Embed Size (px)
Citation preview


EE-EP 2 152 872 B1
LEIUTISE ALA
Käesolev leiutis käsitleb selliste rekombinantsete polüklonaalsete valkude produtseerimist,
nagu seda on immunoglobuliini sugukonna valgud, näiteks nende lahustuvad või
membraaniga seotud vormid, või T-raku retseptorid, milles kasutatakse
produktsioonisüsteeme, mis on sõltumatud saitspetsiifilisest integratsioonist. 5
LEIUTISE EELDUS
Paljude haiguste puhul, näiteks selliste, nagu infektsioonilised haigused ja erinevad
vähivormid, puuduvad efektiivsed ravimeetodid. Monoklonaalsed antikehad ei ole
tavaliselt efektiivsed kõikide nende märklaudade vastu, osaliselt nende märklaudade
muutlikkuse ja märklaudvalkude adaptiivsete mutatsioonide tõttu, mis teevadki nad 10
tundmatuteks monoklonaalsetele antikehadele. Teisest küljest, polüklonaalsed antikehad
on võimelised tabama mitut dünaamilist märklauda, näiteks, viirustel, bakteritel või
vähirakkudel. Lisaks sellele, polüklonaalsetel antikehadel on suurim tõenäolisus säilitada
aktiivsus antigeeni mutatsioonide puhul.
On olemas erinevaid kaubanduslikult saadavaid, polüklonaalsetel antikehadel põhinevaid 15
havimeid, kaasaarvatud: 1) normaalne inimese immunoglobuliin, mis on eraldatud
normaalsete inimdoonorite verest; 2) inimese hüperimmuunne immunoglobuliin, mis on
tuletatud individuaasete inimdoonorite verest, mis sisaldab antikehasid konkreetsete
haiguse märklaudade, näiteks, viiruste vastu, millega need inimesed on varem kokku
puutunud, kas infektsiooni või vaktsioneerimise käigus; ja 3) looma hüperimmuunne 20
immunoglobuliin, mis on tuletatud immuniseeritud loomade verest.
Immunoglobuliin, mis oli eraldatud inimese verest, on tõestanud oma efektiivsust selliste
infektsioonitekitajate vastu, nagu seda on hepatiidi B viirus, respiratoorne süntsütiaalne
viirus, tsütomegaloviirus ja teised herpese viirused, marutõve viirus, botuliini toksiin, jne,
aga ka neonataalsel reesus-D profülaktikal. Immunoglobuliini, mis sai puhastatud inimese 25
T-rakkudega immuniseeritud küülikute verest, kasutati, et saavutada T-rakkude
immunosuppression transplantaadi ravil või selle hülgamise ärahoidmisel (nt,
tümoglobuliin). Normaalset inimese immunoglobuliini on kasutatud, et võimendada
immunodefitsiitsete patsientide immuunsüsteemi, aga ka erinevate autoimmuunsete
häirete ravil. 30
Sellele vaatamata, immunoglobuliini laialdast kasutamist piirab doonorivere saadavus,
probleemid, mis on seotud erinevate partiidega ja nende erineva ohutusega. Loomse

EE-EP 2 152 872 B1 2
päritoluga immunoglobuliinide kasutamine põrkub samasuguste immunogeensuste
probleemidega nagu need, mida täheldati loomse päritoluga monoklonaalsete antikehade
kasutamisel 1980-ndatel ja 1990-ndatel aastatel. Ja lõpuks, nagu teistegi verest
valmistatud produktidega, eksiteerib ka siin infektsioonitekitajate, nagu HIV, herpes, või
hepatiidi viirused või prioonid, ülekande oht. Kõige selle tõttu on tekkinud olukord, milles 5
arstid tunnistavad, et polüklonaalsed antikehad on eelistatud ravivahenditeks mõningates
situatsioonides, kuid nende kasutamine on jäänud küllaltki piiratuks.
Transgeensete loomade tehnoloogia kasutamisega on tekkinud uued lähenemisviisid
inimese immunoglobuliinide genereerimisel. On loodud transgeensed hiired, kes kannavad
inimese immunoglobuliini lookuseid (USA patent nr 6111166). Need hiired võivad 10
produtseerida inimese terveid immunoglobuliine ja antikehasid, mis on suunatud
konkreetsete märklaudade vastu, mille puhul kasutatakse tavalisi
immuniseerimismeetoodikaid. Kuid suuremad antikeha saagised on siin piiratud hiirte
suhteliselt väikeste mõõtmetega. Transgeenseteks inimese immunoglobuliini geenide
suhtes on tehtud ka suuremaid loomi, näiteks, lehmi (Kurolwa, Y. et al. Nature 15
Biotechnology; 2002; 20: 889-893). Kuid polüklonaalsete antikehade produtseerimine ravi
tarbeks selliste loomade verest, ei kulge samuti ilma komplikatsioonideta. Esiteks,
konkreetse looma ja inimese immunofüsioloogiad võivad omada olulisi erinevusi, mis
põhjustavad erinevusi tulemusena saadud immuunses spektris, funktsionaalses
ümberkorralduses ja immuunse vastuse mitmekesisuses, antikehade näol. Teiseks, 20
sisestatud immunoglobuliini lookuste mitootiline ebastabiilsus võib mõjustada antikehade
pikaajalist produktsiooni. Kolmandaks, tehniliselt on väga raske eemaldada looma oma
immunoglobuliini lookused nii, et näiteks looma antikeha produktsioon ei ületaks inimese
antikeha produktsiooni. Neljandaks eksisteerib selliste infektsioonitekitajate, nagu
viirused, prioonid või teised patogeenid, ülekandmise risk, mis kaasneb loomades 25
produtseeritud antikehade manustamisega.
Viimasel ajal on välja töötatud uus tüüp polüklonaalseid antikehi, mis on sõltumatud
doonori kätteaadavusest selle produtseerimisel. Need polüklonaalsed antikehad
genereeritakse isoleerides doonoritest, kellel on immuunne reaktsioon soovitava märklaua
vastu, seda antikeha kodeerivad nukleiinhappejärjestused, millele järgneb soovitava 30
märklauaga spetsiifiliselt seostuvate antikehade sõelumine. Need polüklonaalsed
antikehad võivad olla valmistatud imetajatele kohandatud ekspressioonitehnoloogiaga,
mis põhineb ühte antikeha ekspresseeriva plasmiidi saitspetsiifilisel integratsioonil iga

EE-EP 2 152 872 B1 3
raku samasse genoomisaiti nii, nagu on kirjeldatud publikatsioonis WO 2004/061104.
Üheks seda uut tüüpi polüklonaalse antikeha näiteks on rekombinantne polüklonaalne
antikeha, mis on suunatud reesus-D vastu (WO 2006/007850). Saitspetsiifiline
integratsioon annab rakupopulatsiooni, milles iga rakk sisaldab ühte koopiat, ja kus
ekspressioonitasemed ja kasvutempo on oodatavalt suhteliselt ühtlased. 5
WO 2004/029284 avalikustab saitspetsiifilise raku ekspressioonisüsteemi, mis on
võimeline vahetama vähemalt ühe märklaudgeeni, kus see see süsteem sisaldab esimest
integratsioonikassetti, esimest märklaudkassetti ja vähemalt ühte recelementi, mis
kodeerib vähemalt ühe rekombinaasi aktiivsust, mis tunneb ära nende kassettide
rekombinaasi äratundmissaiti. 10
WO 2006/007853 avalikustab meetodid rekombinantse polüklonaalse antikeha või teise
rekombinantse polüklonaalse valgu või sellist valku produtseeriva polüklonaalse rakuliini
struktuurseks iseloomustamiseks, mis on ette nähtud selleks, et kontrollida lõpp-
produktide kokkusobivust partiilt partiile aga ka kompositsioonilist stabiilsust ühe
produktsioonitsükli jooksul. 15
Wiberg et al. (Biotechn. Bioeng. (2006) 94(2):396-405) kirjeldavad reesus-D-vastase
rekombinantse polüklonaalse antikeha ekspressiooni ja produktsiooni, mis sisaldab 25
inimese IgG1 antikehal põhinevat saitspetsiifilist antikeha geeni integratsiooni CHO
rakkudes.
Huang et al. (J. Immunol. Methods (April 2007) 322(1-2):28-39) kirjeldavad 20
märklaudraku vektorgeeni integratsioonisüsteemi, mis on ette nähtud kõrgetasemeliseks
antikeha ekspressiooniks, kasutades positsiooniefektide ületamiseks FRT/FLP strateegiat.
Haurum et al. (IDrugs (2005) 8(5):404-9) annavad ülevaate antikehaterapeutikast alates
esimesest inimese immunoglobuliini generatsioonist kuni teise rekombinantsete
monoklonaalsete antikehade generatsioonini, ja edasi rekombinantsete polüklonaalsete 25
antikehadeni.
LEIUTISE OLEMUS
Käesolev leiutis esitleb alternatiivseid meetodeid rekombinantse polüklonaalse valgu
produtseerimiseks, mis on sõltumatu saitspetsiifilisest integratsioonist ja seetõttu näeb ette
suurendatud paindlikkust rakuliini produktsiooni valiku suhtes, säilitades samal ajal valgu 30
polüklonaalsuse. Lisaks sellele, ekspressiooni tasemed võivad olla kõrgemad, kui see on
võimalik saitspetsiifilise integratsiooni puhul.

EE-EP 2 152 872 B1 4
Käesoleva leiutise kohane lähenemine põhineb huvipakkuvate individuaalsete geenide
juhuslikul integratsioonil peremeesrakkudesse, millele elistatavalt järgneb soovitavate
karakteristikutega üksikute rakkude kloneerimine. Need individuaalsed rakukloonid,
millest igaüks produtseerib individuaalset polüklonaalse valgu liiget, segatakse seejärel, et
tekitada polüklonaalseid valke produtseeriv rakuliin polüklonaalse valgu 5
produtseerimiseks.
Tavaliselt on kõige enam tuntumateks polüklonaalseteks valkudeks, polüklonaalsed
antikehad. Sama käib ka T-raku retseptorite (TcR-d) kohta, mis võivad olla saadud
lahustuval kujul, kui nad on produtseeritud rekombinantses ekspressioonisüsteemis, ja mis
võivad omada, sarnaselt polüklonaalsetele antikehadele, ravipotentsiaali. Kuid käesoleva 10
leiutise kohaste rekombinantsete ekspressioonisüsteemidega on võimalik kokku panna ka
selliseid valke, mis ei ole ilmtingimata homoloogsed, näiteks tuntud tähtsusega erinevad
valgud konkreetse defitsiitsuse või haiguse puhul. Käesolevat leiutist illustreerivad
polüklonaalsete antikehade näited, kuid ette on nähtud hõlmata ka polüklonaalseid TcR-sid
ja teisi polüklonaalseid valke, mis võivad olla soovitavad valmistada koos. Sellisteks 15
valkudeks võivad soovi korral olla ka fusioonvalgud.
Käesolev leiutis võimaldab rekombinantsete polüklonaalsete valkude tööstuslikku tootmist
ühes konteineris, näiteks, nende kasutamiseks farmatseutilistes kompositsioonides.
Käesoleva leiutise üheks tähtsaks tunnusjooneks on see, et tootmise käigus
polüklonaalsete valku moodustavate individuaalsete molekulide nihutatud ekspressiooni 20
on hoitud madalal tasemel, et, minimiseerida ebasoovitavaid muutusi partiilt partiile, ja
vältida polüklonaalse antikeha rühma liikmete kõrvaldamist valmistamise käigus.
Ühest aspektist käsitleb käesolev leiutis meetodit polüklonaalse rakkuliini
genereerimiseks, kus see rakuliin on võimeline ekspresseerima polüklonaalset valku, mis
sisaldab 2 kuni n erisugust liiget, kus nimetatud meetod sisaldab: 25
a) Ekspressioonivektorite komplekti ettevalmistamist, milles iga nimetatud vektor
sisaldab vähemalt ühte koopiat erisugustest nukleiihapetest, mis kodeerivad nimetatud
polüklonaalsete valkude rühma konkreetset liiget;
b) Peremeesrakkude eraldi transfekteerimist iga nimetatud ekspressioonivektoriga
tingimustes, milles hoitakse ära ekspressioonivektorite saitspetsiifiline integratsioon 30
rakkude genoomi, misläbi saadakse 2 kuni n kompositsiooni rakke, milles iga
kompositsioon ekspresseerib ühte distinktset polüklonaalse valgu liiget;

EE-EP 2 152 872 B1 5
c) Nimetatud 2 kuni n rakukompositsiooni kokkusegamist, et saada polüklonaalne
rakuliin.
Eelistatud teostusviisis kasutatakse leiutisekohaseid polüklonaalseid rakuliine, kui
polüklonaalseid tootmisrakuliine, ja külmutatakse, säilitatakse ning kasutatakse kui
polüklonaalset baasrakkude panka (Master Cell Bank - pMCB), millest võetud proovid 5
(näiteks, ampullid) võivad olla üles sulatatud ja kasutatud otse rekombinantse
polüklonaalse valgu valmistamiseks või polüklonaalse töörakkude panga (Working Cell
Bank - pWCB) genereerimiseks.
Ühes teostusviisis on leiutisekohasteks ekspressioonivektoriteks episoomivektorid. Teises
eelistatud teostusviisis on leiutisekohased ekspressioonivektorid stabiilselt ja juhuslikult 10
integreeritud peremeesrakkude genoomi. Need ekspressioonivektorid või olla stabiilselt
integreeritud juhuslikes asendites ühes või enamas peremeesraku kromosoomis.
Etapil b) saadud transfekteeritud rakud eelistatavalt kloneeriti. Ühes teostusviisis teostati
kloneerimine, kasutades FACS-kloneerimist, nagu seda on kirjeldatud käesolevas
dokumendis. 15
Leiutisekohased kloonid võivad olla valitud vähemalt ühe kriteeriumi järgi, mis on valitud
rühmast, millesse kuuluvad: kasvu kiirus, kahekordistumise aeg, ekspressiooni tase,
produktsiooni tase, produtseerimise stabiilsus ajas, elujõulisus, töökindlus, vastupidavus,
morfoloogia ja koopiate arv. Eelistatavalt on need kloonid valitud nii, et nad oleksid
ühtlased vähemalt ühe nimetatud kriteeriumi suhtes. Enam eelistatavalt, on kloonid valitud 20
nii, et nad oleksid ühtlased kahekordistumise aja ja ekspressioonitaseme suhtes.
Igale kindlale polüklonaalsele valguliikmele võib olla valitud üks kloon, või enam, kui üks
kloon. Seega, iga kindla polüklonaalse valguliikme kohta võib olla valitud 2 klooni, või
võib olla valitud 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 35, 40,
45, 50, 60, 70, 80, 90, või 100 klooni. 25
Leiutisekohaste rakkude kompositsioon, mis ekspresseerib erinevaid kindlaid liikmeid,
võib olla segatud proportsioonides 1:1, või suhtes, mis erineb 1:1 suhtest.
Eelistatavalt on leiutisekohased ekspressioonivektorid identsed, väljaarvatud
variatsioonid, mis kodeerivad leiutisekohase polüklonaalse valgu järjestust.
See meetod võib olla rakendatud nii monomeersete kui ka multimeersete polüklonaalsete 30
valkude puhul.

EE-EP 2 152 872 B1 6
Multimeersete valkude üks ekspressioonivektor võib kodeerida ühe kindla polüklonaalse
valguliiksme kõiki subühikuid. Alternatiivselt koosneb leiutisekohane kogum
ekspressioonivektoreid etapil a) kahest või enamast ekspressioonivektorite alakogumist,
kus esimene alakogum sisaldab sellist varianti nukleiinhappe järjestustest, mis kodeerib
ühte valgu subühikut, ja teine alakogumist sisaldab nukleiinhappe järjestuste varianti, mis 5
kodeerib teist valgu subühikut, nii, et iga transfektsioon teostatakse liikmega esimesest
ekspressioonivektorite alakogumist ja liikmega teisest ekspressioonivektorite alakogumist.
See transfektsioon võib olla üheaegne või järjestikune. Konkreetselt võib see kogum
ekspressioonivektoreid etapil a) koosneda kahest alakogumist ekspressioonivektoritest,
kus see esimene alakogum sisaldab varianti nukleiinhappe järjestusest, mis kodeerib 10
antikeha rasket ahelat ja teine alakogum sisaldab varianti nukleiinhappe järjestustest, mis
kodeerib antikeha kerget ahelat nii, et iga transfektsioon teostati liikmega esimesest
ekspressioonivektorite alakogumist ja liikmega ekspressioonivektorite teisest alakogumist.
Leiutisekohane ekspressioonivektor või järgmine ekspressioonivektor kodeeris
eelistatavalt selekteeritavat markerit. Lisaks sellele, rakke kultiveeritakse pidevalt sellistes 15
tingimustes, mis soodustavad leiutisekohast selekteeritavat markerit ekspresseerivate
rakkude kasvu. See on kõige paremini tagatav, kasutades selekteeritava markerina
geeniprodukti, mille suhtes peremeesrakk on defitsiitne. See selekteeritav marker on ühes
teostusviisis kodeeritud transkriptiga, mis kodeerib ka polüpeptiidliiget või nimetatud
polüpeptiidliikme subühikut, eelistatavalt, kui see selekteeritav marker on kodeeritud 20
transkriptiga, mis kodeerib suurimat subühikut.
Üks käesoleva leiutise aspekt käsitleb meetodit polüklonaalse valgu valmistamiseks,
milles nimetatud polüklonaalne valk sisaldab 2 kuni n distinktset liiget, kus nimetatud
meetod sisaldab:
a) polüklonaalse rakuliini ettevalmistamist, mis saadakse, kasutades käesoleva leiutise 25
kohast meetodit;
b) leiutisekohase polüklonaalse rakuliini kultiveerimist tingimustes, mis võimaldavad
polüklonaalse valgu ekspressiooni;
c) polüklonaalse valgu saagi kogumist ja valikulist puhastamist rakkudest või
söötmest; ja valikuliselt 30
d) iga erineva kogutud liikme ja valikuliselt puhastatud polüklonaalse valgu
juuresoleku kontrollimist.

EE-EP 2 152 872 B1 7
Käesoleva leiutise autorid on kindlaks teinud, et eri kloonide jaotuvus säilib üllataval
kombel produktsioonitsükli modelleerimise käigus, mis on omane rekombinantse
ravimprodukti tööstusliku tootmise tingimustele. See on väga üllatav, kuna polüklonaalse
antikeha individuaalsete liikmete ekspressioonivektorid integreeruvad erinevates asendites
ja seetõttu, et koopiate arv erineb. Sellest tulenevad erinevused nii ekspressioonitasemes ja 5
kasvu kiiruses.
Vastupidiselt ootustele, selline produtseerimine ei põhjustanud ühe või enama
polüklonaalse antikeha liikme täielikku või osalist kadu. Isegi väikeste erinevuste puhul
kasvu kiirustes erinevatel rakuliinidel on oodata, et mõne aja möödudes polüklonaalne
kompositsioon kaotab kas täielikult või osaliselt vähemalt ühe liikme sellest 10
polüklonaalsest kompositsioonist. Seega, selles käesoleva leiutise teostusviisis on
kompositsiooniline stabiilsus säilinud enam, kui 10 raku pooldumise kestel pärast
töörakkude panga (Working Cell Bank) ülessulatamist, eelistatavalt enam, kui 15
rakupooldumise, näiteks, enam, kui 20 raku pooldumise, näiteks enam, kui 25 raku
pooldumise, näiteks, enam, kui 30 raku pooldumise, näiteks, enam, kui 40 raku 15
pooldumise, näiteks, enam, kui 50 raku pooldumise, näiteks, enam, kui 75 raku
pooldumise, näiteks, enam, kui 100 raku pooldumise kestel.
Lisatud näited demonstreerivad, et see kompositsiooniline stabiilsus säilib vastuvõetavates
piirides enam, kui 25 raku pooldumise kestel.
Kuigi rõhuvas enamuses juhustest on eelistatud eraldi transfektsiooni, 20
ekspressioonivektorite kogumine enne transfektsiooni on võimalik teatud tingimustes. Kui
leiutisekohaseks polüklonaalseks valguks on monomer, on see tõepoolest valikuvõimalus,
kuna see ei ole probleem, et rakk ekspresseerib mitut erinevat polüklonaalse valgu liiget.
Kui leiutisekohaseks polüklonaalseks valguks on multimeer, siis ekspressioonivektorite
kogumine ühisvaruks enne transfektsiooni on vaid siis võimalik, kui kasutatakse võtteid, 25
mis kindlustavad selle, et rakku saab olema sisestatud ainult üks koopia. Muidu võib
toimuda ebasoovitav subühikute segipaiskamine (scrambling).
Kuna ekspressioonivektorite kogumine ühisvaruks on teatud võimalus, siis on see vähem
eelistatav, kui eraldi transfektsioon, kuna on oodata, et see põhjustab vähem töökindla
tootmissüsteemi kompositsioonilise stabiilsuse säilitamise suhtes. 30
Kummagi käesoleva leiutise kohase meetodi puhul on eelduseks võetud, et polüklonaalne
valk on tavaliselt see, mis on loomulikult assotsieeritud rakkudega, milles ekspressioon

EE-EP 2 152 872 B1 8
toimub.
Käesolevas leiutises on kirjeldatud mõningaid meetodeid, mille abil nukleiinhappe
järjestuse variantide raamatukogu võib olla sisestatud peremeesrakuliini, et genereerida
rakkude kollektsiooni, mis on polüklonaalsete valkude valmistamiseks sobilikuks
rakuliiniks. Nende meetodite hulka kuuluvad rakkude kollektsiooni põhimassi 5
transfektsioon nimetatud raamatukoguga, rakualikvootide transfektsioon raamatukogu
fraktsioonidega või, eelistatavalt, individuaalne transfektsioon, kui peremeesrakud on
transfekteeritud nimetatud raamatukogu individuaalsete liikmetega, millele järgneb
selektsiooni käigus genereeritud kloonide kogumine ühisvaruks. Käesolev leiutis kasutab
peremeesrakuliinina eelistatavalt imetajarakke (rakuliine või rakutüüpe). 10
Käesoleva leiutise ühest aspektist on polüklonaalse valgu individuaalsed liikmed
kodeeritud sõltumatute geenisegmentide paaridega. Polüklonaalsete valkude hulka, kus
need individuaalsed liikmed koosnevad kahest või enamast polüpeptiidahelast, kuuluvad
antikehade ja T-raku retseptorite lahustuv või membraaniga seotud vormid. Järgmises
käesoleva leiutise teostusviisis geeni segmentide paar kodeerib antikeha raske ahela ja 15
kerge ahela variaabelpiirkonda, või T-raku retseptori alfa-ahela ja beeta-ahela
variaabelpiirkonda või T-raku retseptori gamma-ahela ja delta-ahela variaabelpiirkonda.
Käesolev leiutis esitleb edasi polüklonaalset rakuliini, mis sisaldab 2 kuni n populatsiooni
rakku igast populatsioonist, millest igaüks ekspresseerib rekombinantse polüklonaalse
valgu erinevat liiget, milles see rekombinantne polüklonaalne valk sisaldab erinevaid, kuid 20
homoloogseid valgumolekule, kus need rakud sisaldavad vähemalt ühte
ekspressioonikonstrukti, mis on juhuslikult integreeritud genoomi nii, et
integratsioonisaidid varieerivad polüklonaalsete rakuliinide liikmete vahel. Ühes
teostusviisis on vähemalt üks ekspressioonikonstrukt juhuslikult integreeritud
kromosoomivälisesse elementi. Teises teostusviisis on vähemalt üks 25
ekspressioonikonstrukt integreeritud juhuslikku asendisse ühes või enamas peremeesraku
kromosoomis.
Leiutisekohane rakuliin pärineb eelistatavalt sellisest imetajaraku liinist, nagu seda on
hiina hamstri munasarja (Chinese hamster ovary - CHO) rakud, COS rakud, BHK rakud,
müeloomirakud (nt Sp2/0 rakud, NS0, YB2/0), NIH 3T3, fibroblastid või 30
immortaliseeritud inimese rakud, sellised nagu HeLa rakud, HEK 293 rakud või PER.C6.
Kuigi kasutatud võivad olla samuti ka sellised mitteimetaja eukarüootsed rakud või
prokarüootsed rakud, nagu taimerakud, putukarakud, pärmirakud, bakterid, seened jne.

EE-EP 2 152 872 B1 9
Samuti on avalikustatud siin DHFR-negatiivne CHO rakk, mis sisaldab stabiilselt
integreeritud nukleiinhapet, mis kodeerib tüüpi 5 adenoviiruse transaktivaatorit E1A, mis
on toimivalt seostatud konstitutiivse promootoriga.
Selline modifitseeritud CHO rakuliin konstrueeriti siin kirjeldatud eksperimentide tarbeks. 5
See rakuliin osutus erakordselt stabiilseks kasvu kiiruste ühtluse ja ekspressiooni tasemete,
ning stabiilsuse suhtes pika aja jooksul, nagu on näidatud käesolevasse dokumenti
lülitatud näidetes, ja on seetõttu eriti hästi kohandatud kasutamiseks käesolevas leiutises.
Hilisemad eksperimendid näitasid, et E1A mRNA ei olnud tuvastatav kaks korda
alakloonitud rakus. See tähendab, et saadud tulemusi, mis osutavad märkimisväärsele 10
kompositsioonilisele stabiilsusele, ei saa kanda ainult E1A ekspressiooni arvele. Sellest
hoolimata on oodata, et DHFR-negatiivne CHO rakuliin, mis ekspresseerib stabiilselt
E1A-d, põhjustab selle rakuliini väga stabiilse ja kõrge ekspressiooni.
Eelistatud teostusviisis on see rakuliin tuletatud DG44 rakuliinist, mis on homosügootne
DHFR-knockout. See rakuliin võib ellu jääda vaid tümidiinidefitsiitses söötmes, kui need 15
rakud sisaldavad rekombinantset DHFR ekspressioonikonstrukti.
Eelistatud teostusviisis sisaldab käesoleva leiutise kohane rakuliin lisaks vähemalt ühte
koopiat stabiilselt integreeritud ekspresseerivat konstrukti, mis kodeerib huvipakkuvat
polüpeptiidi. Selleks huvipakkuvaks polüpeptiidiks võib olla multimeerne valk ja
eelistatavalt on selleks multimeerseks valguks antikeha. 20
Selleks, et võimaldada selekteerimist tümidiinidefitsiitses söötmes, kodeerib see
huvipakkuvat polüpeptiidi kodeeriv ekspressioonikonstrukt, lisaks ka DHFR-i.
Eelistatavalt on DHFR ja vähemalt üks huvipakkuva polüpeptiidi subühik kodeeritud
sama transkriptiga, enam eelistatavalt on DHFR kodeeritud transkriptis, mis kodeerib
suurimat subühikut. See viib soovitava produkti tugeva seostumiseni huvipakkuva 25
polüpeptiidi ja selektsiooni markeri, DHFR-ga, ja tagab, et ellujäänud rakud
ekspresseeriva huvipakkuvat polüpeptiidi.
Huvipakkuva polüpeptiidi ekspressiooni edasiseks suurendamiseks võib huvipakkuva
polüpeptiidi ekspressioon olla kontrollitud ühe või enama promootoriga, mida on võimalik
aktiveerida transkriptsionaalse aktivaatoriga E1A, eelistatavalt sellisega, milles 30
promootoriks on CMV promootor või promootor, mis on tuletatud CMV promootorist.

EE-EP 2 152 872 B1 10
Definitsioonid
Terminiga "valk" või "polüpeptiid" on peetud silmas ükskõik millist aminohapete ahelat,
hoolimata selle pikkusest või post-translatsionaalsest modifikatsioonist. Valgud võivad
eksisteerida monomeeride või multimeeridena, mis sisaldavad kahte või enamat
kokkupandud polüpeptiidahelat, valgu fragmenti, polüpeptiidi, oligopeptiidi või peptiidi. 5
Kasutatuna käesolevas dokumendis, termin "polüklonaalne valk" või "polüklonaalsus"
viitab valgu kompositsioonile, mis sisaldab erinevaid, kuid homoloogseid valgumolekule,
mis on eelistatavalt valitud immunoglobuliinide sugukonnast. Seega iga valgumolekul on
homoloogne teiste kompositsiooni molekulidega, kuid sisaldab kas ühte või enamat
fragmenti variaabelpolüpeptiidjärjestust, mida iseloomustab(vad) erinevus(ed) 10
polüklonaalse valgu individuaalsete liikmete aminohappejärjestus(t)e vahel. Selliste
polüklonaalsete valkude tuntud näidete hulka kuuluvad antikehade või immunoglobuliini
molekulid või nende derivaadid (sellised nagu Fab Fab2; üheahelalised Fvs jne), T-raku
retseptorid ja B-raku retseptorid. Polüklonaalne valk võib koosneda defineeritud
alakogumist valgumolekulidest, millel on defineeritud ühiseid tunnusjooni, selliseid nagu 15
ühine seostumisaktiivsus soovitava märklaua suhtes, näiteks polüklonaalse antikeha puhul
on see seostumise aktiivsus suunatud soovitava märklaudantigeeni vastu.
Termin " huvipakkuv polüklonaalne valk" hõlmab defineeritud polüklonaalsete valkude
alakogumit, millel on selliseid ühised tunnusjooned, nagu seostumisaktiivsus, mis on
suunatud soovitava märklaua vastu, näiteks, juhul, kui tegemist on kirjeldatud 20
polüklonaalsete antikehadega, millel on seostumisaktiivsus või spetsiifilisus suunatud
märklaudantigeeni vastu, kus nimetatud antigeeniks on üks või enam, näiteks, eraldi
valku, mikroorganismi, parasiiti, rakutüüpi, allergeeni või süsivesikumolekule, või
ükskõik millist teist struktuuri, molekuli või ainet, mis võib olla märklauaks antikeha
spetsiifilisel seostumisel, või nimetatud antigeenide segud. 25
Terminid "rekombinantse polüklonaalse valgu kompositsiooni üks liige" või
"rekombinantse polüklonaalse valgu üks liige" tähendab ühte valgumolekuli valgu
kompositsioonis, mis sisaldab erinevaid, kuid homoloogseid valgumolekule, kus iga
valgumolekul on homoloogne teiste kompositsiooni molekulidega, kuid sisaldab ka ühte
või enamat fragmenti variaabelpolüpeptiidjärjestusest, mida iseloomustavad erinevused 30
polüklonaalse valgu individuaalsete liikmete aminohappejärjestuste vahel.

EE-EP 2 152 872 B1 11
Termineid "variaabelpolüpeptiidjärjestus" ja "variaabelpiirkond" on kasutatud
vahetatavalt.
Terminid "rekombinantse polüklonaalse valgu eristatav liige" tähendab ühte
valgumolekuli valgu kompositsioonist, mis sisaldab erinevaid, kuid homoloogseid
valgumolekule, kus iga valgumolekul on homoloogne teiste kompositsiooni molekulide 5
suhtes, kuid sisaldab ka ühte või enamat variaabelpolüpeptiidjärjestuse fragmenti, mida
iseloomustavad erinevused polüklonaalse valgu individuaalsete liikmete
aminohappejärjestustes vahel.
Termin "antikeha" kirjeldab seerumi funktsionaalset komponenti, ja seda nimetatakse tihti
kas molekulide kogumiks (antikehad või immunoglobuliinid) või üheks molekuliks 10
(antikehamolekul või immunoglobuliinimolekul). Antikeha molekul on võimeline
seostuma või reageerima spetsiifilise antigeense determinandiga (antigeeni või antigeense
epitoobiga), mis omakorda võib käivitada immunoloogiliste efektorite mehhanismid.
Individuaalset antikehamolekuli vaadeldakse tavaliselt kui monospetsiifilist, ja
antikehamolekulide kompositsioon võib olla monoklonaalne (s.o, koosneb identsetest 15
antikehamolekulidest) või polüklonaalne (s.o, koosneb erinevatest antikeha molekulidest,
mis reageerivad sama või erinevate epitoopidega samal antigeenil või isegi distinktsetel,
erinevatel antigeenidel). Igal antikehamolekulil on unikaalne struktuur, mis võimaldab
sellel spetsiifiliselt seostuda vastava antigeeniga, ja kõikidel loomulikel
antikehamolekulidel on samad ühised põhistruktuurid, mis sisaldavad kahte identset kerget 20
ahelat ja kahte identset rasket ahelat. Antikehi tuntakse ka üheskoos kui
immunoglobuliine. Terminid antikeha või antikehad hõlmavad, kasutatuna käesolevas
dokumendis, ka kimäärseid ja üheahelalisi antikehasid, aga ka selliseid antikehade
seostumisfragmente, nagu Fab-, Fv-fragmendid või scFv-fragmendid, aga ka selliseid
multimeerseid vorme, nagu dimeersed IgA molekulid või pentavalentsed IgM molekulid. 25
Termin "polüklonaalne antikeha" kirjeldab kompositsiooni erinevatest
antikehamolekulidest, mis on võimelised seostuma või reageerima mõningate erinevate
spetsiifiliste antigeensete determinantidega samal või erinevatel antigeenidel. Tavaliselt
polüklonaalse antikeha variaablus on arvatavasti lokaliseeritud polüklonaalse antikeha
niinimetatud variaabelpiirkondades. Kuid käesoleva leiutise kontekstis võib 30
polüklonaalsuse all mõista ka kirjeldatud erinevusi individuaalsete antikehamolekulide
vahel, mis paiknevad niinimetatud konstantsetes piirkondades, näiteks, kui tegemist on
antikehade segudega, mis sisaldavad kahte või enamat sellist antikeha isotüüpi, nagu

EE-EP 2 152 872 B1 12
inimese isotüübid IgG1, IgG2, IgG3, IgG4, IgA1, IgA2, IgM, IgD ja IgE, või the hiire
isotüübid IgG1, IgG2a, IgG2b, IgG3, ja IgA.
Termin "huvipakkuv rekombinantne polüklonaalne antikeha" kirjeldab defineeritud
rekombinantsete polüklonaalsete antikehade alakogumit, mida iseloomustab võime
seostuda soovitava märklauaga või soovitavate märklaudade kogumiga, kus nimetatud 5
märklaudadeks on, näiteks, eraldi valk, mikroorganism, parasiit, rakk, allergen või
süsivesikumolekul või teine struktuur, molekul või substants, mis võib olla märklauaks
spetsiifilise antikeha seostumisel, või nende segud.
Terminit "immunoglobuliin" kasutatakse tavaliselt tähistamaks veres või seerumis leitud
antikehade kogu segu, kuid see võib samuti olla kasutatud, et tähistada teistest allikatest 10
tuletatud antikehade segu.
Termin "immunoglobuliinimolekul" tähendab individuaalset antikehamolekuli, näiteks,
kui osa immunoglobuliinist, või osa ükskõik millisest polüklonaalsest või
monoklonaalsest antikehakompositsioonist.
Kui kinnitatakse, et polüklonaalse valgu liige seostub antigeeniga, siis peetakse silmas 15
seostumist seostumiskonstandiga, mis on alla 1 mM, eelistatavalt alla 100 nM või veelgi
enam eelistavalt, alla 10 nM.
Terminit "huvipakkuv variantnukleiinhapemolekulide raamatukogu" kasutatakse, et
kirjeldada nukleiinhappemolekulide kollektsiooni, mis üheskoos kodeerivad
"huvipakkuvat rekombinantset polüklonaalset valku". Kasutatuna transfektsiooniks, see 20
huvipakkuvate variantnukleiinhappe molekulide raamatukogu sisaldub
ekspressioonivektorite raamatukogus. Selline raamatukogu sisaldab tüüpiliselt vähemalt 2,
3, 4, 5, 6, 10, 20, 50, 1000, 104, 105 või 106 erinevat liiget.
Käesolevas dokumendis kasutatud terminit "distinktne nukleiinhappejärjestus" tuleb
mõista kui nukleiinhappe järjestust, mis võib kodeerida erinevaid polüpeptiidahelaid, mis 25
üheskoos moodustavad huvipakkuva valgu. Kui need distinksed nukleiinhappe järjestused
koosnevad enam, kui ühest kodeerivast järjestusest, siis need järjestused võivad olla
bitsistronilise transkriptsiooniühiku kujul, või nad võivad töötada kui kaks eraldi
transkriptsiooniühikut, kui nad on toimivalt seostatud sobivate promootoritega. Samuti on
mõeldav tri- ja kvatrorotsistroniliste transkriptsiooniühikute kasutamine, kui huvipakkuv 30
valk koosneb 3 või 4 subühikust, või kui selektsioonimarker on sisse lülitatud
transkriptsioonaalsesse ühikusse koos nukleiinhappega, mis kodeerib huvipakkuvat valku

EE-EP 2 152 872 B1 13
või selle subühikut. Eelistatavalt on distinktne käesoleva leiutise kohane
nukleiinhappejärjestus osaks nukleiinhappemolekulist, näiteks sellise, nagu vektor.
Mõningate näidete hulka, kus on vaja enam, kui ühte kodeerivat järjestust, et tekitada
huvipakkuva valgu terve molekul, kuuluvad B raku retseptorid, antikehad ja selliseid
antikehade fragmendid, nagu Fab's ja variaabeldomeenid, või T-raku retseptorid. 5
Sisestatuna rakku, need geenid, mis üheskoos kodeerivad tervet kokkupandud
huvipakkuvat valku, paiknevad samas vektoris, olles seega sidestatud kokku ühes
nukleiinhappejärjestuses.
Termin "huvipakkuv geen" kasutatuna käesolevas dokumendis, viitab nukleiinhappe
järjestusele, mis koosneb ühest või enamast geeni segmendist (genoomsest või cDNA-st), 10
mis kodeerivad ühte huvipakkuva valgu liiget. "Huvipakkuvate geenide" mitmene vorm
viitab nukleiinhappejärjestuste raamatukogule, mis kodeerib huvipakkuvat polüklonaalset
valku. Terminit "GOI" kasutatakse kui lühendit huvipakkuv(ad) geen(id) (gene(s) of
interest).
Kasutatuna käesolevas dokumendis, termin "vektor" viitab nukleiinhappemolekulile, mille 15
nukleiinhappejärjestus võib olla selle transportimiseks sisestatud erinevatesse
geneentilistesse keskkondadesse ja/või ekspressiooniks peremeesrakku. Kui see vektor
kannab regulatoorseid elemente, mis on vajalikud vektorjärjestusse inserditud
nukleiinhappe transkriptsiooniks (vähemalt sobivat promootorit), siis seda vektorit
nimetatakse "ekspressioonivektoriks". Kui see nukleiinhappe järjestus, mis on sisestatud 20
eelpool identifitseeritud vektoritesse, kodeerib huvipakkuvat valku, nagu on siin
defineeritud, siis kasutatakse järgmiseid termineid " huvipakkuv vektor" ja " huvipakkuv
ekspressioonivektor". Termin "isotüüpi kodeeriv vektor" viitab vektorile, mis kannab
antikeha isotüüpi kodeerivat nukleiinhappejärjestust. Käesolevas spetsifikatsioonis
kasutatakse termineid "fagemiidi vektor" ja "faagi vektor" vahetatavalt. 25
Terminid "plasmiid" ja "vektor" kasutatakse vahetatavalt. Käesolevasse leiutisse on ette
nähtud lülitada sellised teised vektorite vormid, millel võivad olla ekvivalentsed
funktsioonid, näiteks plasmiidid, fagemiidid ja viiruse genoomid või ükskõik millised
nukleiinhappe molekulid, mis on võimelised suunama soovitava valgu produktsiooni õiges
peremeesorganismis. 30
Terminit "huvipakkuvate vektorite raamatukogu iga liige" kasutatakse, et kirjeldada
individuaalseid vektormolekule, millel on eristatav nukleiinhappe järjestus, mis on
tuletatud huvipakkuvate vektorite raamatukogust, kus see nukleiinhappe järjestus kodeerib
huvipakkuva rekombinantse polüklonaalse valgu ühte liiget.

EE-EP 2 152 872 B1 14
Terminit "massiline ülekanne" kasutatakse, et kirjeldada huvipakkuvate
nukleiinhappejärjestuste ülekannet ühest vektorite populatsioonist teise vektorite
populatsiooni ja teha nii iga DNA-ga üheaegselt, kasutamata abinõuna huvipakkuvate
individuaalsete DNA-de eraldamist. Sellisteks vektorite populatsioonideks võivad olla
raamatukogud, mis sisaldavad näiteks huvipakkuvaid variaabelpiirkondi, promootoreid, 5
liiderjärjestusi või intensiivistavaid elemente. Need järjestused võivad seejärel olla
liigutatud ilma eelneva eraldamiseta näiteks faagi vektorist imetaja
ekspressioonivektorisse. Eriti antikeha järjestuste puhul tagab see metoodika selle, et side
VH ja VL mitmekesisuste vahel ei kao raamatukogude üleviimisel, näiteks,
selektsioonivektorist (nt, faagil eksponeeritavast vektorist) imetaja 10
ekspressioonivektorisse. Siinjuures säilitatakse algne VH ja VL paardumine.
Terminit "transfektsioon" on siin kasutatud laias mõttes võõr-DNA sisestamisel rakku.
See termin hõlmab käesolevas dokumendis ka teisi funktsionaalselt ekvivalentseid
meetodeid võõr-DNA sisestamiseks rakku, selliseid nagu näiteks, transformatsioon,
infektsioon, transduktsioon või doonorraku ja aktseptorraku fusioon. 15
Terminit "selektsioon" kasutatakse, et kirjeldada meetodit, kus rakud on omandanud teatud
tunnusjoone, mis võimaldab neid eristada rakkudest, mis ei ole omandanud seda
tunnusjoont. Selliseks tunnusjooneks või olla resistentsus tsütotoksiliste agensite suhtes
või olulise tähtsusega toitaine, ensüüm või värvaine produktsioon.
Termineid "selekteeritav markergeen", "selektsiooni markergeen", "selektsioonigeen" ja 20
"markergeen" on kasutatud, et kirjeldada geeni, mis kodeerib selekteeritavat markerit
(näiteks, geeni, mis annab resistentsuse mõningate selliste tsütotoksiliste ravimite suhtes,
nagu teatud antibiootikumid, geeni, mis on võimeline produtseerima olulise tähtsusega
toitainet, mis võib olla ammendatud kasvusöötmest, geeni, mis kodeerib ensüümi, mis
produtseerib metaboliite, mis võivad olla analüüsitavad, või geeni, mis kodeerib värvilist 25
valku, mis võimaldab, näiteks, sorteerimist FACS abil), mis on sisestatud rakkudesse koos
huvipakkuva(te) geeni(de)ga.
Terminit "rekombinantne valk" kasutatakse, et kirjeldada valku, mida ekspresseeritakse
rakuliini poolt, mis on transfekteeritud valku kodeeriva nukleiinhappejärjestust sisaldava
ekspressioonivektoriga. 30
Kasutatuna käesolevas dokumendis, viitab termin "toimivalt seostatud" segmendile, mis
on seostatud teise segmendiga, kui see on pandud funktsionaalsesse sõltuvusse teisest

EE-EP 2 152 872 B1 15
segmendist. Näiteks, DNA, mis kodeerib signaaljärjestust, on toimivalt seostatud DNA-
ga, mis kodeerib polüpeptiidi, kui seda ekspresseritakse kui liidrit, mis osaleb polüpeptiidi
ülekandes endoplasmaatiline retiikulumi. Ka promootor või võimendusjärjestus on
toimivalt seostatud kodeeriva järjestusega, kui see stimuleerib selle järjestuse
transkriptsiooni. 5
Termin "valdav osa individuaalseid rakke" viitab sellisele rakkude protsendile, nagu enam,
kui 80%, eelistatavalt enam, kui 85%, enam eelistatavalt 90%, 95%, või isegi 99% või
enam.
Kasutatuna käesolevas dokumendis, terminit "genoom" ei tasu võtta täht-tähelt kui tavalist
kromosoomide täiskomplekti, mis on esindatud rakus, vaid ka kromosoomiväliseid 10
elemente, miks võivad olla sisestatud rakku, ja säilitatakse selles. Selliste
kromosoomiväliste elementide hulka võivad kuuluvda, kuid ei ole nendega piiratud, mini-
kromosoomid, YACs (yeast artificial chromosomes – pärmi kunstlikud kromosoomid),
MACs (mouse artificial chromosomes - hiire kunstlikud kromosoomid), või HACs (human
artificial chromosomes - inimese kunstlikud kromosoomid). 15
Termin "promootor" viitab DNA piirkonnale, millega seostub RNA-polümeraas, et
initsieerida transkriptsiooni.
Termin "head-to-head promootorid" (”pea-pea promootorid”) viitab promootoripaarile,
mis on paigutatud teineteise vahetusse lähedusse nii, et nende kahe geenifragmendi
transkriptsioon juhitakse nimetatud promootorite abil vastassuundades. Head-to-head 20
promootorid võivad samuti olla konstrukeeritud täidisega, mis koosneb mitteasjakohastest
nukleiinhapetest nimetatud kahe promootori vahel. Selline täidisfragment võib kergesti
sisaldada enam, kui 500 nukleotiidi.
"Geeniks, mis vastutab resistentsuse eest antibiootkumide suhtes" on geen, mis kodeerib
valku, mis võib ületada antibiootikumi inhibitoor- või toksilise efekti rakule, kindlustades 25
rakkude ellujäämise ja jätkuva proliferatsiooni antibiootikumi juuresolekul.
Termin "sisemine ribosoomi sisenemissait" või "IRES" (internal ribosome entry site)
kirjeldab struktuuri, mis erineb tavalisest 5' cap-struktuurist mRNA-l. Kumbki neist
struktuuridest või olla ära tuntud ribosoomi poolt, et initsieerida AUG-koodoni
skaneerimist ning, et initsieerida translatsiooni. Kasutades ühte promootorjärjestust ja 30
kahe AUG-koodoni initsieerimist, esimene ja teine polüpeptiidjärjestus võib olla
transleeritud üksikult mRNA-lt. Seega, et võimaldada esimese ja teise

EE-EP 2 152 872 B1 16
polünukleotiidjärjestuse koostranslatsiooni ühelt kahetsistroniliselt mRNA-lt, esimene ja
teine polünukleotiidjärjestus võivad olla transkriptsiooniliselt kokkusulandatud
linkerjärjestuse kaudu, mis sisaldab IRES järjestust ja võimaldab selle
polünukleotiidjärjestuse translatsiooni allavoolu IRES järjestusest. Sel juhul
transkribeeritud kahetsistroniline RNA molekul transleeritakse kahetsistronilise RNA 5
molekuli kummastki capped 5' otsast ja sisemisest IRES järjestusest, produtseerides nii
esimest, kui ka teist polüpeptiidi.
Terminit "indutseeritav ekspressioon" kasutatakse, et kirjeldada ekspressiooni, mis vajab
selleks, et ekspressioon leiaks aset, interaktsiooni indutseeriva molekuliga või
korepressormolekuli ja regulaatorvalgu vabastamist. 10
Termin "konstitutiivne ekspressioon" viitab ekspressioonile, mis tavaliselt ei ole
indutseeritav.
Termin "segipaiskamine" ("scrambling") kirjeldab situatsioone, milles kaks või enam
erinev polüklonaalse valgu liiget, millest igaüks sisaldab kaks või enam erinevat
polüpeptiidahelat, näiteks immunoglobuliini sugukonnast, ekspresseeritakse 15
individuaalsest rakust. Selline olukord võib tekkida, kui individuaalne rakk on
integreerinud genoomi enam, kui ühe paari geenisegmente, kus iga paar geenisegmente
kodeerib erinevat polüklonaalse valgu liiget. Sellistes olukordades võivad tekkida
nimetatud geeni segmentidest ekspresseeritud mitte ettenähtud polüpeptiidahelate
kombinatsioonid. Neil polüpeptiidahelate mitte ettenähtud kombinatsioonidel võib 20
terapeutiline efekt üldse puududa.
Termin "VH-VL ahelate segipaiskamine" on üheks selliseks eelpool määratletud
segipaiskamine näiteks. Selles näites geeni segmendid, mis kodeerivad VH ja VL ahelaid,
moodustavad geenisegmentide paari. Nimetatud segipaiskamine leiab aset, kui VH- ja VL-
polüpeptiidide mitte ettenähtud kombinatsioonid produtseeritakse nende rakkude poolt, 25
millesse on integreeritud kaks erinevat VH- ja VL-polüpeptiididi kodeerivat geenisegmendi
paari. Selline skrambleeritud antikeha molekul tõenäoliselt ei säilita algset spetsiifilisust,
ja seega sellel võib puududa mis tahes terapeutiline toime ja isegi mitte ettenähtud
terapeutiline toime.
Termin "rekombinantne polüklonaalne produtseeriv rakuliin" viitab valku ekspresseerivale 30
rakkude populatsioonile, mis on transfekteeritud huvipakkuva variantnukleiinhappe
järjestuste raamatukoguga nii, et need individuaalsed rakud, mis üheskoos moodustavad

EE-EP 2 152 872 B1 17
rekombinantse polüklonaalse produtseeriva rakuliini, kannavad ühte või enamat
huvipakkuva nukleiinhappe järjestuse erinevat koopiat, mis kodeerib ühte selle
huvipakkuva rekombinantse polüklonaalse valgu liiget, ja et iga koopia on integreeritud
iga raku genoomi. Rakud, mis moodustavad selle rekombinantse polüklonaalse
produtseeriva rakuliini, on valitud nende võime järgi säilitada distinktse huvipakkuva 5
nukleiinhappe järjestuse integreeritud koopia, näiteks selektsiooniga, kasutades
antibiootikumi. Rakkudeks, mis võivad moodustada sellise produtseeriva rakuliini, võivad
olla näiteks bakterid, seened, selliseid eukarüootsed rakud, nagu pärm, putukarakud või
imetajarakud, eriti selliseid surematud imetajaraku liinid, nagu CHO rakud, COS rakud,
BHK rakud, müeloomirakud (nt, Sp2/0 rakks, NS0, YB2/0), NIH 3T3 ja selliseid 10
immortaliseeritud inimese rakud, nagu HeLa rakud, HEK 293 rakud või PER.C6.
Terminit "kõrvalekalle" (”bias”) kasutatakse, et tähistada fenomeni, mis leiab aset
rekombinantse polüklonaalse valgu produktsiooni kestel, milles polüklonaalse vektori,
polüklonaalse rakuliini või polüklonaalse valgu kompositsioon muutub pika aja jooksul,
mille põhjuseks on juhuslikud geneetilised mutatsioonid, erinevused individuaalsete 15
rakkude proliferatsiooni kineetikates vahel, erinevused ekspressioonitasemetes erinevate
ekspressioonikonstruktide järjestuste vahel, või erinevused DNA kloneerimise
efektiivsuses.
Termin "RFLP" viitab "restriktsioonifragmentide pikkuse polümorfismile" (restriction
fragment length polymorphism), meetodile, millega analüüsitakse nukleiinhappe 20
molekulide fragmentide migratsioonimustrit geelis pärast lõikamist
restriktsiooniensüümidega.
Termin "5'-UTR" viitab mRNA 5'-otsa transleerimata piirkonnale (untranslated region).
Termin "tingimused, mis hoiavad ära saitspetsiifilise integratsiooni" viitab
transfektsiooniprotsessile, mis ei sisalda ühtegi võimalikest viisidest, millega on võimalik 25
saavutada saitspetsiifiline integratsioon. Saitspetsiifiline integratsioon võib, näiteks, olla
saavutatud, kasutades rekombinaasi ja rekombinaasi äratundmissaidi kombinatsiooni
peremeesraku kromosoomis. See rekombinaas võib samuti olla kovalentselt seostatud
nukleotiidfragmendiga, mis tunneb ära konkreetse saidi kromosoomis. Saitspetsiifiline
integratsioon võib olla saavutatud ka, kuigi väiksema efektiivsusega, kasutades 30
homoloogset rekombinatsiooni. Saitspetsiifilise integratsiooni vältimine põhjustab sageli
integratsiooni juhuslikes asendites peremeesraku genoomi ulatuses, kui kasutatud on
integratsioonivektoreid.

EE-EP 2 152 872 B1 18
Termin "juhuslik integratsioon" viitab ekspressioonivektori integratsioonile peremeesraku
genoomi, asendites, mis on juhuslikud. Sõnastiku kohaselt sõna ”juhuslik” tähendab, et
igal elemendil on võrdsed võimalused selle integratsiooni saidi puhul. Rakkude
transfekteerimisel ei ole kõigil integratsioonisaitidel absoluutselt võrdsed võimalused
integratsiooniks, kuna mõningad kromosoomide osad on enam altid integratsiooniks, kui 5
teised. Kui midagi ei tehta ekspressioonivektori juhtimiseks konkreetsesse
integratsioonisaiti, integreerub see asendis, mis on juhuslik võimalike integratsioonisaitide
rühmas. Seega, terminit "juhuslik integratsioon" tuleb käesoleva leiutise kontekstis mõista
kui transfektsiooniprotsessi, mille puhul mitte midagi ei tehta, et juhtida
ekspressioonikonstrukti ettemääratud asendisse. Vahendite puudumine 10
ekspressioonivektori ettemääratud asendisse juhtimiseks, on piisav selleks, et kindlustada
"juhuslik integratsioon". Seeläbi integratsioonisait(did) transfekteeritud populatsioonis
varieerub rakult rakule ja täpset(eid) integratsioonisaiti(te) võib vaadelda
ettearvamatutena.
Termin "stabiilselt integreeritud" viitab ekspressioonivektori integratsioonile 15
peremeesraku genoomi, milles see integratsioon jääb stabiilseks vähemalt 20, enam
eelistatavalt 30, enam eelistatavalt 40, enam eelistatavalt 50, näiteks, 75, näiteks, 100
põlvkonna jooksul või kauemgi.
Lühendid: "CMV" = (inimese) tsütomegaloviirus (Cytomegalo Virus). "AdMLP" =
adenoviiruse peamine hiline promootor (Major Late Promoter). SV40 poly A = 20
ahviviiruse 40 (Simian Virus 40) polü-A signaaljärjestus. GFP = rohelised fluorestseeruvad
vagud (Green Flourescent Proteins). TcR = T-raku retseptor. ELISA = ensüümseotud
immunosorbent analüüs (Enzyme-Linked Immunosorbent Assay). LTR= pikk otsmine
kordus (Long Terminal Repeat).
JOONISTE KIRJELDUS 25
Joonis fig 1. Skemaatiline ülevaade polüklonaalsete rakkude panga genereerimisest.
See joonis illustreerib skemaatiliselt etappe, mis on vajalikud polüklonaalsete rakkude
panga, näiteks, polüklonaalse baasrakkude panga (Master Cell Bank) saamiseks. a)
Illustreerib erinevaid ekspressioonivektoreid N.A,1, N.A,2, N.A,3, jne, millest igaüks
kodeerib erinevat ja distinkset polüklonaalse valgu liiget. b) Illustreerib 30
ekspressioonivektoritega, transfekteeritavaid peremeesrakke. c) Illustreerib
ekspressioonivektorite integratsiooni erinevates asendites ja erinevatel koopia arvudel
individuaalsetes rakkudes. d) Illustreerib rakukloonide selektsiooni iga polüklonaalse

EE-EP 2 152 872 B1 19
valgu liikme suhtes. Sellel konkreetsel juhul, illustreerimise lihtsustamiseks, on näidatud
ainult üks kloon polüklonaalse valgu ühe eristatava liikme kohta. Etapp e) illustreerib
kloonide segamist, mis olid valitud etapil d), et genereerida polüklonaalsete rakkude pank.
Joonis fig 2a. Prototüüpvektor, mis kodeerib rasket ja kerget ahelat
Selle vektori elemendid on järgmised: 5
• Kaks identset head-to-head inimese CMV promootorit speiserelemendiga nende
vahel
• rasket (VH + gamma 1 konstantne piirkond) ja kerge ahelat (kappa 02-286)
kodeerivad piirkonnad
• bGH=härja kasvuhormooni polüadenülatsioonijärjestus 10
• SV40 pA=SV40 polüadenülatsioonijärjestus
• raske ja kerge ahela genoomsed liidrid
• IRES + DHFR=ECMV sisemine ribosoomi sisenemissait ja hiire
dihüdrofolaatreduktaasi cDNA
• pUC ori=pUC replikatsiooni algus (origin of replication) 15
• bla, amp=ampitsilliini resistentsuse geen
Joonis fig 2b. E1A ekspressioonivektor pML29
Selle vektori elemendid on järgmised:
See vektor põhineb plasmiidil pcDNA3,1+ (Invitrogen)
CMV=inimese CMV promootor 20
Ela=adenoviiruse tüüpi 5 13S transaktivaatori cDNA
bGH=härja kasvuhormooni polüadenülatsiooni piirkond
SV40EP=SV40 varajane promootor
Neo=neo resistentsuse geen
SV40 polyA=SV40 polüadenülatsiooni piirkond AMP=β-laktamaasi geen, mis kodeerib 25
ampitsilliiniresistentsust
Joonis fig 3. IgG sisaldus segudes 1 - 9 viienädalase perioodi kestel, mil see eksperiment
läbi viidi. Detaile vaata näites 1.

EE-EP 2 152 872 B1 20
Joonis fig 4. Ioonvahetuse kromatogrammid, mis näitavad Mix 8 koostist eksperimendi
alguses (must) ja 5 nädalase kultiveerimisperioodi lõpus (sinine).
Joonis fig 5. Esimene (hall) ja viimane (must) proov kõikidest 9 segust analüüsiti
ioonvahetus-kromatograafiaga ja iga individuaalse antikeha sisaldus on arvutatud välja ja
näidatud graafiliselt 5
Joonis fig 6 Proove neljast segust (näide 5), mis olid võetud külvi ja bioreaktori töö kestel,
analüüsiti ioonvahetuskromatograafiaga ja iga individuaalse antikeha sisaldus arvutati
välja, ja see on näidatud graafiliselt.
Segu 1 A - 3 A sisaldas ühte klooni ühe antikeha kohta. Need rakukloonid, mis
ekspresseerivad igat 6 antikeha, olid erinevad segus 1 A (joonis fig 6a), segus 2 A (joonis 10
fig 6b) ja segus 3 A (joonis fig 6c).
Segu 4 A (joonis fig 6d) sisaldas 3 klooni ühe antikeha kohta.
LEIUTISE DETAILNE KIRJELDUS
Rekombinantne polüklonaalse valgu ekspressioonisüsteem
Käesolev leiutis esitleb meetodeid rekombinantsete polüklonaalsete valkude järjekindlaks 15
tootmiseks, kus need valgud on eelistatavalt valitud immunoglobuliini sugukonnast,
immunoglobuliiniga sarnaseid domeene omavate valkude perekonnast. Suurem osa
nendest liikmetest on kaasatud raku pinnal toimuvate sündmuste äratundmiseks.
Järjestuste analüüs näitab, et antikehad, T-raku retseptorid, MHC-molekulid, mõned
rakuadhesioonimolekulid ja tsütokiinide retseptorid on suure homoloogsusega. Eriti selle 20
perekonna liikmed, mis sisaldavad variaabelpiirkondi, on sobilikud käesoleva leiutise
kohaste rekombinantsete polüklonaalsete valkude genereerimiseks. Selliste liikmete hulka
kuuluvad antikehad, membraaniga seotud antikehad (B-raku retseptorid), Fab-fragmendid,
Fv- fragmendid, üheahelalised Fv-fragmendid (scFv), T-raku retseptorid (TcRs),
lahustuvad TcR-d, TcR variaabeldomeeni fragmendid, TcR variaabeldomeeni fragmendid, 25
mis on seostatud polüpeptiidlinkeri või teise antikeha või TcR-st tuletatud fragmentidega.
Konkreetselt, kaalutakse, kas käesolev leiutis või olla kasutatud rekombinantsete
terapeutiliste polüklonaalsete antikehade ja TcR-de suuremastaabiliseks tootmiseks ja
produktseerimiseks.
Üks käesoleva leiutise kohase tootmise peamistest eelistest on see, et kõik liikmed, mis 30
moodustavad rekombinantse polüklonaalse valgu, võivad olla produtseeritud ühes või

EE-EP 2 152 872 B1 21
mõnes bioreaktoris või nende ekvivalentides. Edasi, see rekombinantne polüklonaalne
valgukompositsioon võib olla eraldatud ja puhastatud reaktorist üksiku preparaadina ilma
eraldi individuaalse liikmeta, mis moodustavad rekombinantse polüklonaalse valgu selle
protsessi kestel. Erinevalt sellest, kui tahetakse imiteerida rekombinantse polüklonaalse
antikeha kompositsiooni, segades kokku puhastatud monoklonaalseid antikehi (nagu, 5
näiteks, on pakutud publikatsioonis WO 91/16074), see nõuab iga kompositsiooni
lülitatava antikeha eraldi valmistamist bioreaktoris ja samuti peaksid need antikehad
olema puhastatud eraldi. Selline monoklonaalse segu tootmine saab olema väga kulukas,
ja peale selle on see aega ja ruumi nõudev, võrreldes siin kirjeldatud rekombinantsete
polüklonaalsete antikehade või teiste polüklonaalsete valkude produtseerimise meetodiga. 10
Seega on meetodil, mida on kirjeldatud publikatsioonis WO 91/16074 loomulikud
praktilised piirangud produtseeritavate monoklonaalsete antikehade arvu suhtes, mis
võiksid olla sisse lülitatud sellisesse segusse, samal ajal kui käesoleva leiutise kohase, siin
kirjeldatud tehnoloogiaga, võib üldiselt produtseerida polüklonaalse antikeha ükskõik
millist individuaalset liiget, põhimõtteliselt ilma ülemiste piiranguteta. 15
Kasutatud peremeesrakuliiniks on eelistatavalt imetajaraku liin, mis sisaldab raku liine,
mida tüüpiliselt kasutatakse biofarmatseutiliseks valgu ekspressiooniks, näiteks, CHO
rakke, COS rakke, BHK rakke, müeloomi rakke (nt, Sp2/0 rakke, NS0, YB2/0), NIH 3T3,
ja immortaliseeritud inimese rakke, selliseid nagu HeLa rakud, HEK 293 rakke või rakke
PER.C6. Käesolevas leiutises kasutati CHO rakke, konkreetsemalt, modifitseeritud DG44 20
klooni. Valik sellele konkreetsele rakuliinile langes seetõttu, et CHO rakke kasutatakse
laialdaselt rekombinantsete antikehade valmistamiseks, ja seetõttu, et DG44 kloon võib
olla kasutatud kombinatsioonis metaboolse selektsioonimarkeriga DHFR, mis lisaks
võimaldab kodeeritud geeni amplifitseerida.
Seda DG44 rakuliini modifitseeriti tranfekteerides E1A transaktivaatoriga, mis kodeerib 25
ekspressioonivektorit. Seda tehti, et suurendada üldsaagist, kui huvipakkuv geen on
operatiivselt seostatud CMV promootoriga. See CMV promootor transaktiveeritakse E1A-
ga. See modifikatsioon annab erakordselt stabiilse rakuliini, mis annab rakukloonid, millel
on ühtlased kasvukiirused ja erinevate antikehade ühtlased ning kõrged
ekspressioonitasemed. Seetõttu usutakse, et see modifikatsioon parandab pika aja jooksul 30
kompositsioonilist stabiilsust. Katsed tuvastada E1A ekspressiooni modifitseeritud
rakuliinis ebaõnnestusid. Seetõttu on vähetõenäoline, et ühtlased kasvu kiirused ja
ühtlased ning kõrged ekspressiooni tasemed võivad olla kantud eranditult E1A

EE-EP 2 152 872 B1 22
ekspressiooni arvele. Kuna antud juhul E1A ekspressioon ei olnud tuvastatav, siis endiselt
arvatakse, et CMV promootori transaktivatsioon, milles kasutatakse E1A ekspressiooni
võib põhjustada isegi kõrgemaid ja stabiilsemaid ekspressioonitasemeid.
Ekspressiooniks kasutatakse eelistatavalt BHK-21 rakke või CHO rakke. Sobivate CHO
rakkude hulka kuuluvad, kuid ei ole nendega piiratud CHO-K1 ja CHO-S rakud. 5
Käesoleva leiutise praktikas on eelistatud selliseid imetajarakud, nagu CHO Dhfr-miinus
mutandid, nagu CHO-DUKX-B11 või DG44. Need rakud on sellel alal hästi tuntud ja
laialt kättesaadavad, näiteks, Ameerika Tüüpkultuuri Kollektsioonist (A.T.C.C. - American
Type Culture Collection) Rockville, Md. (BHK-21) või Dr. Lawrence Chasin, Columbia
University, New York (CHO DUKX-B11 või DG44). Need rakkud kohanevad hästi 10
kasvama suspensioonikultuurides (ka seerumivabades tingimustes) ja/või võivad kasvada
väikeste seerumikontsentratsioonide juures ja võivad olla kasutatud ühenduses DHFR
selektsioonimarkeriga.
Eelistatavalt see rakuliin alakloneeritakse ja selekteeritakse kloonide suhtes, millel on
polüklonaalsete valkude liikmete kõrge ja stabiilne ekspressioon. 15
Järelikult, keskmise kogemusega spetsialist on võimeline asendama DG44 klooni teiste
kloonidega ja asendada CHO rakud teiste imetajarakkudega, nagu on kirjeldatud, või isegi
kasutada teist tüüpi rakke, kaasaarvatud taime rakud, pärmirakud, putukarakud, seened ja
bakterid. Seega, rakutüübi valik ei ole piiravaks käesolevas leiutises.
Saagised, mis on saadavad, kasutades käesoleva leiutise kohaseid meetodeid, sõltuvad 20
suurest hulgast parameetritest, mille hulka kuuluvad, kuid ei ole nendega piiratud, selle
kultuuri kasvatamise tingimused ja kasutatud peremeesrakkude liik. Käesoleva leiutise
teostusviisis ületab see saagis eelistatavalt 50 mg/l valku, näiteks, enam, kui 60 mg/l,
näiteks enam, kui 75 mg/l, näiteks, enam, kui 100 mg/l, näiteks enam, kui 125 mg/l,
näiteks, enam, kui 150 mg/l, näiteks enam, kui 200 mg/l, näiteks, enam, kui 250 mg/l, 25
näiteks enam, kui 300 mg/l, näiteks, enam, kui 400 mg/l, näiteks enam, kui 500 mg/l,
näiteks, enam, kui 600 mg/l, näiteks enam, kui 700 mg/l, näiteks, enam, kui 750 mg/l,
näiteks enam, kui 800 mg/l, näiteks, enam, kui 900 mg/l, näiteks enam, kui 1,000 mg/ l
näiteks, enam, kui 2 g/l, näiteks enam, kui 3 g/l, näiteks, enam, kui 4 g/l, näiteks enam, kui
5 g/l. 30
Silmas on peetud, et käesoleva leiutise kohane rekombinantne polüklonaalne valk hõlmab
valgukompositsiooni, mis sisaldab erinevaid, kuid homoloogseid valgumolekule, mis on

EE-EP 2 152 872 B1 23
looduslikult varieeruvad, mis tähendab seda, et eelistatud teostusviisis, sisaldab
variantnukleiinhapete raamatukogu loomulikult asetleidvat mitmekesisust. Seega iga
valgumolekul on homoloogne teiste kompositsiooni molekulidega, kuid sisaldab ka ühte
või enamat variaabelpolüpeptiidjärjestust, mida iseloomustavad erinevused
aminohappejärjestuses polüklonaalsete valkude individuaalsete liikmete vahel. Erinevused 5
aminohappejärjestus(t)es, mis moodustavad variaabelpolüpeptiidjärjestuse võivad olla nii
väikesed, nagu üks aminohape. Eelistatavalt moodustavad need erinevused
aminohappejärjestuses enam, kui ühe aminohappelise erinevuse.
Tavaliselt arvatakse, et see loomulik polüklonaalse antikeha või TcR variaablus
lokaliseerub nende polüpeptiidahelate niinimetatud variaabelpiirkondades või V-10
piirkondades.
Polüklonaalse valgu individuaalsed liikmed võivad sisaldada variaabelpiirkondi, mis on
ligikaudu 80 kuni 120 aminohappejäägi pikkused. Need variaabelpiirkonnad võivad
sisaldada hüpervariaabeldomeene, näiteks, komplementaarsust määravaid piirkondi
(CDR). 15
Looduslikes TcR-des on neli CDR-i igas variaabelregiooni. Looduslikult leiduvates
antikehades on kolm CDR-i raskes ahelas ja kolm CDR-i kerges ahelas.
Lisaks sellele, need polüklonaalse valgu individuaalsete liikmete variaabelpiirkonnad
võivad sisaldada vähemalt ühte hüpervariaabeldomeeni, mis on 1 kuni 26
aminohappejäägi pikkune, eelistatavalt 4 kuni 16 aminohappejäägi pikkune. See 20
hüpervariaabeldomeen võib vastata CDR3 piirkonnale. Antikehade puhul on iga
variaabelpiirkond eelistatavalt moodustatud kolmest hüpervariaabeldomeenist. Need
võivad vastata CDR1-le, CDR2-le ja CDR3-le. TcR-de iga variaabel piirkond on
eelistatavalt moodustatud neljast hüpervariaabeldomeenist. Need võivad vastata CDR1-le,
CDR2-le, CDR3-le ja CDR4-le. Nimetatud hüpervariaabeldomeenid võivad üksinda 25
moodustada variaabeljärjestusi käesoleva leiutise kohase rekombinantse polüklonaalse
valgu variaabelpiirkonnas.
Käesoleva leiutise kontekstis on polüpeptiidjärjestuse variaablus (polüklonaalsus) samuti
mõistetav, et kirjeldada erinevusi individuaalsete antikehamolekulide vahel, mis paiknevad
niinimetatud konstantsetes piirkondades või antikeha polüpeptiidahelate C- piirkondades, 30
näiteks, antikehade segude puhul, mis sisaldavad kaks või enam erinevat antikeha
isotüüpi, näiteks sellist, nagu inimese isotüübid IgG1, IgG2, IgG3, IgG4, IgA1, IgA2,

EE-EP 2 152 872 B1 24
IgM, IgD ja IgE, või hiire isotüübid IgG1, IgG2a, IgG2b, IgG3, IgM ja IgA. Seega
rekombinantne polüklonaalne antikeha võib sisaldada antikehamolekule, mida
iseloomustavad järjestuste erinevused individuaalsete antikeha molekulide vahel
variaabelpiirkonnas (V-piirkond) või konstantses piirkonnas (C-piirkond) või mõlemas.
Eelistatavalt on need antikehad sama isotüüpi, kuna see kergendab järgnevat puhastamist 5
oluliselt. On samuti mõeldav kombineerida antikehasid, näiteks, isotüüpi IgG1, IgG2, ja
IgG4, kuna nad võivad olla kõik puhastatud üheskoos, kasutades affiinsuskromatograafiat
valguga A. Eelistatud teostusviisis on kõikidel antikehadel, mis moodustavad
polüklonaalse antikeha, sama konstantne piirkond, et nende puhastamist veelgi
hõlbustada. Enam eelistatavalt, on neil antikehadel raske ahela konstantsed piirkonnad 10
samad. Kerge ahela konstantne piirkond võib samuti olla kõikidel distinktsetel antikehadel
sama.
Selleks, saada variantnukleiinhappe järjestusi, mis kodeerivad valke, mis seostvad
konkreetset antigeeni, võib olla kasutatud suur hulk sellel alal tuntud meetodeid. Reeglina
on käesolevas leiutises kasulik kasutada sõelumist, mis võimaldab nende nukleiinhapete 15
identifitseerimist ja/või eraldamist, mis kodeerivad valku, mis seostab konkreetset
antigeeni. Mõned nendest meetoditest sisaldavad rikastamise etappi või niinimetatud
biopanning etappi, mis on tuntud sellistest tehnoloogiatest, nagu Symplex™ (Mejier et al,
2006, J. Mol. Biol, 358:764-772; WO 2005/042774), faagil eksponeerimise (Kang, A. S. et
al. 1991. Proc Natl Acad Sci U S A 88, 4363-4366), ribosoomil eksponeerimise 20
(Schaffitzel, C. et al. 1999. J. Immunol. Methods 231, 119-135), DNA-l eksponeerimise
(Cull, M. G. et al. 1992. Proc Natl Acad Sci U S A 89, 1865-1869), RNA-peptiidil
eksponeerimise (Roberts, R. W., Szostak, J.W., 1997. Proc Natl Acad Sci U S A 94, 12297-
12302), kovalentselt eksponeerimise (WO 98/37186), bakteri pinnal eksponeerimise
(Fuchs, P. et al. 1991. Biotechnology 9, 1369-1372), pärmiraku pinnal eksponeerimise 25
(Boder, E. T., Wittrup, K.D., 1997. Nat Biotechnol 15, 553-557) ja eukarüootsete viirusel
eksponeerimise (Grabherr, R., Ernst, W., 2001. Comb. Chem. High Throughput. Screen. 4,
185-192), meetodid, mis kõik on tuntud sellel alal, ja kõik nad on huvipakkuvateks
abivahenditeks käesoleva leiutise. praktikas FACS ja magnetgraanulitel sorteerimine on
samuti rakendatavad rikastamise (panning) eesmärkidel, milles kasutatakse märgistatud 30
antigeeni. Sellised immunodetektsioonitestid, nagu ELISA (Dreher, M. L. et al. 1991. J.
Immunol. Methods 139, 197-205) ja ELISPOT (Czerkinsky, C. C. et.al,1983. J Immunol
meetods. 65, 109-21) võivad samuti olla kasutatud kas pärast biopanning (“biouhtmise”)
etappi või omaette.

EE-EP 2 152 872 B1 25
Huvipakkuv rekombinantse polüklonaalse valgu kompositsioon sisaldab defineeritud
alakogumit valke, mida iseloomustavad selliseid üldised tunnusjooned, nagu ühine
seostumisaktiivsus soovitavate märklaudadega, näiteks, polüklonaalsete antikehade puhul
soovitava märklaudantigeeni vastu. Reeglina on polüklonaalses valgu kompositsioonis
vähemalt 2, 3, 4, 5, 10, 20, 50, 100, 1000, 104, 105 või 106 erinevat variantliiget. 5
Distinktseste liikmete arv, mida on vaja polüklonaalse valgu kompositsiooniks, sõltub
märklaua komplitseeritusest. Juhul, kui tegemist on antikehadega, siis
märklaudantigeeni(de) komplitseeritus mõjub distinktseste variantliikmete arvule, mis on
vajalik polüklonaalse antikeha kompositsioonis. Väikeste või mitte eriti komplekssete
märklaudade puhul, näiteks väikese valgu puhul, mis sisaldab polüklonaalse antikeha 10
kompositsiooni 2-st või 3-st kuni 100 erineva variantliikmeni, võib olla piisav, ja on
eelistatud, et nende variantide arv ei ületa 90, või isegi 80 või 70. Paljudel juhtudel
distinktsete variantide arv ei ületa 60 või 50, ja on eelistatud, et see variantide arv kõigub
vahemikus 5 kuni 40, näiteks, vahemikus 5 kuni 30. Teisest küljest, komplekssemate
märklaudade puhul võib olla piisav, kui polüklonaalse antikeha kompositsioon sisaldab 20 15
kuni 500 distinkset variantliiget. Väga komplekssete märklaudade puhul, kus antigeen
sisaldab palju erinevaid molekule, võib vaja minna, et polüklonaalse antikeha
kompositsioon sisaldaks 50 kuni 10000 erinevat variantliiget.
Imetajatel on mitmeid tuntud näiteid looduslikult leiduvatest polüklonaalsetest valkudest,
mis kas ringlevad vabalt veres, selliseid nagu antikehad või immunoglobuliini molekulid, 20
või on nad esindatud rakkude pindadel, sellised, nagu T-raku retseptorid ja B-raku
retseptorid. Nende looduslikult leiduvate polüklonaalsete valkude mitmekesisus
mõningates imetajates saavutatakse nende valkude variaabelpiirkondi kodeerivate geenide
geneetilise rekombinatsiooniga. Lisaks, on teada, et antikehade mitmekesisust
suurendavad somaatilised mutatsioonid. Käesolevas leiutises on ära kasutatud antikehade 25
loomulikku mitmekesisust, eraldades nukleiinhappejärjestusi, mis vastutavad nende
(näiteks, variaabeldomeenide või immunoglobuliini molekulide CDR piirkondade või
TcR-de) mitmekesisuse eest, ja genereeridess neist raamatukogu. Valkude puhul, mis on
kodeerid kahe sõltumatu geeni segmendis, näiteks antikeha raske ahela variaabelpiirkond
ja kerge ahela variaabelpiirkond, TcRa-ahel ja β-ahel või TcRδ-ahel ja γ- ahel, iga 30
raamatukogus sisalduv vektor moodustab paari nendest variaabelpiirkondadest, mis
kodeerivad järjestusi. Järjestusi kodeerivate variaabelpiirkondade paaride raamatukogude
genereerimine on sellel alal tuntud.

EE-EP 2 152 872 B1 26
Raamatukogudeks, mis sisaldavad looduslikult aset leidvaid mitmekesisusi on näiteks,
kombinatoriaalsed raamatukogud (järjestusi kodeerivate variaabelpiirkondade juhuslik
paardumine), aga ka suguluses olevate paaride raamatukogud (samast rakust, näiteks, WO
2005/042774 tuletatud variaabel piirkondi kodeerivate järjestuste paarid). Järgmised
raamatukogud, mis on genereeritud eraldatud CDR geeni fragmentidest, mis on sisse 5
lülitatud vastavasse karkassi (nt Soderlind, E. et al., 2000. Nat. Biotechnol. 18, 852-856),
näiteks sellisesse, nagu antikeha või TcR variaabelpiirkond, on rakendatavad käesolevas
leiutises. Need raamatukogud on eelistatavalt välja sõelutud, et saada soovitava
spetsiifilisusega alaraamatukogud (huvipakkuvad raamatukogud).
Valkude mitmekesisut võib tekitada ka kunstlikul teel, näiteks sünteetilisel teel või 10
mutatsioonide abil. Mutatsioonid võivad olla kas juhuslikud või punktmutatsioonid
nukleiinhappejärjestuses, mis kodeerib üksikut valku, mille puhul genereeritakse üksiku
valgu polüklonaalne populatsioon. Kunstlike antikehade raamatukogude genereerimise
teist näidet on kirjeldatud publikatsioonis EP 0 859 841, kus on toodud meetod, mis
põhineb variaabelpiirkondade karkasside raamatukogu genereerimisel, mis võib olla 15
kombineeritud teise CDR-de raamatukoguga.
Käesoleva leiutise eelistatud teostusviisis on selleks rekombinantseks polüklonaalseks
valguks rekombinantne polüklonaalne antikeha või antikeha fragment.
Teises käesoleva leiutise eelistatud teostusviisis on rekombinantseks polüklonaalseks
valguks rekombinantne polüklonaalne TcR- või TcR-fragment. 20
Käesoleva leiutise kohane rekombinantne polüklonaalne valk võib seetõttu olla
moodustatud erinevatest isotüüpidest, või enam eelistatult, erinevatest alaklassidest.
Immunoglobuliinide polüklonaalsus võib aset leida immunoglobuliinimolekuli
konstantses osas või selle variaabeldomeenis, või nii konstantses osas kui ka
variaabeldomeenis. 25
Polüklonaalsus niinimetatud konstantses piirkonnas ja konkreetselt, antikehade raskes
ahelas, on huvipakkuv antikehade terapeutilise rakendamise seisukohalt. Erinevatel
immunoglobuliini isotüüpidel on erinevad bioloogilised funktsioonid (mis on
summeeritud tabelis 1), mis võiksid olla soovitavalt kombineeritud antikehade kasutamisel
ravi eesmärgil, kuna loomulikes immuunsetes vastustes võivad osaleda erinevad 30
immunoglobuliini isotüübid (Canfield ja Morrison 1991. J.Exp.Med. 173, 1483-91;
Kumpel et al. 2002. Transfus.Clin.Biol. 9, 45.-53; Stirnadel et al. 2000. Epidemiol.
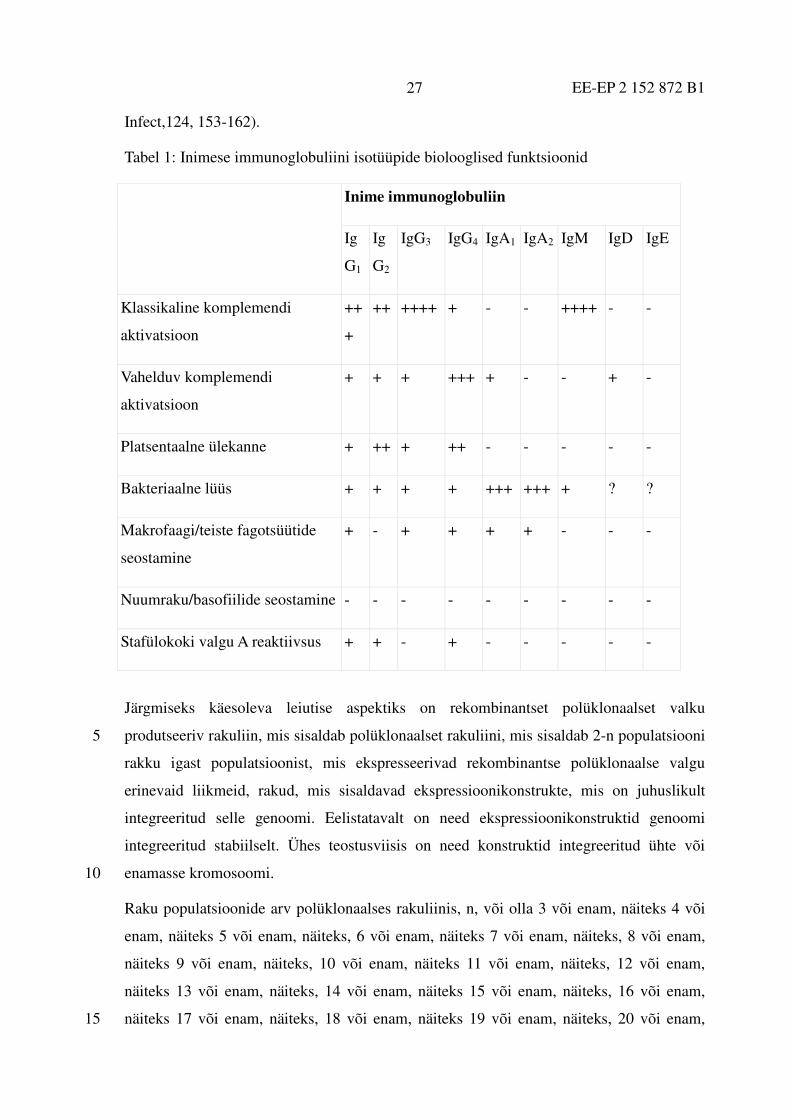
EE-EP 2 152 872 B1 27
Infect,124, 153-162).
Tabel 1: Inimese immunoglobuliini isotüüpide biolooglised funktsioonid
Inime immunoglobuliin
Ig
G1
Ig
G2
IgG3 IgG4 IgA1 IgA2 IgM IgD IgE
Klassikaline komplemendi
aktivatsioon
++
+
++ ++++ + - - ++++ - -
Vahelduv komplemendi
aktivatsioon
+ + + +++ + - - + -
Platsentaalne ülekanne + ++ + ++ - - - - -
Bakteriaalne lüüs + + + + +++ +++ + ? ?
Makrofaagi/teiste fagotsüütide
seostamine
+ - + + + + - - -
Nuumraku/basofiilide seostamine - - - - - - - - -
Stafülokoki valgu A reaktiivsus + + - + - - - - -
Järgmiseks käesoleva leiutise aspektiks on rekombinantset polüklonaalset valku
produtseeriv rakuliin, mis sisaldab polüklonaalset rakuliini, mis sisaldab 2-n populatsiooni 5
rakku igast populatsioonist, mis ekspresseerivad rekombinantse polüklonaalse valgu
erinevaid liikmeid, rakud, mis sisaldavad ekspressioonikonstrukte, mis on juhuslikult
integreeritud selle genoomi. Eelistatavalt on need ekspressioonikonstruktid genoomi
integreeritud stabiilselt. Ühes teostusviisis on need konstruktid integreeritud ühte või
enamasse kromosoomi. 10
Raku populatsioonide arv polüklonaalses rakuliinis, n, või olla 3 või enam, näiteks 4 või
enam, näiteks 5 või enam, näiteks, 6 või enam, näiteks 7 või enam, näiteks, 8 või enam,
näiteks 9 või enam, näiteks, 10 või enam, näiteks 11 või enam, näiteks, 12 või enam,
näiteks 13 või enam, näiteks, 14 või enam, näiteks 15 või enam, näiteks, 16 või enam,
näiteks 17 või enam, näiteks, 18 või enam, näiteks 19 või enam, näiteks, 20 või enam, 15

EE-EP 2 152 872 B1 28
näiteks 21 või enam, näiteks, 22 või enam, näiteks 23 või enam, näiteks, 24 või enam,
näiteks 25 või enam, näiteks, 26 või enam, näiteks 27 või enam, näiteks, 28 või enam,
näiteks 29 või enam, näiteks, 30 või enam, näiteks 35 või enam, näiteks, 40 või enam,
näiteks 45 või enam, näiteks, 50 või enam, näiteks 60 või enam, näiteks, 70 või enam,
näiteks 80 või enam, näiteks, 90 või enam, näiteks 100 või enam. 5
Suuremal osal eesmärkidest võib n olla vähem, kui 50, näiteks, vähem kui45, näiteks
vähem kui 40, näiteks, vähem kui 35, näiteks vähem kui 30.
Üheks käesoleva leiutise tähtsaks teostusviisiks on raku kloneerimise etapp, mis on
teostatud enne lõplikku polüklonaalse rakuliini lõplikku segamist. Selle etapi tulemusena
paraneb saadud polüklonaalsete rakkude pankade saagis ja stabiilsus, mis leiab aset 10
klonaalse nihke minimiseerimise tagajärjel. Rakud, mis ekspresseerivad ühte
rekombinantse polüklonaalse valgu distinktset liiget, võivad olla tuletatud 1 või enamast
kloneeritud rakust, näiteks, 2-st või enamast, näiteks 3-st või enamast, näiteks, 4-st või
enamast, näiteks 5-st või enamast, näiteks, 6-st või enamast, näiteks 7-st või enamast,
näiteks, 8-st või enamast, näiteks 9-st või enamast, näiteks, 10-st või enamast, näiteks, 11 15
või enamast, näiteks, 12-st või enamast, näiteks 13-st või enamast, näiteks, 14 või
enamast, näiteks 15-st või enamast, näiteks, 16-st või enamast, näiteks 17-st või enamast,
näiteks, 18 või enamast, näiteks 19-st või enamast, näiteks, 20-st või enamast, näiteks 21-
st või enamast, näiteks, 22-st või enamast, näiteks 23-st või enamast, näiteks, 24-st või
enamast, näiteks 25-st või enamast, näiteks, 26-st või enamast, näiteks 27-st või enamast, 20
näiteks, 28-st või enamast, näiteks 29-st või enamast, näiteks, 30-st või enamast, näiteks
35-st või enamast, näiteks, 40-st või enamast, näiteks 45-st või enamast, näiteks, 50-st või
enamast, näiteks 60-st või enamast, näiteks, 70-st või enamast, näiteks 80-st või enamast,
näiteks, 90-st või enamast, näiteks 100-st või enamast kloneeritud rakust. Suurema osa
eesmärkide puhul klooneritud rakkude arv on väiksem kui 50, näiteks väiksem kui 20, 25
näiteks, väiksem kui 15, näiteks väiksem kui 10.
Eelmises teostusviisi lisateostusviisis need variantnukleiinhappejärjestused, mis
kodeerivad polüklonaalset valku (eelistatavalt immunoglobuliinide sugukonnast), on kõik
tuletatud looduslikult leiduvatest järjestustest, näiteks on eraldatud doonorist.
Klonaalne mitmekesisus 30
Polüklonaalse valgu üheks karakteristikuks on see, et see on moodustatud hulgast
individuaalsetest valgumolekulidest, kus iga valgumolekul on homoloogne teiste

EE-EP 2 152 872 B1 29
polüklonaalae valgu molekulidega kuid on samuti variaabel, see tähendab, et seda
iseloomustavad polüklonaalse valgu individuaalsete liikmete erinevused
aminohappejärjestustes. Eelistatavalt, need polüklonaalse valgu üldstruktuuri erinevused
on piiratud erinevate piirkondadega. Sellisteks piirkondadeks on, näiteks, antikeha või
TcR-i variaabelpiirkond ja, võimalik, et need erinevused on piiratud CDR piirkondadega 5
nendel aladel. See variaablus võib samuti olla kirjeldatud kui mitmekesisus, mis võib olla
identifitseeritud nii nukleiinhappe tasemel kui ka valgu funktsionaalsel tasemel, näiteks,
spetsiifilisuse ja afiinsuse erinevustes märklaua suhtes.
Rakuliini klonaalne mitmekesisus võib olla analüüsitud RFLP-ga kloonidel, mis on
eraldatud rekombinantset polüklonaalset valku ekspresseerivate rakkude ühisvarust. 10
Teiseks võimaluseks analüüsida klonaalset mitmekesisust, on (RT)-PCR-produktide
sekveneerimine. See mitmekesisus võib samuti olla analüüsitud selle rakuliini poolt
produtseeritud rekombinantse polüklonaalse valgu funktsionaalsete testidega (nt, ELISA).
WO 2006/007853 avalikustab meetodid, mis on ette nähtud polüklonaalse rakuliini ja
polüklonaalse valgu iseloomustamiseks. Need meetodid võivad olla kasutatud rakuliini 15
klonaalse mitmekesisuse ja tulemusena saadud polüklonaalse valgu analüüsimiseks.
Klonaalne nihe (s.o, järk-järguline muutus polüklonaalseid antikehi moodustavates
individuaalsetes antikehades), kui see on olemas, siis või olla hinnatud võrreldes
transfektsiooniks kasutatud algse raamatukogu klonaalset mitmekesisust,
mitmekesisusega, mis leiti rekombinantset polüklonaalset valku ekspresseerivate rakkude 20
ühisvarus (rakuliinis).
Rakuliinist ekspresseeritud polüklonaalse valgu klonaalne mitmekesisus võib olla
hinnatud kui polüklonaalvalgu märklaudulatus. Juhul, kui ligikaudu 25-100%
soovitavatest märklaudmolekulidest on seotud polüklonaalse valguga, siis loetakse, et see
on omandatud piisava mitmekesisuse. Näiteks, polüklonaalse antikeha puhul, antikeha 25
seostumine vähemalt 25% mitteidentsete epitoopidega märklaudantigeeni pinnal räägib
kompositsiooni piisavast mitmekesisusest. Eelistatavalt on klonaalne mitmekesisus
märklaudulatuses vähemalt 50%, ja isegi enam, eelistatavalt vähemalt 75%. Antikehade
puhul, sellist märklaudulatust võib hinnata, näiteks,epitoopide kaardistamisega.
Alternatiivselt, klonaalset mitmekesisust võib hinnata, kui polüklonaalse kompositsiooni 30
individuaalsete liikmete jaotuvust. See jaotuvus võib olla hinnatud kui erinevate
individuaalset liikmete üldarv lõplikus polüklonaalse valgu kompositsioonis, võrreldes

EE-EP 2 152 872 B1 30
erinevate kodeerivate järjestuste arvuga, mis olid sisestatud rakuliini transfektsiooni
kestel. Sel juhul piisavaks omandatud mitmekesisuseks on, kui vähemalt 50%
kodeerivatest järjestustest, mida kasutati algselt transfektsiooniks, võib olla
identifitseeritud polüklonaalsete valkude erinevate individuaalsete liikmetena, eelistatavalt
vähemalt 75%, enam eelistatavalt vähemalt 80%, enam eelistatavalt vähemalt 90%, 5
näiteks, vähemalt 95%, 97%, 98% või 99%. Väljendatuna teisel viisil võib klonaalne
mitmekesisus olla vaadeldud piisavana, kui produtseerimise kestel on kaotatud ainult üks
polüklonaalse valgu liige, või kui kaotatud on 2, 3, 4 või 5 liiget.
Polüklonaalse kompositsiooni individuaalsete liikmete jaotuvust võib hinnata ka üksikute
individuaalsete liikmete vahelise vastastikuse jaotuvuse suhtes. Sel juhul loetakse 10
omandatuks piisavat klonaalset mitmekesisust, kui mitte ükski kompositsiooni üksik liige
ei moodusta enam, kui 75 % polüklonaal valgu kompositsiooni lõplikust üldkogusest.
Eelistatavalt, ükski individuaalne liige ei ületa enam, kui 50%, veelgi enam eelistatult 25
% ja enim eelistatult 10% kogu individuaalsete liikmete arvust lõplikus polüklonaalses
kompositsioonis. Klonaalse mitmekesisuse hindamine, mis põhineb polüklonaalse 15
kompositsiooni individuaalsete liikmete jaotusel võib olla teostatud RFLP analüüsiga,
järjestuste analüüsiga ja valkude analüüsiga, näiteks, kasutades selliseid lähenemisviise,
mida on käesolevas dokumendis hiljem kirjeldatud seoses polüklonaalse kompositsiooni
iseloomustamisega.
Klonaalne mitmekesisus võib väheneda tekkida võiva klonaalse nihke tulemusena a) 20
erinevate ekspressioonitasemete tõttu, b) rakuproliferatsioonis aset leidvate muutuste tõttu.
Kui sellised nihked tekivad, siis iga selline klonaalse mitmekesisuse kadu on kegesti
parandatav väikeste modifikatsioonidega, mis on sisse viidavad siin kirjeldatud
meetoditesse.
On võimalik, et variatsioonid individuaalsete rakkude proliferatsiooni kiirustes 25
konkreetses rakuliinis võivad pika aja jooksul viia sisse nihke rekombinantse
polüklonaalse valgu ekspressiooni, suurendades või vähendades rakuliini poolt
ekspresseeritud mõningate rekombinantsete polüklonaalse valgu liikmete esindatust. Kuna
käesoleva leiutise kohased meetodid võivad põhineda juhuslikul integratsioonil
peremeesraku genoomi, siis nii asend kui ka koopiate arv varieeruvad polüklonaalse 30
rakuliini liikmelt liikmele. Tõenäolislt see põhjustada erinevusi kloonide proliferatsiooni
kiirustes ja ekspressiooni tasemetes. Valides rakukloone, millel on ühesugused
proliferatsioonikiirused, võib seda probleemi minimiseerida. Järgmiseks võimaluseks on

EE-EP 2 152 872 B1 31
kasutada enam, kui ühte klooni iga polüklonaalse valgu liikme kohta. Näited
demonstreerivad, et kompositsiooniline stabiilsus kasvab, kui kasutada nii 3 kuni 5 klooni,
mis ekspresseerivad polüklonaalse valgu üht liiget võrreldes ainult ühe klooni
kasutamisega ühe polüklonaalse valgu liikme kohta.
Teine moodus selle probleemi lahendamiseks, on ühe või enama selektsioonikriteeriumi 5
kasutamine, tagamaks, et rakud oleksid ühtlased teatud etteantud piirides ühe või enama
kriteeriumi suhtes, mis on valitud rühmast, milllesse kuuluvad kasvu kiirus,
kahekordistumise aeg, ekspressiooni tase, produktsioonitase, produtseerimise stabiilsus
ajas, elujõulisus, töökindlus, vastupidavus, morfoloogia ja koopiate arv.
Üheks põhjuseks, mispärast muutuvad rakkude proliferatsiooni kiirused, võib olla see, et 10
selle populatsiooni rakkud, mis moodustavad algseks transfektsiooniks kasutatud rakuliini,
on heterogeensed. On teada, et individuaalsed rakud arenevad rakuliinis pikema
ajavahemiku jooksul erinevalt. Et tagada homogeensem algmaterjal, võib olla läbi viidud
rakuliini alakloneerimine või korduv alakloneeirmine enne seda transfektsiooni
huvipakkuva ekspressioonivektoriga, kasutades selle rakuliini piiratud lahjendamist kuni 15
üksiku raku tasemeni ja iga üksiku raku kasvatamist kuni uue rakkude populatsiooni
saamiseni (niinimetatud rakkude alakloneerimine piiratud lahjendamisega).
Alternatiiviks ja eelistatud meetodiks üksiku raku kloneerimisel hästi defineeritud
rakupopulatsiooni tagamiseks, on fluorestsetselt aktiveeritud rakkude sorteerimine (FACS)
pärast nende transfektsiooni. See võib olla teostatud eelnevalt selektsiooniprotsessile. 20
Fluorestsentselt märgistatud antikehad võivad olla kasutatud rikastamiseks suure
produktiivsusega rakkudega, mis on tuletatud rakkude ühisvarust, mis on transfekteeritud
IgG-konstruktidega (Brezinsky et al. J. 2003. Immunol meetods 277, 141-155). FACS-
sorteerimise eeliseks on see, et selles meetodis on kombineeritud üksiku raku
kloneerimine (üksikute rakkude sorteerimisel süvikutesse) samaaegse informatsiooni 25
saamisega iga üksiku raku ekspressioonitasemest. Et veelgi parandada seda
sorteerimismetoodikat, võib olla söötmesse sisse viidud värv, mis näitab raku elujõulisust
nii, et surnud ja surevad rakud saavad olema eemaldatud. FACS-metoodika puhul
sorteeritavad rakud puutuvad kokku küllaltki karmide tingimustega, kaasaarvatud
tangentsiaalpinge. See tähendab, et rakud selekteeritakse kaudselt ka vastupidavuse suhtes 30
sellistes tingimustes. Lisaks sellele, see FACS-metoodika on automatiseeritud, mis
võimaldab sorteerida suurt arvu üksikuid rakke.

EE-EP 2 152 872 B1 32
See FACS meetod võib olla samuti kasutatud, rakkude sorteerimiseks, mis ekspresseerivad
samasuguseid tasemeid immunoglobuliini, moodustades produktiivsuse suhtes
homogeense rakupopulatsiooni. FACS-meetoditega võivad olla välja selekteeritud rakud,
millel on ühesugused proliferatsioonikiirused, ka kasutades märgistamist fluorestsentse
värviga, 5,6-karboksüülfluorestseiindiatsetaadi suktsiinimidüülestriga (CFSE - 5,6-5
carboxylfluorescein diacetate succinimidyl ester).
Käesoleva leiutise tähtsaks teostusviisiks on ühe või enama kloneeritud rakuliini
genereerimine polüklonaalse antikeha iga liikme kohta. Üksikute rakukloonide
genereerimine võib olla läbi viidud, kasutades ükskõik millist ühte suurest hulgast
meetoditest. Kuigi, tuli välja, et FACS-rakkude sorteerimine, kus rakud valitakse 10
elujõulisuse ja IgG tasemete järgi, ning sorteeritakse individuaalsetesse süvikutesse,
osutus samavõrd stabiilseks kloonide saamisel, mis sobivad polüklonaalsete baasrakkude
panga (Master Cell Bank) ja seejärel polüklonaalse töörakkude panga (Working Cell Bank)
valmistamiseks. Selektsiooni surve, näiteks Mtx poolt, võib olla kõrvaldatud enne FACS-
sorteerimist või selle kestel, kuid pidev selektsioonisurve, näiteks, kultiveerimisel 15
nukleosiidivabas söötmes, eelistatavalt säilitatakse. Individuaalsed kloonid on eelistatavalt
valitud pärast teatud päevade arvu kestnud kultiveerimist selektsioonisurve all, mis leidis
aset pärast rakkude sorteerimist. Kuna kloonid on valitud samal päeval pärast sorteerimist,
siis nende kloonide kasvu kiirus on suhteliselt ühtlane. Lisaks sellele, neid kolooniaid
kontrolliti visuaalselt, et eemaldada kloonid, milles on suured morfoloogilised muutused 20
ja madalad kasvu kiirused võrreldes algse transfekteerimata rakuliiniga. Ja lõpuks,
antikeha ekspressioonitase võib olla hinnatud, kasutades, näiteks, ELISA-testi või teist
analüütilist meetodit, ja nende abil võivad olla välja valitud kloonid, millel on kõrge ja
suhteliselt ühtlane ekspressiooni tase.
Isegi kui proliferatsioonikiiruse poolt indutseeritud nihe ei arene, siis individuaalsete 25
liikmete kadu või üleekspressioon võib olla mittekriitiline, sõltuvalt nõuetest lõpliku
rekombinantse polüklonaalse valguprodukti mitmekesisuse ja selle mitmekesisuse
stabiilsuse suhtes pika aja jooksul.
Peremeesrakk
Peremeesrakud võivad olla genereeritud ükskõik millistest rakkudest, mis võivad 30
integreerida DNA nende kromosoomidesse või säilida selle sellistes kromosoomivälistes
elementides, nagu minikromosoomid, YAC-d (pärmi kunstlikud kromosoomid), MAC-d

EE-EP 2 152 872 B1 33
(hiire kunstlikud kromosoomid) või HAC-d (inime kunstlikud kromosoomid). MAC-e ja
HAC-e on detailselt kirjeldatud publikatsioonis WO 97/40183. Eelistatavalt on
imetajarakkudena kasutatud selliseid rakke, nagu CHO rakke, COS rakke, BHK rakke,
müeloomi rakke (nt, Sp2/0, YB2/0 või NS0 rakke), selliseid fibroblastid, nagu NIH 3T3 ja
selliseid immortaliseeritud inimese rakke, nagu HeLa rakud, HEK 293 rakud, või PER.C6. 5
Kuid kasutatud võivad olla ka sellised mitteimetaja eukarüootsed või prokarüootsed rakud,
nagu taimerakud, putukarakud, pärmirakud, seened, E. coli jne.
Ühes käesoleva leiutise teostusviisis algmaterjalina kasutatav rakuliin alakloneeriti,
teostades rakuliini niinimetatud piiratud lahjendust kuni üksiku raku tasemeni, millele
järgneb iga üksiku raku kultiveerimine uueks rakupopulatsiooniks enne transfektsiooni 10
huvipakkuvate vektorite raamatukoguga. Soovi korral selline alakloneerimine võib olla
teostatud ka hiljem, õige rakuliini selekteerimise protsessis. Teiste, üksiku raku
kloneerimise meetodite hulka kuuluvad: FACS kloneerimine (Brezinsky et al. J. 2003.
Immunol Methods 277, 141-155), LEAP™ tehnoloogia (Cyntellect, San Diego, California,
USA), ja ClonePix (Genetix, UK). 15
Integreerimiseks vajalik vektor
Alljärgnev kirjeldus keskendub imetaja ekspressioonisüsteemide kasutamisele. Kuigi,
käesoleva leiutise kohaseid meetodeid on võimalik kasutada ka ekspressiooniks bakterites,
kasutades sobivaid modifikatsioone.
Sobiv vektor sisaldab sobivat selektsioonigeeni. Selektsioonigeenide hulka, mis sobivad 20
kasutamiseks imetajaraku ekspressioonil, kuuluvad, kuid ei ole nendega piiratud, sellised
geenid, mis võimaldavad läbi viia toitumisega seotud selektsiooni, nagu tümidiinkinaasi
geen (TK), glutamiinsüntetaasi geen (GS), trüptofaansüntetaasi geen (trpB) või
histidinooldehüdrogenaasi geen (hisD). Järgmisteks selektsiooni markeriteks on sellised
antimetaboliitide resistentsusegeenid, mis teevad rakud ravimiresistentseks, nagu 25
dihüdrofolaatreduktaasi geen (dhfr), mis võib olla välja selekteeritud, kasutades
hüpoksantiini ja tümidiinidefitsiitset söödet ja edasi selekteeritud, kasutades metotreksaati;
ksantiin-guaniinfosforibosüültransferaasi geeni (gpt), mis võib olla selekteeritud,
kasutades mükofenoolhapet, neomütsiini fosfotransferaasi geeni (neo), mis võib olla
selekteeritud kasutades G418, eukarüootsetes rakus, ja neomütsiini või kanamütsiini 30
prokarüootsetes rakkudes; hügromütsiini B fosfotransferaasi (hyg, hph, hpt) geen, mis
võib olla selekteeritud kasutadesc hügromütsiini; puromütsiin N-atsetüültransferaasi geen

EE-EP 2 152 872 B1 34
(pac), mis võib olla selekteeritud, kasutades puromütsiini; või blastitsidiin S deaminaasi
geeni (Bsd), mis võib olla selekteeritud, kasutades blastitsidiini; seotsiiniresistentsuse geen
(Sh ble), mis vahendab resistentsust zeotsiini ja bleomütsiini vastu. Ja lõpuks, geenid, mis
kodeerivad valke, mis võimaldavad sorteerimist, näiteks, läbivoolutsütomeetriaga, võivad
samuti olla kasutatud kui selektsioonimarkerid, mille hulka kuuluvad selliseid nagu 5
roheline fluorestseeruv valk (GFP - green fluorescent protein), närvikasvufaktori retseptor
(NGFR - nerve growth factor receptor) või teised membraanivalgud, või
beetagalactosidaas (LacZ).
See selektsioonimarker võib paikneda eraldi ekspressioonivektoril, seega
kaastransfektsioon teostatakse ekspressioonivektoriga, mis kodeerib selektsioonimarkerit 10
ja ühte või enamat ekspressioonivektor(it), mis kodeerib huvipakkuvat valku või
huvipakkuva valgu subühikut. See selektsioonimarker võib samuti paikneda
ekspressioonivektoril, mis kodeerib huvipakkuvat valku. Viimasel juhul see
selektsioonimarker paikneb eelistatavalt transkriptil, mis kodeerib ka huvipakkuvat valku
või selle ühte subühikut. See võib olla tehtud, näiteks, kasutades IRES-konstrukti. 15
Multimeerse valgu, nagu näiteks antikeha puhul, see selektsioonimarker paikneb
eelistatavalt transkriptil, mis kodeerib suurimat subühikut, näiteks sellist nagu antikeha
raske ahel.
Vektor, mis on ette nähtud huvipakkuva geeni integreerimiseks, sisaldab lisaks DNA-d,
mis kodeerib huvipakkuva rekombinantse polüklonaalse valgu ühte liige, millele eelneb 20
oma, imetaja promootor, mis suunab selle valgu ekspressiooni. Kui huvipakkuva
rekombinantse polüklonaalse valgu liige sisaldab enam, kui ühte valguahelat, näiteks, kui
see liige on antikeha, või T-raku retseptor, siis see DNA, mis kodeerib selle valgu ahelaid,
võib sisaldada eelnevalt oma, imetaja promootorit, mis suunab iga selle ahela
kõrgetasemelist ekspressiooni (kahesuunalist või ühesuunalist). Kahesuunalise 25
ekspressiooni puhul võib ekspressioonivektoris olla kasutatud head-to-head promootori
konfiguratsiooni, ja ühesuunalisel ekspressioonil võib ekspressioonil olla kasutatud kaks
promootorit või üks promootor, mis on kombineeritud, näiteks, IRES järjestusega. Samuti
on mõeldav kahetsistroniline ekspressioonivektor, milles on kodeeritud kaks erinevat
subühikut samas transkriptis, mis on eraldatud IRES-järjestusega. 30
Sobivateks head-to-head promootori konfiguratsioonideks on näiteks AdMLP promootor
hiire metallotioneiin-1 promootoriga kummaski orientatsioonis, kus see AdMLP
promootor koos elongatsioonifaktori-1 promootoriga kummaski orientatsioonis või CMV

EE-EP 2 152 872 B1 35
promootor koos MPSV promootoriga kummaski orientatsioonis, või nimetatud CMV
promootor, kasutatna kummaski orientations, kuid ei ole nendega piiratud.
Juhul, kui tegemist on antikehadega, siis kogemused on näidanud, et raske ahela kogused,
mida ekspresseerib rakk, ei või ületada kerge ahela koguseid. Seetõttu, promootor, mis
juhib kerge ahela ekspressiooni, on eelistatavalt vähemalt sama tugev, kui promootor, mis 5
suunab raske ahela ekspressiooni.
Nukleiinhappejärjestus, mis kodeerib funktsionaalset liiderjärjestust või
translokatsioonisignaali või olla sisse lülitatud ekspressioonivektorisse, et suunata seda
geeniprodukti endoplasmaatilisse retiikulumi või sellisesse konkreetsesse paika rakus,
nagu organell. Tugev polüadenülatsioonisignaal võib asetseda valku kodeeriva DNA 10
järjestuse 3'-otsas. See polüadenülatsioonisignaal tagab tekkiva RNA transkripti
terminatsiooni ja polüadenülatsiooni ja on korrelatsioonis selle signaali stabiilsusega.
DNA, mis kodeerib huvipakkuva rekombinantse polüklonaalse valgu liiget, võib, näiteks,
kodeerida nii antikeha rasket kui ka kerget ahelat või antikeha fragmente, kus iga geeni
järjestusele eelnevad valikuliselt nende oma imetaja promootori elemendid ja/või neile 15
järgnevad tugevad polü-A signaalid, mis suunavad kummagi kahe ahela kõrgetasemelist
ekspressiooni.
Integreerimiseks ette nähtud ekspressioonivektor võib kanda selliseid lisa
transkriptsionaalseid regulaatorelemente, nagu võimendusjärjestused, anti-repressoreid või
UCOE-d (ubiquitous chromatin opening elements - kõikjalolevad kromatiini avavad 20
elemendid), ekspressiooni ja ekspressiooni stabiilsuse suurendamiseks
integratsioonisaidis. Võimendusjärjestused on nukleiinhappejärjestused, mis
interageerivad spetsiifiliselt tuumavalkudega, mis on kaasatud transkriptsiooni. UCOE-d
avavad kromatiini või säilitavad kromatiini avatud olekus ja hõlbustavad toimivalt
sidestatud geeni reprodutseeritavat ekspressiooni (seda on detailsemalt kirjeldatud 25
publikatsioonides WO 00/05393 ja Benton et al, Cytotechnology 38:43-46, 2002). Lisa
võimendusjärjestuste või võimenduselementide hulka kuuluvad maatriksiga kinnitumise
piirkonnad (MARs - Matrix Attachment Regions), nagu on kirjeldanud, näiteks, Girod &
Mermod 2003 ("Chapter 10: Use of scaffold/matrix-attachment regions for protein
production", pp 359-379 In Gene Transfer and Expression in Mammalian Cells, SC 30
Makrides (ed), 2003, Elsevier Science BV). Antirepressor elementide hulka kuuluvad,
kuid ei ole nendega piiratud, STAR elemendid (Kwaks et al Nat Biotechnol. 2003 May;
21(5):553-8). Kui üks või enam eelpool kirjeldatud regulaatorelementi on integreeritud

EE-EP 2 152 872 B1 36
peremeesraku kromosoomi, siis neid nimetatakse heteroloogsseteks
regulaatorelementideks.
Polüklonaalsed tööraku pangad ja baasrakkude pangad
Käesoleva leiutise eelistatud teostusviisis kasutatakse polüklonaalste baasrakkude
pankasid (pMCB - polyclonal Master Cell Banks) ja polüklonaalseid töörakkude pankasid 5
Working Cell Banks (pWCB – polyclonal Working Cell Bank).
pMCB-d, mis võivad olla kasutatud polüklonaalse produtseeriva rakuliini loomiseks
üheainsa ampulli sisu ülessulatamise ja paljundamisega, võivad olla genereeritud
külmutatud varust, mis koosneb individuaalsetest rakuliinidest. Individuaalsed rakuliinid,
mida on kasutatud selliste pMCB genereerimiseks, on saadud kas i) üksikust kloonist 10
(nagu on kirjeldatud näites 2), ii) üksikute kloonide segust (nagu on kirjeldatud näites 2),
või iii) kloonide ühisvarust (üksikute selektsioonil saadud kolooniate ühisvarust). Need
kloonid saadi peremeesrakkudest, mis olid individuaalselt transfekteeritud, ja selekteeritud
polüklonaalse valgu individuaalse liikme stabiilse ekspressiooni suhtes. Selektsiooni
stabiilse ekspressiooni suhtes teostati sellel alal tuntud meetoditega, näiteks, kasutades 15
selektsiooni markergeene. Käesoleva leiutise eelistatud teostusviisis on individuaalsed
rakuliinid saadud rakkude kloonidest või alakloonidest, näiteks, kasutades punktides i) ii)
või iii) pärit rakuliini lahjendamist või üksiku raku FACS analüüsi ja selektsiooni, või
selekteerides kõrge ekspressiooniga kloone, näiteks kasutades sellist robotit, nagu
ClonePix FL (vaata allpool). Individuaalsed rakuliinid, mida on kasutatud pMCB 20
genereerimisks, nagu eelpool kirjeldatud, võivad olla eelnevalt säilitatud külmutatud
individualsetest rakuliinidest raamatukogu varuna, millest iga individuaalse rakuliini
ampull sulatatakse üles ja paljundatakse enne pMCB genereerimist. Eelistatavalt, need
individuaalsed rakuliinid ekspresseerivad täispikki antikehasid, mille omadused erinevad
nende antikehade omadustest, mis on produtseeritud pMCB teise liikme poolt, näiteks, 25
nad võivad erineda antigeeni spetsiifilisuse poolest, affiinsuse poolest, erinevate variaabel-
või CDR-piirkondade ja/või erinevte konstantsete piirkondade poolest.
Iga rakuliin, mida kasutati pMCB geenreerimiseks, produtseerib polüklonaalse valgu
erinevaid liikmeid. Eelistatavalt seostub polüklonaalse valgu erinev liige konkreetse
antigeeniga. Lisaks sellele on eelistatav, et igat erinevat liiget produtseeritakse mitmest 30
integrandist, mis paiknevad iga peremeesraku genoomi juhuslikes saitides. pMCB
genereeritakse segades kokku ettemääratud arv rakke igast individuaalsest rakuliinist.

EE-EP 2 152 872 B1 37
Eelistatavalt, segatakse need rakud kokku võrdsetes kogustes (suhtes 1:1), kuigi
soovitavad või olla ka teised suhted (vaata allpool). Rakkude segu, mis oli külmutatud
alikvootidena, millena nad on jaotatud suurde hulka viaalidesse, ettemääratud rakkude
arvuga igas viaalis. Need viaalid külmutati ja säilitati pMCB-na hilisemaks kasutamiseks.
Eelistatavalt on nende viaalide arv, mis moodustavad pMCB, suurem, kui 10, 25, 50, 75, 5
100, 200, 500 või 1000 viaali. Üksikud viaalid pMCB-s võivad olla üles sulatatud
erinevatel ajapunktidel, et genereerida erinevaid partiisid polüklonaalseid
produktsioonirakuliine, mis on võimelised produtseerima polüklonaalset valku partiilt
partiile praktiliselt sama kompositsiooniga.
Käesoleva leiutise alternatiivses lähenemisviisis võib polüklonaalne produtseeriv rakuliin 10
olla paljundatud pWCB-st, mis on tuletatud pMCB-st. pWCB genereeritakse sulatades
üksik viaal pMCB-st ja paljundades neid rakke mitmeks genereerimiseks, mis on piisav, et
produtseerida selline arv rakke, mis võib olla külmutatud uues alikvootide seerias
(leiutisekohane pWCB), ligikaudu sama arv rakke igas pWCB alikvoodis, nagu oli pMCB
viaalis, mida on algselt kasutatud pWCB genereerimiseks. Kui pWCB on ammendatud, on 15
võimalik pMCB alikvoodist geenreerida uus pWCB. See lähenemisviis on seetõttu
oluliselt väiksema töömahuga, kui võib vaja minna individuaalse rakuliini paljundamiseks
külmutatud raamatukogu varust ja uue pMCB kokkusegamist. Edasi, juhul, kui pWCB on
ammendatud, siis on suurem võimalus produtseerida järgmised partiid polüklonaalse
töörakuliini rakke, mis on võimeline produtseerima polüklonaalseid valke, ja mis on 20
partiilt partiile praktiliselt sama koostisega.
pMCB produtseerimisel üksikute individuaalse transfektsiooniga saadud rakuliinide
kokkusegamise teel on see eelis, et see annab võimaluse teostada individuaalselt
transfekteeritud rakuliini lisa analüüsi ja selektsiooni enne pMCB genereerimist. See võib
tagada stabiilsema polüklonaalse produktsioonirakuliini, vastab mitmekesisuse nõuetele, 25
mida on juba kirjeldatud. Lisaks polüklonaalne valk võib olla valmistamid enam
reprodutseeritaval teel.
Kõik mis on siin öeldud pMCB kohta, kehtib ka pWCB puhul.
Käesoleva leiutise lisateostusviisis on individuaalseid rakuliine, mis on valitud eelpool
kirjeldatud polüklonaalse valgu individuaalse liikme stabiilseks ekspressiooniks, edasi 30
iseloomustatud nende proliferatsioonikiiruste ja/või produktiivsuse suhtes enne pMCB
genereerimist. Eelistatud teostusviisis on pMCB genereerimiseks valitud rakuliinid, millel

EE-EP 2 152 872 B1 38
on samasugusd proliferatsioonikiirused või produktiivsus. Veelgi enam eelistatavalt,
pMCB genereerimiseks on valitud rakuliinid, millel on samasugune produktiivsus, aga ka
samasugused proliferatsioonikiirused. Eelistatavalt, need rakuliinid on kohandatud
kultiveerimiseks suspensioonina seerumivabas söötmes, enne, kui nende
proliferatsioonikiirusi ja/või produktiivsust iseloomustati. Alternatiivselt on 5
transfektsiooniks kasutatud emarakud kohandatud enne nende transfektsiooni,
kultiveerimisele suspensioonina, seerumivabas söötmes.
Proliferatsiooni kiirused võivad olla hinnatud sellel alal tuntud meetoditega. Imetaja
rakuliinide proliferatsioonikiirused peaksid olema 18 kuni 100 tundi, eelistatavalt 22 kuni
40 tundi ja enim eelistatavalt 24 kuni 32 tundi. Nende rakkude produktiivsus peaks olema 10
suurem, kui 0,5 pg valku raku kohta päevas (pg/(rakk*päev)), eelistatavalt peaks see
ületama 1, 1,5, 3, 5, 8, 10, 15, 20, 30, 40, 50, 75, või 100 pg/(rakk*päev). Edasi, see
rakuliin peaks koosnema ekspressiooni suhtes homogeensete rakkude populatsioonist,
hinnatuna rakusisese värvimismeetodiga. Kui soovitakse homogeensemat
rakupopulatsiooni igale individuaalsele rakuliinile, siis võib see olla saadud 15
kloneerimisega, näiteks, sorteerimisega siin kirjeldatud FACS-sorteerimise meetoditega.
Järgmises käesoleva leiutise teostusviisis sorteeritakse need individuaalsed rakuliinid
FACS-sorteerimismeetodiga, et identifitseerida homogeenne ekspressioonitasemega rakke,
pärast siin kirjeldatud transfektsiooni ja selektsiooni protsessi.
Fluorestsentselt märgistatud antikehasid võib kasutada rakkude sorteerimiseks, mis 20
ekspresseerivad kõrgel tasemel soovitavat valku, näiteks, antikeha või TcR-i, mis
võimaldab luua produktiivsuse suhtes homogeense rakupopulatsiooni. See tehnoloogia
põhineb tähelepanekul, et eritunud valgud võivad olla tuvastatud neid valke eritavate
rakkude pinnal, ja pinnavalgu kogus vastab nähtavasti individuaalse raku
ekspressioonitasemetele. Kõrge produktiivsusega rakkudeks võivad seega olla sorteeritud 25
üksikud märgistatud antikehadega värvitud rakud, millele järgneb analüüs FACS-
meetodiga. Kasutatud tehnoloogiat on kirjeldanud Brezinsky (Brezinsky et al. J. 2003.
Immunol Methods 277, 141-155).
Alternatiivne sorteerimismeetod põhineb rakkude poolt ekspresseeritava valgu suhtes
spetsiifilise ligandi sidestamisel rakkude pinnale. Näiteks Fc-vastane antikeha või 30
idiotüübivastane antikeha või olla sidestatud valku eritava raku pinnaga
rakupopulatsioonis, läbi biotiini. Antikehad, mis on eritatud individuaalsete rakkude poolt,

EE-EP 2 152 872 B1 39
püütakse seejärel kinni Fc-vastaste antikehadega selle raku pinnal. Pärast seda võivad
kõrge produktiivsusega rakud olla sorteeritud FACS abil märgistatud antikehade värvi
põhjal. Seda meetodit on kirjeldatud patendis EP 667 896.
Et saada rakuliinid, millel on homogeensed ja kõrged ekspressioonitasemed, üksikuid
rakke, millel on kõrge ekspressioonitase, analüüsiti põhinedes FACS profiilile, mis on 5
saadud ühe siin kirjeldatud tehnoloogiaga. Seejärel neid individualsete rakkude kloone
paljundati ja, võimalik, analüüsiti proliferatsiooni kiiruste ja produktiivsuse suhtes nii,
nagu on kirjeldatud eelpool. Alternatiivselt, sorteerimisega koguti rakkude alaühisvaru,
millel olid kõige kõrgemad FACS profiili abil identifitseeritud ekspressioonitasemed. See
rakkude alaühisvaru individuaalsest rakuliinist võib samuti soovi korral olla analüüsitud 10
proliferatsioonikiiruse ja produktiivsuse suhtes.
Käesoleva leiutise alternatiivses teostusviisis kasutatakse sellist robotit nagu ClonePixFL
robot (Genetix, UK), et valida kloone, millel on kõrged ekspressioonitasemed ja/või
samasugused kasvuomadused. Seda tehakse järgmiselt: kolooniad, mis on saadud pärast
transfektsiooni ja selektsiooni, kasvatati pooltahkes söötmes, mis võimaldas tuvastada 15
kõrge produktiivsusega kolooniaid, püüdes eritatud valguprodukti koloonia vahetus
läheduses. Iga koloonia produktsioonitase määratakse rakkude poolt ekspresseeritud valgu
immunofluorestsentsmärgistuse abil, millele järgnes parimate kloonide selektsioon,
kasutades kujutise tarkvara selliste ettemääratud kriteeriumide põhjal, nagu ekspressiooni
tase ja kasvuomadused. Lisaks sellele, iga koloonia suurus (mis peegeldab selle raku 20
proliferatsioonikiirust) võib olla hinnatud selle robotiga, kasutades valgusega tuvastatavat
visualiseerimist (light detection imaging). Seejärel, kolooniad, millel on soovitavad
produktiivsuse ja/või kasvuomadused, eraldatakse robotiga ja kantakse üle 96-
süvikulistele plaatidele nende eadasiseks paljundamiseks.
Eelistatavalt, valitakse pMCB genereerimiseks välja samasuguse produktiivsusega 25
individuaalsed rakuliinid. Eelistatud teostusviisis, individuaalsed rakuliinid, mis
moodustavad pMCB, genereeritakse kloneeritud rakkudest, näiteks nendest, mis on
saadud üksiku raku kaupa sorteerimisega, piiratud lahjendamise või robotiga
väljanoppimisega, kõrge ekspressioonitasemega rakkudest või kõrge
ekspressioonitasemega rakkude ühisvarust. 30
Käesolevas leiutises, nii individuaalsed rakuliinid, mis saadi üksikust rakkude kolooniast,
mis eraldati pärast transfektsiooni ja selektsiooni, kui ka individuaalsed rakuliinid, mis on

EE-EP 2 152 872 B1 40
saadi kloonist, mis on saadud, näiteks, ühe raku kaupa FACS-sorteerimisel, nimetatakse
kloneeritud rakuliinideks. Eelistatud teostusviisis selliseid kloneeritud rakuliine
kasutatakse pMCB genereerimiseks.
Käesoleva leiutise järgmises teostusviisis, pMCB genereerimisel need individuaalsed
rakuliinid segatakse kokku erinevates proportsioonides. Need üksikud rakuliinid võivad 5
olla segatud vastavalt ettemääratud kriteeriumidele, mis põhinevad üksikute rakuliinide
omadustel ja/või sõltuvalt nimetatud rakuliini poolt ekspresseeritud individuaalse
valguprodukti omadustest, näiteks, spetsiifilisest produktiivsusest või seostumise
afiinsusest.Näiteks, eraldi rakuliinid, mis ekspresseerivad teatud antikehasid, mis
seostuvad eriti tähtsate antigeenide või epitoopidega, võivad olla varutud liiaga, võrreldes 10
teiste pMCB rakuliini liikmetega, näiteks, 2-kordselt, 3-kordselt, 5- kordselt või 10-
kordselt suuremates kogustes. Üks rakuliini liige võib olla näiteks lisatud suhtes 2:1
võrreldes kõigi teiste liikmetega, näiteks 1. liikme rakke on 4 x 106 ja igast ülejäänud
liikme rakuliinist 2 x 106 rakku.
See lähenemisviis diferentseeritud individuaalsete rakuliinide proportsioonidega pMCB-s 15
võib samuti olla tarvitusele võetud, et vältida erinevusi individuaalsete rakuliinide
proliferatsioonikiirustes ja produktiivsustes, eriti, kui nad ei olnud välja valitud just nende
tunnusjoonte põhjal. Järelikult, kui üks või enam individuaalset rakuliini on madalama
proliferatsioonikiirusega, s.o pikema kahekordistumise ajaga, võrreldes teiste
polüklonaalse tööraku panga liikmetega, mida iseloomustab suurem proliferatsioonikiirus, 20
kuid see madalam proliferatsioonikiirus ei ole seotud konkreetse kõrge produktiivsusega,
see (need) konkreetne(sed) liige(kmed) või(vad) olla lisatud pMCB-le suuremates
kogustes, kompenseerimaks nende aeglast kasvu. Näiteks, rakuliin
proliferatsioonikiirusega 50 tundi, võib olla lisatud proportsioonides 2:1, kui ülejäänud
rakuliinide, mis moodustavad pMCB, proliferatsioonikiirused jäävad vahemikku 22 ja 30 25
tundi. Peale selle, lühikeste kahekordistumise aegadega rakuliinide suhe võib olla
vähendatud, et tagada, et need ei saaks prevaleerima tootmise käigus. Peale selle, erinevate
individuaalsete rakuliinide suhe pMCB-s võib olla korrigeeritud pMCB-st genereeritud
polüklonaalsete tootmisrakuliinide poolt produtseertavate polüklonaalsete valguproduktide
analüüsil. Selline korrigeerimine võib, näiteks, olla tehtud, põhinedes IEX profiilidele või 30
ekvivalentsetele iseloomustamise vahenditele. Kui selline analüüs näitab, et ühte või
enamat konkreetset valguliiget produtseeritakse suurendatud kogustes võrreldes ülejäänud
liikmetega, siis võib olla genereeritud uus pMCB, milles nende, liigset valku

EE-EP 2 152 872 B1 41
produtseerivate rakuliinides, osa võib olla vähendatud. Ja vastupidi, kui see või teine
konkreetne liige produtseeritakse väikestes kogustes, siis võib olla genereeritud pMCB,
milles on seda liiget produtseeriva rakuliini osa on suurendatud.
Kultiveerimise süsteemid
Käesoleva leiutise kohane rekombinantne polüklonaalne valk võib olla valmistatud 5
kasutades ükskõik millist sobivat kultivatsiooniviisi, mille hulka kuuluvad, kuid ei ole
nendega piiratud partii kaupa teostatavad (bach), poolpidevad (fed-batch-) ja
perfusiooniprotsessid.
Ekspressioonisüsteemi loomine kõrgetasemeliseks valkude ekspressiooniks
Meetodid nukleiinhappejärjestuse sisestamiseks rakku on teada. Need meetodid sisaldavad 10
tüüpiliselt DNA vektori kasutamist huvipakkuva järjestuse sisestamiseks rakku, selle
genoomi või kromosoomivälisesse elementi. Rakkude transfektsioon võib olla teostatud
suure hulga selle ala spetsialistile teada olevate meetoditega, kaasaarvatud lipofektsioon,
keemiliselt vahendatud transfektsioon, sadestamine kaltsiumfosfaadiga, elektroporatsioon,
mikroinjektsioon, liposoomi fusioon, punaste vererakkude varjude fusioon (RBC ghost 15
fusion), protoplasti fusioon, viiruse transduktsioon ja muu taoline.
Peremeesrakuliini transfektsiooniks kasutatakse huvipakkuvate vektorite raamatukogu,
milles iga vektor sisaldab ainult ühte nukleiinhappe järjestuse koopiat, mis kodeerib
huvipakkuva rekombinantse polüklonaalse valgu ühte liiget. See huvipakkuvate
ekspressioonivektorite raamatukogu kodeerib kollektiivselt huvipakkuvat rekombinantset 20
polüklonaalset valku. Integratsiooniks sobivaid vektoreid on kirjeldatud eelmises lõigus.
Rekombinantse polüklonaalse produtseeriva rakuliini genereerimine ja rekombinantse
polüklonaalse valgu produktsioon sellisest rakuliinist võib olla saadud mitmete erinevate
transfektsiooni ja valmistamise strateegiatega.
Eelistatud transfektsiooniviis, mida illustreerib joonis fig 1, on suure jõudlusega meetod, 25
milles peremeesrakud transfekteeritakse eraldi, kasutades individuaalseid vektoreid, mis
moodustavad huvipakkuvate vektorite raamatukogu. Seda meetodit nimetatakse
individuaalseks transfektsiooniks. Need individuaalselt transfekteeritud peremeesrakud on
eelistatavalt eraldi valitud. Kuid nad võivad olla ka kogutud varuks enne selektsiooni.
Individuaalsete rakkude kloonid, mis genereeritakse selektsiooni käigus, võivad olla 30
analüüsitud ekspressioonitaseme, proliferatsioonikiiruse ja integratsioonipildi suhtes, ja
eelistatavalt need ühesuguste kasvu kiirustega, ühesuguse koopiate arvuga, ühesuguse

EE-EP 2 152 872 B1 42
ekspressiooni ja/või ühesuguse töökindluse tasemetega kloonid võivad olla kasutatud
polüklonaalse GOI raamatukogude varu geenreerimiseks. Individuaalsed rakukloonid
võivad olla kokku segatud, et saada soovitav polüklonaalne rakuliin enne varu
genereerimist, vahetult pärast seda, nad saadakse varust, või pärast lühikest
proliferatsiooni ja adaptatsiooniperioodi. See lähenemisviis võib veelgi parandada 5
kompositsioonilist stabiilsust.
Transfektsiooni asjaloudes, mis võimaldavad enam, kui ühe koopia integratsiooni igasse
rakku, võib olla teostatud kõigi rakkude transfektsioon, kasutades ekspressioonivektorite
segusid, kui selleks polüklonaalseks valguks on monomer. Multimeersevalgu puhul,
selline kõigi rakkude transfektsioon, mis võimaldab mitmekordset integratsiooni 10
peremeesraku genoomi, põhjustab subühikute ”segipaiskamist”. Paljudel juhtudel, nagu
näiteks rekombinantse polüklonaalse antikeha valmistamisel farmatseutiliseks
kasutamiseks, peab selline ”segipaiskamine” olema välistatud. Multimeersete valkude
puhul võib kõigi rakkude transfektsioon olla teostatud, kui ”segipaiskamine” on
vastuvõetav, või kui transfektsioon on läbi viidud tingimustes, mis tagavad ainult ühe 15
koopia integratsiooni peremeesraku genoomi. Selliste meetodite näidete hulka kuuluvad
retroviiruslik transduktsioon ja sferoblasti fusioon. Individuaalsed vektorid, mis
moodustavad huvipakkuvate variantnukleiinhappejärjestuste raamatukogu, võib olla
kokku segatud ühte ainsasse kompositsiooni, või eelistatavalt, võivad individuaalsed
vektorid, mis kodeerivad iga raamatukogu liiget, olla hoitud eraldi kompositsioonides, või 20
segus, mis koosneb ligikaudu 5 kuni 50 selle raamatukogu individuaalsest vektorist.
Teiseks mooduseks on vektorite raamatukogu fraktsioonideks jaotamise kasutamine, kus
need fraktsioonid sisaldavad transfektsioonis ligikaudu 5 kuni 50 individuaalset
raamatukogusse kuuluvat vektorit. Eelistatavalt sisaldab see raamatukogu fraktsioon 10
kuni 20 individuaalset vektorit. Iga kompositsioon transfekteeritakse seejärel 25
peremeesrakkude alikvootidesse. Seda meetodit nimetatakse semi-bulk transfektsiooniks.
Transfekteeritud alikvootide arv sõltub raamatukogu suurusest ja individuaalsete vektorite
arvust igas fraktsioonis. Näiteks, kui see raamatukogu koosneb 100 erinevast
variantliikmest, mis on jagatud fraktsioonideks, mis sisaldavad kompositsioonis 20
erinevat variantliiget, siis on vaja, et 5 alikvooti peremeesrakke oleks transfekteeritud 30
raamatukogu kompositsioonga, mis on moodustatud algse raamatukogu erinevatest
frakstioonidest. Peremeesrakkude alikvoodid on valitud pärast transfektsiooni.
Eelistatavalt, need erinevad alikvoodid valitakse eraldi. Kuigi, nad võivad olla ühisvarusse

EE-EP 2 152 872 B1 43
kogutud ka enne selektsiooni. Nimetatudd alikvoodid võivad olla analüüsitud nende
klonaalse mitmekesisuse suhtes ja polüklonaalse GOI raamatukogu varu genereerimiseks
kasutatakse ainult neid alikvoote, millele on omane piisav mitmekesisus.
Külmutatud polüklonaalse rakuliini varu või olla genereeritud enne rekombinantse
polüklonaalse valgu valmistamise initsieerimist. Et saada produtseerimiseks soovitav 5
polüklonaal rakuliin, need kloonid võivad olla enne külmutatud varu genereerimist
segatud, vahetult pärast seda, kui olid välja võetud varust, või pärast lühikest
proliferatsiooni ja adaptatsiooniaega.
Ühiseks tunnusjooneks eelpool kirjeldatud tootmisstrateegiatel on see, et kõik
individuaalsed liikmed, mis moodustavad rekombinantse polüklonaalse valgu, võivad olla 10
produtseeritud ühes või piiratud koguses konteinerites, näiteks bioreaktoris, mida on
maksimaalselt ligikaudu 10.
Kui ekspressioonitasemeid on vaja suurendada, siis geeni amplifikatsioon või olla
teostatud, kasutades selektsiooni DHFR geeni suhtes või glutamiinsüntetaasi (GS) geeni
suhtes, hprt (hüpoksantiinfosfooribosüültransferaasi) või trüptofaansntetaasi geeni suhtes. 15
See vajab niisuguste vektorite kasutamist, mis sisaldavad sellist selektsioonimarkerit.
Üheks käesoleva leiutise konkreetseks tunnusjooneks on see, et rakkude stabiilsuse
hoidmiseks kõrgel tasemel on vaja koopiate arvu hoida suhteliselt madalana. Seetõttu viidi
eelistatavalt läbi rakkude üks selektsioonitsükkel suhteliselt tagasihoidliku
selektsioonisurve all nukleosiidivabas söötmes, mis sisaldas madalat MTX (nt, 1-10 nM) 20
kontsentratsiooni konstrukti tüübi puhul, mida kasutati lisatud näidetes. Arvatakse, et
suhteliselt madala arvu koopiate põhjuseks on selline tagasihoidlik selektsioonirõhk.
Arvatakse, et mõõdukas selektsioonisurve põhjustab balanseeritud koopiate arvu, mis
annab tulemusena kõrge ekspressioonitaseme, hoides ära nende rakkude ebastabiilsuse,
millel on väga suur koopiate arv. 25
Selleks, et saavutada kõrgemaid ekspressioonitasemeid, ekspressiooniks kasutatav rakuliin
sisaldab eelistatavalt heteroloogset transaktivaatorit, mis on võimeline tõstma
polüklonaalse valgu promootoriga kontrollitavat ekspressiooni. Sobivate transaktivaatori
ja promootori kombinatsioonide näiteid on mainitud järgnevas tabelis (tabel 2).
30

EE-EP 2 152 872 B1 44
Tabel 2. Näited transaktivaatori/promootori paaridest
Transaktivaator Promootori näited Kommentaarid
lentiviirus Tat pikk lõpukordus (LTR)
adenoviirus E1A HCMV peamine IE
võimendusjärjestus/promootor
herpes simpleksi viirus VP16 herpes simpleksi viiruse geeni
promootoriks on IE175
US 6635478
hepatiidi B viiruse X valk (HBx) SV40 varajane
Sünteetilise Zn-sõrme valgud Sünteetiline Sangamo inc
SV40 suur antigeen SV40 hiline promootor
tetratsükliiniga-kontrollitavad
transaktivaatorid (tTA)
Sünteetiline Sünteetilised
fusioonid
inimese tsütomegaloviirus IE2p86 HCMV peamine IE
võimendusjärjestus/promootor
inimese tsütomegaloviirus IE1p72 HCMV peamine IE
võimendusjärjestus/promootor
Epstein-Barri viiruse R
transaktivaator (Rta)
EBV promootor
kilpnäärme hormooni retseptorid kasvuhormooni promootor
glükokortikoidhormooni retseptorid piimanäärme tuumori viiruse
(MMTV) promootor
Eelistatavalt on rakuliin transfekteeritud ekspressioonikonstruktiga, mis kodeerib
transaktivaatorit, ja kloonid on valitud, kasutades piiratud lahjendust või teist meetodit
üksiku raku kloneerimiseks. Ekspressioonivektor võib sisaldada selliseid elemente, nagu 5
promootor, selektsioonimarker jne, nii nagu seda on siin kirjeldatud seoses

EE-EP 2 152 872 B1 45
ekspressioonivektoritega. Eelistatavalt on promootor, mis kontrollib transaktivaatori
ekspressiooni, selline konstitutiivne promootor, nagu elongatsioonifaktori 1 promootor,
CMV promootor, metallotioneiin-1 promootor või analoogne sellega. Eelistatud
teostusviisis on selleks promootoriks CMV promootor.
Eriti eelistatud on kasutada adenoviiruse E1A transaktivaatorit, mis paistab stabiliseerivat 5
rakke iseenesest, lisaks CMV promootori transaktiveerimisele, mis kontrollib huvipakkuva
geeni ekspressiooni. Nagu on juba märgitud, E1A ekspressioon ei olnud tuvastatav
esimese produtseeriva rakuliini, ECHO puhul. Seetõttu side E1A ja rakkude
stabilisastiooni vahel ei ole käesolevates eksperimentides tõestatud.
Polüklonaalse valgu valmistamiseks, kus valgu iga liige sisaldab enam, kui kahte 10
polüpeptiidahelat, nende ahelate kombinatsioon võib olla tähtis selle valgu afiinsuse,
spetsiifilisuse ja aktiivsuse seisukohalt, mille nad moodustavad. Seda võib näha, näiteks
antikehade ja TcR-de peal. Näiteks, on teada, et antikeha raske ahela variaabelpiirkonna ja
kerge ahela variaabelpiirkonna kombinatsioon mõjutab nende ahelate poolt moodustatud
antikeha afiinsust ja spetsiifilisust. Seega, kui antikeha raamatukogu, mis kodeerib 15
järjestusi, on valitud nende võime järgi produtseerida antikehi, afiinsusega teatud
märklaua suhtes, siis on soovitav tagada, et see raske ahela variaabelpiirkonna ja kerge
ahela variaabelpiirkonna kombinatsioon lõpp-produktis ka vastab sellele. Sel põjusel on
polüpeptiidahelad, mis moodustavad polüklonaalse valgu individuaalse liikme,
eelistatavalt paigutatud samasse vektorisse, mida on kasutatud integratsiooniks, tagades 20
sellega, et nad saavad olema koos kogu protsessi jooksul. Alternatiivselt, peremeesrakud
võivad olla transfekteeritud ekspressioonivektorite paaridega, mis kodeerivad raske ja
kerge ahela suguluspaare.
Järgmine kirjeldus on üheks näiteks sellest, kuidas saada rekombinantset polüklonaalset
antikeha ekspresseeriv rakuliin. 25
Võib olla konstrueeritud universaalne promootorikassett, mis on ette nähtud
konstitutiivseks ekspressiooniks, milles on kaks promootorit, mis on paigutatud
vastupidistesse transkriptsiooni suundadesse, näiteks pea-pea(head-to-head)-
konstruktsioonina, mis on ümbritsetud raske ahela variaabelpiirkonna ja terve kappa või
lambda kerge ahelaga, võimaldades kogu konstrukti ülekannet vektorisse, mis sisaldab 30
selektsioonimarkerit ja raske ahela konstantset piirkonda. On kaalutud, kas
indutseeritavaks ekspressiooniks võib olla kasutatud ka promootorkassett. Lisaks sellele,

EE-EP 2 152 872 B1 46
need promootorid võivad olla paigutatud saba-saba (tall-to-tall) orientatsioonis, mis
põhjustab transkriptsiooni vastassuundades, või saba-pea (tall-to-head) orientatsioonis,
ühesuunaliseks transkriptsiooniks. Ekspressiooni kontrolliks võib olla kasutatud ka
indutseeritav promootor. Pärast transfektsiooni, neid rakke kultiveeriti eelistatavalt
selektiivsetes tingimustes, et välja valida stabiilseid tranformante. 5
Rakud, mis jäävad ellu nendes tingimustes, võivad järgnevalt kasvada erinevates
kultuurisüsteemides, näiteks sellistes, nagu traditsionaalsed väikesed kultuurikolvid,
Nunci mitmekihilised rakuvabrikud, väikesed, suure saagisega bioreaktorid (MiniPerm,
INTEGRA-CELLLine), loksutajad, ja segurkolvidest kuni kiud- ja bioreaktorite WAVE
kottideni (Wave Biotech, Tagelswangen, Switzerland) või teiste ühekordsete 10
nõude/konteineriteni. Need rakud võivad olla testitud antikeha produktsiooni suhtes,
kasutades ELISA-testi. Eelistatavalt valitakse polüklonaalsed rakuliinid, mis on
elujõulisused kultiveerimisel suspensioonis, seerumivabas söötmes, selektsiooni survel,
pikendatud ajavahemike jooksul.
Polüklonaalsuse säilitamise hindamine ekspressioonisüsteemis. 15
Vastavalt käesolevale leiutisele on sageli tähtis kindlustada, et polüklonaalsus
ekspressioonisüsteemis ei muutuks tõsiselt produktsiooni kestel nii, et oleks võimalik
peatada produktsioon, kui polüklonaalsus tõepoolest on muutunud. Seda tehakse vastavalt
käesolevale leiutisele jälgides variantnukleiinhappe järjestuste suhtelisi
ekspressioonitasemeid. Neid ekspressioonitasemeid võib, näiteks, jälgida mRNA tasemete 20
kaudu, kasutades näiteks RFLP- analüüsi, maatriks- või reaalaja PCR-i, või valgu taseme
kaudu, kasutades, näiteks, kahedimensionaalset polüakrüülamiidgeelelektroforeesi, mass-
spektromeetriat või erinevaid kromatograafiameetodeid. Nende meetoditega on võimalik
kindlaks teha kõikide individuaalsete ekspressioonitasemete baasväärtused ja seejärel
võtta välja proovid kultuurist produktsiooni kestel selleks, et hinnata, kas 25
ekspressioonitasemed on muutunud (nii üldine, kui ka suhteline). Käesoleva leiutise
tavalises praktikas võib baasväärtustele lähedaste väärtuste diapasoon olla fikseeritud, ja
kui on leitud, et suhtelised ekspressioonitasemed väljuvad nendest piiridest, siis
produktsioon lõpetatakse.
Rakkude kultivatsioon ja rekombinantse polüklonaalse antikeha produtseerimine 30
Leiutisekohane polüklonaalne rakuliin valmistati nii, nagu eelpool kirjeldatud, ja see või
olla kultiveeritud sobivas söötmes ning tingimustes, mis sobivad huvipakkuva, rakkude

EE-EP 2 152 872 B1 47
genoomi sisestatud variantnukleiinhappe järjestuses kodeeritud polüklonaalse valgu
ekspresseerimiseks. Nende rakkude kultiveerimine võib olla teostatud mitmes etapis. Kui
kasutatakse imetajarakke, siis valitud rakud on eelistatavalt adapteeritud kultiveerimiseks
suspensioonis, aga ka seerumivabades tingimustes. Adaptatsioon kultiveerimiseks
seerumivabas söötmes võib eelistatavalt olla teostatud ka enne kloneeritud rakuliinide 5
segamist polüklonaalse rakuliini saamiseks. Adaptatsioon või olla teostatud ühes või kahes
etapis ja selektsiooni survel või ilma selleta. Eelistatavalt kasutatakse sellist
selektsioonisüsteemi, mis võimaldab teostada selektsiooni kogu produktsiooniperioodi
vältel, kahjustamata valmistava raviprodukti puhtust. Üldiselt, farmatseutiliseks
kasutamiseks ette nähtud rekombinantsete valkude valmistamisel on eelistatud mitte 10
kasutada selektsioonisurveks, näiteks, antibiootikume või teisi madalmolekulaarseid
ravimeid, kuna siis on tarvis kinnitust, et lõpp-produkt ei sisalda mis tahes antibiootikumi
jälgi.
Kui polüklonaalne rakuliin on juba kohandatud vastavate tingimustega, siis võib alustada
produtseemimise mõõtkava suurendamisega. Selles punktis võib polüklonaalsete 15
töörakkude varu ( pWCB) ja/või polüklonaalne baasrakkude pank (pMCB) olla
külmutatud. Eelistatavalt võivad olla kasutatud bioreaktorid, mahuga 30 kuni 100 liitrit,
kuid väiksemad (5-10 liitrit) või suuremad (kuni 1000, 5000, 10000, 15000 liitrit, või
veelgi suuremad) bioreaktorid võivad samuti olla kasutatud. Sobiv produktsiooniaeg ja
bioreaktori suuruse valik sõltuvad valgu soovitavast saagisest partiis ja rakuliini 20
ekspressioonitasemetest. Ajad võivad varieerida paarist päevast kuni kolme kuuni.
Ekspresseeritud rekombinantne polüklonaalne valk võib olla kogutud kas rakkudest või
rakukultuuri supernatandist. See rekombinantne valk või olla puhastatud iseloomustatud
selle ala spetsialistile tuntud meetoditega. Näiteid puhastamise metoodikatest on loetletud
allpool. Näiteid iseloomustamise metoodikatest võib leida näiteks, publikatsioonis WO 25
2006/007853.
Rekombinantse polüklonaalse valgu eraldamine ja puhastamine kultuuri
supernatandist
Spetsiifiliste valkude eraldami kultuuri supernatantidest on võimalik, kasutades erinevaid
kromatograafiameetodeid, milles kasutatakse ära valkude erinevusi füüsikalis-keemilistes 30
omadustes, näiteks, erinevusi molekulkaaludes, summaarses laengus, hüdrofoobsuses või
afiinsuses spetsiifilise ligandi või valgu suhtes. Seega, valgud võivad olla eraldatud
vastavalt molekulkaaludele, kasutades geelfiltratsioonkromatograafiat, või vastavalt

EE-EP 2 152 872 B1 48
summaarsele laengule, kasutades ioonvahetus(katioon/anioon)kromatograafiat või
alternatiivselt, kasutades kromatofookustamist. Analoogselt sellele võivad valgud olla
eraldatud vastavalt nende hüdrofoobsusele, kasutades hüdrofoobseid interaktsioone või
laengu indutseerimisel põhinevat kromatograafiat või afiinsuskromatograafiat, milles
kasutatakse afiinsuse erinevusi spetsiifiliste immobiliseeritud ligandide või valkude 5
suhtes. Keeruliste valkude segude eraldamine võib seega olla saavutatud kasutades
järjestikku erinevate kromatograafiliste meetodite kombinatsiooni. Valkude segu võib
seega olla eraldatud algselt, näiteks, vastavalt selle molekulide summaarsele laengule
ioonvahetuskromatograafiaga, ja valgud, millel on ühesugune laeng, võivad olla edasi,
lahutatud vastavalt molekulkaalule, kasutades geelfiltratsioonkromatograafiat või 10
kasutades hüdrofoobsetel interaktsioonidel põhinevat kromatograafiat valitud
soolakontsentratsioonide juuresolekul.
Afiinsuskromatograafiat, kombinatsioonis selliste järgnevate puhastamisetappidega, nagu
ioonvahetuskromatograafia, hüdrofoobsed interaktsioonid ja geelfiltratsion, kasutatakse
sageli IgG (polüklonaalsete aga ka monoklonaalsete) ja TcR eraldamiseks ja 15
puhastamiseks erinevatest allikatest, näiteks, astsiidivedelikest, rakukultuuri
supernatantidest ja seerumist. Afiinne puhastamine, kus eraldamine põhineb valgu(kude)
ja kromatograafilisele maatriksile sidestatud spetsiifilise ligandi vahelisel pöörduval
interaktsioonil, on lihtne ja kiire meetod, mis võimaldab saavutada suurt selektiivsust,
tavaliselt suurt jõudlust ja kontsentratsiooni väiksemas mahus. Kahel bakteriaalse 20
rakupinna valgul, valgul A ja valgul G, on suur afiinsus antikehade Fc piirkonna suhtes, ja
seda kasutatakse immobiliseeritud kujul tavalistel, väga erinevatel rakendustel,
kaasaarvatud polüklonaalse IgG ja selle alaklasside puhastamine erinevatest molekulidest
ja immuunsete kompleksite absorptsioon ja puhastamine.
Afiinsuskromatograafiale järgnevad kromatograafiaetapid, milleks on näiteks ioonvahetus 25
ja/või hüdrofoobsetel interaktsioonidel põhinev kromatograafia, võivad olla teostatud
peremeesraku valkude, lekkinud valgu A ja DNA eemaldamiseks.
Geelfiltratsioon, kui lõplik puhastamise etapp, võib olla kasutatud selliste saastavate
lisandimolekulide, nagu dimeerid ja teised agregaadid, eemaldamiseks, ja proovi
ülekandeks säilituspuhvrisse. Sõltuvalt allikast ja ekspressioonitingimustest võib olla 30
vajalik sisse lülitada lisa puhastamise etappe, et saavutada antikeha nõutav puhtuse tase.
Seega terapeutiliseks kasutuseks ette nähtud antikehade puhastamisel on tihti
kasutatavateks meetoditeks, kombinatsioonis valgu A kromatograafia ja

EE-EP 2 152 872 B1 49
gelfiltratsioonkromatograafiaga, hüdrofoobsel interaktsioonil põhinev kromatograafia või
ioonvahetuskromatograafia.
Selleks, et lihtsustada puhastamist, on eelistatav, et kõigil polüklonaalse antikeha liikmetel
oleks sama raske ja/või kerge ahela konstantne piirkond.
Selleks, et puhastada teisi antikehade klasse, kasutatakse alternatiivseid 5
afiinsuskromatograafia meetodeid, kuna valgud A ja G ei seostu antikehadega IgA ja IgM.
Kasutatud võivad olla immunoaffiinsed puhastamismeetodid (tahke faasiga sidestatud
anti-IgA või anti-IgM monoklonaalsed antikehad) või, alternatiivselt võivad olla kasutatud
mitmeetapilise puhastamise strateegiad, kaasaarvatud ioonvahetus ja hüdrofoobsel
interaktsioonil põhinev kromatograafia. 10
Struktuurne iseloomustus
Selliste polüklonaalsete valkude, nagu antikehad ja TcR-d, struktuurne iseloomustus, vajab
suurt lahutusvõimet seoses nende segu keerukusega (klonaalne mitmekesisus ja
glükosüleerimine). Traditsioonilised lähenemisviisid, nagu näiteks geelfiltratsioon,
ioonvahetuskromatograafia või elektroforees, ei võimalda saavutada piisavat 15
lahutusvõimet, et eraldada teineteisest individuaalse antikehad. Komplekssete valgusegude
analüüsil on kasutatud kahedimensionaalset elektroforeesi polüakrüülamiidgeelis (2D-
PAGE), millele järgneb mass- spectromeetria (MS) või vedelikkromatograafia (LC)-MS
(näiteks, proteoomika). 2D-PAGE, milles on kombineeritud valgu lahutus algul selle
molekulide laengute põhjal, seejärel aga masside põhjal, on tõestanud oma kasulikkust 20
seerumiproovides sisalduvate polüklonaalsete, oligoklonaalsete ja monoklonaalsete
immunoglobuliinide diferentseerimisel. Kuigi, ka sellel meetodil on mõningad piirangud.
Üha enam leiavad keerukate peptiidisegude analüüsil rakendust kromatograafilised
meetodid, konkreetselt, kapillaar- ja vedelikkromatograafia, kombinatsioonis
elektrospreiionisataioon-MS-ga. LC-MS on kasutatud monoklonaalsete antikehade 25
iseloomustamiseks. Väga keerukate proovide analüüs vajab suurema lahutusvõimega
kromatograafilist süsteemi, mis võib olla saadud lahutamisel kahes (või enamas)
dimensioonis. Selline lähenemisviis võiks põhineda ioonvahetusel esimese mõõtmes ja
pöördfaasi kromatograafial (või hüdrofoobsel interaktsioonil) teises mõõtmes, valikuliselt
ühenduses MS-ga. 30

EE-EP 2 152 872 B1 50
Funktsionaalne iseloomustus
Polüklonaalne valk võib näiteks olla iseloomustatud funktsionaalselt, võrdlevates
uuringutes polüklonaalsete valkudega, millel on spetsiifilisus mõningate samasuguste
märklaudade suhtes, või millel on analoogne aktiivsus. Sellised uuringud võivad olla
teostatud in vitro, aga ka in vivo. 5
Polüklonaalse antikeha funktsionaalseks iseloomustamiseks in vitro võiks näiteks olla
kasutatud immunopretsipitatsioon, mis on väga spetsiifiline meetod märklaudantigeenide
analüütiliseks eraldamiseks töötlemata rakulüsaatidest. Kombineerides
immunopretsipitatsiooni teise, sellise meetodiga, nagu SDS-PAGE, millele järgneb valgu
värvimine (Coomassie Blue, värvimine hõbedaga või märgistamine biotiiniga) ja/või 10
immunoblotting, on võimalik tuvastada ja kvantifitseerida antigeene, näiteks, ja seega
hinnata antikehade mõningaid funktsionaalseid omadusi. Kuigi, see meetod ei anna hinnet
ei antikehamolekulide arvule ega nende seostumise afiinsusele, kuid annab võimaluse
visualiseerida märklaudvalke ja seega ka nende spetsiifilisust. See meetod võib samuti olla
kasutatud selleks, et jälgida antikehade potentiaalseid erinevusi antigeenivastastes 15
omadustes (klonaalse mitmekesisuse terviklikkust) ekspressiooniprotsessi kestel.
Polüklonaalse antikeha in vivo funktsionaalne iseloomustamine võiks olla läbi viidud
uurides infektsioone. Sellised eksperimentaalsed loomad, nagu hiir, võivad, näiteks, olla
nakatatud konkreetse viirusega, mille vastu polüklonaalne antikeha on välja töötatud.
Uuritava polüklonaalse antikeha funktsionaalsust näitab see, millisel määral nimetatud 20
infektsioon saab olema maha surutud.
Terapeutilised kompositsioonid
Farmatseutiline kompositsioon, mis on valmistatud vastavalt käesolevale leiutisele,
sisaldab rekombinantset polüklonaalset valku, mis on valitud immunoglobuliinide
sugukonnast, kuna see on toimeaine, mis on ette nähtud haiguse raviks või ärahoidmiseks 25
imetajas.
See farmatseutiline kompositsioon sisaldab toimeainena eelistatavalt rekombinantset
polüklonaalset antikeha või selle antikeha fragmenti, ja farmatseutiliselt vastuvõetavat
täidisainet.
Eelistatud on ka farmatseutiline kompositsioon, mis sisaldab toimeainena rekombinantset 30
polüklonaalset T-raku retseptorit või T-raku retseptori fragmenti ja farmatseutiliselt
vastuvõetavat täidisainet.

EE-EP 2 152 872 B1 51
Leiutisekohased farmatseutilised kompositsioonid valmistatakse tuntud viisil, näiteks,
kasutades traditsioonilise lahustamise, külmkuivatamise, segamise, granuleerimise või
kokkusegamise protsesse. See farmatseutiline kompositsioon võib olla formuleeritud
vastavalt traditsioonilisele farmatseutilisele praktikale (vaata, näiteks, käsiraamatus
Remington: The Science ja Practice of Pharmacy (20th ed.), ed. A.R. Gennaro, 2000, 5
Lippincott Williams & Wilkins, Philadelphia, PA ja Encyclopedia of Pharmaceutical
Technology, eds. J. Swarbrick ja J. C. Boylan, 1988-1999, Marcel Dekker, New York,
NY).
Lüofiliseeritud kompositsioonide valmistamisel, mis on ette nähtud toimeaine lahuste või
suspensioonide valmistmiseks vahetult enne nende kasutamist, kasutatakse eelistatavalt 10
selle toimeaine lahuseid ja suspensioone, ning eriti isotoonilisi vesilahuseid või
suspensioone, mis võivad sisaldadada peale toimeaine ka kandjat, näiteks mannitooli. See
farmatseutiline kompositsioon võib olla steriliseeritud ja/või võib sisaldada täidisaineid,
näiteks säilitusaineid, stabilisaatoreid, niiskusesäilitajaid ja/või emulgeerivaid agenseid,
solubilisaatoreid, soolasid osmootilise rõhu reguleerimiseks ja/või puhvreid ja 15
valmistatakse tuntud viisil, näiteks, kasutades traditsioonilisi lahustamise või
külmkuivatamise protsesse. Nimetatud lahused või suspensioonid võivad sisaldada
viskoossust suurendavaid aineid, näiteks selliseid nagu naatriumkarboksümetüültselluloos,
karboksümetüültselluloos, dekstraan, polüvinüülpürrolidoon või želatiin.
Leiutisekohased injektsiooniks ette nähtud kompositsioonid valmistatakse tavalisel viisil, 20
steriilsetes tingimustes; sama käib ka nende kompositsioonide ampullidesse või
viaalidesse sisestamise ja nende mahutite hemetiseerimise kohta.
Leiutisekohased farmatseutilised kompositsioonid võivad sisaldada alates ligikaudu 1%
kuni ligikaudu 95%, eelistatavalt alates ligikaudu 20% kuni ligikaudu 90% toimeainet.
Farmatseutilised kompositsioonid võivad olla, näiteks, ühikdoosi kujul, näiteks ampullide, 25
viaalide, suposiitide, dražeede, tablettide või kapslite kujul.
Leiutisekohaste kompositsioonide terapeutilised kasutused
Farmatseutiline kompositsioon, mis on valmistatud vastavalt käesolevale leiutisele, võib
olla kasutatud haiguse raviks, leevendamiseks või ärahoidmiseks imetajas. Haiguste hulka,
mis võivad olla ravitud käesoleva farmatseutilise kompositsiooniga, kuuluvad vähk, 30
nakkushaigused, põletikulised haigused, allergia, astma ja teised respiratoorsed haigused,
autoimmunsed haigused, kardiovaskulaarsed haigused, kesknärvisüsteemi haigused,

EE-EP 2 152 872 B1 52
metaboolsed ja endokriinsed haigused, transplantaadi hülgamisreaktsioon ja ebasoovitav
rasedus.
Leiutisekohased kompositsioonid võivad olla kasutatud meetodis, mis on ette nähtud
haiguse raviks, leevendamiseks või profülaktikaks loomal, mille puhul sellele
manustatakse efektiivne kogus rekombinantset polüklonaalset antikeha või selle antikeha 5
fragmenti, või milles manustatakse efektiivne kogus rekombinantset polüklonaalset T-raku
retseptorit või T-raku retseptori fragmenti.
Lisaks sellele võivad rekombinantne polüklonaalne antikeha või rekombinantne
polüklonaalne T-raku retseptor või antikehade, või T-raku retseptorite fragmendid olla
kasutatud kompositsiooni valmistamisel, mis on ette nähtud selliste haiguste raviks, mis 10
on valitud rühmast, millesse kuuluvad vähk, nakkushaigus, põletikulisne haigus, allergia,
astma ja teine respiratoorne haigus, immunoloogilised häired, autoimmuunne haigus,
kardiovaskulaarne haigus, kesknärvisüsteemi haigus, metaboolsed ja endokriinsed
haigused, transplantaadi hülgamisreaktsioon ja ebasoovitav rasedus.
Diagnostiline kasutamine ja kasutamine keskkonnauuringutes 15
Diagnostilised komplektid ja komplektid, mis on ette nähtud kasutamiseks
keskkonnauuringutes võivad sisaldada rekombinantset polüklonaalset valku, mis on
valmistatud vastavalt käesolevale leiutisele, kusjuure see valk võib olla märgistatud
tuvastatava märgisega või mittemärgistatud, märgiseta tuvastamiseks. Kui käesoleva
leiutise kohane rekombinantne polüklonaalne valk on märgistatud, siis võib see olla 20
lisatud proovidele, mis oletatavasti sisaldavad märklaudmolekule ja märgise olemasolu või
selle puudumine osutab märklaudmolekuli olemasolule või selle puudumisele. Testitav
proov võib olla võetud sellisest keha vedelikust, nagu veri, seerum, plasma, seljaaju
vedelik, lümf või uriin, või mitteimetajalt võetud proovist, näiteks sellisest, mis on võetud
keskkonna sellisest allikast, mida kahtlustatake märklaudsaasta sisaldamises. 25
Mitteimetajalt võetud proovideks võivad olla vesi, õhk või reostatud muld.
Mittemärgistatud detekteerimine hõlmab järskude muutuste mõõtmist BIAcore-s
seostamisel, milles rekombinantste polüklonaalset valku kasutatakse märklaudmolekuli
kinnihaaramiseks.
30

EE-EP 2 152 872 B1 53
NÄITED
NÄIDE 1
Antikehadega CHO rakukloonide tuletamine
Ekspressioonivektor
Kasutatud IgG ekspressioonivektor on toodud joonisel fig 2a. 5
E1A ekspressioonivektor on toodud joonisel fig 2b.
Rakuliin
Kasutatud rakuliin on DHFR-negatiivse CHO rakuliini DG44 derivaat, mis on saadud
Lawrence Chasin’ilt Columbia Ülikoolist (saadaval ka Gibco kategoorias # 12613-014).
DG44 rakke transfekteeriti cDNA-ga, et saada 13S versioon 5. tüübi adenoviiruse 10
transaktivaatorist E1A (NCBI kood AY339865, cDNA järjestus:
vektoris pcDNA3.1+ (Cat # V790-20, invitrogeen). Valitud transfektandid sisaldasid
genetitsiini (invitrogeeni) kontsentratsioonis 500 µg/ml. Pärast selekteerimist klooniti
rakke ükshaaval lahjendamise piiramise abil. Kloonidel kontrolliti katseliselt CMV 15
promootori transaktiveerimist (kõrgenenud ekspressiooni) lühiajalise transfektsiooniga
antikeha plasmiidiga (näidatud ülalpool). Üksikklooni ekspressioonitase lühiajalisel
analüüsil oli kõrgenenud faktoriga 3, võrreldes transfekteerimata DG44 rakuliiniga.
Kõrgenenud ekspressioonitase ei kujuta endast tõendit tegelikust transaktiveerimisest ning
seda võis põhjustada eriti kõrge ekspressiooniga alamklooni valik. Stabiilse 20
transfektsiooniga läbi viidud võrdluste korral oli valitud kogumites ekspressioonitase
looduslikku tüüpi DG44 rakuliiniga võrreldes 4-5 korda kõrgem. Seda klooni (nimega
ECHO) klooniti omakorda veel kaks korda, ning see näis olevat CMV promootori
transaktiveerimise (kõrgenenud ekspressiooni) osas stabiilne. CMV promootori tegelikku
transaktiveerimist ei mõõdetud, kuid klooni puhul väljendus sellegipoolest stabiilselt 25

EE-EP 2 152 872 B1 54
kõrge antikeha ekspressioon CMV promootori kontrolli all.
Antikeha ekspressiooni plasmiidid
Kasutatud antikeha ekspressiooni plasmiidid moodustati nii, nagu näidatud ülalpool.
Selleks valiti 6 erinevat antikeha, mis olid suunatud erinevate vaktsiiniaviiruse
pinnavalkude vastu. Need valiti põhjusel, et igaühel neist on ioonivahetuskromatograafias 5
väga iseloomulik profiil, mis võimaldab teostada identifitseerimist ja kvantifitseerimist
erinevate antikehade segudes. Antikehad (mida on kirjeldatud samuti vastust ootavas
taotluses PCT/DK2006/000686, esitatud 4. detsembril 2006, nimetusega "Orthopox-
viiruse vastane rekombinantne polüklonaalne antikeha", avaldatud koodiga WO
2007/065433) olid järgmised: 10
• Sym002-037 (kloon 002-037)
• Sym002-186 (kloon 002-186)
• Sym002-235 (kloon 002-235)
• Sym002-286 (kloon 002-286)
• Sym002-303 (kloon 002-303) 15
• Sym002-482 (kloon 002-482)
IgG ELISA
IgG mõõtmiseks kasutati kihttehnikal põhinevat ELISA katset. Lühidalt kirjeldades kaeti
96 avaga plaadid (Maxisorp, NUNC) kitse inimesevastase Fc-ga (Serotec, STAR106),
millele järgnes inkubeerimine proovide ja standardiga (puhastatud inimese kapa-tüüpi 20
monoklonaalne IgG1 antikeha). Tuvastamise teostamiseks konjugeeriti kitse
inimesevastaseid kapa-tüüpi ahelaid mädarõika peroksidaasiga (Serotec STAR100P).
ECHO rakkude transfektsioon
ECHO rakud külvati T80 kolbidesse tihedusega 0,30*106 rakku kolvi kohta MEM alfa
söötmes nukleosiididega (invitrogeen kat. nr. 32571) koos 10% vasika looteseerumiga 25
(FCS) (Invitrogeen). Tunni aja jooksul pärast külvi transfekteeriti rakke Fugene6-ga
(Roche):
• 10 µl Fugene6 segatakse 490 µl Dulbecco muudetud Eagle’i söötmega ja lastakse
5 minutit toatemperatuuril seista

EE-EP 2 152 872 B1 55
• Lisatskse 5 µg ekspressiooniplasmiidi ja segu inkubeeritakse veel 15 minutit
toatemperatuuril
• Segu lisatakse rakukultuuri kolbi
Järgmisel päeval transfektsioonireagente sisaldav sööde aspireeriti, iga kolbi pesti üks
kord 5 ml MEM alfa söötmega (ilma nukleosiidideta) koos 10% dialüüsitud FCS-iga 5
(Invitrogeen) (MEMalfa-) ning lisati 10 ml sama söödet koos metotreksaadiga
kontsentratsioonis 2 nM. Seejärel vahetati söödet kaks korda nädalas.
15 päeva pärast rakud trüpsineeriti ning kõik rakud viidi üle uutesse kolbidesse. Veel 2
päeva hiljem vahetati sööde ning järgmisel päeval sööde aspireeriti, rakud loendati ning
tootlikkust mõõdeti IgG ELISA katsega. Tulemused on toodud tabelis 3. Tootlikkus on 10
arvutatud pikogrammides raku kohta päevas, kasutades rakkude arvu korje ajal.
Tabel 3
Antikeha IgG conc.,
µg/ml
Kokku
IgG
Rakkude arv,
*106
Tootlikkus, pikogrammi raku
kohta päevas
Sym002-
037
3,65 36,5 3,0 12,2
Sym002-
186
8,15 81,5 6,0 13,6
Sym002-
235
5,71 57,1 3,3 17,3
Sym002-
286
1,39 13,9 0,8 17,4
Sym002-
303
11,4 114 9,4 12,1
Sym002-
482
17,0 170 12,5 13,6
15

EE-EP 2 152 872 B1 56
Rakkude arv, söötme IgG sisaldus ning konkreetsed rakkude tootlikkused
transfekteeritud vannides
Üherakuliste kloonide tootmiseks peitsiti vannides olevaid rakke pinnaga seotud antikeha
määramiseks ning sorteeriti üherakuliselt 96 avaga plaatidesse, mis sisaldasid 50%
ECHO-rakkudele kohandatud söödet (MEMalpha-) ning 50% sama söödet ilma 5
kohandamata. Lühidalt oli peitsimisprotokoll järgmine:
1. Rakud trüpsineeritakse ja loendatakse
2. Asetage pipetiga 1-5 x 106 rakku steriilsesse FACS torusse
3. Tsentrifuugige rakke 1 min 250 g 4 °C juures ja eemaldage supernatant
4. Peske rakke 2 ml steriilses FACS PBS-s (PBS + 2% FCS) (5ml) 10
5. Peitsige rakke (Kitse F(ab)2 fragmentne inimesevastane IgG H+L- PE (Beckman-
Coulter, IM1626) lahjendatud 1:20 100 µl lahjendatud Ab/106 rakkudesegus ja
inkubeerige 20 min (4 °C pimedas)
6. Peske rakke kaks korda 2 ml FACS PBS-s (5ml)
7. Suspendeerige uuesti tasemeni 1-5 x 106/ml FACS PBS-s (2ml) 15
8. Lisage propiidiumjodiid, 10 µg/ml 1:100
Loodi korrektne lüüs ja rakud sorteeriti üherakuliselt 96 avaga plaatidesse (5 plaati
antikeha kohta) FACS-Aria abil (Beckton-Dickinson).
Ligikaudu 1 nädala pärast kontrolliti mikroskoobiga üksikkloonide olemasolu avades.
Ligikaudu 2 nädala pärast analüüsiti üksikklooniga avade supernatante eraldi lahustes 20
ELISA katse abil ning ELISA väärtuse ja avade visuaalse ülevaatuse põhjal valiti kultuuri
jätkamiseks 24 klooni, mis esindasid iga antikeha. Kloonide valimisel kasutati visuaalset
kontrolli rakkude arvu ja morfoloogia osas koos antikeha ekspressioonitaseme valikuga.
Valitud kloonidega viidi läbi täiendav ammendumisanalüüs: lühidalt kirjeldades külvati
rakud 24 avaga plaatidele ja lasti kasvada kuni enamiku rakkude surmani. Supernatante 25
analüüsiti ELISA katsega ning seerumivaba suspensioonikultuuri jaoks kohandamiseks
valiti välja peamised 10 klooni iga antikeha kohta.
Kohandamine seerumivaba suspensioonikultuuriga
Rakud trüpsineeriti ja loendati. 5*106 rakku tsentrifuugiti ja suspendeeriti uuesti 10 ml

EE-EP 2 152 872 B1 57
ProCHO4 seerumivabas söötmes (Cambrex). Rakud viidi üle 50 ml rakukultuuri torudesse
(TRP, Shveits) ja inkubeeriti loksutusseadmel 37°C juures. Rakutihedust mõõdeti kaks
korda nädalas ning iga kord lahjendati kultuurid tasemeni 0,5*106 rakku ml kohta
(esimesel 2 nädalal) või 0,3*106 rakku ml kohta (ülejäänud aja vältel). 4-5 nädala pärast
lähenes enamiku kloonide kahekordistumisaeg 30 tunnile, ning selles ajapunktis loeti nad 5
seerumivaba kultuuri jaoks kohanenuks.
Kohanemisperioodi lõpus teostati rakkudega ELISA katse, külmutati nad kultuurisöötmes
10% DMSO-ga ning neid kasutati ekspressioonikatseteks (vt Näide 2 allpool).
NÄIDE 2
Ekspressioonikatsed 10
Segakultuuride koostise püsivuse kontrollimiseks pika aja vältel valmistati mitu
kloonisegu. Kohanemisajal tehtud loenduste alusel võeti võimaluse korral arvesse
kahekordistumisaega. Omavahel püüti sobitada sarnase kahekordistumisajaga kloone.
Kokku valmistati 9 segu:
• Segud 1-5: igaühes neist kasutati üht klooni iga antikeha kohta 15
• Segu 6: kasutati kaht klooni iga antikeha kohta
• Segu 7: kasutati viit klooni iga antikeha kohta
• Segu 8: kasutati 3 klooni iga antikeha kohta
• Segu 9: kasutati kõiki saadaval olevaid kloone, 5-7 klooni iga antikeha kohta
Kloonid segati nii, et iga antikeha esindav rakkude arv (1-7 klooni iga antikeha 20
kohta) moodustas 1/6 rakkude koguarvust segus.
Katse viidi läbi 50 ml kultuuritorudes nii, nagu kirjeldatud näites 1. Kasutatud söötmeks
oli ProCHO4 ja kultuuri kogumaht 10 ml. Katset alustati kontsentratsiooniga 0,3*106
rakku ml kohta. Kultuurid lahjendati tasemeni 0,3*106 rakku kaks korda nädalas 3- ja 4-
päevase intervalliga. Kord nädalas võeti proove ELISA ja ioonivahetuskromatograafia 25
analüüsi jaoks. Esimesed proovid võeti 4. päeval ja viimased 35. päeval.
9 segu ELISA väärtused on toodud joonisel fig 3.
Arvutatud rakkude jagunemise arv algusest lõpuni erines segudes umbes 25-st kuni umbes
27-ni. See tähendab, et kui kultuure oleks lahjendatud kaks korda nädalas nii, nagu
kirjeldatud, kuid hoides kogumahu igas punktis sama, oleks kogumaht lõpuks varieerunud 30

EE-EP 2 152 872 B1 58
43 000 liitrist 152 000 liitrini. Suurte tööstuslike imetajate rakkude kultuuride maht on
tavaliselt kuni 15 000 liitrit, mis tähendab, et siinkirjeldatud segamiskatsed kujutavad
katseaja ning generatsioonide arvu poolest endast kultuuri tõepärast simulatsiooni
tööstuslikus mastaabis.
Rakkude tootlikkus näib olevat 5 nädala pikkuse perioodi vältel suhteliselt konstantne, 5
kaldudes mõnedes segudes langema.
IgG koostise analüüs
5-10 ml 0,22 µm filtreeritud söötme supernatanti viidi 1 ml MabSelect Sure kolonni (GE
Healthcare). Kolonni pesti 10 ml PBS pH 7,4-ga ja elueeriti 0.1 M glütsiiniga pH 2,7 nii,
nagu tootja poolt kirjeldatud. Vanni paigutatud valgumaterjali dialüüsiti kaks korda 40 10
mM NaCl, 50 mM Na-atsetaadi pH 5,0 suhtes, ja IgG üldkontsentratsioon määrati
kindlaks absorptsiooni mõõtmisega 280 nm juures.
60 µg IgG segu viidi nõrga katioonivahetusega kolonni PolyCat A (100x4, 6 mm, 3 µm,
1500 Å) firmalt PolyLC. Valgu elueerimiseks rakendati gradienti 150 kuni 500 mM NaCl
Na-atsetaadi pH 5,0 puhvris vooluhulgaga 1 ml/min 72 minuti vältel. Jälgiti eluaadi 215 15
nm absorptsiooni ja eraldi IgG-de suhtelised kogused määrati kindlaks signaali
integratsiooni abil.
Joonisel fig 3 on toodud segu 8 esimese ja viimase saagi IgG koostise katioonivahetuse
analüüsi kromatogrammid pärast MabSelect puhastust. Nagu näha, on eraldi Ab-d
korralikult eraldatud, algstaadiumis võib näha nelja eraldust. Seetõttu võimaldab UV 20
signaali integratsioon proovi suhtelisi IgG kontsentratsioone väga täpselt määrata.
Kõiki segusid analüüsiti nii, nagu kirjeldatud ülalpool, ning arvutati iga antikeha sisaldus
igas segus.
Algproovides võib näha üsna ühtset jaotust 6 erineva antikeha lõikes. Nähtud erinevused
võivad kajastada erinevat ekspressioonitaset segudes kasutatud kloonide vahel. Iga proovi 25
antikeha sisaldus on toodud graafiliselt joonisel fig 5. Üllataval moel saab kõiki antikehi
kõikides viimastes provides selgelt tuvastada. Samuti võib näha, et segudes, kus iga
antikeha esindas rohkem kui üks kloon (segud 6-9), esineb viimastes proovides kalduvus
ühtlasema antikehade jaotuse suunas.
30

EE-EP 2 152 872 B1 59
NÄIDE 3, E1A ekspressioon ECHO-s
Et uurida E1A ekspressiooni ECHO-s, määrati E1A mRNA tase kindlaks kvantitatiivse
pöördtranskribeeritud reaalajas PCR (qRT-PCR) meetodiga SYBR rohelise abil. qRT-PCR
meetod põhineb sihtjärjestuse PCR võimendusel. PCR teostatakse DNA siduva pigmendi,
SYBR rohelise abil, mis eritab kaheahelalise DNA-ga sidumisel rohelist valgust. See 5
võimaldab kaheahelalise DNA reaalajas kvantifitseerimist iga võimendusvooru järel.
Kui kaheahelalise DNA kogus on väike, pole PCR algusjärgus seotud SYBR rohelise
pigmendi signaali võimalik taustamürast eristada. Kuid võimendusprotsessis tugevneb
signaal mürast tugevamaks ning kaheahelalise DNA tootmist on võimalik jälgida. Nii saab
määrata proovides algselt sisalduva sihtaine suhtelise koguse, võttes aluseks tsüklite arvu, 10
mida vajab SYBR rohelise signaal konkreetse piiri ületamiseks. Täpne punkt, milles piir
ületatakse, on Ct väärtus, mida võib kasutada algse sihtkoguse suhtelise võrdluse jaoks.
Materjalid ja meetodid
Üldine RNA ekstraheeriti 3.0E+06 ECHO ja Hek293 rakkudest, kasutades vahendit
RNeasy Mini Kit, kat. nr. 74104, firmalt Qiagen vastavalt tootja juhistele. RNA proovide 15
kontsentratsioonide ja terviklikkuse määramiseks kasutati firma Agilent 2100
bioanalüsaatorit koos Eukaryote üldise RNA nanoseeria II analüüsiga. Terviklikkus on
määratud RNA terviklikkuse numbrina (RIN) vahemikus 1-10, kus 1 on degradeerunud ja
10 puutumatu RNA. ECHO RNA RIN oli 9,5 ja HEK293 RIN oli 8,9. Iga proovi cDNA
mõõtmiseks kasutati QuantiTect pöördtranskriptsiooni komplekti, kat. nr. 205311, firmalt 20
Qiagen, kasutades algmaterjalina 800 ng RNA-d. Hek293 cDNA lahjendati 25x, ECHO
cDNA aga lahjendati kaks korda. qRT-PCR viidi läbi seadmega Stratagene Mx3005P,
kasutades segu Brilliant SYBR Green QPCR Master Mix, kat. nr. 600548, firmalt
Stratagene. Termotsükli tingimused: 10 min seisuaega 95 °C juures; 40 tsüklit 15s
denatureerimisega 95 °C juures, 1 min kaksikahelaks seondamise aega 60 °C juures ja 30s 25
pikendusaega 72 °C juures; sulamiskõvera analüüs temperatuuril 55-95 °C. Kasutatud
praimerid (E1A-696bpF: 5'-TGACTCCGGTCCTTCTAACACA-'3, E1A-772bpR: 5'-
TCACGGCAACTGGTTTAATGG-'3) on suunatud 77bp fragmendile E1A geeni 3' otsas.
Tulemused
E1A ekspressiooni analüüsimiseks ECHO rakkudes kasutati qRT-PCR katset ning see 30
näitas, et E1A mRNA-d ei saa antud analüüsis võimendada ja tuvastada, viidates oluliselt
võimalusele, et ECHO rakuliinil E1A valku ei väljendu. Positiivse kontrollivahendina

EE-EP 2 152 872 B1 60
kasutati inimese 293 rakuliini, mis muteerub adenoviiruse DNA fragmendi mõjul ja
väljendab teadaolevalt E1A-d suhteliselt kõrgel tasemel.
qRT-PCR näitab võimendust Hek293 proovis, positiivne kontrolltulemus Ct = 18.74, kuid
NTC (aluskontrollita) ega ECHO proovis võimendust ei täheldatud.
Proov Ct
Hek293 18.74
ECHO Ct puudus
NTC Ct puudus
5
Näide 4. Rakupankade koostamine bioreaktori katsete jaoks
Koostise stabiilsuse uurimiseks vajalike bioreaktori katsete teostamiseks koostati pea- ja
töörakupangad. Katse jaoks kasutati 6 erineva antikehaga ECHO raku kloone, nagu
kirjeldatud näites 1. Kasutatud kloonid olid kohandatud seerumivaba suspensioonikultuuri
jaoks kultuursöötmes ProCHO4. 10
Pearakupank
Kloonid sulatati üles ja neil lasti suspensioonikultuuris ühe perioodi jooksul taastuda.
Eksponentsiaalse kasvuga rakkudest koostati 4 erinevat pearakupanka vastavalt
järgmistele spetsifikatsioonidele:
• Rakud külmutati logaritmilise kasvu olekusse 15
• Igas rakupangas oli esindatud kõik 6 antikeha
• Omavahel segati võrdsel arvul iga antikeha esindavaid rakke
• Segud 1 A -3 A sisaldasid üht klooni antikeha kohta
• Segu 4 A sisaldas 3 klooni antikeha kohta
• Segu kohta külmutati 10 ampulli, igaühes 20*106 rakku 20
• Külmutussööde: kultuursööde (ProCHO4), 10% DMSO

EE-EP 2 152 872 B1 61
Töörakupank
Üks külmutatud ampull segu kohta sulatati üles. Segusid hoiti kultuuris 8-10 päeva enne
töörakupanga koostamist:
• Rakud külmutati logaritmilise kasvu olekusse
• Segu kohta külmutati 10 ampulli, igaühes 20*106 rakku 5
• Külmutussööde: kultuursööde (ProCHO4), 10% DMSO
Näide 5: Koostise stabiilsus bioreaktoris
Üks külmutatud ampull töörakupanga segu kohta sulatati üles ning rakke hoiti kultuuris
külvijadana 14 päeva, mille möödumise ajahetkel pandi algus bioreaktori kultuurile:
Iga külvijada kasutades inokuleeriti kaks bioreaktorit 0,6*106 rakuga/ml 250 ml 10
algsöötmes (ProCHO4 + 5mM glutamiini + 1/100 asendatavaid aminohappeid).
Kultuure toideti alates 2. päevast kuni 14. päevani pärast inokulatsiooni toitesöötmega
(ProCHO4 + 6 g/L glükoosi + 5mM glutamiini + 1/100 asendatavaid aminohappeid), mille
tulemusena saadi koguseks ∼ 585 ml 16. korjepäeval.
DASGIP bioreaktorites kontrolliti järgnevaid parameetreid (üksused 1-8): 15
Tabel 4. Üldised bioreaktori protsessi parameetrid.
Kogus 250 ml pideva toitega, alates 2. päevast kuni 14. päevani
Temp. seadistuspunkt: 36.8 °C, üleminekuga 32,0 °C-le 120 tunni möödudes
pH seadistuspunkt: 6.95
pH kontroll: Kasutatakse steriilset filtreeritud 0,25 M Na2CO3.
Segamine: 80 p/min.
pO2 seadistuspunkt: 30 % (reguleeritakse gaasi O2 sisalduse kaudu)
Gaasivool: 0,1 sl/h
CO2 tase: Reguleeritakse DASGIP süsteemi poolt pH säilitamiseks

EE-EP 2 152 872 B1 62
Iga päev võeti 5 ml proove elujõulisuse, elusrakkude osakaalu, IgG toodangu ja
metaboliitide analüüsimiseks. Kõik kultuurid toimisid ootustepäraselt elujõulisuse ja
elusrakkude osakaalu osas, mis olid sarnased. Veel 10 ml võeti IgG koostise
analüüsimiseks ioonivahetuskromatograafia abil külvijadast (9. päeval ja 14. päeval) ning
igast bioreaktorist (üksus 1-8) 20., 24., 28. ja 30. päeval pärast ampullide ülessulatamist. 5
Läbi viidi IgG koostise analüüs nii, nagu kirjeldatud ülalpool näite 2 juures.
Kõiki segusid analüüsiti nii, nagu kirjeldatud ülalpool, ning välja arvutati iga antikeha
sisaldus 4 segu igas proovis. IgG kogusaagis kultuuridest jäi vahemikku ligikaudu 150
kuni 250 mg/L.
Iga proovi antikeha sisaldus on toodud graafiliselt joonisel fig 5. Üllataval moel on kõik 10
antikehad kõikides proovides selgelt tuvastatavad. Koostis näib olevat tugev, ükski kloon
pole kadunud ega asendunud 14 päeva kestnud külviahela kultiveerimisperioodi ning
sellele järgnenud 16-päevase bioreaktori tööperioodi vältel. Koostised on erinevad
sõltuvalt sisendkloonidest.
Samuti näib olevat võimalik määrata segu jaoks konkreetseid kloone valides kindlaks 15
individuaalsete antikehade suhteline jaotus lõppsaaduses.

EE-EP 2 152 872 B1 63
Patendinõudlus
1. Meetod polüklonaalse rakuliini genereerimiseks, mis suudab toota 2 kuni n erinevat
elementi sisaldavat polüklonaalset valku, kusjuures nimetatud meetod hõlmab järgmist:
a) ekspressioonivektorite komplekti moodustamine, kus iga nimetatud vektor sisaldab
vähemalt üht eraldiseisva nukleiinhappe eksemplari, milles on kodeeritud nimetatud 5
polüklonaalse valgu eraldiseisev element;
b) eraldi peremeesrakkude transfekteerimine iga nimetatud ekspressioonivektoriga
tingimustes, milles välditakse ekspressioonivektorite kohaspetsiifilist integreerimist
rakkude genoomi, mille tulemusena saadakse 2 kuni n rakusegu, millest igaüks annab ühe
eraldiseisva polüklonaalse valgu elemendi; 10
c) nimetatud 2 kuni n rakusegu segamine polüklonaalse rakuliini saamiseks.
2. Punktile 1 vastav meetod, kus ekspressioonivektorid on stabiilselt ja juhuslikkuse alusel
integreeritud ühte või mitmesse peremeesrakkude kromosoomi.
3. Punktile 1 või 2 vastav meetod, kus punktis b) saadud transfekteeritud rakud
kloonitakse, näiteks FACS kloonimise teel, ning kus eelistatavalt valitakse kloonid 15
vähemalt ühe kriteeriumi alusel, mis on valitud rühmast, kuhu kuuluvad kasvukiirus,
kahekordistumisaeg, ekspressioonitase, tootmistase, tootmise stabiilsus ajas, elujõulisus,
vastupidavus, tugevus, morfoloogia ja eksemplaride arv.
4. Punktile 3 vastav meetod, kus kloonid valitakse ühtlaselt vähemalt ühe kriteeriumi
alusel, näiteks kus kloonid valitakse ühtlaselt vastavalt kahekordistumisajale ja/või 20
ekspressioonitasemele, võimaliku variandina, kus rohkem kui üks kloon on valitud iga
erineva polüklonaalse valgu elemendi kohta.
5. Mistahes eelnevale punktile vastav meetod, kus nimetatud erinevaid eraldiseisvaid
elemente tootvad rakusegud segatakse vahekorras 1:1, või kus nimetatud rakusegud
segatakse muus vahekorras kui 1:1. 25
6. Mistahes eelnevale punktile vastav meetod, kus ekspressioonivektorid on identsed, välja
arvatud variatsioonide osas polüklonaalse valgu kodeerimisjärjestuses.
7. Mistahes eelnevale punktile vastav meetod, kus peremeesrakud saadakse ühest kloonist
enne transfekteerimist.
8. Mistahes eelnevale punktile vastav meetod, kus polüklonaalne valk on oligomeerne 30
valk, nt kus üks ekspressioonivektor kodeerib kõiki ühe eraldiseisva polüklonaalse valgu

EE-EP 2 152 872 B1 64
elemendi allüksusi, või kus ekspressioonivektorite komplekt punktis a) koosneb kahest või
enamast ekspressioonivektorite alamhulgast, millest esimene alamhulk sisaldab
varieeruvaid nukleiinhappe järgnevusi, mis kodeerivad üht valgu alamhulka, nt antikeha
raske ahel, ning teine alamhulk sisaldab varieeruvaid nukleiinhappe järgnevusi, mis
kodeerivad teist valgu alamhulka, nt antikeha kerge ahel, mille korral iga transfektsioon 5
teostatakse ekspressioonivektorite esimese alamhulga ühe elemendi ja teise alamhulga ühe
elemendiga.
9. Mistahes eelnevale punktile vastav meetod, kus ekspressioonivektor või täiendav
ekspressioonivektor kodeerib valitavat markerit, nt kus rakke kultiveeritakse pidevalt
tingimustes, mis soodustavad valitavat markerit tootvate rakkude kasvu, nt kus valitavaks 10
markeriks on geeniprodukt, milles peremeesrakk on puudulik, näiteks kus valitavat
markerit kodeerib transkribeering, mis kodeerib ühtlasi polüpeptiidset elementi või
nimetatud polüpeptiidse elemendi alamhulka, ning kus eelistatavalt kodeerib valitavat
markerit suurimat alamhulka kodeeriv transkribeering.
10. Mistahes eelnevale punktile vastav meetod, kus polüklonaalne valk on polüklonaalne 15
antikeha või polüklonaalse antikeha fragment; eelistatavalt, kus kõikidel polüklonaalse
antikeha elementidel on sama konstantne raske ja/või kerge ahela, eelistatavalt raske ahela
piirkond.
11. Mistahes eelnevale punktile vastav meetod, kus peremeesrakud on päristuumsed
rakud, eelistatavalt imetajarakud, nt imetajarakud, mis on valitud rühkast, kuhu kuuluvad 20
Hiina hamstri munarakud (CHO), COS rakud, BHK rakud, müeloomirakud (nt. Sp2/0
rakud, NSO, YB2/0), NIH 3T3 rakud, fibroblastsed või põlistatud inimese rakud, nagu
näiteks HeLa rakud, HEK293 rakud või PER.C6 rakud.
12. Punktile 11 vastav meetod, kus peremeesrakk annab rekombinantse transaktivaatori,
mis suudab transaktiveerida ergutava kodeeringu polüklonaalse valgu tootmiseks. 25
13. Meetod polüklonaalse valgu tootmiseks, mis hõlmab järgmist:
a) polüklonaalse rakuliini loomine, mille saamiseks on kasutatud mistahes punkti 1 kuni
12 meetodit;
b) polüklonaalse rakuliini kultiveerimine tingimustes, mis võimaldavad polüklonaalse
valgu tootmist; 30
c) polüklonaalse valgu taastamine ja võimalik puhastamine rakkudest või söötmest; ja,
lisavariandina,

EE-EP 2 152 872 B1 65
d) iga eraldiseisva elemendi olemasolu kontrollimine taastatud ja soovi korral puhastatud
polüklonaalses valgus.
14. Polüklonaalne rakuliin, mis koosneb 2 kuni n rakupopulatsioonist, kusjuures iga
populatsioon annab rekombinantse polüklonaalse valgu eraldiseisva elemendi, kus
rekombinantne polüklonaalne valk sisaldab erinevaid, kuid homoloogseid valgumolekule, 5
ning rakud sisaldavad vähemalt üht ekspressiooniskeemi, mis on integreeritud
juhuslikkuse põhimõttel genoomi nii, et integratsioonipaigad võivad polüklonaalse
rakuliini elementide vahel varieeruda.
15. Punkti 14 polüklonaalne rakuliin, kus vähemalt üks ekspressiooniskeem on
integreeritud ühte või mitmesse kromosoomi. 10
16. Punkti 14 või 15 polüklonaalne rakuliin, kus n on vähem kui 50, näiteks vähem kui 45,
näiteks vähem kui 40, näiteks vähem kui 35, näiteks vähem kui 30.
17. Mis tahes punkti 14-16 polüklonaalne rakuliin, kus rekombinantse polüklonaalse valgu
üht eraldiseisvat elementi tootvad rakud saadakse 2 või enamast kloonitud rakust ja kus
kloonitud rakkude arv on väiksem kui 50, näiteks vähem kui 20, näiteks vähem kui 15, 15
näiteks vähem kui 10.
18. Mis tahes punkti 14-17 polüklonaalne rakuliin, kus polüklonaalne valk on oligomeerne
valk, nt kus iga ekspressiooniskeem kodeerib oligomeerse valgu allüksusi, nt kus
allüksuste tootmine on samade või identsete promootorite kontrolli all.
19. Mistahes punkti 14-18 polüklonaalne rakuliin, kus ekspressiooniskeemid kodeerivad 20
valitavat markerit, kus valitav marker võidakse kodeerida transkribeeringuga, mis
kodeerib ühtlasi polüpeptiidset elementi või nimetatud polüpeptiidse elemendi allüksust.
20. Mis tahes punkti 14-19 polüklonaalne rakuliin, kus polüklonaalne valk on
polüklonaale antikeha või polüklonaalse antikeha fragment, kus eelistatavalt on kõik
polüklonaalse antikeha elemendid sama isotüübiga. 25
21. Mis tahes punkti 14-20 polüklonaalne rakuliin, kus peremeesrakud on päristuumsed
rakud, eelistatavalt imetajarakud.
22. Mis tahes punkti 14-21 polüklonaalne rakuliin, kus rakud sisaldavad stabiilselt
integreeritud ekspressiooniskeemi, kodeerides transaktivaatori, mis suudab
transaktiveerida ergutava kodeeringu polüklonaalse valgu elementide jaoks.30

EE-EP 2 152 872 B1
1/8
5

EE-EP 2 152 872 B1
2/8

EE-EP 2 152 872 B1
3/8
Fig. 4
(muudetud 230107 seguga 5
(muudetud 230207 seguga

EE-EP 2 152 872 B1
4/8

EE-EP 2 152 872 B1
5/8

EE-EP 2 152 872 B1
6/8

EE-EP 2 152 872 B1
7/8

EE-EP 2 152 872 B1
8/8
5





























