Embed Size (px)
Citation preview

1994, 68(8):5216. J. Virol.
B Delmas, J Gelfi, E Kut, H Sjöström, O Noren and H Laude distinct from the enzymatic site.within a domain of aminopeptidase-N that is
residegastroenteritis virus-receptor interaction Determinants essential for the transmissible
http://jvi.asm.org/content/68/8/5216Updated information and services can be found at:
These include:
CONTENT ALERTS more»cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new articles
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on February 23, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jvi.asm
.org/D
ownloaded from

JOURNAL OF VIROLOGY, Aug. 1994, p. 5216-5224 Vol. 68, No. 80022-538X/94/$04.00+0Copyright © 1994, American Society for Microbiology
Determinants Essential for the Transmissible GastroenteritisVirus-Receptor Interaction Reside within a Domain
of Aminopeptidase-N That Is Distinctfrom the Enzymatic Site
BERNARD DELMAS,' JACQUELINE GELFI,' EMMANUEL KUT,' HANS SJOSTROM,2OVE NOREN,2 AND HUBERT LAUDEl*
Unite de Virologie et Immunologie Moleculaires, Institut National de la Recherche Agronomique, 78850 Jouy-en-Josas,France, ' and Department of Medical Biochemistry and Genetics, Panum Institute, 2200 Copenhagen N, Denmark2
Received 21 March 1994/Accepted 13 May 1994
The swine-specific coronavirus transmissible gastroenteritis virus (TGEV) uses pig aminopeptidase-N(pAPN) as a cellular receptor. We showed that the human aminopeptidase-N (hAPN) cannot substitute forpAPN in this respect, although the two enzymes have 80% amino acid sequence identity. In order to map theTGEV binding site on pAPN, we constructed a series of APN cDNA chimeras between pAPN and hAP?N andanalyzed them for their capacity to confer infectivity. The region between residues 717 and 813 was found tobe essential for infectivity. This region also contains the epitopes for three TGEV-blocking monoclonalantibodies directed against pAPN. These data support the view that the catalytic site and the TGEV receptorsite are located in different domains. Moreover, APN inhibitors and mutations in the catalytic site had noobvious effect on permissiveness for virus, thus providing evidence that the APN enzymatic activity is notinvolved in the process of infection.
Aminopeptidase-N (APN) has been shown to act as a majorreceptor for three coronaviruses belonging to the same geneticsubset, i.e., porcine transmissible gastroenteritis virus (TGEV),porcine respiratory coronavirus, a variant of TGEV, and thehuman respiratory coronavirus HCV-229E (3, 4, 25). APN hasalso been reported to mediate human cytomegalovirus infec-tion by a process that increases virus binding on the cell surface(21).APN is a member of a membrane-bound metallopeptidase
family with wide distribution on the surfaces of diverse celltypes. APN is expressed at a high level on the brush bordermembrane of the enterocytes, where it participates in the finalsteps of digestion by cleaving peptides preferentially afterN-terminal neutral amino acids. Less is known about itsphysiological role in other tissues, although it has been sug-gested that APN is involved in modulating signals of bioactivepeptides in the brain. More generally, APN is expressed inepithelial cell systems such as those in the brush border of thekidney and in the lung epithelium (10, 17, 20). The interest inAPN was reinforced when CD13 was identified as human APN(hAPN) (14). CD13 is expressed by normal human granulo-cyte-monocyte progenitors and by their more differentiatedprogeny. CD13 is one of the surface markers on leukemicblasts in acute myeloblastic and, less frequently, lymphoidleukemia (see reference 14 for additional references). APN isa type II glycoprotein, and the pig enzyme can be cleaved bytrypsin into subunit B (N-terminal part) and subunit C (C-terminal part). The catalytic site, assumed to be HExxH byhomology with other Zn2+ metalloproteases such as endopep-tidase 24.11 and thermolysin, was localized within the Bsubunit (9).The introduction of the species-related APN cDNA into
* Corresponding author. Fax: 33 1 3465 2621. Electronic mailaddress: [email protected].
cultured cells nonpermissive for the TGEV and HCV-229Ecoronaviruses is sufficient to render these cells susceptible toviral infection (3, 25). Moreover, a direct and specific interac-tion between TGEV virions and purified APN has beendemonstrated. Altogether, these results support the view thatthe virus infection is initiated by the binding of these corona-viruses to APN. The question of whether or not the enzymaticactivity plays any role at a subsequent step of the infectiousprocess has not been resolved. By studying the effect ofcurrently available APN inhibitors, it was found that 1,10-phenanthroline and 2,2'-dipyridyl inhibited HCV-229E repli-cation, whereas bestatin had no effect on HCV-229E or TGEVreplication (3, 25). In addition, anti-hAPN antibodies whichblock HCV-229E infection were found to be potent inhibitorsof APN activity (25).The aim of the present study was to provide information
about the physical locations of determinants essential for theTGEV receptor activity of APN and to investigate the possibleinvolvement of the enzymatic function in the overall process ofinfection.
MATERIALS AND METHODS
DNA sequencing. Shotgun DNA sequencing was performedby standard procedures (19a). Briefly, sonicated plasmid frag-ments were subcloned into the dephosphorylated M13mpl8SmaI-cut vector, such that the porcine APN (pAPN) cDNAsequence was determined on both strands. Sequencing reac-tions were done with the Sequenase kit (United States Bio-chemicals) and the -21M13 primer.
Construction of chimeric APN cDNAs. The positions ofrestriction sites will be given according to the porcine nucle-otide sequence, with + 1 corresponding to the A of the startcodon. To construct the chimeric APN cDNAs AP7 to AP11,
5216
on February 23, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jvi.asm
.org/D
ownloaded from

DOMAIN OF APN INVOLVED INTGEV RECEPTOR FUNCTION 5217
a segment ofcDNA was removed by using unique or doublerestriction sites located at the same position in the two parentalcDNAs and was replaced by the reciprocal human or porcinefragment. When needed, a second restriction site within theplasmid sequence was used. AP7 was constructed by using twoNcoI sites (+ 1 and + 1043), AP8 was constructed by using theKp,nI site (+2519), and AP11 was constructed by using twoPvuII sites (+ 1924 and +2740). A similar strategy was used todesign chimeric cDNAs AP13 to AP15 and AP18 after PCR-directed mutagenesis (6) was used to create or destroy restric-tion sites. Mutageneses of restriction sites were designed tomake silent mutations. An SphI site was put into the hAPNcDNA at a position analogous to that of pAPN cDNA(+ 1497). AP13 was constructed by using the Sphl site and theSmaI site (+2792), and AP14 was constructed by using theSphI site and the KpnI site (+2519). To construct AP15, thePvuII site (+2449) in the porcine cDNA was destroyed and thehuman PvuIl segment (+1924 to +2740) was replaced by itsporcine homolog. To construct AP18, the PvuII site (+2740) inthe human cDNA was destroyed and the human PvuII-KpnIsegment (+1924 to +2519) was replaced by its porcine ho-molog mutated at thePvuII site (+2449). AP16 and AP17 wereconstructed by replacing the human PvuII segment (+ 1924 to+ 2740) with the PvuII porcine segment (+ 1924 to + 2449 and+2449 to +2740, respectively). For chimeric APN cDNAsAP19 to AP26, the unique restriction sites PvuII (+ 1924) andKpnI (+2519) remaining in the human cDNA were used toclone different hybrids generated by PCR-directed mutagene-sis between these two restriction sites. Oligonucleotides 2184and 2187 (5'-GGAGGAAGATTCAGACTCAGCTGCAGAand 5'-GGGTGTAACCCAGGTACCTGTTCAGGA, respec-tively) with internal PvuIl and KpnI sites, respectively, weresystematically used for PCR amplification and relevant cloningsteps. The internal pairs of oligonucleotides used to amplifyporcine or human segments were as follows: for AP19 andAP20, 2186 (5'-CTCCGAGGTCTATGGCCCCATGAAGAA)and 2185 (5'-TTYCTTCATGGGGCCATAGACCTCGGAG)with porcine and human cDNA as templates; for AP21 andAP22, 2285 (5'-ACCCGATCCACCCCAACCTGCGGTCCAC) and 2286 (5'-GTGGACCGCAGGTTGGGGTGGATCGGGT) with AP20 and human cDNA as templates; for AP23,2424 (5'-CCCAGAAAATCTGATGGACCAGTACAG) and2425 (5'-CTGTACTGGTCCATCAGAlTTTTCTGGG) withporcine and human cDNA as templates; for AP24, 2422(5'-AGTGGGACTTTGCCTGGGGGCAGTT) and 2423 (5'-AACTGCCCCCAGGCAAAGTCCCACT) with AP20 andhuman cDNA as templates; and for AP26, 2424 and 2425 withAP24 and human cDNA as templates. AP27 was constructedby replacing the porcine PvuII segment (+ 1924 to + 2740) withits human homolog.The PCRs were carried out by using Taq polymerase (Pro-
mega) in a DNA thermal cycler (Perkin-Elmer Cetus orHybaid). The structure of each chimeric cDNA was confirmedby restriction mapping with enzymes diagnostic of human orporcine APN cDNAs and, in several cases, by DNA sequencedetermination for the complete inserted segment (automatedsequencing on an Applied Biosystems 373A apparatus). Thechimeric cDNAs were then cloned in expression vectorpTEJ-4, under the control of the ubiquitin promoter andcontaining the polyadenylation sequence of simian virus 40 (8).
Expression of chimeric APN cDNAs. For transient andstable expression, the pTEJ-4 vector carrying one of thedifferent chimeric cDNAs was transfected into MDCK cells byCaPO4 precipitation (AP7 to AP15) or by lipofection (AP16 toAP27) according to the manufacturer's procedures (BethesdaResearch Laboratories). To obtain stable MDCK clones, cells
were cotransfected with pSV2neo and cell clones resistant tothe neomycin analog G418 were selected and then screened forAPN activity by using the chromogenic substrate leucinep-nitroanilide at a 1.6 mM concentration.
Construction and expression of enzymatically defectivepAPN. The pAPN cDNA was subcloned into pcDNA1 (In-vitrogen) by using BamHI and EcoRV restriction sites toproduce pcDNAP2. For the construction of the mutatedpAPN, pcDNAP2 was cut by XbaI and self-ligated to deletepart of the polylinker and to obtain EcoRI (+526) and SphI(+1497) as unique cloning sites in the pAPN cDNA. PCR-directed mutagenesis was performed between these two re-striction sites with oligonucleotides 2918 and 2919 (5'-GTACGAGATGGAGAGTGAATTCCA and 5'-GGCAAAGGCATGCAAGTAGGAC, respectively) carrying internal EcoRIand SphI sites, respectively. The internal oligonucleotides 2915(5'-GATTGCTCTCGAGCTGGCCCTCCAGTGG) and 2916(5'-CCACTGGAGGGCCAGCTCGAGAGCAATC) wereused in a nested PCR to replace the two histidines (+383 and+387) with two leucines and to carry a silent XhoI site forvalidation of the pcDNAP* construct. Expression of the twopcDNA1 derivatives was obtained by lipofection of COS-7cells. Enzymatic activity was determined as follows. A total of4 x 106 cells were lysed 48 h posttransfection by using 50 mMTris (pH 7.3) with 1% Triton X-100; enzymatic activity wasmeasured after incubation of the cell extracts in the presenceof leucine p-nitroanilide for 10 min at 20°C.
Receptor activity assays. For APN-expressing MDCK cellclones, monolayers were infected at a multiplicity of infectionof 10 PFU of the TGEV Purdue strain per cell. MDCK cellsand a pAPN-expressing MDCK clone were used as negativeand positive controls, respectively. At 24 h postinfection, cellsurvival was measured by a colorimetric assay (11) as follows.Monolayers were fixed and stained with a crystal violet solu-tion; the dye incorporated into cells surviving the viral cyto-pathic effect was determined by measuring optical absorbanceat 590 nm after solubilization in acetic acid.For APN transient expression assays, cell monolayers were
infected at 48 h postlipofection and then fixed at 24 hpostinfection. The monolayers were processed for immunofluo-rescence analysis to detect newly synthesized viral antigens byusing appropriate monoclonal antibodies (13).APN inhibitors. Three drugs provided by Marion Merrell
Dow Laboratory (Strasbourg, France) and designated 74,465,73,534, and 73,794 were tested. The structures of the first twohave been reported previously (19b). Drug stock solutionswere prepared in 20% dimethyl sulfoxide at a concentration of10 mM. ST cell line monolayers were pretreated for 2 h at 37°Cwith various concentrations of the drugs. For the plaquereduction assay, monolayers were infected for 1 h with 100 to150 PFU of virus per well, added with the same concentrationsof compounds. Then the inoculum containing virus and inhib-itor was withdrawn, and appropriate amounts of medium,inhibitor, and agarose were added so as to allow plaqueformation. The plaques were scored at 40 h postinfection.
Nucleotide sequence accession number. The nucleotide se-quence data reported in this paper will appear in the EMBLdatabase under accession number Z29522.
RESULTS
Predicted amino acid sequence of pAPN. The completenucleotide sequence of APN cDNA was determined from twoclones, which were isolated from a pig intestine cDNA libraryas previously described (3). A 2,889-bp-long open readingframe was found to encode a 963-amino-acid-long polypeptide
VOL. 68, 1994
on February 23, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jvi.asm
.org/D
ownloaded from

5218 DELMAS ET AL.
CCCTGCCCGTCCTGAGCCTCCCCGAGCTCCCTTCTCAC'CCTCACCATGGCCAAGGGATTCTACATTTCCAAGGCCCTGGGCATCCTGGGCM A K G F Y I S K A L G I L G 15
ATCCTCCTCGGCGTGGCGGCCGTGGC'CACCATCATCGCTCTGTCTGTGGTGTACGCCCAGGAGAAGAACAAGAATGCCGAGCATGTCCCCI L L G V A A V A T I I A L S V V Y A Q E K N K N A E H V P 45
CAGGCCCCCACGTCGC'CCACCATCAC'CAC'CAC'AGCCGC'CATCAC('CTTGGACCAGAGCAAGCCGTGGAACCGGTACCGCCTACCCACAACGQ A P T S P T I T T T A A I T L D Q S K P W N R Y R L P T T 75CTGTTGCCTGATTCCTAC'TTC'COTC-AC'C,CTGAGAC'C'CTACCTCACTCCCAACGC'GGATGGCCTGTACATCTTCAAGGGCAAAAGCATCGTCL L P D S Y F V T L R P Y L T P N A [) G L Y I F K G K S I V 105CGCTTACTCTGCCAGGAGCC'CAC'CGA'iTGTC'ATC'ATC'ATC'CATAGCAAGAAGCTCAAC'TACACCACCCAG-'GGGCACATGGTGGTCCTGCGGR L L C' Q E P' T D) V I I I H S K K L N T Q G H M V V L R 135GGCGTGGGGGACTC'CCAGGTC'C'CAGAC-ATC0GAC'ACCGA(CTGAGCTC;'GTA0-'AGCTC'ACTC0AGTACCTGGTGGTC'CACC'TCAA0GGGCTCGCTGG V G L) S Q V Pl E I F) R T E L V E L T E Y L V V H L K G S L 165CAGCCCGGCCACATGTAC'GAGATGGAGAGTGAATTCC'AGGGGGAACTTGCCGACGACCTGGCAGGCTTCTACCGCAGCGAGTACATGGAGQ P G H M Y E M E S E F Q G E L A D D L A G F Y R S E Y M E 195GGCAACGTCAAAAAGGTGCTGGCCAC'GACACAGATGCAGTCTACAGATGCCCGGAAATCCTTCCCATGCTTTGACGAGCCAO,CCATGAAGG N V K K V L A T T Q M Q S T D A R K S F P C F D E P A M K 225GCCACGTTCAACATCACTCTCATCCAC'CCTAACAAC'CTCACGGCCCTGTCCAATATGCCGCCCAAAGGTTCCAGCACCCCACTTGCAGAAA T F N IT L I H P' N *N.I. A L S N N P P K G S S T P L A E 255GACCCCAACTGGTCTGTC'ACTGAG;TTCGAAACCAC'ACCTGTGATOGTCCACGTAC'CTTCTG;GCCTACATCGTGAGCGAGTTCCAGAGCGTGD P N W.S V T E F E T T P' V M S T Y L L A Y I V S E F Q S V 285AATGAAACGGCCCAAAATGGCGTC'CTGATC'CGGATCTO,GGCTCGGCCTAATGC'AATTGCAGAGGGCCATGGC'ATGTATGCCCTGAATGTGN E T A Q N G V L I R I W A R P N A I A E G H C N Y A L N V31SACAGGTCCCATCCTAAACTTCTTTGC'CAATCATTATAATACATCCTACCCACTCCCCAAATCCGACCAGATTGCCTTGCCCGACTTCAAT
G P' I L N F F A N H Y N T...S Y P L El K S F) Q I A L P D F N 345GCCCGGTGCCCATGGAGAACTGOOOCT,G,GOC',"GT GAC'CTACC CAOAA -'OCOCCTGOCTGTTTGACCCACAGTCCTCCTCCATCAGCAACAAAGAGA G A M E N W G L V T Y R E N A L L F D Pl Q S S S I S N K E 375
R V V T V I A H E L A H Q'- W F O'N L V T L AW W N D)L W L N 405
E G F A Sc' Y V E Y L 0-, A [) H A K P T W N L K F) L I V P G D V 435TACCGAGTGATGGCTO,TC-OCOATGC'TCTOOC,C'k-TTC'CTC'CC'AC'C'CGCCT ;'ACCACCCCTGCTGA(-'GOAGGTCAACACACCTO-'CCCAGATCAGCGAGY R V N A V ED A L A S S H P' L T T P) A E E V N T P) A Q I S E 465
M F D S I S Y S K 0 A 'S V I R M L S N F L T E D L F K K C L 495GCGTCCTACTTGCATGC'CTTTGC-C((TATC'ACOAAC'AC'CAC'CTA(C'T,CL ACC'TGTGGGAGCACCTGCAGAAGGCTGTGGATGCTCAG-ACGTCCA S Y L H A F A Y Q N.T.T Y L F) L W E H L Q K A V D A Q T 5 525ATCAGGCTGCCCAGACACTOTGAGAGC'CATC'ATGGATC'GATGGAC"C'TGCAGATCGGGCTTCCCCGTC'ATCACCGTGGACACCAAGACAGGAI R L P D T V R A I N D) R N T L Q N G F P) V I T V D) T K T G 555AACATCTCACAGAAGCAC'TTC'CTC'CTC'GAC'TCCCGAATC'CAACOGT 'ACCCCGCTC'CTCAGCGTTCCGACTACCTCTC,GATTGTTCCCATCTCAN.I Q K H F L L D)S E S N V T R*S S A F D Y L W I V P I S 585
S I K N G0 V M 2 D) H Y W L R D) V S Q A Q N D L F K T A S D D 6 15c7TGGGTCTTGCTGAACO(-'TC'AA(-('C-TOA( 'AOOC','.TATTTQ\('AOO''-,TO~,AA"-TACGACOGAGCGACAACTGGAGGATGATTC'AGC'ATCAGCTGCAGACAW V L L N V N V TOGY FQ V N Y DE EE) N W RNM I Q H Q L Q T 645AACCTGTCGGTC'ATCC(-CTGTC'I(:ATC'AA-TCOOOC,'-'(.'(~.T("A(K4O,,TCATCTA ~'GAC'AGCTTCAACCTGGCCCACTGCCCCACATGGTCCCTGTCACCCTGN.LS V I P) V I N R AYC V I Y E) S F N L A T A H M V P V T L 675GCTCTGGACAAC'AC'CC'TCTTC'CTCOAA;COCOC,AO:,AOAAACOACOTAC'AT- -;CCC'TGCCCAGC'C'CCCCTGAGCAGCCCTGAGCTACTTCAGCCTCATGA L D N T L F L N 0 K 1K E Y M Pl W Q A A L S S L ~S Y F S L M 705TTCGACCGCTCCGACO-'TC'TATOC,C,(CC'CATO-'AAO,AAATACC'TC'AC -AAGCAGGTCGAACC'CCTCTTCCAACATTTCGAAACTCTCACTAAAF D R S K V Y 0 P) N K K Y L R K Q V K P) L F Q H F K T L T K 735AACTGGACCGAGCCGCCC'AGAAAATCTGATGCOACC'AGTAC'AO:,TCOA A.ATTAATC,CCATCAC,C'ACTGCCTGCTCCAATC-C,ATTGCCTCAATC,TN W.T KR P) K N L NE1)Q Y SKI N A I1ST A C S N C L P Q C 765
K N L A K T L F F) Q) W N S F) Pl K N N P I H P N L R S, T I Y C 795AATGCCATAGCCCAGGCOO"COOC'C'AGGAC'CAC-OTCOOO'-AC'TTTOC,(C'TGO;OOOGCACTTACAACAAGCCCACCTGGTAAATGAGCCGCACAAACTCN A I A Q 0 0 Q F) Q W F) F A W 0 Q L Q Q A Q L V N K A D K L 825CCCTCAGCGCTGGCC'TOC-CAOCAACOGAGOTC'TOC-C,'TC'CTO,AAC'AGO TACCTCGGTTACACCCTGAACC'CGGACCTCATTCGCAAGCAAGACR S A L A C' S2 N K V W L L N K Y L C Y T L N P D L I R K Q D 855GCCAC'CTCCACTATTAAC:AOC'(ATTG("'CAOC,AATO,TC'ATCOOC,''-,CA, ;'C'CTCTCOO-CCTOCCGATTTTGTCCAGAGCAACTOCGAAGAAGCTCTTTA T S T I N I A S N V I 0 PF L A W D F V Q S N W K K L F 885CAGGACTATGOCGGTG-.C'TT ('-TTC T '.C'TTrh,TCC(AACC(-TC~.ATI-CCA ;oO.'C,TO,TCOACC'CGAAO:,ATTC'TC'CTCTO-'AGTTTCOAC'CTGCAC'CAGCCTGQ D Y 0' 0' C F F N L I 2 V T R R F S S K F K L Q Q L 915GAGCAOTTCAAOAAOAAC' ,-AT( t,"ATC;'T,-,(''C;OTT,~-(.'.,COIO--CT(-CCOO'AC'C'COOOCGCTCTC-C,AGCAAGCCCCTGGCAOAAGAC'CAACGCCCAACATCK Q F K K N N N D) V0 F 0 T R A L K Q A L K K T K A N I 945AAGTGGGTGAAGGAOAAC AA -'C'AGo0TOO'C,TCOTTCOAATTOO-C,TTC'AT",AGAC'ACAGCCTAATAGTGCCTCC',TC'CTTCCCGCCCACCTCCGCCCCCCK W V K K N F K V V L N N F I K H S / 963GCACAAGATOC'CCGCATCE TTCT AT(-CCACAOGOCE''CACGOC,C'CACOOC,G 'C'CATC-'TTCC'TGAAGCCCGCAGGCACCTGCCCTC'CTCCCTTTAGGGACAAAGCCTGTGGCCCCATC- TTATC TC C ATTC'TC;(C'CCTOOC-O'GC,('C'AATPCC'AGTTTCTGGTC'AC'CAGACTGTCCAGGTGTCTCCCAGCCACTC,CCCCTTCTCCCAACCCC ACC'CT,C,GOCCTGGC'C'CAOCCGCCCTTC'TC'AC'GCAACTCCACCTCCACCCCCACATCACCACAACCCCTTCATCCA
CTATATTTTTTTTCTAGGAO-'TAAAT(O,TAAATAAAGOC,ATTTCT"IAGtA,TO-,AAACCAATTC
FIG. 1. Nucleotide and deduced amino acid sequences of the pAPN cDNA. The single-letter amino acid code is used throughout. Amino acidsof the membrane-spanning domain are doubly underlined. Potential sites of N glycosylation conserved in the human APN sequence are underlinedwith solid lines; other potential sites are underlined with dotted lines. Amino acids involved in the enzymatic site are in bold characters. Thepotential cleavage site after arginine 572 is indicated by an asterisk. Amino acids in italic characters (573 to 585) are the residues previouslyidentified as corresponding to the N terminus of the C subunit.
J. VIROL.
on February 23, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jvi.asm
.org/D
ownloaded from
DOMAIN OF APN INVOLVED IN TGEV RECEPTOR FUNCTION 5219
/ 4
-
*- w
n.O_-'
1_
-C]
0
0
4, 4,of.
MDCK pAPN-MDCK hAPN-MDCK
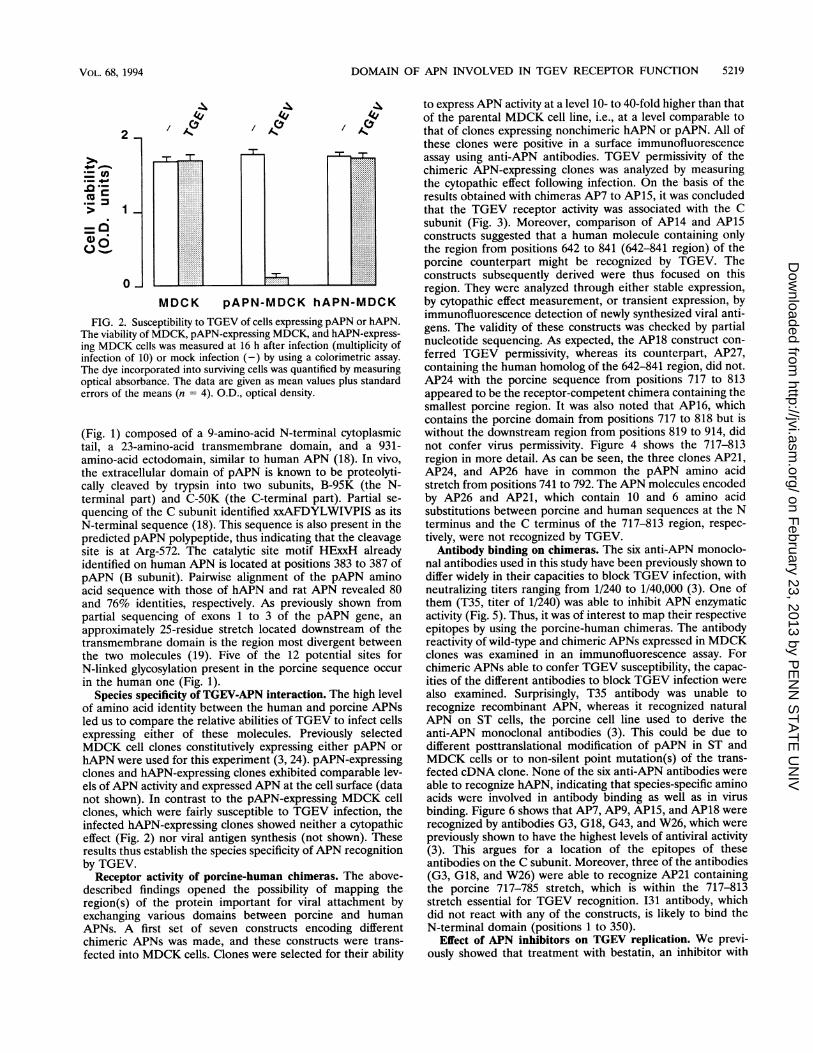
FIG. 2. Susceptibility to TGEV of cells expressing pAPN or hAPN.The viability of MDCK, pAPN-expressing MDCK, and hAPN-express-ing MDCK cells was measured at 16 h after infection (multiplicity ofinfection of 10) or mock infection (-) by using a colorimetric assay.
The dye incorporated into surviving cells was quantified by measuringoptical absorbance. The data are given as mean values plus standarderrors of the means (n = 4). O.D., optical density.
(Fig. 1) composed of a 9-amino-acid N-terminal cytoplasmictail, a 23-amino-acid transmembrane domain, and a 931-amino-acid ectodomain, similar to human APN (18). In vivo,the extracellular domain of pAPN is known to be proteolyti-cally cleaved by trypsin into two subunits, B-95K (the N-terminal part) and C-50K (the C-terminal part). Partial se-
quencing of the C subunit identified xxAFDYLWIVPIS as itsN-terminal sequence (18). This sequence is also present in thepredicted pAPN polypeptide, thus indicating that the cleavagesite is at Arg-572. The catalytic site motif HExxH alreadyidentified on human APN is located at positions 383 to 387 ofpAPN (B subunit). Pairwise alignment of the pAPN aminoacid sequence with those of hAPN and rat APN revealed 80and 76% identities, respectively. As previously shown frompartial sequencing of exons 1 to 3 of the pAPN gene, an
approximately 25-residue stretch located downstream of thetransmembrane domain is the region most divergent betweenthe two molecules (19). Five of the 12 potential sites forN-linked glycosylation present in the porcine sequence occur
in the human one (Fig. 1).Species specificity of TGEV-APN interaction. The high level
of amino acid identity between the human and porcine APNsled us to compare the relative abilities of TGEV to infect cellsexpressing either of these molecules. Previously selectedMDCK cell clones constitutively expressing either pAPN or
hAPN were used for this experiment (3, 24). pAPN-expressingclones and hAPN-expressing clones exhibited comparable lev-els of APN activity and expressed APN at the cell surface (datanot shown). In contrast to the pAPN-expressing MDCK cellclones, which were fairly susceptible to TGEV infection, theinfected hAPN-expressing clones showed neither a cytopathiceffect (Fig. 2) nor viral antigen synthesis (not shown). Theseresults thus establish the species specificity of APN recognitionby TGEV.
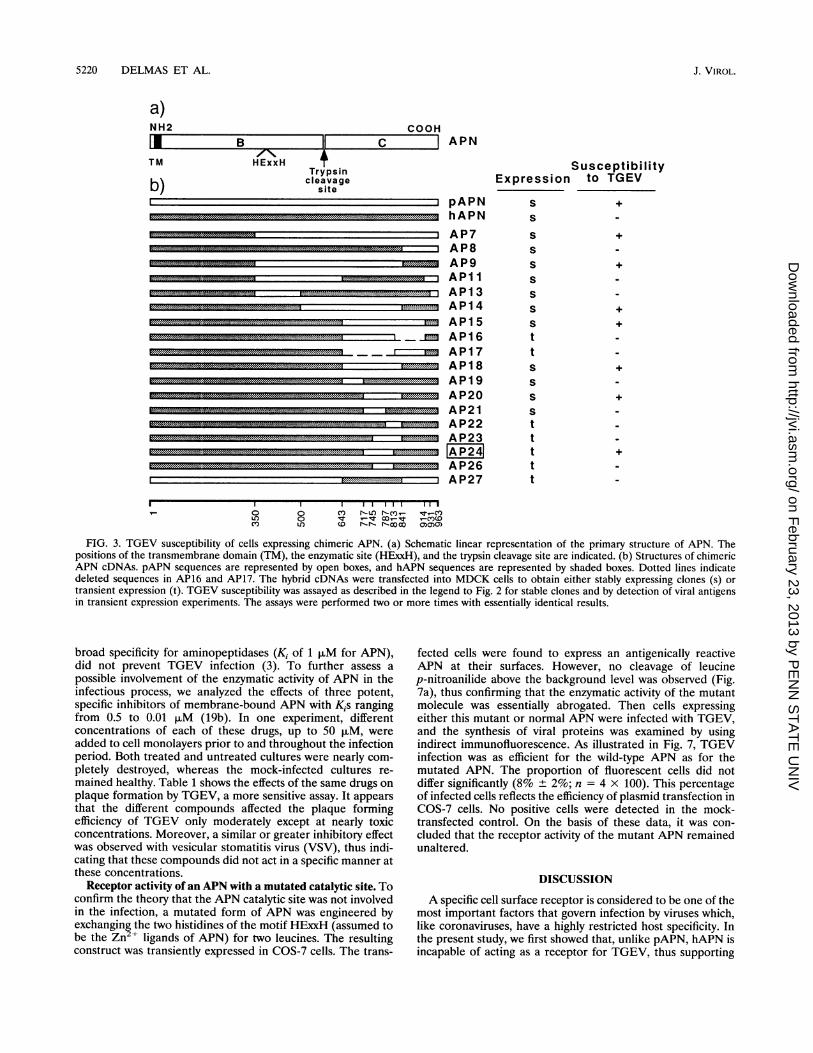
Receptor activity of porcine-human chimeras. The above-described findings opened the possibility of mapping theregion(s) of the protein important for viral attachment byexchanging various domains between porcine and humanAPNs. A first set of seven constructs encoding differentchimeric APNs was made, and these constructs were trans-
fected into MDCK cells. Clones were selected for their ability
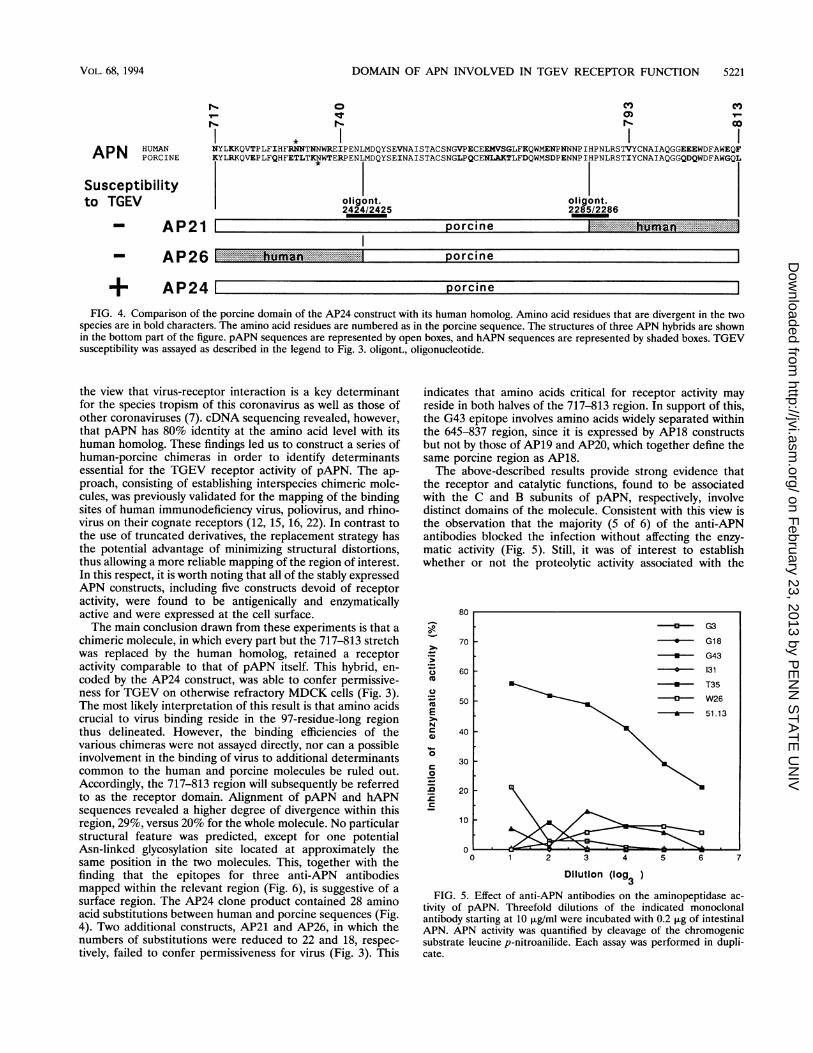
to express APN activity at a level 10- to 40-fold higher than thatof the parental MDCK cell line, i.e., at a level comparable tothat of clones expressing nonchimeric hAPN or pAPN. All ofthese clones were positive in a surface immunofluorescenceassay using anti-APN antibodies. TGEV permissivity of thechimeric APN-expressing clones was analyzed by measuringthe cytopathic effect following infection. On the basis of theresults obtained with chimeras AP7 to AP15, it was concludedthat the TGEV receptor activity was associated with the Csubunit (Fig. 3). Moreover, comparison of AP14 and AP15constructs suggested that a human molecule containing onlythe region from positions 642 to 841 (642-841 region) of theporcine counterpart might be recognized by TGEV. Theconstructs subsequently derived were thus focused on thisregion. They were analyzed through either stable expression,by cytopathic effect measurement, or transient expression, byimmunofluorescence detection of newly synthesized viral anti-gens. The validity of these constructs was checked by partialnucleotide sequencing. As expected, the AP18 construct con-ferred TGEV permissivity, whereas its counterpart, AP27,containing the human homolog of the 642-841 region, did not.AP24 with the porcine sequence from positions 717 to 813appeared to be the receptor-competent chimera containing thesmallest porcine region. It was also noted that AP16, whichcontains the porcine domain from positions 717 to 818 but iswithout the downstream region from positions 819 to 914, didnot confer virus permissivity. Figure 4 shows the 717-813region in more detail. As can be seen, the three clones AP21,AP24, and AP26 have in common the pAPN amino acidstretch from positions 741 to 792. The APN molecules encodedby AP26 and AP21, which contain 10 and 6 amino acidsubstitutions between porcine and human sequences at the Nterminus and the C terminus of the 717-813 region, respec-tively, were not recognized by TGEV.
Antibody binding on chimeras. The six anti-APN monoclo-nal antibodies used in this study have been previously shown todiffer widely in their capacities to block TGEV infection, withneutralizing titers ranging from 1/240 to 1/40,000 (3). One ofthem (T35, titer of 1/240) was able to inhibit APN enzymaticactivity (Fig. 5). Thus, it was of interest to map their respectiveepitopes by using the porcine-human chimeras. The antibodyreactivity of wild-type and chimeric APNs expressed in MDCKclones was examined in an immunofluorescence assay. Forchimeric APNs able to confer TGEV susceptibility, the capac-ities of the different antibodies to block TGEV infection werealso examined. Surprisingly, T35 antibody was unable torecognize recombinant APN, whereas it recognized naturalAPN on ST cells, the porcine cell line used to derive theanti-APN monoclonal antibodies (3). This could be due todifferent posttranslational modification of pAPN in ST andMDCK cells or to non-silent point mutation(s) of the trans-fected cDNA clone. None of the six anti-APN antibodies wereable to recognize hAPN, indicating that species-specific aminoacids were involved in antibody binding as well as in virusbinding. Figure 6 shows that AP7, AP9, AP15, and AP18 wererecognized by antibodies G3, G18, G43, and W26, which werepreviously shown to have the highest levels of antiviral activity(3). This argues for a location of the epitopes of theseantibodies on the C subunit. Moreover, three of the antibodies(G3, G18, and W26) were able to recognize AP21 containingthe porcine 717-785 stretch, which is within the 717-813stretch essential for TGEV recognition. 131 antibody, whichdid not react with any of the constructs, is likely to bind theN-terminal domain (positions 1 to 350).
Effect of APN inhibitors on TGEV replication. We previ-ously showed that treatment with bestatin, an inhibitor with
VOL. 68, 1994
on February 23, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jvi.asm
.org/D
ownloaded from
5220 DELMAS ET AL.
a)NH2 COOH
A| B C ITM HExxH
Trypsincleavageb) site
APN
pAPNhAPN
= AP7X A P8
~m. A P9API I
= AP15l--~ AP16
W AP17
~~ AP18_ ~AP1 9~~ AP2O
EY AP21~~ AP22~~ AP23
~~ AP26~~ AP27
SusceptibilityExpression to TGEV
s +
s -
+
+
++
ss
5
5sss
ttsss
tSt
tt
t
+
+
FIG. 3. TGEV susceptibility of cells expressing chimeric APN. (a) Schematic linear representation of the primary structure of APN. Thepositions of the transmembrane domain (TM), the enzymatic site (HExxH), and the trypsin cleavage site are indicated. (b) Structures of chimericAPN cDNAs. pAPN sequences are represented by open boxes, and hAPN sequences are represented by shaded boxes. Dotted lines indicatedeleted sequences in AP16 and AP17. The hybrid cDNAs were transfected into MDCK cells to obtain either stably expressing clones (s) ortransient expression (t). TGEV susceptibility was assayed as described in the legend to Fig. 2 for stable clones and by detection of viral antigensin transient expression experiments. The assays were performed two or more times with essentially identical results.
broad specificity for aminopeptidases (Ki of 1 ,uM for APN),did not prevent TGEV infection (3). To further assess apossible involvement of the enzymatic activity of APN in theinfectious process, we analyzed the effects of three potent,specific inhibitors of membrane-bound APN with Kis rangingfrom 0.5 to 0.01 puM (19b). In one experiment, differentconcentrations of each of these drugs, up to 50 ,uM, wereadded to cell monolayers prior to and throughout the infectionperiod. Both treated and untreated cultures were nearly com-pletely destroyed, whereas the mock-infected cultures re-mained healthy. Table 1 shows the effects of the same drugs onplaque formation by TGEV, a more sensitive assay. It appearsthat the different compounds affected the plaque formingefficiency of TGEV only moderately except at nearly toxicconcentrations. Moreover, a similar or greater inhibitory effectwas observed with vesicular stomatitis virus (VSV), thus indi-cating that these compounds did not act in a specific manner atthese concentrations.
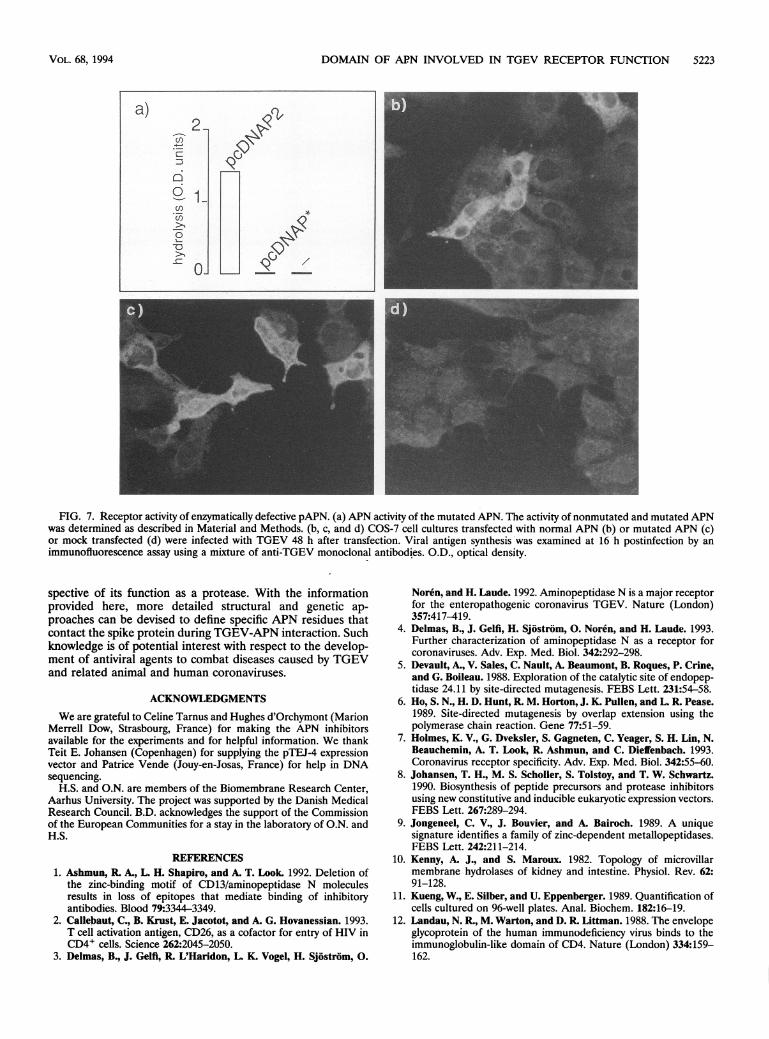
Receptor activity of an APN with a mutated catalytic site. Toconfirm the theory that the APN catalytic site was not involvedin the infection, a mutated form of APN was engineered byexchanging the two histidines of the motif HExxH (assumed tobe the Zn + ligands of APN) for two leucines. The resultingconstruct was transiently expressed in COS-7 cells. The trans-
fected cells were found to express an antigenically reactiveAPN at their surfaces. However, no cleavage of leucinep-nitroanilide above the background level was observed (Fig.7a), thus confirming that the enzymatic activity of the mutantmolecule was essentially abrogated. Then cells expressingeither this mutant or normal APN were infected with TGEV,and the synthesis of viral proteins was examined by usingindirect immunofluorescence. As illustrated in Fig. 7, TGEVinfection was as efficient for the wild-type APN as for themutated APN. The proportion of fluorescent cells did notdiffer significantly (8% ± 2%; n = 4 x 100). This percentageof infected cells reflects the efficiency of plasmid transfection inCOS-7 cells. No positive cells were detected in the mock-transfected control. On the basis of these data, it was con-cluded that the receptor activity of the mutant APN remainedunaltered.
DISCUSSION
A specific cell surface receptor is considered to be one of themost important factors that govern infection by viruses which,like coronaviruses, have a highly restricted host specificity. Inthe present study, we first showed that, unlike pAPN, hAPN isincapable of acting as a receptor for TGEV, thus supporting
I.i I I.I I I..0 0 CY) rI_Lf)In -c--be_ tc)LO) 0) - S', O- r--c'C COU') co r-r- N-00 0)0)0)
J. VIROL.
u-
on February 23, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jvi.asm
.org/D
ownloaded from
DOMAIN OF APN INVOLVED IN TGEV RECEPTOR FUNCTION 5221
N- 0-
NYLKKQVTPLFIHFRNNTNNWREIPENLMDQYSEVNAISTACSNGVPECEEMVSGLFKQWMENPNNNPIHPNLRSTVYCNAIAQGGEEEWDFAWEQFA N PORCINE K YLRKQVEPLFQHFETLTKNWTERPEN MQYSEINAISTACSNGLPQCENLAKTLFDQWMSDPENNPIHPNLRSTIYCNAIAQGGQDQWDFAWGQL
Susceptibilityto TGEV oligont. oligont.
2424/2425 2285/2286
- AP21 I orcinei
AP26
+ AP24
po rc in e
norcine
FIG. 4. Comparison of the porcine domain of the AP24 construct with its human homolog. Amino acid residues that are divergent in the twospecies are in bold characters. The amino acid residues are numbered as in the porcine sequence. The structures of three APN hybrids are shownin the bottom part of the figure. pAPN sequences are represented by open boxes, and hAPN sequences are represented by shaded boxes. TGEVsusceptibility was assayed as described in the legend to Fig. 3. oligont., oligonucleotide.
the view that virus-receptor interaction is a key determinantfor the species tropism of this coronavirus as well as those ofother coronaviruses (7). cDNA sequencing revealed, however,that pAPN has 80% identity at the amino acid level with itshuman homolog. These findings led us to construct a series ofhuman-porcine chimeras in order to identify determinantsessential for the TGEV receptor activity of pAPN. The ap-proach, consisting of establishing interspecies chimeric mole-cules, was previously validated for the mapping of the bindingsites of human immunodeficiency virus, poliovirus, and rhino-virus on their cognate receptors (12, 15, 16, 22). In contrast tothe use of truncated derivatives, the replacement strategy hasthe potential advantage of minimizing structural distortions,thus allowing a more reliable mapping of the region of interest.In this respect, it is worth noting that all of the stably expressedAPN constructs, including five constructs devoid of receptoractivity, were found to be antigenically and enzymaticallyactive and were expressed at the cell surface.The main conclusion drawn from these experiments is that a
chimeric molecule, in which every part but the 717-813 stretchwas replaced by the human homolog, retained a receptoractivity comparable to that of pAPN itself. This hybrid, en-coded by the AP24 construct, was able to confer permissive-ness for TGEV on otherwise refractory MDCK cells (Fig. 3).The most likely interpretation of this result is that amino acidscrucial to virus binding reside in the 97-residue-long regionthus delineated. However, the binding efficiencies of thevarious chimeras were not assayed directly, nor can a possibleinvolvement in the binding of virus to additional determinantscommon to the human and porcine molecules be ruled out.Accordingly, the 717-813 region will subsequently be referredto as the receptor domain. Alignment of pAPN and hAPNsequences revealed a higher degree of divergence within thisregion, 29%, versus 20% for the whole molecule. No particularstructural feature was predicted, except for one potentialAsn-linked glycosylation site located at approximately thesame position in the two molecules. This, together with thefinding that the epitopes for three anti-APN antibodiesmapped within the relevant region (Fig. 6), is suggestive of asurface region. The AP24 clone product contained 28 aminoacid substitutions between human and porcine sequences (Fig.4). Two additional constructs, AP21 and AP26, in which thenumbers of substitutions were reduced to 22 and 18, respec-tively, failed to confer permissiveness for virus (Fig. 3). This
indicates that amino acids critical for receptor activity mayreside in both halves of the 717-813 region. In support of this,the G43 epitope involves amino acids widely separated withinthe 645-837 region, since it is expressed by AP18 constructsbut not by those of AP19 and AP20, which together define thesame porcine region as AP18.The above-described results provide strong evidence that
the receptor and catalytic functions, found to be associatedwith the C and B subunits of pAPN, respectively, involvedistinct domains of the molecule. Consistent with this view isthe observation that the majority (5 of 6) of the anti-APNantibodies blocked the infection without affecting the enzy-matic activity (Fig. 5). Still, it was of interest to establishwhether or not the proteolytic activity associated with the
80
70 -
1-
0
C-U
m
Cu
m
EN
._
._
0
._
0
3-
60 -
50 F
40
30
20
10
0
1 2 3 4 5 6
Dilution (log )3
FIG. 5. Effect of anti-APN antibodies on the aminopeptidase ac-
tivity of pAPN. Threefold dilutions of the indicated monoclonalantibody starting at 10 ,ug/ml were incubated with 0.2 jig of intestinalAPN. APN activity was quantified by cleavage of the chromogenicsubstrate leucine p-nitroanilide. Each assay was performed in dupli-cate.
-Cu G3
-9- G18
-U* G43
-v. 131. T35
- a--- W26
51.13
VOL. 68, 1994
""In
II.-
on February 23, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jvi.asm
.org/D
ownloaded from
5222 DELMAS ET AL.
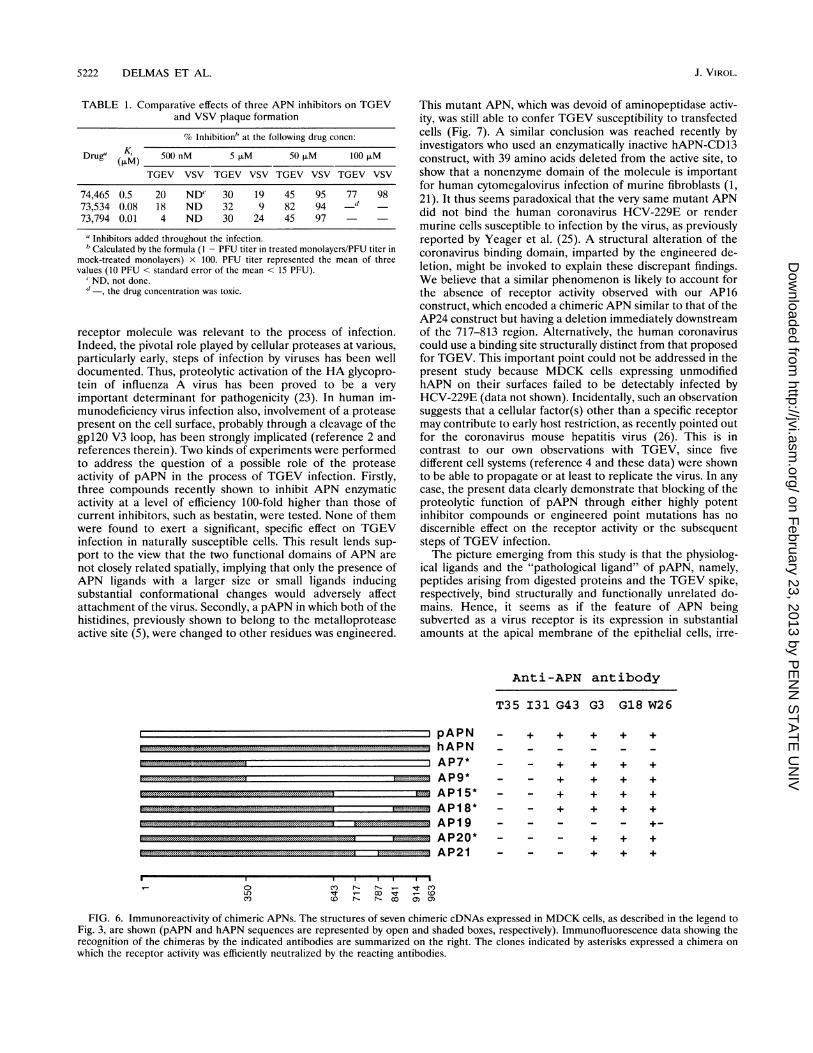
TABLE 1. Comparative effects of three APN inhibitors on TGEVand VSV plaque formation
% Inhibition" at the following drug concn:
Drug" (KIAM 500 nM 5 ,uM 50 ,uM 100 ,uM
TGEV VSV TGEV VSV TGEV VSV TGEV VSV
74,465 0.5 20 NDC 30 19 45 95 77 9873,534 0.08 18 ND 32 9 82 94 -d73,794 0.01 4 ND 30 24 45 97
aInhibitors added throughout the infection.'Calculated by the formula (1 - PFU titer in treated monolayers/PFU titer in
mock-treated monolayers) x 100. PFU titer represented the mean of threevalues (10 PFU < standard error of the mean < 15 PFU).
' ND, not done."-, the drug concentration was toxic.
receptor molecule was relevant to the process of infection.Indeed, the pivotal role played by cellular proteases at various,particularly early, steps of infection by viruses has been welldocumented. Thus, proteolytic activation of the HA glycopro-tein of influenza A virus has been proved to be a veryimportant determinant for pathogenicity (23). In human im-munodeficiency virus infection also, involvement of a proteasepresent on the cell surface, probably through a cleavage of thegpl20 V3 loop, has been strongly implicated (reference 2 andreferences therein). Two kinds of experiments were performedto address the question of a possible role of the proteaseactivity of pAPN in the process of TGEV infection. Firstly,three compounds recently shown to inhibit APN enzymaticactivity at a level of efficiency 100-fold higher than those ofcurrent inhibitors, such as bestatin, were tested. None of themwere found to exert a significant, specific effect on TGEVinfection in naturally susceptible cells. This result lends sup-port to the view that the two functional domains of APN arenot closely related spatially, implying that only the presence ofAPN ligands with a larger size or small ligands inducingsubstantial conformational changes would adversely affectattachment of the virus. Secondly, a pAPN in which both of thehistidines, previously shown to belong to the metalloproteaseactive site (5), were changed to other residues was engineered.
This mutant APN, which was devoid of aminopeptidase activ-ity, was still able to confer TGEV susceptibility to transfectedcells (Fig. 7). A similar conclusion was reached recently byinvestigators who used an enzymatically inactive hAPN-CD13construct, with 39 amino acids deleted from the active site, toshow that a nonenzyme domain of the molecule is importantfor human cytomegalovirus infection of murine fibroblasts (1,21). It thus seems paradoxical that the very same mutant APNdid not bind the human coronavirus HCV-229E or rendermurine cells susceptible to infection by the virus, as previouslyreported by Yeager et al. (25). A structural alteration of thecoronavirus binding domain, imparted by the engineered de-letion, might be invoked to explain these discrepant findings.We believe that a similar phenomenon is likely to account forthe absence of receptor activity observed with our AP16construct, which encoded a chimeric APN similar to that of theAP24 construct but having a deletion immediately downstreamof the 717-813 region. Alternatively, the human coronaviruscould use a binding site structurally distinct from that proposedfor TGEV. This important point could not be addressed in thepresent study because MDCK cells expressing unmodifiedhAPN on their surfaces failed to be detectably infected byHCV-229E (data not shown). Incidentally, such an observationsuggests that a cellular factor(s) other than a specific receptormay contribute to early host restriction, as recently pointed outfor the coronavirus mouse hepatitis virus (26). This is incontrast to our own observations with TGEV, since fivedifferent cell systems (reference 4 and these data) were shownto be able to propagate or at least to replicate the virus. In anycase, the present data clearly demonstrate that blocking of theproteolytic function of pAPN through either highly potentinhibitor compounds or engineered point mutations has nodiscernible effect on the receptor activity or the subsequentsteps of TGEV infection.The picture emerging from this study is that the physiolog-
ical ligands and the "pathological ligand" of pAPN, namely,peptides arising from digested proteins and the TGEV spike,respectively, bind structurally and functionally unrelated do-mains. Hence, it seems as if the feature of APN beingsubverted as a virus receptor is its expression in substantialamounts at the apical membrane of the epithelial cells, irre-
Anti-APN antibody
T35 I31 G43 G3 G18 W26
_ + + + + +
_ _ + + + +
_ _ + + + +
- + + + +_ _ + + + +
_ _ - + + +
- + + +
FIG. 6. Immunoreactivity of chimeric APNs. The structures of seven chimeric cDNAs expressed in MDCK cells, as described in the legend toFig. 3, are shown (pAPN and hAPN sequences are represented by open and shaded boxes, respectively). Immunofluorescence data showing therecognition of the chimeras by the indicated antibodies are summarized on the right. The clones indicated by asterisks expressed a chimera onwhich the receptor activity was efficiently neutralized by the reacting antibodies.
pAPNhAPNAP7*AP9*AP15*AP18*AP19AP20*AP21
0CO
CY) C- - N-
cv N- N- O:)t C0)ca) a)
J. VIROL.
on February 23, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jvi.asm
.org/D
ownloaded from
DOMAIN OF APN INVOLVED IN TGEV RECEPTOR FUNCTION 5223
FIG. 7. Receptor activity of enzymatically defective pAPN. (a) APN activity of the mutated APN. The activity of nonmutated and mutated APNwas determined as described in Material and Methods. (b, c, and d) COS-7 cell cultures transfected with normal APN (b) or mutated APN (c)or mock transfected (d) were infected with TGEV 48 h after transfection. Viral antigen synthesis was examined at 16 h postinfection by animmunofluorescence assay using a mixture of anti-TGEV monoclonal antibodies. O.D., optical density.
spective of its function as a protease. With the informationprovided here, more detailed structural and genetic ap-proaches can be devised to define specific APN residues thatcontact the spike protein during TGEV-APN interaction. Suchknowledge is of potential interest with respect to the develop-ment of antiviral agents to combat diseases caused by TGEVand related animal and human coronaviruses.
ACKNOWLEDGMENTS
We are grateful to Celine Tarnus and Hughes d'Orchymont (MarionMerrell Dow, Strasbourg, France) for making the APN inhibitorsavailable for the experiments and for helpful information. We thankTeit E. Johansen (Copenhagen) for supplying the pTEJ-4 expressionvector and Patrice Vende (Jouy-en-Josas, France) for help in DNAsequencing.
H.S. and O.N. are members of the Biomembrane Research Center,Aarhus University. The project was supported by the Danish MedicalResearch Council. B.D. acknowledges the support of the Commissionof the European Communities for a stay in the laboratory of O.N. andH.S.
REFERENCES1. Ashmun, R. A., L. H. Shapiro, and A. T. Look. 1992. Deletion of
the zinc-binding motif of CD13/aminopeptidase N moleculesresults in loss of epitopes that mediate binding of inhibitoryantibodies. Blood 79:3344-3349.
2. Callebaut, C., B. Krust, E. Jacotot, and A. G. Hovanessian. 1993.T cell activation antigen, CD26, as a cofactor for entry of HIV inCD4+ cells. Science 262:2045-2050.
3. Delmas, B., J. Gelfi, R. L'Haridon, L. K. Vogel, H. Sjostrom, 0.
Noren, and H. Laude. 1992. Aminopeptidase N is a major receptorfor the enteropathogenic coronavirus TGEV. Nature (London)357:417-419.
4. Delmas, B., J. Gelfi, H. Sjostrom, 0. Noren, and H. Laude. 1993.Further characterization of aminopeptidase N as a receptor forcoronaviruses. Adv. Exp. Med. Biol. 342:292-298.
5. Devault, A., V. Sales, C. Nault, A. Beaumont, B. Roques, P. Crine,and G. Boileau. 1988. Exploration of the catalytic site of endopep-tidase 24.11 by site-directed mutagenesis. FEBS Lett. 231:54-58.
6. Ho, S. N., H. D. Hunt, R. M. Horton, J. K. Pullen, and L. R. Pease.1989. Site-directed mutagenesis by overlap extension using thepolymerase chain reaction. Gene 77:51-59.
7. Holmes, K. V., G. Dveksler, S. Gagneten, C. Yeager, S. H. Lin, N.Beauchemin, A. T. Look, R. Ashmun, and C. Dieffenbach. 1993.Coronavirus receptor specificity. Adv. Exp. Med. Biol. 342:55-60.
8. Johansen, T. H., M. S. Scholler, S. Tolstoy, and T. W. Schwartz.1990. Biosynthesis of peptide precursors and protease inhibitorsusing new constitutive and inducible eukaryotic expression vectors.FEBS Lett. 267:289-294.
9. Jongeneel, C. V., J. Bouvier, and A. Bairoch. 1989. A uniquesignature identifies a family of zinc-dependent metallopeptidases.FEBS Lett. 242:211-214.
10. Kenny, A. J., and S. Maroux. 1982. Topology of microvillarmembrane hydrolases of kidney and intestine. Physiol. Rev. 62:91-128.
11. Kueng, W., E. Silber, and U. Eppenberger. 1989. Quantification ofcells cultured on 96-well plates. Anal. Biochem. 182:16-19.
12. Landau, N. R., M. Warton, and D. R. Littman. 1988. The envelopeglycoprotein of the human immunodeficiency virus binds to theimmunoglobulin-like domain of CD4. Nature (London) 334:159-162.
a)2-
co
66
0-o
VOL. 68, 1994
on February 23, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jvi.asm
.org/D
ownloaded from

5224 DELMAS ET AL.
13. Laude, H., J.-M. Chapsal, J. Gelfi, S. Labiau, and J. Grosclaude.1986. Antigenic structure of transmissible gastroenteritis virus. I.Properties of monoclonal antibodies directed against virion pro-teins. J. Gen. Virol. 67:119-130.
14. Look, A. T., R. A. Ashmun, L. H. Shapiro, and S. C. Peiper. 1989.Human myeloid plasma membrane glycoprotein CD13 (gplS0) isidentical to aminopeptidase N. J. Clin. Invest. 83:1299-1307.
15. McClelland, A., J. De Bear, S. C. Yost, A. M. Meyer, C. W. Marlor,and J. M. Greve. 1991. Identification of monoclonal antibodyepitopes and critical residues for rhinovirus binding in domain 1 ofintercellular adhesion molecule 1. Proc. Natl. Acad. Sci. USA88:7993-7997.
16. Morrison, M. E., and V. R. Racaniello. 1992. Molecular cloningand expression of a murine homolog of the human poliovirusreceptor gene. J. Virol. 66:2807-2813.
17. Noren, O., H. Sjostrom, E. M. Danielsen, G. M. Cowell, and H.Skovbjerg. 1986. The enzymes of the enterocyte plasma mem-brane, p. 335-365. In P. Desnuelle (ed.), Molecular and cellularbasis of digestion. Elsevier/North Holland, Amsterdam.
18. Olsen, J., G. Cowell, E. Konigshofer, E. M. Danielsen, J. MollerLaustsen, L. 0. C. Hansen, K. Welinder, J. Engberg, W. Hunziker,M. Spiess, H. Sjostrom, and 0. Noren. 1988. Complete amino acidsequence of human intestinal aminopeptidase N as deduced fromcloned cDNA. FEBS Lett. 238:307-314.
19. Olsen, J., H. Sjostrom, and 0. Noren. 1989. Cloning of the pigaminopeptidase N gene. FEBS Lett. 251:275-281.
19a.Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecularcloning: a laboratory manual, 2nd ed. Cold Spring Harbor Labo-ratory, Cold Spring Harbor, N.Y.
19b.Schallz, C., H. D'Orchymont, M.-F. Jauch, and C. Tarnus. 3-Ami-no-2-tetralone derivates. Novel potent and selective inhibitors ofaminopeptidase-M (EC.3.4.11.2). Arch. Biochem. Biophys., inpress.
20. Semenza, G. 1986. Anchoring and biosynthesis of stalked brushborder membrane proteins: glycosidases and peptidases of entero-cytes and renal tubuli. Annu. Rev. Cell Biol. 2:255-313.
21. Soderberg, C., T. D. Giugni, J. A. Zaia, S. Larsson, J. M.Wahlberg, and E. Moller. 1993. CD13 (human aminopeptidase N)mediates human cytomegalovirus infection. J. Virol. 67:6576-6585.
22. Staunton, D. E., M. L. Dustin, H. P. Erickson, and T. A. Springer.1990. The arrangement of the immunoglobulin-like domains ofICAM-1 and the binding sites for LFA-1. Cell 61:243-254.
23. Webster, R. G., and R. Rott. 1987. Influenza virus A pathogenicity:the pivotal role of hemagglutinin. Cell 50:665-666.
24. Wessels, H. P., G. H. Hansen, C. Fuhrer, A. T. Look, H. Sjostrom,0. Noren, and M. Spiess. 1990. Aminopeptidase N is directlysorted to the apical domain in MDCK cells. J. Cell Biol. 111:2923-2930.
25. Yeager, C. L., R. A. Ashmun, R. K. Williams, C. B. Cardellichio,L. H. Shapiro, A. T. Look, and K. V. Holmes. 1992. Humanaminopeptidase N is a receptor for human coronavirus 229E.Nature (London) 357:420-422.
26. Yokomori, K., M. Asanaka, S. A. Stohlman, and M. M. C. Lai.1993. A spike protein-dependent cellular factor other than theviral receptor is required for mouse hepatitis virus entry. Virology196:45-56.
J. VIROL.
on February 23, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jvi.asm
.org/D
ownloaded from





























