Embed Size (px)
Citation preview

Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Medicine 960
_____________________________ _____________________________
DNA Fragmentation in Cultured Cells Exposed to High Linear
Energy Transfer Radiation
BY
ERIK HÖGLUND
ACTA UNIVERSITATIS UPSALIENSISUPPSALA 2000

Dissertation for the Degree of Doctor of Philosophy, Faculty of Medicine, in BiomedicalRadiation Sciences presented at Uppsala University in 2000
ABSTRACT
Höglund, E. 2000. DNA fragmentation in cultured cells exposed to high linear energytransfer radiation. Acta Universitatis Upsaliensis. Comprehensive Summaries of UppsalaDissertations from the Faculty of Medicine 960. 58 pp. Uppsala. ISBN 91–554–4822–4.
The DNA double-strand break (DSB) is a critical lesion which, if not completely restored,can have serious biological consequences. The relative biological effectiveness (RBE) ofmany severe end-points are closely related to radiation quality, with increasedeffectiveness at elevated ionization density. Data presented provide information aboutthe influence of radiation quality on the initial processes causing DNA damage, and themechanisms leading to its restoration. Such information will increase the understandingof radiation action mechanisms in mammalian cells.
Human cells were irradiated with accelerated ions having linear energy transfer (LET)values in the range 40-225 keV/µm, and 60Co-photons. Detailed analyses of the DNAfragment distributions were performed in the size-range 5 kilobasepairs to 6megabasepairs by pulsed-field gel electrophoresis.
A non-random fragmentation of DNA was evident, with an elevated number of smalland medium-sized fragments for ion irradiation, and the total number of breaksincreased by 80–110% when these fragments were included in the analyses. The RBE forDSB induction was 1.2–1.5. A two-fold increase of the number of breaks induced pernitrogen ion passing the cell nuclues was found when LET was increased from 80 to 225keV/µm, indicating a possible role of particle track structure in DSB induction.Furthermore, the ability to repair DNA was closely related to radiation quality, with anincreased proportion of unrejoined breaks for densely ionizing radiation. Surprisingly,the majority of breaks were rapidly rejoined even following exposure to high-LETradiation. The proportion of breaks restored by the slow phase showed a five-foldincrease for the highest LET tested, compared with photons. The results presentednominates the complexity of breaks as one determining factor for reduced reparabilityreported following high-LET exposure.
Key words: DNA fragmentation, high linear energy transfer, DNA repair, double-strandbreak, complexity of breaks
Erik Höglund, Division of Biomedical Radiation Sciences, Department of Oncology, Radiologyand Clinical Immunology, Rudbeck Laboratory, Uppsala University, SE-751 85 Uppsala, Sweden
© Erik Höglund 2000
ISSN 0282–7476ISBN 91–554–4822–4
Printed in Sweden by Eklundshofs Grafiska AB, Uppsala 2000

To
Johanna and Alexander

This thesis is based on the following paper, which will be referred to in the text
by their roman numerals I–IV.
I. STENERLÖW, B., HÖGLUND, E. AND CARLSSON, J. Induction and rejoining of
large DNA fragments after ion irradiation. Radiation Research, 151, 642-648
(1999).
II. HÖGLUND, E., BLOMQUIST, E., CARLSSON, J. and STENERLÖW, B., DNA
damage induced by radiation of different linear energy transfer: initial
fragmentation. International Journal of Radiation Biology, 76, 539-547 (2000).
III. STENERLÖW, B., HÖGLUND, E., CARLSSON, J. and BLOMQUIST, E., Rejoining of
DNA fragments produced by radiations of different linear energy transfer.
International Journal of Radiation Biology, 76, 549-557 (2000).
IV. HÖGLUND, E. AND STENERLÖW, B., Induction and rejoining of DNA damage
after exposure to radiation of different linear energy transfer. Possible roles
of track structure and chromatin organisation. Radiation Research,
Conditionally accepted.
Reprints were made with kind permission from The Radiation Research Society
(I), and Taylor & Francis Ltd. (http://www.tandf.co.uk) (II and III).

CONTENTS
ABBREVIATIONS_____________________________________________ 7
INTRODUCTION _____________________________________________ 8
Tumour therapy by high-LET irradiation ____________________________ 9
Internal irradiation ________________________________________________ 9
External radiation ________________________________________________ 10
Radiation safety________________________________________________ 12
DNA damage and repair _________________________________________ 13
Cellular processing of the damage___________________________________ 14
AIMS OF THE STUDY ________________________________________ 16
METHODS __________________________________________________ 17
Cells _________________________________________________________ 17
Irradiation ____________________________________________________ 17
Low-LET photons ________________________________________________ 17
High-LET ions ___________________________________________________ 17
Quantification of induced DNA damage ____________________________ 20
Pulsed-field gel electrophoresis, PFGE_______________________________ 20
Conventional assay - the FAR assay ________________________________ 21
DNA fragmentation analysis_______________________________________ 22
RESULTS & DISCUSSION ____________________________________ 25
Induction _____________________________________________________ 25
FAR assay_______________________________________________________ 25
Fragment analysis ________________________________________________ 27
Rejoining _____________________________________________________ 35
Kinetics _________________________________________________________ 35
Rejoining after exposure to the same number of particles_______________ 40

Chromatin organization _________________________________________ 41
SUMMARY AND CONCLUSIONS ____________________________ 45
REFERENCES ________________________________________________ 47
ACKNOWLEDGEMENTS _____________________________________ 56

7
ABBREVIATIONS
bp base pair
δMi width of gel segment i
DNA deoxyribonucleic acid
DSB double-strand break
Fi fraction of DNA in gel segment i
Fslow fraction of breaks restored by the slow phase of repair
F<k fraction of DNA smaller than the threshold size k
FAR fraction of activity released
GCR galactic cosmic rays
HR homologous recombination
HZE high (H) atomic number (Z) and energy (E) particles
kbp kilobasepair
LET linear energy transfer
LMDS locally multiply damaged sites
Mbp megabasepair
Mi average fragment size in gel segment i
NHEJ non-homologous end-joining
OER oxygen enhancement ratio
PCC premature chromosome condensation
PFGE pulsed-field gel electrophoresis
RBE relative biological effectiveness
SEP solar energetic particles
τ½ DSB repair half-time

8
INTRODUCTION
Researchers have been fascinated by the nature of radiation, and its effects on
the environment and man, ever since the discovery of X-rays (Röntgen, 1895),
and of radioactive elements, polonium and radium, by Pierre and Marie Curie
in 1898. Although radiobiological research started in the context of medical
radiology, it gradually progressed to more fundamental questions concerning
the effects of different types of radiation on all types of biological systems.
Consequently, radiobiological research came to involve investigators from
various fields, not only biologists and clinicians, but also physicists and
chemists.
Already in the early 20th century, ionizing radiation was used in the hope of
eradicating tumours. The pioneers in the field explored its possibilities by a trial
and error approach, but as the available technology and scientific knowledge
improved, more sophisticated methods evolved. Developments — mainly in
physics and dosimetry — resulted in the concept of conformal radiotherapy, i.e.
the possibility of shaping radiation fields to accord with treatment volumes by
crossing several X-ray beams. It was soon realized that the physical properties
of charged particles, and their stopping in matter, could be used to improve the
therapeutic effects of radiation. At present, considerable efforts are being made
in the field of radiobiology, to elucidate the mechanisms governing how DNA
damage occurs and how it is repaired. Such knowledge could provide the
means to modify radiation response in both healthy and malignant tissues,
thereby improving the therapeutic effects still further.
It was also realized early on that ionizing radiation could also have adverse
health effects. An association between skin cancer and radiation among
radiologists were found as early as 1902, and much has been learned since then
about the risks of radiation, mainly from epidemiological studies on atomic
bomb survivors and radiation workers (see review by Ron (1998) and references
therein). Present-day radiation safety research is focused on risk assessment, e.g.
studies on the influence of low dose/long duration exposure to radiation of
different qualities.
Ionizing radiation can be grouped into two main categories: sparsely and
densely ionizing radiation. The qualitative property that distinguishes the two

9
is the spatial distribution of energy transfer in the surrounding matter. The
initial events, eventually leading to radiation induced biological effects, are the
ionization and excitation of atoms and molecules of the irradiated matter.
X-rays and gamma photons deposit their energy via secondary electrons, set in
motion by the incident photons at all depths in the target matter, and are
therefore sparsely ionizing. In contrast, the energy deposition of charged
particles such as α-particles and accelerated ions, is restricted to a limited
volume adjacent to the primary particle track, and a more densed ionization
pattern occurs. Linear energy transfer (LET) and related concepts have been
introduced to determine the energy deposited by charged particles in
microscopic regions. In 1962 the International Commission on Radiological
Units (ICRU) defined this quantity as follows:
"The linear energy transfer of charged particles in medium is the
quotient of dE/dl, where dE is the average energy locally imparted to the
medium by a charged particle of specific energy in traversing a distance of dl."
Tumour therapy by high-LET irradiation
Tumour therapy by high-LET radiation can be performed either internally, i.e.
endoradiotherapy — irradiation with nuclides located directly in the tumour
tissue — or by external beams of accelerated particles such as heavy ions, e.g.
carbon ions. Endoradiotherapy using radioactive nuclides holds promise for the
treatment of spread cancer disease, while external irradiation with high-LET
beams is more suitable for deap-seated, voluminous tumours.
Internal irradiation
The use of nuclides emitting high-LET particles in tumour therapy will
probably increase significantly in the future. Boron neutron capture therapy
(BNCT) has already been in use for a long time (reviewed by Carlsson et al.,
1992, and Barth, 1999). The rationale for BNCT is the likelihood that thermal
(slow) neutrons will be captured by the stable boron which is thereby activated.
A nuclear reaction that is initiated by the capture of neutrons, results in the
emission of short-range high-LET nuclear fragments — an α-particle and a
lithium ion — which deposit their energy within a few micrometres of the

10
decay (a distance comparable to the radius of a mammalian cell) with
considerable likelihood of cell killing.
Naturally radioactive nuclides can also be used for therapy. For instance,
α-particle emitters (e.g. 211At and 212Bi) have shown promising results in
pre-clinical studies, using a variety of labelled compounds (Vaidyanathan and
Zalutsky, 1996). Furthermore, nuclides emitting cascades of low-energy
electrons, viz. Auger emitters, can be useful for treatment (Daghighian et al.,
1996; Welt et al., 1996) by virtue of their proven ability to cause severe DNA
damage and thereby cell death, when situated within cell nuclei (Kassis et al.,
1987). The Auger cascade resembles in many ways the clustering of ionizations
that is characteristic of high-LET radiation.
Common to all of the internal radiation treatment modes outlined above, is the
need for effective targeting of the tumour cells by some kind of carrier
molecule. Of course, without strictly selective targetting at the malignant cells,
damage to healthy tissue will be unacceptable high. The prospects for cancer
therapy using radioactive nuclides were reviewed by Kampf (1990).
External radiation
One benefit of radiotherapy using external beams of light- and heavy ions, is
their proven high relative biological effectiveness (RBE) for reproductive cell death.
More important, the possibility of controlling the site of energy deposition of
charged particles, thereby inflicting radiation damage restricted to the tumour
area alone while sparing the surrounding healthy tissue, further improves the
therapeutic ratio (Tobias et al., 1982; Blakely et al., 1984). Furthermore, the effect
of oxygen on radiation sensitivity works in favour of tumour treatment using
radiation having high-LET, the explanation being that cells exposed to low-LET
radiation, such as X-rays, show a significantly increased radiation sensitivity
when molecular oxygen is present during the exposure. This effect is called
oxygen enhancement ratio (OER), defined as the ratio of hypoxic to aerated doses
needed to achieve the same biological effect. OER decreases with increasing
LET, from about 3–3.5 for cell killing with X-rays, to 1, i.e. no oxygen effect, for
α-particles at 150–200 keV/µm (Barendsen et al., 1966; Prise et al., 1990).
Irradiation with high-LET therefore overcomes the reduced sensitivity to
radiation damage expected of poorly oxygenated tumour cells.

11
External radiation for tumour treatment, using charged particles, started with
protons more than half a century ago, and proton therapy centres are now
operating all over the world. The first proposal to use protons for tumour
treatment was put forward as early as 1946 (Wilson, 1946), and pioneer
treatment started in the mid 1950s at the Lawrence Berkeley Laboratory
(California, USA) (Tobias et al., 1955). The historical development of proton
radiobiology and therapy was reviewed by Raju (1995), amongst others.
The use of charged particles in radiation therapy makes possible a better dose
distribution in tissue, compared with photons and electrons, due to their
inverse dose profile (i.e. the Bragg peak, see Figure 1). This increases the tumour
dose, while reducing the dose to the surrounding healthy tissue — a crucial
attribute, since the tolerance of the healthy tissue around the tumour is the
major limiting factor for tumour control. Furthermore, the narrow Bragg peak
can be modulated by means of absorbers, to achieve increased conformity with
the tumour tissue. This technique, with a modulated spread-out Bragg peak,
was first used in the late 1950s for treatment of human cancers by means of
protons (Larsson, 1962).
0
0.5
1
1.5
0 0.5 1 1.5 2
Abs
orbe
d do
se (a
rbitr
ary
units
)
Depth (mm)
Figure 1. Theoretical Bragg curve. Depth–dose distribution for a heavy charged
particle, stopped in water.

12
Most clinical experience with particles other than protons comes from
treatments at the BEVALAC accelerator in Berkeley (Calif., USA), using
helium, carbon and neon ions (Castro, 1995). Treatment centres where
accelerated ions are used (or are going to be used) have been set up at
numerous locations around the world, e.g. in Japan (Tsujii et al., 1997), in
Germany (Eickhoff et al., 1999), and Italy (Amaldi, 1998).
Regarding all treatment modalities, the dose to the tumour volume must be
sufficiently high for tumour control, while sparing healthy tissues in order to
prevent late side effects. For this purpose, beams of accelerated particles may be
an effective tool for external therapy of deep-seated tumours. The effect of the
treatment is governed mainly by its ability to produce irreparable damage to
the DNA. The LET dependence on induction and repair of DNA damage, in
both normal and malignant cells, following exposure to different particles
needs to be thoroughly explored in order to ascertain the most suitable
radiation quality to use. Too heavy a particle could produce irreparable damage
not only at the site of the tumour, but also in the channel of entry on its passage
through non-malignant tissue in front of the tumour, while lighter particles, e.g.
protons, may be insufficiently effective in the target volume. Extensive
investigations into the biological effects, in relation to radiation quality, may
help to optimize the outcome of tumour treatments using particle beams (Kraft,
1990; Blakely and Kronenberg, 1998; Kraft et al., 1999).
Radiation safety
Besides application to clinical treatment, high-LET radiation and its effects on
biological systems is highly relevant for radiation protection and safety. Several
epidemiological studies, performed mainly in mineworkers, have confirmed a
link between radon exposure (high-LET α-particles) and cancer in the lungs, as
reviewed by Jostes (1996). The relevance of such studies regarding the lower-
exposure home environment has been disputed and more information is
needed about the effect of low doses of high-LET radiation, regarding DNA
damage induction and repair. Complex damage, and subsequent mis-rejoining
or deletions, following high-LET radiation do cause mutations and
chromosomal rearrangements that in unfortunate cases lead to malignant
transformations. Knowledge about these mechanisms is vital for adequate
quantification of radiation risk.

13
Radiation safety is also discussed within the framework of space research.
Crews of manned space-craft are exposed to various kinds of ionizing radiation.
Two main sources of radiation in space are protons emitted as solar energetic
particles (SEP), and the galactic cosmic rays (GCR) consisting of protons and other
energetic nuclei — high (H) atomic number (Z) and energy (E) particles (HZE).
Since the physical aspects of the SEP protons and their interaction with matter
are fairly well established, measures can be taken to monitor their presence and
adequate estimation of the risk is possible. Thus, they do not constitute a
serious radiation protection problem, whereas the HZE particles present
significant problems for radiation protection. They have an energy of several
hundred MeV per nucleon, and high-LET nuclear fragments, produced on
impact with the shielding material, are highly penetrating and may severely
damage DNA in a large number of cells in the exposed crew. Further
information about the effects of HZE particles may make future risk estimation
more accurate and reliable, and help to define radiation limits for human
exploration of space (Schimmerling, 1992; Schimmerling, 1995).
DNA damage and repair
Ionizing radiation produces many different types of damage to macromolecular
species within cells, and the critical targets for radiation-induced cell killing are
located in the cell nucleus. This became evident in several experimental studies
where the cytoplasm and the nucleus were selectively irradiated. It is also
generally accepted that the critical target for generation of biological effects is
the DNA, where different types of lesions, including base damage, cross-links
(within the DNA molecule or with associated proteins) and strand breaks are
produced by the radiation. Of these lesions, the DNA double-strand break (DSB)
stands out as the critical one for radiobiological effects (reviewed by Iliakis,
1991). Furthermore, there is a close relation between radiation-induced DSBs,
chromosomal aberration production and cell transformation (Bryant, 1984;
Yang et al., 1989; Kiefer, 1992).
The consequences of an isolated lesion in DNA is not significant in terms of the
ultimate biological end-point, e.g. cell killing, because of effective cellular repair
mechanisms. However, a DSB is often associated with additional damage in the
immediate vicinity of the break, on either the same or the opposite strand,

14
forming more complex lesions caused by local clusters of ionizations. These
lesions have been called locally multiply damaged sites (LMDS) (Ward, 1981) and
their composition is closely related to radiation quality (Prise et al., 1994).
These qualitative differences in DSBs may provide some explanation for the
close relation between ionization density, i.e. LET, and a variety of biological
end-points. For instance, the RBE for reproductive cell death has been shown to
depend on LET, with RBE in the range 3.5–5 for LET values around 100–300
keV/µm (Wulf et al. 1985; Raju et al. 1991; Stenerlöw et al., 1995). Moreover, in
support of the proposed role of break complexity on the microscopic level, the
initial number of DSBs induced was found not to increase with LET (reviewed
by Prise et al., 1998). However, evidence of elevated levels of initially induced
DSBs has been reported by some investigators, using improved electrophoresis
protocols. These revealed a significant contribution from smaller DNA
fragments (200 kbp – 1 Mbp) that were not resolved in earlier studies (Löbrich
et al, 1996; Newman et al., 1997; Kraxenberger et al., 1998).
The smaller fragments have been suggested to derive from interactions between
the radiation, e.g. the track of a charged particle, and the higher order chromatin
structures of the genome. Even shorter fragments (<1 kbp), related to the basic
organization of DNA around the nucleosomes in the 30 nm chromatin fibre,
were predicted by theoretical analysis (Holley and Chatterjee, 1996). Such
fragments have also been detected experimentally by Rydberg et al. (1996). An
extensive exploration of DNA fragmentation, in relation to radiation quality
and genomic organization, may facilitate adequate modelling of the
development of DNA damage. This could help extrapolation to effects from low
doses, or even single events, and such information would be valuable for risk
estimation in radiation safety (Friedland et al., 1998; Sachs et al., 1998).
Cellular processing of the damage
The repair of DNA damage is a critical step that could lead from the initial
damage, to broken or rearranged chromosomes, cell death, or cancer. In order
to prevent this, the cells have evolved various ways to restore the genome, and
early evidence of the occurrence of repair of X-ray induced damage in
mammalian cells was provided by Elkind and Sutton (1960).

15
It is well known that high-LET irradiation can cause damage that is more
difficult to repair, and that this largely explains the high RBE values for severe
outcome. The explanation of the restricted repair capacity is believed to be a
greater complexity of induced DSBs, due to clustered damage (Roots et al., 1979;
Ward, 1985; Goodhead, 1989; Goodhead, 1994; Ward, 1994). Indeed, slower
repair and a larger proportion of residual damage following high-LET exposure
have frequently been reported (Ahnström and Edvardsson, 1974; Ritter et al.,
1977; Coquerelle et al., 1987; Blöcher, 1988; Heilmann et al., 1993; Jenner et al.,
1993; Weber and Flentje, 1993; Stenerlöw et al., 1996), and non-repairable DSBs
and unrejoined chromatin fragments are closely correlated to cell killing.
The increased complexity may lead to misrejoining of breaks, i.e. two ‘wrong’
ends are joined together by the cellular repair mechanism. Several end-points
that are believed to require misrejoining events, e.g. chromosome exchanges
(Durante et al., 1992; Durante et al., 1995) and interstitial deletions (Thacker,
1992), are induced at increased frequencies by increasing LET values. A possible
explanation may be found in the clustering of breaks seen for high-LET. The
numerous breaks within a cluster, or in the immediate vicinity of a high-LET
particle track, are more likely to be rejoined with a ‘wrong’ end, suggesting the
involvement of higher order chromatin structures in the formation of
chromosome aberrations derived from induced DSBs (Sachs et al., 1997).

16
AIMS OF THE STUDY
The DNA double-strand break is a critical lesion which, if not completely
restored, can have serious biological consequences, including cell death,
chromosomal aberrations, mutations, and cellular transformation. Moreover,
the relative biological effectiveness (RBE) of these end-points has been shown to
be closely related to radiation quality, with increased effectiveness at elevated
ionization density. Better knowledge of the initial processes causing DNA
damage, and the mechanisms leading to its restoration, should help us to
identify critical lesions and increase our understanding of radiation action
mechanisms in mammalian cells. The principal aim of this work was to
investigate the influence of radiation quality on the induction and processing of
DNA damage in mammalian cells in vitro, taking DNA fragment distribution
into account. In greater detail:
• to investigate the initial fragmentation of DNA following exposure to the
same type of ions having different LET values;
• to investigate the rejoining of DNA fragments following exposure to the
same type of ions having different LET values;
• to investigate the induction and rejoining of DNA double-strand breaks
following exposure to different ions having similar LET values;
• to investigate the possible involvement of particle track structure and
higher order chromatin organization in the induction and rejoining of DNA
double–strand breaks.

17
METHODS
Cells
Low-passage normal human fibroblast cells (GM5758, Human Genetic Mutant
Cell Repository, Camden, NJ, USA) were used as a model system in most of the
presented work (Papers II–IV). The cells, which were labelled with 14C or 3H
and grown to confluence prior to irradiation, constituted a homogeneous
system, suitable for investigation of DNA damage. Furthermore, since they are
not very specialized, they share many characteristics with other kinds of cells in
the body. In the first paper (Paper I) two human cell lines were used, U-343MG
glioma and K562 erythroleukemia cells.
Cells were irradiated as monolayer cultures (DNA DSB rejoining and
induction), or embedded in agarose plugs (DNA DSB induction). All irradiation
was performed on ice to prevent any repair processes during exposure.
Irradiation
Low-LET photons
Throughout this work we have used photons from a conventional 60Co
apparatus (1.2 MeV photons, LET ~0.5 keV/µm) as reference radiation. The
photon dose rate was 1–1.3 Gy per minute, as determined by ionization
chamber and thermoluminiscence dosimetry.
High-LET ions
The Gustaf Werner synchrocyclotron at the The Svedberg Laboratory (Uppsala,
Sweden) provided the accelerated ions. The different ions (14N6+, 14N7+, 10B5+ and20Ne9+) were extracted from external ion sources and accelerated to energies of
about 32–48 MeV/u. Beam properties are listed in Table 1. The ion beams were
directed, from the accelerator to the Biomedical Unit, through a high vacuum
beam transport system.

18
ED C B A
beam direction
Figure 2. Ion-beam delivery. Experimental set-up for ion irradiation at the
Biomedical Unit, The Svedberg Laboratory, Uppsala University (see Stenerlöw
et al., 1996 for details). Gold foil (A), transmission chamber (B), plastic
absorber (C), scanning device for Markus chamber (D) and ice-cooled sample
holder (E). [By courtesy of T. Hartman, The Svedberg Laboratory, Uppsala,
Sweden.]
On entering the experimental area the beams were quite narrow (~ 1 cm in
diameter) and had to be scattered laterally by a thin gold foil (A, Figure 2) to
achieve a uniform dose distribution at the position of the sample (E). The beam
profile was analysed by vertically and horizontally scanning an ionization
chamber (D) over the target area, and the dose distribution in the target was
uniform ±5% within a radius of 18 mm. Absolute dosimetry was performed
using a transmission chamber (B) together with the ionization chamber (D) (see
Stenerlöw et al. (1996) for details.)
When heavy charged particles traverse matter, they lose energy mainly by
electromagnetic interaction with the electrons in the target material. The rate of
energy loss for the particle is proportional to the square of its charge (Z2) and
inversely proportional to the square of its velocity (β2). Thus, the magnitude of
the dose absorbed by the medium increases with decreasing particle speed, and
the resulting depth–dose distribution has the shape of the classical Bragg curve
(Figure 1). The depth–dose distribution and the maximum range of the particles
was determined by placing plastic absorbers (C) of varying thickness in front of
the ionization chamber and plotting the registered dose vs the thickness of the
absorber.

19
Cells were irradiated at different positions on the Bragg curve by placing
absorbers of differing thickness in front of the samples. The corresponding ion
energies and LET values (Table 1) were obtained from measurements of the
residual beam range and calculations of the stopping of ions in water (Ziegler
and Manoyan, 1988).
Table 1. Beam properties
Radiation
Ion energy at cells
(MeV/u)LETa at cells
(keV/µm) Paper
60Co photons - < 0.5 I-IV
Helium ions 0-22 40 (35-55)b II
Boron ions 37
16
40
80
unpublished*
Nitrogen ions 36
16-25
14
10
80
125 (110-150)c
175
225
II-IV
I-IV
II-IV
II-IV
Neon ions 24
13
225
300unpublished*
a Unrestricted.b Extended Bragg peak. The LET range stated covers a 68% confidence interval.c Cells in agarose plug (1 mm thick).
* Manuscript in preparation, Höglund, E. and Stenerlöw, B., 2000.

20
Quantification of induced DNA damage
Pulsed-field gel electrophoresis, PFGE
PFGE has found a broad application in biochemistry and genetics. One of its
greatest advantages has been its sensitivity in the analysis of long DNA
molecules. Fragments up to 13 Mbp long have been separated (Orbach et al.,
1988), and the ability to separate such large fragments makes it possible to
study radiation-induced DNA fragmentation, and cellular processing of
damage, following exposure to doses similar to those used in cell survival
experiments. For instance, rejoining of radiation-induced DSBs has been
measured in cultured cells after irradiation with doses as low as 5 Gy
(Stenerlöw et al., 1996).
In order to separate megabasepair-length DNA fragments by size, pulsed
electric fields with periodically changing direction were applied. The optimal
angle between the two field directions has been shown to be near 120°
(Schwartz and Cantor, 1984; Southern et al., 1987; Cantor et al., 1988) which was
the angle used in the present work. The exact mechanisms involved in the
separation have not yet been extensively described, but some intuitive
reasonings might be illustrative. Since the fragments are much longer than the
average size of the pores in the agarose matrix, the retarding effect is of minor
importance in a static field, i.e. two fragments of different length will migrate
with the same velocity. But, when the pulsed fields were applied, the fragments
aligned alternately with the two field directions, always with their trailing ends
first because of the lower force of friction experienced by the fragment ends at
the obtuse-angled corner. Thereby shorter fragments started from a more
advanced position and gained ground in every pulse, thus resulting in a size
separation of the fragments.
By changing various parameters, e.g. pulse duration, field strength or agarose
concentration, the PFGE protocol can be optimized for separation of differently
sized fragments. Longer pulse times and weaker electric fields favour longer
fragments, while an increased agarose concentration favours shorter ones.
Three different protocols were used in this work. First, the very long fragments
(Paper I) were separated using a 74-hour run at 1.4 Volts per cm. In the more
detailed analyses of the fragmentation, performed in the succeding
investigations (Papers II-IV), we needed distinct separation within a wide

21
sizerange. To achieve this we chose two shorter schemes, 45.7 and 17 hours (see
Paper II and Table 2 for details) which, when combined, provided separation of
fragments in the range 5 kbp up to about 6 Mbp with good resolution.
Table 2. Pulsed-field gel electrophoresis protocols
Optimal
size range
Field
(V/cm)
Pulses
(length/duration)
Total time
(h) Paper
< 10.5 Mbp 1.4 180-60 mina 74 I
1 - 6 Mbp 2 10 min/ 3 h
20 " / 5.33 h
30 " / 8 h
40 " / 9.33 h
60 " / 20 h
45.67 II-IV
5 kbp – 1.5 Mbp 7 10 s / 7 h
40 " / 5 h
70 " / 5 h
17 II-IV
a Pulses were ramped, i.e. they were continuously shortened from 180 to 60 minutes during the 74 h
electrophoresis run
Conventional assay — the FAR assay
Various techniques have been used to measure the yields of DNA DSBs
following exposure to radiation (reviewed by Prise et al., 1998). One of the most
widely used methods is the conventional FAR assay, in which the Fraction of
Activity Released from the sample plug is used as a measure of the degree of
damage. The FAR value designates the amount of radioactively labelled DNA
that enters the gel during the electrophoresis run. Only DNA molecules smaller
than a certain exclusion size can enter the gel, leaving very large fragments and
undamaged DNA in the plug. Specific DNA size markers, e.g. the largest
chromosome of Schizosaccharomyces pombe (5.75 Mbp), were also used to
determine the FAR value. The PFGE protocols used are optimized for
separation of Mbp-sized fragments, and the size distribution of the migrated
fragments is disregarded. The number of induced DSBs is related to the FAR

22
value in a non-linear fashion, and an assumption of randomly distributed
breaks along the DNA molecule gives the breakage yield, expressed as breaks
per unit length of DNA and dose (Blöcher, 1990; Cook and Mortimer, 1991;
Cedervall et al., 1995). The random breakage model used throughout this work
is Blöcher’s model (1990) :
))(( nk
nrk
k 11e1F nrk
−−−−++++−−−−==== −−−−<<<< , (1)
where kF<<<< is the fraction of DNA whose size is smaller than threshold size k, r
is the mean number of DSBs per chromosome, and n is the average size of a
chromosome.
DNA fragmentation analysis
Two major drawbacks of the FAR assay are the assumption of random
fragmentation, and the poor resolution for separation of smaller DNA
fragments. These are problems of great importance when analysing DNA
damage after exposure to high-LET radiation, due to an increased likelihood
that short and medium-sized DNA fragments will be produced, compared with
predictions based on random breakage models (Prise et al., 1998). The shorter
fragments are a direct result of the interaction between the particle track
structure and all levels of chromatin organization in the cell (Rydberg, 1996).
The conventional assays, which do not include these smaller fragments, thereby
tend to underestimate the RBE for DSB induction. This is also evident in the
literature (Löbrich et al., 1996; Newman et al., 1997). The RBE is defined by the
quotient of the yields for the test radiation and the 60Co reference radiation.
Thus, a careful investigation of the DNA fragment size distribution is necessary
for correct scoring of the damage.

23
S. cerevisiae
λ DNA-PFGE
Migrated DNA
48 97 145 225 375 680 9301100
Size
Mig
rate
d D
NA
Figure 3. Gel slicing. The lanes of migrated DNA were cut into different size
segments, according to DNA size markers loaded in the gels. Sizes are listed in
Table 3. The figure illustrates the gel optimized for separation in the range 5
kbp – 1.5 Mbp, and the markers S. cerevisiae and Lambda DNA-PFGE were
used.
The lanes of migrated DNA were cut into ten different size segments according
to DNA size markers (S. pombe, S. cerevisiae and Lambda DNA-PFGE markers)
loaded in the gels (listed in Table 3). In Figure 3, the slicing of the gel optimized
in the range 5 kbp–1.5 Mbp is illustrated. The number of DNA fragments (n) in
each size zone is proportional to the mass of DNA in the corresponding gel
segment. By dividing the fraction of DNA in a gel segment (Fi) by the average
fragment size in that zone (Mi), the number of fragments in each size zone can
be calculated according to Eq. 2 (see Paper II for details).
(((( ))))i
ii
M
FMn ≅≅≅≅ (2)
24
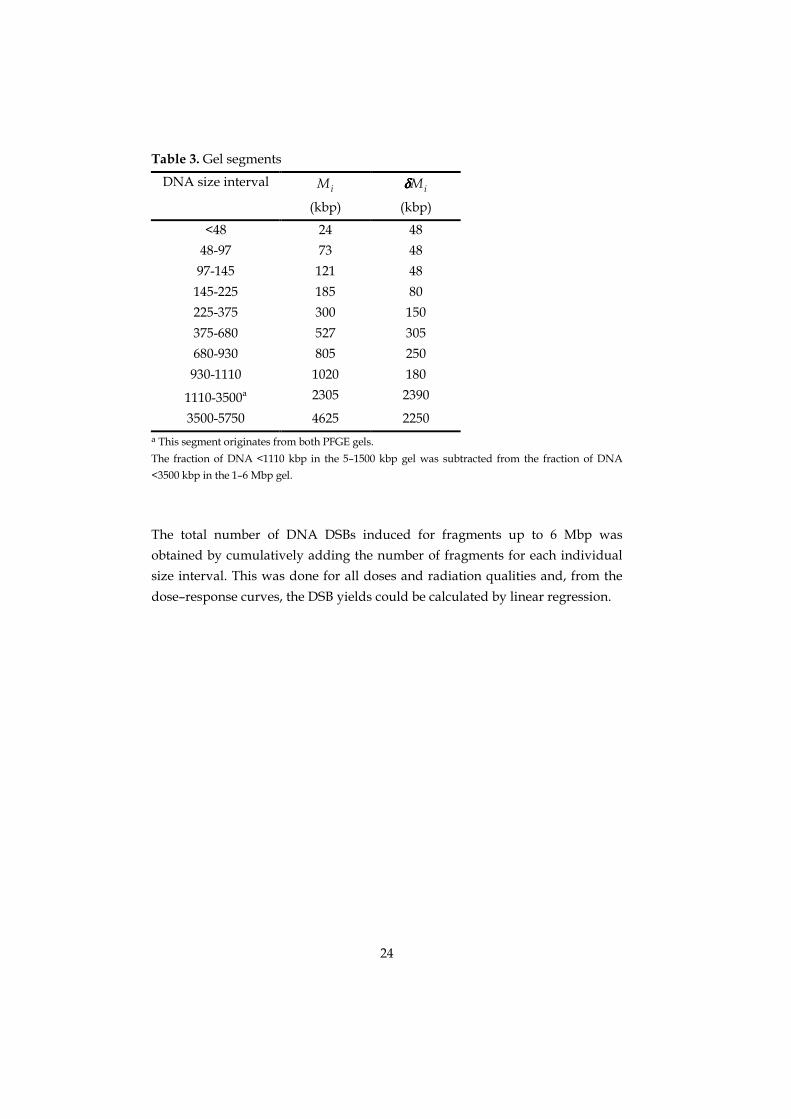
Table 3. Gel segments
DNA size intervaliM
(kbp)iMδδδδ
(kbp)
<48 24 48
48-97 73 48
97-145 121 48
145-225 185 80
225-375 300 150
375-680 527 305
680-930 805 250
930-1110 1020 180
1110-3500a 2305 2390
3500-5750 4625 2250a This segment originates from both PFGE gels.
The fraction of DNA <1110 kbp in the 5–1500 kbp gel was subtracted from the fraction of DNA<3500 kbp in the 1–6 Mbp gel.
The total number of DNA DSBs induced for fragments up to 6 Mbp was
obtained by cumulatively adding the number of fragments for each individual
size interval. This was done for all doses and radiation qualities and, from the
dose–response curves, the DSB yields could be calculated by linear regression.

25
RESULTS & DISCUSSION
Induction
FAR assay
The dose response for the the FAR value, i.e. the amount of DNA that migrates
into the electrophoresis gel, depends on several conditions. Besides the
electrophoretic parameters, such as agarose concentration and field strength,
the threshold size chosen for the analysis can also affect the outcome (Paper I).
Further, the mode of DSB induction determines the dose response. If the breaks
are distributed randomly along the chromosome, an induction of DSBs linear
with dose will result in a non-linear fraction of extractable DNA (Blöcher, 1990).
0
0.05
0.1
0.15
0.2
0 5 10 15 20
nitrogen, U-343MGγ rays, U-343MGnitrogen, K562γ rays, K562
Frac
tion
of D
NA
rele
ased
<6
Mbp
Dose (Gy)
Figure 4. Dose response. The fraction of DNA released for the molecular weights <6
Mbp, is shown for K562 and U-343MG cells irradiated with nitrogen ions
(125 keV/µm) and 60Co-photons. Data for both cell lines were fitted to one
single random-breakage curve after photon irradiation. Following nitrogen ion
exposure, separate linear curves were fitted for each cell line.
(Data from Paper I.)

26
The dose response, as determined by the fraction of DNA smaller than 6 Mbp,
was compared for two human cell lines (U-343MG and K562) after exposure to
low-LET gamma and high-LET nitrogen ions (Figure 4). The data representing
the ion irradiation showed a linear dose response for doses below 30 Gy, in
contrast to the sigmoid-shaped random-breakage curves seen for gamma. The
ion data could not be satisfactorily described by a model assuming random
breakage and were best fitted to linear expressions. Instead, the γ and ion
curves intersected at 12 and 18 Gy for the K562 and the U-343MG cells,
respectively. This phenomenon has not been noted earlier by investigators
using either pulsed or constant-field gel electrophoresis, probably because the
fraction of DNA migrating out of the plug — without any specific DNA size
marker — was used as a measure of the damage, resulting in a relatively large
threshold in their analysis (e.g. Rydberg et al., 1994). In some studies where the
neutral filter elution technique was used, similar linear dose responses were
reported for 238Pu α-particles (Prise et al., 1987) and nitrogen ions (Nygren and
Ahnström, 1996). The reason why the intersections were found at different
doses for the two cell lines in Paper I is unclear. One possible explanation might
be differences in higher-order chromatin structures and organization between
the cell lines. Nevertheless, it is evident from the present work that
nonrandomness of DSB induction by high-LET radiation can also be detected
by the conventional FAR assay.
Low random yield and clustered breaks
The relative yields for fragment sizes smaller than 6 Mbp, calculated by
dividing the ion dose–response curves by the gamma ditto, significantly
exceeded unity (~ 3 at 1.5 Gy) for doses up to about 10 Gy, but decreased at
higher doses (~ 0.8 at 30 Gy). For sizes 6–10.5 Mbp the correlation was reversed,
with a constant ratio around 0.7, i.e. the ions were less effective at producing
large fragments. A possible interpretation of the results could be that the
induction of DSBs following high-LET irradiation includes not only a random
breakage mode, but also a non-random component. It can be assumed that, for
each randomly induced break, one (or more) additional associated break formes
within a certain distance from the original break. If this distance is smaller than
the chosen threshold, these fragments can enter the electrophoresis gel and
thereby contribute a linear dose response to the fraction of DNA released. Based
on these simple assumptions, the data for the nitrogen ions were modelled. It
was found that when a relatively low random induction yield was used (about
50% of the yield measured for 60Co-photons) together with s associated breaks

27
within x Mbp from the initial random break, the nitrogen data could be fitted
for doses up to 30 Gy when sx = 2 Mbp (Paper I). The product sx could be
interpreted as a release of either one single fragment of size 2 Mbp, or several
smaller fragments of total length 2 Mbp. Although no extensive conclusions
should be drawn from such a rough analysis, the result supports the idea of
clustered DSBs within higher orders of chromatin structure, as proposed by
many others (Jin et al., 1995; Löbrich et al., 1996; Newman et al., 1997;
Kraxenberger et al., 1998; Frankenberg et al., 1999). The relative influence of the
non-random component will increase for decreasing doses, because at lower
doses the random breakage produces fragments that are too large to be released
below the chosen threshold. This may also explain why the linear response for
high-LET at low doses has not been recognized earlier — the analyses have
usually been performed using higher doses (e.g. Heilmann et al., 1995). In order
to further analyse the significance of these small and medium-sized fragments
for the DSB induction yield, it is essential to take the size distribution of
fragments into account.
The significant non-random contribution to the induction yields for high-LET
was also confirmed in Paper II, in which a more detailed investigation of the
LET dependence of DNA fragmentation in normal human fibroblast cells (GM
5758) was performed. For this cell line too, a linear dose response was found
when DNA sizes below 1.11 Mbp were measured with the FAR assay for
particle irradiation up to 50 Gy, while the 60Co data were consistent with a
random breakage model.
Fragment analysis
To further examine the dependence on radiation quality, cells were exposed to
accelerated nitrogen ions at four different LET values in the range 80–225
keV/µm. Helium ions having an average LET of 40 keV/µm were also used,
but their results will not be discussed further in this presentation, as the helium
beam consists of a mixture of low and high-LET particles.
Non-random fragmentation
The frequency of DNA fragments over the molecular weight range investigated
was calculated for several doses in the range 0–200 Gy for all radiation qualities
tested. Examples of the results for 125 keV/µm nitrogen ions at 50, 70 and 100
Gy are plotted in Figure 5a. Here the data calculated for each size interval (Eq.

28
2) were divided by δMi (see Table 3) and plotted against fragment size, so that
the area under the curves corresponds to the number of fragments induced per
basepair. The high-LET fragmentation patterns did not show any resemblance
to the distributions expected when the breaks had been induced randomly
along the DNA molecules (dotted curves in Figure 5a). Further, the mode of
DSB induction was examined for the two radiation qualities by calculating the
random DSB yield for measured levels of the fraction of DNA smaller than
different threshold sizes (Figure 5b). The resulting yields for the different sizes
could intuitively be perceived as follows; to produce the measured amount of
DNA smaller than these sizes by random breakage alone, the calculated yields
would be needed. Thus, if the breaks were induced randomly over the genome,
DSB yields would be independent of the fragment sizes studied and the result
would be a horizontal line in Figure 5b. With small deviations, the result for
photons followed a similar curve, with a DSB yield around 6 × 10-9 DSBs bp-1
Gy-1, regardless of threshold size (cf. Table 4), while the high-LET curve
revealed an increased likelihood that shorter DNA fragments would result. For
instance, an RBE of about 2.5 for DSB induction was measured when analysing
fragments smaller than ~100 kbp, and the RBE decreased to below unity when
increasing the threshold to sizes greater than 2–3 Mbp.

29
0.01
0.1
1
10 100 1000 104
125 keV/µm (100 Gy)125 keV/µm (70 Gy)125 keV/µm (50 Gy)
Freq
uenc
y of
DNA
frag
men
ts (x
10-1
2 bp-2
)
Fragment size (kbp)
a
0
5
10
15
20
100 1000
125 keV/µm N-ionsCo-60 photonsDN
A d
sb y
ield
( x1
0-9 d
sb b
p-1 G
y-1)
Threshold size (kbp)
b
Figure 5. Excess of short and medium-sized fragments. (a) Frequency of DNA
fragments as a function of mean fragment size, following exposure to different
doses of 125 keV/µm nitrogen ions. Dotted lines represent the corresponding
fragment distributions, as expected from random breakage (Eq. 1) with a DSB
yield of 5.8 × 10-9 DSBs bp-1 Gy-1. (b) Induction yields calculated with the
random-breakage formula (Eq. 1), using varying threshold sizes (see main text
for details). (Data from Paper II.)

30
Due to the evident excess of short and medium-sized fragments following
exposure to densely ionizing radiation, the total number of breaks has to be
integrated over all fragment sizes, i.e. by fragment analysis. The risk of
underestimating the number of induced DNA DSBs could thereby be
diminished. The results are listed in Table 4 (data from Papers II and IV), and
Table 5 (data from Höglund and Stenerlöw*)
Table 4. DNA double-strand break induction yields after irradiation at different
linear energy transfer (LET)
Radiation LET(keV/µm)
Yield±SEa
(10-9 DSB/bp/Gy)
Yield±SEb
(DSB/traversal)
RBEc
DSB induction60Co < 0.5 5.8±0.1 - 1
N ions 80 8.6±0.9 4.5±0.2 1.5
125 8.6±0.3 6.7±0.2 1.5
175 7.6±1.0 8.4±0.9 1.3
225 6.9±0.7 10.0±0.3 1.2a Calculated using DNA fragmentation analysis.
b The yield as expressed by the number of breaks induced per particle traversal per cell, assuming a
cross-sectional area of the cell nucleus of 150 µm2 and 6x109 bp per diploid cell in G0/G1.c RBE for DSB induction was defined as the quotient between the yield for the test irradiation and
the 60Co-reference yield.
31
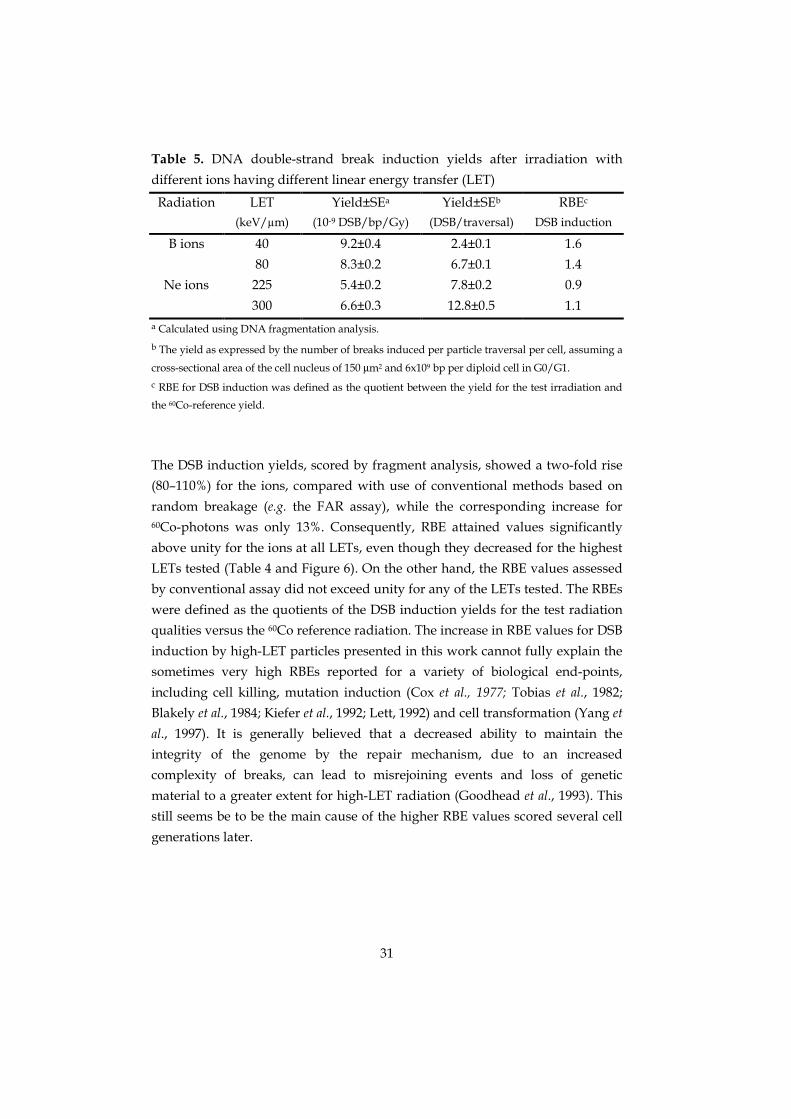
Table 5. DNA double-strand break induction yields after irradiation with
different ions having different linear energy transfer (LET)
Radiation LET(keV/µm)
Yield±SEa
(10-9 DSB/bp/Gy)
Yield±SEb
(DSB/traversal)
RBEc
DSB induction
B ions 40 9.2±0.4 2.4±0.1 1.6
80 8.3±0.2 6.7±0.1 1.4
Ne ions 225 5.4±0.2 7.8±0.2 0.9
300 6.6±0.3 12.8±0.5 1.1a Calculated using DNA fragmentation analysis.
b The yield as expressed by the number of breaks induced per particle traversal per cell, assuming a
cross-sectional area of the cell nucleus of 150 µm2 and 6x109 bp per diploid cell in G0/G1.c RBE for DSB induction was defined as the quotient between the yield for the test irradiation and
the 60Co-reference yield.
The DSB induction yields, scored by fragment analysis, showed a two-fold rise
(80–110%) for the ions, compared with use of conventional methods based on
random breakage (e.g. the FAR assay), while the corresponding increase for60Co-photons was only 13%. Consequently, RBE attained values significantly
above unity for the ions at all LETs, even though they decreased for the highest
LETs tested (Table 4 and Figure 6). On the other hand, the RBE values assessed
by conventional assay did not exceed unity for any of the LETs tested. The RBEs
were defined as the quotients of the DSB induction yields for the test radiation
qualities versus the 60Co reference radiation. The increase in RBE values for DSB
induction by high-LET particles presented in this work cannot fully explain the
sometimes very high RBEs reported for a variety of biological end-points,
including cell killing, mutation induction (Cox et al., 1977; Tobias et al., 1982;
Blakely et al., 1984; Kiefer et al., 1992; Lett, 1992) and cell transformation (Yang et
al., 1997). It is generally believed that a decreased ability to maintain the
integrity of the genome by the repair mechanism, due to an increased
complexity of breaks, can lead to misrejoining events and loss of genetic
material to a greater extent for high-LET radiation (Goodhead et al., 1993). This
still seems be to be the main cause of the higher RBE values scored several cell
generations later.

32
0
0.5
1
1.5
2
0.1 1 10 100
RB
E, d
sb in
duct
ion
LET (keV/µm)
Figure 6. RBE for DSB induction. The RBE as a function of ionization density for60Co-photons and 80-225 keV/µm nitrogen ions, calculated by fragment
analysis (�). The corresponding RBE values calculated without taking the
fragmentation into account, based on the fraction of DNA <6 Mbp and
assuming random distribution of breaks, are also included (�). The lines
connecting the points are plotted for guidance only, and do not assume any
particular behaviour of the RBE/LET relation. RBE determined by fragment
analysis and published by others is included for comparison. Nitrogen ions at
97 keV/µm (�) and iron ions at 150 keV/µm (�) (Löbrich et al, 1996), and
110 keV/µm α-particles (�)(Newman et al. 1997).
The influence of single-hit events, or intra-track effect, is more pronounced for
high-LET tracks than for the more dispersed ionization produced by photons.
Consequently, additional fragments are produced in the same DNA molecule
by the same particle traversal. Thus, for the same dose, the ions prove to be less
efficient for production of fragments of greater molecular weight (>1 Mbp),
compared with photons, or than expected from random breakage alone (e.g.
Figure 5b). The explanation is probably to be found in the nature of the particle
tracks, with different track structures at different LET, and their interaction
with the genomic structure on several levels in the cell. The importance of DNA
structure inside the 30-nm chromatin fibre for the production of very small

33
fragments (80 bp – 2 kbp) has been shown in both theoretical (Holley and
Chatterjee, 1996) and experimental work (Rydberg, 1996). Such small fragments
could not be detected by the methods used in the present work, due to a lower
limit of about 5 kbp for fragments not migrating out of the electrophoresis gels,
but the contribution to the total yield of DSBs from these small fragments is
limited (see discussion in Paper III and Rydberg, 1996). However, the
higher-order chromatin structure may not be the chief cause of the release of
small and medium-sized fragments. Theoretical presentations, assuming a
random-coil structure of the DNA molecule, have also predicted a significant
increase in correlated breaks, i.e. breaks induced within the same molecule by
one particle track, compared with more sparsely ionizing radiation
(Hutchinson, 1996; Sachs et al., 1995). Experimental data on fragmentation of
naked DNA may provide important information on the role of higher-order
chromatin structure.
Efficiency for DSB induction per particle
Furthermore, when delivering the same dose to the cells for different LET
values, the numbers of traversing ions differ, e.g. a dose of 1 Gy to the cell
nucleus is given by approximately 12 ions at 80 keV/µm, while the same dose is
received when 5 ions traverse the nucleus at 175 keV/µm. To account for this,
and to further clarify LET dependence on DSB induction, the cells were exposed
to nitrogen ions at four different LET values (80–225 keV/µm, Paper IV), and
with boron ions (40 and 80 keV/µm) and neon ions (225 and 300 keV/µm) (data
from Höglund and Stenerlöw*). The effectiveness to cause DNA damage was
analysed for each particle traversal. In contrast to yields expressed as a function
of dose (Paper II), the effectiveness to induce DSBs per particle traversal per cell
increased with increasing LET for the same type of ion (Table 4, 5 and Figure 7).
Further, the heavier neon ions were less effective DSB inducers, compared with
nitrogen ions at the same LET (225 keV/µm, Figure 7). An immediate
conclusion from this is that the slower ions having higher LET values (or the
lighter ions having the same LET value) are more effective DSB inducers due to
the more dense ionization pattern in their tracks. This is in accordance with
earlier studies where low energy protons were more effective mutation
inducers than were α-particles (Belli et al., 1992) or neon ions (Durante et al.,
1998) at the same LET. Also singly charged protons and deutrons, matched in
LET, have proved to be more effective for cell inactivation than doubly charged
particles of the same LET (Folkard et al., 1996). However, at 80 keV/µm, no
significant difference was detected between irradiation with boron or nitrogen

34
ions. This type of analysis and presentation provides information about DNA
fragmentation at the low-dose limit of a single particle traversal per cell.
0
2
4
6
8
10
12
14
50 100 150 200 250 300
Boron ionsNitrogen ionsNeon ions
Yiel
d (D
SB/p
artic
le/c
ell)
LET (keV/µµµµm)
Figure 7. DSB induction per ion traversal, in this instance for boron, nitrogen and
neon ions having different LET values. (Data from Paper IV. Boron and neon
data from Höglund and Stenerlöw*).

35
Rejoining
Kinetics
As described in the previous section, the significant increase in DSB induction
yield for high-LET, reported in this work and elsewhere, cannot fully explain
the relatively high RBE values reported for many other end-points. In the
analysis of DNA repair, such additional breaks might have influenced the
rejoining kinetics. It could be argued that the increased number of DNA
fragments, originating from regional clusters of breaks following high-LET
exposure, will impede the cellular repair mechanism in effectively rectifying the
damage due to enzymatic saturation, but only by virtue of a too large number
of breaks to be repaired. Furthermore, a greater complexity of breaks (Ward,
1985; Goodhead, 1989; Goodhead, 1994; Ward, 1994) may restrict repair
capacity (Roots et al., 1979), with consequent misrepair events, which in turn
would promote, e.g. cell death and mutation induction. Many studies on DSB
repair have shown a slower rejoining of fragments, and a larger proportion of
residual breaks following high-LET irradiation (Ritter et al., 1977; Coquerelle et
al., 1987; Blöcher, 1988; Heilmann et al., 1993; Jenner et al., 1993; Weber and
Flentje, 1993; Stenerlöw et al., 1996). However, in order to detect a possible effect
on DSB rejoining from smaller DNA fragments and/or break complexity in
relation to LET, the fragment size distributions have to be included also in
investigations on rejoining.
The way in which DSB rejoining kinetics and the level of unrejoined DSBs was
dependent on LET in human fibroblast cells, following 0–20 hours of
post-irradiation incubation was investigated by fragment analysis (Paper III).
The fragment analysis has been described above. Briefly, the fragment size
distributions were taken into account by summing up the contributions of all
size intervals, and the total number of DSBs for each time point was calculated.
Rapid rejoining following high-LET
As expected from earlier investigations, the rejoining of DSBs showed a
biphasic behaviour for all radiation qualities tested, but surprisingly, and in
contrast to previous investigations, a pronounced rapid rejoining component
which accounted for most of the rejoining could be detected also following
exposure to densely ionizing radiation (Figure 8). Half-times were similar (~15
minutes) for all radiation qualities tested. More consistent with previous

36
studies, both the proportion of unrejoined DSBs and the fraction of breaks
rejoined by the slow phase (Fslow) increased with LET, and the half-time for the
slow kinetics was 2–3h. Some of the rejoining parameters from Paper III are
summarized in Table 6.
Table 6. DSB rejoining parameters following irradiation with 100 Gy of
different radiation qualitiesa
Radiation LET
(keV/µm)
τ1/2b (fast)
(min)
Fslowc Unrejoined (%)d
60Co <0.5 19 ± 8 0.07 ± 0.03 4 ± 3
N ions 80 17 ± 2 0.16 ± 0.04 5 ± 2
125 13 ± 2 0.36 ± 0.10 7 ± 4
175 15 ± 7 0.29 ± 0.02 12 ± 4
225 12 ± 4 0.38 ± 0.05 12 ± 2a Based on fragment analysis for sizes up to 5.7 Mbp (±SE). The parameters were obtained from
double-exponentional curves fitted to the data.b Time to halve DSB level for the rapid phase of rejoining. The number of unrejoined DSBs at 20–22
hours was subtracted to measure the kinetics of rejoinable breaks only.c Fraction of DSBs rejoined by the slow phase.
d Proportion of initially induced DSBs, measured by fragmentation analysis.
Although Fslow increased significantly with LET vis-à-vis photon irradiation,
showing a five-fold increase at 225 keV/µm, the fractions measured were
smaller than earlier reported results (e.g. Stenerlöw et al., 1996). The main cause
of the difference is that the smaller fragments (<1 Mbp), induced by spatially
correlated DSBs, were revealed and included in the present work by fragment
analysis. This yielded a significant increase in the degree of initial damage
(Paper II) that was not detected by earlier conventional assays. However, within
a few hours of repair, both methods showed similar levels of damage (Figure 8),
and it was clear that the majority of the breaks were rejoined in the rapid phase
even for the high-LET ions. This may also explain the rather low levels of
unrejoined breaks presented (Table 6).

37
0
200
400
600
800
1000
0 1 2 3 4 5 6 7
125 keV/µµµµm nitrogen ionsBlöcher repair
Time (h)
Num
ber o
f DSB
(x10
9 bp-1
)
22
Figure 8. Rapid rejoining for high-LET. The number of DSB per Gbp (109 bp) as a
function of post-irradiation incubation time. Cells were irradiated with 125
keVµm nitrogen ions and the number of breaks was determined using
fragmentation analysis (see main text for details). The dotted curve is included
for comparison, representing the number of breaks as determined by Blöcher’s
conventional formula (Eq. 1).
The rapid rejoining described that was evident after high-LET exposure should
not be interpreted as an indication of a quicker rejoining of short DNA
fragments, in comparison with the rejoining of longer fragments. When large
numbers of short fragments are joined to other shorter fragments, or to larger
pieces of DNA, longer fragments are formed. In this way they gradually move
through the fragment size distribution into gel segments representing
increasing fragment sizes, with the net effect of an apparent rapid removal of
shorter fragments. Furthermore, in fragment analysis, the number of fragments
smaller than 6 Mbp is used to estimate the number of DSBs. This is a high-dose
approximation, and at low doses and/or long repair times, there is a risk of
underestimating the number of DSBs. The reason is that the longer fragments,
produced when shorter fragments are rejoined during repair, are not included
in the analysis. To evaluate the contribution of these ‘lost fragments’ we have to
find ways to measure much longer fragments, but also a detailed theoretical

38
modelling of DNA size distributions following high-LET would be helpful
(Sachs et al., 1998).
The tendency toward slower repair as LET values increase, evident in Table 6, is
clearly visualized in Figure 9, where the 80 and 225 keV/µm data are
compared. It can be concluded from the figure that neither the initial number of
fragments, nor their initial size distribution, can explain the obvious slower
repair at higher LET values. The initial level of fragmentation (t=0) below 2
Mbp was the same, or even higher, for the 80 keV/µm ions, compared with the
higher LET. After 4 hours of incubation for DNA repair the situation was the
reverse. Altogether, the results presented here imply that increased DSB
complexity is the determining factor for the reduced repair capacity seen after
exposure to high-LET radiation.
10-8
10-7
100 1000
80 keV/µm, 0h80 keV/µm, 4h225 keV/µm, 0h225 keV/µm, 4h
fract
ion
of D
NA/
δM
(bp-1
)
Fragment size (kbp)
Figure 9. Distribution pattern and rejoining. The fraction of DNA at t=0h and t=4h
was normalized using the width of each size interval (gel segment) and plotted
versus fragment size for 80 and 225 keV/µm nitrogen ions.
(Data from Paper III.)

39
Rejoining and end-points
The prevailing mode for DNA DSB repair in G0/G1 mammalian cells is the
homology-independent end-joining pathway (Wang et al., 1997), a pathway
initiated by the binding of a Ku70/Ku86 protein complex to the DNA ends,
followed by association of the DNA-dependent protein kinase catalytic subunit
(DNA-PKcs). The Ku86 protein is closely related to the fast kinetics (Okayasu
and Iliakis, 1994). However, the biphasic rejoining (see Figure 8) suggests that at
least one additional DNA repair pathway is present, probably representing a
slower process. The present work presents evidence nominating the complexity
of breaks as the determining factor for reduced reparability. The increasing
fraction of DSBs rejoined by the slow phase, together with the elevated level of
remaining DSBs measured after irradiation at high-LET values, could reflect the
failure of the rapid repair processes to rejoin such complex lesions, thereby
increasing the likelihood of misrejoning events. However, recent studies on
misrejoining in a 3.2 Mbp restriction fragment did not show any significant
change with LET (Löbrich et al., 1998). In contrast, mutation analyses have
shown an LET dependency with increased yields for partial or total deletions
ranging from about 5–45 kbp at the HPRT locus for high-LET (Amundson et al.,
1996; Metting et al., 1992). Biophysical modelling also predicts more total
deletions of sizes below 500 kbp (Wu et al., 1998) after high-LET irradiation. It is
conceivable that the fragments produced by correlated breaks within the same
loop domain may be lost in the repair process because they are freely diffusible,
resulting in an increased induction of deletions. Together with data presented
for fragment analysis, this supports the hypothesis that high-LET radiation
damage is related to higher order structures of chromatin. A comparison with
published data on the rejoining of chromosomal fragments as measured by the
premature chromosome condensation technique (PCC) cannot be done as
directly as in the reasoning above. First, the excess of fragments <1 Mbp
observed by fragmentation analysis in the present study cannot be detected by
the PCC technique, and second, a considerable amount of rejoining takes place
during the 20–60 minutes of fusion with the PCC technique. Both these events
lead to an underestimation of the initial number of induced breaks, and will
considerably affect the measured level of unrejoined breaks.

40
Rejoining after exposure to the same number of particles
Complexity of breaks
As regards DSB induction, the rejoining of clustered DSBs after irradiation with
the same number of nitrogen ions traversing the samples was investigated
(Paper IV). The results further supported the complexity of the breaks as the
most important parameter. A slower overall rejoining was found for the higher
LET values tested, with still substantial levels of fragmentation following 1 hour
incubation for repair. For the lowest LET, 80 keV/µm, a major part of the
induced fragments was rejoined already after 30 min (Figure 10).
0
0.2
0.4
0.6
0.8
1
100 1000
0 h0.5 h1 h4 h20 h
80 keV/µµµµm
30 100 1000
175 keV/µµµµm
30
Fragment size (kbp)
Frac
tion
of D
NA/ δδ δδ
M (a
rbrit
ary
units
)
Figure 10. Fragmentation and repair. Fragment size distributions as a function of
mean fragment size after rejoining 0–20h. The cells were irradiated with 5
nitrogen ions per µm2 at 80 and 175 keVµm. Both curves representing the
initial damage (t=0) were normalized by setting the maximum values to unity.
(Data from Paper IV.)
When studying the fragment distributions following different repair times, it
was found that although the smaller fragments were rejoined relatively quickly
even for the 175 keV/µm ions, the number of fragments larger than 2 Mbp did
not change significantly during the first hour of repair at the higher LET. An
alternative explanation for the slower rejoining at higher LET, besides the
increased complexity of the breaks, could be the fact that more breaks are
induced at higher LETs, and that the cellular repair mechanism needs more
time to process all these breaks, due to saturation. However, the results
described above (Paper III) showed that for the same initial number of breaks,

41
and with similar fragment size distributions, those induced by the higher LET
rejoined more slowly. A complementary explanation could be that the slower
rejoining is caused by DSBs induced within compact chromatin structures. Such
breaks might be produced predominantly by direct ionizations as from
high-LET particles, and if there is limited access for repair enzymes/proteins to
such condensed structures, this could result in a reduced capacity of repair
(Ahnström et al., 2000). More experimental work is needed to confirm (or
exclude) this.
Chromatin organization
There is both theoretical and experimental evidence that chromatin
organization on the sub-kilobasepair level, e.g. from nucleosomal winding of
DNA in the 30 nm chromatin fibre, plays a significant role in the induction of
very short DNA fragments (≤2 kbp) — sometimes called “Rydberg fragments”
— for both low and high-LET radiation (Holley and Chatterjee, 1996; Rydberg,
1996). Correlated breaks over much larger distances are also expected due to
the high compaction of the genomic DNA. The packing of DNA is believed to
involve chromatin loops attached to a nuclear matrix (Bodnar, 1988). However,
the possible influence on DNA fragmentation of higher orders of genomic
organization in interphase cells has not been fully elucidated in the literature. It
is possible that such influence could be detected by investigating fragmentation
patterns within a wide molecular weight range, induced by radiation of varying
quality.
When charged particles of the same type (but with differing ionization density)
pass through matter, they show different patterns of energy deposition around
the primary track, due to differences in kinetic energy. The higher velocity of
the ions having lower LET values generates comparatively long-range
secondary electrons, δ-electrons; consequently the energy is distributed more
sparsely throughout the genome. In the present study it is suggested that these
differences in track structure, in combination with target structures, e.g. loop
domains, did influence the yield as well as the size distribution of the induced
damage.

42
Substructures due to chromatin organization?
As illustrated in Figure 11 (left panels), the high-LET ion irradiation tilted the
fragment size distributions in favour of smaller fragments. Furthermore,
substructures were introduced in the distributions, that were not expected from
random breakage (dotted lines). These substructures, or ‘peaks’, were found at
fragment sizes in the range 50–200 kbp and 0.5–1.5 Mbp. As mentioned, the
interphase chromatin is believed to be folded into loops. The exact sizes of these
loops have not yet been accurately characterized, but some evidence of
Mbp-sized loops does exist (Sachs et al., 1995; Yokota et al., 1995). Also smaller
loops, in the range 10–200 kbp, have been suggested in the literature (Bodnar,
1988; Filipski et al., 1990; Jackson et al., 1990; Vogelstein et al., 1980). The peak
structures observed in the present work could be the result of spatially
correlated and clustered breaks within such loops. To isolate the role of
high-order chromatin loops from correlated breaks originating from random-
coiled structures of the genome only, as predicted by some theoretical works
(Hutchinson, 1996; Sachs et al., 1995), detailed analysis of fragmentation
following high-LET irradiation of e.g. naked DNA could be performed, but such
investigations were beyond the scope of this work.

43
0
1 10-7
2 10-7
3 10-7
4 10-7
100 1000
a) 60Co photons
-3 10-7
-2 10-7
-1 10-7
0
1 10-7
2 10-7
100 1000
exp - rand 8 DSB bp-1 Gy-1
exp - rand 6 DSB bp-1 Gy-1
0
1 10-7
2 10-7
3 10-7
100 1000
b) 125 keV/µµµµm
-3 10-7
-2 10-7
-1 10-7
0
1 10-7
2 10-7
100 1000
exp - rand 8 DSB bp-1 Gy-1
exp - rand 6 DSB bp-1 Gy-1
exp - rand 3 DSB bp-1 Gy-1
0
1 10-7
2 10-7
3 10-7
4 10-7
5 10-7
6 10-7
100 1000
c) 225 keV/µµµµm
-3 10-7
-2 10-7
-1 10-7
0
1 10-7
2 10-7
100 1000
exp - rand 8 DSB bp-1 Gy-1
exp - rand 6 DSB bp-1 Gy-1
exp - rand 4 DSB bp-1 Gy-1
Frac
tion
of D
NA/ δδ δδM
(bp-1
)
Non-
rand
om c
ompo
nent
(bp-1
)
Fragment size (kbp)
Figure 11. Fragmentation patterns. The fraction of DNA as a function of fragment
size for (a) 150 Gy 60Co-photons, (b) 100 Gy 125 keV/µm nitrogen ions, and
(c) 200 Gy 225 keV/µm nitrogen ions. Dotted lines represent theoretical
distributions assuming corresponding doses and random breakage, with the
DSB yields 6 and 8 × 10-9 DSB bp-1 Gy-1 (panel a), 3, 6 and 8 × 10-9 DSB bp-1
Gy-1 (panel b) and 4, 6 and 8 × 10-9 DSB bp-1 Gy-1 (panel c). Corresponding
panels (right) illustrate the non-random component of the fragment
distributions (see main text for details). (Data from Paper IV.)

44
Even though not pronounced, a small peak around 100 kbp was seen also for
the photons, indicating that a limited but significant non-random contribution
was present. Such single-hit behaviour, producing smaller DNA fragments
(<1 Mbp), has been reported for low-LET X-ray (Löbrich et al., 1996; Newman et
al., 1997; Rydberg, 1996) and high-energy electrons (Sak et al., 1996).
In Figure 11 (right panels), the non-random contribution to the breakage
patterns is illustrated by subtracting the random curves suggested in the left
panels (from the experimental data). From the appearance of the resulting
fragment size distributions, a close relation to LET is evident regarding the
magnitude of the non-random component, but involvement of higher orders of
DNA folding is also indicated. Due to lower velocity, a particle having a higher
LET value deposits more of its energy by ionization in the direct vicinity of its
path than do particles at a lower LET, which increases the likelihood of
correlated breaks within loop domains, and this may explain the increased non-
random shape of the curves. Thus, the random yield may decrease with
increasing LET, while the number of associated breaks increases. The random
yields chosen in Figure 11 may be too high, as some published data suggest a
formation of more than two additional DSBs for each randomly induced break,
due to clustering of breaks within a few Mbp for high-LET radiation (Sachs et
al., 1998). In fact, the model presented by Sachs et al. 1998 yielded an average
multiplicity of 4 breaks, i.e. 1 random break and 3 non-random breaks, in each
cluster when tested on published data for ions at ~100 keV/µm (Löbrich et al.,
1996; Newman et al., 1997). The same multiplicity, if applied to ion data
presented in Paper II (see Table 4) would give a random yield of around 2–2.5 ×10-9 DSBs bp-1 Gy-1.
This yield is similar to the one used in paper I, where a very simple theoretical
description of this phenomenon was presented. The dose response for the
amount of DNA smaller than 6 Mbp that was released into the electrophoresis
gel was reconstructed, and a close agreement with experimental data was
achieved by applying a relatively low random yield (3 × 10-9 DSBs bp-1 Gy-1) in
combination with additional breaks within 2 Mbp (Paper I). Naturally, in order
to fully isolate the random from the non-random component of the DNA
fragmentation, and to elucidate the relation to chromatin organization, far more
elaborate examinations and detailed theoretical modelling will need to be
performed; this was beyond the scope of the present work.

45
SUMMARY AND CONCLUSIONS
A substancial proportion of non-randomly induced DNA double-strand breaks
was evident when large radiation–induced DNA fragments were analyzed by
pulsed-field gel electrophoresis (PFGE) following exposure to 125 keV/µm
nitrogen ions in the dose-range 0.5–30 Gy. This was in contrast to results for60Co-photons. The non-random contribution to the fraction of DNA released
into the gel may be explained by introducing one or several correlated breaks
within 2 Mbp from a randomly induced break.
The significant non-random contribution to the number of radiation-induced
DNA DSBs for high-LET irradiation, was further confirmed by detailed
fragmentation analysis after exposing normal human fibroblast cells to nitrogen
ions in the LET range 80-225 keV/µm. The radiation quality had a strong
influence on the initial DNA fragment distribution, with an elevated number of
small and medium-sized fragments (5 kbp – 1 Mbp) following high-LET
irradiation. When the fragment size distributions were taken into account, the
ion yields (DSBs bp-1 Gy-1) were some 80–110% higher than those calculated
when disregarding the distributions, i.e. assuming random breakage (FAR
assay). For gamma irradiation the corresponding increase were 13%, and
consequently, the relative yields for DSB induction increased to values
significantly above unity (1.2–1.5) for the high-LET irradiation, however
decreasing for increasing LET values. On the other hand, the relative yields
assessed by conventional assay did not exceed unity for any of the LETs tested.
The DNA fragmentation pattern, i.e. how the DNA DSBs are distributed along
the chromosomes, showed to have an influence on the DSB rejoining in normal
human fibroblasts. DNA fragment analysis detected LET dependent differences
in the amount of residual damage following 20h of incubation for repair, while
no influence of LET on the half-times of the biphasic repair kinetics was found.
In contrast to earlier investigations, the data presented here show a significant
proportion of repair by the rapid phase also for high-LET irradiaion. In fact, the
majority of breaks were restored by the rapid phase for all radiation qualities
tested. However, the fraction of breaks rejoined by the slow phase were closely
related to the radiation quality, with a five-fold increase for the highest LET
values investigated compared to the sparsely ionizing gamma photons. The

46
present work presents evidence nominating the complexity of breaks as one
important determining factor for reduced reparability reported following
high-LET exposure.
The number of breaks induced per particle traversal through a cell nucleus was
calculated for nitrogen ions in the LET range 80–225 keV/µm, boron ions at 40
and 80 keV/µm, and neon ions at 225 and 300 keV/µm. A close relation to LET
was evident, with a continously growing efficiency for fragment induction
when ions pass the cell nuclei at increasing LET values. A nitrogen ion at 225
keV/µm induced more than twice as many DSBs on its passage through the cell
nucleus, compared to an 80 keV/µm ion, and the elevated efficiency for the
higher LET value is probably due to differences in particle track structure. This
was further supported by the result on the heavier neon ions, having similar
LET value (225 keV/µm), which showed a decreased effectiveness for DSB
induction, compared with the nitrogen ions. Furthermore, the shape of the
induced DNA fragment size distributions, following the nitrogen ion exposure,
was highly correlated to the LET, showing sub-structures that may reveal a role
of the higher order of chromatin organisation in the induction and rejoining of
radiation-induced DNA damage.
Data presented in this thesis provide strong evidence for a non-random
induction of DNA DSBs in mammalian cells following high-LET irradiation,
with a probable relation to correlated breaks within the genomic structure.
Futher, the presented work clearly demonstrates the importance of including
detailed information about the fragment distributions in the analyses.
It is believed that data on initial fragmentation and subsequent rejoining, like
the data presented here, may serve as useful input to biophysical modelling
trying to explain the processes causing severe radiation-induced biological
effects. Moreover, such data can increase the general understanding of initial
processes causing radiation-induced DNA damage, and the mechanisms
leading to its restoration.

47
REFERENCES
AHNSTRÖM, G. AND EDVARDSSON, K-A., Radiation-induced single-strand breaks
in DNA determined by rate of alkaline strand separation and
hydroxylapatite chromotography: an alternative to velocity
sedimentation. Internatioal Journal of Radiation Biology, 26, 493–97, (1974).
AHNSTRÖM, G., NYGREN, J. AND ERIKSSON, S., The effect of dimethyl sulphoxide
on the induction and repair of double-strand breaks in human cells
after irradiation with gamma-rays and accelerated ions: rapid or slow
repair may depend on accessibility of breaks in chromatin of different
compactness. International Journal of Radiation Biology, 76, 533–8 (2000).
AMALDI, U., Hadrontherapy in the world and the programmes of the TERA
Foundation. Tumori, 84, 188–99, (1998).
AMUNDSON, S.A., CHEN, D.J. AND OKINAKA, R.T., Alpha particle mutagenesis of
human lymphoblastoid cell lines. International Journal of Radiation
Biology, 70, 219–26 (1996).
BARENDSEN, G.W., KOOT, C.J., VAN KERSEN, G.R., BEWLEY, D.K., FIELD, S.B. AND
PARNELL, C.J., The effect of oxygen on impairment of the proliferative
capacity of human cells in culture by ionizing radiations of different
LET. International Journal of Radiation Biology, 10, 317–27 (1966).
BARTH, R.F., SOLOWAY, A.H., GOODMAN, J.H., GAHBAUER, R.A., GUPTA, N.,
BLUE, T.E., YANG, W. AND TJARKS, W., Boron neutron capture therapy of
brain tumors: an emerging therapeutic modality. Neurosurgery, 44,
433-51, (1999).
BELLI, M., GOODHEAD, D., IANZINI, F., SIMONE, G. AND TABOCCINI, M., Direct
comparison of biological effectiveness of protons and alpha-particles of
the same LET. II. Mutation induction at the HPRT locus in V79 cells.
International Journal of Radiation Biology, 61, 625–29, (1992).
BLAKELY, E.A., NGO, F.Q.H., CURTIS, S.B. AND TOBIAS, C.A., Heavy ion
radiobiology: Cellular studies. In: Advances in Radiation Biology 11.
Edited by J.T. Lett, pp295–389 (Academic Press, New York, 1984)
BLAKELY, E.A. AND KRONENBERG, A., Heavy-ion radiobiology: new approaches
to delineate mechanisms underlying enhanced biological effectiveness.
Radiation Research, 150, S126–45 (1998).
BLÖCHER, D., DNA double-strand break repair determines the RBE of alpha-
particles. International Journal of Radiation Biology, 54, 761–71 (1988).

48
BLÖCHER, D., In CHEF electrophoresis a linear induction of dsb corresponds to
a nonlinear fraction of extracted DNA with dose. International Journal of
Radiation Biology, 57, 7–12 (1990).
BODNAR, J.W., A domain model for eukaryotic DNA organization: a molecular
basis for cell differentiation and chromosome evolution. Journal of
Theoretical Biology, 132, 479–507 (1988).
BRYANT, P.E., Enzymatic restriction of mammalian cell DNA PVU II and Bam
H1: evidence for the double-strand break origin of chromosomal
aberrations. International Journal of Radiation Biology, 46, 57–65, (1984).
CANTOR, C.R., SMITH, C.L. AND MATHEW, M.K., Pulsed-field gel electrophoresis
of very large DNA molecules. Annual Review of Biophysics and
Biophysical Chemistry, 17, 287–304 (1988).
CARLSSON, J., SJÖBERG, S. AND LARSSON, B.S., Present status on boron neutron
capture therapy. Acta Oncologica, 31, 803–13, (1992).
CASTRO, J.R., Results of heavy ion radiotherapy. Radiation and Environmental
Biophysics, 34, 45–48, (1995).
CEDERVALL, B., WONG, R., ALBRIGHT, N., DYNLACHT, J., LAMBIN, P. AND DEWEY,
W.C., Methods for the quantification of DNA double-strand breaks
determined from the distribution of DNA fragment sizes measured by
pulsed-field gel electrophoresis [published erratum appears in Radiat
Res 1995 Oct;144(1):122]. Radiation Research, 143, 8–16 (1995).
COOK, V.E. AND MORTIMER, R.K., A quantitative model of DNA fragments
generated by ionizing radiation, and possible experimental
applications. Radiation Research, 125, 102–6 (1991).
COQUERELLE, T.M., WEIBEZAHN, K.F. AND LÜCKE-HUHLE, C., Rejoining of
double strand breaks in normal human and ataxia-telangiectasia
fibroblasts after exposure to 60Co gamma-rays, 241Am alpha-particles
or bleomycin. International Journal of Radiation Biology, 51, 209–18 (1987).
COX, R., THACKER, J., GOODHEAD, D.T. AND MUNSON, R.J., Mutation and
inactivation of mammalian cells by various ionizing radiations. Nature,
267, 425–27, (1977).
DAGHIGHIAN, F., BARENDSWAARD, E., WELT, S., HUMM, J., SCOTT, A.,
WILLINGHAM, M.C., MCGUFFIE, E., OLD, L.J. AND LARSON, S.M.,
Enhancement of radiation dose to the nucleus by vesicular
internalization of iodine-125-labeled A33 monoclonal antibody. Journal
of Nuclear Medicine, 37, 1052–7 (1996).

49
DURANTE, M., GROSSI, G.F., NAPOLITANO, M., PUGLIESE, M. AND GIALANELLA,
G., Chromosome damage induced by high-LET alpha-particles in
plateau-phase C3H 10T1/2 cells. International Journal of Radiation
Biology, 62, 571–80 (1992).
DURANTE, M., GROSSI, G.F., GIALANELLA, G., PUGLIESE, M., NAPPO, M. AND
YANG, T.C., Effects of alpha-particles on survival and chromosomal
aberrations in human mammary epithelial cells. Radiation and
Environmental Biophysics, 34, 195–204 (1995).
DURANTE, M., CELLA, L., FURUSAWA, Y., GEORGE, K., GIALANELLA, G., GROSSI,
G., PUGLIESE, M., SAITO, M. AND YANG, T.C.,The effect of track structure
on the induction of chromosomal aberrations in murine cells.
International Journal of Radiation Biology, 73, 253–62 (1998).
EICKHOFF, H., HABERER, T., KRAFT, G., KRAUSE, U., RICHTER, M., STEINER, R. AND
DEBUS, J., The GSI cancer therapy project. Strahlentherapie und Onkologie,
175, 21–4, (1999).
ELKIND, M.M. AND SUTTON, H., Radiation response of mammalian cells grown
in culture. I. Repair of x-ray damage in surviving Chinese hamster cells.
Radiation Research, 13, 556–93, (1960).
FILIPSKI, J., LEBLANC, J., YOUDALE, T., SIKORSKA, M. AND WALKER, P.R.,
Periodicity of DNA folding in higher order chromatin structures.
EMBO Journal, 9, 1319–27 (1990).
FOLKARD, M., PRISE, K.M., VOJNOVIC, B., NEWMAN, H.C., ROPER, M.J. AND
MICHAEL, B.D., Inactivation of V79 cells by low-energy protons,
deutrons and helium-3 ions. International Journal of Radiation Biology, 69,
729–38, (1996).
FRANKENBERG, D., BREDE, H.J., SCHREWE, U.J., STEINMETZ, C., FRANKENBERG-
SCHWAGER, M., KASTEN, G. AND PRALLE, E., Induction of DNA double-
strand breaks by 1H and 4He lons in primary human skin fibroblasts in
the LET range of 8 to 124 keV/microm. Radiation Research, 151, 540–9
(1999).
FRIEDLAND, W., JACOB, P., PARETZKE, H. G. AND STORK, T., Monte Carlo
simulation of the production of short DNA fragments by low-linear
energy transfer radiation using higher-order DNA models. Radiation
Research, 150, 170–82, (1998).
GOODHEAD, D.T., The initial physical damage produced by ionizing radiations.
International Journal of Radiation Biology, 56, 623–34 (1989).

50
GOODHEAD, D.T., THACKER, J. AND COX, R., Weiss Lecture. Effects of radiations
of different qualities on cells: molecular mechanisms of damage and
repair. International Journal of Radiation Biology, 63, 543–56 (1993).
GOODHEAD, D.T., Initial events in the cellular effects of ionizing radiations:
clustered damage in DNA. International Journal of Radiation Biology, 65,
7–17 (1994).
HEILMANN, J., RINK, H., TAUCHER SCHOLZ, G. AND KRAFT, G., DNA strand break
induction and rejoining and cellular recovery in mammalian cells after
heavy-ion irradiation. Radiation Research, 135, 46–55 (1993).
HEILMANN, J., TAUCHER SCHOLZ, G. AND KRAFT, G., Induction of DNA double-
strand breaks in CHO-K1 cells by carbon ions. International Journal of
Radiation Biology, 68, 153–62 (1995).
HOLLEY, W. AND CHATTERJEE, A., Clusters of DNA induced by ionizing
radiation: formation of short DNA fragments. I. Theoretical modeling.
Radiation Research, 145, 188–99 (1996).
HUTCHINSON, F., Formation of two double-strand breaks in the same DNA
molecule by a single high-energy photon or ionizing particle.
International Journal of Radiation Biology, 70, 505–12 (1996).
ICRU, Radiation Quantities and Units, ICRU report, 10a (1962).
ILIAKIS, G., The role of DNA double strand breaks in ionizing radiation-induced
killing of eukaryotic cells. Bioessays, 13, 641–8 (1991).
JACKSON, D.A., DICKINSON, P. AND COOK, P.R., The size of chromatin loops in
HeLa cells. EMBO Journal, 9, 567–71 (1990).
JENNER, T.J., DELARA, C.M., O'NEILL, P. AND STEVENS, D.L., Induction and
rejoining of DNA double-strand breaks in V79-4 mammalian cells
following gamma- and alpha-irradiation. International Journal of
Radiation Biology, 64, 265–73 (1993).
JIN, Y., YIE, T. AND CAROTHERS, A., Non-random deletions at the dihydrofolate
reductase locus of Chinese hamster ovary cells induced by alpha-
particles simulating radon. Carcinogenesis, 16, 1981–91 (1995).
JOSTES, R.F., Genetic, cytogenetic, and carcinogenic effects of radon: a review.
Mutation Research, 340, 125–39 (1996).
KAMPF, G., Steps towards cancer therapy with radionuclides – a review
including radiation biophysical aspects. Radiobiologia, Radiotherapia, 31,
215–29 (1990).

51
KASSIS, A.I., SASTRY, K.S.R. AND ADELSTEIN, S.J., Kinetics of uptake, retention,
and radiotoxocity of 125IUdR in mammalian cells: implications of
localized energy deposition by Auger processes. Radiation Research, 109,
78–89, (1987).
KIEFER, J., Heavy ion effects on cells: chromosomal abberations, mutations and
neoplastic transformations. Radiation and Environmental Biophysics, 31,
279–88, (1992).
KRAFT, G., The radiobiological and physical basis for radiotherapy with protons
and heavier ions. Strahlentherapie und Onkologie, 166, 10–3 (1990).
KRAFT, G., SCHOLZ, M. AND BECHTHOLD, U., Tumor therapy and track structure.
Radiation and Environmental Biophysics, 38, 229–37 (1999).
KRAXENBERGER, F., WEBER, K.J., FRIEDL, A.A., ECKARDT-SCHUPP, F., FLENTJE, M.,
QUICKEN, P. AND KELLERER, A.M., DNA double-strand breaks in
mammalian cells exposed to gamma-rays and very heavy ions.
Fragment-size distributions determined by pulsed-field gel
electrophoresis. Radiation and Environmental Biophysics, 37, 107–15
(1998).
LARSSON, B., On the application of a 185 MeV proton beam to experimental
cancer therapy and neurosurgery. Thesis, (1962).
LETT, J.T., Damage tocellular DNA from particulate radiations, the efficacy of its
processing and the radiosensitivity of mammalian cells. Emphasis on
DNA double strand breaks and chromatin breaks. Radiation and
Environmental BiophysicsI, 31, 257–77, (1992).
LÖBRICH, M., COOPER, P.K. AND RYDBERG, B., Non-random distribution of DNA
double-strand breaks induced by particle irradiation. International
Journal of Radiation Biology, 70, 493–503 (1996).
LÖBRICH, M., COOPER, P.K. AND RYDBERG, B., Joining of correct and incorrect
DNA ends at double-strand breaks produced by high-linear energy
transfer radiation in human fibroblasts. Radiation Research, 150, 619–26
(1998).
METTING, N.F., PALAYOOR, S.T., MACKLIS, R.M., ATCHER, R.W., LIBER, H.L. AND
LITTLE, J.B., Induction of mutations by bismuth-212 alpha particles at
two genetic loci in human B-lymphoblasts. Radiation Research, 132,
339-45 (1992).
NEWMAN, H.C., PRISE, K.M., FOLKARD, M. AND MICHAEL, B.D., DNA double-
strand break distributions in X-ray and alpha-particle irradiated V79
cells: evidence for non-random breakage. International Journal of
Radiation Biology, 71, 347–63 (1997).

52
NYGREN, J. AND AHNSTRÖM, G., DNA double- and single-strand breaks induced
by accelerated He2+ and N6+ ions in human cells: relative biological
effectiveness is dependent on the relative contribution of the direct and
indirect effects. International Journal of Radiation Biology, 70, 421–7 (1996).
OKAYASU, R. AND ILIAKIS, G., Evidence that the product of the xrs gene is
predominantly involved in the repair of a subset of radiation-induced
interphase chromosome breaks rejoining with fast kinetics. Radiation
Research, 138, 34–43 (1994).
ORBACH, M.J., VOLLRATH, D., DAVIS, R.W. AND YANOFSKY, C., An
electrophoretic karyotype of Neurospora crassa. Molecular Cell Biology,
8, 1469–73 (1988).
PRISE, K.M., DAVIES, S. AND MICHAEL, B.D., The relationship between radiation-
induced DNA double-strand breaks and cell kill in hamster V79
fibroblasts irradiated with 250 kVp X-rays, 2.3 MeV neutrons or 238Pu
alpha-particles. International Journal of Radiation Biology, 52, 893–902
(1987).
PRISE, K.M., FOLKARD, M., DAVIES, S. AND MICHAEL, B.D., The irradiation of V79
mammalian cells by protons with energies below 2 MeV. Part II.
Measurement of oxygen enhancement ratios and DNA damage.
International Journal of Radiation Biology, 58, 261–77 (1990).
PRISE, K.M., FOLKARD, M., NEWMAN, H.C. AND MICHAEL, B.D., Effect of
radiation quality on lesion complexity in cellular DNA. International
Journal of Radiation Biology, 66, 537–42 (1994).
PRISE, K.M., AHNSTRÖM, G., BELLI, M., CARLSSON, J., FRANKENBERG, D., KIEFER,
J., LÖBRICH, M., MICHAEL, B.D., NYGREN, J., SIMONE, G. AND STENERLÖW,
B., A review of dsb induction data for varying quality radiations.
International Journal of Radiation Biology, 74, 173–84 (1998).
RITTER, M.A., CLEAVER, J.E. AND TOBIAS, C.A., High-LET radiations induce a
large proportion of non-rejoining DNA breaks. Nature, 266, 653–5
(1977).
RAJU, M. R., EISEN, Y., CARPENTER, S. AND INKRET, W. C., Radiobiology of αparticles. III. Cell inactivation by α-particle traversals of the cell
nucleus. Radiation Research, 128, 204–209, (1991).
RAJU, M.R., Proton radiobiology, radiosurgery and radiotherapy. International
Journal of Radiation Biology, 67, 237–59, (1995).
RON, E., ionizing Radiation and Cancer Risk: Evidence from Epidemiology.
Radiation Research, 150, S30–S41 (1998).

53
RÖNTGEN, W.C., Uber eine neue Art von Strahlen. Vorläufige (1.) Mitteilung.
Sitzungsberichte das Physikalisch-Medizinischen Gesellschaft, Würzburg
138–48, (1895).
ROOTS, R., YANG, T.C., CRAISE, L., BLAKELY, E.A. AND TOBIAS, C.A., Impaired
repair capacity of DNA breaks induced in mammalian cellular DNA by
accelerated heavy ions. Radiation Research, 78, 38–49 (1979).
RYDBERG, B., LÖBRICH, M. AND COOPER, P.K., DNA double-strand breaks
induced by high-energy neon and iron ions in human fibroblasts. I.
Pulsed-field gel electrophoresis method. Radiation Research, 139, 133–41
(1994).
RYDBERG, B., Clusters of DNA damage induced by ionizing radiation: formation
of short DNA fragments. II. Experimental detection. Radiation Research,
145, 200–9 (1996).
SACHS, R.K., VAN DEN ENGH, G., TRASK, B., YOKOTA, H. AND HEARST, J.E., A
random-walk/giant-loop model for interphase chromosomes.
Proceedings of the National Academy of Sciences USA, 92, 2710–4 (1995).
SACHS, R.K., CHEN, A.M. AND BRENNER, D.J., Review: proximity effects in the
production of chromosome aberrations by ionizing radiation.
International Journal of Radiation Biology, 71, 1–19 (1997).
SACHS, R.K., BRENNER, D.J., HAHNFELDT, P.J. AND HLATKYS, L.R., A formalism
for analysing large-scale clustering of radiation-induced breaks along
chromosomes. International Journal of Radiation Biology, 74, 185–206
(1998).
SAK, A., STUSCHKE, M., STAPPER, N. AND STREFFER, C., Induction of DNA
double-strand breaks by ionizing radiation at the c-myc locus
compared with the whole genome: a study using pulsed-field gel
electrophoresis and gene probing. International Journal of Radiation
Biology, 69, 679–85 (1996).
SCHIMMERLING, W., Radiobiological problems in space. An overview. Radiation
and Environmental Biophysics, 31, 197–203 (1992).
SCHIMMERLING, W., Space and radiation protection: scientific requirements for
space research. Radiation and Environmental Biophysics, 34, 133–7 (1995).
SOUTHERN, E.M., ANAND, R., BROWN, W.R. AND FLETCHER, D.S., A model for the
separation of large DNA molecules by crossed field gel electrophoresis.
Nucleic Acids Research, 15, 5925–43 (1987).
STENERLÖW, B., PETTERSSON, O-A., ESSAND, M., BLOMQUIST, E. AND CARLSSON, J.,
Irregular variations in radiation sensitivity when the linear energy
transfer is increased. Radiotherapy and Oncology, 36, 133–142, (1995).

54
STENERLÖW, B., BLOMQUIST, E., GRUSELL, E., HARTMAN, T. AND CARLSSON, J.,
Rejoining of DNA double-strand breaks induced by accelerated
nitrogen ions. International Journal of Radiation Biology, 70, 413–20 (1996).
SCHWARTZ, D.C. AND CANTOR, C.R., Separation of yeast chromosome-sized
DNAs by pulsed field gradient gel electrophoresis. Cell, 37, 67–75,
(1984).
THACKER, J., Radiation-induced mutation in mammalian cells at low doses and
low dose rates. Advances in Radiation Biology, 16, 77–124 (1992).
TOBIAS, C.A., ROBERTS, J.E., LAWRENCE, L.H., LOW-BEER, B.V.A., ANGER, H.O.,
BORN, J.L., MCCOMBS, R. AND HUGGINS, C., Irradiation
hypophysectomy and related studies using 340 MeV protons and 190
MeV deutrons. Peaceful Uses of Atomic Energy, 10, 95–106, (1955).
TOBIAS, C.A., BLAKELY, E.A., ALPEN, E.L., CASTRO, J.R., AINSWORTH, E.J., CURTIS,
S.B., NGO, F.Q.H., RODRIGES, A., ROOTS, R.J., TENFORDE, T. AND YANG,
T.C.H., Molecular and cellular radiobiology of heavy ions. International
Journal of Radiation Oncology, Biology and Physics, 8, 2109–20, (1982).
TSUJII, H., MORITA, S., MIYAMOTO, T., MIZOE, JE., MUKAI, M., NAKANO, T., KATO,
T., KAMADA, T., ISHIKAWA, A. AND MATSUOKA, Y., Preliminary results
of hase I/II carbon-ion therapy at the National Institute of Radiological
Sciences. Journal of Brachytherapy, 13, 1–8, (1997).
VAIDYANATHAN, G. AND ZALUTSKY, M.R., Targeted therapy using alpha
emitters. Physics in Medicine and Biology, 41, 1915–31, (1996).
VOGELSTEIN, B., PARDOLL, D.M. AND COFFEY, D.S., Supercoiled loops and
eucaryotic DNA replication. Cell, 22, 79–85 (1980).
WANG, P., ZHOU, R.H., ZOU, Y., JACKSON COOK, C.K. AND POVIRK, L.F., Highly
conservative reciprocal translocations formed by apparent joining of
exchanged DNA double-strand break ends. Proceedings of the National
Academy of Sciences USA, 94, 12018–23 (1997).
WARD, J.F., Some biochemical consequences of the spatial distribution of
ionizing radiation-produced free radicals. Radiation Research, 86, 185–95
(1981).
WARD, J.F., Biochemistry of DNA lesions. Radiation Research, 8, S103–11 (1985).
WARD, J.F., The complexity of DNA damage: relevance to biological
consequences. International Journal of Radiation Biology, 66, 427–32 (1994).
WEBER, K.J. AND FLENTJE, M., Lethality of heavy ion-induced DNA double-
strand breaks in mammalian cells. International Journal of Radiation
Biology, 64, 169–78 (1993).

55
WELT, S., SCOTT, A.M., DIVGI, C.R., KEMENY, N.E., FINN, R.D., DAGHIGHIAN, F.,
GERMAIN, J.S., RICHARDS, E.C., LARSON, S.M. AND OLD, L.J., Phase I/II
study of iodine 125-labeled monoclonal antibody A33 in patients with
advanced colon cancer. Journal of Clinical Oncology, 14, 1787–97 (1996).
WILSON, R.R., Radiological use of fast protons. Radiology, 47, 487–91, (1946).
WU, H., SACHS, R.K. AND YANG, T.C., Radiation-induced total-deletion
mutations in the human hprt gene: a biophysical model based on
random walk interphase chromatin geometry. International Journal of
Radiation Biology, 73, 149–56 (1998).
WULF, H., KRAFT-WEYRATHER, W., MILTENBURGER, H. G., BLAKELY, E. A.,
TOBIAS, C. A. AND KRAFT, G., Heavy-ion effects on mammalian cells:
Inactivation measurements with different cell lines. Radiation Research,
104, S122–S134 (1985).
YANG, T.C., CRAISE, L.M., MEI, M. AND TOBIAS, C.A., Neoplastic cell
transformation by high-LET radiation: Molecular Mechanisms.
Advances in Space Research, 9, 131–40, 1989).
YANG, T.C., GEORGY, K.A., CRAISE, L.M. AND DURANTE, M., Initiation of
oncogenic transformation in human mammary epithelial cells by
charged particles. Radiation Oncology Investigations, 5, 134–8 (1997).
YOKOTA, H., VAN DEN ENGH, G., HEARST, J.E., SACHS, R.K. AND TRASK, B.J.,
Evidence for the organization of chromatin in megabase pair-sized
loops arranged along a random walk path in the human G0/G1
interphase nucleus. Journal of Cell Biology, 130, 1239–49 (1995).
ZIEGLER, J.F. AND MANOYAN, J.M., The stopping of ions in compounds. Nuclear
Instruments and Methods in Physics Research, B35, 215–228 (1988).

56
ACKNOWLEDGEMENTS
“Some folk are wise, and some are otherwise.”
(Tobias George Smollet, 1721–1771)
The studies presented in this thesis were carried out at the Division of
Biomedical Radiation Sciences, Uppsala University, with the main financial
support from the Swedish Cancer Society. A large number of people have
contributed in various ways to this work and I want to express my deepest
gratitude to you all.
I am greatly indebted to my supervisor, the “Rising Star” Ph.D. Bo Stenerlöw for
excellent guiding. I am truly impressed by your knowledge and your working
capacity. Further, I want to thank you for being patient during my frequent
periods of indolence…
My co-supervisors:
Professor Jörgen Carlsson, for providing a positive, open and stimulating
atmosphere at BMS. I also thankfully accepted the very few sets given away on
the badminton course. My ego needed that!
MD, Ass. Professor Erik Blomquist, not only for financial support and useful
comments on manuscripts, but also for fruitful “high-LET” meetings and joyous
acclamations.
Mina vänner & kollegor:
Peter Frisk och Anna Sjöström, mina rumskamrater under mina sista dagar på
BMS. – Tack till er båda för att ni inte knorrat då jag spridit ut mig över hela
vårt rum de senaste 6 månaderna…eller var det därför du flyttade över till
onkologkorridoren Anna….?
….och Peter, jag hoppas att du kan uppehålla den gamla goda och hederliga
“little-NASDAQ” känslan som vi grundade! Hav tröst! MultiQ kommer te’baks!
Semi-seniorerna; Lars Gedda and Pär Olsson.
Lars, för din värdefulla hjälp under min korta historia som Mac-användare,
utan din godhet hade mitt förhållande till Mac-världen varit än mer
komplicerat. Pär, jag hoppas vi snart ses över en sip Isle of Arran — Slainte!

57
Nina Tilly, jag har alltid uppskattat glada middagar och fräscha Gingroggar i
goda vänners lag. Dessutom vill jag tacka för din hjälpsamhet då jag ställt
dumma dosimetrifrågor.
Torbjörn Hartman — BMS store pedagog — vill jag tacka för den fina
illustrationen (Figure 2 i avhandlingen).
To all members of INRA — for adopting me into your mysteries…
Mark Lubberink — aldrig omöjlig när det gäller dator och nätverksinsatser.
Alltid “a nice guy” som dessutom befriade mig från min gamla bil. – Tack och
lycka till…!!
Anna and Vladimir for your friendship and hospitality. I thank you both also for
the frequent delivery of Russian Beluga…
Maria Östh-Eklind, tack för att du återställt ordningen, och för att du alltid varit
hjälpsam.
Veronica Asplund Eriksson, Agneta Johansson och Ulla Johansson, för all hjälp med
praktiska detaljer på labbet.
Tack även till alla er andra som jag mött under min tid på BMS — personal,
seniorer, doktorander och alla andra — en fascinerande skara människor.
Alla berörda på The Svedberg Laboratoriet är också värda ett tack, för alla de joner
ni stressat upp åt mig och sedan levererat vid den biologiska bänken.
Jag vill även tacka mina högt värderade vänner från “andra sidan”, det vill
säga från den riktiga världen utanför den ibland inskränkta forskningsdito;
Staffan “Mäklarn” Dufbäck och hans Anna-Karin, jag uppskattar verkligen allt tok
vi hittat på genom åren. Jag är säker på att nya upptåg väntar runt hörnet…
Mikael Ludvigsson, utan din hjälp hade mitt handicap aldrig nosat på
singel…snart ger jag dig en match – slagspel, scratch – “man to man”.

58
Min familj:
Utan ert starka och kärleksfulla stöd hade detta aldrig lyckats.
Mamma och Pappa, min bror Hans och hans fru Katarina och deras härliga
ungar – Ludvig och Elin.
Stort tack till “Kumla—Eriksbergsgänget” för att ni lånat ut er dotter/syster till
mig, och för att ni så gästvänligt inkluderat mig i er familj.
Slutligen – men absolut viktigast – vill jag tacka min alldeles egna Johanna.
Min älskade fru och min allra bästa vän. För ditt oändliga engagemang och din
aldrig sinande positiva livsyn – och för att du aldrig kallar mig “Skrubbilubben”
när nå’n hör….