Embed Size (px)
Citation preview

Biochemistry 201 Biological Regulatory Mechanisms: Lecture 1
January 5, 2015
DNA POLYMERASE AND IN VIVO ANALYSIS OF REPLICATION TEXTBOOKS/REVIEWS *Stent, G.S. (1978). Molecular Genetics, 2nd edition, W.H. Freeman, San Francisco. Chapters 8 (DNA Structure and Replication), pp. 199-213; 225-250. Kornberg, A. Baker, T.A. (1992). DNA Replication (Second Edition), Freeman, San Francisco. Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts, K. and Walter, P. (2007). Molecular Biology of the Cell (5th editions). Chapter 5 (DNA Replication Mechanisms) pp. 266-281. Garland Press, New York. *Watson, J.D., Baker, T.A., Bell, S.P., Gann, A., Levine, M., Losick, R. (2007). Molecular Biology of the Gene, (7th edition). Chapter 9 (The Replication of DNA). Benjamin Cummings, New York. PRIMARY LITERATURE 1) Watson, J.D. and Crick, F.H.C. (1953). Genetical implications of the structure of deoxyribonucleic
acid. Nature 171, 964-967. The prediction, based on the just-discovered structure of the double helix, that complementary base
pairing underlies the replication process. ***2) Meselson, M. and Stahl, F.W. (1958). The replication of DNA in E. coli. Proc. Natl. Acad. Sci.
USA 44, 671-682. The demonstration--using sedimentation of density-labeled DNA to equilibrium in CsC1 density
gradients--that DNA replication is semi-conservative, as predicted by Watson and Crick. If you thought you knew this paper, check out the discussion questions.
3) Cairns, J. (1963). The bacterial chromosome and its manner of replication as seen by
autoradiography. J. Mol. Biol. 6, 208-213. First direct evidence of a moving locus of DNA synthesis at the replication fork. 4) Josse, J., Kaiser, A.D., and Kornberg, A. (1961). J. Biol. Chem. 236:864.

Nearest-neighbor analysis to show that DNA synthesized by DNA Polymerase I was instructed by the template. Also gave first evidence that two strands are antiparallel. Analysis was rather arcane, but this is what you had to do before DNA sequencing was invented.
*5) De Lucia, P. and Cairns, J. (1969). Isolation of an E. coli strain with a mutation affecting DNA
polymerase. Nature 224, 1164. Demonstration that E. coli mutants that lack the DNA Pol I polymerase activity are still viable. I
recommend this paper because Cairns is remarkable precise about the assumptions in his analysis and what he can and cannot conclude from his data. For example, near the end of his paper he says he cannot conclude that DNA Pol I is unnecessary for DNA replication, and that proves to be prescient, because although it is not the replicative polymerase, it is important for okazaki fragment maturation.
6) Okazaki, R., Okazaki, T., Sakabe, K., Sugimoto, K. and Sugino, A. (1968). Mechanism of DNA
chain growth. I. Possible discontinuity and unusual secondary structure of newly synthesized chains. Proc. Natl. Acad. Sci. USA 59, 598-605.
Discovery of Okazaki fragments by the Okazakis. But does the evidence make sense? 7) Inman, R.B. and Schnos, M. (1971). Structure of branch points in replicating DNA: presence of
single-stranded connections in lambda DNA branch parts. J. Mol. Biol. 56, 319-325. Independent evidence from electron microscopy convinces the doubters that the replication fork is
asymmetric. 8) Reichard, P., Eliasson, R. and Soderman, G. (1974). Initiator RNA in discontinuous polyoma DNA
synthesis. Proc. Natl. Acad. Sci. USA 71, 4901-4905. The first convincing evidence that short RNA oligonucleotides (RNA primers) are linked to the 5'
end of Okazaki fragments. 9) Brutlag, D. and Kornberg, A. (1972). J. Biol. Chem. 247, 241 Demonstration that 3' to 5' exonuclease activity could help DNA polymerase I proofread mistakes at
the 3' end of the primer. 10) Steitz, T.A. (1999). DNA Polymerases: Structural Diversity and Common Mechanisms. J. Biol.
Chem. 274, 17395-98. Minireview of the structure of DNA polymerases. *11) Kunkel, T.A. and Bebenek, K. (2000). DNA Replication Fidelity. Annu. Rev. Biochem. 69:497-
529 Overview of how polymerases maintain replication fidelity.

SOME QUESTIONS TO THINK ABOUT THE LECTURE 1) Formally DNA polymerases could have added nucleotides onto either (A) the 3’ OH end or (B) the 5’-phosphate end of a growing DNA strand. You are Arthur Kornberg, with no structural information about the enzyme. What type of modified nucleotide analog can you feed to the enzyme to distinguish between there two possibilities? How would you monitor the fate of this nucleotide? How would you determine whether any further normal nucleotides could be incorporated? What results would you expect for either possibility A or B? 2) How would you draw the free energy reaction diagram for DNA polymerization to illustrate the importance of pyrophosphate release in driving the reaction forward? What would you add to your polymerase reaction tube if you wanted to reverse the polymerization (assume the exonuclease activity has been destroyed by mutation)? 3) The basic principle of purifying a biochemical activity is straightforward: for each fractionation step account for where all the input activity ended up and where all the input protein ended up. Hopefully you will be able to identify fractions that have increased activity per protein (i.e. specific activity) relative to the input. Such accounting requires accurate quantitative assays for both activity and protein. Specifically you want to make sure that if your assay tube has 2-, 4-, or 10-fold more activity, that your assay readout increases by 2-, 4- or 10-fold, respectively, i.e. you are in the “linear range” of the assay for your activity. For an enzyme like DNA polymerase what should the assay concentration ranges be for its substrates (primer-template and nucleotides) relative to their Km, and why? 4) John Cairns started his search for DNA Pol I mutants with a suspicion that Pol I was not responsible for the bulk of replicative DNA synthesis. Suppose he had suspected the opposite? What type of mutants would he have needed to obtain? How would you show that you had mutated DNA Pol I activity? How would you show that this activity was essential for viability? Is that sufficient to establish that DNA Pol I activity is essential for cellular DNA replication, or do you need to do something further? 5) In the polymerization reaction cycle, nucleotide binding is considered a rapid reversible step relative to later irreversible catalytic steps. Hence, although there is not a true equilibrium between the nucleotide free and nucleotide bound states of the DNA polymerase, we can approximate their relationship by a pseudo dissociation constant. The nucleotide specificity of the DNA polymerase arises in part from the ratio between the dissociation constants for the correct versus the incorrect nucleotides. Why is this primarily determined by the ratios of the off rate constants for the correct versus incorrect nucleotides? 6) A central requirement for kinetic proofreading is that the discard pathway be irreversible. This requires some free energy input into the system, either coupled to the discard pathway itself or stored up from some earlier step. For DNA polymerase proofreading, where is the energy input that makes the exonuclease step irreversible?

7) How does the requirement for DNA polymerase to have primers contribute to its fidelity? How do you explain its contribution in terms of the kinetic parameters that determine fidelity? 8) The idea that the exonuclease activity of replicative DNA polymerases is important for replication fidelity was inferred from kinetic analysis of purified DNA polymerases? What experimental perturbations and readouts would be needed to show that this is indeed the case in cells? 9) Autoradiographic analysis of intact replicating E. coli DNA revealed large circular DNA with single bubbles of various sizes as shown in the lecture handout. Based on that type of data, what can you conclude about the following: (1) number of initiation sites per replication event, (2) position of initiation sites, (3) number of moving replication forks per replication event. What additional information would allow you to derive more conclusions about these issues? 10) Okazaki concluded that newly synthesized DNA that was small became converted into much larger DNA, i.e. that small DNA fragments (later named Okazaki fragments) were an intermediate in the DNA replication pathway. Why is this conclusion not really justified by his experimental design? How might you improve on his design? 11) Okazaki’s experiment is usually cited as one of the key evidence for semidiscontinuous DNA replication. But Okazaki himself concluded that DNA synthesis is fully discontinuous. Why did he conclude this from his results? Can you speculate why he got this result? POINTS RAISED IN LECTURE 1) Genetics and biochemistry are both powerful entry points into the study of a biological problem. In the case of DNA replication, biochemistry dominated the early days. (In other cases--e.g. the development of the operon model for transcriptional regulation—genetics led the way.) As the replication field matured, the two approaches were used together to great advantage. For example, genetics indicated that DNA polymerase I was unlikely to be the primary replicative polymerase, and provided the mutants that allowed biochemists (specifically, our own Tom Kornberg) to uncover and purify DNA polymerase III, the true replicative polymerase. Biochemistry allows you to delineate the exact mechanism by which a biological tasks is performed. Genetics allows you to establish the functional relevance of a mechanism to the biological task of interest.
2) Function is determined by phenotypes and assays. The basic currency of genetics is the mutant phenotype; it allows one to define, observe, and follow mutations. The inability of a mutant to perform a certain task suggests that the wild-type version of the mutant protein normally carries out this task. The linchpin of biochemistry is the assay; it allows one to define, observe, and follow biochemical activities. Understanding the exact nature of the assay is important, since slight changes in the assay may change the nature of the activity being followed, and result in purification of different proteins or complexes of proteins. In well-studied systems this somewhat artificial distinction between genetic phenotypes and biochemical assays becomes blurred. For example, mutant cells can be phenotypically characterized by biochemical assays of extracts made from the cells. Another example is the precise mutagenesis of a specific activity of a protein to determine the mechanistic role of this activity in the protein’s function.

3) Divide and Conquer. The reductionist approach at the core of our scientific process is to break things down into their component parts, figure out how the parts work individually, then determine how the parts work together. This approach is difficult to pursue biochemically when fundamental functions are carried out by complex assemblies of subunits and are not readily assigned to separable individual subunits. Replication has been particularly amenable to this approach because specific functions can be attributed to individual proteins, even though those functions are often greatly enhanced in the presence of the correct neighbors or setting. In contrast, the role of individual protein or RNA molecules in the ribosome is currently best studied in the context of the full ribosome, using mutations that perturb specific proteins or RNAs. 4) Structure informs but is not sufficient to establish function. Based on the Watson-Crick structural model of DNA, it was predicted that DNA would replicate semi-conservatively. But actual proof required the Meselson-Stahl experiment. Also, the W-C model did not anticipate the idea of a replication fork, nor did it predict any features of the replication machinery. Later, EM studies on replication intermediates provided further structural insight leading to the idea of replication forks. However, it was Kornberg’s unswerving faith (yes, faith has a role in science) that proteins must carry out virtually all chemical reactions in the cell that paved the way for biochemical identification of replication proteins and eventual understanding of how those forks function. 5) Crude extract assays may be quite complex. Original DNA polymerization assay was not just assaying the polymerization reaction, but also the conversion of DNA to nucleosides, the conversion of nucleosides to nucleotides (i.e. phosphorylation by kinases), and the conversion of DNA to primer-template junctions. Fractionation of extract led to separation of these multiple activities and eventual discovery of the true substrate requirements for the polymerization reaction itself. Thus even though the initial assay had the wrong substrates and required multiple activities, having some detectable signal allowed Kornberg to tease apart the complexity and eventually focus on the activity of interest. 6) "Purity is in the eye of the beholder"-Nick Cozzarelli. There is no absolute standard for knowing that your activity is "pure". The 5% impurity in your 95% pure protein could be responsible for the activity you are following or could be associated with an additional activity you don’t yet realize you are assaying. The classic approach is to show that a protein(s) repeatedly and tightly cofractionates with your activity over many separation steps, giving you increasing (though never absolute) confidence that the protein is responsible for the activity. Molecular biology has provided several powerful new ways to help associate an activity with a gene product(s), but does not provide a better method for assessing purity. An example where sophisticated molecular biology could not rescue a bad conclusion based on poor purification is provided by a high profile Science paper stating that yeast Trf4, which shows some sequence homology to DNA polymerases, has very weak DNA polymerase activity. Affinity purified Trf4 from E. coli had the activity, whereas affinity purified Trf4 mutated in conserved polymerase residues did not. Seems like a slam dunk, but couldn’t be reproduce and was later refuted. Presumably, the prep of affinity purified wild-type Trf4 was contaminated with a tiny amount of DNA polymerase, whereas the mutant prep was not. Hence, the classic approach to demonstrating purification is still highly relevant in this era of high-powered molecular biology.

7) Holoenzymes allow a core activity to be used in various settings and processes. Although some complex assemblies such as ribosomes are held tightly together, others--as exemplified by the proteins at the T4 replication fork--only loosely associate. Often many different degrees of association exist among proteins involved in the same process. For example, the catalytic core of DNA Pol III contains three tightly associated subunits. The gamma complex (clamp loader), which itself contains five tightly associated subunits, is bound less tightly to the catalytic core and often disassociates during purification. The concept of a "holoenzyme" was initially introduced to incorporate the idea that large complexes containing loosely associated subunits that modify or regulate the core catalytic activity can be reproducibly purified (although usually under "gentle" conditions). These holoenzymes are thought to work as functional units in vivo. Thus, the complex containing two catalytic Pol III cores tethered to each other to one gamma complex via a tau protein dimer is called the Pol III holoenzyme and is thought to be the form of polymerase that works at the replication fork. Given the continuous range of association strengths found among proteins, the definition of a holoenzyme is somewhat arbitrary and now includes some complexes that cannot be purified together but are still thought to function as a unit (compare the concept of holoenzyme for eukaryotic RNA polymerases to that of DNA polymerases). Importantly, the loose association of components in holoenzymes provide a modular way for these components to assemble into more than one type of holoenzyme so as to perform different functions. For example, only a small minority of the Pol III core in a cell is part of the Pol III holoenzyme and participates in DNA replication. The bulk of the Pol III core is likely associated with other proteins in holoenzmes involved in DNA repair. 8) Limitations of loss of function (LOF) genetic analyses. Loss of function genetic analysis is a powerful way to determine the function of genes and proteins. It uses the general scientific strategy of asking whether “A is required for B” by disrupting A and determining whether B is affected. If a wild-type function is impaired when a gene is disrupted, one can usually conclude that the gene is required in some manner for that function. However, things are not as straightforward when a function is not measureably affected, as there are several possible reasons for “false negative” results. For example, initial genetic analysis using the polA1 mutation may have missed a replicative role for DNA Pol I in okazaki fragment maturation because of three possible limitations of the analysis: (1) incomplete disruption of Pol I activity (2) weak redundant function provided by other polymerases (3) indirect and insensitive readout of cell viability instead of direct analysis of okazaki fragment maturation. 9) Replicative DNA polymerases are designed for extremely high fidelity. All replicative polymerases polymerize 5’ to 3’, are instructed by the template, require a primer, and use deoxynucleotide triphosphates as precursors. Almost all also possess a 3' to 5' exonuclease activity, which allow them to proofread their own mistakes. The requirement for a properly based paired primer in effect allows the polymerase to monitor whether an incorporation mistake has recently been made. The 5’ to 3’ polymerization direction ensures that the high energy bond for each polymerization step is contributed by the incoming monomer and not by the polymer (“tail” extension; contrast to “head” extension during translation). Hence, exo “deletion” of any polymerization mistake does not compromise the ability to continue polymerization (contrast this to protein translation). 10) Fundamentals of fidelity. Fidelity requires first the ability to distinguish between correct and incorrect reaction settings. Secondly, fidelity requires having a molecular choice to either advance a reaction (forward reaction) or abort the reaction (discard reaction). Third, fidelity requires that ability to shift the choice of reaction based on the setting that is sensed. Correct settings have to be preferentially

shunted to the forward reaction, and incorrect setting have to be preferentially shunted to the discard reaction. For replicative DNA polymerases the important distinction in reaction setting is between having a correct or incorrect nucleotide opposite a specific template nucleotide. At the core of the ability to make this distinction is the polymerase’s induced fit requirement for catalysis. For example, only proper base pairing of the incoming nucleotide allows the conformational change that “fits” the polymerase snugly around the nucleotide-primer-template. This induced fit also couples the base pairing of the incoming nucleotide identity to the choice of reaction, because the fit allowed by the correct base pairing positions catalytic groups properly to promote polymerization. In contrast, an if the incoming nucleotide is incorrectly base-paired, the induced fit will be much harder, catalysis will be much slower, and the competing discard reaction of the nucleotide leaving the active site will be favored. Fidelity is not always a desirable thing, since highly faithful polymerases will stall if they face damaged or unusual nucleotides in the template strand. In this setting no incoming nucleotide will be “correctly” base-paired with the template, as defined by the ability to satisfy a proper induced fit. In the past ten years a large class of nonreplicative polymerases with low fidelity have been identified. Many of them allow the cell to contend with damaged template nucleotides or promote mutagenesis when genetic diversity might be needed. These enzymes do not have a tight induced fit and/or have little or no 3’ to 5’ exonuclease activity. 11) Kinetic manipulation of molecular choice: The fact that the induced fit also requires proper base pairing of the primer nucleotides provides a second opportunity to test base-pairing, this time of the recently incorporated nucleotide. However, there is no intrinsic discard pathway, such as the diffusion of a free nucleotide away from the active site. So for replicative DNA polymerases which must maintain high fidelity, nature has designed a special discard pathway that aborts the extension from an incorrectly base-paired primer without aborting the entire polymerization process. This discard pathway involves the excision of the incorrectly base-paired primer nucleotide with a 3’>5’ exonuclease activity. In principle, both correctly and incorrectly incorporated nucleotides can be removed by this discard pathway or be extended further in a polymerization reaction. The key is to change the relative kinetics of these two reactions depending on whether the incorporated nucleotide is correct or incorrect. This relative kinetics determines whether the substrate preferentially partitions to the forward pathway of polymerization (correctly incorporated nucleotide) or the discard pathway of exonucleolytic cleavage (incorrectly incorporated nucleotide). Hence the term kinetic partitioning. The separate and distant exonuclease catalytic site places the exonuclease reaction at an inherent disadvantage when the correct nucleotide is incorporated. Moreover, the proper primer-template base-pairing resulting from this incorporation allows the induced fit to facilitate rapid catalysis of the forward polymerization reaction. Thus, almost all the molecules in this setting are partitioned toward polymerization. However, when an incorrect nucleotide is incorporated, the disrupted base pairing of the primer-template inhibits the induced fit of the polymerase and greatly slows the polymerization reaction. The mispairing at the primer-template junction also slightly increases the rate of exonuclease reaction. The net result is that the kinetic competition is shifted in favor of the discard pathway when an incorrect nucleotide is incorporated. 12) Structural analysis of in vivo replication intermediates established that: (1) replication was localized to moving replication forks; (2) synthesis at forks occurs in a semidiscontinuous manner; (3) discontinuous synthesis involves synthesis of short 100-2000 bp okazaki fragments that are ligated to

form the daughter strands on one side of the fork. These observations suggest the presence of a number of activities needed at a replication fork in addition to the replicative DNA polymerase.

Lect
ure
1:
Fide
lity/
Spec
ifici
ty: b
iore
gula
tion
thro
ugh
subs
trat
e co
ntro
l of m
olec
ular
cho
ice
Use
of b
ioch
emis
try
(ass
ays)
and
gen
etic
s (m
utan
t phe
noty
pes)
to d
efin
e fu
nctio
n
DN
A Po
lym
eras
e an
d th
e R
eplic
atio
n Fo
rk
Bre
akin
g do
wn
com
plex
pro
cess
es b
y st
ruct
ural
ana
lysi
s of
inte
rmed
iate
s

Usin
g bo
th B
ioch
emist
ry a
nd G
enet
ics t
o un
ders
tand
func
tion
reve
rse
gene
tics!
(targ
eted
gen
e di
srup
tion
by!
HR
, ZFN
, TAL
EN, C
RIS
PR/C
as9)!
expr
essi
on in
terfe
renc
e!(R
NAi
, siR
NA,
CR
ISPR
i)!

DN
A Re
plic
atio
n: T
he T
ask
and
Chal
leng
e
Spee
d: v
ery
rapi
d du
plic
atio
n of
eve
ry n
ucle
otid
e
Fide
lity:
ext
rem
ely
low
erro
r rat
e
(ex:
6 x
109 b
p in
8 h
rs in
hum
ans)
(~1/
109 n
ucle
otid
e er
ror r
ate)
Coun
t: ex
actly
two
copi
es o
f eve
ry se
quen
ce p
er c
ell c
ycle
Regu
latio
n: c
oord
inat
ion
with
oth
er c
hrom
osom
al e
vent
s (e
g.m
itosis
, rep
air,
reco
mbi
natio
n, tr
ansc
riptio
n, c
hrom
atin
pac
kagi
ng)
Sem
icon
serv
ativ
e D
uplic
atio
n

Enzy
mol
ogy
of D
NA
Synt
hesis
: DN
A Po
lym
eras
es
dNTP
pre
curs
or
Instr
ucte
d by
sing
le-s
trand
ed te
mpl
ate
Prim
er re
quire
men
t*
5’ >
3’ p
olym
eriz
atio
n of
f prim
er*
* en
hanc
es fi
delit
y by
allo
wing
err
or c
orre
ctio
n
- pyr
opho
spha
te re
leas
e pr
ovid
es e
nerg
y
- sen
ses c
ompl
emen
tarit
y of
new
nuc
leot
ide
- ext
ensio
n of
f 3’ h
ydro
xyl
- mov
ing
3’>
5’ o
n te
mpl
ate
- sen
ses c
ompl
emen
tarit
y of
prim
er

In p
rinci
ple:
Mon
itor i
ncor
pora
tion
of ra
dioa
ctiv
e nu
cleo
tide
prec
urso
rs (
)
in
to a
cid
inso
lubl
e fo
rm (p
hysi
cally
sep
arat
e pr
oduc
t fro
m p
recu
rsor
)
Ass
ayin
g D
NA
Poly
mer
ase A
ctiv
ity
In p
ract
ice:
Can
be
diffi
cult
to d
evis
e th
e rig
ht a
ssay
con
ditio
ns w
hen
you
do
not k
now
the
prec
ise
natu
re o
f the
act
ivity
E. c
oli e
xtra
ct -
sour
ce o
f pol
ymer
ase
activ
ity b
ut a
lso
kina
se a
nd n
ucle
ase
activ
ity
3 H T
hym
idin
e -
conv
erte
d to
thym
idin
e tr
ipho
spha
te b
y ki
nase
s in
ext
ract
D
NA
-
inte
nded
as
nucl
ease
dec
oy b
ut n
ucle
ases
con
vert
to p
rimer
-tem
plat
e
and
sou
rce
of A
,G,C
nuc
leot
ides
Initi
al c
ondi
tions
use
d w
ere
real
ly a
ssay
ing
a co
mpl
ex m
ixtu
re o
f act
iviti
es:
Firs
t Exp
erim
ent:
50 o
ut o
f 1 m
illio
n cp
m in
solu
ble
Ten
Year
s La
ter:
pur
ify D
NA
Poly
mer
ase
I, sh
ow it
is te
mpl
ate-
dire
cted
and
figu
re o
ut e
nzym
e re
quire
men
ts

DN
A Po
lym
eras
e St
ruct
ure
and
Cata
lysis
Stru
ctur
e re
sem
bles
a r
ight
han
d
Two
Mg++
ions
pos
ition
ed b
y co
nser
ved
acid
ic re
sidue
s cat
alyz
e re
actio
n
Cry
stal
stru
ctur
e of
bac
teri
opha
ge T
7 D
NA
Poly
mer
ase
com
plex
ed w
ith p
rim
er-te
mpl
ate
and
dNTP
Res
t of e
nzym
e po
sitio
ns p
rim
er-te
mpl
ate
and
dNTP
and
ensu
res c
atal
ysis
only
occ
urs w
ith p
rope
r “fit
” Prim
er
Tem
plat
e

DN
A Po
l I h
as 3
’ >
5’
Exo
nucl
ease
Act
ivity
Exo
Ass
ay:
T AA
AA
AA
AA
T T
T T A
AA
AA
AA
A T
T T
DN
A Po
l I
no d
TTP
5’
3’
5’
3’
5’
3’
5’
3’
exo
activ
ity is
slo
w re
lativ
e to
pol
act
ivity
ex
o ac
tivity
is e
nhan
ced
by s
talli
ng p
ol a
ctiv
ity
o
r mak
ing
3’ e
nd s
ingl
e-st
rand
ed
3’ m
ism
atch
gen
erat
es b
oth
cond
ition
s
* *
* Pr
oofr
ead
Ass
ay:
T AA
AA
AA
AA
T T
T T A
AA
AA
AA
A T
T D
NA
Pol I
+
dTTP
5’
3’
5’
3’
5’
3’
5’
3’
*
T T T
T T
T AA
AA
AA
AA
T T C
T A
AA
AA
AA
A T
T D
NA
Pol I
+
dTTP
5’
3’
5’
3’
5’
3’
5’
3’
*
T T T
T
T
C *
mis
mat
ch s
peci
fic e
xo a
ctiv
ity u
nder
nor
mal
pol
con
ditio
ns
both
pol
and
exo
act
iviti
es a
re s
ensi
ng p
rimer
-tem
plat
e pa
iring
Care
ful q
uant
itativ
e an
alys
is of
bio
chem
ical
act
ivity
can
sugg
est b
iolo
gica
l fun
ctio
n

The
Poly
mer
ase
and
Exon
ucle
ase A
ctiv
ities
of
Repl
icat
ive
DN
A Po
lym
eras
es R
esid
e in
Dist
inct
Dom
ains
Poly
mer
ase
Act
ive
Site
Exon
ucle
ase
Act
ive
Site
~ 30
Å
2- M
ode
Mod
el fo
r Pol
ymer
ase
Func
tion
Poly
mer
izin
g Ed
iting
Mov
emen
t bet
wee
n P
and
E sit
es re
quire
s p
rimer
-tem
plat
e un
win
ding
tran
sloca
tion
of 3
’ e
nd

mut
agen
ize
plat
e
E. c
oli
mut
ant E
. col
i
extr
acts
from
sin
gle
mut
ant c
olon
ies
phen
otyp
e:
assa
y dN
TP
inco
rpor
atio
n in
to D
NA
mut
ant 3
473
(pol
A1)
ha
s <1
% w
t act
ivity
DN
A Po
l I is
not
the
repl
icat
ive
DN
A po
lym
eras
e in
E. c
oli
Use
bio
chem
ical
ass
ay to
scr
een
for m
utan
ts la
ckin
g D
NA
poly
mer
ase
activ
ity
Illus
trat
es im
port
ance
of g
enet
ics
for e
stab
lishi
ng fu
nctio
nal r
elev
ance
in c
ell
norm
al g
row
th
repa
ir de
ficie
nt
Purif
icat
ion
of re
sidu
al p
olym
eras
e ac
tivity
from
pol
A1
yiel
ds D
NA
Pol I
I and
Pol
III
Gen
etic
s an
d bi
oche
mis
try
late
r sho
w:
- D
NA
Pol I
II is
the
repl
icat
ive
poly
mer
ase
- D
NA
Pol I
is im
port
ant f
or o
kaza
ki fr
agm
ent m
atur
atio
n

Purifi
catio
n of
DN
A Po
l III:
Diff
eren
t Tem
plat
e, D
iffer
ent A
ssay
, Diff
eren
t Act
ivity
In
trodu
cing
the
conc
ept o
f hol
oenz
ymes
and
mod
ular
enz
yme
suba
ssem
blie
s

Fide
lity
Ove
rvie
w
Intr
insic
Fid
elity
(pol
ym)
Exon
ucle
ase
Proo
frea
ding
(pol
ym)
Mism
atch
Rep
air (
post
pol
ym)
10-3
- 10
-4
10-2
- 10
-3
(sen
sing
dNTP
com
plem
enta
rity
to te
mpl
ate)
(sen
sing
prim
er c
ompl
emen
tarit
y to
tem
plat
e)
(sen
sing
com
plem
enta
rity
of tw
o str
ands
)
(dist
ingu
ishin
g pa
rent
al a
nd d
augh
ter s
trand
s)
10-2
Ove
rall
Rep
licat
ion
Fide
lity
10-8
- 10
-9
Erro
r rat
e
Con
trib
utio
ns to
E c
oli
DN
A R
eplic
atio
n Fi
delit
y Fi
delit
y C
ompa
riso
ns
DN
A
Rep
licat
ion
RN
A
Tran
scri
ptio
n
Prot
ein
Tran
slatio
n
Spee
d Er
ror
Rat
e Pr
oduc
t Si
ze
10-9
- 10
-10
5 x
106
6 x
109
1 x
1011
(E. c
oli)
(hum
ans)
(lily
)
500
bp/se
c
50 b
p/se
c
(Pro
kary
otes
)
(Euk
aryo
tes)
30 b
p/se
c
20 a
a/se
c
10-4
10-4
103 -
106
102 -
103

How
to D
istin
guish
Mism
atch
ver
sus C
orre
ct B
ase
Pair
Geo
met
ry F
rom
Cry
stal
Str
uctu
re
Glo
bal s
truct
ure
of h
elix
is n
ot g
reat
ly p
ertu
rbed
But t
here
are
:
di
ffere
nces
in C
1’ -
C1’ d
istan
ce a
nd C
1’ b
ond
angl
es
prot
rusio
ns o
f bas
es in
to m
ajor
gro
ove
loss
of u
nive
rsal
H a
ccep
tor p
ositi
ons i
n m
inor
gro
ove
WC
bp
WC
bp
mis
mat
ch
mis
mat
ch
mis
mat
ch
Sour
ces o
f Pol
ymer
ase
Disc
rim
inat
ion
Ster
ic C
onst
rain
ts (s
truc
ture
/geo
met
ry)
H-b
ondi
ng (b
indi
ng e
nerg
etic
s)
O
utsid
e th
e ac
tive
site,
unp
aire
d nu
cleo
tides
ar
e H
-bon
ded
to H
2O.
Insid
e th
e ac
tive
site
thes
e H
-bon
ds c
an b
e re
plac
ed b
y W
C ba
se
pairi
ng b
ut o
nly
inco
mpl
etel
y re
plac
ed b
y m
ismat
ch p
airin
g
Mism
atch
H b
ondi
ng c
an a
lso e
xace
rbat
e ste
ric a
nd st
acki
ng c
lash
es (s
ee b
elow
)
Im
pose
d by
enz
yme’s
“in
duce
d fit
”,
whi
ch c
an te
st fo
r pre
cise
bas
e pa
ir ge
omet
ry, p
rope
r bas
e sta
ckin
g, a
nd c
orre
ct pr
imer
-tem
plat
e fit
.

Intr
insic
Fid
elity
: Pot
entia
l Bas
e Pa
ir D
iscrim
inat
ion
for d
NTP
at
Thre
e St
ages
Of t
he D
NA
Poly
mer
izat
ion
Reac
tion
Cycl
e
E!D
NA! N!
E!D
NA! N!
dNTP!C!
E!D
NA! N!dNTP!I!
E!D
NA! N!
dNTP!C!
*
E!D
NA! N!dNTP!I!
*
E!D
NA! N+
1!C!
PP! i
E!DNA! N+1!
I!PP! i
Rea
ctio
n pa
thw
ay fo
r cor
rect
nuc
leot
ide!
Rea
ctio
n pa
thw
ay fo
r inc
orre
ct n
ucle
otid
e!
K D!C!
k conf
C!
k pol
C!
k conf
I!k pol
I!
Rap
id d
NTP
Bin
ding
Pseu
do-e
quili
briu
m
Slow
Con
form
atio
nal C
hang
e “In
duce
d Fi
t”
1!2!
3!
Poly
mer
izat
ion
Rea
ctio
n
K D!I!
K D!I!
K D!C!
20 µ
M!
> 8m
M!
~!~!
400x!
k conf
C!
k conf
I!~!
~!R
apid
and
Not
Mea
sure
d!0.
3 s!
300
s!-1! -1!
1000
x!
Exam
ple:
T7
DN
A po
lym
eras
e (a
rrow
thic
knes
s rou
ghly
cor
resp
onds
to r
ate
cons
tant
s) O
ther
pol
ymer
ases
disc
rim
inat
e di
ffere
ntly
at e
ach
stag
e

Erro
r Cor
rect
ion:
Exo
nucl
ease
act
ivity
allo
ws t
he p
olym
eras
e’s
kine
tic d
iscrim
inat
ion
to le
ad to
diff
eren
t prim
er fa
tes
E!D
NA! N!
dNTP!C!
E!D
NA! N!
dNTP!C!
* E!
DN
A! N+
2!C!
PP! i
E!DNA! N+1!
I!
Fast
reac
tion
path
way
s fo
r cor
rect
prim
er w
ith c
orre
ct n
ucle
otid
e!
Slow
reac
tion
path
way
s fo
r inc
orre
ct p
rimer
with
cor
rect
nuc
leot
ide!
K D!C!
k pol
C!
k conf
C!
I!
1!2!
3!
E!D
NA! N+
1!C!
dNTP!C!
E!DNA! N+1!
I!E!*
dNTP!C!
DNA! N+1!
I!E!DNA! N+2!
I!PP! i
k conf
C!
pol
k C!
I!
K D!C!
I!
E!D
NA! N!C!
E!D
NA! N!C!
k exo
I!k exo
Arr
ow th
ickn
ess r
ough
ly c
orre
spon
ds to
rat
e co
nsta
nt
E!D
NA! N!
dNTP!C!
E!D
NA! N!
dNTP!C!
* E!
DN
A! N+
2!C!
PP! i
K D!C!
k pol
C!
E!D
NA! N+
1!C!
k conf
C!

Erro
r Cor
rect
ion:
Kin
etic
man
ipul
atio
n of
mol
ecul
ar c
hoic
e ba
sed
on c
ompl
emen
tarit
y of
prim
er
E!D
NA! N+
2!C!
PP! i
E!DNA! N+1!
I!
E!D
NA! N+
1!C!
E!DNA! N+2!
I!PP! i
E!D
NA! N!C!
E!D
NA! N!C!
pol
exo
pol
exo
Whe
n a
corr
ect n
ucle
otid
e is
inco
rpor
ated
, 3’>5
’ e
xonu
clea
se a
ctiv
ity is
muc
h slo
wer
tha
n 5’
>3’ p
olym
eras
e ac
tivity
. A
dditi
on o
f the
nex
t nuc
leot
ide
is ki
netic
ally
favo
red.
Whe
n an
inco
rrec
t nuc
leot
ide
is in
corp
orat
ed, d
isrup
tion
of th
e pr
imer
gre
atly
slo
ws 5
’>3
’ p
olym
eras
e ac
tivity
for t
he n
ext n
ucle
otid
e (a
nd sl
ight
ly in
crea
ses 3
’>5
’
exo
nucl
ease
act
ivity
). E
xcisi
on o
f the
inco
rrect
nuc
leot
ide
is ki
netic
ally
favo
red.
Arr
ow th
ickn
ess r
ough
ly c
orre
spon
ds to
rat
e co
nsta
nt

Erro
r Cor
rect
ion:
Kin
etic
man
ipul
atio
n of
mol
ecul
ar c
hoic
e ba
sed
on c
ompl
emen
tarit
y of
prim
er
pol
exo
pol
exo
5’-TAGCTTCG
3’-ATCGAAGCTCATG!
5’-TAGCTTCGA
3’-ATCGAAGCTCATG!
5’-TAGCTTC
3’-ATCGAAGCTCATG!
5’-TAGCTTC A
3’-ATCGAAGCTCATG!
A!
5’-TAGCTTC
3’-ATCGAAGCTCATG!
A!
5’-TAGCTTC
3’-ATCGAAGCTCATG!
Blac
k ar
row
thic
knes
s rou
ghly
cor
resp
onds
to re
lativ
e ra
te c
onst
ant
Ligh
t blu
e ar
row
thic
knes
s rou
ghly
cor
resp
onds
to re
lativ
e flu
x
k T7pol
C p
rimer!
300
s!-1!
k T7exo
C p
rimer!
0.2
s!-1!
k T7pol
I prim
er!
0.01
s!-1!
k T7exo
I prim
er!
2.3
s!-1!
1500!
k T7pol
C p
rimer!
k T7exo
C p
rimer!
k T7pol
I prim
er!
k T7exo
I prim
er!
230!
1!
~! ~!
Exam
ple:!
T7 D
NA
Poly
mer
ase!

A D
iscar
d St
rate
gy fo
r Fid
elity
: Kin
etic
man
ipul
atio
n of
mol
ecul
ar
choi
ce b
etw
een
irrev
ersib
le fo
rwar
d an
d di
scar
d pa
thw
ays
forw
ard
disc
ard
forw
ard
disc
ard
Cog
nate
Sub
stra
te
Cor
rect
Pro
duct
Non
cogn
ate
Subs
trat
e In
corr
ect P
rodu
ct
Elim
inat
ion
Elim
inat
ion
The
choi
ce is
ulti
mat
ely
dete
rmin
ed b
y th
e re
lativ
e flu
x of
mol
ecul
es
that
pro
ceed
dow
n th
e tw
o co
mpe
ting
path
way
s (li
ght b
lue
arro
w)
In p
rinci
pal,
just
one
or b
oth
path
way
s co
uld
disc
rimin
ate
betw
een
cogn
ate
and
nonc
ogna
te s
ubst
rate
s, i.
e. c
hang
e ra
te c
onst
ants
with
su
bstra
te.
In p
ract
ice,
nat
ure
ofte
n di
scrim
inat
es w
ith b
oth.
For e
ach
subs
trate
, the
mol
ecul
ar fl
ux (a
nd h
ence
mol
ecul
ar c
hoic
e) is
de
term
ined
by
the
ratio
of t
he fo
rwar
d to
dis
card
rate
con
stan
ts (b
lack
ar
row
s) fo
r tha
t sub
stra
te.
For c
ogna
te s
ubst
rate
s th
is ra
tio s
houl
d fa
vor t
he fo
rwar
d re
actio
n. F
or n
onco
gnat
e su
bstra
tes,
the
ratio
sho
uld
“fli
p” to
favo
r the
dis
card
pat
hway
.
Pat
hway
irre
vers
ibili
ty u
sual
ly re
quire
s so
me
chem
ical
ene
rgy
expe
nditu
re (e
.g d
NTP
hyd
roly
sis)
, whi
ch c
ould
be
coup
led
to e
ither
pa
thw
ay o
r to
a re
actio
n st
ep p
rece
ding
thes
e pa
thw
ays

5’-TTCGA
3’-AAGCTCA!
3’-AAGCTCA!
5’-TTC A
A!
5’-TTCG
3’-AAGCTCA!
3’-AAGCTCA!
5’-TTC A!
5’-TTC
3’-AAGCTCA!
G!
3’-AAGCTCA!
5’-TTC
A!
5’-TTCG
3’-AAGCTCA!
A!
3’-AAGCTCA!
5’-TTC A!A!
5’-TTC
3’-AAGCTCA!
k pol
C p
rimer!
k pol
I prim
er!
k pol
C d
NTP!
k pol
I dN
TP!k exo
C p
rimer!
k exo
I prim
er!
K D!C
dN
TP!
K D!I d
NTP!
K D!C
prim
er!
K D!I p
rimer!
K D!
pseu
doeq
uilib
rium
con
stan
t fo
r nuc
leot
ide
bind
ing
k pol
com
posi
te ra
te c
onst
ant f
or
poly
mer
izat
ion
reac
tion
com
posi
te ra
te c
onst
ant f
or
exon
ucle
ase
reac
tion
k exo
prim
er-te
mpl
ate
5’-TTC
3’-AAGCTCA!
DN
A po
lym
eras
e
5’-TTC
3’-AAGCTCA!
G!
prim
er-te
mpl
ate
DN
A po
lym
eras
e
boun
d dN
TP
Intr
insic
Fid
elity
+ E
rror
Cor
rect
ion

How
DN
A Po
lym
eras
es C
heck
for P
rope
r Bas
e Pa
iring
Geo
met
ry
Cry
stal
stru
ctur
e su
gges
tion
for “
Indu
ced
Fit”
Poly
mer
ase
+ Pr
imer
-Tem
plat
e Po
lym
eras
e +
Prim
er-T
empl
ate
+ dN
TP
DN
A Po
lym
eras
e co
ntac
ts m
inor
gro
ove
of p
rim
er-te
mpl
ate
Tem
plat
e
Prim
er
Purp
le: I
nter
actio
n su
rface
with
DN
A po
lym
eras
e G
reen
: Uni
vers
al H
-bon
d ac
cept
ors
H
-bon
ding
with
DN
A po
lym
eras
e
Base
pai
r fit i
s stil
l “te
sted
” a
fter p
olym
eriz
atio
n Ba
se p
air fi
t is “
test
ed” b
efor
e po
lym
eriz
atio
n
Stac
king
Inte
ract
ion
Onl
y W
-C b
ase
pairs
allo
w p
rope
r sta
ckin
g In
duce
d fit
pos
ition
s nuc
leot
ide,
prim
er 3
’, m
etal
ions

Mos
t DN
A po
lym
eras
es in
the
cell
have
NO
N-re
plic
ativ
e ro
les
Pol I
Po
l II (
Din
A)
Pol I
II ho
loen
zym
e Po
l IV
(Din
B)
Pol V
(Um
uC, U
muD
’2C
)
Prok
aryo
tic D
NA
Poly
mer
ases
(E. c
oli)
DN
A R
eplic
atio
n (R
NA
prim
er re
mov
al);
DN
A re
pair
DN
A re
pair
DN
A R
eplic
atio
n D
NA
repa
ir; T
LS; a
dapt
ive
mut
agen
esis
TL
S (tr
ansl
esio
n sy
nthe
sis)
Euka
ryot
ic D
NA
Poly
mer
ases
Pol α
Po
l β
Pol γ
Po
l δ
Pol ε
Po
l θ
Pol ζ
Pol λ
Po
l µ
Pol κ
Po
l η
Pol ι
R
ev1
DN
A R
eplic
atio
n (P
rimer
Syn
thes
is)
Bas
e ex
cisi
on re
pair
Mito
chon
dria
l DN
A re
plic
atio
n/re
pair
DN
A R
eplic
atio
n; n
ucle
otid
e an
d ba
se e
xcis
ion
repa
ir D
NA
Rep
licat
ion;
nuc
leot
ide
and
base
exc
isio
n re
pair
DN
A cr
ossl
ink
repa
ir TL
S M
eios
is-a
ssoc
iate
d D
NA
repa
ir So
mat
ic h
yper
mut
atio
n TL
S Er
ror-
free
TLS
pas
t cyc
lobu
tane
dim
ers
TLS,
som
atic
hyp
erm
utat
ion
TLS
Man
y of
the
nonr
eplic
ativ
e po
lym
eras
es h
ave
low
fide
lity
and
are
erro
r-pr
one
bec
ause
they
tole
rate
non
-WC
bp
and
lack
3’>5
’ e
xo a
ctiv
ity
Low
fide
lity
is n
eede
d to
byp
ass
tem
plat
e le
sion
s th
at a
re s
talli
ng re
plic
atio
n Lo
w fi
delit
y m
ay b
e us
ed to
incr
ease
gen
etic
var
iatio
n in
spe
cial
circ
umst
ance
s

Sem
i-Disc
ontin
uous
DN
A Sy
nthe
sis a
t Rep
licat
ion
Fork
s Le
adin
g da
ught
er st
rand
: pol
ymer
ase
mov
es c
ontin
uous
ly in
sam
e di
rect
ion
as re
plic
atio
n fo
rk
Lagg
ing
daug
hter
stra
nd: p
olym
eras
e m
oves
disc
ontin
uous
ly in
opp
osite
dire
ctio
n as
repl
icat
ion
fork
5’
5’
3’
3’
5’
3’ le
adin
g
lagg
ing
A
B C
Fork
Mov
emen
t
Synt
hesis
with
disc
ontin
uous
lagg
ing
stran
d pi
eces
(oka
zaki
frag
men
ts) r
equi
res r
epea
ted:
A. p
rimin
g B.
repl
acem
ent o
f prim
ed se
quen
ce
C. li
gatio
n
Oka
zaki
frag
men
t len
gth:
pro
kary
otes
100
0 - 2
000
nt; e
ukar
yote
s 100
-200
nt

Det
ectio
n an
d A
naly
sis o
f In
Vivo
DN
A Re
plic
atio
n In
term
edia
tes
Rep
licat
ion
is lo
caliz
ed to
fork
s (A
utor
adio
grap
h E.
col
i DN
A)
fork
fork
daug
hter
daug
hter
pare
nt
Sho
rt (5
- 10
sec
) pul
se
thou
ght t
o re
pres
ent i
nitia
l nu
cleo
tide
inco
rpor
atio
n
Sem
i-dis
cont
inuo
us s
ynth
esis
(E
M re
plic
atin
g λ
DN
A)
EM c
an d
istin
guis
h SS
from
DS
DN
A
New
DN
A sy
nthe
sis
is s
mal
l (p
ulse
labe
l the
n si
ze)
smal
l
alka
line
sucr
ose
grad
ient
(siz
e)
HL
HL
HH
Infe
r H (l
abel
ed) a
nd L
(unl
abel
ed)
from
film
gra
in d
ensi
ty
SS
regi
ons
in tr
ans,
som
e in
terr
upte
d by
sm
all D
S s
egm
ents
SS D
NA
(lagg
ing)
SS D
NA
(lagg
ing)
D
S D
NA
(lead
ing)
DS
DN
A (le
adin
g)
cons
iste
nt w
ith d
isco
ntin
uous
ok
azak
i fra
gmen
t syn
thes
is o
nly
on
stra
nd o
f spe
cific
pol
arity
(lag
ging
)

Repl
icat
ion
Fork
Tas
ks a
nd th
e Act
iviti
es T
hat P
erfo
rm T
hem
sepa
rate
par
enta
l str
ands
begi
n D
NA
synt
hesis
stab
ilize
SS
DN
A
synt
hesiz
e D
NA
ensu
re p
roce
ssiv
ity
unlin
k pa
rent
al st
rand
s
Task
Act
ivity
helic
ase
prim
ase
SSBP
poly
mer
ase
clam
p lo
ader
/cla
mp
topo
isom
eras
e
conn
ect o
kaza
ki fr
agm
ents
repl
ace
prim
er
ligas
e
nucl
ease
/pol
ymer
ase

Bio
reg
2015
Lec
ture
1 R
eplic
atio
n
AP
PE
ND
IX

Usin
g Bi
oche
mic
al A
ssay
s to
Defi
ne B
ioch
emic
al F
unct
ions
Ass
ay m
ust d
istin
guish
or p
hysic
ally
sepa
rate
pro
duct
s fro
m su
bstr
ates
Smal
l diff
eren
ces i
n as
say
cond
ition
s can
defi
ne d
iffer
ent a
ctiv
ities
prim
ed si
ngle
-stra
nded
tem
plat
e de
fines
Pol
III h
oloe
nzym
e ac
tivity
Poly
mer
izat
ion:
con
vers
ion
of ra
dioa
ctiv
e nu
cleo
tide
from
aci
d so
lubl
e to
aci
d pr
ecip
itabl
e
can
be d
etec
ting
mul
tiple
type
s of a
ctiv
ities
and
be
affe
cted
by
mul
tiple
com
petin
g ac
tiviti
es
Com
plic
atio
ns o
f ass
ayin
g cr
ude
extr
acts
(bew
are
of w
astin
g cl
ean
thou
ghts
on d
irty
enzy
mes
)
Nuc
leas
e: c
onve
rsio
n of
inco
rpor
ated
radi
oact
ive
nucl
eotid
e fro
m a
cid
prec
ipita
ble
to a
cid
solu
ble
Qua
ntifi
catio
n is
impo
rtan
t for
infe
rrin
g bi
olog
ical
rele
vanc
e
3’ >
5’ e
xonu
clea
se n
egat
es th
e po
lym
eriz
atio
n re
actio
n, b
ut is
gen
eral
ly m
uch
slow
er
gapp
ed/n
icke
d te
mpl
ate
defin
es P
ol II
I cor
e ac
tivity
can
be d
etec
ting
mul
tiple
sim
ilar a
ctiv
ities
DN
A Po
l I’s p
oor p
olym
eriz
atio
n ra
ised
the
poss
ibili
ty th
at it
was
not
the
repl
icat
ive
helic
ase

The A
wes
ome
Chal
leng
es o
f Gen
etic
s po
lA m
utan
t rev
isite
d Le
ctur
e 1:
pol
A1
mut
ant w
ith <
1% a
ssay
able
DN
A Po
l1 a
ctiv
ity h
ave
rela
tivel
y no
rmal
repl
icat
ion
Cai
rns
conc
lude
s D
NA
Pol1
is n
ot im
port
ant f
or D
NA
repl
icat
ion
Lect
ure
2: D
NA
Pol1
pla
ys a
role
in o
kaza
ki fr
agm
ent m
atur
atio
n, a
n im
port
ant p
art o
f rep
licat
ion
Wha
t hap
pene
d to
the
awes
ome
pow
er o
f gen
etic
s?
Cav
eats
abo
ut g
ene
anal
ysis
C
avea
ts a
pplie
d to
pol
A1
mut
ants
Lim
itatio
ns in
Phe
noty
pic
Ana
lysi
s P
ol I
activ
ity in
livi
ng p
olA
1 ce
lls m
ay b
e gr
eate
r tha
n th
at m
easu
red
in
vitro
(in
extra
cts)
as
mut
ant p
rote
in m
ay b
e m
ore
labi
le o
r inh
ibita
ble
in
the
hars
her i
n vi
tro s
ettin
g th
an in
viv
o.
Exc
ess
Act
ivity
/Lea
ky A
llele
E
.col
i has
an
estim
ated
300
mol
ecul
es o
f DN
A P
ol I,
mos
t use
d in
DN
A re
pair.
Few
er m
olec
ules
are
nee
ded
for t
he 2
repl
icat
ion
fork
s, s
o th
e re
sidu
al a
ctiv
ity in
a p
olA
1 m
utan
t may
be
suffi
cien
t. N
ote,
alth
ough
po
lA1
has
an e
arly
non
sens
e m
utat
ion,
read
-thro
ugh
of th
e no
nsen
se
codo
n is
sus
pect
ed o
f gen
erat
ing
the
resi
dual
Pol
I ac
tivity
Red
unda
ncy
We
can
elim
inat
e th
e fir
st tw
o ca
veat
s w
ith a
nul
l mut
ant,
but t
he
polA∆
mut
ant i
s st
ill v
iabl
e in
min
imal
med
ia (a
lthou
gh n
ot in
rich
m
edia
, whe
re th
e de
man
ds fo
r rap
id D
NA
repl
icat
ion
are
grea
ter)
. In
this
mut
ant P
ol II
or P
ol II
I is
thou
ght t
o su
bstit
ute
(poo
rly b
ut
suffi
cien
tly) f
or P
ol I
in O
F m
atur
atio
n
With
all
thes
e ca
veat
, wha
t is
the
evid
ence
that
DN
A P
ol1
is im
porta
nt fo
r OF
mat
urat
ion
and
DN
A re
plic
atio
n?
polA
12 ts
mut
ant a
ccum
ulat
es in
crea
sed
OFs
at r
estri
ctiv
e te
mp
(sim
ilar t
o th
e ts
lig4
mut
ant)
polA
12 li
g4 d
oubl
e m
utan
t not
onl
y ac
cum
ulat
es O
Fs b
ut ra
pidl
y ce
ases
DN
A sy
nthe
sis
at re
stric
tive
tem
p

Gen
eral
Str
ateg
ies
for I
sola
ting
DN
A Sy
nthe
sis
Mut
ants
1) S
cree
n: a
ssay
mac
rom
olec
ular
syn
thes
is in
viv
o in
ts-le
thal
mut
ants
soo
n af
ter s
hift
to re
stric
tive
tem
p
D
NA
synt
hesi
s
–
(3 H
Thy
min
e in
corp
.)
RN
A sy
nthe
sis
+
P
rote
in s
ynth
esis
+
Nuc
leot
ide
synt
hesi
s +
2)
Sel
ectiv
e en
richm
ent:
ts m
utan
ts th
at fa
il to
inco
rpor
ate
pois
onou
s
n
ucle
otid
e an
alog
dur
ing
tran
sien
t shi
ft to
rest
rictiv
e te
mp
w
ash
out
5-B
U
5-B
U
UV
Rep
licat
ion
mut
ants
poo
rly in
corp
orat
e 5-
BU
- su
rviv
e
32°
42°
42°
32°
WT
inco
rpor
ate
5-br
omou
raci
l (5-
BU
) - U
V se
nsiti
ve
Can
recy
cle
surv
ivor
s to
furt
her e
nric
h

Mism
atch
Rep
air:
Cor
rect
ion
of R
eplic
atio
n Er
rors
E. c
oli m
ismat
ch re
pair
Mut
S bo
und
to m
ispai
red
DN
A
Mut
H -
reco
gniz
es n
earb
y G
ATC
Mut
L - a
ssoc
iatio
n w
ith M
utS
and
Mut
H st
imul
ates
Mut
H to
nic
k
un
met
hyla
ted
daug
hter
stra
nd (b
asis
of st
rand
bia
s)
Exon
ucle
ase
and
helic
ase
II, d
irect
ed b
y M
utS
and
Mut
L
ex
cise
dau
ghte
r stra
nd fr
om n
ick
to m
ispai
red
bp
DN
A po
lIII,
clam
p, c
lam
p-lo
ader
, and
SSB
synt
hesiz
e
re
plac
emen
t DN
A
DN
A!
Dam
met
hyla
se e
vent
ually
fully
met
hyla
tes
GAT
C s
ites
so b
oth!
stra
nds
are
mar
ked
as p
aren
tal f
or n
ext r
ound
of r
eplic
atio
n !
DN
A- b
oth
pare
ntal
stra
nds m
ethy
late
d at
GAT
C sit
es
d
augh
ter s
trand
tran
sient
ly u
nmet
hyla
ted
afte
r rep
licat
ion
Mut
S - r
ecog
nize
s misp
aire
d bp
by
susc
eptib
ility
to k
inki
ng

Dem
onst
ratin
g 5’→
3’ C
hain
Gro
wth
IF 5
’→
3’
corr
ect
P-P-
P O
H
5’
3’
+ O
H
5’
3’
OH
poly
mer
gro
wth
+ PP
i
IF 3
’→
5’ c
orre
ct +
OH
5’
3’
PPi
poly
mer
gro
wth
Test
: U
se d
ideo
xy n
ucle
otid
e to
ask
if 3
’ O
H re
quire
d fo
r mon
omer
inco
rpor
atio
n?
Res
ult:
Did
eoxy
is in
corp
orat
ed b
ut s
ubse
quen
t pol
ymer
izat
ion
is b
lock
ed
P-P-
P
P-P-
P P
P P
P-P-
P O
H
5’
3’
P P
P
P-P-
P P
P P
P
OH
P P
P P-
P-P
P
+
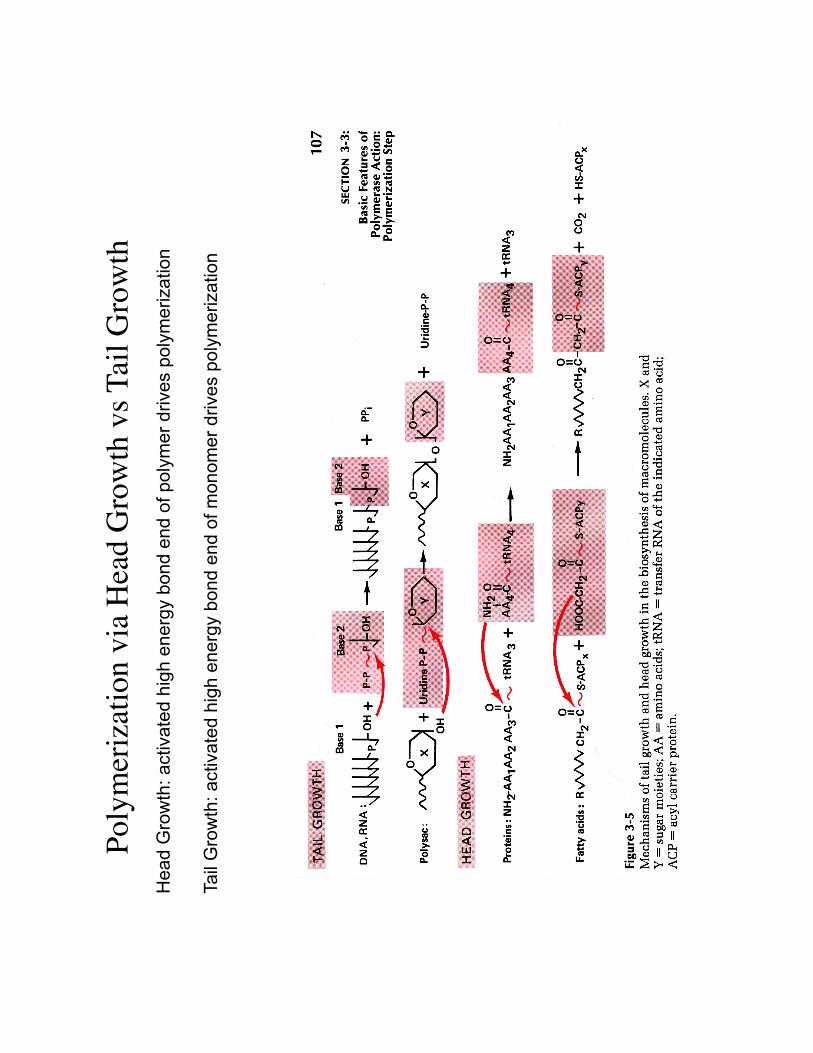
Poly
mer
izat
ion
via
Hea
d G
row
th v
s Tai
l Gro
wth
H
ead
Gro
wth
: act
ivat
ed h
igh
ener
gy b
ond
end
of p
olym
er d
rives
pol
ymer
izat
ion
Tail
Gro
wth
: act
ivat
ed h
igh
ener
gy b
ond
end
of m
onom
er d
rives
pol
ymer
izat
ion
























