Embed Size (px)
Citation preview

DIMERIZATION CONTROLS THE ACTIVITY OF FUNGAL ELICITORS THAT TRIGGER SYSTEMIC RESISTANCE IN PLANTS†
Walter A. Vargas1, Slavica Djonović1 *, Serenella A. Sukno2, and Charles M. Kenerley1
1Department of Plant Pathology and Microbiology, Texas A&M University, College Station TX 77843 2Centro Hispano-Luso de Investigaciones Agrarias (CIALE), Departamento de Microbiología y Genética,
Universidad de Salamanca, 37007 Salamanca, Spain. Running title: Post-translational modifications of elicitors that activate disease resistance
To whom correspondence should be addressed: Charles M. Kenerley, 413C LF Peterson, Texas A&M University, College Station, TX 77843. Tel 979-8457-544 Fax: 979-845-6483; Email: [email protected]
The soilborne fungus Trichoderma virens secretes a small protein (Sm1) that induces local and systemic defenses in plants. This protein belongs to the ceratoplatanin protein family and is mainly present as a monomer in culture filtrates. However, Hypocrea atroviride (the telomorph form of T. atroviride) secretes a Sm1 homologous protein, Epl1, with high levels of dimerization. Nonetheless, the molecular mechanisms involved in recognition and the signaling pathways involved in the induction of systemic resistance in plants are still unclear. In this report, we demonstrate that Sm1 and Epl1 are mainly produced as monomer and a dimer, respectively, in the presence of maize seedlings. The results presented show that the ability to induce plant defenses reside only the monomeric form of both Sm1 and Epl1 and we demonstrate for the first time that the monomeric form of Epl1, likewise Sm1, induces defenses in maize plants. Biochemical analyses indicate that monomeric Sm1 is produced as a glycoprotein, but the glycosyl moiety is missing from its dimeric form and Epl1 is produced as a non-glycosylated protein. Moreover, for Sm1 homologues in various fungal strains, there is a negative correlation between the presence of the glycosylation site and their ability to aggregate. We propose a subdivision in the ceratoplatanin protein family according to the presence of the glycosylation site and the ability of the proteins to aggregate. The data presented suggest that elicitor’s aggregation may control the Trichoderma-plant molecular dialogue and block the activation of induced systemic resistance in plants.
Rhizosphere colonization by certain bacterial strains results in a state of heightened resistance in plants, locally and systemically, to subsequent pathogen attack (1, 2). During the colonization process, a complex molecular dialogue is established between plants and the microorganisms in which signaling molecules (elicitors) play an essential role (3-6). Various molecules associated with the basic metabolism of microbes, such as cell wall glucans, chitin oligomers, and glycopeptides, have been described elicitor activity (7-9). In addition, certain structural proteins and enzymes encoded in the invader genome (such as Sm1, chitinase and flagellin) have also been shown to act as signal molecules (10-15).
One important outcome of some plant-microbe interactions is the induction of plant defenses mediated by the activation of Induced Systemic Resistance (ISR). ISR confers systemic protection in plants by mechanisms different from those of the well-known and well-studied systemic acquired resistance [reviewed by Vallard and Goodman (2)]. However, the mechanisms that underlie the molecular crosstalk between plants and the microbes that initiate ISR are not fully understood. This type of induced resistance has primarily been linked to the colonization of plant roots by plant growth promoting rhizobacteria (PGPR) and recently to some species of the filamentous fungus Trichoderma (13, 14, 16-18). The characterization of two ISR elicitors secreted by T. virens Gv29-8 was recently described. Peptaibols (peptides with antimicrobial activity) produced by T. virens were demonstrated to have ISR elicitor effects and they systemically induce defenses in cucumber leaves (18). The second ISR elicitor produced by T. virens is the extracellular
1
http://www.jbc.org/cgi/doi/10.1074/jbc.M802724200The latest version is at JBC Papers in Press. Published on May 15, 2008 as Manuscript M802724200
Copyright 2008 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

small protein Sm1, whose gene expression was demonstrated to be up-regulated in the presence of cotton plants. This elicitor is able to locally and systemically induce defense-responses in plants (13). Further in vivo studies, using reverse genetic analyses, demonstrated that expression of Sm1 is essential for triggering ISR in maize plants and providing protection against the foliar pathogen Colletotrichum graminicola (14). In maize, the metabolic pathways that lead to the establishment of Sm1-mediated ISR involve the signaling networks associated with salicylic acid, green leafy volatiles and jasmonic acid metabolism, and seem to be independent of PR proteins (14). However, the molecular mechanisms relevant to Sm1 recognition and signal transduction have not been described yet.
Sm1 was the first and the only (to our knowledge) proteinaceous elicitor, with no enzymatic activity, involved in ISR responses described to date. Sm1 is a secreted hydrophobin-like protein, with a predicted molecular mass of 12,545.8 Da. According to structural and sequence analyses, Sm1 belongs to the cerato-platanin (CP) protein family (13, 19), which consists mainly of proteins produced by plant and human fungal pathogens, and associated with toxicity and infection processes (20-24). Paradoxically, characterization of Sm1 demonstrated a lack of phytotoxicity towards various plants, including cotton, rice, tobacco and peanut (13). Sm1 was previously purified from T. virens Gv29-8 culture filtrates as a monomeric protein (13, 14). However, in some growth conditions, a faint band corresponding to the SDS-resistant dimeric form was detected in western blots (CMK, unpublished). The existence of Sm1 monomer and dimer forms could be important for its biological activity. For example, there are reports of key regulators of defense responses whose activity in vivo is regulated by redox changes (25, 26). Recently, Epl1, the Sm1 homologue produced by Hypocrea atroviride was also detected in two forms (monomer and dimer) in fungal culture filtrates (27). To gain novel insights into ISR and the molecular mechanisms underlying the elicitor recognition and response induction, we have investigated the relevance of the oligomerization states of Sm1 and Epl1 for ISR. In this report we demonstrate that Epl1 indeed elicits the activation of plant defense
mechanism, but only the monomeric forms of Sm1 and Epl1 are able to induce such responses, while both dimers fail to protect maize plants against a leaf pathogen. Further biochemical characterization of Sm1 and Epl1 demonstrates that the monomeric form of Sm1 is produced as a glycoprotein. However, glycosylation was not present in either dimer forms of Sm1 or Epl1. The occurrence of Sm1/Epl1 homologues was analyzed in different fungal strains by sequence analysis and immunodetection of the polypeptides. The results of this work led us to conclude that the glycosylation state may be controlling the protein oligomerization status and altering the recognition of the elicitor and the induction of defenses in plants. We discuss the importance of the glycosylation state of Sm1 in the molecular dialogue during plant-Trichoderma associations and in the stimulation of ISR in plants. A novel subdivision on the CP protein superfamily, based on to the protein’s ability to aggregate and the presence of glycosylation sites, is also proposed.
EXPERIMENTAL PROCEDURES
Fungal and plant materials- The Trichoderma species used in this study were, T. virens Gv29-8, T. virens G6, T. virens G9, T. virens Gv29-8-ΔSm1 (14), T. atroviride IMI206040, T. reesei 6 and T. viride 21. Also for this research we used the filamentous fungi Magnaporthe grisea 70-15, Aspergillus fumigatus FGSC1152 and Neurospora crassa 74A. A GFP-tagged Colletotrichum graminicola M1.001-BH isolate (28) was used as a foliar pathogen for disease development studies in maize. The strains were routinely maintained on potato dextrose agar (PDA, Difco). Maize (Zea mays inbred line B73) seedlings used in this study were grown in a hydroponic system (13) or planted in plastic containers (3.81 × 20.9 cm) containing a soil-less mix (Metromix 366) and incubated in a growth chamber at 25 °C, with a 14 h photoperiod and 60% humidity.
Analysis of fungal secreted proteins from the hydroponic growth system- A hydroponic system (13) was used to evaluate the plant defense response of maize seedlings when inoculated with T. virens or T. atroviride strains. Maize seeds were surface sterilized using 10% hydrogen peroxide as previously described (13). The mycelial inoculum of the Trichoderma strains was aseptically added into the hydroponic systems
2
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

containing 300 mL of fresh Murashige and Skoog (MS) medium and 4-day-old maize seedlings. Two days later, the medium from each treatment was collected and successively filtered through a 10 μm NITEX nylon cloth and a 0.45 μm filter. Culture filtrates (CFs) were then treated with a protease inhibitor cocktail (0.05% v/v, Sigma, St. Louis, MO), allowed to incubate at room temperature for 20 min, and then 300 mL of each treatment were concentrated by using 10 kDa cutoff Millipore filter devices (Bedford, MA). Protein extracts were used to determine enzymatic activity and also used for SDS-PAGE or western blotting analyses.
Enzymatic activity assays and protein quantification- The activity of β-1,3-glucanase, β-1,6-glucanase, endochitinase and proteinase secreted by T. virens and T. atroviride, in various culture conditions was compared. Enzymatic activities of both strains when grown in liquid Vogel’s minimal medium (29) (VM), VM supplemented with 1.5% sucrose (VMS), or cultured in MS media in the presence or absence (control) of maize seedlings, were assayed. The activity of β-1,3, β-1,6-glucanase was determined in the presence of pustulan and laminarin, respectively, by detecting the reducing sugars released after incubation (30). Protease and endochitinase activities were determined in the presence of Suc-Ala-Ala-Pro-Phe-pNA (Sigma) and 4-methylumbelliferyl-β-D-N,N’,N’’-triacetylchitotriose (Sigma), respectively, as recently described (30). Protein concentration was determined in a micro-plate assay using the Protein Reagent (Bio-Rad) according to the manufacturer’s instructions or by detecting absorbance at 280 nm, using BSA as a standard. The specific activities were calculated as Units/mg protein.
SDS-PAGE and western blot assays- Polypeptides were separated on SDS-PAGE (15% polyacrylamide) (31) and stained with Coomassie blue or blotted onto a nitrocellulose membrane (HyBond C®, Amersham) for immunoassays (32). Protein blots were probed with specific antibodies raised against Sm1 (13).
Purification of Sm1 and Epl1 isoforms- For Sm1 and Epl1 purification, T. virens Gv29-8 and T. atroviride IMI206040 were grown for 7-days in VMS medium, and CFs were collected by filtration through a 10 μm NITEX nylon cloth. Proteins were precipitated with 80% ammonium
sulfate, collected by centrifugation, dissolved in 20 mM NH4HCO3 and dialyzed against the same solution. Total protein was loaded onto High Q support columns (0.5X20 cm) [Bio-Rad] pre-equilibrated with 20 Tris-NaOH (pH 7.5) (anion exchange chromatography, AEC). Proteins were eluted with a 0–0.5 M NaCl linear gradient in the equilibration buffer. The elution of Sm1 and Epl1 was followed by SDS-PAGE and protein immunodetection. The fractions containing the protein of interest were pooled, dialyzed against 20 mM NH4HCO3, and vacuum-dried. The dried protein was dissolved in 50 mM Tris-NaOH (pH 7.5), 150 mM NaCl and further purified by gel filtration (GFC) through Bio-Gel P30 columns (1.0X100 cm) [BioRad] pre-equilibrated with the same solution. The elution of Sm1 and Epl1 was followed by SDS-PAGE and western blotting. While the monomeric form of Epl1 was not detected in the column eluant, the monomeric form of Sm1 eluted pure after this purification step. The fractions containing pure Sm1 were pooled, dialyzed (20 mM NH4HCO3) and vacuum-dried for further experiments. For the purification of the dimeric forms of Sm1 and Epl1, an immunoaffinity purification assay was required. The fractions containing the dimers were pooled dialyzed and concentrated to continue their purification. Both dimers were purified through an Affi-Gel column where antibodies anti-Sm1 (13) had been immobilized according to the manufacturer’s directions (Affi-Gel® Hz Immunoaffinity Kit, Bio-Rad). In order to obtain the dissociated form of Sm1 and Epl1, the purified dimers were sonicated in trifluoroacetic acid (TFA) at 0 °C (33). The dissociated fraction was then dialyzed, dried under vacuum and dissolved in water.
Sm1 tryptic digestion and MS-fingerprint analysis-The bands corresponding to both isoforms of Sm1 after separation on SDS-PAGE were excised manually and subjected to in gel trypsin digestion. The digestion products were extracted and prepared for MALDI-MS. The peptide mass fingerprint was analyzed in a Shimadzu/Kratos MALDI-TOF mass spectrometer at the Protein Chemistry Lab (Texas A&M University). Protein glycosylation assays- Glycosylation state was assayed using a Glycoprotein Detection Reagent (Pierce Biotechnology) according to the manufacturer suggestions. Different amounts of
3
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

purified Epl1 (dimer) and both isoforms of Sm1 were oxidized with sodium meta-periodate and subjected to reaction with freshly made 0.5% solution of Glycoprotein Detection Reagent in 1 N NaOH. A negative (bovine serum albumin, BSA) and a positive control (ovalbumin, OVA) were also included in the assay. In gel glycoprotein staining was performed after SDS-PAGE protein separation, using the GelCode Glycoprotein Staining Kit (Pierce Biotechnology) as suggested by the manufacturer.
RNA extraction and gene expression analyses-Total RNA from maize or fungal tissue was prepared using the TRIZOL® reagent (Gibco-BRL). Previous to the extraction, plant tissues were ground with a mortar and a pestle in the presence of liquid nitrogen; in the case of the fungal tissue it was lyophilized and crushed prior to the extraction process. RNA quality and PCR products were analyzed by electrophoresis on agarose gels. In all cases, total RNA was DNAse treated before expression assays using a DNA-Free Kit (Ambion). For Sm1 and Epl1 expression assays, quantitative real-time RT-PCR experiments were performed. The constitutively expressed gene for T. reesei actin (X75421) was used to design primers to amplify its homologue in T. virens and T. atroviride as a control reference (13). In maize the expression of allene oxide synthase, AOS; hydroperoxide lyase, HPL; 12-oxo-phytodienoic acid reductase 7, OPR7; phenylalanine ammonia-lyase, PAL was evaluated by quantitative real-time RT-PCR using glyceraldehyde-3-phosphate-dehydrogenase GAPc, as the control reference. The experiments were performed using the QuantiTect® SYBR® green RT-PCR kit (QIAGEN). The reactions were carried out in a 20-μl reaction 1X QuantiTect SYBR Green Master Mix, 1X RT QuantiTect Mix, 200 nM primers (Table 1) and 100 ng total RNA. The reactions were performed in a 7500 Fast Real-Time PCR System (Applied Biosystems) with the conditions suggested by the manufacturer. The absence of primer-dimers was confirmed in reactions without RNA. The experiments were independently repeated two times (with similar results) and each reaction was performed in triplicate using relative quantification analysis. The expression of each specific gene was normalized versus the control reference with the formula 2-∆∆CT where ∆CT = CTspecific gene-CTGAPc gene; ∆∆CT = ∆CT -
arbitrary constant (the highest ∆CT) (PE Applied Biosystems Sequence Detector User Bulletin #2). Statistical analysis was performed by one-way analysis of variance (ANOVA) followed by Turkey's HSD (honestly significant difference) test using VassarStat: (http://faculty.vassar.edu/lowry/VassarStats.html).
Infection of Trichoderma-treated maize plants
with C. graminicola- Cultures of C. graminicola strain M1.001-BH expressing GFP gene were grown for 14 days on PDA at room temperature under constant light. Spores were scraped from plates, filtered through Miracloth (Calbiochem, San Diego, CA), and washed three times with water by centrifugation for 1 min at 10,000 rpm. Spores were counted by using a hemacytometer, and a spore suspension of 6.5 × 104 spores/ml was prepared in a solution of 0.02% Tween-20 in water. Inoculation with T. virens Gv29-8, T. virens-ΔSm1 and T. atroviride IMI206040 strains was performed by coating maize seeds with a latex sticker (Rohm and Haas, Philadelphia, PA), and then coating with fine granules of chlamydospore preparations (34). Control (non-treated) and coated seeds were planted in plastic containers (3.81 × 20.9 cm) containing a soil-less mix (Metromix 366) and incubated in a growth chamber at 25 °C, with a 14 h photoperiod and 60% humidity for two weeks. Fourteen-day-old plants (at the V4 developmental stage) were inoculated with C. graminicola by placing the plants in trays and proceeding as previously described (14). The third leaf from all plants was inoculated with six droplets (10 μl) of the spore suspension, placed on the adaxial side, away from mid vein of the leaf. The trays were sealed and incubated for 24 h at room temperature. After 48 h of incubation in growth chambers, inoculated leaves were used for microscopy or immediately frozen for gene expression assays. For microscopy assays five plants were infected in each of three independent experiments. Forty-eight hours after infection five infection sites were excised form each leaf and analyzed with an Olympus BX51 fluorescence microscope (Olympus America Inc., Mellville).
Infection of maize plants with C. graminicola after elicitor infiltration- The purified monomer of Sm1, dissociated Epl1 and dimers of both, Sm1
4
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

and Epl1 (1 nmol), were syringe infiltrated into the third leaf of 14-day-old plants (13). The infiltrated plants were incubated for 24 h in growth chamber and then, leaves were harvested for RNA extraction or infected with the GFP-tagged C. graminicola strain as described above. In the case of the infection with the GFP-tagged pathogen, three independent experiments were performed infecting five plants in each case. Forty-eight hours after inoculation five infection sites were excised form each leaf and analyzed with a fluorescence microscope as above.
Sequence Analyses- Sequence comparisons were performed using deduced amino acid sequences available in the databases at the National Center for Biotechnology Information (NCBI, http://www.ncbi.nlm.nih.gov/BLAST/) and those previously reported by Seidl et al. (27). Sequence alignments were generated with the CLUSTAL X (version 1.8) software (35) and graphic representations of phylogenetic trees were performed using the TREEVIEW16 software. The presence of putative glycosylation sites was predicted using the resources available at ExPASy proteomics tools (http://www.expasy.ch/tools/).
RESULTS
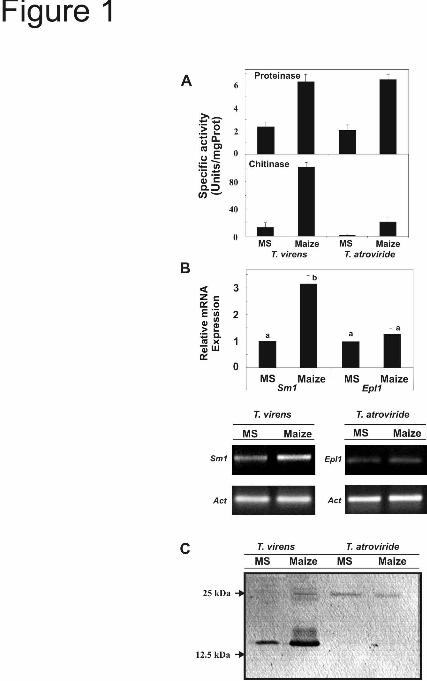
Proteins secreted by T. virens Gv29-8 and T. atroviride IMI206040 in the presence of maize seedlings- Fungal cell wall degrading enzymes (e.g. chitinases, glucanases) are known to be important factors in plant-microbe interactions (36). We determined the level of activity of chitinase, proteinase, β-1,3 and β-1,6 glucanase secreted by both strains in the presence of maize plants. While no glucanase activity was detected in the CF of either strain, similar levels of proteinase activity were detected for both of them. Although T. atroviride presented lower chitinase activity than T. virens, both strains produced higher levels of the activity when cultured in the presence of maize plants (Fig. 1A). The activity of the hydrolytic enzymes was compared also from CFs collected after 4-days culture in VM or VMS media. In both conditions, activity of all enzymes assayed was detected in T. virens and T. atroviride, in either culture condition. In both strains, proteinase and chitinase were down-regulated in VMS medium, the activity of the β-1,6-glucanase was up-regulated in the presence of sucrose, and no differences were detected between the strains for the activity of β-1,3-glucanase
(supplemental Fig. 1). The production of Sm1 and its homologue in T. atroviride (Epl1) was also compared. As previously described (13, 14), Sm1 expression was up-regulated in the presence of maize plants grown hydroponically. However, no significant difference (P<0.01) in the steady state for Epl1 mRNA was evident when T. atroviride was cultured in the presence of plants or under control conditions (Fig. 1B). Polypeptide levels of both Sm1 and Epl1 reflected the differences detected in mRNA levels. In addition, in the presence of plants, Sm1 is mainly produced with a mass suggestive of a monomer, but no monomeric form was detected for Epl1 (Fig. 1C).
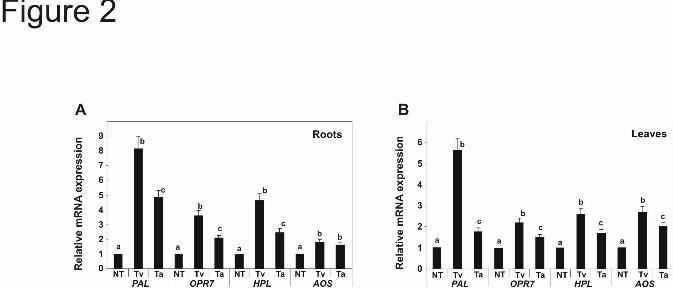
Defense induction and systemic protection in maize seedlings mediated by T. virens Gv29-8 and T. atroviride IMI206040- Previously, we demonstrated that the presence of Sm1 systemically up-regulates genes for defense mechanisms (allene oxide synthase, AOS; hydroperoxide lyase, HPL; 12-oxo-phytodienoic acid reductase 7, OPR7; phenylalanine ammonia-lyase, PAL) in maize plants inoculated with T. virens (14). To compare the effects of T. virens and T. atroviride on the induction of resistance, we tested (locally and systemically) the expression of those genes in maize plants grown in the hydroponic systems and inoculated with fungal tissue. After 48 h inoculation, expression levels of HPL, OPR7 and PAL were assayed in roots and leaves by quantitative real-time RT-PCR. Significant differences in the modulation of gene expression in planta, mediated by T. virens and T. atroviride, were detected (P<0.01). In roots, PAL, HPL, and OPR7 were up-regulated by both strains, with higher levels of expression detected in plants inoculated with T. virens (Fig. 2A). A similar effect of the strains was detected in leaves. The expression of all genes was systemically up-regulated by both strains, with higher levels of expression induced by the presence of T. virens (Fig. 2B). The PCR products were also analyzed on agarose gel electrophoresis after ethidium bromide staining (supplemental Fig. 2). To assess the in vivo relevance of T. virens Gv29-8 and T. atroviride IMI206040 to induce disease resistance, the systemic protection of 2 week-old maize seedlings against the foliar pathogen C. graminicola was assayed (as a reference C. graminicola development, on maize leaves, is presented in supplemental Fig. 3). We also included a T. virens
5
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

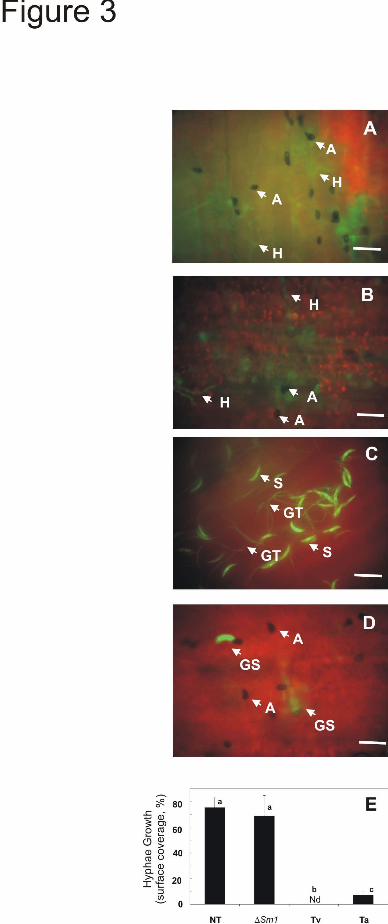
Gv29-8 strain impaired in the expression of Sm1 (ΔSm1) (14). Fluorescent micrographs of the leaf areas inoculated with C. graminicola M1.001-BH-GFP are presented in Figure 3. The pathogen applied to non-Trichoderma treated (control plants) and in plants treated with the ΔSm1 strain showed normal development. The germinated spores produced melanized appressoria and the invading hyphae were colonizing the leaf epidermis and mesophyl (Fig. 3A, B). However, on plants treated with T. virens or T. atroviride, the infection cycle was delayed (Fig. 3C, D) but a distinct difference was observed in the developmental stage of C. graminicola. On plants treated with T. atroviride, the pathogen had already developed melanized appressoria and some primary hyphae already developed (developmental stage 3), but on plants treated with T. virens appressoria had not developed yet (developmental stage 1) and the presence of elongated germ tubes was noticeable (Fig. 3C, D). The quantitative comparison of the pathogen hyphal growth is presented in Figure 3E indicating a significative difference (P<0.05) between T. virens and T. atroviride.
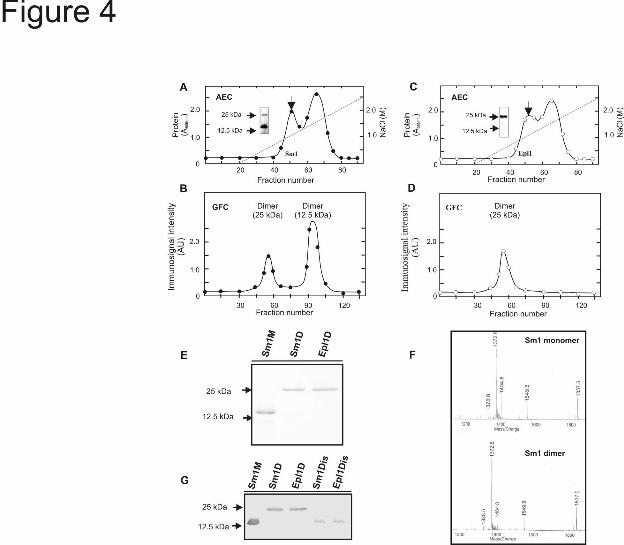
Purification and biochemical characterization of Sm1 and Epl1 isoforms- Since T. virens Gv29-8 and T. atroviride IMI206040 presented differences in the resistance response in maize, we purified and further characterized both isoforms. Both isoforms of Sm1 and Epll eluted from AEC in a single peak at a NaCl concentration of 100 mM (Fig. 4A, C, indicated by arrow). With the second chromatographic procedure (GFC) the monomer of Sm1 was purified to homogeneity but the monomer of Epl1 was not detected (Fig. 4B, D). After GFC the dimer of Sm1 and Epl1 co-eluted with other proteins of similar of molecular weight (not shown). To further purify the dimers of both proteins, an immunoaffinity chromatography using anti-Sm1 antibodies, was implemented. The fractions containing the dimers of Sm1 and Epl1 after GFC, were collected, concentrated and loaded onto the immunospecific column. This purification step allowed the isolation of both dimers to homogeneity as determined by SDS-PAGE (Fig. 4E). To confirm that Sm1 can self-associate to assemble a homodimer, as was demonstrated for Epl1 (27), the purified monomer and dimer were subjected to in gel tryptic digestion. The peptide mass fingerprint was assayed and both isoforms
presented similar patterns (Fig. 4F). We also succeeded in dissociating the purified dimeric isoforms of the Sm1 and Epl1 proteins. Dissociation was achieved by sonicating the protein in the TFA as described by Peña et al. (33). The dissociation of both dimers was determined by SDS-PAGE and western blotting. The immunoassay revealed that the molecular masses of the dissociation products were in the range of the monomeric forms and no dimer was detected by western blotting (Fig. 4G). Dissociation of both Sm1 and Epl1 dimers was also attempted in the presence of a variety of detergents and reducing agents (SDS, mercaptoethanol, DTT, Nonidep-40, Triton-100) with various heating times and combinations of them . However no dissociation was achieved in any of these conditions (not shown).
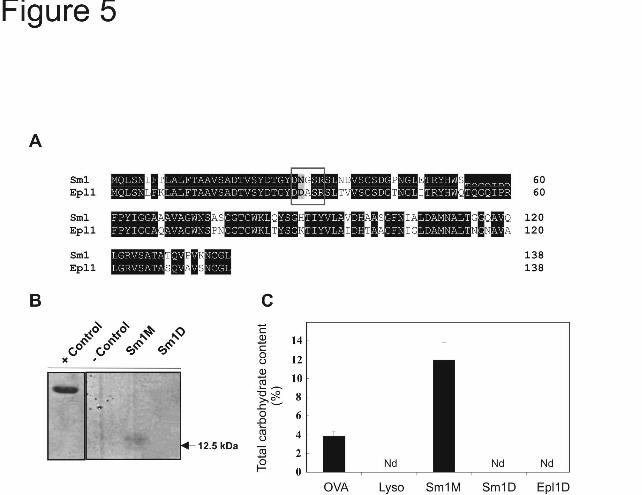
Glycosylation status of Sm1 and Epl1- Deduced amino acid sequences of Sm1 and Epl1 were compared after sequence alignment demonstrating 82% sequence identity (Fig. 5A). Previously, the presence of putative post-translational sites on Sm1 have been described (13), and we compared the presence of these putative sites on Sm1 and Epl1. The N29 residue in Sm1 defines a putative glycosylation site that was replaced by D29 in Epl1. The N29 residue is in the motif DNGSR which is the putative motif recognized by the glyscosylation machinery. Because of this modification in the putative glycosylation motif (N29→D29), no glycosylation site was predicted for Epl1 (Table 2). We performed in gel glycoprotein staining and carbohydrate content quantification using purified Sm1 (monomer and dimer) and Epl1 (dimer). Only the monomeric form of Sm1 was detected in gel as a glycoprotein (Fig. 5B), and the quantitative assay revealed that the monomeric form of Sm1 displays a carbohydrate content of about 14% on a weight basis. On the other hand, no reaction was detected for the dimeric forms of Sm1 or Epl1 in either of the tests (Fig. 5B, C).
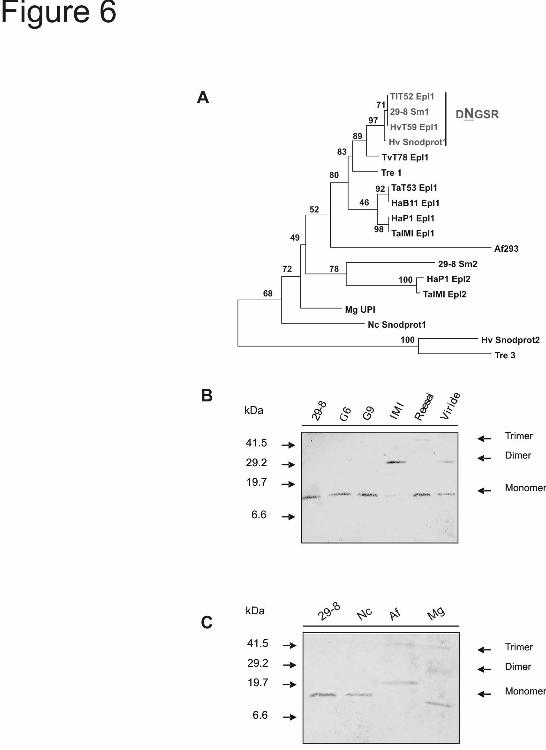
To compare the occurrence of the N-glycosylation site in members of the Epl1, Epl2 and Epl3 clusters reported by Seidl et al. (27) (Table 2), we performed a neighbor-joining analysis including Sm1 from T. virens Gv29-8 and homologous sequences belonging to those clusters. All of the sequences that presented a putative N-glycosylation site belonged to strains of Trichoderma/Hypocrea virens and grouped
6
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

together in a cluster including a sequence from T. longibrachiatum (TlT52 Epl1) (Fig. 6A). We compared the oligomerization state of Sm1/Epl1 proteins in the CFs of some strains of Trichoderma spp. by western blotting assays. In the CF of T. virens Gv29-8, G6 and G9, Sm1 was immunodetected as a single band with a molecular weight similar to that previously described for the monomeric isoform (13). On the other hand in CF of T. viride and T. atroviride the antibodies detected bands corresponding to the monomer and the dimer, and in the case of T. reesei the presence of bands with Mr corresponding to the monomer and a trimer were detected (Fig. 6B). Table 2 lists homologous sequences from Neurospora crassa (Snodprot1), Aspergillus fumigatus (XP_750502.1), and Magnaporthe grisea (UniParc accession number UPI000021A10F). Among those sequences, the presence of a putative glycosylation site was predicted only in N. crassa Snodprot1, and western blotting revealed a single band with Mr corresponding to a monomer (Fig. 6C, “Nc”). Seidl et al. (27) reported that the dimeric form of Epl1 from T. atroviride presented an oxidized W residue in the motif YHWQTQGQIPR. The oxidation of W residues is involved in the aggregation and inactivation of some proteins (37, 38). We compared this motif in Sm1/Epl1 sequences from the different Trichoderma strains analyzed previously (Table 3). This motif is highly conserved in all Trichoderma species, and only Trichoderma/Hypocrea virens sequences presented a point difference (Q→S) next to the W residue to be oxidized. This motif is less conserved in N. crassa, A. fumigatus and M. grisea; however the W residue is still conserved in this region of the protein among these fungi (data not shown).
Differential effect of monomer and dimer forms of Sm1 and Epl1 on defenses in maize leaves-We have previously demonstrated that Sm1 induces plant defense reactions when infiltrated into leaves (13). In the present report we adopted this strategy as a rapid and reliable method to compare the responses that Sm1 and Epl1 induced in maize leaves. To test the effect of the oligomerization state of Sm1 and Epl1 on the expression of plant-defense related genes, the purified forms of the Sm1 monomer and dimer, the Epl1 dimer, and its dissociated form were syringe-infiltrated (1
nmol/leaf) into leaves of 14-days-old maize plants.
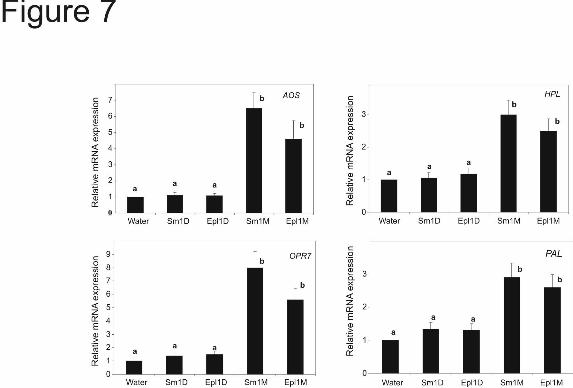
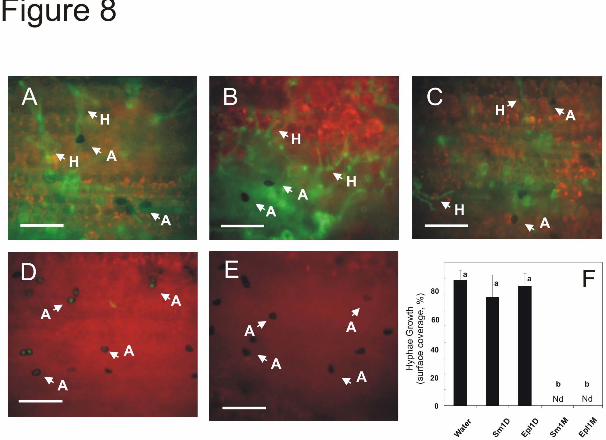
Quantitative real-time PCR experiments indicated that the expression of AOS, HPL, OPR7 and PAL was significatively (P<0.01) up-regulated 24 h after the monomer of either Sm1 or Epl1 was infiltrated (Fig. 7). These experiments indicated that only the monomeric form of either protein up-regulates the expression of the defense-related genes. These data were confirmed by analyzing the PCR amplification products after separation on agarose gels (supplemental Fig. 4). As previously described for other plant species (13), none of the treatments presented phytotoxicty toward maize since no visible lesions on leaves were observed (not shown). The effect of the infiltrated proteins on disease protection was evaluated by inoculating plants with an isolate of the foliar maize pathogen C. graminicola expressing GFP as a reporter (28). Forty-eight hours after inoculation, the leaves that had been infiltrated with water or the dimeric forms of Sm1 or Epl1 (Fig. 8A-C) displayed an extensive hyphal development of the pathogen (similar to that of the non-Trichoderma treated plants, Fig. 3A). Contrastingly, the leaves that had been infiltrated with the monomeric form of either elicitor displayed a more delayed pathogenic development. The spores had only developed mature appressoria but no hyphal colonization was detected (Fig. 8D, E). The quantitative comparison of the pathogen hyphal growth is presented in Figure 8F. DISCUSSION
Protein aggregation or self-association is a widespread phenomenon that occurs in different organisms (39-41). Moreover, recent findings show that protein dimerization or oligomerization is a key factor in the regulation of proteins such as enzymes, ion channels, receptors and transcription factors [Reviewed by Marianayagam et al. (39)]. In this report, we demonstrate that the dimerization of an elicitor protein (Sm1) involved in plant-microbe interactions results in the loss of its eliciting properties, blocking the signaling transduction pathways that lead to the activation of resistance mechanisms in its host plant (maize).
Recently, Sm1 was described as essential for the induction of ISR in plants (13, 14) but only the monomeric form of the elicitor was examined. The characterization of Epl1, a homologue of
7
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

Sm1, was limited to gene expression assays and to the description of structural traits of the protein (27), but elicitor assays were not performed. Here, the ability Epl1 to induce plant defenses is demonstrated for the first time. The functional comparison of both isoforms from Sm1 and Epl1 highlighted that only the monomeric isoform of these small proteins was able to elicit the resistance mechanisms in maize leaves, and no activity was detected for the dimer (Fig. 7, 8, supplemental Fig. 4). As T. virens Gv29-8 and T. atroviride IMI206040 displayed different effects on systemic protection in maize, this result was attributed to variations in the production of the elicitor. Both strains secreted similar levels of hydrolytic enzymes important for Trichoderma-plant interactions, but T. virens produced higher levels of the monomeric form of Sm1. The production of the monomeric form of Epl1 by T, atroviride was at such low levels as not to be immunodetected. However, it was described that various Trichoderma species also produce other elicitors that induce resistance mechanisms in plants (18, 42). Then the fact that T. atroviride still displays ISR might be ascribed to the presence of additional elicitors other than monomeric Epl1.
The dimeric form of Sm1 was only detected in the presence of maize seedlings and Epl1 was only immunorevealed as a dimer in the hydroponic experiment (Fig. 1C). However, when T. virens was grown in VM supplemented with sucrose, glucose, or glycerol, as well as in GYEC medium (Fig. 6B and supplemental Fig. 5), the presence of only the monomer was immunodetected. These data suggest that aggregation of the elicitors may be influenced by the presence of the plant. These observations lead us to hypothesize that as part of the Trichoderma-plant interaction, the plant may alter the aggregation state of Sm1/Epl1 and ultimately affects its ability to induce defenses.
The glycosylation state of pathogenic bacterial cell surface proteins such as flagellin and pilin has been recognized as an important factor in host-pathogen interactions (43, 44). Recent reports describe the importance of the glycosylation state of flagellin from Pseudomonas syringae during plant-pathogen associations (45-47). Glycosylation of flagellin was demonstrated to be strongly related to the pathogen’s ability to cause a hypersensitive reaction leading to cell
death, and could be the specific determinant of compatibility between phytopathogenic bacteria and plant species (45). For Sm1/Epl1 proteins, glycosylation appears to be important in controlling the elicitor’s activity during the plant-microbe interaction. We conclude that the glycosyl moieties might be associated with structural functions, where they contribute to control the monomer-dimer dynamic offering a steric impediment to subunit association. Our results illustrate molecular and biochemical mechanisms that control the manner in which ISR, activated by proteinaceous elicitors, are triggered in plants. We speculate the existence of a very specific mechanism for the recognition of elicitors by plant. This mechanism could either enable the plant to discriminate between monomeric and dimeric forms, or detect special features of the monomeric form that are not longer exposed when the subunits aggregate.
The intermolecular interaction between monomers of Sm1 or Epl1 is resistant to SDS/β-mercaptoethanol treatment (and other detergents at various temperatures), suggesting an inter-subunit covalent interaction other than disulfide-bridges. In the case of hydrophobins secreted by Trichoderma spp. and other fungi, different aggregation states with strong interactions (similar to Sm1 and Epl1) have been described (33, 48-50). However, the molecular mechanisms involved in such strong interactions were not further studied. In humans, plants and fungi, it has been reported that oxidative cross-linking can control activity and aggregation states of proteins generating post-translational covalent linkages resistant to SDS/β-mercaptoethanol treatment (37, 38, 51-57). In T. reesei and Agaricus bisporus, an enzymatically-driven protein cross-linking in which tyrosines are involved, has been characterized (57). An oxide-reduction mechanism controlling the activity and the aggregation state of human superoxide dismutase 1 (hSOD1) has also been described (37). This mechanism involves the oxidation of a tryptophan residue to its peroxyl radical to form kynurenine, a highly reactive species that drives the reaction between both monomers and generates a covalent bond between the molecules (37). Peptide mass fingeprinting and peptide analysis of the dimer form of Epl1 demonstrated the presence of one peptide (YHWQTQGQIPR) with two different oxidation states. It was proposed that the
8
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

9
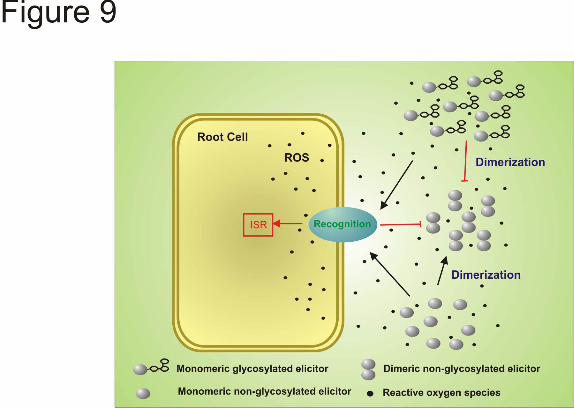
oxidations occurred on the tryptophan residue to form N-fromylkynurenine (27). Likewise, in the amino acid sequence of Sm1, we identified a segment of the sequence (YHWSTQGQIPR) very similar to that for Epl1 presenting only one substitution (Q→S) (Table 3). This motif was 100% conserved in all Sm1-homologues from all Trichoderma/Hypocrea virens sequences analyzed, and in the homologous sequence identified in T. viride T78 (Table 3). When Sm1 is in contact with plant cells, an oxidative burst is elicited (13). This accumulation of reactive oxygen species in the plant cells likely stimulates an oxidative-driven dimerization of the monomeric proteins, altering the concentration of the active elicitor in the environment. However, in the case of T. virens strains, the presence of a glycosyl moiety on the elicitor may prevent that dimerization, keeping in solution a protein with high elicitor activity (Fig. 9).
We also analyzed the sequences of Sm1-homologues in various fungal strains to identify differences in the glycosylation motif in protein from different species. The glycosylation site was only present among members of a closely-related cluster containing sequences from Trichoderma/Hypocrea virens strains and a Sm1-homologue from T. longibrachiatum T52 (Fig. 6, Table 2). According to phylogenetic reconstruction of the genus Trichoderma using parsimony analysis of ITS1-, ITS2-, mitSSU
DNA-, tef1- and ech42-sequences, it was demonstrated that T. longibrachiatum is distantly related to T. virens strains (58). This phylogenetic difference coupled with a high similarity between T. virens Sm1 and T. longibrachiatum Epl1 sequences might suggest functional and molecular characteristics conserved throughout the evolution of both species. Remarkably, we also found the presence of the putative glycosylation site in some other sequences of CP proteins from various fungal species (Table 2, not shown). The aggregation and oligomerization patterns of Sm1 homologues in different filamentous fungus strains (Fig. 6B, C) correlate with the prediction that oligomerization may be prevented by the glycosylation of the polypeptides. According to these results, we suggest that the CP protein family be divided into two sub-families according to the presence or absence of the glycosylation site and their ability to self-aggregate.
Our future research is focused on characterizing modified proteins lacking the glycosylation site and/or the tryptophan residue putatively involved in an oxidative cross-linking of the monomers. Further structural analyses, such as 3-D structure determination and carbohydrate moiety characterization, will contribute to our understanding of the processes involved in the recognition of elicitors and the initiation of ISR in plants.
REFERENCES
1. Cook RJ, Thomashow LS, Weller DM, Fujimoto D, Mazzola M, Bangera G, Kim D (1995) Proc Natl Acad Sci USA 92, 4197-4201 2. Vallard GE, Goodman RM (2004) Crop Sci 44, 1920-1934 3. Nimchuk Z, Eulgem T, Holt BF3rd, Dangl JL (2003) Annu Rev Genet 37, 579-609 4. Nürnberger T, Brunner F, Kemmerling B, Piater L (2004) Immunol Rev 198, 249-266 5. Bais HP, Park SW, Weir TL, Callaway RM, Vivanco JM (2004) Trends Plant Sci 9, 26-32 6. Pozo MJ, Van Loon LC, Pieterse CMJ (2005) J Plant Growth Regul 23, 211-222 7. Reymond P, Grünberger S, Paul K, Müller M, Farmer EE (1995) Proc Nat Acad Sci USA 92,
4145-4149 8. Ramonell KM, Zhang B, Ewing RM, Chen Y, Xu D, Stacey G, Somerville S (2002) Molec Plant
Pathol 3, 301-311 9. Kaku H, Nishizawa Y, Ishii-Minami N, Akimoto-Tomiyama C, Dohmae N, Takio K, Minam E,
Shibuya N (2006) Proc Nat Acad Sci USA 103, 11086-11091 10. Felix G, Duran JD, Volko S, Boller T (1999) Plant J 18, 265-276 11. Brunner F, Rosahl S, Lee J, Rudd JJ, Geiler C, Kauppinen S, Rasmussen G, Scheel D,
Nürnberger T (2002) EMBO J 21, 6681-6688 12. Kunze G, Zipfel C, Robatzek S, Niehaus K, Boller T, Felix G (2004) Plant Cell 16, 3496-3507 13. Djonović S, Pozo MJ, Dangott LJ, Howell CR, Kenerley CM (2006) Mol Plant Microbe Interact
19, 838-853
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

14. Djonović S, Vargas WA, Kolomiets MV, Horndeski M, Wiest A, Kenerley CM (2007) Plant Physiol 145, 875-889
15. Ferrari S, Galletti R, Denoux C, De Lorenzo G, Ausubel FM, Dewdney J (2007) Plant Physiol 144, 367-379
16. Yedidia I, Shoresh M, Kerem K, Benhamou N, Kapulnik Y, Chet I (2003) Appl Environ Microbiol 69, 7343-7353
17. Shoresh M, Yedidia I, Chet I (2005). Phytopathology 95, 76-84 18. Viterbo A, Wiest A, Brotman Y, Chet I, Kenerley C (2007) Mol Plant Pathol 8,737-746 19. Wosten HAB (2001) Annu Rev Microbiol 55, 625-646 20. Pan SC, Cole GT (1995) Infect Immun 63, 3994-4002 21. Hemmann S, Blaser K, Crameri R (1997) Am J Respir Crit Care Med 156, 1956-1962 22. Hall N, Keon JPR, Hargreaves JA (1999) Physiol Mol Plant Pathol 55, 69-73 23. Pazzagli L, Cappugi G, Manao G, Camici G, Santini A, Scala A (1999) J Biol Chem 274, 24959-
24964 24. Wilson LM, Idnurm A, Howlett BJ (2002) Mol Plant Pathol 3, 487-493 25. Mou Z, Fan W, Dong X (2003) Cell 113, 935-944 26. Durrant WE, Dong X (2004) Annu Rev Phytopathol 42, 185-209 27. Seidl V, Marchetti M, Schandl R, Allmaier G, Kubicek CP (2006) FEBS J 273, 4346-4359 28. Sukno SA, García VM, Shaw BD, Thon MR (2008) Appl Environ Microbiol 74, 823-832 29. Vogel HJ (1956) Microbiol Genet Bull 13, 42-43 30. Djonovic S, Vittone G, Mendoza-Herrera A, Kenerley C (2007) Mol Plant Pathol 8, 469-480 31. Laemmli UK (1970) Nature 227, 680-685 32. Renart J, Sandoval IV (1984) In: Methods in enzymology vol. 104, Jacoby WB ed,. pp. 455-460.
Academic, New York 33. Peñas MM, Ásgeirsdóttir SA, Lasa I, Culiañez-Macià FA, Pisabarro AG, Wessels JGH, Ramírez
L (1998) Appl Environ Microbiol 64, 4028-4034 34. Weaver M, Kenerley C (2005) Biocontrol Sci Technol 15, 847-857 35. Harman GE (2006) Overview of mechanisms and uses of Trichoderma spp. Phytopathology 96,
190-194 36. Thompson JD, Gibson TJ, Plewniak FM, Jeanmougin F, Higgins DG (1997) Nucleic Acids Res
15, 4876-4882 37. Zhang H, Andrekopoulos C, Joseph J, Chandran K, Karoui H, Crow JP, Kalyanaraman B (2003)
J Biol Chem 278, 24078-24089 38. Fu X, Kao JLF, Bergt C, Kassim SY, Huq NP, d'Avignon A, Parks WC, Mecham RP, Heinecke
JW (2004) J Biol Chem 279, 6209-6212 39. Marianayagam NJ, Sunde M, Matthews JM (2004) Trends Biochem Sci 29, 618-625 40. Paul R, Abel S, Wassmann P, Beck A, Heerklotz H, Jenal U (2007) J Biol Chem 282, 29170-
29177 41. Wayne N, Bolon DN (2007) J Biol Chem 282, 35386-35395 42. Brotman Y, Briff E, Viterbo A, Chet I (2008) Plant Physiol, in press
(doi:10.1104/pp.108.116243) 43. Benz I, Schmidt MA (2002) Mol Microbiol 45, 267-276 44. Logan SM (2006) Microbiology 152, 1249-1262 45. Taguchi F, Shimizu R, Inagaki Y, Toyoda K, Shiraishi T, Ichinose Y (2003) Plant Cell Physiol
44, 342-349 46. Takeuchi K, Taguchi F, Inagaki Y, Toyoda K, Shiraishi T, Ichinose Y (2003) J Bacteriol 185,
6658-6665 47. Takeuchi K, Ono H, Yoshida M, Ishii T, Katoh E, Taguchi F, Miki R, Murata K, Kaku H,
Ichinose Y (2007) J Bacteriol 189, 6945-6956 48. Wessels JGH (1994) Annu Rev Phytopathol 32, 413-437 49. Torkkeli M, Serimaa R, Ikkala O, Linder M (2002) Biophys J 83, 2240-2247 50. Linder MB, Szilvay GR, Nakari-Setälä T, Penttilä ME (2005) FEMS Microbiol Rev 29, 877-896
10
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

51. Bradley DJ, Kjellborn P, Lamb CJ (1992) Cell 70, 21-30 52. Heinecke JW, Li W, Francis GA, Goldstein JA (1993) J Clin Invest 91, 2866-2872 53. Brisson LF, Tenhaken R, Lamb C (1994) Plant Cell 6, 1703-1712 54. Brown I, Trethowan J, Kerry M, Mansfield J, Bolwell GP (1998) ) Plant J 15, 333-343 55. Ziouzenkova O, Asatryan L, Akmal M, Tetta C, Wratten ML, Wich GL, Jürgens G, Heinecke J,
Sevanian A (1999) J Biol Chem 274, 18916-18924 56. Larios JM, Budhiraja R, Fanburg BL, Thannickal VJ (2001) J Biol Chem 276, 17437-17441 57. Mattien M-L, Lantto R, Selinheimo E, Kruus K, Buchert, J (2008) J Biotechnol 133, 395-402 58. Kullnig-Gradinger CM, Szakacs G, Kubicek CP (2002) Mycol Res 106, 757-767
FOOTNOTES * Present address: Department of Molecular Biology, Massachusetts General Hospital and Department of
Genetics, Harvard Medical School, Boston, MA 02114.
# The abbreviations used are: AEC anion exchange chromatography; AOS, allene oxide synthase; CF, culture filtrate; GAPc, glyceraldehyde-3-phosphate-dehydrogenase; GFC, gel filtration chromatography; GFP, green fluorescent protein; HPL, hydroperoxide lyase; ISR, induced systemic resistance; OPR7, 12-oxo-phytodienoic acid reductase 7; PAGE, polyacrylamide gel electrophoresis; PAL, phenylalanine ammonia-lyase.
† This work was supported by grants of the U.S. Department of Agriculture National Research Initiative (2003-35316-13861) and the National Research Foundation (IOB0445650) to CMK.
Acknowledgements-We thank to E. Monte (Universidad de Salamanca, Spain), A. Herrera-Estrella (CINVESTAV, Mexico), C. Howell (USDA, College Station, Texas) and H. Wilkinson (Texas A&M University, College Station, Texas) for providing fungal isolates for this study.
FIGURE LEGENDS Fig. 1. Comparison of secreted proteins from T. virens and T. atroviride incubated in the presence of maize plants in a hydroponic system . As a control both strains were cultured in MS medium supplemented with 0.05% sucrose, (13). After 48 h incubation, mycelia were collected for gene expression, and culture filtrates (CFs) were used to assay enzymatic activities. A, Specific activity of chitinase and proteinase in CF. The values are the mean value ± standard error of two independent experiments. B, (Upper panel) Quantitative RT-PCR analysis of Sm1 and Epl1 from total RNA of T. virens Gv29-8 and T. atroviride IMI206040, respectively, cultured in MS or with maize plants. The gene expression data presented were normalized to a reference control and depict the mean value ± standard error of two independent experiments, Columns with different letters differ significantly according to Turkey’s HSD test at a significant level of 1%. In the lower panel, PCR products visualized after agarose gel electrophoresis and ethidium bromide-staining. Amplification of the actin gene was used as an internal loading control. C, Sm1 and Epl1 immunodetection in CF from hydroponic systems. Proteins were separated by SDS-PAGE on 15% polyacrylamide gels and blotted onto nitrocellulose membranes. Blots were probed with anti-Sm1 antibodies. Fig. 2. Quantitative RT-PCR analysis of defense-related genes in roots and leaves of T. virens- or T. atroviride-treated maize plants. Plants grown without Trichoderma were included as controls. Two days after inoculation, qRT-PCR assays were performed with 100 ng total RNA extracted from: A, roots or B, leaves. The expression of AOS, HPL, OPR7 and PAL was analyzed using the primers described in Table 1. GAPc was used as the reference internal control and the expression results were normalized to its abundance. The values shown are the average of two independent experiments ± standard error. Columns with different letters (for every gene) differ significantly according to Turkey’s HSD test at a significant
11
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

level of 1%.NT, non-treated; Tv, plants inoculated with T. virens GV29-8; Ta, plants inoculated with T. atroviride IMI206040. Fig. 3. Systemic disease protection mediated by T. virens Gv29-8 or T. atroviride IMI206040 in maize plants. The development of the GFP-tagged C. graminicola was assayed on leaves of: A, non-Trichoderma treated maize plants; B, plants that had been root-inoculated with T. virens ΔSm1; C, T. virens Gv29-8 or D, T. atroviride IMI206040. Fourteen-day old plants were challenged with the pathogen and analyzed 48 h after infection. E, quantification of C. graminicola growth on leaves. The data presented show the percentage of the inoculation area covered by the pathogen hyphae. The bars depict the mean value ± SD determined in pictures taken in three independent experiments. Columns with different letters differ significantly according to Turkey’s HSD test at a significant level of 5% Fluorescent micrographs of the pathogen were taken with an Olympus BX-51 fluorescent microscope with excitation from 470 to 490, emission from 510 to 550. The areas on the micrographs were determined using ImageJ software. A, appressoria; S, spore; GS, germinating spore; GT, germ tube; H, hypha; Nd, no detected. Bars = 50 μm. Fig. 4. Purification and molecular studies of Sm1 and Epl1. Sm1 and Epl1 were isolated from CF of T. virens Gv29-8 or T. atroviride IMI602040 cultured in VMS liquid medium for seven days. A and B, Purification of Sm1 through anion exchange and gel filtration chromatography, respectively. C and D, Purification of Epl1 through anion exchange and gel filtration chromatography, respectively. The protein elution profile was assayed by determining protein concentration, and the elution of Sm1 or Epl1 was followed by western blot. The inserts in panels (A) and (C) show the immunodetection of both proteins after concentration of the peak from anion exchange chrormatography. E, Sm1 and Epl1 dimeric isoforms were purified to homogeneity by immunoaffinity chromatography and the purified proteins were analyzed on SDS-PAGE. F, Comparison of the MS polypeptide fingerprint of the Sm1 monomer and dimer after tryptic digestion. Both isoforms were in gel digested and the polypeptides were analyzed by MS-MALDI. G, Comparison of dissociated Epl1 and Sm1 isoforms by immunoassay. Proteins were electroblotted onto nitrocellulose membranes, and protein blots were probed with antibodies to Sm1. AEC, anion exchange chromatography, GFC, gel filtration chromatography, Sm1M, monomer form of Sm1; Sm1D, dimer form of Sm1; Sm1Dis, dissociated form of dimer Sm1; Epl1D, dimer form of Epl1; Epl1Dis, dissociated form of Epl1. Fig. 5. Protein analysis and post-translational modifications on Sm1 and Epl1. A, Alignment of deduced amino acid sequences of Sm1 and Epl1. Immature protein sequences were aligned using Clustal X software. Black-shading indicates residues conserved in both sequences. The putative motif for glycosylation is denoted by the grey frame and the residue predicted to be glycosylated is grey-shaded. B, In gel glycoprotein staining. The purified monomer and dimer of Sm1 were separated in a 15% SDS-PAGE gel, and carbohydrate-containing proteins were stained. C, Carbohydrate detection and quantification on Sm1 and Epl1 isoforms. The assay was performed in triplicate, and using two batches of proteins purified independently. The data are the mean value ± standard error. Sm1M, monomer form of Sm1; Sm1D, dimer form of Sm1; Epl1D, dimer form of Epl1; OVA, ovalbumin; Lys, lysozyme; Nd, no detected. Fig. 6. Analysis of the occurrence of Sm1 and Epl1 in different fungal strains. A, Phylogenetic reconstruction of Sm1/Epl1 homologue proteins. Protein sequences were selected according to the study performed by Seidl et al. (27). Sequence designation is presented in Tables 2 and 3. The tree was constructed with MEGA 3.1 after sequence alignment using Clustal X with a BLOSSUM matrix and bootstrap trial of 1000. B, Immunodetection of Sm1/Epl1 homologues in different Trichoderma strains. Polypeptides were separated by SDS-PAGE and blotted onto nitrocellulose membranes. Membranes were probed with anti-Sm1 antibodies. C, Immunodetection of Sm1/Epl1 homologues in non-Trichodema species. Polypeptides in CF of A. fumigatus (Af), N. crassa (Nc) and M. grisea (Mg), were electrophoresed on denaturing 15% acrylamide gels, transferred to nitrocellulose membranes and probed
12
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

with anti-Sm1 antibodies. 29-8, T. virens Gv29-8; G6, T. virens G6; G9, T. virens G9; IMI, T. atroviride IMI206040; Reesei, T. reesei; Viride, T. viride. Fig. 7. Analysis of the expression of defense-related genes (AOS, HPL, OPR7 and PAL) in maize leaves after Sm1 and Epl1 infiltration. The third leaves of 14-day-old maize plants were infiltrated with 1 nmol/leaf of Sm1 monomer (Sm1M), Sm1 dimer (Sm1D), dissociated Epl1 (Epl1M), Epl1 dimer (Epl1D), or water as a control (Water). After 24 h treatment, total RNA was extracted and used for quantitative real-time RT-PCR assays. Expression of GAPc was used as quantitative internal control. The gene expression data presented were normalized to the reference control. The data depicted are the mean values ± standard error of two independent experiments. Columns with different letters differ significantly according to Turkey’s HSD test at a significant level of 1% Fig. 8. Effect of Sm1 and Epl1 isoforms on the development of the foliar pathogenic fungus C. graminicola. Leaves were infiltrated with water (A), dimeric Sm1 (B), dimeric Epl1 (C), monomeric Sm1 (D) or dissociated Epl1 (E) and inoculated with GFP-tagged C. graminicola M1.001-BH strain 24h post-infiltration. F, quantification of C. graminicola growth on leaves. The data presented show the percentage of the inoculation area covered by the pathogen hyphae. The bars depict the mean value ± SD determined in pictures taken in three independent experiments. Columns with different letters differ significantly according to Turkey’s HSD test at a significant level of 1%. Pathogen development was compared 48h after inoculation with an Olympus BX-51 fluorescent microscope with excitation from 470 to 490, emission from 510 to 550. The areas on the micrographs were determined using ImageJ software. Bars = 50 μm. A, appressoria; H, hypha; Nd, no detected. Bars =50 μm. Fig. 9 Proposed biochemical processes involved in the control of ISR induction mediated by proteinaceous elicitors. When the elicitors are produced as monomer they are recognized in the cell interface, the defense responses are activated and ROS reaction is elicited. The monomeric form of the elicitors in their non-glycosylated state would be susceptible to oxidative-driven dimerization and not longer active to induce ISR. However, when the monomeric elicitor is produced in its glycosylated form, the presence of the glycosyl moiety will not allow to the monomers to dimerize and they will remain in their active form for activating the defense responses in plants.
13
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from
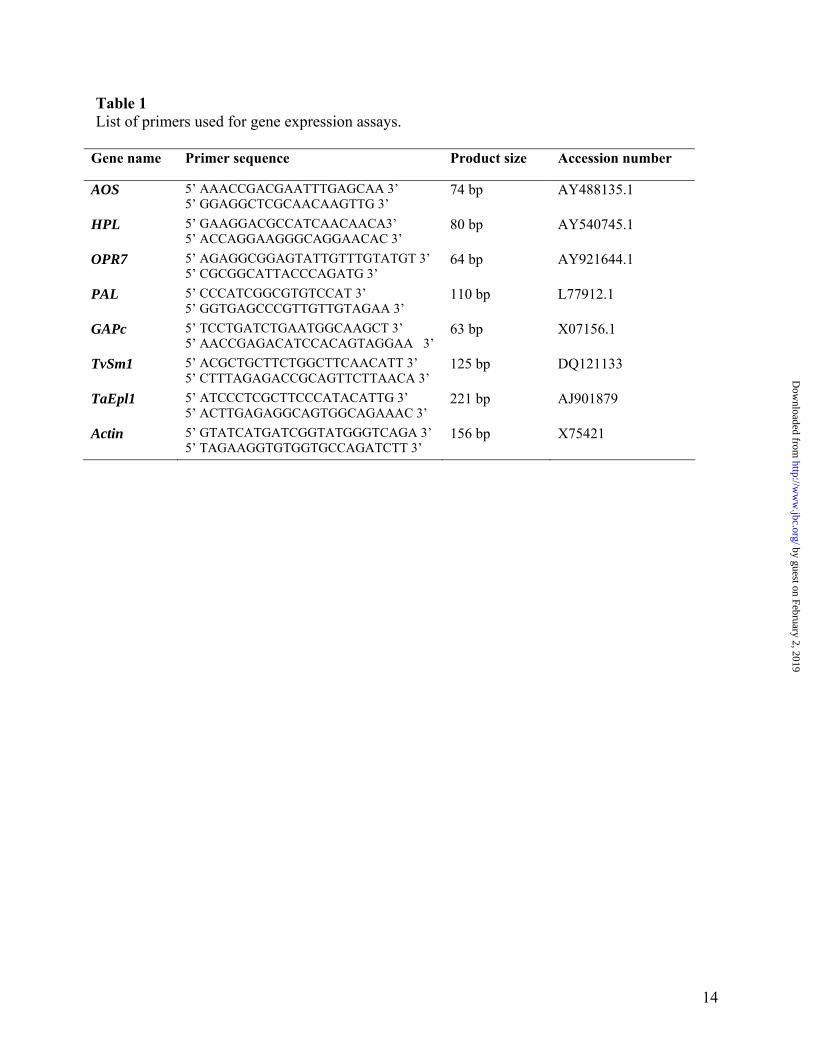
Table 1 List of primers used for gene expression assays.
Gene name Primer sequence Product size Accession number
AOS 5’ AAACCGACGAATTTGAGCAA 3’ 5’ GGAGGCTCGCAACAAGTTG 3’
74 bp AY488135.1
HPL 5’ GAAGGACGCCATCAACAACA3’ 5’ ACCAGGAAGGGCAGGAACAC 3’
80 bp AY540745.1
OPR7 5’ AGAGGCGGAGTATTGTTTGTATGT 3’ 5’ CGCGGCATTACCCAGATG 3’
64 bp AY921644.1
PAL 5’ CCCATCGGCGTGTCCAT 3’ 5’ GGTGAGCCCGTTGTTGTAGAA 3’
110 bp L77912.1
GAPc 5’ TCCTGATCTGAATGGCAAGCT 3’ 5’ AACCGAGACATCCACAGTAGGAA 3’
63 bp X07156.1
TvSm1 5’ ACGCTGCTTCTGGCTTCAACATT 3’ 5’ CTTTAGAGACCGCAGTTCTTAACA 3’
125 bp DQ121133
TaEpl1 5’ ATCCCTCGCTTCCCATACATTG 3’ 5’ ACTTGAGAGGCAGTGGCAGAAAC 3’
221 bp AJ901879
Actin 5’ GTATCATGATCGGTATGGGTCAGA 3’ 5’ TAGAAGGTGTGGTGCCAGATCTT 3’
156 bp X75421
14
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from
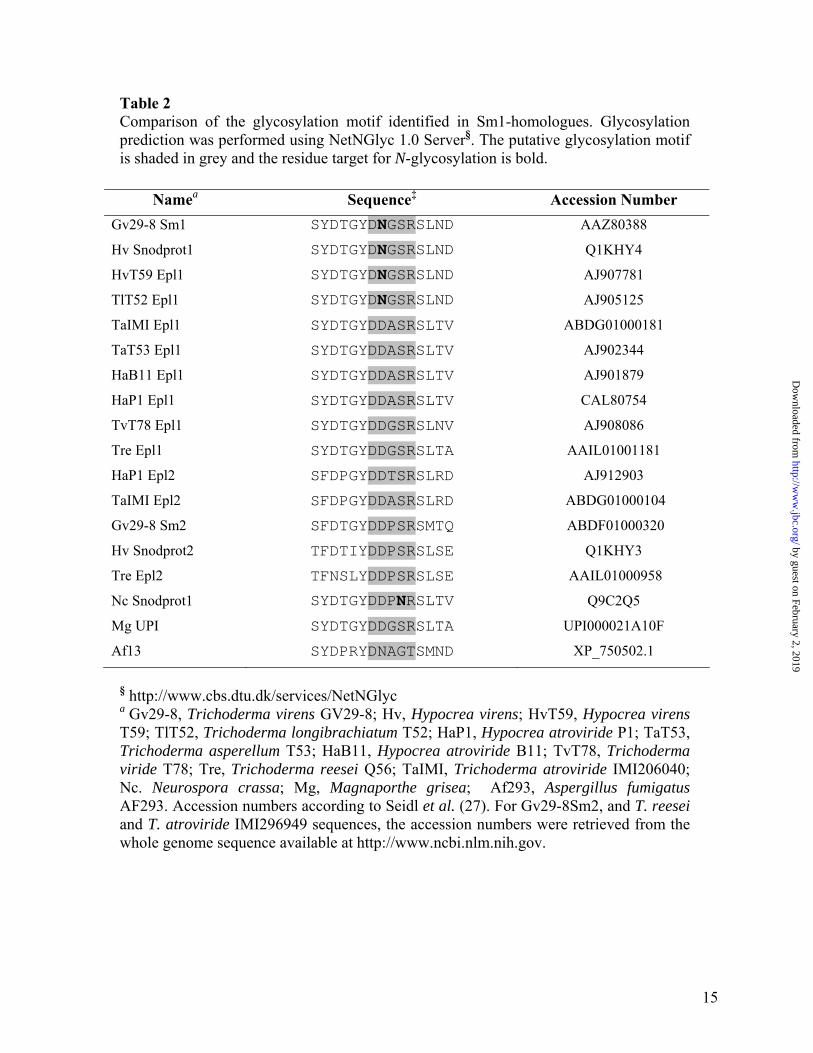
Table 2 Comparison of the glycosylation motif identified in Sm1-homologues. Glycosylation prediction was performed using NetNGlyc 1.0 Server§. The putative glycosylation motif is shaded in grey and the residue target for N-glycosylation is bold.
Namea Sequence‡ Accession Number Gv29-8 Sm1 SYDTGYDNGSRSLND AAZ80388
Hv Snodprot1 SYDTGYDNGSRSLND Q1KHY4
HvT59 Epl1 SYDTGYDNGSRSLND AJ907781
TlT52 Epl1 SYDTGYDNGSRSLND AJ905125
TaIMI Epl1 SYDTGYDDASRSLTV ABDG01000181
TaT53 Epl1 SYDTGYDDASRSLTV AJ902344
HaB11 Epl1 SYDTGYDDASRSLTV AJ901879
HaP1 Epl1 SYDTGYDDASRSLTV CAL80754
TvT78 Epl1 SYDTGYDDGSRSLNV AJ908086
Tre Epl1 SYDTGYDDGSRSLTA AAIL01001181
HaP1 Epl2 SFDPGYDDTSRSLRD AJ912903
TaIMI Epl2 SFDPGYDDASRSLRD ABDG01000104
Gv29-8 Sm2 SFDTGYDDPSRSMTQ ABDF01000320
Hv Snodprot2 TFDTIYDDPSRSLSE Q1KHY3
Tre Epl2 TFNSLYDDPSRSLSE AAIL01000958
Nc Snodprot1 SYDTGYDDPNRSLTV Q9C2Q5
Mg UPI SYDTGYDDGSRSLTA UPI000021A10F
Af13 SYDPRYDNAGTSMND XP_750502.1
§ http://www.cbs.dtu.dk/services/NetNGlyc a Gv29-8, Trichoderma virens GV29-8; Hv, Hypocrea virens; HvT59, Hypocrea virens T59; TlT52, Trichoderma longibrachiatum T52; HaP1, Hypocrea atroviride P1; TaT53, Trichoderma asperellum T53; HaB11, Hypocrea atroviride B11; TvT78, Trichoderma viride T78; Tre, Trichoderma reesei Q56; TaIMI, Trichoderma atroviride IMI206040; Nc. Neurospora crassa; Mg, Magnaporthe grisea; Af293, Aspergillus fumigatus AF293. Accession numbers according to Seidl et al. (27). For Gv29-8Sm2, and T. reesei and T. atroviride IMI296949 sequences, the accession numbers were retrieved from the whole genome sequence available at http://www.ncbi.nlm.nih.gov.
15
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

Table 3 Comparison of the oxidized peptide of Epl1 with homologue sequences from various Trichoderma species.The W residue reported to be oxidized in the dimeric form of Epl1 is shown in bold, and Q→ S subtitution in T. virens strains is underlined. Identical residues are grey-shaded.
16
aGv29-8, Trichoderma virens GV29-8; Hv, Hypocrea virens; HvT59, Hypocrea virens T59; TlT52, Trichoderma longibrachiatum T52; HaP1, Hypocrea atroviride P1; TaT53, Trichoderma asperellum T53; HaB11, Hypocrea atroviride B11; TvT78, Trichoderma viride T78; Tre, Trichoderma reesei Q56; TaIMI, Trichoderma atroviride IMI206040. For accession numbers see Table 2.
Namea Sequence Gv29-8 Sm1 YHWSTQGQIPR
Hv Snodprot1 YHWSTQGQIPR
HvT59 Epl1 YHWSTQGQIPR
TlT52 Epl1 YHWSTQGQIPR
TaIMI Epl1 YHWQTQGQIPR
TaT53 Epl1 YHWQTQGQIPR
HaB11 Epl1 YHWQTQGQIPR
HaP1 Epl1 YHWQTQGQIPR
TvT78 Epl1 YHWSTQGQIPR
Tre Epl1 YHWQTQGQIPK
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

Walter A. Vargas, Slavica Djonovic, Sukno A. Serenella and Charles M. Kenerleyplants
Dimerization controls the activity of fungal elicitors that trigger systemic resistance in
published online May 15, 2008J. Biol. Chem.
10.1074/jbc.M802724200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2008/05/20/M802724200.DC1
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from