Embed Size (px)
Citation preview

Behaviour 154 (2017) 1123–1144 brill.com/beh
Do songbirds attend to song categories when selectingbreeding habitat? A case study with a wood warbler
Janice K. Kelly ∗ and Michael P. Ward
Department of Natural Resources and Environmental Sciences, University of Illinois,Urbana, IL 61801, USA
*Corresponding author’s e-mail address: [email protected]
Received 19 October 2017; accepted 28 October 2017; published online 14 November 2017
AbstractBreeding habitat selection strongly affects reproduction and individual fitness. Among birds, usingsocial cues from conspecifics to select habitat is widespread, but how different types of conspecificsocial cues influence breeding habitat selection remains less understood. We conducted a play-back experiment evaluating if the yellow warbler (Setophaga petechia), a species with two songcategories linked to pairing status, uses categories differently when selecting breeding habitat. Wehypothesized that yellow warblers use second-category singing mode, which is mostly sung bypaired males, over first-category singing mode for habitat selection, as successfully paired malesshould indicate higher-quality habitat. We broadcast yellow warbler first-category singing mode,second-category singing mode, and silent controls at sites in Illinois. Yellow warblers were moreabundant at sites treated with second-category singing mode compared other sites. Our resultsdemonstrate that yellow warblers use social cues informing successful pairing over other types ofsocial cues to select breeding habitat.
Keywordshabitat selection, social information, conspecific attraction, social cue, song category, yellowwarblers.
1. Introduction
Breeding habitat selection is a critical decision-making process becauseof its direct effects on fitness (Holmes et al., 1996; Arlt & Pärt, 2007).To maximize reproductive success, individuals can collect information onhabitat quality and choose the highest-quality habitat available (Koops,2004; Stamps & Krishnan, 2005). Often times, individuals base their de-cisions on reproductive experience in a habitat (e.g., win-stay lose-switch:
© Koninklijke Brill NV, Leiden, 2017 DOI 10.1163/1568539X-00003461

1124 Song categories are social cues for habitat selection
Schmidt, 2004; Chalfoun & Martin, 2010), directly assessing predation risk(e.g., predator presence/absence: Emmering & Schmidt, 2010; Buxton et al.,2017), or by sampling environmental cues linked to successful reproduction(e.g., vegetation cues: Rosenzweig, 1973; Cody, 1981; Martin, 2001; foodabundance: Murkin et al., 1997). Although these cues provide informationfor decision-making, collecting this information can be time consuming andcostly (Valone, 2007). Alternatively, individuals can use social informationto assess habitat quality by sampling cues from other individuals settled inthe environment (Seppänen et al., 2007). Social information use can removethe cost of search time typically incurred when acquiring personal infor-mation or directly sampling the environment (Valone & Templeton, 2002).Moreover, social cues can enhance fitness by steering individuals towardhigher-quality habitat where reproductive success is high (Fletcher, 2006).Using intraspecific social cues for habitat selection, termed conspecific at-traction (Stamps, 1988; Reed & Dobson, 1993; Ahlering et al., 2010), isparticularly beneficial due to similar breeding habitat requirements amongconspecifics (Holt, 2009). Indeed, conspecific attraction is a common phe-nomenon observed across taxa (arthropods: Fletcher & Miller, 2008; am-phibians: Buxton et al., 2015; molluscs: Lecchini, 2011; reptiles: Scott et al.,2013; birds: Ahlering et al., 2010; mammals: Matthysen, 2005).
It is clear that many species use conspecific attraction to select breed-ing habitat, but what remains less understood is whether the type of socialcue present affects settlement decisions (Betts et al., 2008; Szymkowiak etal., 2016). Location-based social cues, such as the position of other indi-viduals or visual presence, contain information about conspecific presencein a prospective habitat (Betts et al., 2008). These social cues, however,do not provide much information on reproductive performance and successof those settled in the habitat (Betts et al., 2008; Rushing et al., 2015). Incontrast, performance-based social cues, such as offspring presence, directlyinform habitat quality because the reproductive consequences of settlementdecisions are made apparent by presence of the cue (Danchin et al., 2004;Valone, 2007; Betts et al., 2008). Females of some insect species, for ex-ample, select oviposition sites using the presence of conspecific eggs andnymphs rather the presence of adult conspecifics (Rudolf & Rödel, 2005;Fletcher & Miller, 2008; Raitanen et al., 2014). Many bird species, particu-larly among songbirds, use performance-based social cues to select breedinghabitat (Brown et al., 2000; Ward, 2005; Betts et al., 2008; Pärt et al., 2011;

J.K. Kelly, M.P. Ward / Behaviour 154 (2017) 1123–1144 1125
Rushing et al., 2015), with one study demonstrating that performance-basedsocial cues elicit more individuals to settle than location-based social cues(fledgling vocalizations versus adult song; Betts et al., 2008). Similarly, the-oretical models suggest that strategies using cues of conspecific reproductivesuccess outperform other strategies for breeding site selection (e.g., conspe-cific presence: Doligez et al., 2003). With all else being equal, individualsshould use performance-based social cues over location-based cues becauseof their high reliability for assessing breeding habitat quality (sensu Parejoet al., 2007).
Most studies consider signs of reproductive success as performance-basedsocial cues (e.g., Brown et al., 2000; Betts et al., 2008; Kelly & Schmidt,2017). These cues, however typically can only be sampled at the end of thebreeding season and used after a one-year delay. Migratory songbirds coulduse the singing behaviour of territorial males as a performance-based so-cial cue available directly upon spring arrival at the breeding habitat (e.g.,Rushing et al., 2015). In some songbirds, males have two distinctive singingmodes for specific contexts: one mode is mostly sung while unpaired formate attraction (i.e., ‘first-category singing mode’; Catchpole, 1983; Spector,1992; Titus, 1998), and the second is mostly sung after successfully pairing(i.e., ‘second-category singing mode’; Spector, 1991, 1992; Proppe & Ritchi-son, 2008). Depending on the species, first- and second-category singingmodes are referred to as either ‘Type I and Type II’, where singing behaviourdistinguishes the two categories (Spector et al., 1989; Spector, 1991; Staicer,1996; Beebee, 2002; Jane & Ryker, 2011), or ‘accented and unaccented,’where the aural/phonetic characteristics distinguish the categories (Morse,1966; Weary et al., 1994a; Byers, 1996; Hof & Podos, 2013). In both namingsystems, first- and second-category singing modes may contain different in-formation about reproductive opportunities within the habitat. First-categorysinging mode would inform prospectors of where territorial males are lo-cated, while second-category singing mode informs prospectors of whichsites contain successfully paired males.
For socially monogamous songbird species, individuals could expectmore extra-pair copulation opportunities in habitat where many males arepaired (e.g., Mayer & Pasinelli, 2013 and references therein). Thus, song-birds may treat first- and second-category singing modes as location-basedand performance-based social cues to select habitat with, and choose siteswhere second-category singing mode is more prevalent because knowing

1126 Song categories are social cues for habitat selection
pairing status is a step closer towards reproductive success. To date, most re-search addresses distinction between singing mode categories in the contextof sexual selection or territoriality (reviewed in Byers & Kroodsma, 2009;Reichard & Anderson, 2015), but rarely do studies consider singing modecategories in terms of different social cues useful for habitat selection (e.g.,Szymkowiak et al., 2016).
We used experimental playbacks to test if the yellow warbler (Se-tophaga petechia), a wood warbler with well-studied singing mode cate-gories (Morse, 1966; Fickens & Fickens, 1970; Spector, 1991; Weary et al.,1994b; Beebee, 2002), select breeding habitat based on how much first- andsecond-category singing mode is present at the breeding site. In yellow war-blers, singing mode functions as a sex-specific signal: males engage in morein first-category singing mode when interacting with females before pairing,and typically sing more in second-category singing mode when interactingwith males after successfully pairing (Spector, 1991; Weary et al., 1994b;but see Beebee, 2004). Thus, second-category singing mode contains usefulinformation about potential mating opportunities that individuals can usurpwhen making settlement decisions. We expected that because the second-category singing mode indicates pairing success, yellow warblers would treatsecond-category singing mode as a performance-based social cue and selecthabitat with more second-category singing mode present than first-category.We further discuss the costs and benefits of settling near conspecifics singingsecond-category song from the perspective of both males prospecting forhabitat and settled males singing second-category song.
2. Material and methods
2.1. Study species
The yellow warbler is a long-distant migratory songbird that arrives at mostbreeding grounds between early to mid-May (Lowther et al., 1999). Males ar-rive a week earlier than females in our system (J.K.K., personal observation),and up to ten days earlier in other systems (Lowther et al., 1999). Yellowwarblers breed in a wide variety of shrublands, such as overgrown fields(Lowther et al., 1999), shrubby wetlands and uplands (Humple & Burnett,2010), as well as in developed areas such as along roadsides or in suburbanparks (Rousseau et al., 2015). Yellow warblers have moderate site fidelitywith return rates around 0.30 (Yezerinac et al., 1996; but see Cilimburg et

J.K. Kelly, M.P. Ward / Behaviour 154 (2017) 1123–1144 1127
al., 2002 for return rates up to 0.42) such that immigration/emigration be-tween breeding sites is likely in this species (Anders & Marshall, 2005).Thus, we expect that adults make settlement decisions based on social cues(specifically song categories) sampled upon arrival, as these cues should bemore accessible than environmental cues (Valone, 2007).
2.2. Study sites
We established 44 study sites at shrublands throughout central and northernIllinois on both state and private land (see Figure A1 in the Appendix forlocations by county). Sites were selected opportunistically. We used satel-lite imagery on Google Earth PRO 7.1 (Google Inc. 2013) to locate andmeasure the area of shrublands, and ground-truthed locations to verify thateach shrubland contained habitat suitable for yellow warblers. Honeysuckle(Lonicera spp.), multi-flora rose (Rosa multiflora), autumn olive (Elaeagnusumbellate), young dogwood (Cornus spp.) and young willows (Salix spp.)were the dominant shrubs at each site. A mix of forest, agricultural fields,and suburban landscapes surrounded shrublands. All shrublands were >1 kmapart to ensure independence. We measured site area by visually delineat-ing shrublands in Google Earth PRO 7.1 using the Polygon option withinthe Ruler tool (Google Inc. 2013). Sites ranged considerably in size (1.16–41.76 ha, mean: 8.87 ha), but the variation was comparable among sites foreach treatment (in ha; first-category sites: max 33.74; min 1.56; mean 8.09;second-category sites: max 47.96; min 1.95; mean 11.18; silent control sites:max 32.88: min 1.16; mean 7.49).
2.3. Playback experiment
During the 2014 breeding season, we conducted a playback experiment andrandomly assigned study sites one of three treatments: (1) first-categorysinging mode (location-based social cue, n = 15), (2) second-categorysinging mode (performance-based social cue, n = 14), or (3) silent controlswith no playback equipment or stimulus (n = 15). We did not broadcastprocedural sounds (e.g., white noise) at silent control sites because simi-lar experiments found no difference in response between procedural con-trols and control sites without any playback equipment (e.g., Emmering &Schmidt, 2011; Hua et al., 2013). At sites with song treatments, we estab-lished stations comprised of one FOXPRO model NX3 or NX4 game caller(FOXPRO, Lewiston, PA, USA) powered by a 12 V deep-cycle car battery

1128 Song categories are social cues for habitat selection
and operated by a Diehl 12V DC digital timer (Diehl Stiftung, Nuremberg,Germany). Equipment was housed in a plastic bin with mesh-covered open-ings for sound transmission and camouflaged with spray-paint. Playbackstations were placed directly on the ground. Broadcast started mid-April be-fore males arrived at sites (the first male at any site was detected on 30 April).Broadcast ran daily until early-July for approx. 10 weeks total, and encom-passed both the earliest and peak portions of the breeding season (late Mayto early June; J.K.K., personal observation). Playback was broadcast in themorning (0500–0800 h) and late in the afternoon (1700–1900 h) to mimicnatural vocal activity of yellow warblers (Lowther et al., 1999), as well astwo periods at night (2200–0000 and 0200–0300 h) to simulate the vocalactivity of migrants known to attract conspecifics (Alessi et al., 2010).
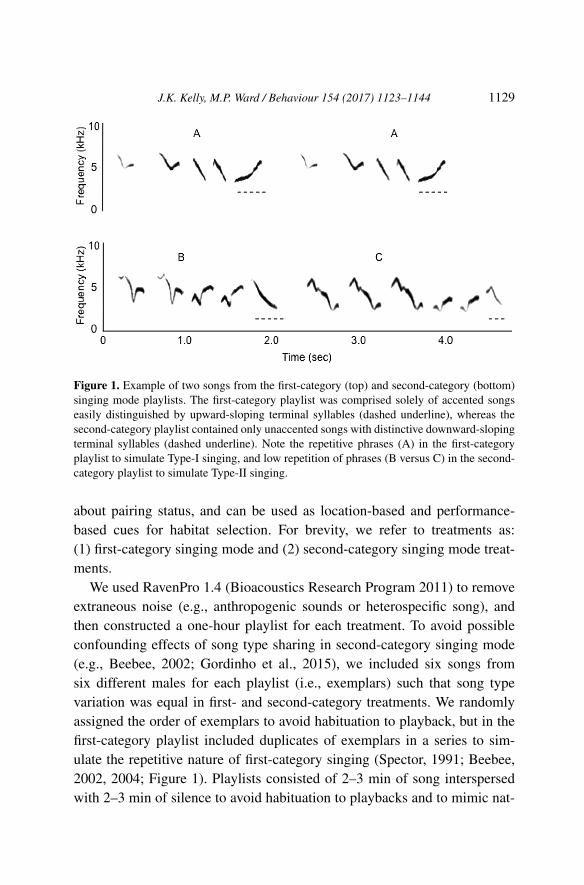
We used recordings of yellow warblers provided by the Borror Labora-tory of Bioacoustics from the Midwestern United States (specifically Illinois,Indiana and Ohio) to avoid confounding effects of song dialect. Yellow war-blers singing modes are described as both Type I versus Type II (character-ized, respectively, by high and low repetition of songs throughout the male’srepertoire; Spector, 1991; Weary et al., 1994b; Beebee, 2002), as well as ac-cented versus unaccented (characterized aurally by the downward or upward-sloping structure of a song’s last syllable; Morse, 1966; Fickens & Fickens,1970; Bankwitz & Thompson, 1979). Thus we incorporated both charac-terization systems (Type I/Type II and accented/unaccented) to constructfirst-/second-category singing mode playlists for our treatments (Figure 1).As per Spector’s distinction between singing mode categories (1991), weselected songs for our first-category singing mode playlist that had higher-frequency first syllables with less variation among first syllable frequenciesacross songs compared to the second-category singing mode playlist (meanfrequency for first-category playlist ± SD: 6562.50 ± 290.47 Hz; second-category: 5625.00 ± 660.26 Hz). We also made our first-category singingmode playlist more repetitive than our second-category singing mode playlist(0.50 vs. 0.23, where repetition is the proportion of songs on the playlist fol-lowed by the same song; Spector, 1991).
Considering how we constructed playlists for our treatments, we rec-ognize the alternative way to refer to treatments are: (1) accented songswith less immediate variation (location-based cue) and (2) unaccentedsongs with more immediate variation (performance-based cue). Nonethe-less, we think the playlists deliver two types of social cues with information
J.K. Kelly, M.P. Ward / Behaviour 154 (2017) 1123–1144 1129
Figure 1. Example of two songs from the first-category (top) and second-category (bottom)singing mode playlists. The first-category playlist was comprised solely of accented songseasily distinguished by upward-sloping terminal syllables (dashed underline), whereas thesecond-category playlist contained only unaccented songs with distinctive downward-slopingterminal syllables (dashed underline). Note the repetitive phrases (A) in the first-categoryplaylist to simulate Type-I singing, and low repetition of phrases (B versus C) in the second-category playlist to simulate Type-II singing.
about pairing status, and can be used as location-based and performance-based cues for habitat selection. For brevity, we refer to treatments as:(1) first-category singing mode and (2) second-category singing mode treat-ments.
We used RavenPro 1.4 (Bioacoustics Research Program 2011) to removeextraneous noise (e.g., anthropogenic sounds or heterospecific song), andthen constructed a one-hour playlist for each treatment. To avoid possibleconfounding effects of song type sharing in second-category singing mode(e.g., Beebee, 2002; Gordinho et al., 2015), we included six songs fromsix different males for each playlist (i.e., exemplars) such that song typevariation was equal in first- and second-category treatments. We randomlyassigned the order of exemplars to avoid habituation to playback, but in thefirst-category playlist included duplicates of exemplars in a series to sim-ulate the repetitive nature of first-category singing (Spector, 1991; Beebee,2002, 2004; Figure 1). Playlists consisted of 2–3 min of song interspersedwith 2–3 min of silence to avoid habituation to playbacks and to mimic nat-

1130 Song categories are social cues for habitat selection
ural singing behaviour of territorial males (e.g., Ward & Schlossberg, 2004;Andrews et al., 2015). Playlists were adjusted to peak amplitude in order tostandardize amplitude across exemplars so that all broadcast at similar vol-umes; we did not use filters on either playlist.
We quantified yellow warbler abundance at each site by conductingweekly 10-min point counts after playbacks started (5 May through 1 July2014, n = 8 surveys per site) to correspond with breeding activity of yel-low warblers at our sites. At each site, we conducted two point counts, oneat the playback station and one >300 m away in a random direction to en-sure independence. Sites were surveyed simultaneously by four people, withone person per site conducting both counts. To improve detection, we turnedplaybacks off during counts. Detection likely did not differ between treat-ments, as yellow warblers do not seem to change vocal behaviour in responseto different singing mode categories (such as song rate or song type sung;Beebee, 2004). We conducted counts from sunrise until 1000 h and recordedall yellow warblers seen or heard within 100 m of each sampling point.During counts, we also recorded whether individuals within earshot usedaccented (location-based social cue) or unaccented singing (performance-based social cue) to quantify the amount of natural second-category singing.Of the 613 detections where males sang, 20% used both accented and unac-cented singing. For these cases, we categorized the song as a performance-based social cue because mixed accents likely indicates successful pairing(Fickens & Fickens, 1965; Morse, 1966; Bankwitz & Thompson, 1979). Weadded the total number of individuals singing accented and unaccented songsduring the two counts at each site, and then calculated the ratio of unaccentedto accented singing per site visit to represent relative amounts of unaccentedsinging at sites in analyses (herein referred to as ‘males using unaccentedsong’). Sampling order was rotated each week to ensure all sites were sam-pled early in the morning at least once. We did not conduct point countsduring high winds or heavy rain. For statistical analyses, we used averagenumber of yellow warblers observed per site visit as the metric for abun-dance.
2.4. Statistical analyses
Count data were not normally distributed and variance was unequal acrosstreatments. Thus, we built a generalized linear mixed model fitted with aPoisson distribution with site treated as a random factor. We used individual

J.K. Kelly, M.P. Ward / Behaviour 154 (2017) 1123–1144 1131
counts at sites as the response variable and treatment, site area, males usingunaccented song, and all 2-way interactions (treatment × unaccented song,treatment × area, area × unaccented song) as main effects. Initially, we alsoincluded sampling week in the model to assess if responses to treatmentschanged over time, but removed the term and all interactions as none weresignificant (F1-2,669 = 0.01–1.38, p = 0.24–0.98). To determine if our threetreatments significantly differed from one another in yellow warbler abun-dance, we contrasted least squares mean estimates for each treatment, whilecentring area and unaccented song (Schielzeth, 2010). We built all models inSAS 9.3 (SAS Institute, Cary, NC, USA).
3. Results
Yellow warbler abundance was greater at sites where second-category songwas played (Figure 2), as well as where a higher proportion of males weresinging unaccented song (Figure 3, Table 1). There was a significant dif-ference in yellow warbler abundance between the treatments (least-squaresmean estimate ± SE; silent: −1.35 ± 0.20, p < 0.001; first-category:−0.64 ± 0.11, p < 0.001; second-category: −0.59 ± 0.11, p < 0.001;Figure 2). There were twice as many yellow warblers at second-category
Figure 2. Mean yellow warbler abundances, measured via weekly point counts at 44 play-back treatment sites in 2014. The bar graph is of actual mean estimates from the data. Barsrepresent standard error.

1132 Song categories are social cues for habitat selection
Figure 3. Regression of mean yellow warbler counts, measured via weekly point counts at 44playback treatment sites in 2014, on the proportion of males at sites using unaccented song(with regression lines, R2 coefficients), where x-marks indicate silent controls sites with noplayback (dashed–dotted line), circles first-category song treatment sites (dashed line) andtriangles second-category song treatments sites (solid line). Points are actual mean estimatesfrom the data. Bars represent standard error.
singing mode treatment sites than silent control sites; warblers were ∼20%more abundant at second-category song treatment sites than first-categorysinging mode sites (Figure 2). There were also significant interactive ef-fects between treatment and males using unaccented song on yellow warblerabundance. Specifically, yellow warblers abundance was highest at second-category singing mode treatment sites where more males sang unaccentedsong (i.e., natural second-category singing) compared to other treatments(Figure 3). Yellow warbler abundance was not correlated with the area ofa site (R = 0.16, p = 0.28; Table 1). However, interactions between areaand treatment, as well as between area and males using unaccented song didinfluence yellow warbler abundance (Table 1). Yellow warblers were mostabundant at sites where more unaccented songs are present (Figure 3), andof these sites, the largest second-category song treatment areas had the mostyellow warblers (Figure 4).

J.K. Kelly, M.P. Ward / Behaviour 154 (2017) 1123–1144 1133
Table 1.Generalized linear mixed model results regressing counts of yellow warblers at sites onplayback treatment (first-category song, second-category song or silent controls), relativeproportion of males singing unaccented song at sites (unaccented song) and site area (ha).
Effect Estimate SE df(num, denom) F p
Intercept 1.25 0.16 – – –Treatment – – 671, 2 7.96 <0.001Silent −0.84 0.21 – – –First-category −0.48 0.21 – – –Unaccented song −2.28 0.22 671, 1 222.17 <0.001Area −0.05 0.01 671, 1 0.22 0.64Area × Treatment – – 671, 2 20.78 <0.001Silent 0.13 0.02 – – –First-category 0.04 0.01 – – –Unaccented song × Area 0.05 0.01 671, 1 18.77 <0.001Unaccented song × Treatment – – 671, 2 4.92 0.01Silent 0.78 0.26 – – –First-category 0.25 0.24 – – –
Regression estimates, standard error (SE) and degrees of freedom (df: numerator, denom-inator) are given for each model. Second-category song treatments served as the referencelevel for regression estimates.
Figure 4. Modelled estimates from regression of mean yellow warbler counts on the area ofthe site (ha), where x-marks indicate silent controls sites with no playback (dashed–dottedline), circles first-category song treatment sites (dashed line) and triangles second-categorysong treatments sites (solid line). Points are mean estimates from the regression model. Barsrepresent standard error.

1134 Song categories are social cues for habitat selection
4. Discussion
Yellow warblers appear to distinguish singing mode categories as differ-ent sources of social information and select breeding habitat based on thecategory present. Sites with high numbers of males using unaccented songhad more yellow warblers, providing further support for the hypothesis thatyellow warblers select habitat based on the type of social cue present, specif-ically by choosing habitat with performance-based social cues, rather thanlocation-based social cues.
Many studies demonstrate that songbirds use both location-based andperformance-based social cues to select breeding habitat with (reviewedin Szymkowiak, 2013). Moreover, both theoretical models (Doligez et al.,2003) and other playback experiments (e.g., Betts et al., 2008) demonstratethat songbirds rely more on performance-based social cues than location-based social cues for habitat selection. Often times, however, performance-based social information is only available during the post-breeding seasonand is inaccessible to most individuals (Nocera et al., 2006; Betts et al.,2008; Rushing et al., 2015). Male yellow warblers arrived at our sites inlate-April, and some males paired with females as soon as they arrivedthe first week of May. Males continue to arrive/pair throughout May, withpeak-nesting/territory defence occurring late May through early June. Nest-ing slows throughout June, and the breeding season ends with territoriesbreaking down in early July (J.K.K., personal observation). Thus, our resultssuggest that performance-based social cues are as available during the pre-breeding season as location-based social cues are for habitat selection (lateApril through early May in our system), specifically in the form of second-category singing mode.
Second-category singing mode may not be available to first-arrivingmales, and performance-based social cues will become more prevalent laterin the season. As the breeding season progresses, individuals may switchto using other social cues more readily available later in the season, suchas fledgling activity (Betts et al., 2008; Kelly & Schmidt, 2017) or post-breeding singing activity (Nocera et al., 2006; Betts et al., 2008; Farrellet al., 2012). These other social cues would be particularly useful to indi-viduals prospecting for future breeding locations during the post-breedingseason (e.g., Betts et al., 2008; Pärt et al., 2011). With our results, we can-not determine if yellow warblers prioritized pre-breeding performance-basedsocial cues (second-category singing mode, in this case) over other social

J.K. Kelly, M.P. Ward / Behaviour 154 (2017) 1123–1144 1135
cues available throughout the entire breeding season. In several instances,however, we detected males at our treatment site before any nestlings orfledgling were present (J.K.K., personal observation). Future studies couldtest if second-category singing mode ‘trumps’ other sources of social infor-mation indicative of habitat quality (sensu Betts et al., 2008), or if songbirdsprioritize pre- over post-breeding performance-based social cues for habitatselection.
We only altered one song characterization, singing mode category, toserve as location-based and performance-based social cues, but male songcontains even greater variation that songbirds undoubtedly recognize andrespond to. Many male songbirds attract mates by changing acoustic charac-teristics (e.g., frequency modulation, song length, song matching; Beecher etal., 2000; Searcy & Beecher, 2009; Searcy et al., 2014), and switch betweensong types to signal aggression towards territorial intruders (Byers, 1996;Hof & Podos, 2013). For example, a similar experiment found that songbirdstreat male song quality as a social cue for habitat selection and preferentiallysettle where poor-quality song is broadcast (Szymkowiak et al., 2016). In ef-fort to control for all other song characteristics that could affect settlementdecisions (e.g., Szymkowiak et al., 2016), we used the same playlist at allsites within a treatment. By using the same playlists, there could have beenqualities other than singing mode category specific to the songs we chosethat we were unaware of. A more robust approach could be to play a vari-ety of exclusive second-category and first-category singing mode playlistsat sites (see Kroodsma et al., 2001). This design, however, inevitably intro-duces greater variation due to the many different acoustic characteristics, andmakes it challenging to associate social information with particular song fea-tures. Future studies might work to resolve this issue by using species withtwo simple song categories whose song has been intensively studied. Simi-larly, with our design we cannot tease apart whether individuals settled usedunaccented singing or the Type-II singing mode as the performance-basedsocial cue in our second-category song treatment. Although more researchis needed to address the specific acoustic characteristics songbirds use forhabitat selection, our experiment takes the first step towards understandinghow song variation may be distinguished as location-based and performance-based social cues during the habitat selection process.
Another area of study worth investigating is sex-specific habitat selectionin response to songs. This study focused solely on adult breeding males, as

1136 Song categories are social cues for habitat selection
only 7% of the 712 yellow warbler detections were female. Females eventu-ally settle in the same location as males, but may show different settlementresponses to song categories due to sex-specific audiences commonly linkedto each singing mode category (e.g., first-category for females, and second-category for males; Kroodsma et al., 1989; Spector, 1991; Byers, 1996).For example, if females prioritize their own pairing success with a socialmate, then habitat with more first-category singing mode prevalent couldbe more attractive to females looking for unpaired males. Similarly, age-specific differences in habitat selection could also occur. Second-categorysinging mode, for example, may be more readily available to second-yearsindividuals (i.e., individuals experiencing their first breeding season), whichoften arrive 1–2 weeks later than adults (e.g., Lozano et al., 1996; Stewartet al., 2002), presumably when adults have paired and switched to second-category singing mode. It also possible that some individuals do not attendto singing mode categories at all when selecting breeding habitat, but insteaduse site-fidelity and return to the same breeding sites regardless of socialcues present. Site-fidelity likely influenced settlement decisions at our sitesgiven that up to 40% of the bird may be returning individuals (Yezerinac etal., 1996; Cilimburg et al., 2002). Determining how sex, age and personalexperience (particularly site-fidelity) influence the types of social cues usedfor habitat selection would be a fruitful direction in this field.
From the perspective of a male prospecting for a breeding location, thereare potential costs and benefits to using second-category singing mode asa performance-based cue for settlement decisions. For benefits, if a male ispresent and successfully attracted a female, then the habitat is leastwise highenough quality for a female to settle and support a nesting attempt. In ad-dition, the presence of other females could provide extra-pair mating oppor-tunities that often increase with increasing female densities (e.g., Westneat& Sherman, 1997). Extra-pair mating is common among yellow warblers(Hobson & Sealy, 1989; Yezerinac & Weatherhead, 1997; Grunst & Grunst,2014), as well as other species with two singing mode categories (e.g., woodwarblers: reviewed in Spector, 1992; sparrows: Groschupf, 1985; Lohr et al.,2013) that exhibit some level of extra-pair paternity (reviewed in Westneat& Stewart, 2003). In terms of costs, established males could also cuckold themale settling nearby (e.g., Balsby & Dabelsteen, 2005). Likewise, competi-tion for space, food, and mates (e.g., Rodenhouse & Holmes, 1992; Rich-mond et al., 2011) could be greater at sites with more performance-based

J.K. Kelly, M.P. Ward / Behaviour 154 (2017) 1123–1144 1137
social cues such that later arriving males may be at a competitive disadvan-tage (sensu the ‘mate opportunity’ and ‘rank advantage’ hypotheses on malearrival times; reviewed in Kokko et al., 2006).
The male broadcasting second-category song also faces potential costsand benefits of males settling near them. The cost of attracting more malesin the vicinity that in turn use second-category singing mode is likely lowerthan cost of not using second-category singing mode while the male’s femaleis nesting. In larger patches of habitat, second-category singing mode shouldnot be costly due to fewer crowding effects (Fretwell & Lucas, 1970; Holmeset al., 1996). That is, with ample space for territories, males could afford tocompete with males settling in response to second-category singing. Indeed,we found significant interactions between area and song category treatment,and between area and amount of unaccented singing by males (Table 1).These interactions could indicate that second-category singing mode, eitherproduced by singing males or used by males assessing social cues for set-tlement decisions, mostly occurs when there enough space to avoid negativedensity-dependent effects (Fretwell & Lucas, 1970; Holmes et al., 1996).
We found a positive correlation between yellow warbler abundance andthe amount of unaccented singing by males (i.e., natural second-categorysinging), with highest abundances and amounts of unaccented song occur-ring at second-category treatment sites (Figure 3). With our data, we cannotdetermine if increased abundances at second-category singing mode treat-ment sites caused males to use unaccented singing, or if our second-categorysinging mode treatment caused males to sing more unaccented song, whichturn attracted more conspecifics. Other studies on wood warblers, however,could shed light on a mechanistic explanation for this correlation as wellas the settlement responses our experiment induced. In other wood war-blers, males use second-category singing during male-male aggressive in-teractions (Byers, 1996; Hof & Podos, 2013). Thus, either crowding effectsor ‘perceived male-male aggression’ from the playback could have causedmales to sing more unaccented song. There may be a positive feedback loopto second-category singing mode: more individuals result in more second-category singing due to male-male territorial interactions, and more second-category singing attracts more males and lead to more second-categorysinging. This feedback could ultimately lead to clustered distributions ofyellow warblers, a pattern that has been observed in this species (Clark &

1138 Song categories are social cues for habitat selection
Robertson, 1979). In conclusion, we believe this study is the first step in un-derstanding how specific song variants, in this case different singing modecategories, influence habitat selection.
Acknowledgements
We are grateful to Allison Hanna, Vicki Morgan and Hayley Stansell fortheir help in the field, and to an anonymous reviewer for valuable com-ments on earlier drafts of this manuscript. We thank the University of IllinoisCommittee on Natural Areas, Macon and Vermilion County ConservationDistricts, Forest Preserve Districts of Champaign, DuPage, Kane and Win-nebago Counties, McLean County Park District, Bourbonnais Township ParkDistrict, The Nature Conservancy, and the Illinois Department of Natural Re-sources for use of their properties. We thank the Borror Lab of Bioacousticsfor providing sound files used in our experiment. This research was con-ducted under the University of Illinois IACUC protocol No. 10127 and wasfunded by the Army Corps of Engineers, Construction Engineering ResearchLaboratory.
References
Ahlering, M.A., Arlt, D., Betts, M.G., Fletcher Jr., R.J., Nocera, J.J. & Ward, M.P. (2010).Research needs and recommendations for the use of conspecific attraction methods in theconservation of migratory songbirds. — Condor 112: 252-264.
Alessi, M.G., Benson, T.J. & Ward, M.P. (2010). Nocturnal social cues attract migratingyellow-breasted chats. — Wilson J. Ornithol. 122: 780-783.
Anders, A. & Marshall, M. (2005). Increasing the accuracy of productivity and survivalestimates in assessing landbird population status. — Conserv. Biol. 19: 66-74.
Andrews, J.E., Brawn, J.D. & Ward, M.P. (2015). When to use social cues: conspecificattraction at newly created grasslands. — Condor 117: 297-305.
Arlt, D. & Pärt, T. (2007). Nonideal breeding habitat selection: a mismatch between prefer-ence and fitness. — Ecology 88: 792-801.
Balsby, T.J. & Dabelsteen, T. (2005). Simulated courtship interactions elicit neighbor intru-sion in the whitethroat Sylvia communis. — Anim. Behav. 69: 161-168.
Bankwitz, K. & Thompson, W. (1979). Song characteristics of the yellow warbler. — WilsonBull. 91: 533-550.
Beebee, M. (2002). Song sharing by yellow warblers differ between two modes of singing:implications for song function. — Condor 104: 146-155.
Beebee, M. (2004). The functions of multiple singing modes: experimental tests in yellowwarblers, Dendroica petechia. — Anim. Behav. 67: 1089-1097.

J.K. Kelly, M.P. Ward / Behaviour 154 (2017) 1123–1144 1139
Beecher, M., Campbell, S., Burt, J., Hill, C. & Nordby, J. (2000). Song-type matching be-tween neighboring song sparrows. — Anim. Behav. 59: 21-27.
Betts, M.G., Hadley, A.S., Rodenhouse, N. & Nocera, J.J. (2008). Social information trumpsvegetation structure in breeding-site selection by a migrant songbird. — Proc. Roy. Soc.Lond. B: Biol. Sci. 275: 2257-2263.
Brown, C., Brown, M. & Danchin, E. (2000). Breeding habitat selection in cliff swallows:the effect of conspecific reproductive success on colony choice. — J. Anim. Ecol. 69:133-142.
Buxton, V.L., Ward, M.P. & Sperry, J.H. (2015). Use of chorus sounds for location of breedinghabitat in 2 species of anuran amphibians. — Behav. Ecol. 26: 1111-1118.
Buxton, V.L., Ward, M.P. & Sperry, J.H. (2017). Frog breeding pond selection in response topredators and conspecific cues. — Ethology 123: 397-404.
Byers, B. (1996). Messages encoded in the songs of chestnut-sided warblers. — Anim. Behav.52: 691-705.
Byers, B. & Kroodsma, D. (2009). Female mate choice and songbird song repertoires. —Anim. Behav. 77: 13-22.
Catchpole, C. (1983). Variation in the song of the great reed warbler Acrocephalus arundi-naceus in relation to mate attraction and territorial defence. — Anim. Behav. 31: 1217-1225.
Chalfoun, A.D. & Martin, T.E. (2010). Facultative nest patch shifts in response to nest preda-tion risk in the Brewer’s sparrow: a “win-stay, lose-switch” strategy? — Oecologia 163:885-892.
Cilimburg, A.B., Lindberg, M.S., Tewksbury, J.J. & Hejl, S.J. (2002). Effects of dispersal onsurvival probability of adult yellow warblers (Dendroica petechia). — Auk 119: 778-789.
Clark, K. & Robertson, R. (1979). Spatial and temporal multi-species nesting aggregations inbirds as anti-parasite and anti-predator defenses. — Behav. Ecol. Sociobiol. 5: 359-371.
Cody, M. (1981). Habitat selection in birds — the roles of vegetation structure, competitors,and productivity. — Bioscience 31: 107-113.
Danchin, E., Giraldeau, L.-A., Valone, T.J. & Wagner, R.H. (2004). Public information: fromnosy neighbors to cultural evolution. — Science 305: 487-491.
Doligez, B., Cadet, C., Danchin, E. & Boulinier, T. (2003). When to use public informa-tion for breeding habitat selection? The role of environmental predictability and densitydependence. — Anim. Behav. 66: 973-988.
Emmering, Q.C. & Schmidt, K.A. (2011). Nesting songbirds assess spatial heterogeneityof predatory chipmunks by eavesdropping on their vocalizations. — J. Anim. Ecol. 80:1305-1312.
Farrell, S.L., Morrison, M.L., Campomizzi, A.J. & Wilkins, R.N. (2012). Conspecific cuesand breeding habitat selection in an endangered woodland warbler. — J. Anim. Ecol. 81:1056-1064.
Fickens, M. & Fickens, W. (1965). Comparative ethology of the chestnut-sided warbler,yellow warbler, and American redstart. — Wilson Bull. 77: 363-375.
Fickens, M. & Fickens, W. (1970). Response of four warbler species to playback of their twosong types. — Auk 87: 296-304.

1140 Song categories are social cues for habitat selection
Fletcher Jr., R.J. (2006). Emergent properties of conspecific attraction in fragmented land-scapes. — Am. Nat. 168: 207-219.
Fletcher Jr., R.J. & Miller, C.W. (2008). The type and timing of social information altersoffspring production. — Biol. Lett. 4: 482-485.
Fretwell, S.D. & Lucas, H.L.J. (1970). On territorial behavior and other factors influencinghabitat distribution in birds part 1 — theoretical development. — Acta Biotheor. 19: 16-36.
Gordinho, L.D.O., Matheu, E., Hasselquist, D. & Neto, J.M. (2015). Song divergence be-tween subspecies of reed bunting is more pronounced in singing styles under sexualselection. — Anim. Behav. 107: 221-231.
Groschupf, K. (1985). Changes in 5-striped sparrow song in intrasexual and intersexualcontexts. — Wilson Bull. 97: 102-106.
Grunst, A. & Grunst, M. (2014). Multiple sexual pigments, assortative social pairing, andgenetic paternity in the yellow warbler (Setophaga petechia). — Behav. Ecol. Sociobiol.68: 1451-1463.
Hobson, K. & Sealy, S. (1989). Mate guarding in the yellow warbler Dendroica petechia. —Ornis Scand. 20: 241-249.
Hof, D. & Podos, J. (2013). Escalation of aggressive vocal signals: a sequential playbackstudy. — Proc. Roy. Soc. Lond. B: Biol. Sci. 280: 20131553.
Holmes, R., Marra, P. & Sherry, T. (1996). Habitat-specific demography of breeding black-throated blue warblers (Dendroica caerulescens): implications for population dynamics.— J. Anim. Ecol. 65: 183-195.
Holt, R.D. (2009). Bringing the Hutchinsonian niche into the 21st century: ecological andevolutionary perspectives. — Proc. Natl. Acad. Sci. USA 106: 19659-19665.
Hua, F., Fletcher Jr., R.J., Sieving, K. & Dorazio, R.M. (2013). Too risky to settle: avian com-munity structure changes in response to perceived predation risk on adults and offspring.— Proc. Roy. Soc. Lond. B: Biol. Sci. 280. DOI:10.1098/rspb.2013.0762.
Humple, D.L. & Burnett, R.D. (2010). Nesting ecology of yellow warblers (Dendroica pe-techia) in montane chaparral habitat in the northern Sierra Nevada. — West N. Am. Nat.70: 355-363.
Jane, S. & Ryker, L. (2011). Geographic variation in type I songs of black-throated graywarblers. — Wilson J. Ornithol. 123: 339-346.
Kelly, J.K. & Schmidt, K.A. (2017). Fledgling calls are a source of social information forconspecific, but not heterospecific, songbird territory selection. — Ecosphere 8. DOI:10.1002/ecs2.1512.
Kokko, H., Gunnarsson, T., Morrell, L. & Gill, J. (2006). Why do female migratory birdsarrive later than males? — J. Anim. Ecol. 75: 1293-1303.
Koops, M.A. (2004). Reliability and the value of information. — Anim. Behav. 67: 103-111.Kroodsma, D., Bereson, R., Byers, B. & Minear, E. (1989). Use of song types by the chestnut-
sided warbler — evidence for both intra-sexual and inter-sexual functions. — Can. J. Zool.67: 447-456.
Kroodsma, D., Byers, B., Goodale, E., Johnson, S. & Liu, W.-C. (2001). Pseudoreplicationin playback experiments, revisited a decade later. — Anim. Behav. 61: 1029-1033.

J.K. Kelly, M.P. Ward / Behaviour 154 (2017) 1123–1144 1141
Lecchini, D. (2011). Visual and chemical cues in habitat selection of sepioid larvae. — C.R.Biol. 334: 911-915.
Lohr, B., Ashby, S. & Wakamiya, S. (2013). The function of song types and song componentsin grasshopper sparrows (Ammodramus savannarum). — Behaviour 150: 1085-1106.
Lowther, P.E., Celada, C., Klein, N.K., Rimmer, C.C. & Spector, D.A. (1999). Yellow warbler(Setophaga petechia). — In: The birds of North America (Rodewald, P.G., ed.). CornellLab of Ornithology, Ithaca, NY. DOI:10.2173/bna.454.
Lozano, G.A., Perreault, S. & Lemon, R.E. (1996). Age arrival date and reproductive successof male American redstarts Setophaga ruticilla. — J. Avian Biol. 27: 164-170.
Martin, T. (2001). Abiotic vs. biotic influences on habitat selection of coexisting species:climate change impacts? — Ecology 82: 175-188.
Matthysen, E. (2005). Density-dependent dispersal in birds and mammals. — Ecography 28:403-416.
Mayer, C. & Pasinelli, G. (2013). New support for an old hypothesis: density affects extra-pair paternity. — Ecol. Evol. 3: 694-705.
Morse, D. (1966). Context of songs in the yellow warbler. — Wilson Bull. 78: 444-455.
Murkin, H., Murkin, E. & Ball, J. (1997). Avian habitat selection and prairie wetland dynam-ics: a 10-year experiment. — Ecol. Appl. 7: 1144-1159.
Nocera, J., Forbes, G. & Giraldeau, L. (2006). Inadvertent social information in breeding siteselection of natal dispersing birds. — Proc. Roy. Soc. Lond. B: Biol. Sci. 273: 349-355.
Parejo, D., White, J., Clobert, J., Dreiss, A. & Danchin, E. (2007). Blue tits use fledglingquantity and quality as public information in breeding site choice. — Ecology 88: 2373-2382.
Pärt, T., Arlt, D., Doligez, B., Low, M. & Qvarnstrom, A. (2011). Prospectors combine socialand environmental information to improve habitat selection and breeding success in thesubsequent year. — J. Anim. Ecol. 80: 1227-1235.
Proppe, D. & Ritchison, G. (2008). Use and possible functions of the primary and sustainedsongs of male grasshopper sparrows. — Am. Midl. Nat. 160: 1-6.
Raitanen, J., Forsman, J.T., Kivela, S.M., Maenpaa, M.I. & Valimaki, P. (2014). Attraction toconspecific eggs may guide oviposition site selection in a solitary insect. — Behav. Ecol.25: 110-116.
Reed, J. & Dobson, A. (1993). Behavioral constraints and conservation biology — conspe-cific attraction and recruitment. — Trends Ecol. Evol. 8: 253-256.
Reichard, D. & Anderson, R. (2015). Why sing softly? The structure, function and evolution-ary significance of low-amplitude signals. — Anim. Behav. 105: 253-265.
Richmond, S., Nol, E. & Burke, D. (2011). Avian nest success, mammalian nest predatorabundance, and invertebrate prey availability in a fragmented landscape. — Can. J. Zool.89: 517-528.
Rodenhouse, N.N. & Holmes, R.T. (1992). Results of experimental and natural food reduc-tions for breeding black-throated blue warblers. — Ecology 73: 357-372.
Rosenzweig, M.L. (1973). Habitat selection experiments with a pair of coexisting heteromyidrodent species. — Ecology 54: 111-117.

1142 Song categories are social cues for habitat selection
Rousseau, J.S., Savard, J.L. & Titman, R. (2015). Shrub-nesting birds in urban habitats: theirabundance and association with vegetation. — Urban Ecosyst. 18: 871-884.
Rudolf, V. & Rödel, M. (2005). Oviposition site selection in a complex and variable environ-ment: the role of habitat quality and conspecific cues. — Oecologia 142: 316-325.
Rushing, C.S., Dudash, M.R. & Marra, P.P. (2015). Habitat features and long-distance dis-persal modify the use of social information by a long-distant migratory bird. — J. Anim.Ecol. 84: 1469-1479.
Schielzeth, H. (2010). Simple means to improve the interpretability of regression coefficients.— Meth. Ecol. Evol. 1: 103-113.
Schmidt, K. (2004). Site fidelity in temporally correlated environments enhances populationpersistence. — Ecol. Lett. 7: 176-184.
Scott, M.L., Whiting, M.J., Webb, J.K. & Shine, R. (2013). Chemosensory discriminationof social cues mediates space use in snakes, Cryptophis nigrescens (Elapidae). — Anim.Behav. 85: 1493-1500.
Searcy, W. & Beecher, M. (2009). Song as an aggressive signal in songbirds. — Anim. Behav.78: 1281-1292.
Searcy, W., Akçay, C., Nowicki, S. & Beecher, M. (2014). Aggressive signaling in songsparrows and other songbirds. — Adv. Stud. Behav. 46: 89-125.
Seppänen, J., Forsman, J.T., Mönkkönen, M. & Thomson, R.L. (2007). Social informationuse is a process across time, space, and ecology, reaching heterospecifics. — Ecology 88:1622-1633.
Spector, D. (1991). The singing behavior of yellow warblers. — Behaviour 117: 29-52.
Spector, D. (1992). Wood-warbler song systems. A review of paruline singing behaviors. —Curr. Ornithol. 9: 199-238.
Spector, D., McKim, L. & Kroodsma, D. (1989). Yellow warblers are able to learn songs andsituations in which to use them. — Anim. Behav. 38: 723-725.
Staicer, C. (1996). Acoustical features of song categories of the Adelaide’s warbler (Den-droica adelaidae). — Auk 113: 771-783.
Stamps, J. (1988). Conspecific attraction and aggregation in territorial species. — Am. Nat.131: 329-347.
Stamps, J. & Krishnan, V. (2005). Nonintuitive cue use in habitat selection. — Ecology 86:2860-2867.
Stewart, R.L.M., Francis, C.M. & Massey, C. (2002). Age-related differential timing of springmigration within sexes in passerines. — Wilson Bull. 114: 264-271.
Szymkowiak, J. (2013). Facing uncertainty: how small songbirds acquire and use socialinformation in habitat selection process? — Springer Sci. Rev. 1: 115-131.
Szymkowiak, J., Thomson, R. & Kuczynski, L. (2016). Wood warblers copy settlementdecisions of poor quality conspecifics: support for the tradeoff between the benefit ofsocial information use and competition avoidance. — Oikos 125: 1561-1569.
Titus, R. (1998). Short-range and long-range songs: use of two acoustically distinct songclasses by dark-eyed juncos. — Auk 115: 386-393.

J.K. Kelly, M.P. Ward / Behaviour 154 (2017) 1123–1144 1143
Valone, T. & Templeton, J. (2002). Public information for the assessment of quality:a widespread social phenomenon. — Philos. Trans. Roy. Soc. B: Biol. Sci. 357: 1549-1557.
Valone, T.J. (2007). From eavesdropping on performance to copying the behavior of others:a review of public information use. — Behav. Ecol. Sociobiol. 62: 1-14.
Ward, M.P. (2005). Habitat selection by dispersing yellow-headed blackbirds: evidence ofprospecting and the use of public information. — Oecologia 145: 1432-1939.
Ward, M.P. & Schlossberg, S. (2004). Conspecific attraction and the conservation of territorialsongbirds. — Conserv. Biol. 18: 519-525.
Weary, D., Lemon, R. & Perreault, S. (1994a). Different responses to different song types inAmerican redstarts. — Auk 111: 730-734.
Weary, D., Lemon, R. & Perreault, S. (1994b). Male yellow warblers vary use of song typesdepending on pairing status and distance from nest. — Auk 111: 727-729.
Westneat, D.F. & Sherman, P.W. (1997). Density and extra-pair fertilizations in birds: a com-parative analysis. — Behav. Ecol. Sociobiol. 41: 205-215.
Westneat, D.F. & Stewart, I.R.K. (2003). Extra-pair paternity in birds: causes, correlates, andconflict. — Annu. Rev. Ecol. Evol Syst. 34: 365-396.
Yezerinac, S.M. & Weatherhead, P.J. (1997). Extra-pair mating, male plumage colorationand sexual selection in yellow warblers (Dendroica petechia). — Proc. Roy. Soc. Lond.B: Biol. Sci. 264: 527-532.
Yezerinac, S.M., Weatherhead, P.J. & Boag, P.T. (1996). Cuckoldry and lack of parentage-dependent paternal care in yellow warblers: a cost–benefit approach. — Anim. Behav. 52:821-832.

1144 Song categories are social cues for habitat selection
Appendix
Figure A1. Map of study sites located throughout central and northern Illinois during 2014(lines indicate county boundaries), where circles indicate first-category song treatments, tri-angles second-category song treatments, and x-marks silent controls with no playback. Sitesample sizes by county were as follows: Champaign (n = 4), DeKalb (n = 2), DeWitt (n = 1),DuPage (n = 9), Fulton (n = 1), Iroquois (n = 1), Kane (n = 4), Kankakee (n = 3), LaSalle(n = 1), Lee (n = 1), Ogle (n = 1), Macon (n = 1), McHenry (n = 2), McLean (n = 1), Ver-milion (n = 3), Will (n = 3) and Winnebago (n = 6). The map was created using ArcGISversion 10.4 (ESRI, Redlands, CA, USA). Map source: Illinois Natural Resources GeospatialData Clearinghouse, Illinois State Geological Survey, https://clearinghouse.isgs.illinois.edu/data/reference/illinois-county-boundaries-polygons-and-lines (accessed 14 April 2017).