Embed Size (px)
Citation preview

Draft
Genome-wide analysis of the SBP-box gene family in
Chinese cabbage (Brassica rapa ssp. pekinensis)
Journal: Genome
Manuscript ID gen-2015-0074.R2
Manuscript Type: Article
Date Submitted by the Author: 10-Sep-2015
Complete List of Authors: Tan, Hua-Wei; Nanjing Agricultural University, State Key Laboratory of Crop Genetics and Germplasm Enhancement / Key Laboratory of Biology and Germplasm Enhancement of Horticultural Crops in East China, Ministry of Agriculture / College of Horticulture Song, Xiao-Ming; Nanjing Agricultural University, State Key Laboratory of Crop Genetics and Germplasm Enhancement / Key Laboratory of Biology
and Germplasm Enhancement of Horticultural Crops in East China, Ministry of Agriculture / College of Horticulture; North China University of Science and Technology, Center of Genomics and Computational Biology, College of Life Sciences Duan, Wei-Ke; Nanjing Agricultural University, State Key Laboratory of Crop Genetics and Germplasm Enhancement / Key Laboratory of Biology and Germplasm Enhancement of Horticultural Crops in East China, Ministry of Agriculture / College of Horticulture Wang, Yan; Nanjing Agricultural University, State Key Laboratory of Crop Genetics and Germplasm Enhancement / Key Laboratory of Biology and Germplasm Enhancement of Horticultural Crops in East China, Ministry of Agriculture / College of Horticulture
Hou, Xilin; Nanjing Agricultural University, State Key Laboratory of Crop Genetics and Germplasm Enhancement / Key Laboratory of Biology and Germplasm Enhancement of Horticultural Crops in East China, Ministry of Agriculture / College of Horticulture
Keyword: Genome-wide analysis, SBP-box gene family, expression profile, <i>Brassica rapa</i>
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
1
Genome-wide analysis of the SBP-box gene family in Chinese cabbage (Brassica rapa
ssp. pekinensis)
Hua-Wei Tan1, Xiao-Ming Song
1,2, Wei-Ke Duan
1, Yan Wang
1, Xi-Lin Hou
1*
1. State Key Laboratory of Crop Genetics and Germplasm Enhancement / Key Laboratory of Biology and Germplasm
Enhancement of Horticultural Crops in East China, Ministry of Agriculture / College of Horticulture, Nanjing
Agricultural University, Nanjing, 210095, China
2. Center of Genomics and Computational Biology, College of Life Sciences, North China University of Science and
Technology, Tangshan, 063000, China.
*Please address all correspondence to: Xi-Lin Hou ([email protected])
__________________
Dr. Xi-Lin Hou
Professor
State Key Laboratory of Crop Genetics and Germplasm Enhancement/Key Laboratory of Biology and Germplasm
Enhancement of Horticultural Crops in East China, Ministry of Agriculture
Nanjing Agricultural University, Nanjing, 210095, China
Tel: +86 25 84395332
Email: [email protected]
Page 1 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
2
Highlights
Twenty-nine SBP-box genes were identified in the Chinese cabbage genome.
Phylogenetic and evolutionary relationships of the SBP gene family were illustrated.
MiR156/157 members may regulate the coding regions or 3′-UTR regions of SBP-box genes.
The expression pattern of Chinese cabbage SBP genes was analyzed.
Abstract
The SQUAMOSA PROMOTER BINDING PROTEIN (SBP)-box gene family contains highly conserved plant-specific
transcription factors that play an important role in plant development, especially in flowering. Chinese cabbage (Brassica
rapa ssp. pekinensis) is a leafy vegetable grown worldwide, and is used as a model crop for research in genome duplication.
The present study aimed to characterize the SBP-box transcription factor genes in Chinese cabbage. Twenty-nine SBP-box
genes were identified in the Chinese cabbage genome and classified into six groups. We identified twenty-three orthologous
and five co-orthologous SBP-box gene pairs between Chines cabbage and Arabidopsis, and constructed an interaction
network among these genes. Sixteen SBP-box genes were found more abundant in flowers than in other tissues, suggesting
their involvement in flowering. We show that the MiR156/157 family members may regulate the coding regions or 3′-UTR
regions of Chinese cabbage SBP-box genes. As SBP-box genes were found to potentially participate in some plant
development pathways, quantitative real-time PCR analysis was performed and showed that Chinese cabbage SBP-box
genes were also sensitive to exogenous hormone methyl jasmonic acid and salicylic acid. The SBP-box genes have
undergone gene duplication and loss, evolving a more refined regulation for diverse stimulation in plant tissues. Our
comprehensive genome-wide analysis provides insights into the SBP-box gene family of Chinese cabbage.
Keywords
Brassica rapa; Genome-wide analysis; SBP-box gene family; expression profile
Abbreviation List
SBP, SQUAMOSA PROMOTER BINDING PROTEIN; SPL, SQUAMOSA PROMOTER BINDING PROTEIN; bp, base
pair(s); UTR, untranslated regions; HMMs, hidden Markov models; PCR, polymerase chain reaction; LF, least fractionated
blocks; MF1, the medium fractionated blocks; MF2, the most fractionated blocks; BRAD, Brassica rapa Database;
Page 2 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
3
1. Introduction
Transcription factors regulate plant development and stress responses by temporarily and spatially regulating the
transcription of their target genes. Transcription factors are usually classified into different families and subfamilies based
on their DNA-binding domain sequences, the evolutionary components of sequence-specific transcription factors that
mediate the specificity of its interaction (Charoensawan et al., 2010; Jin et al., 2014). A genome-wide analysis of plant
transcription factors was first performed in Arabidopsis thaliana soon after the completion of its genome sequencing in
2000 (Initiative, 2000). SQUAMOSA PROMOTER BINDING PROTEIN (SBP)-box genes encode a highly conserved
family of plant-specific transcription factors, which were first identified as nuclear proteins interacting within the promoter
region of the Antirrhinum majus floral meristem gene SQUAMOSA (Klein et al., 1996). SBP-box genes encode proteins
sharing a conserved DNA-binding domain of 79 amino acids. SBP proteins contain a zinc finger motif with two
zinc-binding sites (Cys-Cys-His-Cys and Cys-Cys-Cys-His) and most of them have a three-stranded antiparallel beta-sheet
(Yamasaki et al., 2004). SBP proteins play important roles in leaf development, vegetative phase change, flower and fruit
development, plant architecture, sporogenesis, gibberelic acid signaling and toxin response (Chen et al., 2010). In
Arabidopsis, AtSPL3 is involved in the floral transition, and is under the regulation of miR156 during the vegetative
development stage (Cardon et al., 1997). SBP-box genes are also responsible for defense against biotic and abiotic stresses
in grape (Hou et al., 2013). Given that SBP-box proteins play important roles in plant development, their further
characterization in other plant species is warranted.
In plants, miRNAs form a near-ubiquitous class of short (21–24 nt), endogenous single-strand RNA molecules that mainly
perform post-transcriptional gene regulation (Zhang et al., 2013). The SBP-box gene family exhibits different regulatory
patterns among different species: some members are only regulated by miR156 and others by miR156 and miR529 (Ling
and Zhang, 2012). Nineteen rice SPL (the SBP-box genes of rice) (OsSPL) genes and 12 rice miRNA156 (OsmiR156)
precursors were identified in the rice genome. Eleven of these OsSPL genes were putative targets of OsmiR156, which
indicated OsmiR156 and OsSPL target genes are involved in various developmental processes, especially flower
development in rice (Xie et al., 2006).
Chinese cabbage (Brassica rapa ssp. pekinensis) is a leafy vegetable grown worldwide. The genome of Chinese cabbage
was sequenced and assembled in 2011, and valuable genomics and bioinformatics information is available at BRAD
database (http://brassicadb.org/brad/) (Cheng et al., 2011; Wang et al., 2011). Genome-wide analyses have been performed
for transcription factors such as AP2 (Li et al., 2013; Liu et al., 2013; Song et al., 2013), GRAS (Song et al., 2014b),
HSF(Ma et al., 2014b; Song et al., 2014b), Aux/IAA (Mun et al., 2012), NAC (Liu et al., 2014; Ma et al., 2014a), WAKY
(Tang et al., 2014) and bHLH (Song et al., 2014a), providing a systematic view of post-transcriptional regulation in Chinese
Page 3 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
4
cabbage. SBP-box genes have been identified through whole-genome analysis in several plants, including Arabidopsis (Guo
et al., 2008; Yang et al., 2008), rice (Xie et al., 2006; Yang et al., 2008), grape (Hou et al., 2013) and castor bean (Zhang and
Ling, 2014). However, the specific biological and functional roles of each SBP-box transcription factor gene in Chinese
cabbage remain unknown. Here, we performed a genome-wide identification of SBP-box genes in the Chinese cabbage
genome, and then analyzed gene classification through an examination for exon-intron structure, gene phylogeny and
synteny, and their interaction with microRNAs.
2. Materials and methods
2.1. Identification, location and chemical characteristics of Chinese cabbage SBP-box genes
Characterization of sequence similarity within a conserved domain has usually been used to identify transcription factors.
Based on the sequence similarity, a large collection of transcription factor families have been collected in the Pfam protein
family database, and each of the families represented by multiple sequence alignments with hidden Markov models (HMMs)
(Finn et al., 2014). The HMMER3 software package has been recommended for searching transcription factors based on
HMM models (Eddy, 2011). In addition to search tools, several databases have been developed that focus on the
identification and annotation of plant transcription factors. The PlantTFDB (http://planttfdb.cbi.pku.edu.cn) is a
comprehensive resource for functional and evolutionary research of 58 plant transcription factor families. There are 129,288
transcription factors from 83 plant species deposited in this database, of which 67 species have whole-genome sequences,
providing detailed annotation for several lineages of green plants (Jin et al., 2014). Moreover, several databases contain
comprehensive data on transcription factors from a single species, such as the DATF database (Guo et al., 2005), DRTF
database (Gao et al., 2006), DPTF database (Zhu et al., 2007), TOBFAC database (Rushton et al., 2008) and SoyDB
database (Wang et al., 2010), providing solid support for research on Arabidopsis, rice, poplar, tobacco, grape and soybean,
respectively.
The genome, gene and protein sequences of Chinese cabbage were downloaded from the BRAD database (version 1.5).
PF03110.9 from the Pfam database was used as an SBP-domain search model in SBP-box gene searching against all 41019
Chinese cabbage genes using HMMER, with a cut-off E-value of 10-5
. To confirm the HMMER results, all 29 candidates
were checked against the proteomes in the SMART database (http://smart.embl-heidelberg.de/). The position of each
SBP-box gene on Chinese cabbage chromosomes or scaffolds was obtained according to information from BRAD database.
A Perl script was developed to mark 29 genes on the chromosome and construct a diagram. The boundaries of the
untranslated regions (UTRs) were obtained from an RNA-seq transcriptome research, and transformed into sequences using
Page 4 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
5
a Perl script. Protparam (http://web.expasy.org/protparam/) was employed in the physical and chemical characteristics
analysis of the SBP proteins. To estimate the divergence of duplicated Chinese cabbage SBP-box genes, MUSCLE was used
in protein sequence alignment of duplicated pairs of SBP proteins. Subsequently, synonymous rate (Ks), nonsynonymous
rate (Ka), and evolutionary constraint (Ka/Ks) between potential pairs of genes were calculated with KaKs_calculator tool.
2.2. Phylogenetic analyses construction
SBP-domain sequences for phylogenetic analyses were gathered from seven plant species (Table S1). Chlamydomonas
reinhardtii, Physcomitrella paten and Vitis vinifera SBP-box genes were gathered from a previous report (Hou et al., 2013).
A. thaliana, Populus trichocarpa and Oryza sativa SBP-box genes were downloaded from the DATF, DPTF and TIGR
databases, respectively (Table S2). Only the longest transcript was used if alternative splicing existed. For the phylogenetic
analysis, only the regions aligned to the SBP-domain were considered, and sequences consistent with a previous report
(Hou et al., 2013) were retained. ClustalX2 was used to generate the multiple sequence alignment of the SBP domains with
default settings according to previously report (Liu et al., 2014b). MEGA (version 6.06) was used to construct
maximum-parsimony phylogenetic trees with 2000 bootstrap replicates.
2.3. Gene structure and motif analysis
The Gene Structure Display Server tool (http://gsds1.cbi.pku.edu.cn/) was used to analyze the exon-intron structures.
MEME software (http://meme.nbcr.net/meme/) was used to search for motifs in all 29 SBP-box genes, with the motif length
set as at least 20 bp up to a maximum of 200 bp. The number of motifs were set to 16 and motifs were reported in this study.
2.4. Identification of orthologous and paralogous genes
OrthoMCL (version 2.0.3) was used to search for orthologous and paralogous genes in Chinese cabbage, Arabidopsis and
rice using the entire protein sequences of SBP. We used an E-value of 1e-5
for BLASTP and OrthoMCL analyses, gathered
the orthologous and paralogous relationships and displayed them using the Circos software (http://circos.ca/).
2.5. Identification miRNAs related to SBP-box genes
A collection of 157 Chinese cabbage miRNAs from the public miRBase database (version 21) were used to identify
SBP-box gene-related miRNAs. TargetFinder software (v1.6) was employed to predict the targets of miRNAs in Chinese
cabbage SBP-box genes, with the maximum mis-pairing set at 3.
2.6. SBP-box gene expression analysis in tissues
We used the Chiifu-401-42 RNA-seq transcriptions with the related fragments per kilobase per million measure (FPKM)
Page 5 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
6
values to study SBP-box genes expression patterns in six Chinese cabbage tissues from eight samples. According to
previous report, callus tissue was gathered via tissue culture; root, stem and leaf tissues were collected from seven-week old
plants, while root and stem were obtained from different batches; flower tissue was collected from blooming plants; silique
tissue was collected from 15-day plant after pollination (Tong et al., 2013). The SBP-box gene expression profile from each
sample was analyzed via the HemI program (http://hemi.biocuckoo.org/) with the average hierarchical clustering method.
2.7. Plant material and hormone treatment
The Chinese cabbage cultivar Chiifu-401-42 was used. Seeds were germinated and grown in pots containing a soil :
vermiculite mixture (2:1) in a controlled-environment growth chamber programmed for 14/10 h at 23/18°C for day/night,
and a relative humidity of 55–60% (Song et al., 2014b). At the five-leaf stage, hormone treatments were conducted by
spraying leaves with 100 µM salicylic acid (SA) or 50 µM methyl jasmonic acid (MeJA). The young leaf samples were
collected at 0 h, 6 h and 12 h after treatment, and then snap-frozen in liquid nitrogen before storage at -75°C for further
analysis.
2.8. SBP-box gene expression analysis by qPCR
An RNA kit (Tiangen, Beijing, China) was used to extract total RNA from isolated plant leaves, according to the
manufacturer’s instructions. The Prime Script RT reagent kit (Takara, Kyoto, Japan) reverse transcribed the RNAs into
cDNA. The Chinese cabbage actin gene (Bra028615) was used as an internal control to normalize the expression level of
the target genes among different samples. Primer 5.0 software was used to design specific primers according to the SBP-box
gene sequences (Table S3). Three biological and three technical replicates were performed. Each reaction was performed in
a 20-µL reaction mixture containing diluted cDNA sample as the template, SYBR Pre-mix Ex Taq (2×) (Takara) and
gene-specific primers. Quantitative real-time PCR (qPCR) was performed using a CFX96 Single color Real-Time PCR
Detection System (BioRad, Hercules, CA, USA) with the following cycling profile: 94°C for 30 s; followed by 40 cycles at
94°C for 10 s, 58°C for 30 s; and then a melting curve (61 cycles at 65°C for 10 s) was generated to check the specificity of
the amplification. The comparative Ct value method was employed to analyze the relative gene expression level. The RNA
level was expressed relative to the actin gene expression level as 2−∆∆CT
, according to previous analyses (Pfaffl, 2001). The
SBP-box gene expression cluster from each stress treatment was analyzed via the HemI software.
3. Results
3.1. Identification of SBP-box genes in Chinese cabbage
Bioinformatics analysis identified 29 SBP-box transcription factors in the Chinese cabbage genome among 41019 coding
Page 6 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
7
genes. All 29 candidates were confirmed and retained after checking against the SMART database. All of the Chinese
cabbage SBP-box genes were well conserved, since the E-value of the full sequence aligned with the SBP-box domain
varied from 4e-40
to 5.1e-19
. We named these genes “BrSBP” with a serial number, sorted by E-value (Tables 1 and S4). The
average length of the coding sequence (CDS) of the SBP-box genes (~1224 bp) is longer than that of all Chinese cabbage
genes (~1173 bp), while the average whole gene length (~1702 bp) is shorter than all Chinese cabbage genes (~2019 bp). In
other worlds, the SBP-box genes have longer exon and smaller introns. To determine the closest SBP-box genes in
Arabidopsis, we searched against the TAIR database (version 10) and found the most similar gene pair for each Chinese
cabbage SBP-box gene. Twenty-six Chinese cabbage SBP-box genes could be classified to LF, MF1 and MF2 subgenomes
while three genes were unclear. The present B. rapa genome (version 1.5) is annotated only with CDSs of genes; however,
RNA-seq transcriptome data provided the 5′-UTR sequence of 12,879 genes and the 3′-UTR sequence of 12485 genes. The
5′- and 3′-UTRs of eight SBP-box genes were extracted and are shown in Table S5. The average length of the 5′-UTRs was
326 bp, much longer than in all detected 5′-UTRs in the transcriptome sample. This suggested that transcription factor genes
with binding domains seem to have longer 5′-UTRs. However, the average length of the 3′-UTRs was about 129 bp, shorter
than the average of all estimated 3′-UTRs. To be more specific, the 5’-UTR of BrSBP17 was 599 bp, and this gene also had
the longest coding region among the SBP-box genes. BrSBP17 may have similar functions to AtSPL14, playing a role in
sensitivity to FB1 and the development of normal plant architecture.
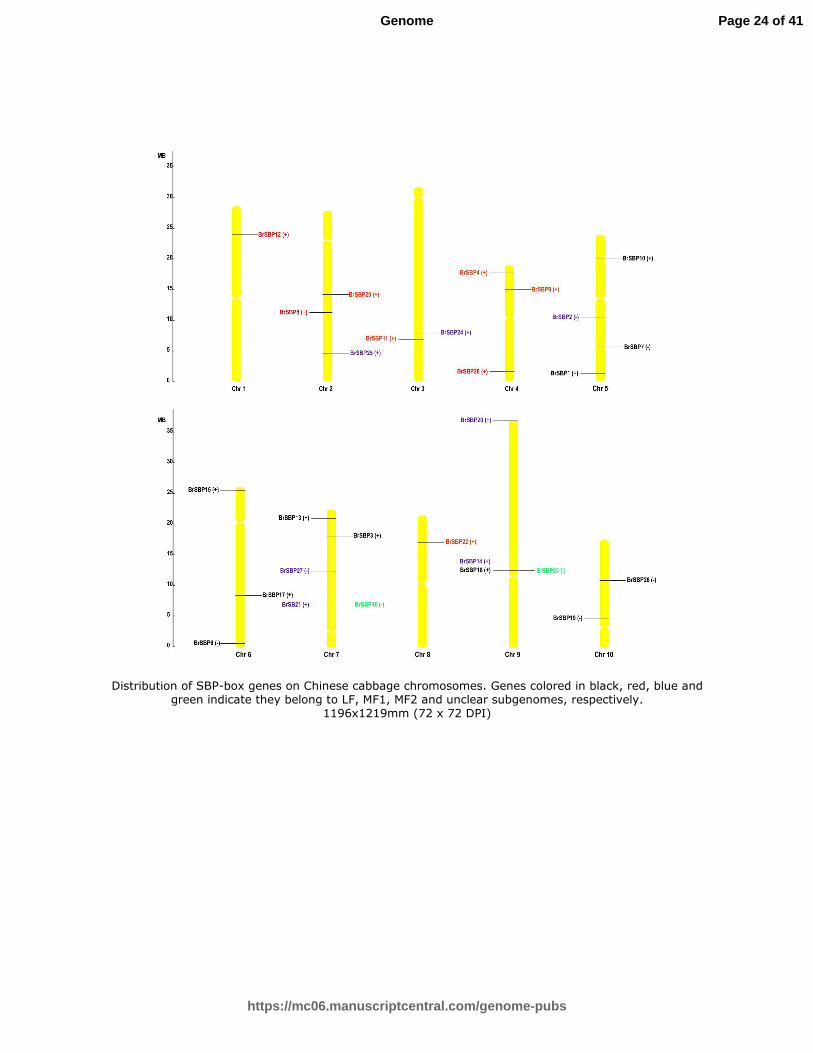
According to the most recently assembled Chinese cabbage genome resources, the 29 Chinese cabbage SBP-box genes were
distributed unevenly on the 10 chromosomes. Five Chinese cabbage SBP-box genes were located on chromosome 7; four on
chromosomes 4, 5 and 9; three on chromosomes 2 and 6; two on chromosomes 3 and 10; one each on chromosomes 1 and 8;
and one on Scaffold000403. The positions of the 28 genes located on the chromosomes are shown in Fig. 1, providing a
visual insight into SBP-box gene distribution.
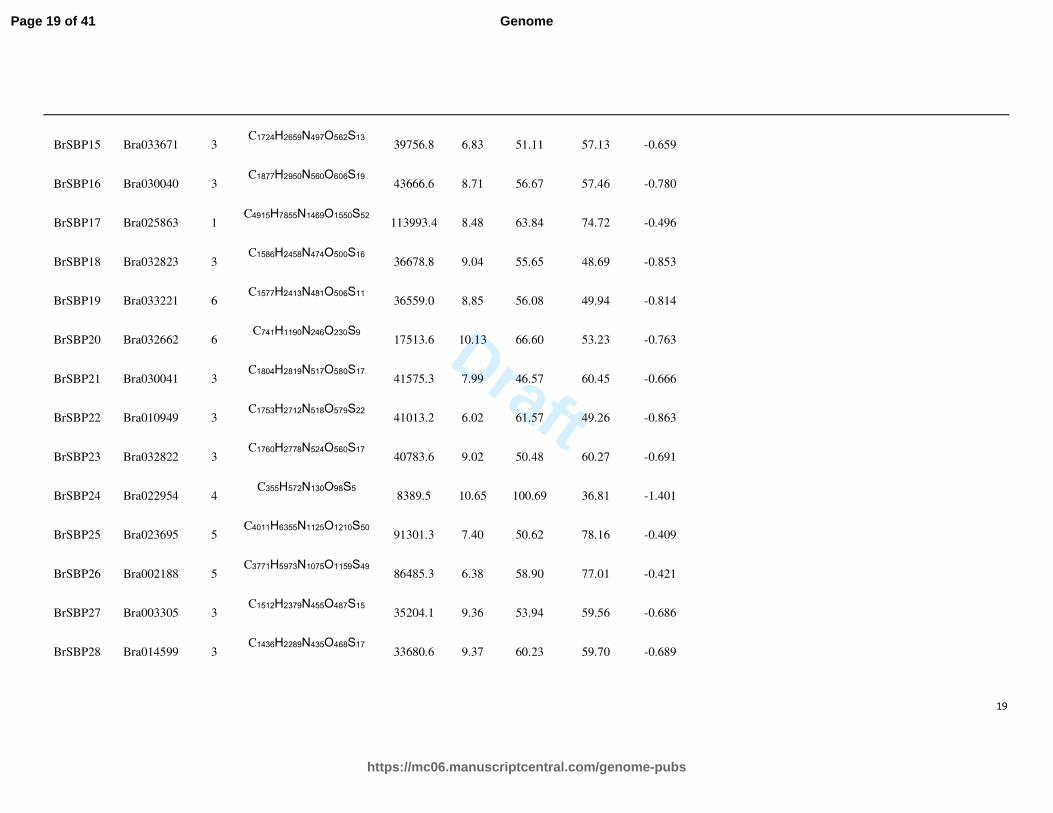
Physical and chemical characteristics of SBP proteins were analyzed and the result was summarized in Table 2. The
molecular formula, molecular weight, pI (theoretical isoelectric point), instability index, aliphatic index and GRAVY (grand
average of hydropathicity) were characterized for each SBP protein. Most proteins were alkaline, which is opposite to the
GRAS or HSF families. The instability index of all the proteins ranged from 46.57 to 111.23, revealing that all of them are
unstable molecules. The GRAVY scores were all less than 0, indicating that the SBP proteins are hydrophilic (Table 2).
3.2. Phylogenetic relationship and evolutionary divergence of SBP-domain family genes
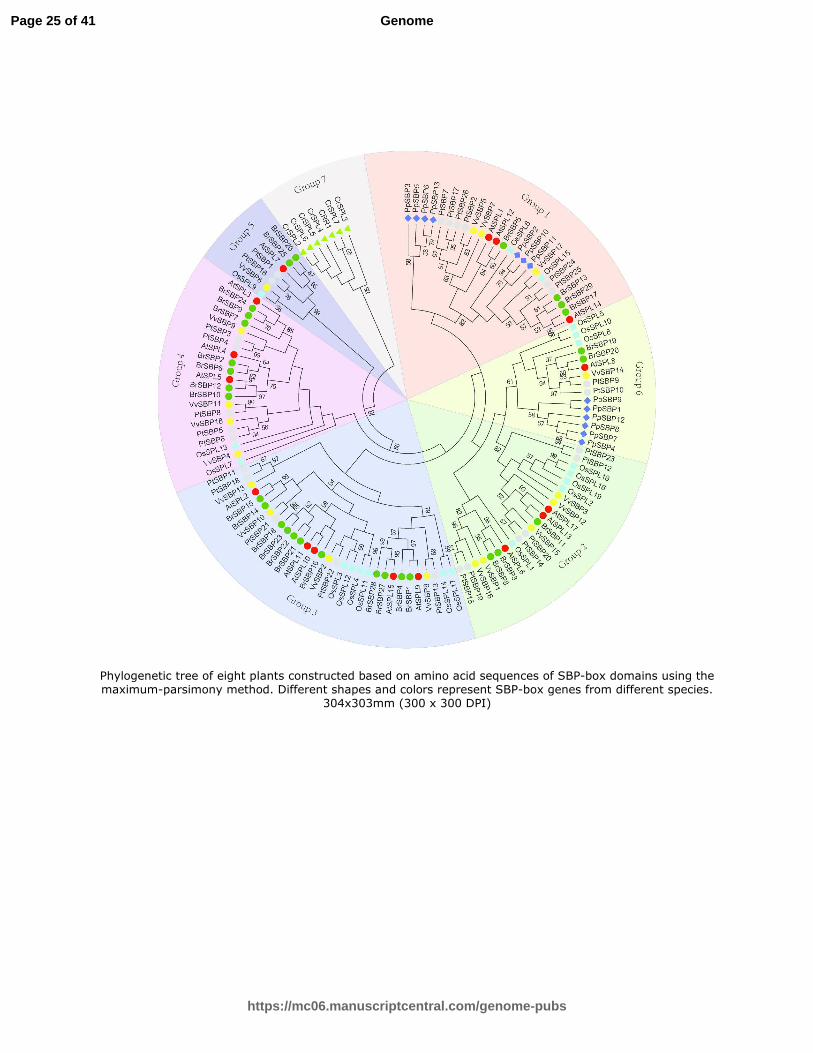
For the phylogenetic analysis, we selected seven well-studied and representative plant species, including green alga (C.
reinhardtii), moss (Physcomitrella patens), four eudicots (Arabidopsis, Chinese cabbage, grape and poplar), as well as a
Page 7 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
8
monocotyledonous angiosperm, rice. To obtain a more precise phylogenetic tree of plant SBP proteins, we trimmed three
amino acids and used the more conserved SBP domains (76 amino acids). Certain SBP proteins were not included in the
analysis because their conserved domains contained too few amino acids to perform the phylogenetic analysis. All of the
Chinese cabbage SBP proteins were retained for further phylogenetic analysis (Table S2).
According to the phylogenetic analysis, the 128 SBP-box genes could be classified into seven groups (Fig. 2). Similar to
other higher plants, none of the Chinese cabbage SBP-box genes were present in Group 7, which only contains seven alga
genes. Six and seven SBP-box proteins from moss were classified into Group 6 and Group 1, respectively, suggesting that
Group 6 and Group 1 are secondary ancient groups along with Group 7.
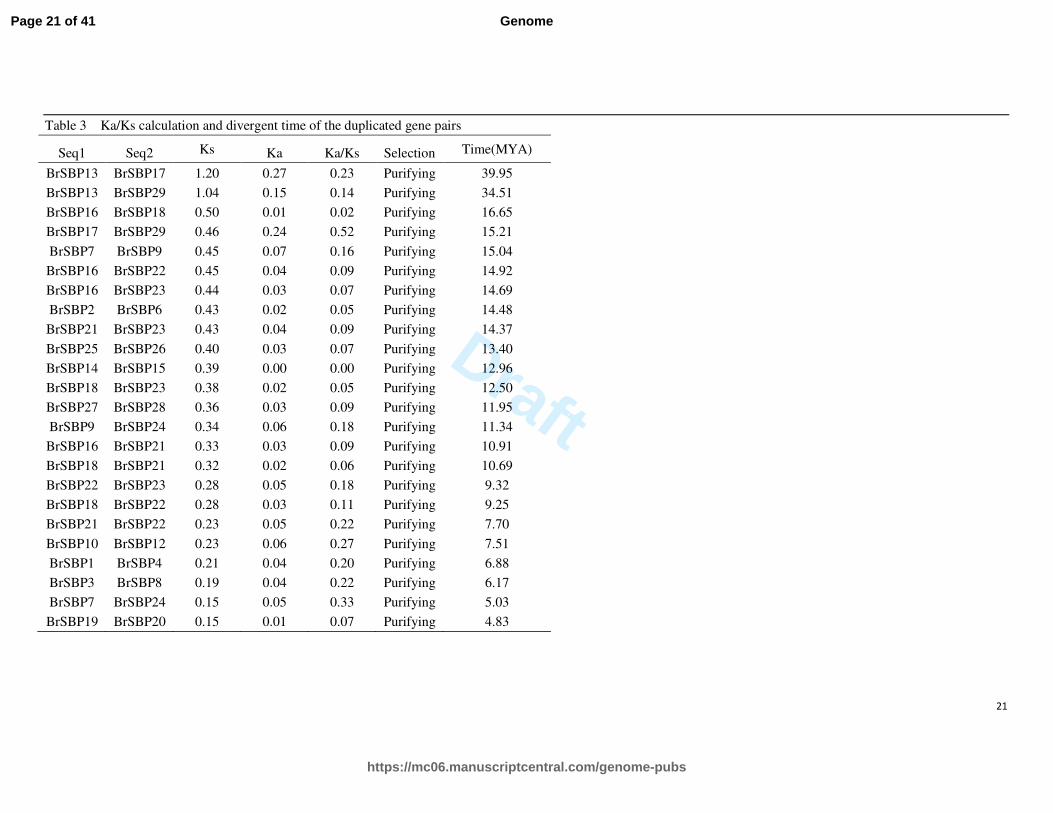
The selection types and divergence timings of duplicated genes were estimated by calculating the Ks, Ka and Ka/Ks of gene
pairs. Among 193 gene pairs, only 2 were underwent neutral selection, 52 and 139 gene pairs underwent positive and
purifying selection, respectively. The divergent time is varied from 174.82 MYA (Million Years Ago) to 4.83 MYA.
Interesting, the Ka/Ks values of all 24 duplicated gene pairs were less than 1, indicates that they underwent purifying
selection. Besides, the divergent time is much closer than other gene pairs, with a range of 39.95 MYA to 4.83 MYA (Table
3).
3.3. Gene structure and motif analysis
To compare the 29 Chinese cabbage SBP-box genes, their exon-intron structures were predicted (Fig. 3). In general, genes
in the same group have similar exon and intron numbers, and even intron phases. Group 1 and Group 5 genes contain nine
or ten exons. The similarity of splicing patterns in Group 1 and Group 5 showed the close relationship between these two
groups. Group 3 was the biggest branch, containing genes with three or four exons. Group 4 and Group 6 have more
conserved splicing patterns of their two and three exons, respectively. Interestingly, according to the phylogenic analysis,
both BrSBP3 and BrSBP8 are related to AtSPL6 (three exons), while BrSBP3 has only one exon and BrSBP8 has three
exons. To validate the sequence information of BrSBP3, we performed a genomic DNA PCR and transcriptional RNA PCR.
The same sequence obtained from the DNA and RNA confirmed that BrSBP3 is a single-exon gene. We hypothesized that
an intron loss event occurred after the divergence of Arabidopsis and Chinese cabbage.
We searched for motifs to analyze the conserved features of Chinese cabbage SBP proteins using MEME (Fig. 4). The
majority of SBP proteins contains Motif 1, indicating that they have both Cys-Cys-Cys-His and Cys-Cys-His-Cys type zinc
finger, and the former seems less conserved than the latter. BrSBP24, BrSBP27, BrSBP28 and BrSBP29 contain Motif 2,
showing that they only have Cys-Cys-His-Cys type zinc finger. Motif 8 was only found in Group 3, indicating that the
Page 8 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
9
proteins in Group 3 might share a unique function. Moreover, some motifs only exists one unique group but were shared by
all the members within the group. This is the case of Motif 6, Motif 9, Motif 14 in Group 1 and Motif 10 in Group 3.
3.4. Identification of orthologous and paralogous SBP-box genes in plants
Comparative analysis was performed to identify the orthologous and paralogous gene pairs. We identified 23 orthologous
and four coorthologous SBP-box gene pairs among 21 Chinese cabbage genes and 16 Arabidopsis genes using OrthoMCL
with default settings. Moreover, nine orthologous and four coorthologous gene pairs among Chinese cabbage and rice genes
were identified, providing comparative genomics information of SBP-box genes between eudicots and monocots. Nine
paralogous pairs were found in Chinese cabbage, more than four pairs in Arabidopsis and six in rice. The relationships of
orthologous, coorthologous and paralogous SBP-box genes among the three species were visualized using the Circos
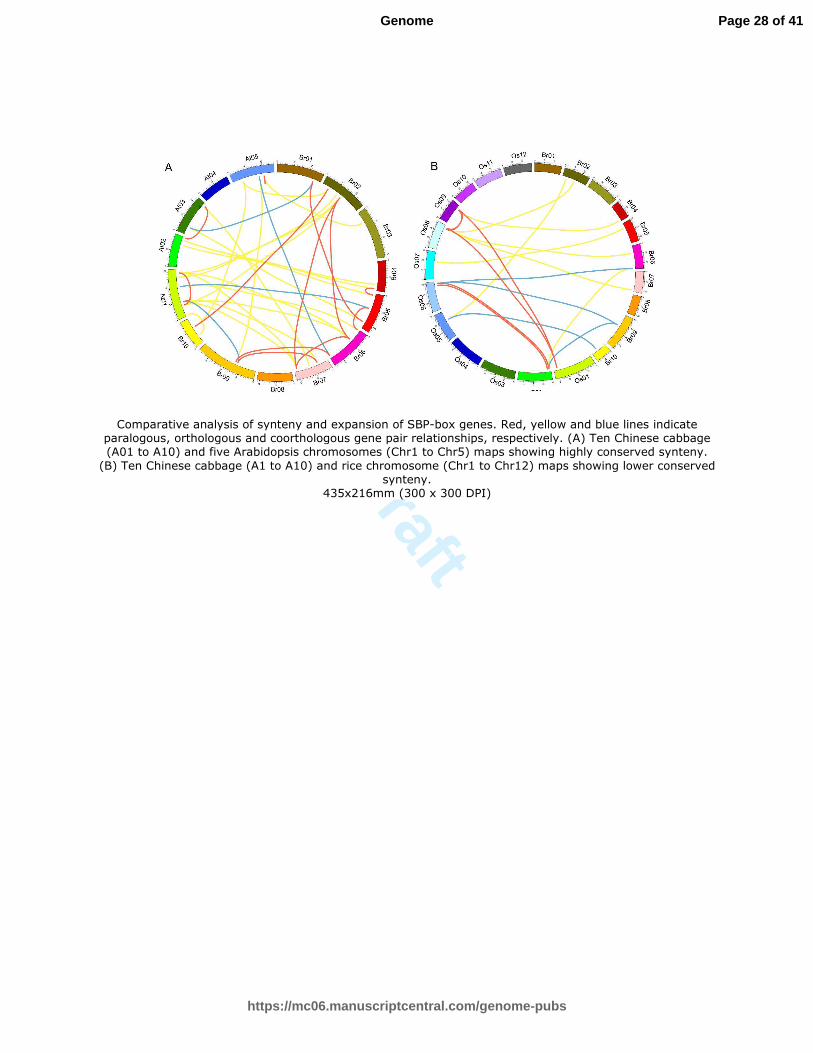
program (Table S7, Fig. 5).
An interaction network of Chinese cabbage SBP-box genes was constructed to improve our understanding of the
genome-wide regulation network. The Arabidopsis Interactions Viewer
(http://bar.utoronto.ca/interactions/cgi-bin/arabidopsis_interactions_viewer.cgi) is a database of 70944 predicted and 36306
confirmed Arabidopsis interacting proteins, providing deep insight and high confidence interactions among Brassicaceae
plants. We replaced Arabidopsis genes with corresponding orthologous and coorthologous Chinese cabbage genes after
constructing an Arabidopsis gene interaction network (Fig. 6). The PCC (Pearson correlation coefficients) of 22 gene pairs
were less than zero, whereas that of two genes were more than zero, which revealed SBP proteins mainly have a negative
interaction with other proteins in Chinese cabbage. Twenty-eight gene pairs were not calculated, indicating that many of
their regulation patterns remain unknown.
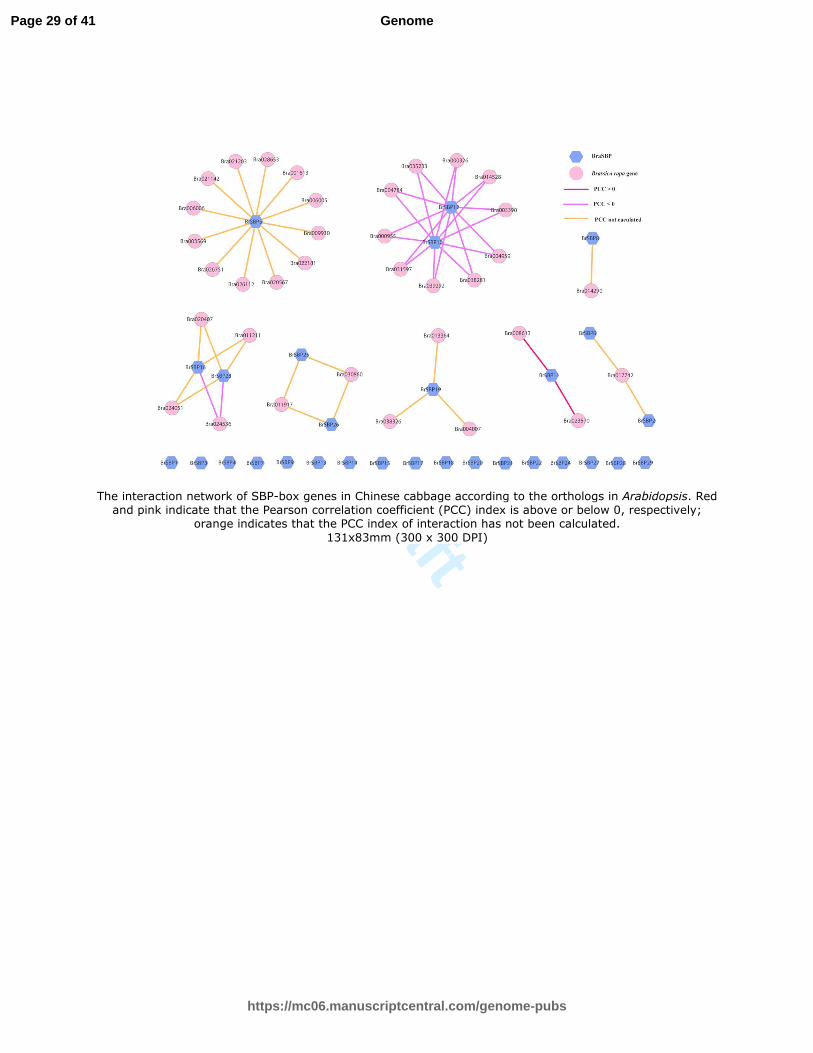
The interaction network showed that the number of proteins regulated by each SBP-box gene was significantly different.
For instance, BrSBP5, BrSBP10 and BrSBP12 potentially interacted with at least 10 proteins, suggesting their importance in
transcriptional-level regulation. BrSBP11 has a positive interaction with Bra008613 and Bra023570, both of which encode a
KANADI protein (KAN) that regulates organ polarity. It has been suggested that BrSBP11 is required for the development
of carpel and the outer integument of the ovule (Ha et al., 2007). Both BrSBP16 and BrSBP23 had opposite interactions
with Bra024536, which participates in a circadian clock-controlled flowering pathway under long days. Moreover, these
two SBP-box genes also have unclear interactions with three other genes. The gene pair of BrSBP12 and BrSBP10 also has
opposite interactions with 10 other Chinese cabbage genes that are orthologous genes with AT4G11330. These 12 genes
may play an import role in the protein phosphorylation process, and have protein kinase activity, protein serine/threonine
kinase activity, ATP binding, MAP kinase activity, transferase activity and transferring phosphorus-containing groups.
Page 9 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
10
3.5. Identification of SBP-box genes regulated by miRNAs
The UTR sequences of Chinese cabbage SBP-box genes were obtained through previously reported transcriptome (Table
S5). To study transcriptional regulation of SBP-box genes, we searched 29 Chinese cabbage SBP-box genes with all 157
available relevant miRNAs. We identified only the miR156/157 family and all positive 5p mature members could
potentially regulate the coding regions or 3′-UTRs of Chinese cabbage SBP-box genes (Table S8). In grape, miR156/157
sequences were complementary with nine SBP-box genes in their coding regions. Three members of Group 4, except for
VvSBP4, contain an miR156/157 target site in their 3′-UTR. In Arabidopsis, AtSPL3, AtSPL4 and AtSPL5 in Group 4 also
showed similar complementarity with miR156/157 members (Hou et al., 2013). All SBP-box genes from Group 2 and
Group 3, except for BrSBP18, were potential targets of miR156/157 members, and the target site is located in the last exon.
The loss of a potential exon of BrSBP18 leads to loss of regulation interaction with miRNAs. Interestingly, BrSBP7 from
Group 4 contains a target site in the 3′-UTR like AtSPL3, while other members have no target site in their largely different
3′-UTRs.
3.6. Expression patterns of Chinese cabbage SBP-box genes in different tissues
We searched expression values for each SBP-box gene using RNA-seq data. The FPKM method was performed to eliminate
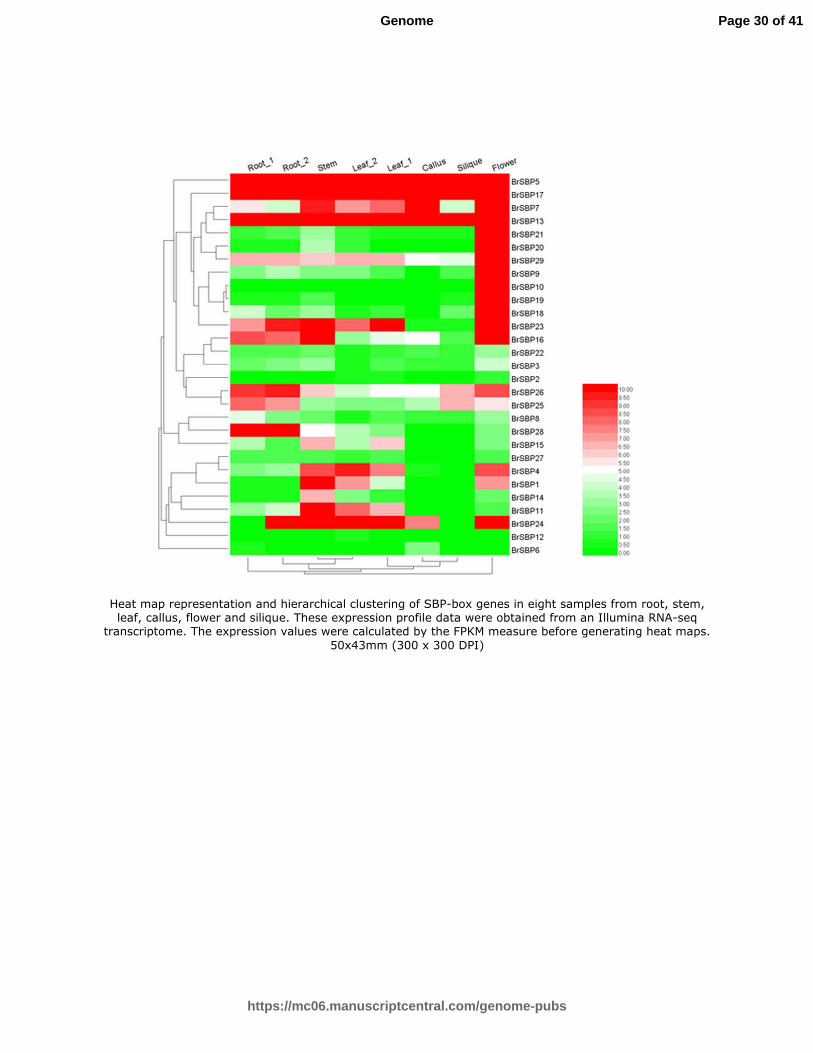
errors of sequencing data size. According to the FPKM values, all the BrSBP genes were expressed in at least one tissue,
and 28 (96.6%) of these genes were expressed at relatively high levels (FPKM>1) in at least one tissue, including root, stem,
leaf, callus, flower and silique (Table S8, Fig. 7). Sixteen BrSBP genes expressed more abundantly in flowers than in any
other tissues, especially BrSBP9, BrSBP10 and BrSBP19, which suggested their necessity in the flowering process.
SBP-box genes in Group 1 had similar expression quantities in six tissues, suggesting that these four genes have similar
transcription patterns. Similar results were obtained for Group 3 and Group 5. The expression profile in callus is similar to
that in silique tissue, and is of more difference with stem and root, and seems big difference with flower.
3.7. Expression patterns of Chinese cabbage SBP-box genes treated with exogenous plant hormones
As SBP-box genes were found to potentially participate in some plant development pathways, quantitative real-time PCR
analysis was performed and showed that Chinese cabbage SBP-box genes are also sensitive to exogenous hormone methyl
jasmonic acid and salicylic acid (Hou et al., 2013). To increase our understanding of the expression profiles of the SBP-box
genes in Chinese cabbage under exogenous plant hormones, we performed qPCR with primer pairs within non-conserved
regions for each Chinese cabbage SBP-box gene, to ensure the specificity of PCR amplification..
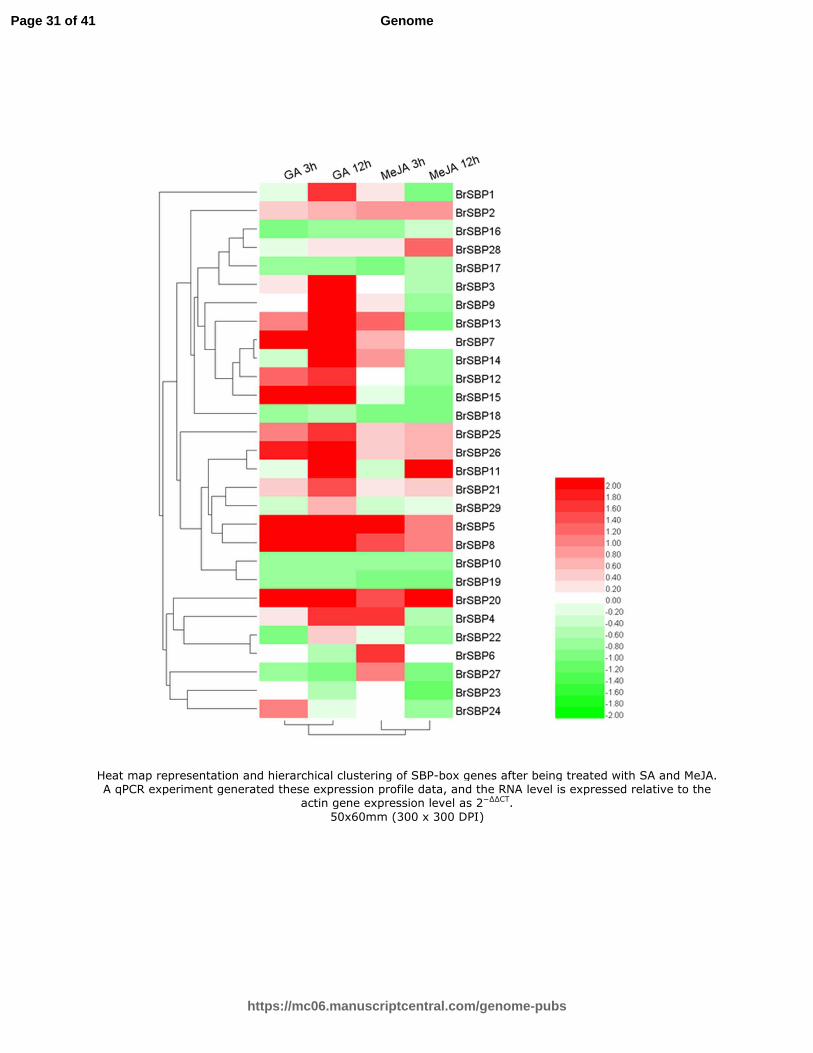
Under SA treatment, the expressions of 16 genes increased and those of 13 decreased after 3 h, and the expressions of 20
Page 10 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
11
genes increased and nine decreased after 12 h compared with control. The BrSBP8 gene increased over 20 times compared
with the control, suggesting that it is a response acceptor. BrSBP1 decreased over 10 times after 3 h treatment compared
with the control, and then increased to almost 1.5 times higher than the control.
Under MeJA treatment, the expressions of 20 genes increased and those of 9 decreased after 3 h, while the expression of 11
genes increased and 18 decreased after 12 h, compared with the control. The expression of the SBP-box genes under the
exogenous hormone varied considerably among each group (Table S10, Fig. 8).
4. Discussion
In the present study, we analyzed the SBP-box transcription factor family in Chinese cabbage and seven other well-studied
plants, and investigated their expression profiles during Chinese cabbage developmental stages.
Our results confirmed that during the vegetative period, the expressions of two members from Group 3 that are similar to
AtSPL9 and AtSPL15 increased progressively and reached their highest levels at the heading stage, revealing that SBP-box
genes play an important role in heading time (Wang et al., 2014). Besides, a number of previous reports also found that
SBP-box genes play an important role in sporogenesis, gibberelic acid signaling and toxin response. A recent study on grape
SBP-box genes supported that some grape SBP-box genes responded to the pathogen as well as hormone (Hou et al., 2013).
In this study, Chinese cabbage SBP-box genes supported its widely different responses to exogenous hormone MeJA and
SA; however, the mechanism and pathway require further investigation.
A. thaliana has been thoroughly studied as a model plant, and deep insights have been gained into its metabolism and
development. As another plant in the Brassicaceae family, Chinese cabbage is similar to Arabidopsis phenotypically and
genetically (Wang et al., 2011). As expected, we found there is a high similarity between the Chinese cabbage SBP-box
proteins with those from Arabidopsis, suggesting that SBP-box genes from Arabidopsis and Chinese cabbage are closely
related thus share more similar expression profiles compared with other plants. Every SBP-box in Chinese cabbage has a
highly similar counterpart in Arabidopsis; thus, during evolution, no Chinese cabbage species-specific SBP-box genes have
evolved or emerged. Moreover, previous report showed that a lot of Chinese cabbage has three subgenomes originating
from the Arabidopsis genome, called least fractionated blocks (LF), the medium fractionated blocks (MF1) and the most
fractionated blocks (MF2). On the genome-wide scale, the LF subgenome retains 70% of the genes found in A. thaliana,
while the MF1 and MF2 subgenomes retain substantially lower proportions of retained genes (46% and 36%) (Wang et al.,
2011; Cheng et al., 2014). We search each Chinese cabbage SBP-box gene in the BRAD database, and 26 of them can be
Page 11 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
12
classified as subgenomes while three are unclear. Compared with 16 Arabidopsis SBP-box genes, the Chinese cabbage
retains 69% (11 genes), 50% (eight genes) and 44% (seven genes) of the genes in LF, MF1 and MF2 subgenomes; the actual
ratio would be higher when considering the other three unclassified SBP-box genes. SBP-box genes in the LF genome
showed consistency in their statistical data compared with all genes, while the ratio of SBP-box genes from MF1 and MF2
was higher. We observed that in three and eight homologous groups, one Arabidopsis protein has two or three homologous
genes in Chinese cabbage, respectively. Interestingly, in Group 1, only BrSBP5 was identified to be homologous with
AtSPL1 and AtSPL12. Similarly, only BrSBP11 was deduced to be homologous with AtSPL13 and AtSPL17. We infer that
some of the Chinese cabbage SBP-box genes underwent gene loss after genome triple duplication. The ratio of gene loss is
slightly lower than for other genes, indicating their importance in Chinese cabbage plant development.
The genome locations of the 29 Chinese cabbage SBP-box genes are far from each other, except BrSBP18 and BrSBP23.
The SBP-box domains of these two genes are very similar, but their gene structures are different. In the subgroup of
BrSBP18, BrSBP21, BrSBP22 and BrSBP23, BrSBP23 has four exons, BrSBP21 and BrSBP22 have three introns, and
BrSBP18 has three exons, two introns and a downstream sequence, according to the B. rapa genome. BrSBP18, BrSBP21
and BrSBP22 belong to LF, MF2, MF1 subgenomes, respectively, while BrSBP23 is still unknown. We hypothesized that
BrSBP18 and BrSBP23 may have undergone a tandem duplication, and that BrSBP23 belongs to the LF subgenome like
BrSBP18. Given the gene structure, the BrSBP23 retains an ancient gene structure, and BrSBP18 has evolved after tandem
duplication.
Changes in gene structures also cause changes in expression profiles. In the SBP-box gene group including BrSBP18,
BrSBP21 and BrSBP22, the tissue expression value of BrSBP18 was similar to BrSBP21, both of which were significantly
higher in flower and callus than BrSBP22. After treatment with exogenous hormone, the expression profiles became more
complicated, and BrSBP18 and BrSBP22 seemed more similar than BrSBP21. The diversification and complexity of
SBP-box genes expression under multiple circumstances suggested that the genes that have been retained after gene
duplication and loss have evolved to provide a more refined regulation of diverse stimuli in plant tissues.
In conclusion, we have performed a comprehensive analysis of SBP-box transcription factors in Chinese cabbage. The
physical and chemical characteristics of SBP proteins provide useful information for protein extraction. The results revealed
the SBP proteins participate in plant tissue development and are sensitive to exogenous hormones, suggesting that the SBP
proteins play an important role in plant development, signaling pathway regulation and defense responses under stress
conditions via interactions with other proteins. Our bioinformatics analysis provides new insights in the evolution and
diversity of SBP-box transcription factors, thus can serve as a basic resource for developing new cultivars via manipulation
Page 12 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
13
of SBP-box genes.
Acknowledgments
This work was supported by the National High Technology Research and Development Program of China (863 Program,
No. 2012AA100101), China Agriculture Research System CARS-25-A-12, Jiangsu Science and Technology Support
Program (BE2013429) and a Project Funded by the Priority Academic Program Development of Jiansu Higher Education
Institutions. We thank Dr. Dapeng Zhang from Sustainable Perennial Crops Laboratory, USDA-ARS, Beltsville Agricultural
Research Center, Beltsville, MD, USA for his advice and revision.
References
Cardon, G.H., Hohmann, S., Nettesheim, K., Saedler, H., and Huijser, P. 1997. Functional analysis of the Arabidopsis thaliana
SBP-box gene SPL3: a novel gene involved in the floral transition. Plant Journal, 12(2): 367-377. doi:
10.1046/j.1365-313X.1997.12020367.x.
Charoensawan, V., Wilson, D., and Teichmann, S.A. 2010. Genomic repertoires of DNA-binding transcription factors across the
tree of life. Nucleic Acids Research, 38(21): 7364-7377. doi: 10.1093/Nar/Gkq617.
Chen, X., Zhang, Z., Liu, D., Zhang, K., Li, A., and Mao, L. 2010. SQUAMOSA promoter-binding protein-like transcription factors:
star players for plant growth and development. Journal of Integrative Plant Biology, 52(11): 946-951. doi:
10.1111/j.1744-7909.2010.00987.x.
Cheng, F., Liu, S.Y., Wu, J., Fang, L., Sun, S.L., Liu, B., Li, P.X., Hua, W., and Wang, X.W. 2011. BRAD, the genetics and genomics
database for Brassica plants. BMC Plant Biology, 11(136). doi: 10.1186/1471-2229-11-136.
Cheng, F., Wu, J., Wang, X.W. 2014. Genome triplication drove the diversification of Brassica plants. Horticulture Research, 1:
14024. doi: 10.1038/hortres.2014.24.
Eddy, S.R. 2011. Accelerated profile HMM searches. PLoS Computational Biology, 7(10): e1002195. doi:
10.1371/journal.pcbi.1002195.
Finn, R.D., Bateman, A., Clements, J., Coggill, P., Eberhardt, R.Y., Eddy, S.R., Heger, A., Hetherington, K., Holm, L., Mistry, J., et al.
2014. Pfam: the protein families database. Nucleic Acids Research, 42(Database issue): D222-D230. doi: 10.1093/nar/gkt1223.
Gao, G., Zhong, Y., Guo, A., Zhu, Q., Tang, W., Zheng, W., Gu, X., Wei, L., and Luo, J. 2006. DRTF: a database of rice transcription
factors. Bioinformatics, 22(10): 1286-1287. doi: 10.1093/bioinformatics/btl107.
Guo, A., He, K., Liu, D., Bai, S., Gu, X., Wei, L., and Luo, J. 2005. DATF: a database of Arabidopsis transcription factors.
Bioinformatics, 21(10): 2568-2569. doi: 10.1093/bioinformatics/bti334.
Guo, A.Y., Zhu, Q.H., Gu, X.C., Ge, S., Yang, J., and Luo, J.C. 2008. Genome-wide identification and evolutionary analysis of the
plant specific SBP-box transcription factor family. Gene, 418(1-2): 1-8. doi: 10.1016/j.gene.2008.03.016.
Page 13 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
14
Ha, C.M., Jun, J.H., Nam, H.G., and Fletcher, J.C. 2007. BLADE-ON-PETIOLE1 and 2 control Arabidopsis lateral organ fate through
regulation of LOB domain and adaxial-abaxial polarity genes. The Plant Cell, 19(6): 1809-1825. doi:
http://dx.doi.org/10.1105/tpc.107051938.
Hou, H.M., Li, J., Gao, M., Singer, S.D., Wang, H., Mao, L.Y., Fei, Z.J., and Wang, X.P. 2013. Genomic organization, phylogenetic
comparison and differential expression of the SBP-Box family genes in grape. PLoS ONE, 8(3): e59358. doi:
10.1371/journal.pone.0059358.
Initiative, A.G. 2000. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature, 408(6814): 796-815.
doi: 10.1038/35048692.
Jin, J.P., Zhang, H., Kong, L., Gao, G., and Luo, J.C. 2014. PlantTFDB 3.0: a portal for the functional and evolutionary study of plant
transcription factors. Nucleic Acids Research, 42(Database Issue): D1182-D1187. doi: 10.1093/Nar/Gkt1016.
Klein, J., Saedler, H., and Huijser, P. 1996. A new family of DNA binding proteins includes putative transcriptional regulators of the
Antirrhinum majus floral meristem identity gene SQUAMOSA. Molecular and General Genetics, 250(1): 7-16. doi:
10.1007/Bf02191820.
Li, M.Y., Wang, F., Jiang, Q., Li, R., Ma, J., and Xiong, A.X. 2013. Genome-wide analysis of the distribution of AP2/ERF transcription
factors reveals duplication and elucidates their potential function in Chinese cabbage (Brassica rapa ssp. pekinensis). Plant
Molecular Biology Reporter, 31(4): 1002-1011. doi: 10.1007/s11105-013-0570-7.
Ling, L.Z., and Zhang, S.D. 2012. Exploring the evolutionary differences of SBP-box genes targeted by miR156 and miR529 in
plants. Genetica, 140(7-9): 317-324. doi: 10.1007/s10709-012-9684-3.
Liu, T.K., Song, X.M., K., D.W., Huang, Z.N., Liu, G.F., LI, Y., and Hou, X.L. 2014a. Genome-wide analysis and expression patterns of
NAC transcription factor family under different develepment stages and abiotic stresses in Chinese cabbage. Plant Molecular
Biology Reporter, 32(5): 1041-1056. doi: 10.1007/s11105-014-0712-6.
Liu, T.K., Qian Y., Duan W.K., Ren J., Hou, X.L. and Li Y. 2014b. BcRISP1, isolated from non-heading Chinese cabbage, decreases the
seed set of transgenic Arabidopsis. Horticulture Research, 1: 14062. doi: 10.1038/hortres.2014.62.
Liu, Z.N., Kong, L.J., Zhang, M., Lv, Y.X., Liu, Y.P., Zou, M.H., Lu, G., Cao, J.S., and Yu, X.L. 2013. Genome-wide identification,
phylogeny, evolution and expression patterns of AP2/ERF genes and cytokinin response factors in Brassica rapa ssp. pekinensis.
PLoS ONE, 8(12): e83444. doi: 10.1371/journal.pone.0083444.
Ma, J., Wang, F., Li, M.Y., Jiang, Q., Tan, G.F., and Xiong, A.S. 2014a. Genome wide analysis of the NAC transcription factor family
in Chinese cabbage to elucidate responses to temperature stress. Scientia Horticulturae, 165: 82-90. doi:
10.1016/j.scienta.2013.11.005.
Ma, J., Xu, Z.S., Wang, F., Tan, G.F., Li, M.Y., and Xiong, A.X. 2014b. Genome-wide analysis of HSF family transcription factors and
their responses to abiotic stresses in two Chinese cabbage varieties. Acta Physiologiae Plantarum, 36(2): 513-523. doi:
10.1007/s11738-013-1432-5.
Mun, J.H., Yu, H.J., Shin, J.Y., Oh, M., Hwang, H.J., and Chung, H. 2012. Auxin response factor gene family in Brassica rapa:
genomic organization, divergence, expression, and evolution. Molecular Genetics and Genomics, 287(10): 765-784. doi:
10.1007/s00438-012-0718-4.
Pfaffl, M.W. 2001. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Research, 29(9): e45.
doi: 10.1093/nar/29.9.e45.
Rushton, P.J., Bokowiec, M.T., Laudeman, T.W., Brannock, J.F., Chen, X., and Timko, M.P. 2008. TOBFAC: the database of tobacco
Page 14 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
15
transcription factors. BMC Bioinformatics, 9 :53. doi: 10.1186/1471-2105-9-53.
Song, X.M., Huang, Z.N., Duan, W.K., Ren, J., Liu, T.K., Li, Y., and Hou, X.L. 2014a. Genome-wide analysis of the bHLH transcription
factor family in Chinese cabbage (Brassica rapa ssp. pekinensis). Molecular Genetics and Genomics, 289(1): 77-91. doi:
10.1007/s00438-013-0791-3.
Song, X.M., Li, Y., and Hou, X.L. 2013. Genome-wide analysis of the AP2/ERF transcription factor superfamily in Chinese cabbage
(Brassica rapa ssp. pekinensis). BMC Genomics, 14: 573. doi: 10.1186/1471-2164-14-573.
Song, X.M., Liu, T.K., Duan, W.K., Ma, Q.H., Ren, J., Wang, Z., Li, Y., and Hou, X.L. 2014b. Genome-wide analysis of the GRAS gene
family in Chinese cabbage (Brassica rapa ssp. pekinensis). Genomics, 103(1): 135-146. doi: 10.1016/j.ygeno.2013.12.004.
Tang, J., Wang, F., Hou, X.L., Wang, Z., and Huang, Z.N. 2014. Genome-wide fractionation and identification of WRKY transcription
factors in Chinese cabbage (Brassica rapa ssp. pekinensis) reveals collinearity and their expression patterns under abiotic and
biotic stresses. Plant Molecular Biology Reporter, 32(4): 781-795. doi: 10.1007/s11105-013-0672-2.
Tong, C.B., Wang, X.W., Yu, J.Y., Wu, J., Li, W.S., Huang, J.Y., Dong, C.H., Hua, W., and Liu, S.Y. 2013. Comprehensive analysis of
RNA-seq data reveals the complexity of the transcriptome in Brassica rapa. BMC Genomics, 14: 689. doi:
10.1186/1471-2164-14-689.
Wang, X.W., Wang, H.Z., Wang, J., Sun, R.F., Wu, J., Liu, S.Y., Bai, Y.Q., Mun, J.H., Bancroft, I., Cheng, F., et al. 2011. The genome of
the mesopolyploid crop species Brassica rapa. Nature Genetics, 43(10): 1035-U1157. doi: 10.1038/Ng.919.
Wang, Y.L., Wu, F.J., Bai, J.J., and He, Y.K. 2014. BrpSPL9 (Brassica rapa ssp. pekinensis SPL9) controls the earliness of heading time
in Chinese cabbage. Plant Biotechnology Journal, 12(3): 312-321. doi: 10.1111/Pbi.12138.
Wang, Z., Libault, M., Joshi, T., Valliyodan, B., Nguyen, H.T., Xu, D., Stacey, G., and Cheng, J. 2010. SoyDB: a knowledge database of
soybean transcription factors. BMC Plant Biology, 10: 14. doi: 10.1186/1471-2229-10-14.
Xie, K., Wu, C., and Xiong, L. 2006. Genomic organization, differential expression, and interaction of SQUAMOSA
promoter-binding-like transcription factors and microRNA156 in rice. Plant Physiology, 142(1): 280-293. doi:
10.1104/pp.106.084475.
Yamasaki, K., Kigawa, T., Inoue, M., Tateno, M., Yamasaki, T., Yabuki, T., Aoki, M., Seki, E., Matsuda, T., Nunokawa, E., et al. 2004.
A novel zinc-binding motif revealed by solution structures of DNA-binding domains of Arabidopsis SBP-family transcription
factors. Journal of Molecular Biology, 337(1): 49-63. doi: 10.1016/j.jmb.2004.01.015.
Yang, Z.F., Wang, X.F., Gu, S.L., Hu, Z.Q., Xu, H., and Xu, C.W. 2008. Comparative study of SBP-box gene family in Arabidopsis and
rice. Gene, 407(1-2): 1-11. doi: 10.1016/j.gene.2007.02.034.
Zhang, R., Marshall, D., Bryan, G.J., and Hornyik, C. 2013. Identification and characterization of miRNA transcriptome in potato by
high-throughput sequencing. PLoS ONE, 8(2): e57233. doi: 10.1371/journal.pone.0057233.
Zhang, S.D., and Ling, L.Z. 2014. Genome-wide identification and evolutionary analysis of the SBP-box gene family in castor bean.
PLoS ONE, 9(1): e86688. doi: 10.1371/journal.pone.0086688.
Zhu, Q.H., Guo, A.Y., Gao, G., Zhong, Y.F., Xu, M., Huang, M.R., and Luo, J.C. 2007. DPTF: a database of poplar transcription factors.
Bioinformatics, 23(10): 1307-1308. doi: 10.1093/bioinformatics/btm113.
Page 15 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome
Draft
16
Tables
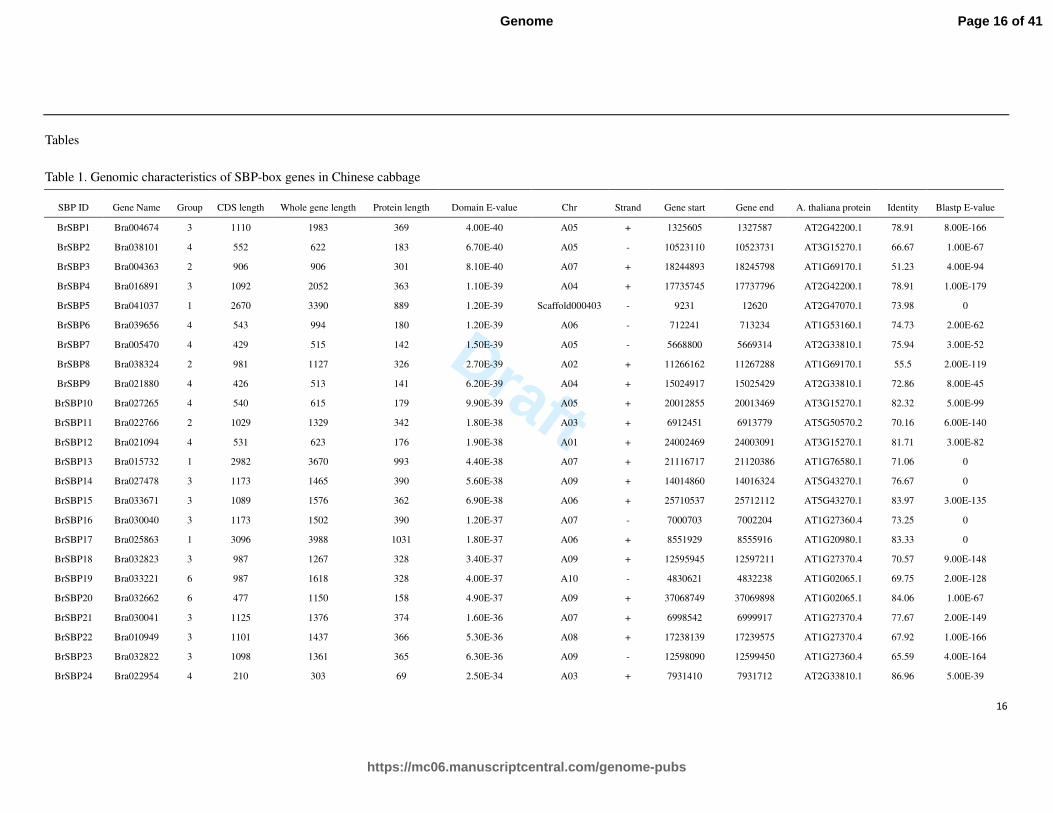
Table 1. Genomic characteristics of SBP-box genes in Chinese cabbage
SBP ID Gene Name Group CDS length Whole gene length Protein length Domain E-value Chr Strand Gene start Gene end A. thaliana protein Identity Blastp E-value
BrSBP1 Bra004674 3 1110 1983 369 4.00E-40 A05 + 1325605 1327587 AT2G42200.1 78.91 8.00E-166
BrSBP2 Bra038101 4 552 622 183 6.70E-40 A05 - 10523110 10523731 AT3G15270.1 66.67 1.00E-67
BrSBP3 Bra004363 2 906 906 301 8.10E-40 A07 + 18244893 18245798 AT1G69170.1 51.23 4.00E-94
BrSBP4 Bra016891 3 1092 2052 363 1.10E-39 A04 + 17735745 17737796 AT2G42200.1 78.91 1.00E-179
BrSBP5 Bra041037 1 2670 3390 889 1.20E-39 Scaffold000403 - 9231 12620 AT2G47070.1 73.98 0
BrSBP6 Bra039656 4 543 994 180 1.20E-39 A06 - 712241 713234 AT1G53160.1 74.73 2.00E-62
BrSBP7 Bra005470 4 429 515 142 1.50E-39 A05 - 5668800 5669314 AT2G33810.1 75.94 3.00E-52
BrSBP8 Bra038324 2 981 1127 326 2.70E-39 A02 + 11266162 11267288 AT1G69170.1 55.5 2.00E-119
BrSBP9 Bra021880 4 426 513 141 6.20E-39 A04 + 15024917 15025429 AT2G33810.1 72.86 8.00E-45
BrSBP10 Bra027265 4 540 615 179 9.90E-39 A05 + 20012855 20013469 AT3G15270.1 82.32 5.00E-99
BrSBP11 Bra022766 2 1029 1329 342 1.80E-38 A03 + 6912451 6913779 AT5G50570.2 70.16 6.00E-140
BrSBP12 Bra021094 4 531 623 176 1.90E-38 A01 + 24002469 24003091 AT3G15270.1 81.71 3.00E-82
BrSBP13 Bra015732 1 2982 3670 993 4.40E-38 A07 + 21116717 21120386 AT1G76580.1 71.06 0
BrSBP14 Bra027478 3 1173 1465 390 5.60E-38 A09 + 14014860 14016324 AT5G43270.1 76.67 0
BrSBP15 Bra033671 3 1089 1576 362 6.90E-38 A06 + 25710537 25712112 AT5G43270.1 83.97 3.00E-135
BrSBP16 Bra030040 3 1173 1502 390 1.20E-37 A07 - 7000703 7002204 AT1G27360.4 73.25 0
BrSBP17 Bra025863 1 3096 3988 1031 1.80E-37 A06 + 8551929 8555916 AT1G20980.1 83.33 0
BrSBP18 Bra032823 3 987 1267 328 3.40E-37 A09 + 12595945 12597211 AT1G27370.4 70.57 9.00E-148
BrSBP19 Bra033221 6 987 1618 328 4.00E-37 A10 - 4830621 4832238 AT1G02065.1 69.75 2.00E-128
BrSBP20 Bra032662 6 477 1150 158 4.90E-37 A09 + 37068749 37069898 AT1G02065.1 84.06 1.00E-67
BrSBP21 Bra030041 3 1125 1376 374 1.60E-36 A07 + 6998542 6999917 AT1G27370.4 77.67 2.00E-149
BrSBP22 Bra010949 3 1101 1437 366 5.30E-36 A08 + 17238139 17239575 AT1G27370.4 67.92 1.00E-166
BrSBP23 Bra032822 3 1098 1361 365 6.30E-36 A09 - 12598090 12599450 AT1G27360.4 65.59 4.00E-164
BrSBP24 Bra022954 4 210 303 69 2.50E-34 A03 + 7931410 7931712 AT2G33810.1 86.96 5.00E-39
Page 16 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome
Draft
17
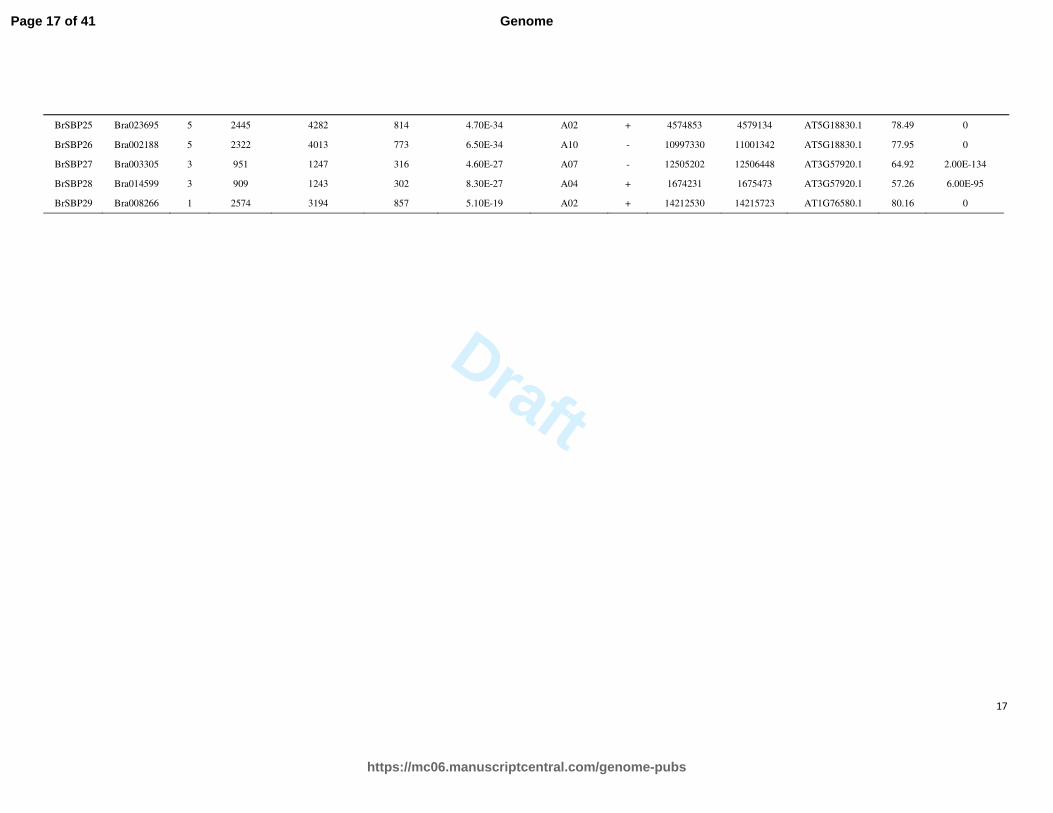
BrSBP25 Bra023695 5 2445 4282 814 4.70E-34 A02 + 4574853 4579134 AT5G18830.1 78.49 0
BrSBP26 Bra002188 5 2322 4013 773 6.50E-34 A10 - 10997330 11001342 AT5G18830.1 77.95 0
BrSBP27 Bra003305 3 951 1247 316 4.60E-27 A07 - 12505202 12506448 AT3G57920.1 64.92 2.00E-134
BrSBP28 Bra014599 3 909 1243 302 8.30E-27 A04 + 1674231 1675473 AT3G57920.1 57.26 6.00E-95
BrSBP29 Bra008266 1 2574 3194 857 5.10E-19 A02 + 14212530 14215723 AT1G76580.1 80.16 0
Page 17 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome
Draft
18
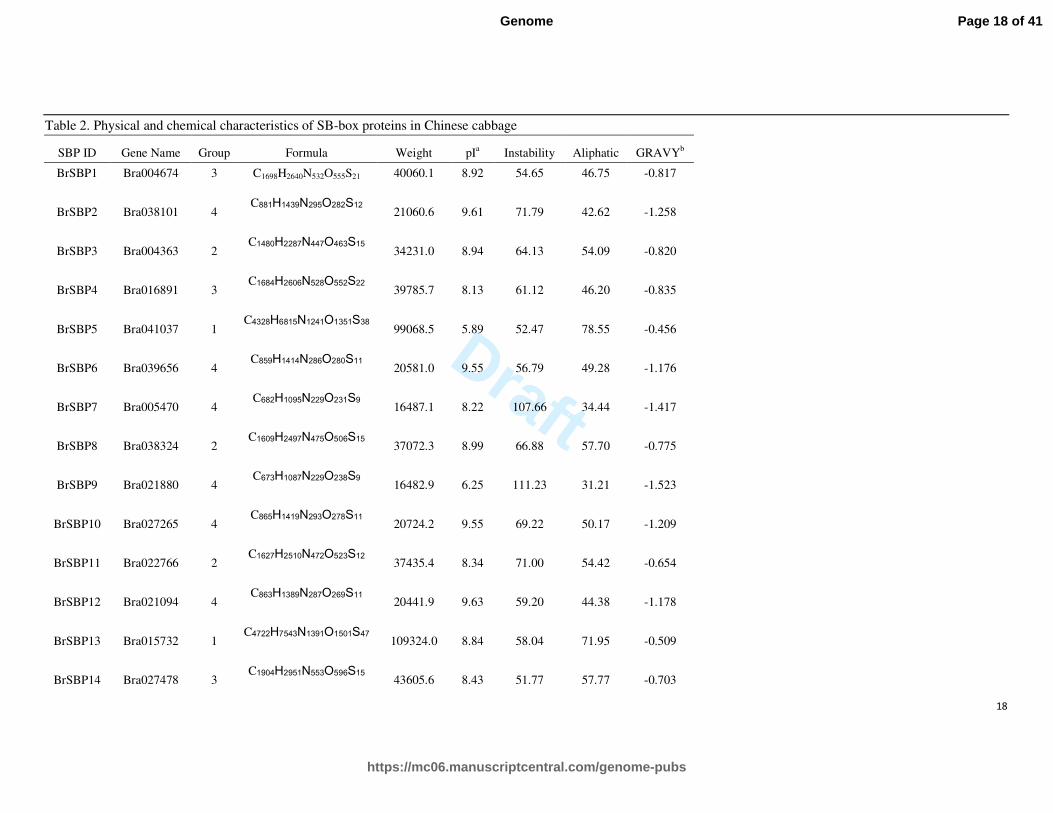
Table 2. Physical and chemical characteristics of SB-box proteins in Chinese cabbage
SBP ID Gene Name Group Formula Weight pIa Instability Aliphatic GRAVY
b
BrSBP1 Bra004674 3 C1698H2640N532O555S21 40060.1 8.92 54.65 46.75 -0.817
BrSBP2 Bra038101 4 C881H1439N295O282S12
21060.6 9.61 71.79 42.62 -1.258
BrSBP3 Bra004363 2 C1480H2287N447O463S15
34231.0 8.94 64.13 54.09 -0.820
BrSBP4 Bra016891 3 C1684H2606N528O552S22
39785.7 8.13 61.12 46.20 -0.835
BrSBP5 Bra041037 1 C4328H6815N1241O1351S38
99068.5 5.89 52.47 78.55 -0.456
BrSBP6 Bra039656 4 C859H1414N286O280S11
20581.0 9.55 56.79 49.28 -1.176
BrSBP7 Bra005470 4 C682H1095N229O231S9
16487.1 8.22 107.66 34.44 -1.417
BrSBP8 Bra038324 2 C1609H2497N475O506S15
37072.3 8.99 66.88 57.70 -0.775
BrSBP9 Bra021880 4 C673H1087N229O238S9
16482.9 6.25 111.23 31.21 -1.523
BrSBP10 Bra027265 4 C865H1419N293O278S11
20724.2 9.55 69.22 50.17 -1.209
BrSBP11 Bra022766 2 C1627H2510N472O523S12
37435.4 8.34 71.00 54.42 -0.654
BrSBP12 Bra021094 4 C863H1389N287O269S11
20441.9 9.63 59.20 44.38 -1.178
BrSBP13 Bra015732 1 C4722H7543N1391O1501S47
109324.0 8.84 58.04 71.95 -0.509
BrSBP14 Bra027478 3 C1904H2951N553O596S15
43605.6 8.43 51.77 57.77 -0.703
Page 18 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome
Draft
19
BrSBP15 Bra033671 3 C1724H2659N497O562S13
39756.8 6.83 51.11 57.13 -0.659
BrSBP16 Bra030040 3 C1877H2950N560O606S19
43666.6 8.71 56.67 57.46 -0.780
BrSBP17 Bra025863 1 C4915H7855N1469O1550S52
113993.4 8.48 63.84 74.72 -0.496
BrSBP18 Bra032823 3 C1586H2458N474O500S16
36678.8 9.04 55.65 48.69 -0.853
BrSBP19 Bra033221 6 C1577H2413N481O506S11
36559.0 8.85 56.08 49.94 -0.814
BrSBP20 Bra032662 6 C741H1190N246O230S9
17513.6 10.13 66.60 53.23 -0.763
BrSBP21 Bra030041 3 C1804H2819N517O580S17
41575.3 7.99 46.57 60.45 -0.666
BrSBP22 Bra010949 3 C1753H2712N518O579S22
41013.2 6.02 61.57 49.26 -0.863
BrSBP23 Bra032822 3 C1760H2778N524O560S17
40783.6 9.02 50.48 60.27 -0.691
BrSBP24 Bra022954 4 C355H572N130O98S5
8389.5 10.65 100.69 36.81 -1.401
BrSBP25 Bra023695 5 C4011H6355N1125O1210S50
91301.3 7.40 50.62 78.16 -0.409
BrSBP26 Bra002188 5 C3771H5973N1075O1159S49
86485.3 6.38 58.90 77.01 -0.421
BrSBP27 Bra003305 3 C1512H2379N455O487S15
35204.1 9.36 53.94 59.56 -0.686
BrSBP28 Bra014599 3 C1436H2289N435O468S17
33680.6 9.37 60.23 59.70 -0.689
Page 19 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
20
BrSBP29 Bra008266 1 C4102H6538N1194O1292S41
94568.7 8.38 54.21 76.23 -0.408
apI, theoretical isoelectric point
bGRAVY, grand average of hydropathicity
Page 20 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome
Draft
21
Table 3 Ka/Ks calculation and divergent time of the duplicated gene pairs
Seq1 Seq2 Ks Ka Ka/Ks Selection Time(MYA)
BrSBP13 BrSBP17 1.20 0.27 0.23 Purifying 39.95
BrSBP13 BrSBP29 1.04 0.15 0.14 Purifying 34.51
BrSBP16 BrSBP18 0.50 0.01 0.02 Purifying 16.65
BrSBP17 BrSBP29 0.46 0.24 0.52 Purifying 15.21
BrSBP7 BrSBP9 0.45 0.07 0.16 Purifying 15.04
BrSBP16 BrSBP22 0.45 0.04 0.09 Purifying 14.92
BrSBP16 BrSBP23 0.44 0.03 0.07 Purifying 14.69
BrSBP2 BrSBP6 0.43 0.02 0.05 Purifying 14.48
BrSBP21 BrSBP23 0.43 0.04 0.09 Purifying 14.37
BrSBP25 BrSBP26 0.40 0.03 0.07 Purifying 13.40
BrSBP14 BrSBP15 0.39 0.00 0.00 Purifying 12.96
BrSBP18 BrSBP23 0.38 0.02 0.05 Purifying 12.50
BrSBP27 BrSBP28 0.36 0.03 0.09 Purifying 11.95
BrSBP9 BrSBP24 0.34 0.06 0.18 Purifying 11.34
BrSBP16 BrSBP21 0.33 0.03 0.09 Purifying 10.91
BrSBP18 BrSBP21 0.32 0.02 0.06 Purifying 10.69
BrSBP22 BrSBP23 0.28 0.05 0.18 Purifying 9.32
BrSBP18 BrSBP22 0.28 0.03 0.11 Purifying 9.25
BrSBP21 BrSBP22 0.23 0.05 0.22 Purifying 7.70
BrSBP10 BrSBP12 0.23 0.06 0.27 Purifying 7.51
BrSBP1 BrSBP4 0.21 0.04 0.20 Purifying 6.88
BrSBP3 BrSBP8 0.19 0.04 0.22 Purifying 6.17
BrSBP7 BrSBP24 0.15 0.05 0.33 Purifying 5.03
BrSBP19 BrSBP20 0.15 0.01 0.07 Purifying 4.83
Page 21 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
22
Figure Legends
Fig. 1 Distribution of SBP-box genes on Chinese cabbage chromosomes. Genes colored in black, red, blue and green indicate they belong to LF, MF1, MF2 and unclear subgenomes,
respectively
Fig. 2 Phylogenetic tree of eight plants constructed based on amino acid sequences of SBP-box domains using the maximum-parsimony method. Different shapes and colors represent
SBP-box genes from different species
Fig. 3 Gene structure of Chinese cabbage SBP-box genes. Green boxes indicate the exon regions and black and blue lines indicate introns and downstream regions, respectively. The
lengths of the boxes and lines are scaled based on the length of the genes
Fig. 4 Distribution of conserved motifs of SBP-box genes in Chinese cabbage. (A) The phylogenetic tree and motifs located on each gene with relative combined P-values. (B) Amino
acid sequences of each motif
Fig. 5 Comparative analysis of synteny and expansion of SBP-box genes. Red, yellow and blue lines indicate paralogous, orthologous and coorthologous gene pair relationships,
respectively. (A) Ten Chinese cabbage (A01 to A10) and five Arabidopsis chromosomes (Chr1 to Chr5) maps showing highly conserved synteny. (B) Ten Chinese cabbage (A1 to A10)
Page 22 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
23
and rice chromosome (Chr1 to Chr12) maps showing lower conserved synteny
Fig. 6 The interaction network of SBP-box genes in Chinese cabbage according to the orthologs in Arabidopsis. Red and pink indicate that the Pearson correlation coefficient (PCC)
index is above or below 0, respectively; orange indicates that the PCC index of interaction has not been calculated
Fig. 7 Heat map representation and hierarchical clustering of SBP-box genes in eight samples from root, stem, leaf, callus, flower and silique. These expression profile data were
obtained from an Illumina RNA-seq transcriptome. The expression values were calculated by the FPKM measure before generating heat maps
Fig. 8 Heat map representation and hierarchical clustering of SBP-box genes after being treated with SA and MeJA. A qPCR experiment generated these expression profile data, and
the RNA level is expressed relative to the actin gene expression level as 2−∆∆CT
Page 23 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome
Draft
Distribution of SBP-box genes on Chinese cabbage chromosomes. Genes colored in black, red, blue and green indicate they belong to LF, MF1, MF2 and unclear subgenomes, respectively.
1196x1219mm (72 x 72 DPI)
Page 24 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome
Draft
Phylogenetic tree of eight plants constructed based on amino acid sequences of SBP-box domains using the maximum-parsimony method. Different shapes and colors represent SBP-box genes from different species.
304x303mm (300 x 300 DPI)
Page 25 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
Gene structure of Chinese cabbage SBP-box genes. Green boxes indicate the exon regions and black and blue lines indicate introns and downstream regions, respectively. The lengths of the boxes and lines are
scaled based on the length of the genes.
Page 26 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
Distribution of conserved motifs of SBP-box genes in Chinese cabbage. (A) The phylogenetic tree and motifs located on each gene with relative combined P-values. (B) Amino acid sequences of each motif.
271x509mm (300 x 300 DPI)
Page 27 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome
Draft
Comparative analysis of synteny and expansion of SBP-box genes. Red, yellow and blue lines indicate paralogous, orthologous and coorthologous gene pair relationships, respectively. (A) Ten Chinese cabbage (A01 to A10) and five Arabidopsis chromosomes (Chr1 to Chr5) maps showing highly conserved synteny.
(B) Ten Chinese cabbage (A1 to A10) and rice chromosome (Chr1 to Chr12) maps showing lower conserved synteny.
435x216mm (300 x 300 DPI)
Page 28 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome
Draft
The interaction network of SBP-box genes in Chinese cabbage according to the orthologs in Arabidopsis. Red and pink indicate that the Pearson correlation coefficient (PCC) index is above or below 0, respectively;
orange indicates that the PCC index of interaction has not been calculated.
131x83mm (300 x 300 DPI)
Page 29 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome
Draft
Heat map representation and hierarchical clustering of SBP-box genes in eight samples from root, stem, leaf, callus, flower and silique. These expression profile data were obtained from an Illumina RNA-seq
transcriptome. The expression values were calculated by the FPKM measure before generating heat maps.
50x43mm (300 x 300 DPI)
Page 30 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome
Draft
Heat map representation and hierarchical clustering of SBP-box genes after being treated with SA and MeJA. A qPCR experiment generated these expression profile data, and the RNA level is expressed relative to the
actin gene expression level as 2−∆∆CT.
50x60mm (300 x 300 DPI)
Page 31 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
Type Gene 1 Gene 2 Index
paralog Arabidopsis|AT1G20980 Arabidopsis|AT1G76580 1
paralog Arabidopsis|AT1G27360 Arabidopsis|AT1G27370 1
paralog Arabidopsis|AT2G47070 Arabidopsis|AT3G60030 1
paralog Arabidopsis|AT5G50570 Arabidopsis|AT5G50670 1
paralog rapa|Bra002188 rapa|Bra023695 108
paralog rapa|Bra004674 rapa|Bra016891 108
paralog rapa|Bra008266 rapa|Bra015732 108
paralog rapa|Bra008266 rapa|Bra025863 108
paralog rapa|Bra015732 rapa|Bra025863 108
paralog rapa|Bra021094 rapa|Bra027265 0.646
paralog rapa|Bra027478 rapa|Bra033671 108
paralog rapa|Bra030040 rapa|Bra032822 108
paralog rapa|Bra038101 rapa|Bra039656 0.599
paralog rice|LOC_Os01g69830 rice|LOC_Os08g41940 0.466
paralog rice|LOC_Os01g69830 rice|LOC_Os09g32944 0.405
paralog rice|LOC_Os02g04680 rice|LOC_Os06g49010 1.261
paralog rice|LOC_Os02g07780 rice|LOC_Os06g45310 0.529
paralog rice|LOC_Os08g39890 rice|LOC_Os09g31438 1.098
paralog rice|LOC_Os08g41940 rice|LOC_Os09g32944 175
ortholog Arabidopsis|AT1G02065 rapa|Bra033221 0.835
ortholog Arabidopsis|AT1G20980 rapa|Bra008266 118
ortholog Arabidopsis|AT1G20980 rapa|Bra015732 118
ortholog Arabidopsis|AT1G20980 rapa|Bra025863 118
ortholog Arabidopsis|AT1G27360 rapa|Bra030040 118
ortholog Arabidopsis|AT1G27370 rapa|Bra030040 118
ortholog Arabidopsis|AT1G53160 rapa|Bra039656 0.524
ortholog Arabidopsis|AT1G69170 rapa|Bra038324 0.858
ortholog Arabidopsis|AT1G76580 rapa|Bra008266 118
ortholog Arabidopsis|AT1G76580 rapa|Bra015732 118
ortholog Arabidopsis|AT1G76580 rapa|Bra025863 118
ortholog Arabidopsis|AT2G33810 rapa|Bra021880 0.371
ortholog Arabidopsis|AT2G42200 rapa|Bra004674 118
ortholog Arabidopsis|AT2G42200 rapa|Bra016891 118
ortholog Arabidopsis|AT2G47070 rapa|Bra041037 118
ortholog Arabidopsis|AT3G15270 rapa|Bra027265 0.659
ortholog Arabidopsis|AT3G57920 rapa|Bra003305 0.925
ortholog Arabidopsis|AT3G60030 rapa|Bra041037 118
ortholog Arabidopsis|AT5G18830 rapa|Bra002188 118
ortholog Arabidopsis|AT5G18830 rapa|Bra023695 118
ortholog Arabidopsis|AT5G43270 rapa|Bra027478 118
ortholog Arabidopsis|AT5G50570 rapa|Bra022766 1.031
ortholog Arabidopsis|AT5G50670 rapa|Bra022766 1.031
ortholog Arabidopsis|AT1G20980 rice|LOC_Os08g40260 1.422
ortholog Arabidopsis|AT1G76580 rice|LOC_Os08g40260 1.422
ortholog Arabidopsis|AT2G42200 rice|LOC_Os09g31438 0.376
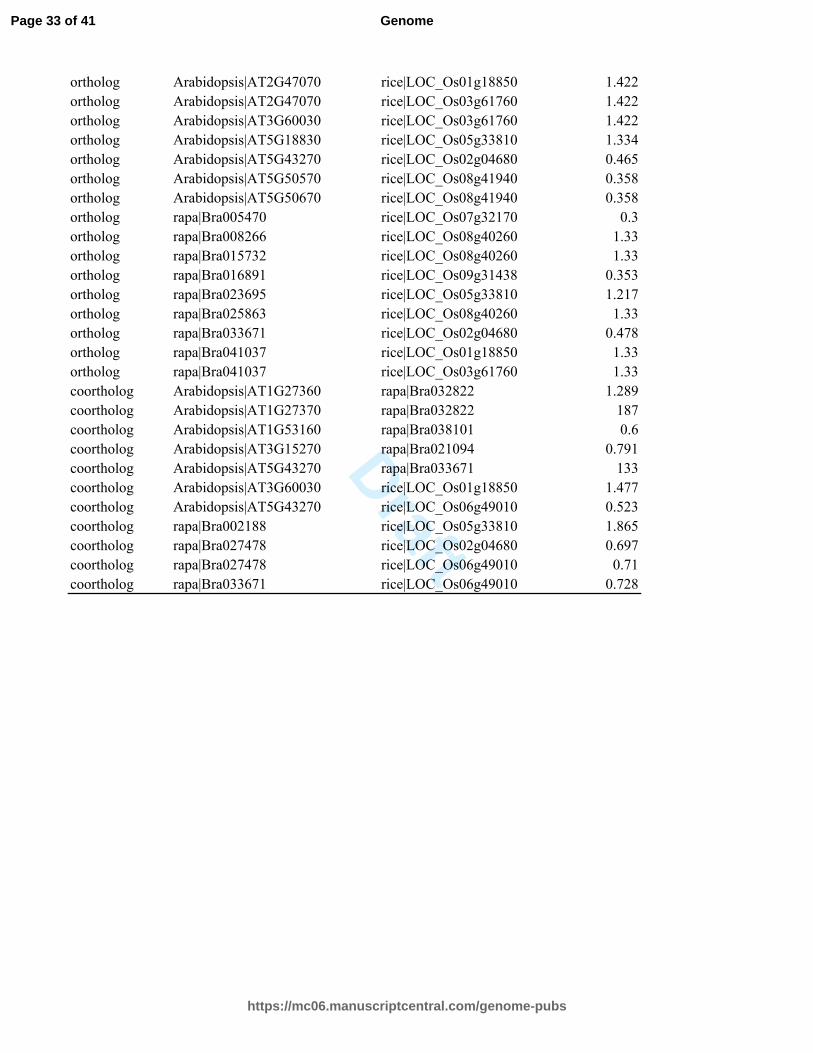
Table S7. Orthologs, coorthologs and paralogs information
Page 32 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome
Draft
ortholog Arabidopsis|AT2G47070 rice|LOC_Os01g18850 1.422
ortholog Arabidopsis|AT2G47070 rice|LOC_Os03g61760 1.422
ortholog Arabidopsis|AT3G60030 rice|LOC_Os03g61760 1.422
ortholog Arabidopsis|AT5G18830 rice|LOC_Os05g33810 1.334
ortholog Arabidopsis|AT5G43270 rice|LOC_Os02g04680 0.465
ortholog Arabidopsis|AT5G50570 rice|LOC_Os08g41940 0.358
ortholog Arabidopsis|AT5G50670 rice|LOC_Os08g41940 0.358
ortholog rapa|Bra005470 rice|LOC_Os07g32170 0.3
ortholog rapa|Bra008266 rice|LOC_Os08g40260 1.33
ortholog rapa|Bra015732 rice|LOC_Os08g40260 1.33
ortholog rapa|Bra016891 rice|LOC_Os09g31438 0.353
ortholog rapa|Bra023695 rice|LOC_Os05g33810 1.217
ortholog rapa|Bra025863 rice|LOC_Os08g40260 1.33
ortholog rapa|Bra033671 rice|LOC_Os02g04680 0.478
ortholog rapa|Bra041037 rice|LOC_Os01g18850 1.33
ortholog rapa|Bra041037 rice|LOC_Os03g61760 1.33
coortholog Arabidopsis|AT1G27360 rapa|Bra032822 1.289
coortholog Arabidopsis|AT1G27370 rapa|Bra032822 187
coortholog Arabidopsis|AT1G53160 rapa|Bra038101 0.6
coortholog Arabidopsis|AT3G15270 rapa|Bra021094 0.791
coortholog Arabidopsis|AT5G43270 rapa|Bra033671 133
coortholog Arabidopsis|AT3G60030 rice|LOC_Os01g18850 1.477
coortholog Arabidopsis|AT5G43270 rice|LOC_Os06g49010 0.523
coortholog rapa|Bra002188 rice|LOC_Os05g33810 1.865
coortholog rapa|Bra027478 rice|LOC_Os02g04680 0.697
coortholog rapa|Bra027478 rice|LOC_Os06g49010 0.71
coortholog rapa|Bra033671 rice|LOC_Os06g49010 0.728
Page 33 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
miRNA name Mature miRNA Sequence Pre-miRNAbra-miR156a-5p UGACAGAAGAGAGUGAGCAC BrSBP1,BrSBP3,BrSBP4,BrSBP8,BrSBP11,BrSBP14,BrSBP15,BrSBP16,BrSBP21,BrSBP22,BrSBP23,BrSBP27,BrSBP28,BrSBP7-3UTRbra-MIR156a
bra-miR156a-3p GCUUACUCUCUCUCUGUCACC bra-MIR156a
bra-miR156b-5p UGACAGAAGAGAGUGAGCAC BrSBP1,BrSBP3,BrSBP4,BrSBP8,BrSBP11,BrSBP14,BrSBP15,BrSBP16,BrSBP21,BrSBP22,BrSBP23,BrSBP27,BrSBP28,BrSBP7-3UTRbra-MIR156b
bra-miR156b-3p GCUCACUCUCUAUCUGUCACC bra-MIR156b
bra-miR156c-5p UGACAGAAGAGAGUGAGCAC BrSBP1,BrSBP3,BrSBP4,BrSBP8,BrSBP11,BrSBP14,BrSBP15,BrSBP16,BrSBP21,BrSBP22,BrSBP23,BrSBP27,BrSBP28,BrSBP7-3UTRbra-MIR156c
bra-miR156c-3p GCUCACUGCUCUAUCUGUCAGA bra-MIR156c
bra-miR156d-5p UGACAGAAGAGAGUGAGCAC BrSBP1,BrSBP3,BrSBP4,BrSBP8,BrSBP11,BrSBP14,BrSBP15,BrSBP16,BrSBP21,BrSBP22,BrSBP23,BrSBP27,BrSBP28,BrSBP7-3UTRbra-MIR156d
bra-miR156d-3p GCUCACUCUCUAUCUGUCACC bra-MIR156d
bra-miR156e-5p UGACAGAAGAGAGUGAGCAC BrSBP1,BrSBP3,BrSBP4,BrSBP8,BrSBP11,BrSBP14,BrSBP15,BrSBP16,BrSBP21,BrSBP22,BrSBP23,BrSBP27,BrSBP28,BrSBP7-3UTRbra-MIR156e
bra-miR156e-3p UGCUCACCUCUCUUUCUGUCAGU bra-MIR156e
bra-miR156f-5p UGACAGAAGAGAGUGAGCAC BrSBP1,BrSBP3,BrSBP4,BrSBP8,BrSBP11,BrSBP14,BrSBP15,BrSBP16,BrSBP21,BrSBP22,BrSBP23,BrSBP27,BrSBP28,BrSBP7-3UTRbra-MIR156f
bra-miR156f-3p UGCUCACUGCUCUUUCUGUCAGA bra-MIR156f
bra-miR156g-5p UGACAGAAGAGAGUGAGCAC BrSBP1,BrSBP3,BrSBP4,BrSBP8,BrSBP11,BrSBP14,BrSBP15,BrSBP16,BrSBP21,BrSBP22,BrSBP23,BrSBP27,BrSBP28,BrSBP7-3UTRbra-MIR156g
bra-miR156g-3p GCUCACUGCUCUAUCUGUCAGA bra-MIR156g
bra-miR157a UUGACAGAAGAUAGAGAGCAC BrSBP1,BrSBP4,BrSBP8,BrSBP14,BrSBP15,BrSBP16,BrSBP21,BrSBP22,BrSBP23,BrSBP27,BrSBP28bra-MIR157a
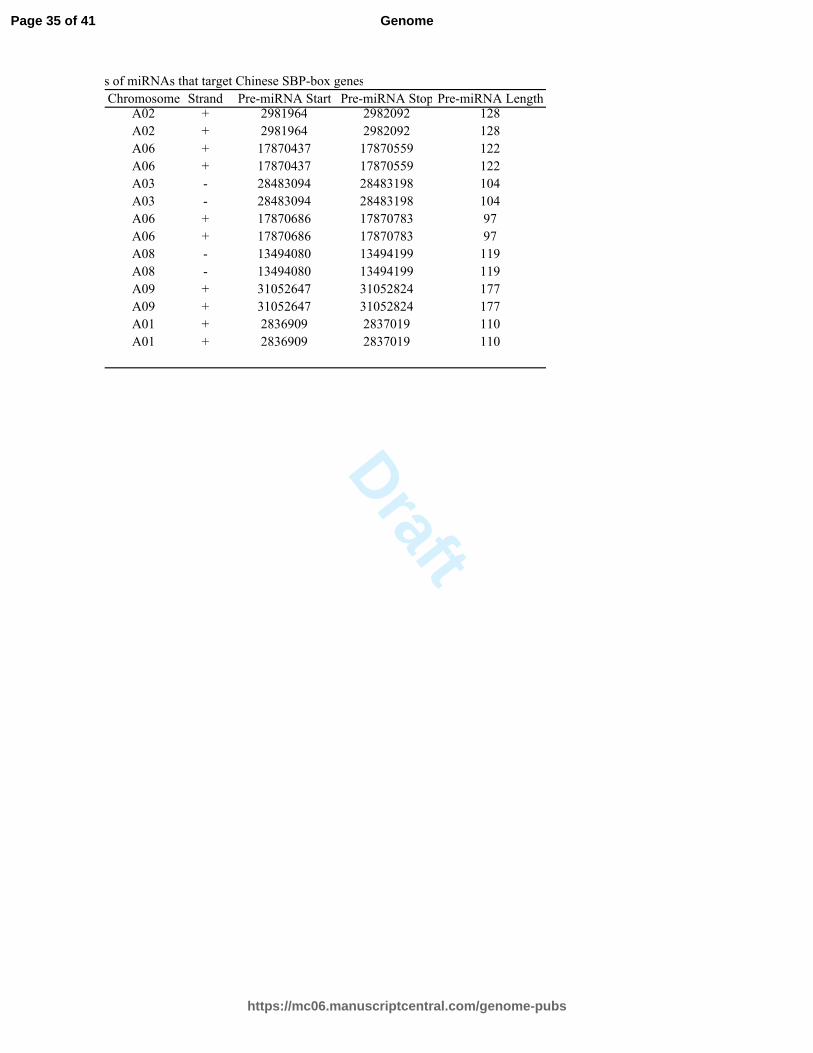
Table S8. Characterisitics of miRNAs that target Chinese SBP-box genes
Page 34 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome
Draft
Chromosome Strand Pre-miRNA Start Pre-miRNA Stop Pre-miRNA LengthA02 + 2981964 2982092 128
A02 + 2981964 2982092 128
A06 + 17870437 17870559 122
A06 + 17870437 17870559 122
A03 - 28483094 28483198 104
A03 - 28483094 28483198 104
A06 + 17870686 17870783 97
A06 + 17870686 17870783 97
A08 - 13494080 13494199 119
A08 - 13494080 13494199 119
A09 + 31052647 31052824 177
A09 + 31052647 31052824 177
A01 + 2836909 2837019 110
A01 + 2836909 2837019 110
Table S8. Characterisitics of miRNAs that target Chinese SBP-box genes
Page 35 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
Pre-miRNA SequenceAAGGUAAGGGGAGGUGACAGAAGAGAGUGAGCACACAUGGUGGCUUUCUUGCAUGCUUUUUUCAAUUAGGGUUUCAUGCUUGAAGCUAUGUGUGCUUACUCUCUCUCUGUCACCCCUUCUCUCUCUCU
AAGGUAAGGGGAGGUGACAGAAGAGAGUGAGCACACAUGGUGGCUUUCUUGCAUGCUUUUUUCAAUUAGGGUUUCAUGCUUGAAGCUAUGUGUGCUUACUCUCUCUCUGUCACCCCUUCUCUCUCUCU
GAGUGCUGAGGAAUUGAUGGUGACAGAAGAGAGUGAGCACACAUGGUGGCUUUCUUGCAUAUUUGAAGAUGUCAUGCUUGAAGCUAUGUGUGCUCACUCUCUAUCUGUCACCGCCUUCUCUC
GAGUGCUGAGGAAUUGAUGGUGACAGAAGAGAGUGAGCACACAUGGUGGCUUUCUUGCAUAUUUGAAGAUGUCAUGCUUGAAGCUAUGUGUGCUCACUCUCUAUCUGUCACCGCCUUCUCUC
CGCAUAGAAACUGACAGAAGAGAGUGAGCACACAAAGGCACUUUGCAUGUUCGAUGCAUUUGCUUCUCUUGCGUGCUCACUGCUCUAUCUGUCAGAUUCCGGCU
CGCAUAGAAACUGACAGAAGAGAGUGAGCACACAAAGGCACUUUGCAUGUUCGAUGCAUUUGCUUCUCUUGCGUGCUCACUGCUCUAUCUGUCAGAUUCCGGCU
UGGUGACAGAAGAGAGUGAGCACACAUGGUGGCUUUCUUGCAUAUUGGAAGAUCUCAUGCUUGAAGCUAUGUGUGCUCACUCUCUAUCUGUCACCCC
UGGUGACAGAAGAGAGUGAGCACACAUGGUGGCUUUCUUGCAUAUUGGAAGAUCUCAUGCUUGAAGCUAUGUGUGCUCACUCUCUAUCUGUCACCCC
GAAAUGCAACAGAGAAAACUGACAGAAGAGAGUGAGCACUUGCAAGGCACUGUUAUGUGUCAAUAACUUUGCGUGUGCGUGCUCACCUCUCUUUCUGUCAGUUGCCUGACUUCUCGCUU
GAAAUGCAACAGAGAAAACUGACAGAAGAGAGUGAGCACUUGCAAGGCACUGUUAUGUGUCAAUAACUUUGCGUGUGCGUGCUCACCUCUCUUUCUGUCAGUUGCCUGACUUCUCGCUU
UUGUGAGUGAAUGAGCUGGGGCAAAAGAAACACACAGAAACUGACAGAAGAGAGUGAGCACACAAAAGUAAAUUGCAUAUGUUGCAUUUGCUUCUCUUGCGUGCUCACUGCUCUUUCUGUCAGAUUCCCGUGCUGAUCUCUUUGGCCUGUCCUUGUUCCUCUUUGUCUCAGUCUCUC
UUGUGAGUGAAUGAGCUGGGGCAAAAGAAACACACAGAAACUGACAGAAGAGAGUGAGCACACAAAAGUAAAUUGCAUAUGUUGCAUUUGCUUCUCUUGCGUGCUCACUGCUCUUUCUGUCAGAUUCCCGUGCUGAUCUCUUUGGCCUGUCCUUGUUCCUCUUUGUCUCAGUCUCUC
CGCAAAGAAACUGACAGAAGAGAGUGAGCACACAAAGGCACUUUGCAUGUUCGAUGCAUUUGCUUCUCUUGCGUGCUCACUGCUCUAUCUGUCAGAUUCCGGCUCCGAUU
CGCAAAGAAACUGACAGAAGAGAGUGAGCACACAAAGGCACUUUGCAUGUUCGAUGCAUUUGCUUCUCUUGCGUGCUCACUGCUCUAUCUGUCAGAUUCCGGCUCCGAUU
GUGAUGCUGGUUGUUGACAGAAGAUAGAGAGCACUAAGGAUGACAUGCAAGUACAUAUGUAUGUAUCAUCACACCGCCUGUGGAUGAUUACAAAAAUAGCCUGCAUGUCUACUCUUUCGUGCUCUCUAUACUUCUGUCACCACCAUUAU
Page 36 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
AAGGUAAGGGGAGGUGACAGAAGAGAGUGAGCACACAUGGUGGCUUUCUUGCAUGCUUUUUUCAAUUAGGGUUUCAUGCUUGAAGCUAUGUGUGCUUACUCUCUCUCUGUCACCCCUUCUCUCUCUCU
AAGGUAAGGGGAGGUGACAGAAGAGAGUGAGCACACAUGGUGGCUUUCUUGCAUGCUUUUUUCAAUUAGGGUUUCAUGCUUGAAGCUAUGUGUGCUUACUCUCUCUCUGUCACCCCUUCUCUCUCUCU
GAGUGCUGAGGAAUUGAUGGUGACAGAAGAGAGUGAGCACACAUGGUGGCUUUCUUGCAUAUUUGAAGAUGUCAUGCUUGAAGCUAUGUGUGCUCACUCUCUAUCUGUCACCGCCUUCUCUC
GAGUGCUGAGGAAUUGAUGGUGACAGAAGAGAGUGAGCACACAUGGUGGCUUUCUUGCAUAUUUGAAGAUGUCAUGCUUGAAGCUAUGUGUGCUCACUCUCUAUCUGUCACCGCCUUCUCUC
CGCAUAGAAACUGACAGAAGAGAGUGAGCACACAAAGGCACUUUGCAUGUUCGAUGCAUUUGCUUCUCUUGCGUGCUCACUGCUCUAUCUGUCAGAUUCCGGCU
CGCAUAGAAACUGACAGAAGAGAGUGAGCACACAAAGGCACUUUGCAUGUUCGAUGCAUUUGCUUCUCUUGCGUGCUCACUGCUCUAUCUGUCAGAUUCCGGCU
UGGUGACAGAAGAGAGUGAGCACACAUGGUGGCUUUCUUGCAUAUUGGAAGAUCUCAUGCUUGAAGCUAUGUGUGCUCACUCUCUAUCUGUCACCCC
UGGUGACAGAAGAGAGUGAGCACACAUGGUGGCUUUCUUGCAUAUUGGAAGAUCUCAUGCUUGAAGCUAUGUGUGCUCACUCUCUAUCUGUCACCCC
GAAAUGCAACAGAGAAAACUGACAGAAGAGAGUGAGCACUUGCAAGGCACUGUUAUGUGUCAAUAACUUUGCGUGUGCGUGCUCACCUCUCUUUCUGUCAGUUGCCUGACUUCUCGCUU
GAAAUGCAACAGAGAAAACUGACAGAAGAGAGUGAGCACUUGCAAGGCACUGUUAUGUGUCAAUAACUUUGCGUGUGCGUGCUCACCUCUCUUUCUGUCAGUUGCCUGACUUCUCGCUU
UUGUGAGUGAAUGAGCUGGGGCAAAAGAAACACACAGAAACUGACAGAAGAGAGUGAGCACACAAAAGUAAAUUGCAUAUGUUGCAUUUGCUUCUCUUGCGUGCUCACUGCUCUUUCUGUCAGAUUCCCGUGCUGAUCUCUUUGGCCUGUCCUUGUUCCUCUUUGUCUCAGUCUCUC
UUGUGAGUGAAUGAGCUGGGGCAAAAGAAACACACAGAAACUGACAGAAGAGAGUGAGCACACAAAAGUAAAUUGCAUAUGUUGCAUUUGCUUCUCUUGCGUGCUCACUGCUCUUUCUGUCAGAUUCCCGUGCUGAUCUCUUUGGCCUGUCCUUGUUCCUCUUUGUCUCAGUCUCUC
CGCAAAGAAACUGACAGAAGAGAGUGAGCACACAAAGGCACUUUGCAUGUUCGAUGCAUUUGCUUCUCUUGCGUGCUCACUGCUCUAUCUGUCAGAUUCCGGCUCCGAUU
CGCAAAGAAACUGACAGAAGAGAGUGAGCACACAAAGGCACUUUGCAUGUUCGAUGCAUUUGCUUCUCUUGCGUGCUCACUGCUCUAUCUGUCAGAUUCCGGCUCCGAUU
GUGAUGCUGGUUGUUGACAGAAGAUAGAGAGCACUAAGGAUGACAUGCAAGUACAUAUGUAUGUAUCAUCACACCGCCUGUGGAUGAUUACAAAAAUAGCCUGCAUGUCUACUCUUUCGUGCUCUCUAUACUUCUGUCACCACCAUUAU
Page 37 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
AAGGUAAGGGGAGGUGACAGAAGAGAGUGAGCACACAUGGUGGCUUUCUUGCAUGCUUUUUUCAAUUAGGGUUUCAUGCUUGAAGCUAUGUGUGCUUACUCUCUCUCUGUCACCCCUUCUCUCUCUCU
AAGGUAAGGGGAGGUGACAGAAGAGAGUGAGCACACAUGGUGGCUUUCUUGCAUGCUUUUUUCAAUUAGGGUUUCAUGCUUGAAGCUAUGUGUGCUUACUCUCUCUCUGUCACCCCUUCUCUCUCUCU
GAGUGCUGAGGAAUUGAUGGUGACAGAAGAGAGUGAGCACACAUGGUGGCUUUCUUGCAUAUUUGAAGAUGUCAUGCUUGAAGCUAUGUGUGCUCACUCUCUAUCUGUCACCGCCUUCUCUC
GAGUGCUGAGGAAUUGAUGGUGACAGAAGAGAGUGAGCACACAUGGUGGCUUUCUUGCAUAUUUGAAGAUGUCAUGCUUGAAGCUAUGUGUGCUCACUCUCUAUCUGUCACCGCCUUCUCUC
GAAAUGCAACAGAGAAAACUGACAGAAGAGAGUGAGCACUUGCAAGGCACUGUUAUGUGUCAAUAACUUUGCGUGUGCGUGCUCACCUCUCUUUCUGUCAGUUGCCUGACUUCUCGCUU
GAAAUGCAACAGAGAAAACUGACAGAAGAGAGUGAGCACUUGCAAGGCACUGUUAUGUGUCAAUAACUUUGCGUGUGCGUGCUCACCUCUCUUUCUGUCAGUUGCCUGACUUCUCGCUU
UUGUGAGUGAAUGAGCUGGGGCAAAAGAAACACACAGAAACUGACAGAAGAGAGUGAGCACACAAAAGUAAAUUGCAUAUGUUGCAUUUGCUUCUCUUGCGUGCUCACUGCUCUUUCUGUCAGAUUCCCGUGCUGAUCUCUUUGGCCUGUCCUUGUUCCUCUUUGUCUCAGUCUCUC
UUGUGAGUGAAUGAGCUGGGGCAAAAGAAACACACAGAAACUGACAGAAGAGAGUGAGCACACAAAAGUAAAUUGCAUAUGUUGCAUUUGCUUCUCUUGCGUGCUCACUGCUCUUUCUGUCAGAUUCCCGUGCUGAUCUCUUUGGCCUGUCCUUGUUCCUCUUUGUCUCAGUCUCUC
GUGAUGCUGGUUGUUGACAGAAGAUAGAGAGCACUAAGGAUGACAUGCAAGUACAUAUGUAUGUAUCAUCACACCGCCUGUGGAUGAUUACAAAAAUAGCCUGCAUGUCUACUCUUUCGUGCUCUCUAUACUUCUGUCACCACCAUUAU
Page 38 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
UUGUGAGUGAAUGAGCUGGGGCAAAAGAAACACACAGAAACUGACAGAAGAGAGUGAGCACACAAAAGUAAAUUGCAUAUGUUGCAUUUGCUUCUCUUGCGUGCUCACUGCUCUUUCUGUCAGAUUCCCGUGCUGAUCUCUUUGGCCUGUCCUUGUUCCUCUUUGUCUCAGUCUCUC
UUGUGAGUGAAUGAGCUGGGGCAAAAGAAACACACAGAAACUGACAGAAGAGAGUGAGCACACAAAAGUAAAUUGCAUAUGUUGCAUUUGCUUCUCUUGCGUGCUCACUGCUCUUUCUGUCAGAUUCCCGUGCUGAUCUCUUUGGCCUGUCCUUGUUCCUCUUUGUCUCAGUCUCUC
Page 39 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome
Draft
Gene Root_1 Root_2 Stem Leaf_1 Leaf_2 Callus Flower Silique
BrSBP1 0.756616 0.764038 15.0976 4.25738 7.05537 0 7.33216 0
BrSBP2 0 0 0.13992 0.559191 0.599698 0 1.36194 0
BrSBP3 2.35488 2.46055 3.12274 1.72164 0.703342 1.20368 4.2021 1.38701
BrSBP4 2.86584 3.27663 8.74825 7.54017 9.78902 0.71844 8.70391 0
BrSBP5 40.4507 38.7602 40.2669 34.422 28.4204 31.9049 45.2483 24.8002
BrSBP6 0.555228 0 0.431904 0 0 2.71212 0.279091 0
BrSBP7 5.67669 4.19335 9.7713 7.92492 7.41201 18.475 24.3286 3.98694
BrSBP8 4.64453 2.86966 2.1524 1.56066 0.75868 0.954481 2.92099 1.07831
BrSBP9 2.46272 3.78704 2.76902 1.68866 2.51034 0 75.9181 1.72907
BrSBP10 0.279937 0.147213 0.29074 0 0 0 14.07 0
BrSBP11 3.07888 3.98698 14.9462 6.87185 8.01645 0.257272 1.43811 0
BrSBP12 0.287145 0 0.149744 0.296271 0.482139 0 0 0
BrSBP13 17.8825 19.6114 22.0673 17.5282 20.8956 18.1867 46.329 10.1702
BrSBP14 0.708541 0.712533 6.56756 0.944426 2.86279 0.109963 2.05023 0
BrSBP15 3.87062 1.67019 6.5319 6.30419 3.58726 0.360407 2.7989 0.158493
BrSBP16 8.60371 8.21788 11.3742 4.61719 3.31481 5.38817 23.6801 1.45159
BrSBP17 30.716 39.6156 50.4626 32.8997 39.04 49.6573 57.9488 21.3327
BrSBP18 4.23615 2.31291 3.02501 1.03271 0.752593 0.270679 13.6143 2.14069
BrSBP19 0.747556 0.711665 1.7201 0 0 0.135339 23.1948 0.713562
BrSBP20 0.679233 0.558269 3.83746 0 1.18322 0 10.5569 0
BrSBP21 1.17007 1.90281 2.90863 0.771821 1.2707 0.577771 11.3088 0.609959
BrSBP22 1.63719 1.80136 2.42208 1.24443 0.816303 1.77853 3.42554 0.93864
BrSBP23 7.33671 9.50304 13.6003 10.6689 8.13694 0.832696 26.1633 0.784741
BrSBP24 0 73.711 59.0046 16.7625 89.1882 7.51926 28.0411 0
BrSBP25 7.93662 7.16234 3.46062 2.53186 2.74284 3.80629 5.53642 6.44741
BrSBP26 8.95531 9.84367 6.12856 5.35434 4.23337 4.99299 8.66192 6.7492
BrSBP27 1.43464 1.49524 1.43252 1.75641 1.18598 0 1.45116 0
BrSBP28 26.0727 35.8825 5.29174 2.85761 3.92107 0.299681 2.78992 0
BrSBP29 6.50194 6.84278 5.98547 6.88159 6.58872 5.19653 14.2673 4.7719
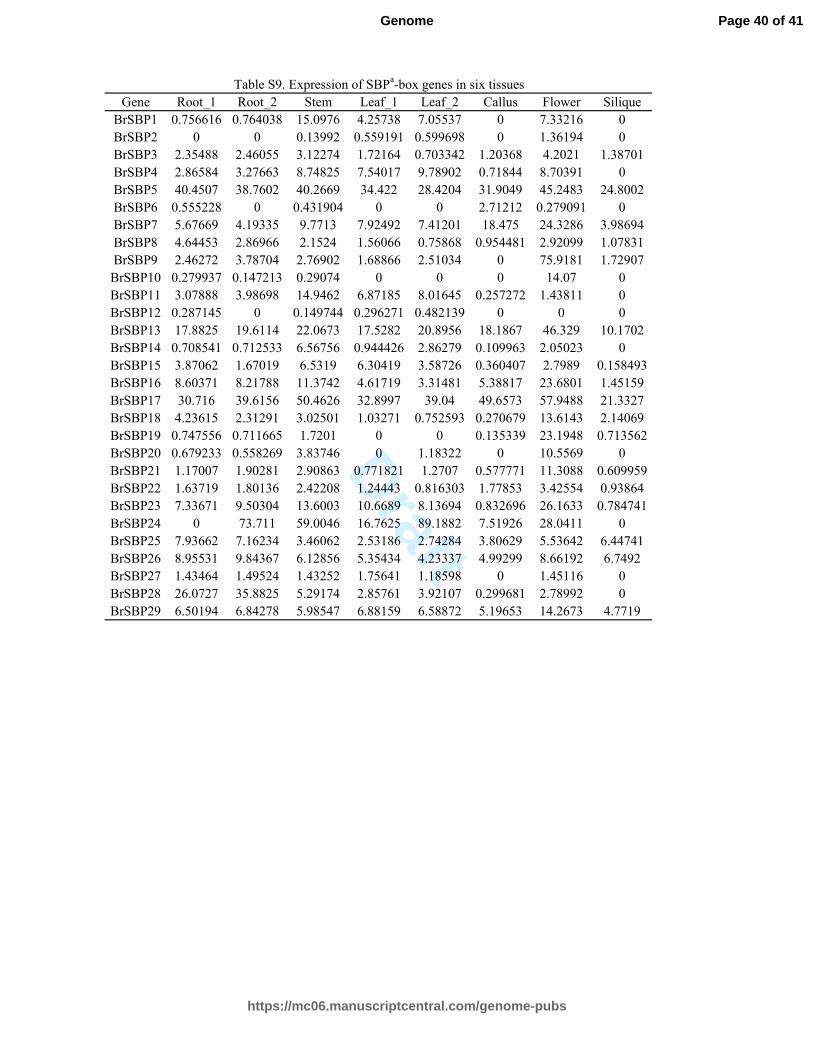
Table S9. Expression of SBPa-box genes in six tissues
Page 40 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome

Draft
Gene SAa 3 h SA 12 h MeJA
b 3 h MeJA 12 h
BrSBP1 -0.082048307 1.586593272 0.332001533 -0.90575646
BrSBP2 0.525150304 0.719921539 0.95001895 0.971563887
BrSBP3 0.312020043 4.423922723 0.06135397 -0.566176672
BrSBP4 0.208623456 1.71874819 1.651550653 -0.521829746
BrSBP6 0.062747502 -0.499256332 1.590944301 0.077065533
BrSBP5 2.668475078 2.544081718 2.092812845 1.126650148
BrSBP7 2.470618073 7.208109924 0.783347664 0.000275066
BrSBP8 28.70696521 24.5346532 1.482512546 1.119120912
BrSBP9 0.104856098 3.159062877 0.310480183 -0.722355355
BrSBP10 -0.619223541 -0.610073783 -0.790971084 -0.798871297
BrSBP11 -0.143460329 4.678674599 -0.258900661 2.172925509
BrSBP12 1.263635695 1.74716407 0.067990426 -0.601012806
BrSBP13 1.174687987 7.375553646 1.238127636 -0.932895878
BrSBP14 -0.312662225 4.400034028 0.962358765 -0.616150842
BrSBP15 4.414024618 7.581301757 -0.077750295 -0.94278136
BrSBP16 -0.836699415 -0.731858467 -0.621660137 -0.32936298
BrSBP17 -0.728251999 -0.621780637 -0.870466133 -0.436144243
BrSBP18 -0.698474835 -0.470774363 -0.89126174 -0.96548447
BrSBP19 -0.745750025 -0.798319215 -0.926347165 -0.960513984
BrSBP20 3.772063028 3.827317022 1.536205581 2.631789293
BrSBP21 0.423286204 1.554348403 0.308550563 0.404381329
BrSBP22 -0.885036902 0.430875183 -0.066454462 -0.781981998
BrSBP23 -0.003496521 -0.586740279 0.169355929 -0.987418482
BrSBP24 1.137888567 -0.136454808 0.169476325 -0.643889687
BrSBP25 1.077347451 1.676471886 0.570154493 0.717097986
BrSBP26 1.897665857 3.257541604 0.545055564 0.677839091
BrSBP27 -0.819749777 -0.880469972 1.049237284 -0.951634171
BrSBP28 -0.049813968 0.340789281 0.260477554 1.25567975
BrSBP29 -0.292123776 0.649723862 -0.377716823 -0.106192825
asalicylic acid
bmethyl jasmonic acid
Table S10. Expression of SBP-box genes under the treatment of plant hormone
Page 41 of 41
https://mc06.manuscriptcentral.com/genome-pubs
Genome