals) at pH 6.4 for 5 min and washed with distilled water, as
described (Shukunami, et al., 1997).
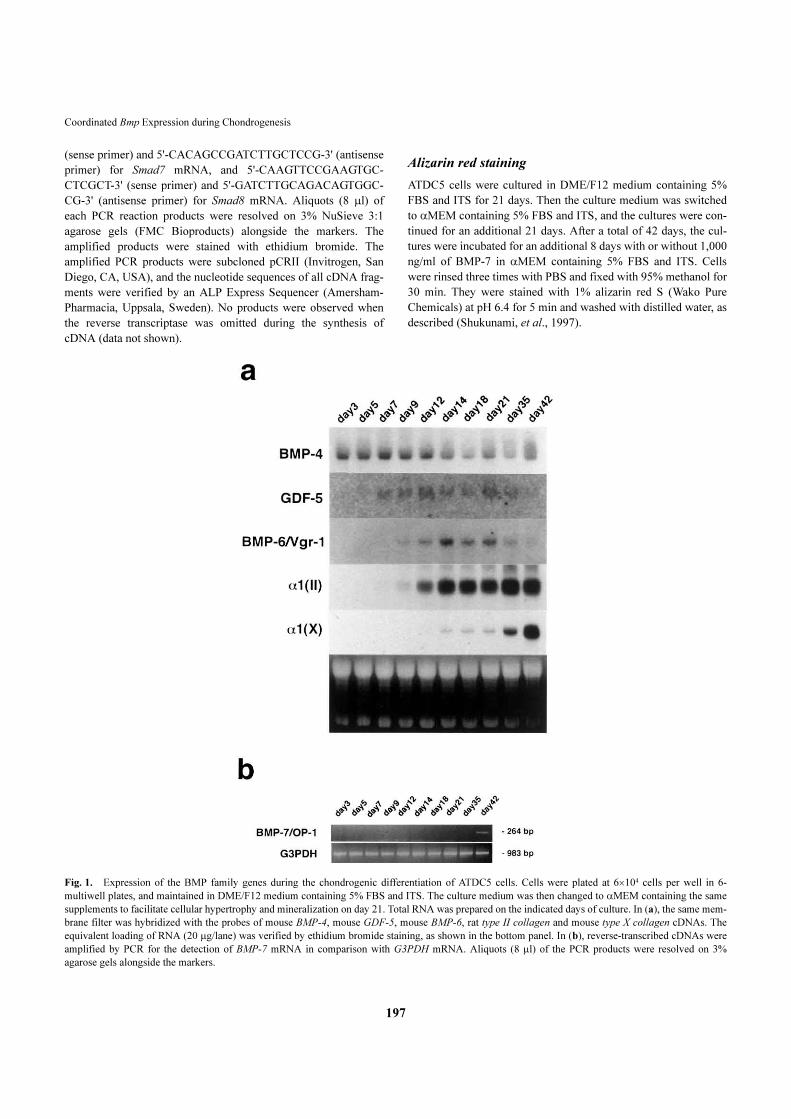
Fig. 1. Expression of the BMP family genes during the chondrogenic differentiation of ATDC5 cells. Cells were plated at 6�104 cells per well in 6-
multiwell plates, and maintained in DME/F12 medium containing 5% FBS and ITS. The culture medium was then changed to �MEM containing the same
supplements to facilitate cellular hypertrophy and mineralization on day 21. Total RNA was prepared on the indicated days of culture. In (a), the same mem-
brane filter was hybridized with the probes of mouse BMP-4, mouse GDF-5, mouse BMP-6, rat type II collagen and mouse type X collagen cDNAs. The
equivalent loading of RNA (20 �g/lane) was verified by ethidium bromide staining, as shown in the bottom panel. In (b), reverse-transcribed cDNAs were
amplified by PCR for the detection of BMP-7 mRNA in comparison with G3PDH mRNA. Aliquots (8 �l) of the PCR products were resolved on 3%
agarose gels alongside the markers.

H. Akiyama et al.
198
Results
Expression of BMP-4, BMP-6, BMP-7 and GDF-5 mRNAs during chondrogenic differentiation
The transcripts for the BMP family genes are expressed spa-
tially and temporally at the sites of skeletogenesis such as
limb buds and vertebrae during embryonic development
(Francis West, et al., 1995; Lyons, et al., 1990). ATDC5
cells express type I collagen in the undifferentiated stage.
However, the cells undergo differentiation to become type II
collagen-expressing chondrocytes in the presence of insulin
(Shukunami, et al., 1996). As shown in Fig. 1a, ATDC5
cells expressed transcripts for BMP-4 in the undifferentiated
(day 3) and condensation stages (day 5), prior to conversion
into chondrocytes. When the growth of differentiated cells
ceased by day 21 of culture, cells in cartilage nodules then
undergo late-phase differentiation to become hypertrophic
and calcifying chondrocytes (Shukunami, et al., 1997). The
BMP-4 mRNA level declined as the cartilage nodules
stopped growing after two weeks. The GDF-5 transcripts
were first detected in a cellular condensation stage on day 5
and continuously expressed in these cells, and declined as
differentiated cells began to calcify.
When the regions of cellular condensation are formed in
ATDC5 cell cultures from day 3 to day 6, type II collagen-
expressing chondrocytes are generated in the condensing
regions (Shukunami, et al., 1996). Chondrocytes thus
formed gradually proliferate to expand cartilage nodules in
the culture for another 10 days, as previously reported
(Shukunami, et al., 1996). As the number of chondrocytes
increased during this time period, the type II collagen
mRNA level increased and became maximal on day
14 or later. Type X collagen-expressing hypertrophic
chondrocytes then appeared in cartilage nodules, and mark-
edly increased in number from day 21 when the growth of
chondrocytes in the nodules completely ceased.
Interestingly, the BMP-6 and BMP-7 genes exhibited a
distinct temporal pattern of expression along with the
phenotypic transitions of ATDC5 cells. Transcripts for the
BMP-6 gene were absent until mature chondrocytes
appeared in the culture. The BMP-6 mRNA level increased
in parallel with the induction of type II collagen mRNA lev-
el in the culture, and peaked at around day 14 (Fig. 1a). The
level of BMP-6 mRNA was kept high during the maturation
of chondrocytes (from day 14 to day 21), and then declined
until the cells became calcified. In contrast, transcripts for
the BMP-7 gene were not detected until the cells became
calcified on day 42 or later (Fig. 1b). However, the level of
BMP-7 mRNA was so low as to be detected by northern
blotting.
Expression of BMP receptor mRNA during differenti-ation
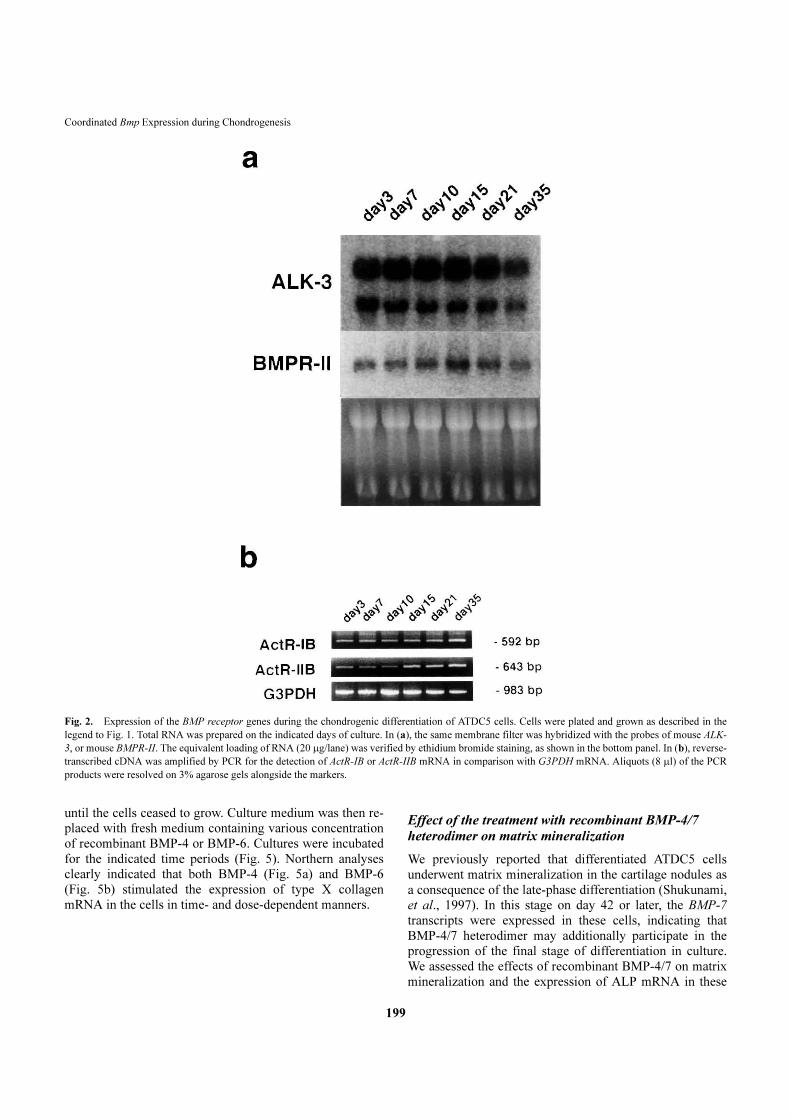
As shown in Fig. 2a, ATDC5 cells expressed the transcripts
for the ALK-3 gene, encoding the BMP type IA receptor.
The level of these transcripts did not change significantly
during chondrogenic differentiation of ATDC5 cells. Simi-
larly, transcripts for the BMP type II receptor gene were
detected in all stages of differentiation of ATDC5 cells,
although the level of the transcripts upregulated during
maturation of differentiated chondrocytes from day 10 to
day 21 of culture. However, mRNA for these receptors were
detected in all stages. The transcripts for the BMP type IB
receptor (ALK-6) gene were not detected in the ATDC5
cells (data not shown). Transcripts for the type IB and type
IIB receptors for activin (ActR-IB and ActR-IIB) were also
found in all stages of differentiation of ATDC5 cells (Fig.
2b). ATDC5 cells similarly expressed transcripts for the
type I receptor for activin (ActR-I or ALK-2) (data not
shown).
The Smad family proteins play an important role in the
transduction of BMP signals from the cell-surface receptors
(Heldin, et al., 1996). As shown in Fig. 3, ATDC5 cells
clearly expressed transcripts for the Smad family genes in
all stages of differentiation.
Effect of the treatment with recombinant BMP-4 on the early-phase differentiation in ATDC5 cells
BMP-4 is expressed in undifferentiated ATDC5 cells,
indicating that BMP-4 participates in the early-phase differ-
entiation of these cells. When ATDC5 cells were cultured in
DME/F12 medium containing 5% FBS and ITS for 11 days,
cartilage nodules were apparently formed (Fig. 4a), as
reported previously (Shukunami, et al., 1996). Recombinant
BMP-4 (1,000 ng/ml) was added to the confluent monolayer
culture in the medium containing 5% FBS and ITS on day 5,
and the culture was maintained for additional 6 days. By
skipping out the condensation stage, almost all cells in cul-
ture underwent differentiation and formed a nearly continu-
ous sheet of chondrocytes (Fig. 4a) accompanying alcian
blue-positive matrix. As shown in Fig. 4b, recombinant
BMP-4 apparently induced the expression of type II
collagen gene within 24 hours in a dose dependent manner
in the confluent monolayer culture of ATDC5 cells.
Effect of the treatment with recombinant BMP-4 and BMP-6 on the late-phase differentiation
When type II collagen-expressing differentiated ATDC5
cells cease to grow in cartilage nodules, type X collagen-
expressing cells begin to appear in the nodules (Shukunami,
et al., 1997). We assessed by northern analyses the effects of
recombinant BMP-4 and BMP-6 on the late-phase differen-
tiation of ATDC5 cells. The cells were first cultured in
DME/F12 medium containing 5% FBS and ITS for 21 days
Coordinated Bmp Expression during Chondrogenesis
199
until the cells ceased to grow. Culture medium was then re-
placed with fresh medium containing various concentration
of recombinant BMP-4 or BMP-6. Cultures were incubated
for the indicated time periods (Fig. 5). Northern analyses
clearly indicated that both BMP-4 (Fig. 5a) and BMP-6
(Fig. 5b) stimulated the expression of type X collagen
mRNA in the cells in time- and dose-dependent manners.
Effect of the treatment with recombinant BMP-4/7
heterodimer on matrix mineralization
We previously reported that differentiated ATDC5 cells
underwent matrix mineralization in the cartilage nodules as
a consequence of the late-phase differentiation (Shukunami,
et al., 1997). In this stage on day 42 or later, the BMP-7
transcripts were expressed in these cells, indicating that
BMP-4/7 heterodimer may additionally participate in the
progression of the final stage of differentiation in culture.
We assessed the effects of recombinant BMP-4/7 on matrix
mineralization and the expression of ALP mRNA in these
Fig. 2.�Expression of the BMP receptor genes during the chondrogenic differentiation of ATDC5 cells. Cells were plated and grown as described in the
legend to Fig. 1. Total RNA was prepared on the indicated days of culture. In (a), the same membrane filter was hybridized with the probes of mouse ALK-
3, or mouse BMPR-II. The equivalent loading of RNA (20 �g/lane) was verified by ethidium bromide staining, as shown in the bottom panel. In (b), reverse-
transcribed cDNA was amplified by PCR for the detection of ActR-IB or ActR-IIB mRNA in comparison with G3PDH mRNA. Aliquots (8 �l) of the PCR
products were resolved on 3% agarose gels alongside the markers.

H. Akiyama et al.
200
cells. Differentiated cultures of ATDC5 cells were incu-
bated with BMP-4/7 (1,000 ng/ml) from day 42 to day 50.
As shown by alizarin red staining (Fig. 6a), mineralization
was clearly promoted by the treatment. The induction of a
high level of ALP activity in chondrocytes is closely associ-
ated with matrix mineralization (Shukunami, et al., 1997).
When the ATDC5 cell culture reached in the initial calcify-
ing stage on day 42, the culture was further incubated for
the indicated time periods in the presence of various
concentrations of recombinant BMP-4/7. As indicated by
northern blot analysis, the treatment with BMP-4/7 resulted
in an increase of the level of ALP mRNA in a dose-depend-
ent manner within 12 hours (Fig. 6b).
Discussion
ATDC5 cells retain the properties of chondroprogenitor
cells well (Shukunami, et al., 1996). Taking on a fibro-
blastic morphology, undifferentiated cells express the type
I collagen gene transcripts, and they actively proliferate
with a short doubling time of 16 hours. In the presence of
insulin, regions of cellular condensation appear transiently
in the culture, from which proliferating chondrocytes (with
a longer doubling time of 48 hours) are generated to form
cartilage nodules (Shukunami, et al., 1996). When the
formation of cartilage nodules is completed, the cells then
become hypertrophic and calcified as the late-phase differ-
entiation progresses (Shukunami, et al., 1997). These order-
ly transitions of cellular phenotype were positively and
negatively modulated by the exogenous growth and differ-
entiation factors added to the culture (Shukunami, et al.,
1998). Recombinant BMP-2 positively regulated the pheno-
typic transitions during early-phase and late-phase differen-
tiation. Parathyroid hormone (PTH) negatively regulated the
progression of differentiation (Shukunami, et al., 1997;
Shukunami, et al., 1996). These results strongly suggested
the presence of a complex network of endogenous growth/
differentiation signaling molecules underlying the orderly
transition of cellular phenotype in the ATDC5 cultures.
In the developing cartilaginous bone precursors, BMP-4
mRNA is expressed first in the perichondral mesenchymal
cells and late in perichondrium from which undifferentiated
mesenchymal cells are recruited (Wozney, 1993). The BMP-
4 transcripts were constitutively expressed in ATDC5 cells
irrespective of their differentiation stages. However, the
mRNA level slightly decreased prior to the induction of the
late-phase differentiation (day 21 or later). In contrast, the
GDF-5 mRNA were expressed after the onset of cellular
condensation. As shown in Fig. 2, ATDC5 cells also
expressed the BMP signaling counterparts, BMP type IA
receptor (ALK-3) and BMP type II receptor, in all stages of
differentiation. Intracellular signal transducing components
Smads were also expressed in the cells (Fig. 3). Thus,
ATDC5 cells retained their responsiveness to BMP-4
signaling.
Zou and coworkers demonstrated the distinct functions of
BMP receptors type IA and type IB (Zou, et al., 1997).
BMP type IB receptor (ALK-6) expression preceded the
early chondrogenic differentiation. Signaling from the
receptor is necessary for the early steps of mesenchymal
condensation and cell death. In contrast, BMP type IA
receptor (ALK-3) was essential for proper progression of
the cartilage differentiation program. In agreement with
these observations, ATDC5 cells properly kept track of the
early and late-phase differentiation program despite the lack
of BMP type IB receptor expression. ATDC5 cells are com-
mitted chondroprogenitor cells. In fact, when the confluent
cultures of undifferentiated ATDC5 cells were treated with
recombinant BMP-4, almost all of the cells were induced to
become chondrocyte-like without the formation of cellular
condensation (Fig. 4a). GDF-5 has also been shown to stim-
ulate chondrogenic differentiation (Tsumaki, et al., 1999).
ATDC5 cells expressed GDF-5 mRNA as well as mRNAs
for its type II receptors, BMP type II receptor and ActR-II
(Nishitoh et al., 1996). However, its type I receptor (ALK-6)
mRNA was not detected in these cells. Thus, it remained to
be elucidated the specific role of GDF-5 in differentiation of
ATDC5 cells.
Interestingly, BMP-6 and BMP-7 genes exhibited the
distinct and transient patterns of expression along with the
late-phase differentiation of ATDC5 cells. The expression
Fig. 3. Expression of the genes for the Smad family members during the
chondrogenic differentiation of ATDC5 cells. Cells were plated and grown
as described in the legend to Fig. 1. Total RNA was prepared on the indi-
cated days of culture. Reverse-transcribed cDNAs were amplified by PCR
for the detection of Smad1, Madr2, Smad3, DPC4, Smad5, Smad6, Smad7,
or Smad8 mRNA in comparison with G3PDH mRNA. Aliquots (8 �l) of
the PCR products were resolved on 3% agarose gels alongside the markers.

Coordinated Bmp Expression during Chondrogenesis
201
of BMP-6 mRNA increased in parallel with the increase of
type II collagen mRNA in the ATDC5 cells, and then
declined (Fig. 1a), suggesting that the BMP-6 expression
was induced in association with nodule formation. When
the cultures of differentiated ATDC5 cells were treated with
recombinant BMP-6, it facilitated the phenotypic conver-
sion of proliferating cells to hypertrophic cells, as evidenced
by the upregulation of type X collagen expression (Fig. 5b).
The stimulatory effects of recombinant BMP-6 on the late-
phase differentiation of ATDC5 cells is similar to those of
BMP-4 (Fig. 5a). Differentiated ATDC5 cells express ALK-
3, but do not express ALK-6. Thus, the stimulatory effects of
BMP-6 on differentiation of ATDC5 cells may possibly be
mediated by ALK-3 in ATDC5 cells.
We previously showed that the ATDC5 cultures
underwent matrix mineralization in the differentiated carti-
lage nodules without �-glycerophosphate supplement
(Shukunami, et al., 1997). BMP-7 mRNA was expressed in
a matrix mineralization stage in these cells, and recombi-
nant BMP-4/7 actually promoted a marked upregulation of
ALP mRNA and matrix mineralization in these cells (Fig.
6), suggesting that BMP-7 plays a role in the progression of
Fig. 4. Effects of recombinant BMP-4 on the early-phase chondrogenic differentiation of ATDC5 cells. Cells were plated at 6�104 cells per well in a
6-multiwell plate and grown to confluency on day 5 in DME/F12 medium containing 5% FBS and ITS. Cells were then incubated in the absence or the pres-
ence of recombinant BMP-4 (1,000 �g/ml) for another 6 days. Cells were then assessed for cartilaginous nodule formation under phase-contrast microscopy
(a) (Bar denotes 100 µm). (b) After 5 days, medium was replaced with the fresh medium containing various doses of BMP-4. For the time-dependence, cells
were treated with 1,000 ng/ml of BMP-4 for the various time periods indicated. For the dose-dependence, cells were treated with 100 or 1,000 ng/ml of
BMP-4 for a total of 48 hours. Total RNA was prepared and subjected for northern analysis of type II collagen mRNA. The equivalent loading of RNA (20
�g/lane) was verified by ethidium bromide staining, as shown in the bottom panel. Three independent experiments were performed and gave similar results.

H. Akiyama et al.
202
mineralization in ATDC5 cells.
Chondrogenic differentiation includes a series of cellular
events. which enable cells to undergo orderly transitions of
the cartilage phenotype, as characterized by the sequential
expression of unique cartilage matrix. The present results
indicated that, in addition to the basal expression of BMP-4,
the differentiation stage-specific expression of BMP-6 and
BMP-7 contributed to the progressive multistep differentia-
tion after induction of cellular condensation by insulin in
ATDC5 cells. Thus, the mouse clonal cell line ATDC5 is a
useful in vitro model for molecular analysis of the multistep
chondrogenic differentiation during endochondral bone for-
mation.
Acknowledgements. We thank Dr. J. M. Wozney (Genetics Institute Inc.)
for his kind provision of human recombinant BMP-6. This work was partly
Fig. 5. Effects of recombinant BMP-4 or BMP-6 on the late-phase chondrogenic differentiation of ATDC5 cells. Cells were plated at 6�104 cells per well
in a 6-multiwell plate and cultured for 21 days in DME/F12 medium containing 5 %FBS and ITS. After 21 days, medium was replaced with the fresh medi-
um containing various doses of recombinant BMP-4 (a) or BMP-6 (b). For the time-dependence, cells were treated with 1,000 ng/ml of BMP-4 or BMP-6
for the various time periods indicated. For the dose-dependence, cells were treated with 100 or 1,000 ng/ml of BMP-4 or BMP-6 for a total of 48 hours. Total
RNA was prepared and subjected for northern analysis of the expression of type X collagen mRNA. The equivalent loading of RNA (20 �g/lane) was veri-
fied by ethidium bromide staining, as shown in the bottom panel. Three independent experiments were performed and gave similar results.

Coordinated Bmp Expression during Chondrogenesis
203
supported by The Naito Foundation and Terumo Life Science Foundation
to YH.
References
Apte, S.S., Seldin, M.F., Hayashi, M., and Olsen, B.R. 1992. Cloning of
the human and mouse type X collagen genes and mapping of the mouse
type X collagen gene to chromosome 10. Eur. J. Biochem., 206: 217–
224.
Carey, D.E., and Liu, X. 1995. Expression of bone morphogenetic protein-
6 messenger RNA in bovine growth plate chondrocytes of different size.
J. Bone Miner. Res., 10: 401–405.
Chomczynski, P., and Sacchi, N. 1987. Single-step method of RNA isola-
tion by acid guanidinium thiocyanate-phenol-chloroform extraction.
Anal. Biochem., 162: 156–159.
Francis West, P.H., Robertson, K.E., Ede, D.A., Rodriguez, C., Izpisua
Belmonte, J.C., Houston, B., Burt, D.W., Gribbin, C., Brickell, P.M., and
Tickle, C. 1995. Expression of genes encoding bone morphogenetic
proteins and sonic hedgehog in talpid (ta3) limb buds: their relationships
in the signalling cascade involved in limb patterning. Dev. Dyn., 203:
187–197.
Heldin, C.H., Miyazono, K., and ten Dijke, P. 1997. TGF-beta signalling
from cell membrane to nucleus through SMAD proteins. Nature, 390:
465–471.
Kimura, T., Mattei, M.-G., Stevens, J.W., Goldring, M.B., Ninomiya, Y.,
and Olsen, B.R. 1989. Molecular cloning of rat and human type IX
collagen cDNA and localization of the �1(IX) gene on the human
chromosome 6. Eur. J. Biochem., 179: 71–78.
Lyons, K.M., Pelton, R.W., and Hogan, B.M.L. 1990. Organogenesis and
Fig. 6. Effects of recombinant BMP-4/7 on the matrix mineralization of ATDC5 cells. Cells were plated at 6�104 cells per well in a 6-multiwell plate and
cultured for 42 days as described in the methods section. (a) Cells were then incubated in the absence or the presence of recombinant BMP-4/7 (1,000 ng/
ml) for an additional 8 days and stained with 1% alizarin red S. (b) After 42 days, medium was replaced with the fresh medium containing various doses of
BMP-4/7. For the time-dependence, cells were treated with 1,000 ng/ml of BMP-4/7 for the various time periods indicated. For the dose-dependence, cells
were treated with 100 or 1,000 ng/ml of BMP-4/7 for a total of 48 hours. Total RNA was prepared and subjected for northern analysis of ALP mRNA. The
equivalent loading of RNA (20 �g/lane) was verified by ethidium bromide staining, as shown in the bottom panel. Three independent experiments were per-
formed and gave similar results.

H. Akiyama et al.
204
pattern formation in the mouse: RNA distribution patterns suggest a role
for bone morphogenetic protein-2A (BMP-2A). Development, 109: 833–
844.
Nishitoh, H., Ichijo, H., Kimura, M., Matsumoto, T., Makishima, F.,
Yamaguchi, A., Yamashita, H., Enomoto, S., and Miyazono, K. 1996.
Identification of type I and type II serine/threonine kinase receptors for
growth/differentiation factor-5. J. Biol. Chem., 271: 21345–21352.
Noda, M., Yoon, K., Thiede, M., Buenaga, R., Weiss, M., Henthorn, P.,
Harris, H., and Rodan, G.A. 1987. cDNA cloning of alkaline
phosphatase from rat osteosarcoma (ROS 17/2.8) cells. J. Bone Miner.
Res., 2: 161–164.
Rosen, V., and Thies, S. 1992. The BMP proteins in bone formation and
repair. Trends in Genetics, 8: 97–102.
Shukunami, C., Ishizeki, K., Atsumi, T., Ohta, Y., Suzuki, F., and Hiraki, Y.
1997. Cellular hypertrophy and calcification of embryonal carcinoma-
derived chondrogenic cell line ATDC5 in vitro. J. Bone Min. Res., 12:
1174–1188.
Shukunami, C., Ohta, Y., Sakuda, M., and Hiraki, Y. 1998. Sequential
progression of the differentiation program by bone morphogenetic
protein-2 in chondrogenic cell line ATDC5. Exp. Cell Res., 241: 1–11.
Shukunami, C., Shigeno, C., Atsumi, T., Ishizeki, K., Suzuki, F., and
Hiraki, Y. 1996. Chondrogenic differentiation of clonal mouse embry-
onic cell line ATDC5 in vitro: differentiation-dependent gene expression
of parathyroid hormone (PTH)/PTH-related peptide receptor. J. Cell
Biol., 133: 457–468.
Storm, E.E., Huynh, T.V., Copeland, N.G., Jenkins, N.A., Kingsley, D.M.,
and Lee, S.-J. 1994. Limb alterations in brachypodism mice due to
mutations in a new member of the TGFb-superfamily. Nature, 368: 639–
643.
Tsumaki, N., Tanaka, K., Arikawa-Hirasawa, E., Nakase, T., Kimura, T.,
Thomas, J.T., Ochi, T., Luyten, F.P., and Yamada, Y. 1999. Role of
CDMP-1 in skeletal morphogenesis: promotion of mesenchymal cell
recruitment and chondrocyte differentiation. J. Cell Biol., 144: 161–173.
Vortkamp, A., Lee, K., Lanske, B., Segre, G.V., Kronenberg, H.M., and
Tabin, C.J. 1996. Regulation of Rate of Cartilage Differentiation by
Indian Hedgehog and PTH-Related Protein. Science, 273: 613–622.
Vukicevic, S., Latin, V., Chen, P., Batorsky, R., Reddi, A.H., and Sampath,
T.K. 1994. Localization of osteogenic protein-1 (bone morphogenetic
protein-7) during human embryonic development: high affinity binding
to basement membranes. Biochem. Biophys. Res. Commun., 198: 693–
700.
Wozney, J.M. 1993. Bone morphogenetic proteins and their gene expres-
sion. In Cellular and Molecular Biology of Bone (M. Noda, eds.). Aca-
demic Press, San Diego, pp.131–167.
Zou, H., Wieser, R., Joan, M., and Niswander, L. 1997. Distinct roles of
type I bone morphogenetic protein receptors in the formation and differ-
entiation of cartilage. Genes Dev., 11: 2191–2203.
(Received for publication, June 20, 2000
and accepted, June 30, 2000)





























