Embed Size (px)
Citation preview

INSTITUTO POLITÉCNICO NACIONAL
CENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS
DIVERSIDAD PARASITARIA DE Octopus
hubbsorum (BERRY, 1953) EN LA COSTA DE
BAJA CALIFORNIA SUR, MÉXICO.
TESIS QUE PARA OBTENER EL GRADO DE
MAESTRÍA EN CIENCIAS
EN
MANEJO DE RECURSOS MARINOS
PRESENTA
ELISA TÉLLEZ CHÁVEZ
LA PAZ, B.C.S., JUNIO DE 2016



DEDICATORIA
A MIS PADRES, POR LA VIDA, POR NUNCA DEJAR DE CREER EN MI, POR TODO SU INFINITO
APOYO Y AMOR.
A MIS QUERIDOS HERMANOS, MIS AMIGOS DE TODA LA VIDA, PORQUE DE USTEDES Y CON
USTEDES HE APRENDIDO TANTO.
A LOS QUE YA NO ESTÁN.
A TI, QUIEN NUNCA ME DEJÓ TIRAR LA TOALLA, QUIEN ME TOMÓ DE LA MANO HASTA EL FINAL
PORQUE POR USTEDES VALIÓ LA PENA . . .

AGRADECIMIENTOS Principalmente a mi familia, mis padres Alfonso Téllez Fuentes y Epifania Chávez
Suárez, mis hermanos JuanPablo y César Leonardo, por sacrificarse a extrañarme
día a día. A mi tía Gloria, tío Dago y mis primos Joel y Andy.
A mi codirectora, por toda su paciencia, apoyo y guía durante la identificación de
parásitos, gracias por hacerlo interesante. A mi director, por las salidas amenas en
campo, las oportunidades profesionales y sus valiosos comentarios. A mi comité
tutorial Dr. Andrés Abitia Cárdenas, Dr. Agustín Hernádez Herrera, M.C. Marcial
Trinidad Villalejo Fuerte y Dra. Patricia Ceballos Vázquez por sus valiosas
observaciones y consejos en el manuscrito.
A todos mis queridos compañeros de generación (winnies pa la banda): Juan Carlos,
Diego, Ania, Juanita, Rubén (chico plancton), Iván, Paquito, Kenny, Ilse, Noemí,
Rafa, Sharon, Bianca, Elena y Karen Pabon por haber compartido tantas
experiencias y aunque reunirnos era un desmadre, nunca faltó el pretexto de un
pastel de cumpleaños, un pozole, una ida a la playa, bienvenida o una posada para
juntarnos. A mis queridos amigos del instituto Yaraomir (Yaro) y Antonino (manito),
por las experiencias y siempre tenr un oído para cualquier tema y siempre sacarme
una sonrisa. Nefertiti por tu apoyo incondicional en campo y amistad. Cynthia por la
buena charla durante el trabaj en el lab.
A mis fieles amigos que de lejos nunca dejaron de creer en mi, Chuy, Gaby, Giovis,
Alex, Gera, Octavio, Daniel, Alfonso (Costeño), Dalia, Greben, Roger y que siempre
me demostraron su apoyo con más de un mensaje.
A ti Juan Carlos Castro Salgado, que fuiste parte de esta gran aventura, experiencia
profesional y personal, por todo tu cariño, tu fuerza, paciencia mucha paciencia y por
ser ese amigo incondicional. Gracias por todo y más, porque sin tu motivación y
palabras de aliento no se como lo habría conseguido. Gracias por dejarme compartir
el otoño a tu lado. Espero vengan muchos más.

A la B.M. Alma Rivera Camacho por todos tus consejos, aprendizaje y amistad. Al Dr.
Francisco Domínguez-Contreras por sus comentarios y orientación en el desarrollo
profesional. Así mismo, a los pescadores Martín Osuna de Santa Rosalía y a la
Sociedad Cooperativa de Pescadores de Bahía Magdalena. A Humberto Ceseña
Amador y César Casas Núñez por su valioso tiempo y paciencia en todos los
trámites desde el inicio del posgrado. A los administrativos de CICIMAR. A Blanca de
Jesús Carball Carballo, por su amable atención relacionado con trámites
económicos.
A los que sin formar parte del proyecto nunca me negaron su apoyo ante cualquier
duda: Dr. Rodrigo Moncayo Estrada, por su asesoría en los análisis estadísticos y
sabios consejos; Lic. Armando Hernández López, por la asesoría en la edición de
imágenes; al Dr. Mauricio Ramírez Rodríguez, por su orientación en la definición del
área de muestreo; Dr. Carlos Cedillo Peláez (INP) por sus consejos y apoyo en el
análisis histológico; Farallon Broughton (Santa Barbara, CA) por su oreintación en la
identificación de protozoarios. A todos mis profesores de clase de la maestría,
gracias por sus enseñanzas, especialmente al Dr. Arturo Tripp Quezada, por
permitirme conocer las maravillas del mar, hizo que la meta se cumpliera gracias por
la travesía.
Al Centro Interdisciplinario de Ciencias Marinas y al Instituto Politécnico
Nacional por ser parte de mi formación académica y profesional, así como al Consejo
Nacional de Ciencia y Tecnología, a la Comisión de Operación y Fomento de
Actividades Académicas del I.P.N., a la Beca de Estímulo Institucional de Formación
de Investigadores por los apoyos económicos y académicos para realizar el trabajo
de tesis.

ÍNDICE LISTA DE FIGURAS .................................................................................................... I
LISTA DE TABLAS ..................................................................................................... III
GLOSARIO DE TÉRMINOS ....................................................................................... IV
RESUMEN ................................................................................................................ VII
ABSTRACT .............................................................................................................. VIII
1. INTRODUCCIÓN .................................................................................................... 1
2. ANTECEDENTES ................................................................................................... 3
2.1 Parasitismo ........................................................................................................ 3
2.2 Parásitos en cefalópodos ................................................................................... 4
2.3 Mecanismos de defensa ante agentes parasitarios ........................................... 7
2.4 Parásitos en cefalópodos distribuidos en México ............................................... 7
2.5 Octopus hubbsorum ........................................................................................... 8
2.5.1 Biología general ........................................................................................... 8
2.5.2 Distribución .................................................................................................. 8
2.6 Importancia del pulpo como recurso pesquero .................................................. 9
3. JUSTIFICACIÓN ................................................................................................... 10
4. OBJETIVOS .......................................................................................................... 11
4.1 Objetivo general ............................................................................................... 11
4.2 Objetivos específicos ....................................................................................... 11
5. MATERIAL Y MÉTODOS ...................................................................................... 12
5.1 Áreas de estudio .............................................................................................. 12
5.2 Obtención de hospederos ................................................................................ 13
5.3 Búsqueda de parásitos .................................................................................... 14

5.3.1 Toma de muestras en campo .................................................................... 14
5.3.2 Toma de muestras en el laboratorio .......................................................... 14
5.4 Observación e identificación de parásitos ........................................................ 15
5.4.1 Tratamiento de nemátodos ........................................................................ 15
5.4.2 Identificación taxonómica de nemátodos ................................................... 15
5.4.3 Análisis histológico ..................................................................................... 16
5.4.4 Identificación taxonómica de protozoarios ................................................. 16
5.5 Prevalencia, intensidad media y abundancia ................................................... 17
5.6 Análisis estadísticos ......................................................................................... 17
6. RESULTADOS ...................................................................................................... 19
6.1 Análisis biométricos ......................................................................................... 19
6.2 Análisis histológico de protozoarios ................................................................. 21
6.2.1 Descripción de protozoarios ...................................................................... 21
6.2.2 Resumen taxonómico ................................................................................ 26
6.2.3 Observaciones del género Aggregata ........................................................ 26
6.3 Análisis microscópico de nemátodos ............................................................... 28
6.3.1 Descripción de nemátodos ........................................................................ 28
6.3.2 Resumen taxonómico ................................................................................ 33
6.3.3 Observaciones del género Hysterothylacium ............................................. 33
6.4 Análisis del hábitat de parásitos ....................................................................... 35
6.4.1 Preferencia de hábitat de protozoarios ...................................................... 35
6.4.2 Preferencia de hábitat de nemátodos ........................................................ 39
6.5 Prevalencia, intensidad media y abundancia ................................................... 40
6.6 Relación de tallas del hospedero con el número de parásitos ......................... 41
6.7 Lesiones al hospedero ..................................................................................... 42

6.7.1 Lesiones por protozoarios .......................................................................... 42
6.6.2 Lesiones por nemátodos ............................................................................ 46
7. DISCUSIÓN .......................................................................................................... 47
8. CONCLUSIONES .................................................................................................. 54
9. RECOMENDACIONES ......................................................................................... 55
10. LITERATURA CITADA ........................................................................................ 56
11. ANEXO ................................................................................................................ 64

I
LISTA DE FIGURAS Figura Página
1 Esquema del ciclo de vida directo e indirecto de un parásito……….. 4
2 Esquema del ciclo de vida del protozoario Aggregata……………….. 5
3 Serie histórica de la producción de pulpo en México………………... 9
4 Áreas de captura de Octopus hubbsorum…………………………….. 13
5 Presencia de ooquistes de Aggregata en Octopus hubbsorum……. 21
6 Microgamontes de Aggregata sp. en Octopus hubbsorum………….. 23
7 Macrogamontes de Aggregata sp. en Octopus hubbsorum……….... 24
8 Formación del cigoto y esporonte de Aggregata sp. en Octopus
hubbsorum…………………………………………………………………
24
9 Desarrollo de esporoblastos de Aggregata sp. en Octopus
hubbsorum…………………………………………………………………
25
10 Formación de esporozoitos de Aggregata sp. en Octopus
hubbsorum…………………………………………………………………
25
11 Extremo anterior de Hysterothylacium sp……………………………... 30
12 Extremo anterior de Hysterothylacium sp. larva L3 de Octopus
hubbsorum…………………………………………………………………
31
13 Esófago y apéndices de Hysterothylacium sp. larva L3 de Octopus
hubbsorum……………………………………………………..................
32
14 Esófago y apéndices de Hysterothylacium sp. larva L3 de Octopus
hubbsorum…………………………………………………………………
33
15 Número de quistes en los órganos más infectados en Octopus
hubbsorum, por mes de muestreo en la región de Santa Rosalía….
36
16 Número de quistes en los órganos más infectados en Octopus
hubbsorum, por zona de muestreo……………………………………..
37
17 Dendograma construido a partir del análisis de cluster para mostrar
la preferencia de hábitat de protozoarios en los órganos de Octopus
hubbsorum……………………………………………………..................
38
18 Número de nemátodos en Octopus hubbsorum, por zonas de
captura……………………………………………………………………...
39

II
19 Presencia de nemátodos en el intestino de Octopus hubbsorum…... 39
20 Relación del tamaño del hospedero y el número de quistes en
pulpos de Bahía Magdalena……………………………………………..
41
21 Relación del tamaño del hospedero y el número de quistes en
pulpos de la región de Santa Rosalía…………………………………..
42
22 Lesiones macroscópicas por Aggregata sp. sobre diversos órganos
de Octopus hubbsorum…………………………………………………..
42
23 Localización de quistes de Aggregata sp. en la región de la mucosa
y serosa del ciego de Octopus hubbsorum…………………………….
43 24 Efecto de quistes de Aggregata sp. en el tejido infectado de
Octopus hubbsorum………………………………………………………
44
25 Evidencia de mecanismos de defensa de Octopus hubbsorum en
presencia de quistes de Aggregata sp………………………………….
44
26 Evidencia de daño por protozoarios en el buche y ciego de Octopus
hubbsorum…………………………………………………………………
45
27 Evidencia histológica del manto de Octopus hubbsorum infectado
por Aggregata sp………………………………………………………….
45
28 Nemátodo atravesando el intestino de Octopus hubbsorum………… 46

III
LISTA DE TABLAS Tabla Página
1 Comparación de las variables biométricas de Octopus hubbsorum,
por mes de muestreo en Santa Rosalía………………………………..
19
2 Comparación de las variables biométricas de Octopus hubbsorum,
capturados en febrero en ambas áreas de estudio…………………...
20
3 Medidas de los caracteres morfológicos de la larva L3 de
Hysterothylacium sp. encontrados en Octopus hubbsorum……........
29
4 Número de quistes en los órganos poco parasitados de Octopus
hubbsorum, por zona y mes de muestreo………………………………
38
5 Datos totales de prevalencia, intensidad media y abundancia de
quistes de Aggregata sp. en Octopus hubbsorum, por mes de
muestreo……………………………………………………………............
40
6 Datos totales de prevalencias, intensidad media y abundancia del
nemátodo Hysterothylacium sp. en Octopus hubbsorum, por mes
de muestreo………………………………………………………..............
41

IV
GLOSARIO DE TÉRMINOS Coccidios. Protozoos intracelulares de importancia médica ubicados en el Phylum
Aplicomplexa. Parásitos de intestino y otros órganos y sistemas (Universidad
Nacional Autónoma de México, 2011. http://www.facmed.unam.mx/deptos/microbiolo
gia/glosario.html).
Diciémido. Orden de invertebrados parásitos del Phylum Mesozoa. Son animales
pequeños, con aspecto vermiforme, que viven como endoparásitos de moluscos
cefalópodos generalmente en los órganos renales de la sepia y del pulpo (Micronet,
2016). Endoparásito. Parásito cuyo hábitat se encuentra dentro del cuerpo de su
hospedero (Universidad Nacional Autónoma de México, 2011.
http://www.facmed.unam.mx/deptos/microbiologia/glosario.html).
Esporoquiste. En algunos protozoos del Phylum Apicomplexa, forma de resistencia
que contiene esporozoitos y se encuentra dentro del ooquiste (Universidad Nacional
Autónoma de México, 2011. http://www.facmed.unam.mx/deptos/microbiologia/glosar
io.html).
Esporozoíto. Organismo delgado, fusiforme, móvil, haploide, resultante de la
esporogonia en algunos protozoos del Phylum Apicomplexa. Forma invasiva de
algunos organismos del Phylum Apicomplexa (Universidad Nacional Autónoma de
México, 2011. http://www.facmed.unam.mx/deptos/microbiologia/glosario.html).
Heteroxeno. Parásito que requiere dos o más hospederos para completar su ciclo de
vida (Universidad Nacional Autónoma de México, 2011. http://www.facmed.unam.mx/
deptos/microbiologia/glosario.html).
Hospedero definitivo o final. Organismo que aloja las formas adultas o
sexualmente maduras del parásito (Universidad Nacional Autónoma de México,
2011. http://www.facmed.unam.mx/deptos/microbiologia/glosario.html).
Hospedero intermediario. Organismo que aloja las formas larvarias, asexuales, o
inmaduras del parásito (Universidad Nacional Autónoma de México, 2011.
http://www.facmed.unam.mx/deptos/microbiologia/glosario.html).

V
Hospedero de transferencia o paraténico. Hospedero facultativo, innecesario para
que el agente patógeno completa su ciclo vital (Universidad Nacional Autónoma de
México, 2011. http://www.facmed.unam.mx/deptos/microbiologia/glosario.html).
Infección. Colonización y multiplicación de agentes patógenos en un organismo.
Puede o no dar lugar a una enfermedad manifiesta (Universidad Nacional Autónoma
de México, 2011. http://www.facmed.unam.mx/deptos/microbiologia/glosario.html).
Metazoario. Organismos pluricelulares del reino Animalia, subreino Metazoa, con
sus células organizadas para formar tejidos; incluye a todos los animales, excepto los
protozoarios (Cruz-Reyes & Camargo-Camargo, 2001).
Metacíclico. Formas infestantes de un parásito protozoario (Gállego-Berenguer,
2006).
Nemátodo. Gusano cilíndrico no segmentado, con simetría bilateral, cavidad
celómica y tubo digestivo completo, dioico (Universidad Nacional Autónoma de
México, 2011. http://www.facmed.unam.mx/deptos/microbiologia/glosario.html).
Ontogenia. Secuencia de fases fisiológicas y morfológicas que los organismos
experimentan a medida que avanzan sus ciclos vitales. Historia del desarrollo de un
organismo individual desde su origen hasta su muerte (Universidad Nacional
Autónoma de México, 2011. http://www.facmed.unam.mx/deptos/microbiologia/
glosario.html).
Ooquiste. Forma quística resultante de la esporogonia en los Apicomplexa, que
contiene esporozoítos, los cuales pueden estar cubiertos adicionalmente por una
envoltura, denominada esporoquiste o estar desnudos (Universidad Nacional
Autónoma de México, 2011. http://www.facmed.unam.mx/deptos/microbiologia/
glosario.html).
Parasitismo. Estado de simbiosis en el cual el simbionte se beneficia de la
asociación, y el hospedador es dañado de manera “metabólica” y/o “ecológica”
(Cruz-Reyes & Camargo-Camargo, 2001).
Parásito. Organismo que vive en o sobre un hospedero, adquiere de él sus
nutrientes durante una parte o toda su vida. Causa daño de diferente grado

VI
(Universidad Nacional Autónoma de México, 2011. http://www.facmed.unam.mx/
deptos/microbiologia/glosario.html).
Patógeno. Organismo capaz de producir enfermedad (Universidad Nacional
Autónoma de México, 2011. http://www.facmed.unam.mx/deptos/microbiologia/
glosario.html).
Potencial zoonótico. Es la capacidad de transmitir de manera natural aquellas
enfermedades e infecciones entre animales y el hombre (Gállego-Berenguer, 2006).
Prevalencia. El número de hospederos infectados por 1 ó más individuos de una
especie parásita dividido por el número de hospederos examinados para dicha
especie (Universidad Nacional Autónoma de México, 2011. http://www.facmed.unam.
mx/deptos/microbiologia/glosario.html).
Protozoarios. Organismos unicelulares eucarióticos con uno o más núcleos del
subreino Protozoa; cada célula realiza las funciones necesarias de metabolismo y
reproducción para vivir (Cruz-Reyes & Camargo-Camargo, 2001).
Quiste. Organismo (u organismos) encapsulado o cubiertos por una capa protectora,
en ocasiones en estadio latente, se forma en respuesta a condiciones ambientales
adversas (Cruz-Reyes & Camargo-Camargo, 2001).

VII
RESUMEN El conocimiento de la carga parasitaria en pulpos permite comprender el
impacto económico de las enfermedades que los parásitos pueden causar al
hospedero, ya sea reduciendo poblaciones o la calidad del producto, y el de su
posible efecto en el humano. Octopus hubbsorum es la principal especie de pulpo
capturada en Baja California Sur, y la información acerca de su carga parasitaria y su
papel como hospedero es poco conocido. En este estudio se analizó la diversidad
parasitaria de O. hubbsorum de Bahía Magdalena (BM) y de la región de Santa
Rosalía (SR) B.C.S., México. Se identificaron las principales características
morfológicas de los parásitos, así como su estadio ontogénico y se determinó y
comparó la prevalencia, intensidad media y abundancia, así como la evidencia de
daño del parásito a su hospedero. El análisis histológico permitió identificar diversos
estadios del ciclo de vida de protozoarios como micro y macrogamontes
representativos de la fase gamogónica, así como esporontes, esporoblastos y
esporozoitos pertenecientes a la fase esporogónica, tales características son
específicas del género Aggregata. El análisis microscópico permitió identificar larvas
de nemátodos en estadio L3 pertenecientes al género Hysterothylacium, por la
presencia de un ciego intestinal y un apéndice ventricular, así como del poro excretor
localizado inmediatamente después del anillo nervioso. Se observó una preferencia
de hábitat de los protozoarios en cada zona de captura, donde el buche fue el órgano
más parasitado en los pulpos de BM y el ciego en los de la región de SR, mientras
que los nemátodos siempre se localizaron en el intestino del hospedero. La
prevalencia total de protozoarios fue del 90%, aunque la intensidad media y la
abundancia fue mayor en los organismos de BM. La prevalencia total de nemátodos
fue del 7%, y en BM se registraron los valores máximos de prevalencia, intensidad
media y abundancia. Finalmente, mediante el análisis histológico también se
observaron evidencias de ruptura del tejido por la presencia de ambos parásitos, así
como alteración en la estructura celular de los órganos más parasitados por
protozoarios. En este trabajo se demuestra que el pulpo O. hubbsorum se
desempeña como hospedero definitivo para protozoarios y como hospedero
intermediario para nemátodos, ambos descritos por primera vez en esta especie.

VIII
ABSTRACT The knowledge of the parasites in octopuses allows to comprise the economic impact
of diseases in the host, reducing populations or the product quality, and its posible
effect on the human. Octopus hubbsorum is the main species of octopus captured in
Baja California Sur, and the information about their parasites and its role as host is
little known. In this study, we analyzed the parasitic diversity of O. hubbsorum of
Bahía Magdalena (BM) and Santa Rosalía (SR) region B.C.S., Mexico. the main
morphological characteristics of the parasites and their ontogenetic stage were
identified and we determined and compared the prevalence, mean intensity and
abundance, as well as evidence of damage in the host by the parasite. Whit
histological analysis we identified various stages of the life cycle of protozoan as
micro and macrogamonts, of the gamogonic phase, and sporonts, sporoblasts and
sporozoites belonging to the esporogonic phase, such features are specific of
Aggregata genus. Microscopic analysis identified nematode larvae in L3 stage
belonging to the genus Hysterothylacium, by the presence of a caecum intestinal and
atrial appendage, and the excretory pore located immediately after the nerve ring. A
preference habitat protozoans in each capture zone was observed, where the crop
was the most parasitized organ in octopuses of BM and the caecum in the octopuses
of SR region, while nematodes always located in the host intestine. The prevalence of
protozoan was 90%, although the mean intensity and abundance was higher in the
octopuses of Bahía Magdalena. The prevalence of nematodes was 7%, and the
maximum prevalence, mean intensity and abundance were in BM. Finally, by
histological analysis also evidence tissue breakdown were observed by the presence
of both parasites and alteration in the cellular structure of the organs parasitized by
protozoa. This paper shows that the octopus O. hubbsorum serves as the definitive
host for protozoans and as intermediate host for nematodes, both described for the
first time in this species.

1
1. INTRODUCCIÓN Los parásitos representan más de la mitad de todas las especies del planeta
(González et al., 2003; Rhode, 2005). En la actualidad, el conocimiento de la
parasitología en moluscos como los cefalópodos aún se considera escaso
(Castellanos-Martínez & Gestal, 2013), sin embargo, su importancia económica en la
pesca y acuicultura ha favorecido el interés por conocer agentes zoonóticos
patógenos y entender las relaciones parásito hospedero (González et al., 2003;
Castellanos-Martínez & Gestal, 2013); debido a las enfermedades que causan en
otros animales (Estévez et al., 1998). En ocasiones, las enfermedades infecciosas
pueden alterar el sistema inmune del hospedador y facilitar la multiplicación de
parásitos que en condiciones normales estaría controlada; al mismo tiempo, la
presencia de organismos parasitarios puede favorecer la infección del hospedador
por agentes infecciosos similares o de otro tipo (Gállego-Berenguer, 2006).
Hace 20 años, se estimó que alrededor de 150 especies de protozoarios y
metazoarios parasitan alrededor de 650 especies de cefalópodos (Pascual et al.,
1996), de los cuales, algunos nemátodos provocan enfermedades en el humano al
ser consumidos (Audicana & Kennedy, 2008; Food and Agriculture Organization,
2014). Dentro del ecosistema marino, los pulpos participan como hospederos
definitivos en el ciclo de vida de algunos parásitos heteroxenos (Pascual et al., 1997),
tal es el caso de coccidios del género Aggregata (Gestal et al., 2002a), los cuales no
generan un daño letal en los cefalópodos como tal, sino que esto los hace
vulnerables ante factores bióticos o abióticos de estrés (Castellanos-Martínez &
Gestal, 2013), llegando a afectar el potencial pesquero de este recurso (González et
al., 2003).
La importancia económica en la pesca y acuacultura de pulpos en México, ha
favorecido el interés por conocer su fauna parasitaria, así como entender sus
relaciones parásito-hospedero (González et al., 2003). Sin embargo, estos estudios
hasta ahora se han centrado en la descripción y definición taxonómica de las
especies (Castellanos-Martínez et al., 2011). En Baja California Sur, Octopus
hubbsorum es una de las principales especies de pulpo capturada (López-Uriarte et
al., 2005) y para la cual la información acerca de su diversidad parasitaria es escasa,

2
por lo que hasta ahora sólo se ha descrito una especie de diciémido para la Bahía de
La Paz (Castellanos-Martínez et al., 2011).
Por lo anterior, la necesidad por conocer la carga parasitaria en O.
hubbsorum, no sólo radica en ampliar el conocimiento de la biología y ecología de
esta especie, sino que permitirá comprender mejor las relaciones interespecíficas
con otros organismos, para que se puedan generar estrategias ante patógenos con
impacto humano, ya sea en la salud o económicamente. Evidentemente, no es
posible comprender a fondo la magnitud del problema para los hospederos sin antes
conocer los tipos de patógenos parásitos, así como el sitio que habitan dentro del
hospedero, lo cual es importante para definir el grado de infección e identificar
especies. Con base a lo anterior, la presente investigación pretende ampliar el
conocimiento de la carga parasitaria en O. hubbsorum, así también proporcionar una
breve descripción del daño tisular en los órganos principalmente dañados.
Este trabajo registra por primera vez parásitos protozoarios y nemátodos en O.
hubbsorum. En los resultados se señalan las características morfológicas de los
parásitos utilizando técnicas histológicas y de microscopía compuesta, con el fin de
reconocer el estadio ontogénico, el papel del hospedero, y asimismo su identificación
taxonómica a partir de claves especializadas. Por otro lado, se identifican los
principales órganos infectados y se determinan la prevalencia, intensidad media y
abundancia, mediante el conteo de nemátodos y quistes, para analizar diferencias
entre zonas de captura y su relación con la talla de los hospederos. Finalmente, se
señalan lesiones en el tejido del hospedero, empleando nuevamente técnicas
histológicas y de microscopía compuesta, para tener un conocimiento previo del
impacto de parásitos en O. hubbsorum.

3
2. ANTECEDENTES En México, el estudio de pulpos se enfoca principalmente en su importancia
ecológica y biológica, a pesar de esto, el conocimiento de su parasitismo y las
relaciones parásito – hospedero, así como las enfermedades que le generan al
pulpo, han sido poco estudiadas. A pesar de la importancia comercial del pulpo O.
hubbsorum en Baja California Sur, esta información aún es limitada.
2.1 Parasitismo
De manera natural, los seres vivos establecen asociaciones interespecíficas
durante una parte o la totalidad de sus vidas (Gállego-Berenguer, 2006), tal es el
caso del parasitismo, una asociación constituida por hospedador y parásito, donde el
segundo ocupa al hospedero como su hábitat y se alimenta de sus tejidos o
sustancias nutricias, haciendo que resulte o no perjudicado (Rhode, 2005; Gállego-
Berenguer, 2006).
Durante su ciclo de vida, en los parásitos pluricelulares se identifican estadios
adultos y juveniles, mientras que en los organismos unicelulares se reconocen
formas sexuadas o asexuadas, esta separación permite distinguir el papel del
hospedero en definitivo o intermediario (Fig. 1A) (Gállego-Berenguer, 2006). El
hospedero definitivo es indispensable para la culminación del ciclo del parásito y es
el único que participa cuando dicho ciclo es directo, mientras que aquellos
organismos que tienen un ciclo de vida indirecto, requieren de uno o más hospederos
intermediarios que son utilizados por las formas juveniles o asexuales (Gállego-
Berenguer, 2006). Por otro lado, existen hospederos paraténicos (Fig. 1B), los cuales
no son indispensables para el desarrollo del parásito pero facilitan su llegada hacia el
hospedero definitivo (Gállego-Berenguer, 2006).

4
Figura 1. Esquema del ciclo de vida directo (A) e indirecto (B) de un parásito. En ocasiones, las fases juvenil o asexual ocurren en vida libre. 2.2 Parásitos en cefalópodos
Los cefalópodos, tienen un papel importante durante el ciclo de vida de
algunos parásitos, pues son importantes como hospederos definitivos para
protozoarios, diciémidos y crustáceos, y al mismo tiempo sirven como hospederos
intermediarios a larvas de digéneos, céstodos, acantocéfalos y nemátodos (Kinne,
1990). Kinne (1990) y Pascual et al. (1996) mencionaron que alrededor de 150
especies de organismos protistas y metazoarios son capaces de parasitar al menos
650 especies de cefalópodos, pero en los últimos 20 años estos datos no se han
actualizado. Entre los agentes patógenos más comunes se encuentran las bacterias,
protozoarios, mesozoarios, céstodos, tremátodos, nemátodos y crustáceos (Forsythe
et al., 1991; Castellanos-Martínez & Gestal, 2013).
Los principales agentes bacterianos descritos en cefalópodos, pertenecen al
género Vibrio, y generalmente aparecen en condiciones de cultivo tanto en piel como
en ojos (Kinne, 1990). Algunas especies donde se han registrado estos vibrios son
O. vulgaris (Fichi et al., 2015), O. bimaculoides, O. joubini y O. briareus, así como en
la sepia Seppia officinalis (Kinne, 1990).
Dentro del grupo de protozoarios, los apicomplexos del género Aggregata son
parásitos generalmente localizados en la cutícula del tracto digestivo, en branquias y
en el manto de cefalópodos bentónicos. Los cefalópodos parasitados por este

5
género, participan como hospederos definitivos (Fig. 2), pues en estos se desarrollan
las fases de gamognia (gamogónica o sexual) y esporogonia (esporogónica)
(Pascual et al., 1996), mientras que la fase de Merogonia (agomogónica o asexual)
se desarrolla dentro de crustáceos, (Castellanos-Martínez & Gestal, 2013), dichas
características permiten diferenciar entre especies de acuerdo a las diferencias
morfológicas en su último hospedero (Estévez et al., 1998). Se han descrito especies
como A. eberthi en S. officinalis, A. octopiana en O. vulgaris, entre otras especies en
Todaropsis eblanae y Todarodes sagittatus (Pascual et al., 1996).
Figura 2. Esquema del ciclo de vida del protozoario Aggregata. Modificado de
Gállego-Berenguer (2006).
Los mesozoarios diciémidos son parásitos con una amplia distribución a nivel
mundial que se alojan en los órganos excretores, generalmente dentro de los sacos
renales (nefronas) de cefalópodos bentónicos (Pascual et al., 1996; Castellanos-
Martínez et al., 2011) y epibentónicos (Kinne, 1990). Entre sus formas parasitarias se

6
pueden identificar estadios vermiformes maduros y embrionarios, así como
infusorígenos y larvas infusoriformes (Pascual et al., 1996). Se ha descrito una gran
diversidad de especies en sepias como S. officinalis y S. orbignyana, y en pulpos
como Bathypolypus sponsalis y O. vulgaris (Pascual et al., 1996).
En el caso de los Platelmintos, dentro del tracto digestivo de algunas especies
de calamares neríticos y oceánicos, se albergan comúnmente larvas plerocercoides
de Tetraphyllidea y metacéstodos de Trypanorhyncha (Kinne, 1990), estos últimos
también han sido localizados en sepias (Pascual et al., 1996). La presencia de dichas
larvas permite comprender mejor las interrelaciones tróficas en ambientes marinos
(Kinne, 1990; Pascual et al., 1996). Por otro lado, los cefalópodos participan como
hospederos paraténicos de tremátodos monogéneos; en loligínidos se han hallado
dentro de la cavidad del manto, vasos sanguíneos, branquias y brazos (Kinne, 1990).
Asimismo, los cefalópodos son capaces de albergar larvas de digéneos, participando
como el hospedero final o el segundo intermediario, ante el consumo de
invertebrados o peces (Kinne, 1990). Por otra parte, el conocimiento de nemátodos
en cefalópodos se resume a estadios larvarios, tal es el caso de espirúridos
encontrados en el estómago de O. vulgaris (Pascual et al., 1996). Referente a los
ascaroideos, Anisakis simplex es un parásito localizado en su fase larvaria 3, dentro
del tracto digestivo y gónadas principalmente de calamares como Loligo vulgaris,
Illex coindetii, T. eblanae y T. sagittatus (Pascual et al., 1996), estos nemátodos
llegan incidentalmente al cefalópodo facilitando el paso del parásito al ser humano,
mediante su consumo, llegando a provocar una zoonosis y enfermedades
gastrointestinales (Audicana & Kennedy, 2008; Food and Agriculture Organization,
2014).
En relación a crustáceos parásitos, se han registrado copépodos de
Octopicola en branquias del pulpo O. vulgaris (Pascual et al., 1996). También se han
hallado estadios postembrionarios de Pennella varians en las laminillas branquiales y
en la cavidad del manto de Eleodone moschata, S. officinalis, S. elegans, L. vulgaris
y T. eblanae (Pascual et al., 1996).

7
2.3 Mecanismos de defensa ante agentes parasitarios
El estudio de mecanismos ante agentes parasitarios en organismos marinos
requiere de especial atención ya sea por su potencial zoonótico o especialmente
para establecer medidas preventivas de enfermedades que pueden afectar su
producción (Carella et al., 2015).
Los cefalópodos, carecen de una respuesta inmune que les permita atacar a
agentes infecciosos como bacterias, virus y parásitos (Kinne, 1990); por lo tanto, no
adquieren memoria inmunológica (Castellanos-Martínez & Gestal, 2013). Se ha
observado, que los coccidios originan la ruptura de la membrana basal y de células
epiteliales, así como la disminución en la actividad enzimática y absorción de las
mismas (Gestal et al., 2002b).
Respecto a la presencia de diciémidos en cefalópodos, Kinne (1990)
mencionó que no se ha observado que estos organismos dañen de manera
considerable a su hospedero, mientras que Castellanos-Martínez & Gestal (2013)
indicaron que ayudan a eliminar el amonio de la orina, llegando a saturar los ductos
del animal en presencia de una sobrepoblación de individuos.
Por otro lado, los protozoarios del género Aggregata son parásitos capaces de
alterar la absorción de nutrientes en cefalópodos infectados (Poynton et al., 1992;
Gestal et al., 2002a; Gestal et al., 2002b). Generalmente infectan el tracto digestivo,
donde se origina el desprendimiento de las células digestivas y un daño a nivel
celular (Castellanos-Martínez & Gestal, 2013); sin embargo, no se les considera
como la principal causa de muerte en pulpos, pero sí son un factor que los vuelve
vulnerables ante otros agentes de estrés (Gállego-Berenguer, 2006; Storero &
Narvarte, 2013), pues una vez que dañan la epidermis se ha observado la presencia
de infecciones bacterianas en heridas (Forsythe et al., 1991; Castellanos-Martínez &
Gestal, 2013).
2.4 Parásitos en cefalópodos distribuidos en México
En México los registros de parásitos en cefalópodos están representados sólo
por mesozoarios en pulpos. El primer registro que se tiene es del diciémido Dicyema
shorti en el pulpo O. burryi, de Veracruz, Golfo de México, donde únicamente se

8
registran embriones vermiformes y nematógenos dentro de los pliegues de los sacos
renales, los demás estadios de su ciclo de vida no han sido descritos (Furuya et al.,
2002).
Por otro lado, en el sur de la Bahía de La Paz, Baja California Sur, se tiene el
registro del diciémido Dicyema guaycurense en el pulpo O. hubbsorum, dentro del
término anterior entre los pliegues de los apéndices renales de los sacos renales,
donde se encontraron nematógenos, embriones vermiformes, rombógeneos,
infusorígenos y embriones infusiformes (Castellanos-Martínez et al., 2011).
2.5 Octopus hubbsorum
2.5.1 Biología general
Octopus hubbsorum es una especie que habita en sustratos rocosos (Roper et
al., 1995; López-Uriarte et al., 2005), entre las zona submareal e intermareal a
profundidades de hasta 30m (Keen, 1971). Su cuerpo es redondeado a oval, no
presenta ocelos, sus brazos son robustos y musculosos, el segundo par aún más
que los demás, y presentan una longitud de 3 a 4 veces el manto (López-Uriarte,
2006). Son organismos dioicos (Roper et al., 1995), con hectocótilo diminuto y
cuentan con 8 a 10 laminillas branquiales (López-Uriarte, 2006). Su color va del gris
oscuro, verde, café y rojizo (Domínguez-Contreras et al., 2013), y su alimentación se
basa principalmente en crustáceos, moluscos, peces óseos, así como de pulpos de
su misma especie (Roper et al., 1995; López-Uriarte, 2006).
2.5.2 Distribución
Su distribución contempla el litoral del Pacífico Mexicano desde Bahía San
Carlos, Sonora, hasta Salina Cruz, Oaxaca (Berry, 1953; Keen, 1971; Aguilar &
Godínez-Domínguez, 1997; López-Uriarte et al., 2005), contemplando también las
islas Revillagigedo (Gotshall, 1987). Asimismo, se distribuye en la costa de la
Península de Baja California, del lado oriental, desde Bahía de los Ángeles (López-
Uriarte et al., 2005) hasta Cabo San Lucas (Gotshall, 1987), y por el occidente de la
península en Bahía Magdalena (Domínguez-Contreras et al., 2013).
9
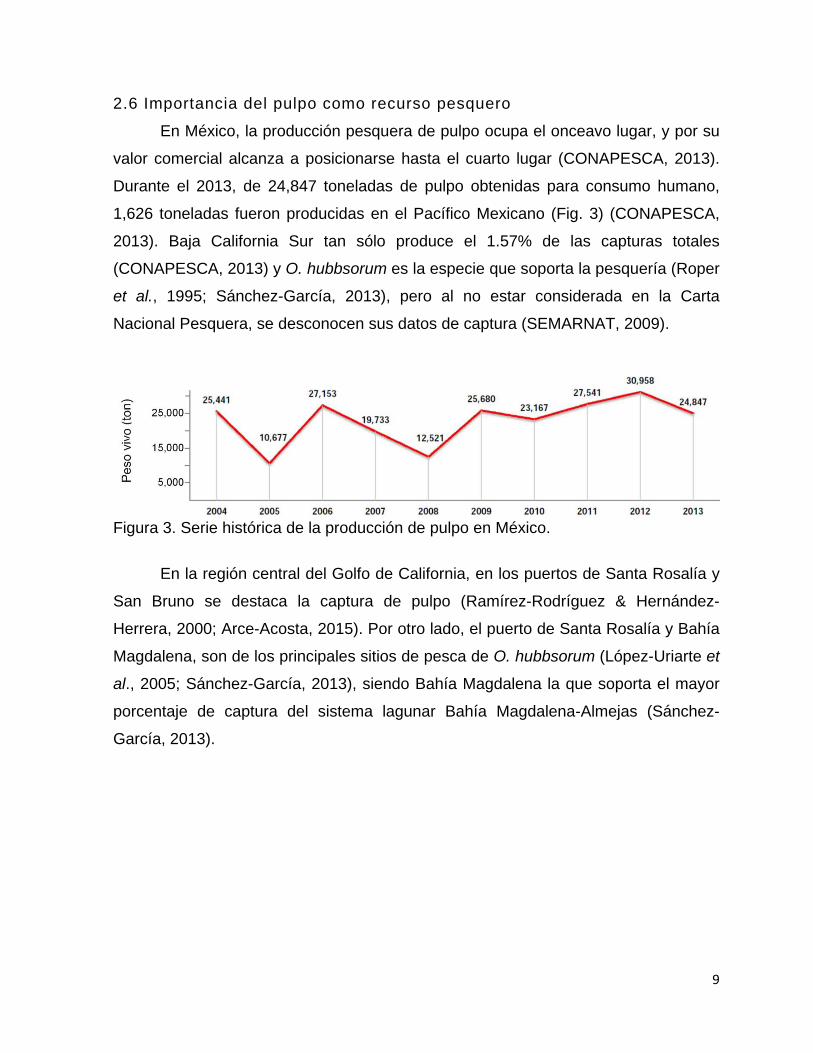
2.6 Importancia del pulpo como recurso pesquero
En México, la producción pesquera de pulpo ocupa el onceavo lugar, y por su
valor comercial alcanza a posicionarse hasta el cuarto lugar (CONAPESCA, 2013).
Durante el 2013, de 24,847 toneladas de pulpo obtenidas para consumo humano,
1,626 toneladas fueron producidas en el Pacífico Mexicano (Fig. 3) (CONAPESCA,
2013). Baja California Sur tan sólo produce el 1.57% de las capturas totales
(CONAPESCA, 2013) y O. hubbsorum es la especie que soporta la pesquería (Roper
et al., 1995; Sánchez-García, 2013), pero al no estar considerada en la Carta
Nacional Pesquera, se desconocen sus datos de captura (SEMARNAT, 2009).
Figura 3. Serie histórica de la producción de pulpo en México.
En la región central del Golfo de California, en los puertos de Santa Rosalía y
San Bruno se destaca la captura de pulpo (Ramírez-Rodríguez & Hernández-
Herrera, 2000; Arce-Acosta, 2015). Por otro lado, el puerto de Santa Rosalía y Bahía
Magdalena, son de los principales sitios de pesca de O. hubbsorum (López-Uriarte et
al., 2005; Sánchez-García, 2013), siendo Bahía Magdalena la que soporta el mayor
porcentaje de captura del sistema lagunar Bahía Magdalena-Almejas (Sánchez-
García, 2013).

10
3. JUSTIFICACIÓN La mayoría de los estudios parasitarios, son referidos a organismos marinos
como la principal fuente de investigación, pues son un reflejo de la estructura del
ecosistema. De igual modo, los parásitos sirven como indicadores de cambios en la
cadena trófica, la estabilidad del ecosistema y en ocasiones, del cambio climático.
Puesto que los cefalópodos son organismos distribuidos y consumidos
mundialmente, el estudio de su parasitología debe consistir en la importancia sobre
el daño morfológico hacia su hospedero, así como el de su posible efecto en el
humano. Sin embargo, para alcanzar estos objetivos es necesario primero
comprender la función de los cefalópodos como hospederos e identificar descriptiva y
morfológicamente sus parásitos.
Los pulpos son un importante recurso pesquero y han adquirido un gran
potencial en estudios ecológicos, etológicos y de acuacultura, debido al aumento en
su consumo dado que son una fuente considerable de proteína para el humano.
Dicho potencial, hace que surja la necesidad por evitar y prevenir enfermedades,
tanto en cautiverio como en vida libre, abriendo un nuevo panorama en el estudio del
parasitismo en cefalópodos, así como del conocimiento del papel que desempeñan
en el ecosistema marino. En México, el conocimiento de parásitos en pulpos está
representado por sólo dos trabajos, uno para el Golfo de México en O. burryi (Furuya
et al., 2002) y otro para el Golfo de California en O. hubbsorum (Castellanos-
Martínez et al., 2011), en ambos casos se registra un parásito mesozoario.
En Bahía Magdalena, la información acerca de sus aspectos ecológicos aún
es escasa, y en la región de Santa Rosalía no se han llevado a cabo estudios de este
organismo, lo cual las hace zonas adecuadas para su investigación. Asimismo,
ambos sitios tienen características diferentes entre sí por presentarse una al oriente y
otra al occidente de la Península de Baja California, por lo que son sitios ideales para
realizar análisis comparativos del parasitismo en O. hubbsorum.

11
4. OBJETIVOS 4.1 Objetivo general
Analizar la diversidad parasitaria del pulpo Octopus hubbsorum en dos
principales áreas de pesca en Baja California Sur, México.
4.2 Objetivos específicos
• Determinar el estadio ontogénico del parásito en el hospedero por medio de
técnicas histológicas y de microscopía compuesta.
• Describir las características morfológicas relevantes para la identificación
taxonómica de los parásitos.
• Identificar los principales órganos infectados y conocer si existen diferencias
para cada área de estudio.
• Determinar la prevalencia, intensidad media y abundancia en cada área de
estudio.
• Comparar el número de parásitos de cada zona entre tallas.
• Determinar las lesiones en el tejido del hospedero, por efecto de los parásitos.

12
5. MATERIAL Y MÉTODOS 5.1 Áreas de estudio
Las áreas de recolecta son importantes zonas de pesca de pulpo en Baja
California Sur (Fig. 4). La región de Santa Rosalía, que en este estudio, incluye los
puertos de Santa Rosalía y San Bruno, se localiza en el corredor Santa Rosalía-
Mulegé, dentro de la región central-occidental del Golfo de California (Arce-Acosta,
2015). Esta región es considerada mesotrófica debido a la distribución promedio de
clorofila (Espinosa-Carreón & Valdez-Holguín, 2007), y a la generación de surgencias
a lo largo de la costa, influenciadas por vientos del norte, prevalentes en invierno, y
por los vientos del sur, presentes en verano (Rodríguez-Meza, 2004; Espinosa-
Carreón & Valdez-Holguín, 2007). Por otro lado, es una región minera caracterizada
por tener depósitos de Cobre (Cu), Cobalto (Co), Zinc (Zn) y manganeso-yeso
(Coordinación General de Minería, 2008), y también es caracterizada por su actividad
pesquera (Shumilin et al., 2013; Arce-Acosta, 2015), donde el pulpo comprende el
cuarto lugar en orden de importancia de especies objetivo (Arce-Acosta, 2015).
Bahía Magdalena conforma la zona central del complejo lagunar Bahía
Magdalena-Almejas, está ubicada en la costa sur-occidental de la península de Baja
California. Se comunica con mar abierto a través de una amplia boca de acceso de
5.6 km de ancho y 40m de profundidad máxima (Funes-Rodríguez et al., 2007). En
esta bahía, se obtiene el mayor porcentaje de captura de O. hubbsorum del complejo
lagunar Bahía Magdalena-Almejas, B.C.S. (Sánchez-García, 2013). El sistema
lagunar está provisto de una gran cantidad de nutrientes generalmente en primavera,
debido al flujo de marea por parte del área adyacente al sistema lagunar, generado
por las corrientes a través de la boca de acceso (Funes-Rodríguez et al., 2007), y
puede presentar variaciones debido a la batimetría y al efecto de los vientos
(Rodríguez-Meza, 2004), por su gran productividad, es el centro de varias pesquerías
importantes (Lluch-Belda et al., 2000). La salinidad es alta en la zona de canales y
esteros ubicados al noroeste, mientras que en la región sureste la salinidad es baja
debido a la alta tasa de evaporación, la variación en la salinidad se relaciona con los
cambios de marea (Rodríguez-Meza, 2004).

13
Figura 4. Áreas de captura de Octopus hubbsorum
5.2 Obtención de hospederos
Por medio de buceo libre, se obtuvieron organismos de O. hubbsorum en la
región de Santa Rosalía en septiembre y noviembre del 2014, y febrero y octubre del
2015, mientras que en Bahía Magdalena sólo se capturaron en febrero del 2015.
Debido a que no fue posible registrar la temperatura del agua durante la obtención de
los pulpos, se obtuvieron datos de la temperatura superficial del mar por medio de
imágenes satelitales para cada día de muestreo (http://es.magicseaweed.com).
En campo, los ejemplares fueron colocados en una hielera para su
preservación. De cada organismo, se registraron con una cinta métrica (±0.5mm de
precisión), la longitud total (LT, distancia de la punta de la cabeza hasta el brazo
más largo) y la longitud del manto dorsal (LMD, distancia de la punta de la cabeza
hasta el ojo); también se registraron el peso total (PT) y el peso de la masa visceral
(PMV), con una balanza electrónica (±0.1 g). Asimismo, los organismos fueron
sexados por medio del hectocótilo en el tercer brazo derecho y mediante la
observación de gónadas masculinas y femeninas.

14
5.3 Búsqueda de parásitos
5.3.1 Toma de muestras en campo
Los pulpos fueron revisados externamente en búsqueda de parásitos o
posibles formas metacíclicas, anotando el tipo de órgano en que se localizaban, así
como alguna otra evidencia de daño o infección en el organismo. También se
tomaron muestras de músculo del manto (de 5 a 10 mm3), cuya apariencia mostró
alteraciones como abultamientos pequeños y coloración diferente. Estas muestras de
tejido fueron colocadas en Finefix (Anexo) para su preservación. Asimismo, se llevó a
cabo una inspección exhaustiva de los órganos de los pulpos en búsqueda de
endoparásitos, se procedió con la disección de la masa visceral que contiene los
órganos internos y fueron preservados en el fijador para su posterior análisis
histológico.
5.3.2 Toma de muestras en el laboratorio
En el laboratorio, las muestras preservadas en campo se colocaron al chorro
de agua para lavar el exceso de fijador. Después, la masa visceral fue colocada en
una caja petri de vidrio para llevar a cabo la separación de órganos. Los órganos se
revisaron con ayuda de un microscopio estereoscópico Olympus SZX9® en
búsqueda de parásitos o posibles formas metacíclicas, anotando el tipo de órgano en
que se localizaba, así como alguna otra evidencia de daño o infección en el
organismo, tal como se hizo cuando los pulpos se revisaron en campo.
De acuerdo al acomodo de los órganos internos y para facilitar su
manipulación e inspección, primero se separaron branquias, corazones branquiales,
gónadas, sacos renales y corazón central, seguido de las glándulas salivales
anteriores y posteriores, la boca y la glándula digestiva, y finalmente separando el
buche, estómago, ciego pilórico, intestino y ano. A cada órgano, se le realizó una
incisión a lo largo y su contenido fue removido poco a poco con un pincel, para evitar
desechar algún tipo de parásito sumergido en cada espacio. Cuando se encontraron
estructuras metacíclicas (quistes) o parásitos, estos fueron contabilizados y, a su
vez, se tomó una porción del tejido aparentemente alterado y una porción de tejido
sin evidencia de daño, para su procesamiento histológico.

15
Para la extracción de nemátodos parásitos, se realizó una disección sagital del
intestino y se observó minuciosamente con un microscopio estereoscópico. Los
nemátodos obtenidos, fueron lavados con agua de la llave para eliminar el fijador y
colocados en alcohol etílico anhidro 70% para su preservación y posterior
observación.
5.4 Observación e identificación de parásitos
5.4.1 Tratamiento de nemátodos
Con el fin de facilitar la observación de estructuras internas de los nemátodos
con el microscopio compuesto, se procedió a su deshidratación en una serie gradual
de alcoholes etílico anhidro, colocándolos en un portaobjetos cóncavo y
sumergiéndolos en gotas de alcohol al 70%, en tres tiempos de 5 minutos cada uno,
seguido de 7 minutos en alcohol al 96% y 5 minutos en alcohol al 100%. Finalmente,
se sumergieron en glicerina por al menos 10 minutos para su transparentación, y así
llevar a cabo su identificación y toma de fotografías.
5.4.2 Identificación taxonómica de nemátodos
Los nemátodos, una vez transparentados, se observaron con un microscopio
compuesto Zeiss® equipado con una cámara clara. Con un micrómetro ocular marca
Reichert® se tomaron medidas de la longitud total, del esófago (región muscular y
glandular) y de los apéndices (ventricular e intestinal), de la anchura corporal máxima
y del anillo nervioso, así como de la longitud del extremo anterior al anillo nervioso y
del extremo anterior al poro excretor. La identificación de los nemátodos se realizó
empleando los trabajos de Kinne (1990) y Anderson (2000). Todas las medidas
fueron expresadas en micrómetros, a menos que se indique lo contrario. Después de
su análisis, los nemátodos se sumergieron, en tiempos de cinco minutos, en una
serie de alcoholes etílico anhidro de concentración decreciente (100% - 96%) y
finalmente en alcohol al 70% para ser almacenados.

16
5.4.3 Análisis histológico
Para el análisis microscópico de los protozoarios, las secciones de tejido
extraídas fueron procesadas mediante la técnica histológica convencional (Humason,
1979), que consiste en la deshidratación del tejido a través de una serie de alcoholes
de concentraciones crecientes (70% - 80% - 90% - 100%), aclarado con Hemo-De®
e inclusión en Paraplast-Xtra®. Posterior a esto, se hicieron cortes de 4µm de grosor
con un micrótomo marca LEICA RM2245®, y los tejidos se colocaron en laminillas
para teñirlos con hematoxilina y eosina en un aparato automático marca Tissue-Tek
DRS®. Finalmente, los tejidos se observaron con un microscopio compuesto marca
Olympus BX41®.
5.4.4 Identificación taxonómica de protozoarios
Para la identificación taxonómica se caracterizaron las etapas de desarrollo de
los protozoarios (por medio de fotografías), considerando la forma y el tamaño del
esporocisto, así como el número y la longitud de los esporoblastos, y el tamaño de
los ooquistes de acuerdo con Poynton et al. (1992). Las imágenes obtenidas fueron
digitalizadas y analizadas por medio del programa Corel Draw Graphics Suite X3 y
Sigma Scan Pro 5. Los protozoarios fueron identificados utilizando claves
especializadas (Lom & Dyková, 1992; Estévez et al., 1996; Pascual et al., 1996;
Gestal et al., 1999b; Lee et al., 2000; Sardella et al., 2000; Ibáñez et al., 2005).
Todas las medidas se tomaron con un micrómetro ocular marca Edmund Optics® y
fueron expresadas en micrómetros, a menos que se indique lo contrario.
Para determinar la preferencia de hábitat de los protozoarios, los quistes
fueron contabilizados para cada uno de los órganos infectados. Por otro lado, el
número de individuos de protozoarios utilizado para el posterior cálculo de la
intensidad media y la abundancia, se sustituyó por el número de quistes de
protozoarios.

17
5.5 Prevalencia, intensidad media y abundancia
La frecuencia con que un parásito afecta a una población determinada, es
elemental para seguir la evolución de la parasitosis (Gállego-Berenguer, 2006). Por
lo tanto, se calculó la prevalencia, la intensidad media y la abundancia.
La prevalencia expresa la frecuencia del parasitismo en una población, y se
define como el número de hospederos infectados con uno o más individuos de un
parásito en particular, dividido por el número de hospederos revisados para esa
especie (Morales et al., 1991; Bush et al., 1997):
Prevalencia= número de hospederos infectados por una especie en particularnúmero de hospederos revisados para una especie en particular x 100
La intensidad media, se calculó dividiendo el número total de parásitos de un
taxa en particular entre el número de hospederos parasitados con ese parásito (Bush
et al., 1997).
Intensidad media =número de parásitos de una especie
número de hospederos parasitados con una especie
La abundancia, es el número promedio de parásitos que se encuentran por
hospedero, incluyendo a los parasitados como a los no parasitados (Morales et al.,
1991; Bush et al., 1997).
Abundancia = número de parásitos de una especie en particular
total de hospederos revisados
5.6 Análisis estadísticos
Se realizaron análisis de varianza de una vía para examinar las variaciones
mensuales de las variables biométricas (LM, LT, PT, PMV) en la región de Santa
Rosalía, seguido de pruebas a posteriori de comparación de medias (Tukey) para
determinar el mes más significativo. También se utilizaron ANDEVAs de una vía para

18
observar diferencias entre las variables biométricas y las prevalencias totales de las
dos localidades. Por otro lado, se llevaron a cabo correlaciones lineales simples, para
averiguar la relación entre el número de quistes y el tamaño de los individuos
(hospederos) capturados en cada zona. Las comparaciones entre zonas únicamente
fueron llevadas a cabo con el número de organismos capturados en febrero, debido a
que en Bahía Magdalena no fue posible obtener pulpos en los otros meses.
Finalmente para visualizar la preferencia de hábitat donde se desarrolla el ciclo de
vida de los protozoarios en O. hubbsorum, considerando todos los órganos
analizados, se construyó un dendograma a partir de un análisis de agrupación
jerárquico, empleando el algoritmo de agrupación por el método de aglomeración de
Ward 2 y el coeficiente de distancia de Bray-Curtis. Los análisis estadísticos se
diseñaron con el programa Rstudio® con un nivel de significancia de 0.05.
19
6. RESULTADOS
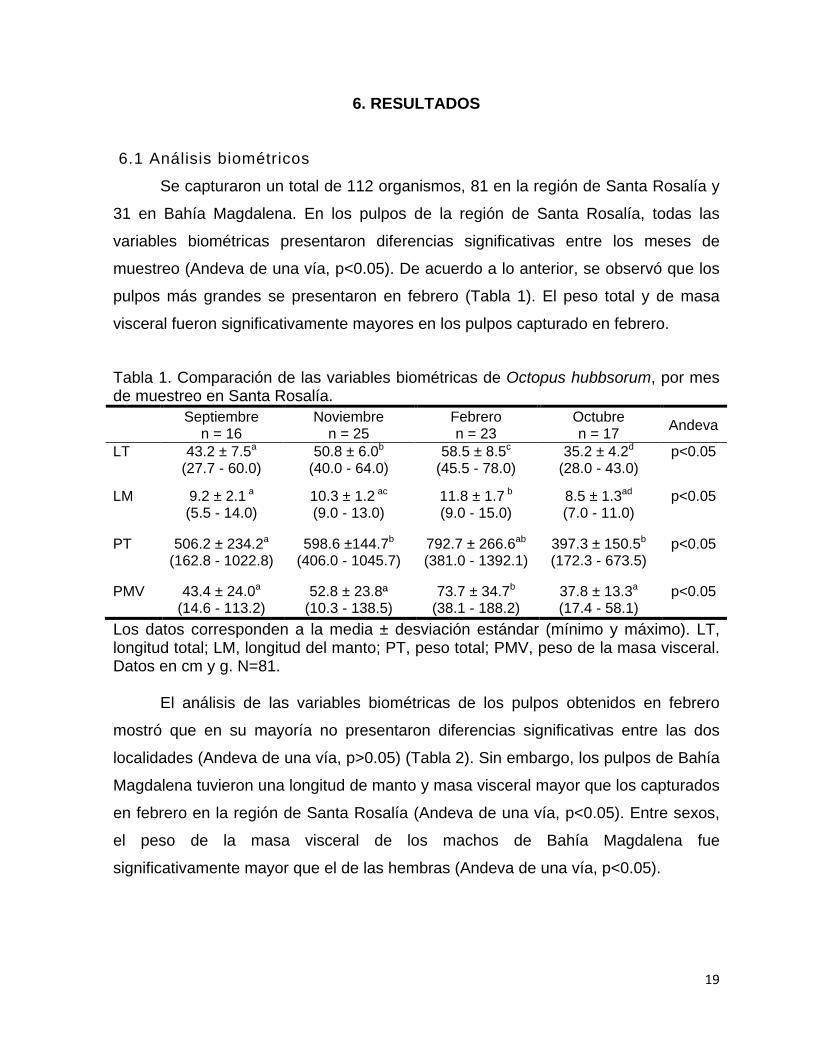
6.1 Análisis biométricos
Se capturaron un total de 112 organismos, 81 en la región de Santa Rosalía y
31 en Bahía Magdalena. En los pulpos de la región de Santa Rosalía, todas las
variables biométricas presentaron diferencias significativas entre los meses de
muestreo (Andeva de una vía, p<0.05). De acuerdo a lo anterior, se observó que los
pulpos más grandes se presentaron en febrero (Tabla 1). El peso total y de masa
visceral fueron significativamente mayores en los pulpos capturado en febrero.
Tabla 1. Comparación de las variables biométricas de Octopus hubbsorum, por mes de muestreo en Santa Rosalía.
Septiembre n = 16
Noviembre n = 25
Febrero n = 23
Octubre n = 17 Andeva
LT 43.2 ± 7.5a
(27.7 - 60.0) 50.8 ± 6.0b
(40.0 - 64.0) 58.5 ± 8.5c
(45.5 - 78.0) 35.2 ± 4.2d
(28.0 - 43.0) p<0.05
LM 9.2 ± 2.1 a (5.5 - 14.0)
10.3 ± 1.2 ac (9.0 - 13.0)
11.8 ± 1.7 b (9.0 - 15.0)
8.5 ± 1.3ad (7.0 - 11.0)
p<0.05
PT 506.2 ± 234.2a (162.8 - 1022.8)
598.6 ±144.7b
(406.0 - 1045.7) 792.7 ± 266.6ab
(381.0 - 1392.1) 397.3 ± 150.5b (172.3 - 673.5)
p<0.05
PMV 43.4 ± 24.0a (14.6 - 113.2)
52.8 ± 23.8ª (10.3 - 138.5)
73.7 ± 34.7b (38.1 - 188.2)
37.8 ± 13.3a (17.4 - 58.1)
p<0.05
Los datos corresponden a la media ± desviación estándar (mínimo y máximo). LT, longitud total; LM, longitud del manto; PT, peso total; PMV, peso de la masa visceral. Datos en cm y g. N=81.
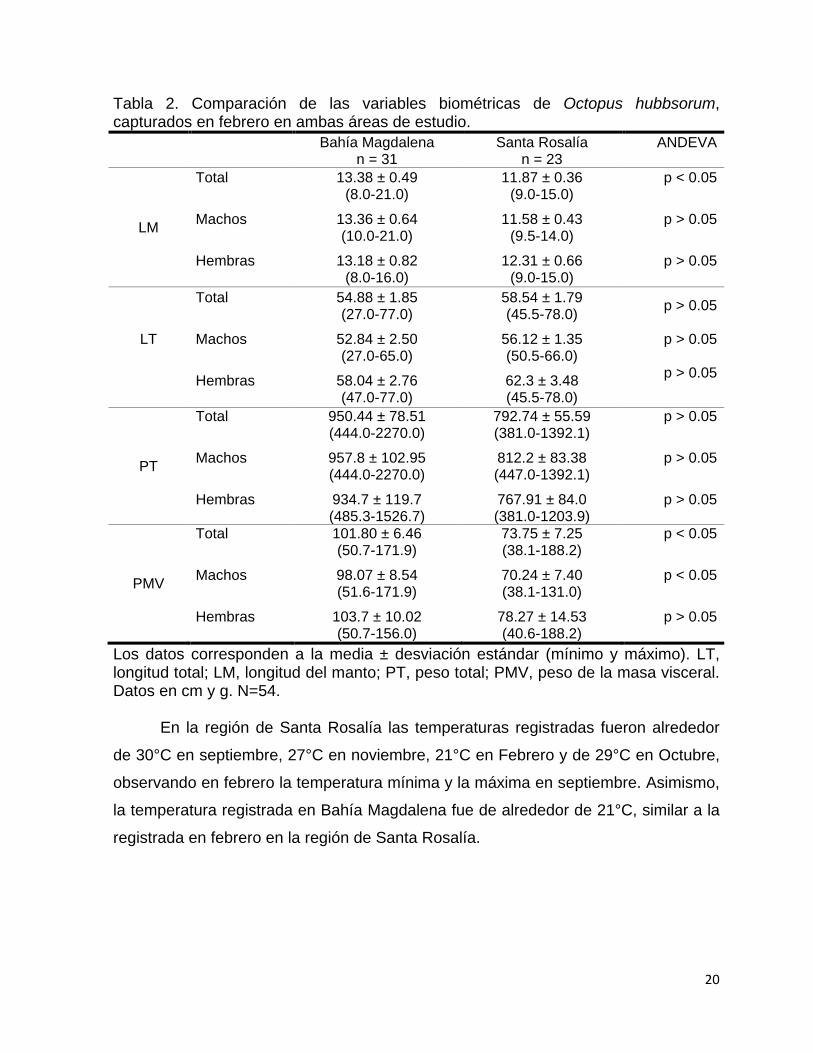
El análisis de las variables biométricas de los pulpos obtenidos en febrero
mostró que en su mayoría no presentaron diferencias significativas entre las dos
localidades (Andeva de una vía, p>0.05) (Tabla 2). Sin embargo, los pulpos de Bahía
Magdalena tuvieron una longitud de manto y masa visceral mayor que los capturados
en febrero en la región de Santa Rosalía (Andeva de una vía, p<0.05). Entre sexos,
el peso de la masa visceral de los machos de Bahía Magdalena fue
significativamente mayor que el de las hembras (Andeva de una vía, p<0.05).
20
Tabla 2. Comparación de las variables biométricas de Octopus hubbsorum, capturados en febrero en ambas áreas de estudio.
Bahía Magdalena n = 31
Santa Rosalía n = 23
ANDEVA
LM
Total Machos Hembras
13.38 ± 0.49 (8.0-21.0)
13.36 ± 0.64 (10.0-21.0)
13.18 ± 0.82 (8.0-16.0)
11.87 ± 0.36 (9.0-15.0)
11.58 ± 0.43 (9.5-14.0)
12.31 ± 0.66 (9.0-15.0)
p < 0.05
p > 0.05
p > 0.05
LT
Total Machos Hembras
54.88 ± 1.85 (27.0-77.0)
52.84 ± 2.50 (27.0-65.0)
58.04 ± 2.76 (47.0-77.0)
58.54 ± 1.79 (45.5-78.0)
56.12 ± 1.35 (50.5-66.0) 62.3 ± 3.48 (45.5-78.0)
p > 0.05
p > 0.05
p > 0.05
PT
Total Machos Hembras
950.44 ± 78.51 (444.0-2270.0) 957.8 ± 102.95 (444.0-2270.0) 934.7 ± 119.7 (485.3-1526.7)
792.74 ± 55.59 (381.0-1392.1) 812.2 ± 83.38 (447.0-1392.1) 767.91 ± 84.0 (381.0-1203.9)
p > 0.05
p > 0.05
p > 0.05
PMV
Total Machos Hembras
101.80 ± 6.46 (50.7-171.9) 98.07 ± 8.54 (51.6-171.9)
103.7 ± 10.02 (50.7-156.0)
73.75 ± 7.25 (38.1-188.2) 70.24 ± 7.40 (38.1-131.0)
78.27 ± 14.53 (40.6-188.2)
p < 0.05
p < 0.05
p > 0.05
Los datos corresponden a la media ± desviación estándar (mínimo y máximo). LT, longitud total; LM, longitud del manto; PT, peso total; PMV, peso de la masa visceral. Datos en cm y g. N=54.
En la región de Santa Rosalía las temperaturas registradas fueron alrededor
de 30°C en septiembre, 27°C en noviembre, 21°C en Febrero y de 29°C en Octubre,
observando en febrero la temperatura mínima y la máxima en septiembre. Asimismo,
la temperatura registrada en Bahía Magdalena fue de alrededor de 21°C, similar a la
registrada en febrero en la región de Santa Rosalía.
21
6.2 Análisis histológico de protozoarios
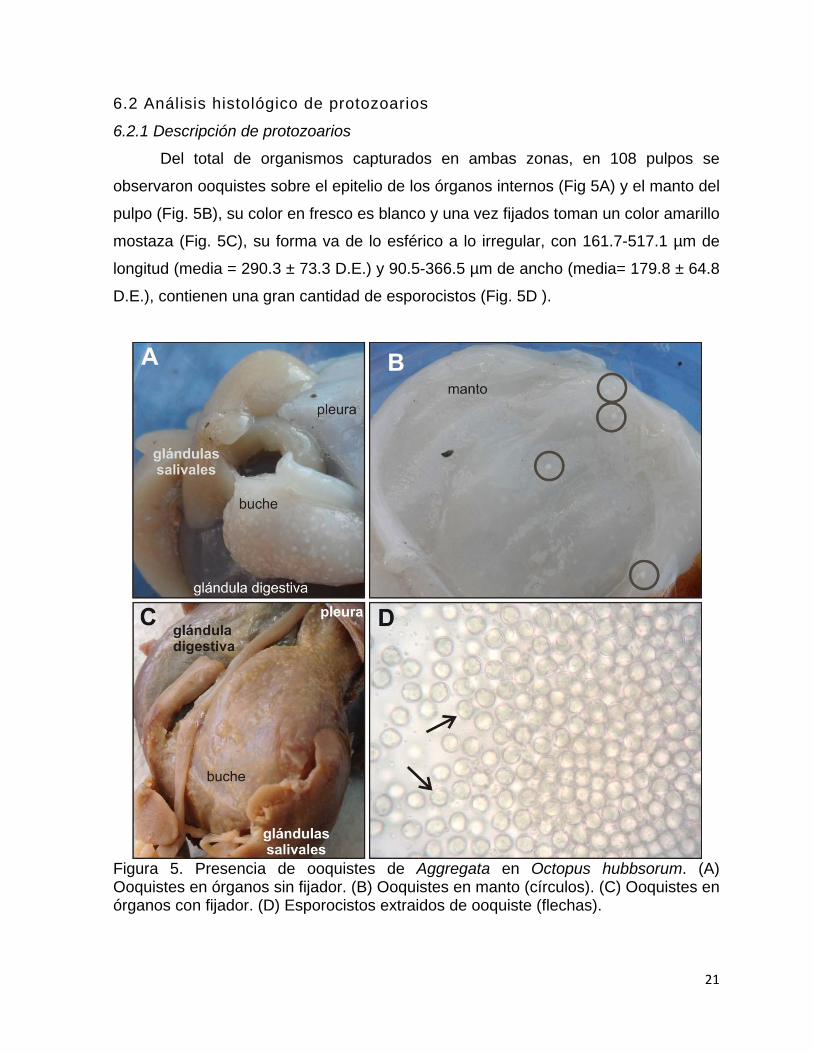
6.2.1 Descripción de protozoarios
Del total de organismos capturados en ambas zonas, en 108 pulpos se
observaron ooquistes sobre el epitelio de los órganos internos (Fig 5A) y el manto del
pulpo (Fig. 5B), su color en fresco es blanco y una vez fijados toman un color amarillo
mostaza (Fig. 5C), su forma va de lo esférico a lo irregular, con 161.7-517.1 µm de
longitud (media = 290.3 ± 73.3 D.E.) y 90.5-366.5 µm de ancho (media= 179.8 ± 64.8
D.E.), contienen una gran cantidad de esporocistos (Fig. 5D ).
Figura 5. Presencia de ooquistes de Aggregata en Octopus hubbsorum. (A) Ooquistes en órganos sin fijador. (B) Ooquistes en manto (círculos). (C) Ooquistes en órganos con fijador. (D) Esporocistos extraidos de ooquiste (flechas).

22
Por medio del análisis microscópico de los diferentes órganos de O. hubbsorum
capturados en Bahía Magdalena y la región de Santa Rosalía se identificaron
protozoarios del género Aggregata sp. cuya ubicación taxonómica es:
Phylum Apicomplexa Levine, 1970
Clase Conoidasida Levine, 1988
Subclase Coccidiasina Leuckart, 1879
Orden Eucoccidiorida Léger y Duboscq, 1910
Suborden Eimeriorina Léger, 1911
Familia Aggregatidae Labbé, 1899
Género Aggregata Frenzel, 1885
Aggregata sp. (Figs. 6, 7, 8, 9, 10)
Se identificaron estructuras características de la fase gamogónica
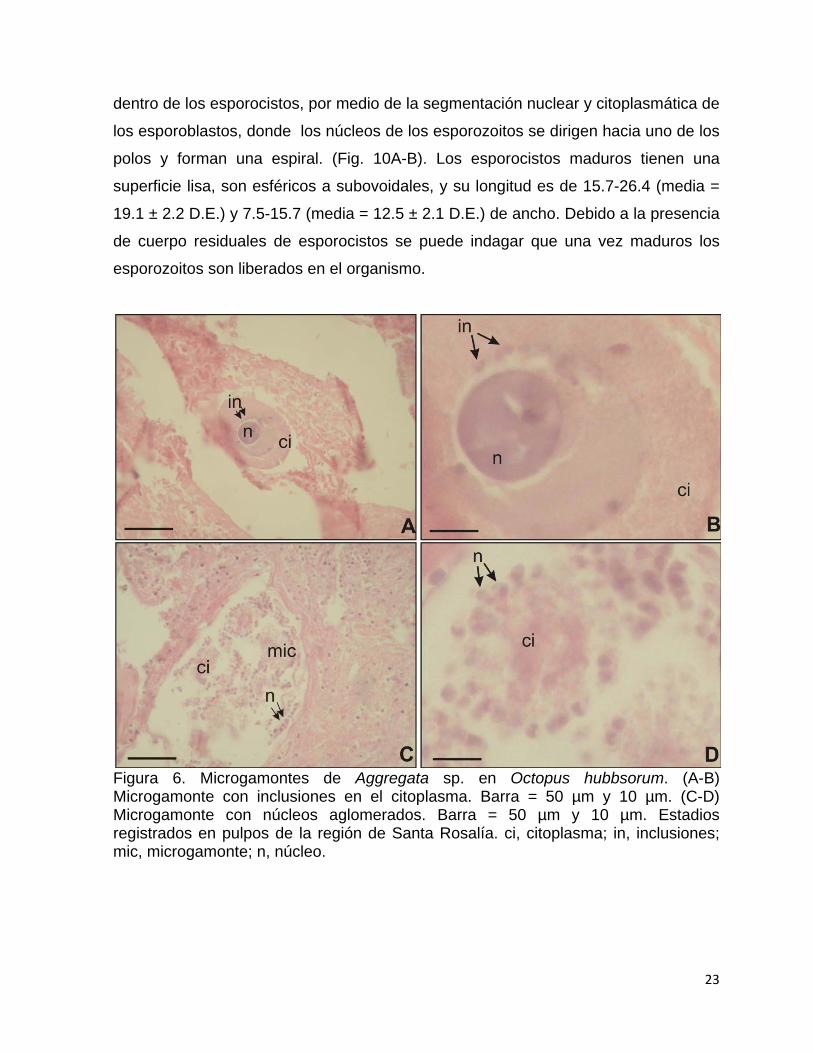
(Gamogonia), donde se observaron microgamontes (célula sexual masculina
inmadura) (Fig. 6A-B) con un núcleo central rodeado por un área poco teñida y
numerosas inclusiones en el interior del citoplasma. Los microgamontes en desarrollo
(Fig. 6C-D) presentaron una serie de plegamientos citoplasmáticos y diversos
núcleos alargados rodeando el citoplasma. No se observaron microgametos en los
organismos capturados en Bahía Magdalena. Por otro lado, los macrogamontes
(célula sexual femenina inmadura) (Fig. 7) se distinguen de los microgamontes por
ser más grandes, no presentan inclusiones en el citoplasma y presenta un citoplasma
de apariencia poroso (esponjoso).
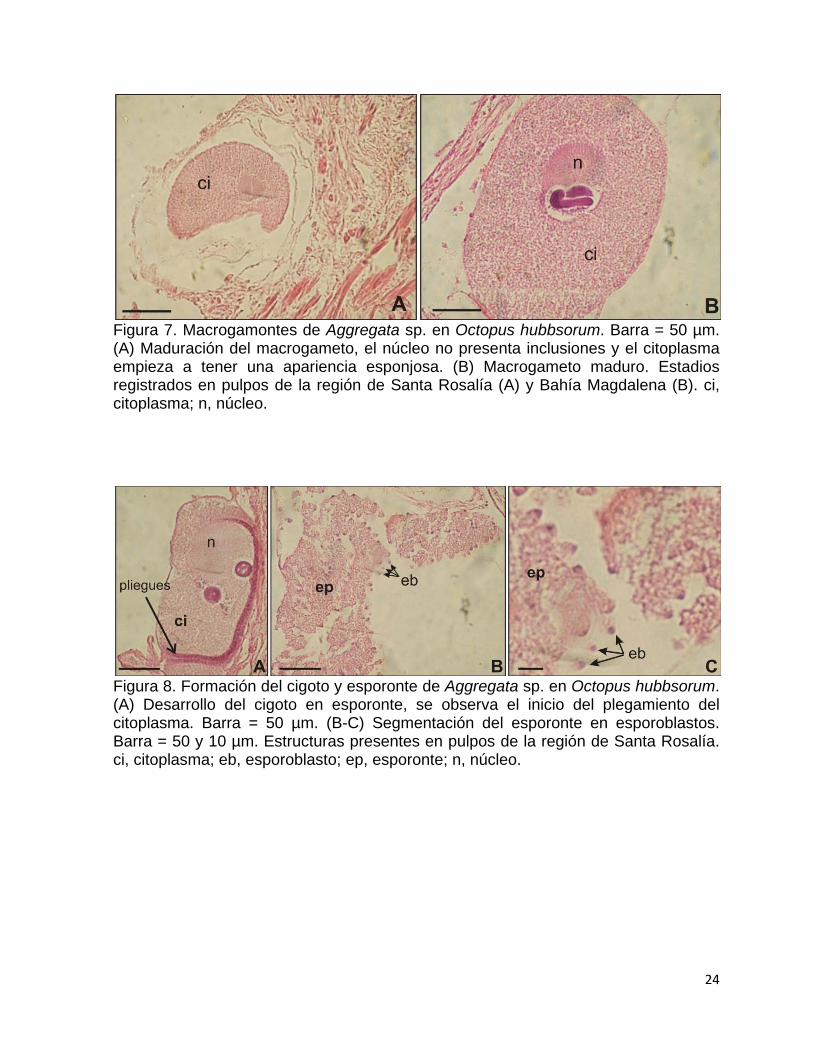
La fase esporogónica (Esporogonia), inicia una vez que se forma el cigoto
(Fig. 8A) como producto de la unión del macro y microgametos. En esta fase, el
cigoto da origen al esporonte mediante una serie de plegamientos citoplasmáticos y
la presencia de núcleos en la periferia del citoplasma (Fig. 8B-C). Los núcleos se
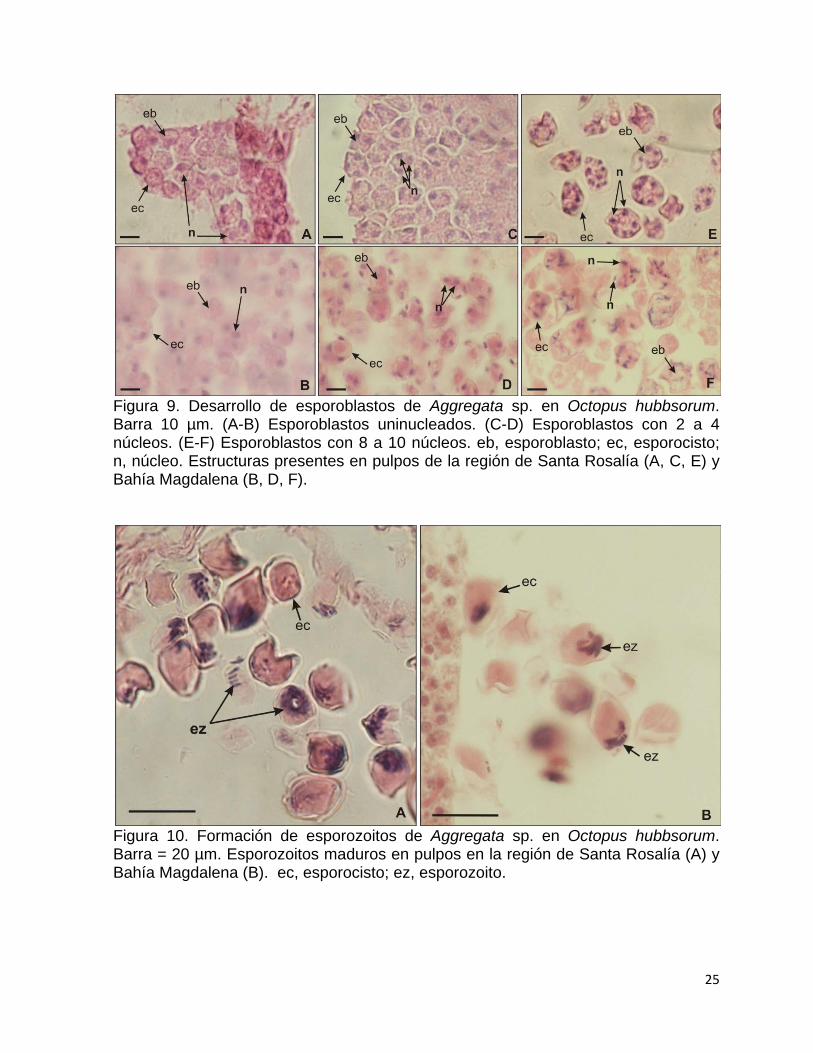
desprenden del citoplasma del esporonte y forman esporoblastos uninucleados (Fig.
9A-B), y durante su desarrollo el número de núcleos aumenta de 2 a 4 (Fig. 9C-D)
hasta 6 a 8 (Fig. 9E-F) por esporocisto. El desarrollo de los esporozoitos ocurre
23
dentro de los esporocistos, por medio de la segmentación nuclear y citoplasmática de
los esporoblastos, donde los núcleos de los esporozoitos se dirigen hacia uno de los
polos y forman una espiral. (Fig. 10A-B). Los esporocistos maduros tienen una
superficie lisa, son esféricos a subovoidales, y su longitud es de 15.7-26.4 (media =
19.1 ± 2.2 D.E.) y 7.5-15.7 (media = 12.5 ± 2.1 D.E.) de ancho. Debido a la presencia
de cuerpo residuales de esporocistos se puede indagar que una vez maduros los
esporozoitos son liberados en el organismo.
Figura 6. Microgamontes de Aggregata sp. en Octopus hubbsorum. (A-B) Microgamonte con inclusiones en el citoplasma. Barra = 50 µm y 10 µm. (C-D) Microgamonte con núcleos aglomerados. Barra = 50 µm y 10 µm. Estadios registrados en pulpos de la región de Santa Rosalía. ci, citoplasma; in, inclusiones; mic, microgamonte; n, núcleo.
24
Figura 7. Macrogamontes de Aggregata sp. en Octopus hubbsorum. Barra = 50 µm. (A) Maduración del macrogameto, el núcleo no presenta inclusiones y el citoplasma empieza a tener una apariencia esponjosa. (B) Macrogameto maduro. Estadios registrados en pulpos de la región de Santa Rosalía (A) y Bahía Magdalena (B). ci, citoplasma; n, núcleo.
Figura 8. Formación del cigoto y esporonte de Aggregata sp. en Octopus hubbsorum. (A) Desarrollo del cigoto en esporonte, se observa el inicio del plegamiento del citoplasma. Barra = 50 µm. (B-C) Segmentación del esporonte en esporoblastos. Barra = 50 y 10 µm. Estructuras presentes en pulpos de la región de Santa Rosalía. ci, citoplasma; eb, esporoblasto; ep, esporonte; n, núcleo.
25
Figura 9. Desarrollo de esporoblastos de Aggregata sp. en Octopus hubbsorum. Barra 10 µm. (A-B) Esporoblastos uninucleados. (C-D) Esporoblastos con 2 a 4 núcleos. (E-F) Esporoblastos con 8 a 10 núcleos. eb, esporoblasto; ec, esporocisto; n, núcleo. Estructuras presentes en pulpos de la región de Santa Rosalía (A, C, E) y Bahía Magdalena (B, D, F).
Figura 10. Formación de esporozoitos de Aggregata sp. en Octopus hubbsorum. Barra = 20 µm. Esporozoitos maduros en pulpos en la región de Santa Rosalía (A) y Bahía Magdalena (B). ec, esporocisto; ez, esporozoito.

26
6.2.2 Resumen taxonómico
Hospederos: O. hubbsorum (Berry), ‘pulpo de Hubb’, (Octopodidae).
Localidades: Costa de Bahía Magdalena y región de Santa Rosalía, Baja
California Sur, México.
Sitio de infección. La gamogonia y esporogonia se desarrollan en la región no
cuticularizada del ciego pilórico, intestino, buche, glándula digestiva, ductos del
segundo par de glándulas salivales, branquias, músculo del manto, ano, boca,
gónada, corazones branquiales, estómago, hectocótilo, ojos y sacos renales. Se
localizan generalmente en la región de la mucosa y serosa del tejido.
6.2.3 Observaciones del género Aggregata
En los últimos 20 años, el género Aggregata Frenzel, 1885 ha sido sujeto de
diversas revisiones taxonómicas (Hansson, 1997), y hasta ahora se han descrito
alrededor de 25 especies. Las especies reconocidas de acuerdo a Hansson (1997),
Perkins et al. (2000) y las páginas de internet Global Biodiversity Information Facility,
Global Names Index y World Register of Marine Species (WoRMS Editorial Board)
son:
Aggregata andresi Gestal, Nigmatullin, Hochberg, Guerra & Pascual,
2005; A. coelomica Léger, 1901; A. dobelli Poynton, Reimschussel & Stoskopf,
1992; A. eberthi (Labbé, 1895) Léger & Duboscq, 1906; A. inachi (Smith, 1905)
Léger & Duboscq, 1906; A. jacquementi Moroff, 1906; A. kudoi
Narasimhamurti, 1979; A. labbei Moroff, 1908; A. leandri (Goodrich, 1950)
Théodoridès & Desportes, 1975; A. legeri Moroff, 1908; A. máxima
Théodoridès & Desportes, 1975; A. millerorum Poynton, Reimschussel &
Stoskopf, 1992; A. octopiana (Schneider, 1875) Frenzel, 1885; A. ovata Moroff,
1908; A. patagonica Sardella, Re & Timi, 2000; A. portunidarum Frenzel, 1885;
A. reticulosa Morof, 1908; A. sagittata Gestal, Guerra, Abollo & Pascual; A.
schneideri Moroff, 1908; A. sepiae Lankester, 1863; A. siedleckii Moroff, 1908;
A. spinosa Moroff, 1906; A. stellata Moroff, 1908; A. vagans Léger & Duboscq,
1903; A. valdessensis Sardella, Re & Timi, 2000.

27
Son sinónimos Benedenia Schneider, 1875, pro parte; Legeria Blanchard,
1900; Eucoccidium Lühe, 1902, pro parte: Legendaria Jacquement, 1903 (Perkins et
al., 2000). También se ha considerado que A. portunidarum Frenzel, 1885 podría ser
un sinónimo junior de A. eberthi Labbé, 1895 o de A. octopiana Schneider, 1875 por
ser una especie tipo. Por otro lado, A vagans Léger & Duboscq, 1903 fue
considerada como el gregarínido Cephaloidophora ocellata (Sprague & Couch,
1971).
Los parásitos del género Aggregata requieren de hospederos intermediarios
para completar su ciclo de vida, donde la fase merogónica (asexual) ocurre en el
intestino de crustáceos decápodos (Olsen, 1974; Hansson, 1997; Perkins et al.,
2000) y las fases gamogonia y esporogonia se desarrollan principalmente en el tracto
digestivo de cefalópodos (Olsen, 1974), convirtiéndolos en su hospedero definitivo.
Algunas especies de coccidios pueden generar enfermedades en vertebrados e
invertebrados, sin embargo, para Aggregata no existen evidencias de daño hacia el
ser humano.
En este trabajo, se registra por primera vez en el pulpo O. hubbsorum el
género Aggregata en dos localidades de la costa oriental y occidental de Baja
California Sur, México, con lo que se consigue conocer su distribución en México y
ampliar su distribución mundial.

28
6.3 Análisis microscópico de nemátodos
6.3.1 Descripción de nemátodos
Dentro del intestino de 112 individuos de O. hubbsorum se encontraron 19
ejemplares de nemátodos del género Hysterothylacium sp. en fase larvaria L3,
ubicados taxonómicamente de la siguiente manera:
Phylum Nematoda (Rudolphi, 1808) Lankester, 1877
Clase Secernentea Von Linstow, 1905
Orden Ascaridida Skrjabin y Schulz, 1938
Superfamilia Ascaridoidea Railliet y Henry, 1915
Familia Anisakidae (Railliet y Henry, 1912) Skrjabin y Karokhin, 1945
Género Hysterothylacium Ward y Magath, 1917
Hysterothylacium sp. (Larvas) (Figs. 11, 12, 13, 14)
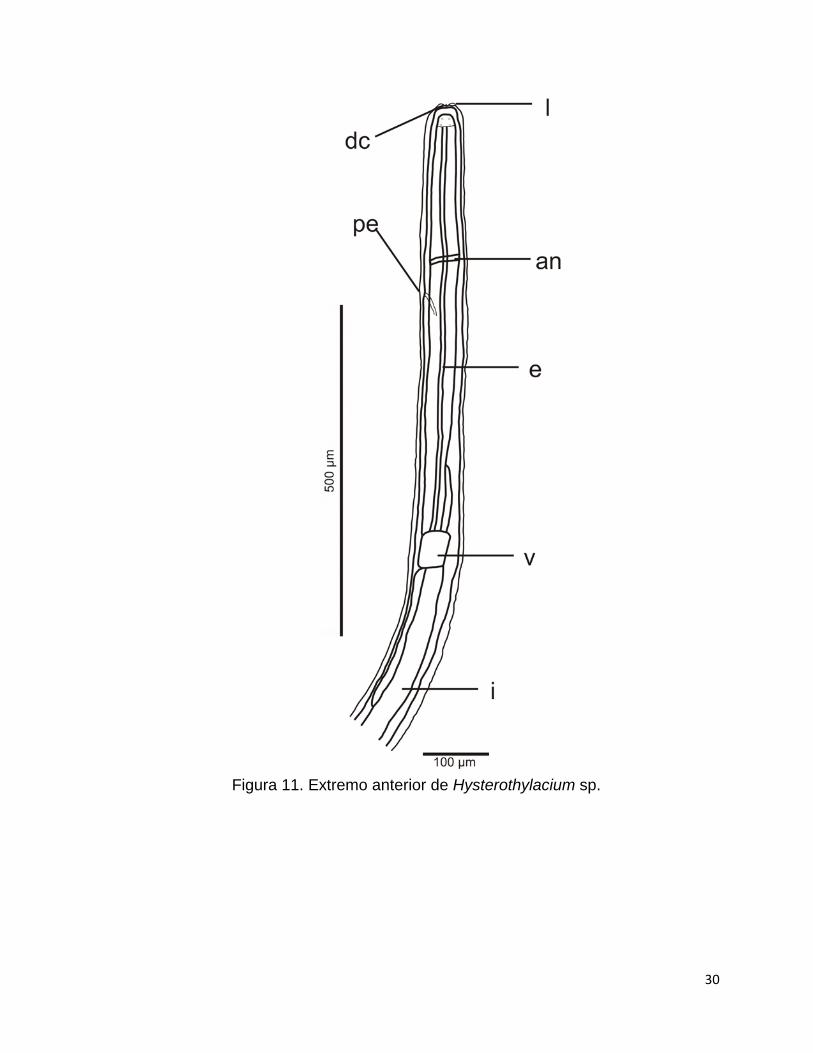
Los especímenes son de color blanco, de cuerpo alargado y cilíndrico, de 2.5
a 3.9 mm de longitud por 83.2 a 99.3 de anchura mayor corporal (Tabla 3). Es
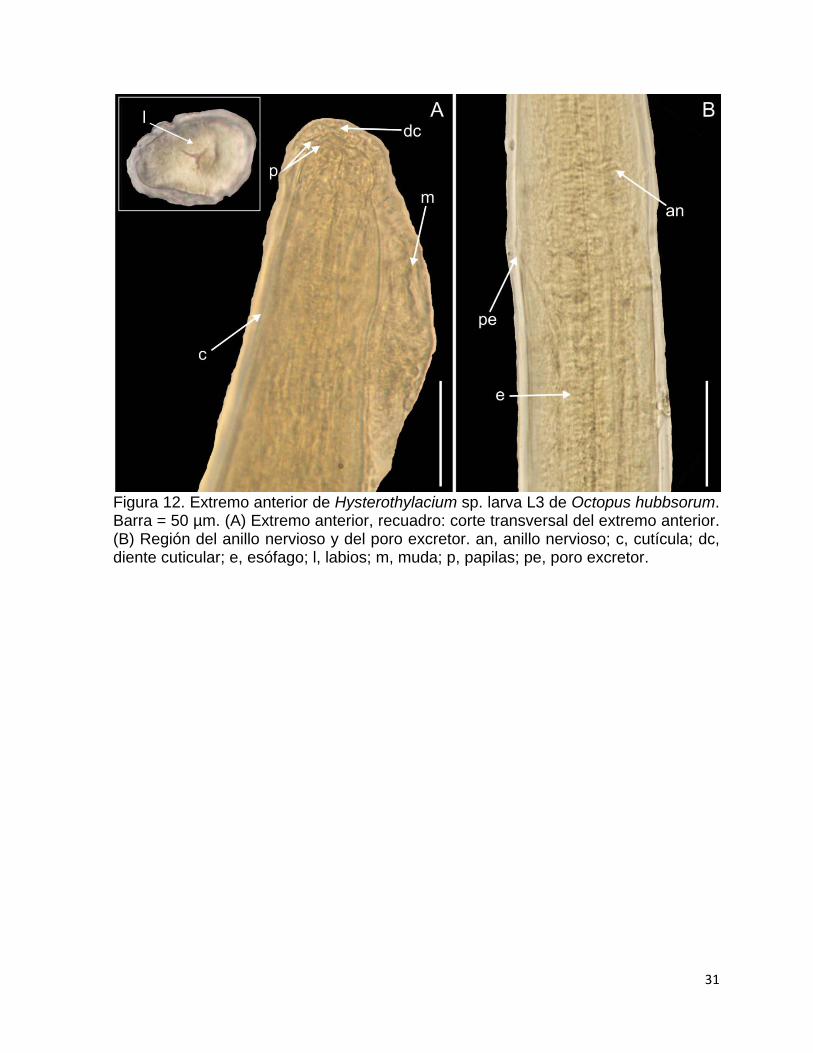
evidente la presencia de una cutícula y muda (Fig. 12A). El extremo anterior es romo,
tiene tres labios gruesos y un diente cuticular anteroventral, así como un par de
papilas en el lado dorsal (Fig. 12A). El anillo nervioso rodea la región anterior del
esófago muscular y por debajo de éste, se sitúa el poro excretor (Fig. 12B).
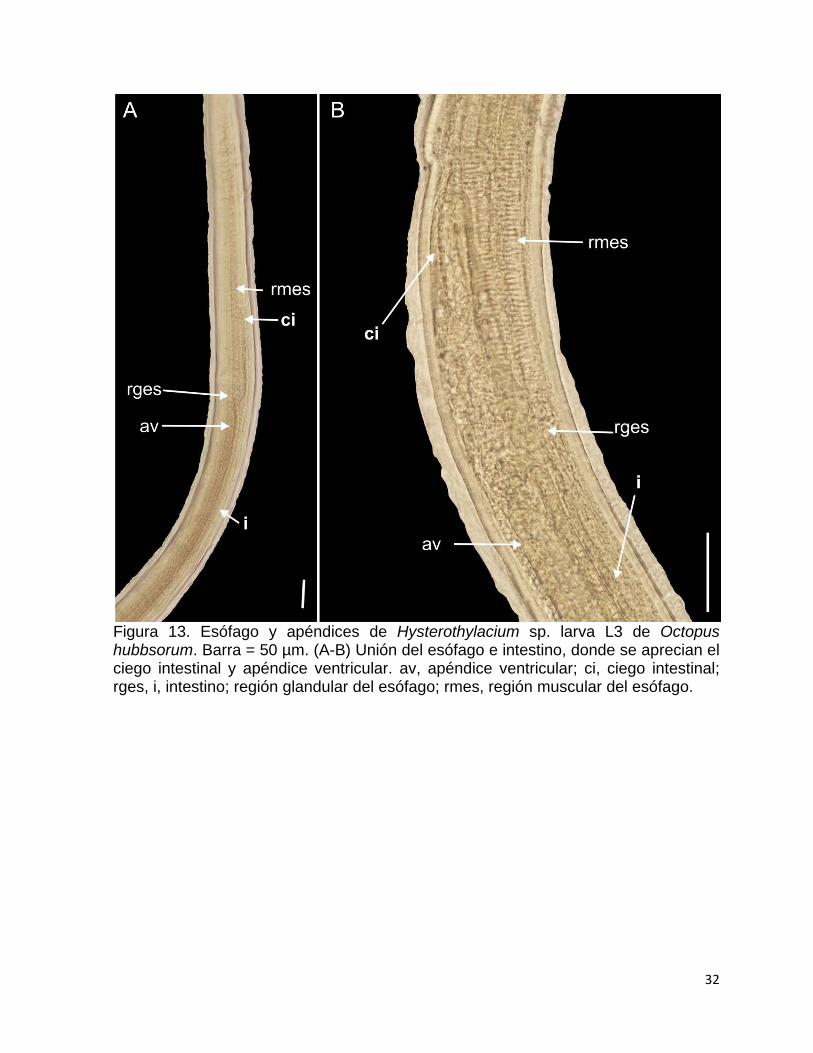
El esófago tiene una longitud de 577.9 a 630, es largo y cilíndrico, y está
conformado por una región muscular casi en su totalidad de 535.6 a 588.2, y una
glandular en una última porción que mide de 41.8 a 42.2, la cual constituye un
pequeño ventrículo (Fig. 13). Se aprecian dos apéndices, uno intestinal (ciego) de
89.7 a 105 de longitud y otro ventricular en forma de saco y más grande que el
apéndice intestinal, con una longitud de 164.6 a 181 (Fig. 13). El intestino se conecta
con la región glandular del esófago (Fig. 13) y ocupa el resto del cuerpo, con una
longitud de 2.4 a 3 mm y termina en la cloaca en organismos machos o en el ano en
hembras (Figura 14).
29
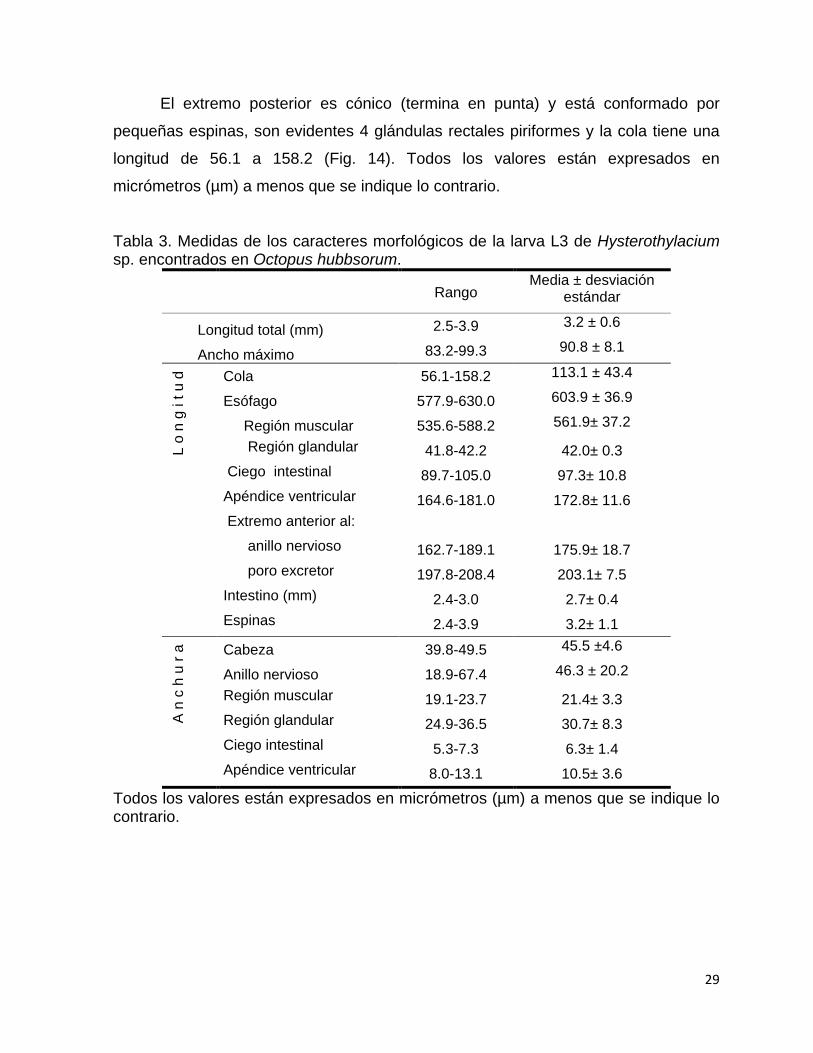
El extremo posterior es cónico (termina en punta) y está conformado por
pequeñas espinas, son evidentes 4 glándulas rectales piriformes y la cola tiene una
longitud de 56.1 a 158.2 (Fig. 14). Todos los valores están expresados en
micrómetros (µm) a menos que se indique lo contrario.
Tabla 3. Medidas de los caracteres morfológicos de la larva L3 de Hysterothylacium sp. encontrados en Octopus hubbsorum.
Rango
Media ± desviación estándar
Longitud total (mm) 2.5-3.9 3.2 ± 0.6
Ancho máximo 83.2-99.3 90.8 ± 8.1
L o
n g
i t u
d
Cola 56.1-158.2 113.1 ± 43.4
Esófago 577.9-630.0 603.9 ± 36.9
Región muscular 535.6-588.2 561.9± 37.2 Región glandular 41.8-42.2 42.0± 0.3 Ciego intestinal 89.7-105.0 97.3± 10.8 Apéndice ventricular 164.6-181.0 172.8± 11.6 Extremo anterior al: anillo nervioso 162.7-189.1 175.9± 18.7 poro excretor 197.8-208.4 203.1± 7.5 Intestino (mm) 2.4-3.0 2.7± 0.4 Espinas 2.4-3.9 3.2± 1.1
A n
c h
u r a
Cabeza 39.8-49.5 45.5 ±4.6
Anillo nervioso 18.9-67.4 46.3 ± 20.2 Región muscular 19.1-23.7 21.4± 3.3 Región glandular 24.9-36.5 30.7± 8.3 Ciego intestinal 5.3-7.3 6.3± 1.4 Apéndice ventricular 8.0-13.1 10.5± 3.6
Todos los valores están expresados en micrómetros (µm) a menos que se indique lo contrario.
30
Figura 11. Extremo anterior de Hysterothylacium sp.
31
Figura 12. Extremo anterior de Hysterothylacium sp. larva L3 de Octopus hubbsorum. Barra = 50 µm. (A) Extremo anterior, recuadro: corte transversal del extremo anterior. (B) Región del anillo nervioso y del poro excretor. an, anillo nervioso; c, cutícula; dc, diente cuticular; e, esófago; l, labios; m, muda; p, papilas; pe, poro excretor.
32
Figura 13. Esófago y apéndices de Hysterothylacium sp. larva L3 de Octopus hubbsorum. Barra = 50 µm. (A-B) Unión del esófago e intestino, donde se aprecian el ciego intestinal y apéndice ventricular. av, apéndice ventricular; ci, ciego intestinal; rges, i, intestino; región glandular del esófago; rmes, región muscular del esófago.

33
Figura 14. Extremo posterior de Hysterothylacium sp. larva L3 de Octopus hubbsorum. Barra = 50 µm. En el recuadro se aprecian las espinas en la punta de la cola del nemátodo. a, ano; c, cloaca; es, espinas; gr, glándulas rectales; i, intestino; p, papilas.
6.3.2 Resumen taxonómico
Hospederos: Octopus hubbsorum (Berry) (Octopoda: Octopodidae).
Localidad: Santa Rosalía y Bahía Magdalena, B.C.S.
Sitio de infección: Mucosa del intestino.
6.3.3 Observaciones del género Hysterothylacium
El género Hysterothylacium, antes contemplado como Thynnascaris Dollfus,
1933 o Contracecum sensu lato, fue revisado por Deardorff & Overstreet (1981), y en
los últimos 10 años, se han registrado nuevas especies del género Hysterothylacium
en organismos marinos (Angelucci et al., 2011). Las siguientes especies son
reconocidas de acuerdo a la página de internet Global Names Index:
Hysterothylacium aduncum (Rudolphi, 1802) Deardorff y Overstreet, 1981; H.
aetobatum Lakshmi, 2005; H. amoyense Hsu, 1933; H. analarum Rye & Baker,
1984; H. arnoglossi (Lebre & Petter, 1983) Petter & Maillard, 1988; H. auctum
Rudolphi, 1802; H. bidentatum Linstow, 1899; H. bifidalatum Petter & Maillard,
1988; H. brachyurum Ward & Magath, 1916; H. carutti Rajyalakshmi,
Hanumantha Rao & Shyamasundari, 1993; H. cayugensis Wigdor, 1918; H.

34
cenaticum Bruce & Cannon, 1989; H. cenotae Pearse, 1936; H. channai
Rajyalakshmi, 1995; H. chaunaxi, Olsen 1952; H. chorinemi Parukhin, 1966; H.
chrysostomi Bruce, 1990; H. clavatum (Rudolphi, 1809) Deardoff & Overstreet,
1981; H. collare Cobb, 1929; H. cornutum (Stossich, 1904) Derdoff &
Overstreet, 1981; H. corrugatum Deardorff & Overstreet, 1981; H.
coryphaenoidi Parukhin, 1989; H. cyclopteri Kreis, 1952; H. elurensis
Rajyalakshmi & Lakshmi, 1995; H. eurycheilum Olsen, 1952; H. fabri
(Rudolphi, 1819) Derdoff & Overstreet, 1981; H. fluviatile Moravec & Sey,
1988; H. fortalezae Klein 1973; H. fossilii Rajyalakshmi 1996; H. gadi (O.F.
Müller, 1777) Derdoff & Overstreet, 1981; H. ganeshi Rajya Lakshmi &
Sreeramulu 2007; H. geschei Torres, Andrade & Silva 1998; H. histiophori
Yamaguti 1935; H. hospitum Solovjeva & Pozdnjakov 1984; H. increscens
(Molin, 1858) Derdoff & Overstreet, 1981; H. incurvum (Rudolphi, 1819)
Derdoff & Overstreet, 1981; H. japonicum Rajyalakshmi 1996; H. kiranii
Rajyalakshmi 1993; H. krishnai Rajyalakshmi 1992; H. legendrei Dollfus, 1933;
H. leptaspi Bruce 1990; H. liparis Liang Li, Zhen Xu & Luping Zhang 2007; H.
makairi Bruce & Cannon 1989; H. marinum Linnaeus, 1767; H. muraenesoxin
Luo 1999; H. narayanensis Rajyalakshmi 1997; H. nellorensis Rajyalakshmi
1996; H. neocornutum Rajyalakshmi, Hanumantha Rao & Shyamasundari
1992; H. nipponense Moravec & Nagasawa 1998; H. ogcocephali Olsen 1952;
H. patagonense Moravec, Urawa & Coria 1997; H. pelagicum Deardorff &
Overstreet 1982; H. perezi Gopar-Merino, Osorio-Sarabia & Garcia-Prieto,
2005; H. petteri Sheenko 1991; H. physiculi Moravec & Nagasawa 2000; H.
poecilurai Rajyalakshmi & Sreeramulu 2005; H. psettodi Parukhin 1989; H.
pseudotumbili Rajya Lakshmi, Hanumantha Rao & Shyamasundari 1991; H.
punctati Rajyalakshmi 1995; H. reliquens Norris & Overstreet, 1975; H.
rhacodes Deardorff & Overstreet, 1978; H. rhamdiae Brizzola & Tanzola 1995;
H. rigidum Rudolphi, 1809; H. scomberoidei Bruce & Cannon 1989; H.
scomberomori Yamaguti 1941; H. sebae Bruce 1990; H. seriolae Yamaguti,
1941; H. sinense Li, An & Zhang 2007; H. tasmaniense Johnston & Mawson,

35
1945; H. tetrapteri Bruce & Cannon 1989; H. thalassini Bruce 1990; H. trichiuri
Thwaite 1927; H. winteri Torres & Soto, 2004 y H. zenopsis, Yamaguti, 1941.
Los nemátodos adultos del género Hysterothylacium generalmente se
localizan en el tracto digestivo de peces, mientras que las larvas son capaces de
parasitar diversos tejidos tanto de peces como de invertebrados (Deardorff &
Overstreet, 1981), mismos que pueden actuar como hospederos definitivos,
intermediarios o paraténicos (Norris & Overstreet, 1976). Por otro lado, los
nemátodos del género Hysterothylacium y Contracecum son registrados
principalmente en cefalópodos con hábitos alimenticios bentónicos (Cannon, 1977).
Con este trabajo, se consigue ampliar el conocimiento de la distribución de
Hysterothylacium sp., registrado por primera vez en O. hubbsorum en dos
localidades de ambas costas de Baja California Sur, México.
6.4 Análisis del hábitat de parásitos
6.4.1 Preferencia de hábitat de protozoarios
En general, los órganos más parasitados fueron el intestino, el ciego pilórico,
la glándula digestiva y el buche. En los pulpos de la región de Santa Rosalía, se
observaron quistes sobre el ciego y el intestino en todos los meses de captura,
mientras que la glándula digestiva y el buche, no presentaron quistes en todos los
meses de captura (Fig. 15).
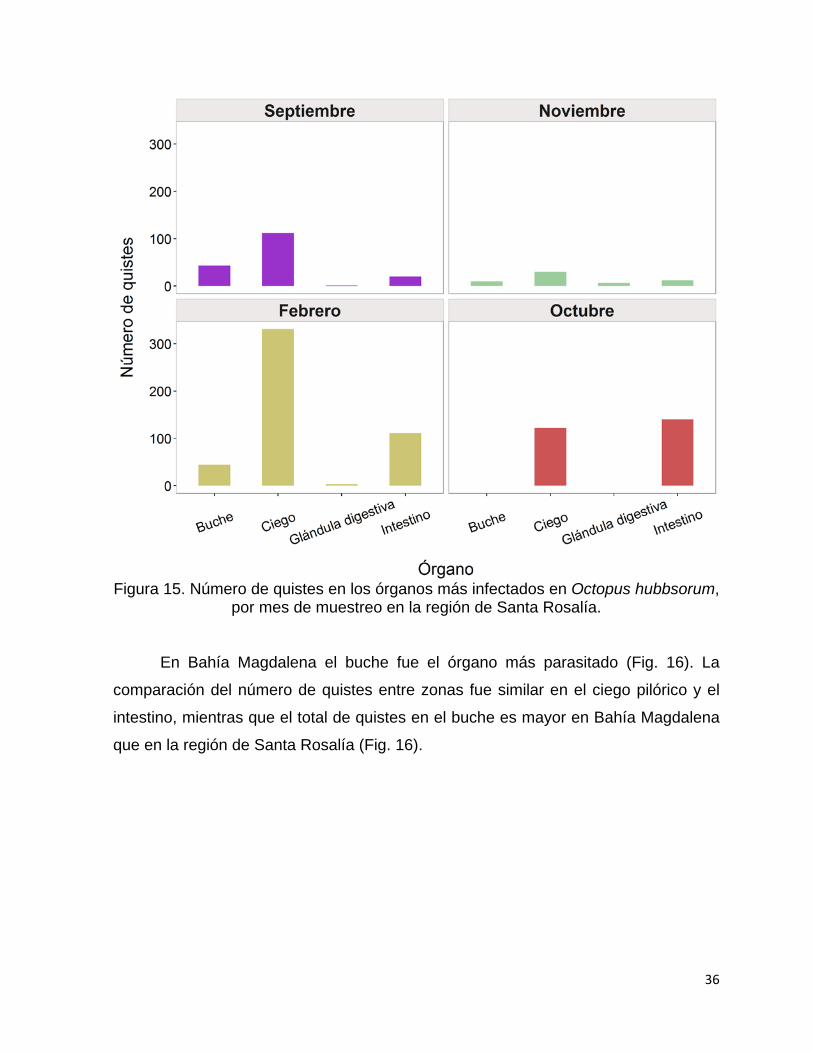
36
Figura 15. Número de quistes en los órganos más infectados en Octopus hubbsorum,
por mes de muestreo en la región de Santa Rosalía.
En Bahía Magdalena el buche fue el órgano más parasitado (Fig. 16). La
comparación del número de quistes entre zonas fue similar en el ciego pilórico y el
intestino, mientras que el total de quistes en el buche es mayor en Bahía Magdalena
que en la región de Santa Rosalía (Fig. 16).
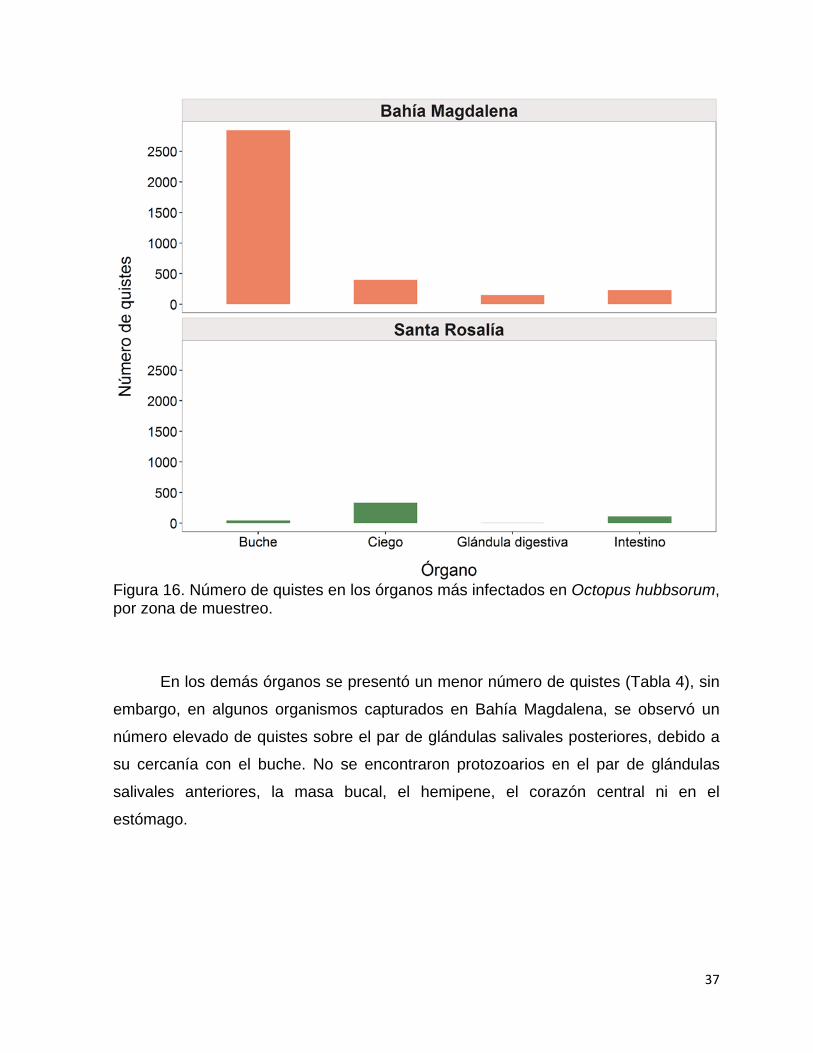
37
Figura 16. Número de quistes en los órganos más infectados en Octopus hubbsorum, por zona de muestreo.
En los demás órganos se presentó un menor número de quistes (Tabla 4), sin
embargo, en algunos organismos capturados en Bahía Magdalena, se observó un
número elevado de quistes sobre el par de glándulas salivales posteriores, debido a
su cercanía con el buche. No se encontraron protozoarios en el par de glándulas
salivales anteriores, la masa bucal, el hemipene, el corazón central ni en el
estómago.
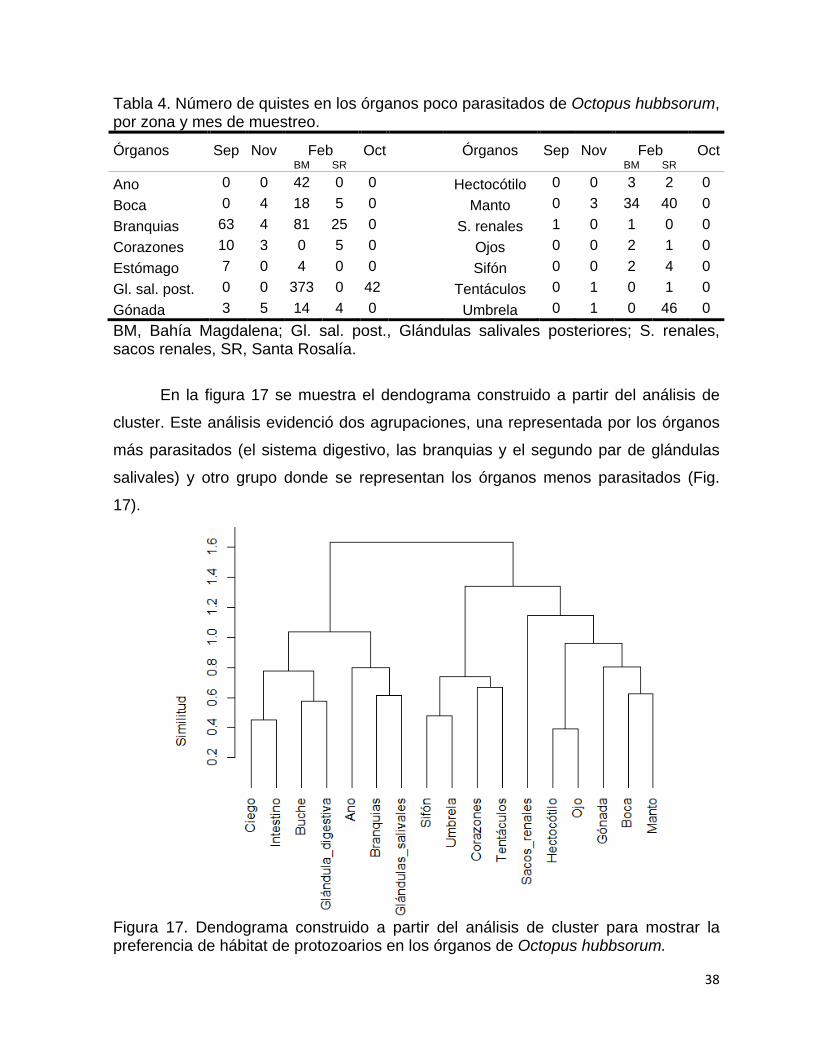
38
Tabla 4. Número de quistes en los órganos poco parasitados de Octopus hubbsorum, por zona y mes de muestreo.
Órganos Sep Nov Feb Oct Órganos Sep Nov Feb Oct BM SR BM SR
Ano 0 0 42 0 0 Hectocótilo 0 0 3 2 0 Boca 0 4 18 5 0 Manto 0 3 34 40 0 Branquias 63 4 81 25 0 S. renales 1 0 1 0 0 Corazones 10 3 0 5 0 Ojos 0 0 2 1 0 Estómago 7 0 4 0 0 Sifón 0 0 2 4 0 Gl. sal. post. 0 0 373 0 42 Tentáculos 0 1 0 1 0 Gónada 3 5 14 4 0 Umbrela 0 1 0 46 0 BM, Bahía Magdalena; Gl. sal. post., Glándulas salivales posteriores; S. renales, sacos renales, SR, Santa Rosalía.
En la figura 17 se muestra el dendograma construido a partir del análisis de
cluster. Este análisis evidenció dos agrupaciones, una representada por los órganos
más parasitados (el sistema digestivo, las branquias y el segundo par de glándulas
salivales) y otro grupo donde se representan los órganos menos parasitados (Fig.
17).
Figura 17. Dendograma construido a partir del análisis de cluster para mostrar la preferencia de hábitat de protozoarios en los órganos de Octopus hubbsorum.
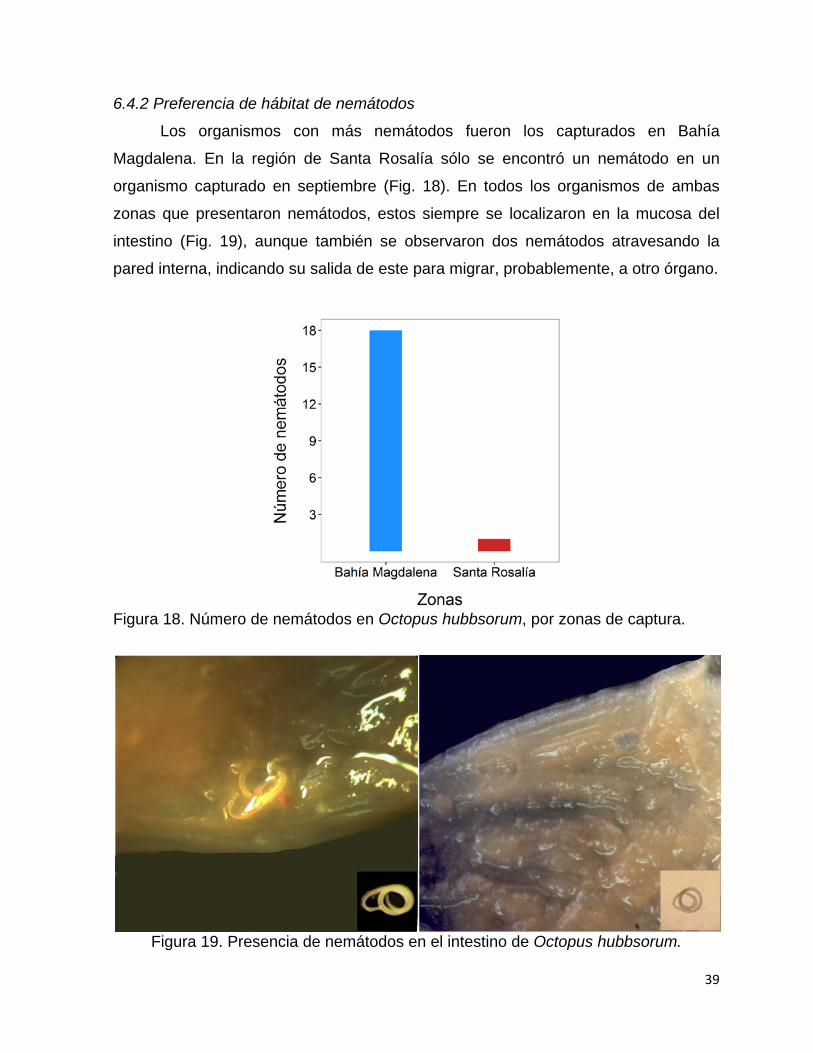
39
6.4.2 Preferencia de hábitat de nemátodos
Los organismos con más nemátodos fueron los capturados en Bahía
Magdalena. En la región de Santa Rosalía sólo se encontró un nemátodo en un
organismo capturado en septiembre (Fig. 18). En todos los organismos de ambas
zonas que presentaron nemátodos, estos siempre se localizaron en la mucosa del
intestino (Fig. 19), aunque también se observaron dos nemátodos atravesando la
pared interna, indicando su salida de este para migrar, probablemente, a otro órgano.
Figura 18. Número de nemátodos en Octopus hubbsorum, por zonas de captura.
Figura 19. Presencia de nemátodos en el intestino de Octopus hubbsorum.
40
6.5 Prevalencia, intensidad media y abundancia
De los 112 organismos capturados en ambas zonas, la prevalencia total de
protozoarios fue del 96%, mientras que la prevalencia total de nemátodos fue del 7%.
Los organismos capturados en la región de Santa Rosalía presentaron las
prevalencias más altas de protozoarios, principalmente en febrero, sin embargo, su
abundancia total fue menor que la reportada para los organismos de Bahía
Magdalena (Tabla 5).
Tabla 5. Datos totales de prevalencia, intensidad media y abundancia de quistes de Aggregata sp. en Octopus hubbsorum, por mes de muestreo.
Santa Rosalía Bahía
Magdalena
Septiembre Noviembre Febrero Octubre Febrero
Hospederos revisados 16 25 23 17 31
Hospederos parasitados 15 24 23 16 30
Número de parásitos 718 174 1587 1322 29922
Prevalencia (%) 93.75 96 100 94.12 96.77
Intensidad media 47.87 7.25 71.2 82.63 997.4
Abundancia 44.87 6.96 71 77.76 965.22
Por otro lado, los valores de prevalencia de nemátodos en ambas áreas de
estudio fueron muy bajos, sin embargo, en Bahía Magdalena la abundancia fue
mayor que el total obtenido en los cuatro meses de muestreo en Santa Rosalía,
donde únicamente se localizó un nemátodo en septiembre (Tabla 6).
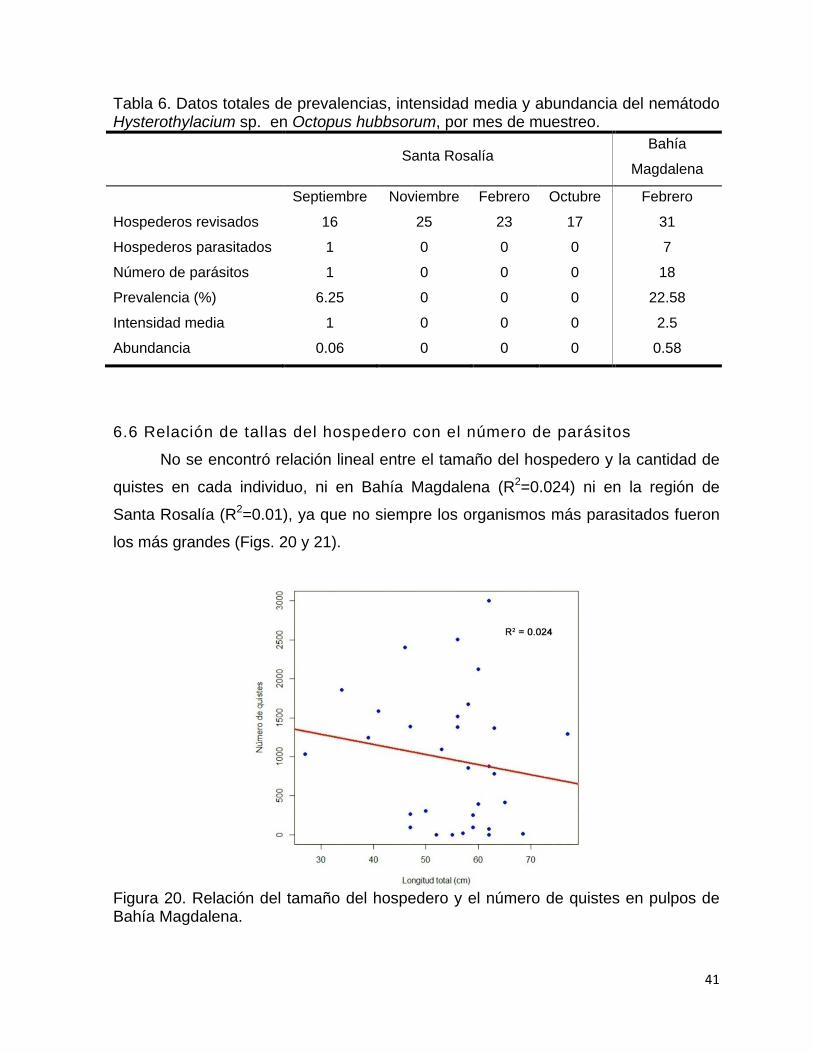
41
Tabla 6. Datos totales de prevalencias, intensidad media y abundancia del nemátodo Hysterothylacium sp. en Octopus hubbsorum, por mes de muestreo.
Santa Rosalía Bahía
Magdalena
Septiembre Noviembre Febrero Octubre Febrero
Hospederos revisados 16 25 23 17 31
Hospederos parasitados 1 0 0 0 7
Número de parásitos 1 0 0 0 18
Prevalencia (%) 6.25 0 0 0 22.58
Intensidad media 1 0 0 0 2.5
Abundancia 0.06 0 0 0 0.58
6.6 Relación de tallas del hospedero con el número de parásitos
No se encontró relación lineal entre el tamaño del hospedero y la cantidad de
quistes en cada individuo, ni en Bahía Magdalena (R2=0.024) ni en la región de
Santa Rosalía (R2=0.01), ya que no siempre los organismos más parasitados fueron
los más grandes (Figs. 20 y 21).
Figura 20. Relación del tamaño del hospedero y el número de quistes en pulpos de Bahía Magdalena.
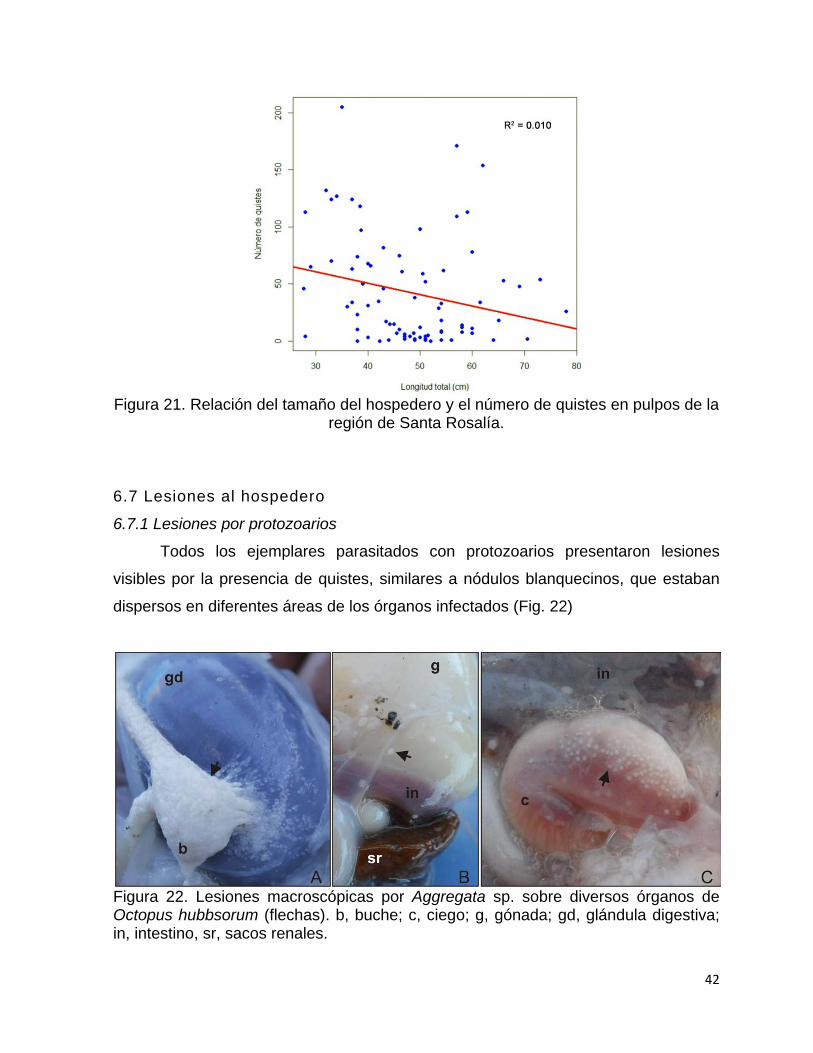
42
Figura 21. Relación del tamaño del hospedero y el número de quistes en pulpos de la
región de Santa Rosalía.
6.7 Lesiones al hospedero
6.7.1 Lesiones por protozoarios
Todos los ejemplares parasitados con protozoarios presentaron lesiones
visibles por la presencia de quistes, similares a nódulos blanquecinos, que estaban
dispersos en diferentes áreas de los órganos infectados (Fig. 22)
Figura 22. Lesiones macroscópicas por Aggregata sp. sobre diversos órganos de Octopus hubbsorum (flechas). b, buche; c, ciego; g, gónada; gd, glándula digestiva; in, intestino, sr, sacos renales.
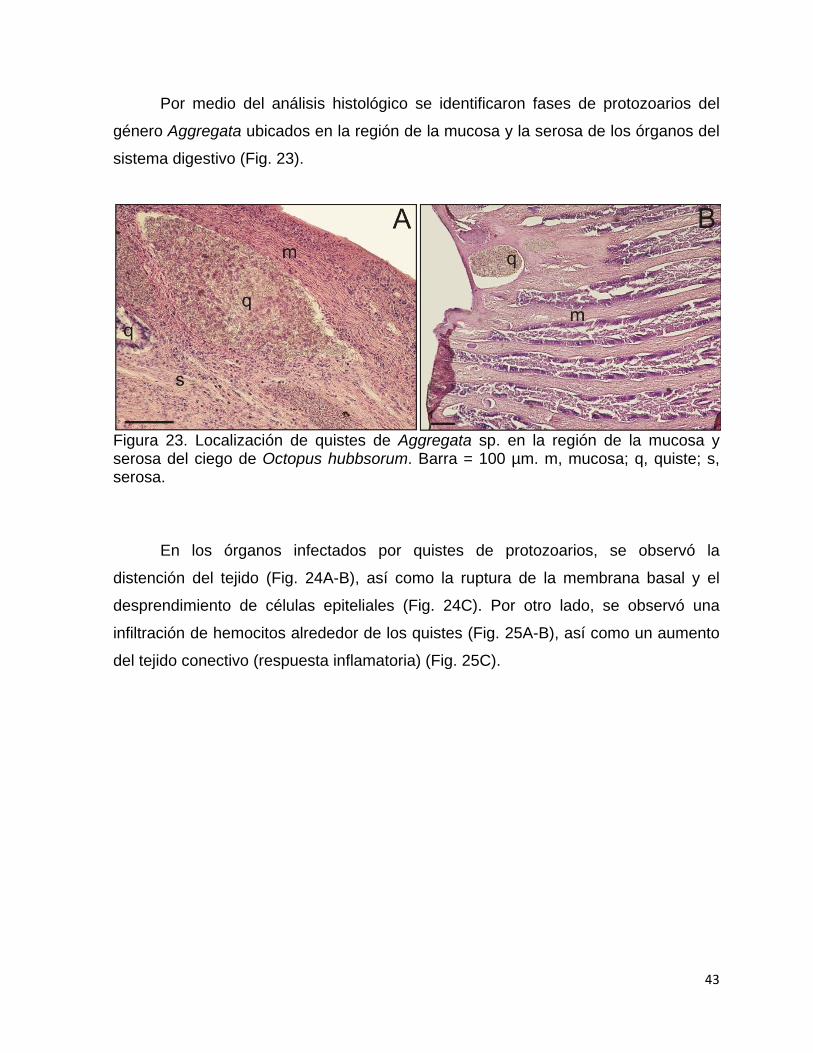
43
Por medio del análisis histológico se identificaron fases de protozoarios del
género Aggregata ubicados en la región de la mucosa y la serosa de los órganos del
sistema digestivo (Fig. 23).
Figura 23. Localización de quistes de Aggregata sp. en la región de la mucosa y serosa del ciego de Octopus hubbsorum. Barra = 100 µm. m, mucosa; q, quiste; s, serosa.
En los órganos infectados por quistes de protozoarios, se observó la
distención del tejido (Fig. 24A-B), así como la ruptura de la membrana basal y el
desprendimiento de células epiteliales (Fig. 24C). Por otro lado, se observó una
infiltración de hemocitos alrededor de los quistes (Fig. 25A-B), así como un aumento
del tejido conectivo (respuesta inflamatoria) (Fig. 25C).
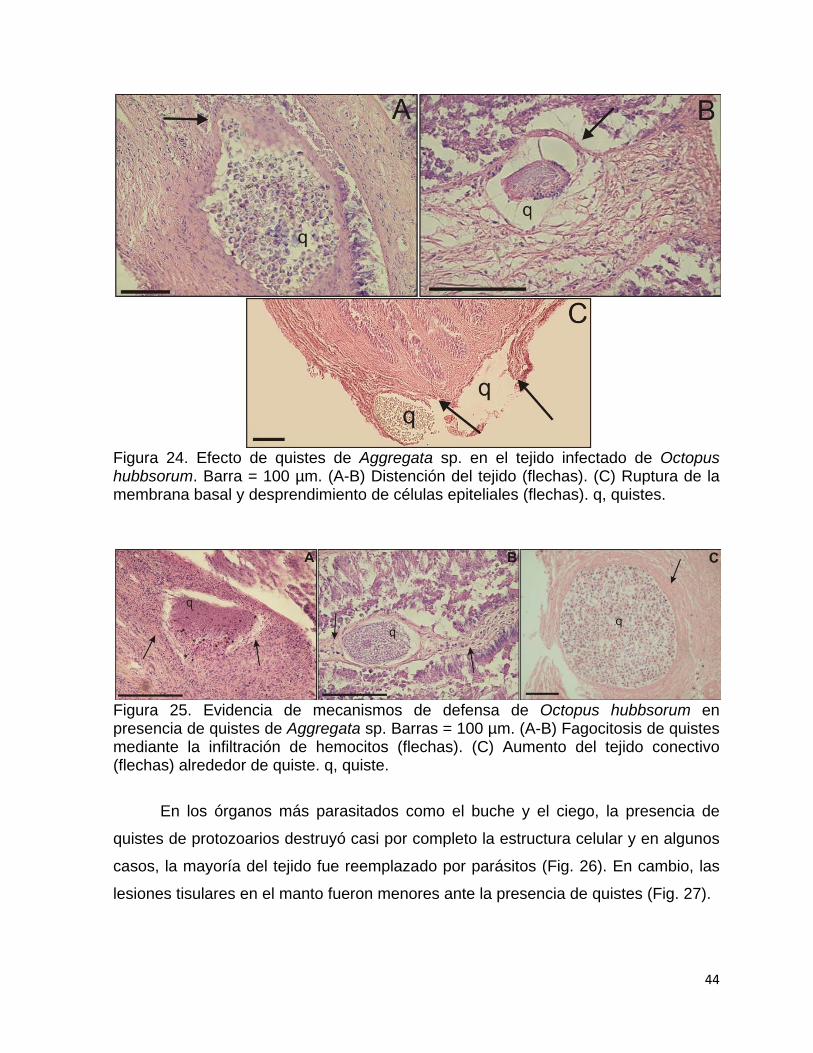
44
Figura 24. Efecto de quistes de Aggregata sp. en el tejido infectado de Octopus hubbsorum. Barra = 100 µm. (A-B) Distención del tejido (flechas). (C) Ruptura de la membrana basal y desprendimiento de células epiteliales (flechas). q, quistes.
Figura 25. Evidencia de mecanismos de defensa de Octopus hubbsorum en presencia de quistes de Aggregata sp. Barras = 100 µm. (A-B) Fagocitosis de quistes mediante la infiltración de hemocitos (flechas). (C) Aumento del tejido conectivo (flechas) alrededor de quiste. q, quiste.
En los órganos más parasitados como el buche y el ciego, la presencia de
quistes de protozoarios destruyó casi por completo la estructura celular y en algunos
casos, la mayoría del tejido fue reemplazado por parásitos (Fig. 26). En cambio, las
lesiones tisulares en el manto fueron menores ante la presencia de quistes (Fig. 27).
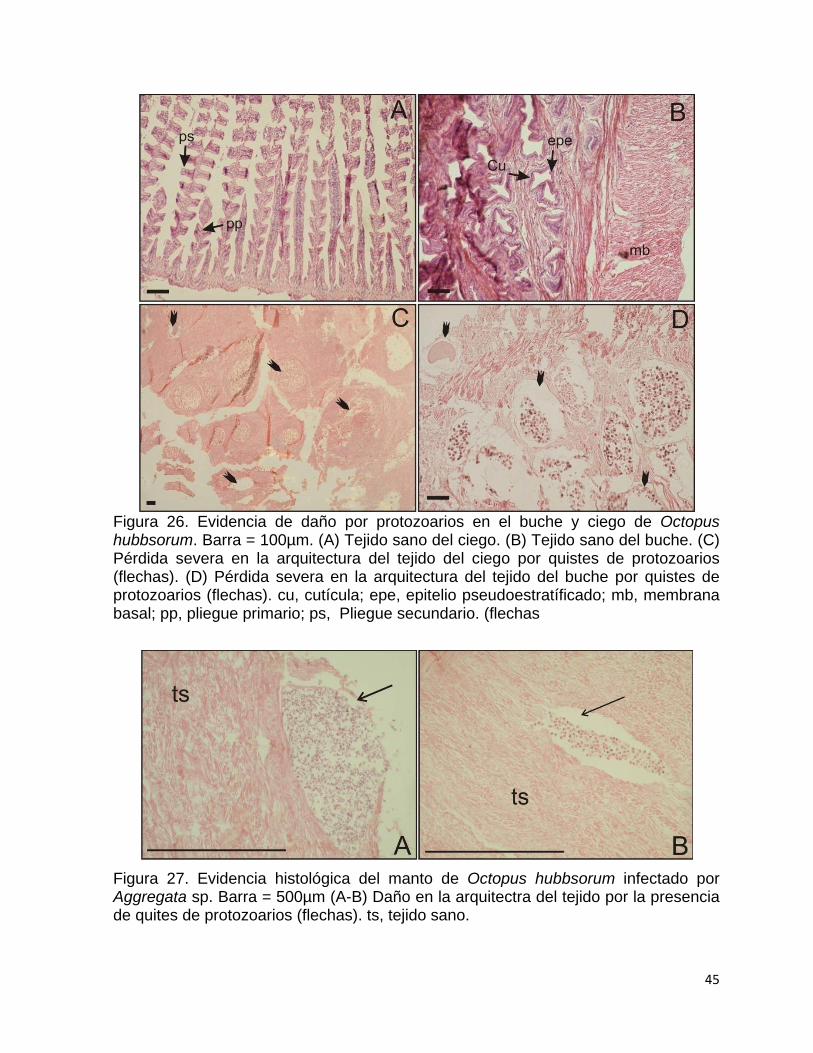
45
Figura 26. Evidencia de daño por protozoarios en el buche y ciego de Octopus hubbsorum. Barra = 100µm. (A) Tejido sano del ciego. (B) Tejido sano del buche. (C) Pérdida severa en la arquitectura del tejido del ciego por quistes de protozoarios (flechas). (D) Pérdida severa en la arquitectura del tejido del buche por quistes de protozoarios (flechas). cu, cutícula; epe, epitelio pseudoestratíficado; mb, membrana basal; pp, pliegue primario; ps, Pliegue secundario. (flechas
Figura 27. Evidencia histológica del manto de Octopus hubbsorum infectado por Aggregata sp. Barra = 500µm (A-B) Daño en la arquitectra del tejido por la presencia de quites de protozoarios (flechas). ts, tejido sano.
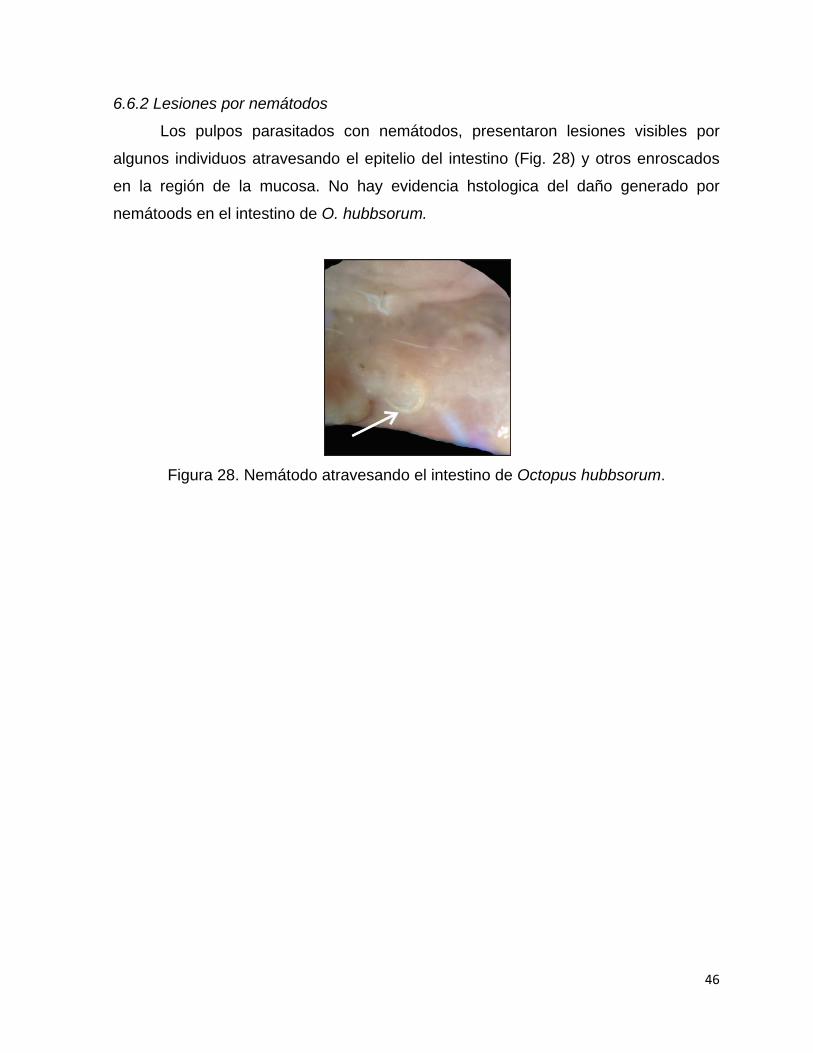
46
6.6.2 Lesiones por nemátodos
Los pulpos parasitados con nemátodos, presentaron lesiones visibles por
algunos individuos atravesando el epitelio del intestino (Fig. 28) y otros enroscados
en la región de la mucosa. No hay evidencia hstologica del daño generado por
nemátoods en el intestino de O. hubbsorum.
Figura 28. Nemátodo atravesando el intestino de Octopus hubbsorum.

47
7. DISCUSIÓN En relación a los meses de captura en la región de Santa Rosalía, la constante
del mal tiempo debido a fenómenos naturales evitó que los muestreos fueran
mensuales o temporales, lo que obligó a tomar en cuenta solo los meses donde se
facilitó la captura de organismos. Por otro lado, los elevados costos para las capturas
en Bahía Magdalena, hicieron obligada una sola visita a la isla.
Las principales zonas de captura de O. hubbsorum, forman parte del área de
distribución de pulpos como O. alecto, O. chierchia, O. digueti, O. penicillifer y O.
bimaculatus en el Golfo de California y de O. rubescens, O. veligero y O. bimaculatus
en la costa Pacífico de Baja California Sur (Roper et al., 1995; Norman et al., 2014), y
de las cuales no se tienen datos de su carga parasitaria. Por otro lado, Enteroctopus
dofleini es una especie de la que se tienen registros de parásitos protozoarios
(Poynton et al., 1992) y a su vez, comparte su distribución con O. rubescens y O.
bimaculatus al norte de California (Norman et al., 2014), debido a este tipo de
interacciones geográficas entre hospederos, los protozoarios del género Aggregata
son capaces de distribuirse cosmopolitamente y habitar en un gran número de
hospederos (Anexo).
Se han descrito alrededor de 17 especies de protozoarios del género
Aggregata, asimismo, se ha registrado la fase de merogonia en crustáceos
decápodos (Hansson, 1997) y portúnidos (Dobell, 1925) (Anexo), y la fase de
gametogonia y esporogonia en octópodos como O. vulgaris, en omastrépidos como I.
argentinus (paralarvas y juveniles) (Vidal, 1999), T. sagittatus (Gestal et al., 2000), y
en sépidos como S. officinalis (Hansson, 1997; Gestal et al., 2002a; Gestal et al.,
2002b), lo que concuerda con la presencia de estructuras representativas de las
fases de gametogonia y esporogonia en O. hubbsorum. En la fase esporogónica, las
características morfológicas como el tamaño y superficie de los esporocistos, así
como la longitud de los esporozoitos son determinantes de cada especie del género
Aggregata (Estévez et al., 1996). En este estudio, los esporocistos fueron más
grandes (15.7 a 26.4 µm) que los encontrados en A. octopiana (Estévez et al., 1996)
y T. sagittatus (Gestal et al., 2000), lo cual podría indicar que se trata de una especie
diferente. Por otro lado, para diferenciar entre especies, también es necesario tomar

48
en cuenta las características morfológicas de los esporozoitos, sin embargo, en este
trabajo no fue posible aislarlos. Los protozoarios descritos en la región de Santa
Rosalía y Bahía Magdalena, podrían pertenecer a la misma especie debido a la
preferencia del mismo hospedero (Rensch, 1938) y porque presentaron
características similares. Incluso, podría tratarse de una nueva especie, puesto que
los coccidios del género Aggregata, asociados al tracto digestivo de cefalópodos,
también son identificados de acuerdo al hospedero que habitan, debido a su alta
especificidad hospedatoria (Estévez et al., 1996; Peñalver et al., 2008).
El ingreso de coccidios en los cefalópodos, se debe al consumo de presas
infectadas, como crustáceos (Dobell, 1925), las cuales son necesarias para
completar el ciclo de vida del parásito. En el contenido visceral de O. hubbsorum se
observaron restos muy fragmentados de pulpos, valvas de moluscos, escamas y
espinas de peces, así como de anfípodos, copépodos y cangrejos, estos últimos
posiblemente pertenecientes a la familia de los porcelánidos de acuerdo a la dieta
descrita para O. hubbsorum en el Pacífico Mexicano (Alejo-Plata et al., 2009), ya que
se ha descrito el desarrollo de la fase de merogonia en porcelánidos (Dobell, 1925),
éstos podrían ser los transmisores del parásito en O. hubbsorum, sin embargo, no
se tomaron muestras de estos organismos.
Los principales órganos de O. hubbsorum parasitados por protozoarios fueron
el ciego, el intestino, el buche y la glándula digestiva, lo cual concuerda con lo
registrado en O. tehuelchus (Sardella & Ré, 1990), O. vulgaris (Estévez et al., 1996;
Gestal et al., 2002b; Cavaleiro, 2013; Linares et al., 2015), I. argentinus (Vidal, 1999),
T. sagittatus (Gestal et al., 2000), E. megalocyathus (Sardella et al., 2000; Ibáñez et
al., 2005), y S. officinalis (Linares et al., 2015), donde se observa que las principales
infecciones por coccidios en cefalópodos son desarrolladas en el tracto digestivo
(Estévez et al., 1996). Durante la digestión en pulpos, una vez que el alimento entra
en la boca, pasa a través de los órganos del sistema digestivo, donde por medio de
la liberación de enzimas digestivas, se llevará a cabo la absorción de nutrientes
(Linares et al., 2015), debido a esto, el sistema digestivo es el sitio ideal para el
establecimiento de Aggregata en cefalópodos. Por otro lado, de acuerdo a las
características del hábitat de los cefalópodos, se pueden presentar variaciones en los

49
procesos digestivos (Linares et al., 2015), por ejemplo, en S. officinalis la comida es
almacenada principalmente en el buche, mientras que en O. vulgaris el alimento se
almacena en el ciego, por lo que la absorción de nutrientes, será llevada a cabo en
estos órganos durante la mayor parte del proceso digestivo (Linares et al., 2015).
En la región de Santa Rosalía el mayor número de quistes se registró en el
ciego de los organismos capturados en febrero, debido a que los eventos de
surgencia aumentan en invierno (Rodríguez-Meza, 2004; Espinosa-Carreón &
Valdez-Holguín, 2007), lo cual traería como consecuencia mayor disponibilidad de
aliento. Por otro lado, el menor número de quistes se registró en noviembre, cuando
no se presentan grandes eventos de surgencias y la temperatura aumenta, sin
embargo, es necesario realizar muestreos mensuales para corroborarlo. En
comparación con los organismos capturados en la región de Santa Rosalía, el buche
de los pulpos de Bahía Magdalena fue el principal órgano parasitado, puesto que la
temperatura y la evidencia de surgencias estacionales fue similar en ambas zonas,
durante el mes de febrero, sugiriendo que los organismos se están alimentando y la
absorción de nutrientes se lleva a cabo principalmente en el buche.
Por otro lado, la presencia de quistes en el tejido muscular del manto de O.
hubbsorum es inusual, de acuerdo a lo registrado en O. vulgaris en el Río de Vigo,
España (Estévez et al., 1996) y en el Mar Adriático al sur de Europa (Mladineo &
Jozič, 2005). Por otra parte, debido a la frecuencia de infección de Aggregata sp. en
las branquias y las glándulas salivales de O. hubbsorum, en el análisis de clúster
fueron agrupados junto con los órganos del sistema digestivo. La presencia de
quistes en las branquias, concuerda con lo registrado en O. vulgaris (Peñalver et al.,
2008), y de acuerdo a Cavaleiro (2013) se debe a que funcionan como sitios
secundarios de infección por coccidios, cuando el ciego y el intestino están altamente
infectados, sin embargo, en la mayoría de los casos, las branquias estuvieron más
parasitadas cuando la infección en el intestino y el ciego fue menor, mientras que la
infección de las glándulas salivales está principalmente relacionada por su cercanía
al buche de O. hubbsorum y a la liberación de enzimas cuando el buche se
encuentra lleno de alimento (Linares et al., 2015). Por lo anterior, en órganos como
los sacos renales, los ojos, el sifón y tentáculos, la presencia de quistes puede ser

50
accidental por el contacto con otros órganos o fluidos del mismo pulpo. Asimismo, la
presencia de quistes en la gónada puede estar relacionada con su cercanía al tracto
digestivo, pues en la mayoría de los organismos con gónadas parasitadas, se
observó que la abundancia en el intestino era alta.
La elevada prevalencia (93%) de parásitos protozoarios fue común en ambos
lados de la península, similar a lo registrado por otros autores en octópodos del
Atlántico (Pascual et al., 1996), en costas españolas (Estévez et al., 1996; Peñalver
et al., 2008), y de Croacia (Mladineo & Bočina, 2007). Se ha observado que las
poblaciones de parásitos protozoarios prosperan de acuerdo al sistema inmune del
hospedero (Marcogliese, 2005), el cual se encuentra limitado en cefalópodos (Gestal
et al., 2002b), mientras que en otras poblaciones el incremento o decremento de
estos parásitos son el resultado de la contaminación directa de los hospederos
intermediarios (Marcogliese, 2005). En Santa Rosalía los valores de abundancia e
intensidad media fueron menores que en los organismos capturados en Bahía
Magdalena, posiblemente se deba a la contaminación por los residuos de la actividad
minera, los cuales más que tener un efecto negativo en los pulpos, podría estar
afectando a los hospederos intermediarios, evitando así la llegada de los parásitos al
hospedero definitivo.
Por otro lado, en paralarvas y juveniles de I. argentinus, Aggregata sólo se
observó en invierno (Vidal, 1999), en contraste con O. hubbsorum, donde se
encontraron en otoño e invierno, registrándose enla segunda temporada, los
menores valores de ntensidad y abundancia. Finalmente, en este trabajo, Aggregata
sp. es considerada especie núcleo por presentar valores de prevalencia mayores al
70% (Bush & Holmes, 1986). No hubo evidencia de especies secundarias, con
prevalencias entre el 40 y el 70% (Bush & Holmes, 1986), ni de especies satélite, con
valores de prevalencia menores al 40% (Bush & Holmes, 1986).
No se observó relación entre el número de quistes de protozoarios y la talla de
los organismos capturados en ambas zonas. Esto puede estar relacionado con los
hábitos alimentarios de cada individuo, donde se ha observado que las crías de
algunos pulpos se alimentan de zooplancton como isópodos, anfípodos y otras larvas

51
de crustáceos durante los primeros meses de vida (Rodríguez-Serna & Carmona-
Osalde, 2008).
Debido a la importancia pesquera que tienen los pulpos, es importante
conocer las relaciones parásito-hospedero que presentan en medio silvestre y con
ello poder describir posibles alteraciones en la salud del hospedador. La presencia
del parásito protozoario Aggregata en cefalópodos, está relacionada con daños a
nivel histopatológico los cuales varían de acuerdo al grado de infección (Gestal et al.,
2002a) y generan una degradación e hipertrofia celular, así como la ruptura de la
membrana basal y las células epiteliales (Gestal et al., 2002b). En O. hubbsorum, el
daño generado por el coccidio Aggregata sp. es similar al registrado en O. vulgaris
(Gestal et al., 2002a; Peñalver et al., 2008), por lo que es posible suponer que por el
alto grado de infección, en órganos como el buche y el ciego, le causan una
alteración en la absorción de nutrientes, así como una disminución en la actividad
enzimática (Gestal et al., 2002b), haciéndolo vulnerable a otros factores de estrés
(Castellanos-Martínez & Gestal, 2013; Storero & Narvarte, 2013). Por otra parte, ante
la presencia de patógenos, en el tejido del hospedero se llevan a cabo mecanismos
de defensa como la inflamación por aumento del tejido conectivo y la infiltración de
hemocitos que fagocitan a los patógenos (Gestal et al., 2002a), estos mecanismos
fueron observados en O. hubbsorum y son similares a los registrados en O. vulgaris
(Gestal et al., 2002b; Fichi et al., 2015).
La mayoría de los caracteres utilizados para la identificación de ascaroideos
se basa en estructuras de nemátodos adultos, y la información de larvas en
nemátodos de peces e invertebrados marinos, se centra en registros de presencia o
ausencia de individuos (Hochberg, 1990), lo que hizo complicada la identificación a
especie de los nemátodos encontrados en O. hubbsorum.
La presencia del nemátodo Hysterothylacium en cefalópodos ha sido
registrada en el Golfo de México (Anexo), donde se encontraron larvas de H.
reliquens en el calamar Lolliguncula brevis (Deardorff & Overstreet, 1981; Hochberg,
1990). La morfología de las larvas de nemátodos descritas en O. hubbsorum
concuerda con lo descrito por Anderson (1988) para larvas L3 del mismo género,
debido a la presencia de un ciego intestinal y un apéndice ventricular bien

52
desarrollados, y sobre todo porque el poro excretor se sitúa inmediatamente después
del anillo nervioso (Anderson, 1988). Por otro lado, O. hubbsorum participa como
hospedero intermediario o paraténico de Hysterothylacium sp., debido a que el
desarrollo del tercer o cuarto estadio larvario de ascaroideos ocurre principalmente
en hospederos paraténicos invertebrados, a excepción de algunas especies como
Hysterothylacium aduncum, donde los peces también participan como hospederos
intermediarios o paraténicos (Anderson, 1988).
Los estadios anteriores a L3 de Hysterothylacium, generalmente son ingeridos
por crustáceos como misidáceos, copépodos o isópodos (Anderson, 1988), sin
embargo, el conocimiento de su desarrollo y transmisión aún es escasa (Anderson,
1988). De acuerdo a lo anterior, en el contenido estomacal de O. hubbsorum se
observaron restos muy fragmentados de pulpos, valvas de moluscos, escamas y
espinas de peces, así como de crustáceos: anfípodos, copépodos y posiblemente
porcelánidos (Alejo-Plata et al., 2009), mismos que podrían corresponder al primer
hospedero intermediario del nemátodo.
La abundancia de nemátodos en los pulpos de Bahía Magdalena fue mayor
que la registrada en los organismos de la región de Santa Rosalía, esto se debe a
que Bahía Magdalena es un sitio rico en nutrientes debido a eventos de surgencia
costera (Álvarez-Borrego et al., 1975; Zaitsev et al., 2003), producidos por el flujo de
marea que genera corrientes a través de la boca de acceso en el complejo lagunar
Bahía Magdalena-Almejas (Funes-Rodríguez et al., 2007). Se ha observado que la
eutrofización en un ecosistema mejora la transmisión de algunos helmintos en
hospederos intermediarios, debido a que el sedimento aumenta y facilita la
transmisión entre hospederos, cuando este proceso es invertido los niveles de
infección disminuyen (Esch et al., 1986). La baja abundancia de parásitos nemátodos
en Santa Rosalía, puede deberse a factores de contaminación, debido a la actividad
minera, sobre los hospederos intermediarios, ya que en la mayoría de los parásitos
helmintos, se requiere de más de un hospedero para completar su ciclo de vida
(Marcogliese, 2005), asimismo, tales factores puede influir directamente sobre la fase
larvaria L2, pues estos son más vulnerables a cambios ambientales que sus
hospederos (Williams & MacKenzie, 2003 & Marcogliese, 2004).

53
O. hubbsorum podría tan solo participar como un hospedero ocasional de
nemátodos en Santa Rosalía, debido a la baja prevalencia (Sardella & Martorelli,
1997), sin embargo para confirmar esto, sería necesario llevar a cabo muestreos
mensuales por al menos un año. En cambio, la temperatura, la humedad, las
precipitaciones y la radiación solar, son factores que limitan la presencia, el
desarrollo y la distribución geográfica de parásitos, así como de los hospederos
(Gállego-Berenguer, 2006), por lo que son factores que tienen que ser tomados en
cuenta a futuro. Asimismo, la casi ausencia de parásitos nemátodos en O.
hubbsorum, puede deberse a que el tiempo de manipulación de los organismos en
campo fue demasiado, lo que pudo hacer que los organismos salieran del hospedero
debido a señales químicas relacionadas con el estrés y la muerte del pulpo. En este
trabajo, Hysterothylacium sp. es considerada especie satélite por tener valores de
prevalencia menores al 40% (Bush & Holmes, 1986). No hubo evidencia de especies
secundarias (prevalencia 40~70%), ni de especies núcleo (prevalencia > 70%) (Bush
& Holmes, 1986).
Ante la presencia de un patógeno, en el tejido celular se llevan a cabo
procesos inflamatorios como mecanismo de defensa (Marcogliese, 2005), mismos
que no fueron observados en el intestino en O. hubbsorum por la presencia de
nemátodos.
A pesar de que los cefalópodos participan como hospederos definitivos de
protozoarios, diciémidos y crustáceos, e intervienen como reservorios de larvas de
digéneos, céstodos, acantocéfalos y nemátodos (Kinne, 1990), en este trabajo no se
localizaron otros tipos de parásitos mesozoarios o metazoarios, posiblemente por el
tiempo de manipulación de los hospederos en campo y a la respuesta de estrés ante
la captura.

54
8. CONCLUSIONES El pulpo cumple un papel como hospedero definitivo para Aggregata sp., pues
en éste se lleva a cabo la fase sexual, evidenciada por la formación de
gamontes y la formación del cigoto.
El pulpo participa como hospedero intermediario o paraténico de
Hysterothylaciuim sp., pues en éste se desarrolla la fase larvaria L3 del
nemátodo.
Los protozoarios y nemátodos son considerados endoparásitos de O.
hubbsorum.
Aggregata sp. habita principalmente en el ciego de los pulpos de Santa
Rosalía y en el buche de los organismos de Bahía Magdalena.
Los nemátodos Hysterothylacium sp. habitan en el intestino de O. hubbsorum.
La prevalencia de protozoarios fue mayor en todos los meses de captura, la
intensidad media y la abundancia en febrero fue mayor en Bahía Magdalena,
mientras que en Santa Rosalía fueron mayores en octubre.
La prevalencia, intensidad media y la abundancia de nemátodos fue mayor en
Bahía Magdalena.
No existe una relación entre la cantidad de parásitos y el tamaño de los
pulpos.
La presencia de protozoarios causaron daño celular en los diversos tejidos de
O. hubbsorum.

55
9. RECOMENDACIONES
Para comprender mejor la participación de parásitos en el pulpo, es necesario
mejorar la metodología llevada a cabo, así como las condiciones para facilitar el
análisis de los organismos conforme a los resultados obtenidos. Por lo que se
sugiere:
Llevar a cabo muestreos mensuales durante un año, con la finalidad de que se
pueda reflejar el comportamiento estacional de los parásitos.
Se recomienda transportar los organismos completos para su revisión
detallada en laboratorio.
Analizar el contenido donde permanecieron los pulpos en campo, pues
durante el tiempo post mortem es posible perder evidencia de helmintos
dentro de éste.
Considerar la influencia de las variables ambientales como la temperatura, ya
que con esto se puede facilitar la interpretación de los datos de prevalencia en
las áreas de estudio.
Se requiere calcular el número de esporozoitos por quiste, para tener mejores
datos de abundancia e intensidad media parasita, utilizando una cámara
Neubauer.

56
10. LITERATURA CITADA Aguilar, S.C. & E. Godínez-Domínguez. 1997. Presencia del pulpo Octopus
hubbsorum (Cephalopoda: Octopoda) en el Pacífico central mexicano. Rev.
Biol. Trop., 45:253-260.
Alejo-Plata, M.C., J.L. Gómez-Márquez, S. Ramon-Carrillo & J.E. Herrera-Galindo.
2009. Reproducción, dieta y pesquería del pulpo Octopus (Octopus)
hubbsorum (Mollusca: Cephalopoda) en la costa de Oaxaca, México. Rev. Bio.
Trop., 57:63-78.
Álvarez-Borrego, S., A. Galindo-Beet & C. Chee-Barragan. 1975. Características
hidroquímicas de Bahía Magdalena, B. C. S. Cienc. Mar., 2:94-109.
Anderson, R.C. 1988. Nematode transmission patterns. J. Parasitol., 74:30-45.
Anderson, R.C. 2000. Nematode Parasites of Vertebrates: Their development and
transmission. CABI Publishing, London. 650 p.
Arce-Acosta, M. 2015. Interacciones entre pesquerías ribereñas en el corredor Santa
Rosalía-Mulegé, Baja California Sur, México. Tesis de Maestría. CICIMAR-
IPN, México. 89 p.
Audicana, M. T. & M. W. Kennedy. 2008. Anisakis simplex: from Obscure Infectious
Worm to Induce of Immune Hypersensitivity. Clin Microbiol Rev., 21:360-379.
Berry, S.S. 1953. Preliminary diagnoses of six west American species of Octopus.
Leafl. Malacol., 1:51-58.
Bush, A.O. & J.C. Holmes, 1986. Intestinal helmints of lesser scaup ducks: patterns
of association. Can. J. Zool., 64:132-141.
Bush, A.O., K.D. Lafferty, J.M. Lotz & A.W. Shostak. 1997. Parasitology metes
ecology on its own terms: Margolis et al. revised. J. Parasitol., 83:575-583.
Cavaleiro, F. 2013. Parasite fauna of Octopus vulgaris (Cephalopoda: Octopodidae)
and Platichthys flesus (Actinopterygii: Pleuronectidae): morphology,
systematics, life history strategies and ecology. Tesis de doctorado. PORTO,
Portugal. 188 p.
Cannon, L.R.G. 1977. Some ecological relationships of larval ascaroids from south-
eastern Queensland marine fishes. Int. J. Parasitol., 7:227-232.

57
Carella, F., S.W. Feist, J.P. Bignell & G. De Vico. 2015. Comparative pathology in
bivalves: Aetiological agents and disease processes. J. Invertebr. Pathol.,
131:107-120.
Castellanos-Martínez, S., M.C. Gómez, F.G. Hochberg, C. Gestal & H. Furuya. 2011.
A new dicyemid from Octopus hubbsorum (Mollusca: Cephalopoda:
Octopoda). J. Parasitol., 97:265-269.
Castellanos-Martínez, S. & C. Gestal. 2013. Pathogens and immune response of
cephalopods. J. Exp. Mar. Biol. Ecol., 447:14-22.
CONAPESCA. 2013. Anuario Estadístico de Acuacultura y Pesca 2013. Comisión
Nacional de Acuacultura y Pesca, México. 295 p.
Cook, T., M. Folli, J. Klinck, S. Ford & J. Miller. 1998. The relationship between
increasing sea-surface temperatura and the northward spread of Perkinsus
marinus (Dermo) disease epizootics in oysters. Estuar. Coast. Shelf Sci.,
46:587-597.
Coordinación General de Minería. 2008. Panorama minero del estado de Baja
California Sur. Servicio Geológico Mexicano, México. 39 p.
Cruz-Reyes, A. & B. Camargo-Camargo. 2001. Glosario de términos en parasitología
y ciencias afines. Plaza y Valdés, S. A. de C. V., México. 349 p.
Deardorff, T.L. & R.M. Overstreet. 1981. Larval Hysterothylacium (=Thynnascaris)
(Nematoda: Anisakidae) from Fishes and Invertebrates in the Gulf of Mexico.
Proc. Helminthol. Soc. Wash., 48:113-126.
Dobell, C. 1925. The life-history and chromosome cycle of Aggregata eberthi
[Protozoa: sporozoa: coccidia]. J. Parasitol., 17:1-136.
Domínguez-Contreras, J.F. 2011. Reproducción del pulpo Octopus hubbsorum Berry,
1953 en Bahía Magdalena, B.C.S., México. Tesis de Maestría. UNAM, México.
70 p.
Domínguez-Contreras, J.F., B.P. Ceballos-Vázquez, F.G. Hochberg & M. Arellano-
Martínez. 2013. A new record in a well-established population of Octopus
hubbsorum (Cephalopoda: Octopodidae) expands its know geographic
distribution range and maximum size. Am. Malacol. Bull., 31:95-99.
Micronet (Ed.). 2016. ENCICLONET. Micronet S.A. http://www.enciclonet.com

58
Esch, G.W., T.C. Hazen, D.J. Marcogliese, T.M. Goater & A.E. Crews. 1986. A long-
term study on the population biology of Crepidostomum cooperi (Trematoda:
Allocreadidae) in the Burrowing Mayfly, Hexageia limbata (Ephemeroptera).
Am. Midl. Nat., 116:304-314.
Espinosa-Carreón, T.L. & J.E. Valdez-Holguín. 2007. Variabilidad interanual de
clorofila en el Golfo de California. Ecol. Apl., 6:83-92.
Estévez, J., S. Pascual, C. Gestal, M. Soto, H. Rodríguez & C. Arias. 1996.
Aggregata octopiana (Apicomplexa: Aggregatidae) from Octopus vulgaris off
NW Spain. Dis. Aquat. Org., 27:227-231.
Estévez, J., H. Rodríguez, M. Soto & C. Arias. 1998. Cross-reactivity between
Aggregata octopiana Schneider, 1875 and Aggregata eberthi Labbé, 1895
(Apicomplexa, Aggregatidae): two coccidian parasites of cephalopods. J Fish
Dis., 21:309-312.
Fichi, G., G. Cardeti, S. Perrucci, A. Vanni, A. Cersini, C. Lenzi, T. De Wolf, B. Fronte,
M. Guarducci & F. Susini. 2015. Skin lesion-associated pathogens from
Octopus vulgaris: first detection of Photobacterium swingsii, Lactococcus
garvieae and betanodavirus. Dis. Aquat. Org., 115: 147-156.
Food and Agriculture Organization of the United Nations. 2014. Multicriteria-based
ranking for risk management of food-borne parasites. Microbiol. Risk Asst.
Series, 23:63-65.
Forsythe, J.W., R.T. Hanlon, R.A. Bullis & E.J. Noga. 1991. Octopus bimaculoides
(Pickford & McConnaughey, 1949): a marine invertebrate host for ectoparasitic
protozoans. J. Fish Dis., 14:431-422.
Funes-Rodríguez, R., J. Gómez-Gutiérrez & R. Palomares-García. 2007. Estudios
ecológicos en Bahía Magdalena. Instituto Politécnico Nacional, México. 311 p.
Furuya, H., R.T. Damian & F.G. Hochberg. 2002. Dicyema shorti n. sp. (Phylum
Dicyemida) from Octopus burry (Mollusca: Cephalopoda: Octopodidae) in the
Gulf of Mexico. J. Parasitol., 88:325-329.
Gállego-Berenguer, J. 2006. Manual de parasitología: Morfología y biología de los
parásitos de interés sanitario. Universitat de Barcelona, Barcelona. 516 p.

59
Gestal, C., E. Abollo & S. Pascual. 1999. Evaluation of a method for isolation and
purification of sporocysts of the cephalopod coccidian parasite Aggregata
Frenzel, 1885 (Apicomplexa: Aggregatidae). Iberus, 17:115-121.
Gestal, C., A. Guerra, E. Abollo & S. Pascual. 2000. Aggregata sagittata n. sp.
(Apicomplexa: Aggregatidae), a coccidian parasite from the European flying
squid Todarodes sagittatus (Mollusca: Cephalopoda). Syst. Parasit., 47:203-
206.
Gestal, C., E. Abollo & S. Pascual. 2002a. Observations on associated histopathology
with Aggregata octopiana infection (Protista: Apicomplexa) in Octopus vulgaris.
Dis. Aquat. Org., 50:45-49.
Gestal, C., M. Páez de la Cadena & S. Pascual. 2002b. Malabsorption syndrome
observed in the common octopus Octopus vulgaris infected with Aggregata
octopiana (Protista: Apicomplexa). Dis. Aquat. Org., 51:61-65.
Global Names Index (Ed.). 2016. Index of Scientific Names. http://gni.globalnames.
org/namestrings?search_term= aggregata&commit=Search.
González, A.F., S. Pascual, C. Gestal, E. Abollo & A. Guerra. 2003. What makes a
cephalopod a suitable host for parasite?. Fish Res., 60:177-183.
Gotshall, D.W. 1987. Marine animals of Baja California: a guide to the common fishes
and invertebrates. Sea Challenger, Monterey. 112 p.
Hansson, H.G. 1997. South Scandinavian marine protoctista. TJÄRNÖLAB,
Strömstad. 106 p.
Hochberg, F.G. 1990. Diseases caused by protistans and metazoans. 47-202. En: O.
Kinne (Ed.). Diseases of marine animals, Volume III. Biologische Anstalt
Helgoland, Hamburg. 696 pp.
Humason, G.L. 1979. Animal Tissue Techniques. W.H. Freeman y Co., San
Francisco. 661 p.
Ibáñez, C.M., M.C. Pardo-Gandarillas & M.G. Nascimento. 2005. Uso del
microhábitat por el protozoo parásito Aggregata patagónica Sardella, Ré &
Timi, 2000 (Apicomplexa: Aggregatidae) en hospedador definitivo, el pulpo
Enteroctopus megalocyathus (Gould, 1852) (Cephalopoda: Octopodidae) en el
surde Chile. Rev. Chil. de Hist. Nat., 78:41-450.

60
Keen, A.M. 1971. Sea shells of tropical west America. Stanford University Press,
Stanford. 1064 p.
Kinne, O. 1990. Diseases of marine animals, Volume III. Biologische Anstalt
Helgoland, Hamburg. 696 p.
Lee, J.J., G.F. Leedale & P. Bradbury. 2000. An illustrated guide to the protozoa.
Society of Protozoologists, Lawrence Kansas. 689 p.
Linares, M., C. Caamal-Monsreal, A. Olivares, A. Sánchez, S. Rodríguez, O. Zúñiga,
C. Pascual, P. Gallardo & C. Rosas. 2015. Timing of digestion, absorption and
assimilation in octopus species from tropical (Octopus maya) and subtropical-
temperate (O. mimus) ecosystems. Aquat. Biol., 24:127-140.
Lluch-Belda, D., M.E. Hernández-Rivas, R. Saldierna-Matínez & R. Guerrero-
Caballero. 2000. Variabilidad de la temperatura superficial del mar en Bahía
Magdalena, B. C. S. Oceánides, 15:1-23.
Lom, J. & I. Dyková. 1992. Protozoan parasites of fishes. Dev. Aquac. Fish. Sci.,
26:1-315.
López-Uriarte, E., E. Ríos-Jara & M. Pérez-Peña. 2005. Range extension for Octopus
hubbsorum (Mollusca: Octopodidae) in the Mexican Pacific. Bul. Mar. Sci.,
77:171-176.
López-Uriarte, E. 2006. Ciclo vital y pesca del pulpo Octopus hubbsorum Berry 1953
(Cephalopoda: octopodidae) en la costa de Jalisco, México. Tesis DE
Doctorado. CUCBA-UDG., México. 175 p.
Magic sea weed, 2016. http://es.magicseaweed.com/Mexico-Pacific-MSW-Surf-
Charts/19/
Marcogliese, D.J. 2004. Parasites: small players with crucial roles in the ecological
theatre. EcoHealth., 1:151-164.
Marcogliese, D.J. 2005. Parasites of the superorganism: Are they indicators of
ecosystem health? Int. J. Parasitol., 35:705-716.
Mladineo, I. & I. Bočina. 2007. Extraintestinal gamogony of Aggregata octopiana in
the reared common octopus (Octopus vulgaris) Cephalopoda: Octopodidae). J.
Invertebr. Pathol., 96:261-264.

61
Mladineo, I. & M- Jozič. 2005. Aggregata infection in the common octopus, Octopus
vulgaris (Linnaeus, 1758), Cephalopoda: Octopodidae, reared in a flow-
through system. Acta Adriática, 44:193-199.
Morales, G.A., L.A. Pino, D. Graterol & L. Perdomo. 1991. Técnica de muestreo para
la estimación de la abundancia y la prevalencia de nematodos parásitos en
bovinos. Revista Científica FCV de LUZ, 2:46-50.
Norman, M.D., J.K. Finn & F.G. Hochberg. 2014. Family Octopodidae d’Orbigny,
1840. 36-216. En: P. Jereb, C.F.E. Roper, M.D. Norman & J.K. Finn. (Ed.)
Cephalopods of theworld. An annotated and illustrated catalogue of
cephalopod species know to date. Volume 3. Octopods and Vampire Squids.
FAO species catalogue for fishery purposes, 4. FAO, Rome. 370 p.
Norris, D.E. & R.M. Overstreet. 1976. The public health implications of larval
Thynnascaris nematodes from shellfish. J. Milk Food Technol., 38:769-773.
Olsen, O.W. 1974. Animal parasites, their life cycles and ecology. Dover Publications,
Inc., New York, New York. 562 p.
Pascual, S., C. Gestal, J.M. Estévez, H. Rodríguez, M. Soto, E. Abollo & C. Arias.
1996. Parasites in commercially-exploited cephalopods (Mollusca,
Cephalopoda) in Spain: an update perspective. Aquaculture, 142:1-10.
Pascual, S., C. Gestal & E. Abollo. 1997. Effect of Pennella sp. (Copepoda,
Penellidae) on the condition of Illex coindetii and Todaropsis eblanae
(Cephalopoda, Ommastrephidae). Bull. Eur. Ass. Fish Pathol., 17:91-95.
Peñalver, J., E. MaríaDolores, P. Muñoz, J. Cerezo, B. García & E. Viuda. 2008.
Valoración sobre la presencia y el control sanitario del coccidio Aggregata
octopiana en el pulpo común procedente de acuacultura. An. Vet., 24:57-62.
Perkins, F.O., J.R. Barta, R.E. Clopton, M.A. Peirce & S.J. Upton. 2000. Phylum
Apicomplexa. 190-369. En: J.J. Lee, G.F. Leedale & P. Bradbury. (Eds.). An
Illustrated guide to the protozoa, Volume I. Society of Protozoologists,
Lawrences. 689 p.
Poynton, S.L., R. Reimschuessel & M.K. Stoskopf. 1992. Aggregata dobelli n. sp. and
Aggregata millerorum n. sp. (Apicomplexa: Aggregatidae) from two species of

62
Octopus (Mollusca: Octopodidae) from the Eastern North Pacific Ocean. J.
Protozool., 39:248-256.
Ramírez-Rodríguez, M. & A. Hernández-Herrera. 2000. Pesca artesanal en la costa
oriental de Baja California Sur, México (1996-1997). 18-29. En: O. Aburto-
Oropeza & C. Sánchez-Ortíz. (Eds.). Recursos arrecifales del Golfo de
California. UABCS, México.
Rensch, B. 1938. Some problems of geographical variation and species-formation.
Proc. Linn. Soc. Lond., 150:275-285.
Rhode, K. 2005. Marine Parasitology. CSIRO Publishing, New England. 592 p.
Rodríguez-Meza, G.D. 2004. Caracterización geoquímica por componentes mayores
y elementos traza de sedimentos de los ambientes marinos costeros
adyacentes a la Península de Baja California. Tesis de Doctorado. CICIMAR-
IPN, México. 227 p.
Rodríguez-Serna, M. & C. Carmona-Osalde. 2008. Hidrobiológica, 18:209-214.
Roper, C.F.E., M.J. Sweeney & F.G. Hochberg. 1995. Cefalópodos. 305-353. En: W.
Fischer, F. Krupp, W. Schneider, C. Sommer, K.E. Carpenter & V.H. Niem.
(Ed.). Guía FAO para la identificación de especies para los fines de la pesca.
Pacífico centro-oriental, I, plantas e invertebrados. FAO, Roma. 646 p.
Rosa, R., P.R. Costa & M.L. Nunes. 2004. Effect of sexual maturation on the tissue
biochemical composition of Octopus vulgaris and O. defilippi (Mollusca:
Cephalopoda). Mar. Biol., 145:563-574.
Sánchez-García, A. M. 2013. Estudio biológico pesquero de Octopus spp. en el
complejo lagunar Bahía Magdalena-Almejas, B.C.S. México. Tesis de
Licenciatura. Universidad Autónoma de Baja California Sur, México. 70 p.
Sardella, N.H. & M.E. Ré. 1990. Parasitosis por coccidios del género Aggregata en
pulpos costeros patagónicos. I. Aggregata sp. en Octopus tehuelhus
D’Orbigny. Physis. A., 46: 51-60.
Sardella, N.H. & S.R. Martorelli. 1997. Ocurrence of merogony of Aggregata Frenzel
1885 (Apicomplexa) in Pleoticus muelleri and Artemesia longinaris (Crustacea:
Natantia) form Patagonian Waters (Argentina). J. Invertebr. Pathol., 70:198-
202.

63
Sardella, N.H., M.E. Ré & J.T. Timi. 2000. Two new Aggregata species (Apicomplexa:
Aggregatidae) infecting Octopus tehuelchus and Enteroctopus megalocyathus
(Mollusca: Octopodidae) in Patagonia, Argentina. J. Parasitol., 86:1107-1113.
SEMARNAT, 2009. Programa de conservación y manejo de la Reserva de la
Biósfera: Alto Golfo de California y Delta del Río Colorado. D.O.F., México.
135 p.
Shumilin, E., Á.R. Jiménez-Illescas & S. López-López. 2013. Anthropogenic
contamination of metals in sedimens of the Santa Rosalía Harbor, Baja
California Peninsula. Bull. Environ. Contam. Toxicol., 90:333-337.
Sprague, V. & J. Couch. 1971. An annotated list of protozoan parasites,
hyperparasites, and commensals of decapod crustacea. J. Protozool., 18: 526-
537.
Storero, L.P. & M.A. Narvarte. 2013. Coccidian infection may explain the differences
in the life history of Octopus host populations. J. Invertebr. Pathol., 114:222-
225.
Universidad Nacional Autónoma de México (Ed.). 2011. Glosario. Departamento de
microbiología y parasitología.
http://www.facmed.unam.mx/deptos/microbiologia/glosario.html
Vidal, E.A.G. 1999. Digestive tract parasites in rhynchoteuthion squid paralarvae,
particulary in Illex argentinus (Cephalopoda: Ommastrephidae). Fish. Bull.,
97:402-405.
Williams, H.H. & K. Mackenzie. 2003. Marine parasites as pollution indicators: an
update. J. Parasitol., 126:S27-S41.
WoRMS Editorial Board (Ed.). 2016. World Register of Marine Species. VLIZ.
http://www.marinespecies.org/aphia.php?p=taxlist.
Zaitsev, O., R. Cervantes-Duarte, O. Sánchez-Montante & A. Gallegos-García. 2003.
Coastal upwelling activity of the Pacific shelf of Baja California península.
Oceanography, 59:489-502.

64
11. ANEXO Preparación de Finefix:
- FineFix 280 ml
- Alcohol absoluto 720 ml
Agregar el finefix al alcohol absoluto. Mezclar suavemente y almacenar etiquetado. Hospederos y distribución de protozoarios del género Aggregata:
Hospederos: Eledone massyae (Voss), ‘pulpo desflecado’; E. dofleini martini
(Pickford), ‘pulpo gigante’; E. megalocyathus (Gould), ‘pulpo gigante de Patagonia’;
O. bimaculoides (Pickford and McConnaughey), ‘pulpo con dos manchas’; O.
tehuelchus (d’Orbigny), ‘pulpo tehuelche’; O. vulgaris (Cuvier), ‘pulpo común’;
(Octopodidae). Martialia hyadesi (Gestal, Nigmatullin, Hochberg, Guerra &
Pascual); Todarodes sagittatus (Lamark), ‘calamar volador común’; Todaropsis
eblanae (Ball), ‘calamar volador menor’, (Ommastrephidae). Sepia officinalis
(Férussac), ‘sepia común’; S. elliptica (Hoyle), (Sepiidae). Atelecyclus rotundus
(Olivi), (Atelecyclidae). Carcinus aestuarii (C. mediterraneus) (Nardo), ‘cangrejo
mediterraneo común’, (Carcinidae). Corystes cassivelaunus (Pennant), (Corystidae).
Dromia personata (Linnaeus), (Dromiidae). Goneplax rhomboides (Linnaeus),
(Goneplacidae). Pachygrapsus marmoratus (Fabricius), ‘cangrejo corredor o
zapatero’, (Grapsidae). Acanthephrya eximia (J.J. Sm), (Hoplophoridae). Inachus
communissimus (Rizza); I. dorsettensis (Pennant), ‘araña de mar’; Macropodia
rostrata (Linnaeus), ‘cangrejo araña’, (Inachidae). Eupagurus prideauxi (Leach); E.
sp. (Frenzel), (Paguroidea). Leander squilla (Rathke), ‘camarón báltico’;
(Palaemonidae). Parthenope angulifrons (Latreille), (Parthenopidae). Artemesia
longinaris (Bate), ‘camarón argentino’; Solenocera membranácea (Risso);
Parapeneopsis sculptilis (Heller), (Penaeidae). Pilumnus spinifer (H. Milne Edwards);
P. hirtellus (Linnaeus), (Pilumnidae). Pinnotheres pinnotheres (Bosc); P. pisum
(Linnaeus)’ cangrejo guisante’, (Pinnotheridae). Macropipus depurator (Linnaeus); M.
corrugatus (Pennant); M. puber (Linnaeus); M. vernalis (Risso), ‘cangrejo nadador
gris’, (Polybiidae). Carcinus maena (Linnaeus), ‘cangrejo verde’; Portunus arcuatus

65
(Leach); P. depurator (Linnaeus); P. latipes (Pennant), (Portunidae). Sergestes
robustus (Smith), (Sergestidae). Pleoticus muelleri (Bate), ‘langostino patagónico’,
(Solenoceridae). Xantho poressa (Olivi), (Xanthidae).
Localidades: Costa de Bahía Magdalena y región de Santa Rosalía, Baja
California Sur, México. Allin Island, Long Island y Tacoma en Washington, USA.
Aguas del Pacífico americano, California y Canadá. Golfo San Matias, Golfo San
José, Golfo Nuevo y puerto Rawson en Argentina. Ancud y Quellón, Chiolé, X
Región, Chile. Island of Brač, Mar Adriático. Ría of Vigo, zona marítima limítrofe
entre Murcia y Alicante, costa mediterránea, noroeste y noreste de España.
Suroeste del Océano Atlántico. Nápoles, Italia. English Chanel y Banyuls Francia.
Bombai, India.
Hospederos, distribución y sitios de infección de nemátodos del género Hyterothylacium.
Hospederos: Paralichthys albigutta (Jordan & Gilbert), ‘lenguado del golfo’; P.
lethostigma (Jordan & Gilbert 1884), ‘lenguado de sur’ (Bothidae); Caranx crysos
(Mitchill), corredor azul; Trachinotus carolinus (Linnaeus), ‘pompano Florida’,
(Carangidae); Fundulus grandis (Baird & Girard), Gulf killifish, F. heterodltus
(Linnaeus), mummichog (Cyprinodontidae); Dactyloscopus moorei (Fowler),
stargazer (Dactyloscopidae); Anchoa hepsetus (Linnaeus), striped anchovy,
(Mitchilli) (Valenciennes), anchoa de bahía (Engraulidae); Chaetodipterus faber
(Broussonet), Atlantic spadefish (Ephippidae); Eucinostomus argenteus (Baird &
Girard), spotfin mojarra (Gerreidae); Lutjanus campechanus (Poey), red snapper
(Lutjanidae); Mugil cephalus (Linnaeus), striped mullet (Mugilidae); Ophichthus
gomesi (Castelnau), shrimp eel (Ophichthidae); Polydactylus octonemus (Girard),
Atlantic threadfin (Polynemidae); Oncorhynchus kisutch (Walbaum), salmón coho, O.
mykiss (Walbaum), trucha arcoíris, Salmo salar (Linnaeus), salmón del Atlántico
(Salmonidae); Cynoscion arenarius (Ginsburg), trucha de mar y arena, C. nebulosus
(Cuvier), spotted seatrout, Menticirrhus americanus (Linnaeus), pez real del sur,
Micropogonias imdulatus (Linnaeus), corvina del atlántico, Sciaenops ocellata
(Linnaeus), tambor rojo, Stellifer lanceolatus (Holbrook), tambor estrella

66
(Sciaenidae); Scomberomorus maculatus (Mitchill), jurel español (Scombridae);
Centropristis philadelphica (Linnaeus), lubina roca, Diplectrum formosiim (Linnaeus),
perca de arena, Epinephalus morio (Valenciennes), mero rojo (Serranidae);
Archosargus probatocephalus (Walbaum), cabeza de oveja (Sparidae); Peprilus
alepidotus (Linnaeus), harvestfish, P. burti (Fowler), Gulf butterfish (Stromateidae);
Trichiurus lepturus (Linnaeus), Atlantic cutlassfish (Trichiuridae); Prionotus scitulus
(Jordan and Gilbert), leopard searobin (Triglidae). Invertebrates: Cantharus
cancellarius (Conrad), cancellate cantharus (Gastropoda); Lolliguncula brevis
(Blainville), brief thumbstall squid (Cephalopoda); Callinectes sapidus (Rathbun),
cangrejo azul, Clibanarius vittatus (Bosc), cangrejo hermitaño rayado, Penaeus
aztecus (Ives), Brown shrimp, P. duorarum (Burkenroad), camarón rosa, P. setifems
(Linnaeus), camarón blanco (Decapoda); Luidia clathrata (Say), starfish (Asteroidea);
Sagitta hispida (Conant), S. tennis (Conant), Sagitta sp., gusanos flecha
(Chaetognatha).
Localidades: Santa Isla Sapelo, Georgia; Bahía Vizcaína, Florida (costa este);
Bahía de Tampa, costa de Tampa & Destin, Florida (costa oeste); Mississippi Sound
and Golfo de México, Mississippi; sureste de Louisiana; Bahía Galveston, Texas.
Sitio de infección: Mucosa del intestino. Mesenterio de peces, cavidad
corporal y órganos digestivos de invertebrados. Larvas enquistadas en la cavidad
del cuerpo, vísceras y musculatura de peces de escama y mariscos. Estómago,
intestino y ciego pilórico de truchas criadas





























