Embed Size (px)
Citation preview

Distribution of Prostasomes in NeoplasticEpithelial Prostate Cells
B. Ove Nilsson,1 Lars Egevad,2 Meishan Jin,1 Gunnar Ronquist,3* andChrister Busch2
1Department of Human Anatomy, University of Uppsala, Uppsala, Sweden2Department of Pathology, University of Uppsala, Uppsala, Sweden
3Department of Clinical Chemistry, University of Uppsala, Uppsala, Sweden
BACKGROUND. Prostasomes are a secretory product from the prostate. We aimed to in-vestigate whether the distribution and amount of prostasomes in normal prostate epitheliumwere influenced by the dedifferentiation occurring in adenocarcinomas of the human prostategland.METHODS. Transurethrally resected material from 11 patients with prostatic carcinoma ofvarious malignancy grades, material from two lymph node metastases, and benign tissuefrom 10 total prostatectomies were subjected to immunohistochemical staining, using a mousemonoclonal antibody against human prostasomes (mAb 78).RESULTS. Immunostaining of low-grade carcinoma was similar to that of normal prostategland which displayed a cytoplasmic granular staining of the apical (luminal) aspects of thesecretory epithelial cells. In moderately well and poorly differentiated adenocarcinoma, theamount of stained components decreased, and the staining pattern became more heteroge-neous. In multilayered glandular structures, the staining was concentrated at the lumen,leaving most other cells negative. The neoplastic cells of lymph node metastases of prostatecarcinoma differed in amount and distribution of immunostained prostasomes.CONCLUSIONS. The antigen recognized in the prostasomes by mAb78 was expressed inbenign prostate tissue, prostate cancer, and to a lesser degree in lymph node metastases. Therewas a tendency towards decreased expression with increasing tumor grade. Prostate 39:36–40,1999. © 1999 Wiley-Liss, Inc.
KEY WORDS: prostate; prostate adenocarcinoma; prostasomes; monoclonal antibodies;immunohistochemistry
INTRODUCTION
Prostasomes are small vesicles with a size of about50–500 nm [1,2] appearing in human seminal plasma.They are derived from the prostatic secretory epithe-lial cells [3]. The prostasomes are located in the apicalparts of these cells, where they occur within storagevesicles. The prostasomes are released into the glandducts by exocytosis when a storage vesicle fuses withthe apical cell plasma membrane [3]. This is a processwhich loads the prostatic secretion in the gland ductswith prostasomes. When the prostatic secretion is dis-charged at ejaculation, the prostasomes will appear inthe seminal plasma in free form and to some extentcoating the spermatozoa [4].
The distribution of prostasomes in the prostatic
secretory epithelial cells and the gland ducts wasexamined with immunohistochemistry, using a mono-clonal anti-prostasome antibody (mAb78) [5]. SincemAb 78 is a marker for prostasomes in normally se-creting prostatic epithelium, we examined the ex-pression of prostasomes, as indicated by mAb 78-immunohistochemistry, in neoplastic epithelial pros-tate cells and in lymph node metastases of prostate
Grant sponsor: Swedish Medical Research Council; Grant number:Project 00070; Grant sponsor: Swedish Association for Medical Re-search.*Correspondence to: Gunnar Ronquist, M.D., Department of Clini-cal Chemistry, University Hospital, S-751 85 Uppsala, Sweden.Received 3 November 1997; Accepted 15 September 1998
The Prostate 39:36–40 (1999)
© 1999 Wiley-Liss, Inc.

carcinoma. The aim of the investigation was to exam-ine the expression and distribution pattern of prosta-somes in prostatic tissues of different grades of malig-nancy.
MATERIALS AND METHODS
Human Tissues
Normal prostate gland tissue was obtained from 10total prostatectomy specimens and included seminalvesicle and transition zone tissue. Transurethrally re-sected material (TUR-P) was secured from 11 patientswith prostatic carcinoma of various malignancygrades based on the Gleason score [6]: 2 patients hada Gleason score (GS) of 4, another 2 had GS 5, 2 wereGS 6, 1 was GS 7, 1 was GS 8, 2 were GS 9, and 1patient had GS 10. Lymph node metastases were de-rived from 2 cases of prostate cancer. Also, lymphnode material without prostate cancer metastasis wasobtained from 10 cases in which preoperative lymph-adenectomy was performed, and from cancer-negative lymph nodes from 3 breast cancer patients.The age of patients ranged from 61–84 years. The tis-sues were fixed in 4% neutral-buffered formaldehyde,paraffin-embedded, and sectioned for immunohisto-chemical staining.
Immunohistochemical Localizationof Prostasomes
Sections were deparaffinized and rehydrated bystandard procedures. They were placed in Coplin jarswith citrate buffer (pH 6.0) and boiled for 2 × 5 minin a microwave oven. After cooling, the sectionswere blocked using 10% bovine serum albumin(BSA) (Sigma Chemical Co., St. Louis, MO) and 3%normal goat serum for 30 min and processed as fol-lows:
1. Mouse anti-human prostasome mAb78, diluted 1:5with phosphate-buffered saline (PBS), overnight in-cubation at 4°C in a moist chamber.
2. Biotin-conjugated rabbit immunoglobulin againstmouse IgG (Sigma Chemical Co.), diluted 1:200with 3% BSA in PBS, for 30 min.
3. Avidin-biotin-alkaline phosphatase-complex(DAKO, Copenhagen, Denmark), diluted 1:50 with3% BSA in PBS for 30 min.
4. Detection system: Ni-DAB (1% NiSO4 × 6 H2O;Merck, Darmstadt, Germany) and 0.05% DAB(Sigma Chemical Co.) in 0.1 M acetate buffer,pH 7.2 for 20 min. Between incubations, sectionswere rinsed three times with PBS (pH 7.6) for 5min each. They were then counterstained with
Kernechtrot (Apoteksbolaget, Stockholm, Sweden)for 10 min. To visualize macrophages a secondstaining was performed with antibodies KP1 orMAC 387 (both from DAKO). In this double stain-ing, no microwave pretreatment was used, butthe sections were treated with protease (1:2,Sigma Chemical Co.) at room temperature for 10min.
Staining was performed as follows:
1–4. As reported above.5. Sections were then blocked with 10% BSA for 20
min.6. MAC 387 antibody diluted 1:100 or KP1 antibody
diluted 1:50 with 3% BSA in PBS for 30 min.7. Biotin-conjugated rabbit antimouse immunoglobu-
lin (DAKO) diluted 1:200 with 3% BSA in PBS for 30min.
8. Avidin-biotin-alkaline phosphatase -complex(DAKO) diluted 1:50 with 3% BSA in PBS for 30min.
9. Substrate for alkaline phosphatase (Kit 1, VectorRed, Vector Laboratories, Burlingame, CA) withTris-buffered saline, pH 8.2, for 20 min. Betweenincubations the sections were rinsed three timeswith PBS (pH 7.6) for 5 min each in the doublestaining. They were then counterstained with LightGreen (Merck) for 1 min. The negative control con-sisted of a human prostate section incubated with-out the primary antibody.
RESULTS
Prostasome Distribution in NormalProstatic Epithelium
Three cell types are present in the prostatic epithe-lium: secretory, basal, and endocrine-paracrine cells[7,8]. The mAb 78 bound exclusively to the ducto-acinar secretory cells of the prostate gland. Thestaining was cytoplasmic and most intense in theapical (luminal) aspect of the secretory epithe-lium, whereas the basal cells were negative (Fig. 1).Some of the inspissated secretion (corpora amylacea)showed granular staining with mAb78. All compo-nents of the stroma were negative with the exceptionof rather numerous macrophages, which were con-stantly stained. Double immunostaining also using themacrophage marker MAC 387 revealed the macro-phage identity of the stained cells. Macrophages inother locations such as normal skin, granulation tis-sue, and reactive lymph nodes were also labelled withmAb78.
Prostasomes and Prostate Cancer 37
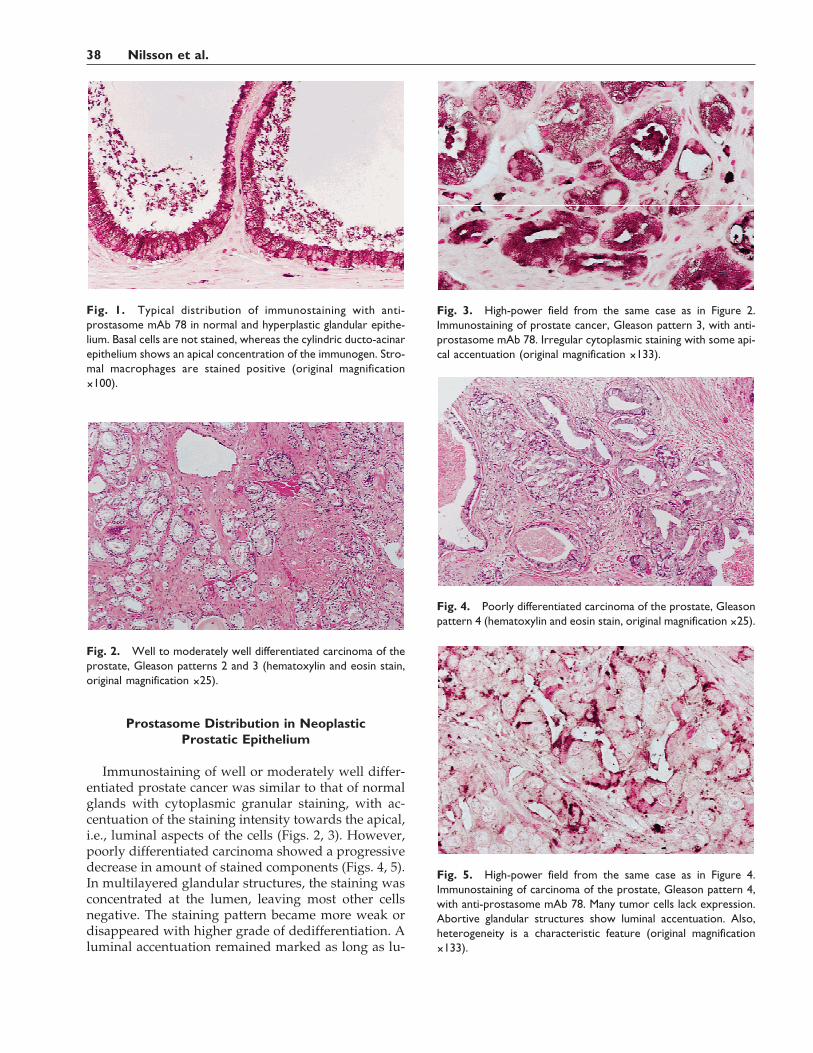
Prostasome Distribution in NeoplasticProstatic Epithelium
Immunostaining of well or moderately well differ-entiated prostate cancer was similar to that of normalglands with cytoplasmic granular staining, with ac-centuation of the staining intensity towards the apical,i.e., luminal aspects of the cells (Figs. 2, 3). However,poorly differentiated carcinoma showed a progressivedecrease in amount of stained components (Figs. 4, 5).In multilayered glandular structures, the staining wasconcentrated at the lumen, leaving most other cellsnegative. The staining pattern became more weak ordisappeared with higher grade of dedifferentiation. Aluminal accentuation remained marked as long as lu-
Fig. 1. Typical distribution of immunostaining with anti-prostasome mAb 78 in normal and hyperplastic glandular epithe-lium. Basal cells are not stained, whereas the cylindric ducto-acinarepithelium shows an apical concentration of the immunogen. Stro-mal macrophages are stained positive (original magnification×100).
Fig. 2. Well to moderately well differentiated carcinoma of theprostate, Gleason patterns 2 and 3 (hematoxylin and eosin stain,original magnification ×25).
Fig. 3. High-power field from the same case as in Figure 2.Immunostaining of prostate cancer, Gleason pattern 3, with anti-prostasome mAb 78. Irregular cytoplasmic staining with some api-cal accentuation (original magnification ×133).
Fig. 4. Poorly differentiated carcinoma of the prostate, Gleasonpattern 4 (hematoxylin and eosin stain, original magnification ×25).
Fig. 5. High-power field from the same case as in Figure 4.Immunostaining of carcinoma of the prostate, Gleason pattern 4,with anti-prostasome mAb 78. Many tumor cells lack expression.Abortive glandular structures show luminal accentuation. Also,heterogeneity is a characteristic feature (original magnification×133).
38 Nilsson et al.

mina prevailed. In fact, the immunostaining of re-maining, secreted prostasomes tended to highlight thelumina, even of abortive glandular formations.
Prostasome Distribution in Lymph NodeMetastases of Prostate Cancer
The prostate carcinoma metastases in lymph nodesshowed the same distribution pattern as that of pri-mary tumours, i.e., the degree of immunostaining de-creased with decreasing grade of differentiation.
DISCUSSION
Diagnosis and prognostication of prostate carci-noma are important issues in diagnostic uropathol-ogy. Various histochemical markers for prostatic can-cer cells have been evaluated by, among other things,correlating the staining reactions with combinations ofhistopathological grades, presence of metastases, orsurvival [7–11]. We examined the changes in distribu-tion of prostasomes, which are secretory componentsin the prostate epithelial cells and involved in the pro-cess of reproduction [3].
Many functional effects are ascribed to the prosta-somes. For instance, having attached onto washed,prostasome-free spermatozoa they are able to exert astimulatory influence on the progressive motility ofsperm cells [12,13]. They have an immunosuppressivecapacity by inhibiting mitogen-induced proliferationof lymphatic cells [2,14] and phagocytosis by macro-phages [15,16]. The prostasomes also regulate thecomplement activation cascade [17]. For instance, theprostasomes contain CD46 [18], a factor for proteolyticinactivation of C3b and C4b, and CD55 [17], the decay-accelerating factor. Furthermore, CD59, which is aninhibitor of the membrane attack complex (MAC), hasbeen demonstrated in prostasomes [19]. It has beensuggested that the prostasomes, by coating the sper-matozoa [4], assist them in escaping from the immu-nological defense system in the female genital tract [2].Considering the immunosuppressive capacity of theprostasomes, it has even been discussed whether theirpresence in the prostatic gland and excretory ductscould contribute to the frequent occurrence of variousinfections and prostatic carcinoma [2].
The immunomarker mAb78 for prostasomes wasproduced by splenic immunization with purifiedprostasomes [20]. Electrophoresis and Western blotsdemonstrated that the antibody detected a 35-kDaband in prostasomes, ejaculated prostasome-coatedsperm cells, and prostatic tissues [21]. The mAb 78 didnot bind to the prostate specific antigen (PSA) or pros-tatic acid phosphatase (PAP) bands, indicating that itwas different from anti-PSA and anti-PAP antibodies
[21]. The antibody, however, was not specific for pros-tasomes but cross-reacted with antigen epitopes in theapical parts of cells of the liver lobuli and the exocrinepancreas, in some cells of the pancreatic islets, in first-trimester syncytial trophoblast, in the monocyte/macrophage lineage, and in the apical parts of theseminal gland epithelium. Hence, not only our mono-clonal antibody [22] but also a polyclonal antibody[23] against prostasomes react dispersedly with someantigens on other cell structures, both inside and out-side the urogenital tract. Notwithstanding this, there isa pure prostatic origin of the prostasomes [1,24–27],and there are no indications of a production and se-cretion of any similar structures in other accessorygenital glands in man [25,26]. The prostate gland asthe sole source of prostasomes also tallies with thefindings obtained by immunoelectron miscroscopydemonstrating tissue factor antigen located on the sur-faces of prostasomes, while no tissue factor was foundin seminal vesicle secretions in man [28].
Immunostaining of prostate glands with mAb78 re-vealed a substance, located apically in the prostateepithelium and in the gland ducts. Since this reason-ably represented a prostatic secretion, it was con-cluded that the antibody recognized the prostasomesof the epithelium in normal glands [5]. Therefore, itwould be expected that the prostasomes, when normalprostate cells are transformed to a neoplastic state andbecome less differentiated, would decrease in number.We found that mAb 78 labelled the epithelial cells inwell and moderately well differentiated carcinomas,but that the staining pattern became weaker or disap-peared in less well differentiated carcinomas. Themarker showed a distinct concentration at the glandu-lar lumen, even at very small and sometimes abortivelumina. Thus, there was a covariance between theamount and distribution of immunostaining and thegrade of differentiation of prostate neoplastic cells.This relationship to differentiation was retained inlymph node metastases of prostate carcinoma, i.e., theprostate neoplastic cells contained varying amounts ofimmunostained prostasomes.
CONCLUSIONS
The expression of an antigen recognized in prosta-somes by mAb78 covaries with the functional differ-entiation of the prostate epithelium. It may be usefulas a marker and prognostic tool in prostate cancer andshould be further evaluated and compared with othermarkers.
ACKNOWLEDGMENTS
The authors thank Ms. Barbro Einarsson and Kata-rina Engstrøm for their excellent technical assistance.
Prostasomes and Prostate Cancer 39

The study was supported by the Swedish Medical Re-search Council (project 00070 to B.O.N.) and by theSwedish Association for Medical Research (to M.J.).
REFERENCES
1. Brody I, Ronquist G, Gottfries A. Ultrastructural localization ofthe prostasome—an organelle in human seminal plasma. Ups JMed Sci 1983;88:63–80.
2. Kelly RW, Holland P, Skibinski G, Harrison C, McMillan L,Hargreave T, James K. Extracellular organelles (prostasomes)are immunosuppressive components of human semen. Clin ExpImmunol 1991;86:550–556.
3. Ronquist G, Brody I. The prostasome: its secretion and functionin man. Biochim Biophys Acta 1985;822:203–218.
4. Ronquist G, Nilsson BO, Hjerten S. Interaction between prosta-somes and spermatozoa from human semen. Arch Androl 1990;24:147–157.
5. Nilsson BO, Jin M, Ronquist G. Immunolocalization of prosta-somes in the human prostate. Ups J Med Sci 1996;101:149–158.
6. Gleason DF. Histologic grading and clinical staging of prostaticcarcinoma. In: Tannenbaum M, editor. Urologic pathology: theprostate. Philadelphia: Lea & Febiger; 1977. p 171–197.
7. Bonkhoff H, Stein U, Remberger K. Multidirectional differentia-tion in the normal, hyperplastic, and neoplastic human prostate:simultaneous demonstration of cell-specific epithelial markers.Hum Pathol 1994;25:42–46.
8. Dhom G, Seitz G, Wernert N. Histology and immunohistochem-istry studies in prostate cancer. Am J Clin Oncol 1988;11:537–542.
9. Chu TM. Prostate cancer-associated markers. Immunol Res1990;53:339–356.
10. Isaacs JT. Molecular markers for prostate cancer metastasis. De-veloping diagnostic methods for predicting the aggressivenessof prostate cancer. Am J Pathol 1997;150:1511–1521.
11. Shi X-B, Gumerlock PH, de Vere White RW. Molecular biologyof prostate cancer. World J Urol 1996;14:318–328.
12. Stegmayr B, Ronquist G. Promotive effect on human sperm pro-gressive motility by prostasomes. Urol Res 1982;10:253–257.
13. Fabiani R, Johansson L, Lundkvist O, Ulmsten U, Ronquist G.Promotive effect by prostasomes on normal human spermato-zoa exhibiting no forward motility due to buffer washings. EurJ Obstet Gynecol Reprod Biol 1994;57:181–188.
14. Kelly RW. Immunosuppressive mechanisms in semen: implica-tions for contraception. Hum Reprod 1995;10:1686–1693.
15. Skibinski G, Kelly RW, Harkiss D, James K. Immunosuppres-sion by human seminal plasma—extracellular organelles (pros-tasomes) modulate activity of phagocytic cells. Am J ReprodImmunol 1992;28:97–103.
16. Lazarevic M, Skibinski G, Kelly RW, James K. Immunomodu-latory effects of extracellular secretory vesicles isolated frombovine semen. Vet Immunol Immunopathol 1995;44:237–250.
17. Rooney IA, Heuser JE, Atkinson JP. GPI-anchored complementregulatory proteins in seminal plasma. An analysis of theirphysical condition and the mechanisms of their binding to ex-ogenous cells. J Clin Invest 1996;97:1675–1686.
18. Kitamura M, Namiki M, Matsumiya K, Tanaka K, MatsumotoM, Hara T, Kiyohara H, Okabe M, Okuyama A, Seya T. Mem-brane cofactor protein (CD46) in seminal plasma is a prosta-some-bound form with complement regulatory activity andmeasles virus neutralizing activity. Immunology 1995;84:626–632.
19. Rooney IA, Atkinson JP, Krul ES, Schonfeld G, Polakoski K,Saffitz JE, Morgan BP. Physiologic relevance of the membraneattack complex inhibitory protein CD59 in human seminalplasma: CD59 is present on extracellular organelles (prosta-somes), binds cell membranes, and inhibits complement-mediated lysis. J Exp Med 1993;177:1409–1420.
20. Nilsson BO, Svalander PC, Larsson A. Immunization of miceand rabbits by intrasplenic deposition of nanogram quantities ofprotein attached to sepharose beads or nitrocellulose paperstrips. J Immunol Methods 1987;99:67–75.
21. Jin M, Nilsson BO, Larsson A, Hellman U, Ronquist G. Theanti-human prostasome mAb 78 binds to an antigen distinctfrom PSA and PAP. J Urol 1997;157:1932–1936.
22. Nilsson BO, Jin M, Einarsson B, Persson BE, Ronquist G. Mono-clonal antibodies against human prostasomes. Prostate 1998;35:178–184.
23. Renneberg H, Konrad L, Dammshauser I, Seitz J, Aumuller G.Immunohistochemistry of prostasomes from human semen.Prostate 1997;30:98–106.
24. Ronquist G, Brody I, Gottfries A, Stegmayr B. An Mg2+ andCa2+-stimulated adenosine triphosphatase in human prostaticfluid: part I and part II. Andrologia 1977;10:261–272, 427–433.
25. Brody I, Ronquist G, Gottfries A, Stegmayr B. Abnormal defi-ciency of both Mg2+ and Ca2+-dependent adenosine triphospha-tase and secretory granules and vesicles in human seminalplasma. Scand J Urol Nephrol 1981;15:85–90.
26. Stegmayr B, Ronquist G, Kollberg H, Brody I. Analysis of ejacu-lates from patients with cystic fibrosis. Ultrastruct Pathol 1981;2:357–363.
27. Stegmayr B, Busch C, Fritjofsson Å, Ronquist G. Biochemicaland morphologic studies of the prostate gland in men subjectedto radical cystectomy. Ups J Med Sci 1985;90:139–145.
28. Fernandez JA, Heeb MJ, Radtke KP, Griffin JH. Potent bloodcoagulant activity of human semen due to prostasome-boundtissue factor. Biol Reprod 1997;56:757–763.
40 Nilsson et al.



















![Neuroendocrine cells in the normal, hyperplastic and neoplastic … · 2016. 3. 10. · 1970s [10, 57], but only recently have NE cells in prostate cancer gained increasing attention,](https://img.pdfslide.net/doc/110x75/60a7f8be0d1a990af03a59fd/neuroendocrine-cells-in-the-normal-hyperplastic-and-neoplastic-2016-3-10-1970s.jpg)









