Embed Size (px)
Citation preview

Journal of the Peripheral Nervous System 18:168–176 (2013)
RESEARCH REPORT
Disruption of nodal architecture in skin biopsiesof patients with demyelinating neuropathies
Kathrin Doppler, Christian Werner, and Claudia Sommer
Department of Neurology, University of Wurzburg, Wurzburg, Germany
Abstract Cutaneous nerves represent the most distal part of the sensory nervoussystem. We took advantage of the good discernibility of longitudinal myelinated fibers inskin biopsies to analyze the distribution of nodal and paranodal proteins in neuropathiesand to assess nodal disorganization as a diagnostic marker of demyelinating neuropathy(NP). We analyzed myelinated nerve fibers in skin biopsies from the finger and the proximalleg of 52 prospectively recruited patients with different peripheral neuropathies and 17controls. We performed immunohistochemical double labeling with anti-MBP, anti-PGP9.5,anti-caspr, anti-pan-neurofascin, and anti-pan-sodium-channel. Three potential features ofdemyelinating NP could be established: elongated nodes of Ranvier and dispersion ofcontactin-associated protein (caspr) staining were found more often in demyelinating thanin axonal neuropathies (p < 0.05) and were not detectable in normal controls. Broadening ofneurofascin staining was detectable more often in demyelinating neuropathies comparedwith normal controls (p < 0.05). Our data suggest that pathological changes of nodalarchitecture can be visualized in skin biopsies and that the detection of elongated nodesof Ranvier and alterations in the distribution of paranodal proteins may be useful in thediagnostic assessment of peripheral NP.
Key words: demyelination, node of Ranvier, peripheral neuropathy, skin biopsy
IntroductionNodes of Ranvier are important sites of disease
initiation in demyelinating disorders. Myelinatednerve fibers consist of nodes of Ranvier, paranodalregions, juxtaparanodal regions, and internodes(Dupree et al., 1999). Each compartment containscharacteristic proteins that are constituents of eitherthe axolemma or the myelin sheath (Susuki andRasband, 2008). Nodes are characterized by voltage-gated sodium channels, ankyrin G, and by neurofascin-186 (Black et al., 1989; Arroyo and Scherer, 2000;Sherman et al., 2005), paranodes by neurofascin-155
Address correspondence to: Dr. Kathrin Doppler, Neurologi-sche Klinik, Universitatsklinikum Wurzburg, Josef-Schneider-Str.11, 97080 Wurzburg, Germany. Tel: 004993120123641; Fax:004993120123697; E-mail: [email protected]
and contactin-associated protein (caspr), and thejuxtaparanodal regions by potassium channels andcontactin-2 (Arroyo and Scherer, 2000; Scherer andArroyo, 2002; Traka et al., 2003; Sherman et al.,2005). Demyelination leads to a disruption of thischaracteristic architecture (Arroyo et al., 2004; Liet al., 2005; Coman et al., 2006; Howell et al.,2006; Lonigro and Devaux, 2009; Cifuentes-Diaz et al.,2011). Information on the organization of paranodaland nodal proteins in pathologic conditions of thehuman peripheral nervous system (PNS) is limited,because nerve biopsy is an invasive technique and theexamination of nodes requires longitudinal sectionsor teased fiber preparations. Skin biopsy has beenused as a minimally invasive tool for the morphologicalassessment of myelinated nerve fibers (Nolano et al.,2003; Provitera et al., 2007; Saporta et al., 2009;Doppler et al., 2012). We used skin punch biopsy
© 2013 Peripheral Nerve Society 168

Doppler et al. Journal of the Peripheral Nervous System 18:168–176 (2013)
samples from the index finger and from the upperleg to identify changes in nodal length and in thedistribution of nodal and paranodal proteins in glabrousand hairy skin. We describe the normal appearance ofnodal architecture in skin in the largest thus far reportedgroup of human controls, and explore the potentialutility of the analysis of dermal Ranvier nodes forthe differential diagnosis of axonal and demyelinatingneuropathy (NP).
Patients and MethodsSubjects
The study was approved by the ethics committeeof the Medical Faculty of the University of Wurzburgand was performed in accordance with the ethicalstandards of the Declaration of Helsinki of 1964. Allpatients and controls gave written informed consentto take part in the study.
Between 2010 and 2012, 110 patients whoattended the Department of Neurology of the Uni-versity of Wurzburg for diagnostic work-up of sus-pected peripheral NP were prospectively recruited.Sixty-five patients were asked to take part in thisstudy, 59 patients gave written informed consent, andskin biopsy was performed. Additionally, 33 healthypersons were asked, resulting in the recruitment of 17normal controls. Seven patients were excluded afterobtaining skin biopsy because their biopsies did notreach the dermis and could therefore not be evaluated.Complete data could be collected from the remaining52 patients and all 17 controls.
A neurological evaluation and nerve conductionstudies were performed in all cases. Thirty-sevenpatients obtained sural nerve biopsy. Biopsies wereclassified as demyelinating (n = 21) if onion bulbs,naked fibers, or >20% thinly myelinated fibers weredetectable and as axonal (n = 16) if there were no signsof demyelination, but clusters of regenerating fibers,severe axonal loss, or Wallerian degeneration. Nerveconduction studies were categorized as demyelinatingif they showed features of demyelination according tothe INCAT criteria (Hughes et al., 2001). On the basisof the results of nerve biopsy and nerve conductionstudies, 21 patients were classified as demyelinatingNP and 21 patients as axonal NP. In 10 cases, thefindings in electrophysiology and nerve biopsy werenot congruent or did not result in a clear diagnosis ofdemyelinating or axonal NP. Those cases, in analogy tohereditary NP, were classified as ‘‘intermediate NP’’(Reilly, 2007). Seventeen patients did not have anysymptoms of the upper limbs and had normal nerveconduction studies of the median and ulnar nerve. Fivepatients had only motor symptoms. Etiologic work-up
of all patients resulted in the diagnoses of chronicpolyneuritis (15), including 5 cases of chronic inflam-matory demyelinating polyradiculoneuropathy (CIDP)according to the INCAT criteria, 1 case of multifo-cal acquired demyelinating sensory and motor NP(MADSAM), and 9 cases of clinically suspected CIDP,non-systemic vasculitic NP (5), diabetic NP (4, type IIdiabetes), hereditary NP (2, 1 adrenomyeloneuropathyand 1 Charcot-Marie-Tooth disease [CMT] with hyper-trophic nerve roots), NP associated with motoneurondisease (2), NP with monoclonal gammopathy (1, IgA),and amyloid NP (1, associated with monoclonal gam-mopathy). The origin remained unknown in 23 cases.One of the diabetic NP and three of the NP of unknownetiology were small fiber neuropathies. Controls(n = 17) were included in this study if they did not showsymptoms of NP and had normal intraepidermal nervefiber density (IENFD) in the proximal thigh comparedwith an age-matched sample from our skin biopsy lab-oratory (n = 49, mean age 55 ± 14; IENFD 13.9 ± 3.8).
Skin biopsy and immunohistochemistry
Skin biopsies were taken from the proximalthigh using a 5-mm disposable punch (Stiefel GmbH,Offenbach, Germany) and from the glabrous skin ofthe lateral index finger between the interphalangealand palmophalangeal joints using a 3-mm punch,as previously described (Li et al., 2005; Uceyleret al., 2010). All biopsies were taken under localanesthesia. After fixation in 4% paraformaldehyde,immunohistochemical double labeling was performedon 50-μm frozen sections using anti-PGP9.5 (1:1,000,Ultraclone, Isle of Wight, UK) as an axonal marker, anti-MBP (mouse monoclonal, 1:800, Ultraclone and rabbitpolyclonal, 1:200, Millipore, Billerica, Massachusetts)as a marker for myelin, anti-pan-neurofascin (1:400,Abcam, Cambridge, UK), anti-caspr (1:100, Abcam),and anti-pan-sodium channels (1:100, Sigma, St.Louis, Missouri) using the free-floating method withfluorescent secondary antibodies (Dylight, Cy3, 1:200,Dianova, Hamburg, Germany), as described previously(Saporta et al., 2009; Doppler et al., 2012).
Fluorescence microscopy
The sections were analyzed using a fluorescentmicroscope (Zeiss Axiophot 2) with an AxiocamMRm camera (Zeiss, Oberkochen, Germany) andSPOT software (Diagnostic Instruments, Inc., SterlingHeights, Michigan), a confocal microscope Leica TCSSP2, and a fluorescent microscope (Zeiss Ax10) withCARVII-system and Visiview software (Visitron GmbH,Puchheim, Germany). Images were taken with theconfocal microscope and the fluorescent microscopewith CARVII-system. The examiner was blinded for theclinical or histological diagnosis of the biopsies.
169
Doppler et al. Journal of the Peripheral Nervous System 18:168–176 (2013)
(A) (B)
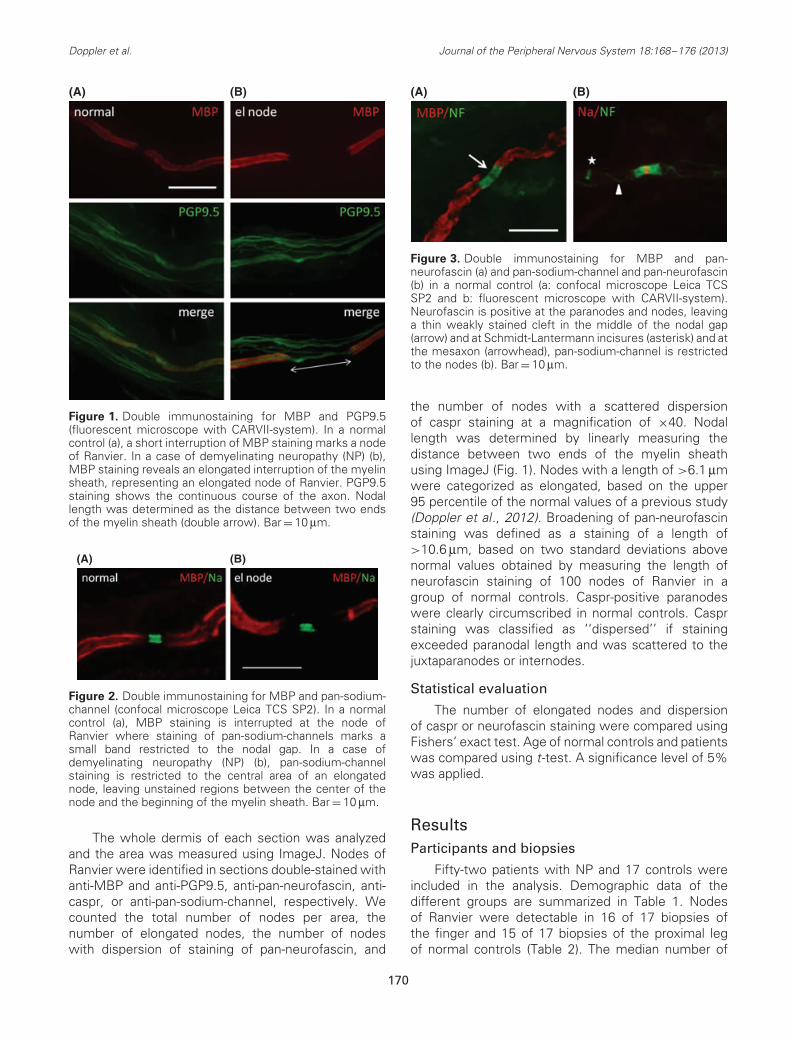
Figure 1. Double immunostaining for MBP and PGP9.5(fluorescent microscope with CARVII-system). In a normalcontrol (a), a short interruption of MBP staining marks a nodeof Ranvier. In a case of demyelinating neuropathy (NP) (b),MBP staining reveals an elongated interruption of the myelinsheath, representing an elongated node of Ranvier. PGP9.5staining shows the continuous course of the axon. Nodallength was determined as the distance between two endsof the myelin sheath (double arrow). Bar = 10 μm.
(A) (B)
Figure 2. Double immunostaining for MBP and pan-sodium-channel (confocal microscope Leica TCS SP2). In a normalcontrol (a), MBP staining is interrupted at the node ofRanvier where staining of pan-sodium-channels marks asmall band restricted to the nodal gap. In a case ofdemyelinating neuropathy (NP) (b), pan-sodium-channelstaining is restricted to the central area of an elongatednode, leaving unstained regions between the center of thenode and the beginning of the myelin sheath. Bar = 10 μm.
The whole dermis of each section was analyzedand the area was measured using ImageJ. Nodes ofRanvier were identified in sections double-stained withanti-MBP and anti-PGP9.5, anti-pan-neurofascin, anti-caspr, or anti-pan-sodium-channel, respectively. Wecounted the total number of nodes per area, thenumber of elongated nodes, the number of nodeswith dispersion of staining of pan-neurofascin, and
(A) (B)
Figure 3. Double immunostaining for MBP and pan-neurofascin (a) and pan-sodium-channel and pan-neurofascin(b) in a normal control (a: confocal microscope Leica TCSSP2 and b: fluorescent microscope with CARVII-system).Neurofascin is positive at the paranodes and nodes, leavinga thin weakly stained cleft in the middle of the nodal gap(arrow) and at Schmidt-Lantermann incisures (asterisk) and atthe mesaxon (arrowhead), pan-sodium-channel is restrictedto the nodes (b). Bar = 10 μm.
the number of nodes with a scattered dispersionof caspr staining at a magnification of ×40. Nodallength was determined by linearly measuring thedistance between two ends of the myelin sheathusing ImageJ (Fig. 1). Nodes with a length of >6.1 μmwere categorized as elongated, based on the upper95 percentile of the normal values of a previous study(Doppler et al., 2012). Broadening of pan-neurofascinstaining was defined as a staining of a length of>10.6 μm, based on two standard deviations abovenormal values obtained by measuring the length ofneurofascin staining of 100 nodes of Ranvier in agroup of normal controls. Caspr-positive paranodeswere clearly circumscribed in normal controls. Casprstaining was classified as ‘‘dispersed’’ if stainingexceeded paranodal length and was scattered to thejuxtaparanodes or internodes.
Statistical evaluation
The number of elongated nodes and dispersionof caspr or neurofascin staining were compared usingFishers’ exact test. Age of normal controls and patientswas compared using t-test. A significance level of 5%was applied.
ResultsParticipants and biopsies
Fifty-two patients with NP and 17 controls wereincluded in the analysis. Demographic data of thedifferent groups are summarized in Table 1. Nodesof Ranvier were detectable in 16 of 17 biopsies ofthe finger and 15 of 17 biopsies of the proximal legof normal controls (Table 2). The median number of
170
Doppler et al. Journal of the Peripheral Nervous System 18:168–176 (2013)
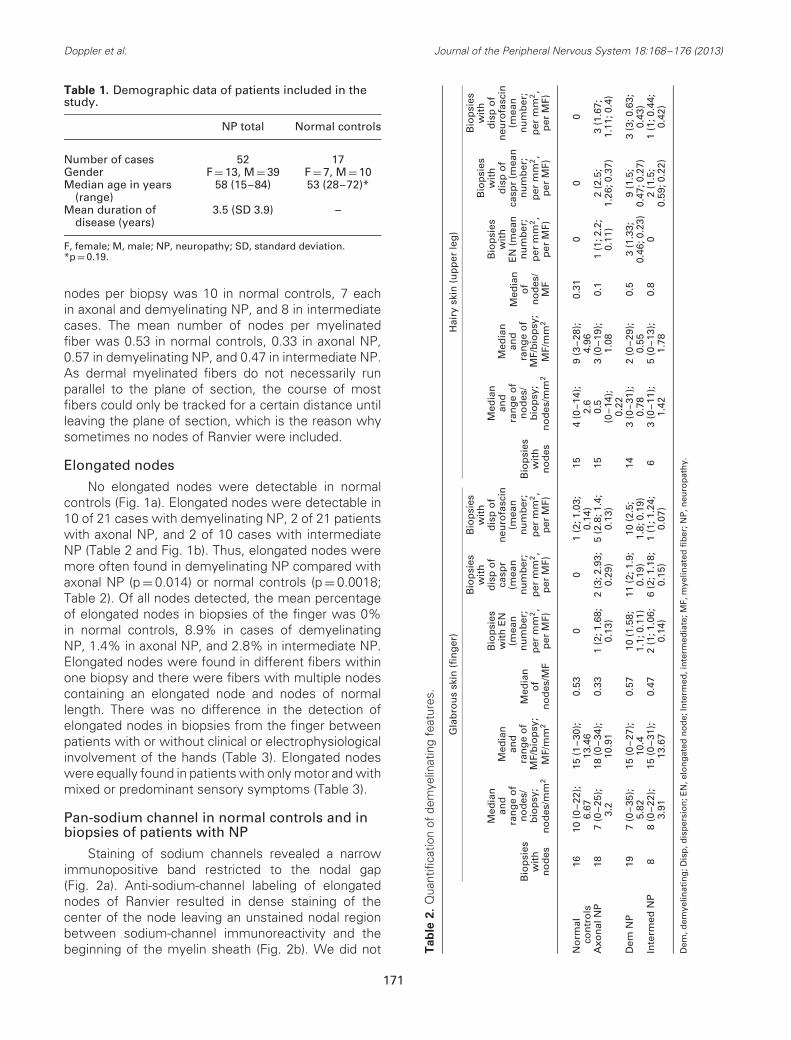
Table 1. Demographic data of patients included in thestudy.
NP total Normal controls
Number of cases 52 17Gender F = 13, M = 39 F = 7, M = 10Median age in years
(range)58 (15–84) 53 (28–72)*
Mean duration ofdisease (years)
3.5 (SD 3.9) –
F, female; M, male; NP, neuropathy; SD, standard deviation.*p = 0.19.
nodes per biopsy was 10 in normal controls, 7 eachin axonal and demyelinating NP, and 8 in intermediatecases. The mean number of nodes per myelinatedfiber was 0.53 in normal controls, 0.33 in axonal NP,0.57 in demyelinating NP, and 0.47 in intermediate NP.As dermal myelinated fibers do not necessarily runparallel to the plane of section, the course of mostfibers could only be tracked for a certain distance untilleaving the plane of section, which is the reason whysometimes no nodes of Ranvier were included.
Elongated nodes
No elongated nodes were detectable in normalcontrols (Fig. 1a). Elongated nodes were detectable in10 of 21 cases with demyelinating NP, 2 of 21 patientswith axonal NP, and 2 of 10 cases with intermediateNP (Table 2 and Fig. 1b). Thus, elongated nodes weremore often found in demyelinating NP compared withaxonal NP (p = 0.014) or normal controls (p = 0.0018;Table 2). Of all nodes detected, the mean percentageof elongated nodes in biopsies of the finger was 0%in normal controls, 8.9% in cases of demyelinatingNP, 1.4% in axonal NP, and 2.8% in intermediate NP.Elongated nodes were found in different fibers withinone biopsy and there were fibers with multiple nodescontaining an elongated node and nodes of normallength. There was no difference in the detection ofelongated nodes in biopsies from the finger betweenpatients with or without clinical or electrophysiologicalinvolvement of the hands (Table 3). Elongated nodeswere equally found in patients with only motor and withmixed or predominant sensory symptoms (Table 3).
Pan-sodium channel in normal controls and inbiopsies of patients with NP
Staining of sodium channels revealed a narrowimmunopositive band restricted to the nodal gap(Fig. 2a). Anti-sodium-channel labeling of elongatednodes of Ranvier resulted in dense staining of thecenter of the node leaving an unstained nodal regionbetween sodium-channel immunoreactivity and thebeginning of the myelin sheath (Fig. 2b). We did not T
ab
le2
.Q
uant
ifica
tion
ofde
mye
linat
ing
feat
ures
.
Gla
bro
us
skin
(fin
ger
)H
airy
skin
(up
per
leg
)
Bio
psi
esw
ith
no
des
Med
ian
and
ran
ge
of
no
des
/b
iop
sy;
no
des
/mm
2
Med
ian
and
ran
ge
of
MF/
bio
psy
;M
F/m
m2
Med
ian
of
no
des
/MF
Bio
psi
esw
ith
EN
(mea
nn
um
ber
;p
erm
m2,
per
MF)
Bio
psi
esw
ith
dis
po
fca
spr
(mea
nn
um
ber
;p
erm
m2,
per
MF)
Bio
psi
esw
ith
dis
po
fn
euro
fasc
in(m
ean
nu
mb
er;
per
mm
2,
per
MF)
Bio
psi
esw
ith
no
des
Med
ian
and
ran
ge
of
no
des
/b
iop
sy;
no
des
/mm
2
Med
ian
and
ran
ge
of
MF/
bio
psy
;M
F/m
m2
Med
ian
of
no
des
/M
F
Bio
psi
esw
ith
EN
(mea
nn
um
ber
;p
erm
m2,
per
MF)
Bio
psi
esw
ith
dis
po
fca
spr
(mea
nn
um
ber
;p
erm
m2,
per
MF)
Bio
psi
esw
ith
dis
po
fn
euro
fasc
in(m
ean
nu
mb
er;
per
mm
2,
per
MF)
No
rmal
con
tro
ls16
10(0
–22)
;6.
6715
(1–3
0);
13.4
60.
530
01
(2;1
.03;
0.14
)15
4(0
–14)
;2.
69
(3–2
8);
4.96
0.31
00
0
Axo
nal
NP
187
(0–2
5);
3.2
18(0
–34)
;10
.91
0.33
1(2
;1.6
8;0.
13)
2(3
;2.9
3;0.
29)
5(2
.8;1
.4;
0.13
)15
0.5
(0–1
4);
0.22
3(0
–19)
;1.
080.
11
(1;2
.2;
0.11
)2
(2.5
;1.
26;0
.37)
3(1
.67;
1.11
;0.4
)
Dem
NP
197
(0–3
5);
5.82
15(0
–27)
;10
.40.
5710
(1.5
8;1.
1;0.
11)
11(2
;1.9
;0.
19)
10(2
.5;
1.8;
0.19
)14
3(0
–31)
;0.
782
(0–2
9);
0.55
0.5
3(1
.33;
0.46
;0.2
3)9
(1.5
;0.
47;0
.27)
3(3
;0.6
3;0.
43)
Inte
rmed
NP
88
(0–2
2);
3.91
15(0
–31)
;13
.67
0.47
2(1
;1.0
6;0.
14)
6(2
;1.1
8;0.
15)
1(1
;1.2
4;0.
07)
63
(0–1
1);
1.42
5(0
–13)
;1.
780.
80
2(1
.5;
0.59
;0.2
2)1
(1;0
.44;
0.42
)
Dem
,dem
yelin
atin
g;D
isp
,dis
per
sio
n;E
N,e
lon
gat
edn
od
e;In
term
ed,i
nte
rmed
iate
;MF,
mye
linat
edfi
ber
;NP
,neu
rop
ath
y.
171

Doppler et al. Journal of the Peripheral Nervous System 18:168–176 (2013)
Table 3. Features of demyelination in finger skinbiopsies of patients with motor neuropathy and withoutinvolvement of the upper limbs.
Allpatients(n = 52)
Motorneuropathy
(n = 5)
Noinvolvement of
upper limbs(n = 17)
Elongated nodes (n;%)
13; 25% 2; 40% 4; 24%
Dispersion of caspr(n; %)
19; 37% 2; 40% 6; 35%
Dispersion ofneurofascin (n; %)
16; 31% 4; 80% 5/17; 29%
observe any spreading of sodium channels or anyother difference in the distribution of sodium channelsin patients with demyelinating or axonal NP comparedwith normal controls.
Pan-neurofascin in normal controls and inbiopsies of patients with NP
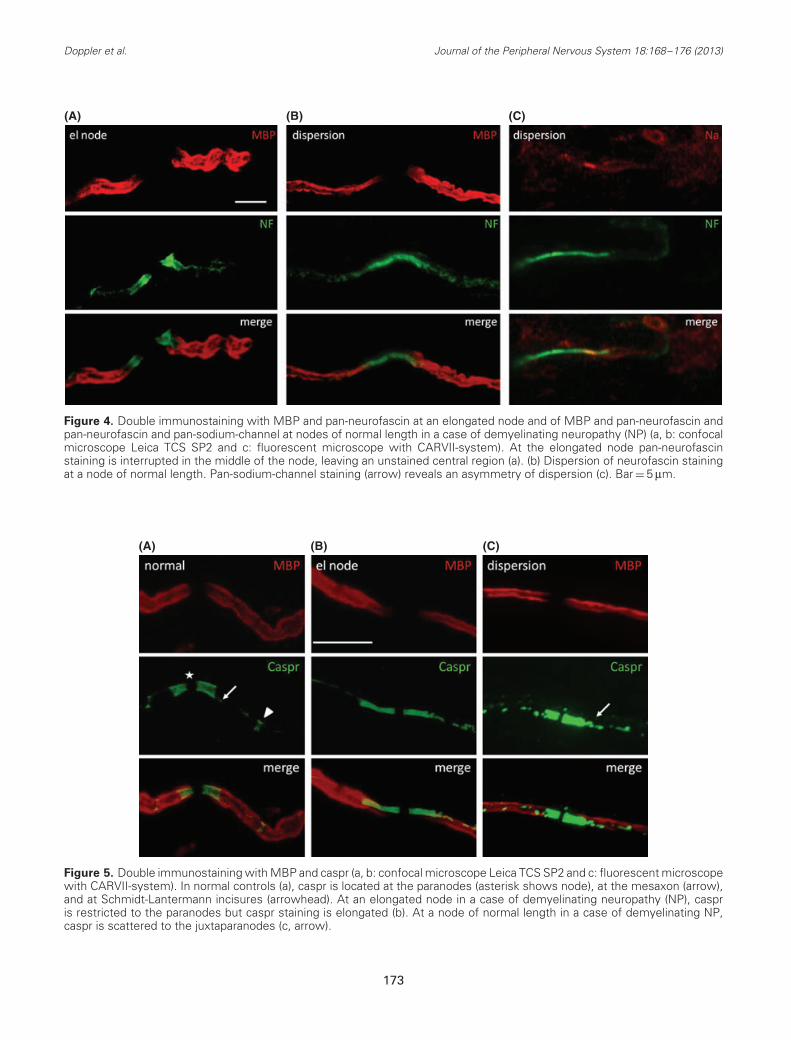
In normal controls, pan-neurofascin immunolabel-ing resulted in staining of the paranodal and nodalregions (Figs. 3a and 3b). In 41.8% (36 of 116) ofall nodes of the finger, a small central cleft remainedunstained (Fig. 3a). These clefts were mainly foundin nerve fibers innervating Meissner corpuscles. Inbiopsies of the proximal leg, neurofascin was alsodetectable at the mesaxon and at Schmidt-Lantermannincisures (Fig. 3b), but only 1.2% (2 of 59) of all nodeshad an unstained central cleft. Some elongated nodesshowed an interruption of the neurofascin stainingin the center of nodes (Fig. 4a). In nodes of normallength, broadening of neurofascin staining (Figs. 4b and4c) was observed in 11 of 21 patients with demyeli-nating NP, 6 of 21 cases with axonal NP, and 1 of 10cases with intermediate NP. In normal controls, broad-ening of neurofascin staining was observed only in onebiopsy of the finger. This feature was more often foundin demyelinating NP than in normal controls (p = 0.005,Table 2). There were nerve fibers containing a nodewith dispersion of neurofascin and nodes with normaldistribution of neurofascin.
Caspr in normal controls and in biopsiesof patients with NP
In normal controls, caspr immunolabeling wassymmetrically detectable at the paranodal regionson both sides of the node of Ranvier and at themesaxon, resulting in a thin line emanating fromthe paranodal region and running along the nervefiber (Fig. 5a). In some fibers, short extranodal caspr-positive and pan-sodium-negative bands could befound, probably corresponding to Schmidt-Lantermann
incisures (Fig. 5a). They were abundant in myelinatedfibers innervating Meissner corpuscles. Staining ofelongated nodes with anti-caspr revealed an elongationof the caspr- and MBP-negative gap between theparanodes and an elongation of the caspr-positiveparanodes (Fig. 5b). At nodes of normal length, adispersion of caspr was observed at some paranodes.Caspr staining was scattered at these paranodes incontrast to paranodes with broadening of neurofascinstaining, which remained clearly circumscribed. Thiswas detectable in 11 of 21 cases of demyelinating NP(Fig. 5c, Table 2), but only in 2 of 21 cases of axonalNP and 5 of 10 cases of intermediate NP. It wasnot found in any normal controls. Thus, dispersion ofcaspr staining was more often seen in demyelinatingcompared with axonal NP (p = 0.006). Again, therewere myelinated fibers containing both nodes withdispersion of caspr and normal nodes. Dispersionof neurofascin and caspr staining was detectable inpatients with motor as well as in patients with mixedor predominant sensory symptoms and was found inbiopsy specimens from the finger of patients with andwithout involvement of the upper limbs (Table 3).
Combination of nodal changes in biopsies ofdemyelinating and axonal NP
As elongated nodes and dispersion of casprand neurofascin were mainly found in demyelinatingNP, we tested for an overlap of these features indemyelinating NP. All three features were found in 5 of21 biopsies of demyelinating NP, but not in any axonalNP or normal control. Two features were identified in12 of 21 cases of demyelinating NP, 3 of 21 cases ofaxonal NP, and in 1 intermediate case. Only one featurecould be detected in 2 of 21 cases of demyelinating NPand 4 of 21 biopsies of patients with axonal NP. Thus,the combination of at least two demyelinating featuresappears to be most suitable to predict demyelination.
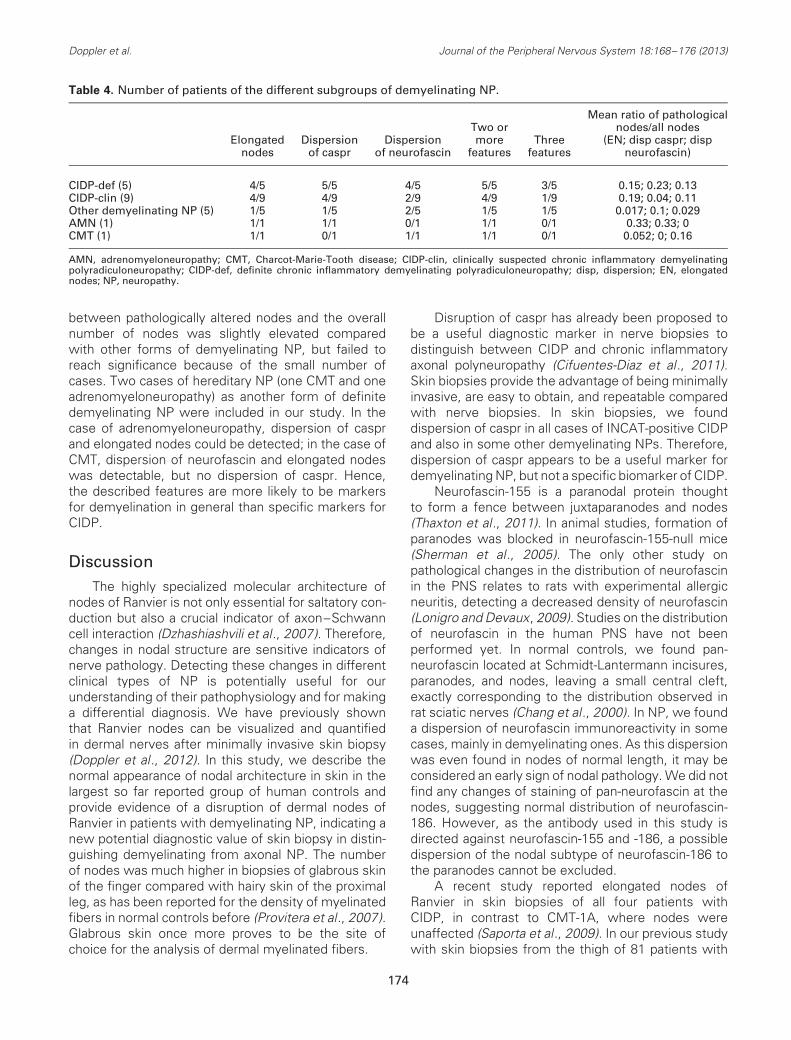
Elongated nodes and nodal changes in CIDP
CIDP is an acquired, treatable type of demyelinat-ing NP. To find out whether any of the demyelinatingfeatures described above might predict a diagnosisof CIDP, we performed an additional analysis of alldemyelinating NP cases (n = 21). Among these, fivecases of CIDP fulfilling the INCAT criteria (Hugheset al., 2001) were identified (definite CIDP, CIDP-def)and nine cases of clinically suspected CIDP (CIDP-clin)not fulfilling these criteria. Scattered dispersion ofcaspr was detectable in all CIDP-def cases, andelongated nodes and dispersion of neurofascin each infour of five cases. Thus, all cases of CIDP-def showedat least two demyelinating features. In CIDP-clin,four of nine cases had two or more demyelinatingfeatures (Table 4). In CIDP-clin and CIDP-def, the ratio
172
Doppler et al. Journal of the Peripheral Nervous System 18:168–176 (2013)
(A) (B) (C)
Figure 4. Double immunostaining with MBP and pan-neurofascin at an elongated node and of MBP and pan-neurofascin andpan-neurofascin and pan-sodium-channel at nodes of normal length in a case of demyelinating neuropathy (NP) (a, b: confocalmicroscope Leica TCS SP2 and c: fluorescent microscope with CARVII-system). At the elongated node pan-neurofascinstaining is interrupted in the middle of the node, leaving an unstained central region (a). (b) Dispersion of neurofascin stainingat a node of normal length. Pan-sodium-channel staining (arrow) reveals an asymmetry of dispersion (c). Bar = 5 μm.
(A) (B) (C)
Figure 5. Double immunostaining with MBP and caspr (a, b: confocal microscope Leica TCS SP2 and c: fluorescent microscopewith CARVII-system). In normal controls (a), caspr is located at the paranodes (asterisk shows node), at the mesaxon (arrow),and at Schmidt-Lantermann incisures (arrowhead). At an elongated node in a case of demyelinating neuropathy (NP), caspris restricted to the paranodes but caspr staining is elongated (b). At a node of normal length in a case of demyelinating NP,caspr is scattered to the juxtaparanodes (c, arrow).
173
Doppler et al. Journal of the Peripheral Nervous System 18:168–176 (2013)
Table 4. Number of patients of the different subgroups of demyelinating NP.
Elongatednodes
Dispersionof caspr
Dispersionof neurofascin
Two ormore
featuresThree
features
Mean ratio of pathologicalnodes/all nodes
(EN; disp caspr; dispneurofascin)
CIDP-def (5) 4/5 5/5 4/5 5/5 3/5 0.15; 0.23; 0.13CIDP-clin (9) 4/9 4/9 2/9 4/9 1/9 0.19; 0.04; 0.11Other demyelinating NP (5) 1/5 1/5 2/5 1/5 1/5 0.017; 0.1; 0.029AMN (1) 1/1 1/1 0/1 1/1 0/1 0.33; 0.33; 0CMT (1) 1/1 0/1 1/1 1/1 0/1 0.052; 0; 0.16
AMN, adrenomyeloneuropathy; CMT, Charcot-Marie-Tooth disease; CIDP-clin, clinically suspected chronic inflammatory demyelinatingpolyradiculoneuropathy; CIDP-def, definite chronic inflammatory demyelinating polyradiculoneuropathy; disp, dispersion; EN, elongatednodes; NP, neuropathy.
between pathologically altered nodes and the overallnumber of nodes was slightly elevated comparedwith other forms of demyelinating NP, but failed toreach significance because of the small number ofcases. Two cases of hereditary NP (one CMT and oneadrenomyeloneuropathy) as another form of definitedemyelinating NP were included in our study. In thecase of adrenomyeloneuropathy, dispersion of casprand elongated nodes could be detected; in the case ofCMT, dispersion of neurofascin and elongated nodeswas detectable, but no dispersion of caspr. Hence,the described features are more likely to be markersfor demyelination in general than specific markers forCIDP.
DiscussionThe highly specialized molecular architecture of
nodes of Ranvier is not only essential for saltatory con-duction but also a crucial indicator of axon–Schwanncell interaction (Dzhashiashvili et al., 2007). Therefore,changes in nodal structure are sensitive indicators ofnerve pathology. Detecting these changes in differentclinical types of NP is potentially useful for ourunderstanding of their pathophysiology and for makinga differential diagnosis. We have previously shownthat Ranvier nodes can be visualized and quantifiedin dermal nerves after minimally invasive skin biopsy(Doppler et al., 2012). In this study, we describe thenormal appearance of nodal architecture in skin in thelargest so far reported group of human controls andprovide evidence of a disruption of dermal nodes ofRanvier in patients with demyelinating NP, indicating anew potential diagnostic value of skin biopsy in distin-guishing demyelinating from axonal NP. The numberof nodes was much higher in biopsies of glabrous skinof the finger compared with hairy skin of the proximalleg, as has been reported for the density of myelinatedfibers in normal controls before (Provitera et al., 2007).Glabrous skin once more proves to be the site ofchoice for the analysis of dermal myelinated fibers.
Disruption of caspr has already been proposed tobe a useful diagnostic marker in nerve biopsies todistinguish between CIDP and chronic inflammatoryaxonal polyneuropathy (Cifuentes-Diaz et al., 2011).Skin biopsies provide the advantage of being minimallyinvasive, are easy to obtain, and repeatable comparedwith nerve biopsies. In skin biopsies, we founddispersion of caspr in all cases of INCAT-positive CIDPand also in some other demyelinating NPs. Therefore,dispersion of caspr appears to be a useful marker fordemyelinating NP, but not a specific biomarker of CIDP.
Neurofascin-155 is a paranodal protein thoughtto form a fence between juxtaparanodes and nodes(Thaxton et al., 2011). In animal studies, formation ofparanodes was blocked in neurofascin-155-null mice(Sherman et al., 2005). The only other study onpathological changes in the distribution of neurofascinin the PNS relates to rats with experimental allergicneuritis, detecting a decreased density of neurofascin(Lonigro and Devaux, 2009). Studies on the distributionof neurofascin in the human PNS have not beenperformed yet. In normal controls, we found pan-neurofascin located at Schmidt-Lantermann incisures,paranodes, and nodes, leaving a small central cleft,exactly corresponding to the distribution observed inrat sciatic nerves (Chang et al., 2000). In NP, we founda dispersion of neurofascin immunoreactivity in somecases, mainly in demyelinating ones. As this dispersionwas even found in nodes of normal length, it may beconsidered an early sign of nodal pathology. We did notfind any changes of staining of pan-neurofascin at thenodes, suggesting normal distribution of neurofascin-186. However, as the antibody used in this study isdirected against neurofascin-155 and -186, a possibledispersion of the nodal subtype of neurofascin-186 tothe paranodes cannot be excluded.
A recent study reported elongated nodes ofRanvier in skin biopsies of all four patients withCIDP, in contrast to CMT-1A, where nodes wereunaffected (Saporta et al., 2009). In our previous studywith skin biopsies from the thigh of 81 patients with
174

Doppler et al. Journal of the Peripheral Nervous System 18:168–176 (2013)
NP, we found elongated nodes only in demyelinatingNP. In this study, we could confirm that elongatednodes were present more often in NP classified asdemyelinating, and also in some that were of theaxonal type according to neurophysiological and suralnerve biopsy criteria. Thus, the analysis of Ranviernodes in very distal nerves, that is, in skin biopsy, mayhelp to identify an otherwise undetected demyelinatingpathology. Interestingly, pathologic findings in nodallength or architecture in skin samples from thefinger could even be detected in patients withoutclinical or neurophysiological involvement of the hands,indicating that these findings give complementaryinformation about the state of distal myelinated nerves.
We did not observe any changes in the distributionof sodium channels in dermal myelinated fibers ofpatients with demyelinating NP, a finding that isconsistent with previous studies analyzing nodal archi-tectures in skin biopsies of patients with hereditaryperipheral neuropathy (PNP) (Li et al., 2005; Saportaet al., 2009). In contrast, an altered distribution ofsodium channels has been described in nerve biopsiesof patients with CIDP and in multiple sclerosis(Howell et al., 2006; Cifuentes-Diaz et al., 2011),indicating potential differences in the mechanisms ofdemyelination in more proximal parts of nerve fibersand in the central nervous system.
Recent studies have suggested nodes and paran-odes to be a target of autoimmune attack by detectingauto-antibodies against neurofascin, gliomedin, andcontactin-1 in different demyelinating disorders likeGuillain-Barre syndrome, CIDP, multiple sclerosis,or experimental autoimmune neuritis (EAN) (Matheyet al., 2007; Lonigro and Devaux, 2009; Pruss et al.,2011; Devaux, 2012; Devaux et al., 2012; Ng et al.,2012; Querol et al., 2013). Our findings of a disruptionof nodal architecture in CIDP support the notionthat nodes of Ranvier and paranodal regions are animportant site of action in this disease. However,the pathogenetic link between auto-antibodies anda possible disruption of these proteins remains tobe established. Furthermore, an immune attack maynot be the only mechanism leading to paranodaldisruption, as we also found it in non-immune NP.
In summary, we identified three potential featuresof demyelination in skin biopsies of hairy and glabrousskin, namely elongated nodes and dispersion ofcaspr or neurofascin immunoreactivity. By studyingdifferent forms of NP, we found that the features ofdemyelination were present more often in patientsfulfilling the INCAT criteria of CIDP. For patients withsuspected CIDP not fulfilling these diagnostic criteria,the detection rate was lower. However, such patientsare the reason why biomarkers of CIDP are urgentlyneeded, and larger cohorts of patients should be
studied using the parameters described here. As ourpatients only obtained skin biopsy once at the time ofdiagnostic work-up, this study does not provide anyinformation on the development of nodal changes dur-ing the course of disease or in response to treatment.Further studies are needed to elucidate these aspects.
AcknowledgementsWe thank Barbara Dekant for technical assistance
and Alexandra Geßner and Waldemar Kafke forperforming skin biopsies. This study was supportedby Research Funds of the University of Wurzburg.
ReferencesArroyo EJ, Scherer SS (2000). On the molecular architecture of
myelinated fibers. Histochem Cell Biol 113:1–18.Arroyo EJ, Sirkowski EE, Chitale R, Scherer SS (2004).
Acute demyelination disrupts the molecular organizationof peripheral nervous system nodes. J Comp Neurol479:424–434.
Black JA, Friedman B, Waxman SG, Elmer LW, AngelidesKJ (1989). Immuno-ultrastructural localization of sodiumchannels at nodes of Ranvier and perinodal astrocytes inrat optic nerve. Proc R Soc Lond B Biol Sci 238:39–51.
Chang BJ, Cho IJ, Brophy PJ (2000). A study on theimmunocytochemical localization of neurofascin in rat sciaticnerve. J Vet Sci 1:67–71.
Cifuentes-Diaz C, Dubourg O, Irinopoulou T, Vigny M, LachkarS, Decker L, Charnay P, Denisenko N, Maisonobe T, LegerJM, Viala K, Hauw JJ, Girault JA (2011). Nodes of ranvierand paranodes in chronic acquired neuropathies. PLoS One6:e14533.
Coman I, Aigrot MS, Seilhean D, Reynolds R, Girault JA, ZalcB, Lubetzki C (2006). Nodal, paranodal and juxtaparanodalaxonal proteins during demyelination and remyelination inmultiple sclerosis. Brain 129:3186–3195.
Devaux JJ (2012). Antibodies to gliomedin cause peripheraldemyelinating neuropathy and the dismantling of the nodesof Ranvier. Am J Pathol 181:1402–1413.
Devaux JJ, Odaka M, Yuki N (2012). Nodal proteins are targetantigens in Guillain-Barre syndrome. J Peripher Nerv Syst17:62–71.
Doppler K, Werner C, Henneges C, Sommer C (2012). Analysisof myelinated fibers in human skin biopsies of patients withneuropathies. J Neurol 259:1879–1887.
Dupree JL, Girault JA, Popko B (1999). Axo-glial interactionsregulate the localization of axonal paranodal proteins. J CellBiol 147:1145–1152.
Dzhashiashvili Y, Zhang Y, Galinska J, Lam I, Grumet M, SalzerJL (2007). Nodes of Ranvier and axon initial segments areankyrin G-dependent domains that assemble by distinctmechanisms. J Cell Biol 177:857–870.
Howell OW, Palser A, Polito A, Melrose S, Zonta B, ScheiermannC, Vora AJ, Brophy PJ, Reynolds R (2006). Disruption ofneurofascin localization reveals early changes precedingdemyelination and remyelination in multiple sclerosis. Brain129:3173–3185.
175

Doppler et al. Journal of the Peripheral Nervous System 18:168–176 (2013)
Hughes R, Bensa S, Willison H, Van den Bergh P, ComiG, Illa I, Nobile-Orazio E, van Doorn P, Dalakas M,Bojar M, Swan A (2001). Randomized controlled trial ofintravenous immunoglobulin versus oral prednisolone inchronic inflammatory demyelinating polyradiculoneuropathy.Ann Neurol 50:195–201.
Li J, Bai Y, Ghandour K, Qin P, Grandis M, Trostinskaia A,Ianakova E, Wu X, Schenone A, Vallat JM, Kupsky WJ,Hatfield J, Shy ME (2005). Skin biopsies in myelin-relatedneuropathies: bringing molecular pathology to the bedside.Brain 128:1168–1177.
Lonigro A, Devaux JJ (2009). Disruption of neurofascin andgliomedin at nodes of Ranvier precedes demyelination inexperimental allergic neuritis. Brain 132:260–273.
Mathey EK, Derfuss T, Storch MK, Williams KR, Hales K,Woolley DR, Al-Hayani A, Davies SN, Rasband MN, OlssonT, Moldenhauer A, Velhin S, Hohlfeld R, Meinl E, LiningtonC (2007). Neurofascin as a novel target for autoantibody-mediated axonal injury. J Exp Med 204:2363–2372.
Ng JK, Malotka J, Kawakami N, Derfuss T, Khademi M, OlssonT, Linington C, Odaka M, Tackenberg B, Pruss H, SchwabJM, Harms L, Harms H, Sommer C, Rasband MN, Eshed-Eisenbach Y, Peles E, Hohlfeld R, Yuki N, Dornmair K,Meinl E (2012). Neurofascin as a target for autoantibodies inperipheral neuropathies. Neurology 79:2241–2248.
Nolano M, Provitera V, Crisci C, Stancanelli A, Wendelschafer-Crabb G, Kennedy WR, Santoro L (2003). Quantification ofmyelinated endings and mechanoreceptors in human digitalskin. Ann Neurol 54:197–205.
Provitera V, Nolano M, Pagano A, Caporaso G, Stancanelli A,Santoro L (2007). Myelinated nerve endings in human skin.Muscle Nerve 35:767–775.
Pruss H, Schwab JM, Derst C, Gortzen A, Veh RW (2011).Neurofascin as target of autoantibodies in Guillain-Barresyndrome. Brain 134:e173 author reply e174.
Querol L, Nogales-Gadea G, Rojas-Garcia R, Martinez-HernandezE, Diaz-Manera J, Suarez-Calvet X, Navas M, Araque J,Gallardo E, Isabel I (2013). Antibodies to contactin-1 inchronic inflammatory demyelinating polyneuropathy. AnnNeurol 73:370–380.
Reilly MM (2007). Sorting out the inherited neuropathies. PractNeurol 7:93–105.
Saporta MA, Katona I, Lewis RA, Masse S, Shy ME, Li J(2009). Shortened internodal length of dermal myelinatednerve fibres in Charcot-Marie-Tooth disease type 1A. Brain132:3263–3273.
Scherer SS, Arroyo EJ (2002). Recent progress on the molecularorganization of myelinated axons. J Peripher Nerv Syst7:1–12.
Sherman DL, Tait S, Melrose S, Johnson R, Zonta B, CourtFA, Macklin WB, Meek S, Smith AJ, Cottrell DF, Brophy PJ(2005). Neurofascins are required to establish axonal domainsfor saltatory conduction. Neuron 48:737–742.
Susuki K, Rasband MN (2008). Molecular mechanisms of nodeof Ranvier formation. Curr Opin Cell Biol 20:616–623.
Thaxton C, Pillai AM, Pribisko AL, Dupree JL, Bhat MA(2011). Nodes of Ranvier act as barriers to restrict invasionof flanking paranodal domains in myelinated axons. Neuron69:244–257.
Traka M, Goutebroze L, Denisenko N, Bessa M, Nifli A, HavakiS, Iwakura Y, Fukamauchi F, Watanabe K, Soliven B, GiraultJA, Karagogeos D (2003). Association of TAG-1 with Caspr2is essential for the molecular organization of juxtaparanodalregions of myelinated fibers. J Cell Biol 162:1161–1172.
Uceyler N, Kafke W, Riediger N, He L, Necula G, ToykaKV, Sommer C (2010). Elevated proinflammatory cytokineexpression in affected skin in small fiber neuropathy.Neurology 74:1806–1813.
176





























