Embed Size (px)
Citation preview

Distribution of FMRFamide-likeImmunoreactivity in the Amphibian
Brain: Comparative Analysis
CLAUDIA PINELLI,1 BIAGIO D’ANIELLO,1 MARIA FIORENTINO,1 PAOLA CALACE,1
MARIA DI MEGLIO,1 LUISA IELA,1 DIETRICH L. MEYER,2 JOSEPH T. BAGNARA,3
AND RAKESH K. RASTOGI1*1Dipartimento di Zoologia, Universita di Napoli Federico II, 80134 Naples, Italy
2Department of Neuroanatomy, School of Medicine, University of Gottingen,37025 Gottingen, Germany
3Department of Cell Biology and Anatomy, College of Medicine, The University of ArizonaHealth Science Center, Tucson, Arizona 85724
ABSTRACTFMRFamide is a small neuropeptide present in particular neurons of the basal forebrain
and midbrain of the vertebrate groups studied, especially fishes and mammals. In order toassess interspecies variation, the distribution of FMRFamide-like immunoreactivity wasstudied in the brains of 13 species of amphibian. Although FMRFamide-immunoreactive (IR)terminals occurred throughout much of the brain, IR cell groups were noted in circumscribedregions of the CNS. In the eight anuran species studied, two major populations of labeledperikarya were observed: one in the septopreoptic area and another one in the caudal portionof the diencephalon. The rostrocaudal extent of both and the number of labeled somata in eachneuronal group displayed species-specific differences. In urodeles and gymnophiones, labeledperikarya were located in the diencephalon, but there were remarkable species differences inthe number of such cells. It is discussed whether sex or season of collection may account forsome of the differences observed. The distribution of FMRFamide-IR perikarya, fibers, andpathways in the brain of anurans, urodeles, and gymnophiones was compared. The existenceof FMRFamide perikarya in the anterior preoptic neuropil and medial septum appeared to bea feature common to all anurans; labeled neurons in the dorsal thalamus, however, may bepresent only in the (viviparous) gymnophione Typhlonectes compressicauda. Cerebrospinalfluid contacting FMRFamide neuronal cell bodies and fibers were observed in each of the threetaxonomic orders. The data are compared with those previously obtained for other groups ofvertebrates. J. Comp. Neurol. 414:275–305, 1999. r 1999 Wiley-Liss, Inc.
Indexing terms: anura; urodela; gymnophiona; neuropeptide; neuroanatomy
A variety of neuropeptides are synthesized by the cen-tral nervous system (CNS) of vertebrates. FMRFamide(Phe-Met-Arg-Phe-amide) has been known for more thantwo decades. It was originally isolated from the nervoussystem of a mollusc, Macrocallista nimbosa (Price andGreenberg, 1977). There is a considerable amount ofevidence indicating the presence of several FMRFamide-related and/or FMRFamide-like peptides in the nervoussystem of vertebrates and invertebrates (Price and Green-berg, 1989; Greenberg and Price, 1992). The immunohisto-chemical localization of this small peptide and/or itsequivalent and related forms has been described in theCNS of species of all classes of vertebrates, from mammalsto cyclostomes (see Dockray et al., 1986; Wicht and North-cutt, 1992; Boersma et al., 1993; Vallarino et al., 1991,
1994, 1995; D’Aniello et al., 1996; Eisthen and Northcutt,1996; Pestarino and Vallarino, 1996; Wright and Demski,1996; Oelschlager et al., 1998, and references therein).FMRFamide is widely distributed in the CNS of manyvertebrates and it has been suggested that FMRFamide-like peptides may play a variety of physiological rolesacting as neurotransmitters, neuromodulators, or neuro-
Grant sponsors: MURST, German Science Foundation, Vigoni Program.Dr. Dietrich L. Meyer passed away on June 8, 1999.*Correspondence to: Dr. Rakesh K. Rastogi, Dipartimento di Zoologia,
Universita di Napoli Federico II, 80134 Napoli, Italy.E-mail: [email protected]
Received 10 May 1999; Revised 6 July 1999; Accepted 21 July 1999
THE JOURNAL OF COMPARATIVE NEUROLOGY 414:275–305 (1999)
r 1999 WILEY-LISS, INC.

hormones (Tang et al., 1984; Wong et al., 1985; Raffa et al.,1986; Roth et al., 1987; Raffa, 1988; Yang and Majane,1990). However, little is known about the roles ofFMRFamide in the amphibian brain.
Immunohistochemical techniques have often been usedto delineate neuroanatomical profiles of several neuropep-tides in the amphibian brain (see Andersen et al., 1992).However, in contrast to other vertebrate groups such asfish and mammals, amphibians have been largely over-looked with respect to the distribution of FMRFamide-likepeptides in the brain. To our knowledge, a rough descrip-tion of the distribution of FMRFamide-like immunoreactiv-ity in the amphibian brain is available only for Ranacatesbeiana (Uchiyama et al., 1988) and R. esculenta(Ekstrom et al., 1990; Vallarino et al., 1993; D’Aniello etal., 1996). Some investigators focused on FMRFamide-immunoreactive (IR) neurons or fibers in the terminalnerve and their placodal origin (Muske and Moore, 1988;
Northcutt and Muske, 1991). One study was directedtoward unveiling FMRFamide-IR retinopetal fibers in R.pipiens (Wirsig-Wiechmann and Basinger, 1988), and an-other one attempted the same in R. catesbeiana andXenopus laevis (Uchiyama et al., 1988). Hitherto, it hasbeen demonstrated in some anurans that FMRFamide-IRand mammalian gonadotropin-releasing hormone (mGnRH)-IR neuronal somata and fibers form a continuum acrossthe terminal nerve-medial basal forebrain-anterior preop-tic area (Muske and Moore, 1988; Rastogi et al., 1996).Furthermore, in the anterior preoptic area of R. esculentamany GnRH neurons were found to be innervated byFMRFamide-IR projections (Rastogi et al., 1996). Thepresence of an FMRFamide innervation of GnRH neuronssuggests a neuromodulatory role for FMRFamide on thereproductive axis via GnRH neurons. At the same time,the existence of numerous FMRFamide terminals in themedian eminence may be indicative of a hypophysiotropic
Abbreviations
A anterior thalamic nucleusAAC lateral acoustic (acousticolateralis) areaAC anterior commissureAMY amygdalaAMYl lateral amygdalaAMYm medial amygdalaAOB accessory olfactory bulbAON anterior olfactory nucleusAPOA anterior preoptic areaAQ aqueductAU rhombencephalic auricleB neuropil of BellonciBNCP bed nucleus of the pallial commissureBNST bed nucleus of the stria terminalisCB cerebellumCC central canalCG central grayCGT thalamic geniculate bodyCP pallial commissureDB nucleus of the diagonal band of BrocaDIE diencephalonE epiphysisEN entopeduncular nucleusES ependymal septumGL glomerular layerGR granular layerHB habenulaHR hindbrain reticular formationHYd dorsal hypothalamusHYv ventral hypothalamusIN interpeduncular nucleusINF infundibulumIR infundibular recessL lateral thalamic nucleusLC locus coeruleusLFB lateral forebrain bundleLIR lateral infundibular recessLL lateral line lobeLR lateral recessLV lateral ventricleMES mesencephalonMFB medial forebrain bundleML mitral cell layerMLB medial longitudinal bundleMSA medial septal areaMSI median sulcusNA nucleus accumbensNB nucleus of BellonciNI nucleus isthmiNO oculomotor nucleusNPM deep midbrain nucleusNT nucleus nervi trochlearisOB olfactory bulbOC optic chiasma
ON optic nerveOT optic tectumOV optic ventricleP posterior thalamic nucleusPC posterior commissurePd dorsal palliumPD pars distalis of the pituitaryPG pretectal grayPl lateral palliumPm medial palliumPOA preoptic areaPOC postoptic commissurePON preoptic nucleusPONa anterior preoptic nucleusPONm magnocellular preoptic nucleusPONp posterior preoptic nucleusPOR preoptic recessPRT pretectumPT posterior tuberclePTN posterior tectal nucleusPVO paraventricular organRB retroflex bundleRHOM rhombencephalonRN raphe nucleusSB solitary bundleSCN suprachiasmatic nucleusSCO subcommissural organSH sulcus limitans of HisSl lateral septumSL spinal lemniscusSm medial septumSM stria medullarisST striatumTE thalamic eminenceTEG tegmentumTEGd dorsal tegmentumTEGi isthmic tegmentumTEGr rostral tegmentumTEGt trigeminal tegmentumTEGv ventral tegmentumTEL telencephalonTHd dorsal thalamusTHN lateral thalamic neuropilTHv ventral thalamusTO optic tractTS torus semicircularisTSc central nucleus of the torus semicircularisTSL laminar nucleus of the torus semicircularisVL ventrolateral thalamic nucleusVM ventromedial thalamic nucleusVN vomeronasal nerveVST vestibulospinal tractIII third ventricleIV fourth ventricle
276 C. PINELLI ET AL.

activity of FMRFamide (Rastogi et al., 1996). In conjunc-tion, this observation and the assumption that FMRF-amide-like peptides play multiple roles in the vertebratebody have triggered this investigation.
A detailed knowledge of the distribution of FMRFamide-like immunoreactivity in the CNS of amphibians mayfacilitate insights into the functional roles played byFMRFamide. This knowledge will also be useful for com-parisons with other neuropeptide systems. The amphibianbrain exhibits a multitude of anatomical differences, inparticular, between the three subgroups (see Roth et al.,1993). This morphological variety has been used for thecomparative investigation of species representing the threeamphibian orders, Anura, Urodela, and Gymnophiona.
We have investigated the localization of FMRFamide-like peptide in the brain of eight species of anurans, threespecies of urodeles, and two species of gymnophiones byimmunohistochemical protocol. Except for the viviparousgymnophione Typhlonectes compressicauda, all other spe-cies discussed below are oviparous and exhibit strikinglydifferent reproductive behaviors. They occupy diverse eco-logical niches.
MATERIALS AND METHODS
Animals
The following species were studied: common toad, Bufobufo (Anura, Bufonidae; collected in the field, Italy); greenfrog, R. esculenta (Anura, Ranidae; collected in the field,Italy); Mexican leaf frog, Pachymedusa dacnicolor (Anura,Hylidae, Phyllomedusinae; from a breeding colony inTucson, Az); ornated horned frog, Ceratophrys ornata(Anura, Leptodactylidae, Ceratophryinae; from a commer-cial dealer); Puerto Rican frog, Eleutherodactylus coqui(Anura, Leptodactylidae, Telmatobiinae; gift of Dr. E.Kicliter, University of Puerto Rico); African clawed frog,Xenopus laevis (Anura, Pipidae, Xenopodinae; from abreeding colony, Germany); Surinam toad, Pipa pipa(Anura, Pipidae, Pipinae; from a commercial dealer); treefrog, Hyla crucifer (Anura, Hylidae; from a commercialdealer); crested newt, Triturus carnifex (Urodela, Sala-mandridae; collected in the field, Italy); smooth newt, T.vulgaris (Urodela, Salamandridae; collected in Poland,permit OP. 4072/174/92 issued to Prof. Rybo, Polish Acad-emy of Sciences, Warsaw); tiger salamander, Ambystomatigrinum (Urodela, Ambystomatidae; from a commercialdealer); burrowing oviparous caecilian, Ichthyophis bed-domei (Gymnophiona, Ichthyophiidae, Ichthyophiinae;collected in South India); viviparous aquatic caecilian,Typhlonectes compressicauda (Gymnophiona, Typhlonec-tidae; from a commercial dealer). The sample size was 10,16, 4, 2, 2, 5, 2, 2, 5, 3, 2, 4, and 2, respectively. Animalswere anesthetized by immersion in MS 222 (tricainemethanesulfonate, Sigma, St. Louis, MO) and either thebrains were quickly removed and immersed in Bouin’sfixative overnight at room temperature, or animals weretranscardially perfused with 4% cold paraformaldehyde in0.1 M phosphate buffer saline (PBS; pH 7.4). Preliminaryexamination in R. esculenta and B. bufo had shown thatboth fixatives yielded similar results as far as the distribu-tion of FMRFamide-IR elements was concerned. Brainsfixed in Bouin’s fluid were processed for paraffin embed-ding, whereas paraformaldehyde-perfused brains wereremoved from the skull and postfixed for 4 hours at 4°C,rinsed in PBS, and cryoprotected overnight in cold 30%
sucrose solution in PBS before they were placed in embed-ding medium (Tissue-Tek O.C.T. Compound, Miles Inc.,Elkhart, IN) for 1 hour for cryostat sectioning at 224°C.Serial sections (7 µm for paraffin-embedded brains and 15or 20 µm for paraformaldehyde-fixed brains) were mountedon glass slides. From some brains, two series of alternatesections were prepared. Transverse sections of the brainwere used for all species. In addition, one brain each of B.bufo, R. esculenta, P. dacnicolor, T. carnifex, I. beddomei,and T. compressicauda was sectioned sagittally, and one ofE. coqui was sectioned in the horizontal plane. The entireseries of sections of each species used for cell counts wasanalyzed to determine the number of labeled neurons.
Immunohistochemistry
Paraffin sections were rehydrated. All sections weretreated with 1% normal goat serum in PBS for 30 minutesfollowed by 1% H2O2 in PBS for 3 minutes. After a rinse inPBS they were incubated with primary antiserum (rabbitantiserum raised against FMRFamide; dilution between1:10,000 and 1:20,000) overnight at 4°C. The succeedingsteps consisted of treatments with biotinylated goat anti-rabbit IgG (1:150) and avidin-biotinylated horseradishperoxidase complex, using ImmunoPure ABC Staining Kit(Pierce Co., Rockford, IL). The reaction product was visual-ized with diaminobenzidine. In several cases the immuno-reaction was intensified by addition of nickel chloride tothe incubation medium. Sections were lightly counter-stained with either Mayer’s haemalum or neutral red.After dehydration sections were mounted with eitherEukitt or Permount. In addition, one series of paraffinsections from two P. dacnicolor were also processed withfluorescein isothiocyanate-conjugated goat anti-rabbitg-globulin (1:200) for fluorescence microscopy. These sec-tions were mounted in glycerol-PBS (1:4) and examinedwith a Leica DMRB fluorescence microscope. All photomi-crographs are original prints, without alteration.
Anti-FMRFamide was obtained from two differentsources: Phoenix (Mountain View, CA) and Incstar Corp.(Stillwater). Both antisera were screened on B. bufo and R.esculenta sections (paraffin and cryostat), with and with-out metal intensification of the reaction product. Evenusing different methods, the spatial distribution of immu-noreactive elements appeared substantially similar withineach species tested. Besides B. bufo and R. esculenta, bothantisera were used for P. dacnicolor and I. beddomei,whereas only Incstar antiserum was used for the remain-ing species.
Specificity of immunoreaction
To analyze the specificity of immunoreaction, sectionsfrom B. bufo, R. esculenta, P. dacnicolor, T. carnifex, and I.beddomei brains were processed as follows: primary antise-rum replaced by PBS or normal goat serum; omission ofsecondary antibody; incubation with primary antiserum(1:10,000) preabsorbed overnight with syntheticFMRFamide (5 and 1 µM; Peninsula Labs, Belmont, CA).It has previously been suggested that anti-FMRFamidemay cross-react, to some degree, with neuropeptide Y(NPY) (Dockray and Williams, 1983; O’Donahue et al.,1984; Triepel and Grimmelikhuijzen, 1984; Chen et al.,1989; Shen and Cheng, 1989; Fischer et al., 1996). To testthis assumption, some sections from R. esculenta, P. dac-nicolor, and I. beddomei brains were incubated withprimary antiserum (1:10,000), preabsorbed overnight with
FMRFAMIDE IN AMPHIBIAN BRAIN 277

synthetic NPY (5, 1, and 0.1 µM; Peninsula), and alternatesections were reacted with anti-NPY (Affiniti, Notting-ham, UK) as well as with anti-FMRFamide. We alsoconducted a heterologous preabsorption test with syn-thetic GnRH (5LHRH, 5 µM, Sigma) in some sections of R.esculenta, P. dacnicolor, and I. beddomei brains, sinceFMRFamide and GnRH immunoreactivities coexist interminal nerve and forebrain of some vertebrates (Stell etal., 1984; Muske and Moore, 1988; Wright and Demski,1996; Oelschlager et al., 1998) and in some neurons of theanterior preoptic area of the R. esculenta brain (Rastogi etal., 1996).
Nomenclature
The identification of possibly homologous brain struc-tures in species belonging to different amphibian sub-groups is still debated. We used several sources, includingHerrick (1948), Kemali and Braitenberg (1969), Northcuttand Kicliter (1980), Wada et al. (1980), Neary and North-cutt (1983), Frontera (1952), Clairambault and Anteıno(1970), Opdam et al. (1976), Neary (1990), Lazar et al.(1993), Puelles et al. (1996), Marın et al. (1997) for anuranbrains; Opdam and Nieuwenhuys (1976), Wicht and Him-stedt (1988), Clairambault and Timmel (1990), Lowry etal. (1997) for urodele brains; Zilles et al. (1981), Clairam-bault et al. (1994), Naujoks-Manteuffel and Meyer (1996),Pinelli et al. (1997) for the caecilian brains. This studyprotocol complied with legislation on the care and use ofanimals in research.
RESULTS
Specificity tests
The lack of immunostaining during control proceduresin which the primary antiserum was replaced by normalgoat serum or PBS, or when the secondary antiserum wasomitted, clearly indicates that the procedure is specific andvisualizes FMRFamide-like material in the brain. Screen-ing of homologous and heterologous preabsorption testsrevealed that preabsorption with a homologous antigen(FMRFamide) completely eliminated the immunostaining.Preabsorption with GnRH did not affect immunostaining,but antisera activity was reduced when preabsorbed with5 µM concentration of NPY, and hardly any reduction wasnoted with a 0.1 µM concentration of NPY. In spite of thepossibility of unspecific staining, we feel that the antibod-ies (anti-FMRFamide) used did not visualize NPY-likeneuropeptide. When alternate sections were incubatedwith anti-FMRFamide and anti-NPY different sets ofneurons were stained. Thus, we conclude that cells immu-nostained with anti-FMRFamide express a FMRFamide-like molecule. We do not know whether we have visualizedthe same molecule against which the antibody was raisedand hence we define it as FMRFamide-like molecule. Forconvenience we apply the term ‘‘FMRFamide’’.
Distribution of FMRFamide-likeimmunoreactivity
In the following description no reference is made to sexor the period of animals’ collection. It may, however, beworth mentioning that an examination of brains fromadult males and females of R. esculenta and B. bufocollected in summer, late autumn, and early spring re-vealed no difference of the distribution of FMRFamide-IRelements during different seasons and among genders.Furthermore, within each order the relative gross size of
the brain is about the same except that it is slightly biggerin B. bufo among anurans, and A. tigrinum among urodels.
Anura.
Rana esculenta [Figures 1 and 2, and Table 1]. Thedistribution of FMRFamide-IR neurons and fibers in there-examined green frog was identical to that reportedpreviously (Ekstrom et al., 1990; Vallarino et al., 1993;D’Aniello et al., 1996). However, the earlier studies pro-vided only a rough outline of the distribution profile.The present analysis was intended to be detailed.FMRFamide-IR neurons formed two well-defined aggre-gates. The rostral subpopulation of IR neurons consisted ofa continuum starting in the medial septum and thediagonal band of Broca that continued caudal to theanterior preoptic area, immediately rostral to the anteriorcommissure, in the region corresponding to the laminaterminalis (Fig. 1C,D). Labeled fusiform and bipolar neu-rons were located in the innermost part of the medialseptum and along the dorsolateral and ventral borders ofthe diagonal band of Broca and a few were positioned atthe ventromedial surface of the telencephalon. In theneuropil of the anterior preoptic area, just dorsal to theanterior preoptic nucleus, IR neurons were more numer-ous. The majority was elongated and bipolar. Togetherwith the rostral labeled neurons about 200 (between 203and 219 in four brains) were counted. The more caudalsubpopulation of FMRFamide-IR neurons was compar-atively bigger, both numerically (474–509 labeled cellbodies) as well as regarding its rostrocaudal extent. Thisincludes the suprachiasmatic nucleus rostrally, and thepostchiasmatic, dorsal and ventral, infundibular hypothala-mus caudally (Figs. 1G–J, 2). Neurons of this group weresituated in a periependymal position on each side of thethird ventricle, and several of them were observed incontact with the ventricular lumen. Labeled somata weremost frequent in the suprachiasmatic nucleus becomingprogressively less numerous caudally in the ventral hypo-thalamus. No labeled neurons were observed in the mid-and hindbrain.
While some scattered fibers were present in the olfactorybulb (mainly in the granular cell layer), in the telencepha-lon IR fibers innervated the medial septum, the diagonalband of Broca, the dorsal and the lateral pallium, and theventral striatum (Fig. 1A–D). The caudalmost portion ofthe septum, overlying the rostralmost anterior preopticnucleus, was heavily innervated by labeled fibers. Thesefibers formed a continuum with those located ventrolater-ally in the medial amygdala which continued furthercaudal ventrolaterally to the preoptic nucleus. We did notobserve labeled fibers in the anterior preoptic nucleus,that is, in the periventricular cell column of the anteriorpreoptic recess. From the lateral preoptic area, IR fibersjoined those innervating the ventral striatum and thelateral forebrain bundle. Some labeled fibers were locatedin the bed nucleus of the pallial commissure, and somewere observed in the anterior commissure (Fig. 1E). In thediencephalon a dense plexus of thick beaded fibers inner-vated the area corresponding to the nucleus of Bellonci(Fig. 1F). Some thin fibers were present in the retroflexbundle, and many were located in the periependymalstrips of white matter innervating many unlabeled neu-rons of the thalamic nuclei. The ventrally located magno-cellular part of the preoptic nucleus was innervated, andmany IR fibers were observed in the ventrolateral neuro-pil. They formed a continuum with those in the nucleus of
278 C. PINELLI ET AL.

Bellonci through thin fibers running along the lateral wallof the diencephalon. Another plexus of labeled fibers wasobserved in the neuropil just lateral and ventral to thesuprachiasmatic nucleus (Fig. 1G). In transverse sectionsthis plexus of fibers continued with those located in theperipheral neuropil which, in turn, joined those present inthe area adjacent to the epiphysis (the epiphysial paren-chyma had little FMRFamide-immunoreactivity). An-terodorsal and dorsolateral thalamic nuclei were moder-ately innervated. Some scattered FMRFamide fibers werepresent in the optic chiasma and optic tract (Fig. 1G). Thepretectal gray was rich in IR fibers. The posterior tuberclewas lightly innervated and, in its vicinity, labeled cellbodies were observed in the dorsal hypothalamic nucleus(Fig. 1H). The rostral part of torus semicircularis, corre-sponding to the laminar nucleus, was heavily innervated,and a dense innervation in the optic tectum showed thelaminated pattern in reflecting its layered organization(Fig. 1I,J). The rostral tegmental area contained numer-ous labeled fibers and a dense network of IR fibers was
formed more caudally, in the central nucleus of the torussemicircularis (Fig. 1J,K). In the isthmic region, isthmicand interpeduncular nuclei were not innervated, whereasa light innervation of ventral tegmentum and ventrolat-eral mesencephalon was observed. The cerebellum dis-played moderate FMRFamide innervation. In the rhomb-encephalon, the central gray contained many labeledfibers, and fiber tracts were observed across the lateralreticular formation and the ventrolateral rhombencepha-lon (Fig. 1L,M). The solitary bundle and the lateralacoustic area showed hardly any IR fibers.
Pachymedusa dacnicolor [Figures 1, 2, and 3, and Table1]. The distribution of FMRFamide-IR perikarya andfibers in the brain of the Mexican leaf frog was very similarto that of R. esculenta (Figs. 1A–M, 2; Table 1). However,labeled neurons were less numerous, and IR fibers weresparse. In the one brain analyzed, the number ofFMRFamide neurons was 68 in the septopreoptic groupand 129 in the posterior diencephalon. Rostrally,FMRFamide neurons were situated in the medial septum
TABLE 1. Summary Distribution of FMRFamide-like Immunoreactivity in the Brain of Eight Anurans1
Brain areaR.
esculentaP.
dacnicolorP.
pipaX.
laevisH.
cruciferB.
bufoE.
coquiC.
ornata
ForebrainOlfactory bulb – 1 – 1 – 11 – 11 – 11 – 11 – – – 1Accessory olfactory bulb – – – – – – – – – – – – – – – –
TelencephalonDorsal pallium – 1 – – – 1 – 1 – 1 – 1 – – – –Medial pallium – – – – – 11 – – – 1 – – – – – 1Lateral pallium – 1 – – – 1 – 1 – 11 – 1 – 1 – 1Striatum – 1 – 1 – 1 – 1 – 1 – 1 – 1 – 11Medial septum o 11 oo 1 oo 11 oo 11 oo 1 o 1 o 1 o 1Lateral septum – – – – – 11 – 1 – 1 – 1 – – – 1Diagonal band of Broca o 1 o 1 – 11 – 1 o 1 o 1 – 1 – 1Mediobasal telencephalon o 1 – 1 – 11 – 11 o 11 o 1 – 1 – 1Anterior commissure – 1 – 1 – 11 – 11 – – – – – – – 1Amygdala
Lateral – – – – – 11 – 1 – – – 1 – – – 11Medial – 1 – 1 – 11 – 1 – 1 – 1 – 1 – 11
DiencephalonEpiphysis – 1 – – – – – 1 – – – – – – – –Habenulae – – – – – 1 – 1 – – – – – – – –Habenular commissure – 1 – – – 1 – 1 – 1 – 1 – 1 – 1Dorsal thalamus – 1 – 1 – 1 – 1 – 1 – 1 – 1 – 1Ventral thalamus – 1 – – – 1 – 1 – 1 – 1 – 1 – 1Lateral thalamic neuropil – 11 – 11 – 11 – 1 – 1 – 1 – 1 – 1Nucleus of Bellonci – 111 – 11 – 111 – 11 – 111 – 11 – 11 o 11Neuropil of Bellonci – 111 – 11 – 111 – 111 – 111 – 111 – 11 – 11Anterior preoptic area oo 11 oo 11 ooo 111 oo 111 ooo 111 oo 11 o 11 o 111Preoptic nucleus
Anterior – – – – – 1 oo 11 – 11 – 11 – 1 – 1Magnocellular/posterior – 1 – 11 – – – 1 o 1 o 1 o 1 – 1
Suprachiasmatic nucleus ooo 11 ooo 1 oo 1 ooo 1 ooo 11 ooo 111 oo 1 ooo 11Entopeduncular nucleus – 1 – 1 – 11 – 1 – 1 – – – – – –Geniculate body – 11 – 11 – 11 – 11 – 11 – 1 – 1 – 1Dorsal hypothalamus oo 1 oo 1 – 1 – 11 o 1 oo 11 – 1 o 1Ventral hypothalamus o 1 – 1 – 11 – 11 oo 1 ooo 11 o 1 o 1Optic chiasma – 1 o 1 o 11 – – – 11 o 1 – – – –Optic tract – 1 – 1 – 1 – – – 11 – – – – – –Median eminence – 11 – 1 – 11 – 111 – 1 – 11 – 1 – 1
Posterior tubercle – 1 – – – 1 – 1 – 1 – 1 – 1 – 1Pretectum – 11 – 1 – 11 – 1 – 1 – 1 – 1 – 1Midbrain
Optic tectum – 11 – 1 – 111 – 11 – 11 – 11 – 1 – 11Torus semicircularis – 11 – 11 – 1 – 1 – 111 – 11 – 1 – 11Tegmentum
Dorsal – 11 – 1 o 1 – 1 – 11 – 1 – 1 – 1Ventral – 11 – 1 – 11 – 1 – 11 – 1 – 1 o 1
Interpeduncular nucleus – – – – – – – 1 – 11 – 1 – – – 1Oculomotor nucleus – 11 – – – 11 – 11 – 1 – 1 – 1 – 1Isthmic nucleus – – – – – – – – – – – – – – – –
HindbrainCerebellum – 1 – 1 – 11 – – – 11 – 1 – 1 – 1Raphe nuclei – – – – – 1 – 1 – – – 1 – – – –Central gray – 1 – 1 – 11 – 1 – 11 – 1 – 1 – 1Reticular formation – 1 – 11 – 11 – 1 – 11 – 1 – 1 – 1Acousticolateral area – 1 – 1 – – – 1 – 11 – 1 – – – 1Solitary bundle – – – – – – – – – 11 – 1 – – – –
1Density of immunostained perikarya is indicated by o’s, fibers by 1’s. Dash denotes absence.
FMRFAMIDE IN AMPHIBIAN BRAIN 279

and in the diagonal band of Broca. They formed a con-tinuum with cells located in the rostralmost portion of theanterior preoptic area and a few labeled cell bodies withinthe periventricular cell layers of the anterior preopticrecess (Fig. 1C,D). The more caudal group of labeledneurons comprised those in the suprachiasmatic nucleus,adjacent to the ependyma. It continued caudal into thedorsal hypothalamus, just behind the optic chiasma (Fig.1G,H). Further caudal, the infundibulum lacked IR cellbodies (Fig. 1I). Labeled groups of neurons on each side ofthe suprachiasmatic nucleus merged below the third ven-tricle (Figs. 1G, 3). Some had fibers contacting the thirdventricle (Fig. 3). Occasionally, FMRFamide cells wereobserved in the optic chiasma (Fig. 1G). The olfactory bulb,the telencephalon, and the anterior preoptic area were notas densely innervated as in R. esculenta (Fig. 1A–E). Thepallium was without FMRFamide innervation. At habenu-lar levels, there were random fibers in the retroflex bundle,whilst the neuropil of Bellonci was richly innervated by amixture of thin and thick IR fibers. More ventrally, numer-ous such fibers were located close to the ependyma of thethird ventricle, and a few fibers were present in the lateralpreoptic neuropil (Fig. 1F). The dorsal and ventral tegmen-tum were sparsely innervated and the optic tectum showedscattered fibers between cell layers and in the superficial
neuropil (Fig. 1I–K). The rostral tegmentum, the centralnucleus of the torus semicircularis, and the area dorsome-dial to the isthmic nuclei were densely innervated by fineand coarse varicose fibers. The cerebellum was sparselyinnervated. A few fibers were observed among the periven-tricular cell layers of the fourth ventricle (Fig. 1L–M).Further caudal, however, the reticular formation in themedulla and the dorsomedial gray, adjacent to raphenuclei, were densely innervated by thin and sparselyramified fibers.
Pipa pipa [Figures 1, 4, 5, and 6A, and Table 1]. TheFMRFamide system of this frog was one of the most diffuseof all amphibians studied, regarding the density of neu-rons as well as fibers. Two major groups of FMRFamideperikarya were distinguished. The rostral group spannedthe medial septum as well as the dorsal and lateralanterior preoptic area (Fig. 1C,D). The caudal groupwas restricted to regions at the level of the suprachias-matic nucleus (Fig. 1G). In the anterior preoptic area,FMRFamide neurons were distributed in the neuropildorsal and dorsolateral to the preoptic recess (Figs. 1D,4A). In the suprachiasmatic nucleus, they were mostlyfound in the periventricular cell layers, with some neuronsbeing located in the ependyma (Figs. 1G, 4B). In the onebrain analyzed, the FMRFamide perikarya totaled more
Fig. 1. A–M: Camera lucida drawings of transverse sections (rostro-caudal sequence) of the brain of four species of anuran amphibiansgiving the distribution of FMRFamide-IR cell bodies (large dots) andfibers (dashes and fine dots). The density of large dots and dashes aswell as fine dots corresponds to low, moderate, and high densities as
described in the results section. Thin lines represent the boundariesbetween neuropil and cell columns. The levels of sections A–M of R.esculenta, P. dacnicolor, and H. crucifer are indicated in the sketch of alateral view of the brain in Figure 2; those of the P. pipa brain are atsimilar levels. For abbreviations, see list.
280 C. PINELLI ET AL.

than 650 in the septopreoptic group and 316 in thesuprachiasmatic group. In contrast to some other anurans,but resembling P. dacnicolor, the postchiasmatic infundibu-lar hypothalamus did not contain any IR neurons (Fig.1I,J). However, several (as many as 13) solitary IR neuronswere observed in the neuropil lateral to the suprachias-matic nucleus in the vicinity of the lateral forebrain bundle(Fig. 1G) and, occasionally, in the optic chiasma (Fig. 4C)as well as in the neuropil of the rostrolateral tegmentum,just below the torus semicircularis (Fig. 1J). Long andthick-beaded IR fibers were distributed in all layers of theolfactory bulb (Figs. 1A, 5A) as well as in the lateral,dorsal, and medial pallium, in the rostral- and midtelen-cephalon (Figs. 1B,C). Caudal to the anterior preopticarea, the pallium was devoid of such fibers. The medialpallium, medial septum, lateral septum, diagonal band ofBroca, medial amygdala, anterior lateral and dorsal preop-tic area were heavily innervated by FMRFamide fibers.Only in the rostral half of the telencephalon, FMRFamidefibers innervated the ependyma as well as the unlabeledperikarya of the cell columns around the lateral ventricles(Figs. 1A–E, 5B). Dense fiber nets were present in theentire neuropil of the anterior preoptic area, the nucleus ofBellonci (Fig. 5C), caudal ventral hypothalamus, pretec-tum, tectum, ventral tegmentum, cerebellar gray, raphenuclei, and lateral rhombencephalic area (Fig. 1D–M). Themedian eminence was packed with FMRFamide-IR termi-nals (Fig. 5D). The torus semicircularis, which is not asconspicuous as in other anurans, was also well innervated
(Fig. 1J). Unlike other anurans studied, the area aroundthe subcommissural organ was heavily innervated in thisspecies (Figs. 1H, 5E). The optic tectum did not exhibit thelaminated pattern typical of most anurans; cells werenot organized into layers, and heavy caliber beadedFMRFamide fibers were found across its entire extent.They were distributed in a haphazard fashion (Fig. 6A).
Xenopus laevis [Figures 6B and 7, and Table 1]. TheAfrican clawed frog with its 189 FMRFamide neurons inthe septopreoptic group and 448 neurons in the suprachias-matic-infundibular cluster was another anuran speciessupplied with a vast network of FMRFamide elements.Few labeled neurons were observed in the medial septum,but they were numerous in the dorsolateral cell column ofthe anterior preoptic nucleus (Fig. 7A). The neurites ofthese cells were long and innervated the entire lateralanterior preoptic neuropil. Few FMRFamide neurons wereobserved in subependymal positions, and the ependymal
Figure 1 (Continued)
Fig. 2. Camera lucida diagram of a near midline sagittal section ofR. esculenta, P. dacnicolor, and H. crucifer. The brain’s profile isoutlined by dashed lines. The distribution of FMRFamide-IR peri-karya (large dots) is viewed from lateral. Arrows indicate areasinnervated by FMRFamide-IR fibers. Uppercase letters above thebrains refer to levels of transverse sections given in Figure 1. Forabbreviations, see list.
FMRFAMIDE IN AMPHIBIAN BRAIN 281

FMRFamide innervation around the preoptic recess wasrather weak. In the rostral suprachiasmatic nucleusFMRFamide neurons occupied a ventrolateral subependy-mal position. They were furnished with long processesinnervating the ependymal cell lining (Fig. 7B). In thecaudalmost portion of the suprachiasmatic nucleus manyFMRFamide neurons were observed amidst unlabeledependymal cells; they contacted the ventricle. Like inP. pipa, some multipolar FMRFamide-IR neurons withlong arborized processes were observed lateral to thesuprachiasmatic nucleus (Fig. 7C). The accessory olfactorybulb was devoid of IR fibers, as in R. esculenta and otheranurans, but the olfactory bulb was richly supplied. Cau-dally, the fibers were contiguous with IR processes in thelateral pallium, striatum, nucleus accumbens, medial sep-tum, and the diagonal band of Broca. In the anteriorpreoptic area, conspicuous fibers encircled the preopticnucleus. They penetrated the nucleus to its innermost celllayer, but a direct contact with the lumen of the preopticrecess was not found. The optic tectum showed somedegree of lamination and FMRFamide fibers were distrib-uted between as well as across cell layers (Fig. 6B). Thehindbrain was very poorly innervated by IR fibers. Thecerebellum lacked such fibers, and there were only a fewendings in the raphe nuclei, rhombencephalic central gray,lateral acoustic area, and reticular formation. The oculomo-tor nuclei were richly innervated.
Hyla crucifer [Figures 1, 2, 6C, 8, and 9, and Table 1].The one brain of this hylid that was analyzed contained252 FMRFamide neurons in the septopreoptic aggregateand 458 in the posterior diencephalon. Rostrocaudally, IRneurons were observed in the medial septum, diagonalband of Broca, and ventromedial midtelencephalon. Theyformed a continuum with somata that were located in the
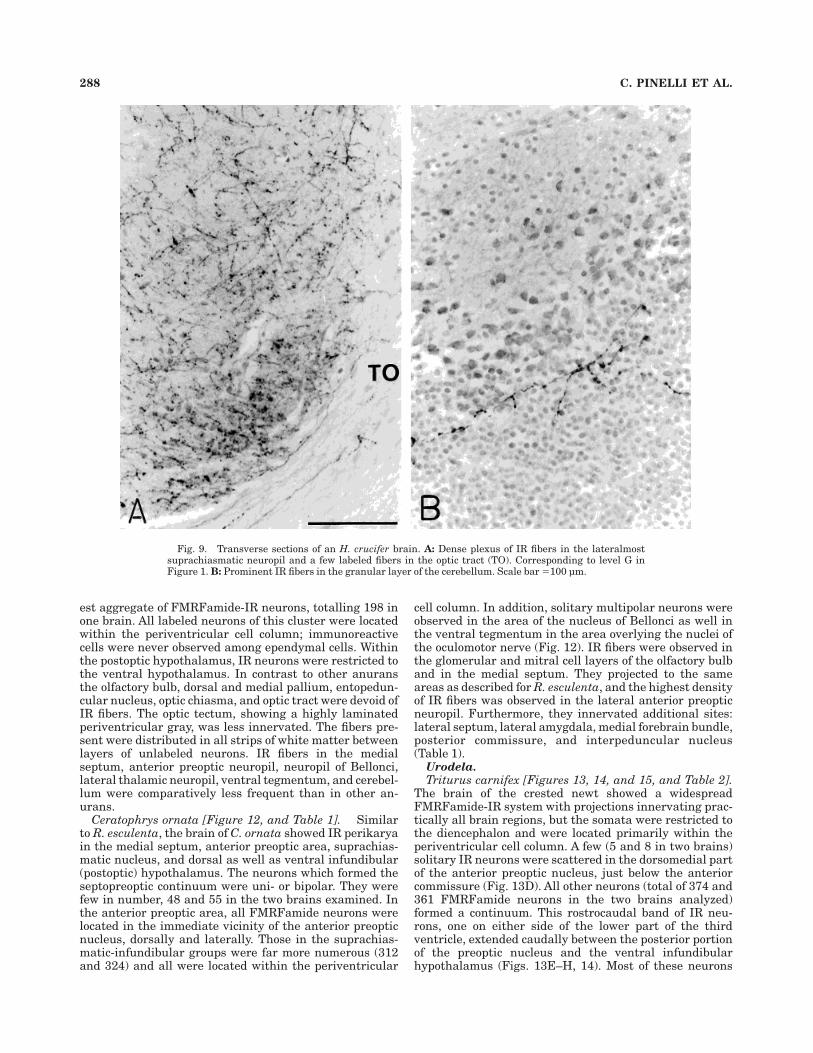
neuropil overlying the anterior preoptic nucleus (Fig.1C,D). This neuronal band extended to the level of theanterior commissure (Fig. 1E). Ir neurons in the medialseptum were bipolar with long dorsoventral projections(Fig. 8A). Labeled perikarya in the ventromedial telen-cephalon were located adjacent to the brain’s surface. Atthe caudal level of the medial septum, just rostral to theanterior commissure, IR neurons were unipolar with longprocesses oriented caudally and ventrally (Fig. 8B). Thesefibers joined those that emerged from neurons positionedin the rostralmost anterior preoptic area, encircled theentire preoptic nucleus, and extended below the preopticrecess (Fig. 8C). Caudal to this level, mostly bipolar IRneurons were present in the neuropil just above thepreoptic recess (Fig. 8D). No labeled perikarya were ob-served in the periventricular cell layers of this area, but IRfibers innervated many unlabeled cells in the vicinity ofthe preoptic recess. In its caudal part, there was a conspicu-ous accumulation of IR fibers in subependymal positions(Fig. 8C,D). At the level of the suprachiasmatic nucleus, IRneuronal groups located laterally in the medial and lateralregion of the periventricular cell column were displacedventrally (Fig. 1G). In this nucleus, as in the preopticnucleus, we observed a dense accumulation of immunore-activity around the ependymal cell layer. Numerous IRneurons were observed in the ventral infundibular hypo-thalamus, and a few were noted in the dorsal infundibularhypothalamus; all neurons were positioned in the periven-tricular cell layers (Fig. 1I). The olfactory bulbs and therostral portion of the telencephalon were rich in IR fibers(Fig. 1A–C); the caudalmost part of the telencephalon wasdevoid of them (Fig. 1D). In the optic tectum, whichshowed a high degree of lamination, FMRFamide fiberswere observed between the cell layers and in the superfi-cial neuropil (Figs. 1I–K, 6C). The lateral suprachiasmaticneuropil showed a heavy innervation by IR fibers, prob-ably the densest of all anurans studied. A few long and thinfibers were noted among optic tract fibers (Fig. 9A). Almostall other major midbrain areas had FMRFamide innerva-tion. In the hindbrain, the cerebellum showed long beadedfibers crossing the entire extent of the granular layer (Fig.9B), and there was a heavy innervation of the lateralacoustic area and solitary bundle (Fig. 1L,M).
Bufo bufo [Figures 6D and 10, and Table 1]. As inR. esculenta, H. crucifer, and other anurans, the rostral-most IR neurons in the common toad were observed in themedial septum and the diagonal band of Broca (Table 1).Most of these cells were bipolar and had pyriform somatawith long processes. These scattered neurons formed acontinuum with more numerous IR neurons in the ante-rior preoptic area. As assessed in four brains (two malesand two females) they totalled 263 to 279. In the anteriorpreoptic area, besides many IR cells in the dorsal andlateral anterior preoptic neuropil, many labeled cells werelocated beneath the preoptic recess, within the subependy-mal cell layers (Fig. 10A). Fibers of these neurons inner-vated the underlying and ventrolateral neuropil, and weretraced as far deep as the ependymal cell lining. Anothersubpopulation of IR neurons was observed in the caudalpart of the diencephalon. As in R. esculenta, these formed amagnocellular preoptic–suprachiasmatic–caudal infun-dibular–hypothalamic continuum. This group, containingmore IR somata (between 348 and 367 neurons) than therostrally located set, encompassed bipolar and unipolar aswell several irregularly shaped neurons. In the magnocel-
Fig. 3. FMRFamide-immunofluorescent neurons in the P. dac-nicolor suprachiasmatic nucleus; transverse section. The third ven-tricle is depicted by an arrow. Section corresponds to level G in Figures1 and 2. Scale bar 5 50 µm.
282 C. PINELLI ET AL.

lular preoptic nucleus, the FMRFamide neurons wereprimarily located in close proximity to the ependyma, withsome IR cell bodies among ependymal cells (Fig. 10B).Those in the suprachiasmatic nucleus were positioned
somewhat more distantly from the ependymal lining andshowed fiber projections contacting the ventricular lumen(Fig. 10C). In the infundibular hypothalamus, the IRneurons were by far the most numerous of all anurans
Fig. 4. Transverse sections of a P. pipa brain. A: FMRFamide-IRneurons are distributed dorsal and dorsolateral to the preoptic recess(asterisk); corresponds to level D in Figure 1. The periventricular cellcolumn is innervated by a few IR fibers. B: FMRFamide-IR neurons inthe suprachiasmatic nucleus, with a few cell bodies located amidst
ependymal cells and a neuron with its neurite (arrow) contacting theventricle. Corresponds to level G in Figure 1. C: FMRFa-IR neurons(arrows) and fibers in the optic chiasma. Corresponds to the same levelas panel B. Scale bars 5 100 µm in A,C; 50 µm in B.
FMRFAMIDE IN AMPHIBIAN BRAIN 283

investigated. They had different shapes and were foundprimarily around the lateral infundibular recess, at vari-able distances from the ependymal cell lining (Fig. 10D).Some scattered IR neurons were observed in the opticchiasma. Like in other anurans labeled fibers were notedin various brain areas. FMRFamide innervation of thefollowing forebrain areas was observed: olfactory bulb(glomerular, mitral, and granular cell layers), dorsal aswell as lateral pallium, medial septum, striatum, diagonalband of Broca, amygdala, habenular commissure, dorsalas well as ventral thalamus, neuropil of Bellonci, lateraland dorsal as well as ventrolateral anterior preoptic area,lateral thalamic neuropil, geniculate body, and dorsal aswell as ventral hypothalamus. In addition, a few labeledfibers were observed in the optic chiasma and optic tract,and IR material was abundant in the median eminence.The pretectum and posterior tubercle contained only fewlabeled fibers. In the midbrain, the optic tectum displayedFMRFamide innervation in a laminar organization, andIR fibers were observed between the inner cell layers and,
rarely, in the outer neuropil (Fig. 6D). Ir fibers werefrequent in different zones of the midbrain tegmentum.The central tegmental nucleus was the most heavilyinnervated structure. FMRFamide fibers also reached theinterpeduncular and the oculomotor nuclei. A dense net-work of labeled fibers was observed in the dorsolateral andmedioventral area around the isthmic nucleus. The cerebel-lum and other hindbrain structures contained few labeledelements.
Eleutherodactylus coqui [Figure 11, and Table 1]. Likeother anurans, in E. coqui IR perikarya were located in thecaudalmost medial septum, the dorsal and dorsolateralanterior preoptic area, the suprachiasmatic nucleus, andthe infundibular hypothalamus (Table 1). Labeled neuronsin the medial septum and anterior preoptic area weresparse, totalling 107 in one brain. These neurons weremostly bipolar and were situated outside the periventricu-lar cell layers. As compared to the seven other anuranspecies studied, in this species the suprachiasmaticnucleus-infundibular hypothalamus contained the small-
Fig. 5. Transverse sections of a P. pipa brain. A: Prominent IRfibers in glomerular and molecular layers of the olfactory bulb.B: Concentration of IR fibers in and around the ependyma (arrows) oflateral telencephalon. C: Dense plexus of IR fibers in the areacorresponding to the nucleus of Bellonci (large arrow) and sparsefibers in the periependymal area beneath the left habenular nucleus
(HB). Corresponds to level F in Figure 1. The third ventricle (III) is onthe right. D: Dense accumulation of FMRFamide-IR material in themedian eminence. E: FMRFamide innervation around the subcommis-sural organ (SCO) in the dorsocaudal diencephalon. Corresponds tolevel H in Figure 1. Scale bars 5 50 µm in A,B; 100 µm in C–E.
284 C. PINELLI ET AL.

Fig. 6. Transverse sections of the anuran optic tectum. A: P. pipa.Haphazard distribution of thick fibers in unlaminated tectum.B: Distribution of IR fibers in the optic tectum of X. laevis. Ependymais on the bottom. C: H. crucifer. Ir fibers are distributed in the
periventricular cell layers of the tectum; a few fibers are present in thesuperficial neuropil. D: B. bufo. Ir fibers distribution in the optictectum resembles that in H. crucifer. Scale bars 5 100 µm in A,D; 50µm in B,C.
FMRFAMIDE IN AMPHIBIAN BRAIN 285

Fig. 7. Transverse sections of a X. laevisbrain. A: Ir perikarya in the periventricularcell column of the anterior preoptic nucleuswith long and ramified fibers in the surround-ing neuropil. B: Ir neurons in the suprachias-matic nucleus, ventrolateral to the third ven-tricle (III), showing long process innervatingthe ependyma. Scale bar, same as panel A.C: A multipolar FMRFamide neuron (arrow)in the lateral suprachiasmatic area. Scalebar 5 100 µm (applies to all).
286 C. PINELLI ET AL.

Fig. 8. Transverse sections of an H. crucifer brain. A: A fusiformbipolar FMRFamide-IR neuron in the medial septum. B: Unipolar IRneurons in caudalmost medial telencephalon with long fibers extend-ing toward the ventromedial anterior preoptic area. C: Ir neurons aredistributed as a continuum between the caudalmost medial septum(arrow) and the rostralmost anterior preoptic area (bottom). Asterisk
depicts preoptic recess. Corresponding to level D in Figure 1. D: Bipolarfusiform FMRFamide-IR neurons located dorsal to the caudal preopticrecess (asterisk). Note punctate immunoreactivity in vicinity of theependymal cell layer. Scale bars 550 µm in A; 100 µm in B,D; 200 µmin C.
est aggregate of FMRFamide-IR neurons, totalling 198 inone brain. All labeled neurons of this cluster were locatedwithin the periventricular cell column; immunoreactivecells were never observed among ependymal cells. Withinthe postoptic hypothalamus, IR neurons were restricted tothe ventral hypothalamus. In contrast to other anuransthe olfactory bulb, dorsal and medial pallium, entopedun-cular nucleus, optic chiasma, and optic tract were devoid ofIR fibers. The optic tectum, showing a highly laminatedperiventricular gray, was less innervated. The fibers pre-sent were distributed in all strips of white matter betweenlayers of unlabeled neurons. IR fibers in the medialseptum, anterior preoptic neuropil, neuropil of Bellonci,lateral thalamic neuropil, ventral tegmentum, and cerebel-lum were comparatively less frequent than in other an-urans.
Ceratophrys ornata [Figure 12, and Table 1]. Similarto R. esculenta, the brain of C. ornata showed IR perikaryain the medial septum, anterior preoptic area, suprachias-matic nucleus, and dorsal as well as ventral infundibular(postoptic) hypothalamus. The neurons which formed theseptopreoptic continuum were uni- or bipolar. They werefew in number, 48 and 55 in the two brains examined. Inthe anterior preoptic area, all FMRFamide neurons werelocated in the immediate vicinity of the anterior preopticnucleus, dorsally and laterally. Those in the suprachias-matic-infundibular groups were far more numerous (312and 324) and all were located within the periventricular
cell column. In addition, solitary multipolar neurons wereobserved in the area of the nucleus of Bellonci as well inthe ventral tegmentum in the area overlying the nuclei ofthe oculomotor nerve (Fig. 12). IR fibers were observed inthe glomerular and mitral cell layers of the olfactory bulband in the medial septum. They projected to the sameareas as described for R. esculenta, and the highest densityof IR fibers was observed in the lateral anterior preopticneuropil. Furthermore, they innervated additional sites:lateral septum, lateral amygdala, medial forebrain bundle,posterior commissure, and interpeduncular nucleus(Table 1).
Urodela.
Triturus carnifex [Figures 13, 14, and 15, and Table 2].The brain of the crested newt showed a widespreadFMRFamide-IR system with projections innervating prac-tically all brain regions, but the somata were restricted tothe diencephalon and were located primarily within theperiventricular cell column. A few (5 and 8 in two brains)solitary IR neurons were scattered in the dorsomedial partof the anterior preoptic nucleus, just below the anteriorcommissure (Fig. 13D). All other neurons (total of 374 and361 FMRFamide neurons in the two brains analyzed)formed a continuum. This rostrocaudal band of IR neu-rons, one on either side of the lower part of the thirdventricle, extended caudally between the posterior portionof the preoptic nucleus and the ventral infundibularhypothalamus (Figs. 13E–H, 14). Most of these neurons
Fig. 9. Transverse sections of an H. crucifer brain. A: Dense plexus of IR fibers in the lateralmostsuprachiasmatic neuropil and a few labeled fibers in the optic tract (TO). Corresponding to level G inFigure 1. B: Prominent IR fibers in the granular layer of the cerebellum. Scale bar 5100 µm.
288 C. PINELLI ET AL.

Fig. 10. Transverse sections of a B. bufo brain. A: Labeled peri-karya in the subependymal cell column below the preoptic recess(asterisk). Dorsal to the left. Scale bar 5 100 µm. B: Labeled perikaryain and near the ependymal lining of the magnocellular preopticnucleus. Third ventricle is to the right. C: FMRFamide neurons in the
periependymal cell layers of the suprachiasmatic nucleus. Some of thefibers contact the third ventricle (III). Scale bar, same as panel B.D: Labeled neurons distributed in the periventricular cell column ofmidinfundibular recess (asterisk). Scale bars 5 100 µm in A, B (bar inB applies to C,D).
FMRFAMIDE IN AMPHIBIAN BRAIN 289

appeared unipolar; bipolar neurons were observed onlyoccasionally (Fig. 15A). They had thick axons which pen-etrated into the surrounding neuropil and exhibited long
beaded ramifications, some of which were very thick (Fig.15B). Although fewer than the unipolars, the bipolarneurons were comparatively more conspicuous, and eachshowed a short projection ending in contact with theventricular lumen (Fig. 15A). Thin IR fibers were presentin the rostral telencephalon, medial pallium, basal pal-lium, and striatum. Most of the IR fibers were located inthe neuropil and only some fibers invaded the periventric-ular cell column. Ir fibers were observed in the medialseptum, lateral septum, and particularly dense, in theventromedial area of the caudal portion of the telencepha-lon (Fig. 13B,C). FMRFamide fibers also innervated thelateral and medial amygdala, and many of them wereobserved in the anterior commissure (Fig. 13D). At thesame rostrocaudal level, the anterior preoptic area wasrichly supplied with FMRFamide fibers which continuedcaudally in the ventrolateral neuropil through the entirelength of the third ventricle and reached the suprachias-matic and the ventral infundibular (postoptic) hypothala-mus (Fig. 13E–H). The lateral forebrain bundle containedIR fibers, but the medial forebrain bundle was totallydevoid of them. In the diencephalon, at the base of thehabenulae, fiber aggregates were observed in the retroflexbundle, and, somewhat ventral and lateral in the neuropilof Bellonci, we observed a high density of thick and beadedFMRFamide fibers (Figs. 13E,F, 15C). At the level of thehabenulae, many IR fibers were noted running parallel tothe ependymal lining of the third ventricle. The network ofIR fibers in the neuropil of Bellonci formed a continuumwith IR fibers positioned ventrolaterally in the diencepha-lon; fiber projections were observed in the dorsal thala-mus, ventral thalamus, and lateral forebrain bundle (Fig.13E). Further caudal, the dorsoventrally oriented tractsoccupied a position just lateral to the periventricular gray,whereas the lateralmost thalamic neuropil was devoid ofsuch fibers (Fig. 13F). In this area, many fibers reached theventral hypothalamus, and several of them crossed to thecontralateral side just above the postoptic commissure.
Fig. 11. Camera lucida drawing of horizontal sections, at twolevels, with the left drawing ventral to the right drawing, of an E.coqui brain giving the distribution of FMRFamide-IR perikarya (blackdots) and fibers (dashes and fine dots). The rostral group of neurons islocated in the neuropil of the anterior preoptic area (POA). The caudalgroup of neurons is restricted to the periventricular cell column of theinfundibular recess (IR).
Fig. 12. A solitary neuron in the ventral midbrain tegmentum (onthe right side) in a transverse section of a C. ornata brain. Scale bar 5100 µm.
290 C. PINELLI ET AL.

The pretectal white was crossed by some IR fibers whichdid not innervate areas around the subcommissural organ(Fig. 13G). In the midbrain, the optic tectum was weaklyinnervated, and only occasionally were IR fibers observedin the unlaminated periventricular gray (Fig. 13H). Thedorsal tegmentum and the tegmentum isthmi were ratherheavily innervated, except for the peripheral neuropil.A major amount of fibers reached areas of the oculo-motor and interpeduncular nuclei. More caudally, thecerebellar neuropil contained a few IR fibers, and in therostral rhombencephalon, a ventrally running bundle ofFMRFamide fibers emerged lateral to the raphe nuclei andinnervated the ventrolateral area of the spinal lemniscus(Fig. 13I,J). The lateral acoustic area, the medial longitudi-nal bundle, and the central gray were moderately inner-vated by such fibers.
Triturus vulgaris [Figures 13 and 16A, and Table 2].Like in T. carnifex, in this species all FMRFamide-IR cellbodies were located within the diencephalon. They totaled
298 in the one brain analyzed. FMRFamide neurons werepositioned in the hypothalamus where they formed arostrocaudal continuum, one on each side of the thirdventricle. This net of neurons spanned through the poste-rior part of the preoptic nucleus and caudally along thesuprachiasmatic area to the ventral postoptic infundibularhypothalamus (Fig. 13E–H). All FMRFamide perikaryawere distributed within the multilayered periventricularcell lining (Fig. 16A). Each IR cell body was either veryclose to or in direct contact with the ventricle. Axons ofthese cells were relatively thick and projected laterallyinto the adjacent neuropil. Once in the neuropil, eachaxonal filament showed moderate to intense ramifications.In the suprachiasmatic area immunostained neurons weredistributed in the central part of the pars posterior of thepreoptic nucleus. Further caudal, similar neurons wereobserved in the dorsal portion of the ventral infundibularhypothalamus (ventral infundibular nucleus) (Fig. 13F–H).
FMRFamide fibers projected in all directions innervat-ing practically all brain regions. A few terminals wereobserved in the mitral cell layer of the olfactory bulb. Justbehind the olfactory bulbs, within anterior and mid-telencephalon, FMRFamide fibers innervated all regionsof the pallium. They were quite abundant in the area of themedial septum, accumbens nucleus, and amygdala (Fig.13B,C). Mostly, these fibers were located in the neuropiland many of them invaded the lateral and medial fore-brain bundles. Stained fibers continued caudally in thetransition area between telencephalon and diencephalonand were abundant in the neuropil of the anterior preopticarea (Fig. 13D). Further caudal, in the thalamus and in theunderlying suprachiasmatic area, the entire neuropil wasrichly endowed with FMRFamide fibers and terminals.These accumulated dorsolaterally and projected throughthe stria medullaris to reach the habenulae (Fig. 13E). Adense fiber network was observed in the lateralmostventral neuropil of the postoptic infundibular hypothala-mus (Fig. 13F). These fibers continued caudally into themedian eminence. In the mesencephalon, stained fiberswere distributed through the entire dorso-ventral exten-sion of the neuropil and innervated areas such as the optictectum, midbrain tegmentum, isthmic tegmentum, andinterpeduncular nucleus (Fig. 13H,I). The neuropil of thecerebellum was richly supplied with FMRFamide fibersand a dense network of fibers was present in the ventrolat-eral neuropil of the anterior rhombencephalon, particu-larly in the area of the trigeminal tegmentum. These fiberbundles continued further caudal into the spinal lemnis-cus (Fig. 13J).
Ambystoma tigrinum [Figures 13 and 16B, and Table 2].In contrast to crested and smooth newts, the brain of thetiger salamander had a relatively large aggregate ofFMRFamide neurons in the anterior preoptic nucleus, oneon each side of the preoptic recess (Fig. 13D). There were89 IR perikarya in this nucleus. No such neurons werepresent in the posterior (suprachiasmatic) portion of thepreoptic nucleus (Fig. 13E). The caudal aggregate ofFMRFamide neurons was located in the postchiasmatichypothalamus containing 164 neurons in the one brainanalyzed (Fig. 13F–H). The fiber network was quite simi-lar to that of the two species of Triturus (Fig. 13A2J; Table2). In the neuropil of Bellonci, we observed a matrix ofparticularly prominent beaded fibers (Fig. 16B). Someminor differences included the presence of fewer fibers inthe pallium and their complete absence in the gray matter
TABLE 2. Summary Distribution of FMRFamide-like Immunoreactivityin the Brain of T. carnifex, T. vulgaris,and Ambystoma tigrinum (Urodela)1
Brain areaT.
carnifexT.
vulgarisA.
tigrinum
ForebrainOlfactory bulb – 1 – 1 – 1Accessory olfactory bulb – – – – – –
TelencephalonDorsal pallium – 1 – 1 – 1Medial pallium – 1 – 1 – 1Lateral pallium – 1 – 1 – –Striatum (Basal pallium) – 1 – 1 – 1Medial septum – 1 – 1 – 1Lateral septum – 1 – 1 – 1Mediobasal telencephalon – 11 – 1 – 11Anterior commissure – 1 – – – 1Amygdala
Lateral – 11 – 1 – 1Medial – 1 – 1 – 1
Lateral forbrain bundle – 1 – 1 – 1Medial forebrain bundle – – – 1 – 1
DiencephalonEpiphysis – – – – – –Habenulae – 1 – 1 – 1Habenular commissure – 1 – – – –Thalamic eminence – 1 – 1 – 1Dorsal thalamus – 1 – 1 – 1Ventral thalamus – 1 – 1 – 1Neuropil of Bellonci – 111 – 111 – 11Lateral thalamic neuropil – 11 – 11 – 11Lateral forebrain bundle – 1 – 1 – 1Medial forebrain bundle – – – 1 – –Lateral preoptic area – 11 – 11 – 111Preoptic nucleus, anterior o 1 – – ooo 11Preoptic nucleus, posterior oo 1 ooo 1 – 11Postchiasmatic hypothalamus oo 1 ooo 1 oo 1Infundibular hypothalamus oo 11 oo 11 o 11Optic chiasma – 11 – 1 – 1Optic tract – 1 – 1 – 1Median eminence – 1 – 1 – 1
Posterior tubercle – 1 – 1 – 1Pretectum – 1 – 1 – 1Midbrain
Optic tectum – 1 – 1 – 1Posterior tectal nucleus – – – 1 – 1Tegmentum
Dorsal – 11 – 1 – 1Isthmi – 111 – 11 – 11
Lateral forebrain bundle – 1 – 1 – 11Medial longitudinal bundle – 1 – – – –Interpeduncular nucleus – 1 – 1 – –
HindbrainCerebellum – 1 – 11 – 1Raphe nuclei – 1 – – – –Solitary bundle – 1 – – – –Acousticolateral area – 1 – – – –Spinal lemnisco – 11 – 11 – 1
1Density of immunostained perikarya is indicated by o’s, fibers by 1’s. Dash denotesabsence.
FMRFAMIDE IN AMPHIBIAN BRAIN 291

Fig. 13. Rostrocaudal sequence of camera lucida drawings of T.carnifex, T. vulgaris, and A. tigrinum brains (transverse sections)showing the distribution of FMRFamide-IR perikarya (large dots) andfibers (dashes and fine dots). Section levels A–J are indicated in a
lateral view of T. carnifex brain in Figure 14; those of T. vulgaris and A.tigrinum are at similar levels. Other details: as in Figure 1. Forabbreviations, see list.

of the dorsal and isthmic tegmentum. A relatively smallnumber of fibers innervated the ventrolateral area of therhombencephalon (Fig. 13J).
Gymnophiona.
Typhlonectes compressicauda [Figures 17, 18, and 19,and Table 3]. The telencephalon lacked FMRFamide-IRcell bodies. All labeled perikarya were located in thediencephalon and formed two distinct groups, one in thedorsal half and another in the ventral half of the diencepha-lon (Figs. 17D,E, 18, 19A,B). FMRFamide neurons of thefirst cluster were primarily located along the border be-tween the multilayered thalamic periventricular cell col-umn and the surrounding neuropil. Occasional IR peri-karya were positioned in the nearby neuropil or amidstunlabeled cells, but never adjacent to the ventricle (Fig.19A,C). Most of the neurons appeared unipolar. Fibers ofthese neurons were observed along the boundary of theperiventricular cell column; many of them projected to the
lateral thalamic wall and continued ventrally. The rostro-caudal extent of this group of neurons, one on each side ofthe third ventricle, was approximately 280 µm. Bilaterally,up to 31 labeled neurons were counted in a 20 µmtransverse section, and up to 168 neurons formed thisthalamic group. Vertically, these neurons were positionedalong the entire extend of the dorsal thalamus. They werenoted as far ventral as the dorsalmost portion of theventral thalamus. A few, far dorsally positioned, neuronswere in contact with the dorsolateral surface of the subcom-missural organ. The other set of FMRFamide neurons,located in the ventral diencephalon, extended rostrocau-dally, and the labeled cell bodies were located within theperiventricular cell column, primarily in subependymalpositions (Fig. 19B). In each section, one or two labeledneurons were observed in lateral locations. Ir somata, indirect contact with the third ventricle were rare. Thisgroup of neurons extended rostrocaudally from just behind
Figure 13 (Continued)
FMRFAMIDE IN AMPHIBIAN BRAIN 293

the preoptic recess to the midventral infundibular hypo-thalamus, up to the caudalmost level of the posteriortubercle (Figs. 17D,E, 18). At the rostrocaudal level atwhich they were most frequently observed up to 65 labeledperikarya were counted in a 20 µm transverse section and,altogether, 365 immunostained perikarya were present inthis group. These neurons formed two separate aggre-gates, one on each side of the third ventricle (see Fig. 19B).
A few thin scattered fibers were observed in the dorsalpallium; in the medial septum they were more frequent(Fig. 17A). The labeled fibers noted in the medial septumcontinued ventrally and ventrolaterally in the neuropilunderlying the accumbens nucleus, the medial and antero-lateral part of amygdaloid complex, and the lower part ofthe striatum. Further caudal, at the level of the thalamiceminence, some IR fibers were present in the adjacentneuropil, and, occasionally, some fine caliber fiberswere observed crossing the anterior and pallial commis-sures (Fig. 17B). At this telencephalic level, some IR fiberswere observed in the dorsal pallium, but more caudally(the cerebral hemispheres extent far caudally and, in alateral profile, cover the entire diencephalon and most ofthe midbrain; Fig. 18) all pallial zones were withoutFMRFamide immunoreactivity (Fig. 17B–F). The anteriorpreoptic nucleus was totally devoid of IR perikarya andfibers, whereas many IR fibers were present in the lateralpreoptic neuropil, extending as far lateral as the neuropilunderlying the medial amygdala and the lateral forebrainbundle (Fig. 17B). The medial part of the amygdaloidcomplex did not display FMRFamide immunoreactivity.The rostral subhabenular neuropil (in contrast to allother amphibians investigated, in this species the habe-nulae appear symmetrical) was well innervated byFMRFamide-IR fibers which continued caudally into theretroflex bundle (Fig. 17C). Only occasionally, IR fiberswere seen in the habenular commissure. In the mid-diencephalon, long and thick FMRFamide-IR fibers werelocated laterally in the neuropil and there was a densenetwork of thick fibers in the ventrolateral corner of themid-diencephalon continuing caudally into the ventral
hypothalamus (Fig. 17D,E). Thick labeled fibers wereobserved along the lateral thalamic walls. They appearedto stem from the thalamic FMRFamide neurons, whereasfibers from the ventral diencephalic group of FMRFamideneurons contributed rostrally to the posterior preopticarea fiber network and caudally to ventrolaterally locatedhypothalamic net. Dorsally, in the posterior diencephalon,some labeled fibers crossed the posterior commissure and,at this level, a massive plexus of IR fibers was observed inthe neuropil, far lateral to the posterior tubercle (Fig.17E). In the mesencephalon, a few long and thin fibersinnervated the optic tectum (neuropil as well as periepen-dymal cell layers). Ir fibers were also observed along thelateral wall of the rostral midbrain tegmentum (Fig. 17F).These axons continued ventrally to merge with a distinctassembly of IR fibers in the ventrolateral position, whichwas also reached by FMRFamide fibers fanning out of theretroflex bundle. The interpeduncular nucleus area wasonly lightly innervated. At the level of mid-mesencephalonthe rostral rhombencephalon was raised upward, formingthe so-called rhombomesencephalic flexure (see Fig. 18),where the rhombencephalic-auricular roof comes to liejuxtapposed to the midbrain floor (Fig. 17F). At this level,the dorsolateral area showed the densest plexus of IRfibers; another dense plexus of IR fibers was present in aventrolateral position, and there were a few labeled fibersbetween the two areas (Fig. 17F). Caudal to this level,the midbrain tectum and tegmentum contained IR fibersin the neuropil as well as amidst unlabeled periventricu-lar cells (Fig. 17G); ventrally, the interpeduncular nu-cleus and the surrounding neuropil received a moderateFMRFamide innervation. The dorsolateral cerebellar ex-tensions were richly supplied with IR fibers (Fig. 17G). Inthe rhombencephalon, rostrocaudally running IR fibersinnervated the gray matter at the level of sulcus limitansof His and around the raphe nuclei (Fig. 17G). No IR fiberswere observed in the vicinity of the lateral acoustic area.Further caudal, in the medulla, IR fibers running aroundthe raphe fanned out into the neuropil as small bundlesoriented ventrally and ventrolaterally (Figs. 17H, 19D).
Fig. 14. T. carnifex brain. Schematic lateral profile, based on camera lucida tracing of serial sagittalsections, depicting the distribution of FMRFamide-IR neurons (black dots) and their presumptiveterminal fields (arrows). Alphabet on top line indicates levels of transverse sections shown in Figure 13.
294 C. PINELLI ET AL.

The overlying cerebellum was richly innervated (Figs.17H, 19E). Unlabeled cell layers around the solitary tractand the central canal were moderately innervated (Fig.17H,I). Longitudinally running thin IR fibers appeared toconstitute two pairs of bundles in the neuropil, one justlateral to the central canal and the other one in theventrolateral area encompassing part of the medial longi-tudinal bundle (Fig. 17I).
Ichthyophis beddomei [Figures 17, 18, 20, and 21, andTable 3]. The distribution of FMRFamide-IR fibers in thebrain of I. beddomei resembled that of T. compressicauda,but there were major differences in the distribution of IR
perikarya. In contrast to their distribution in T. compressi-cauda, all IR cell bodies in the Ichthyophis brain werelocated within the periventricular cell column and themajority of the perikarya (293 FMRFamide neurons in theone brain analyzed) was positioned in the anterior preopticnucleus. Up to 76 labeled neurons were observed in theperiventricular cell column in the caudal dorsal and ven-tral hypothalamus (Figs. 17B,C,E, 18). Neurons of bothgroups were located in a subependymal position. Thoselocated in the anterior preoptic nucleus, on each side of thepreoptic recess, were slightly elongated laterally, whereaslabeled neurons in the infundibular hypothalamus weremostly spherical (Fig. 20A,B). As compared to T. compressi-cauda, some minor differences in the distribution profile ofthe FMRFamide fiber network existed: a slightly greaterdensity of IR fibers was noted in the lateral preopticneuropil and in the ventrolateral caudal hypothalamus(Fig. 17B,C,E). Labeled fibers were frequent in the lateralthalamic neuropil. The unlabeled periventricular cell col-umn was devoid of IR fibers (Fig. 17D,E). Furthermore,unlike any other brain region, in the caudal part of themedial septum, in the area of transition from septum tothe medial basal forebrain and just rostral to the pallialand anterior commissures, fine caliber IR fibers wereobserved around unlabeled neurons in the outer layers ofthe periventricular cell column (Fig. 21A). The habenularnuclei were innervated, and fine punctate FMRFamideimmunoreactivity was always observed at the base of thehabenulae (Fig. 21B). At the level of the caudal midbrain,the lateral cerebellar extensions received dense FMR-Famide innervation, both in the neuropil as well as amidstunlabeled cells (Fig. 21C). In the rostral rhombencephalon,caudally running FMRFamide-IR fibers were restricted tomediolateral and ventrolateral positions (Fig. 17G); fur-ther caudal, however, many additional fibers were ob-served in progressively more dorsal and dorsolateral posi-tions (Fig. 21D).
DISCUSSION
Except for an incomplete description on the brain of R.esculenta (Vallarino et al., 1993; D’Aniello et al., 1996) anda fragmentary report on the forebrain of H. regilla, Tari-cha granulosa, and R. pipiens (Muske and Moore, 1988;Wirsig-Wiechmann and Basinger, 1988), this is the firststudy providing a detailed analysis of FMRFamide-likeimmunoreactivity in the brain of several species of an-urans as well as in that of some urodeles and gymnophi-ones. Re-examination of the R. esculenta brain confirmedthe results of Vallarino et al. (1993); however, we neverobserved FMRFamide perikarya in the olfactory bulbs asdescribed by Wirsig-Wiechmann and Basinger (1988) in R.pipiens. In spite of the fact that only eight species ofanurans and only three species of urodeles and two speciesof caecilians were studied, our results reveal a number ofinterspecific similarities, as well as differences, within thesame order and among the three orders. On the basis of thedata available a neuroanatomical profile of FMRFamide-IRneuronal system that is shared by all amphibians, is notyet obvious (see Table 4).
Comparative analysis among anurans
Labeled perikarya were present in the medial septum ofthe eight anuran species examined, but in only four species(R. esculenta, P. dacnicolor, H. crucifer, and B. bufo)
Fig. 15. Transverse sections of a T. carnifex brain. A: FMRFamideneurons in the anterior preoptic nucleus. The lateral neuropil isstrongly innervated by IR fibers. B: Higher magnification of axonalprojection of a neuron located in the preoptic nucleus. Note thicknessand beaded appearance (cf. panel C). C: Dense plexus of IR fibers inthe neuropil of Bellonci. Scale bars 5100 µm in A; 50 µm in B (appliesto C).
FMRFAMIDE IN AMPHIBIAN BRAIN 295

FMRFamide neurons were found in the diagonal band ofBroca (Table 1; Fig. 1). FMRFamide-IR perikarya wereobserved in the dorsal and dorsolateral neuropil of ante-rior preoptic area in seven of our eight species. Only in X.laevis labeled cell bodies were located between unlabeledneurons of the dorsolateral periventricular gray. Further-more, only in B. bufo FMRFamide perikarya were ob-served in the ventral periventricular cell column, beneaththe preoptic recess. The suprachiasmatic nucleus was theonly location where labeled perikarya were observed in allof the eight anuran species (Table 1); also in the postchias-matic (infundibular) hypothalamus, which is a site ofFMRFamide-IR perikarya in some representatives of eachvertebrate group, labeled neurons were totally absent intwo (P. pipa and X. laevis) of eight anuran species exam-ined. It is noteworthy that both of these genera belong tothe same family, the Pipidae. In spite of the fact that insome species labeled solitary neurons were observed in‘‘unusual’’ locations, like the optic chiasma (in P. dacnicolorand P. pipa) and midbrain lateral tegmentum (in P. pipa),it is evident that there are two major subgroups ofFMRFamide perikarya. One extends from the septum tothe preoptic region, the other one is located between thesuprachiasmatic nucleus and the infundibular hypothala-mus. We never observed FMRFamide-IR neurons in thehindbrain.
Irrespective of sex and reproductive status, in R. escu-lenta as well as in B. bufo the number of IR neurons is verysimilar in the specimens of either species. P. pipa hasabout 1,000 FMRFamide neurons, but the P. dacnicolorbrain contains just over 200 neurons. This variation in thenumber of IR neurons could correlate with the expansionof the IR fiber network. However, this does not seem to bethe case. In fact, in the brain of P. pipa and P. dacnicolor,FMRFamide innervation was noted in all major brainareas; a species difference exists with regard to the relativedensity of IR fibers in a given brain area (see Table 1; Fig.1). In relation to the number of neurons in the twoneuronal subpopulations, in seven of eight species thesuprachiasmatic-infundibular subpopulation was numeri-cally higher than the septopreoptic subpopulation, whereasonly in P. pipa did the latter subgroup exceed by far thenumber in the former.
The densest accumulation of IR fibers was observed inthe neuropil of Bellonci in all species. However, there wereremarkable interspecific differences in the density of IRfibers in several areas of the brain. For example, theolfactory bulbs were heavily innervated in P. pipa and H.crucifer, but much less in P. dacnicolor (Fig. 1A). Incontrast to other species, in the optic tectum of P. pipa, theunlabeled neurons did not reveal a laminar organizationand were distributed homogeneously throughout the in-
Fig. 16. A: FMRFamide neurons in the multilayered periventricular cell column of the posteriorpreoptic nucleus overlying the optic chiasma in transverse section of T. vulgaris. A few neurons contact thethird ventricle (III). One neuron sends a fiber to the ventricle (arrow). Corresponding to level E in Figure13. B: Network of IR fibers in the neuropil of Bellonci in A. tigrinum. Scale bars 5 100 µm in A; 50 µm in B.
296 C. PINELLI ET AL.
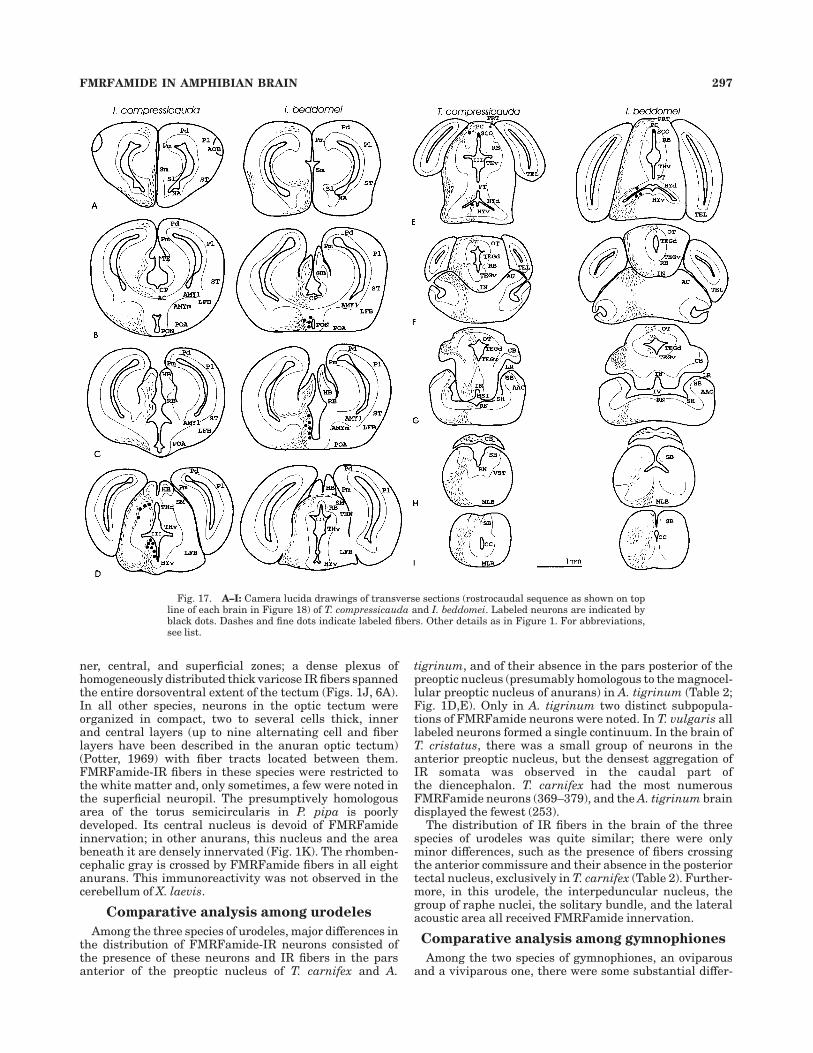
ner, central, and superficial zones; a dense plexus ofhomogeneously distributed thick varicose IR fibers spannedthe entire dorsoventral extent of the tectum (Figs. 1J, 6A).In all other species, neurons in the optic tectum wereorganized in compact, two to several cells thick, innerand central layers (up to nine alternating cell and fiberlayers have been described in the anuran optic tectum)(Potter, 1969) with fiber tracts located between them.FMRFamide-IR fibers in these species were restricted tothe white matter and, only sometimes, a few were noted inthe superficial neuropil. The presumptively homologousarea of the torus semicircularis in P. pipa is poorlydeveloped. Its central nucleus is devoid of FMRFamideinnervation; in other anurans, this nucleus and the areabeneath it are densely innervated (Fig. 1K). The rhomben-cephalic gray is crossed by FMRFamide fibers in all eightanurans. This immunoreactivity was not observed in thecerebellum of X. laevis.
Comparative analysis among urodeles
Among the three species of urodeles, major differences inthe distribution of FMRFamide-IR neurons consisted ofthe presence of these neurons and IR fibers in the parsanterior of the preoptic nucleus of T. carnifex and A.
tigrinum, and of their absence in the pars posterior of thepreoptic nucleus (presumably homologous to the magnocel-lular preoptic nucleus of anurans) in A. tigrinum (Table 2;Fig. 1D,E). Only in A. tigrinum two distinct subpopula-tions of FMRFamide neurons were noted. In T. vulgaris alllabeled neurons formed a single continuum. In the brain ofT. cristatus, there was a small group of neurons in theanterior preoptic nucleus, but the densest aggregation ofIR somata was observed in the caudal part ofthe diencephalon. T. carnifex had the most numerousFMRFamide neurons (369–379), and the A. tigrinum braindisplayed the fewest (253).
The distribution of IR fibers in the brain of the threespecies of urodeles was quite similar; there were onlyminor differences, such as the presence of fibers crossingthe anterior commissure and their absence in the posteriortectal nucleus, exclusively in T. carnifex (Table 2). Further-more, in this urodele, the interpeduncular nucleus, thegroup of raphe nuclei, the solitary bundle, and the lateralacoustic area all received FMRFamide innervation.
Comparative analysis among gymnophiones
Among the two species of gymnophiones, an oviparousand a viviparous one, there were some substantial differ-
Fig. 17. A–I: Camera lucida drawings of transverse sections (rostrocaudal sequence as shown on topline of each brain in Figure 18) of T. compressicauda and I. beddomei. Labeled neurons are indicated byblack dots. Dashes and fine dots indicate labeled fibers. Other details as in Figure 1. For abbreviations,see list.
FMRFAMIDE IN AMPHIBIAN BRAIN 297

ences in the distribution of FMRFamide neurons. In theoviparous I. beddomei the rostral subpopulation of labeledneurons was located in the preoptic nucleus and the caudalsubgroup in the infundibular hypothalamus. In the vivipa-rous T. compressicauda, besides IR cells in the hypothala-mus, IR neurons were observed in the dorsal thalamus(Figs. 17, 18; Table 3). These neurons were primarilylocated outside the periventricular cell column. By con-trast, the anterior preoptic area and hypothalamic neu-rons were distributed amidst the unlabeled neurons of theperiventricular cell column. The number of labeled neu-rons in the hypothalamus of T. compressicauda was muchhigher (365) than in I. beddomei (only 76). In view of thefact that the gross size of the brains in these species isrelatively similar, this difference is indeed dramatic. Alsothe number of FMRFamide neurons in the entire brain
was higher in T. compressicauda. Except for small differ-ences in the relative density of IR fibers in a given brainarea, their distribution pattern was similar in the telen-cephalon of the two species. In the diencephalon, theanterior preoptic nucleus was totally devoid of immunore-activity in T. compressicauda and, only in this species, thegroup of raphe nuclei was moderately innervated (Fig. 1;Table 3).
Comparison between Anura, Urodela,and Gymnophiona
Among amphibians, the presence of FMRFamide-IRneuronal cell bodies in the telencephalon (medial septumand diagonal band of Broca) was a feature unique to theanurans (Table 4). Another observation concerned thepresence of labeled neurons in the dorsal thalamus of T.
Fig. 18. Camera lucida diagram of a quasi-midline sagittal section of T. compressicauda and I.beddomei brains. The lateral profile of each brain is indicated by dashed lines. Based on serial sagittalsections, the overall distribution of FMRFamide neurons is given by black dots and their presumptiveprojection fields are marked by arrows. Other details as in Figure 2. For abbreviations, see list.
298 C. PINELLI ET AL.

Fig. 19. Transverse sections of a T. compressicauda brain.A: Labeled neurons distributed at the periphery of periventricular cellcolumn at the level of dorsal thalamus. Corresponding level D inFigure 17. B: Labeled neurons in periependymal cell layers of thehypothalamus. Same level as panel A. C: Labeled neurons in the
dorsal thalamic periventricular cell column. D: Labeled fiber bundlesrunning ventrally in the mediolateral vestibulospinal tract in caudalrhombencephalon. Corresponding to level H in Figure 17. E: Punctateimmunoreactivity in the cerebellum. Scale bars 5100 µm in A (appliesto B); 50 µm in C (applies to D,E).
FMRFAMIDE IN AMPHIBIAN BRAIN 299

compressicauda; it was not noted in the other speciesinvestigated. If it were not for P. pipa, in which occasionalFMRFamide-IR neurons were observed as far caudal asthe lateral midbrain tegmentum, the absence of IR neu-rons in the mid- and hindbrain could be considered anubiquitous feature among all amphibian orders (Table 4).Thus, it is evident that, with the exception of anurans,FMRFamide perikarya are primarily located in the dien-cephalon.Although Wirsig-Wiechmann and Basinger (1988)described FMRFamide perikarya in the olfactory bulb of R.pipiens, we never observed them in any of our species (seealso Vallarino et al., 1993; D’Aniello et al., 1996). Notwith-standing interspecific differences in the precise location ofFMRFamide perikarya, that is, in the periventricular cellcolumn or outside of it in the surrounding neuropil inanurans and gymnophiones, the entire population of la-beled neurons was subdivided into two major subgroupsbut, in some urodeles, there was just one aggregate oflabeled perikarya on each side of the third ventricle. Thus,the highest density of FMRFamide perikarya was found inthe diencephalon, that is, the posterior preoptic nucleus–suprachiasmatic nucleus–infundibular hypothalamus con-tinuum representing the biggest cluster in most amphib-ians (present study; Muske and Moore, 1988; Wirsig-Wiechmann and Basinger, 1988; Vallarino et al., 1993;D’Aniello et al., 1996). Species of all amphibian orders
possess either cerebrospinal fluid-contacting somata orlabeled, presumably dendritic, fiber projections in contactwith the ventricular lumen. Furthermore, some anuransand urodeles show a high density of immunoreactivityaround the ependymal cell lining, which we consider as aterminal field of FMRFamide fiber projections. In spite ofremarkable interspecific differences in the number ofFMRFamide neurons per brain among and within thethree orders, the present study shows that in all amphib-ians FMRFamide immunoreactivity is widely distributedwithin the brain. An exception is the order Gymnophionain which the olfactory bulbs do not receive FMRFamideinnervation.
Comparison with other vertebrate groups
Some technical problems, such as the use of antibodiesfrom different sources or differences in fixation and immu-nohistochemical techniques, are intrinsic to this type ofanalysis. In spite these limitations, an attempt was madeto compare the distribution of FMRFamide-like immunore-activity in the amphibian brain with the one described inother vertebrates.
Homologies of brain nuclei and areas between differentgroups of vertebrates are a matter of continuous debate,and we have tried to apply ascertained facts (embryology-,hodology-, or biochemistry-based) as well as presumptivehomologies (see Butler and Hoods, 1996). In cyclostomesFMRFamide perikarya were observed in the diencephalon(preoptic area, infundibular hypothalamus, mammillaryregion) and midbrain tegmentum (Jirikowski et al., 1984;Ohtomi et al., 1989; Chiba and Honma, 1992; Wicht andNorthcutt, 1992; Eisthen and Northcutt, 1996). While incartilaginous fishes and in African lungfish these peri-karya were observed exclusively in the forebrain (Chiba etal., 1991; Vallarino et al., 1991; Vallarino et al., 1995), inteleosts they have been described in the fore-, mid-, andhindbrain (Bonn and Konig, 1988, 1989a, 1989b; Batten etal., 1990; Ostholm et al., 1990; Vecino and Ekstrom, 1990,1992; Fujii and Kobayashi, 1992b; Rama Krishna andSubhedar, 1992; Pestarino and Vallarino, 1996; Wrightand Demski, 1996). In reptiles and birds labeled perikaryawere described in the fore- and midbrain (ruin lizard:Vallarino et al., 1994; caiman and turtle: D’Aniello et al.,1999; Japanese quail: Fujii and Kobayashi, 1992a). Inmammals these perikarya were found in the fore-, mid-,and hindbrain of the rat (Triepel and Grimmelikhuijzen,1984), while in the Formosan monkey they were noted onlyin the forebrain (Chen et al., 1989). However, as inamphibians, species differences in the distribution ofFMRFamide perikarya and fibers within each class ofvertebrates have become obvious.
In the forebrain of amphibians labeled perikarya in theolfactory bulbs have been described in R. pipiens (Wirsig-Wiechmann and Basinger, 1988). In other vertebrategroups, IR perikarya have been reported in the olfactorybulb of the teleost Xenotoca eisenii (Bonn and Konig, 1988)and the Japanese quail (Fujii and Kobayashi, 1992a), inthe olfactory tubercle of the rat and monkey (Weber et al.,1981; Chen et al., 1989), and in the olfactory peduncle ofthe ruin lizard (Vallarino et al., 1994). As in anuranamphibians, the presence of FMRFamide perikarya in theseptal area has been observed in Scyliorhinus canicula(Vallarino et al., 1991), Protopterus annectens (Vallarino etal., 1995), Podarcis sicula (Vallarino et al., 1994), Coturnixcoturnix japonica (Fujii and Kobayashi, 1992a), the rat
TABLE 3. Summary Distribution of FMRFamide-like Immunoreactivityin the Brain of T. compressicauda and I. beddomei1
Brain areaT.
compressicaudaI.
beddomei
ForebrainOlfactory bulb – – – –Accessory olfactory bulb – – – –
TelencephalonDorsal pallium – 1 – 1Medial pallium – – – –Lateral pallium – – – –Striatum – 1 – 1Medial septum – 11 – 11Lateral septum – 1 – 1Mediobasal telencephalon – 11 – 111Amygdala – – – –Anterior commissure – 1 – 1
DiencephalonThalamic eminence – 1 – 1Habenulae – 1 – 1Habenular commissure – 1 – 1Retroflex bundle – 1 – 1Dorsal thalamus oo 11 – 1Ventral thalamus – 1 – 1Stria medullaris/dorsolateral thalamic neuropil – 11 – 11Lateral forebrain bundle – 1 – 1Lateral preoptic area – 1 – 11Preoptic nucleus – – ooo 11Dorsal hypothalamus – 1 o 1Ventral hypothalamus ooo 11 o 111Posterior commissure – 1 – 1Median eminence – 1 – 1
Pretectum – 1 – 1Posterior tubercle – – – –Midbrain
Subcommissural organ – – – –Optic tectum – 1 – 1Tegmentum – 1 – 1Interpeduncular nucleus – 1 – 1Retroflex bundle – 1 – 1
HindbrainCerebellum – 111 – 111Auricle – 111 – 111Solitary bundle – 1 – 1Raphe nuclei – 1 – –Medial longitudinal bundle – 11 – 11Reticular formation – 11 – 11Vestibulospinal tract – 11 – 1
1Density of immunostained perikarya is indicated by o’s, fibers by 1’s. Dash denotesabsence.
300 C. PINELLI ET AL.

(Williams and Dockray, 1983; Chronwall et al., 1984), andthe Formosan monkey (Chen et al., 1989). A unique featureof teleosts is the presence of FMRFamide perikarya in theterminal nerve ganglion, termed olfactoretinal nucleus,located in the rostral telencephalon (Boer et al., 1980;Bonn and Konig, 1988, 1989a, 1989b; Ekstrom et al., 1988;Batten et al., 1990; Ostholm et al., 1990; Chiba et al., 1991;Vallarino et al., 1991, 1995; Fujii and Kobayashi, 1992b;Rama Krishna and Subhedar, 1992; Magliulo-Cepriano etal., 1993; Pinelli, 1999). The existence of IR perikarya inthe mid- and caudal telencephalon is also a feature sharedby many vertebrate groups, that is, fish, amphibians,reptiles, and mammals.
The presence of FMRFamide neurons in the diencepha-lon (preoptic, suprachiasmatic, and/or postoptic areas) is acommon feature of all vertebrate groups, from mammals tocyclostomes. FMRFamide perikarya in the thalamus of theviviparous gymnophione is a feature shared with only afew other vertebrates, a cartilaginous fish (Scyliorhinuscanicula, Vallarino et al., 1991), some teleosts (Bonn andKonig, 1989a, 1989b; Rama Krishna and Subhedar, 1992;Vecino and Ekstrom, 1992), a dipnoan (Vallarino et al.,1995), and the rat (Williams and Dockray, 1983). Whereasthe anterior preoptic area and hypothalamus containFMRFamide perikarya in species belonging to all fiveclasses of vertebrates, the presence of similar neurons inthe suprachiasmatic region has only been described infishes (Vallarino et al., 1991; Rama Krishna and Subhedar,1992), amphibians (present study), and reptiles (Vallarinoet al., 1994; D’Aniello et al., 1999).
As mentioned, the presence of FMRFamide-IR peri-karya in the amphibian midbrain was rarely observed:occasionally, neurons were found in the midbrain tegmen-tum of P. pipa. However, in teleosts labeled perikarya weredescribed in the midbrain of about 70% of the speciesinvestigated (Bonn and Konig, 1988, 1989a, 1989b; Battenet al., 1990; Ostholm et al., 1990; Fujii and Kobayashi,1992b; Vecino and Ekstrom, 1992; Vallarino et al., 1995;
Pestarino and Vallarino, 1996). In cyclostomes mesence-phalic FMRFamide neurons were, so far, described exclu-sively in two of five species studied (Chiba and Honma,1992; Eisthen and Northcutt, 1996). In reptiles, only theruin lizard displayed labeled neurons in the same position(Vallarino et al., 1994). In the Japanese quail, Fujii andKobayashi (1992a) did not describe FMRFamide neuronsin the midbrain; in mammals, the existence of suchneurons in the midbrain is uncertain (Triepel and Grimme-likhuijzen, 1984; Chen et al., 1989). Also in the hindbrainFMRFamide perikarya were observed only in some speciesof cyclostomes (Wicht and Northcutt, 1992), teleosts (Bonnand Konig, 1988, 1989a,b; Vecino and Ekstrom, 1992; Fujiiand Kobayashi, 1992b), and rat (Triepel and Grimmelikhui-jzen, 1984). It is obvious that there are differences in thedistribution of FMRFamide neurons in any given brainarea within each class of vertebrates.
Considerations on sites of embryonic origin
Our knowledge on the embryonic origin of FMRF-amide-IR neurons in amphibians is limited to a fewspecies, namely, T. granulosa (Northcutt and Muske, 1994)and B. bufo (Fiorentino et al., 1999): in these species,belonging to two different orders, the origin of the septopre-optic continuum of FMRFamide neurons was confirmed byablation (uni- and bilateral) of the olfactory placode,identifying this structure as the source of these neurons.In R. esculenta and B. bufo this result has been confirmedby ontogenetic analysis of FMRFamide-IR system(D’Aniello et al., 1996; our unpublished data). The placodalorigin of FMRFamide neurons in amphibians is shared byother vertebrates: fish (ontogenesis: Danio rerio; Pinelli,1999), birds (heterologous olfactory placode transplanta-tion in quail and chick embryos; Yamamoto et al., 1996),and reptiles (Chalcides chalcides; Pinelli et al., 1999).These studies reveal an extracranial origin of FMRFamideneurons located in the telencephalon and anterior preopticarea of adult specimens. The contention that the terminal
Fig. 20. Sagittal sections of a I. beddomei brain. A: Spherical FMRFamide neurons in the infundibularhypothalamus (IR, infundibular recess). B: A labeled bipolar neuron in the preoptic nucleus. Scale bars 550 µm.
FMRFAMIDE IN AMPHIBIAN BRAIN 301

Fig. 21. Sagittal sections of an I. beddomei brain. A: PunctateFMRFamide immunoreactivity (presumed terminal field) surround-ing unlabeled neurons in the anterior preoptic nucleus. Third ventricleis on top right of figure. B: FMRFamide immunoreactivity in the lefthabenular nucleus (HB). C: Heavy FMRFamide immunorectivity in
the unlabeled fiber and cell layers of lateral cerebellar extension.Caudal is to the right. D: FMRFamide-IR fiber tract in the dorsolateralrombencephalon. Caudal is to the bottom. Scale bars 5 100 µm in A(applies to B,D); 50 µm in C.
302 C. PINELLI ET AL.

nerve–septopreoptic continuum of FMRFamide neurons inanurans and urodeles has an extracranial origin re-sembles the situation described for the terminal nerve-located or terminal nerve–preoptic region continuum ofGnRH-IR neurons in fish (see Parhar et al., 1998), amphib-ians (see Murakami et al., 1992; Northcutt and Muske,1994; D’Aniello et al., 1994, 1995; Muske, 1997), birds(Akutsu et al., 1992), and mammals (see Quanbeck et al.,1997; Schwanzel-Fukuda, 1997). The source of FMRF-amide neurons located at the level of thalamus, posteriorpreoptic nucleus, and postchiasmatic hypothalamus re-mains to be determined. The proximity of hypothalamicneurons and the ventricular ependyma as well as theirpresence in the ependyma, suggests an ependymal origin.Thus we presume that FMRFamide neuronal systems mayhave multiple sources (see Chen et al., 1990). This re-sembles the presumed multiple embryonic origins of otherneuronal systems, for example, the GnRH-IR system(D’Aniello et al., 1994, 1995; Northcutt and Muske, 1994;Muske, 1997; Parhar et al., 1998; White and Fernald,1998).
Putative functions
Although the extensive distribution of FMRFamide-likeimmunoreactivity suggests that this peptide plays mul-tiple roles, it may have specific functions in different brainregions. For example, the high density of FMRFamideperikarya and fibers in the hypothalamus and perivascu-lar areas of the median eminence hints at hypophysio-tropic roles of FMRFamide exerted in these regions.Colocalization of FMRFamide-like and GnRH immunore-activities within the same neuron as well as FMRF-amide-IR innervation of some GnRH neurons in the R.
esculenta anterior preoptic area (Rastogi et al., 1996)support the assumption that FMRFamide-like peptidesare involved in GnRH-regulated functions that may berelated to reproduction.
CONCLUSIONS
Although it is arduous to make interspecific compari-sons of brain structures on the basis of proven andpresumed homologies between brain areas, it is obviousthat the FMRFamide-IR system is widely distributed in allmajor brain areas of all vertebrates. Homologies betweenbrain structures of anurans, urodeles, and gymnophionesare still far from certain; nevertheless, a comparativeanalysis of the immunohistochemical localization of neuro-peptides may provide a useful tool to identify homologies.Anurans have a well-developed visual system and showdense FMRFamide innervation of the optic tectum. Thetectum of gymnophiones is the least developed and has avery poor FMRFamide innervation. Of the anurans stud-ied, two arboreal species, namely, P. dacnicolor and H.crucifer, display a remarkable difference in the density ofFMRFamide-IR fibers in the optic tectum; H. crucifer isactive during the day and has a heavy FMRFamideinnervation of the tectum, whereas P. dacnicolor is noctur-nal and shows only a moderate FMRFamide innervation.In summary, the present study indicates the presence ofFMRFamide-like immunoreactivity in the brain of allamphibian groups which, judging by its distribution, maybe related to the control or integration of sensory (olfac-tory, visual, auditory, and motor), autonomic, and pituitaryfunctions.
Amphibians and all other vertebrates have FMRFamideneurons in the diencephalon, in particular, in the hypo-thalamus. However, unique features like the presence oflabeled neurons in the dorsal thalamus of a viviparousgymnophione or in the medial septum and in the diagonalband of Broca of anurans make the class Amphibia notablyheterogeneous. Furthermore, within the class of the Am-phibia, the absence of FMRFamide neurons in the telen-cephalon of adult urodeles and gymnophiones is a uniquefeature among all gnathostome vertebrates. Currently, weare unable to provide a plausible explanation for thisdifference between the three amphibian orders.
ACKNOWLEDGMENTS
This work was supported by grants from M.U.R.S.T. andthe University of Naples Federico II (R.K.R.), from theGerman Science Foundation (D.L.M.), and the VigoniProgram. Expert technical assistance of Ms. SabineSchmidt is acknowledged.
LITERATURE CITED
Akutsu S, Takada M, Ohki-Hamazaki H, Murakami S, Arai Y. 1992. Originof luteinizing hormone-releasing hormone neurons in the chick embryo:effect of the olfactory placode ablation. Neurosci Lett 142:241–244.
Andersen AC, Tonon MC, Pelletier G, Conlon JM, Fasolo A, Vaudry H. 1992.Neuropeptides in the amphibian brain. Int Rev Cytol 138:89–210.
Batten TFC, Cambre ML, Moons L, Vandesande F. 1990. Comparativedistribution of neuropeptide-immunoreactive systems in the brain ofthe green molly, Poecilia latipinna. J Comp Neurol 302:893–919.
Boer HH, Schot LPC, Veenstra JA, Reichelt D. 1980. Immunocytochemicalidentification of neural elements in central nervous systems of a snail,some insects, a fish and a mammal with an antiserum to the molluscancardioexcitatory tetrapeptide FMRFamide. Cell Tissue Res 231:21–27.
TABLE 4. Distribution of FMRFamide-ir Neuronal Elements(o, perikarya; 1, fibers; –, absence) in the Amphibian Brain
Brain area Anura Urodela Gymnophiona
ForebrainOlfactory bulb – 1 – 1 – 1
TelencephalonDorsal pallium – 1 – 1 – 1Medial pallium – 1 – 1 – –Lateral pallium – 1 – 1 – 1Striatum – 1 – 1 – 1Medial septum o 1 – 1 – 1Lateral septum – 1 – 1 – 1Mediobasal telencephalon o 1 – 1 – 1Amygdala – 1 – 1 – –Diagonal band of Broca o 1 – 11 – 11
Anterior commissure – 1 – 1 – 1Diencephalon
Habenulae – 1 – – – 1Dorsal thalamus – 1 – 1 o 1Ventral thalamus – 1 – 1 – 1Lateral thalamic neuropil – 1 – 1 – 1Anterior preoptic neuropil o 1 – 1 – 1Anterior preoptic nucleus o 1 o 1 o 1Posterior preoptic nucleus o 1 o 1 o 1Suprachiasmatic nucleus o 1 o 1 ?Dorsal hypothalamus o 1 o 1 o 1Ventral hypothalamus o 1 o 1 o 1Median eminence – 1 – 1 – 1Optic chiasma o 1 – 1 ?Optic tract – 1 – 1 – 1
MidbrainOptic tectum – 1 – 1 – 1Tegmentum o2 1 – 1 – 1Interpeduncular nucleus – 1 – 1 – 1
HindbrainCerebellum – 1 – 1 – 1Raphe nuclei – 1 – 1 – 1Reticular formation – 1 – 1 – 1
1Presumptively homologous (at least neuroanatomically corresponding area) to that inanurans ?, identification uncertain.2Occasional perikarya in P. pipa.
FMRFAMIDE IN AMPHIBIAN BRAIN 303

Boersma CJC, Sonnemans MAF, van Leeuwen FW. 1993. Immunocytochemi-cal localization of neuropeptide FF (FMRFamide-like-peptide) in hypo-thalamus-neurohypophyseal system of Wistar and Brattleboro rats bylight and electron microscopy. J Comp Neurol 336:555–570.
Bonn U, Konig B. 1988. FMRFamide-like immunoreactivity in brain andpituitary of Xenotoca eisenii: (Cyprinidontoformes, Teleostei). J Hirn-forsch 29:121–131.
Bonn U, Konig B. 1989a. FMRFamide-like immunoreactivity in the brainand pituitary of the teleost, Eigenmannia lineata (Gymnotiformes). ZMikrosk-Anat Forsch 103:221–236.
Bonn U, Konig B. 1989b. FMRFamide-like immunoreactivity in the brainand pituitary of Carassius auratus (Cyprinidae, Teleostei). J Hirnforsch30:361–370.
Butler AB, Hoods W. 1996. Comparative vertebrate neuroanatomy: evolu-tion and adaptation. New York: Wiley-Liss.
Chen ST, Tsai MS, Shen CL. 1989. Distribution of FMRFamide-likeimmunoreactivity in the central nervous system of the Formosa mon-key (Macaca cyclopsis). Peptides 10:825–834.
Chen ST, Tsai MS, Shen CL. 1990. Ontogeny of the FMRFamide-immunoreactivity in the rat forebrain and diencephalon. Proc Natl SciCounc Repub China (B) 14:91–97.
Chiba A, Honma Y. 1992. FMRFamide-immunoreactive structures in thebrain of the brown hagfish, Paramyxine atami: relationship withneuropetide Y-immunoreactive structures. Histochemistry 98:33–38.
Chiba A, Oka S, Y. Honma S, 1991. Immunocytochemical distribution ofFMRFamide-like substances in the brain of the cloudy dogfish, Scyliorhi-nus torazame. Cell Tissue Res 265:243–250.
Chronwall BM, Olschowska JA, O’Donohue TL. 1984. Histochemical local-ization of FMRFamide-like immunoreactivity in the rat brain. Peptides5:569–584.
Clairambault P, Anteino F. 1970. Anatomie comparee du diencephale desanoures. J Hirnforsch 12:143–160.
Clairambault P, Timmel JF. 1990. Developmental organization of theamphibian pallium. In: Schwerdtfeger WK, Germroth P, editors. Theforebrain in nonmammals: new aspects in structure and development.Berlin: Springer-Verlag. p 29–41.
Clairambault P, Christophe N, Pairault C, Herbin M, Ward R, Reperant J.1994. Organization of the serotoninergic system in the brain of twoamphibian species, Ambystoma mexicanum (Urodela) and Typhlonectescompressicauda (Gymnophiona). Anat Embryol 190:87–99.
D’Aniello B, Masucci M, di Meglio M, Iela L, Rastogi RK. 1994. Immunohis-tochemical localization of GnRH in the crested newt (Triturus carnifex)brain and terminal nerve: a developmental study. J Neuroendocrinol6:167–172.
D’Aniello B, Pinelli C, Di Fiore MM, Iela L, King JA, Rastogi RK. 1995.Development and distribution of gonadotropin-releasing hormone neu-ronal system in the frog (Rana esculenta) brain: immunohistochemicalanalysis. Brain Res Dev Brain Res 89:281–288.
D’Aniello B, Fiorentino M, Pinelli C, di Meglio M, Vallarino M, Rastogi RK.1996. Distribution of FMRFamide-like immunoreactivity in the brainand pituitary of Rana esculenta during development. Brain Res DevBrain Res 95:194–204.
D’Aniello B, Pinelli C, Jadhao AG, Rastogi RK, Meyer DL. 1999. Compara-tive analysis of FMRFamide-like immunoreactivity in caiman (Caimancrocodilus) and turtle (Trachemys scripta elegans) brains. Cell TissueRes. (in press).
Dockray GJ, Williams RG. 1983. FMRFamide-like immunoreactivity in ratbrain: development of a radioimmunoassay and its application instudies of distribution and chromatographic properties. Brain Res266:295–303.
Dockray GJ, Sault C, Holmes S. 1986. Antibodies to FMRF-amide, and therelated pentapeptide LPLRF-amide, reveal two groups of immunoreac-tive peptides in chicken brain. Regul Pept 16:27–37.
Eisthen HL, Northcutt RG. 1996. Silver lampreys (Ichthyomyzon unicus-pis) lack a gonadotropin-releasing hormone- and FMRFamide-immuno-reactive terminal nerve. J Comp Neurol 370:159–172.
Ekstrom P, Honkanen T, Ebbesson SOE. 1988. FMRFamide-like immunore-active neurons of the nervus terminalis of teleosts innervate both retinaand pineal organ. Brain Res 460:68–75.
Ekstrom P, Ostholm T, Meissl H, Bruun A, Richards JG, Mohler H. 1990.Neural elements in the pineal complex of the frog, Rana esculenta. II:GABA-immunoreactive neurons and FMRFamide-immunoreactive ef-ferent axons. Vis Neurosci 4:399–412.
Fiorentino M, Pinelli C, D’Aniello B, Meyer DL, Rastogi RK. 1999. Placodalorigin of FMRFamide-immunoreactive neurons in the telencephalon of
the common toad, Bufo bufo. In: Elsner N, Eysel U, editors: GottingenNeurobiology Report 1999. Stuttgart: George Thiem Verlag. p 590.
Fischer AJ, Reisch HM, Kyle AL, Stell WK. 1996. Characterization of theRFamide-like neuropeptides in the nervus terminalis of the goldfish(Carassius auratus). Regul Pept 62:73–87.
Frontera JG. 1952. A study of the anuran diencephalon. J Comp Neurol96:1–67.
Fujii K, Kobayashi H. 1992a. FMRFamide-like immunoreactivity in thebrain of the Japanese quail Coturnix coturnix japonica. NeuroendocrLett 14:97–102.
Fujii K, Kobayashi H. 1992b. FMRFamide-like immunoreactivity in thebrain and pituitary of the goldfish, Carassius auratus. Ann Anat174:217–222.
Greenberg MJ, Price DA. 1992. Relationships among the FMRFamide-likepeptides. Progr Brain Res 92:25–37.
Herrick CJ. 1948. The brain of the salamander. Chicago: University ofChicago Press.
Jirikowski G, Erhart G, Grimmelikhuijzen CJP, Triepel J, Patzer RA. 1984.FMRFamide-like immunoreactivity in the brain and pituitary of thehagfish, Eptatretus burgeri (Cyclostomata). Cell Tissue Res 237:363–366.
Kemali M, Braitenberg V. 1969. Atlas of the frog’s brain. Berlin: Springer-Verlag.
Lazar G, Maderdrut JL, Trasti SL, Liposits Z, Toth P, Kozics T, Merchentha-ler I. 1993. Distribution of proneuropeptide Y-derived peptides in thebrain of Rana esculenta and Xenopus laevis. J Comp Neurol 327:551–571.
Lowry CA, Richardson CF, Zoeller TR, Miller LJ, Muske LE, Moore FL.1997. Neuroanatomical distribution of vasotocin in a urodele amphib-ian (Taricha granulosa) revealed by immunohistochemical and in situhybridization techniques. J Comp Neurol 385:43–70.
Magliulo-Cepriano L, Schreibman MP, Blum V. 1993. The distribution ofimmunoreactive FMRFamide, neurotensin, and galanin in the brainand pituitary gland of three species of Xiphophorus from birth to sexualmaturity. Gen Comp Endocrinol 92:269–280.
Marın O, Gonzales A, Smeets WJAJ. 1997. Basal ganglia organization inamphibians: efferent connections of the striatum and the nucleusaccumbens. J Comp Neurol 380:23–50.
Murakami S, Kikuyama S, Arai Y. 1992. The origin of the luteinizinghormone-releasing hormone (LHRH) neurons in the newts (Cynopspyrrhogaster): the effect of olfactory placode ablation. Cell Tissue Res269:21–27.
Muske LE. 1997. Ontogeny, phylogeny and neuroanatomical organizationof multiple molecular forms of GnRH. In: Parhar IS, Sakuma Y, editors.GnRH neurons, gene to behavior. Tokyo: Brain Shuppan. p 145–180.
Muske LE, Moore FL. 1988. The nervus terminalis in amphibians: anatomy,chemistry and relationship with the hypothalamic gonadotropin-releasing hormone system. Brain Behav Evol 12:141–158.
Naujoks-Manteuffel C, Meyer DL. 1996. Glial fibrillary acidic protein in thebrain of the caecilian Typhlonectes natans (Amphibia, Gymnophiona):an immunocytochemical study. Cell Tissue Res 283:151–158.
Neary TJ. 1990. The pallium of anuran amphibians. In: Jones EG, Peters A,editors. Cerebral cortex, vol 8A: comparative structure and evolution ofcerebral cortex, part I. New York: Plenum. p 107–138.
Neary TJ, Northcutt RG. 1983. Nuclear organization of the bullfrogdiencephalon. J Comp Neurol 213:262–278.
Northcutt RG, Kicliter E. 1980. Organization of the amphibian telencepha-lon. In: Ebbesson SOE, editor. Comparative neurology of the telencepha-lon. New York: Plenum. p 203–255.
Northcutt RG, Muske LE. 1991. Experimental embryological evidence ofthe placodal origin of GnRH and FMRFamide neurons of the terminalnerve and preoptic area in salamanders. Soc Neurosci Abstr 17:321.
Northcutt RG, Muske LE. 1994. Multiple embryonic origins of gonadotropin-releasing hormone (GnRH) immunoreactive neurons. Brain Res DevBrain Res 78:279–290.
O’Donohue TL, Bishop JF, Chronwall BM, Groome J, Watson WH. 1984.Characterization and distribution of FMRFamide immunoreactivity inthe rat central nervous system. Peptides 5:563–568.
Oelschlager HA, Helpert C, Northcutt RG. 1998. Coexistence of FMRFamide-like and LHRH-like immunoreactivity in the terminal nerve andforebrain of the big brown bat, Eptesicus fuscus. Brain Behav Evol52:139–147.
Ohtomi M, Fujii K, Kobayashi H. 1989. Distribution of FMRFamide-likeimmunoreactivity in the brain and neurohypophysis of the lamprey,Lampetra japonica. Cell Tissue Res 256:581–584.
304 C. PINELLI ET AL.

Opdam P, Nieuwenhuys R. 1976. Topographical analysis of the brain stemof the axolotl Ambystoma mexicanum. J Comp Neurol 165:285–306.
Opdam P, Kemali M, Nieuwenhuys R. 1976. Topological analysis of thebrain stem of the frog Rana esculenta and Rana catesbeiana. J CompNeurol 165:307–332.
Ostholm T, Ekstrom P, Ebbesson OE. 1990. Distribution of FMRFamide-like immunoreactivity in the brain, retina and nervus terminalis of thesockeye salmon parr, Oncorhynchus nerka. Cell Tissue Res 261:403–418.
Parhar IS, Soga T, Ishikawa Y, Nagahama Y, Sakuma Y. 1998. Neuronssynthesizing gonadotropin-releasing hormone mRNA subtypes havemultiple developmental origins in the medaka. J Comp Neurol 401:217–226.
Pestarino M, Vallarino M. 1996. Immunoreactive FMRFamide-like pep-tides in the brain of the Antarctic icefish, Chionodraco hamatus.Neurosci Lett 213:21–24.
Pinelli C. 1999. Ontogenesi ed analisi comparativa del sistema FMRFa-immunoreattivo nell’encefalo di pesci, anfibi, rettili ed uccelli. Doctoralthesis. Universita degli Studi di Napoli ‘‘Federico II,’’ Napoli, Italy.
Pinelli C, D’Aniello B, Fiorentino M, Bhat G, Saidapur SK, Rastogi RK.1997. Distribution of gonadotropin-releasing hormone immunoreactiv-ity in the brain of Ichthyophis beddomei (Amphibia: Gymnophiona). JComp Neurol 384:283–92.
Pinelli C, D’Aniello B, Guarino FM, Fiorentino M, Angelini F, Rastogi RK.1999. Ontogenetic profile of FMRFamide-like immunoreactivity in thecentral nervous system of the viviparous skink, Chalcides chalcides. In:Elsner N, Eysel U, editors: Gottingen Neurobiology Report 1999.Stuttgart: George Thiem Verlag. p 591.
Potter HD. 1969. Structural characteristics of cell and fiber populations inthe optic tectum of the frog (Rana catesbeiana). J Comp Neurol136:203–232.
Price DA, Greenberg MJ. 1977. Structure of a molluscan cardioexcitatoryneuropeptide. Science 197:670–671.
Price DA, Greenberg MJ. 1989. The hunting of the FaRPs: the distributionof FMRFamide-related peptides. Biol Bull 177:198–205.
Puelles L, FJ Milan M. MartInez-de-la-Torre 1996. A segmental map ofarchitectonic subdivisions in the diencephalon of the frog Rana perezi:acetylcholinesterase-histochemical observations. Brain Behav Evol 47:279–310.
Quanbeck C, Sherwood NM, Millar RP, Terasawa E. 1997. Two populationsof luteinizing hormone-releasing hormone neurons in the forebrain ofthe Rhesus macaque during embryonic development. J Comp Neurol379:1–17.
Raffa RB. 1988. The action of FMRFamide (Phe-Met-Arg-Phe-NH2) andrelated peptides on mammals. Peptides 9:915–922.
Raffa RB, Heyman J, Porreca F. 1986. Intrathecal FMRFamide (Phe-Met-Arg-Phe-NH2) induces excessive grooming behavior in mice. NeurosciLett 65:94–98.
Rama Krishna NS, Subhedar NK. 1992. Distribution of FMRFamide-likeimmunoreactivity in the forebrain of the catfish, Clarias batrachus(Linn.). Peptides 13:183–191.
Rastogi RK, D’Aniello B, Fiorentino M, Pinelli C, di Meglio M. 1996.Coexistence of FMRFamide and gonadotropin-releasing hormone immu-noreactivities in the anterior preoptic area and median eminence of frogbrain. In: Joss J, editor. Environmental and conservation endocrinology.Sydney: Macquarie University. p 179–180.
Roth BL, Disimone J, Majane EA, Yang HYT. 1987. Elevation of arterialpressure in rats by two new vertebrate peptides FLFQPQRF-NH2 andAGEGLSSPFWSLAAPQRF-NH2 which are immunoreactive to FMRF-NH2 antiserum. Neuropeptides 10:37–42.
Roth G, Nishikawa KC, Naujoks-Manteuffel C, Schmidt A, Wake DB. 1993.Paedomorphosis and simplification in the nervous system of sala-manders. Brain Behav Evol 42:137–170.
Schwanzel-Fukuda M. 1997. The origin and migration of LHRH neurons inmammals: a comparison between species including humans. In: ParharIS, Sakuma Y, editors. GnRH neurons, gene to behavior. Tokyo: BrainShuppan. p 221–242.
Shen CL, Cheng JT. 1989. Cross-reaction of FMRFamide with a neuropep-tide Y (NPY) antiserum: histological and radioimmunological evidence.Biomed Ref 10:517–520.
Stell WK, Waller SE, Chohan KS, Ball AK. 1984. The goldfish nervus
terminalis: a luteinizing-hormone-releasing hormone and molluscancardioexcitatory peptide immunoreactive olfactoretinal pathway. ProcNatl Acad Sci USA 81:940–944.
Tang J, Yang HY, Costa E. 1984. Inhibition of spontaneous and opiate-modified nociception by an endogenous neuropeptide with Phe-Met-Arg-Phe-NH2-like immunoreactivity. Proc Natl Acad Sci USA 81:5002–5005.
Triepel J, Grimmelikhuijzen Y. 1984. Mapping of neurons in the centralnervous system of the guinea pig by use of antisera specific to themolluscan neuropeptide FMRFamide. Cell Tissue Res 237:575–586.
Uchiyama H, Reh TA, Stell WK. 1988. Immunocytochemical and morpho-logical evidence for a retinopetal projection in anuran amphibians. JComp Neurol 274:48–59.
Vallarino M, Salsotto-Cattaneo M, Feuiolloley M, Vaudry H. 1991. Distribu-tion of FMRFamide-like immunoreactivity in the brain of the elasmo-branch fish Scyliorhinus canicula. Peptides 12:1321–1328.
Vallarino M, Salsotto-Cattaneo M, Vaudry H. 1993. Immunohistochemicallocalization of FMRFamide-like peptides in the brain of the frog, Ranaesculenta. In: Facchinetti F, Henderson IW, Pierantoni R, Polzonetti-Magni A, editors: Cellular Communication in Reproduction. J Endocri-nol 1993:45–48.
Vallarino M, Feuilloley M, D’Aniello B, Rastogi RK, Vaudry H. 1994.Distribution of FMRFamide-like immunoreactivity in the brain of thelizard Podarcis sicula. Peptides 15:1057–1065.
Vallarino M, M, Thoumas JL, Demorgny R, Masini MA, Vaudry H. 1995.Distribution of FMRFamide-like immunoreactivity in the brain of thelungfish Protopterus annectens. Peptides 16:1187–1196.
Vecino E, Ekstrom P. 1990. Distribution of met-enkephalin, leu-enkephalin,substance P, neuropeptide Y, FMRFamide, and serotonin immunoreac-tivities in the optic tectum of the Atlantic salmon (Salmo salar L.). JComp Neurol 299:229–241.
Vecino E, Ekstrom P. 1992. Colocalization of neuropeptide Y (NPY)-like andFMRFamide-like immunoreactivities in the brain of Atlantic salmon(Salmo salar). Cell Tissue Res 270:435–442.
Wada M, Urano A, Gorbman A. 1980. A stereotaxic atlas for diencephalicnuclei of the frog Rana pipiens. Arch Histol Jpn 43:157–173.
Weber E, Evans CJ, Samuelsson SJ, Bachas JD. 1981. Novel peptideneuronal system in the rat brain and pituitary. Science 214:1248–1251.
White RB, Fernald RD. 1998. Ontogeny of gonadotropin-releasing hormone(GnRH) gene extression reveals a distinct origin for GnRH-containingneurons in the midbrain. Gen Comp Endocrinol 112:322–329.
Wicht H, Himstedt W. 1988. Topological and connectional analysis of thedorsal thalamus of Triturus alpestris (Amphibia, Urodela, Salamandri-dae). J Comp Neurol 267:545–561.
Wicht H, Northcutt RG. 1992. FMRFamide-like immunoreactivity in thebrain of the Pacific hagfish, Eptatetrus stouti (Myxinoidea). Cell TissueRes 270:443–449.
Williams RG, Dockray GJ. 1983. Immunohistochemical studies ofFMRFamide-like immunoreactivity in rat brain. Brain Res 276:213–219.
Wirsig-Wiechmann CR, Basinger SF. 1988. FMRFamide-immunoreactiveretinopetal fibers in the frog, Rana pipiens: demonstration by lesion andimmunocytochemical techniques. Brain Res 449:116–134.
Wong TM, Greenberg MJ, Tse SYH. 1985. Cardiovascular effects ofintraventricular injection of FMRFamide, met-enkephalin and theircommon analogues in the rat. Comp Biochem Physiol 81:175–179.
Wright DE, Demski LS. 1996. Organization of GnRH and FMRFamidesystems in two primitive bony fishes (order Polypteriformes). BrainBehav Evol 47:267–278.
Yamamoto N, Uchiyama H, Ohki-Hamazaki H, Tanaka H, Ito H. 1996.Migration of GnRH-immunoreactive neurons from the olfactory placodeto the brain: a study using avian embryonic chimeras. Brain Res95:234–244.
Yang HYT, Majane EA. 1990. Mammalian Phe-Met-Arg-Phe-NH2-likepeptides: structure, biological activity and distribution. In: Epple A,Scanes CG, Stetson MH, editors. Progress in comparative endocrinol-ogy. New York: Wiley-Liss. p 86–91.
Zilles K, Wetsch U, Schleicher A. 1981. The telencephalon of Ichthyophispaucisulcus (Amphibia, Gymnophiona (5Caecilia)): a quantitative cyto-architectonic study. Z Mikrosk-Anat Forsch Leipzig 95:943–962.
FMRFAMIDE IN AMPHIBIAN BRAIN 305





























