Embed Size (px)
Citation preview

Disk-Elektrophoretische Untersuchungen zur A rtcharakterisierung paludicoler Tricladen 263
- 1969: Paradigm for an evolutionary process. I n : Towards a theoretical biology 2 (ed. WAD- WADDINGTON, C. H. , 1968: Does evolution depend on random search? Towards a theoretical
biology 1, 111-119 (ed. WADDINGTON, C. H.). Edinburgh University Press, Edinburgh. DINGTON, C. H.). Edinburgh University Press, Edinburgh.
WAHLERT, G. v., 1965: The role of ecological factors in the origin of higher levels of organi- zation. Syst. Zool. 14, 288-300.
WATSON, J. D., 1969: Die Doppelhelix. Rowohlt-Verlag, Reinbek. WEBER, H. , 1954: Stellung und Aufgaben der Morphologie der Gegenwart. Verhdlg. dtsch.
- 1958: Konstruktionsmorphologie. Zool. Jb. Abt. allg. Zool. 68, 1-112. WICKLER, W., 1968: Mimikry. Kindlers Universitits-Bibliothek, Miinchen. ZIMMERMANN, W., 1967: Methoden der Evolutionswissenschaft (= Phylogenetik). I n : HELIERER,
ZUSI, R. L., 1971 : Functional anatomy in systcmatics. Taxon 20, 75-84. ZWICK, P., 1969 : Das phylogenetische System dcr Plecopteren als Ergebnis vergleichend-ana-
2001. Ges. 18, 137-158.
G. (Hrsgb.) : Die Evolution der Organismen I, 61-160. G. Fischer-Verlag, Stuttgart.
tomischer Untersuchungen. Inauguraldiss. Natwiss. Fakul t i t d. Univ. Kiel.
Anschrift der Verfasser: Dr. DIETER STEFAN PETERS und Dr. WOLFGANG FRIEDRICH GUTMANN, Forschungs-Institut Senckenberg, 6 Frankfurt a. M., Senckenberg- Anlage 25
Aus dem I . Zoologischen Institut der Universitat Wien
Disk-Elektrophoretische Untersuchungen zur Frage der Artcharakterisierung paludicoler Tricladen
(Platyhelminthes: Turbellaria)
Von ROLAND PETER
Mit 22 Abbildungen
Eingegangen a m 6 . Mai 1971
I. Einleitung
Datieren die ersten vergleichenden Proteinuntersuchungen an Tieren von der Jahrhundert- wende (NUTTALL 1904), so brachte die Aufklarung des genetischen Code 1961 und 1962 (NI- RENLIERG, MATTHAEI e t al. 1963; YANOFSKY et al. 1963) die experimentelle Bestatigung dafiir, dai3 die Primarstruktur der Proteine in den Basenabfolgen der Desoxyribonukleinsauren (DNS) eindeutig vorgegeben und somit cine getreue Ubersetzung der genetischen Information ist. Eine Analyse der Proteinstrukturen von Organismen erfagt also ihre genetische Konstitution und sollte sich ebenso wie morphologische und andere Methoden fur die Abgrenzung taxono- mischer Einheiten und ihre verwandtschaftliche Gruppierung eignen. SILILEY (1962) gibt dazu cine iibersichtliche Begriindung und zeigt darin auch die moglichen Methoden einschlienlich ihrer Problematik und historischen Entwicklung auf. Ein exakter Vergleich ist selbstverstandlich nur iiber die Aufkl l rung der Primirstrukturen, also der Aminosauresequenzen, einzelner Proteine moglich, wie dies bereits mehrfach festgestellt worden ist (so etwa von SIBLEY 1962 und von BIANCHI 1967). Entsprechende Untersuchungen sind insbesondere a n Mammaliern durchgefiihrt worden und fuhrten auch zur Aufstellung phylogenetischer Beziehungen; FLORKIN (1966, pp. 144-160) berichtet dariiber zusammenfassend. Man mug aber erkennen, dai3 solche Analysen, sollen sie nicht auf wenige Artcn und somit von vornherein in ihrer phylogenetischen Aussage beschrinkt bleiben, beim gegenwartigen Stand der Methodik augerst aufwendig sind. Es er- scheint dahcr sinnvoll, auch weniger exakte Methoden anzuwenden, die Proteine zwar meist
2. zool. Sysr. Evolut.-forsch. 9 (1971) 263-318 @ 1971 Vcrlag Paul Parey, Hamburg und Berlin

264 R. Peter
nicht eindeutig charakterisieren, dafur aber Gemische von Eiweiflstoffen mit relativ geringem Aufwand trennen und so einem Vergleich zuganglich machen.
Seit der Beschreibung der Papierelektrophorese durch KONIG (1937) wurden zahlreiche andere Elektrophoresemethoden entwickelt, welhe verschiedene Tragermaterialien beniitzcn und eine Verbesserung des Trennvermogens Zuni Ziel haben. Sie fanden auf vershiedenste Gebiete ausgedehnte Anwendung, vor allem auch zur Trennung und Kennzeihnung von Pro- teinen. Auch systematische und phylogenetische Probleme wurden damit bearbeitet. Der von LEONE (1964) herausgegebene Symposiumbericht ,,Taxonomic Biochemistry and Serology" vermittelt einen Uberblidc uber die bis dahin publizierten Untersuchungen, deren Schwerge- wicht entschieden bei den Vertebraten liegt. Darauf sol1 wie auch auf ahnliche Publikationen neueren Datums in der Diskussion Bezug genommen werden, urn verschiedene Ergebnisse unter- einander und mit denen der vorliegenden Arbeit vergleichen zu konncn.
Die von ORNSTEIN und DAVIS entwickelte Disk-Elektrophorese (DAVIS 1962, 1964; ORN- STEIN 1962, 1964) brachte eine entscheidende Erhohung des Auflosungsvermogens, nachdem schon zuvor die Trennmedien Starkegel nach SMITHIES (1955) und RAYMONIX Polyacrylamidgel (RAYMOND and WEINTRAUB 1959) eine bedeutende Verbesserung in dieser Hinsicht gebracht hatten, indem sie zusatzlich zur unterschiedlichen elektrischen Ladung der Proteinmolekule deren verschiedene Grofle und Gestalt zur Trennung im elektrischen Feld ausniitzen (Molekular- siebeffekt). Zum einfachen Trenngel der Polyacrylamidgel-Elektrophorese tritt nun ein Sam- melgel mit grofleren Poren und anderem pH-Wert. Diese Diskontinuitat gab der Methodc wie auch die scheibchenformige Gestalt der aufgetrennten Fraktionen (englisch ,,disc") in den urspriinglich zylindrischen Gelen den Namen und fiihrt zu einer Konzentrierung der Sub- stanzlosung und einer Verengung der ins Trenngel wandernden Proteinzone. Ergibt die Papier- elektrophorese bei menschlichem Serum fiinf Fraktionen, so erhalt man disk-elektrophoretisch bis zu dreiflig. Die theoretischen Grundlagen der Methode sind bei ORNSTEIN (1964), ihre prak- tische Ausfuhrung ist bei DAVIS (1964) und einschlicfllich wesentlicher Varianten und Weiter- entwicklungen bei MAURER (1968) nachzulesen.
Wahrend die parasitischen Klassen der Platyhelminthes bereits in beschranktem Mafl elek- trophoretisch untersucht worden sind, findet man in der Literatur keine Hinweise auf entspre- chende Studien an Turbellarien. GEYER (1967) analysierte Totalextrakte von drei Trematoden- sowie je einer Cestoden- und Nematodenart mit verschiedenen Elektrophoresemethoden ein- schliefllich der Disk-Elektrophorese und konnte dabei sowohl Arten kennzeichnen als auch Hinweise fur die Verwendbarkeit immunelektrophoretischer Untersuchungen zur Klarung phylogenetischer Fragen gewinnen. YOSHIhlURA (1969a, b) gelang es, mit Hilfe der Disk-Elek- trophorese drei Arten der Trematoden-Gattung Paragonimus zu charakterisieren.
Aufgabe d e r vorliegenden Arbeit ist es, z u ermitteln, ob sich Ex t rak te paliducoler Tricladen disk-elektrophoretisch in Proteinfraktionen t rennen lassen, ob d ie so erhal- tenen Muster reproduzierbar u n d artcharakteristisch sind, welche Schwankungen dabei auftreten und ob sich die erhaltenen Spekt ren allenfalls neben den morphologischen Merkmalen verwenden lassen, u m Art- ode r Gattungsverwandtschaflen festzustellen.
Dazu mussen mindestens zwei Gattungen einer Familie, in mindestens einer Gattung wenig- stens zwei Arten und bei zumindest einer Art zwei oder mehrere Populationen untersucht werden. Griff die Wahl des Materials auch uber diese Minimalforderungen hinaus, so konnen freilich aus den Resultaten schon angesichts der nicht sehr groflen Individuenzahlen keine end- gultigen Schluflfolgerungen erwartet werden. Wie in der Diskussion erortert wird, 1aRt sich aus den bisher erzielten Ergebnissen an anderen Tiergruppen nicht vorhersagen, mit welchem Erfolg eine bestimmte Elektrophoresemethode bei einer taxonomischen Gruppe angewendet werden kann. D a zudem die Zahl der untersuchten Arten, von den Vertebraten abgesehen, gering ist und vor allem vergleichende Studien an mehreren Arten nicht haufig sind, erschien es lohnend, die Beantwortung der oben erwahnten Fragestellungen zu versuchen.
11. Material
Die untersuchten Tiere gehorten 9 Arten der beiden Familien Planariidae (4 Genera) und Dendrocoelidae (3 Genera) an und entstammten Biotopen in Usterreich (6 Arten), im englischen Lake District (3 Arten) und in Japan (3 Arten). Die folgende Ubersicht gibt die Fundorte an (vgl. dazu Abb. 1 bis 3).

Disk-Elektrophoretische Untersuchungen zur Artcharakterisierung paludicoler Tricladen 265
Abb. 1. Ubersichtskarte und Kartenskizzen zu den Fundorten in Usterreich. 1 : Schreiber Bach, 2 : Hainbach, 3: Primelquelle im Wassergspreng, 4: Wassergsprengbach, 5: Laxenburg - Graben im Schloflpark, 6 : Quellbach am Ostufer des Gleinker Sees, 7: Quellaustritt des Notscher Baches,
8 : Lunz am See - Biologische Station und Urngebung

266 R. Peter
200 km - Abb. 2 . Kartenskizzen zu den Fundorten in England. 1 : Out Gate, 2: Ees Bridge, 3: The Ferry
House
Planariidae
Dugesia gonocephala (Dugbs 1830) : drei Bache im Wienerwald; a. Wassergsprengbach bei Mod- ling (GieShubl), Niederosterreich. b. Hainbach, Wien, 14. Bezirk (an der westlichen Peripherie). c. Schreiber Bach bei Grinzing, Wien, 19. Bezirk (an der nordlichen Peripherie). Die Fund- orte sind durch Wasserscheiden voneinander getrennt, die Entfernung zwischen a) und b) be- tragt ca. 16 km.
Dugesia tigrina (Girard 1850) : eingeschleppte nordamerikanische Art pus zwei Warmwasser- aquarien in Wien; es kann nichts daruber ausgesagt werden, wie writ die beiden Populationen voneinander isoliert sind. Die Art ist aus Freilandbiotopen bei Wien bisher nicht bekannt.
Dugesia lugrbris (0. Schmidt 1861): a. Biologische Station Lunz am See, Niederosterreich: Wasserbcdten im Gewachshaus. MEIXNER (191 5, S. 575) beschreibt sie als ,Planaria polychroa". b. Ees Bridge bei Ambleside, Westmorland, England.
Crenobia alpina (Dana 1766): a. Population aus der Primelquelle bei GieShubl im Wasser- gspreng (Niederosterreich), isoliertes Vorkommen (vgl. KUEHN 1938). b. Population voni Quellaustritt des Notscher Baches, Villacher Alpe bei Villach, Karnten. c. Tiere von verschiede- nen Fundorten in der Umgebung von Lunz am See, Niederosterreich, wurden vergleichend zu den oben genannten Tieren untersucht, die Resultate konnten aber in diese Arbeit nur z. T. einbezogen werden.
Polycelis felina Dalyell 1814: a. Population aus einem Quellbach am Ostufer des Gleinkcr Secs bei Windischgarsten, Oberosterreich. b. Tiere aus dem Lake District bei Ambleside (West- morland, England), bei Out Gate und Ash Landing aufgesammelt.
Phagocata vivida Ijima et Kaburaki 1916: Quelle in Hirosaki City, Aomori-Prefecture (Aomori-Ken), HonshQ, Japan (s. KAWAKATSU, TESHIROGI und YAGIHASHI 1967).

Disk-Elektropboretiscbe Untersucbwngen zur Artcbarakterisierung paludicoler Tricladen 267
Dendrocoelidae
Dendrocoelum lacteum 0. F. M i i h 1773: a. Laxenburg bei Wien, Nicderosterreich, Graben (Arm der Schwcchat) im Schloflpark. b. Windermerc Lake unweit ,The Ferry House" bei Ambleside, Wcstmorland, England.
Dendrocoelopsis ezensis Ichikawa et Okugawa 1958 : Tomikawa-chB, Hidaka District, Hokkaid6, Japan.
Bdellocephala brunnea Ijima e t Kaburaki 1916: Quclle in Hirosaki City, Aomuri-Prefect- ure, Honshb, Japan (s. KAWAKATSU, TESHIROCI und YACIHASHI 1967).
Die europ2ischen Tiere wurden nach BOHMIG (1909) und REYNOLDSON (1967) bestimmt; die Bezeichnungen folgen den Angaben des zuletzt genannten Autors. Herr Prof. Dr. M. KAWAKATSU und Herr Dr. W. TESHIROCI iibernahmcn dankcns- werterwcisc die Beschaff ung und Determi- nation des japanischen Materials. Einen Obcrblick uber die japanischen Artcn und weitere Literaturangaben gibt KAWAKATSU (1969). Die untcrsuchten Dendrocoeliden pflanzen sich ausschliefllich sexuell fort (s. STEINMANN-BRESSLAU 1913, KAWA- KATSU, YAMADA und I W A K I 1967). Von den Planariidcn wicscn Dugesia tigrina, Polycelis felina aus England (REYNOLDSON 1967) ascxuelle, Phagocata wiwida sexu- elle und asexuclle (KAWAKATSU, YAMADA und IWAKI 1967), die iibrigen Arten nur sexuelle Fortpflanzung auf (vgl. dazu auch DAHM 1958, REYNOLDSON 1967). Die oberosterreichische Population von Poly- celis felina konnte nur iiber kurzc Zeit beobachtet werden, ihre Fortpflanzungs- a r t ist dem Verfasser daher lcidcr unbe- kannt.
Die teils zu verschiedcner Jahrcszeit aufgesammelten Tiere von unterschiedli- chcr Grofle wurden vor den Elektropho- rese-Experimenten im Kiihlschrank bei 4-8T mindcstens eine Woche lang ohne Fiitterung gchalten. Es waren dann mit Sicherheit keinc Nahrungseinflussc mehr festzustellcn; nach PICKAVANCE (1970) und nach REYNOLDSON und DAVIES (1970) ist mit dcr vie1 empfindlicheren immuno- logischen Mcthode nach langstcns 8 Tagen bei Dugesia tigrina bzw. Dendrocoelum lacteum dic aufgenommene Nahrung nicht mehr feststellbar. Ebensowenig konnten OSBORNE und MILLER (19621
Abb. 3. Kartenskizze zu den Fundorten in Japan. 1 : Hirosaki City, 2: Tomikawa
nach diesem Zeitraum bei Dugesia' t igrha verfiitterte Peroxidasc histochemisch nachweisen. Kannibalismus wurde nie beobachtet, so dafl es sich eriibrigtc, die Tiere langere Zeit vor den Experimenten zu isolicren. Die Groflenkennzeichnung erfolgte nach dem annahernden Frisch- gewicht. Altersangaben sind nicht moglich, weil nur Freilandtiere untersucht wurden.
111. Methoden
1. Disk-Elektrophorese
Moglichst rasche gleichzeitige Analyse mehrerer Individuen ohne zcitraubcnde Vorbehandlung der stets frisch hergestellten Extrakte einzelner Tiere war das Ziel methodischer Vorarbeiten,

268 R. Peter
die zu dem beschriebenen Verfahren fiihrten. Die Disk-Elektrophorese bot sich hierbei nicht nur wegen ihres hohen Auflosungsvermogens an, sondern auch deshalb, weil selbst stark ver- dunnte Proteinlosungen aufgetrennt werden konnen, ohne dai3 die Ergebnisse von deren Kon- zentration merklich beeinflufit werden.
Probenbereitung Zum Wiegen befanden sich die Tiere auf einem Plastikplattchen; Wasser war mit Filtrierpapier vorsihtig abgesaugt worden, ein Alpha-Strahler knapp uber der Waagschale der Sartorius- Analysenwaage verhinderte Fehlwiegen durch elektrostatische Aufladung. Die Gewichtsangaben auf Zehntel Milligramm sind freilich nicht sehr genau, weil die Wiegekammer keine definierte Lufifeuchtigkeit besai3 und von den Tieren standig Wasser verdunstete (daher mui3te auch sehr schnell gewogen werden), erwiesen s i h aber als ausreichend. Die Tiere wurden unter Eiskuh- lung bei 0-5 OC in Homogenisatoren aus Glas nach DOUNCE (1955, S. 102-103) homogenisiert, deren MaBe, der Kleinheit der Objekte ents rechend, im Vergleich zu den Originalangaben reduziert waren (Innendurchmesser der Zyl inLr 4,3-4,9 mm). Dabei wurde das kugelige Pistil1 30- bis 50mal in dem nur um wenige Zehntel Millimeter groi3eren Zylinder auf- und abbe- wegt, der auger dem Objekt eine dessen GroSe entsprechende Menge von Sammelgel-Puffer rnit 5O/oigem Saccharosezusatz (Tris-HC1 p H 6,7: 0,7475 g Tris, 6 ml 1 nHC1, 5 g Saccharose auf 100 ml; vgl. MAURER 1968) enthielt. Es wurde darauf geachtet, dai3 keine groaeren Gewe- bestiicke ubri blieben. Diese Methode schliei3t nach DOUNCE (1955) die Zellen ohne nennens- werten Angriff auf Kerne und Mitohondrien auf.
Die Puffermenge wurde in der Weise bestimmt, dai3 das Gewicht der Tiere in Milligrammen mit 19 multipliziert und die so erhaltene Zahl als Strecke in Millimetern auf jeweils einer Hamatokrit-Kapillare von etwa 1 mm2 innerem Querschnitt und 75 mm Lange aufgetragen wurde (Clay-Adams ,,Yankee" Micro-Hematocrit Tubes No. A-2931, I. D. 1,l-1,Z mm). 1 mg Frischsubstanz wurde somit in ca. 20 yl Gesamtvolumen homogenisiert. Mit Hilfe einer 2-ml- Injektionsspritze, die uber eine gebogene und abgeschnittene Nadel und ein passendes Stuckchen Silikonschlauch mit der Kapillare verbunden war (Abb. 4a), konnten die Puffervolumina be- quem pipettiert werden. D a zum Zentrifugieren und Auftragen der Proben jeweils dasselbe Rohrchen Verwendung fand und auch die Probenmengen nach Millimetern abgemessen wur- den, konnten sich Schwankungen im Kapillardurchmesser nicht auf die schliefllich verwendete Menge an Frischsubstanz auswirken und nur deren Konzentration leicht verandern. Im all- gemeinen wurden die Tiere einzeln, nur in einigen Fallen mehrere kleine Individuen zugleich homogenisiert. Die innerhalb etwa 5 Minuten hergestellten eisgekuhlten Homogenate fur ein Elektrophorese-Experiment wurden mit der erwahnten Injektionsspritze in die entsprechen- den Kapillaren aufgesaugt, diese beidseitig vorsichtig verschmolzen (dies dauerte insgesamt ca. 15-20 Minuten) und unter Kiihlung 20-25 Minuten lang in einer entsprechend adaptierten Laborzentrifuge bei 700-900 g (2300-2600 UpM) zentrifugiert. Die Homogenate waren her- nach in eine mehr oder weniger dunkle Sedimentschicht, einen opaleszierenden Uberstand und eine diinne Lipoidschicht dariiber getrennt. Mit einer Ampullenfeile wurden die Kapillaren in passende Stucke geteilt, der darin befindliche Uberstand gelangte auf die inzwischen vor- bereiteten Gele. Vom Homogenisieren bis zum Beginn der Elektrophorese verstrichen i. a. 60- 70 Minuten; ein genaues Zeitprotokoll erfai3te alle wesentlichen Arbeitsgange, so dai3 es mog- lich war, Unterschiede in den experimentellen Bedingungen und deren allfallige Auswirkungen auf die Ergebnisse zu iiberprufen (vgl. dazu den Abschnitt ,Untersuchungen zur Methodik").
Gelbereitung
Die angewandte Methode entsprach der von DAVIS (1964) angegebenen in der Durchfiihrung nach MAURER (1968, S. 39-50) mit den nachfolgend erwahnten Einzelheiten und Modifika- tionen. Die Fluka AG, Buchs (Schweiz), lieferte N,N,N,N-Tetramethyl-aethylendiamin (purum), Acrylamid (purum) und N,N-Methylen-bis-acrylamid (pract.); die beiden zuletzt genannten Substanzen wurden nach LOENING (1967) aus Chloroform bzw. Aceton umkristal- lisiert. Glycin (puriss. bzw. Erg. B. 6) und Tris-(hydroxymethy1)-aminomethan (puriss. p. a. bzw. p.a.) stammten von Fluka oder von Merck (Darmstadt), die ubrigen Reagenzien (mit dem jeweils hochsten Reinheitsgrad, i. a. zur Analyse) von Merck. Gelsystem Nr. 1 nach MAURER (1968, S. 42-43), welches ein 2,5O/oiges Sammelgel (pH 6,7) und ein 7,5O/oiges Trenngel (pH 8,9) urnfafit, wurde unter Verwendung des angegebenen Tris-HCI-Puffers und einer O,ZO/oigen Ammoniumperoxodisulfat-Losung statt der Riboflavin-Losung fur das Sammelgel hergestellt. Die Konzentrationen der Losungen fur das Trenngel wichen etwas von den angege- benen ab, fiihrten jedoch zur gleichen Endzusammensetzung. Alle Losungen wurden durch Sartorius Membranfilter Nr. SM 11306 (,MF 50", Porengrofie 0,6/0,45 pm) filtriert. Die che- misch katalysierte Polymerisation erwies sich als der photochemisch katalysierten gleichwertig

Disk-Elektrophoretische Untersuchungen zur Artcharakterisierung paludicoler Tricladen 269
Abb. 4. Gerate. a: Injektionsspritze mit Kapillare fur die Proben; b: Gelrohrchenhalter aus Plexiglas; c : ,,MikrometersDritze" aus einer Tuberkulinsprirze (1 ml), einem Stuck Plexirohr, zwei durchbohrtcn Korken und einer einfachen Schraubspindel mit Drehknopf; d-f: Betriebs-
bereite Anordnung aus Netzgerat (d), Elektrophoresezelle (e) und MeBgerat (f)

270 R. Peter
und war unter den gegebenen Laborbedingungen besser zu handhaben. Es war nicht erforder- lich, iiber der an der Grenze zur Saccharoselosung gebildeten Sammelgeloberflache eine zweite Schicht zu polymerisieren. Auf ein Substanzgel wurde verzichtet (,Alternative Procedure" nach DAVIS 1964); wie Versuche ergaben, hemmen die verwendeten Extrakte sowohl die chemisch als auch die photochemisch katalysierte Polymerisation.
75 mm lange numerierte Glasrohrchen rnit einem Innendurchmesser von 3,8 mm (gegen- seitige Abweichung maximal 0,2 mm) dienten als Gunformen fur die Gele; ihr Querschnitt ist um etwa 40 O/o kleiner als bei Rohrchen von 5 mm Durchmesser nach DAVIS (1964). Strich- marken im Abstand von 19,s und 36 mm vom unteren Ende bezeidmeten die Fullhohen fur die Saccharose- bzw. Sammelgel-Losung. Der verwendete Rohrchenhalter aus Plexielas
Substanzlosung Sammelgel
Trenngel
Gelrohrchen
10 c m
Halterung fur den Steckanschluss
Abb. 5. Manstabgerechter Querschnitt durch die betriebsbereite Elektrophoresetelle. Die Teile aus Plexiglas sind mit einem Polymerisationskleber zusammengefiigt. Die Elektroden aus Platindraht von 0,3 mm Starke sind an einem Ende in die Sei- tenwand der Zelle eingeklebt und gegeniiber mit einem Steck- anschlui3 verbunden, welcher in ein auf der Aunenwand der Zelle montiertes Plexiglasrohr eingeklebt ist. Die Mittelpunkte der zehn Bohrungen fur die Rohrchen sind 15 mm voneinander
entfernt, die Zelle ist innen 150 mm lang.
(Abb. 4b) bestand aus G h n etwa 40 mm langen, in ei- ner Reihe auf einer Leiste aufgeklebten Rohrstiicken von 10 mm Innendurch- messer. Passende 0-Ringe dichteten die Gelrohrchen am Boden ab und fixierten sie am Ende der Plexirohre, so dad sie genau normal zur Unterlage standen, die sich uber ein fixierbares Kugel- gelenk in die Horizontale einrichten lien. Kleine Boh- rungen in der Basisplatte unter den Rohrchen waren beim Polymerisieren mit ei- nem Selbstklebeband ver- schlossen, das vor dem Her- ausdrehen der Gelrohrchen abgezogen wurde, damit kein Sog die Adhasion der Gele am Glas storen konn- te. DAVIS (1964) beschreibt einen ahnlichen Gelstander, jedoch ohne Vorrichtung, die Rohrchen in lotrechter Lage zu fixieren und ohne Bohrungen in der Basis- platte. Zum Oberschichten mit Wasser diente eine ein- fache ,,Mikrometerspritze" (Abb. 4c). Das Einfiillen der Sammelgel-Losung nahm durchschnittlich 20 Minuten in Anspruch. Jeweils fiinf Rohrchen wurden knapp nacheinander mit einer Plq- stik-Injekrionsspritze mit lan er Nadel aus V-2-A- Stat1 mit Gellosung gefiillt und dann mit Wasser iiber- schichtet, wahrend ein Ma- gnetriihrer die restliche, in einem kleinen Becherglas be- findliche Losung am Poly- merisieren hinderte. Gerin- ge Unterschiede der Gele im Trennverhalten waren nicht mit der Reihenfolge der Sammelgelbereitung korre- lierbar. Das Trenngel wurde in einer mit Stickstoff ge- fiillten Kammer polymeri- siert (dam geniigte eine am

Disk-Elektrophoreliscbe Untersucbungen zur Artcharakterisierung paludicoler Tricladen 271
Arbeitstisch befestigte Dose aus Weichplastik mit dicht schlieflendem Deckel und absperrbarer Zu- und Ableitung); Polymerisieren an der Luff wie auch Abdecken mit Plastikfolien fuhrte zu ungleichmaniger Struktur am Gelende. Die fertigen Gele bestanden aus einer ca. 15 nim hohen Sanimelgelschicht uber durchschnittlich 39,5 mm Trenngel; die Schichtdicken schwankten um maximal etwa 1 mm.
Trennverfahren - apparative Ausriistung Ein mit kleinen Abanderungen nach ADMIRAAL (1953) selbst gebautes Netzgerat mit wihlbarer clcktronischer Strom- bzw. Spannungsstabilisierung (Abb. 4d) lieferte den notigen Gleich- strom. Die Spannung lien sich von 0 bis 320 V, die Stromstarke von 0 bis 150 mA stufenlos einstellen und war jeweils gegen Schwankungen der Netzspannung und des Verbraucher- widerstandes stabilisiert. In allen Experimenten wurde der Strom konstant gehalten. Als An- zeigeinstrument diente ein UniversalmeRgerat in niA-Schaltung (Normameter T) mit Spiegel- skala und einem maximalen Menfehler von 1 O / o der Gesamtskala von 25 mA. Der Strom blieb so gut konstant, daR man auf dieser Skala keine Schwankungen wahrnehmen konnte.
Die aus Plexiglas-Teilen selbst konstruierte Elektrophoresezelle ist in den Abb. 4e und 5 dargestellt. Sie nimmt zehn Rohrchen auf, deren Lange bis zu 150 mm betragen kann, was hier allerdings nicht voll ausgenutzt wurde. Die beschickte und mit vorgekuhlter Pufferlosung gefullte Zelle wurde in einem einfachen Haushaltkiihlschrank betrieben. Sie bewihrte sich recht gut; vor allem gestatten die lineare Anordnung und der geringe Abstand der Gelober- flfchen vom Rand, die Proben rasch in die bereits fertig vorbereitete Anordnung einzubringen und die Wanderung der Farbstoff-Front, wenn notig, leicht zu uberblicken. Die im Betricb gewonnenen Erfahrungen lassen jedoch zwei kleine Abanderungen wunschenswert erscheinen. (1) Die beiden Kammern sollten sich auseinandernehmen lassen, dies wurde die Reinigung erleichtern; dazu genugte es, die fix geklebte Kathodenkammer in die mit geringem Spiel pas- sende, etwas groRere Anodenkammer zu stellen. (2) Eine zweite Plexiglasleiste mit fur die Rohrchen passenden Bohrungen in einigem Abstand von den Dichtungen wurde die vertikale Orientierung der Gele erleichtern. Wird eine noch effektivere Kuhlung gewunscht, so konnte der Anodenpuffer, evtl. zusatzlich der Kathodenpuffer, uber einen Kryostaten zirkulieren.
Trennverfahren - Methode
Die dem Gelstander entnommenen Rohrchen wurden uber der Sammelgeloberflache grundlich mit Substanzpuffer, aui3en mit Elektrodenpuffer gespult und dann mit hangendem Puffer- tropfen am Ende unter Drehen vorsichtig in die Dichtungen der Elektrophoresezelle eingesetzt, deren Anodenkammer mit vorgekuhlter Pufferlosung (Tris-Glycin p H 8,3; 500ioige Stamm- Iosung nach MAURER 1968, S. 43) gefullt war. Aus den in 20 bis 40 mm lange Stucke geschnitte- nen eisgekuhlten Zentrifugenkapillaren gelangten die Proben (i. a. etwa 30 PI, entsprechend 1,5 mg Frischgewicht) mit der beschriebenen Injektionsspritze mit abgewinkelter Nadel auf die Geloberflache. Mindestens ein Rohrchen nahm als Standardprobe Humanserum in 1OOioiger Losung in Substanzpuffer, gef i rbt mit Bromphenolblau, auf. Die so beschickten Gele wurden vorsichtig mit Elektrodenpuffer uberschichtet und gleichmaRig tief in den Anodenpuffer ein- getaucht. Nachdem die Kathodenzelle langsam mit vorgekuhltem Puffer gefullt worden war, erfolgte die Elektrophorese fur 5 Minuten bei 0,5 mA und anschlienend fur 35 Minuten bei 2,5 mA pro Rohrchen. Die Farbstoff-Front der Standardprobe war hernach 5-8 mm vor dem Gelende angelangt. Die Puffertemperatur stieg bis zum Ende der Elektrophorese auf 10-18 OC. Anoden- und Kathodenpuffer gelangten aus der dem Kuhlschrank entnommenen Zelle in getrennte Aufbewahrungsflaschen und wurden bis zu funfmal verwendet. Mit einem Stuck Kunststoffschwamm lienen sich die Rohrchen leicht aus den Dichtungen drehen. U m die Gele aus den Rohrchen zu entfernen, wurde - etwas anders, als von MAURER (1968) beschrieben, - eine 0,8 mm dicke abgefeilte Injektionsnadel auf eine wassergefullte Spritze gestedct und unter Drehen und Kolbendrudi ein Stuck zwischen Trenngel und Glas geschoben; beim vorsichtigen Zuruckziehen der Spritze glitten die Gele in ein mit Wasser gefulltes Becherglas, aus dem sie sogleich zum Fixieren einzeln in ReagenzglIser gebracht wurden.
Fixieren und Farben Von den erprobten Verfahren (s. ,,Untersuchungen zur Methodik") erwies sich Fixieren in einer 2G"icigen wagrigen Losung von 5-Sulfosalicylsaure fur ca. 24 Stunden mit nachfolgender Farbung (cbenfalls etwa 24 Stunden) in einer 1°/oigen Losung von Amidoschwarz 10 B in einem Gemisch aus Wasser, Methanol und Eisessig (5:5:1 Volumsteile) als am besten geeignet. Zwi- schen Fixieren und Farben mufiten die kurz mit Wasser abgespulten Gele in einer zumindest

272 R. Peter
dreimal gewechselten Mischung aus Wasser, Methanol und Eisessig (10:4:1 Volumsteile) zur Entfernung der Sulfosalicylsaure wenigstens 3 Stunden lang belassen werden, damit sich nicht storende Farbstoff niederschlage bildeten. Die gleiche Mischung diente zum Auswaschen iiber- schiissigen Farbstoffs und mit 0,0001 O/o Farbstoffzusatz zum Aufbewahren der Gele. Es ist unbedingt zu vermeiden, die Gele Ianger als einen Tag zu fixieren, da sie sonst in zunehmen- dem Mad irreversibel schrumpfen. Analysenreine Merck-Reagenzien wurden verwendet. Die Farbe- und Waschlosung sind in einer Anleitung (Technical Bulletin No. 134) der E-C Appa- ratus Corporation, Philadelphia, angegeben (vgl. auch RAYMOND und WANC 1960).
Auswertung: Densitometrie
Pherogramme der gefarbten Gele wurden mit einem Vitatron Photometer, Model1 UFD 100 (optische Basiseinheit UFD 200 mit Quecksilber-Spektrallampe) mit Densitometereinheit (UFD 500) und integrierendem Sdireiber (UR loo), bei 578 nm aufgenommen. Das Prinzip des Densi- tometers ist von WIEME (1964) beschrieben worden. Gele aus den wenigen Experimenten mit abweichendem Serumstandard-Muster schieden von vornherein aus. Es kam der langsamste Probenvorschub (1,72 cm/min) in Verbindung mit dem raschesten Papiervorschub (15cmimin) zur Anwendung; der verwendete Spalt war 0,2 mm breit. Die Gele befanden sich in einer selbst geklebten Kiivette aus 1 mm starkem Plexiglas, die mit Aufbewahrungslosung gefiillt und mit einer Plexiglasplatte lufkblasenfrei abgedeckt war. Zwei Marken in der Mitte der Bodenplatte wurden mit der Mittellinie der Transportvorrichtung zur Deckung gebracht. Da das Immersi- onsmedium, welches durch ein Membranfilter (Sartorius Nr. SM 11306) filtriert worden war, annahernd gleichen Brechungsindex wie Polyacrylamidgel besitzt (vgl. MAURER 1968, S. 79), fielen optische Linseneffekte der Gelzylinder praktisch weg. Die Gele hatten in der Kiivette nur einige Zehntel Millimeter Spiel, so dad sie hinreichend gerade lagen, aber nicht durch Dehnung verzerrte Bilder lieferten. Die Extinktionsschwankungen der nur mit Immersionsfliissigkeit gefiillten Kiivette lagen unter jenen proteinfreier Gelbereiche. Schon kleinste Staubteilchen konnten sehr storen, waren aber zum Gliick am unvermittelt steilen Anstieg der verursachten Artefaktmaxima zu erkennen.
Durch eine zweifache Lupe gegen einen indirekt beleuchteten griinlichweiden Hintergrund betrachtet, lieden sich die Banden der in kleinen Glasrohrchen (Polystyrol wurde nach einiger Zeit angegriffen) aufbewahrten Gele sehr gut auflosen. Mit einiger Ubung gelang es leicht, die gefarbten Fraktionen den einzelnen Kurvenmaxima zuzuordnen und schwache, in den Kurven nur als kleine Zacken oder Stufen ausgewiesene Banden von Gelunreinheiten oder ungleichmadiger Farbung zu unterscheiden. In Abb. 6a ist ein Original-Pherogramm dargestellt. Eine Frontbande niedrigmolekularer Proteine, die mit der Grenzschicht, also ebenso rasch wie Bromphenolblau, wanderte und bei pigmentierten Arten durch eine schwach braunliche bis graue Farbung schon am unfixierten Gel leicht zu erkennen war, diente als besonders genaue und stets vorhandene Bezu sfraktion mit der relativen Beweglichkeit Ry = 1,000. Sie war auf allen Pherogrammen afs scharfes Maximum ausgepragt. Die Bandenpositionen wurden nach der Lage der Kurvenmaxima bestimmt. Diese Methode empfiehlt sich nicht nur wegen der starkeren Abszissendehnung gegeniiber der Messung auf Photographien der Gele, sondern auch deshalb, weil die Rp-Werte mit den Positionen der Maxima bis auf wenige Ausnahmen eindeutig anzugeben sind, was bei Photographien meist nur mit vie1 kleinerer Genauigkeit moglich ist. Die Gele mit einem Madstab direkt zu vermessen, wie das DAVIS und LINDSAY (1967) beschrieben, erwies sich schon wegen des kleineren Geldurchmessers als unpraktisch. Wahrend die Linsenwirkung der zylindrischen Aufbewahrungsgefade zusammen mit jener der Gele die Banden deutlich hervortreten ladt, konnen am frei liegenden Gel nur die starker ge- farbten mit einiger Miihe lokalisiert werden. Freil ih ist auch die Auswertung der densito- metrisch erhaltenen Pherogramme nicht ohne Probleme. Infolge der Lichtstreuung im Gel kommt es vor, dad schwache in der Nahe starker oder einander sehr nahe Banden nicht als echte Maxima, sondern nur als Stufen wiedergegeben werden. Sie wurden dann mit der Lage des Knicks angegeben. D a ihr echtes Maximum aber sehr wohl vom Anstieg des benachbarten Gipfels verdeckt sein kann, ergibt sich moglicherweise eine Abweichung von ihrer tatsachlichen Lage, die sich bei Vergleichen dann auswirkt, wenn die gleiche Bande auf anderen Pherogram- men als deutliches Maximum ausgepragt ist. Ferner konnen besonders flache oder sehr unruhige Maxima nur relativ ungenau lokalisiert werden; hier bringt aber weder die Photographie noch die direkte Vermessung bessere Ergebnisse. Selten kam es vor, dad eng benachbarte Doppel- oder Mehrfachbanden nicht aufgelost wurden.
Bei der Ermittlung der relativen Beweglichkeiten (im folgenden als RF-Werte bezeichnet) der einzelnen Proteinfraktionen nach den auf den Abszissen der Pherogramme angezeichneten Maxima ersetzte ein optisches Verfahren umfangreiche Rechenoperationen. Trotz sorgfaltiger Kontrolle der meisten experimentellen Bedingungen traten geringe Unterschiede in den abso- luten Wanderungsstrecken der Standardproben (Humanserum) und der Frontbanden von Gel

Disk-Elektrophoretische Untersuchungen zur Artcharakterisierung palrrdicoler Tricladen 273
zu Gel und auch zwischen verschiedenen Elektrophorese-Experimenten auf. Obwohl die Diffe- renzen auf den Gelen cines Elektrophorese-Experiments 2 mm nicht uberschritten, konnten die Strecken nicht direkt miteinander verglichen werden; deshalb wurde ein Diapositiv eines in 200 Teile gegliederten Manstabs mit einem Vergroflerungsapparat niit automatischer Scharfe- nachstellung so auf das Pherogramm projiziert, daI3 sich seine Gesamtlange mit der Distanz zwischen Star t und Frontbande (bzw. zur Genauigkeitssteigerung mit deren halber Lange) dccktc. Durch Interpolation zwischen den Maflstabmarken konnten die RF-Werte in tausendstcl Einheiten sofort unter den Maxima vernierkt werden.
Auswertung: Photographie
Zur Dokumentation wurden ausgewahlte Gele nach BURNS und POLLAK (1963) durch ein mit einem Gclbgrunfilter versehenes schwaches Tele-Objektiv (Schneider Tele-Xenar 1 :3,5 / f 135 mm) photographiert. Sie befanden sich in gebogenen, mit Aufbewahrungslosung gefull- ten Glasrohrchen von 26 cm Krummungsradius und ca. 4,5 mm Innendurchmesser. Der Krum- mungsmittelpunkt der Rohrchen lag in der Hauptebene des Objektivs. Ein Leuchrkasten mit Mattschcibe unter den Gclen diente als Lichtquelle. Von mehreren erprobten Aufnahmemate- rialien (Agfa Isopan IFF 15, I F 17, Agfa Copex, Ilford Pan F, Kodak Panatomic-X, Perutz- Positiv-Film) erwies sich der Film ,,Agfa Isopan IF 17" in Verbindung mit dem Entwickler ,,Neofin blau" (Tetenal-Photowerk, Hamburg-Berlin) und hartem Photopapier als optimal.
Vergleichsmethoden
Pherogramme von Parallelproben (mit dern gleichen Homogenat beschickten Gelen) konnten direkt miteinandcr verglichen werden, d a der Kurvenverlauf stets sehr ahnlich war . Aus den einzelnen RF-Werten wurde sodann jeweils das arithmetische Mittel berechnet. Es erschien wunschenswert, die so erhaltenen Mittelwerte mit Hilfe eines statistischen Verfahrens mit jenen anderer Proben zu verglcichen. Dies war leider nur zum Teil sinnvoll, d a vielfach schon wegen der Kleinheit der Tiere die Zahl der Parallelproben zu gering war . Jedoch lieflen sich inner- halb der einzelnen Populationen die R p W e r t e in Shnlicher Weise wie zwischen Parallelproben gegenseitig zuordnen. Sie lagen vielfach im Variationsbereich einer anderen Probe, was als Kriterium fur ihre gegenseitige Identitat galt. Wo dies nicht zutraf, durfken einander die Vari- ationsbereiche der RpMit te lwerte in einer Population (Stichprobe) nicht uberdecken, wenn die entsprechenden Einzclwerte als miteinander identisch gelten sollten. Bei Bdellocephala brun- nea und Dendrocoelopsis ezensis wurden die so gewonnencn Zuordnungen mittels des t-Tests (WEBER 1967) uberprufk und fur richtig befunden (Irrtumswahrscheinlichkeit a = 1 O/o bzw. p = 0,Ol). DAVIS und LINDSAY (1967) wenden ebenfalls den t-Test an, um die Identitat oder Verschiedenheit zweicr Banden zu ermitteln. Dcr Vergleich zwischen den Stichproben (Popu- lationen, Arten) erfolgte ausschliefllich nach dem erwahnten Test, weitere Einzelheiten sind zum besseren Verstindnis im Kapitel ,,Ergebnisse" beschrieben.
2. Polyacrylamid-Elektrophorese mit einer Gelschicht
Fur erste orientierende Untersuchungen und auch zu methodischen Vergleichszwecken diente 7,5O/aiges Polyacrylamidgel ohne Diskontinuitaten der Puffer- und Gelzusammensetzung ahn- lich dem von RAYMOND (1964) beschriebenen. Die nach einer Vorschrifk (Technical Bulletin No. 134) der EC-Apparatus Corporation, Philadelphia, bereitete Gel-Losung enthielt in 150 ml 10,6875 g Acrylamid, 0,5625 g N,N'-Methylen-bis-acrylamid, 0,lO ml N,N,N',N'-Tetramethyl- aethylendiamin und 0, l g Ammoniumperoxodisulfat. Der Dimer-Gehalt der Gele von 5 O/o der Monomerengesamtmenge entsprach der Zusammensetzung des angegebenen ,,Cyanogum 41" der American Cyanamid Company, New York (s. WHITE 1960). Als Gel- und Elcktroden- puffer diente Tris-EDTA-Borat-Puffer, p H 9,2, nach der erwahnten Vorschrift (40,O g Tris, 4,O g wasserfreies Dinatriumsalz der Aethylendiamintetraessigsaure und 1,52 g Borsaure auf 4 1). Alle Gerate und Arbeitsgange mit Ausnahme der nachfolgend beschriebenen waren mit jenen dcr Disk-Elektrophorese identisch. Das Verfahren wurde in Anlehnung a n RAYMOND (1964) und CLARKE (1964) entwickelt. Jedoch gofi RAYMOND Platten pus 5°/aigem Gel in einer Plexiglasform, wahrend CLARKE Rohrchen von 5 mm Innendurchmesser ebenfalls zum Gieflcn P/oiger Gele, aber rnit anderem Dimergehalt und Puffer, verwendet.
Ca. 60 mm lange Gele wurden in jeweils drei Rohrchen knapp nacheinander gegossen und mit Wasser uberschichtet, wahrend ein Magnetruhrer die ubrige Losung in Bewegung hielt, und innerhalb von 1-2 Stunden verwendet. Eine Vorelektrophorcse von 20 Minuten bei 2,2 mA

274 R. Peter
pro Rohrchen diente nach RAYMOND (1964) dam, die Diskontinuitaten zu vermindern und eine verzogerte Wanderung der Proteine zu Beginn der Elektrophorese zu vermeiden. Dann erst wurden die inzwischen vorbereiteten und zentrifugierten Homogenate (statt Substanzpuff er gelangte Elektrodenpuffer mit einem Zusatz von 5 O/o Saccharose zur Anwendung) unter der Pufferoberflache vorsichtig auf die Gele pipettiert. Dies war etwas schwieriger als bei der Disk- Elektrophorese, da die Kapillarstuckchen noch mit einer fein ausgezogenen kurzen Kapillare verbunden werden mudten. Ein Losungsvolumen von ca. 10 pl, entsprechend 0,5 mg Frischge- wicht, erwies sich als optimal. GroBere Mengen fiihrten zu einem merkbaren Schrumpfen der Geloberflache und dadurch zu unbrauchbaren Trennungen. Wiederum diente Humanserum als Standard. Nach 5 Minuten Elektrophorese bei 0,5 mA und 25 Minuten bei 2,2 mA pro Rohr- chen gelangten die Gele, wie beschrieben, in die Fixierlosung; ein kombiniertes Fixier- und Farbegemisch (80 mg Amidoschwarz 10B pro Liter 2O/oiger Essigsaure) nach WIEME (1962) reichte fur diese Methode aus. Die Verweildauer betrug mindestens 24 Stunden, doch hatten auch wesentlich langere Zeiten keinen Einflud auf das Ergebnis. Differenziert wurde in mehr- mals gewechselter 7O/oiger Essigsaure, die mit einem Zusatz von 0,0001 O/o Amidoschwarz 10 B auch zur Aufbewahrung in Polystyrolrohrchen diente. Die fur die Disk-Elektrophorese ver- wendete Fixiermethode lieferte bei den deutlich weniger transparenten Gelen keine besseren Ergebnisse. Alle weiteren Schritte waren jenen nach der Disk-Elektrophorese gleich. Als Im- mersionsmedium fur die Densitometrie fungierte jedoch 7O/oige Essigsaure, als Bezugsfraktion das Transferrin (die 1-0-Komponente nach CLARKE 1964) der entsprechenden Serumproben. da keine deutlich lokalisierbaren Frontbanden auftraten.
3. Zymogramme
Bei wenigen Gelen wurden Enzymnachweise versucht. Zum Nachweis der Lactat- bzw. Malat- Dehydrogenasen (LDH, MDH) diente die von SCHRAUWEN (1966) angegebene Methode unter Verwendung von Nitro BT (Tetrazolium-Nitroblau) und 5-Methyl-phenaziniummethylsulfat (Phenazinmethosulfat) der Firma Schuchardt, Miinchen. Die ubrigen Reagenzien stammten von Merck. Die Pherogramme wurden bei 578 nm aufgenommen; sie zeigten meist relativ flache Maxima. Die geringe Farbintensitat der Banden nach der Disk-Elektrophorese machte eine densitometrische Auswertung unmoglich, so dad die Gele mit einem Magstab vermessen werden mudten.
4. Untersuchungen zur Methodik
Vergleich von Disk- und Polyacrylarnidgel-Elektrophorese
Zur Trennung der vorliegenden Proteingemische erwies sich die Disk-Elektrophorese als ein- deutig uberlegen. Zwar sind die Unterschiede in der Zahl der auflosbaren Banden nicht schwer- wiegend : bei Dugesia gonocephala (Tiere aus dem Wassergsprengbach und dem Hainbach) lieaen sich disk-elektrophoretisch insgesamt 40, bei Crenobia alpina (Individuen aus der Pri- melquelle und aus dem Notscher Bach) 32 Fraktionen auftrennen; Polyacrylamidgel-Elektro- phorese fuhrte in beiden Fallen (die Exemplare von Crenobia alpina stammten aus der Um- gebung von Lunz am See) zu 30 Banden. Das Verfahren mit nur einer Gelshimt weist jedoch gegenuber der Disk-Elektrophorese einige Nachteile auf.
1. Die maximal zulassige Probenmenge betragt mit 0,5 mg Frischsubstanz nur rund ein Drit- tel der beim anderen Verfahren optimalen Menge. Die Geloberflache dellt sich dann nur schwach ein. Wird mehr aufgebracht, so schrumpft die Geloberflache so stark, dad sich fur die Proteine eine zusatzliche, ringformige Einwanderungszone ins Gel ergibt. Schlecht reprodu- zierbare und z. T. verwaschene Trennungen sind das Resultat. An Humanserum konnte ein derartiger Effekt nicht beobachtet werden. Dagegen t ra t er auch an den mit Tricladenhomo- genat uberschichteten Samrnelgelen auf; diese halten allerdings nach RITCHIE et al. (1966) auch Storungen von der Trenngeloberflache fern, so dad die Ergebnisse d a d u r h nicht beeintrachtigt werden.
2. Die grodere Lichtstreuung der Gele bedingt zusammen mit der geringeren Farbungsinten- sitat einen merkbar unruhigeren Kurvenverlauf mit wesentlich flacheren Maxima (entspre- chend diffuseren Banden) und daher auch geringerer Auflosung, als dies bei den Pherogrammen nach der Disk-Elektrophorese der Fall ist (Abb. 6).
3. Wahrend die ,,Disk-Gele" in der gefarbten Proteinfront uber eine innere Vergleichs- bande verfugen, fallt diese bei den einschichtigen Polyacrylamidgelen weg. Da sich von Gel zu Gel geringe Abweichungen in den Wanderungsstrecken der Fraktionen ergeben, sind die RF-Werte der Polyarylamidgele weniger genau und schwieriger miteinander zu vergleichen.

Disk-Elektrophoretische Untersuchungen zur A rtcharakterisierung paludicoler Tricladen 275
Abb. 6 . Elektropherogramme von Dugesia gonocephala aus dem Wassergsprengbach. Oben: nach Disk-Elektrophorese (Tier N. 3 : 3,O mg), unten: nach Polyacrylamid-Elektrophorese mit nur eincr Gelschicht (Tier von 7,5 mg). Die sinusformige Aufzeichnung unter der Null-Linie
gibt das jeweilige Kurvenintcgral an.

2 76 R. Peter
Dennoch konnte die Elektrophorese mit einfacher Polyacrylamidgel-Schicht fur den Nach- weis von Enzymen vorzuziehen sein. Die wenigen Ergebnisse weisen darauf hin, dad die Farb- intensitat hier starker ist als bei der Disk-Elektrophorese, bei welcher nach ORNSTEIN (1964, S . 346) der Dissoziationsgrad von Komplexen i. a. grofler sein durfte als bei anderen Methoden. Das konnte ein Grund fur die Inaktivierung vorhandener Enzyme sein. Die geringe Zahl der Zymogramme und vor allem die mangelnde Variation der experimentellen Bedingungen lassen diesbeziiglich aber keinen zwingenden Schlud zu.
Mischproben
Konnten zweierlei Proteingemische auf ein und demselben Gel der Elektrophorese unterwor- fen werden, dann sollten sich die beiden auf Grund der gegenuber den Einzelproben hinzu- tretenden Banden wesentlich genauer und sicherer vergleichen lassen, als das sonst der Fall ist. JOHNSON et al. (1967) geben eine solche Methode fur Getreidesamenalbumine an. Wie sich in einigen Versuchen herausstellte, ladt sich diese aus zwei Grunden fur die Planarienextrakte leider nicht verwenden. Einmal bedingt die Mischung einen allgemein erhohten Farbuntergrund und eine Abnahme der Farbintensitaten der einzelnen Fraktionen (die Gesamtmenge an Pro- tein darf ja ein Maximum nicht uberschreiten, das etwa 30-40 pl Homogenat entspricht). Zu- dem bedingt die relativ enge Bandenabfolge, dad zwei unterschiedliche Banden dann oft mit dem Densitometer gar nicht und selbst mit dem Auge nur schwierig zu trennen sind. Die Auf- trennung zweier Proben in einem Rohrchen (CLARKE 1964) war wegen des relativ geringen Rohrchendurchmessers fur grodere Serien nicht anwendbar.
Verschiedene Fixiermethoden Urn die optimalen Bedingungen zu ermitteln, wurden verschiedene Fixiermittel an disk-elek- trophoretischen Auftrennungen von Dugesia-lugubris-Extrakten erprobt. Eine 0,2O/oige Lo- sung von Ponceau S in 3O/oiger wagriger Trichloressigsaure farbte nur sehr schwach an. Zur Farbung diente sonst immer Amidoschwarz 10 B, teils gleichzeitig mit der Fixierung, teils nach- her. Fur eine statistisrhe Absicherung ist die Zahl der Experimente leider zu gering, doch durf- ten ihre Ergebnisse zumindest einige orientierende Informationen enthalten. Gefarbt wurde etwa ein bis drei Stunden, fixiert z. T. langer. Viele Gemische wurden auch heid verwendet (80-90 OC), es zeigte sich aber nirgends ein merklicher Unterschied zur kalten Fixierung, ausge- nommen die 200/oige wadrige Sulfosalicylsaurelosung, welche, heid angewandt, starke irrever- sible Gelschrumpfung zur Folge hatte.
Unbrauchbar waren: 1. Amidoschwarz-Losungen in Fixiergemischen mit uber 3 O/o 5-Sulfo- salicylsaure; es bildeten sich nicht mehr entfernbare Farbstoffniederschlf e. 2. 5O/oige wadrige Losungen von Phosphorwolframsaure mit bis zu 0,5 O/o Amidoschwarz (feine Farbung). Min- derwertige Ergebnisse lieferte Fixieren in gesattigter wadriger Sublimatlosung bzw. in 7O/oiger Essigsaure, jeweils mit nachfolgender Farbung (vgl. DAVIS 1964, S. 426).
Resultate steigender Qualitat ergaben sich nach gleichzeitigem Fixieren und Farben in Lo- sungen von Amidoschwarz in (1) 20loiger Essigsaure (0,008 O/o Farbstoff), (2) 3,5O/oigem For- mol, (3) 700/oigem Xthanol, (4) 7O/oiger Essigsaure, (5) einem Gemisch aus Methanol, Eisessig und Wasser (5:1:5 Volumsteile).
Noch etwas bessere Ergebnisse als mit der zuletzt genannten Methode lieden sich durch Fixieren der Gele in einer 200/oigen wHBrigen Losung von 5-Sulfosalicylsaure und nachfolgende Farbung in einer 1Oloigen Losung von Amidoschwarz in dem unter (5) genannten Gemisch erzielen. Die verwendeten Farbstoffkonzentrationen erstreckten sich z. T. von 0,Ol O/o bis 1 O/o;
es zeigte sich, dad lO/oige Losungen den verdunnteren eindeutig uberlegen sind, indem sie zu etwas starkerer und weniger diffuser Anfarbung fuhren.
Einflul der Probenbereitung auf die Ergebnisse
Es konnte keine Abhangigkeit der Trennergebnisse von der Verwendung irgendeines bestimm- ten Homogenisators unter sechs gleichartig konstruierten Geraten festgestellt werden. Nicht zentrifugierte Homogenate fuhrten im wesentlichen zu gleichen Ergebnissen wie zentrifugierte, was Abb. 22 an Bdellocephala brunnea illustriert. Unterladt man das Zentrifugieren, dann tritt allerdings etwas erhohte Untergrundfarbung auf. Fur Polyacrylamidgele ohne Sammelgel ist es schon deshalb erforderlich, die Proben zu zentrifugieren, weil sie vom Homogenisieren im Elektrodenpuffer her Luftblasen enthalten, welche sich nicht vollstandig beseitigen lassen und die Homogenate beim Aufbringen auf die Gele an die Pufferoberflache treiben.
Unterschiedliche Lagerzeiten der Tricladenextrakte von 60 bis 240 Minuten fuhrten selbst

Disk-Elektrophoretische Untersuchungen zur Artcharakterisierung paludicoler Tricladen 277
bei 20°C (80 Minuten) zu keinen Unterschieden in den Trennergebnissen, wie Abb. 9 a n Dugesia gonocephala zeigt. Diese Tatsache ist auch deshalb wichtig, weil ja zwischen Homoge- nisieren und Beginn der Elektrophorese etwa eine Stunde verstrich. D a die Homogenate wah- rend dieser Zeit eingekuhlt blieben, ist keine merkbare Verfalschung der Ergebnisse gegenuber knapp nach dem Homogenisieren aufgetrennten Probcn zu befiirchten.
Reproduzierbarkeit der Trennergebnisse Die Abb. 9, 21 und 22 geben einen Eindruck von der gegenseitigen Ubereinstimmung der Er- gebnisse disk-elektrophoretisch untersuchter Parallelproben (auf verschiedenen Gelen in ei- nem Elektrophorese-Experiment aufgetrennter Anteile des gleichen Homogenates). Die durch- schnittliche Streuung der RF-Werte um ihr Mittel betragt hierbei ebenso wie bei wiederholten densitometrischen Aufnahmen desselben Gels 0,004 Einheiten (d. s. rund 0,5 O/o des durch- schnittlichen RF-Wertes der jeweils am weitesten gewanderten Fraktion, ausgenommen die Frontbanden, oder ca. 1,3 o/o des RpWertes von menschlichem Transferrin). Die aus allen untersuchten Populationen berechnete gesamte durchschnittliche Streuung liegt mit 0,006 Ein- heiten (etwa 1,9 O/o des RF-Wertes von Transferrin) nur wenig dariiber, was die gute Repro- duzierbarkeit auch der in verschiedenen Elektrophorcse-Experimenten gewonnenen Resultate unterstreicht, deren Fehler sich somit in derselben Groflenordnung wie die vom Densitomctcr bedingten bewegen. - Die Polyacrylamid-Elektrophorese mit einfacher Gelschicht weist dem- gegeniiber mit durchschnittlich 0,036 Einheiten (d. s. 3,6 O/o des RF-Wertes von Transfcrrin oder 1,3 O/o des mittleren RF-Wertes der jeweils vordersten Bande) e twa die doppelte Gesamt- streuung auf.
Ursachen fur die beobachteten Schwankungen der RF-Werte konnten nicht ermittelt werden, doch diirflen die geringen Abweichungen der gesamten Trennstrecken innerhalb und zwischen Experimenten mit dazu beitragen (vgl. DAVIS und LINDSAY 1967). Die erwahnten Schwan- kungen lieden sich weder zu den leicht differierenden Trenngel-Langen noch zu den Positionen der betreffenden Rohrchen in der Elektrophoresezelle in Beziehung setzen.
5. Kritik der Methode
DAVIS und LINDSAY (1967) legen die Nachteile der Disk-Elcktrophorese dar und stellen fest, dad ihre Verwendung in der Taxonomie nur dann sinnvoll ist, wcnn sie nicht isoliert, sondern zusammen mit anderen Methoden gebraucht wird. Die a n Gastropoden gewonnenen Erfah- rungen der genannten Autoren konnten an der hier untcrsuchten Tiergruppe bestatigt werden. Vor allem wirkt sich die Diskrepanz zwischen der hohen Auflosung und der relativ dazu groflen (wenn auch, absolut betrachtet, geringen) Streuung der RF-Werte dahingehend aus, dad Zufalls- iibereinstimmungen in Wahrheit unterschiedlicher Fraktionen noch wahrscheinlicher werden, als dies von den theoretischen Voraussetzungen her der Fall ist. Dem Vergleich von Individuen stchen, wenn nicht in ausreichendem Made Parallelproben verfiigbar sind, die, wenngleich ge- ringen, Schwankungen im Trenneffekt und die optisch und durch die Untergrundfarbung be- dingten Auflosungsgrenzen des Densitometers und des menschlichen Auges im Wege. Noch bessere Normierung der experimentellen Bedingungen, eventuell durch eine weitgehend auto- matisierte Apparatur, konnte dem zuerst erwahnten Obelstand abhelfen. Ein Densitometer mit erhohter Auflosung wurde auch noch Unterschiede erfassen, die mit der verwendeten An- ordnung nicht darstellbar sind, ware aber nur dann von groderem Wert, wenn uber einen Refe- renzstrahl anderer Wellenlange der Streulichtanteil elektronisch wegkompensiert wurde (ORN- STEIN 1964) und sich somit eine Kontrolle der Pherogramme durch Vergleich mit den Gelen erubrigte.
Die Bereitung der Extrakte stellt eine zusatzliche Fehlerquelle dar. Hat ten z.war die ver- schiedenen Homogenisatoren und die zwischen Homogenisieren und Elektrophorese verstrei- chende Zeit keinen merklichen Einflufl auf die erhaltenen RF-Werte, so konnten Konzentra- tionsschwankungen der Proteinlosungen oder einzelner Komponenten und das nicht kontrol- lierte Ionenmilieu der Individuen doch iiber intermolekulare Wechselwirkungen und Dissozi- ationseffckte von Komplexen in den Resultaten zum Ausdruck kornmen. Diese Faktoren diirf- ten aber hier von untergeordneter Bedeutung sein, d a die Vergleiche ja stets zwischen Stichpro- bcn aus mehrercn Experimenten angestellt wurden. Fur Individuenvergleiche waren sie vor allem dann zu beachten, wenn die Mengen einzclner Fraktionen verglichen werden.
Abschlieflend sei noch betont, dafl die erzielten Resultate von den angewendeten experimcn- tellen Bedingungen, so vor allem vom Extraktionsverfahren und von der Gelzusammenset- zung, abhangen und die einzelnen Trennungen, d a sie unter weitgehend identischen Bedingun- gen erfolgten, zwar untereinander vergleichbar sind, aber keine Aussage dariiber gestatten,

278 R. Peter
Nr.
Tabelle 1 RF-Werte von Dugesia gonocephala (Tiere aus dem Wassergsprengbach)
1 2 3 4 5 6 7 8 9 10 11 12 13 15 16 17 18 19 20 21 22 23 24 25 26 28 29 30 32 33 34 35 36 37 38 39 40
007 025 039 054 073 080 09 1 105 105 117 129 156 168 194 215 231 246 267 278 306 339 358 393 408 434 459 486 515 577 608 641 685 737 782 816 886 949
S
I RyWerte
N.l 2 3 4 5 6 7 8
t 1 3 1 3 1 1 1 1 H% 1 mg 1,0 2,5 3,O 10,O 14,3 15,7 26,2 32,3
I n 1 1 1 2 1 2 1 1 - _ _ 006 3 100 011 004 008 005 005 003 009 006
039 2 100 037 040 040 041 039 038 043 038 055 4 100 049 052 055 060 056 052 + 058 073 4 75 070 + - - 066 073 073 070 080 5 100 083 075 078 081 076 + 085 076
091 - - - + 091 1 25 104 7 100 113 097 101 111 099 101 110 103
- - +
023 5 63 - 027 - 022 + + - +
- - -
- - 105 - - 105 - 25 122 - 13 - - - - - - 122 - 131 6 100 135 127 124 131 124 140 131 128 158 2 75 - 157 - 156 159 159 160 + 168 5 100 168 164 173 160 + 174 + 166 194 10 50 188 205 - - - 189 + 216 5 100 219 215 213 220 210 212 221 218 236 3 25 248 8 100 250 252 236 250 241 247 258 241 267 3 13 276 2 13 307 3 100 310 307 306 307 302 308 310 306 334 6 88 333 335 331 339 328 328 (v) 343
392 5 100 394 394 390 394 393 386 396 400 405 3 50 - 407 404 407 - 403 -
434 5 88 443 - 433 440 433 429 430 432 455 5 75 462 459 459 453 446 456 - - 486 7 50 - - - 481 - 482 496 488 523 13 75 510 - - 516 545 + 523 515 573 12 75 - - 571 572 593 567 564 575 608 5 38 643 5 88 651 - 644 641 637 643 636 648 683 3 100 683 685 688 678 683 685 683 684
- 729 732 730 728 - 746 732 7 63 -
- - 771 785 789 779 10 38 820 9 38 - - 827 - 827 813 - - 886 5 38 - - 889 - 889 882 - - 949 4 25
-
- 237 233 - - - - -
- - 267 - - 276 -
- - - - - - - - -
- 358 + - - - - - 358 - 25
-
612 - 602 609 - - - -
- - -
- 946 950 - - - - -
Erliiwterungen: Nr. = Nummer der Fraktion (es ist jeweils die Reihe der bei allen unter- suchten Tieren bzw. Populationen der Art gefundenen RF-Mittelwerte durchnumeriert, so da8 hier nicht alle Nummern erscheinen). Alle RF-Werte sind in Tausendsteln angegeben und auf die Frontbande mit RF = 1,000 bezogen. GT = Mittelwerte aus allen Einzelwerten (unter AusschluB des einen Exemplars aus dem Schreiber Bah) ; R T = Mittelwerte zu den in dieser Tabelle angegebenen RF-Werten; s = Streuung (Standardabweichung) in Tausend- steln; HO/o = Haufigkeit der entsprechenden Fraktion in Prozenten, bezogen auf die Zahl der untersuchten Individuen. N. = Nummer des Tieres; mg = annaherndes Frischgewicht in mg; t = Haltungszeit ohne Nahrungsaufnahme in Wochen vor dem Experiment (mindestens so lange hat das betreff ende Tier also gehungert); n = Zahl der Parallelproben (der mit dem gleichen Homogenat im selben Elektrophorese-Experiment beshickten Gele) ; + = Fraktion vorhanden, aber den- sitometrisch nicht aufgezeichnet; - = Fraktion nicht vorhanden; v = Bande grenzt an starke Nachbarfraktion, so dai3 sie im Pherogramm nicht aufscheint; (v) = Bande konnte, auch wenn sie vorhanden ware, n i h t wahrgenommen werden, da die benachbarte Bande sie verdecken wiirde. Die in Tab. 1 aufgezahlten Tiere wurden im April gesammelt.

Disk-Elektrophoretische Untersuchungen zur A rtcharakterisierung paludicoler Tricladen 279
wic wcit die erhaltenen Fraktionen gegenuber ihrem Zustand irn lebenden Organismus dena- turicrt sind. Auch kann durch Dissoziation von Kornplexen eine Molekulspecies in rnehreren Banden auftreten (ORNSTEIN 1964).
IV. Ergebnisse
1. Disk-Elektrophorese
Die fertigen Gele zeigten 25 bis 40 densitometrisch trennbare Banden (Proteinfrak- tionen) pro Art, welche meist wie bei den von NAGAI et al. (1964) untersuchten Kol- lagenkomponenten als Ringe auftraten. Dieses Phanomen lie13 sich hier ebensowenig wie von den genannten Autoren erklaren oder beeinflussen. Aus den vorhandenen Daten konnte keine Abhangigkeit des Musters von der Gro13e oder dem Ernahrungs- zustand der Tiere abgeleitet werden. Es fie1 jedoch auf, da13 bei fast allen Arten am Be- ginn der Pherogramme bei den schwersten Tieren besonders hohe Maxima auftraten.
Tabelle 2
RF-Werte von Dugesia gonocephala (Tiere aus dern Hainbach)
RF-Werte I N ~ . ~ & ~ Q I 1 ~ 1 . i ~ . 1 2 3 4 5 6 7 mg 1,8 4,5 5,7 5,8 6,O 37,O 40,5 n 1 2 1 2 1 1 1
1 2 3 4 5 7 8
10 11 12 14 16 17 18 20 21 22 24 25 27 28 30 31 32 34 35 36 37 38
007 025 039 054 073 09 1 105 117 129 156 178 215 231 246 278 306 339 393 408 447 459 515 555 577 64 1 685 737 782 816
008 026 039 054 075 09 1 105 115 127 154 178 214 224 244 279 304 347 395 411 447 465 508 555 583 638 688 743 783 802
3 5 3 5 5 3 4 2 5 5 4 5 6 5
11 4
16 6 8
11 9
12 11 5 6 9
20 7 -
100 86 71 86
100 100 100 71
100 100 100 100 29 86 71
100 57
100 86
100 100 86 71 71 86
100 86 86 14
010 006 026 029 - 041
046 059 073 080 088 + 101 109 115 - 130 131 160 158 175 176 208 223
238 249 280 276 321 302 372 348 391 397 404 414 448 436 + 455 - 518
563 + - 586
628 636 678 680 + 726 + 780
- -
- -
01 1 023 - -
065 093 + 116 131 155 188 213
239 285 306
400 42 1 464 471 488
-
-
- - -
702 753 780 -
005 012
038 041 055 055 075 079 + + 105 109 - + 123 128 148 155 179 176 210 214 220 - 247 239 263 - 300 298
401 391 - 400
453 434 472 473 507 516 + - 578 585 636 639 696 684 721 750 783 783
029 -
332 -
802 -
008 027 038 052 075 + 103 113 122 154 177 213 228 249 293 305
395 419 458 464 510 555 584 644 689
-
~
- -
005 020 035 050 072 + 101 + 120 150 176 210 - - -
304 348 388 408 446 + + 547 585 648 686 766 790 -
Erliiuterungen: wic fur Tab. 1; Haltungszeit ohne Nahrungsaufnahme (t): 2 Wochen. Die Tiere wurden irn April gcsamrnelt.

Tabe
lle 3
R
F-W
erte
von D
uges
ia t
igri
na (
Tie
re a
us z
wei
Wie
ner W
arrn
was
sera
quar
ien)
Nr.
Aqu
ariu
m a
. A
quar
ium
b.
-
RF
-WC
~C
RF-
Wer
re
-
RF
N.l
2 3
RFt
I
H*,
o N
. 1
2
3 4
5 6
7 8
mg
1 1,
0 1,s
mg
1,4
1,9
2,O
2,O
2,l
2,2
2,2
2,9
n2
11
n
21
11
12
11
~ s
1 Helo 1 00
5 -
003
010
008
1 00
5 00
5 1
100
006
004
005
005
2 75
00
4 00
6 -
2 02
1 01
9 -
33
019
-
-
022
3 50
02
3 -
022
023
-
020
-
031
-
3 03
1 03
1 -
13
4 03
9 03
7 4
100
040
036
032
040
3 10
0 04
2 03
7 03
7 03
9 03
3 04
3 04
0 04
0 5
057
050
3 10
0 05
1 05
2 04
8 05
9 6
75
056
053
059
055
-
065
-
+ 6
070
070
5 10
0 06
6 07
6 07
0 7
079
083
-
33
083
-
- 0
78
5 10
0 07
9 07
2 07
8 07
4 07
0 08
4 08
5 07
7 8
096
094
0 67
-
094
094
096
5 10
0 09
6 08
8 09
4 09
8 09
9 +
100
101
9 13
0 12
5 3
100
123
127
126
132
5 88
13
6 12
8 -
139
126
132
130
133
10
152
152
7 10
0 14
7 16
0 15
5 11
16
4 16
4 5
88
159
173
161
164
-
163
163
168
12
183
185
10
100
183
195
175
182
4 50
-
186
-
-
178
185
-
177
13
206
201
3 10
0 20
1 20
4 19
7 20
9 6
88
204
215
205
203
208
-
217
215
-
-
-
- -
-
-
-
216
-
-
-
-
14
215
215
4 25
21
4 -
15
235
228
-
33
228
-
- 2
39
7 63
-
-
238
229
243
245
+ -
16
257
250
1 67
25
0 25
0 -
260
6 38
-
-
-
-
259
265
-
253
17
283
281
6 10
0 27
7 28
8 28
1 28
4 7
100
+ 27
0 28
2 28
9 +
289
289
280
18
295
295
4 75
29
4 29
0 30
2 29
5 -
-
295
293
19
313
307
10
100
309
316
296
315
7 75
32
3 31
3 -
-
305
315
+ 31
3 20
35
8 35
3 7
100
347
362
357
359
5 10
0 36
2 35
5 35
3 35
0 36
5 36
2 36
3 36
0 21
37
0
23
444
440
7 10
0 44
1 44
5 43
0 44
5 7
100
450
443
442
444
437
439
452
455
24
485
485
4 10
0 48
4 49
0 48
1 25
49
2 49
2 3
100
493
494
492
485
491
492
493
496
26
536
536
-
33
-
-
536
535
- 13
53
5 -
27
578
578
4 67
58
0 57
5 -
28
629
626
3 10
0 62
5 62
4 62
9 63
0 9
75
629
623
636
-
618
-
643
629
29
668
669
4 10
0 67
1 66
9 66
4 66
8 5
100
671
672
670
669
666
670
671
680
30
792
811
5 33
81
1 -
-
788
13
88
788
795
814
-
770
784
780
792
Erl
iigte
rung
en:
wie
fur
Tab
. 1.
Hal
tung
sdau
er o
hne
Nah
rung
sauf
nahr
ne
(c):
1 W
oche
. Fu
r di
e fo
lgen
den
Hor
noge
nate
wur
den
jew
eils
rne
hrer
e an
- na
hern
d gl
eich
gro
de T
iere
ver
wen
det:
a) N
.l (
14 I
ndiv
idue
n),
N.2
(5)
; b)
N.l
(4),
N.2
(3)
, N
.6 (
3);
es i
st s
tets
das
Dur
chsc
hnitt
sgew
icht
e
ke
s
Indi
vidu
ums
ange
gebe
n.
-
370
+ 37
0 -
38
-
-
-
+-
22
39
5 40
2 7
100
398
400
410
390
12
75
393
+ 40
7 39
1 -
378
379
-
-
-
-
-
-
-

Disk-Elektrophoretische Untersuchungen zur Artcharakterisierung paludicoler Tricladen 28 1
Dies lafit an einen Zusammenhang mit dem Reifezustand denken. Von Dendrocoel- opsis lactea Ichikawa et Okugawa 1958 (diese Art wird wegen der zu geringen Pro- benzahl nur in diesem Zusammenhang erwahnt) konnte ein Kokon analysiert werden; seine Hauptfraktion war mit jener der groi3ten Individuen identisch. Das Material ist aber zu wenig umfangreich, um aus den erwahnten Beobachtungen giiltige Schliisse zuzulassen. Die Gestalt der densitometrisch erhaltenen Kurven erwies sich als variabel, indcni Fraktionen gleicher Ry-Werte bei derselben Art unterschiedlich (als deutliche Maxima oder als Stufen) ausgepragt waren und auch verschiedene Extinktionswerte aufwiesen. Der Kurvenverlauf lie8 sich nur in beschranktem Mafi benutzen, um Arten zu charakterisieren. Darauf wird im folgenden jeweils hingewiesen. Von einer quanti- tativen Auswertung, die den Anteil der Flachen unter den einzelnen Maxima an der gcsaniten Kurvenflache ermittelt, wurde wegen der zu geringen Zahl an Tieren und Parallelproben abgesehen. Da nicht bei jedem Gel jede Fraktion aufgetrennt und auf dem zugchorigen Pherogramm lokalisierbar ist, ergibt sich auaerdem die Frage, ob ein erhohter Flachenanteil einer Bande einer tatsachlich groReren Menge der betreff en- den Fraktion entspricht oder aber durch mangelnde Trennung bedingt ist. Bei der rela- tiv grofien Zahl mitunter eng gestaffelter Banden konnen daraus ziemliche Ungenauig- keiten resultieren. Somit ware aufier einer groaeren Materialmenge eine noch bessere
Tabelle 4
RF-Werte von Dugesia lugubris: Population aus Lunz am See (RT) sowie ein Tier von Ees Bridge bei Ambleside (RF.))
1 005 - 2 012 010 3 022 018 4 042 (v) 5 057 050 6 085 087 7 106 105 8 120 119 9 149 154
10 166 + 11 183 178 12 220 222 13 236 -
14 254 250 15 291 298 16 326 334 17 363 370 18 403 418 19 464 454 20 493 ~
21 543 549 22 598 610 23 626 -
24 656 660 25 723 724
RF-Werte
N . l 2 3 4 5 6 7 8 9 1 0 1 1 mg2,4 2,4 3,O 3,3 3,4 3,5 3,5 3,7 6,2 11,l 12,5 M VIII IV I1 I1 IV VIIIVIIIIV IV I1 IV t 3 6 2 1 0 9 2 3 4 3 4 1 2 9 1 n 1 1 2 1 1 1 2 1 2 2 1
H%
005 2 91 013 3 82 024 5 91 042 4 82 058 4 100 085 4 91 106 2 91 120 7 73 149 4 91 166 4 64 184 4 55 219 4 91 236 4 36 254 7 82 290 5 100 325 7 100 362 7 91 402 6 100 464 9 91 493 14 73 541 7 18 596 18 82 626 19 55 656 7 73 723 12 100
~~
003 005 003 010 007 003 005 003 005 006 -
022 020 + + 023 - 033 025 021 v v 042 045 046 - (v) 048 042 040 035 043 v
084 086 087 085 085 085 084 080 - 081 v 110 106 v - 106 107 106 102 106 108 102
- 155 147 155 151 152 145 147 149 146 148
- 187 - 178 183 + ~ 185 189
+ + 016 + - 011 - v 012 ~-
056 055 058 058 055 060 057 050 058 + 065
(v) (v) 110 113 (v) 127 123 125 v 125 126
165 - 161 173 - 164 167 170 ~ 166 - - - + 219 221 223 222 220 222 214 217 216 -
259 + 263 259 255 245 252 256 247 - - - 235 - 234 234 2 4 0 - - - - -
287 286 295 296 293 289 290 287 290 288 280 + 328 327 332 325 323 328 323 323 330 306
404 400 407 411 407 403 406 402 397 397 394 358 362 364 - 365 358 369 355 366 365 343
456 447 468 473 470 461 460 - 473 461 464 493 484 504 489 490 - 492 496 - 491 -
- 545 - 5 3 3 - - - - - - -
596 - 598 570 600 - 557 591 603 609 566 - 612 - 640 - 628 622 - - 640 616
661 645 651 659 665 651 661 652 - - 656 719 723 726 723 728 713 749 716 733 706 718
Erluuterungen: wie fur Tab. 1 ; M = Monar der Aufsamnilung

Tabe
lle 5
RF-
Wer
te von C
reno
bia
alpi
na
Popu
latio
n au
s de
r P
rim
elqu
elle
~ass
ergs
pren
g Po
pula
tion
aus
dem
Not
sher
Bar
h
RF-
Wer
te
--I I
I I
RF-
Wer
te
h,
00
N
-
Nr.
N.
l
2 3
4 5
6 7
N.
l
2 3
1 RF 1
1 1 ?I
%
1 mg 1,
7 1,
7 1,
8 2,
5 3,
5 5,
l 10
,5
1 1 '
I *% I mg
0,9
2,O
2,4
t1
31
11
31
t
31
31
31
1 01
0 2
043
3 06
5 4
081
5 08
7 6
102
7 11
0 8
124
9 13
6 10
17
0 11
17
5 12
19
1 13
20
9 14
22
4 15
24
6 16
27
0 17
30
0 18
31
9 19
34
5 20
36
8 21
40
8 22
43
7 23
46
2 24
52
3 25
59
4 26
60
7 27
63
9 28
66
1 29
70
3 30
73
3 31
77
9 32
83
2 E
rlau
teru
ngen
:
009
3 04
2 7
066
3
081
3
100 86
100 86
71
86
86
86
100 86
71
14
100
100 71
86
86
71
86
007
029
071
083
101
110
124
138
180
185 -
-
245
263
319
348
367
395
439
488
523
600 -
005
040
064
0 79
007
035
066
078
007
061
078
101
126
144
174
193
217
245
273
311
325
342
360
402
436
459
527
595 -
-
-
010
041
069
083
010
052
070 -
012
046
065
082
011
046
061
087
103
126
137
171
175
199
208
1 10
0 01
1 04
8 05
9
085
102
126
137
169
176
20 1
208
013
010
047
043
063
-
086
089
103
-
121
130
137
-
+ 17
2
198
197
208
208
174
-
3 10
0 3
67
2 10
0 1
67
5 10
0 1
67
2 10
0 1
67
2 10
0 0
100
2 10
0 3
100
2 10
0
7 10
0
7 10
0
3 10
0 3
100
12
67
4 67
101
2 11
0 4
+ 10
2 10
7 11
5 13
3 16
8
102
106
138
166
187
209
243
271
294
318
341
354
399
430
525
600
641
665
707
787
829 -
-
-
-
-
105
128
172
198
-
-
-
240
266
317
342
388
433
458
517
578
-
-
-
113
135
170
183
210
247
270
310
344
39 1
428
520
582 -
-
-
-
-
ii3
12
0 12
3 5
136
5 17
0 6
188
5 20
9 5
224
2 24
5 3
269
4 29
7 11
31
9 5
343
4 36
8 15
40
6 14
43
7 12
46
8 14
52
2 4
594
8
639
9 66
1 9
703
4 73
3 6
831
9 w
ie f
ur T
ab. 1
787
-
128
171
186
213
245
270 + 321
346
384
423
447
468
523
597
-
-
204
224
247
265
290
316 -
-
425
453
518
592
628
-
-
249
275
308
350
414
454
525
591
607
249
278
306
346
406
452
526
599 -
249
248
273
+ 30
7 31
0
346
358
420
417
453
458
527
521
582
-
610
604
100 57
10
0 10
0 29
43
-
661
703
738
835 -
-
646
698 -
-
820
-
699 -
-
841
100 29
14
57
701
729
705
771
834
4 10
0 33
16
100
-
700
708
706
771
-
823
845
- +

Disk-Elektrophoretische Untersuchungen zur Artcharakterisierung paludicoler Tricladen 283
s
Reproduzierbarkeit der Ergebnisse als Voraussetzung fur quantitative Vergleiche zu forderii (vgl. auch Kapitel 111.5.). Die Gesamtflachen der Pherogramme, welche ein ungefahres Ma6 der aufgetrennten Proteinmengen darstellen, schwankten, konnten aber nicht mit der Gro6e oder dem Ernahrungszustand der Tiere korreliert werden.
HVo
~~
Vergleichsmethode
Der Vergleich zwischen zwei Stichproben erfolgte stets nach der Anzahl von Banden mit identischem Rv-Wert. Identitat wurde immer dann angenommen, wenn der t-Test (Irrtumswahrscheinlichkeit a = 1 O / o ) keine signifikante Differenz ergab. Die Ober- einstimmung wird in Prozenten der zwei Stichproben gemeinsamen Zahl von Banden, gemessen an der Gesamtzahl unterscheidbarer Fraktionen, angegeben und im folgenden mit Ahnlichkeit ( O / o ) bezeichnet. Sie wurde nach der Formel
C 100 (VO) -- -.
a + b - c berechnet, wobei a und b die Bandenzahl jeweils einer Stichprobe und c die Zahl der Paare miteinander identischer Banden bezeichnen. (Diese wird im Nenner subtrahiert, weil auf das gemeinsame Gesamtmuster und nicht auf die Summe der Einzelmuster
Tabelk 6
RF-Werte von Polycelis felina (Population aus einem Quellbach am Gleinker See)
1 007 006 2 018 018 3 035 035 5 047 047 6 066 064 8 086 086 9 091 093
10 118 118 11 133 133 12 167 172 14 196 194 15 223 223 17 260 260 18 279 276 19 307 308 22 386 386 23 407 404 24 420 420 26 502 500 27 538 538 29 617 620 30 671 673 31 693 695 32 769 763 33 836 827
RF-Werte
N . l 2 3 4 5 6 7 8 mg 1,6 1,7 2,2 2,7 3,O 3,3 3,4 3,8 t 1 1 3 1 3 1 1 1 1 n 1 3 1 1 1 1 1 1
- 003 005 007 - 007 011 - - 016 - 018 023 -
- 038 - 053 048 050 042 040 - -
067 063 057 070 + 068 068 058 088 087 083 - - 088 081 085 - - 091 093 097 - - 093
111 110 115 113 121 - 138 123 129 126 131 149 131 127 148 - 180 169 175 - + - 165 - 198 194 183 196 193 200 193 193 226 224 213 227 225 220 221 223 268 257 259 251 248 259 268 270 279 276 270 270 280 - - 283 - 300 - - 307 - 328 314
395 386 393 + 362 393 - -
428 419 420 + - 416 416 -
- - - - 038 035 030 -
- - - - 402 + 407 -
503 503 495 501 503 497 495 498 545 529 529 548 538 541 540 + 603 623 643 610 617 610 631 + 669 677 668 673 668 676 666 672 + 694 - + 696 - $ 7 6 3 - + + - - -
- 827 - - + - - -
- -
Erluuterungen: wie fur Tab. 1. Die Tiere wurden im August gesammelt. Fur das Homogenat N.l wurden 2 Exemplare (1,2 bzw. 1,9 mg), fur Homogenat N.2 vier Individuen zu je etwa 1,7 mg verwendet.
284 R. Peter
- Nr. RF R T
bezogen wird). Eine ahnliche Formel wenden BEDFORD und REID (1969) an. Eine Frak- tion hatte gleiches Gewicht, unabhangig davon, wie oft sie auftrat; von einer Wertung wurde Abstand genommen, weil der nicht methodisch bedingte Anteil der interindivi- duellen Schwankungen unbekannt ist.
Rp-Werte
N. 1 2 3 4 5 6 mg 1,6 1,7 2,7 3,3 3,4 11,0 t 16 1 15 1 1 1 n 1 1 1 1 1 3
s H%
Intraspezifischer Vergleich
Die RF-Werte der bei den untersuchten Tieren aufgetrennten Fraktionen ergeben je- weils ein artcharakteristisches Muster. Dieses ist jedoch nicht allen Tieren einer Art ge- meinsam, sondern setzt sich aus einem bei allen Individuen und auf allen Gelen ver- tretenen Grundmuster und etlichen mehr oder weniger haufig hinzutretenden Banden zusammen. Verschiedene Populationen bzw. Stichproben aus geographisch unterscheid- baren Gebieten weisen hierbei statistisch signifikante Unterschiede auf. Die innerhalb derselben Stichprobe auftretenden Schwankungen durften zum GroBteil methodisch be- dingt sein, indem Doppelbanden nicht uberall getrennt und schwache Banden nur bei optimaler Gel- und Farbungsqualitat vom diff usen Untergrund unterscheidbar sind. Aul3erdem kann eine sehr starke Fraktion die schwache Nachbarbande entweder vollig uberdecken oder aber fur das Densitometer unsichtbar machen. Die mit freiem Auge
Tabelle 7
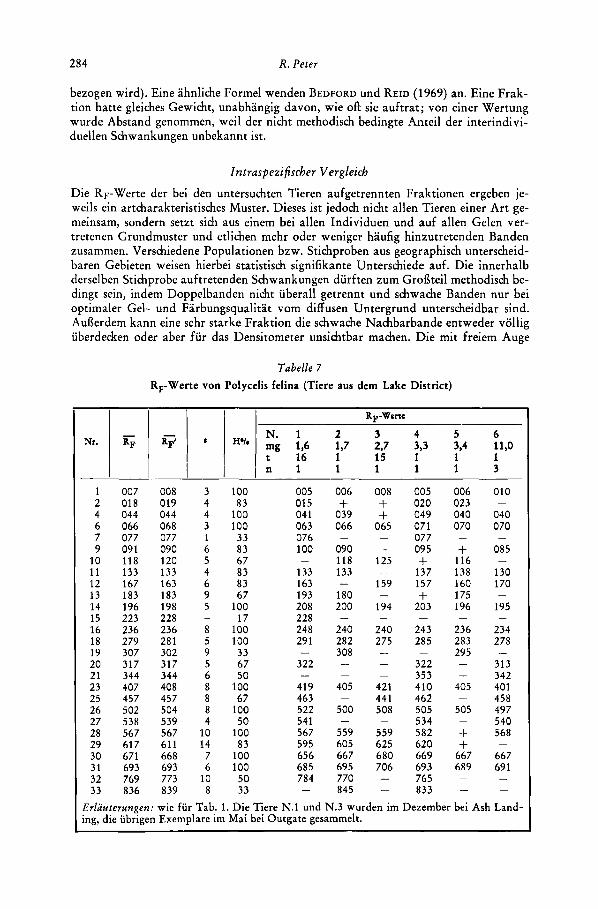
RrWerte von Polycelis felina (Tiere aus dem Lake District)
1 007 008 2 018 019 4 044 044 6 066 068 7 077 077 9 091 090
10 118 120 11 133 133 12 167 163 13 183 183 14 196 198 15 223 228 16 236 236 18 279 281 19 307 302 20 317 317 21 344 344 23 407 408 25 457 457 26 502 504 27 538 539 28 567 567 29 617 611 30 671 668 31 693 693 32 769 773 33 836 839
3 4 4 3 1 6 5 4 6 9 5
8 5 9 5 6 8 8 8 4
10 14 7 6
10 8
-
100 005 83 015
100 04 1 100 063 33 076 83 100 67 83 133 83 163 67 193
100 208 17 228
100 248 100 291 33 67 322 50
100 419 67 463
100 522 50 541
100 567 83 595
100 656 100 685 50 784 33
-
-
-
-
006 + 039 066
090 118 133
180 200
240 282 308
-
-
-
-_ -
405
500
559 605 667 695 770 845
-
-
008 + + 065 - -
125
159
194
240 275
-
-
-
- - -
421 441 508
5 59 625 680 706
-
- -
005 020 049 071 077 09 5 + 137 157 + 203
243 285
322 353 410 462 505 534 582 620 669 693 765 833
-
-
006 023 040 070
+ 116 138 160 175 196
236 283 295 + + 405
505
+ + 667 689
-
-
-
-
- -
010
040 070
085
130 170
195
234 278
313 342 401 458 497 540 568
667 69 1
-
-
-
-
-
-
-
- -
Erlucrterungen: wie fur Tab. 1. Die Tiere N.l und N.3 wurden im Dezember bei Ash Land- ing, die iibrigen Exemplare im Mai bei Outgate gesammelt.

Disk-Elektropboretiscbe Untersucbungen zur Artcbarakterisierung paludicoler Tricladen 285
- Nr. RF
sichtbaren, aber densitometrisch nicht aufgezeichneten Banden sind in den Tabellen entsprechend vermerkt. Reduziert sich die Variabilitat des Musters innerhalb einer Population schon dadurch, so mui3 man ferner bedenken, dai3 RF-Werte uber 0,7 meist schwachen Banden zugehoren und daher i. a. nur bei sehr geringer Untergrundfarbung und niedriger Gelinhomogenitat lokalisierbar sind, ihr Fehlen also nicht sehr signifi- kant ist. Aus dem Gesagten folgt, dai3 keine exakten Aussagen iiber die Variabilitat der Bandenzahl zwischen Individuen moglich sind, wenn auch die Unterschiede zu- meist methodisch bedingt sein durfien. Nur stark hervortretende Maxima vergleichend zu erfassen, scheint sowohl wegen der unvermeidlichen Subjektivitat in der Auswahl als auch deshalb unzulassig, weil die Intensitat einander entsprechender Banden von Tier zu Tier schwanken kann.
Da das Ziel dieser Arbeit Kennzeichnung und Vergleich taxonomischer Gruppen ist, konnte aui3er der Analyse verschieden groi3er Tiere keine bestimmte Auswahl hinsicht- lich ihres Zustandes getrofien werden. Bei Dugesia lugubris erfolgte die Aufsammlung zu verschiedener Jahreszeit; die Ergebnisse zeigen keine damit korrelierbaren Unter- s ch i e d e .
Tabelle 8
RFWerte von Phagocata vivida
RF-Werte
N. 1 2 3 4 mg 1,1 3,1 594 537 t 9 10 10 8 n 1 2 3 1
1 H%
1 006 2 020 3 041 4 057 5 069 6 09 1 7 106 8 122 9 145
10 168 11 177 12 205 13 226 14 241 15 249 16 29 1 17 350 18 377 19 42 1 20 440 21 462 22 476 23 514 24 565 25 61 1 26 640 27 686 28 755 29 793 30 839 31 920
4 2 3 3 1 4 2 2 4 4 5 6 4 5 5
10 15 7 5 5 2 4 9 9
10 4
15 9 7 6 4
- 50 75 020
100 043 50 75 069
100 089 100 104 25 - 50 75 166 25 - 75 25
25
50 50 75 426 75 447 25 75 4 76
100 521 100 571 100 610 75 638 50 25 25 25 25
-
-
- -
100 238
100 307 -
- -
-
- - - - -
012 020 039 059 069 092 106 122 148 - -
198 226 243 249 282 338 381
439 462 478 504 556 619 644 695 755 793 839 920
-
004
040 055 069 094 106
143 170 177 207
243
289 358
420
-
-
-
-
-
- -
475 513 565 602 - - - - - -
- 021 044 - -
084 105 - -
166
213
234
303
370 420 435
-
-
-
-
- -
522 580 623 637 670 - - - -
Erluuterungen: wie fur Tab. 1.

286 R. Peter
Die stets vorhandenen Frontbanden (RF = 1,000) werden ebenso wie die gefarbte Grenze zwischen Sammel- und Trenngel (RF = 0,000) in den Tabellen nicht angegeben, weil beide methodisch bedingt sind und daher keinen Aussagewert hinsichtlich der Identitat der zugehorigen Fraktionen besitzen. Die Auswahl der Abbildungen von Gelen und Pherogrammen erfolgte in der Absicht, jeweils reprasentative Bilder wieder- zugeben, die eine Vorstellung von der Veranderlichkeit des betreff enden Musters ver- mitteln.
Einzelergebnisse
Planariidae
Dugesia gonocephulu (Dug& 1830) (Tab. 1, 2, 12, Abb. 7-9):
Tiere der beiden Fundorte Wassergsprengbach und Hainbach sind einander sehr ahn- lich (Ahnlichkeit 65 O / o ) . Da sich einige Unterschiede auf selten vorkommende Banden und auf die Region uber RF = 0,8 beziehen, konnte die Obereinstimmung noch groi3er sein, als es den Anschein hat. Die beiden Fundorte sind jedoch deutlich durch die fol-
Tabelk 9 RFWerte von Dendrocoelum lacteum (Population aus Laxenburg)
N. 1 2 3 4
t 21 21 21 19 Nr* 1 4 1 * I H*'' 1 mg 4,O 6,5 10,O 15,O
I I I I n 1 2 2 2
1 2 3 4 5 6 7 8 9
10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29
005 01 7 037 060 082 101 114 125 144 171 187 201 220 248 272 296 337 396 440 476 511 540 588 605 625 658 703 773 932
3 7 5 4 8 8 4 4 4 5 5 6 6 5 4 4 8 5
15 3 7 6 8
13 7 6 3 4 3
- 86 71 009 86 041
100 065 57 092 57 100 71 - 86 -
100 138 86 -
100 185 100 + 100 224 100 243 100 267 100 292 86 340
100 394 86
100 471 100 520
-
- 71 57
29 86
- 100 586
- -
100 701 100 766
- 29
003 022 043 058 083 107
124 140 176 185 202 220 248 272 296 332 393 455 475 504 533
590 619 658 703 776
-
-
-
004
041 061 070 097 118 130 149 169 182 208 224 252 277 29 8 335 394 430 477 516 542
600 628 662 706 776 934
-
-
008
032 059
-
- -
112 125 145 167 189 199 215 246 272 295 341 397 429 474 514 539 589 615
656 699 771 932
-
5 6 20,7 25,5 21 2 1 1
003 01 8 036 064 - -
113 122 143 165 188 203 220 248 275 302
388 442 478 505 546 588 621
658 703 770
-
-
-
006 010
060 082 104 116 124 146 174 191 200 232 250 267 298 345 400 443 483 511 544 590 611
666 705 778
-
-
-
7 29,0 16 1
003 023 036 060
-
- -
114 121 145 175 182 194 213 243 273 294 337 403 412 475 507
585 612
651 702 770
-
-
-
Erlduterungen: wie fur Tab. 1. Das Tier N.6 wurde im April, alle anderen Exemplare wur- den im Dezember gesammelt.

Disk-Elektrophoretische Untersuchungen zur Artcharakterisierung paludicoler Tricladen 287
Nr.
genden, bei uber 50 o /o der Individuen vertretenen Fraktionen unterscheidbar: a. Was- sergsprengbach: Nr. 13 (100 O/o), 26 (78 O / o ) , 29 (56 O/O); b. Hainbach: Nr. 14 (100 O / o ) ,
27 (100 O / o ) .
Nur ein Tier aus dem Schreiber Bach wurde (fur methodische Studien) untersucht. Es wies gegenuber dem aus a. und b. erhaltenen Artspektrum nur eine zusatzliche Bande (Nr. 34a) auf. Es ist von Interesse, dai3 die Tiere der genannten Bache nach den An- gaben DAHMS (1958), der fur Individuen aus Grinzing und Modling eine haploide Chromosomenzahl von n = 8 angibt, in diesem Merkmal sehr wahrscheinlich uber- einstimmen.
- RF
Tabelle 10
RF-Werte von Dendrocoelopsis ezensis
1 2 3 4 5 6 7 8 9
10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38
008 020 037 050 059 073 081 091 099 111 131 138 148 152 164 183 208 219 232 249 265 276 296 386 404 430 455 51 1 575 626 654 696 719 732 770 792 840 951
N. 1 2 3 4 5 6 mg 6,8 6,9 8,4 9,5 10,O 12,l t 17 1 16 1 16 15 n 1 1 3 3 4 1
H%
2 50 5 83 4 100 2 67 3 100 2 33 3 83 4 17 5 83 8 33 1 17 3 67 5 67 - 17 5 67 5 100
11 83 2 33 6 83 5 83 0 33 5 67 7 83
10 83 7 67 - 17 7 83 6 100
13 33 11 100 6 83 9 100 4 17 7 67 7 67 4 17
10 67 7 83
- 014 043 050 060
081
104
-
-
- - - - -
159 181 21 1
237 249
275 300
402
454 512
610 649 686 719
766
834 947
-
-
-
-
-
-
-
-
016 03 1 048 062
08 1
106
-
-
- -
137 - -
165 187 - -
224 255
284 304 390 409
447 521 574 630 658 700
731 771
838 948
-
-
-
-
007 025 039
061
080 09 1
108
140 152
170 189 202
227 244
2 72 293 396 410
460 512
637 656 707
732
79 2
-
-
-
-
-
-
-
-
-
-
-
- -
009 01 8 034 050 058 072 080
096 -
- -
135 149 152
179 195
236 253
277 298 381 400
450 510
624 65 1 698
726 765
833 950
-
-
-
-
-
-
-
008 022 036
056 073 082
100 113 131
143
162 183 202 219 237
265
288 387
-
-
-
-
-
-
- -
461 505 575 628 656 695
739 776
840 959
-
-
- -
033 050 062 - - -
099 - -
138 149 - -
176 195 217
250 265
-
- -
384
430
508
639
704
-
-
-
-
- - - - -
956 Erluuterungen: wie fur Tab. 1.

288 R. Peter
Die densitometrisch erhaltenen Pherogramme weisen allgemein besonders viele und meist sehr deutliche Maxima auf, die mitunter Doppel- oder Dreifachbanden entspre- chen. Das starkste Maximum liegt zumeist im Bereich der Fraktionen Nr. 12 bis 16. Bande Nr. 35 bzw. 34a ist immer besonders scharf und deutlich. Das Exemplar aus dern Schreiber Bach (es ist mit 46,7 mg das schwerste der untersuchten Tiere) zeigt ein extrem hohes Maximum Nr . 2.
Dugesia t igrina (Girard 1850) (Tab. 3, Abb. 10, 11):
Die beiden untersuchten Aquarien-Populationen weisen im Kurvenverlauf relativ groi3e Ahnlichkeiten auf. Uberall sind die Fraktionen Nr . 23, 25 und 29 in abnehmen-
Tabelle 1 1 RFWerte von Bdellocephala brunnea
Nr.
1 2 3 4 5 6 7 8 9
10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36
004 1 008 1 023 2 039 3 059 5 0 72 8 086 1 089 1 102 3 114 4 120 2 136 3 148 4 173 4 193 6 205 6 24 1 6 269 5 290 12 307 7 319 6 371 8 396 11 438 10 482 11 512 9 529 11 562 7 606 8 635 5 665 8 686 7 729 23 810 22 937 4 951 3
H%
100 33
100 67
100 67 67 67 67 33 67 67 67 67
100 67
100 33
100 100 100 67
100 100 100 100
67 100 100 100 100 100 67 67 33 33
RF-Werte
3 18,9 6
006
022 028 060 072 085
106
128
159
188
244
293 310 323 3 70 391 432 485 518 546 570 600 633 674 685
-
-
-
-
-
-
-
- - - -
004
021 040 056 072
087
114
134 146 172 187 207 241
283 310 323 373 383 423 483 509
554 605 633 658 681 738 827 937
-
-
-
-
-
-
-
003 008 024
062
086 090 102
120 137
175 194 203 243 269 297 300 316
397 448 48 1 512 518 564 607 635 670 689 708 799
951
-
-
-
-
-
-
Erliiwterwngen: wie fur Tab. 1.

Disk-Elektrophoretische Untersuchungen zur Artcharakterisierung paludicoler Tricladen 289
der Hohe deutlich vertreten. Die Maxima Nr . 17 oder Nr . 18 sind meist a m hochsten. Bei den Tieren N r . 9 und 10 (2,2 und 2,9 mg) ist die Fraktion N r . 4 stark ausgepragt.
Dugesia lugubris (0. Schmidt 1861) (Tab. 4, Abb. 12, 13):
Die erhaltenen Kurven weisen kein einheitliches Charakteristikum auf. Die Tiere N r . 10 und 1 1 aus der Lunzer Population (1 1,l bzw. 12,5 mg) zeigen besonders starke Maxima Nr . 4 bzw. 5, bei jenem aus England (17,9 mg) ist die Fraktion N r . 4 sehr stark ver- treten. Leider konnte aus dem Lake District nur ein Exemplar erhalten werden, wes- halb die ermittelte Ahnlichkeit der beiden Populationen von 80 o/o keinen groi3en Aus- sagewert besitzt. Das englische Tier zeigt im Vergleich zur Lunzer Population keine einzige zusatzliche Bande, die Abweichungen sind nicht groi3er als innerhalb der ge- nannten Population.
Crenobia alpina (Dana 1766) (Tab. 5, Abb. 14, 15):
Alle Pherogramme zeigen deutliche Maxima N r . 24 und 29. Die Tiere Nr . 6 und 7 (5,l und 10,5 mg) besagen einen besonders hohen Anteil an Fraktion Nr . 2. Die zu- satzlich zu den beiden Populationen aus der Primelquelle und von der Villacher Alpe untersuchten Tiere aus der Umgebung von Lunz am See ergaben ein ziemlich unein- heitliches Bild, das mehrere elektrophoretisch abgrenzbare Typen moglich erscheinen lafit. Wegen der zu geringen Individuenzahl sind weitere Experimente erforderlich, um diesen Sachverhalt zu klaren.
Polycelis felina Dalyell 1814 (Tab. 6, 7, Abb. 16, 17):
Die erhaltenen Pherogramme weisen keine erkennbaren allgemeinen morphologischen Charakteristika auf, doch ist der Eindruck der beiden untersuchten Populationen deut- lich verschieden. Die Tiere aus Oberosterreich hatten immer stark hervortretende Maxima Nr . 26 und 30.
Phagocata wiwida Ijima et Kaburaki 1916 (Tab. 8, Abb. 18):
Fraktion Nr. 14 reprasentierte in allen Kurven auger bei Tier Nr . 2 das Hauptmaxi- mum; bei diesem war Bande N r . 16 am starksten entwickelt. Dieses Exemplar zeigte auch auffallend mehr Maxima als die ubrigen Individuen.
Dendrocoelidae
Dendrocoelum lacteum 0. F. Miiller 1773 (Tab. 9, 12, Abb. 19, 20):
Die Tiere aus der Laxenburger Population fuhrten zu morphologisch untereinander z. T. sehr ahnlichen Pherogrammen. Die Maxima Nr . 20, 24 und 27 sind in abnehmen- der Intensitat uberall deutlich ausgepragt. Stets t ra t Bande Nr. 4 hervor, vor allem bei Individuen uber 20 mg.
Das eine Exemplar aus dem Windermere Lake wurde zusammen mit einem Tier aus Laxenburg in einem Experiment untersucht, bei dem die Elektrophoresezeit und moglicherweise auch die Gelzusammensetzung leicht von den sonst angewandten Be- dingungen abwich; es ist daher kein Vergleich mit den ubrigen Laxenburger Tieren moglich, die Fraktionen sind auch unabhangig von diesen numeriert. Zusatzlich zum gemeinsamen Grundmuster von 23 Banden wies das Laxenburger Tier 7, jenes aus Eng- land 3 Fraktionen auf.

290 R. Peter
Nr. ~ Dwgeria gonocepbala Polycrlir felina 1 RF I a) 1 b) I C) d) I e)
Tabelle 12
R,-Mittelwerte von Dugesia gonocephala, Polycelis felina und zweier Exemplare von Den- drocoelurn lacteurn
Dendrocoelum lacteum
f) j 9)
1 2 3 4 5 6 7 8 9
10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 34a 35 36 37 38 39 40
007 025 039 054 073 080 09 1 105 105 117 129 156 168 178 194 215 231 246 267 278 306 339 358 393 408 434 447 459 486 515 555 577 608 64 1
685 737 782 826 886 949
-
006 023 039 055 073 080 09 1 104 105 122 131 158 168
194 216 236 248 267 276 307 334 358 392 405 434
455 486 523
573 608 643
683 732 779 820 886 949
-
-
-
-
008 026 039 054 075
09 1 105
115 127 154
178
214 224 244
279 304 347
395 411
447 465
508 555 583
638
688 743 783 802
-
-
-
-
-
-
-
-
-
-
- -
- 032 - -
072
096 103
-
- -
128 159
177
210
245
-
- -
- -
305 333
385
430 448
479 513 564 596 608 632 666
729 763 807
-
-
-
-
- -
007 018 035 044 047 066 077 086 09 1 118 133 167 183 196 223 236 260 279 307 317 344 386 407 420 457 502 538 567 617 671 693 769 836
006 018 035
047 064
086 093 118 133 172
194 223
260 276 308
-
-
-
-
- -
386 404 420
500 538
620 673 695 763 827
-
-
008 019
044
068 077
090 120 133 163 183 198 228 236
281 302 317 344
408
457 504 539 567 61 1 668 693 773 839
-
-
-
-
-
-
006 027 038 053 065 09 1 122 148
163 185 203 233 251 272 318 369 387 425 460 488 514 529 579 607 638 685 735 754 783 848
932
-
-
014 027 04 1
064 089 109 140 154 169 179 203 232
276 324 368 383 428 459 49 1
551 576 61 1 64 1 683
755
838 910
-
-
-
-
-
-
Erluuterungen: Populationen: a) Wassergsprengbach, b) Hainbach, c) Schreiber Bach (Tier von 46,7 mg), d) Quellbach am Gleinker See, e) Lake District bei Ambleside, f ) Laxenburg (Tier von 15,5 mg), g) Windermere Lake (Tier von 9,7 mg). Hier sind nur die Mittelwerte jener Arten, deren Populationen aus Grunden der Obersicht- lichkeit nicht in einer der vorangegangenen Tabellen einander gegeniibergestellt sind, ange- geben. Die Nummern der Fraktionen (Nr.) sind mit denen der anderen Tabellen der cnt- rprechenden Arten identisch, ausgenommen Dendrocoelum lacteum (s. Text). RF-Werte verschiedener Arten stehen in dieser Tabelle nur durch die Zahlung bedingt neben- einander, gehoren also nicht notwendigerweise identischen Fraktionen an. Die Mittelwertc (RF) von Dugesia gonocephala wurden nur aus den unter a) und b) angegebenen Werten gebildet. Unter c) sind die Mittelwerte aus 4 Parallelproben, unter f) und g) jene aus 3 bzw. 4 Parallelproben jeweils eines Exemplars angegeben.
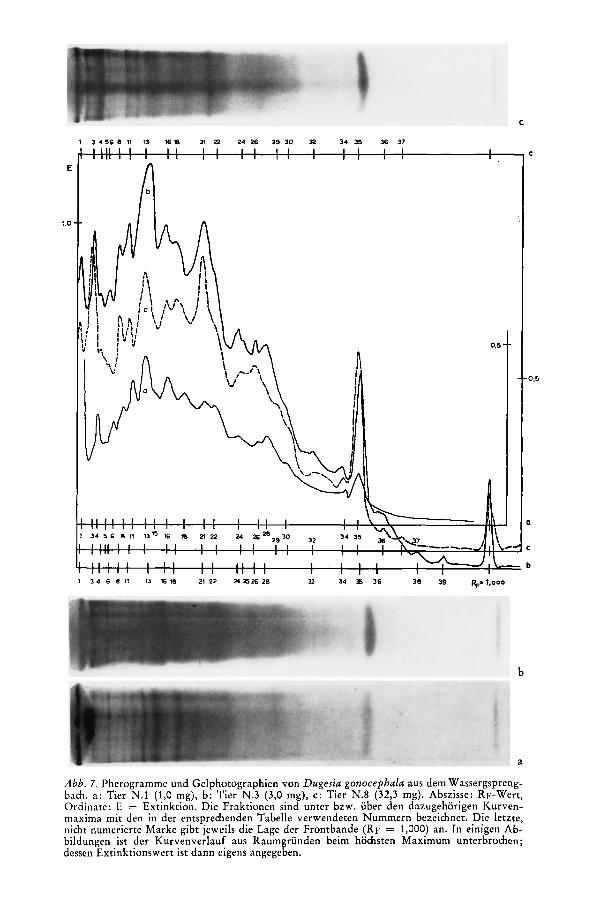
C
3 4 5 6 0 11 13 1 6 8 21 P 24 26 29 30 32 34 35 36 37
*!I I I I I ! I I I I I I I I I I I I I I I 1 I 1 I I 1 I 1 I
A
1 3 4 6 8 1 1 13 1618 2122 2 4 8 2 6 2 8 32 34 35 36 38 39 ~ 1 , 0 0 0
b
a
Abb. 7. Pherograrnrne und Gelphotographien von Dwgesia gonocephala aus dern Wassergspreng- bnch. a : Tier N.l (1,0 mg), b: Tier N.3 (3,O rng), c : Tier N.8 (32,3 rng). Abszisse: RF-Wert, Ordinate: E = Extinktion. Die Fraktionen sind unter bzw. uber den dazugehorigen Kurven- maxima rnit den in der entsprechenden Tabelle verwendeten Numrnern bezeichner. Die letzte, nicht nurnericrte Marke gibt jeweils die Lage der Frontbande (RF = 1,000) an. In einigen Ab- bildungen ist der Kurvenverlauf aus Raumgrunden beirn hochsten Maximum unterbrochen; dessen Extinktionswert ist dann eigens angegeben.
I C
-03
a
C

292 R. Peter
Abb. 8. Pherogramme und Gelphotographien von Dugesia gonocephafa aus dem Hainbach. a: Tier N.4 (5,8 mg), b: Tier N.6 (37,O mg). Erlauterungen wie zu Abb. 7
a
- 0.0
a
Abb. 9 . Pherograrnme und Gelphotographien von 3 Parallelproben von Dugesia gonocephala aus dem Schreiber Bach (Tier von 46,7 mg) zur Illustration der Reproduzierbarkeit der Tren- nungen. Die Hornogenate wurden bei verschiedenen Temperaturen gelagert: a: 90 min bei -20" und 60 min bei 0-5O, b: 150 min bei 0-50, c: 80 min bei 20° und 70 min bei 0-5". Die angegebenen Zeiten sind vom Homogenisieren bis zum Beginn der Elektrophorese gerech-
net. Weitere Erlauterungen wie zu Abb. 7

a
C

294
1 I l l I I I I I I I I I l l E
0.5 A
R. Peter
--0.1
I I l l I I I I 1 1 I b 1 I l l I I I I I 1 I J L ~ a i i I I 1 I I I: I I I I I I : I l l
I l l I I 1 4 5 6 B 9 10 12 U 16 17 19 10 2 2 21 24 27 29 30
Abb. 10. Pherogramme und Gelphotographien von Dugesia tigrina (Population a). a: Tiere N.2 (1,0 mg), b: Tiere N.l (<1 mg). Erlauterungen wie zu Abb. 7
Dendrocoelopsis ezensis Ichikawa et Okugawa 1958 (Tab. 10, Abb. 21):
Der grob morphologische Eindruck der Pherogramme schwankt. Die Banden Nr. 19 oder Nr . 20 treten uberall als hochste Maxima auf, Nr. 31 ist immer markant ausge- pragt.
Bdellocephala brunnea Ijima et Kaburaki 1916 (Tab. 11, Abb. 22):
Die Pherogramme der drei untersuchten Exemplare zeigen hervortretende Maxima Nr. 17 und 30, dazwischen liegt jeweils eine Zone mehr oder weniger stark abgestufter Maxima von abnehmender Hohe. Bei Tier Nr. 3 (d. i. mit 18,9 mg das schwerste Exem- plar) ist die Fraktion Nr. 5 besonders stark vertreten.
Populationsvergleich
Die RF-Werte der einzelnen Populationen sind einander in den Tabellen der entspre- chenden Arten bzw. in Tabelle 12 gegenubergestellt. Ein Ahnlichkeitsvergleich ergibt das folgende Bild.
Die geringste intraspezifische Ahnlichkeit, die nur knapp uber den hochsten beobach- teten, wenn auch eindeutig uber den durchschnittlichen interspezifischen Ahnlichkeiten

Disk-Elektrophoretische Untersuchungen zur Artcharakterisierung paludicoler Tricladen 295
Tabelle 13 Ahnlichkeiten zwischen Populationen
Zahl der Banden Ahnlidy- keit insgesamt I damn
identisdy
Art, Populationen
Dugesia gonocephala: a. Wassergsprengbach, b. Hainbach 40 26 65
Dugesia tigrina: 2 Aquarienpopulationen 30 20 67 Dugesia lugubris: a. Lunz am See,
b. Lake District bei Ambleside 25 20 80 Crenobia alpina: a. PrimelquelleiWassergspreng,
b. Quelle des Notscher Baches 32 20 63 Polycelis felina: a. Quellbach bei Windischgarsten,
b. Lake District bei Ambleside 33 19 58 Dendrocoelum lacteum: a. Laxenburg,
b. Windermere Lake 33 25 76
liegt, weist Polycelis felina auf. Das konnte nicht nur mit der grogeren Distanz der Fundorte im Vergleich zu Dugesia gonocephala, D. tigrina und Crenobia alpina zusam- menhangen, sondern auch mit cytogenetisch erkennbaren Unterschieden korrelierbar sein. Die Chromosomenzahlen weisen nach DAHM (1958) bei der erwahnten Ar t starke, vom Fundort abhangige Schwankungen auf. Leider waren diesbezugliche Untersu- chungen nicht moglich. Dugesia lugubris und Dendrocoelum lacteum, bei denen Tiere von Fundorten etwa gleicher gegenseitiger Entfernung analysiert wurden, zeigen zwar die hochsten intraspezifischen Ahnlichkeiten; da aber von beiden Arten jeweils nur ein Exemplar aus England den Postversand uberlebte und untersucht werden konnte, sind die errechneten Hhnlichkeiten mit Vorbehalt zu betrachten.
Tabelle 14 Interspezifische Ahnlichkeiten in O l o
~
Dugesia gonocephala Dugesia tigrina Dugesia lugubris Crenobia alpina Polycelis felina Phagocata vivida Dendrocoelum lacteum Dendrocoelopsis ezensis
49 35 48 38 58 35 56 55 45 36 50 57 41 42 50
40 42 44 39 34 49 46 51 46 50 49
49 38 48 47 46 50 56
46 48 40

296 R. Peter
I
u.5
I 4 c i a 9 11 1314 18 19 20 22 2 3 x 26 29 29 30 I 1 I 1 1 I I1 I 1 I , I I 1 I 1 I I I I 1 , 9 I1 I I I , I , , I I I +
4 5 7 8 9 11 O H 18 19 2 0 22 23 25 26 -3’ 29 1
(I
0.5
b
1 ,7 3 4 5 ? 9 1’ ‘ 2 14 15 1- 17 2 0 23 25 29 30
Abb. 11. Pherogramme und Gelphotographien von Dwgesia tigrina (Population b). a: Tiere N.l (1,4 mg), b: Tiere N.6 (2,2 mg). Erlauterungen wie zu Abb. 7
Interspezifischer Vergleich
Die Ahnlichkeiten zwischen den untersuchten Arten sind in Tab. 14 angegeben. Wie daraus hervorgeht, ergeben sich weder Abstufungen nach der Familien- noch nach der Gattungszugehorigkeit. Die Artspektren sind jedoch immer eindeutig, und selbst die bei allen Experimenten mit den einzelnen Arten gefundenen ,,minimalen“ Grundmuster (Tab. 15) reichen, von drei Ausnahmen abgesehen, zur Artcharakterisierung aus, so dafl zumeist schon ein Elektrophorese-Experiment mit einem Tier zur Bestinimung einer Art genugen muflte. Damit sol1 freilich, jedenfalls bei den hier behandelten Species, die Disk-Elektrophorese nicht als Mittel zur Artbestimmung empfohlen werden; ihre

Disk-Elektrophoretische Untersuchungen zur Artcharakterisierung paludicoler Tricladen 297
Tabelle 15
Artcharakteristische Grundmuster (bei allen untersuchten Individuen der einzelnen Arten gefundene Fraktionen)
a. Planariidae
Identisrhe Frakrion bei
Dugesia gonocephala 007 105 129 215 306 393 685
Dugesia tigrina 039 + 130 + 358 -1- 444 i- 668
Dugesia lugubris 057 + 29 1 326 403 + 723 +
Crenobia alpina 170 + 246 + 270 + 523 + 703
Polycelis felina
671 Phagocata vivida 041 +
091 + 106 + 241 291 + 514 + 565 + 611 +
+ + + + + + + + + + i- + + + + + + + + + + +
+ i - + + + + + + + + - I - + + + + + + + + +
+ + + + + + + + + + + + + + + + + + + + + + + + + + + + +
+ +
+ + + + + + + + + + + + + + + + + + + + + + + + -I- + + + + + + + + + +
+ + + + + +
+ +
+ +
+
+ + + + + + +
besonderen Aussagemoglichkeiten liegen wohl, bezogen auf die speziell angewandte Methodik, im Bereich intraspezifischer Abgrenzungen. Die Planariiden weisen wie auch die Dendrocoeliden cine allen Arten der Familie gemeinsame Bandenkombinarion auf, die jeweils bei keiner Art der anderen Familie vertreten ist (Tab. 16); allerdings ge- horen nicht alle Fraktionen den Grundmustern der Arten an. Der beschriebenen Fami- liencharakterisierung sollte jedoch wie auch der Eindeutigkeit der ,,minimalen" art-

298 R. Peter
b. Dendrocoelidae
I Idemisdie Fraktion bei I
Dendrocoelum lacteum 060 + + + + + + 144 + + + + 220 + + + + + + + 248 + + + + + 272 + + + 296 + + + + + + 396 + + + + + + 476 + + 511 + + + + 605 + + + + + 703 + + 773 + +
Dendrocoelopsis ezensis 037 + + + + + + + 059 183 + + + + + + +:I + + 511 + 6 2 6 + + + + + + + 696 + + + 023 + + + + + + 059 + + + + + + 193 + + + + + 241 + + + + + 290 + + + + + + 307 + + + + 319 + + + + 3 9 6 + + + + + + + 438 4- + + + + + + 482 + + + + + 512 + + + + 562 + + + + + 606 + + + + + + 635 + + + + + 665 + + + + + 686 + +
-J- Bdellocephala brunnea 004 + +
Erluutermgen: RF = RpMittelwert der Art. In den Tab. 1 bis 1 1 mit ,,+" und ,,v" an- gegebene Fraktionen sind hier nicht berucksihtigt. Die drei Falle vollstandiger Uberein- stimmung einer Bandenkombination mit Fraktionen aus dem Grundmuster einer anderen Art sind umrandet.
charakteristischen Spektren kein allzu groi3es Gewicht beigemessen werden, weil beide stark von der Zahl der untersuchten Arten abhangen diirften, wie ein Vergleich zwi- schen den Fraktionszahlen der Muster der Planariidae (6 Arten) und der Dendro- coelidae (3 Arten) nahelegt. Auch die Gattung Dugesia lal3t sich durch ein den drei untersuchten Arten gemeinsames Grundmuster kennzeichnen (Tab. 17).

Disk-Elektropboretiscbe Untersucbungen zur A rtcbarakterisierung paludicoler Tricladen 299
1 4 6 7 8 9 1 0 1 1 1 2 U 8 16 17 m 79 20 2l 22 23 25
I
1 2 3 4 5 6 7 8 9 10 1213 14 15 16 17 18 20 22 24 25
b
-0.5
b
Abb. 12. Pherogramme und Gelphotographien von Dugesia lugubris aus Lunz am See. a: Tier N.8 (3,7 mg), b: Tier N.10 (11,l mg). Erlauterungen wie zu Abb. 7
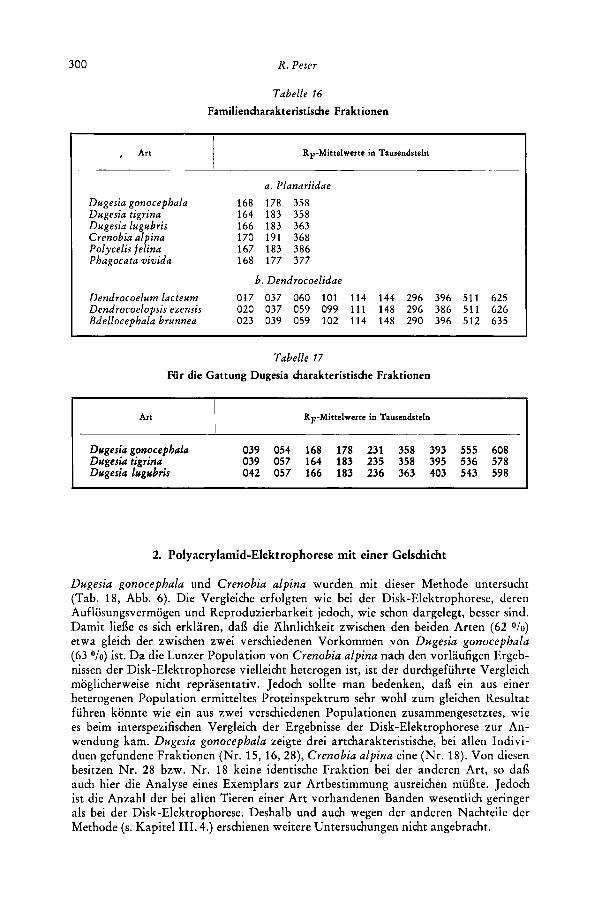
300
Art
R. Peter
RF-Mittelwerte in Tausendsreln
Tabelle 16
Familiencharakteristische Fraktionen
, Art R~-Mirrelwerre in Tausendsteln I Dugesia gonocephala Dugesia tigrina Dugesia lugubris Crenobia alpina Polycelis felina Phagocata vivida
Dendrocoelum lacteum Dendrocoelopsis ezensis Bdellocephala brunnea
~ ~~
a. Planariidae 168 178 358 164 183 358 166 183 363 170 191 368 167 183 386 168 177 377
b. Dendrocoelidae 017 037 060 101 114 144 296 396 511 625 020 037 059 099 1 1 1 148 296 386 511 626 023 039 059 102 114 148 290 396 512 635
I Dngesia gonocephala 039 054 168 178 231 358 393 555 608 Dugesia tigrina 039 057 164 183 235 358 395 536 578 Dngesia lngnbris 042 057 166 183 236 363 403 543 598
2. Polyacrylamid-Elektrophorese mit einer Gelschicht
Dugesia gonocephala und Crenobia alpina wurden mit dieser Methode untersucht (Tab. 18, Abb. 6). Die Vergleiche erfolgten wie bei der Disk-Elektrophorese, deren Auflosungsvermogen und Reproduzierbarkeit jedoch, wie schon dargelegt, besser sind. Damit liefie es sich erklaren, dai3 die Ahnlichkeit zwischen den beiden Arten (62 o/o) etwa gleich der zwischen zwei verschiedenen Vorkommen von Dugesia gonocephala (63 O / o ) ist. Da die Lunzer Population von Crenobia alpina nach den vorlaufigen Ergeb- nissen der Disk-Elektrophorese vielleicht heterogen ist, ist der durchgefuhrte Vergleich nioglicherweise nicht reprasentativ. Jedoch sollte man bedenken, dai3 ein aus einer heterogenen Population ermitteltes Proteinspektrum sehr wohl zum gleichen Resultat fiihren konnte wie ein aus zwei verschiedenen Populationen zusammengesetztes, wie es beim interspezifischen Vergleich der Ergebnisse der Disk-Elektrophorese zur An- wendung kam. Dugesia gonocephala zeigte drei artcharakteristische, bei allen Indivi- duen gefundene Fraktionen (Nr. 15, 16, 28), Crenobia alpina eine (Nr. 18). Von diesen besitzen Nr. 28 bzw. Nr. 18 keine identische Fraktion bei der anderen Art, so daa auch hier die Analyse eines Exemplars zur Artbestimmung ausreichen muate. Jedoch ist die Anzahl der bei allen Tieren einer Art vorhandenen Banden wesentlich geringer als bei der Disk-Elektrophorese. Deshalb und auch wegen der anderen Nachteile der Methode (s. Kapitel 111.4.) erschienen weitere Untersuchungen nicht angebracht.

Disk-Elektrophoretische Untersuchungen zur Artcharakterisierung paludicoler Tricladen 30 1
Tabelle 18
Polyacrylamidgel-Elektrophorese: RF-Mittelwerte von Dugesia gonocephala und Crenobia alpina sowie intra- und interspezifischer Vergleich
~
Dugeria gonocephala
I I a. Wassergsprengbach I b. Hainbach ~
1 2 3 4 5
6 7 8
9 10 11 12 13 14 15
16
17 18 19 20 21 22 23 24
25
26 27
28
29 30
0,115 171 210 253 307
357 432 479
538 583 632 665 733 745 824
1,006
074 155 244 290 366 428 516 609
883
2,040 100
338
569 842
0,118 172 210 249 308
357 435 479
542 585 632 667 736 74 1 821
1,003
~
-
244
367 -
- -
607
885
2,045 100
328
569 825
19 63 17 88 12 75 12 25 30 75
29 88 26 63 22 63
19 63 45 88 17 25 38 38 24 38 27 88 30 100
33 100
74 100
102 50
106 100
107 38
35 38 86 38
59 100
94 38 112 38
0,091 158
265 303
-
-
417 -
512 573
662 720 768 855
1,026
074 155 246 290 363 428 516 645
778
2,020
-
-
415
-
910
~ 50 ~ 50
50 15 100 -
2 100
0 100 18 100
5 100 - 50 11 100 13 100
13 100
3 100 50
- 50 50
- 50 39 100 21 100 - 50
4 100
- 50
-
-
92 100
- 50
Crenobia alpina
c. Lunz am See - I
I 1 2 3 4
5 6 7 8 9
10 11 12
13 14 15 16 1 7 18 19
20
21
22 23 24 25 26
27
28 29 30
, - .
0,112 14 158 16 198 21 273 15
341 21 382 29 420 25 471 21 512 17 550 18 572 12 635 28
729 - 748 24 834 27 906 43 973 21
1,038 27 056 51
211 31
372 34
493 39 625 58 770 -
848 86 920 ~ 32
2,244 71
429 54 542 33 724 83
Ha/Q 1 -
20 40 90 90
90 60 90 50 60 20 30 40
10 90 80 80 20
L 00 20
40
40
60 50 10 60 50
60
80 30 70
Erluuterungen: Nr. = Nummer der Fraktion; s = Streuung (Standardabweichung) in Tau- sendsteln; H"/o = HPufigkeit der entsprechenden Fraktion in Prozenten, bezogen auf die Zahl der untersuchten Tiere. Die R p W e r t e sind auf das Transferrin des menschlichen Se- rums (RI.? = 1,000) bezogen. Nach dem t-Test zu identischen Banden gehorige Werte stehen ieweils in derselben Zeile. Zahl der untersuchten Tiere und der Parallelprobcn: a) 8 Tiere (7,5 bis 31,8 mg), 15 Proben; b) 2 Tiere (24,6 und 29,5 mg), 2 Proben; c) 10 Tiere (2,2 bis ),5 nig), 17 Proben.

302 R. Peter
Tabelle 19
R,-Werte der Zymogramme
Disk- Polyacrylamidgel- I Elektrophorese Elektrophorese
~~ ~
I - - - LDH . - l M D H ~ ~ L D H 1- MDH
Dugesia gonocephala Dugesia lugubris Crenobia alpina O,OlO,Ol
0,02 (034)
Polycelis felina + 0,o (0,02/0,3)
Dendrocoelurn lacteum -
Dendrocoelopsis ezensis - - Bdellocephala brunnea - 0,16
0,44 0,45/0,50 - (1,111,5) 0,54/0,68
1,18 . -
- 0,11/0,17 0,3 1/0,70
- 0,60 ~
Erliiuterungen: Bezugsfraktionen fur die RF-Werte sind, wie auch sonst, die Frontbande bzw. Transferrin. Ihrer geringeren Genauigkeit entsprechend, sind die aus jeweils 1 bis 8 Einzel- werten gebildeten RF-Werte nur auf 1 bis 2 Stellen angegeben. Unterstrichene Werte ge- horen besonders starken Banden an, in Klarnrnern gesetzte traten nicht irnrner auf. Nega- tive Ergebnisse sind durch ein ,,-" verzeichnet. +O,O: Bande irn Trenngel knapp nach dessen Grenze zum Sammelgel.
I l l I l l I 1 I t 1 I I I I I 1 I l l I l l 1 1 I I 1 I I I I I I I 1 3 5 6 7 8 II 12 U 8 I C 17 18 18 If 22 15
Abb. 13. Pherograrnrn und Gelphotographie von Dugesia lugubris von Ees Bridge bei Arnble- side (Tier von 17,9 rng). Erlauterungen wie zu Abb. 7
Abb. 14. Pherograrnrne und Gelphotographien von Crenobia alpina aus der Prirnelquelle. a: Tier N.2 (1,7 mg), b: Tier N.4 (2,5 mg), c: Tier N.7 (10,5 rng). Erlauterungen wie zu Abb. 7
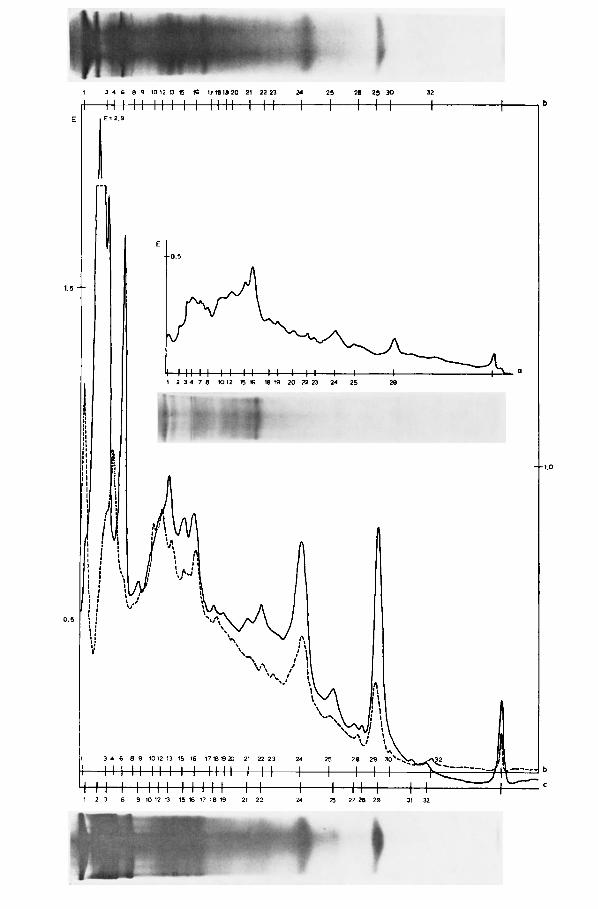
1 3 4 6 8 9 1012 U S l6 17191020 2l 2223 25 1 25 30 32 I l l I 1 I 1 I I 1 I l l 1 I I 1 I l l I I I l l I I I I I I I I I
I 1 I I I I 1 I I l l
i z 3 4 7 8 w12 n s 18 19 20 P P 24 25 20
3 4 6 8 9 101213 15 16 178RaD 21 2223 24 25 28 29 30' I I I I 1 I 1 I I I I I I I I I I I I I I I
I l l I I I 1 1 I I l l 1 I I I I I I I I 1 I I I I I I I I I I I 1
I I I I I I
1 2 3 6 9 10 12 l3 15 6 17 18 19 21 22 24 Z 27 28 29 31 32

304 R. Peter
\
1 23 5 6 8 9 11 QB 15 16 7? 19 21 23 24 25226 29 3l 32
Abb. IT. Pherogramm und Gelphotographie von Crenobia alpina aus dem Notscher Bach (Tier N.2 von 2,O mg). Erlauterungen wie zu Abb. 7
3. Zymogramme (Vorlaufige Ergebnisse)
Erste Versuche zur Lokalisierung von Lactat-Dehydrogenasen (LDH) und Malat-De- hydrogenasen (MDH) ergaben die in Tabelle 19 dargestellten Resultate. Die Farb- intensitat war bei den Polyacrylamidgelen mit nur einer Schicht hoher. Die Oberflachen der Gele und die Frontbanden waren fast immer gefarbt, jedoch konnte bei diesen schwer zwischen ihrer Eigenfarbe und der Farbung des Reaktionsproduktes unter-
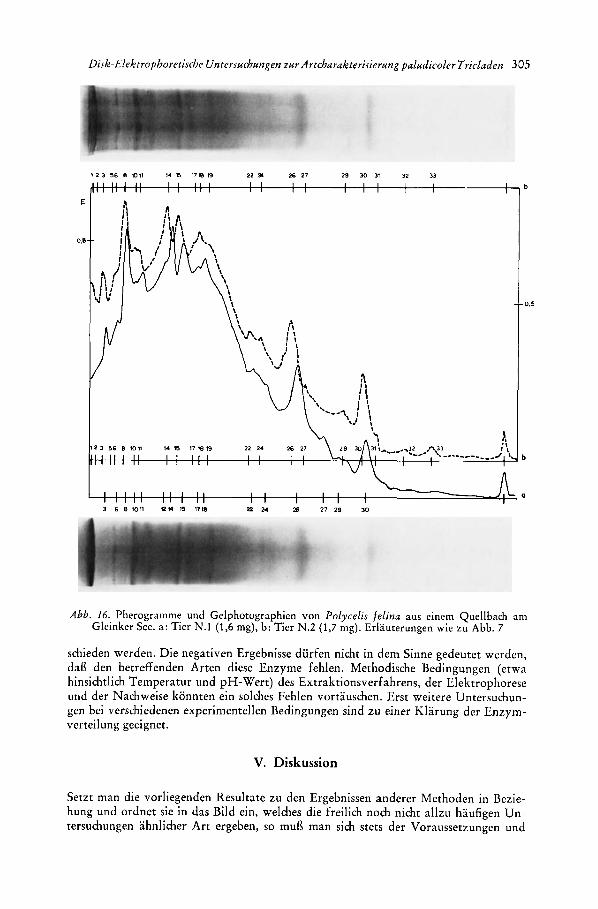
E
0.1
Disk-Elektrophoretische Untersuchungen zur A rtcharakterisierung palxdicoler Tricladen 305
1 2 3 5 6 8 1 0 ~ 1 4 8 17819 22 a 26 2 1 29 30 31 32 33 I I1 I I 1 I I I 1 I I 1 I 1 I I 1 I I I
I I I 1 I I 1 I I 1 I I
3 56 0 1011 14 15 17 10 19 22 24 26 27
- I ! ! / II I I I 1 I I 1 I 1
I I 1 I I I 1 I I 1 I 1 I I I 1 I I I
3 6 0 ion t 2 n 15 010 22 24 P 27 29 30
Abb. 16. Phcrogramme und Gclphotographicn von Polycelis felina aus eincm Quellbach am Glcinker See. a : Tier N.l (1,6 mg), b: Tier N.2 (1,7 nig). Erlauterungen wic zu Abb. 7
schieden werden. Die negativen Ergebnisse diirfen nicht in dem Sinne gedeutet werden, da8 den betreflenden Arten diese Enzyme fehlen. Methodische Bedingungen (etwa hinsichtlich Temperatur und pH-Wert) des Extraktionsverfahrens, der Elektrophorese und der Nachweise konnten ein solches Fehlen vortauschen. Erst weitere Untersuchun- gen bei verschiedenen experimentellen Bedingungen sind zu einer Klarung der Enzym- verteilung geeignet.
V. Diskussion
Setzt man die vorliegenden Resultate zu den Ergebnissen anderer Methoden in Bezie- hung und ordnet sie in das Bild ein, welches die freilich noch nicht allzu haufigen Un- tersuchungen ahnlicher Art ergeben, so mui3 man sich stets der Voraussetzungen und

306 R. Peter
1 4 6 9 afl 014 6 1819 23 25 20 21 30 31 P 33 I I I I I I , I I I I I I I I , I I I I I I 1 1 1 I 1 I 1 1 I I I
3
0.5
1 4 6 9 17 12 14 16 18 20 21 23 25 26 2 7 30 31
Abb. 17. Pherogramme und Gelphotographien von Polycelis felina aus dem Lake District. a: Tier N.2 (1,7 mg), b: Tier N.6 (11,O mg). Erlauterungen wie zu Abb. 7
Abb. 18. Pherogramme und Gelphotographien von Phagocata vizrida. a: Tier N.2 (3,l mg), b: Tier N.4 (5,7 mg). Erlauterungen wie zu Abb. 7

E
O . !
d E z 1 . 8
-I I 1 1 1 I 1 I I I I I 1 I I I I 1 I
2 3 6 7 10 12 14 16 1.9 1920 23 24 nZ6 27

308 R . Peter
E
0.5
4 5 6 8 9 lOnT213 14l5 6 17 2 0 21 22 24 26 27 28
I I I l l I I I I I 1 I I I I I I I I l l 1 I I I I I l l I I I I I
I l l I I I I I a
0.9
a
Abb. 19. Pherogramme und Gelphotographien von Dendrocoelum lacteum aus Laxenburg. a: Tier N.2 (6,5 mg), b: Tier N.4 (15,O mg). Erlauterungen wie zu Abb. 7
Moglichkeiten der Disk-Elektrophorese und der Grenzen und Unsicherheiten der hier im speziellen angewendeten Methode bewui3t bleiben, wie sie im Abschnitt ,,Kritik der Methode"' naher erortert worden sind. Ganz abgesehen von der Tatsache, dai3 jene Probleme, die sonst bei der systematischen Abgrenzung und Einordnung von Arten auftreten, auch durch die biochemische Methodik wenigstens nicht vollig beseitigt wer- den (vgl. SIBLEY 1962), erlaubt die Elektrophorese, wie sie hier und in ahnlichen Arbei- ten angewandt wurde, keine eindeutige Charakterisierung einzelner Proteinmolekul- species. Die Frage nach der Vergleichsbasis fur Elektrophoresespektren beantwortet SIBLEY (1960) dahingehend, dai3 in erster Linie die Position der Banden (also die RF-

E
1.c
Disk-Elektrophoretische Untersuchungen zur Artcharakterisierung paludicoler Tricladen 309
32 2 3 5 6 7 89011 l2 13 15 16 17 zo 21 a 24 r) 26 27 29 I I I 1 I 1 I I I I I 1 I I I I I I I I
,A i I I I : I I I I ; : I I I I I I l l 1 I I
4 : I I I I I 1 I I I I I !I
I I I 1 I I I q \ z L 2 3 5 6 7 8 9 u n 1 2 '13 8 16 17 20 21 P 24 2S 26 27
I I I I I l l I I 1 I I I I I I l l I 1 1 1
33 2 3 5 6 7 8 l O n 1 2 1 3 1 4 1 3 16 1718 2 0 2 1 2 2 a 2 4 1 2 6 27 282930 31
I
0.5
a
Abb. 20. Pherogramme und Gelphotographien von Dendrocoelum lacteum. a: Tier aus Laxen- burg (15,5 mg), b: Tier aus dem Windermere Lake (9,7 mg). Erlauterungen wie zu Abb. 7.
Die Fraktionsnummern entsprechen nicht jenen von Abb. 19 (s. Text)!

310 R. Peter
1 2 3 5 7 8 1 0 1 2 0 8 6 lQ2U2223 24 27 28 P31 34 36 I I
I 1 I 1 I 1 1 1 1 1 I 1 I 1 I I I I I 1 1 1 1 1 I 1 I 1 I I I I
I 1 1 I 1 I I
1
I 1 1 1 1 1 I 1 I 1 I I I I I I 1 2 3 57810 R D D K 1 9 i U n 2 3 24 27 28
1 1 1 1 I I I I I I I I I l l
I 1 I I 1 I I
I I I I I I I I I I I
30 31 32 34 35 1 2 3 4 5 6 7 9 G ? U W 6 1 7 192022D 2425 27 28

Disk-Elektrophoretische Untersuchungen zur Artcharakterisierung paludicoler Tricladen 3 1 1
Werte), dann die Gestalt der densitometrisch erhaltenen Kurven und erst in letzter Hinsicht relative und absolute Hohe der Maxima heranzuziehen sind. Dies entspricht der auch vom Verfasser angenommenen Naherung, eine Proteinzone reprasentiere eine I~!olekiilspecies, die durch ihr elektrophoretisches Verhalten, also durch ihren RF-Wert, charaktcrisiert ist. Zufallsiibereinstimmungen zwischen in Wahrheit unterschiedlichen Fraktionen treten dabei bestimmt auf. Es konnte aber sein, dai3 diese zwar insgesamt cine hohere Identitat zwischen verschiedenen Proteingemischen vortfuschen, als sic tatsachlich vorhanden ist, dai3 aber trotzdem Extrakte mit groi3erer von solchen mit gcringerer Ubereinstimmung unterscheidbar sind. Die Frage dabei ist, innerhalb wel- cher systematischer Kategorien diese Annahme gultig ist. Wie BIANCHI (1967) darlegt, werden zufallige Ubereinstimmungen zwischen Individuen um so wahrscheinlicher, je weitcr entfernt diese in phylogenetischer Hinsicht voneinander sind. Dai3 der Gultig- keitsbereich der erwahnten Annahme, gemessen an der herkommlichen Systematik, nicht universe11 gleich ist, beweisen die in der Literatur bisher beschriebenen Ergebnisse. Er scheint von der speziellen Methode, der Tiergruppe und dem verwendeten Gewebe bzw. Stoffgemisch abzuhangen. Allerdings wird eine Gegenuberstellung der Ergebnisse dadurch problematisch, dal3 die Vergleichsmethoden der einzelnen Autoren unterschied- lich sind; darauf kann hier nicht naher eingegangen werden. Das entworfene Bild ist daher unter dieseni Vorbehalt zu beurteilen und kann zudem auch nicht vollstandig scin; aui3erdem wurden oft nur wenige Arten einer systematischen Gruppe untersucht. Der Obersichtlichkeit wegen seien verschiedene Resultate nach den oben erwahnten Gesichtspunkten verglichen, ehc die eigenen Ergebnisse diskutiert werden. Oberschnei- dungen der erwahnten drei Faktoren sind dabei nicht zu vermeiden.
Ergebnisse anderer Autoren
1. Einflufl der Methode
DESSAUER und Fox (1964) schliei3en aus Untersuchungen an Blutplasma-Proteinen und Hamoglobinen von Amphibien und Reptilien, dal3 Starkegel- und Polyacrylamidgel- Elektrophorese mit Vorteil auf der Populationsebene Verwendung finden, wahrend sich die Papierelektrophorese besser zur Charakterisierung groflerer taxonomischer Grup- pen eignet.
2. Einflafl der untersuchten Tiergruppe
Wie sehr selbst bei offenbar naher verwandten Arten die Anwendbarkeit der Elektro- phorese schwanken kann, zeigen zwei Untersuchungen an Rhizopoden. Wahrend REEVES und BISCHOFP (1968) unter Benutzung von Celluloseacetatfolien verschiedene Stfmnie von Entamoeba histolytica elektrophoretisch unterscheiden konnten, gelang es KATES und GOLDSTEIN (1964) mit der starker auflosenden Disk-Elektrophorese nicht, zwischen zwei Stammen von Amoeba proteus und einem von Amoeba discoides zu unterscheiden; wohl aber bot Chaos chaos ein deutlich anderes Bild. Konnten DAVIS und LINDSAY (1967) durch Disk-Elektrophorese von Fuflmuskelextrakten zwei Ga- stropodenarten unterscheiden und innerhalb einer Art Populationen trennen, so stell- ten WRIGHT und Ross (1966) bei Planorbiden mittels Elektrophorese der Eiproteine auf Celluloseacetatfolien eine hohe Variabilitat zwischen Populationen einzelner Arten fest, die nieist die interspezifischen Unterschiede uberlappten. SALKELD (1969) fand bei
A66. 21. Pherogramme und Gelphotographien von Dendrocoelopsis ezensis. a: Tier N.3 (8,4 mg), b: Tier N.4 (9,5 mg); die zugehorigen Photographien stammcn von Parallelproben.
Erlzutcrungen wie zu Abb. 7

a
E
1. 0 (a)
23 25 26 2728 29 30 332 3 5 9 11 13 15 17 20 I I I I I I I I I I I I I I I I
I I 1 I 1 I I I I I I
E 2.5
!
3 29 30 31 32 34
a
.1.0 b . c )
C b
b
b
C

Disk- Elektropboretiscbe Untersuchungen zur A rtcbarakterisierung paludicoler Tricladen 3 1 3
sieben Insektenordnungen rnit Hilfe der Polyacrylamidgel-Elektrophorese von Eiern artcharakteristische Proteinspektren und vermutet, dai3 diese phylogenetische Ver- wandtschaften wiedergeben konnten; allerdings zeigen sich zwischen drei Familien der Coleopteren keine sichtbaren Whnlichkeiten. Wahrend HEBARD (1964) und FLINDT et al. (1968) bei der Untersuchung von Amphibienseren mit der Papier- bzw. Cellulose- acetat-Elektrophorese Artspezifitat, doch keine Korrelationen zu taxonomischen Grup- pierungen feststellten, konnten bei Fischen mit verschiedenen Elektrophoresemethoden phylogenetische Aussagen auf den Ebenen von Art, Gattung und Familie gemacht wer- den (so z. B. SANDERS 1964, TSUYUKI et al. 1965 und 1966). SIBLEY (1960) gelang es, mit der Papierelektrophorese von Eiklarproteinen zahlreicher Vogelarten einen Beitrag zur Systematik zu leisten.
3. Einflup der untersuchten G e w e b e
BEDFORD und REID (1969) gelangten nach disk-elektrophoretischer Untersuchung der Kristallstiele von funf Bivalvier- und einer Gastropodenart zu einer mit der her- kommlichen Systematik identischen Artengruppierung, wobei keine okologischen Ein- flusse festzustellen waren. DAVIS und LINDSAY (1967), die eine sehr ahnliche Methode verwendeten, erhielten dagegen mit Hilfe von Fui3muskelhomogenaten bei Gastro- poden zwar artcharakteristische, aber nicht phylogenetisch relevante Bilder. Daraus schliei3en die erstgenannten Autoren, dai3 der Kristallstiel ein ,,konservatives Organ" im Sinne der Evolution darstellt. Xhnlich sind nach TSUYUKI et al. (1965) bei den un- tersuchten Fischen (Elasmobranchii, Holocephali, Teleostei) Muskelproteine dem Hamo- globin fur phylogenetische Betrachtungen vorzuziehen. Umfangreiche starkegel-elek- trophoretische Studien an Augenlinsenproteinen (SIBLEY und BRUSH 1967) sowie an Blutplasma von Vogeln (SIBLEY und HENDRICKSON 1970) weisen in eine ahnliche Rich- tung. Wahrend die Proteine der Augenlinsen in der ganzen Klasse ein ziemlich uni- formes Bild ergeben (was die Autoren freilich mit Vorbehalt kommentieren), sind die individuellen Schwankungen bei Plasmaproteinen so hoch, dai3 sich zwar ein gemein- sames Grundmuster, jedoch keine Korrelationen zu taxonomischen Gruppen aufstellen lassen. Beschreiben JOHNSON und WICKS (1 964) die Verwendbarkeit der Elektrophorese von Blutseren fur taxonomische Untersuchungen an Mammaliern, so gelangen BEDNA- ~ I K und CAJTHAMLOVA (1967) hinsichtlich der Milzproteine von Vertebraten (darunter auch Mammaliern) zum gegenteiligen Ergebnis.
Diskussion der eigenen Ergebnisse
Im Rahmen dieser Arbeit war es nicht moglich, aui3er den beschriebenen weitere Me- thoden zu erproben und zu vergleichen. Aus den oben erwahnten Resultaten ware je- doch zu fordern, bei jeweils derselben Art einmal verschiedene Elektrophoresemetho- den auf verschiedene Gewebs- oder Zelltypen anzuwenden, sodann die Auswertung und den Vergleich der Proteinspektren nach mehreren Methoden vorzunehmen, ehe man endgultige Schlusse uber den Wert ihrer Aussage zieht. Kann der Forderung nach Erprobung anderer Elektrophoresemethoden noch relativ leicht entsprochen werden, so ist es ohne umfangreiche Vorarbeit wohl kaum moglich, bei Turbellarien Gewebs- oder Zelltypen in ausreichender Menge zu isolieren. Am ehesten konnten noch Kokons iiber langere Zeit gesammelt und tiefgekuhlt aufbewahrt werden. Auch bei den zitier-
Abb. 22. Pherogramme und Gelphotographien von Bdellocepbala brunnea. a: Tier N.l (5,6 mg), b-c: Tier N.3 (18,9 mg), Parallelproben: b zentrifugiertes, c nicht zentrifugiertes Homo-
genat. Erlauterungcn wie zu Abb. 7

314 R. Peter
ten Arbeiten uber Trematoden (GEYER 1967, YOSHIMURA 1969a, b; s. auch LAMMLER 1969) sowie bei GYSELS’ (1968) Untersuchung an Nematoden gelangten Totalextrakte zur Anwendung. Vielleicht ergaben bestirnrnte Zellfraktionen, etwa Kerne oder Ribo- sornen, besser definierte Extrakte. Was die Methoden der Auswertung b e t r i a , so ist wohl eine Kombination von optischer Betrachtung, Photographie und densitomctrischer Registrierung optimal. Die als MeBdaten objektivierbaren Rv-Werte konnten dabei einem bloBen Gestaltvergleich von Abfolgen gefarbter Proteinzonen, wie er von vielen Autoren durchgefuhrt wurde, uberlegen sein. Zwar enthalten die Intensitaten und Breiten der Farbzonen zusatzliche Information und gewinnen als Ausdruck komplexer biochernischer Gegebenheiten besonders dann an Bedeutung, wenn man bedenkt, wie ofl Banden gleicher Position unterschiedliche Fraktionen reprasentieren konnen, doch ware dann zu erwagen, o b nicht ein Vergleich der densitometrischen Auswertungen mit mathematischen Methoden vorzuziehen ist. DaB in der vorliegenden Arbeit auf quan- titative Vergleiche der Pherograrnme verzichtet wurde, hat, wie fruher erwahnt, seinen Grund in der zu geringen Zahl von Einzelproben und der mangelnden Variation des Materials hinsichtlich definierbarer physiologischer oder okologischer Bedingungen. Es sol1 nicht bestritten werden, daB ein Ahnlichkeitsvergleich der densitometrisch erhal- tenen Kurven, etwa unter Zuhilfenahme eines Computers, wertvolle Aufschliisse liefern konnte, wie auch eine morphologische Beschreibung der Bandenmuster der reinen Cha- rakterisierung taxonomischer Gruppen dienen kann. Unter den gegebenen Bedingungen erschien es jedoch zweckmaBig, die Vergleiche auf die Rp-Werte zu beschranken. Die rneist eng gestaffelte Abfolge von Banden wurde ebenso wie die Variabilitat ihrer In- tensitaten einen rein morphologischen Vergleich zudem sehr schwer ermoglichen. Ein solcher durfte wohl bei Auftrennungen, deren Komponenten wenigstens teil weise auch funktionell definierbar sind, wie etwa bei Blutseren oder bei Hamoglobinen, eher durch- fuhrbar sein. Auch mag eine Reihung der elektrophoretischen Beweglichkeiten funk- tionsgleicher Proteine, so z. B. der Serumalbumine bei Saugern ( JOHNSON und WICKS 1964), phylogenetisch relevante Ergebnisse liefern. Es ist mit ein Nachteil der vorlie- genden und ahnlicher Arbeiten, dai3 die aufgetrennten Fraktionen nur durch relative elektrophoretische Beweglichkeit (RF-Wert) und Intensitat, nicht aber chemisch defi- niert sind; ihre weitere Analyse, etwa in zweidimensionalen Verfahren, konnte zu zu- satzlichen und besser fundierten Aussagen fuhren. Untersuchungen uber die enzymati- schen Aktivitaten bestimmter Fraktionen (Zymogramme) konnten ebenfalls weiteren AufschluB bringen. Sie wurden hier nur in einigen Fallen versuchsweise angewendet.
Wenn nach den dargelegten Argumenten ein endgultiger SchluB hinsichtlich der Ver- wendbarkeit der Disk-Elektrophorese fur die Darstellung von Artverwandtschaflen bei den beiden untersuchten Familien der Tricladen nicht zulassig erscheint, so durfte es doch ungeachtet des noch geringen Materials sehr wahrscheinlich sein, dai3 jede der untersuchten Arten ein von der GroBe und dem physiologischen Zustand der Tiere in bestirnmten Grenzen unabhangiges charakteristisches Spektrum extrahierbarer und disk-elektrophoretisch trennbarer Proteine besitzt und daB selbst geographisch einan- der nahe Populationen oder deren Untereinheiten auch dann unterscheidbar sind, wenn kein Unterschied in der Chromosomenzahl anzunehmen ist. Weitere disk-elektro- phoretische Untersuchungen an groBerem Material waren zusammen mit morphologi- schen und cytologischen Studien zur Kennzeichnung der Variabilitat innerhalb der Arten ebenso interessant wie bei der Untersuchung der Einflusse okologischer und phy- siologischer Faktoren.
Danksagung
An erster Stelle mochte ich meinen lieben Eltern danken, deren ideelle und materielle Unter- stiitzung die Durchfiihrung dieser Arbeit erst ermoglicht hat. Den Vorsranden des I. Zoologi- schen Institutes der Universitlt Wien, Herrn Prof. Dr. WILHELM MARINELLI und Herrn Prof.

Disk-Elektrophoretische Untersuchungen zur Artcharakterisierung paludicoler Tricladen 3 15
Dr. FRIEDRICH SCHALLER, bin ich fur die Arbeitsmoglichkeit, Herrn Prof. Dr. SCHALLER dar- uber hinaus fur die Forderung durch sein reges Interesse, wertvolle Anregungen und die An- schaffung von Geraten sehr zu Dank verpflichtet. Mein herzlicher Dank richtet sich auch an Herrn Prof. Dr. RUPERT RIEDL, derzeit an der University of North Carolina, Chapel Hill, U.S.A., der diese Untersuchung von ihrem Beginn an durch lebhafte Anteilnahme und inter- essante Diskussionen gefordert hat. Besonders danke ich Herrn Prof. Dr. MASAHARU KAWA- KATSU vom Fuji Women's College, Sapporo, und Herrn Dr. WATARU TESHIROGI von der Hiro- saki University, Hirosaki City, fur die Beschaffung und Determination des japanischen Mate- rials. Die Freshwater Biological Association, The Ferry House, Ambleside, sandte dankens- werterweise Material aus England. Ich mochte nicht versaumen, Fraulein HEDWIG KOCKO, Wien, fur wertvolle Hilfe beim Anfertigen der Abbildungen und meinem Freund GOTTPRIED TICHY, Salzburg, fur die Aufsammlung der Karntner Exemplare von Crenobia alpina herzlich zu danken.
Zusammenfassung
Sechs Arten aus der Familie Planariidae (Dugesia gonocephala, D. tigrina, D . lugubris; Creno- bia alpina; Polycelis felina; Phagocata vivida) und drei Arten der Familie Dendrocoelidae (Dendrocoelum lacteum, Dendrocoelopsis ezensis, Bdellocephala brunnea) lieflen sich durch Disk-Elektrophorese von Totalextrakten mit anschlieflender Proteinfarbung eindeutig charak- terisieren. Die erwahnte Methode erwies sich hierbei als der Polyacrylamid-Elektrophorese mit nur einer Gelschicht uberlegen und erbrachte gut reproduzierbare Ergebnisse. Obwohl die Proteinspektren Unterschiede zwischen Individuen und Populationen ergaben, weist jede Art ein charakteristisches, stets vorhandenes Minimalspektrum auf, zu dem - teils methodisch be- dingt - mehr oder weniger Fraktionen hinzutreten konnen. Die erhaltenen Muster wurden nach densitometrischer Auswertung durch die relativen elektrophoretischen Beweglichkeiten (,RF-Werte") ihrer Fraktionen gekennzeichnet und miteinander verglichen. Dabei ergaben sich hohere Ahnlichkeiten zwischen Populationen einer Art als zwischen verschiedenen Arten. Die unterschiedlichen interspezifischen Ubereinstimmungen lieBen sich nicht zur taxonomischen Gruppierung heranziehen. Um das fur einen bestimmten Fundort charakteristishe Protein- spektrum zu ermitteln, muflten mehrere Tiere untersucht werden. Die beschriebene Methode scheint sich zur Kennzeichnung von Fundorten auch geringer geographischer Distanz zu eig- nen, wie die Ergebnisse bei Dugesia gonocephala nahelegen. Weiter voneinander entfernte Populationen von Polycelis felina zeigten demgegenuber geringere Ubereinstimmungen, die nur wenig uber den hochsten beobachteten interspezifischen Ahnlichkeiten lagen; dieses Re- sultat steht im Einklang mit den zahlreichen cytogenetisch definierten taxonomischen Gruppen innerhalb der genannten Art. Die erzielten Ergebnisse werden im Zusammenhang mit den Aussagen einiger ahnlich orientierter Untersuchungen anderer Autoren an anderen Tiergruppen diskutiert. Wahrend die verwendete Methode bei aludicolen Tricladen fur feinere infra- spezifische Differenzierungen geeignet erscheint, mu{ eine endgiiltige Aussage uber ihre Ver- wendbarkeit zur Klarung von Verwandtschaftsverhaltnissen uber die Art hinausgehender taxo- nomischer Einheiten umfangreicheren und auch methodisch reichhaltigeren Untersuchungen vorbehalten bleiben.
Summary
Disc-electrophoretic Studies on the Characterization of Species of Paludicolous Triclads (Platyhelminthes: Turbellaria)
Six species from the family Planariidae (Dugesia gonocephala, D. tigrina, D. lugubris; Creno- bia alpina; Polycelis felina; Phagocata vivida) and three species belonging to the family Den- drocoelidae (Dendrocoelum lacteum, Dendrocoelopsis ezensis, Bdellocephala brunnea) could be characterized distinctly by disc-electrophoresis of whole body extracts followed by protein staining. The mentioned method roved superior to polyacrylamide electrophoresis using a single gel layer and yielded high& reproducible results. Individuals as well as populations could be distinguished by their protein spectra. However, each species shows a characteristic minimum spectrum recognizable in all experiments which is, partly because of methodical reasons, superimposed by a variable number of additional fractions. The obtained patterns were evaluated densitometrically and characterized by the relative electrophoretic mobilities ('I&-values") of their fractions. Comparisons between different populations of a given species yielded higher degrees of similarity than those between different species. T h e varying inter- specific correspondances could not be used for taxonomical grouping. To obtain the protein spectrum characteristic of a locality, at least a few animals had to be studied. The described method appears to be applicable to the characterization of localities even close to each other, as the results with Dugesia gonocephala indicate. Populations of Polycelis felina more distant

316 R. Peter
from each other showed, in contrast, lesser similarity ranging only a little above the highest interspecific similarities observed; this result is in accordance with the fact that numerous taxo- nomic subunits may be distinguished cytogenetically within the mentioned species. The ob- tained results are compared with those of similar studies made by different authors on other taxonomic groups. While infraspecific distinctions of paludicolous triclads may be made with good success using the applied method, more extensive investigations with a variety of methods seem to be necessary for a definitive conclusion concerning the applicability of electrophoresis to elucidating relationships above the species level.
Resume
Recherches dectrophorktiques au sujet de la caractkrisation d’espPces de Triclades paludicoles (Platyhelminthes: Turbellaria)
Six espices de la famille Planariidae (Dugesia gonocephala, D . tigrina, D . lugubris; Creno- bia alpina; Polycelis felina; Phagocata vivida) et trois espices de la famille Dendrocoelidae (Dendrocoelum lacteum, Dendrocoelopsis ezensis, Bdellocephala brunnea) ont CtC caracterisbes distinctement par l’klectrophorise en gel de polyacrylamide suivant la technique d’ORNSTEIN et DAVIS qu’utilise un systirne discontinu de deux couches de gel. Cette mkthode, appliquCe B extraits d’animaux totals et suivie par une coloration des protkines, s’est montrke plus efficace que I’Clectrophorkse en gel de polyacrylamide n’usant qu’une couche de gel, et les rCsultats pou- vaient btre facilement reproduits. Quoique les protkinogrammes rkvilent des diffkrences entre les individues et les populations, chaque espice montre un spectre minimum caractkristique et toujours prksent, auquel s’ajoute un nombre variable de fractions, due parfois B la mkthode. Ces protkinogrammes, classCes par densitomktrie en vue des mobilitks ClectrophorCtiques rela- tives (les valeurs RF) de leurs fractions, ont C t k comparkes entre elles. Les resemblances sont plus marqukes entre les populations d’une espice qu’entre espices diffCrentes. Les divers accords interspbcifiques ne peuvent pas btre utilisCs en faveur d’un groupement taxonomique. La mise en Cvidence d’un spectre caractiristique de prodines pour une mbme localitk a nkcessitk 1’Ctude de plusieurs animaux. La mkthode dCcrite est valable pour caractkriser des localitCs meme peu distantes, comme il en ressort d’aprks les donnCes de Dugesia gonocephala. Les populations de Polycelis felina, venantes de localitCs plus distantes l’une de l’autre que celles d’avant, pos- sident moins d’accords, se situant un peu au-dessus du niveau des plus hautes ressemblances interspbcifiques. Ce rCsultat correspond au fait qu’on peut diviser l’espice citCe en nombreuses groupes cytotaxonomiques. Les rksultats sont comparks avec ceux des travaux analogues d’au- tres auteurs traltant d’autres groups d’animaux. Tandis que la mkthode dkcrite pour ce qui con- cerne Triclades paludicoles peut rCvkler distinctement des differences infraspkcifiques, on ne peut pas encore dire dkfinirivement si elle rCussit B Ctablir d’affinids au-dessus du niveau d’espices.
Literatur
ADMIRAAL, D. J. H., 1953: A stabilized D. C. supply unit for output voltages of 0-300 V. Elec- tronic Application Bulletin 14, 61-71. N. V. Philips’ Gloeilampenfabrieken, Eindhoven.
BEDFORD, J. J., and REID, M. S., 1969: Gel electrophoresis of proteins in the crystalline styles of 6 species of certain Mollusca. Comp. Biochem. Physiol. 29, 659-664.
BEDNAKIK,T, and CAJTHAMLOVA, H., 1967: Spectra of soluble proteins from the spleen of some vertebrates. Physiol. Bohemoslov. 16, 78-88.
BIANCHI, U., 1967: Utilith, significato e limiti dell’elettroforesi applicata a particolari proble- mi tassonomici. Riv. Biol. (Perugia) 60, 227-236.
BOHMIG, L., 1909: Tricladida. In: BRAUER, Die SiiBwasserfauna Deutschlands, Heft 19, 143-176. BURNS, D. A., and POLLAK, 0. J., 1963: Permanent record and scanning of disc electropho-
resis patterns. J. Chromatogr. 11, 559-562. CLARKE, J. T., 1964: Simplified ,,disc“ (polyacrylamide gel) electrophoresis. Ann. N. Y. Acad.
Sci. 121, 428-436. DAHM, A. G., 1958: Taxonomy and ecology of five species groups in the family Planariidac.
Nya Litografen, Malmo. DAVIS, B. J., 1962: Disc electrophoresis - Part 11: Materials and methods. Preprinted by Di-
stillation Products Industries (Eastman Kodak), Rochester. - 1964: Disc electrophoresis - 11. Method and application to human serum proteins. Ann. - -
N. Y. Acad. Sci. l i l , 404-427. DAVIS. G. M.. and LINDSAY, G. K., 1967: Disc electrophoretic analysis of molluscan indivi-
duals and populations. Malacologia 5, 31 1-334.

Disk-Elektrophoretische Untersuchungen zur Artcharakterisierung paludicoler Tricladen 3 1 7
DESSAUER, H. C., and Fox, W., 1964: Electrophoresis in taxonomic studies illustrated by ana- lyses of blood proteins. In: Taxonomic Biochemistry and Serology, 625-647.
DOUNCE, A. L., 1955: The isolation and composition of cell nuclei and nucleoli. In: The Nu- cleic Acids (ed. by E. CHARGAFF and J. N. DAVIDSON), 2, 93-153. Academic Press, New York, London.
FLINDT, R., HEMMER, H., und JAEGER, R., 1968: Das Serumeiweinbild mitteleuropaischer Anu- ren. 2001. Jb. Physiol. 74, 155-163.
FLORKIN, M., 1966: Aspects molCculaires de I’adaptation et de la phyloginie (Les grands pro- blPmes de la biologie, Tome 2) . Masson et Cie., Paris.
GEYER. E.. 1967: Elektrouhoretische Analvsen an Fasciola heoatica-Totalextrakten. 2. Para- sitenk. 28 , 352-366. &
GYSELS. H.. 1968: Electroohoretic observations on the Drotein comoosition of free-living and , I
plant-parasitic nematodes, with a special reference to some components showing a digistive activity. Nematologica 14, 489-496.
HEBARD, W. B., 1964: Serum protein electrophoretic patterns of the Amphibia. In : Taxonomic Biochemistry and Serology, 649-657.
JOHNSON, B. L., BARNHART, D., and HALL, O., 1967: Analysis of genome and species rela- tionships in the polyploid wheats by protein electrophoresis. Amer. J. Bot. 54, 1089-1098.
JOHNSON, M. L., and WICKS, M., 1964: Serum-protein electrophoresis in mammals: significance in the high taxonomic categories. In : Taxonomic Biochemistry and Serology, 681-694.
KATES, J. R., and GOLDSTEIN, L., 1964: A comparison of the protein composition of three species of Amoebae. J. Protozool. 11, 30-35.
KAWAKATSU, M., 1969: An illustrated list of Japanese freshwater planarians in color. Bull. Fuji Women’s College 7, Ser. 11, 45-91.
KAWAKATSU, M., TESHIROGI, W., and YAGIHASHI, M., 1967: Report on the ecological survey of freshwater planarians in the western part of Aomori Prefecture (Hirosaki City, The Mt. Iwaki and The JQniko Districts), HonshG. Jap. J. Ecol. 17, 24-41.
KAWAKATSU, M., YAMADA, T., and IWAKI, S., 1967: Environment and reproduction in Japanese freshwater planarians. Jap. J. Ecol. 17, 263-266.
KONIG, P., 1937: Employment of electrophoresis in chemical experiments with small quan- tities. Actas e Trabalhos do Terceiro Congr. Sud-Amer. Chim. 2, 334.
KUEHN, G., 1938: Zur Ukologie und Biologie der Gewasser (Quellen und Abfliisse) des Was- sergsprengs bei Wien. Diss. Phil. Fak. Univ. Wien.
LAMMLER, G., 1969: Analytische Untersuchungen an Litomosoides carinii-Extrakten. 2. Para- sitenk. 32, 254-265.
LEONE, C. A. (Hsg.), 1964: s. Taxonomic Biochemistry and Serology. LOENING, U. E., 1967: The fractionation of high-molecular-weight ribonucleic acid by poly-
acrylamide-gel electrophoresis. Biochem. J. 102, 251-257. MAURER, H. R., 1968: Disk-Elektrophorese. Theorie und Praxis der diskontinuierlichen Poly-
acrylamidgel-Elektrophorese. De Gruyter, Berlin. MEIXNER, J., 1915: Zur Turbellarienfauna der Ostalpen, insonderheit des Lunzer Seengebietes.
Zool. Jb. Abt. Syst. 38, 459-588. NAGAI, Y., GROSS, J., and PIEZ, K. A., 1964: Disc electrophoresis of collagen components. Ann.
N. Y. Acad. Sci. 121,494-500. NIRENBERG, M. W., MATTHAEI, J. H., JONES, 0. W., MARTIN, R. G., and BARONDES, S. H., 1963:
Approximation of genetic code via cell-free protein synthesis directed by template RNA. Fed. Proc. 22, 55-61.
NUTTALL, G. H. F., 1904: Blood immunity and blood relationships. Cambridge University Press, Cambridge.
ORNSTEIN, L., 1962: Disc electrophoresis - Part I : Theory. Preprinted by Distillation Pro- ducts Industries (Eastman Kodak), Rochester.
- 1964: Disc electrophoresis - I. Background and theory. Ann. N. Y. Acad. Sci. 121, 321-349. OSBORNE, P. J., and MILLER, A. T., jr., 1962: Uptake and intracellular digestion of protein
(peroxidase) in planarians. Biol. Bull. (Woods Hole) 123, 589-596. PICKAVANCE, J. R., 1970: A new approach to the immunological analysis of invertebrate diets.
J. Anim. Ecol. 39, 715-724. RAYMOND, S., 1964: Acrylamide gel electrophoresis. Ann. N. Y. Acad. Sci. 121, 350-365. RAYMOND, S., and WANG, Y., 1960: Preparation and properties of acrylamide gel for use in
RAYMOND, S., and WEINTRAUB, L., 1959: Acrylamide gel as a supporting medium for zone
REEVES, R. E., and BISCHOFF, J. M., 1968: Classification of Entamoeba species by means of
REYNOLDSON, T. B., 1967: A key to the British species of freshwater Triclads. Freshwater Biolo-
electrophoresis. Anal. Biochem. 1, 391-396.
electrophoresis. Science (Washington) 130, 71 1 .
electrophoretic properties of ameba1 enzymes. J. Parasitol. 54, 594-600.
gical Association, Scientific Publication No. 23.

318 R. Peter
REYNOLDSON, T. B., and DAVIES, R. W., 1970: Food niche and co-existence in lake-dwelling Triclads. J. Anim. Ecol. 39, 599-627.
RITCHIE, R. F., HARTER, J. G., and BAYLES, T. B., 1966: Refinements of acrylamide electro- phoresis. J. Lab. Clin. Med. 68, 842-850.
SALKELD, E. H., 1969: Electrophoretic patterns of egg proteins from several insect taxa. Can. Entomol. 101, 1256-1265.
SANDERS, B. G., 1964: Electrophoretic studies of serum proteins of three trout species and the resultine hvbrids within the familv Salmonidae. In: Taxonomic Biochemistrv and Scroloev.
" I ,
New Y&k: 673-679. SCHRAUWEN. T.. 1966: Nachweis von Enzvmen nach elektroohoretischer Trennune an Polv-
acrylamibrSaulchen. J. Chromatogr. 23, '1 77-180. SIBLEY, C. G., 1960: The electrophoretic patterns of avian egg-white proteins as taxonomic
characters. Ibis 102, 215-284. - 1962: The comparative morphology of protein molecules as data for classification. Syst.
2001. 11, 108-118. SIBLEY, C. G., and BRUSH, A. H., 1967: An electrophoretic study of avian eye-lens proteins.
Auk 84,203-219. SIBLEY, C. G., and HENDRICKSON, H. T., 1970: A comparative electrophoretic study of avian
plasma proteins. Condor 72,43-49. STEINMANN, P., und BRESSLAU, E., 191 3 : Die Strudelwiirmer (Turbellaria) (Monographien ein-
heimischer Tiere, Bd. 5). Klinkhardt, Leipzig. Taxonomic Biochemistry and Serology, 1964: Edited by C. A. LEONE. The Ronald Press Com-
pany, New York. TSUYUKI, H., ROBERTS, E., and VANSTONE, W. E., 1965: Comparative zone electropherograms
of muscle myogens and blood hemoglobins of marine and freshwater vertebrates and their application to biochemical systematics. J. Fish. Res. Board Can. 22, 203-213.
TSUYUKI, H., UTHE, J. F., ROBERTS, E., and CLARKE, L. W., 1966: Comparative electrophero- grams of Coregonus clupeaformis, Salvelinus namaycush, S . alpinus, S . rnalma and S . fonti- nalis from the family Salmonidae. J. Fish. Res. Board Can. 23, 1599-1606.
"
WEBER, E., 1967: GrundriB der Biologischen Statistik. 6. Aufl. Fisher, Jena. WHITE, M. L., 1960: The permeability of an acrylamide polymer gel. J. Phys. Chem. 64, 1563
WIEME, R. J., 1962: An integrated procedure for acrylamide gel electrophoresis. In: Protides of the Biological Fluids, Proc. of the 10th Colloquium, Bruges 1962, 309-311. Elsevier, Am- sterdam, London, New York, 1963.
- 1964: A procedure for high voltage electrophoresis in agar gel, with a note on its appli- cation to acrylamide and starch gel. Ann. N. Y. Acad. Sci. 121, 366-372.
WRIGHT, C. A., and Ross, G. C., 1966: Electrophoretic studies on planorbid egg proteins. World Health Organ. Bull. 35, 727-731.
YANOFSKY, C., HENNING, U., HELSINKI, D., and CARLTON, B., 1963: Mutational alteration of protein structure. Fed. Proc. 22, 75-79.
YOSHIMURA, K., 1969a: Paragonimus: Electrophoretic fractionation of whole body proteins as an aid in specific identification of a species from Sado Island, Japan. Exp. Parasitol. 25,
- 1969b: Paragonimus westermani, P. ohirai and P. rniyazakii: Electrophoretic comparison
-1565.
107-117.
of whole body proteins. Exp. Parasitol. 25, 118-130.
Anschrifi des Verfassers: Dr. ROLAND PETER, I. Zoologisches Institut der Universitat Wien, Dr.-Karl-Lueger-Ring 1, A-1010 Wien, Usterreich