Embed Size (px)
Citation preview

Disturbance Ecology: Complex Interactions
Kyle Apigian Christa Dagley Igor Lacan
18 November 2003
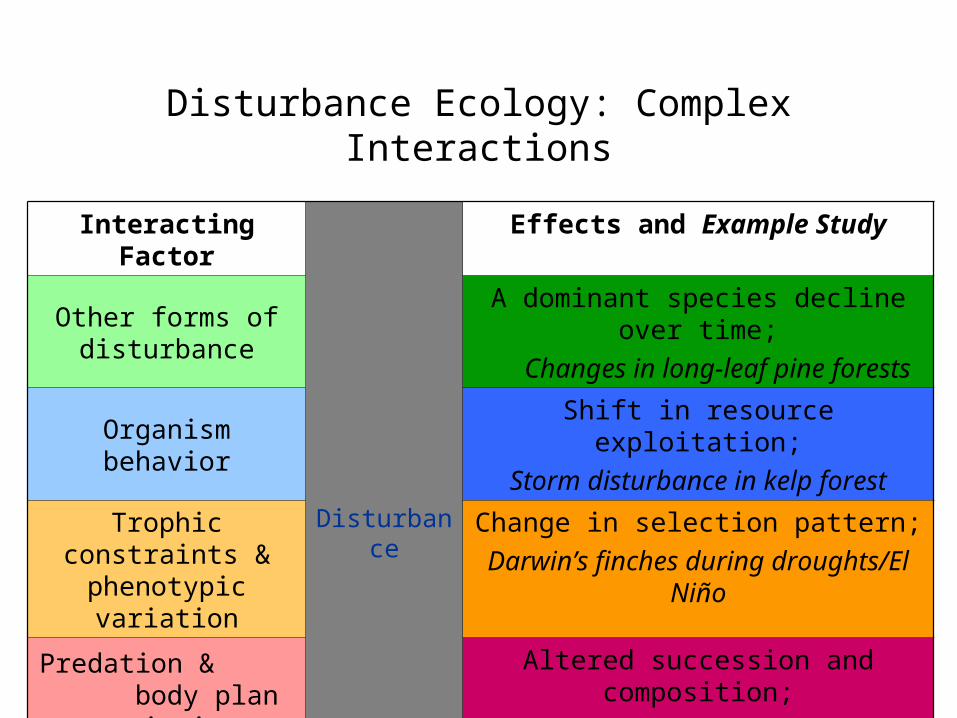
Disturbance Ecology: Complex Interactions
Interacting Factor
Disturbance
Effects and Example Study
Other forms of disturbance
A dominant species decline over time;
Changes in long-leaf pine forests
Organism behaviorShift in resource exploitation;
Storm disturbance in kelp forest
Trophic constraints & phenotypic variation
Change in selection pattern;
Darwin’s finches during droughts/El Niño
Predation & body plan variation
Altered succession and composition;
Fish and storms in tropical creek
CompetitionAltered community composition;
Indirect and toxic effect of a pollutant

• Longleaf Pine
• (Pinus palustris)
Complex Interactions

• Fire-dependent ecosystem– summer lightning fires – 2-8yr FRI
• Most species rich vascular plant communities found in the temperate zone
• Pine savannas - highest level of endemism in N. America
• Today only 3% of its original acreage remains
Complex Interactions

• Contributing factors include:
• naval stores • logging • competition • feral hogs• intensive forestry• fire exclusion
Complex Interactions

Complex Interactions
• First product
• Uses: seal cracks on wooden ships, preserve ropes and sails.

• Contributing factors include:
• naval stores • logging • competition • feral hogs• intensive forestry• fire exclusion
Complex Interactions

• Competition
– loblolly and other pines– large seeds eaten by birds, rodents and
insects – irregular seed production
Complex Interactions

• Contributing factors include:
• naval stores • logging • competition • feral hogs
– 20 seedlings/ha vs. 14,826 s/ha
• intensive forestry• fire exclusion
Complex Interactions

• Contributing factors include:
• naval stores • logging • competition • feral hogs• intensive forestry
– very poor survival– perceived slow early growth
• fire exclusion
Complex Interactions

• Contributing factors include:
• naval stores • logging • competition • feral hogs• intensive forestry• fire exclusion
– grass stage– thick bark– good self pruner
Complex Interactions

• Effects of chronic human activities on invasion of longleaf pine by sand pine
• Florida Panhandle
• Landscape Level: aerial photography to quantify the extent and expansion of sand pine.
• Population Level: 12 stands sampled to verify photo interpretation and assess successional trends
• To assess past vegetation patterns and land-use activities, qualitative data for the area was collected
Complex Interactions McCay (2000)

Complex Interactions
Total = 41,900 ha

• Noninvasive - Stands with no longleaf pine in the canopy.
Complex Interactions Distribution of SP establishment in 5-yr intervals
Grazing discont.
Turpentining ceased Fire suppression

• Conclusions:
• “sand pine invasion occurred b/c of the complex interaction of several factors that increased the susceptibility of LL to enchroachment”
• Turpentining
• competition
• LL crop failures
• fire suppression
• logging
Complex Interactions

• Effects of fire regime and habitat on tree dynamics
• 8 year study• sandhills and flatwoods sites• Fire frequency (annual or biennial) • Fire season (8 different times throughout
the year) • Limited fire temperature and intensity
data taken
• Looked at recruitment, growth, mortality, change in density and BA, and species composition.
Complex Interactions Glitzenstein et. al (1995)

• Results:
• No systematic or predictable effects of season or frequency of burning on LL dynamics.
• Deciduous oaks were most vulnerable to burning in the early growing season (higher mortality, decline in BA and density).
• Spring/summer burns = LL dominated forests
• Dormant season burns = gradual decrease in LL and shift to oak dominated
Complex Interactions

• Take Home Message:
• “Long-term persistence of LL, and perhaps other fire-
adapted species in frequently burned LL-dominated
communities, may be determined by complex
interactions between habitat factors and fire
regimes”.
Complex Interactions

• References
• Glitzenstein, J.S., W.J. Platt, and D.R. Streng. 1995. Effects on fire regime and habitat on tree dynamics in N. Florida longleaf
pine savannas. Ecological Monographs. 65(4):441-476.
• McCay, Deanna. 2000. Effects of chronic human actvities on invasion of longleaf pine forests by sand pine. Ecosystems. 3: 283-
292.
• Outcalt, Kenneth. 2002. The Longleaf pine ecosystem of the south. Native Plants Journal. 1(1) 42-51.
Complex Interactions

Southern California reef – Kelp, sea urchins, and surfperchbehavioral responses of urchins and fish after storms
Galapagos Islands – Darwin’s finchesbehavioral and phenotypic responses to drought and El Nino

Severe storm disturbances and reversal of community structure in a southern California kelp forestA.W. Ebeling, D.R. Laur and R.J. Rowley. 1985. Marine Biology 84, 287-294
From Ebling et al 1985

Post storm 1: Kelp canopy removed – drift kelp lost Urchins left crevices to find food – ate standing kelp and algal turf Urchin population poorly regulated – reef became a barrens
Post storm 2: Storm killed off exposed urchins Kelp resettled in great abundance
Same disturbance resulted different (reverse) effects depending on prior community structure
From Ebling et al 1985

Surfperch feed on benthic arthropods that live in the algal turf
Loss of algal turf caused a decline in surfperch populations
From Ebling et al 1985

Storm effects were not equal across reef, some microhabitats (reef slope, reef crest) retained algal turf
5 surfperch species converged in terms of microhabitat usage
Diet was not significantly affected
Less aggressive, generalist fish left early in season competitive exclusion?
From Stouder 1987

Storm effects were not equal across reef, some microhabitats (reef slope, reef crest) retained algal turf
5 surfperch species converged in terms of microhabitat usage
Diet was not significantly affected
Less aggressive, generalist fish left early in season competitive exclusion?
From Stouder 1987

Darwin’s finches: effects of climactic disturbances on population structure and natural selection
From Grant 1986

Rainfall on the Galapagos
Galapagos islands: high inter-annual variability in rainfalldrought years – little or no rainEl Niño years – excessive rain
From Grant et al 2000

Ex. 1. Geospiza conirostris: large cactus finch
High intraspecific variation in beak size and shape in this species: From long and pointed to shorter and deep.
Beak shape is related to foraging success in one of 3 foraging modes during the dry season:
1. Hammering Opuntia (cactus) fruits long, pointed bills2. Seed cracking long, deep bills3. Stripping bark (to obtain arthropods) deep bills
From Grant and Grant 1989

Ex. 1. Geospiza conirostris: large cactus finch
Extreme rain followed by drought affectedfood supply (cactus fruit and flowers)
Birds with long, pointed bills were at aselective disadvantage.
Birds with deep bills could exploit otherfoods
From Grant and Grant 1989

Ex. 1. Geospiza conirostris: large cactus finch
Primary foraging mode:
Small seeds
Large hard seeds
Ripping Opuntia cactus pads to obtain arthropods
From disruptive to directional selection: Variety of beaks favored in normal years large deep bills favored after disturbance
From Grant and Grant 1989

Ex. 2. Geospiza fortis: medium ground finch - DROUGHT
Drought of 1977: resulted in 85% decline in finch population on Daphne Major
Decline was highly correlated with a decline in seed abundance
From Boag and Grant 1981

Ex. 2. Geospiza fortis: medium ground finch - DROUGHT
Effects of this disturbance on population structure were non-random: large birds more likely to survive than small birds
Large, hard seeds became proportionally more abundant during the drought, as competition for small seeds became intense
Large birds (with large bills) could crack the remaining hard seeds
From Boag and Grant 1981

Ex. 2. Geospiza fortis: medium ground finch - DROUGHT
The drought of 1977 resulted in phenotypic changes to the G. fortis population
From Boag and Grant 1981
Proportional increase in fitness
Body size
Bill “pointedness”

From Grant et al 2000

Ex. 3. Geospiza fortis and G. scandens – EL NINO
Caterpillar abundance is significantly greater during El Niño events
From Grant et al 2000
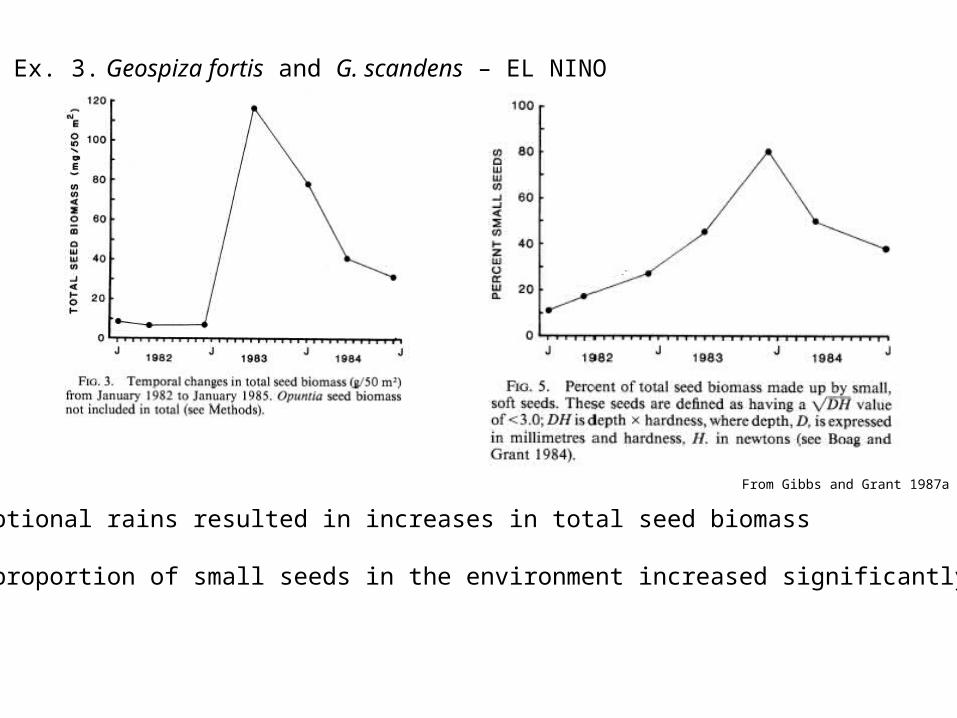
Ex. 3. Geospiza fortis and G. scandens – EL NINO
Exceptional rains resulted in increases in total seed biomass
The proportion of small seeds in the environment increased significantly
From Gibbs and Grant 1987a
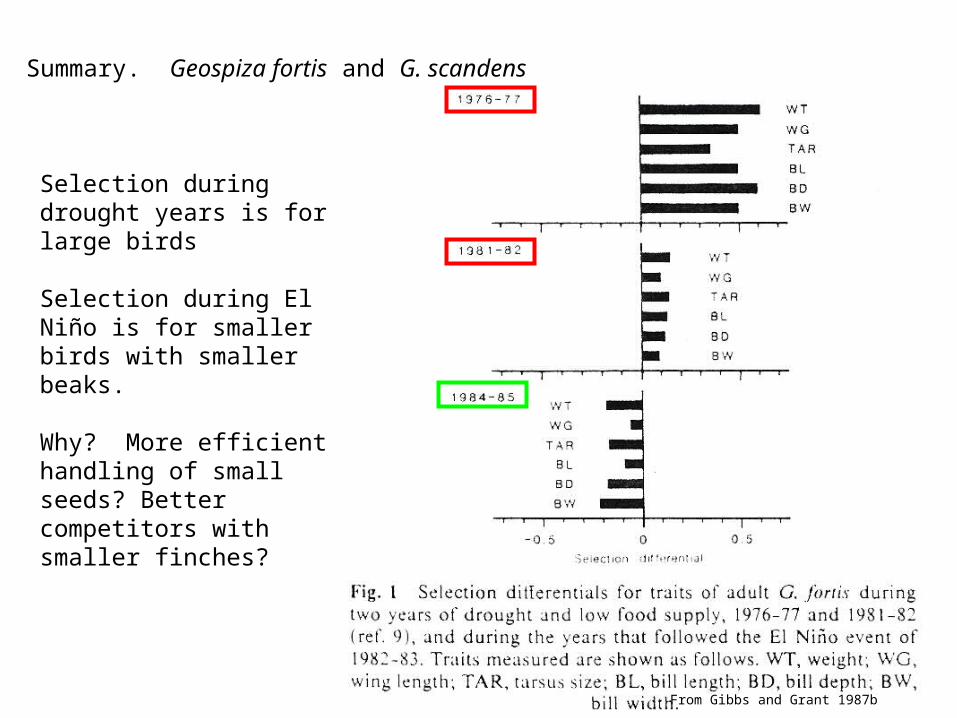
Summary. Geospiza fortis and G. scandens
Selection during drought years is for large birds
Selection during El Niño is for smaller birds with smaller beaks.
Why? More efficient handling of small seeds? Better competitors with smaller finches?
From Gibbs and Grant 1987b

Conclusions:
Behavioral shifts following a disturbance event can have effects on multiple trophic levels
Individuals with phenotypic traits far to one end of the population distribution may be favored following a disturbance
Climactic disturbances, such as droughts and El Nino, can exert strong selection pressure on populations.

References• Boag, P. T. and P. R. Grant. 1981. Intense natural selection in a population of Darwin’s finches. Science 214: 82-85.• Ebeling, A. W., D. R. Laur, and R. J. Rowley. 1985. Severe storm disturbances and
reversal of community structure in a southern California kelp forest. Marine Biology 84: 287-294.• Grant, B. R. 1985. Selection in bill characters in a population of Darwin’s finches: Geospiza conirostris on Isla Genovesa, Galapagos. Evolution 39(3): 523- 532.• Grant, B. R. and P. T. Grant. 1989. Natural selection in a population of Darwin’s finches. American Naturalist 133(3): 377-393.• Grant, P. R. 1986. Ecology and Evolution of Darwin’s Finches. Princeton University Press, Princeton, NJ.• Grant, P.R., B. R. Grant, L. F. Keller, and K. Petren. 2000. Effects of El Nino events on Darwin’s finch productivity. Ecology 81(9): 2442-2457.• Gibbs H. L. and P. R. Grant. 1987. Ecological consequences of an exceptionally strong El Nino event on Darwin’s finches. Ecology 68(6): 1735-1746.• Gibbs H. L. and P. R. Grant. 1987b. Oscillating selection on Darwin’s finches. Nature 327: 511-513.• Price, T. D., P. R. Grant, H. L. Gibbs, and P. T. Boag. 1984. Recurrent patterns of natural selection in a population of Darwin’s finches. Nature 309: 787- 789.• Stouder, D. J. 1987. Effects of a severe- weather disturbance on foraging patterns within a California surfperch guild. Journal of Experimental Biology and Ecology 114: 73-84.17

Complex Interactions: Aquatic Ecosystems - General
Pringle and Hamazaki, 1997
Effects of Fishes on Algal Response to Storms in a Tropical Stream
or“…how trophic factors interact with disturbance to affect
community response.”

Complex Interactions: Aquatic Ecosystems - General
Pringle and Hamazaki, 1997
Methods
• Colonization tiles – For algae, macroinvertebrates– Fish exclosures (electric!) vs. controls
• Sampled tiles for algae, macroinvertebrates• Three large storms during the experimental
period– On days 10, 26, and 39-40
• ANOVA

Complex Interactions: Aquatic Ecosystems - General
Pringle and Hamazaki, 1997
Results 1: Algal Biovolume
“Fig. 2”
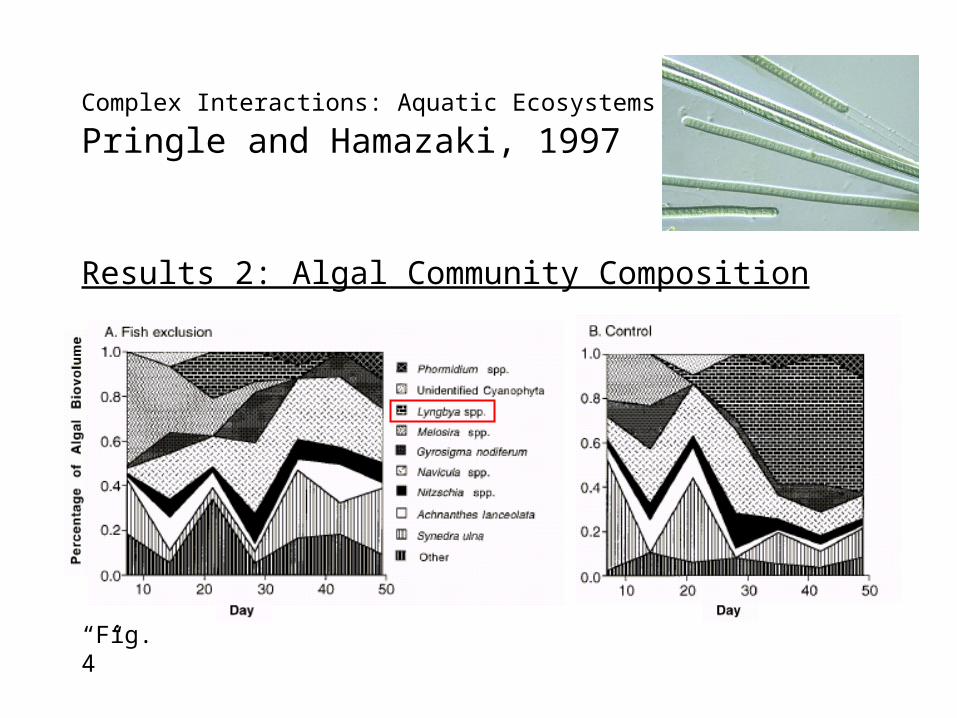
Complex Interactions: Aquatic Ecosystems - General
Pringle and Hamazaki, 1997
Results 2: Algal Community Composition
“Fig. 4”

Complex Interactions: Aquatic Ecosystems - General
Pringle and Hamazaki, 1997
Results 3: Percent Change in Ecosystem Parameters
“Table 4”

Complex Interactions: Aquatic Ecosystems - General
Pringle and Hamazaki, 1997
Discussion• Fish present: diatoms cyanobacteria
• Storms: regularly remove diatoms
• No trophic cascade– Omnivory in fishes…. Or regular storms…
• “Omnivorous fishes play a key role in maintaining stability of benthic algal communities and their resistance to
hydrologic disturbance”

Complex Interactions: Aquatic Ecosystems - Pollutants
Johnston and Keough, 2003
Competition Modifies the Response of Organisms to Toxic Disturbance
orAscidians vs. Serpulids:
Toxic and Indirect
Effects of Copper

Complex Interactions: Aquatic Ecosystems - Pollutants
Johnston and Keough, 2003
Methods
• 2 Experiments– Frequency and Intensity: dose toxicant
• 2 doses, 2 frequencies
– Space: dose toxicant + remove competitors
• Sampled for density of Ascidians and Serpulids
• ANOVA

Complex Interactions: Aquatic Ecosystems - Pollutants
Johnston and Keough, 2003
Results 1: Ascidians reduced
from
“Fig. 2”

Complex Interactions: Aquatic Ecosystems - Pollutants
Johnston and Keough, 2003
Results 2:
Most Serpulids increased
from
“Fig. 3”

Complex Interactions: Aquatic Ecosystems - Pollutants
Johnston and Keough, 2003
Results 3: Space
from
“Fig. 4”

Complex Interactions: Aquatic Ecosystems - Pollutants
Johnston and Keough, 2003
Results Summary

Complex Interactions: Aquatic Ecosystems - Pollutants
Johnston and Keough, 2003
Discussion
• Frequency substitutes for intensity
• Copper effects on serpulids: an integration of– Indirect effects: releases space
– Direct effects: toxicity (species-specific)
• “Good Competitor vs. Good Disperser” tradeoff– Episodic pollution favors …

Complex Interactions: Aquatic Ecosystems – More Pollutants
Courtney and Clements, 2000
Sensitivity to acidic pH in benthic invertebrate assemblages with different histories of exposure
to metals
• A microcosm experiment• Interaction between 2 disturbance forces (pollutants) that
are sequential in time• Previous exposure to metals increases sensitivity to low pH• A species-level response scales up to assemblage level

Complex Interactions – Aquatic EcosystemsReferences
• Courtney, L. A., and W. H. Clements. 2000. Sensitivity to acidic pH in benthic invertebrate assemblages with different histories of exposure to metals. J. N. Am. Benthol. Soc. 19(1): 112-127
• Johnston, E. L., and M. J. Keough. 2003. Competition modifies the response of organisms to toxic disturbance. Mar. Ecol. Prog. Ser. 251: 15-26
• Pringle, C. M., and T. Hamazaki. 1997. Effects of fishes on algal response to storms in a tropical stream. Ecology 78(8): 2432-2442
• Salminen, J., B. T. Anh, and C. A. M. Van Gestel. 2001. Indirect effects of zinc on soil microbes via a keystone Enchytraeid species. Env. Toxicology and Chemistry 20(6): 1167-1174
• Morrison, H. A., D. M. Whittle, and G. D. Haffner. 2000. The relative importance of species invasions and sediment disturbance in regulating chemical dynamics in western Lake Erie. Ecological Modelling 125: 279-294





























