Embed Size (px)
Citation preview

1
RENATA CONDURU RIBEIRO
EFEITO DO ARMAZENAMENTO NA QUALIDADE
FISIOLÓGICA DE SEMENTES DE Gliricidia sepium (J acq.)
Steud. (LEGUMINOSAE PAPILIONOIDEAE).
FEIRA DE SANTANA 2008

2
RENATA CONDURU RIBEIRO
EFEITO DO ARMAZENAMENTO NA QUALIDADE
FISIOLÓGICA DE SEMENTES DE Gliricidia sepium (Jacq.) Steud.
(LEGUMINOSAE PAPILIONOIDEAE).
Dissertação apresentada ao Programa de
Pós Graduação em Botânica, como parte
dos requisitos exigidos para obtenção do
título de MESTRE em BOTÂNICA, na
área de concentração de Fisiologia
Vegetal.
ORIENTADORA: DRA. CLAUDINÉIA REGINA PELACANI COORIENTADOR: DR. RENATO DELMONDEZ DE CASTRO

3
“ Se não houver frutos, valeu a beleza das flores.
Se não houver flores, valeu a sombra das folhas.
Se não houver folhas, valeu a intenção da semente.”
Henfil
À minha mãe, Risete e o meu pai Manoel pelo apoio e amor incondicional,
dedico

4
AGRADECIMENTOS
A força superior que nos protege “Deus”, pela conquista;
Aos meus pais pelo confiança, incentivo, apoio, amor incondicional que me deram toda
vida e que me fez tornar quem sou;
As minhas irmãs pela confiança e pela companhia mesmo estando distante;
A Claudinéia Regina Pelacani pela orientação, alegria, ensinamentos não somente
profissionais, mas também de vida, pode ter certeza que ainda estaremos juntas por um
bom tempo, “minha mãe postiça”;
A minha amiga Cimille que apesar de ter caído de páraquedas em cima de mim, foi
uma aterrissagem muito bem feita. Obrigada pelo companheirismo, cumplicidade, apoio
em vários momentos difíceis que dividimos direitinho;
A meus companheiros, Jonaicon e Hugo podem ter certeza que vocês foram
fundamentais nessa caminhada e os fins de semana perdidos vão valer a pena na frente;
Ao seu José Juracy, seu apoio foi uma peça chave desse trabalho, sem as nossas várias
conversas pelo telefone, sem as coletas, sem a sua disposição o tempo todo nada teria
acontecido com sucesso;
Aos funcionários do Horto pela colaboração, em especial a Dona Zezé pela atenção e
carinho, seus chazinhos sempre me fortaleceram;
A Flavia que entrou como uma luz no meu caminho, uma amiga pra todas as horas, que
será inesquecível;
A Embrapa SemiÁrido pelo apoio, em especial a Dra. Bárbara pela colaboração. Com
certeza estarei presente quando precisar;
Aos amigos, Fabrício, Armando, Zizinho e Alberto que fizeram minhas análises ficarem
tão simples com a alegria contagiante deles o tempo todo;

5
Ao CNPq pela bolsa de estudos concedida;
À FAPESB pelo financiamento do projeto;
A todos que, de alguma maneira contribuíram para que eu conseguisse chegar até aqui,
seja com um sorriso, um incentivo ou simplesmente com a força positiva do
pensamento em mim.
Agradeço a todos, com muito carinho.

6
SUMÁRIO
INTRODUÇÃO........................................................................................................... 8 1.1 Impor tância e Caracter ização da Espécie ........................................................ 9 1.2 Caracter ização do Ambiente........................................................................... 12 1.3 Armazenamento de Sementes ......................................................................... 13 1.4 Aspectos fisiológicos da germinação de sementes........................................... 16
OBJETIVO ............................................................................................................... 20 CAPÍTULO I............................................................................................................. 21 Effects of storage on seed viability in Gliricidia sepium (Jacq.) Steud. (Leguminosae Papilionoideae)............................................................................ 21 ABSTRACT .......................................................................................................... 22 RESUMO .............................................................................................................. 23 INTRODUCTION................................................................................................. 24 MATERIAL AND METHODS ............................................................................ 26 RESULTS.............................................................................................................. 28 DISCUSSION........................................................................................................ 32 ACKNOWLEDGEMENTS .................................................................................. 34 REFERENCES...................................................................................................... 35
CAPÍTULO II ........................................................................................................... 38 Reserve mobilization during imbibition of stored Gliricidia sepium (Jacq.) Steud. seeds. ...................................................................................................................... 38 ABSTRACT .......................................................................................................... 39 RESUMO .............................................................................................................. 40 INTRODUCTION................................................................................................. 41 MATERIAL AND METHODS ............................................................................ 44 RESULTS.............................................................................................................. 47 DISCUSSION........................................................................................................ 53 ACKNOWLEDGMENTS..................................................................................... 56 REFERENCES...................................................................................................... 57
CAPÍTULO III.......................................................................................................... 61 Amylolitc activity and physiological quality of Gliricidia sepium (Jacq.) Steud. (Leguminosae Papilionoideae) seeds dur ing storage. ........................................ 61 ABSTRACT .......................................................................................................... 62 RESUMO .............................................................................................................. 63 INTRODUCTION................................................................................................. 64 MATERIALS AND METHODS .......................................................................... 66 RESULTS.............................................................................................................. 69 DISCUSSION........................................................................................................ 74 ACKNOWLEDGEMENTS .................................................................................. 75 REFERENCES...................................................................................................... 76
CAPÍTULO IV.......................................................................................................... 79 Déficit hídr ico em sementes armazenadas de Gliricidia sepium (Jacq.) Steud. (Leguminosae Papilionoideae)............................................................................ 79 ABSTRACT .......................................................................................................... 80 RESUMO .............................................................................................................. 81 INTRODUÇÃO ..................................................................................................... 82 MATERIAL E MÉTODOS .................................................................................. 84 RESULTADOS E DISCUSSÃO ........................................................................... 86 AGRADECIMENTOS.......................................................................................... 94

7
REFERÊNCIAS BIBLIOGRÁFICAS ................................................................. 95 CONSIDERAÇÕES FINAIS.................................................................................... 99 REFERÊNCIAS BIBLIOGRÁFICAS CITADAS NA INTRODUÇÃO GERAL 100 RESUMO GERAL .................................................................................................. 104 ABSTRACT OF DISSERTATION ........................................................................ 105

8
INTRODUÇÃO
Nos últimos anos tem se intensificado o interesse na propagação de espécies
florestais, ressaltandose a necessidade de recuperação de áreas degradadas e
recomposição da paisagem natural. Entretanto é restrito o conhecimento disponível para
o manejo e análise das sementes da maioria das espécies de modo a caracterizar seus
atributos físicos e fisiológicos. Há também a necessidade de se obter informações
básicas sobre a germinação, o cultivo e as potencialidades dessas espécies nativas ou
introduzidas, visando sua utilização para os mais diversos fins.
Comunidades vegetais como, a caatinga, predominante na região do nordeste
brasileiro, são submetidas à escassez de água por períodos prolongados e tendem a ser
dominadas por plantas nativas ou cultivadas que apresentam características de
tolerância a seca e que sobrevivem à seca através de sementes armazenadas nos bancos
de solos.
Considerandose que os estudos com sementes são o ponto de partida para a
utilização e exploração, de forma racional, de algumas espécies potenciais, o
conhecimento sobre os processos fisiológicos de sementes armazenadas é de
fundamental importância, uma vez que, a longevidade destas é bastante influenciada
pelas condições do armazenamento. A manutenção da viabilidade das sementes durante
o armazenamento depende diretamente do teor de água encontrado nas unidades de
dispersão. Esta dependência está relacionada à intensidade das reações químicas e
metabólicas que caracterizam a atividade fisiológica das sementes, refletindo
diretamente no seu desempenho germinativo.

9
1.1 Impor tância e Caracter ização da Espécie
Gliricidia sepium (Jacq.) Steud. é uma planta nativa da América do Sul e
Central, com distribuição pelas regiões tropicais (SUMBERG, 1985). Segundo Duque
(1998), esta espécie ocorre naturalmente do México até a Colômbia, Venezuela e
Guianas. Desde os tempos précolombianos, G. sepium já era cultivada além das áreas
de ocorrência natural, tendo sido domesticada em Cuba, Jamaica, Havaí, África
Ocidental e Meridional, Índia, Siri Lanka, Tailândia, Filipinas, Indonésia e Austrália
(PARROTTA, 1992).
Vulgarmente conhecida no Brasil como gliricidia, no México e em países da
América Central como “madero negro”, “mata ratón”, “madre de cacao”, G. sepium, é
uma espécie pertencente à família Fabaceae sendo de grande interesse comercial e
econômico para regiões tropicais, devido às suas características de uso múltiplo. É uma
planta perene, que se reproduz sexuada (por semente) e assexuadamente (por estacas).
Apresenta porte arbóreo variando de 12 a 15 metros de altura, com diâmetros de até 30
cm (NATIONAL ACADEMY SCIENCES, 1980) e crescimento cespitoso (touceiras),
formando em média 4 a 5 fustes. Possui casca fina, lisa e esbranquiçada. Sua copa, em
geral, é ampla, entretanto, a forma da árvore é bastante variável, dependendo da
procedência e manejo. As raízes de gliricidia associamse a bactérias do gênero
Rhizobium, com as quais entram em simbiose, originando um grande número de
nódulos, responsáveis pela fixação de nitrogênio (FRANCO, 1988).
As folhas são alternas imparipinadas, constituídas por 7 a 17 folíolos de 3 a 7 cm
de comprimento. As flores estão reunidas em inflorescências terminais, do tipo cacho
ou racemo e apresentam constituição típica das Papilionáceas. As pétalas são
predominantemente de cor lilás, com a porção central de estandarte em tom creme, que
funcionam como guias de néctar. O androceu é formado por onze estames diadelfos e o

10
gineceu apresenta ovário súpero, estilete único e estigma bífido. Os frutos são vagens
chatas, que geralmente apresentam cor verde pálido, podendo apresentar tonalidades
arroxeadas em função da exposição solar (Figura 1).
As vagens variam de 10 a 17 cm de comprimento e contêm três a oito sementes.
As sementes são lisas, com média de 0,9 cm de diâmetro, em geral, de cor marrom e
apresentam dormência tegumentar quando armazenadas por mais de um ano. A floração
e a frutificação desta espécie na região semiárida, geralmente ocorrem no período seco,
nos meses de agosto a novembro, quando as árvores estão parcialmente sem folhas. As
plantas florescem a partir do terceiro ano de idade.
Há vários anos, esta espécie vem sendo cultivada na região sudoeste da Bahia
para o sombreamento do cacaueiro, tendo sido recentemente introduzida nos estados de
Pernambuco e Sergipe. Esta espécie se destaca por apresentar rápido crescimento, alta
capacidade de regeneração, e vem sendo explorada como forrageira pelo seu alto valor
nutritivo, como produtora de estacas vivas, medicinal e, ainda, como alternativa
energética. Entretanto, é necessário o estabelecimento de um programa de
melhoramento de gliricidia visando explorar a sua variabilidade genética, para
melhoraria da espécie, selecionandose indivíduos capazes de suportar as adversidades
climáticas com boa produtividade, e estimular a sua utilização em programas de
recuperação de áreas degradadas na região Nordeste (DRUMOND & CARVALHO
FILHO, 1999).

11
Figura 1. Aspectos da espécie Gliricidia sepium – A Árvore, B Flores, C Frutos,
D Sementes.
A
D
B
C

12
1.2 Caracter ização do Ambiente
O Brasil possui 385 milhões de hectares de florestas nativas (IPEF, 2000), sendo
que a região de Caatinga tem cerca de 800 mil km 2 , totalizando 11% do território
nacional e 70% do território nordestino, abrangendo os estados do Ceará, Rio Grande do
Norte, Paraíba, Pernambuco, Sergipe, Alagoas, Bahia, sul e leste do Piauí e norte de
Minas Gerais (LIMA, 1996). O clima nessa região chamada “Polígono das Secas” é
semiárido, quente com baixa pluviosidade, em torno de 250 a 800 mm anuais. A
quantidade de chuvas pode variar, alcançando em anos de muitas chuvas até 1.000
mm/ano e em anos de seca, apenas 200 mm/ano, em certas regiões. A temperatura
média se situa entre 24 e 26°C e varia pouco durante o ano (MAIA, 2004). Além dessas
condições climáticas rigorosas, a região está submetida a ventos fortes e secos, que
contribuem para a aridez da paisagem nos meses de seca (SAMPAIO & RODAL,
2000).
O bioma Caatinga é caracterizado por uma vegetação xerófila, de fitofisionomia
e florística variada. Esse tipo de formação vegetal tem características bem definidas
como árvores baixas e arbustos que, em geral perdem as folhas na estação das secas
(caducifólias), além da ocorrência de muitas cactáceas. O aspecto geral da vegetação, na
seca, é de uma mata espinhosa e agreste, apresentando três estratos: arbóreo, arbustivo e
o herbáceo (LIMA, 1996).
As espécies vegetais da caatinga possuem adaptações vegetativas ao ambiente
semiárido, tais como folhas transformadas em espinhos, cutículas altamente
impermeáveis, caules suculentos, etc. Todas essas adaptações lhes conferem o aspecto
característico denominado xeromorfismo. Outras adaptações importantes à vida das
plantas na caatinga é a queda das folhas na estação seca, a rápida brotação destas na
estação das chuvas e a presença de um sistema radicular bem desenvolvido. A perda das

13
folhas é uma adaptação para reduzir a perda de água por transpiração e as raízes bem
desenvolvidas aumentam a superfície de absorção de água no solo.
Segundo Drumond et al., (2000), não existe uma lista completa das espécies de
caatinga, no entanto, em trabalhos realizados sobre a flora e vegetação da caatinga,
foram registradas quase 600 espécies arbóreas e arbustivas, sendo 180 endêmicas. Cada
uma delas com características especiais e importantes para a composição deste bioma
(MAIA, 2004).
Nessa região existe uma grande demanda por recursos florestais em pequenas
propriedades, onde cada vez mais, as árvores assumem um importante papel na
economia rural. No entanto, o manejo inadequado da vegetação da caatinga, pela
retirada de madeira, exploração de pecuária e agricultura tem colocado em risco a
biodiversidade deste ecossistema (SAMPAIO, 1995). Desta forma, é importante a
utilização de espécies de uso múltiplo, isto é, cultivadas ou manejadas para mais de uma
finalidade, como a G. sepium.
É significativo o nível de conhecimento referente às características de adaptação
vegetativa das espécies submetidas às condições ambientais do semiárido, entretanto,
pouco se sabe sobre os mecanismos de adaptação das sementes a tais condições, uma
vez que os bancos de sementes no solo constituemse numa das principais estratégias de
sobrevivência dessas espécies (BASKIN & BASKIN, 1998).
1.3 Armazenamento de Sementes
A propagação de espécies florestais ocorre, principalmente, por meio de
sementes. Vários estudos têm sido realizados nos últimos anos sobre a tecnologia de
sementes de espécies florestais nativas ou introduzidas do Brasil, devido à crescente
necessidade de reabilitação de ecossistemas florestais e de conservação de

14
germoplasmas. No entanto, diante da grande diversidade de espécies dos nossos biomas,
as informações relativas ao comportamento fisiológico dessas sementes durante o
armazenamento ainda são deficientes.
O armazenamento das sementes deve ser iniciado na maturidade fisiológica e o
maior desafio é conseguir que as sementes, após certo período, ainda apresentem
elevada qualidade fisiológica (FERREIRA & BORGHETTI, 2004).
A longevidade das sementes é variável de acordo com o genótipo, mas, a
conservação do potencial fisiológico depende, em grande parte, da temperatura e das
condições do ambiente de armazenamento (FERREIRA & BORGHETTI, 2004;
MARCOS FILHO, 2005).
Roberts (1973) sugeriu a classificação fisiológica de armazenamento para
sementes em duas categorias: as que podem ser armazenadas por longos períodos com
baixos teores de água e em baixas temperaturas, denominadas de ortodoxas; e as que
não toleram o dessecamento e o armazenamento em baixas temperaturas, denominadas
de recalcitrantes, as quais nessas condições perdem sua viabilidade mesmo quando
armazenadas por curtos períodos.
As sementes ortodoxas sofrem redução do teor de água após a suspensão da
deposição de reservas, que pode variar conforme as condições climáticas prevalecentes.
Nessas sementes, isso leva a um estado de quiescência metabólica, garantindo a sua
sobrevivência até que as condições favoráveis à germinação sejam restabelecidas
(BEWLEY & BLACK, 1994). Durante esse período, o tipo de armazenamento
fornecido às sementes é de extrema importância para que ocorra a manutenção da
viabilidade destas, sendo o teor de água das sementes e a temperatura de
armazenamento os dois fatores de maior influência (WARD & POWELL, 1983).

15
Portanto, o potencial de conservação das sementes depende diretamente da qualidade
fisiológica das mesmas no início do período de armazenamento.
O alto teor de água das sementes é uma das principais causas da perda do poder
germinativo durante o armazenamento (DESAI et al., 1997). O alto teor de água causa
aumento da taxa respiratória e ação de microrganismos, sendo que umidade acima de
20% do peso inicial das sementes pode promover o aquecimento da massa a
temperatura letal (HARRINGTON, 1972).
A temperatura influencia todas as atividades biológicas. O aumento da
temperatura do ambiente de armazenamento provoca aumento da taxa respiratória da
semente, proliferação de fungos e de insetos que a acompanham (POPINIGIS, 1985).
A preservação da qualidade fisiológica de sementes sob determinadas condições
ambientais de temperatura e umidade relativa do ar é influenciada pelo tipo de
embalagem utilizada (FERREIRA & BORGHETTI, 2004). A escolha da embalagem
depende da espécie, do grau de umidade das sementes, das condições e do período de
armazenamento (MARCOS FILHO, 2005).
As embalagens, quanto à permeabilidade ao vapor de água, podem ser
classificadas em permeáveis, semipermeáveis e impermeáveis (CARNEIRO &
AGUIAR, 1993). As embalagens permeáveis permitem a troca de vapor entre as
sementes e o ambiente externo circundante permitindo que, o teor de água das sementes
sofre flutuações com as variações de umidade relativa do ar. Os principais materiais
empregados comercialmente na confecção de embalagens permeáveis de sementes são
papel, algodão, juta e polipropileno trançado (FERREIRA & BORGHETTI, 2004).
As embalagens semipermeáveis mostramse resistentes à troca de vapor de água
entre as sementes e o ambiente externo circundante. Para a conservação de sementes em
embalagens semipermeáveis, o teor de água das sementes deve ser de 2 a 3 pontos

16
percentuais inferior ao empregado nas embalagens permeáveis. Os materiais utilizados
nesse tipo de embalagem são polietileno de baixa espessura e combinações de lâminas
de papel e outro material, como é o caso dos papéis alumizados, plastificados e com
película de asfalto (FERREIRA & BORGHETTI, 2004).
As embalagens impermeáveis impedem o intercâmbio de vapor de água entre as
sementes e o meio externo. Geralmente, são empregados sacos de polietileno espesso,
de média e alta densidade, envelopes de alumínio, embalagens metálicas de alumínio e
folhas de flandres com sistema de regravação e recipientes de vidro com sistema de
vedação na tampa.
A conservação de sementes de modo geral, é de grande importância, e o
armazenamento, uma vez aplicado de modo adequado, vai diminuir sua velocidade de
deterioração que se caracteriza por ser um processo irreversível (DELOUCHE et al.,
1973; MELO et al., 1998, citados por CABRAL et al., 2003).
A deterioração ocorre em níveis molecular, genético, celular, de tecido e de
população da semente (MATTHEWS, 1985). Esse é um evento que não pode ser
evitado, mas pode ser controlado, sendo esse o principal objetivo do armazenamento,
que está entre as estratégias de conservação ex situ mais utilizadas, por preservar as
características genéticas das sementes até que estas sejam semeadas (CARNEIRO,
1985, NODARI et al., 1998).
1.4 Aspectos fisiológicos da germinação de sementes
A propagação de um grande número de espécies florestais de importância social,
econômica e cultural encontra sérias limitações, em razão do pouco conhecimento que
se dispõe sobre as características fisiológicas, morfológicas e ecológicas de suas
sementes (MACHADO, 2002).

17
Os diversos métodos e procedimentos utilizados para a avaliação da qualidade
de sementes baseiamse na análise dos componentes da qualidade de uma amostra
representativa que retrata o perfil de determinado lote. O teste mais tradicionalmente
utilizado para a avaliação da qualidade de lotes de sementes é o teste de germinação
(OLIVEIRA, 2004).
A germinação da semente ortodoxa é considerada como a retomada das
atividades metabólicas do eixo embrionário, o qual se encontrava paralisado nas fases
finais do processo de maturação; porém, quando estimulado por condições ambientais,
desenvolvese, culminando com o rompimento do tegumento pela radícula. Essa é uma
etapa crítica do biociclo vegetal pelo fato do processo estar associado a vários fatores de
natureza extrínseca (fatores do ambiente físico) e intrínseca, ou seja, a processos fisio
metabólicos (LABORIAU, 1983; POPINIGIS, 1985; ANDRADE & DAMIÃOFILHO,
1989; BIANCHETTI, 1991; BORGES & RENA, 1993; BEWLEY & BLACK, 1994;
SANTOS, 1999).
Como resultado do déficit hídrico, há geralmente elevação de sais solúveis
(salinidade) que podem influenciar, significativamente, na resposta germinativa das
espécies. Dentre os diversos fatores ambientais capazes de influenciar o processo
germinativo de sementes, a indisponibilidade de água e a presença de sais em elevados
potenciais são os mais importantes. Esta condição é vista como um fator limitante à
iniciação da germinação de sementes e está envolvida, direta ou indiretamente em todas
as demais etapas do metabolismo subseqüente, seguindo a ativação do ciclo celular e
crescimento da plântula (ROCHA, 1996).
O processo germinativo se inicia com a absorção de água por embebição,
seguida da retomada das atividades metabólicas, sobretudo da síntese de novas enzimas
e do aumento de atividades das hidrolases préexistentes, visando a mobilização dos

18
compostos de reserva para a retomada de crescimento do eixo embrionário (SALES,
2002). Porém, há necessidade de que a semente alcance um nível adequado de
hidratação o qual permita a reativação dos seus processos metabólicos. Uma análise das
relações hídricas das taxas de germinação de sementes revelou que o início da
germinação está relacionado com a sensibilidade da iniciação do crescimento radicular
ao conteúdo de água no meio. A habilidade do embrião em absorver água do meio e
iniciar o seu crescimento é dependente do potencial osmótico de suas células
(MARCUS FILHO, 1986 apud, FONSECA & PEREZ, 2003).
Durante o processo germinativo as reservas das sementes têm basicamente duas
funções que se relacionam com a manutenção e o desenvolvimento do embrião até a
formação de uma plântula que apresente a capacidade de se manter de forma
autotrófica. Os compostos de carbono normalmente acumulados em sementes podem
ser utilizados tanto para produzir energia como para construir fisicamente as células
(FERREIRA & BORGHETTI, 2004).
Há enorme variação na composição de sementes, mas as substâncias
armazenadas em grande quantidade constituem os carboidratos, os lipídeos e as
proteínas. Os dois primeiros servem como fonte de energia e carbono para a germinação
das sementes e o desenvolvimento das plântulas. As proteínas têm como função
armazenar principalmente nitrogênio e enxofre, essenciais para a síntese de proteínas,
ácidos nucléicos e compostos secundários na plântula, sendo fundamental para o
crescimento inicial (FERREIRA & BORGHETTI, 2004).
Os principais carboidratos que atuam como reservas em sementes são a sacarose,
os oligossacarídeos da série rafinósica, o amido e os polissacarídeos de parede celular.
Enquanto a sacarose é praticamente universal, os oligossacarídeos da série rafinósica
ocorrem em um grande número de sementes de dicotiledôneas. O amido é um dos

19
compostos de reserva de mais larga ocorrência nos vegetais superiores e os
polissacarídeos de parede celular ocorrem em alguns grupos taxonômicos em que
geralmente atuam como reserva, mas preservando funções secundárias importantes
como o controle de absorção e de distribuição da água nos diferentes tecidos das
sementes. Os oligossacarídeos são degradados logo no início da germinação e, acredita
se, assim, que sejam compostos de reserva. Porém, sua principal função tem sido
atribuída à propriedade das sementes ortodoxas de estabilizarem suas membranas e,
com isso, poderem permanecer secas por um longo período, após o qual germinam
normalmente, quando expostas ao meio líquido. Essa hipótese é suportada pelo fato de
haver uma tendência maior de acúmulo de oligossacarídeos da série rafinósica em
sementes ortodoxas em relação às recalcitrantes (FERREIRA & BORGHETTI, 2004).
Apesar do aumento considerável de conhecimentos relativos à análise de
sementes de espécies florestais, gerado pelas pesquisas nestas duas últimas décadas, a
maioria delas necessitam ainda de subsídios básicos referentes às exigências quanto às
condições ótimas de germinação (VARELA et al., 2005).
Assim, como as sementes constituem o principal veículo de multiplicação de
espécies, justificase a prioridade dirigida à concentração de esforços para elucidar ou
aprimorar os conhecimentos sobre o processo de germinação e os efeitos de fatores que
possam beneficiálo ou prejudicálo. Essas informações são fundamentais para o
estabelecimento de diagnósticos e o fornecimento de bases para a adoção de práticas
culturais adequadas, principalmente pelo sertanejo local, podendo ainda ser utilizada na
solução de problemas regionais.

20
OBJETIVO
Objetivouse com esse trabalho avaliar a germinabilidade e longevidade de
sementes de Gliricidia sepium submetidas a diferentes períodos de armazenamento,
visando o ajuste de protocolos para conservação de sementes, da espécie, e sua
utilização pelo pequeno produtor da Caatinga. Para o alcance desse objetivo principal
tevese como objetivos específicos:
ü Avaliar a qualidade fisiológica das sementes de G. sepium, submetidas a
diferentes condições e períodos de armazenamento, visando à escolha da melhor
estratégia de conservação;
ü Avaliar a mobilização de macromoléculas durante a germinação de sementes de
G. sepium verificando as possíveis modificações dessas reservas de
macromoléculas durante o armazenamento;
ü Avaliar as mudanças na qualidade fisiológica de sementes de G. sepium
submetidas ao armazenamento, considerandose a viabilidade e vigor das
sementes pela atividade das enzimas de degradação do amido durante o processo
de germinação.
ü Avaliar o efeito do déficit hídrico sobre a germinação de sementes armazenadas
de G. sepium.

21
CAPÍTULO I
Effects of storage on seed germination in Gliricidia sepium (Jacq.) Steud.
(Leguminosae Papilionoideae)

22
ABSTRACT – (Effects of storage on seed germination in Gliricidia sepium
(Jacq.) Steud. (Leguminosae Papilionoideae)). Studies of seed physiological
processes represent the starting point for the rational utilization of native or cultivated
plant species from the caatinga (dryland) region for which germination studies are still
scarce. In order to determine adequate methods for the conservation of Gliricidia
sepium seeds, the physiological quality of propagules exposed to different conditions
and times of storage were examined. Recently collected seeds of G. sepium were
analyzed immediately while other samples were stored for three, six, nine, or twelve
months in plastic or paper bags under refrigeration or at room temperature. The
germination rates of seeds from each storage regime were determined and these rates
compared to their water content. It was observed that germination capacity was directly
related to changes in seed humidity. Storing seeds in plastic bags yielded the best
conservation, principally when they were stored at low temperatures. However, even at
temperatures near 25°C it was possible to prolong seed viability when their water
content was controlled through the use of impermeable packing.
Key words: germination, water content, physiological quality, gliricidia

23
RESUMO – (Efeito do armazenamento sobre a germinação de sementes de
Gliricidia sepium (Jacq.) Steud. (Leguminosae Papilionoideae)). Estudos dos
processos fisiológicos de sementes são efetivamente o ponto de partida para utilização e
exploração de forma racional de espécies nativas ou cultivadas da caatinga cujos
trabalhos sobre germinação ainda são escassos. Visando contribuir para o
estabelecimento de métodos de conservação de sementes de Gliricidia sepium, este
trabalho foi realizado com o objetivo de avaliar a qualidade fisiológica das sementes,
submetidas a diferentes condições e períodos de armazenamento. Sementes de gliricidia
recém coletadas foram analisadas e o restante armazenadas por três, seis, nove e doze
meses em sacos plásticos e de papel em geladeira e à temperatura ambiente. Para cada
ambiente e período de armazenamento testado foram analisados a taxa de germinação
relacionandoa com o teor de água das sementes. Observouse que a capacidade
germinativa de sementes de gliricidia está totalmente relacionada à tolerância da espécie
ao ganho ou a perda de umidade das sementes. O armazenamento em saco plástico foi o
que proporcionou melhores condições de conservação das sementes, principalmente
quando acondicionadas em temperaturas baixas. No entanto, mesmo em temperaturas
em torno de 25°C é possível prolongar a viabilidade das sementes de gliricidia quando
se controla o teor de água inicial das mesmas através da utilização de embalagens
impermeáveis para o armazenamento.
Palavraschave: germinação, teor de água, qualidade fisiológica, gliricidia

24
INTRODUCTION
“Gliricidia” [(Gliricidia sepium (Jacq.) Steud.) (Leguminosae –
Papilionoideae)] is a native species of Central and South America. It was introduced
into northeastern Brazil in the 1980’s and has since become the object of significant
commercial and economic interest for the region due to its multiple uses which include
its use as a forage species with high nutritive value for grazing animals and bees, and as
a medicinal plant, among others. Drought resistance is one of the most important
characteristics of any plant to be cultivated in the caatinga region, and “gliricidia”
stands out in this respect because of its capacity to readily grow from seed and because
of its rapid growth, high capacity for regeneration, resistance to drought, among other
characteristics (Drumond & Carvalho Filho, 1999). Studies of seed physiological
processes represent the starting point for the rational use of native or cultivated plant
species from the caatinga region for which germination studies are still very scarce.
The seeds of a majority of cultivated species demonstrate orthodox
characteristics, in which an increase in their water content during storage will result in a
rapid loss of viability and a reduced germination percentage. The conservation potential
of any seed depends directly on the physiological quality of these propagules at the start
of the storage period.
Desiccation tolerance is acquired progressively during seed development.
Among orthodox seeds, desiccation tolerance occurs before the seeds experience any
severe reduction in water content but it is not yet possible to determine if this capacity
is acquired beforehand, or if it is directly due to water loss (Bewley & Black, 1994).
Seed storage involves a series of procedures designed to preserve germination
quality; however, good results depend as much on the nature of the seeds themselves as
on the storage conditions (Santana, 2007). The packaging used in storing seeds exerts

25
an important role in the maintenance of their initial vigor. Additionally, environmental
humidity and storage temperature are other factors that can directly influence the
retention of viability (Ward & Powell, 1983).
Information about the water content of seeds allows one to choose the most
adequate procedures – from harvesting to storage that will preserve their physical,
physiological and sanitary qualities, and will aid in the identification of procedural
problems that might arise and in the adoption of adequate responses (Marcos Filho et
al., 1987).
Seed conservation is very important, and adequate storage will diminish the rate
of their deterioration, which is an irreversible process (Delouche et al., 1973; Melo et
al., 1998, cited by Cabral et al., 2003).
Seed deterioration occurs at molecular, genetic, cellular, tissue, and population
levels (Matthews, 1985). It is a process that cannot be avoided, only controlled, and this
is the principal objective of all storage protocols. Seed storage represents one of the
most utilized strategies of ex situ species conservation, as it preserves the genetic
characteristics of the seeds until they are sewn (Carneiro, 1985, Nodari et al., 1998).
With the objective of contributing to the methodology of the conservation of
gliricidia seeds, the present work sought to evaluate the physiological quality of seeds
that were submitted to different storage conditions for different periods of time for
identifying the best conditions for seed storage is of fundamental importance in
determining strategies for their conservation.

26
MATERIAL AND METHODS
Obtaining and storing seeds
Mature seeds were collected in 2006 from G. sepium trees –in Fazenda
Morrinhos, a farm located in the county of Queimadas, state of Bahia (W 39º 40 ׳ S 11º
which ,( ׳ 12 is considered ‘Private Reserve of Natural Patrimony’ under the Brazilian
law. Samples of these recently collected seeds were cleaned and analyzed for their water
content and germination rate, as an initial characterization of the lot.
The remainder of the seeds were placed into paper or plastic bags and stored at
room temperature (25°C ± 3) or in a refrigerator (8°C ± 2) during three, six, nine, or
twelve months in the Germination Laboratory of the Horto Florestal Experimental Unit
UEFS – Feira de Santana, Bahia State, Brazil, for posterior evaluation.
Germination tests
After each experimental storage period, four subsamples of 25 seeds were sown
onto Petri dishes containing two layers of germination paper that had been moistened
with a quantity of distilled water corresponding to 2.5 times the weight of the paper
substrate. The seeds were maintained in a germination chamber at 25°C with a
photoperiod of 14 h for a period of ten days. The germinating seeds were examined
daily and any propagules with its radicule protruding at least 2 mm from the seed coat
was considered to be germinating, and these individuals were counted and subsequently
discarded.

27
Water content
Seed water content was determined by drying them in an oven at 60°C until
obtaining a constant weight, which was then subtracted from the previously determined
fresh weight. Twentyfive seeds were used in each subsample.
Statistical analyses
Data related to water content and germination was expressed in percentages, and
the standard error calculated. The data of germination, average germination time,
average germination velocity, and germination velocity was transformed by the function
(x + 0.5) 0.5 and submitted to variance analysis using the Sisvar 4.3 software program
(Ferreira, 2000). Differences between the averages were compared using the Tukey test
at a 5% probability level.

28
RESULTS
While germination percentages changed in response to storage time and the
packaging used, the average germination time remained essentially constant (Table 1)
and independent of these two variables.
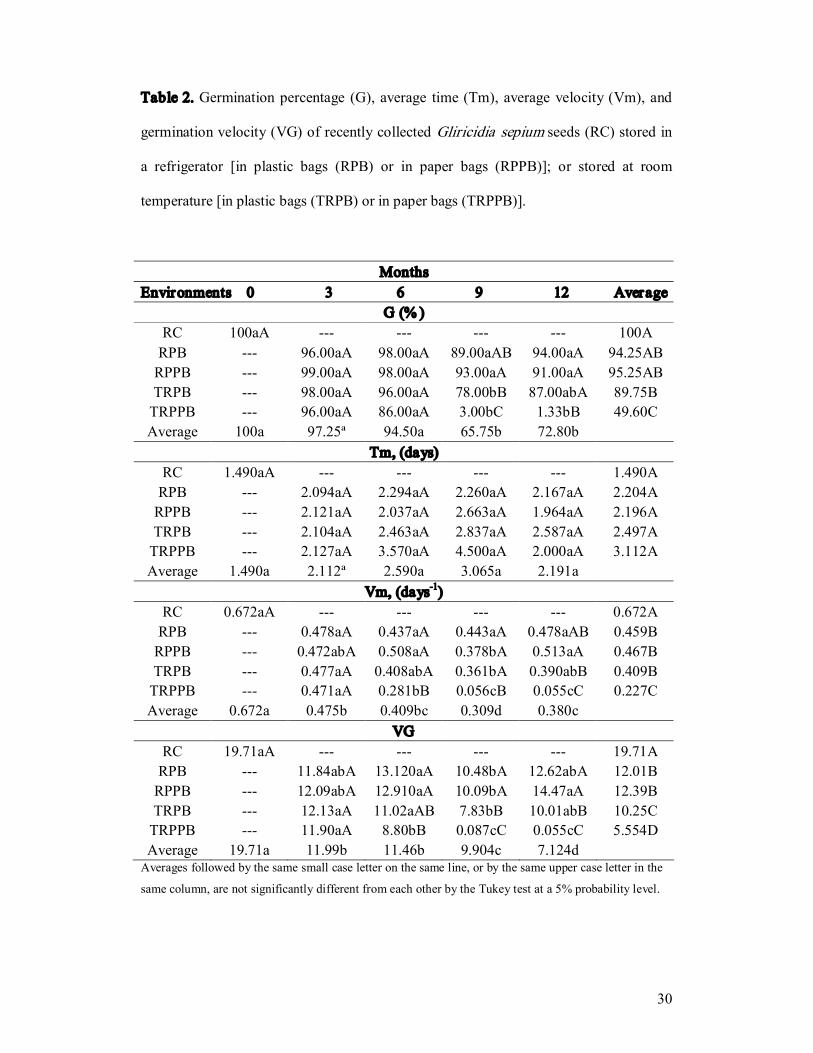
Storage in plastic bags provided the best conditions for seed storage under both
cold and room temperature storage conditions based on percentage germination.
However, seeds stored at room temperature in plastic bags did demonstrate a significant
decrease in germination potential starting at nine months (Table 2).
Changes in water content of Gliricidia sepium seeds over time during storage
depended on the type of packaging used. Initial seed water content was 6.5%, and this
percentage was maintained for the entire period in which the seeds were stored in the
refrigerator in plastic bags. High germination rates were observed (94%) even after 12
months of storage (Figure 1A).
Table 1. Variance analysis of the germination percentage (G), average germination time
(Tm), average germination velocity (Vm), and germination velocity (VG) of stored
Gliricidia sepium seeds.
Var iable G % Tm, days Vm, days 1 VG
Storage time 74.01* 1.143 ns 41.336* 125.77*
Environments 132.97* 0.643 ns 75.075* 216.67*
Time* Environ. 52.46* 0.337 ns 4.569* 36.37*
CV% 7.85 24.26 3.12 5.67 ns = (not significant); * = significant to a 5% level

29
The germination capacity of any species is very dependent on its tolerance to
gaining or losing seed humidity. Figure 1B shows that when the water content of G.
sepium seeds attained lower than initial levels, germination dropped off. Seeds stored in
paper bags at room temperature demonstrated water gains after six months of storage
(Figure 1D) and a concomitant lose of viability. The results point to alterations in seed
carbohydrate reserves due to the increase in their water content.
Under low temperature conditions (refrigeration), gliricidia seeds stored in
impermeable (plastic) bags (Figure 1A) and permeable (paper) bags (Figure 1B)
demonstrated high germination rates and high average germination velocities up to one
year after initial storage (Table 2). Seeds stored at room temperature demonstrated a
decrease in average germination velocity during the year, independent of the packaging
used, as well as a decrease in percent germination (Table 2). Significant reductions in
all of these parameters were observed starting in the sixth month of storage.
Significant differences in the velocity of seed germination were observed when
the seeds were stored at room temperature in paper bags but only after six months of
storage (Table 1). This reduction in germination speed can be attributed to a loss of
vigor in seeds kept under uncontrolled environmental conditions, in comparison to
storage in plastic bags at cold temperatures.
These results suggest that storing gliricidia seeds at low temperatures and in
impermeable packages is much more suitable than holding them at room temperature
and in permeable packaging, for seeds subjected to the latter conditions demonstrated a
much higher loss of viability.
30
Table 2. Germination percentage (G), average time (Tm), average velocity (Vm), and
germination velocity (VG) of recently collected Gliricidia sepium seeds (RC) stored in
a refrigerator [in plastic bags (RPB) or in paper bags (RPPB)]; or stored at room
temperature [in plastic bags (TRPB) or in paper bags (TRPPB)].
Months Environments 0 3 6 9 12 Average
G (% ) RC 100aA 100A RPB 96.00aA 98.00aA 89.00aAB 94.00aA 94.25AB RPPB 99.00aA 98.00aA 93.00aA 91.00aA 95.25AB TRPB 98.00aA 96.00aA 78.00bB 87.00abA 89.75B TRPPB 96.00aA 86.00aA 3.00bC 1.33bB 49.60C Average 100a 97.25ª 94.50a 65.75b 72.80b
Tm, (days) RC 1.490aA 1.490A RPB 2.094aA 2.294aA 2.260aA 2.167aA 2.204A RPPB 2.121aA 2.037aA 2.663aA 1.964aA 2.196A TRPB 2.104aA 2.463aA 2.837aA 2.587aA 2.497A TRPPB 2.127aA 3.570aA 4.500aA 2.000aA 3.112A Average 1.490a 2.112ª 2.590a 3.065a 2.191a
Vm, (days 1 ) RC 0.672aA 0.672A RPB 0.478aA 0.437aA 0.443aA 0.478aAB 0.459B RPPB 0.472abA 0.508aA 0.378bA 0.513aA 0.467B TRPB 0.477aA 0.408abA 0.361bA 0.390abB 0.409B TRPPB 0.471aA 0.281bB 0.056cB 0.055cC 0.227C Average 0.672a 0.475b 0.409bc 0.309d 0.380c
VG RC 19.71aA 19.71A RPB 11.84abA 13.120aA 10.48bA 12.62abA 12.01B RPPB 12.09abA 12.910aA 10.09bA 14.47aA 12.39B TRPB 12.13aA 11.02aAB 7.83bB 10.01abB 10.25C TRPPB 11.90aA 8.80bB 0.087cC 0.055cC 5.554D Average 19.71a 11.99b 11.46b 9.904c 7.124d Averages followed by the same small case letter on the same line, or by the same upper case letter in the
same column, are not significantly different from each other by the Tukey test at a 5% probability level.

31
Figure 1. Water content and germination percentage of the seeds of G. sepium stored in
a refrigerator [in plastic bags (RPB) – A or in paper bags (RPPB) B], or stored at room
temperature [in plastic bags (TRPB) – C or in paper bags (TRPPB) D], during 0, 3, 6,
9, or 12 months. Average of four trials ± ASE.
0 1 2 3 4 5 6 7 8
0 3 6 9 12
Water content (%
)
0 10 20 30 40 50 60 70 80 90 100
WC % G (RPB)
0
2
4
6
8
10
12
0 3 6 9 12 0 10 20 30 40 50 60 70 80 90 100
(%) G
ermination
WC % G (TRPPB)
0 1 2 3 4 5 6 7 8 9 10 11
0 3 6 9 12
Months Storage
Water content (%
)
0 10 20 30 40 50 60 70 80 90 100
WC % G (TRPB)
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14
0 3 6 9 12
Months Storage
0 10 20 30 40 50 60 70 80 90 100
(%) G
ermination
WC % G (TRPPB)

32
DISCUSSION
Variations in water content are of fundamental importance in seed storage,
especially in terms of carbohydrate metabolism, as seed sugar levels are generally
affected by storage temperature. The quantity of sucrose found in seeds of the legume
Caesalpinia echinata Lam. is related to its desiccation tolerance, and high sucrose
levels were encountered in viable seeds stored for 18 months at 7°C (Barbedo et al.,
2002).
Ribeiro et al. (personal communication) observed that an increase in seed water
content in Gliricidia sepium initiated a series of reactions that contributed to their
deterioration by diminishing their sugar content. The accumulation of soluble
carbohydrates in seeds seems to be involved in the desiccation tolerance and longevity
observed in this species. 1
Permeable wrappings that permit water vapor exchange between the seeds and
the atmosphere are more adequate for use in dry environments during short storage
periods. Freitas et al. (1992) reported that seeds maintained in highly permeable
wrappers in humid environments are subject to relatively rapid deterioration.
Cisneiros et al. (2003) observed that seeds of Psidium guineense stored in paper
sacks demonstrated an increase in their water content even when stored in a freezer, and
their ability to germinate became greatly reduced over time.
Maluf & PisciottanoEreio (2005) reported that the best conditions for storing
and conserving seeds of Campomanesia phaea was in plastic bags under controlled cold
conditions. Plastic and glass wrappings (Cisneiros et al., 2003) function as barriers to
gas exchange between the seeds and their external environment, thus reducing their
1 Ribeiro et al. Reserve mobilization during imbibition of stored Gliricidia sepium (Jacq.) Steud. seeds. 2008.

33
metabolism and retarding the processes involved in their deterioration; and these
beneficial effects become more accentuated at lower storage temperatures.
The present work established that the physiological quality of gliricidia seeds
was affected by the permeability of the wrapping material used during their storage. It
is interesting to note, however, that these results are different from those obtained for
other species in the semiarid region, such as Anadenanthera macrocarpa,
Pseudobombax simpliciflolium, Myracrodruon urundeuva (Souza et al., 1980), and
Tabebuia impetiginosa (Cabral et al., 2003) when stored in a cold box in both cloth and
polyethylene bags under conditions of low relative humidity.
According to Carvalho and Nakagawa (2000), the type of packing used for seed
storage is very important in terms of their preservation, viability, and vigor, and the
choice of packing material will depend on the environmental conditions under which
the seeds will be stored. Cold and dry environments are more favorable for storing
orthodox seeds (Villela & Perez, 2004).
Germination velocity is one of the oldest measures of seed vigor (Vieira &
Carvalho, 1994). Seed lots with similar germination percentages frequently
demonstrate differences in germination velocity indicating differences in physiological
vigor (Krzyzanowkski et al., 1999). The changes in germination velocity observed in
gliricidia seeds stored at room temperature thus indicate decreasing overall
physiological vigor.
According to Drumond & Carvalho Filho (1999), gliricidia seeds lose their
viability or enter into secondary dormancy after 12 months in storage. Successful
conservation of gliricidia seeds is intimately related to their initial water content, and
the permanence of that water content at that same level during storage. Maintenance of

34
high viability, and consequently a greater degree of tolerance to storage, was obtained
under low temperature conditions (refrigeration).
The results obtained in the present study are promising in terms of the
conservation of gliricidia seeds for they indicate that these propagules can be conserved
with high viability for reasonably long periods of time (up to 12 months), and even at
temperatures near 25°C, if the initial water content of the seeds is maintained through
the use of impermeable storage containers.
ACKNOWLEDGEMENTS
The authors would like to thank the Conselho Nacional de Desenvolvimento
Científico e Tecnológico (CNPq) and the Fundação de Amparo a Pesquisa do Estado da
Bahia (FAPESB) for financial support of this project, and José Juracy for his help with
plant material.

35
REFERENCES
BABEDO, C.J.; BILIA, D.A.C.; FIGUEIREDORIBEIRO, R.C.L. Tolerância à
dessecação e armazenamento de sementes de Caesalpinia echinata Lam. (paubrasil).
Rev. Bras. de Bot. 25: 431439, 2002.
BEWLEY, J.D.; BLACK, M. Seeds: Physiology of Development and Germination.
2 th ed. Plenum Press, New York, 1994.
CABRAL, E.L.; BARBOSA, D.C.A.; SIMABUKURO, E.A. Armazenamento e
germinação de sementes de Tabebuia áurea (Manso) Benth. & Hook. F. Ex. S. Moore.
Acta. Bot. Bras. 17(4): 609617, 2003.
CARNEIRO, J.G.A. Armazenamento de sementes florestais. curitiba: FUPEF, 35 p.
(Série Técnica, 14), 1985.
CARVALHO, N.M.; NAKAGAWA, A.J. Sementes: ciência, tecnologia e produção. 4
ed. Jaboticabal: FUNEP, 588p, 2000.
CISNEIROS, R.A.; MATOS, V.P.; LEMOS, M.A.; REIS, O.V. DOS; QUEIROZ, R.M.
de. Qualidade fisiológica de sementes de araçazeiro durante o armazenamento. Rev.
Bras. de Eng. Agr.e Amb., 7(3): 513518, 2003.
DRUMOND, M.A.; CARVALHO FILHO, O.M. de. Introdução e avaliação de
Gliricidia sepium na região semiárida do Nordeste Brasileiro. In: QUEIRÓZ, M.A. de;
GOEDERT, C.O.; RAMOS, S.R.R. (ed.) Recursos genéticos e melhoramento de
plantas para o Nordeste brasileiro. Versão 1.0, 1999. PetrolinaPE: Embrapa Semi
Árido /BrasíliaDF: Embrapa Recursos Genéticos e Biotecnologia, nov.
http://www.cpatsa.embrapa.br.

36
FERREIRA, D.F. Análises Estatísticas por meio do Sisvar para Windows versão
4.0. In. 45° Reunião Anual da Região Brasileira da Sociedade Internacional de
Biometria. UFS Car, São Carlos, SP, Julho, p. 255258, 2000.
FREITAS, G.B.; SILVA, R.F.; ARAÚJO, E.F. Influência da condição de
armazenamento na qualidade de sementes de milho. Rev. Bras. de Armaz., Viçosa,
MG. 17(1): 2126, 1992.
KRZYZANOWSKI, F.C.; VIEIRA, R.D.; FRANÇA NETO, J.B. Vigor de Sementes:
conceitos e testes. Londrina: ABRATES, 218 p, 1999.
MALUF, A.M.; PISCIOTTANOEREIO, W.A. Secagem e armazenamento de sementes
de cambuci. Pesq. Agropec. Bras. 40(7): 707714, 2005.
MARCUS FILHO, J.; CÍCERO, S.M.; SILVA, W.R. Avaliação da qualidade das
sementes. Piracicaba: FEALQ, 230p, 1987.
MATTHEWS, S. Physiology of seed ageing. Outlook on Agr iculture, 14(2): 1923,
1985.
NODARI, R.O. et al. Conservação de frutos e sementes de palmiteiro (Euterpe edulis
Matius) sob diferentes condições de armazenamento. Rev. Árvore, 22(1): 110, 1998.
SANTANA, P.J.A. Maturação, secagem e armazenamento de sementes de espécies
de Eugenia (Myr taceae). Dissertação Mestrado, São PauloSP, 2007.
SOUZA, S.M.; PIRES, I.E.; LIMA, P.C.F. Influência da embalagem e condições de
armazenamento na longevidade de sementes florestais. p. 1524. In: Pesquisa Florestal

37
no Nordeste Semiárido: sementes e mudas. Boletim de pesquisa n.2, Embrapa –
CPATSA, Petrolina, 1980.
VIEIRA, R.D.; CARVALHO, N.M. Testes de vigor em sementes. Jaboticabal,
FUNEP, 164p, 1994.
VILLELA, F.A.; PERES, W.B. Coleta, beneficiamento e armazenamento. In:
FERREIRA, A.G.; BORGHETTI, F. (eds), Germinação: do básico ao aplicado.
pp.265281, Porto Alegre: Artmed, 2004.
WARD, F.H.; POWELL, A.A. Evidence for repair processes in onion seeds during
storage at high seed moisture contents. J . Exp. Bot. 34(140):277282, 1983.

38
CAPÍTULO II
Reserve mobilization dur ing imbibition of stored Gliricidia sepium (Jacq.)
Steud. seeds.

39
ABSTRACT – (Reser ve mobilization during imbibition of stored Gliricidia sepium
(Jacq.) Steud. seeds). Gliricidia sepium (Leguminosae Papilionoideae) is a species
which is tolerant to drought and can be easily propagated by seeds. Therefore, it has
been explored by farmers as a source of forage in the semiarid region of the northeast
of Brazil. The present study has been developed to evaluate the effect of seed storage on
the mobilization of reserves during imbibition of Gliricidia seeds. Seeds were freshly
harvested, packed in paper bags and stored for three and six months at the laboratory
under environmental conditions (temperature and humidity). Cotyledons were isolated
from embibed seeds and macerated for the extraction and quantification of total soluble
sugars, reducing sugars, sucrose and starch, as well as of proteins and amino acids.
Storage under such conditions allowed an increase in seed water content, although
maintaining seed germinability at relatively high levels (86%). Seed macromolecule
levels showed significant variation in relation to the period of storage and of imbibition
being associated with the loss of seed viability due to inadequate conditions of storage.
Key words: Gliricidia sepium, seeds, storage, longevity, germination, reserve
mobilization.

40
RESUMO – (Mobilização de reservas durante a embebição de sementes de
Gliricidia sepium (Jacq.) Steud. (Leguminosae Papilionoideae) armazenadas).
Gliricidia sepium é uma espécie que apresenta resistência à seca e facilidade em se
propagar por sementes, sendo intensamente explorada como forrageira no semiárido
nordestino. O presente estudo foi desenvolvido para avaliar o efeito do armazenamento
na mobilização de açúcares e proteínas e as possíveis modificações dessas
macromoléculas de reserva durante a germinação de sementes de gliricidia. Sementes
recém coletadas foram armazenadas por três e seis meses em sacos de papel à
temperatura ambiente. Os cotilédones das sementes embebidas foram extraídos e
macerados para a quantificação dos açúcares solúveis totais, açúcares redutores,
sacarose, amido, proteínas e aminoácidos. O armazenamento em embalagem de papel
em temperatura ambiente propiciou o aumento no teor de água embora a capacidade
germinativa foi mantida em níveis altos (86%). O armazenamento e o tempo de
embebição influenciaram os níveis de macromoléculas nas sementes. A variação nos
teores de macromoléculas esta associada com a perda da viabilidade das sementes
proporcionada pela condição inadequada de armazenamento.
Palavraschave: Gliricidia sepium, sementes, armazenamento, longevidade,
germinação, mobilização de reservas.

41
INTRODUCTION
Gliricidia (Gliricidia sepium (Jacq.) Steud.) (Leguminosae Papilionoideae) is
a native species from the South and Central Americas and it was introduced in the
Brazilian northeast region during the 80´s. It has commercial and economic interests for
the region due to its multipurpose characteristics combined to drought tolerance,
among which a high nutritional value, therefore, an important alternative source of
forage in the composition of living fences and pastures. The rigorous climatic
conditions of the brazilian semiarid is occurs mainly between May and October, due to
lack of rains, strong and dry winds that contribute to dryness and arid landscape
(Sampaio & Rodal, 2000; Teixeira, 2001). During this period, seeds may naturally also
be kept viable in the soil seed bank, e.g. orthodox seeds (Baskin & Baskin, 1998).
Therefore, drought resistance is one of the most important characteristics for
maintaining the viability of seeds during the dry periods and for cultivating this tree
species in the region. In this context, gliricidia stands out as it propagates easily by
seeds, has fast growth, high regeneration capacity, drought resistance, among other
referred characteristics (Drumond & Carvalho Filho, 1999). Although it does exist
significant knowledge relative to plant vegetative adaptation characteristics to semiarid
environmental conditions, much is yet unknown about seed adaptation mechanisms
under such conditions, while soil seed banks consist in one of the main survival
strategies of wild plant species (Baskin & Baskin, 1998).
Maturation of orthodox seeds is characterized by intense deposition of reserves,
after which it undergoes an exclusive process of rapid water reduction, what seems to
vary according to the different species and prevalent climatic conditions. Dehydration
leads these seed species into a state of metabolic quiescence, allowing seed survival
until conditions favorable for germination are reestablished (Bewley & Black, 1985).

42
During this period, seed storage conditions are of extreme importance so that viability is
maintained to the most, e.g. longevity, whereas water uptake and storage temperature
are the most critical factors (Ward & Powell, 1983). Therefore, the potential for seed
conservation or longevity directly depends on the initial seed (physiological) quality
before the storage period.
Dissection tolerance is a common feature of orthodox seeds which is generally
attained during maturation and that allows seeds to be stored for different periods of
time, i.e. many years in some cases, without significant loss of viability (Maluf &
PisciottanoEreio, 2005). Such tolerance has been related to the capacity of an
organism, i.e. seeds, of facing the extreme stress condition of almost complete loss of
water during maturation and its rehydration during the germination process (Hoekstra et
al., 2003).
After maturation, seeds are subjected to a series of degenerative changes of
physiological, physical and biochemical origins which are associated with the reduction
of seed vigor (Alizaga et al., 1990). The occurrence in seeds of some processes such as
the Amadori and Maillard reactions can contribute for deterioration and the loss of seed
viability. These reactions apparently leads to metal complexation, therefore it can
reduce protein digestibility, inhibit the action of digestive enzymes, destroy amino acids
and ascorbic acid and intervene in mineral metabolism (Araújo, 1995). Once these
reactions are triggered by an increase in the water content of quiescent seeds, the
environmental conditions and the type of supplied reserves will significantly influence
the maintenance of physiological quality of the seeds.
The germination process initiates with seed imbibition as a result of water
uptake. This occurs due to a difference in osmotic potential between the dry seed and
the moist environment or substratum. However, it necessary that the seeds reaches an

43
adequate level of hydration, which allows the reactivation of seed metabolic processes
such as reserve mobilization, so that the resulting metabolites can be used by the
germinating embryo and on subsequent seedling growth (Bewley & Black, 1994).
In fact, much is still unknown about the mechanisms acting on the maintenance
of viability, longevity and conservation, and of the processes regarding reserve
mobilization during germination of seeds species that are common to or adapted to the
northeast semiarid region of Brazil. Therefore, it has been carried in the present study
an ex situ evaluation of possible modifications on the composition of reserves as a result
of storage of G. sepium seeds under laboratory controlled conditions, as well as their
subsequent mobilization during germination. In this way, we sought to analyze possible
effects of storage over the initial seed physiological quality in search of comparative
parameters that could apply to the natural conditions in which gliricidia seeds are kept
as viable in soil seed banks of the northeastern semiarid. Ultimately, analyze if those
parameters could contribute as evidences for outstanding rusticity and tolerance of this
species to the adverse draught stress conditions that are common to this region of Brazil.

44
MATERIAL AND METHODS
Seed origin and storage
Mature seeds were collected in 2006 from G. sepium trees –in Fazenda
Morrinhos, a farm located in the county of Queimadas, state of Bahia (W 39º 40 ׳ S 11º
which ,( ׳ 12 is considered ‘Private Reserve of Natural Patrimony’ under the Brazilian
law. Freshly collected seeds were processed and subsequently used for laboratory
analysis or packed in paper bags and submitted to storage for three and six months
under environmental conditions at the Germination Laboratory of the State University
of Feira de Santana (Laboratório de Germinação, Unidade Experimental Horto Florestal
– UEFS, Feira de Santana, Bahia), where water content and laboratory germination tests
were conducted at the end of each storage period.
Germination
Germination analysis was conducted on freshly collected seeds and on seeds
after each period of storage, using four replicates of 25 seeds placed over two layers of
germination paper (Germitest) in 9 mm Petri dishes soaked with an amount of water
equivalent to 2.5 times the weight of the substratum and incubated in germinator
chambers at 25°C and at photoperiod regime of 14 h for a period of 10 consecutive
days. Seeds were considered as germinated when their radicles had protruded at least 2
mm, after when they were scored and discarded.
Water content
Water content was determined on freshly collected seeds and on seeds after each
period of storage by the oven method (fresh weight basis), in which four replicates of 25

45
seeds were weighted before and after being submitted to 60°C, until weight
stabilization.
Extraction and analysis of macromolecules (carbohydrates and proteins)
Macromolecules analysis was performed at the Laboratory of Seed and Plant
Physiology, at the Embrapa SemiArid Research Center, in Petrolina, state of
Pernambuco, Brazil. Four replicates of 25 seeds were incubated in a germination
chamber at 25°C for a period of four days, as for the germination test. Germination was
scored and cotyledons isolated into reaction tubes at 24 h intervals until completing 96 h
and subsequently frozen and stored at 20°C until the macromolecules extraction
procedures.
Macromolecules were extracted from samples of 1 g of fresh cotyledons that had
been homogenized in 5 mL ethanol 80% and centrifuged at 10,000 g during 20 min at
5°C. The procedure was repeated four times and the supernatants collected from each
constituted sample (Seiffert, 2003).
Soluble compounds were quantified spectrophotometrically in aliquots of the
ethanolic extracts. Total soluble sugars (TSS) were quantified by the method of
reactions with antrona (Morris, 1948; Yemm & Willis, 1954); reducing sugars (RS) by
the method of oxidation with 3,5dinitrosalicilic acid (DNS) (Miller, 1959); sucrose
(SUC) through antrona cold reaction, after addition of hot KOH (Passos, 1996); total
proteins (TP) according to Bradford method (1976) and amino acids (AA) according to
Rosen method (1957).
The pellets obtained after each centrifugation were dried in incubator at 60°C for
approximately 18 h (overnight). The dried materials were macerated and 0.1 g samples
put into microtubes for posterior starch digestion (S) with perchloric acid (Allen et al.,

46
1977). The sugars obtained from the starch digestion were quantified through TSS
concentration present in each sample (Morris, 1948; Yemm & Willis, 1954).
Statistical analysis
Germination and water content data were expressed as a percentage and
established the standard mean error. Macromolecule data were analyzed by means of
variance analysis using the Sisvar 4.3 software program (Ferreira, 2000). The averages
were compared by Tukey´s test at 5% probability. For the adjustment of regression
equations, it was selected the significative model which showed the highest
determination coefficient.

47
RESULTS
The contents of macromolecules in seed cotyledons were significantly affected
by variations in time of seed storage and imbibitions, except for the contents of starch
and proteins which was not influenced by the imbibitions time. In general, the contents
of all macromolecules varied when analyzed in relation to the combined effects of
storage and imbibition times (Table 1).
Germination percentage decreased from 100% in freshly collected gliricidia
seeds to 86% in seeds after storage, whereas the water content increased from 6,5% to
13,5% in seeds packed in paper bags after six months of storage under environmental
conditions at the laboratory (Figure1). It observed a decrease of 14% in total percentage
of germination in seeds with higher water content after storage.
Table 1. Values for variance analysis of macromolecule contents present in G. sepium
seeds during stored and imbibition. ST= starch; SUC= sucrose; TSS= total soluble
sugars; RS= reducing sugars; TP= total proteins; AA= amino acids.
ns; *= not significant, significant at 5% respectively
Source of var iation ST SUC TSS RS TP AA
Storage 72.324 * 331.401 * 178.859 * 106.247 * 7.147 * 33.644 *
Imbibition 1.424 ns 108.606 * 81.994 * 10.868 * 1.755 ns 3.233 *
Storage*Imbibition 18.658 * 15.264 * 7.337 * 3.680 * 3.311 * 5.950 *
CV % 25.34 21.02 14.96 18.18 18.71 11.30

48
Figure 1. Water content (%) and Total germination (%) of G. sepium seeds during
storage (m). Average of four replicates ± ASE.
The germinability of stored gliricidia seeds decreased in comparison to the
freashly harvested seeds, as it can be seen from the percentage of seeds that had
germinated during imbibition up to 96 h (Figure 2). Concomitant with this result TSS
content was significantly reduced in cotyledons (Figure 3a). It can be observed in Table
2 that the reduction was significant along storage mainly after six months where the
lowest mean values were observed.
The contents of ST and SUC apparently had a slight decrease after three months
of storage but increased significantly after six months of storage as it can be seen in
seeds imbibed for 24 h (Table 2). Despite that, the contents of SUC decreased during
imbibition of freshly harvested seeds as well as of stored seeds as an evidence of its
mobilization by the embryonic axis (Figure 3B). However, this seems not to be the case
0 2 4 6 8 10 12 14 16
0 3 6
Storage time (m)
Water content (%
)
50 55 60 65 70 75 80 85 90 95 100
Germination (%
)
WC % G

49
for seeds stored for six months which showed reduction in germination percentages
during the same periods (Figure 2) and, therefore, indicating that SUC may have been
distributed to other metabolic pathways.
0 10 20 30 40 50 60 70 80 90 100
24 48 72 96
Imbibition time (h)
Germination (%
)
0 M 3 M 6M
Figure 2. Germinability (%) during imbibition (h, hours) of G. sepium freshly harvested
seeds and after 3 and 6 months of storage (m). Average of four repetitions ± ASE.
Starch content kept about the same level until 72 h of imbibition in cotyledons of
freshly harvested and stored seeds (Figure 3C), after which the contents decreased in
freshly harvested seeds and in seeds stored for three months. This reduction seems also
to coincide with mobilization by the embryonic axis, possibly as a substrate for the
development of the seedling once seeds had initiated the germination process 48 h after
imbibition (Figure 2). On the other hand, a steep increase in ST content was observed in
seeds that were stored for six months, in which an apparent resynthesis of ST occurred
after 96 h of imbibition (Table 2).

50
Figure 3. Macromolecule contentes (µmol/g.DW or µmol/g.FW) – A. Total soluble
sugars; B. Sucrose; C. Starch; D. Reducing sugar; E. Total proteins; F. Amino acids; in
cotyledons during imbibition of G. sepium freshly harvested seeds or after 3 and 6
months of storage (m). Average of six replicates.
0
1000
2000
3000
4000
5000
0 24 48 72 96
Total so
lube su
gars (µ
mol/gFW
)
0 m (y1) 3 m (y2) 6 m (y3) A
0 2000 4000 6000 8000 10000 12000 14000 16000 18000
0 24 48 72 96 Reducing sugars(µmol/gFW
)
D
0
1000
2000
3000
4000
5000
6000
0 24 48 72 96
Starch (µ
mol/gDW)
C
0 200 400 600 800 1000 1200 1400 1600 1800
24 48 72 96
Imbibition time (h)
Total proteins (µm
ol/gFW
)
E
0
500
1000
1500
2000
2500
3000
0 24 48 72 96
Sucrose (m
mol/gFW
)
B
y1= 0.6947x 2 + 64.964x + 1866.4 R 2 = 0.9683
y2= 32.125x + 5124.7 R 2 = 0.7733
y3= 16.638x + 0.8201 R 2 = 0.8201
y1= 10130x + 1E+06 R 2 = 0.9944
y2= 5846.4x + 741504 R 2 = 0.9495
y3= 24297x + 3E+06 R 2 = 0.8891
y1= 11.573x + 2553.2 R 2 = 0.6239
y2= 0.4401x 2 + 54.329x + 254.71 R 2 = 0.9929
y3= 1.191x 2 – 116.74x + 5110.2 R 2 = 0.9484
0
1000 2000
3000 4000
5000
6000
7000
8000
24 48 72 96
Imbibition time (h)
Amino acids (µm
ol/gFW
)
F
y1= 2.0434x 2 – 190.58x + 16143 R 2 = 0.9998
y2= 28.375x + 10928 R 2 = 0.1345
y3= 1.467x 2 – 188x + 10863 R 2 = 0.9293
y1= 25.666x + 3638.3 R 2 = 0.6547 y2= 0.2986x 2 + 37.851x + 5409.6 R 2 = 0.846
y3= 11.078x + 7401.6 R 2 = 0.6484
y1= 0.2459x 2 – 24.583x + 1658.3 R 2 = 0.9324
y2= 3.7153x + 1554.2 R 2 = 0.7043
y3= 0.25x + 1097.5 R 2 = 0.143

51
Table 2. Macromolecule contents (µmol/g.DW or µmol/g.FW) in cotyledons during
imbibition and germination of G. sepium freshly harvested seeds or after storage for 3
and 6 months (m). TSS= total soluble sugars; SUC= sucrose; ST= starch; RS= reducing
sugars; TP= total proteins; AA= amino acids.
Storage (m) Imbibition (h) 0 3 6 Average
TSS 24 2973.92 bB 3845.66 aA 1616.27 cAB 28.11.95 48 3538.41 bA 4217.22 aA 1752.76 cA 3169.46 72 2788.14 aB 3066.81 aB 1120.21 bB 2325.05 96 1752.05 aC 1659.16 aC 492.91 bC 1349.86
Average 2763.12 3197.21 1245.54 SUC
24 883.549 bA 561.580 cA 2191.017 aA 1212.049 48 687.121 bAB 512.337 bA 2126.082 aA 1108.514 72 434.415 bB 336.471 bAB 1441.558 aB 737.481 96 157.359 bC 152.489 bB 421.536 aC 244.480
Average 540.611 390.719 1545.048 ST
24 2083.79 bA 1314.80 cA 2892.13 aB 2096.91 48 2117.75 abA 1819.37 bA 2558.37 aB 2165.17 72 2054.68 aA 1913.98 aA 2571.99 aB 2180.21 96 1178.95 bB 1404.55 bA 5063.28 aA 2414.95
Average 1858.79 1613.17 3195.35 RS
24 12.757 aA 10.874 aB 7.084 bA 10.238 48 11.671 bA 14.750 aA 5.554 cA 10.658 72 13.047 aA 10.258 bB 4.597 cA 9.300 96 16.669 aB 14.640 aA 6.765 bA 12.947
Average 13.536 12.631 5.898 TP
24 1233.333abAB 1500.000 aA 1083.333 bA 1272.22 48 975.000 aB 1283.333 aA 1108.333 aA 1122.22 72 1233.333 aAB 1366.666 aA 1058.333 aA 1219.44 96 1541.666 aA 1175.000 bA 1141.666 bA 1276.47
Average 1245.833 1331.250 1082.609 AA
24 4461.18 cB 6111.57 bA 7306.21 aA 5959.65 48 4972.57 bB 6641.56 aA 6768.15 aA 6127.42 72 4661.09 bB 6483.49 aA 6295.71 aA 5813.43 96 6618.32 aA 6325.43 aA 6864.39 aA 6502.92
Average 5178.30 6390.52 6743.80 Averages followed by the same small letter in row and the same capital letter in a column do not differ between them for Tukey test at 5% of probability.

52
It was observed a significant increase in RS contents in freshly harvested three
months stored seeds after 96 h imbibition (Figure 3D), whereas the content was kept
low in seeds stored for six months throughout the imbibition period (Table 2).
In general, the TP levels of stored gliricidia seeds had little alteration throughout
the period of imbibition. The lowest levels of TP had been verified in seeds stored for
six months and those levels remained low during the period of imbibition (Figure 3E).
The behaviour of seeds stored for three months also showed a significant reduction,
reaching levels of TP similar to the ones of seeds stored for six months 96 h after
imbibition. A different protein profile was observed for freshly harvested gliricidia
seeds, whose levels had reduced in the first 48 h and later they had increased until
completing 96 h after imbibition (Figure 3E). The contents of AA had a significant
increase by the third month and continued to increase in the sixth month of storage
(Figure 3F). There were no significant differences between the hours of imbibition for
those two months (Table 2).

53
DISCUSSION
The longevity of seeds is greatly influenced by storage conditions, mainly by
environmental temperature and humidity which can influence the seed water content
and metabolism. In seeds of Myrtaceae, Campomanesia phaea (Maluf & Pisciottano
Ereio, 2005), it was also verified that seeds stored in natural environment and packed in
paper bags lose total viability after 240 days of storage. Conservation of this seed
species was better provided when storage was done in plastic bags under controlled
conditions in cold chambers. The beneficial effect of this type of storage was observed
also for other species of the same family (Barbedo et al., 1998). Cold and dry
environmental conditions are more favorable for orthodox seed storage (Villela &
Peres, 2004). The association between low temperature and impermeable packing is
responsible for a reduction in cell metabolism and it reflects on seed longevity (Figliolia
et al., 1993; Ferreira & Borgehtti, 2004).
It has been reported that TSS had apparently been used in cell respiration or
released to the medium during the initial imbibition phases, as observed in embryos of
the leguminous plant Senna macranthera (Borges et al., 2001). This corroborates our
observations in which the TSS content decreased in cotyledons of imbibing gliricidia
seeds (Figure 3A). Apparently the mobilization of these reserves occurs following
radicle protrusion in Dalbergia miscolobium seedlings where it was clearly observed an
increase in TSS levels in root as in aboveground parts during the initial development of
seedlings (Sassaki & Felippe, 1992).
Part of RS produced during degradation of reserve carbohydrates is used in
cellular respiration of the storage tissue which is the main source of energy during the
beginning of seed germination. Another part of RS is employed for the SUC production

54
that will be carried to the embryonic axis or S synthesis, when there is RS excess in
storage tissues (Bewley & Black, 1994; Nelson & Coxswain, 2000). Synthesized ST is
stored as energy source for germination and subsequent growth of seedlings. According
to Stone & Gifford (1999), in seeds of Pinus taeda starch accumulation occurred in the
megagametofite tissue during radicle protrusion.
As stated by Zeleny (1954), increase in reducing sugars and posterior reduction
in seeds, as observed with gliricidia seeds in the third month of storage (Figure 3D),
seems to be associated with the loss of seed viability due to the fact that these sugars are
associated with reactions such as the Amadori and Maillard reactions which induce seed
deterioration by means of degradation of proteins and amino acids. As a result, SS
levels tend to diminish; however for the species in study an increase in sucrose content
occurred, possibly as a protection mechanism since the synthesis of this disaccharide
seems to be involved in seed protection systems against deterioration processes (Kigel
& Galili, 1995).
Decrease of reducing sucrose during seed imbibition occurred in rubber tree
seeds, as well as in gliricidia seeds (Figure 3D), partially explaining the process and use
of these sugars in seed metabolic processes. However De Paula et al. (1998), suggest the
participation of RS in Amadori and Maillard reactions.
On the other hand, the increase in hydration levels of gliricidia seeds as observed
in after the third month of storage (Figure 1) may have triggered the activation of
invertases and αgalactosidases. According to Wettlaufer & Leopold (1991), relatively
high levels of seed hydration are necessary so that there is activation of these same
enzymes and consequently promotion of increase in RS levels that will directly
participate in Amadori and Maillard reactions and cause deficiency of these compounds
in cells. As stated by Salisbury & Ross (1992), serious disturbances can occur in cells

55
due to loss of RS. This fact can be triggered since some of these sugars are important
intermediate compounds in photosynthetic metabolic pathways and cellular respiration.
They also constitute basic compounds for the synthesis of many other carbohydrates,
including starch and cellulose.
The increase in AA probably occurred due to activation of proteinases present
in seeds which has the role of degrading reserve proteins resulting in free amino acids.
According to Smith & Berjak (1995), these enzymes can increase their activity with the
seed aging process. Later, amino acid levels tend to diminish through their self
destruction, due to Amadori and Maillard reactions, that according to Wettlaufer &
Leopold (1991), is characterized by non enzymatic attack to amine groups by RS. This
influence can be verified in rubber tree seeds (De Paula et al., 1998), as probably also in
G. sepium seeds (Figure 3F).
On most occasions, the lack of knowledge of ideal storage conditions, the
maintenance of physiological seed quality for long periods become more difficult,
therefore the aging of seeds is a natural process. In view of present results it was
verified that the environmental condition used for storage did not allow good
conservation of gliricidia seeds. The alteration in macromolecular composition in seed
cotyledons was apparently responsible for changes associated with seed deterioration
processes throughout the storage period.
It is not possible by means of the present study to conclude that similar
processes occur and what are the factors responsible for maintaining the viability of G.
sepium seeds while stored under the natural conditions of soil seed banks in the semi
arid region. Therefore, further studies are needed in order to better characterize the
viability, longevity and storage tolerance of G. sepium seeds in order to better

56
understand its conservation and its capacity towards successful seedling establishment
and plant growth under adverse conditions.
ACKNOWLEDGMENTS
We would like to thank the following organizations for their valuable and
generous funding and technical support: Conselho Nacional de Desenvolvimento
Científico e Tecnológico (CNPq) Fundação de Amparo a Pesquisa do Estado da Bahia
(FAPESB), Embrapa SemiÁrido. And José Juracy Pereira for providing access to the
biological material at his property.

57
REFERENCES
ALLEN, S.E.; GRIMSHAW, H.M.; PARKINSON, J.A.; QUARMBY, C. Chemical
analysis of ecological mater iais. Oxford: Blackwell Scientific, 1974.
ALIZAGA, R.L.; MELLO, V.D.C.; SANTOS, D.S.B.; IRIGON, D.L. Avaliações de
testes de vigor em sementes de feijão e suas relações com a emergência em campo. Rev.
Bras. de Sement. Brasília, 12(2):4458, 1990.
ARAÚJO, J.M.A. Química de alimentos: teoria e prática. Viçosa: UFV, Imprensa
Universitária, 335p, 1995.
BASKIN, C.C.; BASKIN, J.M. Seeds, ecology, biogeography, and evolution of
dormancy and germination. Academic Press, New York, 1998.
BEWLEY, J.D.; BLACK, M. Seeds: Physiology of Development and Germination.
2 th ed. Plenum Press, New York, 1994.
BORGES, E.E.L., et al. Crescimento e mobilização de carboidrato em embrião de
sementes de fedegoso (Senna macranthera Irwis et Barneby) durante a germinação.
Rev. Cerne. 7(1), 2001.
DRUMOND, M.A.; CARVALHO FILHO, O.M. de. Introdução e avaliação de
Gliricidia sepium na região semiárida do Nordeste Brasileiro. In: QUEIRÓZ MA de,
GOEDERT CO, RAMOS SRR, (ed.) Recursos genéticos e melhoramento de plantas
para o Nordeste brasileiro. Versão 1.0, 1999. PetrolinaPE: Embrapa SemiÁrido
/BrasíliaDF: Embrapa Recursos Genéticos e Biotecnologia, nov.
http://www.cpatsa.embrapa.br.

58
FERREIRA, D.F. Análises Estatísticas por meio do Sisvar para Windows versão
4.0. In. 45° Reunião Anual da Região Brasileira da Sociedade Internacional de
Biometria. UFS Car, São Carlos, SP, Julho, p. 255258, 2000.
HOEKSTRA, F.A.; GOLOVINA, E.A.; NIJSSE, J. What do we know about desiccation
tolerance mechanism? In: NICOLÁS G, BRADFORD KJ, CÔME D, PRITCHARD HD
(eds) The biology of seeds: resent research advances. pp. 259270, Cambridge: CAB
International, 2003.
KIGEL, J.; GALILI, G. Seed development and germination. New York: Marcel
Dekker, 853p, 1995.
MALUF, A.M.; PISCIOTTANOEREIO, W.A. Secagem e armazenamento de sementes
de cambuci. Pesq. Agropec. Bras. 40(7):707714, 2005.
MILLER, G.L. Use of dinitrosalicilic acid reagent for determination of reducing
sugar . Analytical Chemistry, 31:426428, 1959.
MORRIS, D.L. Quantitative determination of carbohydrates with Drywood’s anthrone
reagent. Science. Washington, 107:254255, 1948.
NELSON, D.L.; COX, M.M. Lehninger pr inciples of biochemistry. 3 th , Worth
Publishers, New York, NY, EUA, 2000.
PASSOS, L.P. Métodos analíticos e laboratoriais em fisiologia vegetal. Coronel
Pacheco: EmbrapaCNPGL, 223p, 1996.
ROSEN, H. A modified ninhydrin calorimetric analysis for amino acids. Arch.
Biochem. Biophys, 67:10–15, 1957.

59
SAMPAIO RODAL, M.J. Fitofisionomias da caatinga. Avaliação e identificação de
ações prioritárias para a conservação, utilização sustentável e repartição de benefícios
da biodiversidade do bioma caatinga. In: Biodiver sidade da caatinga,
http//www.biodiversitas.org/caatinga, Petrolina, 2000.
SASSAKI, R.M.; FELIPPE, G.M. Remoção dos cotilédones e desenvolvimento inicial
de Dalbergia miscolobium. Rev. Bras. de Bot. 5:56, 1992.
SEIFFERT, M. Alguns aspectos fisiológicos e bioquímicos da germinação de
sementes e anatomia foliar de Protium widgrenii Engler . Universidade Federal de
Lavras, Dissertação Mestrado, 2003.
STONE, S.L.; GIFFORD, D.J. Structural and biochemical changes in Loblolly pine
(Pinus taeda) seeds during germination and early seedling growth. ll. Storage
triacylglicerols and carbohydrates. Int. J . Plant Science. 160(4):663671, 1999.
TEIXEIRA, A.H.C. Informações Agrometeorológicas do Pólo Petrolina
PE/J uazeiro BA. Documentos. Embrapa SemiÁrido, PetrolinaPE, 2001.
VILLELA, F.A.; PERES, W.B. Coleta, beneficiamento e armazenamento. In:
FERREIRA, A.G.; BORGHETTI, F. (eds), Germinação: do básico ao aplicado.
pp.265281, Porto Alegre: Artmed, 2004.
WARD, F.H.; POWELL, A.A. Evidence for repair processes in onion seeds during
storage at high seed moisture contents. J . Exp. Bot. 34(140):277282, 1983.
WETTLAUFER, S.H.; LEOPOLD, A.C. Relevance of Amadori and Maillard products
to seed deterioration. Plant Physiol. Rockville, 97(1):165169, 1991.

60
YEMM, E.W.; WILLIS, A.J. The estimation of carbohydrates in plants extracts by
anthrone. Biochem. J . Colchester, 57:508514, 1954.
ZELENY, L. Chemical, physical, and nutrive changes during storage. In: ANDERSON,
J.A.; ALCOCK, A.W. (eds.) Storage of cereal grains and their products. pp. 4676.
Minnesota: American Association of Cereal Chemists, 1954.

61
CAPÍTULO III
Amylolitc activity and physiological quality of Gliricidia sepium (Jacq.) Steud.
(Leguminosae Papilionoideae) seeds during storage.

62
ABSTRACT – (Amylolitc activity and physiological quality of Gliricidia
sepium (Jacq.) Steud. (Leguminosae Papilionoideae) seeds during storage). The
principal objective of seed storage is to maintain propagule quality. Storage is a very
critical procedure, as seeds are subject to a series of degenerative changes very soon
after being dispersed. The present work sought to evaluate the changes in the
physiological quality and in reserve mobilization of Gliricidia sepium seeds during
storage. Recently collected gliricidia seeds were analyzed immediately or stored for
three and six months in paper bags at room temperature. At the end of each storage
period the seeds were analyzed to determine their germination rate, water content, and
amylase activity during swelling. Seeds that had been stored for six months
demonstrated a reduction in vigor that was correlated with an increase in water content
and alterations in their enzymatic activity. The storage conditions tested were not found
to be ideal, for physiological alterations were detected that directly affected seed
germination quality.
Key words: water content, germination, vigor, amylases

63
RESUMO – (Atividade amilolítica e qualidade fisiológica de sementes de
Gliricidia sepium (Jacq.) Steud. (Leguminosae Papilionoideae) submetidas ao
armazenamento). O objetivo principal do armazenamento é manter o nível de
qualidade das sementes. É um procedimento muito importante visto que as sementes
estão sujeitas a uma série de mudanças degenerativas depois de dispersas no ambiente.
Este trabalho visou avaliar as mudanças na qualidade fisiológica de sementes de
Gliricidia sepium submetidas ao armazenamento. Sementes de gliricidia recém
coletadas foram analisadas e o restante armazenadas por três e seis meses em sacos de
papel e em temperatura ambiente de laboratório. Ao final dos períodos de
armazenamento foram analisados a taxa de germinação, o teor de água das sementes e
atividade amilolítica durante a embebição. As sementes armazenadas por seis meses
mostraram redução no vigor que foi correlacionado com o aumento no teor de água e
com a alteração da atividade enzimática durante esse período. As condições de
armazenamento das sementes de gliricidia não foram ideais, uma vez que foram
detectadas alterações fisiológicas que refletiram diretamente na qualidade germinativa
das sementes.
Palavraschave: teor de água, germinação, vigor, amilases

64
INTRODUCTION
Considerable research has been undertaken in recent years that examined the
physiological quality of seeds, and the series of degenerative changes that they suffer in
terms of their biochemistry and their physiology and physical integrity after maturation,
which are all associated with reduced vigor (Alizaga et al., 1990). These degenerative
changes may reflect deficiencies in basic procedures used in seed preservation, such as
storage conditions.
As the principal objective of storage is to maintain high levels of seed quality,
success in that endeavor will be closely associated with the initial quality of the seeds,
their degree of desiccation, and the storage conditions themselves (Carvalho &
Nakagawa, 2000).
In dryland vegetation communities such as the caatinga, which covers most of
northeastern Brazil, seeds are usually subject to prolonged periods of water deficit after
dispersal from the mother plant. As such, native or cultivated species that successfully
inhabit this region have a characteristically high tolerance to drought conditions, and
their seeds can generally survive through the dry season within the soil seed bank.
Gliricidia [Gliricidia sepium (Jacq.) Steud. (Leguminosae Papilionoideae)] is
native to South and Central America, but was recently introduced into northeastern
Brazil. It is of significant commercial and economic interest there due to its multiple
uses as a forage species with high nutritive value, as a source of living fence posts, as
a medicinal plant, and even as an alternative energy source. This species demonstrates
rapid growth, high capacity for regeneration, drought resistance, and is easily
propagated by seeds (Drumond & Carvalho Filho, 1999).

65
As the study of seeds is a common starting point for examining the rational use
of potentially useful plant species, an examination of the physiological processes
affected by storage is of fundamental importance. Seed longevity is greatly influenced
by their storage conditions, and can be measured directly by their germination capacity.
The maintenance of seed viability during storage depends directly on seed water
content, and is reflected by the intensity of the physiological activity of the seeds.
In addition to tissue rehydration that occurs during the first phases of seed
swelling, germination is characterized by the mobilization of accumulated seed
reserves. These reserves are degraded and mobilized in order to promote embryo
development. Starch is one of the many types of seed reserves, and the principal
reserve polymer in legume seeds, and it is metabolized by hydrolytic enzymes, often α
amylase (Buckeridge et al., 2004). This enzyme hydrolyzes the α 1,4 bonds of the
starch molecule, producing various oligosaccharides (Bewley & Black, 1994).
Starch degradation is mediated by enzymes that may or may not be present in
quiescent seeds (Mayer & PoljakoffMayber, 1985). Many preexistent enzymes, such
as the α amylase, can be detected only at low levels in quiescent seeds, and must be re
synthesized during rehydration.
Neves & Moraes (2005) observed that tests that examine the physiological
quality of seeds such as germination, and biochemical analyses such as solubilization,
starch degradation, and hydrolytic enzyme activity, can all serve as measures of the
vigor and viability of seeds that have been submitted to different storage periods.
The present work evaluates the changes in the physiological quality of stored
Gliricidia sepium seeds and evaluates their viability and vigor by analyzing amylase
enzyme activity during germination.

66
MATERIALS AND METHODS
Obtaining and storing seeds
Mature seeds were collected in 2006 from G. sepium trees –in Fazenda
Morrinhos, a farm located in the county of Queimadas, state of Bahia (W 39º 40 ׳ S 11º
which ,( ׳ 12 is considered ‘Private Reserve of Natural Patrimony’ under the Brazilian
law. The freshly collected seeds were cleaned and one subsample was immediately
evaluated while the remaining seeds were placed in paper bags and stored at room
temperature (25°C ± 3) for periods of from three and six months, being newly evaluated
at the end of each of these time periods. All experiments were undertaken at the
Germination Laboratory of the Unidade Experimental Horto Florestal, UEFS, Feira de
Santana, Bahia, Brazil.
Water content
Seed water content was determined by drying them in an oven at 60°C until
obtaining a constant weight, which was then subtracted from the previously determined
fresh weight. Twentyfive seeds were used in each subsample.
Germination tests
After the passing of each experimental storage period, four subsamples of 25
seeds were sewn onto Petri dishes containing two layers of germination paper that had
been moistened with a quantity of distilled water that corresponded to 2.5 times the
weight of the paper substrate. The seeds were maintained in a germination chamber at
25°C under a photoperiod of 14 h for a period of ten days. The germinating seeds were
examined daily and individuals with the radicule protruding at least 2 mm from the seed

67
coat were considered to be germinating, and these seeds were counted and subsequently
discarded.
Enzyme extraction and analysis of enzymatic activity
Enzyme extraction and the subsequent analysis of enzymatic activity were
undertaken at the Seed Analysis Laboratory of Embrapa SemiÁrido, in Petrolina,
Pernambuco State, Brazil. Alphaamylase and total amylase activity were evaluated in
recently collected seeds and in seeds subjected to different storage periods. Seed lots
were divided into four subsamples and one subsample was immediately frozen at
20ºC; the other three subsamples were incubated in a seed germinator at 25ºC for 2, 4,
and 7 days. At the end of these time periods, the cotyledons of the germinating seeds
were removed, without touching the seeds, and stored at 20ºC for later enzymatic
extraction.
Extraction and assay of the amylase enzymes were performed according to the
methods described by Gugelmineti et al. (1995). The cotyledons (1 g) were macerated
in 0.1 mol.L 1 pH 7TRISHCl buffer containing 0.1 mol.L 1 NaCl and 10 mmol.L 1
CaCl2 using a mortal and pestle. The resulting slurry was centrifuged at 12,000 g for 10
minutes at 4 o C. All supernatants were collected and stored at 20 o C until the enzyme
assays were performed. Total amylase and αamylase activity were measured in a
reaction system composed of a reaction buffer (50 mmol.L 1 sodium acetate pH 5.2 and
10 mmol.L 1 CaCl2), with a substrate of soluble potato starch (2.5%). The reaction
system was divided into two parts, and the first portion was incubated at 35 o C for 15
minutes. To inactivate the other amylases (leaving only αamylase), the second portion
of the crude extract was maintained at 70 o C for 15 minutes before initiating enzymatic
analysis. At the end of the reaction periods, free reducing sugars were quantified by the

68
dinitrosalicylic acid (DNS) method using a spectrophotometer (540nm). Total amylase
and αamylase activities were expressed in mmol of reducing sugar produced by the
degradation of starch per g of fresh material per minute.
Statistical analyses
Data related to water content and germination was expressed in percentages, and
their standard errors were calculated. Data of enzymatic activity was submitted to
variance analysis using the Sisvar 4.3 software program (Ferreira, 2000). Differences
between the averages were compared using the Tukey test at a 5% probability level.
The significant model was selected to adjust the regression equations because it
presented the greatest coefficient of determination.

69
RESULTS
Increasing storage time significantly altered enzymatic activity (Table 1). No
significant differences were observed during the period of seed swelling, although a
significant interaction between the two parameters was observed.
The average results of the evaluation of the physiological quality of gliricidia
seeds stored in paper sacks at room temperature are shown in Figure 1.
Seed water content increased during storage, going from an initial value of 6.5%
to 13.5% by the sixth month of storage (Figure 1). These stored seeds demonstrated a
14% reduction in their germination rate over the same time period. This reduction in
seed vigor was significant after six months of storage, and coincided with the period in
which they demonstrated an increase in water content.
Table 1. Variance analysis of total amylase (AT) and αamylase (αA) activity in
Gliricidia sepium seeds during storage.
ns = not significant; * = significant at a 5% probability level
Source of Var iation AT αA
Storage 25.492 * 3.994 *
Imbibition 0.592 ns 0.512 ns
Storage*Imbibition 3.316 * 6.483 *
CV % 9.68 14.10

70
Enzymatic activity during swelling and germination of the gliricidia seeds
followed the same pattern for total amylase activity. The αamylase activity varied
according to the duration of seed storage (Figure 2A,B). In general, it was observed
that total amylase activity of the dry seeds was similar over the entire period of seed
storage (Figure 2B); αamylase activity, however, increased in the third and sixth month
of storage (Figure 2A). This initial increase can be attributed to the effects of the
hydration of the seed tissues (Figure 1) that would cause alterations in seed metabolism
during storage.
Figure 1. Sugar content and percentage of total germination of G. sepium seeds stored
under environmental conditions is paper bags (SPB). Average of four trials ± SE
(Standard Error).
0 2 4 6 8 10 12 14 16
0 3 6
Storage time (m)
Water content (%
)
50 55 60 65 70 75 80 85 90 95 100
Germination (%
)
WC % G

71
Figure 2. Amylase activity in the cotyledons of stored G. sepium seeds during
germination.
0
2.000
4.000
6.000
8.000
10.000
0 2 4 6 8
Activity of α
mylase (µ
mol
RS/g.FW
/min)
0 m (y1) 3 m (y2) 6 m (y3) A
y1= 337.19x + 6312.4 R 2 = 0.6631 y2= 80.983x 2 560.56x + 7682.4 R 2 = 0.3924 y3= 187.32x + 8656.5 R 2 = 0.7889
0
5.000
10.000
15.000
20.000
25.000
30.000
35.000
0 2 4 6 8 Imbibition time (Days)
Activity of total amylase (µ
mol
RS/g.FW
/min)
B
y1= 127.88x + 24230 R 2 = 0.1644 y2= 419.91x + 24948 R 2 = 0.7355 y3= 261.28x 2 + 2114.3x + 25726 R 2 = 0.8185

72
During the swelling of recently colleted gliricidia seeds the levels of αamylase
increased, demonstrating a peak of activity on the seventh day (Figure 2A). Total
amylase activity was little altered during swelling, remaining at high levels; and this
enzymatic activity is correlated with the high germination levels observed in these fresh
seeds (Figure 3).
During the swelling of gliricidia seeds stored for three months, an initial
reduction in αamylase activity was observed (≅ 2,000 µmol RS/g. FW/min), and after
two days of swelling activity levels increased and then remained relatively constant
(Figure 2A). Although there was a reduction in enzymatic activity, germination levels
remained high among threemonth old seeds (Figure 3). Germination levels remained
high even after the increase in enzymatic activity in the other periods of swelling.
0
20
40
60
80
100
120
2 4 7
Imbibition Time (Days)
Germination (%
)
0 M 3M 6M
Figure 3. Germination percentage of stored G. sepium seeds during swelling. Average
of four trials ± SE (Standard Error).

73
The differences seen in germination rates after six months can explain the
patterns of enzymatic activity observed for gliricidia seeds. Note that seeds required
longer swelling period after six months of storage in order to germinate (Figure 3).
Storage for six months altered αamylase activity levels and these remained at a
constant level until the fourth day of swelling, when there was a further reduction,
which reflected the low observed germination levels (Figure 2A). Total amylase levels
during seed swelling were higher in seeds stored for six months than those observed in
the other storage treatment regimes (Figure 2B).
The observed tendency of increasing levels of overall amylase activity during
swelling over time may represent a mechanism for compensating for the loss of vigor
caused by longer periods of storage.

74
DISCUSSION
The preservation of seed physiological quality under any given conditions of
temperature and relative humidity is greatly influenced by the type of packaging
material utilized (Ferreira & Borghetti, 2004). The use of paper bags for storing
gliricidia seeds permitted an increase in their water content over time. Maluf &
PisciottanoEreio (2005) reported that Campomanesia phaea seeds stored under similar
conditions lost their viability and, as such, this storage technique would not be capable
of conserving seeds for long periods of time.
High water content is one of the principal causes of the loss of germination vigor
during storage (Villela & Perez, 2004). High seed humidity causes an increase in the
seed’s respiration rate, mobilizes energy reserves, and accelerates seed deterioration.
The energy liberated in these processes will detract from the stored reserves required to
initiate germination (Marcos Filho, 2005).
An increase in water content in rubber tree seeds likewise initiated seed
deterioration (De Paula et al., 1998). The principal alterations documented for seed
deterioration include the exhaustion of food reserves, alteration of the chemical
composition of seed components, such as the oxidation of lipidic components, the
partial degradation of proteins, the alteration of cell membranes, which results in the
reduction of their integrity and organization, with a concomitant increase in their
permeability, as well as enzymatic alterations, and modifications of the nucleotides
(Ferreira & Borghetti, 2004).
As enzymatic activity is required for germination, any alteration of seed
metabolism caused by an increase in its water content will be deleterious to seed

75
survival as can be observed in the reduced vigor of gliricidia seeds over time (Figure
2).
Alphaamylase demonstrates high activity levels at the start of the germination
process, but its enzymatic activity reduces over time. This behavior can be attributed to
the decrease in starch levels as germination proceeds (Bewley & Black, 1994). Aragão
et. al. (2003) observed this same pattern of amylase activity in sweet corn seeds. Many
studies have correlated a decrease in amylase activity with a loss of seed viability, and
decreasing amylase activity has been reported in cereal grains as a consequence of aging
(Muniz et al., 2007).
The present work verified that the decreasing vigor of gliricidia seeds was
mirrored by alterations in their enzymatic activity during storage (Figure 3). Desai et al.
(1997) observed that seed vigor was affected more rapidly than seed viability during
storage.
As the main objective of seed storage is to maintain seed quality, it can be
concluded that the storage conditions employed here were not ideal, as they resulted in
physiological alterations that directly influenced the quality of gliricidia seed
germination.
ACKNOWLEDGEMENTS
The authors would like to thank the Conselho Nacional de Desenvolvimento
Científico e Tecnológico (CNPq), the Fundação de Amparo a Pesquisa do Estado da
Bahia (FAPESB), and Embrapa SemiÁrido Petrolina (PE) for their support in
undertaken this work, as well as José Juracy.

76
REFERENCES
ARAGÃO, C.A.; DANTAS, B.F.; ALVES, E.; CATANEO, A.C.; CAVARIANI, C.;
NAKAGAWA, J. Atividade amilolítica e qualidade fisiológica de sementes
armazenadas de milho super doce tratadas com ácido giberélico. Rev. Bras. de Sem.
25(1): 4348, 2003.
BEWLEY, J.D.; BLACK, M. Seeds: Physiology of Development and Germination.
2 th , ed. Plenum Press, New York, 1994.
CARVALHO, N.M.; NAKAGAWA, A.J. Sementes: ciência, tecnologia e produção.
4 th , Jaboticabal: FUNEP, 588p, 2000.
BUCKERIDGE, M.S.; SANTOS, H.P.; TINÉ, M.A.S. et al. Mobilização de reservas.
In: FERREIRA, A.G.; BORGHETTI, F. (Orgs.) Germinação do básico ao aplicado.
Porto Alegre: ARTMED. Cap. 11, p. 163185, 2004.
DE PAULA, N.F.; BORGES, E.E.L. DE; BORGES, R.C.G. de, DE PAULA, R.C.
Avaliações bioquímicas e fisiológicas em sementes de seringueira (Hevea brasiliensis
Muell. Arg.). Rev. Bras. de Sem., 20(2): 110, 1998.
DESAI, B.B.; KOTHECA, P.M.; SALUNKE, D.K. Seeds handbook: biology,
production, processing and storage. New York: Marcel Dekker, 627p, 1997.
DRUMOND, M.A.; CARVALHO FILHO, O.M. de. Introdução e avaliação de
Gliricidia sepium na região semiárida do Nordeste Brasileiro. In: QUEIRÓZ, M.A. de;
GOEDERT, C.O.; RAMOS, S.R.R. (ed.) Recursos genéticos e melhoramento de
plantas para o Nordeste brasileiro. Versão 1.0, 1999. PetrolinaPE: Embrapa Semi

77
Árido /BrasíliaDF: Embrapa Recursos Genéticos e Biotecnologia, last consulted in
November, 2007. http://www.cpatsa.embrapa.br.
FERREIRA, D.F. Análises Estatísticas por meio do Sisvar para Windows versão
4.0. In. 45° Reunião Anual da Região Brasileira da Sociedade Internacional de
Biometria. UFS Car, São Carlos, SP, last consulted in July, 2007, p. 255258, 2000.
FERREIRA, A.; BORGHETTI, F.(Orgs.). Germinação Do básico ao aplicado. Porto
Alegre, Ed. Artmed, 2004.
GUGELMINETTI, L.; YAMAGUCHI, J.; PERATA, P.; ALPI, A. Amilolytic activities
in cereal seeds under aerobic and anaerobic conditions. Plant Physiology, Rockville,
109(1):10691076, 1995.
MALUF, A.M.; PISCIOTTANOEREIO, W.A. Secagem e armazenamento de sementes
de cambuci. Pesq. Agropec. Bras. 40(7):707714, 2005.
MARCOS FILHO, J. Fisiologia de sementes de plantas cultivadas. Piracicaba: Fealq,
495 p, 2005.
MAYER, A.M.; POLJAKOFFMAYBER, A. The germination of plants. 4 th . Oxford:
Pergamon Press, 270p, 1978.
MUNIZ, F.R.; CARDOSO, M.G.; PINHO, E.V.R.V.; VILELA, M. Qualidade
fisiológica de sementes de milho, feijão, soja e alface na presença de extrato de tiririca.
Rev. Bras. de Sem. 29(2): 195204, 2007.

78
NEVES, L.A.S. das; MORAES, D.M. de. Análise do vigor e da atividade da αamilase
em sementes de cultivares de arroz submetidas a diferentes tratamentos com ácido
acético. Rev. de Ciênc. Agrovet. Lages. 4(1): 3543, 2005.
VILLELA, F.A.; PERES, W.B. Coleta, beneficiamento e armazenamento. In:
FERREIRA, A.G.; BORGHETTI, F. (eds), Germinação: do básico ao aplicado. pp.
265281, Porto Alegre: Artmed, 2004.

79
CAPÍTULO IV
Déficit hídrico em sementes armazenadas de Gliricidia sepium (Jacq.) Steud.
(Leguminosae Papilionoideae).

80
ABSTRACT – (Water deficit of storage seeds Gliricidia sepium (Jacq.) Steud.
(Leguminosae Papilionoideae)) Among the various environmental factors capable of
influencing the germinative process of seeds, the unavailability of water is one of the
most important. In view of the lack of physiological germination information
concerning the of seeds of many native or introduced species of the caatinga, this work
to verify the effect of water deficit on the germination of stored seeds of Gliricidia
sepium, multiple use species, which has been widely used in this region. The seeds used
were packed in paper bags and stored at room temperature for six, nine and twelve
months for further evaluation of the water deficit at the end of each period. For
simulation of the water deficit was used solution of polyethylene glycol (PEG6000) in
different potential osmotic (0.0; 0.2; 0.4; 0.6; 0.8; 1.0; 1.2; 1.4; 1.6; 1.8; 2.0
MPa). Seed germination was influenced by the osmotic conditioning during storage.
The tolerance of gliricidia seeds was between 0.8 and 1.0 MPa when not stored. The
limits of tolerance of stored seeds were influenced by the conditions of storage which
did not promote its conservation of these affecting its response water deficit.
Key Words: vigour, germination, water deficit, storage

81
RESUMO (Déficit hídr ico em sementes armazenadas de Gliricidia sepium (Jacq.)
Steud. (Leguminosae Papilionoideae)). Dentre os diversos fatores ambientais
capazes de influenciar o processo germinativo de sementes, a indisponibilidade de água
é um dos mais importantes. Tendo em vista a carência de informações fisiológicas
concernentes à germinação de sementes de grande parte das espécies nativas ou
introduzidas na caatinga, objetivouse com este trabalho verificar o efeito do déficit
hídrico sobre a germinação de sementes armazenadas de Gliricidia sepium, uma espécie
de uso múltiplo e que vem sendo muito utilizada nessa região. As sementes utilizadas
foram acondicionadas em sacos de papel e armazenadas em temperatura ambiente
durante seis, nove e doze meses, para posterior avaliação do déficit hídrico ao final de
cada período. Para simulação do déficit hídrico foi utilizada solução de polietileno
glicol (PEG6000) em diferentes potenciais osmóticos (0,0; 0,2; 0,4; 0,6; 0,8; 1,0;
1,2; 1,4; 1,6; 1,8; 2,0 MPa) durante a germinação. A porcentagem de germinação foi
influenciada pelo condicionamento osmótico durante o armazenamento. O limite de
tolerância das sementes de gliricidia encontrouse em entre 0,8 e 1,0 MPa quando não
armazenadas. Os limites de tolerância das sementes armazenadas foram influenciados
pelas condições de armazenamento que neste caso não promoveu a conservação destas
afetando na sua resposta à indisponibilidade hídrica do meio.
Palavraschave: vigor, germinação, déficit hídrico, armazenamento.

82
INTRODUÇÃO
O Brasil possui 385 milhões de hectares de florestas nativas (IPEF, 2000), sendo
que a região de Caatinga tem cerca de 800 mil km 2 , totalizando 11% do território
nacional e 70% do território nordestino. Essa comunidade vegetal predominante nessa
região é submetida à escassez de água por períodos prolongados e tendem a ser
dominadas por plantas nativas ou cultivadas que apresentam características de
tolerância à seca e suas sementes sobrevivem a essas condições, armazenadas nos
bancos de solos.
A germinação de sementes é um processo complexo, e seus mecanismos variam
entre espécies ou mesmo entre populações de uma mesma espécie (Khatri et al., 1991).
É considerada como a retomada das atividades metabólicas do eixo embrionário, o qual
se encontrava paralisado nas fases finais do processo de maturação; porém, quando
estimulado por condições ambientais, promove o rompimento do tegumento pela
radícula. Essa é uma etapa crítica do biociclo vegetal pelo fato do processo estar
associado a vários fatores de natureza extrínseca e intrínseca (fatores do ambiente
físico), ou seja, a processos fisiometabólicos (Laboriau, 1983; Popinigis, 1985;
Andrade & DamiãoFilho, 1989; Borges & Rena, 1993; Bewley & Black, 1994).
Dentre os diversos fatores ambientais capazes de influenciar o processo
germinativo de sementes, a indisponibilidade de água é um dos mais importantes. Este
fator abiótico pode ser considerado como limitante à iniciação da germinação de
sementes, assim como está envolvida, direta ou indiretamente em todas as demais
etapas do metabolismo subseqüente, seguindo a ativação do ciclo celular e
conseqüentemente crescimento da plântula.

83
Segundo Borges et al. (2002), a condição hídrica é um dos fatores que exerce
influência decisiva na manutenção do banco de sementes no solo. A indução de
estresses osmóticos vem sendo oportunamente utilizada nas ultimas décadas como
princípio benéfico em aplicações tecnológicas visando à melhoria da qualidade de
sementes e estabelecimento de plântulas referindose ao termo priming ou
envigoramento de muitas espécies cultivadas (Heydecker & Coolbear, 1977; Haigh,
1988; Roberts & Ellis, 1989).
A solução de polietileno glicol (PEG) tem sido usada como meio osmótico para
simular o estresse hídrico que ocorre em campo, segundo Baskin & Baskin (1998), pode
induzir à dormência secundária. Aparentemente, a imposição de estresse osmótico as
sementes pode induzir tolerância a outros tipos de estresses em espécies cultivadas,
incluindo tolerância à seca e a sais (Savino et al., 1979; Kretschmer, 1982; Burgass &
Powell, 1984; Bruggink et al., 1995; Cayuela et al., 1996; Cadman et al., 2006). Além
disso, esse parece ser um fenômeno evolutivo natural em espécies nativas, de adaptação
a condições adversas. Dentro desse enfoque, percebese a importância das pesquisas
relativas a estresses em sementes, como componente essencial e inicial de tolerância à
estresses abióticos que podem ocorrer ao longo do ciclo de vida natural da maioria das
espécies vegetais.
Tendo em vista a carência de informações fisiológicas relativas à germinação de
sementes de grande parte das espécies nativas ou introduzidas na caatinga, o presente
trabalho foi desenvolvido, com o objetivo de verificar o efeito do déficit hídrico sobre a
germinação de sementes armazenadas de Gliricidia sepium, uma espécie de uso
múltiplo e que vem sendo muito utilizada pelo pequeno produtor da caatinga.

84
MATERIAL E MÉTODOS
Obtenção e armazenamento de sementes
Foram utilizadas sementes de G. sepuim provenientes da Reserva do Patrimônio
Natural da Fazenda Morrinhos (QueimadasBA, W 39º ׳ 40 S 11º ( ׳ 12 coletadas em
outubro de 2006. Uma parte das sementes recém coletadas logo após o beneficiamento
foram submetidas ao déficit hídrico. O restante das sementes foram acondicionadas em
sacos de papel e em seguida foram armazenadas em temperatura ambiente (25 °C ± 3),
durante seis, nove e doze meses no Laboratório de Germinação da Unidade
Experimental Horto Florestal (UEFS) – Feira de Santana –BA, para posterior avaliação
do déficit hídrico ao final de cada período.
Avaliação das sementes sobre déficit hídr ico
As condições de déficit hídrico foram testadas em diferentes potenciais
osmóticos na germinação das espécies. Os experimentos foram conduzidos em placas
de Petri, contendo as sementes umedecidas com a solução teste. As sementes
permaneceram em germinador ajustado a temperatura de 25 ºC e fotoperíodo de 14h por
um período de 10 dias.
Para simulação do déficit hídrico foi utilizada solução de polietileno glicol
(PEG6000) em diferentes potenciais osmóticos (0,0; 0,2; 0,4; 0,6; 0,8; 1,0; 1,2;
1,4; 1,6; 1,8; 2,0 MPa), preparadas de acordo com Villela et al. (1991).
A unidade experimental foi composta de quatro repetições de 25 sementes para
cada potencial osmótico testado. Nas avaliações foram consideradas germinadas as
sementes que protrudiram radícula com pelo menos 2 mm de comprimento sendo estas
contadas e posteriormente descartadas.

85
Análise Estatística
Os dados de germinação foram expressos em porcentagem. Os dados de tempo
médio, velocidade média e velocidade de germinação foram transformados pela função
(x + 0,5) 0,5 e analisados mediante análise de variância utilizando o programa Sisvar 4.3
(Ferreira, 2000). As diferenças entre as médias obtidas foram comparadas pelo teste de
Tukey a 5% de probabilidade.

86
RESULTADOS E DISCUSSÃO
A análise de variância dos dados de germinação das sementes de gliricidia
mostrou interação significativa entre o tempo de armazenamento das sementes e o
déficit hídrico. Contudo, não ocorreu influência do armazenamento para o tempo médio
de germinação das sementes (Tabela 1). Os valores de velocidade média e de
germinação sofreram influência significativa para as duas variáveis incidentes
analisadas.
Tabela 1. Análise de variância do tempo médio de germinação (Tm), velocidade média
de germinação (Vm); velocidade de germinação (VG); de sementes armazenadas de
Gliricidia sepium, submetidas a déficit hídrico durante a germinação.
Fonte de var iação Tm, dias Vm, dias 1 VG
Armazenamento 0.799 ns 44.765* 224.606*
Déficit 3.036* 32.739* 120.489*
Armaz.* Déficit 2.214* 8.689* 18.564*
CV% 18.06 3.82 11.16 F= ns; *= não significativo, significativo a 5% respectivamente.
A porcentagem de germinação foi influenciada pelo condicionamento osmótico
durante o armazenamento das sementes. Observouse redução gradual da taxa de
germinação à medida que o potencial osmótico da solução tornavase mais reduzido,
principalmente com o aumento do tempo de armazenamento, ficando evidente a
diferença do grau de sensibilidade das sementes de gliricidia à condição do meio. Na

87
Figura 1 observase que em potenciais inferiores a 1,0 MPa, a geminação das sementes
de gliricidia foi significativamente reduzida, chegando a taxa nula a 1,4 MPa.
Potenciais hídricos reduzidos, especialmente no começo da embebição das
sementes, inviabilizam a seqüência dos eventos germinativos durante a absorção de
água (Torres et al., 1999). O primeiro efeito mensurável do déficit hídrico na
germinação é o impedimento do crescimento, causado pela diminuição da expansão
celular. O processo de alongamento celular e a síntese de parede são altamente sensíveis
ao déficit hídrico (Kramer, 1974 apud Perez & Moraes, 1991).
De acordo com Jeller & Perez (2003), sob condições de déficit hídrico com
polietileno glicol, as sementes de Cassia excelsa apresentaram redução gradativa da
germinabilidade e da velocidade de germinação quando o potencial foi reduzido de 0,2
a 0,8 MPa.
Figura 1. Porcentagem de germinação de sementes recém coletadas e armazenadas de
G. sepium submetidas a déficit hídrico. Média de quatro repetições.
0 10 20 30 40 50 60 70 80 90 100
0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 1.8 2
MPa
0 m 6 m 9 m 12 m
(%) G
erminação

88
Durante o procedimento experimental, as sementes de gliricidia colocadas em
contato com as soluções osmóticas, iniciaram a embebição de água normalmente,
cessando o processo assim que entraram em equilíbrio com o potencial osmótico da
solução externa. Quando o potencial é reduzido, como em 1,4 MPa, a expansão da
radícula de gliricidia foi impedida. As sementes túrgidas mesmo nesses potenciais
osmóticos reduzidos tornamse fisiologicamente ativas, mobilizam reservas e iniciam
seu processo germinativo sem atingir a fase de alongamento celular, mesmo após
algumas semanas de contato com a solução osmótica. Os eventos do processo de
germinação sob condições de priming ou condicionamento osmótico são iniciados
mesmo em conteúdos limitados de água na semente. O processo de germinação não é
completado devido ao requerimento de absorção de água adicional para iniciar a fase III
do processo metabólico, que volta a ocorrer em condições favoráveis (Ferreira &
Borghetti, 2004).
Para as sementes armazenadas durante seis meses, as sementes apresentaram
germinação em potencial osmótico onde previamente não havia germinação (1,2 MPa),
ampliando, portanto, o limite máximo de tolerância ao déficit hídrico simulado com o
PEG 6000. Essa resistência pode ter sido resultado de uma osmorregulação sofrida pelas
sementes durante o período de exposição à solução osmótica ou durante o tempo de
armazenamento.
O déficit hídrico altera a permeabilidade da membrana e as propriedades do
tonoplasto permitindo a interação entre proteínas citoplasmáticas e enzimas
degradativas, ou ainda, aumenta a degradação de proteínas por estimular a síntese de
enzimas proteolíticas (Perez & Moraes, 1991). O acúmulo de solutos no embrião antes
do crescimento radicular diminui o potencial osmótico, gerando pressão de turgescência
suficiente para permitir que o embrião cresça e rompa a barreira do tegumento.

89
Para as sementes armazenadas por doze meses, o armazenamento provocou a
perda do vigor (Figura 1). A taxa germinativa foi significativamente reduzida, mesmo
em condições favoráveis a germinação ou quando as sementes eram colocadas em
déficit hídrico. Considerandose a interação significativa entre os dois fatores, verificou
se que o déficit hídrico contribui muito para a perda de viabilidade das sementes.
A redução do vigor e da viabilidade das sementes de gliricidia pode ter sido
ocasionada também pela grande incidência de fungos observados visualmente
principalmente nas sementes armazenadas por nove e doze meses. A presença de fungos
foi bastante pronunciada quando as mesmas eram expostas aos diferentes potenciais
osmóticos das soluções.
Sabese que durante a hidratação da semente ocorre a liberação de solutos para o
meio circundante tais como, açúcares, ácidos orgânicos, aminoácidos e vários íons, o
que pode estimular o crescimento de patógenos causando a deterioração das sementes
(Bewley & Black, 1994). Ao mesmo tempo, esse acúmulo de solutos nas sementes de
gliricidia ocasionados pelo condicionamento osmótico, pode ter permitido uma possível
osmorregulação das sementes armazenadas por seis meses, proporcionando um aumento
do grau de tolerância ao déficit.
As Tabelas 2 a 4 mostram a diferença entre os valores médios de tempo,
velocidade média e velocidade de germinação para as sementes armazenadas durante o
déficit hídrico.
Com relação aos valores do tempo médio observouse que, com a redução do
potencial hídrico e aumento do período de armazenamento, as sementes de gliricidia
precisaram de um maior tempo para que o processo germinativo ocorresse (Tabela 2).
Sob condições adequadas as sementes requereram 1,490 dias para germinar, enquanto

90
que sob condições de déficit e em potenciais mais reduzidos necessitaram de
aproximadamente 5 dias para que ocorresse a germinação das sementes.
Tabela 2. Tempo médio (Tm) de sementes de Gliricidia sepium recém coletadas e
armazenadas submetidas a déficit hídrico durante a germinação.
Meses
MPa 0 6 9 Média
Tm, (dias)
0,0 1.490 bB 2.715 abA 3.637 aA 2.614 B
0,2 1.562 aB 3.147 aA 3.167 aA 2.625 B
0,4 2.422 aAB 2.792 aA 2.813 aA 2.676 B
0,6 3.567 aAB 3.207 aA 4.950 aA 3.908 AB
0,8 4.713 aA 3.830 aA 4.117 aA 4.220 AB
1,0 4.750 aA 3.795 abA 3.000 bA 3.848 AB
Média 3.074 a 3.433 a 3.614 a Médias seguidas pela mesma letra minúscula na linha e maiúscula na coluna não
diferem entre si pelo teste de Tukey a 5% de probabilidade.
O tempo de armazenamento influenciou de forma significativa nos valores de
velocidade média de germinação das sementes. A velocidade média foi reduzida
principalmente em sementes com nove meses de armazenadas (Tabela 3). Entre os
potenciais osmóticos testados as sementes armazenadas por seis meses não foi
observada diferença significativa para a velocidade média de germinação.
A diminuição do vigor das sementes armazenadas provocou a redução também
da velocidade de germinação (VG) durante o déficit hídrico. Observouse diferença
significativa durante o armazenamento das sementes para os potenciais testados,
principalmente entre 0,0 a 0,4 MPa (Tabela 4). Verificouse uma relação direta entre a

91
redução dos potenciais osmóticos e a germinação das sementes de gliricidia
promovendo um decréscimo acentuado na velocidade de germinação das sementes
armazenadas. O déficit hídrico normalmente diminui a velocidade de germinação e a
porcentagem de germinação, existindo grande variação entre as espécies, desde aquelas
muito sensíveis até as mais resistentes (Bewley & Black, 1994).
Tabela 3. Velocidade média (Vm) de sementes de Gliricidia sepium recém coletadas e
armazenadas submetidas a déficit hídrico durante a germinação.
Meses
MPa 0 6 9 Média
Vm, (dias 1 )
0,0 0.673 aA 0.368 bA 0.280 bAB 0.440 A
0,2 0.650 aA 0.318 bA 0.323 bAB 0.430 A
0,4 0.422 aB 0.360 aA 0.367 aA 0.383 A
0,6 0.283 abC 0.317 aA 0.207 bBC 0.269 B
0,8 0.212 abC 0.260 aA 0.138 bCD 0.203 B
1,0 0.215 aC 0.270 aA 0.095 bD 0.193 B
Média 0.409 a 0.305 b 0.235 c Médias seguidas pela mesma letra minúscula na linha e maiúscula na coluna não
diferem entre si pelo teste de Tukey a 5% de probabilidade.
A presença de diferentes compostos que altera os potenciais osmóticos do meio,
podem nos dar informações valiosas quanto aos limites de tolerância das sementes ao
déficit hídrico. O alto peso molecular do PEG 6000 impede a penetração da substância
através das membranas celulares, podendo reduzir a disponibilidade de oxigênio em
virtude de sua alta viscosidade, afetando o processo germinativo (Bradford, 1995). A
capacidade das sementes de algumas espécies em germinar sob condições de déficit

92
hídrico lhes confere vantagens ecológicas em relação a outras que são sensíveis à seca.
Um dos benefícios do condicionamento osmótico (priming) é a melhoria do
desempenho da semente por ocasionar o seu envigoramento (Ferreira & Borghetti,
2004).
Nas sementes de gliricidia o efeito do condicionamento osmótico não pode ser
verificado nas sementes não germinadas, devido à contaminação por fungos ter
diminuído o vigor das sementes nos potenciais mais negativos. Embora o tratamento do
priming imponha um estresse osmótico (moderado) às sementes, ele permite que
inúmeros processos metabólicos progridam, mas ao mesmo tempo prevenindo a
protrusão radicular. Acreditase que dessa maneira, as sementes de um mesmo lote
possam sincronizar a germinação, resultando na uniformização do processo
germinativo.
Tabela 4. Velocidade de germinação (VG); de sementes de Gliricidia sepium recém
coletadas e armazenadas submetidas a déficit hídrico durante a germinação.
Meses
MPa 0 6 9 Média
VG
0,0 19.708 aA 8.007 bA 5.195 cA 10.970 A
0,2 19.103 aA 6.265 bAB 3.520 AB 9.629 A
0,4 11.768 aB 8.140 bA 1.153 cBC 7.020 B
0,6 6.493 aC 5.207 aB 1.907 bBC 4.536 C
0,8 3.215 aD 4.260 aB 0.563 bC 2.680 D
1,0 0.920 aD 1.767 abC 0.125 bC 0.937 E
Média 10.201 a 4.866 b 2.077 c Médias seguidas pela mesma letra minúscula na linha e maiúscula na coluna não
diferem entre si pelo teste de Tukey a 5% de probabilidade.

93
O uso de sementes de baixa qualidade, aliado à ocorrência de condições
ambientais adversas, podem resultar em baixa porcentagem de germinação e menor
velocidade de germinação. As condições de armazenamento fornecidas às sementes de
gliricidia provocaram redução na qualidade fisiológica destas. Tratamentos como o
priming, tornamse neste caso uma ferramenta de fundamental importância para
aumentar o vigor dessas sementes e obter melhores taxas de germinação, bem como
aumentar a tolerância das sementes às condições adversas durante o processo
germinativo.
Segundo Fanti & Perez (2004) em sementes de Chorisia speciosa (paineira),
houve uma redução da viabilidade das sementes com a redução do potencial osmótico,
sendo o limite máximo de tolerância ao PEG entre 0,6 e 0,7 MPa. Entre as espécies
que também não apresentaram limites elevados de tolerância ao déficit hídrico simulado
com PEG 6000, podese citar Adenanthera pavonina, Leucaena leucocephala (Fonseca
& Perez, 1999) e Senna occidentalis (Delachiave & Pinho, 2003). Nas sementes de
gliricidia esse limite encontrouse em entre 0,8 e 1,0 MPa quando não armazenadas
(Figura 1).
Ribeiro & Pelacani (2006) estudando a influência do déficit hídrico imposto por
manitol verificaram um aumento do tempo médio de germinação apresentado por duas
espécies de leguminosas (Cenostigma gardnerianum e Anadenanthera colubrina),
corroborando com os resultados obtidos em G. sepium para o polietiolenoglicol. Em
condições naturais, esse aumento do tempo para que as sementes germinem poderia
atuar de forma positiva no estabelecimento das plântulas, pois desta forma a germinação
das sementes que permanecem viáveis pode ser distribuída no tempo e no espaço,
aumentando a probabilidade das plântulas encontrarem condições ambientais adequadas
para seu crescimento e desenvolvimento (Bewley & Black, 1994).

94
A longevidade das sementes pode variar de acordo com o genótipo, mas, a
conservação do potencial fisiológico depende, em grande parte, da temperatura, das
condições do ambiente de armazenamento e do grau de umidade das sementes (Ferreira
& Borghetti, 2004; Marcos Filho, 2005). De acordo com os resultados obtidos, os
potenciais em que a germinação parece ser afetada, denominada de limite de tolerância,
foram influenciados pelas condições de armazenamento das sementes de gliricidia.
Baseado na indisponibilidade hídrica do meio, neste estudo verificouse que a
embalagem utilizada para o armazenamento das sementes promoveu a conservação
destas até seis meses de armazenadas.
AGRADECIMENTOS
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), à
Fundação de Amparo a Pesquisa do Estado da Bahia (FAPESB) e a pessoa de José
Juracy pelo suporte de material vegetal.

95
REFERÊNCIAS BIBLIOGRÁFICAS
ANDRADE, V. M. M.; DAMIÃOFILHO, C. F.1989. Morfologia vegetal. Jaboticabal:
FUNEP/UNESP, 259 p.
BASKIN, C.C; BASKIN, J.M. 1998. Seeds, ecology, biogeography, and evolution of
dormancy and germination. Academic Press, New York.
BEWLEY, J. D., BLACK, M. 1994. Seeds: physiology of development and
germination 2.ed. New York. Plenum, 445 p.
BORGES, E. E. L.; RENA, A. B. 1993. Germinação de sementes. In: AGUIAR, I. B.;
PINÃRODRIGUES, F. C. M.; FIGLIOLA, M. B. (Coord..). Sementes florestais
tropicais. Brasílica: ABRATES, 83 p.
BORGES, E.E.L de; PEREZ, S.C.J.G.A.; BORGES, R.C.G de; REZENDE, S.T de;
GARCIA, S.R. 2002. Comportamento fisiológico de sementes osmocondicionadas de
Platymiscium pubescens Micheli (TamborildaMata). Revista Árvore, ViçosaMG,
26(5):603613.
BRADFORD, K. J. 1995.Water relations in seed germination. In: KIGEL, J.; GALILI,
G. (Ed.). Seed devenlopment and germination. New York: Marcel Dekker Inc.,
p.351396.
BRUGGINK, G.T. & VAN DER TOORN, P. 1995. Induction of desiccation tolerance
in germinated seeds. Seed Science Research. 5: 14.
BURGAS, R.W.& POWELL, A.A. 1984. Evidence for repair processes in the
invigoration of seeds by hydration. Annals of Botany. 53: 753757.

96
CADMAN, C.S.C.; TOOROP, P.E.; HILHORST, H.W.M.; FINCHSAVAGE, W.E.
2006. Gene expression profiles of Arabidopsis Cvi seeds during cycling through
dormant and nondormant states indicate a common underlying dormancy control
mechanism. Plant Journal.46: 805822.
CAYUELA, E.; PÉREZALFOCEA, F.; CARO, M.; BOLARÍN, M.C. 1996. Priming
of seeds with NaCl induces physiological changes in tomato plants grown under salt
stress. Physiology Plant. 96: 231236.
DELACHIAVE, M.E.A.; PINHO, S.Z.2003. Germination of Senna occidentalis Link:
seed at different osmotic potential levels. Br azilian Archives of Biology and
Technology, Botucatu, 46(2):163166.
FANTI, S. C.; PEREZ, S. C. J. G. A. 2004. Processo germinativo de sementes de
paineira sob estresses hídrico e salino. Pesquisa Agropecuár ia Brasileira, Brasília,
39(9): 903909, set.
FERREIRA, D.F. 2000. Análises Estatísticas por meio do Sisvar para Windows
ver são 4.0. In. 45° Reunião Anual da Região Brasileira da Sociedade Internacional de
Biometria. UFS Car, São Carlos, SP, Julho, p. 255258.
FERREIRA, A.G.; BORGHETTI, F.(Orgs.). 2004. Germinação Do básico ao
aplicado. Porto Alegre, Ed. Artmed.
FONSECA, A.G.; PEREZ, S.C.J. G. A. 1999. Efeito do estresse hídrico e interferência
de diferentes profundidades de plantio na germinação de sementes de Leucaena
leucocephala (Lam) de wit. Revista Ceres, 46: 371381.

97
FONSECA, S. C. L.; PEREZ, S. C. J. G. A. 2003. Ação do polietileno glicol na
germinação de sementes de Adenanthera pavonina L. e o uso de poliaminas na
atenuação do estresse hídrico sob diferentes temperaturas. Revista Brasileira de
Sementes, 25(1):16.
HAIGH, A.M. 1988. Why do tomato seed pr ime? PhD thesis. Macquarie University,
Sydney.
HEYDECKER, W. & COOLBEAR, P. 1977. Seed treatments for improved
performancesurvey and attempted prognosis. Seed Science & Technology, 5: 353–
425.
IPEF. 2000. Reflorestamento. http.www.ipef.com.br. Acesso em: 12 de janeiro de 2007.
JELLER, H. & PEREZ, S. C. J. de A. 2003. Condicionamento osmótico na germinação
de sementes de cássia do nordeste sob estresse hídrico, térmico e salino. Pesquisa
Agropecuária Brasileira, Brasília, 38(9):10251034.
KHATRI, R.; SETHI, V.; KAUSHIK, A. 1991. Interpopulation variations of Kochia
indica during germination under different stress. Annals of Botany, 67: 413415.
KRETSCHMER, M. 1982. Extension of the temperature tolerance of Lactuca achenes
with PEG and red light. Gartenbauwissenschaft, 47: 152157.
LABOURIAU, L. G. 1983. A germinação da semente. Washington: OEA, 173 p.
MARCOS FILHO, J. 2005. Fisiologia de sementes de plantas cultivadas. Piracicaba:
Fealq, 495 p.

98
PEREZ, S. C. J. G. de A. & MORAES, J. A. P. V. 1991. Influência do estresse hídrico e
do pH no processo germinativo da algarobeira. Pesquisa Agropecuár ia Brasileira,
Brasília, 26(7): 981988.
POPINIGIS, F. 1985. Fisiologia da semente. Brasília: AGIPLAN, 289 p.
RIBEIRO, R.C.; PELACANI, C.R. 2006. Influência do manitol e NaCl na germinação
de sementes de duas espécies de leguminosas com importância no semiárido baiano.
Revista StientibusSérie Ciências Biológicas, 6(2): 105109.
ROBERTS, E.H. & ELLIS, R.H. 1989. Water and seed survival. Annals of Botany. 63:
39–52.
SAVINO G. et al. 1979. Effects of presoaking upon seed vigor and viability during
storage. Seed Science & Technology. 7: 5764.
TORRES, S.B.; VIERA, E.L.; MARCOSFILHO, J. 1999. Efeitos do estresse hídrico
na germinação e no desenvolvimento de plântulas de pepino. Revista Brasileira de
Sementes. 21: 5963.
VILLELA, F.A.; DONI FILHO, L.; SEQUEIRA, E.L. 1991. Tabela de potencial
osmótico em função da concentração de polietileno glicol 6000 e ta temperatura.
Pesquisa Agropecuária Brasileira. Brasília, 26(11/12): 19571968, nov./dez.

99
CONSIDERAÇÕES FINAIS
Os resultados obtidos no presente trabalho permitiram concluir que:
• A manutenção da viabilidade das sementes de Gliricidia sepium durante o
armazenamento depende da manutenção do teor de água inicial que pode ser
alcançado através do tipo adequado de armazenamento das sementes;
• Embora ambientes com temperaturas mais baixas sejam mais adequados para a
conservação das sementes, em temperaturas em torno de 25°C é possível
prolongar a viabilidade das sementes de gliricidia quando se controla o teor de
água inicial das mesmas através da utilização de embalagens impermeáveis para
o armazenamento;
• A variação nos teores de macromoléculas está associada com a perda da
viabilidade das sementes, proporcionada pela condição inadequada de
armazenamento;
• A tolerância da espécie ao déficit hídrico durante a germinação foi influenciada
pelo tipo de armazenamento inadequado das sementes;
• As sementes armazenadas adequadamente apresentaram boa longevidade,
evidenciando seu uso potencial pelo pequeno agricultor da caatinga, ou em
programas de recomposição ambiental.

100
REFERÊNCIAS BIBLIOGRÁFICAS CITADAS NA INTRODUÇÃO GERAL
ANDRADE, V. M. M.; DAMIÃOFILHO, C. F. Morfologia vegetal. Jaboticabal:
FUNEP/UNESP, 259 p. 1989.
BASKIN, C.C.; BASKIN, J.M. Seeds, ecology, biogeography, and evolution of
dormancy and germination. Academic Press, New York, 1998.
BEWLEY, J.D.; BLACK, M. Seeds: Physiology of Development and Germination.
Second Edition. Plenum Press, New York, 445p, 1994.
BIANCHETTI, A. Tratamentos prégerminativo para sementes florestais. In:
SIMPÓSIO BRASILIERO SOBRE TECNOLOGIA DE SEMENTES FLORESTAIS, 2.
1989, Atibaia. Anais... São Paulo: Instituto Florestal, p. 237246, 1991.
BORGES, E. E. L.; RENA, A. B. Germinação de sementes. In: AGUIAR, I. B.; PINÃ
RODRIGUES, F. C. M.; FIGLIOLA, M. B. (Coord..). Sementes florestais tropicais.
Brasílica: ABRATES, 83 p. 1993.
CABRAL, E.L.; BARBOSA, D.C.A.; SIMABUKURO, E.A. Armazenamento e
germinação de sementes de Tabebuia áurea (Manso) Benth. & Hook. F. Ex. S. Moore.
Acta. Bot. Bras. 17(4): 609617, 2003.
CARNEIRO, J.G.A. Armazenamento de sementes florestais. curitiba: FUPEF, 35 p.
(Série Técnica, 14), 1985.
CARNEIRO, J.G.A. de; AGUIAR, I.B. de. Armazenamento de Sementes. In: AGUIAR,
I.B. de; PINÃRODRIGUES, F.C.M.; FLIGLIOLIA, M.B. Sementes Florestais
tropicais. 350p. 1993.
DESAI, B.B.; KOTECHA, P.M.; SALUNKE, D.K. Seeds handbook: biology,
production, processing and storage. New York: Marcel Dekker, 627p. 1997.
DRUMOND, M.A., CARVALHO FILHO, O.M. de. Introdução e avaliação de Gliricidia sepium na região semiárida do Nordeste Brasileiro. In: QUEIRÓZ, M.A. de,
GOEDERT, C.O., AMOS, S.R.R., (ed.) Recursos genéticos e melhoramento de

101
plantas para o Nordeste brasileiro. (on line). Versão 1.0. PetrolinaPE: Embrapa
SemiÁrido /BrasíliaDF: Embrapa Recursos Genéticos e Biotecnologia, nov. 1999.
Disponível via Word Wide Web http://www.cpatsa.embrapa.br. SBN 8574050016.
DRUMOND, M.A.; PIEDADEKILL, L.H.; LIMA, P.C.F.; OLIVEIRA, M.C.;
OLIVEIRA, V.R.; ALBUQUERQUE, S.G.; NASCIMENTO, C.E.S.; CAVALCANTI,
J. Estratégias para o uso sustentável da biodiversidade da caatinga. Avaliação e
identificação de ações pr ioritár ias para a conser vação, utilização sustentável e
repar tição de benefícios da biodiver sidade do bioma caatinga. In: Biodiversidade da
caatinga, http//www.biodiversitas.org/caatinga, Petrolina, 2000.
DUQUE, J.A. Gliricidia sepium (Jacq.) Steud. Disponível: site Hor ticulture Depar tment at Purdue web. URL: http://www.hort.parde.edu/newcrop/duke _
energy/Gliricidia_ sepium palavrachave: Gliricidia sepium. Consultado em 7 de outubro de 2006.
FERREIRA, A.G.; BORGHETTI, F. (Orgs.). GerminaçãoDo básico ao aplicado.
Porto Alegre, Ed. Artmed, 2004.
FONSECA, S. C. L.; PEREZ, S. C. J. G. A. Ação do polietileno glicol na germinação
de sementes de Adenanthera pavonina L. e o uso de poliaminas na atenuação do estresse
hídrico sob diferentes temperaturas. Revista Brasileira de Sementes, 25(1):16, 2003.
FRANCO, A.A. Uso de Gliricidia sepium como moirão vivo. Rio de Janeiro:
EMBRAPAUAPNPBS, 5p. (EMBRAPAUAPNPBS. Comunicado Técnico, 3), 1988.
HARRINGTON, J. F. Seed storage and longevity. In: KOZLOWSKI, T. T. Seed
biology. New York: Academic, 3:145245, 1972.
IPEF. Reflorestamento. http.www.ipef.com.br. Acesso em: 12 de janeiro de 2000.
LABOURIAU, L. G. A germinação da semente. Washington: OEA, 173 p, 1983.
LIMA, J.L.S. Plantas for rageiras das caatingas: usos e potencialidades. Petrolina:
Embrapa SemiArido/ PNE/ RBGKEW , 44p, 1996.

102
MAIA, G.N. Caatinga: árvores e arbustos e suas utilidades. 1° ed. São Paulo. Editora
D & Z, 2004.
MACHADO, C.F. Metodologia para a condução do teste de germinação e utilização
de raiosX para a avaliação da qualidadde de sementes de aroreirabranca (Lithrae molleoides (Vell.) Engl.). Dissertação (Mestrado em Fitotecnia)Escola Superior de
Agriculuta Luiz de Queiroz, Piracicaba, SP. 51p. 2002.
MARCOS FILHO, J. Germinação de sementes. In: Marcos Filho, J. (Ed.). Atualização
em produção de sementes. Campinas: Fundação Cargill, p.1139, 1986.
MARCOS FILHO, J. Fisiologia de sementes de plantas cultivadas. Piracicaba: Fealq,
495 p., 2005.
MATTHEWS, S. Physiology of seed ageing. Outlook on Agr iculture, 14(2): 1923,
1985.
NATIONAL ACADEMY SCIENCES (Washington). Firewood crops: shrub and tree
species for energy production. Washington, 237p. 1980.
NODARI, R.O. et al. Conservação de frutos e sementes de palmiteiro (Euterpe edulis Matius) sob diferentes condições de armazenamento. Rev. Árvore, 22(1): 110, 1998.
OLIVEIRA, L. M. Avaliação da qualidade de sementes de Tabebuia serratifolia Vahl. Nich. E Tabebuia impetiginosa (Martius Ex A. P. De Candolle Standley)
envelhecidas natural e ar tificialmente. Tese (Doutorado em Agronomia. Fitotecnia)
Universidade Federal de Lavras, Lavras MG. 160 p. 2004.
PARROTTA, A.J. Gliricidia sepium (Jacq.) Walp. Glir icidia, mother of cocoa
Leguminosae (Papilionoideae) Legume family. Rio Piedras : Institute of Tropical
Forestry, 7p. (SOITFSM50), 1992.
POPINIGIS, F. Fisiologia da semente. Brasília: AGIPLAN, 289 p. 1985.
ROBERTS, E. H. P redicting the storage life of seeds Seed Science & Technology,
London, 1(1):499514, Jan. 1973.

103
ROCHA, G. R. Efeito da temperatura e do potencial hídrico na germinação de
sementes de doze cultivares de Feijão Mungo Verde [Vigna radiata (L.) Wilczek].
JaboticabalSP. Trabalho apresentado à Faculdade de Ciências Agrárias e Veterinárias
UNESP para graduação em Agronomia, 1996.
SALES, J. de F. Atividade da celulase sobre o processo germinativo de sementes de
cafeeiro (Coffea arabica L.). Dissertação (Mestrado em Agronomia. Fisiologia
Vegetal)Universidade Federal de Lavras, Lavras, MG. 38 p. 2002.
SAMPAIO, & RODAL, M.J. Fitofisionomias da caatinga. Avaliação e identificação de
ações pr ioritár ias para a conservação, utilização sustentável e repartição de
benefícios da biodiversidade do bioma caatinga. In: Biodiversidade da caatinga,
http//www.biodiversitas.org/caatinga ,Petrolina, 2000.
SAMPAIO, E.V.S.B. Overview of Brazilian caatinga. In: MOONEY, H.A.;
BULLOCK, S.H.; MEDINA, E. (eds.) Dry Tropical forests. Cambridge University
Press, Cambridge. p. 3563, 1995.
SANTOS, S. R. G. dos. Efeito da temperatura na germinação de sementes de Sebastiana commersoniana (Baill.) Smith e Downs (Branquilho). Dissertação (Mestrado em Agronomia. Produção e Tecnologia de Sementes) Univerisade Estadual
Paulista, Jaboticabal, SP. 76 p. 1999.
SUMBERG, J.E. Note on flowering and seed production in a yong Gliricidia sepium
seed orchard. Tropical Agriculture, Trinidad, 62(1):1724, 1985.
VARELA, V. P.; COSTA, S. de S.; RAMOS, M. B. P. Influência da temperatura e do
substrato na germinação de sementes de itaubarana (Acosmium nitens (Vog.) Yakovlev)
Leguminosae, Caesalpinoideae. Acta Amazônica, 35(1):35 – 39. 2005.
WARD, F.H.; POWELL, A.A. Evidence for repair processes in onion seeds during
storage at high seed moisture contents. J Exp. Bot. 34(140):277282, 1983.

104
RESUMO GERAL
Efeito do armazenamento na qualidade fisiológica de sementes de Gliricidia sepium
(Jacq.) Steud. (Leguminosae Papilionoideae). Com a crescente demanda por
informações sobre espécies nativas ou introduzidas, visando sua utilização para os mais
diversos fins, tornase fundamental o conhecimento fisiológico da germinação, levando
em consideração que os estudos com sementes são efetivamente o ponto de partida para
a utilização e exploração, de forma racional, dessas espécies potenciais. Objetivouse,
com esta pesquisa, avaliar a germinabilidade e longevidade de sementes de Gliricidia
sepium submetidas a diferentes períodos de armazenamento, visando o ajuste de
protocolos para conservação de sementes, da espécie, e sua utilização pelo pequeno
produtor da Caatinga. A manutenção da viabilidade das sementes durante o
armazenamento dependeu, diretamente, do teor de água encontrado nas unidades de
dispersão. Esta dependência esteve relacionada à intensidade das reações químicas e
metabólicas que caracterizam a atividade fisiológica das sementes, refletindo
diretamente no seu desempenho germinativo e na sua capacidade de tolerância ao déficit
hídrico imposto durante o armazenamento. As melhores condições de conservação das
sementes foram obtidas com o armazenamento em saco plástico, principalmente quando
acondicionadas em temperaturas baixas. No entanto, mesmo em temperaturas em torno
de 25°C é possível prolongar a viabilidade das sementes de gliricidia quando se controla
o teor de água inicial das mesmas através da utilização de embalagens impermeáveis
para o armazenamento.

105
ABSTRACT OF DISSERTATION
Effect of storage on the physiological quality of seeds of Gliricidia sepium (Jacq.)
Steud. (Leguminosae Papilionoideae). With the growing demand for information on
native or introduced species, targeting their use for many different purposes, it is
essential the physiological knowledge of germination, taking into account that studies
with seeds are actually the starting point for the use and exploitation , in a rational way,
of these species potential. The objective is, with this search, evaluate the germination of
seeds and longevity of Gliricidia sepium subjected to different periods of storage,
seeking to adjust protocols for conservation of seeds, the species, and its use by small
producer of the caatinga. Maintaining the viability of the seed during storage depended,
directly, the level of water found in the dispersion units. This dependence was related to
the intensity of the chemical reactions that characterize the metabolic and physiological
activity of seed, reflecting directly into their performance germination and its ability to
tolerance to water deficit imposed during storage. The best conditions for the
conservation of seeds were obtained with the storage in plastic bag, mainly when they
were stored at low temperatures. However, even at temperatures near 25° C it was
possible to prolong seed viability when their water content was controlled through the
use of impermeable packing.